Embed Size (px)
Citation preview

G. Losse, H . Stange und W. Naumann 143
Liebigs Ann. Chem. 1981, 143 - 151
Synthese von Cystinpeptiden mit einer der aktiven Region des Insulins angenaherten Primarstruktur
Giinter Lome *, Hans Stange und Wolfgang Naumann
Sektion Chemie der Technischen Universitat Dresden, MommsenstraDe 13, DDR-8027 Dresden
Eingegangen am 8. April 1980
Die Synthese des unsymmetrischen Disulfids 1 [B 15 -27(Ala 27) - A 19-21], eines Ausschnitts aus der aktiven Insulinregion, wird beschrieben. Hiervon ausgehend wird durch Aufbau der Di- sulfide B 15-27 (Ala 27)-A 20-21, B 15-20-A 19-21, B 15-27 (SEt 19, Ala 27) und [B 15 -27 (Ala 27)], (2-5) die Primarstruktur schrittweise vereinfacht. Die Herstellung der ent- sprechenden Disulfidbindungen erfolgte hierbei durch saurekatalysierten Disulfidaustausch zwi- schen den beiden Teilkomponenten mit SEt-geschutzten Cysteinfunktionen. Die dargestellten Cystinpeptidea) weisen eindeutige, jedoch um den Faktor 10- bis reduzierte Insulinwirkun- gen auf.
Synthesis of Cystine Peptides with a Primary Structure Related to the Active Region of Insulin The synthesis of the unsymmetrical disulfide 1 [B 15 - 27(Ala 27) - A 19 - 211, a section of the active insulin region, is described. Starting from this material, stepwise simplification of the primary structure was performed by preparing the disulfides B 15-27 (Ala 27)-A 20-21, B 15-20-A 19-21, B 15-27 (SEt 19, Ala 27), and [B 15-27 (Ala 27)12 (2 -5) . The corre- sponding disulfide bonds were established by proton catalysed disulfide exchange between the two partial components containing SEt-protected cysteine functions. The synthesized cystine pep- tides exhibit distinct but by the factor of to reduced insulin activities.
In der Struktur-Wirkungsforschung des Insulins wurden bisher zahlreiche Sequenz- variationen und Substitutionsprodukte auf biologische Aktivitat getestet'). Die sequenzvariierten Insuline wurden hierzu entweder durch Kombination einer totalsyn- thetischen oder einer semisynthetischen, durch Einbeziehung biochemischer Methoden abgewandelten Teilkette mit der nativen Gegenkette und weiterhin durch semisyntheti- sche Umwandlungen am Gesamtmolekul erhalten '). Die so gewonnenen Informationen uber die Abhangigkeit der biologischen Aktivitat von der Art der Sequenzanderung wurden durch die Aufklarung der Sequenzen von Insulinen unterschiedlicher tierischer Provenienz erganzt 2,3) . Aus dem Vergleich dieser verschiedenen Nativstrukturen lassen sich die Sequenzbereiche ablesen, wo die Aminosauren variant sind und sich Sequenz- veranderungen am wenigsten aktivitatsmindernd auswirken.
Weitere wesentliche Fortschritte zu diesem Fragenkomplex brachte die rontgenstruk- turanalytische Aufklarung der Raumstruktur des Insu l inm~nomeren~-~) und der fur
a) Nomenklatur und Abkurzungen nach den Regeln der IUPAC-IUP Commission on Biochemi- cal Nomenclature. - J. Biol. Chern. 241, 2496 (1966), 247, 977 (1972).
Liebigs Ann. Chem. 1981
Liebigs Ann. Chem. 1981, Heft 1
0 Verlag Chemie, GmbH, D-6940 Weinheim, 1981 0170-2041/81/0101-0143 $ 02.50/0
10

144 G. Losse, H. Stange und W. Naumann
die biologische Aktivitat verantwortlichen Struktureinheit '). Dadurch wurden die ein- zelnen, fur die Stabilisierung der Raumstruktur mangebenden Bindungsbeitrage be- stimmter Aminosauren erkannt 5,7). Aus den so gewonnenen strukturanalytischen Da- ten gelang es schlieljlich einer internationalen Arbeitsgruppe unter Einbeziehung von CD-Messungen, Rezeptorbindungsversuchen sowie anhand der Aktivitatsabstufungen bekannter Sequenzanaloger die rezeptorbindende Region zu ermitteln4,7). Danach sind an dieser bevorzugt der Sequenzabschnitt A 1 - 7 der A-Kette, insbesondere durch die Aminosauren Gly A 1 und Glu A 5 , sowie der Abschnitt A 19 - 21, speziell durch Tyr A 19, beteiligt. Fur die B-Kette ergeben sich wesentliche Anteile im Bereich B 12-26 durch Val B 12, Tyr B 16, Arg B 22, Gly B 23, Phe B 24, B 25 und Tyr B 26. Zentrale Stellungen in dieser aktiven Region nehmen weiter die Disulfidbrucken A 20 - B 19 und die Salzbindung zwischen Asn A 21 und Arg B 22 ein.
Schema 1. Aktive Region von Insulin nach BIundelf et (stark schematisiert)
1 (2-4) 5 ( 6 ) 7 (8-18) 1 9 20 21
Gly-Glu-Cys-Tyr-Cys- Asp I .
S I
? - - - - - - - - Val-Cys- A>g-Gly- Phe- Phe-Tyr -
12 (13-18) 1 9 (20-21) 22 23 24 2 5 26
Insgesamt erlauben es die angefuhrten Ergebnisse, fur die meisten Aminosaureposi- tionen qualitative Aussagen zum Wirkungsabfall bei Austausch oder Substitution funktioneller Gruppen in dieser Position, zu ihrer Varianz in verschiedenen nativen In- sulinspecies, ihrem Beitrag zur Stabilisierung der Insulinraumstruktur sowie ihrer Be- teiligung an der rezeptorbindenden Region zu geben ').
Vom Standpunkt der Strukturverfeinerung verdeutlichen alle diese Erkenntnisse in guter Ubereinstimmung, darj in der B-Kette der Verlust bestimmter terminaler Positio- nen relativ gut toleriert wird, wobei sich insbesondere das Fehlen der Aminosauren B 1 - 3 und B 27 - 30 nur unwesentlich auf die biologische Aktivitat auswirkt. Umge- kehrt mussen in der A-Kette die beiden terminalen Abschnitte A 1 - 7 und A 19 - 21 einschliefllich der von dort ausgehenden charakteristischen Disulfidanordnungen als es- sentiell fur die Aktivitat angesehen werden.
Wir beschrieben fruher eine neue, verbesserte Synthesekonzeption fur die Insulin-B- Kette, die zu einem einheitlichen, mit Nativkette identischen Endprodukt von voller in- sulinbildender Potenz fuhrte 9,10). Damit waren gleichzeitig die Voraussetzungen fur die Gewinnung neuartiger Analoga in Reinheitsgraden gegeben, die prazisere biologische Aktivitatsangaben zulassen.
Ausgehend von dem geschilderten Stand der Insulin-Struktur-Wirkungsforschung, insbesondere von den Vorstellungen iiber die aktive Region des Insulins sowie Befun- den von Weitzel"), nach denen einfache Argininpeptide aus dem Sequenzabschnitt B 22 - 26 bereits eine - wenn auch sehr schwache - insulinahnliche Aktivitat an iso- lierten Fett- und Leberzellen aufweisen, entschieden wir uns fur die Synthese der asym- metrischen Disulfide 1 bis 4 sowie des symmetrischen Disulfides 5. Als Bezugsverbin-
Liebigs Ann. Chem. 1981

Synthese von insulinahnlichen Cystinpeptiden 145
dung enthalt 1 wesentliche Teile der von BIundeN et al. 4,7) geforderten Struktur fur die aktive Region, jedoch unter Auslassung des N-terminalen A-Kettenabschnittes. Gleich- zeitig wurde der Sequenzabschnitt B 22 - 2611) integriert und die als tolerierbar erkann- te vereinfachende Substitution 12-14) von Thr (27) der Nativstruktur gegen Ala vorge- nommen. Gegenuber 1 werden bei 2 - 4 schrittweise weitere Verkurzungen im B- und A-Kettenteil ausgefiihrt, wobei fur 3 besonders ein Effekt durch das Fehlen der Region B 22 - 26 erwartet wurde. Bei 5 wird schlieRlich der wesentliche B-Kettenausschnitt als symmetrisches Cystinderivat gewonnen.
Schema 2
1 5 16 1 7 1 8 19 2 0 2 1 22 2 3 2 4 2 5 26 2 1
Leu-Tyr-Leu-Val-Cys-Gly-Glu-Arg-Gly-Phe-Phe-Tyr-Ala
I Tyr-Cys-Asn
1 9 2 0 2 1
1: B 15-27 (Ala 27)-A 1S-21
Leu-Tyr-Leu-Val-Cys-Gly-Glu-Arg-Gly-Phe-Phe- Tyr-Ala
I Cys-Asn
2: B 15-27 (Ala 27)-A 20-21
Leu-Tyr-Leu-Val-Cys-Gly I I
Tyr-Cys-Asn
3: B 15-20-A 1S-21
L eu-Tyr-Leu-Val- Cys-Gly-Glu-Arg-Gly- Phe-Phe- Tyr-Ala
I SEt
4: B 15-27 (SEt 19, Ala 27)
Leu-Tyr-Leu-Val-Cys-Gly-Glu-Arg-Gly- Phe- Phe-Tyr- Ala
Leu- Tyr-Leu-Val-Cys-Gly-Glu-Arg-Gly- Phe- Phe-Tyr-Ala
5: [B 15-27 (Ala 27)12
I
Boc-Leu-Tyr(Bzl)-Leu-Val-Cys(SEt)-Gly-Glu(OBzl)-Arg(H+)-
Gly- Phe- Phe- Tyr(Bz1)-Ala-OBzl
6
Boc-Leu-Tyr(Bz1)-Leu-Val-Cys(SEt)-Gly-OH 7
Die Darstellung von B 15 -27 (Ala 27) in der geschutzten Form 6 wurde bereits in einer vorausgegangenen Publikation von uns beschrieben "1. Ebenso wurde fruher der
Liebigs Ann. Chem. 1981
10'
146 G. Losse, H. Stange und W. Naumann
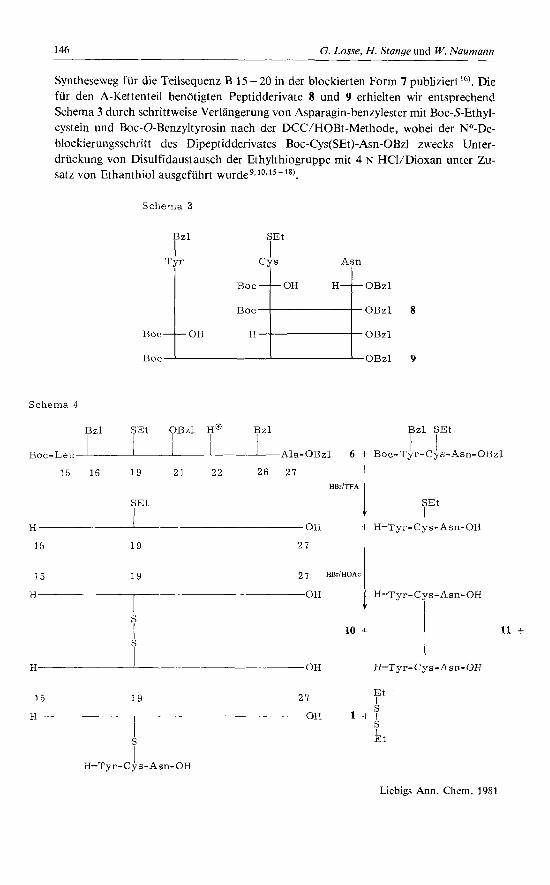
Syntheseweg fur die Teilsequenz B 15 - 20 in der blockierten Form 7 publiziertt6). Die fur den A-Kettenteil benotigten Peptidderivate 8 und 9 erhielten wir entsprechend Schema 3 durch schrittweise Verlangerung von Asparagin-benzylester mit Boc-S-Ethyl- cystein und Boc-0-Benzyltyrosin nach der DCCIHOBt-Methode, wobei der Na-De- blockierungsschritt des Dipeptidderivates Boc-Cys(SEt)-Asn-OBz1 zwecks Unter- druckung von Disulfidaustausch der Ethylthiogruppe mit 4 N HCVDioxan unter Zu- satz von Ethanthiol ausgefuhrt w ~ r d e ~ ~ ~ ~ ~ ' ~ - ~ * ) .
Schema 3 loH ; ; T O H H T O B z I
Boc OBzl
Boc OBzl 9
OBzl 8
Schema 4
p z l 4Et r z l ja f z l B z l SEt
Boc-Leu Ala-OBzl 6 + Boc-Tyr-Cys-Asn-OBzl I t
15 16 1 9 2 1 2 2 2 6 2 7 HBriTFA
SEt I
SEt I
H I on -k H-Tyr-(24s-Asn-OH
15
15
1 9
1 9
27
27 HBr/HOAc
,,doH /, H-Tyr-C
H I OH
15 1 9 2 1
10 +
H-Tyr- C
? s
1 + 1 s Et
s - A s n- OH
s- Asn- OH
11 +
7 H-Tyr-Cys-Asn-OH
Liebigs Ann. Chern. 1981
Synthese von insulinahnlichen Cystinpeptiden 147
Zur Gewinnung des Insulinausschnittes B 15 - 27 (Aia 27) - A 19 - 21 (Verbindung 1) wurden die geschutzten Derivate 6 und 9 mittels HBr/TFA im Gemisch deblockiert. Dabei werden alle Schutzgruppen entfernt, rnit Ausnahme der Ethylthiogruppe, die ohne besondere Vorsichtsmahahmen partiellem Disulfidaustausch unterliegt y,1y,20) (Schema 4).
Demgegenuber fanden wir fruher2'), da8 Deblockierungen rnit 6 N HBr/Eisessig in Gegenwart von Cys-SEt-Gruppen rnit weitgehendem Disulfidaustausch verbunden sind. Fur die eigentliche Deblockierung von 6 und 9 kam diese Methode nicht in Be- tracht, da einerseits die Benzyletherschutzgruppen am Tyrosin nicht vollstandig ent- fernt werdenZ2) und andererseits, im Vergleich zu HBr/TFA, verstarkt Nebenreaktio- nen durch freigesetzte Benzylkationen eintreten. Zur Uberfiihrung der Ethylthiofunk- tion von 6 in das asymmetrische Disulfid 1 schien uns dieses Verfahren aber brauchbar zu sein. Somit wurde das Gemisch der HBr/TFA-Deblockierung einer HBr/Eisessig- Nachbehandlung unterzogen. Um bei den entsprechend Schema 4 theoretisch zu erwar- tenden Disulfiden den Anteil des symmetrischen B-Ketten-Disulfides 10 zugunsten von 1 zuriickzudrangen, wurde das A-Kettenteilstuck 9 im 20fachen molaren UberschuB eingesetzt .
EZBO t
150 2 00 2 50 300 ml
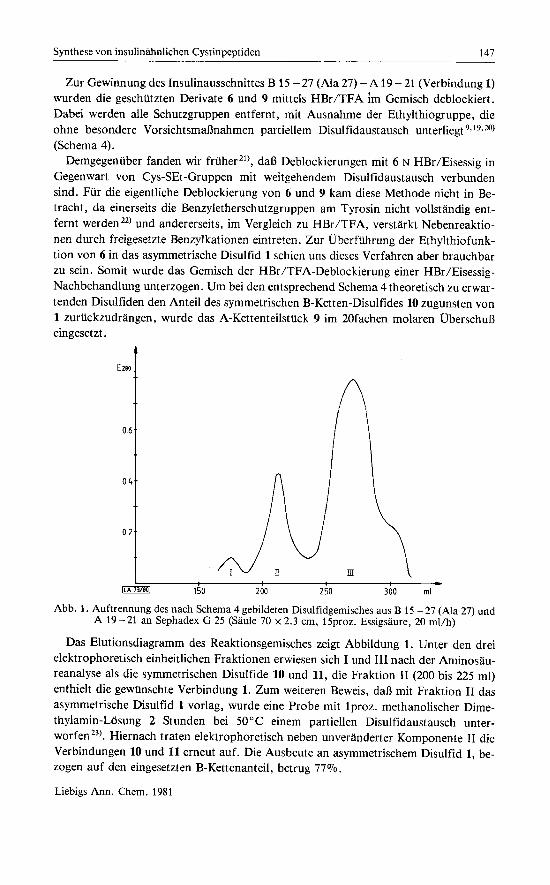
Abb. 1. Auftrennung des nach Schema 4 gebildeten Disulfidgemisches aus B 15 -27 (Ma 27) und A 19 - 21 an Sephadex G 25 (Saule 70 x 2.3 cm, 15proz. Essigsaure, 20 ml/h)
Das Elutionsdiagramm des Reaktionsgemisches zeigt Abbildung 1. Unter den drei elektrophoretisch einheitlichen Fraktionen erwiesen sich I und 111 nach der Aminosau- reanalyse als die symmetrischen Disulfide 10 und 11, die Fraktion I1 (200 bis 225 ml) enthielt die gewunschte Verbindung 1. Zum weiteren Beweis, daB rnit Fraktion I1 das asymmetrische Disulfid 1 vorlag, wurde eine Probe rnit Iproz. methanolischer Dime- thylamin-Losung 2 Stunden bei 50°C einem partiellen Disulfidaustausch unter- ~ o r f e n ~ ~ ) . Hiernach traten elektrophoretisch neben unveranderter Komponente I1 die Verbindungen 10 und 11 erneut auf. Die Ausbeute an asymmetrischem Disulfid 1, be- zogen auf den eingesetzten B-Kettenanteil, betrug 77%.
Liebigs Ann. Chem. 1981

148 G. Losse, H. Stange und W. Naumann
Entsprechend wurden die Verbindungen B 15 - 27 (Ala 27) - A 20 - 21 (2) und B 15 - 20 - A 29 - 21 (3) durch gemeinsame Vordeblockierung der Peptidderivate 6 und 8 bzw. 7 und 9 rnit HBr/TFA und anschliel3enden Disulfidaustausch ihrer Ethyl- thiofunktionen rnit HBrIEisessig bei jeweils uberschussiger A-Kettenkomponente mit 79- bzw. 8lproz. Ausbeute, bezogen auf den B-Kettenanteil, gewonnen.
Fur die Darstellung der vom A-Kettenteil freien Disulfide 4 und 5 bot sich, ausge- hend von der geschiitzten Sequenz 6, ein anderer Weg an. Wie wir fruher an mehreren Beispielen zeigen k ~ n n t e n ~ , ” ~ ~ ~ , ~ ~ ) , unterdriickt der Zusatz von iiberschiissigem Ethan- thiol den H+-katalysierten Disulfidaustausch von Ethylthiogruppen weitgehend, so dal3 bei der HBr/TFA-Acidolyse in Gegenwart des Thiols einheitliche Deblockierungspro- dukte rnit intakter Ethylthiofunktion erhalten werden 19920) . Diese Arbeitsweise ist dem- nach bevorzugt fur die Uberfiihrung von 6-4 geeignet. Andererseits liel3 die thiolfreie Anwendung von HBrIEisessig als Deblockierungsreagens fur 6 eine weitgehende Ein- stellung des Disulfidaustausch-Gleichgewichtes und damit hohe Anteile an symmetri- schem Disulfid 5 erwarten. Wegen der bekannten Komplikationen wahrend derartiger Abspaltungen, bedingt durch die ubrigen Schutzgruppen, wurde jedoch auf diese Va- riante verzichtet. A l s einen Mittelweg wahlten wir die HBr/TFA-Acidolyse ohne Thiol- zusatz, wo der Disulfidaustausch weder unterdriickt noch ausgelost wird. Auf diese Weise wurden aus der geschutzten Teilsequenz 6 gleichzeitig das asymmetrische Disul- fid 4 und das symmetrische Disulfid 5 im Verhaltnis 2.7: 1 (Ausbeute 67 bzw. 25%, be- zogen auf 6) erhalten. Beide konnten an Sephadex G 25 bequem getrennt, elektropho- retisch als einheitliche Verbindung erkannt und durch ihre Fahigkeit zur Disproportio- nierung in methanolischer Amin-Losung unterschieden werden.
Tab. 1. Biologische Aktivitat der Cystinpeptide; Yo-Angaben bezogen auf Insulin als Standard (Fehlergrenze f 10%)
Nr . Sequenzabschnitte Yo
1 B 15-27 (Ala 27)-A 19-21 0.30 2 B 15-27 (Ala 27)-A 20-21 0.09 3 B 15-20-A 19-21 0.01 4 B 15-27 (SEt 19, Ala 27) 0.09 5 [B 15-27 (Ala 27)12 0.04
Die Bestimmung der biologischen Aktivitat fur die Verbindungen 1 - 5 ergab im Mausekrampftest und Radioimmunassay die in Tabelle 1 angefuhrten Werte. Die Insu- linwirkung war durch Glucose aufhebbar und demzufolge nicht durch toxische Effekte bedingt.
Man erkennt daraus deutlich strukturabhangige Wirkungsunterschiede, die sowohl bei Verkurzung des A-Kettenpeptides beim Ubergang 1 +2 -4 als auch des B-Ketten- peptides (1 -3) hervortreten. Verglichen mit den von Weitzel”) an niederen Oligopepti- den des Sequenzbereiches B 22 - 26 aufgefundenen insulinahnlichen Effekten, weisen die untersuchten Verbindungen eine bis zu 3 Gronenordnungen hohere In-vivo-Insulin- wirkung auf.
Liebigs Ann. Chem. 1981

Synthese von insulinahnlichen Cystinpeptiden 149
MaBgebend fur den gegeniiber dem natiirlichen Hormon starken Aktivitatsabfall bei den dargestellten Verbindungen diirfte vor allem das Fehlen des Abschnittes A 1 - 7 aus der aktiven Insulinregion sein.
Der Pharmakologischen Abteilung des VEE Berlin-Chemie danken wir fur die Ausfiihrung der Aktivitatsbestimmungen.
Experimenteller Teil Die Schmelzpunkte sind korrigiert. Zur Dunnschichtchromatographie wurden Silufol-Platten
(Kavalier, CSSR) verwendet. Laufmittel A: n-Butanol/Eisessig/Wasser 4: 1 : 1, Laufmittel B: Aceton/Benzol 5 : 8. Die Papierelektrophoresen wurden auf FN-4-Papier (VEB Spezialpapierfa- brik Niederschlag) im Puffer Ameisensaure/Essigsaure/4 M Harnstoff 3: 2: 15 bei 45 V/cm in 45 min ausgefiihrt; Anfarbung rnit Pauly-Reagens. - Abkiirzungen: DCC = Dicyclohexylcarbo- diimid, HOBt = I-Hydroxybenzotriazol, TFA = Trifluoressigsaure.
Boc-Cys(SEt)-Asn-OEzl(8): 740 mg (2.8 mmol) H-Asn-OBzl . HCI werden unter Zugabe von 280 mg (2.8 mmol) Et3N unter Riihren in 15 ml CH2C12 bei - 10°C gelost, rnit 850 mg (3.0 mmol) Boc-Cys(SEt)-OH und 680 mg (3.2 mmol) DCC, letzteres in 5 ml CH2C12 gelost, versetzt. Danach wird 3 h bei - 10°C und 12 h bei 0°C geriihrt und iiber Nacht bei 20°C stehengelassen. Zur Auf- arbeitung saugt man vom ausgefallenen Niederschlag ab, engt i. Vak. bei 30°C ein, nimmt in Es- sigester auf, filtriert abermals, extrahiert dreimal rnit 5proz. KHS0,- sowie 5proz. NaHC0,- Losung und Wasser, trocknet rnit MgSO, und dampft nach Entfernung des Trockenmittels bei 15 Torr zur Trockne ein. Das so gewonnene dlige Produkt wird aus Essigester/n-Hexan kristallisiert. Ausb. 1.05 g (77070); Schmp. 99-103°C; RF = 0.85 (Laufmittel A), 0.51 (Laufmittel B).
C2,H,,N30,S2 (485.60) Ber. C 51.94 H 6.44 N 8.65 S 13.21 Gef. C 51.81 H 6.45 N8.60 S 13.01
Eoc-Tyr(Ez1)-Cys(SEt)-Asn-OBzl (9): 485 mg (1 .O mmol) Boc-Cys(SEt)-Asn-OBz1 (8) werden nach Zugabe von 1.2 g (20 mmol) C2H,SH rnit 30 ml 4 N HCl/Dioxan 30 min behandelt. An- schlieRend engt man i . Vak. zur Trockne ein, setzt CH2C12 hinzu, destilliert das Losungsmittel bei 10 Torr und 30°C wieder ab und wiederholt diese Operation mehrfach. Der Ruckstand wird 2 h bei 0.1 Torr uber KOH getrocknet; R, = 0.22 (Laufmittel A). - Das so erhaltene Nu- deblockierte Dipeptidderivat wird unter Zugabe von 101 mg (1 .O mmol) Et3N in 10 ml CH2C12 ge- lost und bei - 10°C unter Ruhren rnit 420 mg (1.1 mmol) Boc-Tyr(Bz1)-OH, 270 mg (2.0 mmol) HOBt sowie 229 mg (1.1 mmol) DCC, letzteres in 2 ml CH2C12 gelost, versetzt. Der Ansatz wird 3 h bei - 10°C sowie 15 h bei 0°C geruhrt, iiber Nacht bei 20°C stehengelassen und wie fur Ver- bindung 8 beschrieben aufgearbeitet. Ausb. 0.65 g (88%); Schmp. 148 - 150'C; RF = 0.91 (Lauf- mittel A), 0.71 (Laufmittel B).
C3,H,,N,08S2 (739.90) Ber. C 60.06 H 6.40 N 7.57 S 8.67 Gef. C 60.01 H 6.45 N 7.41 S 8.52
Aminosaureanalyse: Tyr 1.0 (I), Cys 0.85 (I), Asp 1.0 (1).
B 15 -27 (Ala 27) - A 19-21 (1): 145 mg (0.2 mmol) Boc-Tyr(Bz1)-Cys(SEt)-Asn-OBzl (9) und 21 mg (0.01 mmol) Boc-Leu-Tyr(Bzl)-Leu-Val-Cys(SEt)-Gly-Glu(OBzl)-~g(HCl)-Gly-Phe-Phe- Tyr(Bz1)-Ala-OBzl (6)15) werden in 20 ml TFA und 2 ml Anisol geldst und durch Einleiten eines trockenen, bromfreien HBr-Stromes 1.5 h bei 20°C deblockiert. AnschlieDend engt man i. Vak. bei 25 "C auf ca. 2 ml ein, fallt das Produktgemisch durch Zugabe von absol. Ether aus, zentrifu- giert den Niederschlag ab, wlscht mehrfach rnit absol. Ether, trocknet 2 h bei 0.1 Torr iiber KOH
Liebigs Ann. Chem. 1981

150 G. Losse, H. Stanae und W. Naumann
und behandelt 30 min rnit 6 N HBrIHOAc. AnschlieRend engt man erneut i. Vak. ein und trock- net 3 h bei 30°C uber KOH. Zur Auftrennung der Disulfide versetzt man das Gemisch mit 1 ml konz. Na2C03-Losung, lost portionsweise in insgesamt 10 bis 15 ml30proz. Essigsaure, zentrifugiert Unlosliches ab und chro- matographiert entsprechend Abbildung 1 an Sephadex G 25. Der Fraktionsbereich 200 bis 225 ml wird auf 5 ml eingeengt und lyophilisiert. Ausb. 17.5 mg (77%) elektrophoretisch einheitliche Substanz. Aminosaureanalyse: Leu 1.9 (2), Tyr 2.9 (3), Val 1.1 (l), Cys 1.7 (2), Gly 2.0 (2), Glu 1.0 (I), A r g 1.1 (I), Phe 1.9 (2), Ala 1.1 (l), Asp. 1.0 (1).
B 15 -27 (Ala 27) - A 20-21 (2): Die Darstellung erfolgt wie fur die Verbindung 1 beschrieben aus 97 mg (0.2 mmol) Boc-Cys(SEt)-Asn-OBz1 (8) und 21 mg (0.01 mmol) Boc-Leu-Tyr(Bz1)-Leu- Val-Cys(SEt)-Gly-Glu(OBzl)-Arg(HC1)-Gly-Phe-Phe-Tyr(Bzl)-Ala-OBzl (6), ebenso die Aufar- beitung und Sephadex-G-25-Chromatographie, wobei das gewunschte Produkt im Elutionsbe- reich von 160 bis 200 ml erhalten wird. Ausb. 15.3 mg (79%) elektrophoretisch einheitliche Sub- stanz.
Aminosaureanalyse: Leu 1.9 (2), Tyr 1.8 (2), Val 1.1 ( I ) , Cys 1.6 (Z), Gly 2.0 (2), Glu 0.9 (l), Arg 1.0 (l), Phe 2.0 (2), Ala 0.9 (I), Asp 1.0 (1).
B I5 -20-A 19-21 (3): Zur Darstellung wird von 145 mg (0.2 mmol) Boc-Tyr(Bz1)-Cys(SEt)- Asn-OBzl(9) und 19 mg (0.02 mmol) Boc-Leu-Tyr(Bz1)-Leu-Val-Cys(SEt)-Gly-OH (7) ausgegan- gen und wie fur 1 und 2 beschrieben weiterverfahren. Das gewunschte Produkt tritt im Elutions- bereich von 190 bis 230 ml der Sephadex-G-25-Saule aus. Ausb. 17.2 mg (81 %) elektrophoretisch einheitliche Substanz. Aminosaureanalyse: Leu 2.0 (2), Tyr 1.8 (2), Val 1.1 (I), Cys 1.6 (2), Gly 1.0 (l), Asp 1.1 (1).
B 15-27 (SEt 19, Ala 27) (4) und [B 15-27 (Ala 27)J2 (5): 31.5 mg (0.015 mmol) Boc-Leu-Tyr(Bzl)-Leu-Val-Cys(SEt)-Gly-Glu(OBzl)-Arg( HC1)-Gly-Phe-Phe-Tyr(Bz1)-Ala-OBzl (6) werden in 20 ml TFA unter Zusatz von 2 ml Anisol gelost und 2 h ein trockener, bromfreier HBr-Strom eingeleitet. AnschlieRend engt man i. Vak. bei 25 "C auf ca. 2 ml ein, fallt das Roh- produkt mit 30 rnl absol. Ether aus, zentrifugiert den Niederschlag ab, wascht ihn mehrmals mit absol. Ether und trocknet 2 h bei 0.1 Torr uber KOH. Das Disulfidgemisch wird dann rnit 1 ml konz. Na2CO3-Losung versetzt, durch Zugabe von 5 ml 30proz. Essigsaure gelost und unter den in Abbildung 1 gegebenen Bedingungen an Sephadex G 25 chromatographiert. Verbindung 4: Elutionsbereich 180 bis 220 ml. Ausb. 16 mg (6770) elektrophoretisch einheitliche Substanz. Aminosaureanalyse: Leu 2.2 (2), Tyr 1.8 (2), Val 1.1 (I), Cys 0.8 (11, Gly 2.1 (2), Glu 0.9 (l), Arg 1.1 (l), Phe 2.0 (2), Ala 1.0 (1).
Nach 2stdg. Behandlung rnit Iproz. methanolischer Dimethylamin-Losung bei 50 "C laRt sich elektrophoretisch B [15 - 27 (Ala 27)12 (Verbindung 5 ) nachweisen. Verbindung 5: Elutionsbereich 135 bis 170 ml. Ausb. 6 mg (25%); Elektrophoretisch einheitlich und identisch rnit dem durch methanolische Amin-Losung aus Verbindung 4 erhaltenen Dispro- portionierungsprodukt .
Liebigs Ann. Chem. 1981

Svnthese von insulinahnlichen Cvstinueutiden 151
R. Geiger, Chem.-Ztg. 100, 111 (1976). 2, M. 0. Dayhoff, Atlas o f Protein Sequences and Structure, Nat. Biomed. Red. Foundation,
Washington 1972. 3, R. E, Humbel, H. R. Bosshard und H. Zahn, Handbook o f Physiology, Bd. 1 (1972), S. 11,
Washington D.C. Section 7. 4) T. L . Blundell, G. G. Dodson, D. Hodgkin und D. A. Mercola, Adv. Prot. Chem. 26, 279
(1 972). 5 ) T. L. Blundell, G. G. Dodson, E. Dodson, D. Hodgkin und M. Vijavan, Rec. Progr. Hormo-
ne Res. 27, 1 (1971). T. L. Blundell, J. F. Cutfield, S. M. Cutfield, E. Dodson, G. G. Dodson, D. Hodgkin und D. A. Mercola, Diabetes 21, 492 (1972). R. A. Pullen, D. G. Lindsay, S. P. Wood, I. J. Tickle, T. L . Blundell, A. Wollmer, G. Krail, D. Brandenburg, H. Zahn, J. Gliemann und S. Gammeltoff, Nature (London) 259, 369 (1976).
8, H. Stange, Dissertation, Techn. Univ. Dresden 1978. 9, G. Losse, H. Stange, B. Schwenzer und W. Naumann, Tetrahedron 34, 2687 (1978).
lo) G. Losse, Z . Chem. 18, 330 (1978). 11) G. Weitzel, R. Renner und H . Guglielmie, Hoppe-Seyler’s Z . Physiol. Chem. 352, 1617
(1971); G. Weitzel, K. Eisele, V. Schulz und W. Stock, ebenda 354, 321 (1973); G. Weitzel, K. Eisele und W. Stock, ebenda 356, 585 (1975).
12) U. Weber und G. Weitzel, Hoppe Seyler’s Z . Physiol. Chem. 349, 1431 (1968); G. Weitzel, K. Eisele, H. Zollner und U. Weber, ebenda 350, 1480 (1969); G. Weitzel, U. Weber, K. Eise- le, H, Zollner und J. Martin, ebenda 351, 263 (1970); G. Weitzel, F.-U. Bauer und K. Eisele, ebenda 351, 187 (1976); G. Weitzel, F.-U. Bauer und A. Rehe, ebenda 358, 1573 (1977).
l3) P. C. Katsoyannis, J. Ginos, A. Cosmatos und G. Schwartz, J. Am. Chem. SOC. 95, 6427 (1973).
14) R. Gattner, Hoppe-Seyler’s Z. Physiol. Chem. 356, 1397 (1975). G. Losse und H. Stange, J. Prakt. Chem. 321, 308 (1979).
j6) G. Losse, K.-J. Schumacher und H. Stange, Tetrahedron 33, 1817 (1977). 17) G. Losse, M. Mauck und H. Stange, Tetrahedron 33, 1813 (1977). l8) G. Losse, B. Meisegeier, B. Schwenzer, H. Stange und K.-J. Schumacher, Z. Chem. 17, 263
19) G. Losse und W. Naumann, J. Prakt. Chem., 320, 651 (1978). 20) G. Losse, H. Stange, B. Schwenzer, W. Naumann, M. Mauck, K.-J. Schumacher und B. Mei-
segeier, Proc. 15th Europ-Peptide Symp., Sept. 1978, Gdansk, Poland, Edit. Z. Siemion und G. Kupryezewski.
21) G. Berndsen, Dissertation, Techn. Univ. Dresden 1969. 22) E. Schnabel, Liebigs Ann. Chem. 622, 181 (1959). 23) B. Kamber, Helv. Chim. Acta 54, 398 (1971).
(1977).
[73/80]
Liebigs Ann. Chem. 1981

![Proteinstrukturen: Faltung, Vergleich und Vorhersage · Antiparallele Bindung: [Hbond(j,i), Hbond(i,j)], [Hbond(j+2,i+2), Hbond(i+2,j+2)],.. Von der Primärstruktur zur gefalteten](https://img.pdfslide.org/doc/110x75/5ca5b9bc88c993f9768d059a/proteinstrukturen-faltung-vergleich-und-vorhersage-antiparallele-bindung.jpg)



























