Embed Size (px)
Citation preview

U.K.M. DeckingJ. Schrader
Räumliche Heterogenitätvon myokardialer Durchblutungund Stoffwechsel
Z Kardiol 90:970–977 (2001)© Steinkopff Verlag 2001
ZFK
753
Eingegangen: 14. Dezember 2000Akzeptiert: 1. Mai 2001
Hauptvortrag,gehalten auf der 66. Jahrestagungder Deutschen Gesellschaftfür Kardiologie –Herz- und Kreislaufforschung27. bis 29. April 2000 in Mannheim
Priv.-Doz. Dr. Ulrich K.M. Decking ())Prof. Dr. Jürgen SchraderInstitut für Herz- undKreislaufphysiologieHeinrich-Heine-Universität DüsseldorfPostfach 10100740001 Düsseldorf, GermanyTel.: 0211 /81-1 2785Fax: 0211 /81-12785E-Mail: decking@uni-duesseldorf-de
Spatial heterogeneity of perfusionand energy turnover in the heart
n Summary Within the left ven-tricular myocardium, substantialdifferences can be observed interms of both perfusion and en-ergy turnover. In addition to thesmall transmural gradient fromthe subepi- to the subendocar-dium (1:1.2), more recent high-resolution studies reveal a majorpatchwork-pattern, e.g., in terms
of flow. Adjacent 200 �l areas candiffer more than 3-fold in localperfusion. Low flow and highflow areas (<50% or >150% ofmean flow, respectively) representup to 1/5 of the left ventricularmyocardium. This local flow pat-tern is temporally stable for atleast days and possibly weeks.Low and high flow areas also dif-fer in local energy metabolism.High flow areas are characterizedby enhanced glucose phosphory-lation and fatty acid permeability,resulting in increased uptake ofthese substrates. This is the basisfor the recent finding that highflow areas are characterized by anenhanced turnover of the citricacid cycle and thus of local O2consumption. Since local O2 sup-ply and consumption are closelycoupled, low flow areas displayno biochemical signs of ischemia.Reducing local flow by 50% re-sults in a similar rise of adeno-sine or lactate in low and highflow areas. Following completecessation of perfusion, high flowareas display a greater risk of in-farction, indicating enhanced en-ergy demand. Further studies areneeded to elucidate the molecularbasis of this spatial heterogeneityand to test whether the 3-fold dif-ferences in local energy turnoverwithin the myocardial wall alsotranslate into comparable varia-tions of local contractility.
n Key words Coronary flow –myocardial perfusion –energy metabolism – heart –13C NMR spectroscopy – review
n Zusammenfassung Im linkenVentrikel steigen unter Ruhebe-dingungen Durchblutung undStoffwechsel vom Epikard zumEndokard hin um etwa 20% an.Auch darüber hinaus bestehen er-hebliche Heterogenitäten, derenbeobachtetes Ausmaß mit steigen-der räumlicher Auflösung zu-nimmt. Betrachtet man z.B. imlinken Ventrikel den lokalen Flussmit einer Auflösung von 200 �l(=mg), so zeigt sich ein mehr als3facher Flussunterschied zwischenz.T. benachbarten Arealen. Nied-rigflussareale (<50% des Mittel-flusses) und Hochflussareale(>150% des Mittelflusses) machenfast 1/5 des Myokards aus. Daslokale Flussmuster ist über Tagebis Wochen stabil. Da am Ge-samt-Organ O2-Verbrauch(MVO2) und koronarer Fluss enggekoppelt sind, können auch auflokaler Ebene erhebliche Unter-schiede im Energiestoffwechselund -Umsatz erwartet werden.
Hochflussareale zeichnen sichdurch eine höhere Desoxyglukose-und Fettsäure-Aufnahme aus, de-nen eine gesteigerte Glukose-Me-tabolisierung bzw. ein erhöhterFettsäure-Transport zu Grundeliegt. Aktuelle 13C NMR spektro-
MYOKARD

971U. K.M. Decking und J. SchraderHeterogenität von Fluss und Stoffwechsel
skopische Untersuchungen zeigen,dass auch die Umsatzrate des Ci-trat-Zyklus und damit der lokaleMVO2 mit dem lokalen Fluss zu-nehmen, und somit in Hoch- undNiedrigflussarealen das Verhältnisvon O2-Angebot und -Verbrauchvergleichbar ist. Dementsprechendsind in Niedrigflussarealen klassi-sche Ischämieparameter wie Ade-nosin und Laktat nicht erhöht.
Wird die lokale Durchblutungauf die Hälfte reduziert, so kommtes in Hoch- und Niedrigflussarea-
len zu einer vergleichbaren Ade-nosin- und Laktat-Zunahme.Niedrigflussareale sind also nichtstärker Ischämie-gefährdet alsHochflussareale. Im Gegenteil, beivollständiger Unterbrechung derDurchblutung zeichnen sich Hoch-flussareale mit ihrem größerenEnergiebedarf durch eine höhereInfarkt-Wahrscheinlichkeit aus.
Der lokale koronare Fluss vari-iert somit innerhalb des linkenVentrikels um mehr als das 3fache.Dieser Fluss-Heterogenität ent-
spricht die räumliche Verteilungvon Glukose- und Fettsäureauf-nahme, MVO2 und Energieumsatz.Ob die beobachteten Stoffwechsel-unterschiede eine Entsprechung inder lokalen Genexpression finden,und ob sie mit einer unterschied-lichen inotropen Reserve einher-gehen, ist noch ungeklärt.
n SchlüsselwörterKoronarer Fluss –Räumliche Heterogenität –Myokardialer Stoffwechsel
Einleitung
Bei der Untersuchung von Durchblutung, Stoffwech-sel und Funktion des Herzens wurde lange ange-nommen, dass zu mindestens der linke Ventrikel einhomogen strukturiertes Gewebe sei, dessen lokaleFunktion sich durch globale Messungen am ganzenHerzen erschließen lässt.
So zeigten Untersuchungen am ganzen Herzenschon in den klassischen Studien von Eckenhoff eineenge Kopplung von koronarer Durchblutung, myokar-dialem Sauerstoffverbrauch und Herzarbeit (18) unddamit auch eine enge gegenseitige Abstimmung vonEnergiebereitstellung und -Verbrauch. Dabei wirdam gut oxygenierten Herzen mehr als 90% des ATP un-ter Sauerstoffverbrauch nach Substrat-Oxidation imCitrat-Zyklus durch mitochondriale oxidative Phos-phorylierung generiert und nur ein verhältnismäßiggeringer Anteil durch anaerobe Glykolyse (33, 37).Das so gebildete ATP steht dann insbesondere fürdie mit der mechanischen Herzaktion verbundenenProzesse zur Verfügung, wobei die Actomyosin-ATPasedie größte Rolle spielt (76%) (39). Aufgrund der hohenSauerstoffextraktion des Herzens (>70%) kann das ko-ronare O2-Angebot schon bei geringen Einschränkun-gen des Flusses zum limitierenden Faktor werden.
Eine homogene Struktur und Funktion legten auchmorphologische Untersuchungen der myokardialenUltrastruktur nahe (41). Doch zeigten zahlreiche Un-tersuchungen der letzten 25 Jahre (z.B. (4, 7, 9, 11, 20,21, 41, 48)), dass die lokale Durchblutung in einemwesentlich größeren Rahmen variiert, als mit eineruniformen Verteilung des lokalen Energieumsatzesund der kontraktilen Funktion vereinbar ist. Die fol-gende Übersicht wird daher die bisher vorliegendenDaten zur räumlichen (und zeitlichen) Heterogenitätder myokardialen Durchblutung zusammenfassen,und mit den in den letzten Jahren erzielten Ergebnis-
sen zur räumlichen Verteilung von Energieumsatz und-Bedarf am Herzen in Beziehung setzen.
Heterogenität der myokardialen Durchblutung
Lang bekannte Unterschiede in der lokalen myokar-dialen Durchblutung finden sich am Herzen zu-nächst zwischen den verschiedenen Kammern, d.h.zwischen Vorhöfen und Ventrikeln einerseits, undzwischen rechter und linker Kammer andererseits((32), zur Übersicht: (29)). Dies ist angesichts der er-heblichen Unterschiede in der jeweiligen Druck-Vo-lumenarbeit nicht verwunderlich.
Darüber hinaus gibt es innerhalb der Kammerneinen transmuralen Gradienten, wobei z.B. am lin-ken Ventrikel unter physiologischen Bedingungender lokale Fluss vom Subepikard zum Subendokardum 10–30% zunimmt. Dieser Gradient kehrt sich beiStenosen in den großen Leitungsgefäßen um (26),was die größere Infarktwahrscheinlichkeit im Sub-endokard z.T. bedingt.
Schließlich findet sich bei hoher räumlicher Auflö-sung auch innerhalb jeder myokardialen Schicht eineerhebliche, und im Grunde überraschende Heteroge-nität der lokalen Durchblutung. Diese wird umsodeutlicher, je kleiner das betrachtete Volumen istund steht im Mittelpunkt dieser Übersicht.
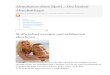
Bestimmt man z.B. am Hundeherzen in vivo dielokale Durchblutung mit radioaktiven Mikrosphären(s.u.) mit einer räumlichen Auflösung von 210 �l(=mg), so zeigt sich eine breite Flussverteilung(Abb. 1A). Während die Mehrzahl der Proben einemittlere Durchblutung aufweisen, erhalten etwa 10%der Proben weniger als 50% des mittleren Sauerstoff-und Substratangebotes und weitere 10% mehr alsdas 1,5fache. Dabei kann die Perfusion zwischen ein-zelnen dieser sogenanten Niedrig- und Hochfluss-areale um mehr als das 10fache variieren.
Die Unterschiede zwischen Niedrig- und Hoch-flussarealen lassen sich nicht auf den transmuralenFlussgradienten zurückführen. Denn während dieDurchblutung von Niedrig- und Hochflussarealen sichum ein mehrfaches unterscheidet, variiert sie zwi-schen Subepi- und Subendokard nur um 10–30%.Und auch innerhalb z.B. des Subepikards findet sicheine erhebliche räumliche Heterogenität der Durch-blutung, wie in Abbildung 1B beispielhaft dargestellt.Diese Heterogenität ist im Prinzip erstaunlich, da wiraus früheren Untersuchungen wissen, dass sich dieseAreale hinsichtlich Kapillar-, Mitochondrien- undMyofibrillendichte nicht unterscheiden (41).
n Messverfahren
Die meisten Versuche zur Erfassung der lokalenDurchblutung wurden mit radioaktiven Mikrosphä-ren durchgeführt. Dieses sind mit �-Strahlern bela-dene, 8–15 �m große, Polystyrol-artige Kügelchen.In den letzten Jahren gewinnen darüber hinaus fluo-reszierende Mikrosphären zunehmend an Bedeutung(24). Mikrosphären werden als Bolus über eine kurzeZeit in den linken Vorhof injiziert, nach Durch-mischung in der linken Kammer mit dem Blutstromin die Peripherie transportiert, und dort aufgrundihrer Größe unmittelbar präkapillär abgelagert. AmVersuchsende wird dann das zu untersuchende Ge-webe entnommen, in einzelne Proben (Areale) ge-teilt, und die Depositionsdichte in jeder einzelnenProbe bestimmt.
Unter idealen Bedingungen entspräche die lokaleMikrosphären-Depositionsdichte dem lokalen Volu-menstrom der Erythrozyten, doch könnten allein dieunterschiedliche Größe, Oberflächenstruktur undDichte einen erheblichen Einfluss auf die lokale Depo-sition ausüben. Um die Validität der Mikrosphären-
technik nachzuweisen, wurden Mikrosphären zusam-men mit molekularen Plasmafluss-Markern (z.B. Des-methylimipramin) injiziert. Dabei korrelierte der mitbeiden Methoden gemessene Fluss eng (r=0,93) (6).Allerdings streute die Depositionsdichte der Mik-rosphären etwas stärker als die des molekularen Fluss-Markers; die Variationskoeffizienten betrugen0,38±0,09 bzw. 0,33±0,10. Da die Verteilung derMikrosphären auf einzelne Gewebsareale ein stochas-tischer Prozess ist, erfordert eine reliable Flussbestim-mung eine bestimmte Anzahl von Mikrosphären (ca.200–400) pro Probe (38), was die räumliche Auflösungdieser Technik auf ca. 50 �l (=mg) begrenzt. Ins-gesamt kann davon ausgegangen werden, dass mit8–15 �m Mikrosphären der lokale Fluss auch in klei-nen Proben adäquat bestimmt werden kann (15).
Prinzipiell ist die Bestimmung der lokalen myo-kardialen Durchblutung auch mit klinisch einsetz-baren Techniken möglich. Doch reicht die räumlicheAuflösung der gebräuchlichen SPECT und PET-Ver-fahren nicht an die o. g. Dimensionen heran. In derMagnetresonanz-Bildgebung kann die lokale Durch-blutung nach Kontrastmittelgabe („first pass“) überdie lokale Intensitätsänderung qualitativ abgeschätztwerden. Doch gelingt es hier bisher nur, 6–8 Seg-mente voneinander zu differenzieren (1). Ein alter-natives MR-Verfahren stellt die räumliche Darstel-lung der sogenannten Relaxationszeit T1 nach selek-tiver Anregung des koronaren Einstroms dar, das ei-ne höhere räumliche Auflösung verspricht, aber bisjetzt nur in wenigen Studien validiert wurde (46).
n Räumliche Heterogenität der Durchblutung
Eine räumliche Heterogenität der Durchblutung findetsich in allen bisher untersuchten Spezies (u. a. Hunde,Schafe, Paviane, Kaninchen) (z.B. (4, 6, 13, 21, 25, 32,
972 Zeitschrift für Kardiologie, Band 90, Heft 12 (2001)© Steinkopff Verlag 2001
Abb. 1 Räumliche Heterogenität dermyokardialen Durchblutung, bestimmtin der freien Wand des linken Ventrikelsam blut-perfundierten Hunde-Herzenin situ in Proben mit einer durchschnitt-lichen Masse von 210 mg. Links: RelativeHäufigkeit, mit der einzelne Proben defi-nierten Fluss-Klassen zugeordnet werdenkonnten. Rechts: Räumliche Darstellungdes lokalen Flusses eines repräsentativenHerzens. Die Graustufen entsprechen de-nen der in A dargestellten Flussvertei-lung

41)). Sie wird häufig über die Breite der Flussvertei-lung charakterisiert (s. Abb. 1A), die durch ihren Va-riationskoeffizienten (CV) beschrieben werden kann.Dieser beträgt z.B. sowohl im Schaf- als auch im Hun-deherzen bei einer Probenmasse von 200 mg etwa 0,35(6, 13, 41). Da in die Breite der Flussverteilung auchder o.g. transmurale Gradient eingeht, ist der Varia-tionskoeffizient in jeder einzelnen transmuralenSchicht, etwa im Midmyokard, verständlicherweise et-was geringer (z.B. 0,24±0,03 (15)).
Wie in Abbildung 1B zu sehen ist, können soge-nannte Niedrigfluss- unmittelbar neben Hochfluss-arealen liegen. Dies legt schon intuitiv nahe, dassdie beobachtete räumliche Heterogenität mit gerin-gerer räumlicher Auflösung allein aufgrund von Mit-telungseffekten abnimmt. Bei einer rein zufälligenVerteilung dürften ab einer bestimmten Arealgrößekeine Unterschiede zwischen den einzelnen Arealenzu beobachten sein. Doch zeigt sich bei genauererAnalyse der lokalen Flussverteilung, dass Niedrig-und Hochflussareale eben nicht rein zufällig verteiltsind, sondern ihre Anordnung durch eine fraktaleGesetzmäßigkeit beschrieben werden kann. D.h.,dass bei verschiedener räumlicher Auflösung(0,05–1 g) sich auf jeder Auflösungsebene bestimmteUnterschiede wiederfinden. In verschiedenen Studienwurde für die räumliche Heterogenität des Blutflus-ses eine fraktale Dimension von 1,2–1,3 bestimmt (5,17, 43), die zwischen einer streng geordneten und ei-ner rein zufälligen Verteilung (D=1,0 bzw. 1,5) liegt.
Entsprechend der nicht-zufälligen Verteilung vonHoch- und Niedrigflussarealen zeigte sich auch eineCluster-artige Anordnung. Hier weisen die unmittel-bar einem Hochflussareal benachbarten Areale mithöherer Wahrscheinlichkeit einen höheren Fluss auf,und ebenso die einem Niedrigflussareal benachbar-ten Areale einen niedrigeren Fluss. Über alle Fluss-bereiche betrug der Korrelationskoeffizient für be-nachbarte Areale zwischen 0,4 und 0,6 (5, 17, 43).
Interessanterweise entsprechen sich die fraktaleDimension der lokalen Durchblutung und des Tei-lungsmusters des arteriellen Gefäßbaumes (45), undes wurde daher versucht, die Heterogenität des Flus-ses über das Teilungsmuster des Gefäßbaumes zu er-klären (45). Doch zeigen Untersuchungen zur loka-len Koronarreserve (4) (s. u.), dass sowohl Niedrig-als auch Hochflussareale noch eine Dilatationsreser-ve haben. Auch finden sich keine Hinweise auf einelokale Ischämie in Niedrigflussarealen (35, 41) (s. u.).Daher ist es nur schwer vorstellbar, dass ein statischverstandener Gefäßbaum die lokale Flussverteilungentscheidend bestimmen sollte. Angesichts des dyna-mischen Charakters der arteriellen Gefäßversorgungam Herzen („Angiogenese“) erscheint es viel wahr-scheinlicher, dass die beobachtete Gefäßstruktur dasMuster eines lokalen Bedarfs widerspiegelt.
n Zeitliche Heterogenität der Durchblutung
Die bisher publizierten Daten zeigen, dass die loka-len Flussmuster (Abb. 1B) über mindestens 24 hkonstant bleiben. Wurde die lokale Durchblutungmit Mikrosphären zu unterschiedlichen Zeiten ge-messen, so korrelierte bei gleichen hämodyna-mischen Bedingungen der lokale Fluss in hohemMaße. Dies galt für Zeitabstände von 5 min (4),30–75 min (13, 34) und 120 min (27). Die längstebisher publizierte Studie konnte eine hohe Stabilitätsogar über 28 h in einem Tier belegen (32). Vorläufi-ge Daten aus unserem eigenen Labor zeigen, dass anwachen Hunden Niedrig- bzw. Hochflussareale übermindestens 2 Wochen Niedrigfluss- bzw. Hochfluss-areale bleiben. Bei einem insgesamt 3fachen Fluss-Unterschied zwischen Hoch- und Niedrigflussarealenfluktuiert hier der lokale Fluss z.B. in Niedrigfluss-arealen nur um etwa 20–30% (31), was die hohezeitliche Stabilität des Flussmusters unterstreicht.
Ein eindrucksvoller Beleg für die geringe zeitlicheVariabilität des lokalen Flusses in Ruhe sind Ver-suche, in denen durch adrenerge Stimulation Herz-arbeit und Durchblutung gesteigert wurden. Hierkam es in jedem einzelnen Areal im Mittel zu einervergleichbaren absoluten Flusszunahme – und90 min nach Ende der Stimulation hatte fast jedesAreal wieder sein Ausgangsniveau erreicht (16).
Räumliche Heterogenitätdes myokardialen Stoffwechsels
Vor dem Hintergrund der zeitlich stabilen räumli-chen Heterogenität des lokalen Flusses liegt der Ver-dacht nahe, dass die erheblichen Unterschiede im lo-kalen Fluss Unterschiede im lokalen Energieumsatzund letztlich Energiebedarf widerspiegeln. Denn dieaußergewöhnlich hohe arterio-venöse Differenz imSauerstoffgehalt am ganzen Herzen schließt aus, dassAreale, deren lokaler Fluss weniger als die Hälfte desmittleren Flusses beträgt, ein ausreichendes O2-An-gebot für einen mittleren Sauerstoffverbrauch haben,und die enge Kopplung von kardialer Arbeit, Sauer-stoffverbrauch und koronarem Fluss am ganzen Her-zen macht eine enge Abstimmung auch auf lokalerEbene wahrscheinlich. Und in der Tat weisen zahlrei-che Untersuchungen der letzten Jahre darauf hin,dass sich Areale mit einer niedrigen bzw. hohenDurchblutung (Niedrig- bzw. Hochflussareale) sub-stantiell in ihren Stoffwechselraten unterscheiden.Jüngste Studien konnten erstmalig belegen, dass derlokale Energieumsatz und O2-Verbrauch der lokalenDurchblutung entspricht und in analoger Weise hete-rogen verteilt ist.
973U. K.M. Decking und J. SchraderHeterogenität von Fluss und Stoffwechsel

n Lokale Metabolitspiegel
Da Niedrigflussareale mit ihrem geringeren O2-Ange-bot Hypoxie- oder Ischämie-gefährdet sein könnten,wurde in ersten Untersuchungen die Beziehung zwi-schen dem lokalen Fluss und lokalen Metabolitspie-geln analysiert. Niedrigflussarealen zeichneten sichdabei durch einen nur geringfügig niedrigeren ATP-Gehalt und keine Unterschiede im Glykogen-Gehaltaus (21). Auch waren weder der lokale Laktat-Gehalt(13, 35) noch die lokale freie Adenosin-Konzentration,gemessen mit der SAH-Technik, erhöht (35, 41). DieseBefunde belegen, dass Niedrigflussareale nicht hypo-xisch sind, d.h. dass in diesen Arealen O2-Angebotund Bedarf im Gleichgewicht miteinander stehen.
n Lokale Enzymaktivitäten
In einem weiteren Ansatz wurde die Aktivität vonSchlüsselenzymen des Energiestoffwechsels in Ab-hängigkeit vom lokalen Fluss bestimmt. Es fand sicheine geringe Zunahme der lokalen Kreatinkinase(CK)- und Lactatdehydrogenase (LDH)-Aktivität mitdem lokalen Fluss (21) sowie eine 30% höhere Hexo-kinase-Aktivität in Hochflussarealen (41). Auch dieAktivität der Succinatdehydrogenase, ein mitochon-driales Enzym, nahm proportional mit dem lokalenFluss zu (9). Hingegen wurde kein Unterschied inder Aktivität der Phosphoglycerat-Kinase sowie dermitochondrial lokalisierten Enzyme Cytochrom-c-Oxidase und Citrat-Synthase beobachtet (41). Da-mit ermöglicht die z.T. erhöhte Enzymaktivität inHochflussarealen u.U. einen erhöhten Umsatz desEnergiestoffwechsels, aber gibt keinen Hinweis aufdas Ausmaß der Unterschiede im Energieumsatzzwischen Niedrig- und Hochflussarealen.
n Lokale Substrataufnahme
Eine Voraussetzung für Unterschiede im Energie-umsatz ist eine entsprechende Substrataufnahme. InÜbereinstimmung mit einem gesteigerten Umsatz inHochflussarealen wurde eine erhöhte Aufnahme vonradioaktiv markierten Fettsäuren (10, 28) in Hoch-flussarealen beobachtet. Auch wurde eine Zunahmeder Desoxyglukose-Aufnahme mit dem lokalen Flussam Hundeherzen (41), nicht aber am Schweineher-zen berichtet (19). Eine geringere Aufnahme mar-kierter Substrate im Niedrigflussbereich könnte prin-zipiell durch ein vermindertes Substrat-Angebot be-dingt sein. Daher wurden mathematische Modellana-lysen durchgeführt, deren wesentliche Parameter dasSubstrat-Angebot, die Membran-Permeabilität unddie intrazelluläre Verstoffwechselung waren. Es zeigte
sich, dass für langkettige Fettsäuren die Membran-Permeabilität, genauer das kapilläre Permeabilitäts-Oberflächenprodukt, mit dem lokalen Fluss anstieg(10), während für Desoxyglukose die intrazellulärePhosphorylierung (durch die Hexokinase) mit demlokalen Fluss zunahm (14), entsprechend der höhe-ren Hexokinase-Aktivität in Hochflussarealen (s. o.).
n Lokaler Energieumsatz
Eine direkte Messung des lokalen Sauerstoffver-brauchs und Energieumsatzes ist schwierig. FrüheArbeiten in den 70er Jahren bestimmten den lokalenPO2 durch intramyokardiale Elektroden (36) und be-richteten einen weiten Bereich von PO2-Werten.Doch blieb offen, ob nicht die Elektroden die lokaleDurchblutung erheblich beeinflussten. In anderenStudien wurde die intrakapilläre oder venöse O2-Sät-tigung erfasst und übereinstimmend eine geringereO2-Sättigung in Endokard-nahen Bereichen gesehen(22, 30). Dies deutete zusammen mit der gesteiger-ten subendokardialen Durchblutung auf eine trans-murale Zunahme im Sauerstoffverbrauch hin, abererlaubte keine Analyse der räumlichen Heterogenitätdes Sauerstoffverbrauchs.
Da mit hoher räumlicher Auflösung (z.B. 200 �l)eine klassische Messung des lokalen O2-Verbrauchsüber die arterio-venöse O2-Gehaltsdifferenz nichtmöglich ist, mussten indirekte Verfahren entwickeltwerden, um die räumliche Heterogenität des O2-Ver-brauchs zu bestimmen.
Ein Ansatz hierbei war die Bestimmung des Um-satzes des Citrat-Zyklus, der die Reduktionsäquiva-lente für die Atmungskette bereitstellt und somit di-rekt an die oxidative Phosphorylierung gekoppelt ist.Hierzu bot sich der Einsatz 13C markierter Substratean, deren Metabolisierung im Citratzyklus mittels13C NMR Spektroskopie verfolgt werden kann.Frühere Studien hatten an salin perfundierten Her-zen gezeigt, dass nach Gabe von 13C markiertemAcetat die Markierung der einzelnen Kohlenstoffato-me des mitochondrialen �-Ketoglutarats sich im re-lativ größeren cytosolischen Glutamat-Pool wieder-spiegelt (47, 49). Die Kinetik der Markierung einzel-ner Glutamat-Kohlenstoffatome war in diesen Studi-en direkt mit der Umsatzrate des Citrat-Zyklusverknüpft. Vor diesem Hintergrund entwickelten un-abhängig voneinander Hans van Beek in Amsterdamund Ulrich Decking in Düsseldorf Verfahren, um auseiner Endpunktbestimmung nach Gabe von 13C-Ace-tat bzw. 13C-Pyruvat in einzelnen Myokard-Probendie Umsatzrate des Citratzyklus zu bestimmen (12,42, 44).
Nach Gabe von an Position C3 markiertem13C-Pyruvat wird im Citrat-Zyklus �-Ketoglutarat zu-
974 Zeitschrift für Kardiologie, Band 90, Heft 12 (2001)© Steinkopff Verlag 2001
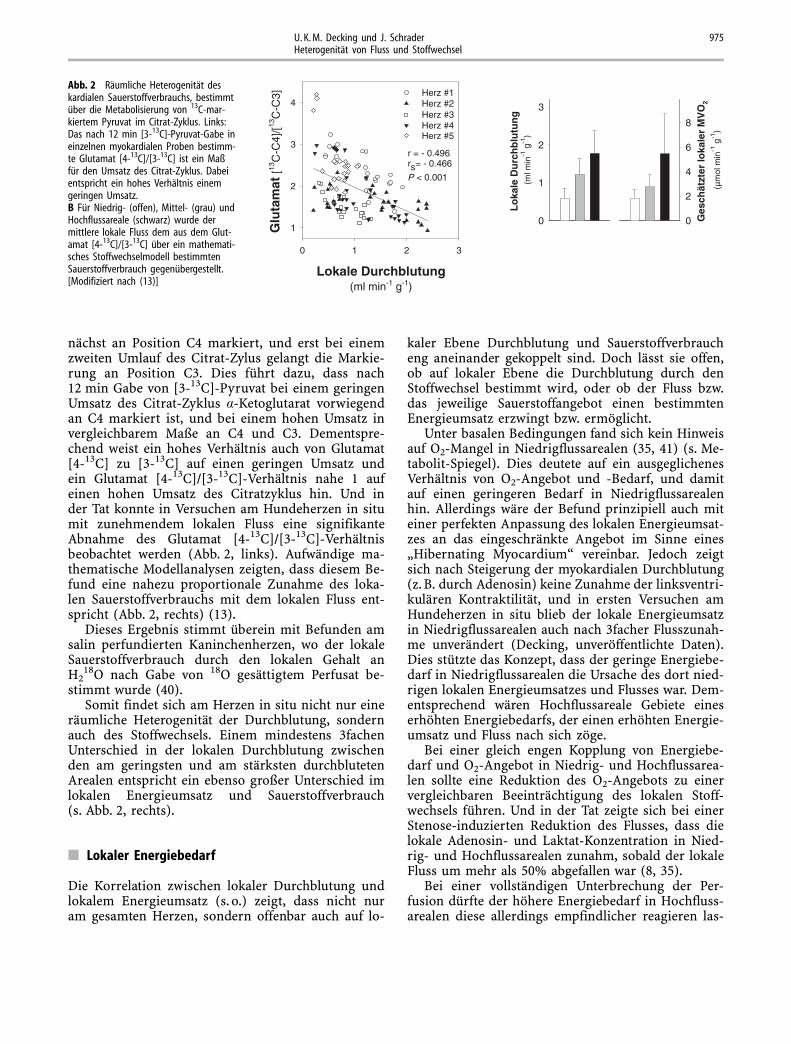
nächst an Position C4 markiert, und erst bei einemzweiten Umlauf des Citrat-Zylus gelangt die Markie-rung an Position C3. Dies führt dazu, dass nach12 min Gabe von [3-13C]-Pyruvat bei einem geringenUmsatz des Citrat-Zyklus �-Ketoglutarat vorwiegendan C4 markiert ist, und bei einem hohen Umsatz invergleichbarem Maße an C4 und C3. Dementspre-chend weist ein hohes Verhältnis auch von Glutamat[4-13C] zu [3-13C] auf einen geringen Umsatz undein Glutamat [4-13C]/[3-13C]-Verhältnis nahe 1 aufeinen hohen Umsatz des Citratzyklus hin. Und inder Tat konnte in Versuchen am Hundeherzen in situmit zunehmendem lokalen Fluss eine signifikanteAbnahme des Glutamat [4-13C]/[3-13C]-Verhältnisbeobachtet werden (Abb. 2, links). Aufwändige ma-thematische Modellanalysen zeigten, dass diesem Be-fund eine nahezu proportionale Zunahme des loka-len Sauerstoffverbrauchs mit dem lokalen Fluss ent-spricht (Abb. 2, rechts) (13).
Dieses Ergebnis stimmt überein mit Befunden amsalin perfundierten Kaninchenherzen, wo der lokaleSauerstoffverbrauch durch den lokalen Gehalt anH2
18O nach Gabe von 18O gesättigtem Perfusat be-stimmt wurde (40).
Somit findet sich am Herzen in situ nicht nur eineräumliche Heterogenität der Durchblutung, sondernauch des Stoffwechsels. Einem mindestens 3fachenUnterschied in der lokalen Durchblutung zwischenden am geringsten und am stärksten durchblutetenArealen entspricht ein ebenso großer Unterschied imlokalen Energieumsatz und Sauerstoffverbrauch(s. Abb. 2, rechts).
n Lokaler Energiebedarf
Die Korrelation zwischen lokaler Durchblutung undlokalem Energieumsatz (s. o.) zeigt, dass nicht nuram gesamten Herzen, sondern offenbar auch auf lo-
kaler Ebene Durchblutung und Sauerstoffverbraucheng aneinander gekoppelt sind. Doch lässt sie offen,ob auf lokaler Ebene die Durchblutung durch denStoffwechsel bestimmt wird, oder ob der Fluss bzw.das jeweilige Sauerstoffangebot einen bestimmtenEnergieumsatz erzwingt bzw. ermöglicht.
Unter basalen Bedingungen fand sich kein Hinweisauf O2-Mangel in Niedrigflussarealen (35, 41) (s. Me-tabolit-Spiegel). Dies deutete auf ein ausgeglichenesVerhältnis von O2-Angebot und -Bedarf, und damitauf einen geringeren Bedarf in Niedrigflussarealenhin. Allerdings wäre der Befund prinzipiell auch miteiner perfekten Anpassung des lokalen Energieumsat-zes an das eingeschränkte Angebot im Sinne eines„Hibernating Myocardium“ vereinbar. Jedoch zeigtsich nach Steigerung der myokardialen Durchblutung(z.B. durch Adenosin) keine Zunahme der linksventri-kulären Kontraktilität, und in ersten Versuchen amHundeherzen in situ blieb der lokale Energieumsatzin Niedrigflussarealen auch nach 3facher Flusszunah-me unverändert (Decking, unveröffentlichte Daten).Dies stützte das Konzept, dass der geringe Energiebe-darf in Niedrigflussarealen die Ursache des dort nied-rigen lokalen Energieumsatzes und Flusses war. Dem-entsprechend wären Hochflussareale Gebiete eineserhöhten Energiebedarfs, der einen erhöhten Energie-umsatz und Fluss nach sich zöge.
Bei einer gleich engen Kopplung von Energiebe-darf und O2-Angebot in Niedrig- und Hochflussarea-len sollte eine Reduktion des O2-Angebots zu einervergleichbaren Beeinträchtigung des lokalen Stoff-wechsels führen. Und in der Tat zeigte sich bei einerStenose-induzierten Reduktion des Flusses, dass dielokale Adenosin- und Laktat-Konzentration in Nied-rig- und Hochflussarealen zunahm, sobald der lokaleFluss um mehr als 50% abgefallen war (8, 35).
Bei einer vollständigen Unterbrechung der Per-fusion dürfte der höhere Energiebedarf in Hochfluss-arealen diese allerdings empfindlicher reagieren las-
975U. K.M. Decking und J. SchraderHeterogenität von Fluss und Stoffwechsel
Abb. 2 Räumliche Heterogenität deskardialen Sauerstoffverbrauchs, bestimmtüber die Metabolisierung von 13C-mar-kiertem Pyruvat im Citrat-Zyklus. Links:Das nach 12 min [3-13C]-Pyruvat-Gabe ineinzelnen myokardialen Proben bestimm-te Glutamat [4-13C]/[3-13C] ist ein Maßfür den Umsatz des Citrat-Zyklus. Dabeientspricht ein hohes Verhältnis einemgeringen Umsatz.B Für Niedrig- (offen), Mittel- (grau) undHochflussareale (schwarz) wurde dermittlere lokale Fluss dem aus dem Glut-amat [4-13C]/[3-13C] über ein mathemati-sches Stoffwechselmodell bestimmtenSauerstoffverbrauch gegenübergestellt.[Modifiziert nach (13)]

sen. Entsprechend dieser Vermutung wurde einehöhere Infarktwahrscheinlichkeit bei Ischämie inHochflussarealen beobachtet (23). Diese räumlichenUnterschiede im Energiebedarf könnten u.U. die kli-nische Beobachtung erklären, dass bei chronischermyokardialer Ischämie (z.B. bei lang bestehenderAngina Pectoris ohne bekanntem Infarkt) sich häufigdisseminierte Nekrosen entwickeln.
Perspektiven
Wie oben gezeigt variiert am Herzen in situ innerhalbder freien Wand des linken Ventrikels die Durchblu-tung in einem beachtlichen Ausmaß, und dies bereitsunter basalen Bedingungen. Dieser räumlichen Hete-rogenität der Durchblutung entspricht eine räumlicheHeterogenität des Sauerstoffverbrauchs und Energie-umsatzes, die offensichtlich (s. o.) durch eine räumli-che Verteilung des lokalen Energiebedarfs bedingt ist.Die bisherigen Forschungsergebnisse lassen nochzahlreiche Fragen offen:1) Angesichts der zeitlichen Stabilität der beobachte-
ten Flussmuster stellt sich die Frage, ob die räum-liche Heterogenität des Energiebedarfs z.B. durchlokale Unterschiede in der Wandspannung auf-grund der funktionellen Anordnung der einzelnenMuskelfasern bedingt ist.
2) Der lokal unterschiedliche Energieumsatz dürftemit einer Variabilität in der lokalen kontraktilen
Funktion gekoppelt sein. Doch ist es bisher nichtgelungen, die lokale myokardiale Arbeit mit aus-reichender räumlicher Auflösung zu untersuchen.Einsatz spezieller, hoch-auflösender Verfahren derMR-Bildgebung (z.B. DENSE; Displacement enco-ding with stimulated echoes) (2, 3) dürfte es inZukunft ermöglichen, die lokale Perfusion und dielokale Wandbewegung in einem Experiment zu er-fassen, und so auf lokaler Ebene Fluss und Funk-tion miteinander zu korrelieren. Mit diesem An-satz könnte auch untersucht werden, ob Arealemit basal niedriger Durchblutung bei Steigerungder Herzarbeit vorzugsweise rekrutiert werden,und so u.U. eine funktionelle Reserve darstellen.
3) Angesichts der nahezu chronischen Unterschiedeim lokalen Fluss und Energieumsatz ist zu ver-muten, dass sich auch auf molekularer Ebene Un-terschiede zwischen Niedrig- und Hochflussarea-len nachweisen lassen. Diese könnten z.B. einerAdaptation an eine lokal unterschiedliche funktio-nelle Beanspruchung entsprechen, oder auch diemolekulare Grundlage für den lokal verschiedenenFluss und Energiebedarf darstellen.
n Danksagung Wir danken allen Doktoranden, Mitarbeitern undKollegen, die gemeinsam mit uns die Heterogenität von myokar-dialer Durchblutung, Stoffwechsel, Gen- und Proteinexpressionuntersuchen, insbesondere Stefan Skwirba, Alexander Janosi, EvaBergschneider sowie Prof. Andreas Deussen und Prof. Volker Thä-mer.
976 Zeitschrift für Kardiologie, Band 90, Heft 12 (2001)© Steinkopff Verlag 2001
Literatur
1. Al-Saadi N, Nagel E, Gross M, Born-stedt A, Schnackenburg B, Klein C,Klimek W, Oswald H, Fleck E (2000)Noninvasive detection of myocardialischemia from perfusion reservebased on cardiovascular magneticresonance. Circ 101:1379–1383
2. Aletras AH, Balaban RS, Wen H(1999) High-resolution strain analysisof the human heart with fast-DENSE.J Magn Reson 140:41–57
3. Aletras AH, Ding S, Balaban RS, WenH (1999) DENSE: displacement en-coding with stimulated echoes in car-diac functional MRI. J Magn Reson137:247–252
4. Austin RE Jr, Aldea GS, Coggins DL,Flynn AE, Hoffman JIE (1990) Pro-found spatial heterogeneity of coron-ary reserve. Circ Res 67:319–331
5. Bassingthwaighte JB, King RB, RogerSA (1989) Fractal nature of regionalmyocardial blood flow heterogeneity.Circ Res 65:578–590
6. Bassingthwaighte JB, Malone MA,King RB, Chan IS, Link JM, KrohnKA (1990) Molecular and particulatedepositions for regional myocardialflows in sheep. Circ Res 66:1328–1344
7. Bassingthwaighte JB, Malone MA,Moffett TC, King RB, Little SE, LinkJM, Krohn KA (1987) Validity of mi-crosphere depositions for regionalmyocardial flows. Am J Physiol 253:H184–H193
8. Bussemaker J, Groeneveld ABJ, Teer-link T, Hennekes M, Westerhof N,Van Beek JHGM (1997) Low- andhigh-blood flow regions in the nor-mal pig heart are equally vulnerableto ischaemia during partial coronarystenosis. Pflügers Arch 434:785–794
9. Bussemaker J, Van Beek JHGM, Groe-neveld ABJ, Hennekes M, Teerlink T,Thijs LG, Westerhof N (1994) Localmitochondrial enzyme activity corre-lates with myocardial blood flow atbasal workloads. J Mol Cell Cardiol26:1017–1028
10. Caldwell JH, Martin GV, RaymondGM, Bassingthwaighte JB (1994) Re-gional myocardial flow and capillarypermeability-surface area productsare nearly proportional. Am J Physiol267:H654–H666
11. Conway RS, Weiss HR (1985) Depen-dence of spatial heterogeneity ofmyocardial blood flow on meanblood flow rate in the rabbit heart.Cardiovasc Res 19:160–168
12. Decking UKM (1998) 13C-MR spec-troscopy – how and why? MAGMA6:103–104
13. Decking UKM, Skwirba S, Zimmer-mann MF, Preckel B, Thämer V,Deussen A, Schrader J (2000) Spatialheterogeneity of energy turnover inthe heart. Pflügers Archiv, Ref Type(In Press)
14. Deussen A (1997) Local myocardialglucose uptake is proportional to, butnot dependent on blood flow. Pflü-gers Arch 433:488–496

977U. K.M. Decking und J. SchraderHeterogenität von Fluss und Stoffwechsel
15. Deussen A (1998) Blood flow hetero-geneity in the heart. Basic Res Cardi-ol 93:430–438
16. Deussen A, Flesche CW, Lauer T,Sonntag M, Schrader J (1996) Spatialheterogeneity of blood flow in thedog heart. II. Temporal stability inresponse to adrenergic stimulation.Pflügers Arch 432:451–461
17. Deussen A, Sonntag M, Flesche CW,Vogel RM (1997) Minimal effects ofnitric oxide on spatial blood flowheterogeneity of the dog heart. Pflü-gers Arch 433:727–734
18. Eckenhoff JE, Hafkenschiel JH, Land-messer CM, Harmel M (1947) Cardi-ac oxygen metabolism and control ofthe coronary circulation. Am J Phy-siol 149:634–649
19. Fallavollita JA (2000) Spatial hetero-geneity in fasting and insulin-stimu-lated (18)F-2-deoxyglucose uptake inpigs with hibernating myocardium.Circ 102:908–914
20. Falsetti HL, Carroll RJ, Marcus ML(1975) Temporal heterogeneity ofmyocardial blood flow in anesthe-tized dogs. Circ 52:848–853
21. Franzen D, Conway RS, Zhang H,Sonnenblick EH, Eng C (1988) Spa-tial heterogeneity of local blood flowand metabolite content in dog hearts.Am J Physiol 254:H344–H353
22. Gamble WJ, LaFarge CG, Fyler DC,Weisul J, Monroe RG (1974) Regionalcoronary venous oxygen saturationand myocardial oxygen tension fol-lowing abrupt changes in ventricularpressure in the isolated dog heart.Circ Res 34:672–681
23. Ghaleh B, Shen Y-T, Vatner SF (1996)Spatial heterogeneity of myocardialblood flow presages salvage versusnecrosis with coronary artery reper-fusion in conscious baboons. Circ94:2210–2215
24. Glenny RW, Bernard S, Brinkley M(1993) Validation of fluorescent-la-beled microspheres for measurementof regional organ perfusion. J ApplPhysiol 74:2585–2597
25. Gonzales F, Bassingthwaighte JB(1990) Heterogeneities in regional vo-lumes of distribution and flows inrabbit heart. Am J Physiol 258:H1012–H1024
26. Griggs DM, Nakamura Y (1968) Ef-fect of coronary constriction on myo-cardial distribution of iodoantipy-rine-131I. Am J Physiol 215:1082–1088
27. Groeneveld ABJ, Van Lambalgen AA,Van den Bos GC, Bronsveld W, NautaJJP, Thijs LG (1991) Maldistributionof heterogenous coronary blood flowduring canine endotoxin shock. Car-diovasc Res 25:80–88
28. Groeneveld ABJ, Visser FC (1993)Correlation of heterogeneous bloodflow and fatty acid uptake in the nor-mal dog heart. Basic Res Cardiol88:223–232
29. Hoffman JIE (1995) Heterogeneity ofmyocardial blood flow. Basic Res Car-diol 90:103–111
30. Holtz J, Grunewald WA, Manz R,Restorff W v, Bassenge E (1977) In-tracapillary hemoglobin oxygen sa-turation and oxygen consumption indifferent layers of the left ventricularmyocardium. Pflügers Arch 370:253–258
31. King RB, Bassingthwaighte JB (1989)Temporal fluctuations in regionalmyocardial flows. Pflügers Arch413:336–342
32. King RB, Bassingthwaighte JB, HalesJRS, Rowell LB (1985) Stability ofheterogeneity of myocardial bloodflow in normal awake Baboons. CircRes 57:285–295
33. Kobayashi K, Neely JR (1979) Controlof maximum rates of glycolysis in ratcardiac muscle. Circ Res 44:166–175
34. Loncar R, Flesche CW, Deussen A(1997) Determinants of the S-adeno-syl-homocysteine (SAH) techniquefor the local assessment of cardiacfree cytosolic adenosine. J Mol CellCardiol 29:1289–1305
35. Loncar R, Flesche CW, Deussen A(1998) Coronary reserve of high- andlow-flow regions in the dog heart leftventricle. Circ 98:262–270
36. Lösse B, Schuchardt S, Niederle N(1975) The oxygen pressure histo-gram in the left ventricular myocar-dium of the dog. Pflügers Arch350:121–132
37. Mallet RT, Hartman DA, Bünger R(1990) Glucose requirement for post-ischemic recovery of perfused work-ing heart. Eur J Biochem 188:481–493
38. Polissar NL, Stanford DC, Glenny RW(2000) The 400 microsphere per piece“rule” does not apply to all bloodflow studies. Am J Physiol 278:H16–H25
39. Schramm M, Klieber H-G, Daut J(1994) The energy expenditure ofactomyosin-ATPase, Ca2+-ATPase andNa+,K+-ATPase in guinea-pig cardiacventricular muscle. J Physiol (Lond)481:647–662
40. Schwanke U, Deussen A, Heusch G,Schipke JD (2000) Heterogeneity oflocal myocardial flow and oxidativemetabolism. Am J Physiol 279:H1029–H1035
41. Sonntag M, Deussen A, Schultz J,Loncar R, Hort W, Schrader J (1996)Spatial heterogeneity of blood flow inthe dog heart. I. Glucose uptake, freeadenosine and oxidative/glycolyticenzyme activity. Pflügers Arch 432:439–450
42. van Beek JH, Csont T, de Kanter FJ,Bussemaker J (1998) Simple modelanalysis of 13C NMR spectra to mea-sure oxygen consumption using fro-zen tissue samples. Adv Exp MedBiol 454:475–485
43. Van Beek JHGM, Roger SA, Bas-singthwaighte JB (1989) Regionalmyocardial flow heterogeneity ex-plained with fractal networks. Am JPhysiol 257:H1670–H1680
44. Van Beek JHGM, van Mil HGJ, KingRB, de Kanter FJJ, Alders DC, Busse-maker J (1999) A 13C NMR double-labeling method to quantitate localmyocardial O2 consumption usingfrozen tissue samples. Am J Physiol277:H1630–H1640
45. VanBavel E, Spaan JAE (1992)Branching patterns in the porcinecoronary arterial tree: Estimation offlow heterogeneity. Circ Res 71:1200–1212
46. Waller C, Kahler E, Hiller KH, Hu K,Nahrendorf M, Voll S, Haase A, ErtlG, Bauer WR (2000) Myocardial per-fusion and intracapillary blood vo-lume in rats at rest and with coron-ary dilatation: MR imaging in vivowith use of a spin-labeling technique.Radiology 215:189–197
47. Weiss RG, Gloth ST, Kalil-Filho R,Chacko VP, Stern MD, Gerstenblith G(1992) Indexing tricarboxylic acid cy-cle flux in intact hearts by carbon-13nuclear magnetic resonance. Circ Res70:392–408
48. Yipintsoi T, Dobbs WA, Scanton PD,Knopp TJ, Bassingthwaighte JB(1973) Regional distribution of diffu-sible tracers and carbonized micro-spheres in the left ventricle of iso-lated dog hearts. Circ Res 33:573–587
49. Yu X, White LT, Doumen C, DamicoLA, LaNoue KF, Alpert NM, Lewan-dowski ED (1995) Kinetic analysis ofdynamic 13C NMR spectra: Metabolicflux, regulation, and compartmenta-tion in hearts. Biophys J 69:2090–2102