Embed Size (px)
Citation preview

Band 101 1 Samec und Dermelj, Studien fiber Pflanzenkolloide, XLVII. 123 Heft 2 (1942)A
Aus dem Chemischen lnstitut tier k6nigl. Universitiit Lubiana, ltalien.
Studien iiber Pflanzenkolloide, XgVII. S~iurehydrolyse jodkolorimetr isch differenzierter St~rkesubstanzen.
Yon M. S a m e c und M. De r me l j (Lub iana , I falien).
Auf (3rund zahlreicher kolloidchemischer Merkmale schlossen wir, dal~ St~irkesubstanzen, welche sich in ihrer Jodfarbe unterscheiden, von- einander strukturchemisch unterschieden sein mtigtenl). Die verschiedenen zur Uberprfifung dieser Annahme unternommenen Arbeiten haben die Richtigkeit der Idee dargetan 2, 3, 4); sie wurde in neuester Zeit auch yon anderen Forschern ange- nommen und experimentell gesttitzt 5, 6, 7).
Ausgehend yon dem Gedanken, dag bei Vor- handensein versehiedener struktureller Bindun- gen auch der Verlauf der S~iurehydrolyse Unter- schiede erkennen lassen kiJnnte, haben wir schon vor l~ingerer Zeit den Verlauf solcher Spaltungen an den einzelnen beim Druckkochen des Kartof- felamylopektins hintereinander erhaltenen Solen verfolgt~). Wenn auch mit 0,5 normaler Schwefel- s~ure bei 800 C erhaltene Zahlenwerte ftir die zeitliche Anderung des Reduktionsvermtigens und der optischen Drehung bei allen beobachteten Solen sehr nahe aneinander lagen, wurden schon damals gewisse Unterschiede beobachtet. Sole, welche vornehmlich Erythrosubstanzen enthiel- ten, wie zum Beispiel das Sol III und Sol IV1), zeigten anfangs ein gr~l~eres Reduktionsverm/Jgen als die Amylosen-Sole (Soll und If), doch stieg wfihrend der Hydrolyse das Reduktionsvermtigen der letzteren Sole rascher an als das der ersteren, so dag man den Eindruck gewann, es besttinde bei den Erythroki~rpern eine gewisse Reaktions- hemmung.
In einem gewissen Widerspruche mit diesen Beobachtungen stand die Angabe yon F r e u d e n- bergS), dag in der Sfiurehydrolyse zwischen den
i) M. Samec und M. Blinc, Kolloid-Beih. 30, 163 (1930).
3) M. Samec, Kolloid-Beih. 33, 103 (1931). 8) M. Samec und E. Waldschmidt-Lei tz ,
Hoppe-Seylers Z. physiol. Chem. 203, 16 (1931). 4) M. Samec und K. Hess, zit. in M. Samec,
Kolloid-Beih. 51, 359 (1940), und zwar SeRe 429. ~) K. Myrb~ck und B. Oertenblad, Skand.
Arch. Physiol. 80, 334 (1938). 6) K.H. Meyer, M.Wertheim und P. Bern-
feld, Helv. chim. acta 23, 865 (1940). 7) K. Myrb~ck und K. Ahlborg, Biochem. Z.
307, 70 (1940). 8) K. Freudenberg, W. Kuhn, W. Dfirr, F.
Boltz und G. Steinbrunn, Ber. Dtsch. chem. Ges. 63, 1510 (1930).
(Eingegangen am 10. Juli 1942)
Amylosen und dem Amylopektin kein Unter- schied feststellbar ist. Aus diesem Grunde fiber- prtiften wit die seinerzeitigen Messungen, und zwar mit 0,5proz. H~SO 4 bei 800 und mit 50proz. Schwefelsfiure bei 400 C an Amylosen, Erythro- amylosen, an tier Erythrogranulose und am Amylodextrin Meyer.
Die beiden ersteren erhielten wir aus Kartofs felstfirke durch Druckkochen und Elektrodekan- tieren. Die Amylosen wurden nach Isttindigem Kochen eines 3proz. Kleisters bei 1200 und Ent- fernen des Amylopektins gewonnen, die Erythro- amylosen resultierten im Sol IV und V nach je 2sttindigem Druekkochen des Amylopektinsl).
Zur Darstellung der Erythrogranulose wurde nach O. A. van K l i n k e n b e r g Lintnerstfirke mit fl-Amylose hydrolysiert. Das Enzym wurde nach B l o m - B a k - B r a a e aus Oerste bereitet und naeh dem Wijsmann 'schen Diffusionsversuch auf die Einheitlichkeit tiberprtift. Die mit Azeton aus- gefiillte ErythrogranuIose zeigte anfangs ein starkes Reduktionsvermt~gen (16,76 Proz. T. M. W.); nach 5 maligem Umf~illen mit Alkohol der Endkonzentration von 60 Proz. resultierte ein Prfiparat vom Reduktionsverm~gen 0,67 T.M.W. Die spezifische Drehung betrug aD = 190,5 bis 191,50 .
Das Amylodextrin Meyer wurde genau nach den Angaben des Autors dargestellt. Wit arbeite- ten mit Portionen yon je l kg, aus welchen ]e 250 g Rohdextrin erhalten wurde. Dieses wurde 3mal mit Azeton und 5mal mit Alkohol umkristal- lisiert. Die optische Drehung, welche anfangs an = 186--1870 betrug, stieg beim Umkristalli- sieren auf aD = 194 ~ AuS 4 kg St~rke resultier- ten 6 g Dextrin, dessert charakteristische Merk- male sich beim Umkristallisieren nicht mehr ~inderten.
Zur Ausftihrung der Hydrolyse mit der 0,5- normalen Sfiure wurden gleiche Teile tier passend verdtinnten S~iure und L~isung der Stfirkesub- stanz auf die Reaktionstemperatur (80 oc) ge- bracht, gemischt, sofort die far die erste Messung notwendige Portion abgezogen und auf Zimmer- temperatur abgektihlt. Vom Augenblick tier Misehung bis zur Ausftihrung tier Messung sind 3--5 Minuten verflossen.
124 Samec und Dermelj, Studien fiber Pflanzenkolloide, XLVII. [" Kolloid- k Z e i t s c h r i f t
Fiir die Hydrolyse mit 50proz. Sfiure wurden die beiden Flfissigkeiten auf 0 o C abgekfihlt und nach dem Mischen sofort auf40 ~ C gebracht. Der Anfangswert wurde nach Erreichen der Tempe- ratur bestimmt (1 Minute nach dem Mischen).
Das ReduktionsvermSgen wurde nach Auer- bach-Bodl~ inder gemessen und auf Glukose ge- rechnet. Es entspricht I ccm einer 1/i on Jod- 15sung 9,005 nag Glukose oder 18,01 mg Maltose.
Die Sole waren 0,5- bis 0,6proz., die Konzen- tration sank nach der Mischung mit S~iure auf 0,25 bis 0,30 Proz.; in 10 ccm - - welche Menge ffir eine Reduktionsbestimmung verwendet wurde - - waren also 0,02 his 0,03 g organische Substanz ent- halten. Die genaue Menge derselben wurde sowohl dutch die Trockengehaltsbestimmung der Aus- gangslSsung als auch titrimetrisch nach totaler Hydrolyse ermittelt. In den folgenden Tabellen ffihren wir unter q den Anfangsgehalt der Mi- schung, unter x den umgesetzten Anteil (als Glu- kosewert), unter a o den Drehungswinkel an.
Die Tabellen I--VI best~itigen die Angab_en yon F r e u d e n b e r g , dab im Verlauf der S~iure- hydrolyse bei den einzelnen Sfiirkesubstanzen keine gr~Beren Unterschiede wahrzunehmen sind. Das optische DrehungsvermSgen nimmt anfangs langsam, sp~iter rascher ab, der Gang ist bei allen untersuchten Substanzen ein analoger. Das Bild findert sich jedoch, wenn man den Gang der nach der Gleichung monomolekularer Reaktionen ge- rechneten Reaktionskonstante betrachtet. Es liegt an der Hand, dab dieser Weft im Verlauf der Hydrolyse nicht konstant bleiben kann, weil ja die Reaktion nicht monomolekular ist. Die Ar t , wie diese Konstante v a r i i e r t , ist jedoch bei den
Tabe l l e I. A m y l o - a m y l o s e n . T = 80 ~ 1/1 n H2S04, a = 0,02944,
1 k = �9 loglo ~ �9 2,302585. 1 2
T a ~ x
15 30 45 60 90
120 150 180 185 240 300
Proz. Spaltung
0 4,62 9,48
14,05 18,35 26,25 33,65 40,90 48,90 50,00 62,70 76,40
a D
180,3 175,2 170,1 165,0 161,6 153,0 146,3 141,2 134,4 132,7 125,8 110,5
0,00136 0,00279 0,00414 0,00540 0,00774 0,00990 0,01205 0,01440 0,01472 0,01845 0,2250
k" 10 a
3,13 3,31 3,36 3,37 3,39 3,41 3,51 3,73 3,74 4,10 4,81
250 500 750
1000 1250 1500 1750 2000 2250 2500
Tabe l le II. A m y l o - A m y l o s e n . T = 80 ~ 1/2 n H2S04, a = 0,03165.
Proz. Spaltung
15,35 27,30 36,95 46,60 56,30 64,25 71,70 78,70 84,30 85,65
x k . 104
0,00486 6,67 0,00864 6,38 0,01170 6,15 0,01477 6,28 0,01780 6,61 0,02035 6,87 0,02270 7,22 0,02490 7,73 0,02670 8,25 0,02708 8,79
Tabe l le Ill. A m y l o - A m y l o s e n . T = 40 ~ 50proz. H2SO~, a = 0,03061 (in 10ccm
Proz. t Spaltung aD X k" 104
0 6O
120 240 360 690
1200 1530 1800 2100 2700 3030 3480 4260 4680 4908 5520 5760 6120 7080
1,47 2,94 5,88 7,33
12,30 25,90 31,80 35,90 40,55 48,80 52,90 58,15 66,20 68,90 71,80 76,10 78,20 79,50 84,10
Tabe l le IV. T
203,3 214,4 0,00045 207,0 0,00090 190,4 0,00180 188,4 0,00261 181,6 0,00477 168,2 0,00792
0,00972 0,01099 0,01242 0,01495 0,01620 0,01782 0,02025 0,02105 0,02195 0,02330 0,02395 0,02430 0,02575
2,46 2,48 2,53 2,47 2,45 2,49 2,49 2,47 2,47 2,48 2,48 2,50 2,54 2,48 2,53 2,59 2,64 2,58 2,60
E r y t h r o - A m y l o s e n . = 40 ~ 50proz. H2SO~, a = 0,03160.
Proz. t Spaltung aD x k . 104
0 60
1035 1500 2400 2940 3550 3840 4380 5220 5820 6840 7260 8220
10050 11430
19,10 25,07 37,35 43,30 50,00 53,00 57,90 64,90 67,25 70,60 72,15 74,35 78,30 79,10
187,7 185,0
122,0
103,5 96,4 87,7 77,2 66,6 63,1 63,1
0,00603 0,00792 0,01180 0,01368 0,01580 0,01675 0,01830 0,02050 0.02125 0,02230 0,02280 0,02350 0,02475 O,O25OO
2,05 1,92 1,95 1,93 1,95 1,97 1,98 2,00 1,92 1,79 !,76
. , 1,66 1,52 1,37
Band 10l "] Samec und Dermelj, Studien fiber Pflanzenkolloide, XLVII. 125 Heft 2 (1942)J
T a b e l l e V . E r y t h r o - G r a n u l o s e . T = 40 ~ 50proz. H~SO~, a = 0,02880.
Proz. t 8paltung x k" 10 ~
0 120 240 480 840
1380 1860 2280 2820 3500 4760 6180 7080
2,19 4,38
7 , 1 9 13,75 21,57 29,40 35,62 42,55 50,00 63,80 76,80 82,95
0,00063 0,00126 0,00207 0,00396 0,00621 0,00846 0,01025 0,01225 0,01440 0,01830 0,02195 0,02390
1,84 1,86 1,55 1,76 1,76 1,86 1,92 1,96 1,98 2,10 2,32 2,50
T a b e l l e VI. A m y l o - D e x t r i n Meyer . T = 40 ~ 50proz. H~SO~, a = 0,2650.
120 180 3O0 480 720
1260 1800 2660 2820 4260 4740 5585 6540 7560 9900
11550
Proz. (a)20 t Spaltung
0 192,1 192;1
4,08 190,2 6,79 190,2
10,86 188,2 15,96 184,3 27,20 174,5 36,75 160,8 50,00 137,0 52,30 131,4 69,3O 107,8 73,75 98,4 80,15 93,5 85,30 86,2 89,30 78,4
70,5 66,6
0,001C 0,001~ 0,0028 0,004~ 0,0072 0,0097 0,0132 0,0138 0,0183 0,0195 0,021~ 0,022E 0,023C
k �9 104
2,31 2,34 2,39 2,41 2,51 2,54 2,60 2,62 2,76 2,82 2,90 2,93 2,95
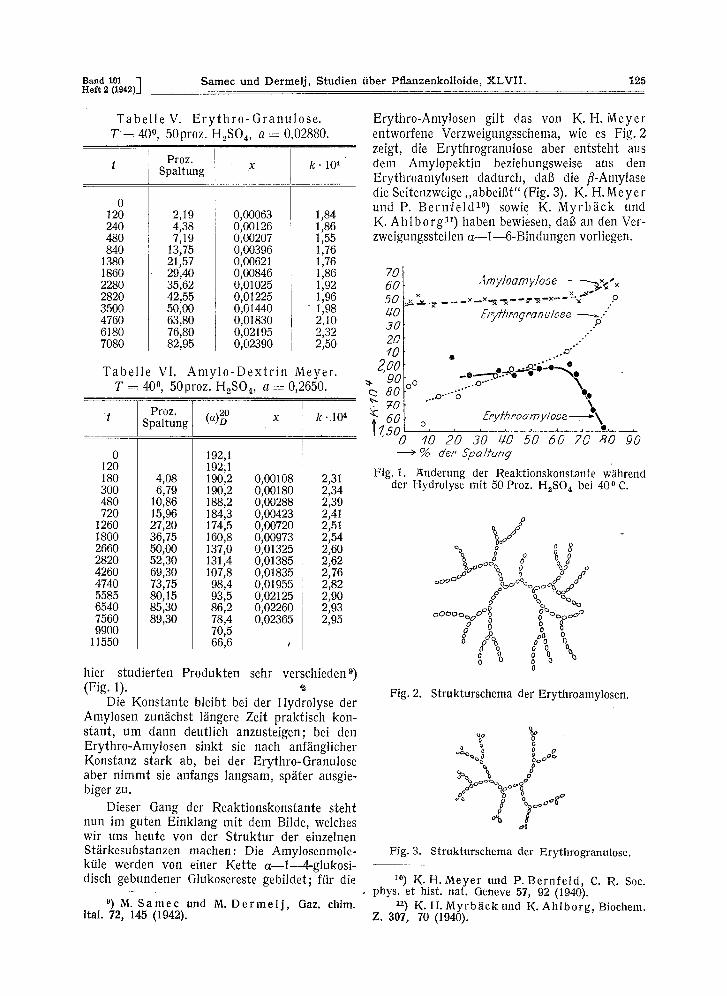
hier studierten Produkten sehr verschieden 9) (Fig. 1).
Die Konstante bleibt bei der Hydrolyse der Amylosen zunfichst lfingere Zeit praktisch kon- stant, um dann deutlich anzusteigen; bei den Erythro-Amylosen sinkt sie nach anffinglicher Konstanz stark ab, bei der Erythro-Granulose aber nimmt sie anfangs langsam, sp~ter ausgie- biger zu.
Dieser Gang der Reaktionskonstante steht nun im guten Einklang mit dem Bilde, welches wit uns heute v o n d e r Struktur der einzelnen Stfirkesubstanzen machen: Die Amylosenmole- ktile werden yon einer Kette a--l--4-glukosi- disch gebundener Glukosereste gebildet; ffir die
9) M. S a m e c und M. D e r m e l j , Oaz. chim. ital. 72, 145 (1942).
Erythro-Amylosen gilt das von K .H. Meyer entworfene Verzweigungsschema, wit es Fig. 2 zeigt, die Erythrogranulose abet entsteht aus dem Amylopektin beziehungsweise aus den Erythroamylosen dadurch, dab die fl-Amylase die Seitenzweige ,,abbeigt" (Fig. 3). K. H .Meyer und P. B e r n f e l d 1~ sowie K. Myrb / ick und K. A h l b o r g n) haben bewiesen, dab an den Ver- zweigungsstellen a~l - -6-Bindungen vorliegen.
70 6O 50 zlO 30 20 r
2,oo ~. 9 0 <b 8O
6 0
r o
Am y/oam y/o se - ----~ ,,.',, :~ x ~,~"?'~ x 0
9~yfhrogranulose --~.p:
. . . "
�9 ~ o .o '~* _e__~. o..~%_,=~,o:o o-~,
C . . . 0 "~
. . ,o-- '%'" %
o ffrythrournylose ", ~ .
?o Jo so #o 6o 7o 9'0 - - ~ % der Spa/ fc , ng~
Fig. 1. Anderung der Reaktionskonstante w~hrend der Hydrolyse mit 50 Proz. H~SO 4 bei 40 0 C.
Fig. 2.
g o o
8oO ~ o g Loo4 Uo
ao ~ ~o o o oo oooo o ~176 oo0 o 0 0 - -
O 0 0 0 C ) ~ O ~ 0 ~ 0 o 0 0
0 0 0 0 o o ? o ~ 0 0 % 0
o~ 0 0 % 0 [3 0 0
0
Strukturschema der Erythroamylosen.
o o a o
~ o o oo
oo>OO%oiO
4 )' o0
Fig. 3. Strukturschema der Erythrogranulose.
lo) K. H. Meyer und P. Bernfeld, C. R. Soc. phys. et hist. nat. Oeneve 57, 92 (1940).
zz) I.(. H. Myrb~ick und K. AMborg, Biochem. Z. 307, 70 (1940).

126 Samec und Dermelj, Studien fiber Pflanzenkolloide, XLVII. 1- Kolloid- kZeitschrift
Bei der Hydrolyse der Amylosen entstehen zunehmend kleinere Teilchen, welche sich immer mehr der Maltose nfihern. Da die Hydrolysen- konstante dieses Zuckers gri313er ist als die der Spaltung a--1 4-glukosidischer Bindungen in der Amylose, ist es verstfindlich, dag man bei dieser St~irkesubstanz einen Anstieg der Hydro- lysengeschwindigkeit beobachtet.
Im Falle der Erythro-Amylose werden die a - - 1 4-Bindungen hydrolysiert, und es h~iufen sich die yon den Zweigstellen stammenden schwe- rer spaltbaren a--l--6-Bindungen an: die Reak- tionsgeschwindigkeit nimmt ab.
Umgekehrt steht es mit der Erythrogranu- lose: hier linden sich zahlreiche endst~indige a--1--6-Bindungen neben dem Gros der a--1 d- Bindungen vor. In dem Mal3e, in welchem die ersferen gespalten werden, schwindet der schwerer hydrolysierbare Anteil aus der Molekel, die Reak- tionsgeschwindigkeit nimmt zu, sie steigt abet fihnlich wie bei den Amylosen auch wegen der N/iherung an die Maltosehydrolyse an.
Von diesem (?esichtspunkte aus deckt also auch die Sfiurehydrolyse Verschiedenheiten zwi- schen den ]odkolorimetrisch differenzierten St/ir-
kesubstanzen auf, und zwar in einer Weise, welche vorzaglich zu dem heute geltenden Strukturbilde der St~irkesubstanzen pal3t.
Z u s a m m e n f a s s u n g . 1. Im Rahmen der Versuche, Unterschiede
zwischen den jodkolorimetrisch differenzierten St~irkesubstanzen zu finden, wurde neuerlich die Sfiurehydrolyse derselben studiert.
2. Im Verlaufe dieser Hydrolyse ~indert sich das Reduktionsvermbgen und die optische Dre- hung aller untersuchten Produkte in einer kaum unterschiedlichen Weise.
3. Deutliche Unterschiede ergeben sich je- doch, wenn man den Gang der Reaktionskon- stante ffir monomolekulare Reaktionen verfolgt. Bei den Amylosen nimmt nach anffinglicher Non- stanz diese t(onstante zu, bei den Erythro-Amy- losen nimmt sie ab, bei der Erythro-Granulose steigt sie anfangs langsam, sp~iter rasch an.
4. Diese Befunde lassen sich durch Vergleich der Hydrolysengeschwindigkeiten der Maltose- bindung in der Maltose, der a--l--4-Bindung in l~ingeren Ketten und 0er a--l--6-Bindung gut verstehen.
Aus clem Laboratorium for Mikrobiologie tier kOnio~l. Universitdt Lubiana, ltaUen.
Versuche zur Anre icherung des Mazeransenzyms . V o n M. B l i n c ( L u b i a n a , I ta l ien) . (Eingegangen am 29. Juli 1942)
Uber die (3ewinnung von EnzymliJsungen aus Kulturen von Bacillus macerans berichten uns E.B. T i lden und C. S. H u d s o n 1,2). Die ge- nannten Forscher zogen den Bazillus auf Kar- toffelbrein/ihrb/Jden bei 38~ und filtrierten darauffolgend die Kulturen durch ein N-Berke- feld-Fil ter . Die so gewonnene Enzymlbsung verflfissigte Kartoffelkleister und ffihrte zur Bil- dung tier typischen Schardinger-Dextr ine , wobei der Reaktionsverlauf im Kleister viskosi- metrisch verfolgt werden konnte.
Im hiesigen Laboratorium wurde die Enzym- gewinnung aus Kulturen yon Bacillus macerans lfingere Zeit bearbeitet und durch zweijfihrige ununterbrochene Untersuchungen mit einigen gut
1) E. B. Tilden und C. S. Hudson, J. Amer. chem. Soc. 61, 2900 (1939).
2) Fast gleichzeitig erhielten K. Freudenberg, E. Schaaf, G. D'umpert und T. Plaetz [Naturwiss. 27, 850 (1939)] Schardinger-Dextrine aus St~irke unter Benutzung bakterienfrei filtrierter Mazerans- Kulturen.
anwendbaren Erfahrungen bereichert~). Wie diese Versuche zeigen, ist zur Enzymgewinnung ein besonderer Weft auf gesunde und starke Bazillusst~imme zu legen. Kommt es zu Dege- nerationserscheinungen - - sei es in mikologi- scher Ht~sicht (Form, (Jr613e, Wachstum), sei es im Hinblick auf die physikochemischen Eigen- schaften des gfirenden Substrates, ist begreiflicher- weise eine befriedigende Enzymanreicherung nicht mehr zu erwarten.
Kr~iftige Mazeransst/imme hfingen vor allem von einem richtig gewfihlten Substrat ab. Wir empfehlen den Bazillus vorerst nach Is Freu- denberg 4) abwechselnd je 10 Tage auf Fleisch- wasser, Kartoffelkeilen und auf Kartoffelbrei bei 380 C zu ziehen. Nach etlichen Wochen ist der Stamm, welcher fortwfihrend mikroskopisch zu beobachten ist, soweit gest~irkt, dab man ihn
~) M. Blinc, Arch. Mikrobiologie 12, 183 (1941). 4) t ( .Freudenberg und R.Jakobi , Liebigs
Ann. 518, 103 (1935).





























