Embed Size (px)
Citation preview

Theorie der Proteinfaltung
Strukturelle Bioinformatik WS15/16
Dr. Stefan Simm, 09.12.2015 [email protected]

EINLEITUNG Proteinfaltung
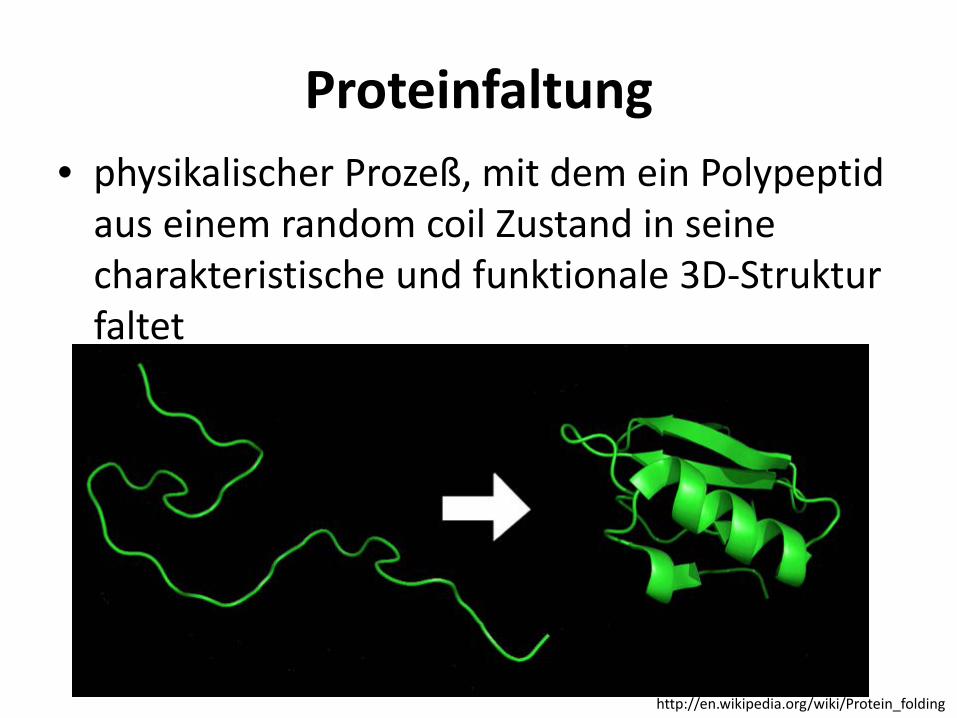
Proteinfaltung • physikalischer Prozeß, mit dem ein Polypeptid
aus einem random coil Zustand in seine charakteristische und funktionale 3D-Struktur faltet
http://en.wikipedia.org/wiki/Protein_folding

Proteinfaltung • physikalischer Prozeß mit dem ein Polypeptid
aus einem random coil Zustand in seine charakteristische und funktionale 3D-Struktur faltet
• Aminosäuren interagieren miteinander, um eine geordnete Tertiärstruktur auszubilden – native state
• die Tertiärstruktur ist von der Aminosäure-sequenz abhängig (Anfinsens Dogma)
http://en.wikipedia.org/wiki/Protein_folding

Anfinsens Dogma
• auch bekannt als thermodynamische Hypothese • bei kleinen, globulären Proteinen wird die Tertiär-
struktur ausschließlich von der Aminosäure-sequenz bestimmt
• unter den Bedingungen (Temperatur, Zusammen-setzung des Lösungsmittels, usw.), die eine Faltung ermöglichen, ist die native Struktur ein einzigartiges, stabiles und kinetisch zugängliches Minimum der freien Energie
http://en.wikipedia.org/wiki/Protein_folding

Levinthals Paradox
• würde ein Protein falten, indem es sequentiell alle möglichen Konformationen durchläuft, würde es astronomisch lange dauern, auch wenn jede Konformation sehr schnell, d.h. im Nano- bzw. Pikosekundenbereich eingenommen würde
10143 Möglichkeiten für ein 150aa Protein
http://www-miller.ch.cam.ac.uk/levinthal/levinthal.html
1080 Atome im sichtbaren Universum

Levinthals Paradox
• da Proteinfaltung sehr viel schneller erfolgt, ist die Suche im Konformationsraum nicht zufällig
• ein Protein faltet in einer Serie von meta-stabilen, intermediären Zuständen
http://www-miller.ch.cam.ac.uk/levinthal/levinthal.html

Proteinfaltung
• gefaltete Proteine besitzen einen hydrophoben Kern
• Minimierung der Anzahl hydrophober Seitenketten, die zum Wasser exponiert sind, ist eine wichtige Triebkraft des Prozesses der Proteinfaltung – hydrophobic collapse
• Stabilisierung des Kerns durch dichte Packung von Seitenketten
http://en.wikipedia.org/wiki/Protein_folding

Proteinfaltung
• intramolekulare H-Brücken tragen ebenfalls zur Stabilisierung der Struktur bei
• Stärke einer H-Brücke abhängig von ihrer Umgebung Proteinkern > Proteinoberfläche
http://en.wikipedia.org/wiki/Protein_folding

Proteinfaltung
• in vivo beginnt Proteinfaltung während der Translation an Ribosomen
• d.h. während der N-terminale Abschnitt schon faltet, wird der C-terminale Abschnitt noch synthetisiert
• Chaperone – verhindern Aggregation von Proteinen – von bestimmten Proteinen benötigt, um in ihre
funktionale Struktur zu falten
http://en.wikipedia.org/wiki/Protein_folding
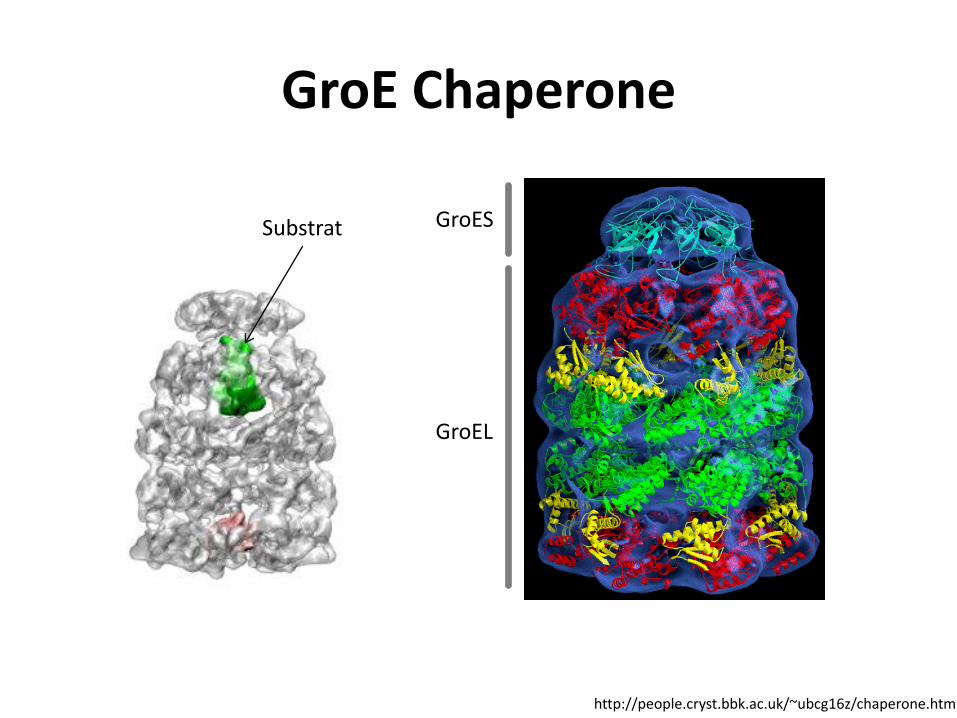
GroE Chaperone
GroES
GroEL
Substrat
http://people.cryst.bbk.ac.uk/~ubcg16z/chaperone.html
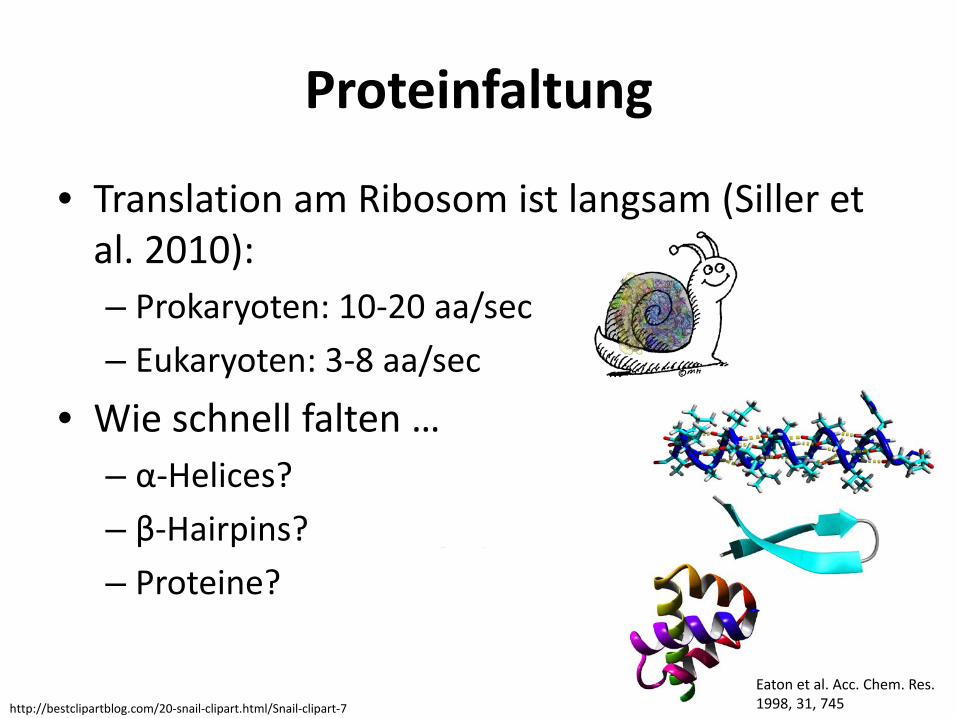
Proteinfaltung
• Translation am Ribosom ist langsam (Siller et al. 2010): – Prokaryoten: 10-20 aa/sec – Eukaryoten: 3-8 aa/sec
• Wie schnell falten … – α-Helices? einige 100 ns (<1µs) – β-Hairpins? wenige µs – Proteine? wenige µs bis ms
Eaton et al. Acc. Chem. Res. 1998, 31, 745 http://bestclipartblog.com/20-snail-clipart.html/Snail-clipart-7
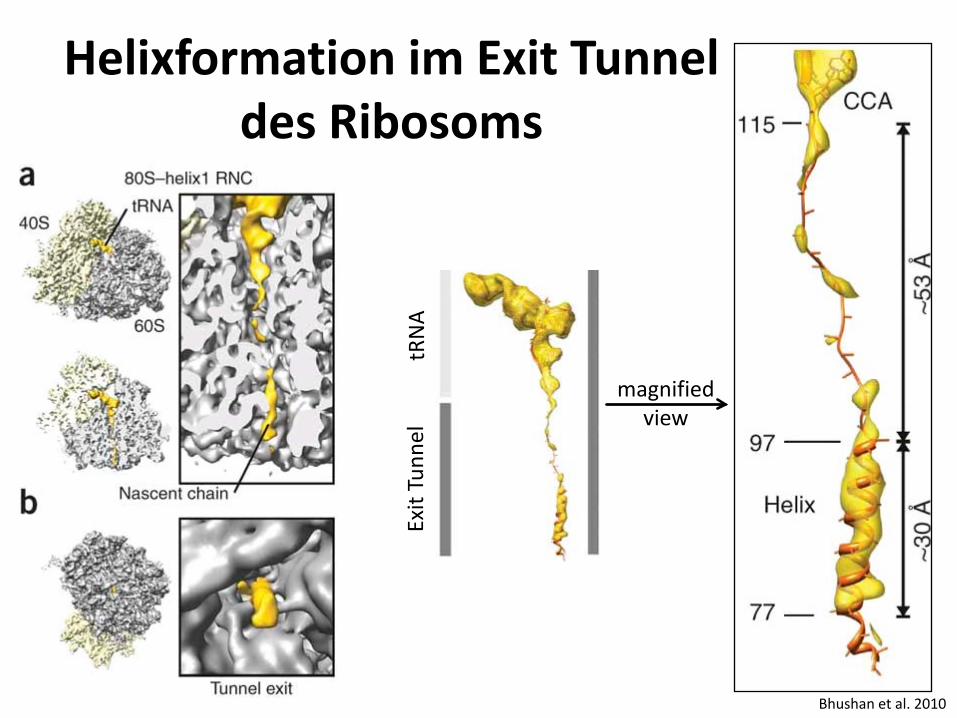
Helixformation im Exit Tunnel des Ribosoms
Bhushan et al. 2010
tRN
A Ex
it Tu
nnel
magnified
view

Theoretische Betrachtungen der Proteinfaltung
• Methoden: – experimentelle – theoretische – computergestützte
• “einfachstes” Problem: – Wie faltet ein globuläres Protein mit einer einzigen
Domäne? – D.h. wie navigiert es durch die rauhe Landschaft der
freien Energie auf dem Weg zu seiner nativen Struktur?
Thirumalai et al. 2010

Theoretische Betrachtungen der Proteinfaltung
• Faltungsmechanismen vielfach mittels einer Approximation auf zwei Zustände beschrieben – nur gefaltete und ungefaltete Zustände als
signifikant häufig eingenommen betrachtet
• aber: – Proteine sind verzweigte Polymere von endlicher
Länge – native Struktur nur marginal stabilisiert durch eine
Anzahl von relativ schwachen Interaktionen
Thirumalai et al. 2010

Theoretische Betrachtungen der Proteinfaltung
• ungefalteter und sogar der gefaltete Zustand sollten – mikroskopisch betrachtet – daher als ein Ensemble von Strukturen angesehen werden
• unter Faltungsbedingungen sind die Fluktuationen im nativen Zustand geringer als im ungefalteten Zustand
Thirumalai et al. 2010
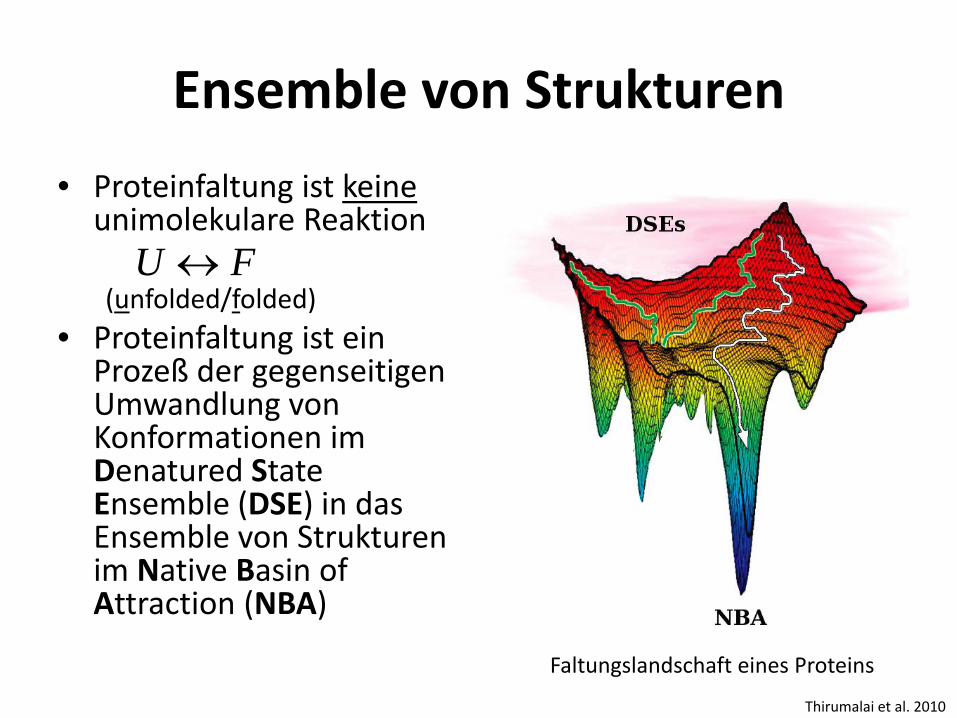
Ensemble von Strukturen • Proteinfaltung ist keine
unimolekulare Reaktion (unfolded/folded)
• Proteinfaltung ist ein Prozeß der gegenseitigen Umwandlung von Konformationen im Denatured State Ensemble (DSE) in das Ensemble von Strukturen im Native Basin of Attraction (NBA)
FU ↔
Thirumalai et al. 2010
Faltungslandschaft eines Proteins

Ensemble von Strukturen
• Proteinfaltung mittels Verteilungsfunktionen beschreibbar
• folgendes wird benötigt, um die Selbstorganisation von Proteinen zu erfassen: – passende “Werkzeuge” aus der statistischen
Mechanik – Konzepte der Polymerphysik
• RNA-Faltung kann ebenfalls auf diese Weise betrachtet werden
Thirumalai et al. 2010

Gyrationsradius
• in der Polymerphysik beschreibt er die Abmessungen einer Polymerkette
• da ein Ensemble von Strukturen vorliegt, ist der Rg der Mittelwert des Ensembles (ensemble average)
( )∑=
−≡N
kmeankg rr
NR
1
22 1
( )∑=
−≡N
kmeankg rr
NR
1
22 1
N = Sequenzlänge rk = Position von Residue k rmean = Schwerpunkt aller Residuen
http://en.wikipedia.org/wiki/Radius_of_gyration

Gyrationsradius
• stark denaturierende Bedingungen Proteine liegen entfaltet als random coil vor
• wenn dem so ist, dann muß – basierend auf der Flory Theorie - der Gyrationsradius (Rg) im ungefalteten Zustand mit skalieren – ad = charakteristische Kuhn Länge – d = denatured state – N = Anzahl Aminosäuren im Protein – ν = 0,6
• gilt für Homopolymere in guten Lösungsmitteln
νNaR ddg ≈
Thirumalai et al. 2010

Gyrationsradius
• gefaltete Proteine sind maximal kompakt • daher skaliert der native state mit
– mit und n=native state
νNaR nng ≈
31 =ν
RG Gyrationsradius ρ Anteil nativer Kontakte Farbverlauf zeigt die Änderung der freien Energie grün=energetisch am ungünstigsten blauer Bereich=native state
Brooks et al. (1998)
Thirumalai et al. 2010

Spingläser und Frustration • Ausgangspunkt ist das „Random
Energy Model“ (REM) aus Theorie der Spingläser
• Spingläser sind ungeordnete, verdünnte Anordnungen magnetischer Atome in diamagnetischen Matrizen
• benachbarte Atome koppeln entweder ferro- oder antiferromagnetisch – ferromagnetisch bevorzugt gleich
orientierte Spins, – antiferromagnetisch hingegen
entgegensetzte Spins – sobald eine Kopplung zwischen zwei
Spins etabliert ist, behält sie ihren Wert bei
• beim Abkühlen einer Spinglasprobe richten sich die Spins so aus, dass insgesamt eine Anordnung mit minimaler Energie entsteht
Weikl 2009
Ferromagnetismus
Antiferromagnetismus

Spingläser und Frustration
• Konflikte: System mit ungerader Zahl antiferromagnetischer Wechselwirkungen – zwei Spins können sich
energetisch günstig einstellen
– dritter kann nur zu jeweils einem der beiden benachbarten energetisch günstig stehen
Antiferromagnetisch wechselwirkende Spins in dreieckiger Anordnung
siehe auch http://www.physics.emory.edu/~weeks/lab/tet/tetrahedra_extra.html http://www.informatik.uni-koeln.de/old-ls_juenger/projects/spinglass.html http://de.wikipedia.org/wiki/Geometrische_Frustration
?

Spingläser und Frustration
• unmöglich, alle lokalen Wechselwirkungen simultan zu optimieren „Frustration“
• Frustration führt im gesamten Spinsystem zu einer Quasi-Entartung des Grundzustandes – viele Konfigurationen mit ähnlicher Energie – durch Abkühlung eingefrorener Mikrozustand hängt
von Vorgeschichte ab – jeder Abkühlzyklus führt zu anderer Spinkonfiguration – außerdem existiert ein Glasübergang:
• Anzahl thermisch zugänglicher Konfigurationen sinkt mit der Temperatur dramatisch ab
• Spinsystem friert unterhalb der Glastemperatur in einer zufälligen Konfiguration ein
Weikl 2009
?

Spingläser und Frustration
• Tendenz der Spins im Spinglas, sich in Bezug auf die Nachbarspins optimal ausrichten zu wollen, entspricht dem Bestreben der Aminosäuren im Protein, sich im kompakten, gefalteten Zustand energetisch optimal zu ihren Nachbarn anzuordnen
• weiterführende Literatur: Binder, Kurt, and A. Peter Young. "Spin glasses: Experimental facts, theoretical concepts, and open questions." Reviews of Modern physics 58.4 (1986): 801.
Weikl 2009
?

folding landscape • Visualisierung des Faltungs-
prozesses als ein Navigieren in einer großen Faltungslandschaft
• Dynamik von Heteropolymeren zufälliger Sequenz zeigt, daß ihre Energielandschaft viel zu rauh ist, um sie in typischen Faltungszeiten (Millisekunden) zu erkunden
• energetische Frustration
Thirumalai et al. 2010

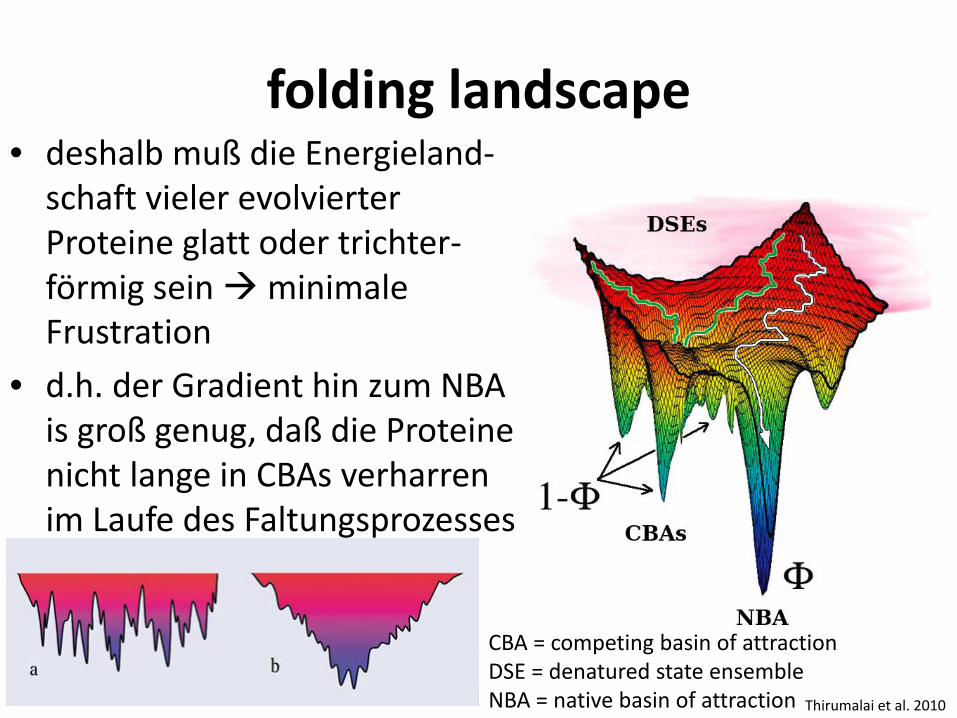
folding landscape • deshalb muß die Energieland-
schaft vieler evolvierter Proteine glatt oder trichter-förmig sein minimale Frustration
• d.h. der Gradient hin zum NBA is groß genug, daß die Proteine nicht lange in CBAs verharren im Laufe des Faltungsprozesses
Thirumalai et al. 2010
CBA = competing basin of attraction DSE = denatured state ensemble NBA = native basin of attraction
folding landscape • deshalb muß die Energieland-
schaft vieler evolvierter Proteine glatt oder trichter-förmig sein minimale Frustration
• d.h. der Gradient hin zum NBA is groß genug, daß die Proteine nicht lange in CBAs verharren im Laufe des Faltungsprozesses
Thirumalai et al. 2010
CBA = competing basin of attraction DSE = denatured state ensemble NBA = native basin of attraction
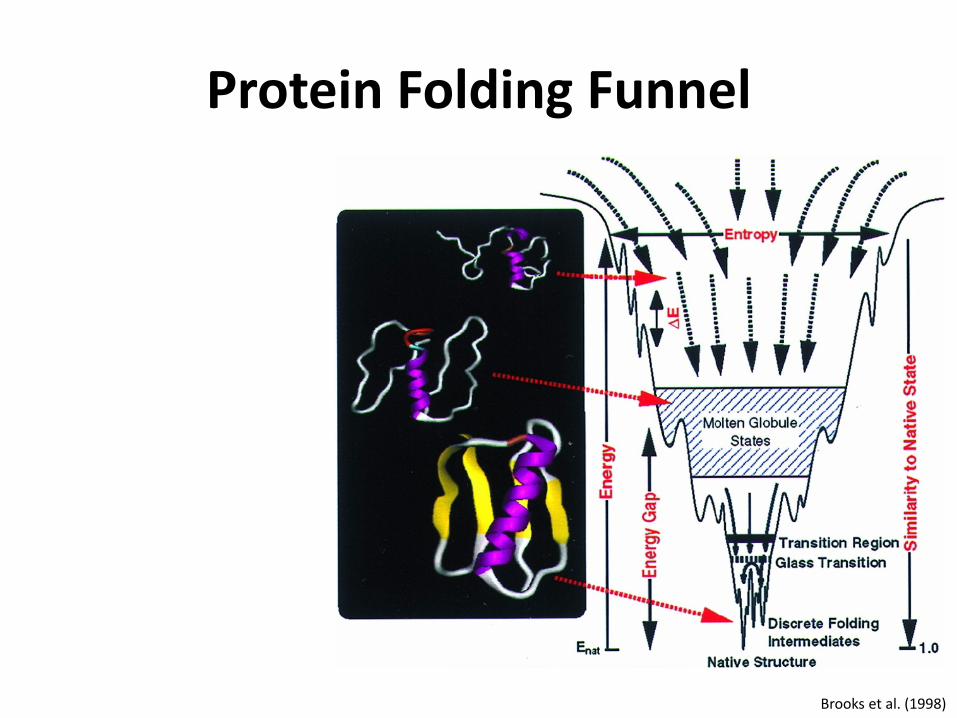
Protein Folding Funnel
Brooks et al. (1998)
Brooks et al. (1998)
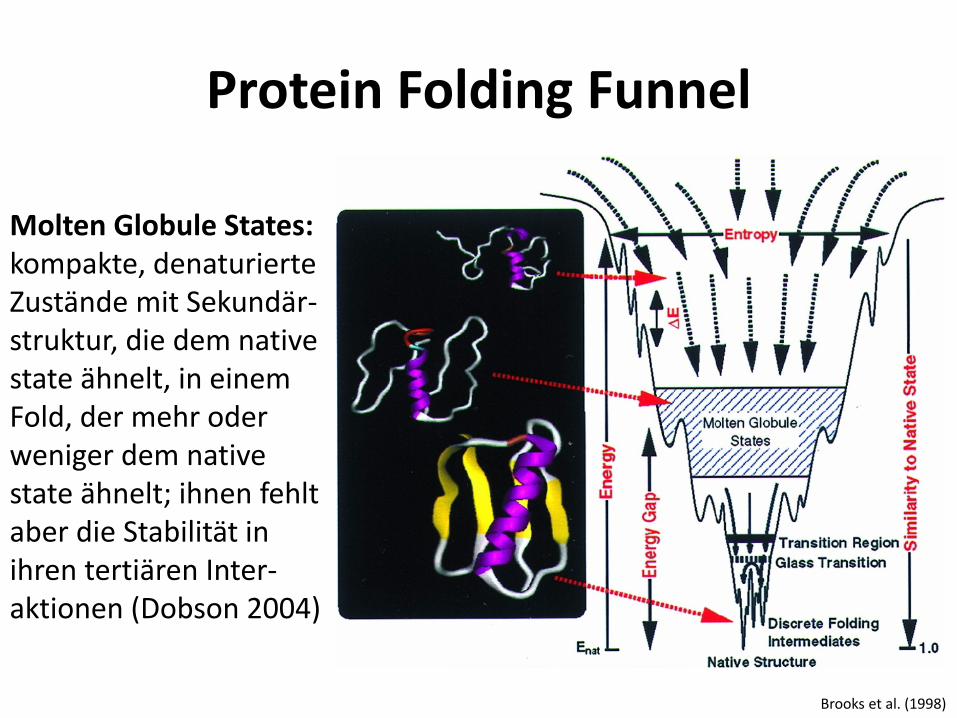
Molten Globule States: kompakte, denaturierte Zustände mit Sekundär-struktur, die dem native state ähnelt, in einem Fold, der mehr oder weniger dem native state ähnelt; ihnen fehlt aber die Stabilität in ihren tertiären Inter-aktionen (Dobson 2004)
Protein Folding Funnel
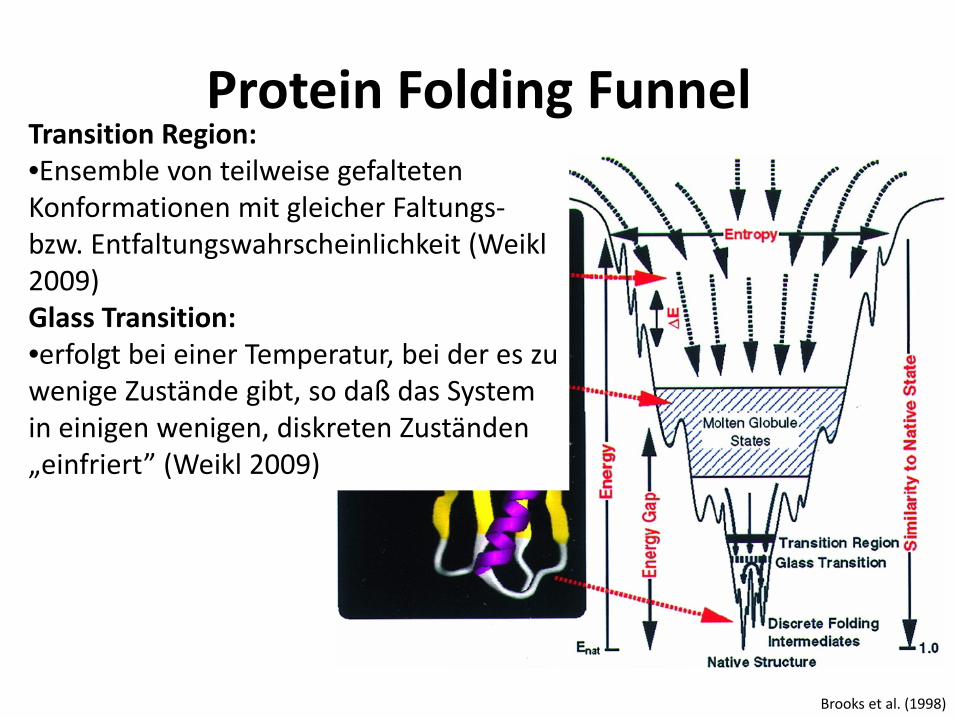
Brooks et al. (1998)
Transition Region: •Ensemble von teilweise gefalteten Konformationen mit gleicher Faltungs- bzw. Entfaltungswahrscheinlichkeit (Weikl 2009) Glass Transition: •erfolgt bei einer Temperatur, bei der es zu wenige Zustände gibt, so daß das System in einigen wenigen, diskreten Zuständen „einfriert” (Weikl 2009)
Protein Folding Funnel
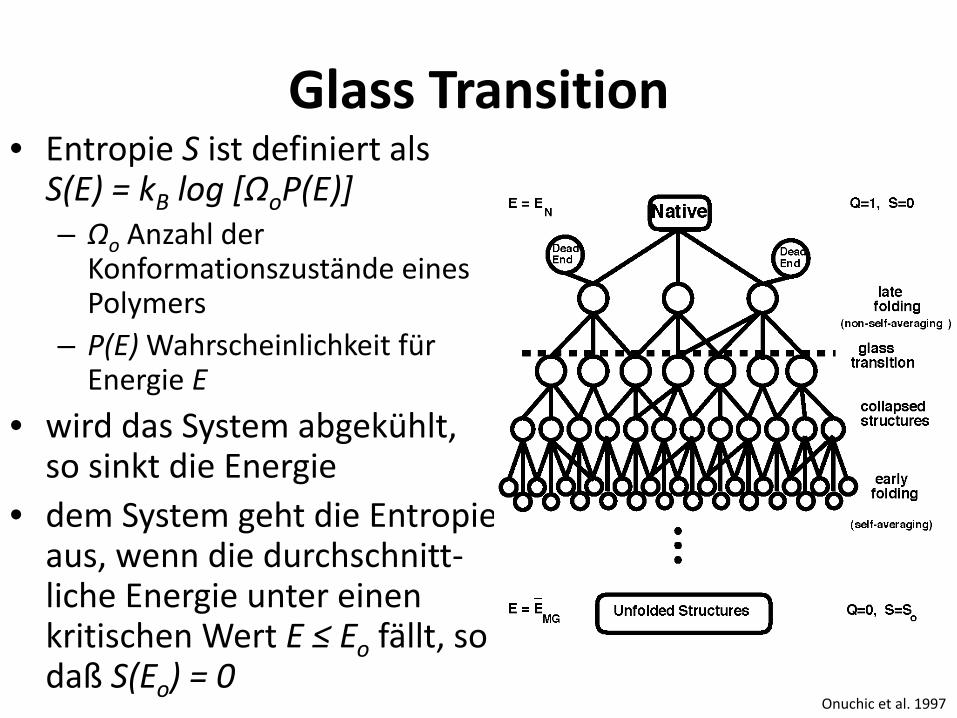
Glass Transition • Entropie S ist definiert als
S(E) = kB log [ΩoP(E)] – Ωo Anzahl der
Konformationszustände eines Polymers
– P(E) Wahrscheinlichkeit für Energie E
• wird das System abgekühlt, so sinkt die Energie
• dem System geht die Entropie aus, wenn die durchschnitt-liche Energie unter einen kritischen Wert E ≤ Eo fällt, so daß S(Eo) = 0
Onuchic et al. 1997
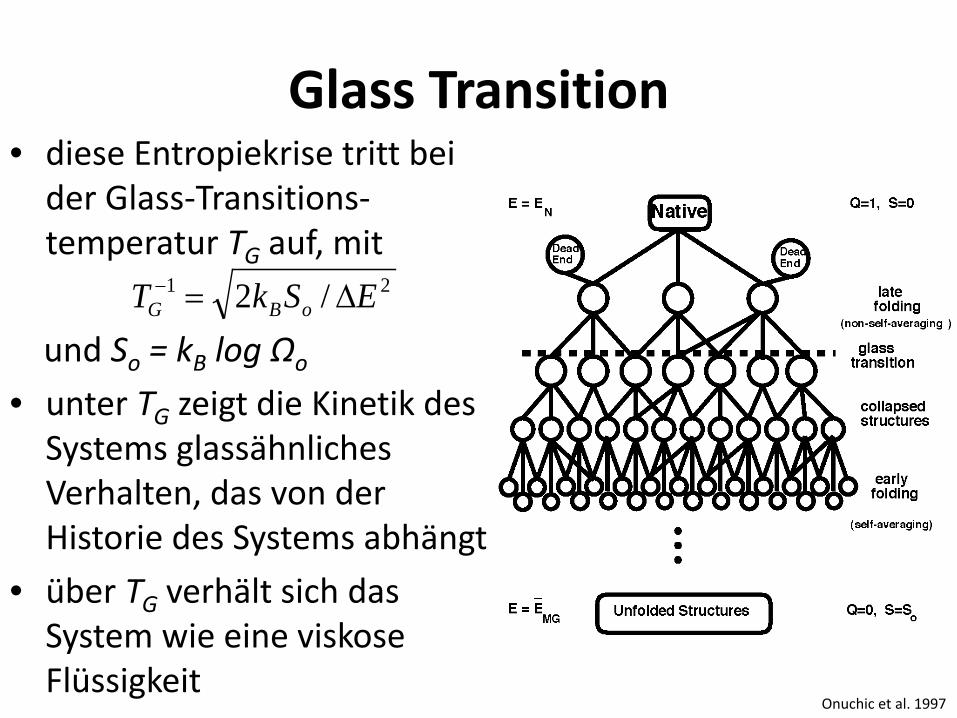
Glass Transition • diese Entropiekrise tritt bei
der Glass-Transitions-temperatur TG auf, mit
und So = kB log Ωo • unter TG zeigt die Kinetik des
Systems glassähnliches Verhalten, das von der Historie des Systems abhängt
• über TG verhält sich das System wie eine viskose Flüssigkeit
21 /2 ESkT oBG ∆=−
Onuchic et al. 1997
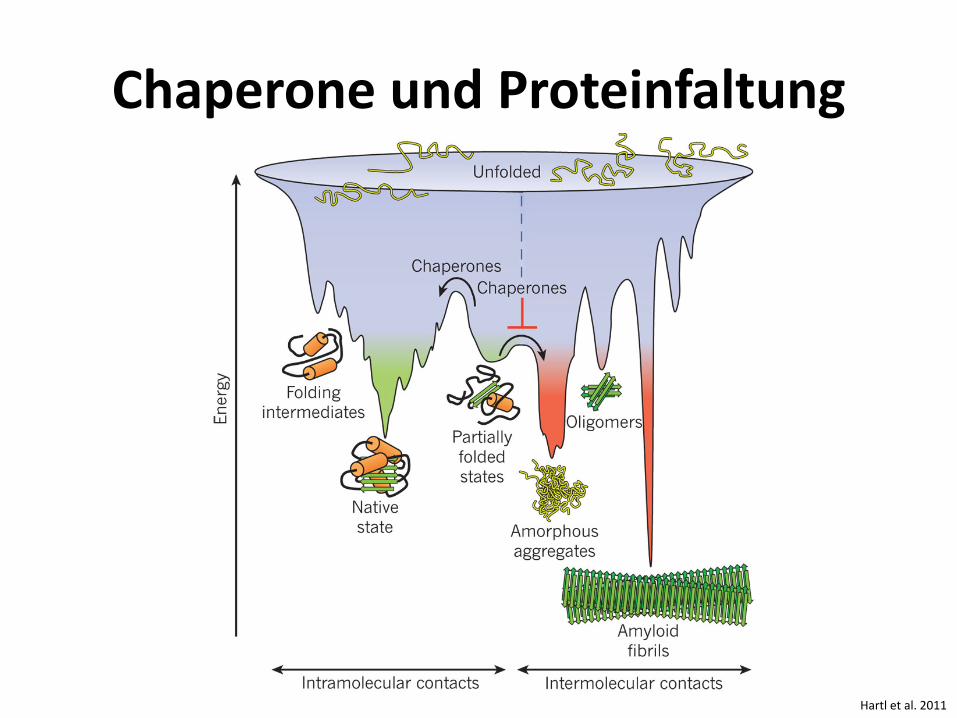
Chaperone und Proteinfaltung
Hartl et al. 2011
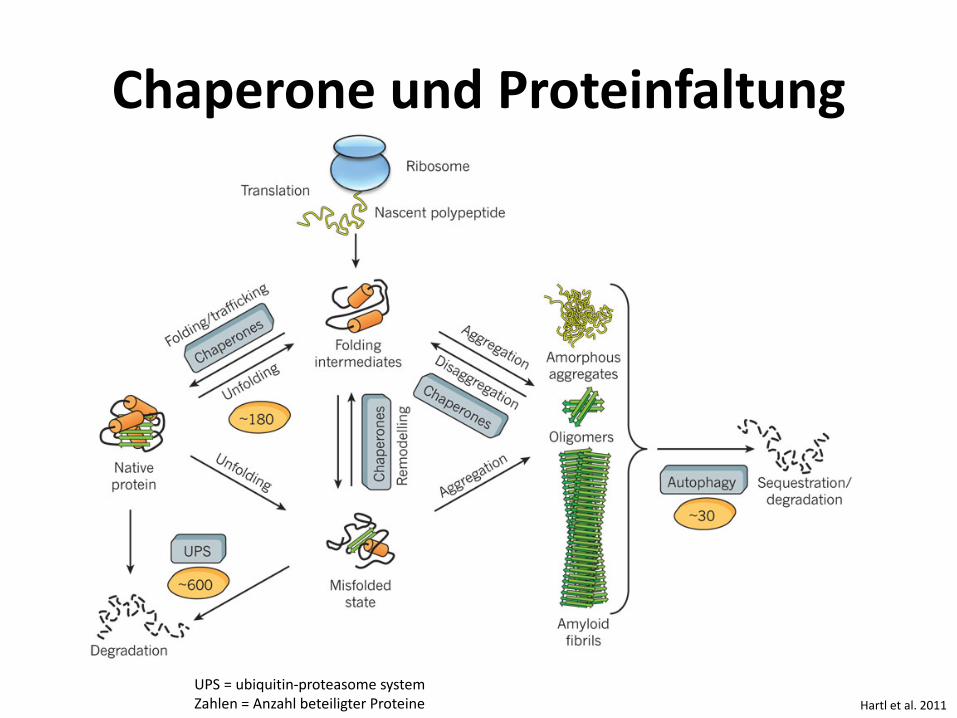
Chaperone und Proteinfaltung
Hartl et al. 2011 UPS = ubiquitin-proteasome system Zahlen = Anzahl beteiligter Proteine

Multiple Folding Nuclei (MFN) Model
• effiziente Faltung von Proteinen erfolgt nach einem NC Mechanismus mit einem raten-limitierenden Schritt, der einhergeht mit der Ausbildung eines der folding nuclei
• Ausbildung des folding nucleus und der collapse der Kette erfolgen nahezu gleichzeitig – NC Mechanismus
• durchschnittliche Größe des wahrscheinlichsten nucleus N*
R für globuläre Proteine mit einer einzigen Domäne beträgt etwa 15-30 Residuen (Chen et al. 2008, J Phys Chem B)
Thirumalai et al. 2010
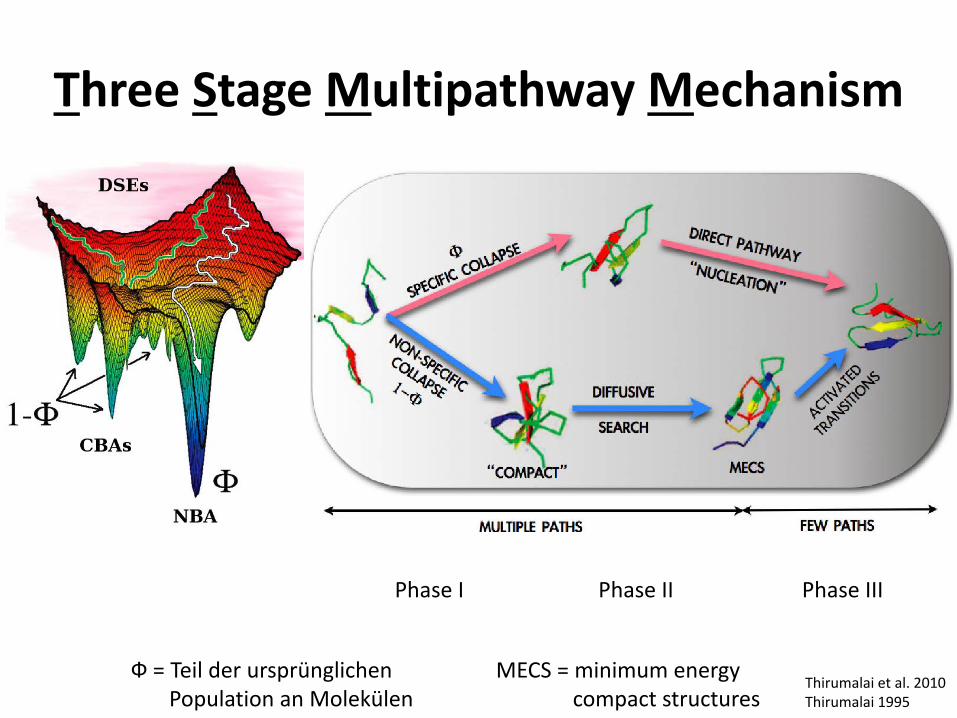
Three Stage Multipathway Mechanism
Thirumalai et al. 2010 Thirumalai 1995
Phase I Phase II Phase III
MECS = minimum energy compact structures
Ф = Teil der ursprünglichen Population an Molekülen
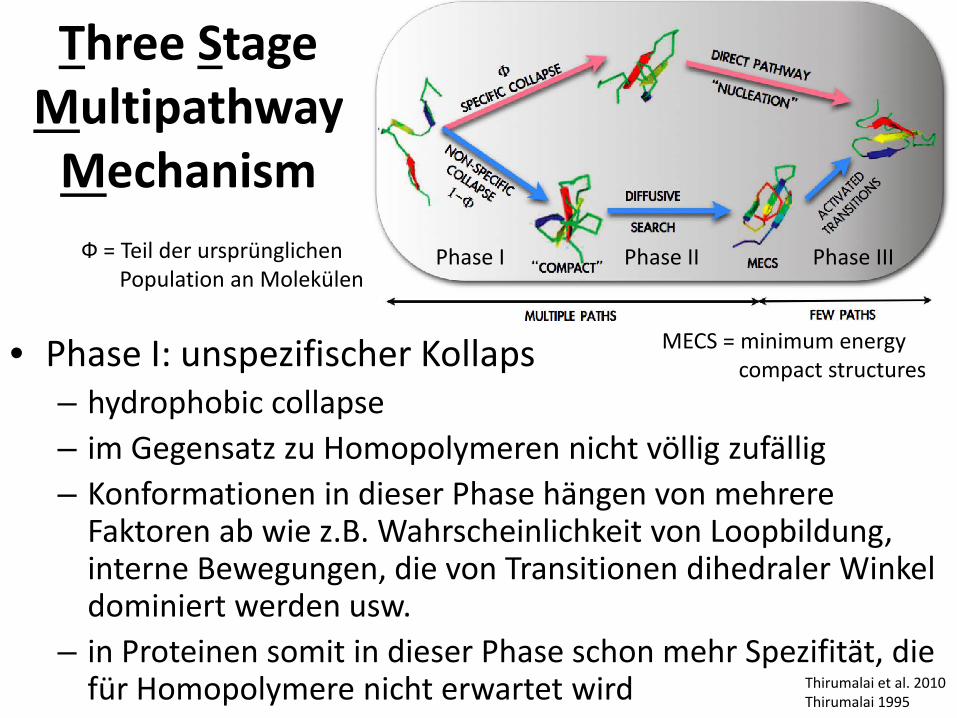
• Phase I: unspezifischer Kollaps – hydrophobic collapse – im Gegensatz zu Homopolymeren nicht völlig zufällig – Konformationen in dieser Phase hängen von mehrere
Faktoren ab wie z.B. Wahrscheinlichkeit von Loopbildung, interne Bewegungen, die von Transitionen dihedraler Winkel dominiert werden usw.
– in Proteinen somit in dieser Phase schon mehr Spezifität, die für Homopolymere nicht erwartet wird
Three Stage Multipathway
Mechanism Phase I Phase II Phase III
MECS = minimum energy compact structures
Ф = Teil der ursprünglichen Population an Molekülen
Thirumalai et al. 2010 Thirumalai 1995
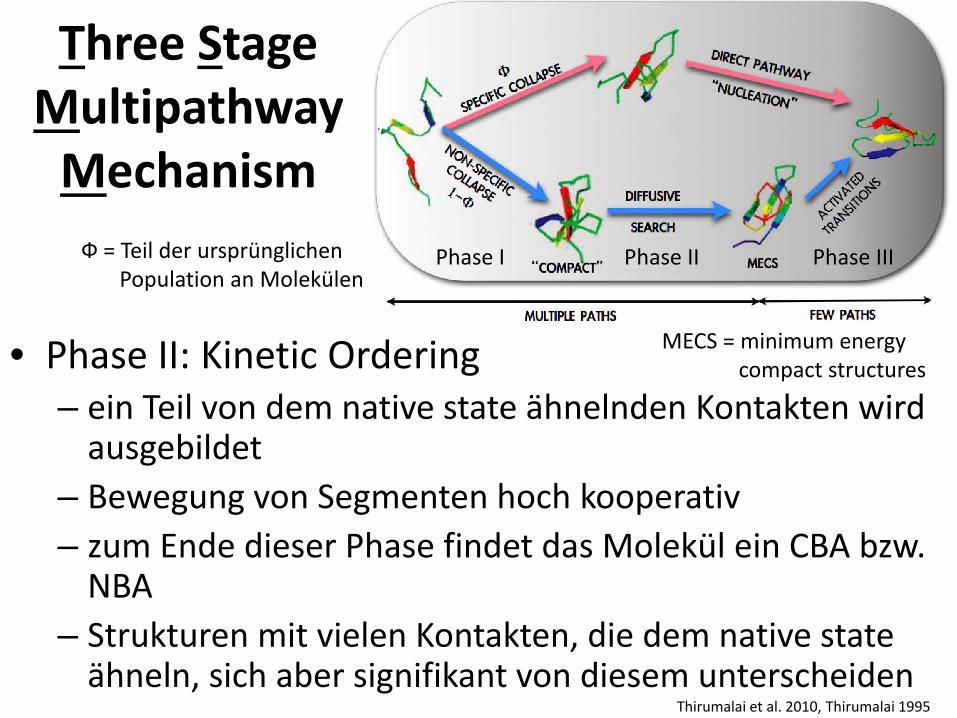
• Phase II: Kinetic Ordering – ein Teil von dem native state ähnelnden Kontakten wird
ausgebildet – Bewegung von Segmenten hoch kooperativ – zum Ende dieser Phase findet das Molekül ein CBA bzw.
NBA – Strukturen mit vielen Kontakten, die dem native state
ähneln, sich aber signifikant von diesem unterscheiden
Three Stage Multipathway
Mechanism Phase I Phase II Phase III
MECS = minimum energy compact structures
Ф = Teil der ursprünglichen Population an Molekülen
Thirumalai et al. 2010, Thirumalai 1995
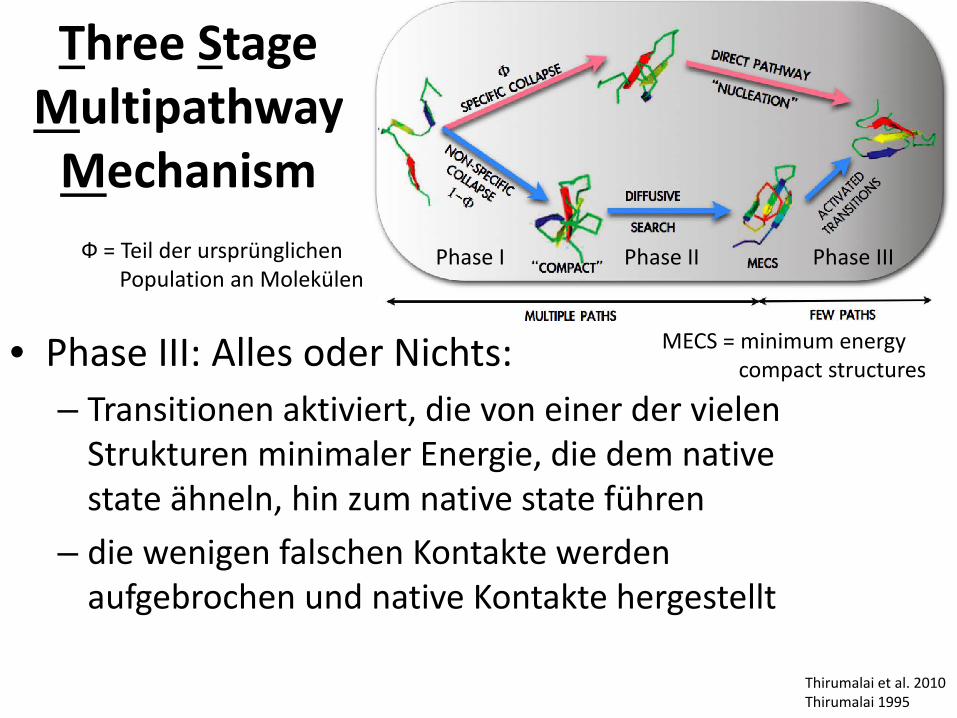
• Phase III: Alles oder Nichts: – Transitionen aktiviert, die von einer der vielen
Strukturen minimaler Energie, die dem native state ähneln, hin zum native state führen
– die wenigen falschen Kontakte werden aufgebrochen und native Kontakte hergestellt
Three Stage Multipathway
Mechanism Phase I Phase II Phase III
MECS = minimum energy compact structures
Ф = Teil der ursprünglichen Population an Molekülen
Thirumalai et al. 2010 Thirumalai 1995
Noé F et al. PNAS 2009;106:19011-19016
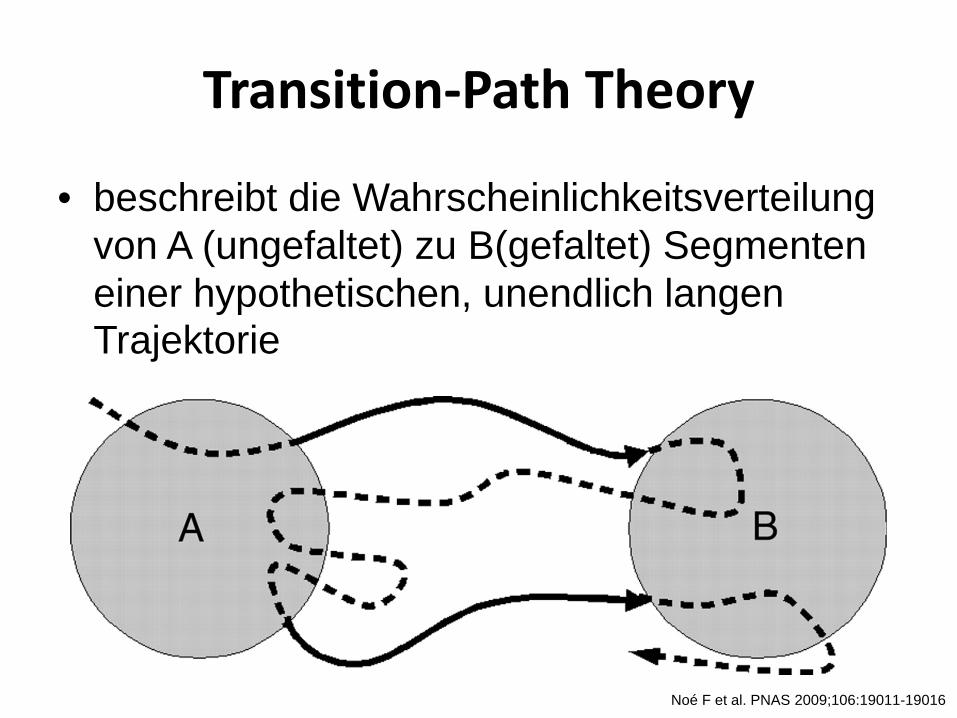
Transition-Path Theory
• beschreibt die Wahrscheinlichkeitsverteilung von A (ungefaltet) zu B(gefaltet) Segmenten einer hypothetischen, unendlich langen Trajektorie
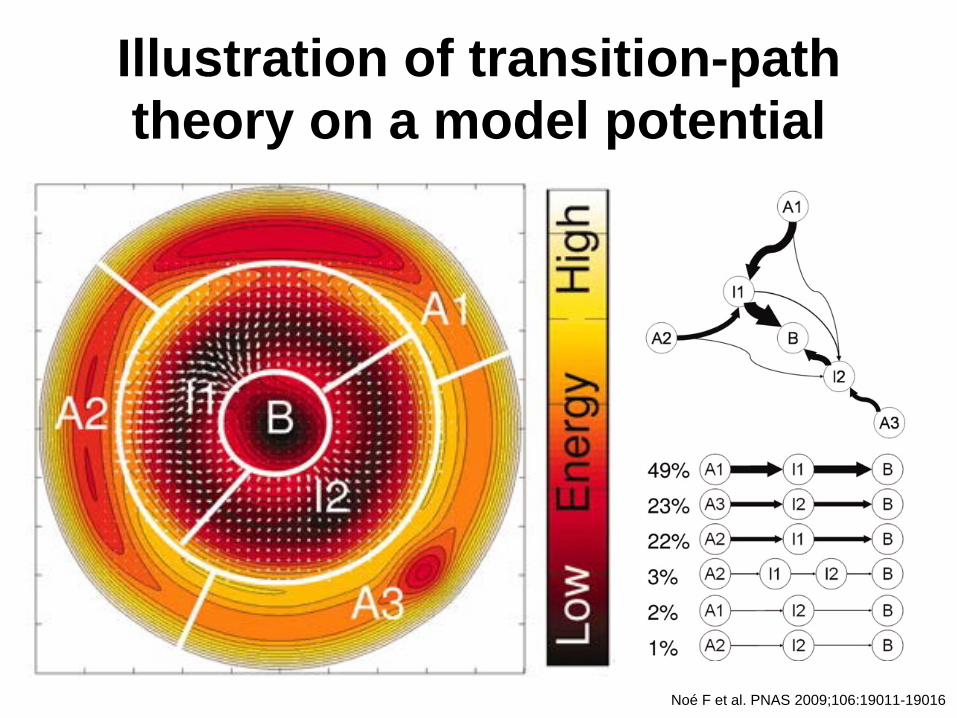
Illustration of transition-path theory on a model potential
Noé F et al. PNAS 2009;106:19011-19016
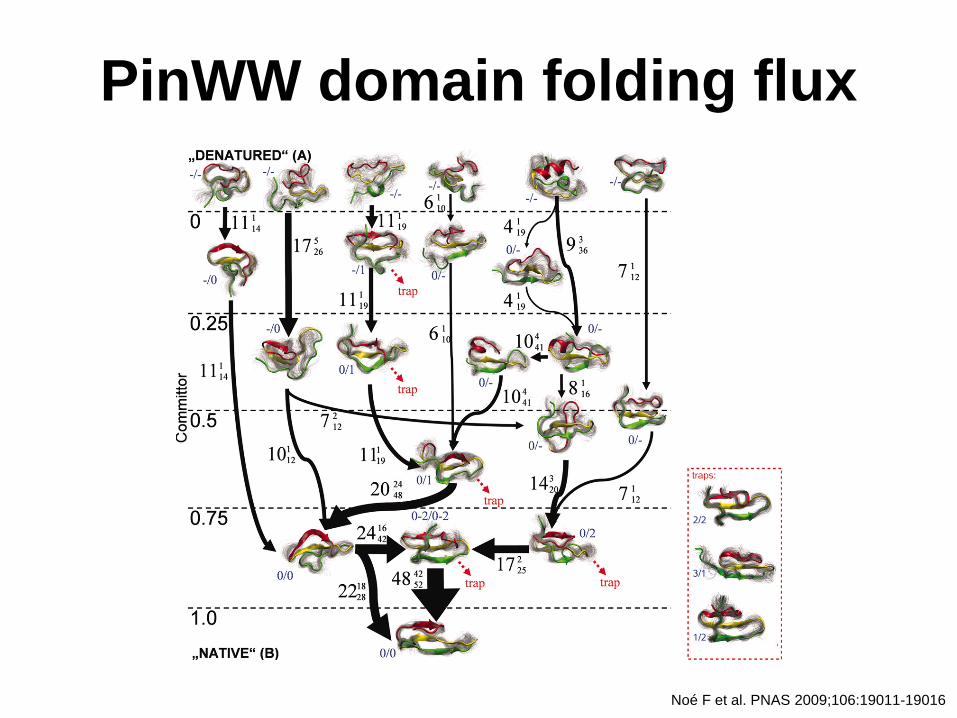
PinWW domain folding flux
Noé F et al. PNAS 2009;106:19011-19016
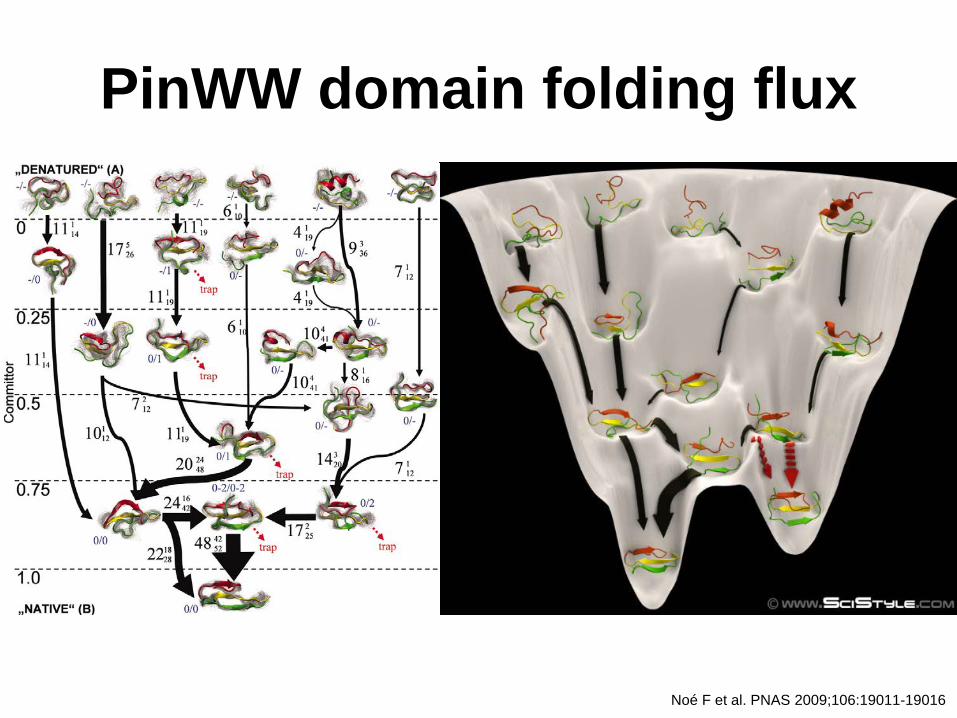
PinWW domain folding flux
Noé F et al. PNAS 2009;106:19011-19016
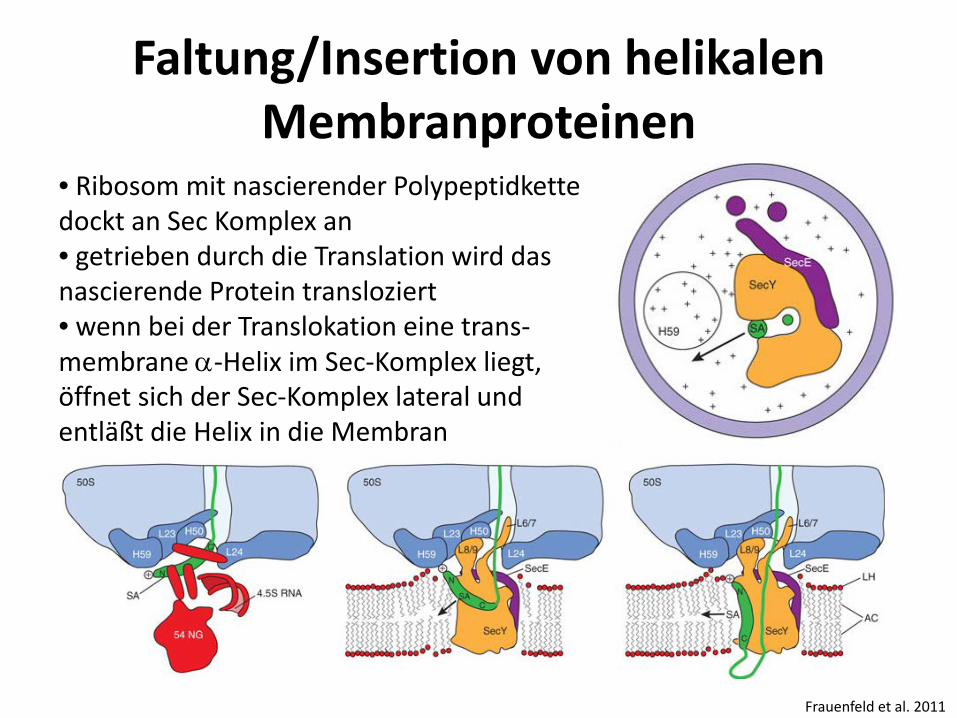
Faltung/Insertion von helikalen Membranproteinen
Frauenfeld et al. 2011
• Ribosom mit nascierender Polypeptidkette dockt an Sec Komplex an • getrieben durch die Translation wird das nascierende Protein transloziert • wenn bei der Translokation eine trans-membrane α-Helix im Sec-Komplex liegt, öffnet sich der Sec-Komplex lateral und entläßt die Helix in die Membran
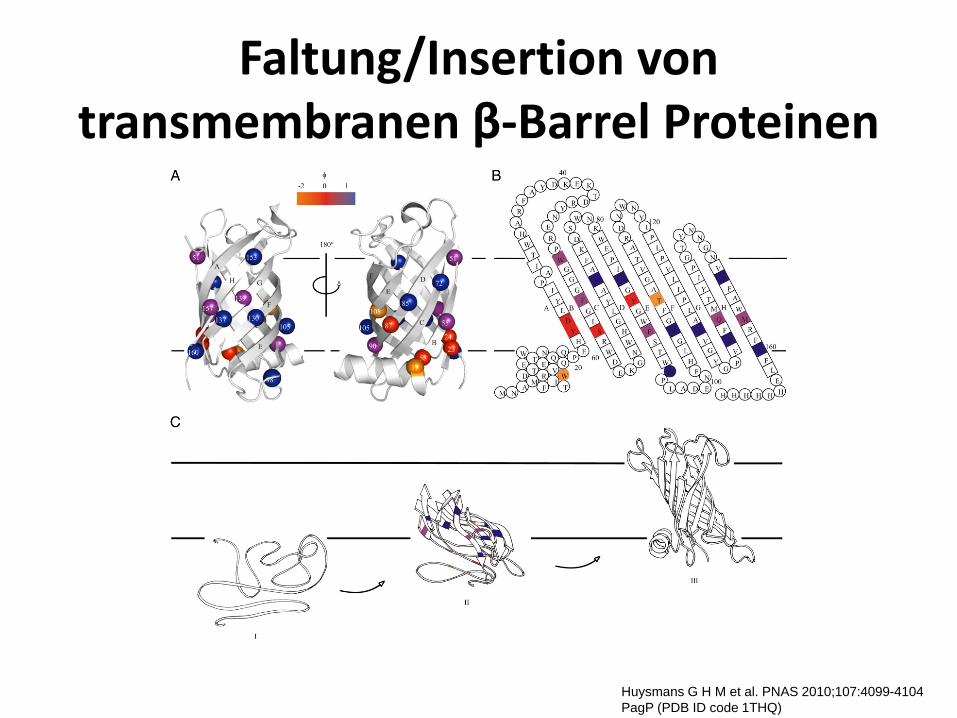
Faltung/Insertion von transmembranen β-Barrel Proteinen
Huysmans G H M et al. PNAS 2010;107:4099-4104 PagP (PDB ID code 1THQ)
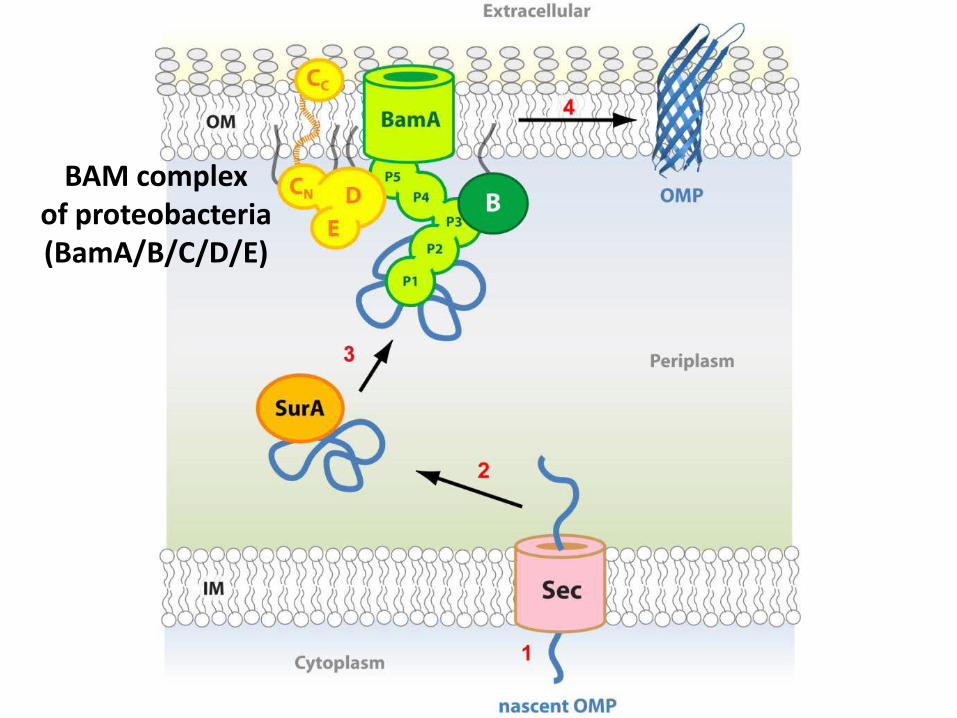
BAM complex of proteobacteria (BamA/B/C/D/E)
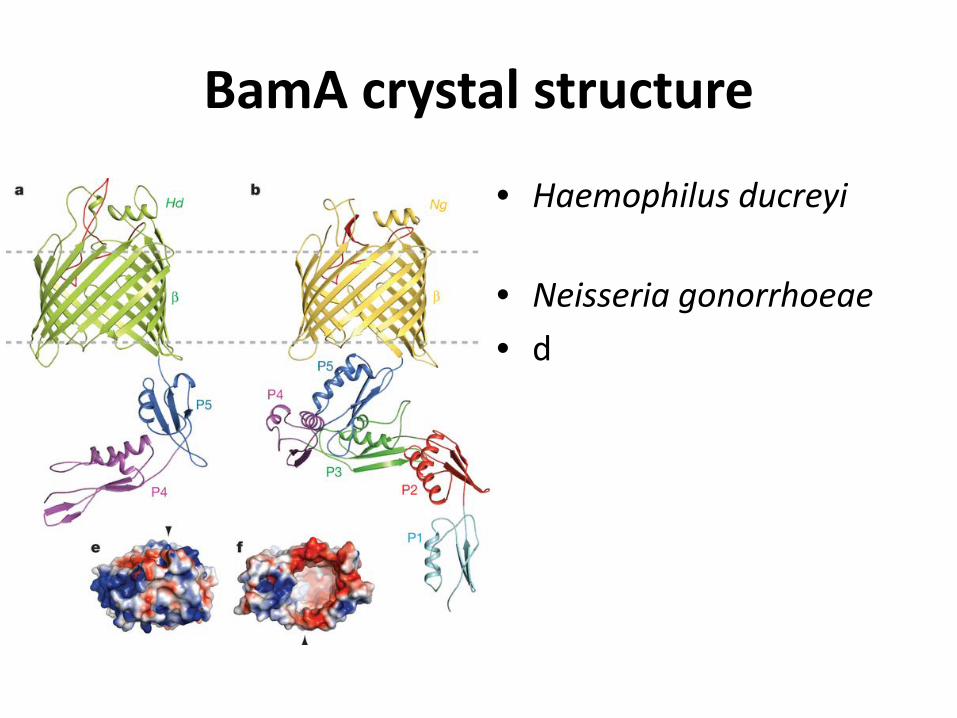
BamA crystal structure
• Haemophilus ducreyi
• Neisseria gonorrhoeae • d
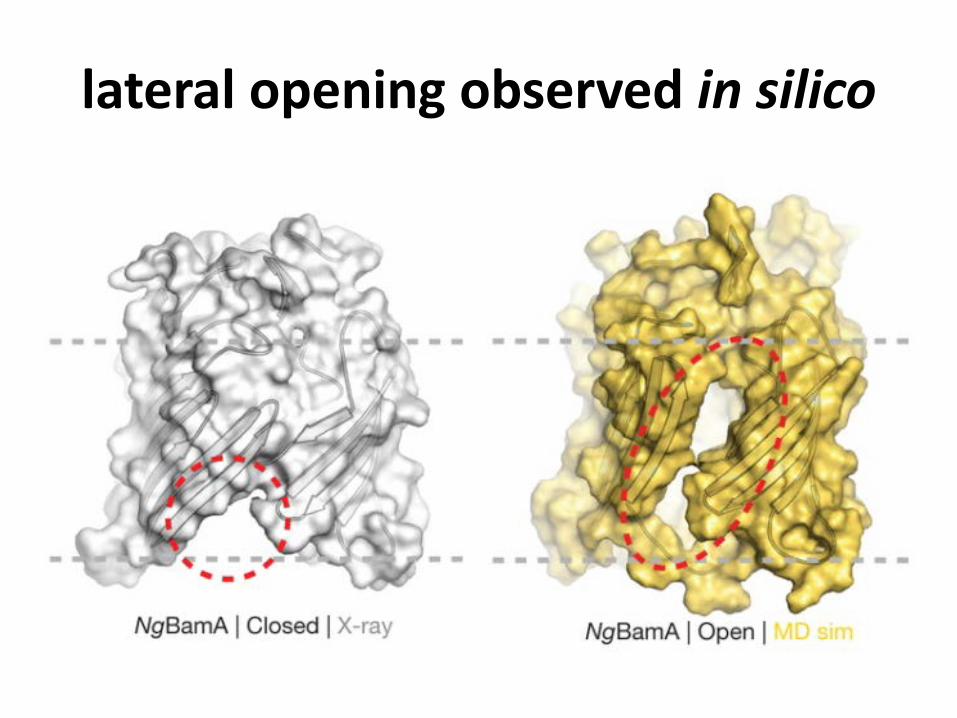
lateral opening observed in silico
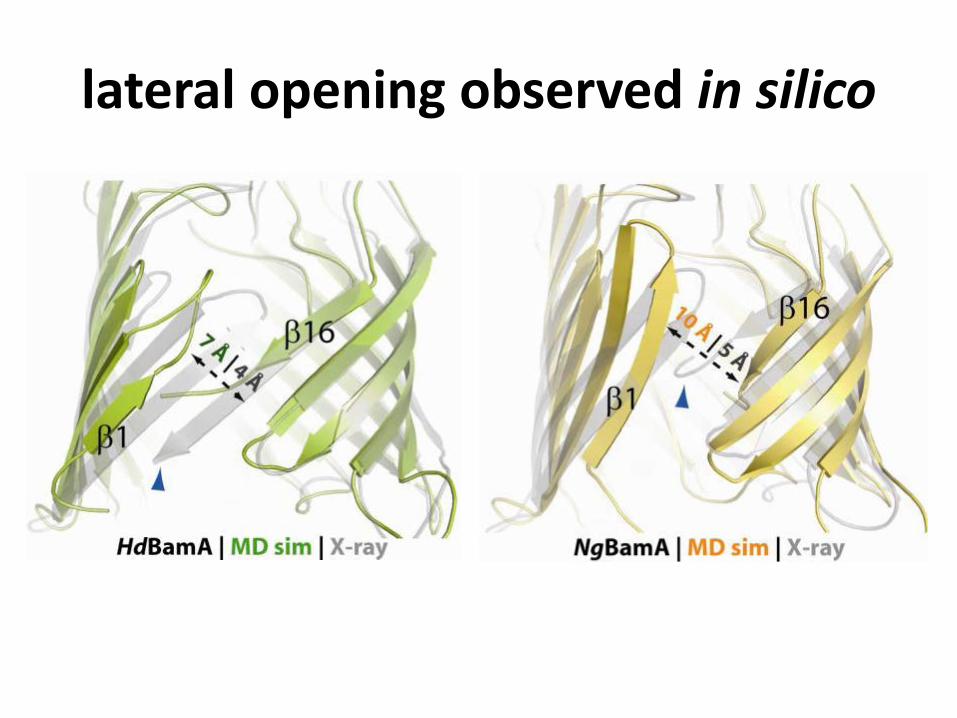
lateral opening observed in silico

Vielen Dank für Eure Aufmerksamkeit!











