Embed Size (px)
Citation preview

1
BIOCHEMISCHES
PRAKTIKUM
für Chemiker und
Wirtschaftsingenieure
Fassung vom September 2012
Universität Kaiserslautern
Fachbereich Chemie
Prof. Dr. W. E. Trommer

2
Inhaltsverzeichnis
Zeitplan zum Biochemie-Praktikum für Chemiker und WI SS 2012 ................................... 3
Anmerkungen zum Praktikum............................................................................................ 4
Enzyme-Linked Immunosorbent Assay (ELISA) ................................................................ 5
Enzymatische Bestimmung von ADP (Adenosindiphosphat) ........................................ 15
Disk-Elektrophorese in Natriumdodecylsulfat (SDS-PAGE) ............................................. 19
Quantitative Proteinbestimmungsmethoden .................................................................... 28
Bestimmung der katalytischen Konstanten und der Michaelis-Konstanten von
Chymotrypsin für N-Succinyl-alanyl-alanyl-prolyl-phenylalanin-4-nitroanilid (Suc-AAPF-
pNA) ........................................................................................................................... 35
3
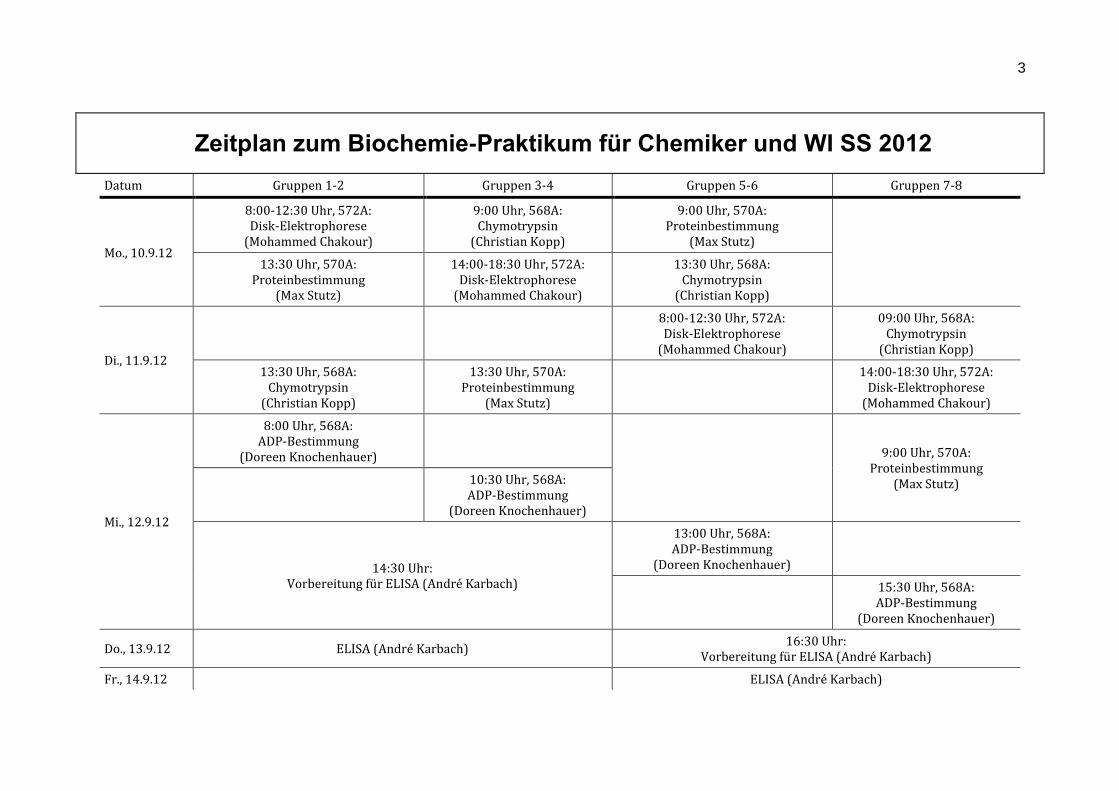
Zeitplan zum Biochemie-Praktikum für Chemiker und WI SS 2012
Datum Gruppen 1-2 Gruppen 3-4 Gruppen 5-6 Gruppen 7-8
Mo., 10.9.12
8:00-12:30 Uhr, 572A: Disk-Elektrophorese
(Mohammed Chakour)
9:00 Uhr, 568A: Chymotrypsin
(Christian Kopp)
9:00 Uhr, 570A: Proteinbestimmung
(Max Stutz)
13:30 Uhr, 570A: Proteinbestimmung
(Max Stutz)
14:00-18:30 Uhr, 572A: Disk-Elektrophorese
(Mohammed Chakour)
13:30 Uhr, 568A: Chymotrypsin
(Christian Kopp)
Di., 11.9.12
8:00-12:30 Uhr, 572A: Disk-Elektrophorese
(Mohammed Chakour)
09:00 Uhr, 568A: Chymotrypsin
(Christian Kopp)
13:30 Uhr, 568A: Chymotrypsin
(Christian Kopp)
13:30 Uhr, 570A: Proteinbestimmung
(Max Stutz)
14:00-18:30 Uhr, 572A: Disk-Elektrophorese
(Mohammed Chakour)
Mi., 12.9.12
8:00 Uhr, 568A: ADP-Bestimmung
(Doreen Knochenhauer)
9:00 Uhr, 570A:
Proteinbestimmung (Max Stutz)
10:30 Uhr, 568A: ADP-Bestimmung
(Doreen Knochenhauer)
14:30 Uhr: Vorbereitung für ELISA (André Karbach)
13:00 Uhr, 568A: ADP-Bestimmung
(Doreen Knochenhauer)
15:30 Uhr, 568A: ADP-Bestimmung
(Doreen Knochenhauer)
Do., 13.9.12 ELISA (André Karbach) 16:30 Uhr:
Vorbereitung für ELISA (André Karbach)
Fr., 14.9.12 ELISA (André Karbach)

4
Anmerkungen zum Praktikum
Protokolle zu den einzelnen Versuchen sind spätestens 1 Woche nach
Beendigung des Versuchs beim Versuchsassistenten / der Versuchsassistentin
abzugeben.
Zulassung zur Praktikumsklausur bzw. Ausstellung des Scheins nur nach
erfolgreicher Abgabe und evtl. Korrektur aller Protokolle.
Alle verwendeten Messwerte etc. sind im jeweiligen Protokoll anzugeben.
Rechenwege sind nachvollziehbar darzulegen.
Protokolle sind in Papierform abzugeben.
Sollten Sicherheitsregeln im Labor nicht befolgt werden, können Praktikant(inn)en
vom Praktikum oder dem Versuchstag ausgeschlossen werden

5
Enzyme-Linked Immunosorbent Assay (ELISA)
1. Einleitung ................................................................................................................... 5
2. Versuchsbeschreibung ............................................................................................. 10
3. Versuchsdurchführung ............................................................................................. 11
3.1 Reagenzien und Chemikalien ............................................................................. 11
3.2 Versuchsablauf ................................................................................................... 12
3.3 Bestückung der Mikrotiterplatte ........................................................................... 13
4. Auswertung .............................................................................................................. 14
5. Literatur .................................................................................................................... 14
1. Einleitung
Die intakte Oberfläche des Körpers stellt eine wirksame Barriere gegenüber den meisten
Mikroorganismen (Viren, Bakterien, Pilzen, Parasiten) dar. Um dennoch eingedrungene
Mikroben unschädlich zu machen, verfügen Wirbeltiere über ein Abwehrsystem aus
Molekülen und Zellen, das Immunsystem.
Alle Zellen des Immunsystems stammen von pluripotenten Stammzellen ab, die sich im
Laufe ihrer Entwicklung in zwei Zellinien, die myeloische und die lymphatische,
differenzieren. Die myeloische Reihe besteht zum gößten Teil aus Phagozyten, welche in
der Lage sind, Fremdorganismen zu endocytieren und zu verdauen. Die Zellen der
lymphatischen Reihe (Lymphocyten) differenzieren, je nach Umgebung (micro-
environment) in der sie heranreifen, in T- und B-Zellen. T-Zellen entwickeln sich dabei im
Thymus, während B-Zellen bei Säugetieren im Knochenmark (bone marrow) entstehen.

6
Neben der zellvermittelten Immunreaktion spielen auch lösliche Faktoren bei der
Immunantwort eine große Rolle (humorale Immunantwort). Den Hauptträger dieser
Abwehr bilden die Immunglobuline oder Antikörper. Diese werden - nach einem Kontakt
des lymphatischen Systems mit fremden immunogenen Molekülen - von Plasmazellen,
die sich aus B-Zellen entwickeln, gebildet. Moleküle, die im Organismus die Bildung
dieser Antikörper induzieren, nennt man Antigene.
Die Antikörper erkennen das infektiöse Agens spezifisch, das heißt, ein bestimmter
Antikörper erkennt nur sein zugehöriges Antigen. Die Bindung erfolgt dabei nicht an das
gesamte Antigen, sondern nur an eine bestimmte Stelle, die Antigendeterminante
(Epitop).
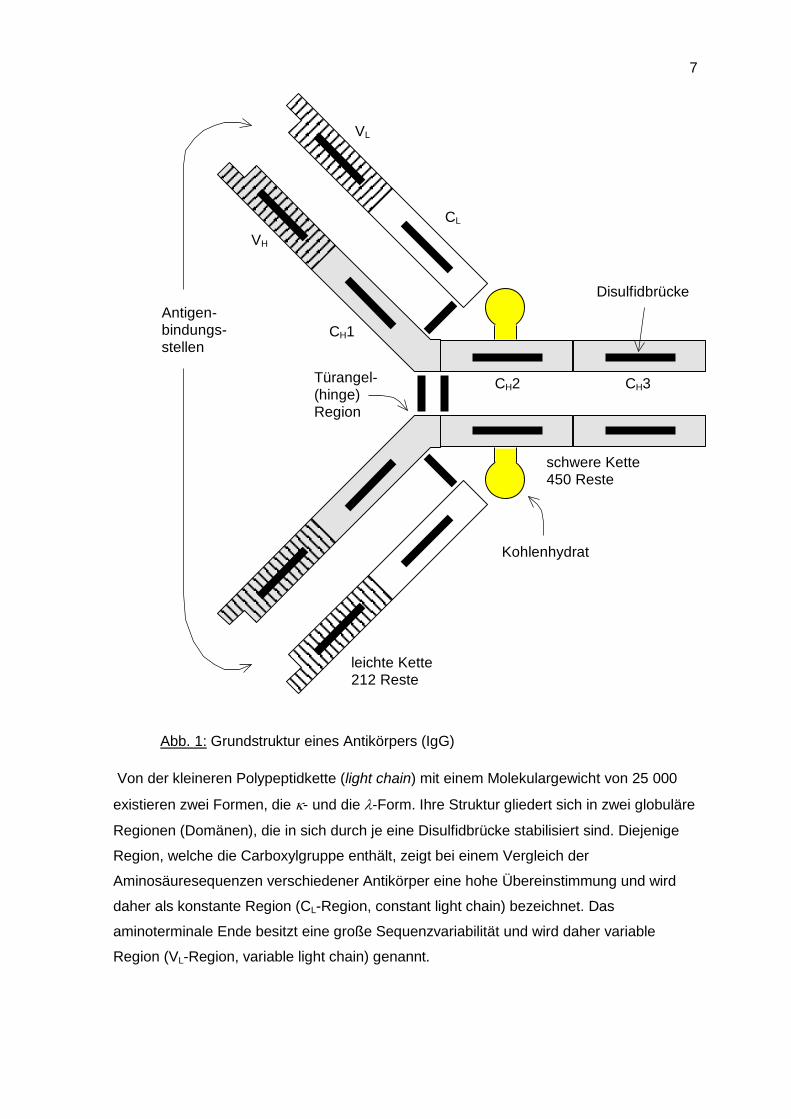
Die Grundstruktur aller Antikörper besteht aus je zwei identischen leichten und schweren
Polypeptidketten, die über Disulfidbrücken miteinander verbunden sind (Abb. 1).
7
Kohlenhydrat
Disulfidbrücke
schwere Kette450 Reste
leichte Kette
212 Reste
CH3CH2
CH1
VH
CL
VL
Türangel-(hinge)
Region
Antigen-
bindungs-stellen
Abb. 1: Grundstruktur eines Antikörpers (IgG)
Von der kleineren Polypeptidkette (light chain) mit einem Molekulargewicht von 25 000
existieren zwei Formen, die - und die -Form. Ihre Struktur gliedert sich in zwei globuläre
Regionen (Domänen), die in sich durch je eine Disulfidbrücke stabilisiert sind. Diejenige
Region, welche die Carboxylgruppe enthält, zeigt bei einem Vergleich der
Aminosäuresequenzen verschiedener Antikörper eine hohe Übereinstimmung und wird
daher als konstante Region (CL-Region, constant light chain) bezeichnet. Das
aminoterminale Ende besitzt eine große Sequenzvariabilität und wird daher variable
Region (VL-Region, variable light chain) genannt.

8
Die schwere Polypeptidkette (heavy chain) existiert in fünf Formen () mit einem
Molgewicht von 50.000-77.000. Jede dieser Formen ist mit einem Leichtkettentyp frei
kombinierbar, wodurch die fünf Immunglobulinklassen (IgA, IgD, IgE, IgG bzw. IgM)
entstehen. Variationen in der Schwerkettenstruktur innerhalb einer Klasse (z. B. 1, 2, 3
u. 4) führt zur Bildung von Subklassen (entsprechend: IgG1, IgG2, IgG3, bzw. IgG4).
Die Struktur der schweren Ketten ist denen der Leichtkette sehr ähnlich. Bedingt durch die
größere Molmasse besitzen sie jedoch neben der variablen Region (VH-Region, variable
heavy chain) drei konstante Domänen (CH1, CH2, CH3). Der Kohlenhydratanteil, den alle
Antikörper besitzen, ist an die CH2-Region gebunden.
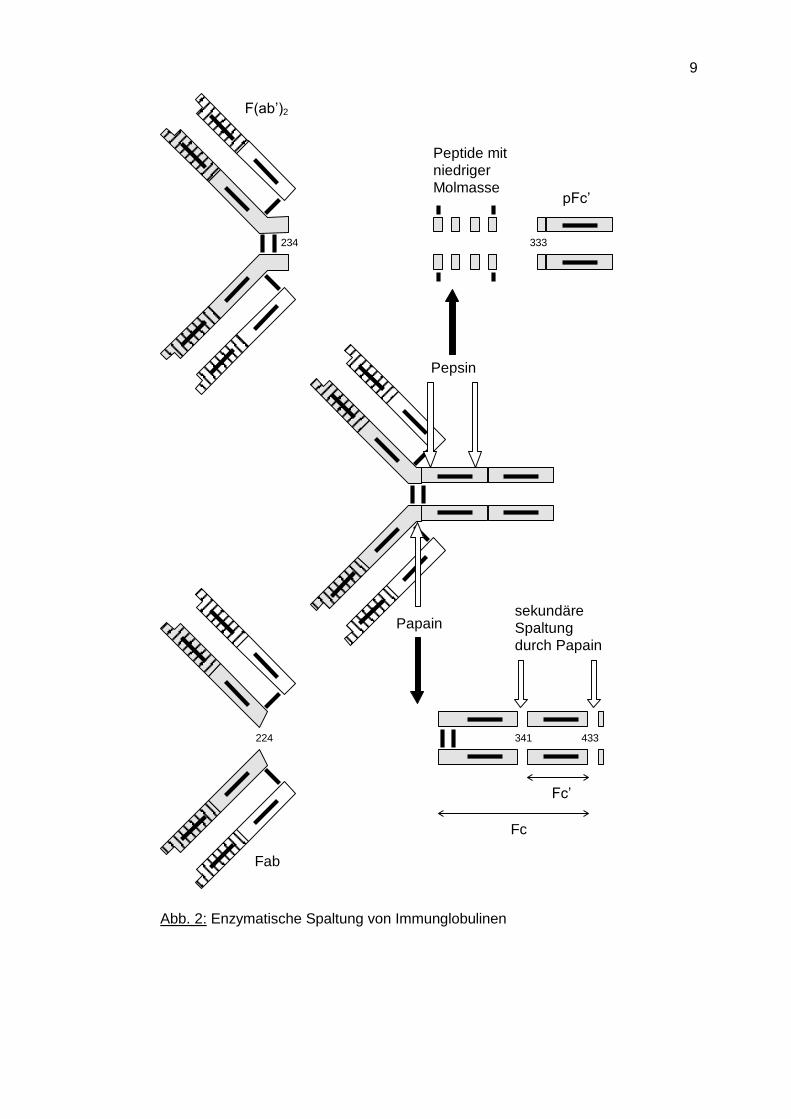
Die pflanzliche Proteinase Papain spaltet Immunglobuline in der Region zwischen den
CH1- und CH2-Domänen, der sogenannten Türangelregion (hinge region), wodurch zwei
identische Fab-Fragmente und ein Fc-Fragment entstehen (Abb. 2). Ein weiteres Enzym
für die Fragmentierung von Antikörpern ist Pepsin, welches zwei größere Fragmente
erzeugt, das F(ab')2-Fragment, das die über die Hinge-Region miteinander verbundenen
Fab-Regionen umfasst, und das pFc'- Fragment, welches den CH3-Domänen des
Antikörpermoleküls entspricht.
Mit diesen Fragmenten konnte gezeigt werden, dass die Antigen-Antikörper-Bindungsorte
in den variablen Bereichen (VL, VH) des Antikörpers liegen, während der Fc-Teil die
Bindung des Immunglobulins an verschiedene Zellen des Immunsystems, sowie
Komplementfaktoren vermittelt. Die hohe Beweglichkeit der Hinge-Region erlaubt eine
Änderung des Abstandes der Antigenbindungsorte, wodurch diese unabhängig
voneinander verfügbar sind.
9
F(ab’)2
Peptide mitniedriger
Molmasse
sekundäre
Spaltung
durch Papain
Papain
Pepsin
pFc’
333234
Fab
Fc’
Fc
433341224
Abb. 2: Enzymatische Spaltung von Immunglobulinen
10
2. Versuchsbeschreibung
Zur Bestimmung von Antikörpertitern gegen bestimmte Antigene existiert mittlerweile eine
Vielzahl von Methoden, wobei an dieser Stelle nur der RIA (radioimmunoassay) und der
ELISA (enzyme-linked immunosorbent assay) genannt seien. Beide Assays sind sehr
einfach in der Durchführung, doch sehr sensitiv und zuverlässig, was zu deren weit
verbreiteter Anwendung geführt hat. Die Grundlage beider Methoden beruht auf der
Tatsache, dass bestimmte Kunststoffoberflächen (z. B. Polystyrol, Polypropylen,
Polycarbonat, Polyvinylchlorid) geringe Mengen der meisten Proteine fest binden können.
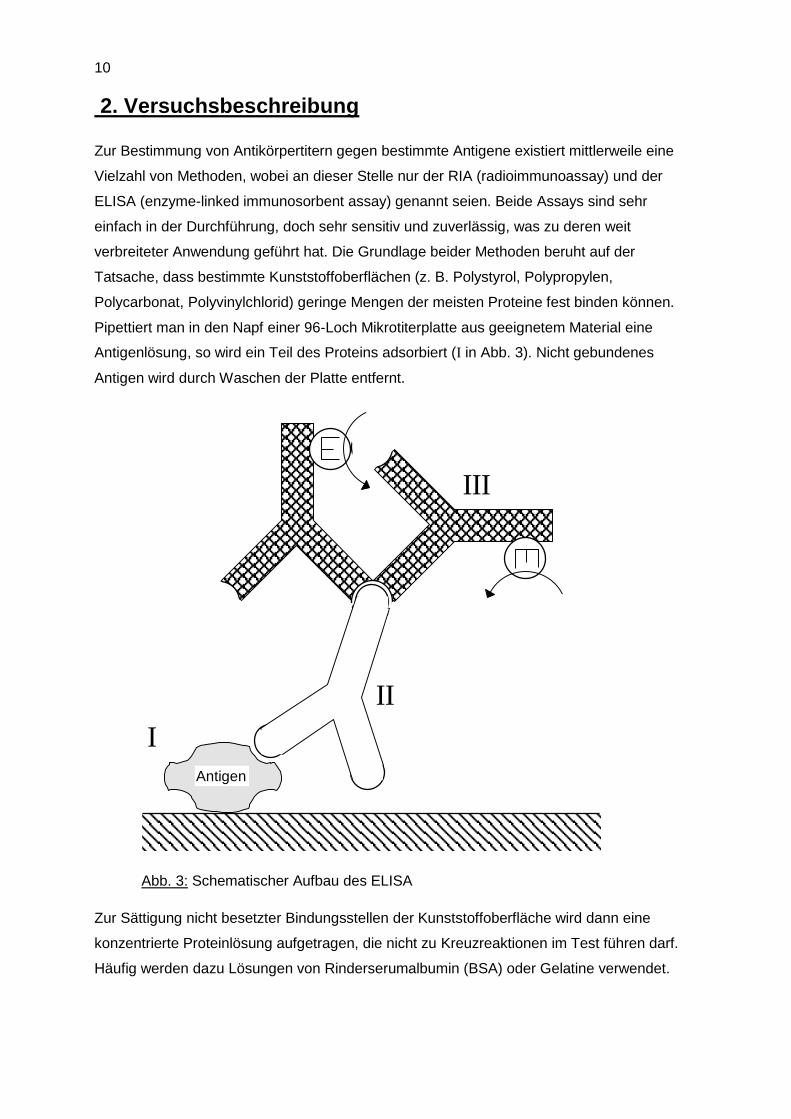
Pipettiert man in den Napf einer 96-Loch Mikrotiterplatte aus geeignetem Material eine
Antigenlösung, so wird ein Teil des Proteins adsorbiert (I in Abb. 3). Nicht gebundenes
Antigen wird durch Waschen der Platte entfernt.
I
II
III
Antigen
Abb. 3: Schematischer Aufbau des ELISA
Zur Sättigung nicht besetzter Bindungsstellen der Kunststoffoberfläche wird dann eine
konzentrierte Proteinlösung aufgetragen, die nicht zu Kreuzreaktionen im Test führen darf.
Häufig werden dazu Lösungen von Rinderserumalbumin (BSA) oder Gelatine verwendet.

11
In die so vorbehandelte Mikrotiterplatte wird dann die zu testende Lösung einpipettiert.
Falls in der Probe antigenspezifischer Antikörper vorhanden ist, bindet dieser an das an
die Festphase fixierte Antigen (II in Abb. 3). Alles nicht gebundene Material wird in einem
anschließenden Waschvorgang entfernt. Um den gebundenen Antikörper detektieren zu
können, wird die Platte dann mit einer Lösung von markiertem Anti-Antikörper behandelt
(III in Abb. 3).
Im Praktikumsversuch soll ein His-Tag markiertes Protein (Gelonin) aus
Expressionsversuchen immunologisch nachgewiesen werden. Es wird deshalb eine
monoclonale Anti-His-Tag Antikörperlösung verwendet. Die Markierung dieses zweiten
Antikörpers kann auf unterschiedliche Art und Weise erfolgen. Beim RIA ist der
Sekundärantikörper radioaktiv markiert, beim ELISA ist er mit einem Enzym (E in Abb. 3)
gekoppelt.
Nach einem letzten Waschvorgang wird das gebundene Enzym-Anti-Maus-Ig-Konjugat
durch eine Farbreaktion nachgewiesen, bzw. beim RIA an die Festphase gebundene
Radioaktivität gemessen. Je nach Anwendung können verschiedene Enzyme zur
Markierung des Anti- Antikörpers verwendet werden. Häufig werden Peroxidase(POD)-
und alkalische Phosphatase(AP)-Konjugate benutzt.
3. Versuchsdurchführung
3.1 Reagenzien und Chemikalien
Testprotein (Antigen) Rekombinantes Gelonin mit His-Tag (20 µg/ml)
Gelonin aus Samen von Gelonium multiflorum (20 µg/ml)
Kontrollen
Negativkontrollen: 20 µg/ml Cytochrom C
Positivkontrolle: 20 µg/ml rekombinantes Gelonin
Primärantikörper Monoklonale, antigenspezifischer Antikörper:
Mouse Anti-His Antikörper
Verdünnung: 1:2000 mit PBS-Tween
Enzymkonjugat Alkalische Phosphatase markierte, polyklonale Antikörper gegen Maus-Immunglobuline:
Anti-mouse lgG Antikörper (AP)
Verdünnung: 1:1000 mit PBS-Tween

12
Coating-Puffer Lösung A: 100 ml 0,2 M Na2CO3 (2,12 g ad 100 ml)
Lösung B: 100 ml 0,2 M NaHCO3 (1,68 g ad 100 ml)
Gebrauchslösung : 8,5 ml Lösung A + 4 ml Lösung B
pH 10,6 (pH-Kontrolle)
ad 50 ml mit bidest. Wasser
Waschpuffer PBS-Tween Na2HPO4 0,92 g
NaH2PO4 * H2O 0,35 g
NaCl 8,18 g
pH 7,2
ad 1 l mit deionisiertem Wasser
100 ml zur Seite stellen (= PBS-Puffer)
zu den restlichen 900 ml:
Tween 20 0,45 ml
Blockierlösung 1% BSA in PBS
250 mg BSA in 25 ml PBS
Substratpuffer (AP) NaN3 0,2 g (0,02 %)
MgCl2 * 6 H2O 0,1 g
DEA (Diethanolamin) 97 ml
pH 9,8
ad 1 l bidest. Wasser
Substratlösung (AP) 15 mg p-Nitrophenylphosphat ad 15 ml Substratpuffer
3.2 Versuchsablauf
a) Das Antigen wird im Coating-Puffer auf eine Konzentration von 20 g/ml eingestellt. Je
Napf werden 100 µl dieser Lösung eingebracht. Die Bindung an die feste Phase erfolgt
über Nacht bei 4 °C.
b) Die Näpfe der Platte werden durch Ausschlagen geleert. Pro Napf werden 200 µl der
Blockierlösung eingefüllt. Die Inkubation erfolgt bei Raumtemperatur für 90 min.
c) 2-maliges Waschen mit jeweils 200 µl PBS-Tween-Puffer. Ausschlagen der Näpfe.
d) Je Napf werden 100 l Primärantikörperlösung eingebracht. Die Inkubation erfolgt bei
Raumtemperatur für 1 h (siehe „Aufteilung der Mikrotiterplatte“, Abschn. 3.3)
e) 3 × Waschen. Näpfe ausschlagen.

13
f) Das Enzymkonjugat wird auf eine geeignete Arbeitskonzentration gebracht, d. h. ca.
1000 × verdünnt (10 l Enzymkonjugatlsg. + 10 ml PBS-Tween) und 100 l pro Napf
eingebracht. Die Inkubation erfolgt bei Raumtemperatur für 1 h.
g) 4 × Waschen. Näpfe ausschlagen.
h) 100 l Substratlösung zugeben. Die Entwicklung erfolgt bei Raumtemperatur. Die
Platte wird nach 10 min mit Hilfe eines EIA-Readers bei 414 nm vermessen.
3.3 Bestückung der Mikrotiterplatte
a) Blank:
Für die Messung benötigt der EIA-Reader eine Spalte der Platte als Blank. In die Spalte 1
dürfen deshalb keine Proben eingebracht werden. Stattdessen wird PBS einpipettiert. Alle
anderen Schritte wie Blockieren, Konjugatzugabe usw. erfolgen wie beschrieben.
b) Negativkontrollen:
Um falsche positive Ergebnisse auszuschließen, müssen in jedem ELISA
Negativkontrollen durchgeführt werden. Zu diesem Zweck werden 2 Spalten der Platte
nicht mit Antigen beschichtet. Das weitere Vorgehen erfolgt wieder wie beschrieben. Als
weitere Negativkontrolle wird in einer antigenbeschichteten Spalte ein Antikörper, der
nicht mit dem Antigen reagiert, als Probe eingebracht.
Die Ursachen für falschpositive Ergebnisse können sein:
Unspezifische Adsorption des Enzymkonjugats an das Plastikmaterial,
Kreuzreaktionen des Enzymkonjugats,
c) Positivkontrollen:
Um sicherzustellen, dass der ELISA richtig durchgeführt wurde, sollte immer eine Spalte
der Platte mit einer Probe bestückt werden, von der bekannt ist, dass sie für das
verwendete Antigen spezifische Antikörper enthält. Diese Löcher müssen sich beim
Entwickeln auf jeden Fall färben.

14
4. Auswertung
Die Auswertung erfolgt zunächst optisch, indem man die Löcher notiert, die deutlich
stärker gefärbt sind als die Negativkontrollen. Die Messwerte des EIA-Readers werden
statistisch ausgewertet.
Dazu werden die Negativkontrollen gemittelt und die Standardabweichung berechnet. Für
jede Probe wird ebenfalls der Mittelwert aus den Messwerten gebildet. Eine Probe ist
dann positiv, wenn dieser Mittelwert größer ist als die Summe aus dem Mittelwert der
Negativkontrollen und der zweifachen Standardabweichung.
5. Literatur
Harlow, E., Lane, D.,
Antibodies. A Laboratory Manual,
Cold Spring Harbor Laboratory, New York, 1988.
Kap. 14, Immunoassays, S. 553-612.
Goding, J. W.,
Monoclonal Antibodies: Principles and Practice, 2nd ed.,
Academic Press, London, 1986.
Kap. 3.10, Screening Assays, S. 76-89.
Peters, J. H., Baumgarten, H., (Hrsg.),
Monoklonale Antikörper. Herstellung und Charakterisierung, 2. Auflage,
Springer-Verlag, Berlin, Heidelberg, 1990.
Kap. 10, Nachweis von monoklonalen Antikörpern, S. 317-458.

15
Enzymatische Bestimmung von ADP (Adenosindiphosphat)
Einleitung zu den photometrischen Bestimmungen ...................................................... 15
Aufgabe ........................................................................................................................ 16
Prinzip der enzymatischen Methode ............................................................................. 17
Reagenzien (M = mol/l) ................................................................................................ 17
Berechnung des Versuchsergebnisses ........................................................................ 17
Durchführung der Bestimmung ..................................................................................... 18
Einleitung zu den photometrischen Bestimmungen
Wird eine Küvette, die mit einer absorbierenden Flüssigkeit oder Lösung gefüllt ist, von
Licht durchstrahlt, dann bewirkt die Absorption des Küvetteninhalts eine
Intensitätsabnahme des Lichtstrahls. Diese ist von der Wellenlänge des Lichts, der
Konzentration der absorbierenden Probe, der Länge des Lichtweges durch die
Messlösung sowie einer Stoffkonstanten (dem Extinktionskoeffizienten ) abhängig. Die
Wellenlängen, bei denen ein Maximum hat (die Absorptionsmaxima) sind für
absorbierende Atomgruppierungen (Chromophore) wie z. B. Nitrogruppen,
Doppelbindungen, Aromaten u. v. a. charakteristisch.
Die Konzentrationsbestimmung durch Messung der Absorption einer monochromatischen
Strahlung nach Durchgang durch die Messlösung bezeichnet man als Photometrie.
Grundlage für die photometrischen Konzentrationsbestimmungen ist das Lambert-
Beersche Gesetz, nach dem sich die Konzentration einer Lichtenergie absorbierenden
Verbindung in verdünnter Lösung berechnen lässt:
cE
d
c = Konzentration (mol/l)
= molarer Extinktionskoeffizient (l mol–1 cm–1)
d = Schichtdicke der Küvette (cm)
E = Extinktion

16
Die Ableitung des Lambert-Beerschen Gesetzes ist den Lehrbüchern zu entnehmen.
In der Regel ist die Skala eines lichtelektrischen Messgerätes linear und logarithmisch
unterteilt. Die lineare Teilung zeigt das Verhältnis der austretenden zur eingestrahlten
Lichtintensität an (Durchlässigkeit, Transmission T ), auf der logarithmischen Teilung wird
die Extinktion E (negativer Logarithmus der Durchlässigkeit) angezeigt.
Viele Substanzen, die farblos oder nur schwach gefärbt sind, können durch Zugabe eines
Reagenzes, mit dem sie unter Bildung stark gefärbter Produkte reagieren, bestimmt
werden. Dabei werden die Reaktionsbedingungen so standardisiert, dass die Menge der
zu bestimmenden Verbindung direkt proportional der gemessenen Extinktion ist.
c F E
c' = Gewichtsmenge der zu bestimmenden Verbindung im Standardtest
F = Proportionalitätsfaktor
Unter diesen Bedingungen wird eine Eichkurve mit bekannten Mengen der zu
bestimmenden Substanz aufgenommen und E gegen c' aufgetragen. Die Kurve sollte,
wenn von den gemessenen Extinktionswerten die Extinktion des Blindwertes (Extinktion
des Ansatzes ohne Analysensubstanz) subtrahiert wird, durch den Koordinatenursprung
gehen und eine Gerade ergeben, deren Steigung F entspricht.
Der ermittelte Proportionalitätsfaktor F dient zur Gehaltsbestimmung bei Lösungen
unbekannter Konzentration.
Aufgabe
In einer ADP-Lösung unbekannter Konzentration ist der Gehalt an ADP (Abb. 1) auf
enzymatischem Wege zu bestimmen.
O
O
ONa
P O
O
O
Na
Na
P O CH2
HO
O
OH
N
N
NH2
N
N
Abb. 1: ADP, Trinatriumsalz (MG 493,15)

17
Prinzip der enzymatischen Methode
Zur enzymatischen Bestimmung von ADP werden folgende Enzymreaktionen benutzt:
ADP + PEP PK
ATP + Pyruvat (1)
Pyruvat + NADH + H+ LDH
Lactat + NAD+ (2)
PEP = Phosphoenolpyruvat
PK = Pyruvatkinase
ATP = Adenosintriphosphat
NAD+ (NADH) = Nicotinamid-Adenin-Dinucleotid, oxidierte (bzw. reduzierte) Form
LDH = Lactatdehydrogenase
Durch Koppeln der Reaktion (1) mit der Indikatorreaktion (2), einem
Dehydrogenasesystem, kann ADP durch einen optischen Test gemessen werden. Da die
Gleichgewichte beider Reaktionen ganz auf der rechten Seite liegen, ist eine quantitative
Umsetzung von ADP in ATP und von Pyruvat in Lactat gewährleistet.
Reagenzien (M = mol/l)
0,1 M Tris-Puffer, mit HCl auf pH 7,5 eingestellt
PEP-Lösung in Wasser (2,5 mg / 0,4 ml)
NADH-Lösung in Wasser (4,5 mg / 0,6 ml)
0,5 M Magnesiumsulfat-Lösung
1 M KCl-Lösung
PK-Lösung in Puffer (ca. 0,1 mg / ml)
LDH-Lösung in Puffer (ca. 0,1 mg / ml)
Berechnung des Versuchsergebnisses
Aus den Reaktionsgleichungen (1) und (2) ist ersichtlich, dass pro mol umgesetztes ADP
1 mol NADH verbraucht wird. Die verbrauchte NADH-Menge kann aus der Extinktions-
änderung der Inkubationslösung berechnet werden:
340 = 6,3 · 103 l mol–1 cm–1
365 = 3,4 · 103 l mol–1 cm–1

18
Durchführung der Bestimmung
In eine Küvette (Schichtdicke 1 cm) werden nacheinander eingefüllt:
720 l Tris-Puffer
40 l MgSO4-Lösung
40 l 1 M KCl-Lösung
16 l PEP-Lösung
24 l NADH-Lösung
80 l der zu untersuchenden ADP-Lösung
40 l LDH-Lösung
Es wird gründlich gemischt und die Extinktion bei 365 nm oder 340 nm über 3 min
verfolgt. Wenn sie konstant bleibt, wird abgelesen, es werden 40 l der PK-Lösung
zugegeben, und es wird umgemischt. Ist die Extinktion erneut konstant, wird E365
bestimmt und daraus die ADP-Konzentration berechnet. Sollte die Menge zu groß oder
zu klein sein, um eine Extinktionsänderung von ca. 0,1 - 0,2 zu erreichen, dann sind die
Menge der ADP-Lösung und entsprechend die Puffermenge zu verändern; das Gesamt-
volumen des Ansatzes sollte dabei konstant bleiben. Es sollen mindestens drei Bestim-
mungen ausgeführt werden.
Tabelle zur Auswertung:
Ergebnis: ADP-Gehalt aufgrund der enzymatischen Bestimmung: ...................
ADP-Lsg [ml] E365 umgesetzte NADH-Menge
ADP-Menge im Ansatz
ADP-Konz. in der Lsg

19
Disk-Elektrophorese in Natriumdodecylsulfat (SDS-PAGE)
Einführung .................................................................................................................... 19
Das Trennmedium ........................................................................................................ 20
Die Elektrophorese ....................................................................................................... 21
Versuchsdurchführung ................................................................................................. 23
Einführung
Neben ihrer großen Bedeutung zur Reinheitskontrolle bei Proteinisolierungen ist die SDS-
Polyacrylamidgelelektrophorese (abgek.: SDS-PAGE) auch eine Methode zur
Molekulargewichtsabschätzung von Proteinen. Zur Trennung von Proteinen im Poly-
acrylamid-Gel wird hauptsächlich der Siebeffekt des Gels ausgenutzt. Um eine gute und
gleichsinnige Beweglichkeit der Proteinmoleküle im elektrischen Feld zu erreichen,
werden diese vor der Elektrophorese mit dem Detergenz Natriumdodecylsulfat (SDS)
beladen. Durch die Bindung von SDS werden Proteine zu Polyanionen, und das Protein
wird denaturiert. Da Proteine mit Disulfidbrücken in der SDS-PAGE häufig ein anomales
Verhalten zeigen, werden die Disulfidbrücken vor der Elektrophorese mit 2-Mercapto-
ethanol reduziert.
Die Vorinkubation der Proteine in SDS und 2-Mercaptoethanol schafft die Voraussetzung
für die Molekulargewichtsabschätzung im SDS-Gel, da jetzt die Beweglichkeit globulärer
Proteine bei gleicher Gelkonzentration (gleicher Porengröße) nur eine Funktion der Größe
des Moleküls ist.
Für die Molekulargewichtsbestimmung wird die relative Beweglichkeit von Proteinen mit
bekanntem Molekulargewicht in Bezug auf einen internen Standard (meist Bromphenol-
blau) verglichen und eine Eichkurve aufgestellt, indem der Logarithmus des Molekular-
gewichtes gegen den Rf-Wert (relative Beweglichkeit) aufgetragen wird. Aus dem Rf-Wert
des unbekannten Proteins kann somit dessen Molekulargewicht abgeschätzt werden.

20
Probleme: Die Beweglichkeit der Proteine im Polyacrylamid-Gel ist nicht allein von ihrem
Molekulargewicht abhängig, sondern zu einem erheblichen Teil auch von der jeweiligen
Form der denaturierten Eiweiße (globuläre Proteine wandern schneller als vollständig
entfaltete, lange Proteinfäden). Außerdem verändern bestimmte, natürlich vorkommende
Modifizierungen der Eiweiße, wie z. B. Glykosylierungen, deren Wanderungstendenz im
elektrischen Feld. Das SDS-Gel kann deshalb nur zur Abschätzung des Molekular-
gewichts dienen.
Das Trennmedium
Durch Kopolymerisation des monomeren Acrylsäureamids,
O
CCHCH2 NH2 ,
und eines quervernetzenden bifunktionellen Reagenzes, meist
N,N’-Methylenbisacrylsäureamid,
O
C CHCH CH2CH2 NH CH2 NH
O
C ,
wird ein dreidimensionales Netzwerk aufgebaut:
y. . .. . .
OC
CHCH2
NH2
OC
CHCH2
NH2
CH2
z
OC
CHCH2
NH2
w
. . .. . .
OC
CHCH2
NH2
OC
CHCH2
NH2 x
OC
CHCH2
NH2

21
Die Porengröße ist in einem recht weiten Bereich variabel, die Bisacrylamidkonzentration
kann in einem Bereich zwischen 2 % und 30 % der Acrylamid-Konzentration liegen. Bei
zu geringer Vernetzerkonzentration zerfließen die Gele, bei zu hoher werden sie brüchig.
Beispielsweise entspricht ein 7,5-%iges Gel einem Porendurchmesser von 50 Å und ein
30-%iges einem solchen von 20 Å. Zum Vergleich sei die räumliche Ausdehnung eines
Serumalbuminmoleküls angegeben: 40 Å × 150 Å. Die Polymerisation geschieht
radikalisch unter Zuhilfenahme von Radikalbildnern (hier: Ammoniumpersulfat).
Die Elektrophorese
Die Ausrüstung für eine Elektrophorese besteht aus zwei Teilen, einem Strom-
versorgungsteil (gewöhnlich ein Gleichspannungsgerät 0 - 550 V bzw. 0 - 200 mA) und
einer Elektrophoresekammer. Abb. 1 zeigt deren prinzipiellen Aufbau. Die Kammer
besteht aus einem oberen und unteren Pufferreservoir, die über die Gelplatte miteinander
verbunden sind. Da bei der SDS-Gelelektrophorese alle Proteine negativ geladen sind,
enthält das untere Reservoir die Anode, das obere die Kathode.
Kathode
Anode
unteres Pufferreservoir
Glasplatten
Trenngel
oberes Pufferreservoir
Sammelgel
Abb. 1: Prizipieller Aufbau einer Vertikal-Elektrophoresekammer

22
In der Praxis verwendet man diskontinuierliche Gele und Puffer. Man spricht deshalb auch
von einer diskontinuierlichen SDS-Polyacrylamidgelelektrophorese (abgekürzt SDS-Disk-
Elektrophorese). Das obere Drittel des Gels besteht aus einem weitporigen Sammelgel
(engl. stacking gel), die unteren zwei Drittel des Gels bestehen aus einem engerporigen
Trenn- oder Laufgel (engl. running gel). Das Sammelgel hat die Aufgabe, die Probe,
sobald sie durch das Gel läuft, zu konzentrieren, sodass sie, sobald sie in das Laufgel ein-
dringt, eine extrem schmale Bande bildet. Im Laufgel findet dann die Trennung des
Proteingemisches statt. Die Wanderungsgeschwindigkeit von Proteinen im Sammelgel
entspricht der in freier Lösung. Der Reservoirpuffer ist ein Tris(hydroxymethyl)-
aminomethan/Glycin-Puffer (Tris/Glycin) pH 8,3, während die Sammel- und Trenngelpuf-
fer aus Tris/HCl bestehen, pH 6,8 bzw. 8,8. Der pH des Sammelgels ist also um zwei pH-
Einheiten niedriger als der des oberen Reservoirs. Bei pH 8,3 liegen nur ca. 5 % des
Glycins als Glycinat vor, das bedeutet, dass die Glycinationen eine relativ geringe effek-
tive Beweglichkeit besitzen. Nach Auftragen der Proteinproben wird das elektrische Feld
angelegt, und die Proteine wandern mit den Glycinationen in das Sammelgel. Die
Chloridionen im Gel fangen natürlich ebenfalls an zu wandern, so dass während der
Elektrophorese zusammen mit den Proteinen zwei verschiedene Ionenarten in
Trennrichtung wandern. Sie werden als Leitionen (Cl–) und als Folgeionen (Glycinat)
bezeichnet.
Zu Beginn der Trennung befinden sich die Leitionen (Cl–) in Sammel- und Trenngel,
während die Folgeionen nur in der Pufferlösung der Elektrodengefäße vorhanden sind.
Durch die Wahl des pH-Wertes von 6,8 für das Sammelgel ordnen sich die effektiven
Beweglichkeiten u (u = Beweglichkeit, = Dissoziationsgrad) folgendermaßen an:
uLeitionen > uProtein > uFolgeionen,
wobei Leitionen, Proteine und Folgeionen dasselbe Ladungsvorzeichen tragen, also zur
gleichen Elektrode wandern.
Im Sammelgel bei pH 6,8 wandert Glycinat etwa 100 mal langsamer als bei pH 8,8 im
Trenngel. Unterwirft man das ganze Gelsystem einem elektrischen Stromfluss, so eilen
die Leitionen infolge höherer Beweglichkeit den Proteinen und Folgeionen voraus und
lassen eine Zone geringerer Leitfähigkeit hinter sich.
Nun ist die spezifische Leitfähigkeit umgekehrt proportional zur Feldstärke. Diese Zone
gewinnt daher eine höhere Feldstärke, welche die Proteine und Folgeionen derart
beschleunigt, dass sie hinter den Leitionen mit gleicher Geschwindigkeit wandern und
sich dadurch ansammeln.

23
Es bewegen sich alle Ionenarten mit gleicher Geschwindigkeit, wenn die Produkte aus
Feldstärke und Beweglichkeit einander gleichen:
v = E · u
(v = Geschwindigkeit, E = Feldstärke, u = Beweglichkeit)
Es stellt sich ein regulierendes Gleichgewicht ein, das die Gleichheit der Produkte
aufrechterhält. Zwischen den Leit- und Folgeionen bildet sich daher eine Grenzschicht
aus, die gleichzeitig die Front zwischen Gebieten niedriger und hoher Feldstärke darstellt.
Man kann sie als eine von oben nach unten wandernde Schlierenfront im praktischen
Versuch wahrnehmen. Diese Grenzschicht wandert nun rasch durch das Sammelgel. Da
die Beweglichkeiten der Proteine zwischen denen der Leit- und Folgeionen liegen, werden
die Proteine von der wandernden Grenzschicht erfasst und zu einer schmalen,
hochkonzentrierten Proteinzone zusammengestaucht.
Erreicht nun die wandernde Proteinzone, eingezwängt zwischen Leit- und Folgeionen, die
Grenze zwischen Sammel- und Trenngel, so trifft sie dort zwei Diskontinuitäten an,
nämlich einen pH-Wechsel und einen Porengrößenwechsel. Der „neue“ pH-Wert im
Trenngel ist so gewählt, dass der Dissoziationsgrad der Folgeionen - und damit deren
Beweglichkeit - um ein Vielfaches zunimmt. Dadurch gewinnen die Folgeionen eine
Beweglichkeit, die der der Leitionen fast gleicht. Die Folgeionen überholen nun alle
Proteine und wandern direkt hinter den Leitionen den Proteinen voraus, die nun in einem
homogenen elektrischen Feld entsprechend ihrer Molekülgröße aufgetrennt werden.
Versuchsdurchführung
SDS-PAGE nach Smith, Methods in Molecular Biology, Volume1 (Proteins), 41-56.
Lösungen
Für alle Lösungen und Puffer entgastes destilliertes Wasser verwenden!!

24
Bemerkung: „Wasser ad 250 ml“ bedeutet, dass die Substanzen in ungefähr 150 ml dest.
Wasser gelöst und anschließend auf 250 ml Gesamtvolumen aufgefüllt werden.
Vorsicht: Acrylamid ist für Haut und Lunge sehr giftig. Abwiegen nur mit Handschuhen
und unter dem Abzug! Nicht mit dem Mund pipettieren! Die mit Acrylamid in Berührung
gekommenen Geräte sofort nach Gebrauch mit Wasser spülen!
2) Trenngelpuffer: SDS 1 g
Tris 45,5 g
HCl bis pH 8,8
Wasser ad 250 ml
Bemerkung: „Wasser ad 250 ml“ bedeutet hier, dass die Substanzen in ungefähr 200 ml
Wasser gelöst werden, der pH eingestellt und anschließend auf 250 ml Wasser aufgefüllt
wird.
Auf 2 l auffüllen. Es stellt sich ein pH von ca. 8,3 ein (Kontrolle!).
1) Acrylamidlösung: Acrylamid 73 g
Bisacrylamid 2 g
Wasser ad 250 ml
3) Sammelgelpuffer: SDS 1 g
Tris 15,5 g
HCl bis pH 6,8
Wasser ad 250 ml
4) Elektrophoresepuffer: Glycin 28,8 g
Tris 6 g
SDS 2 g

25
Bemerkung: Bromphenolblau schädigt die Membran der pH-Elektrode und darf deshalb
erst nach Einstellung des pH-Werts dazugegeben werden.
5) Ammoniumpersulfatlösung: Ammoniumpersulfat 1 g
Auf 10 ml Wasser auffüllen.
6) Trenngel (10 %): Acrylamidlösung 5 ml
Wasser 6,25 ml
Trenngelpuffer 3,75 ml
Gut entgasen!
Ammoniumpersulfatlsg. 25 l
TEMED 10 l
7) Sammelgel (4,5 %): Acrylamidlösung 0,75 ml
Wasser 3 ml
Sammelgelpuffer 1,25 ml
Gut entgasen!
Ammoniumpersulfatlsg. 15 l
TEMED 5 l
8) Probenpuffer (doppelt stark): SDS 0,92 g
Glycerin 4 g
Tris 0,3 g
Mercaptoethanol 2 ml
mit Wasser auf 7 ml
HCl bis pH 6,8
0,1 % Bromphenolblau 2 ml
mit Wasser auf 7 ml

26
9) Probenbereitung:
Proteinlösung wird 1:1 mit Probenpuffer versetzt. Die Lösungen können mit Wasser
verdünnt werden. Proteinmenge einer Auftragung: 5-25 g Protein in 10 l
Gesamtvolumen.
Arbeitsgang:
1) Zusammensetzen der sauberen, mit vergälltem Ethanol abgeriebenen Platten zur
Gelkammer.
2) Nach gründlichem Mischen der Trenngellösung (6) wird die Gelkammer bis 1 cm
unterhalb des Randes mit einer Pasteurpipette gefüllt und vorsichtig mit wasser-
gesättigtem Butanol überschichtet. Dadurch erhält man eine glatte Phasengrenze
zwischen Trenn- und Sammelgel.
3) Nach Beendigung der Polymerisation wird das Butanol mit einer Pasteurpipette
vorsichtig herausgesaugt und mit Wasser nachgespült. Die Sammelgellösung wird auf
das Trenngel aufgetragen und der Kamm vorsichtig in die Lösung eingeschoben.
4) Ist das Sammelgel polymerisiert, zieht man behutsam den Kamm heraus.
5) Die Gele werden mit den Glasplatten in die Elektrophoreseapparatur eingespannt und
diese mit Elektrophoresepuffer aufgefüllt.
6) Zur Elektrophorese werden 20 l der denaturierten Proben mit einer Mikropipette durch
den Elektrophoresepuffer hindurch in die Probentaschen eingebracht, ohne das
Sammelgel zu verletzen. Das Gel sollte insgesamt asymmetrisch beladen werden, da
sonst später keine Auswertung möglich ist.
7) Man setzt den Deckel auf und stellt eine konstante Spannung von 200V ein.
8) Nach erfolgter Elektrophorese und Abschalten der Spannungsquelle wird der
Elektrophoresepuffer dekantiert, und die Gelkammern werden ausgespannt. Das Gel
wird vorsichtig von den Platten abgelöst (nicht mit den Fingern anfassen!) und sofort in
Wasser eingetaucht.

27
9) Gefärbt wird das Gel mit PageBlue, einer Proteinfärbelösung von Fermentas nach
beiliegender Vorschrift. Zunächst wird das Gel in 100ml destilliertes Wasser gelegt und
1min bei höchster Leistung in der Mikrowelle erhitzt. Anschließend verbleibt das Gel für
5min auf dem Schüttler. Das Wasser wird entfernt und der Vorgang noch zwei weitere
Male wiederholt. Dieser erste Waschschritt dient zur Entfernung des SDS, da dieses
die Färbung stört. Anschließend wird das Gel mit genügend Färbelösung bedeckt und
erneut 30sec bei höchster Leistung in der Mikrowelle erhitzt und darauf hin 20min auf
dem Schüttler gefärbt. Danach wird die Färbelösung entfernt und das Gel mit
destilliertem Wasser von Farbresten befreit. Das Gel verbleibt über Nacht in
destilliertem Wasser auf dem Schüttler. Die Gele können so mehrere Tage gelagert
werden.
Aufgabe:
Bestimmung des Molekulargewichts der unbekannten Proteinproben.

28
Quantitative Proteinbestimmungsmethoden
1. Proteinbestimmung mit BCA-Test ............................................................................ 29
Einleitung ................................................................................................................. 29
Methode ................................................................................................................... 30
2. Konzentrationsbestimmung nach Bradford .............................................................. 32
Theorie ..................................................................................................................... 32
Aufgabe .................................................................................................................... 32
Reagenzien .............................................................................................................. 32
Durchführung ........................................................................................................... 33
Auswertung .............................................................................................................. 33
Literatur: ................................................................................................................... 34
Die quantitative Bestimmung von Proteinen gehört wohl zu den wichtigsten Methoden in
der allgemeinen Biochemie. In der Literatur ist eine Vielzahl verschiedener Bestimmungs-
verfahren beschrieben, die jedoch fast alle mit bestimmten Vor- und Nachteilen behaftet
sind.
Es soll hier dringend darauf hingewiesen werden, dass im Allgemeinen mit keinem der
Bestimmungsverfahren absolute Proteinwerte erhalten werden können. In den meisten
Fällen werden zur Bestimmung der unbekannten Proteinkonzentrationen andere, leicht
zugängliche Proteine (wie z. B. Rinderserumalbumin, BSA), als Standard verwendet. Die
über solche Methoden erhaltenen Proteinwerte sind also nur als relative Werte zu dem
jeweiligen Standardprotein zu verstehen.
Absolute Proteinbestimmung kann also nur dann durchgeführt werden, wenn zum einen
das zu untersuchende Protein als Standard verwendet werden kann (nach Lyophilisieren
und genauem Auswiegen kann eine Standardkurve ermittelt werden) oder zum anderen
bei bekanntem Extinktionskoeffizienten des untersuchten Proteins die UV-Absorption bei
einer definierten Wellenlänge bestimmt werden kann.

29
1. Proteinbestimmung mit BCA-Test
Einleitung
Der BCA-Test ist ein Proteinbestimmungs-Assay der Firma Uptima, der sich durch hohe
Empfindlichkeit und geringe Störanfälligkeit auszeichnet. Er ist kompatibel mit vielen
ionischen und nicht-ionischen Detergenzien. Der Test ist einfacher und schneller
durchzuführen als die Proteinbestimmungsmethode nach Lowry. Die Empfindlichkeit
erreicht 5 µg/ml im erweitertem Protokoll. Außerdem kann der Test in angepasster Form
in Microtiterplatten durchgeführt werden.
Bicinchoninsäure (2,2’-Bichinolin-4,4’-dicarbonsäure, bicinchoninic acid, BCA) ist ein Cu+-
Reagenz. Die Farbbildung wird auf die vier Aminosäuren Cystein, Cystin, Tryptophan und
Tyrosin zurückgeführt. In einer Biuret-Reaktion reduziert das Protein Cu2+ in alkalischem
Medium zu Cu+. Anschließend wird das Kupfer(I)-Ion von zwei Molekülen BCA
komplexiert.
Abbildung1: BCA-Reagenz

30
Abbildung 2: Mit BCA-Reagenz komplexiertes Cu+
Methode
Die verwendeten Lösungen enthalten:
Lösung A: - Natriumcarbonat
- Natriumbicarbonat
- BCA
- Natriumtartrat
- in 0,2 N NaOH
Lösung B: - 4%ige Kupfersulfat-Lösung
Lösung A und Lösung B sind kommerziell erhältlich.
Herstellung der Eichgeraden
Es wird eine BSA-Lösung der Konzentration c = 250 µg/ml unter Verwendung einer
Stammlösung (SL) der Konzentration c = 2 mg/ml hergestellt.
Eine Verdünnung von 100 µl SL (2 mg/ml) 200 µg BSA mit 700 µl Wasser ergibt eine
BSA-Lösung von c = 250 µg/ml. Diese Lösung wird zum Erstellen der Eichgerade
verwendet.
Herstellung des BCA-Reagenzes
BCA-Lösung A und BCA-Lösung B werden im Verhältnis 50:1 gemischt. Zum Beispiel
29,4 ml Lösung A und 0,6 ml Lösung B.
31
Das so angesetzte Reagenz kann nur einen Tag lang verwendet werden; immer so viel
Reagenz ansetzen, dass es auch für die zu bestimmenden Proben ausreicht.
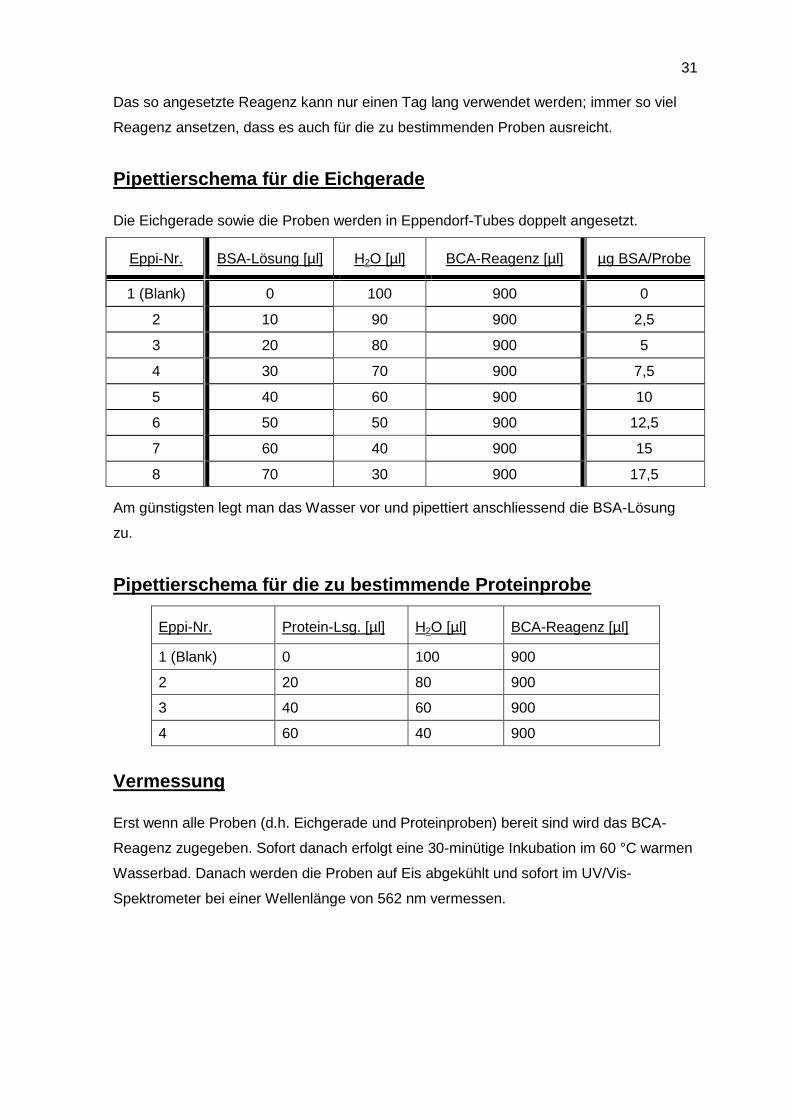
Pipettierschema für die Eichgerade
Die Eichgerade sowie die Proben werden in Eppendorf-Tubes doppelt angesetzt.
Eppi-Nr. BSA-Lösung [µl] H2O [µl] BCA-Reagenz [µl] µg BSA/Probe
1 (Blank) 0 100 900 0
2 10 90 900 2,5
3 20 80 900 5
4 30 70 900 7,5
5 40 60 900 10
6 50 50 900 12,5
7 60 40 900 15
8 70 30 900 17,5
Am günstigsten legt man das Wasser vor und pipettiert anschliessend die BSA-Lösung
zu.
Pipettierschema für die zu bestimmende Proteinprobe
Eppi-Nr. Protein-Lsg. [µl] H2O [µl] BCA-Reagenz [µl]
1 (Blank) 0 100 900
2 20 80 900
3 40 60 900
4 60 40 900
Vermessung
Erst wenn alle Proben (d.h. Eichgerade und Proteinproben) bereit sind wird das BCA-
Reagenz zugegeben. Sofort danach erfolgt eine 30-minütige Inkubation im 60 °C warmen
Wasserbad. Danach werden die Proben auf Eis abgekühlt und sofort im UV/Vis-
Spektrometer bei einer Wellenlänge von 562 nm vermessen.

32
2. Konzentrationsbestimmung nach Bradford
Theorie
Die Bestimmung der Proteinkonzentration nach Bradford ist, ähnlich wie die Methode
nach Lowry, ein Verfahren, das auf einer Farbreaktion basiert. Die Nachweismethode
findet im stark sauren Medium statt, das Coomassie Brilliant Blue G-250, einen
Triarylmethanfarbstoff enthält. Durch Bindung des Farbstoffes an das Protein erfolgt ein
spezifischer Farbwechsel von einem Aborptionsmaximum von 465 nm zu 595 nm. Die
Intensität des Farbkomplexes ist direkt vom Proteingehalt abhängig.
Der Farbkomplex ist nach zwei Minuten gut sichtbar und für ca. eine Stunde stabil.
Diese Proteinbestimmungsmethode ist schnell durchführbar, gut zu reproduzieren und
sehr sensitiv. Sie wird durch Pufferchemikalien und reduzierende Stoffe kaum gestört,
versagt allerdings, wenn in der Proteinlösung Substanzen enthalten sind, die in stark
phosphorsauren Lösungen grobflockige Niederschläge bilden (z. B. Detergenzien wie
Desoxycholat o. ä.). Störungen des Testes werden auch durch Natriumdodecylsulfat
(SDS) und Triton X-100 verursacht.
Der Nachteil der Methode liegt, wie bei allen farb-chemischen Nachweismethoden darin,
dass die Proteinprobe wegen Denaturierung nicht mehr wiederverwendet werden kann.
Aufgabe
Ermittlung der Konzentration einer unbekannten Proteinlösung.
Reagenzien
Bradford-Reagenz
0,1 g Coomassie Brilliant Blue G-250 werden in 50 ml 50-%igem Ethanol (v/v) gelöst.
Dann werden 100 ml 85-%ige Phosphorsäure zugegeben und mit dest. Wasser auf
250 ml aufgefüllt.
Vor Gebrauch wird ein Volumen des Reagenz mit 4 Volumen dest. Wasser gemischt und
filtriert.
Modifizierung nach M. Holtzhauer (1988)

33
Albuminlösung als Standard für die Eichgerade
10 mg Rinderserumalbumin (BSA) werden in einem 10-ml-Messkolben eingewogen und
mit dest. Wasser bis zur Marke aufgefüllt. Die Einwaage muss genau bekannt sein. Lösen
und Mischen durch vorsichtiges Drehen des Kolbens - nicht schütteln!
Durchführung
Erstellen der Eichgerade
Nach folgendem Schema wird der Albuminstandard (1 mg/ml) in einzelne Reagenzgläser
pipettiert.
2 × 0 l (Blank), 2 × 5 l, 2 × 7 l, 2 × 10 l, 2 × 12 l, 2 × 15 l (Dupletts)
Daraufhin werden in jedes Reagenzglas 1 ml Bradford-Reagenz hinzugefügt und auf
einem Whirlmix durchmischt.
Nach ca. 2 Minuten ist der Farbkomplex stabil. Dann erfolgt die Extinktionsmessung in
Kunststoffküvetten bei 595 nm. (Eppendorf-Photometer: 578 nm oder 623 nm)
Konzentrationsbestimmung der unbekannten Proteinlösung
2 × 3 l, 2 × 5 l, 2 × 7 l der Proteinlösung in einzelne Reagenzgläser pipettieren.
Proben mit jeweils 1 ml Bradford-Reagenz versetzen und analog zur Vorschrift der
Eichgeraden vermessen.
Falls die bestimmte Extinktion außerhalb des Bereiches der Eichgeraden liegt, muss die
Stammproteinlösung verdünnt eingesetzt werden.
Achtung: Bei dieser Proteinbestimmungsmethode sind die Eichgeraden nur bis zu einem
maximalen Proteingehalt von 15 g pro Probe proportional zur Proteinmenge, danach
knickt die Eichgerade ab.
Auswertung
Ermittelte Extinktionswerte für die verschiedenen BSA-Gehalte (g) in einem Diagramm
auftragen. Mittels linearer Regression kann daraufhin aus den gemessenen
Extinktionswerten der unbekannten Proteinlösung deren Konzentration bestimmt werden.

34
Literatur:
M. M. Bradford, (1976) Anal. Biochem. 72, 248-254
Holtzhauer M. (1988) Biochemische Labormethoden, Arbeitsvorschriften und Tabellen,
Springer-Verlag, Berlin, Heidelberg, New York, 5-6

35
Bestimmung der katalytischen Konstanten und der Michaelis-Konstanten von Chymotrypsin für
N-Succinyl-alanyl-alanyl-prolyl-phenylalanin-4-nitroanilid (Suc-AAPF-pNA)
Bestimmung der katalytischen Konstanten und der Michaelis-Konstanten von
Chymotrypsin für N-Succinyl-alanyl-alanyl-prolyl-phenylalanin-4-nitroanilid (Suc-AAPF-
pNA) ............................................................................................................................ 35
Graphische Bestimmung von KM und vmax .................................................................... 37
Aufgabe ........................................................................................................................ 39
Herstellung der Lösungen ............................................................................................ 40
Testansatz ................................................................................................................... 40
Auswertung .................................................................................................................. 41
Benötigte Angaben: ...................................................................................................... 41
Anmerkungen: .............................................................................................................. 42
Die Reaktionsgeschwindigkeit einer enzymatisch katalysierten Reaktion hängt u. a.
auch von der Substratkonzentration ab. Bei der maximalen Umsatzgeschwindigkeit
(vmax) ist das Enzym vollständig mit Substrat abgesättigt, während bei geringeren
Substratkonzentrationen nicht alle Enzymmoleküle abgesättigt sind.
KM
[S]
v
vmax
vmax/2

36
Abb. 1
1923 lieferten Michaelis und Menten die mathematische Analyse dieses Verhaltens. Sie
nahmen folgenden Reaktionsverlauf an:
E + S k2
k1 ES
k4
k3 P + E
E = Enzym, S = Substrat, ES = Enzymsubstratkomplex, P = Produkt. [E] ist die
Gesamtkonzentration an Enzym, ([E] - [ES]) die Konzentration an freiem Enzym. Die
Menge an S, die an E gebunden wird, ist, bezogen auf die Gesamtmenge an S, sehr klein
und kann vernachlässigt werden. [S] entspricht dann zu Beginn der Messung der
eingesetzten Substratkonzentration, da noch kein P gebildet wurde. Aus demselben
Grund ist bei Messung der Anfangsgeschwindigkeit die Bildung von ES aus P + E mit der
Geschwindigkeitskonstante k4 vernachlässigbar.
Ist die Bildungsgeschwindigkeit von ES,
k1 · ([E] - [ES]) · [S],
gleich seiner Zerfallsgeschwindigkeit,
k2 · [ES] + k3 · [ES],
so besteht ein Fließgleichgewicht (stationärer Zustand, steady state). Gleichsetzen der
beiden Ausdrücke und Zusammenfassen der drei Geschwindigkeitskonstanten ergibt die
Michaeliskonstante
Kk k
kM
2 3E ES S
ES
1
(1)
Wenn k2 k3, kann k3 vernachlässigt werden, und KM k2/k1 = KD ist die Dissoziations-
konstante des Gleichgewichts E + S ES.
Eine Bestimmung von KM ist dann möglich, wenn [ES] bestimmt werden kann. Dies ist auf
direktem Weg sehr schwierig, aber die Reaktionsgeschwindigkeit ist proportional [ES]:
v = k3 · [ES] (2)
Aus Gleichung (1) wird [ES] errechnet:
ES
E S
SM
K

37
Dieser Wert wird in Gleichung (2) eingesetzt:
vk
K
3
M
E S
S
Wenn [S] so groß ist, dass die gesamte eingesetzte Enzymmenge als Enzymsubstrat-
komplex vorliegt ([E] = [ES]), ist die maximale Geschwindigkeit erreicht:
vmax = k3 · [E]
Daher:
vv
K
max
M
S
S
Dies ist die von Michaelis und Menten entwickelte Gleichung. Bei halbmaximaler
Geschwindigkeit wird KM = [S]. KM hat somit die Dimension einer Konzentration.
Graphische Bestimmung von KM und vmax
Die Michaelis-Menten-Gleichung kann in verschiedener Weise zur Bestimmung von vmax
und KM graphisch dargestellt werden:
a) v wird gegen [S] aufgetragen und ergibt eine Hyperbel. Für v = vmax/2 wird [S] = KM
(Abb. 1).
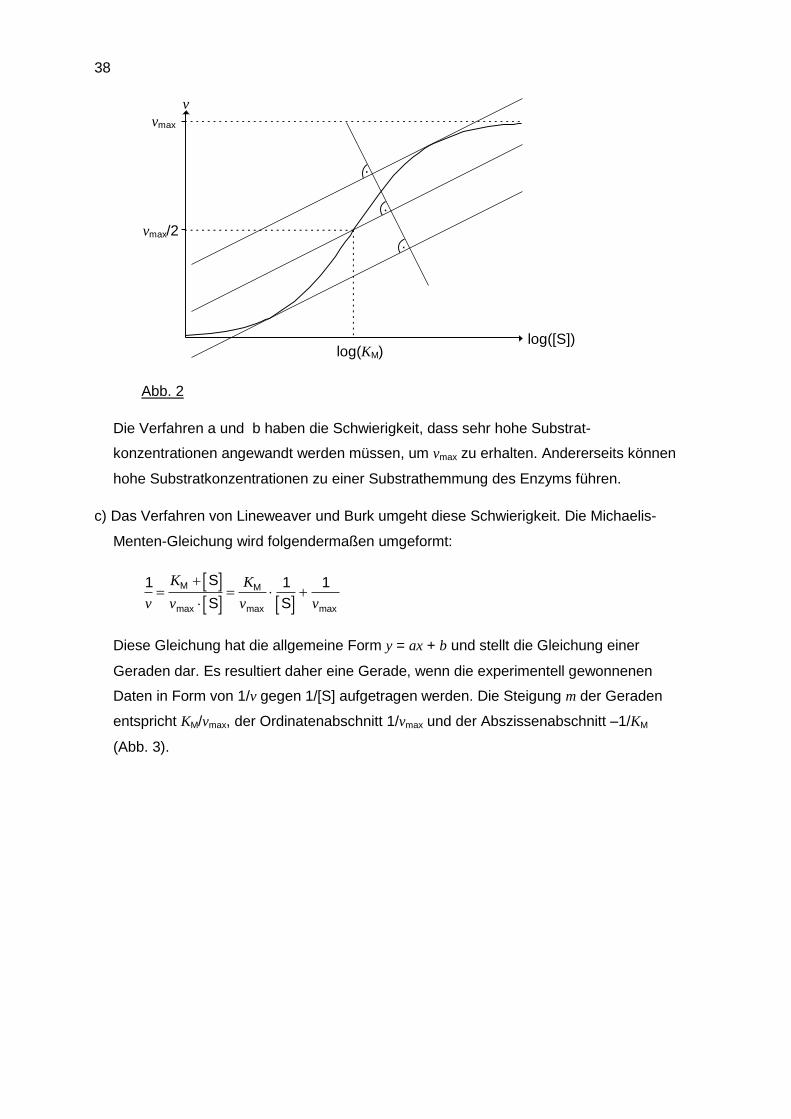
b) v wird gegen log([S]) aufgetragen; es resultiert eine S-förmige Kurve, die Ähnlichkeit mit
der Dissoziationskurve eines Elektrolyten hat. Der Wendepunkt dieser Kurve, der mit
Hilfe zweier paralleler Tangenten und deren Mittelparallelen bestimmt werden kann,
liegt beim Punkt (log(KM), vmax/2) (Abb. 2).
38
log([S])
.
.
.
log(KM)
v
vmax
vmax/2
Abb. 2
Die Verfahren a und b haben die Schwierigkeit, dass sehr hohe Substrat-
konzentrationen angewandt werden müssen, um vmax zu erhalten. Andererseits können
hohe Substratkonzentrationen zu einer Substrathemmung des Enzyms führen.
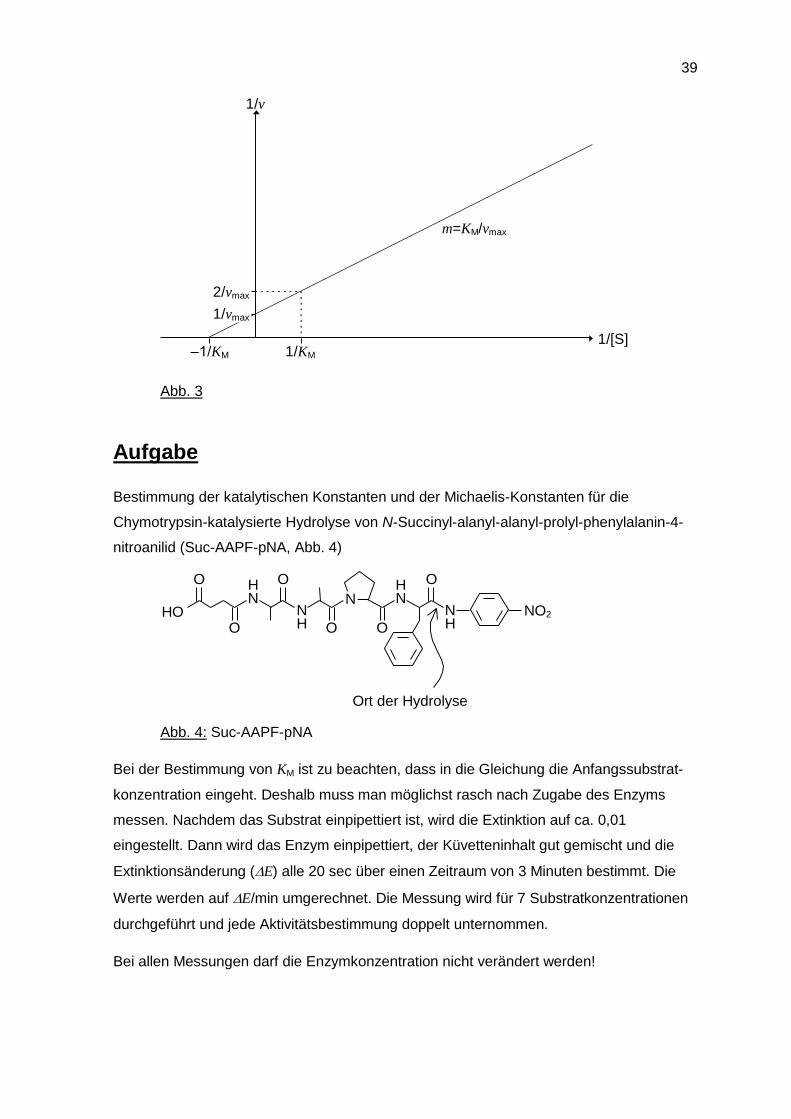
c) Das Verfahren von Lineweaver und Burk umgeht diese Schwierigkeit. Die Michaelis-
Menten-Gleichung wird folgendermaßen umgeformt:
1 1 1
v
K
v
K
v v
M
max
M
max max
S
S S
Diese Gleichung hat die allgemeine Form y = ax + b und stellt die Gleichung einer
Geraden dar. Es resultiert daher eine Gerade, wenn die experimentell gewonnenen
Daten in Form von 1/v gegen 1/[S] aufgetragen werden. Die Steigung m der Geraden
entspricht KM/vmax, der Ordinatenabschnitt 1/vmax und der Abszissenabschnitt –1/KM
(Abb. 3).
39
2/vmax
1/[S]–1/KM
m=KM/vmax
1/KM
1/v
1/vmax
Abb. 3
Aufgabe
Bestimmung der katalytischen Konstanten und der Michaelis-Konstanten für die
Chymotrypsin-katalysierte Hydrolyse von N-Succinyl-alanyl-alanyl-prolyl-phenylalanin-4-
nitroanilid (Suc-AAPF-pNA, Abb. 4)
O HN
HO
Ort der Hydrolyse
NO2
3O
O
NH O
NHN
O
O
NH
Abb. 4: Suc-AAPF-pNA
Bei der Bestimmung von KM ist zu beachten, dass in die Gleichung die Anfangssubstrat-
konzentration eingeht. Deshalb muss man möglichst rasch nach Zugabe des Enzyms
messen. Nachdem das Substrat einpipettiert ist, wird die Extinktion auf ca. 0,01
eingestellt. Dann wird das Enzym einpipettiert, der Küvetteninhalt gut gemischt und die
Extinktionsänderung (E) alle 20 sec über einen Zeitraum von 3 Minuten bestimmt. Die
Werte werden auf E/min umgerechnet. Die Messung wird für 7 Substratkonzentrationen
durchgeführt und jede Aktivitätsbestimmung doppelt unternommen.
Bei allen Messungen darf die Enzymkonzentration nicht verändert werden!

40
Herstellung der Lösungen
Molmassen: Tris: 121,14 g/mol; CaCl2: 147,02 g/mol; Suc-AAPF-pNA: 624,6 g/mol
Verdünnungspuffer (100 mM Tris, 10 mM CaCl2):
1,21 g Tris
147 mg CaCl2
pH 7,8 mit HCl
ad 100 ml Wasser
Substratstammlösung (1 mM):
3,1 mg Suc-AAPF-pNA
ad 5 ml Verdünnungspuffer
Enzymlösung:
2 mg Chymotrypsin ad 1 ml Verdünnungspuffer c = 2 mg/ml;
hiervon 250 l ad 1 ml Verdünnungspuffer c = 0,5 mg/ml;
hiervon 10 l ad 2 ml Verdünnungspuffer c = 0,0025 mg/ml
Pipettierschema:
Für die Messung ist es zweckmäßig, mit stärkster Verdünnung zu beginnen.
Testansatz
0,5 ml der verschiedenen, oben hergestellten Substratverdünnungen
0,1 ml Enzymlösung (0,0025 mg/ml)
Verdünnungspuffer [ml]
1,98 1,96 1,94 1,90 1,80 1,70 1,50
Substratstammlösung [ml]
0,02 0,04 0,06 0,10 0,20 0,30 0,50
Substratkonz. [mM] nach Verdünnen
0,01 0,02 0,03 0,05 0,10 0,15 0,25
Substratkonz. [mM] nach Enzymzugabe
0,0083 0,0167 0,0250 0,0417 0,0833 0,1250 0,2083

41
Auswertung
1) Bestimmen Sie die Anfangsreaktionsgeschwindigkeiten der jeweils vermessenen
Konzentrationen. Auftragung von E gegen die Zeit.
Ermitteln Sie graphisch die Anfangs-Steigung, indem Sie an den Anfang der
Messkurve eine Tangente anlegen.
(Die Messkurve ist eine Hyperbel, die entweder vom Computer an die Messdaten
gefittet oder von Hand näherungsweise eingezeichnet werden kann.)
2) Bestimmen Sie die spezifische Aktivität A für die vermessenen
Substratkonzentrationen nach:
3) Tragen Sie die spezifischen Aktivitäten gegen die Substratkonzentrationen auf
(Michaelis-Menten-Plot) und bestimmen Sie anhand des erhaltenen Graphen KM
und vmax.
4) Ermitteln Sie mit Hilfe der Auftragung nach Lineweaver-Burk ebenfalls KM und die
katalytische Konstante kcat.
Anhand der erhaltenen Geradengleichung lassen sich diese Konstanten
rechnerisch bestimmen.
5) Vergleichen Sie die von Ihnen ermittelten Werte von KM (aus graphischer und
rechnerischer Bestimmung)
6) Ergebnis und Fehlerdiskussion
Benötigte Angaben:
Das bei der Hydrolyse des Substrates freiwerdende 4-Nitroanilin wird in Abhängigkeit von
der Zeit bei 405 nm gemessen. Der molare Extinktionskoeffizient bei dieser Wellenlänge
beträgt 9600 l mol–1 cm–1.
Die spezifische Aktivität A U/ mgmol Substrat
min mg Enzym
errechnet sich nach:
AE V
t d m
106

42
t = Zeit [min]
V = Volumen des Testansatzes in der Küvette [l]
d = Schichtdicke [cm]
m = Enzymmenge in Küvette [mg]
Bei Hydrolasen, zu denen auch Trypsin und Chymotrypsin gehören, verwendet man
häufig anstelle von vmax die katalytische Konstante kcat:
kcat [1/s] = v max [U/mg] · MG [g/mol] / 60 000
MG = Molekulargewicht des Enzyms (hier: 23000 g/mol)
60000: Umrechnungsfaktor von mmol auf mol und von min auf s
In ein Diagramm sind einzutragen:
a) v gegen [S] und
b) v–1 gegen [S]–1,
wobei [S] = jeweilige Substratkonzentration in der Küvette. Anzugeben sind: KM [M],
vmax [U/mg] und kcat [s–1].
Anmerkungen:
Alle Messwerte müssen im Protokoll angegeben werden
Rechenwege müssen klar ersichtlich sein
Evtl. benötigte Pipettierschemata sind anzugeben





























