Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, Apr. 1978, p. 48-590021-9193/78/0134-0048$02.00/0Copyright i 1978 American Society for Microbiology
Vol. 134, No. 1
Printed in U.S.A.
Tryptophan Biosynthesis in Saccharomyces cerevisiae:Control of the Flux Through the Pathway
GIUSEPPE MIOZZARI,t PETER NIEDERBERGER, AND RALF HUTTER*Mikrobiologisches Institut der Eidgenossischen Technischen Hochschule, CH-8092 Zurich, Switzerland
Received for publication 14 October 1977
Enzyme derepression and feedback inhibition of the first enzyme are theregulatory mechanisms demonstrated for the tryptophan pathway in Saccharo-myces cerevisiae. The relative contributions of the two mechanisms to the controlof the flux through the pathway in vivo were analyzed by (i) measuring feedbackinhibition of anthranilate synthase in vivo, (ii) determining the effect of regulatorymutations on the level of the tryptophan pool and the flux through the pathway,and (iii) varying the gene dose of individual enzymes of the pathway at thetetraploid level. We conclude that the flux through the pathway is adjusted tothe rate of protein synthesis by means of feedback inhibition of the first enzymeby the end product, tryptophan. The synthesis of the tryptophan enzymes couldnot be repressed below a basal level by tryptophan supplementation of the media.The enzymes are present in excess. Increasing or lowering the concentration ofindividual enzymes had no noticeable influence on the overall flux to tryptophan.The uninhibited capacity of the pathway could be observed both upon relievingfeedback inhibition by tryptophan limitation and in feedback-insensitive mutants.It exceeded the rate of consumption of the amino acid on minimal medium by afactor of three. Tryptophan limitation caused derepression of four of the fivetryptophan enzymes and, as a consequence, led to a further increase in thecapacity of the pathway. However, because of the large reserve capacity of the"repressed" pathway, tryptophan limitation could not be imposed on wild-typecells without resorting to the use of analogs. Our results, therefore, suggest thatderepression does not serve as an instrument for the specific regulation of the fluxthrough the tryptophan pathway.
Although tryptophan is synthesized along thesame pathway in all microorganisms studied sofar, there are considerable differences in theenzymatic organization of the pathway as wellas in the regulatory mechanisms used by differ-ent organisms to control the level of the endproduct, tryptophan.
In Saccharomyces cerevisiae, the genes cod-ing for the five enzymes of the pathway (Fig. 1)are all unlinked and mostly located on differentchromosomes. Products of the two genes, trp2and trp3, form an aggregate which catalyzes thefirst and fourth steps of the pathway (23). Tryp-tophan synthase, which catalyzes the last stepof the pathway, is coded for by a single gene (6),but it is possible to obtain mutants that eitherexcrete or utilize indole (19).Knowledge of the regulatory mechanisms con-
trolling the level of tryptophan in yeast cells isnot extensive. Lingens (15) and Doy and Cooper(5) showed feedback inhibition of anthranilate
t Present address: Department of Biological Sciences,Stanford University, Stanford, CA 94305.
synthase, the first enzyme of the pathway, bytryptophan. The significance of this mechanismin vivo was confirmed by the isolation ofmutantswith a feedback-insensitive anthranilate syn-thase, which overproduce tryptophan and ex-crete the amino acid into the medium (7, 22). Incontrast to earlier reports (5, 17), Fantes et al.(7) found no repression of the enzymes of thepathway by tryptophan. A two- to threefoldderepression of the tryptophan enzymes can,however, be generated by growth of wild-typecells in the presence of the analogs 5-methyl-tryptophan or indole acrylic acid (7, 21a, 22).These analogs were shown to cause tryptophanlimitation by inhibiting a specific step in thesynthesis of the amino acid (21a).The tryptophan pool in S. cerevisiae, as re-
ported by Fantes et al. (7) and Weiss et al. (27),is extremely small for cells growing exponen-tially in minimal medium. This suggests that theflux through the pathway must be rigorouslycontrolled and poses the question of how thiscontrol is exerted. The present report is an effortto assess the relative importance of feedback
48
on April 27, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

FLUX IN TRYPTOPHAN PATHWAY OF S. CEREVISIAE
90A*,\liz
0.
.C
IL
46
inwo>%In
_
-X
oo
w>
L 0
EEcL .
a:a.
0
0.0
-
cto
0
o I
I c
X 0\p
\E
\0
t
.0
*L.
L.
\
\0\ o\
i
9,.
11.
S o'S~
I.
I e
I
*. [t~~.
inhibition and derepression in controlling theflux through the pathway in vivo.By using completely or partially blocked mu-
tants, by varying the gene dose for individualenzymes, and by adding intermediates or analogsto the growth medium, we generated specificchanges in the flux through the pathway andmeasured the regulatory response of the cell.Further information was obtained from the char-acterization of the regulatory mutants and theireffects on the flux through the pathway in vivo.
MATERIALS AND METHODS
Organisms. All the strains were derived by muta-tion and recombination from the isogenic wild-typestrains of S. cerevisiae X2180-1A (mating type a) andX2180-1B (mating type a). The wild-type strains wereobtained from T. Manney, Manhattan, Kan. The maincharacteristics of the strains used are presented inTable 1.Most of the strains have been described previously
(7, 21a, 22). All auxotrophic strains were isolated aftermutagenic treatment with ethyl methane sulfonate.Strain RH 570 was isolated as a spontaneous proto-trophic revertant of strain RH 21. RH 375 is a 5-methyltryptophan-sensitive mutant isolated after mu-tagenesis with N-methyl-N'-nitro-N-nitrosoguanidine(22). Strains RH 558 and RH 511 were isolated from5-methyltryptophan-resistant revertants of the 5-methyltryptophan-sensitive mutant RH 487 (22); fordetails of strain isolation see Results.
Recombinants between various mutants were con-
structed either by the "spore en masse" technique or
by tetrad analysis as described before (22). They are
designated by the strain number of the parent strains.In parentheses the characteristics of the strain are
indicated. For example, RH 21 511 (trp4, Fbr) standsfor an anthranilate phosphoribosyltransferase-nega-tive, feedback-insensitive segregant from a cross be-tween RH 21 and the feedback-insensitive strain RH511 (Table 1).Media and growth conditions. YEPD medium
(2% Oxoid peptone, 1% Oxoid yeast extract, 2% glu-cose) was used as a complete medium. Minimal me-
dium MV (0.145% Difco yeast nitrogen base withoutamino acids and ammonium sulfate, 0.525% ammo-
nium sulfate, 2% glucose) was buffered to pH 4.0 with1% succinic acid and KOH. Supplements (amino acidsand analogs) were filter sterilized and added to theautoclaved media before use.
Cells were grown at 300C in liquid cultures on a
rotary shaker in Erlenmeyer flasks with several inden-tations to provide adequate aeration. Growth was fol-lowed turbidimetrically at 546 nm with a 1-cm lightpath. The specific growth rate is given as ,u and isdefined as (lnx2 - lnxl)/t2 - ti, where x stands foroptical density at the corresponding time t.Enzyme assays. All enzyme assays were per-
formed at 37°C with Triton X-100-treated cell suspen-
sions (21a).Protein content of the detergent-treated cell sus-
pensions was measured by the method of Herbert etal. (12). Bovine serum albumin (Calbiochem, La Jolla,
VOL. 134, 1978 49
on April 27, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
50 MIOZZARI, NIEDERBERGER, AND HOTTER
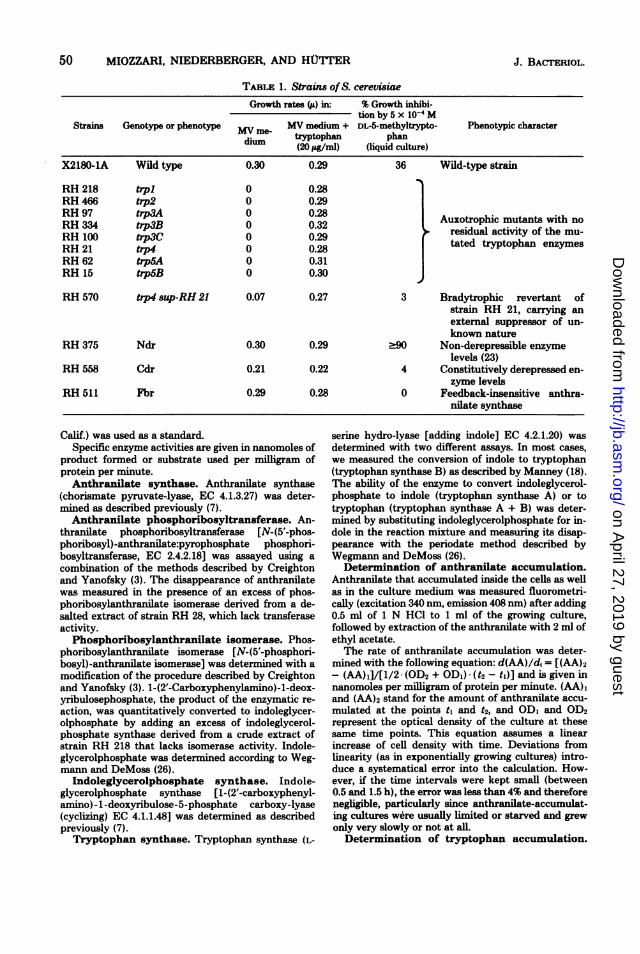
TABLE 1. Strains of S. cerevisiaeGrowth rates (j&) in: % Growth inhibi-
tion by 5 x 10-4 MStrains Genotype or phenotype MV me MV medium + DL-5-methyltrypto- Phenotypic character
dium tryptophan phandim (20 iLg/ml) (liquid culture)
X2180-1A Wild type 0.30 0.29 36 Wild-type strain
RH 218 trpl 0 0.28RH 466 trp2 0 0.29RH 97 trp3A 0 0.28 Auxotrophic mutants with no
RH 100 trp3C 0 0.29 residual activity of the mu-RH 210 trp4C 0 0.29 tated tryptophan enzymesRH 62 trp5A 0 0.31RH 15 tip5B 0 0.30
RH 570 trp4 sup-RH 21 0.07 0.27 3 Bradytrophic revertant ofstrain RH 21, carrying anexternal suppressor of un-known nature
RH 375 Ndr 0.30 0.29 290 Non-derepressible enzymelevels (23)
RH 558 Cdr 0.21 0.22 4 Constitutively derepressed en-zyme levels
RH 511 Fbr 0.29 0.28 0 Feedback-insensitive anthra-nilate synthase
Calif.) was used as a standard.Specific enzyme activities are given in nanomoles of
product formed or substrate used per milligram ofprotein per minute.Anthranilate synthase. Anthranilate synthase
(chorismate pyruvate-lyase, EC 4.1.3.27) was deter-mined as described previously (7).
Anthranilate phosphoribosyltransferase. An-thranilate phosphoribosyltransferase [N-(5'-phos-phoribosyl)-anthranilate:pyrophosphate phosphori-bosyltransferase, EC 2.4.2.18] was assayed using acombination of the methods described by Creightonand Yanofsky (3). The disappearance of anthranilatewas measured in the presence of an excess of phos-phoribosylanthranilate isomerase derived from a de-salted extract of strain RH 28, which lack transferaseactivity.
Phosphoribosylanthranilate isomerase. Phos-phoribosylanthranilate isomerase [N-(5'-phosphori-bosyl)-anthranilate isomerase] was determined with amodification of the procedure described by Creightonand Yanofsky (3). 1-(2'-Carboxyphenylamino)-1-deox-yribulosephosphate, the product of the enzymatic re-action, was quantitatively converted to indoleglycer-olphosphate by adding an excess of indoleglycerol-phosphate synthase derived from a crude extract ofstrain RH 218 that lacks isomerase activity. Indole-glycerolphosphate was determined according to Weg-mann and DeMoss (26).
Indoleglycerolphosphate synthase. Indole-glycerolphosphate synthase [1-(2'-carboxyphenyl-amino)-l-deoxyribulose-5-phosphate carboxy-lyase(cyclizing) EC 4.1.1.48] was determined as describedpreviously (7).Tryptophan synthase. Tryptophan synthase (L-
serine hydro-lyase [adding indole] EC 4.2.1.20) wasdetermined with two different assays. In most cases,we measured the conversion of indole to tryptophan(tryptophan synthase B) as described by Manney (18).The ability of the enzyme to convert indoleglycerol-phosphate to indole (tryptophan synthase A) or totryptophan (tryptophan synthase A + B) was deter-mined by substituting indoleglycerolphosphate for in-dole in the reaction mixture and measuring its disap-pearance with the periodate method described byWegmann and DeMoss (26).
Determination of anthranilate accumulation.Anthranilate that accumulated inside the cells as wellas in the culture medium was measured fluorometri-cally (excitation 340 nm, emission 408 nm) after adding0.5 ml of 1 N HCI to 1 ml of the growing culture,followed by extraction of the anthranilate with 2 ml ofethyl acetate.The rate of anthranilate accumulation was deter-
mined with the following equation: d(AA)/dt = [(AA)2- (AA),]/[1/2 (OD2 + OD,) * (t2 - t1)] and is given innanomoles per milligram of protein per minute. (AA),and (AA)2 stand for the amount of anthranilate accu-mulated at the points ti and t2, and OD, and OD2represent the optical density of the culture at thesesame time points. This equation assumes a linearincrease of cell density with time. Deviations fromlinearity (as in exponentially growing cultures) intro-duce a systematical error into the calculation. How-ever, if the time intervals were kept small (between0.5 and 1.5 h), the error was less than 4% and thereforenegligible, particularly since anthranilate-accumulat-ing cultures were usually limited or starved and grewonly very slowly or not at all.
Determination of tryptophan accumulation.
J. BACTERIOL.
on April 27, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

FLUX IN TRYPTOPHAN PATHWAY OF S. CEREVISIAE
The total amount of tryptophan that accumulatedboth inside the cells and in the culture medium wasdetermined by using a microbiological assay. Portionsof growing cultures were frozen in liquid nitrogen,transferred to a boiling water bath for 20 min, andfilter sterilized. After the addition of 0.1 volumes of 10-times-concentrated MV medium, the trypto'phan con-tent of the extract was determined by its ability topromote incorporation of ['4C]uracil (0.75 pCi/ymol)into acid-insoluble material in the double auxotrophicstrain RH 15-218 (trp5B, tipi). This method allowsthe detection of 0.2 Ag of L-tryptophan per ml (ca. 10-6M).
Determination of the tryptophan pool. The sizeof the tryptophan pool was determined in exponen-tially growing cultures with the microbiological assayusing Lactobacillusplantarum as described by Fanteset al. (7).
Construction of tetraploid gene dosage series.Tetraploid gene dosage series were constructed essen-tially as described by Hilger (13), except that the"spore en masse" technique was employed instead oftetrad analysis. All strains used for the construction oftetraploid gene dosage series had a low reversion rate(- 10-') and showed no residual growth in the absenceof tryptophan (see Table 1).
Chemicals. DL-5-Methyltryptophan was obtainedfrom Fluka AG, Buchs SG, Switzerland. Indole acrylicacid was purchased from Sigma Chemical Co., St.Louis, Mo. Chorismic acid was either commerciallyobtained from Sigma or made according to Gibson(9). 1-(2'-Carboxyphenylamino)-1-deoxyribuloaephos-phate was made by the method of Smith and Yanofsky(24). N-(5'-Phosphoribosyl)-anthranilate was pre-pared immediately before use by following the proce-dure of Creighton and Yanofsky (3).
RESULTSControl of enzyme synthesis. The levels of
the tryptophan enzymes measured in minimalas well as in Casamino Acids-supplemented me-dium could not be further repressed by exoge-nously supplied tryptophan (Table 2; see also
TABLE 2. Activities of tryptophan enzymes in thewild-type strain in the presence and absence of
exogenous tryptophana
Mediumand supple-nthrani Indoleglyc-Medumansupl- rowh at sy- erolphos-mentsb rate (u) thl phate syn-
thaseMV 0.31 1.6 2.5MV + tryptophan 0.32 1.5 2.7MV + Casamino 0.43 1.4 2.2Acids
MV + Casamino 0.43 1.4 2.0Acids + tryptophan
aEnzyme activities were determined in exponentially grow-ing cultures at an optical density between 1.5 and 2.5; they aregiven as specific activities.
bWild-type strain X2180-1A was grown in MV mediumsupplemented with L-tryptophan (50 jg/ml) and CasaminoAcids (Difco, 0.5%) as indicated.
reference 7). Moreover, the basal levels of thetryptophan enzymes provided the capacity forsupporting considerably higher growth ratesthan those observed in minimal medium. In awild-type culture growing in minimal mediumsupplemented with Casamino Acids, all theamino acids with the exception of tryptophanwere provided in the medium. This resulted ina 40% increase in growth rate. Not only didtryptophan not become growth limiting underthese conditions, the culture also failed to re-spond to the increased rate of tryptophan con-sumption by an increase in the enzyme levels.While variations in the level ofthe tryptophan
pool are not reflected in the enzyme levels aslong as the amino acid does not become growthlimiting, it is possible to elicit an increase in thespecific activities of the tryptophan enzymesunder the influence of analogs such as 5-meth-yltryptophan or indole acrylic acid (21a) or incertain bradytrophic strains carrying leaky mu-tations in one ofthe five enzymes ofthe pathway(data not shown). In all cases, this derepressionof the enzyme levels is the result of tryptophanlimitation of the culture and is coupled with amore or less pronounced inhibition of proteinsynthesis and growth (21a).
In a previous publication, we reported theisolation of a class of 5-methyltryptophan-sen-sitive mutants that have lost the ability to de-repress their tryptophan enzymes under the in-fluence of the analog (22). As shown in Table 3for strain RH 375, these mutants showed thesame enzyme levels as the wild-type strain onMV medium. However, they failed to increasetheir enzyme levels in the presence of 5-meth-yltryptophan and were therefore severely in-hibited in growth (>65% at 10-4 M 5-methyl-tryptophan). Starting from another non-dere-pressible mutant strain (RH 487; 22), we isolateda number of 5-methyltryptophan-resistant re-vertants. Strain RH 558 was derived by recom-bination of one such revertant with the wild-type strain and carries a mutation that resultsin constitutively derepressed enzyme levels. Thismutation is recessive to the wild-type allele andsegregates independently from the original mu-tation in strain RH 487. Strain RH 558 grew atabout 70% of the wild-type rate and retainedelevated enzyme levels in all media (includingYEPD medium). In contrast to similar muta-tions described by Wolfner et al. (28), strain RH558 is not temperature sensitive.By adding intermediates of tryptophan bio-
synthesis such as anthran4te or indole to thewild-type strain as well as to suitably blockedmutants, or by the addition of analogs, we triedto determine whether any of the enzymes of the
51VOL. 134, 1978
on April 27, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

52 MIOZZARI, NIEDERBERGER, AND HOTTER
TABLE 3. Specific activities of the tryptophan enzymes in regulatory mutants
Anthranilate Anthranilate Phosphoribo- Indoleglycer- Tryptophana Medium and sup- synthase phosphoribo sylanthrani olphosphate synthase GrowthStrains syltransferase late isomerase synthase rate )
SA' DF SA DF SA DF SA DF SA DF
X2180-1A (wild MV 1.5 1.1 1.2 1.5 4.3 0.30type)
MV + 5-methyl- 3.9 2.7 1.5 1.4 1.3 1.0 3.8 2.5 6.3 1.5 0.19tryptophan
RH 375 (Ndr) MV 1.4 1.0 1.3 1.6 4.2 0.30MV + 5-methyl- 1.5 1.1 1.0 1.0 1.2 0.9 1.6 1.0 4.0 0.9 0.11
tryptophan
RH 558 (Cdr) MV 4.7 3.2 3.0 2.8 1.4 1.1 5.3 3.4 11.1 2.6 0.21
a Ndr, Non-derepressible enzyme levels; Cdr, constitutively derepressed enzyme levels.b5-Methyltryptophan concentration used was 5 X 10-4 M for strain X2180-1A and 10-4 M for strain RH 375.c Derepression factor (DF) is the ratio of the specific enzyme activity (SA) under limiting conditions versus specific activity
under nonlimiting conditions; with strain RH 558 it indicates the ratio of enzyme activity of this strain as compared with thewild-type strain on MV medium. Values given represent the average of at least two separate experiments.
tryptophan pathway are subject to separate reg-ulation by the level of intermediates accumu-lated within the cell. In no case did this result inan alteration of the enzyme activities (data notshown). In particular, we found no induction oftryptophan synthase by indoleglycerolphos-phate. The intermediate was accumulatedwithin the cell along with indole under the influ-ence of the analog indole acrylic acid. Whileindole acrylic acid does cause a slight derepres-sion of tryptophan synthase (21a), this effect isnot larger than the derepression of the enzymegenerated by tryptophan limitation. Moreover,growth of the non-derepressible strain RH 375in the presence of indole acrylic acid did notresult in any derepression of tryptophan syn-thase despite the accumulation of more than 30nmol of indoleglycerolphosphate per mg dryweight (data not shown). These data are incontrast to the findings in Neurospora crassa,where indoleglycerolphosphate has been re-ported to induce tryptophan synthase (14, 20,25).
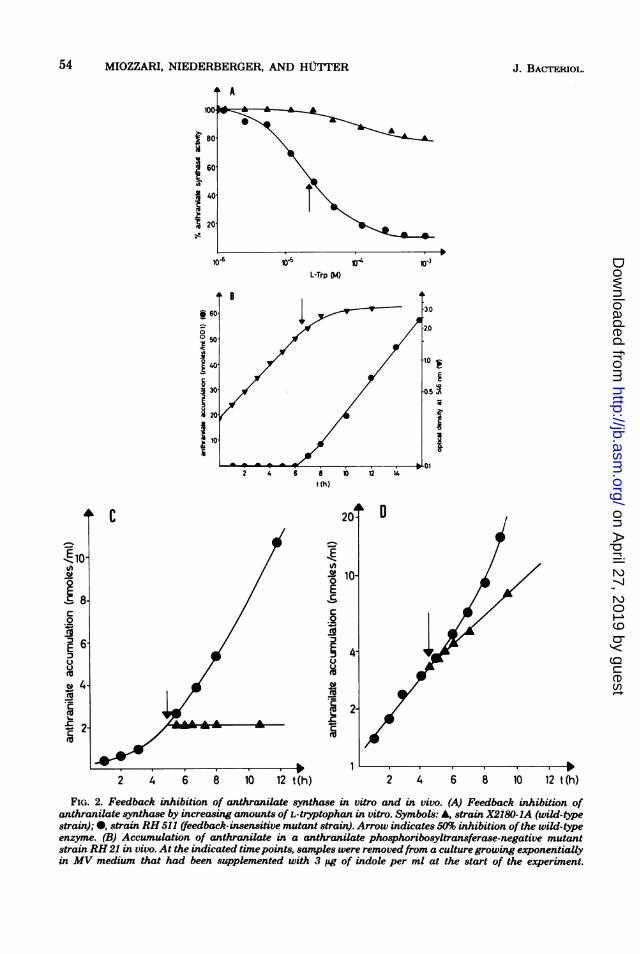
Control of enzyme activity. Feedback in-hibition of anthranilate synthase, the first en-zyme of the pathway, by tryptophan has beendemonstrated repeatedly in vitro (5, 16, 22).Figure 2A illustrates the inhibition of the en-zyme by increasing amounts of tryptophanadded to extracts of the wild-type strain or ofmutant strain RH 511, which has a feedback-insensitive anthranilate synthase. Whereas thewild-type enzyme was inhibited to about 90% at5 x 10-4 M L-tryptophan, the mutant enzymeretained more than 80% of its activity.The in vivo activity of anthranilate synthase
could be determined directly by following thetotal amount of anthranilate accumulated in
samples of a growing culture of a strain thatlacks anthranilate phosphoribosyltransferase,the second enzyme of the pathway. The fluoro-metric assay for anthranilate is very sensitive;accumulation of the intermediate was not sub-ject to product inhibition up to concentrationsof at least 40 ,umol/ml (data not shown). StrainRH 21, a mutant strain lacking anthranilatephosphoribosyltransferase activity, grew with awild-type-like growth rate when supplied withtryptophan or indole (which is endogenouslyconverted to tryptophan by tryptophan syn-thase). A culture of strain RH 21 accumulatedno detectable anthranilate as long as it grew inthe presence of nonlimiting amounts of indole(Fig. 2B). As soon as the availability of trypto-phan became growth limiting, however, as evi-denced by a reduction in growth rate, the cellsstarted to accumulate large amounts of anthra-nilate. Identical results were obtained whencomparable concentrations of L-tryptophan orDL-tryptophan were used instead of indole (datanot shown). It thus appears that tryptophanaccumulated within the cell, whether exoge-nously supplied as tryptophan or endogenouslyderived from indole, acts to prevent any furtherflux through the pathway by inhibiting the ac-tivity of anthranilate synthase. This conclusionis confirmed by the experiment presented in Fig.2C, where anthranilate accumulation in a tryp-tophan-starved culture of strain RH 21 wasshown to stop immediately when indole (or tryp-tophan) was added to the growth medium. An-thranilate accumulated at the time of indoleaddition was very stable, and no reduction influorescence was observed after 8 h of incubationat 30°C. In contrast to this, the recombinantstrain RH 21 511, which carries a feedback-
J. BACTERIOL.
on April 27, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

FLUX IN TRYPTOPHAN PATHWAY OF S. CEREVISIAE
insensitive anthranilate synthase, continued toaccumulate anthranilate even in the presence ofhigh concentrations of indole (Fig. 2D).From the amount ofanthranilate accumulated
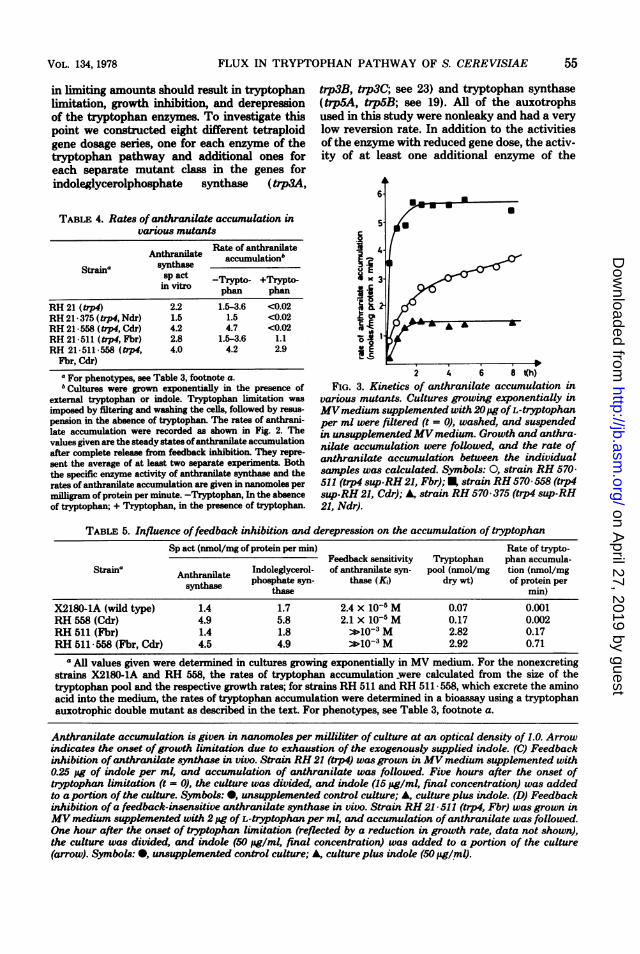
in a given time interval, it is possible to approx-imate the rate of anthranilate accumulation ofa culture and, hence, the in vivo activity ofanthranilate synthase (see Materials and Meth-ods). The steady-state rates of anthranilate ac-cumulation in recombinants of the anthranilatephosphoribosyltransferase-negative strain RH21 with various regulatory mutants are summa-rized in Table 4. In the presence of tryptophanor indole in the growth medium, only strainswith a feedback-insensitive anthranilate syn-thase accumulated anthranilate. In these strains,the rate of accumulation in the presence of tryp-tophan equaled about 65 to 70% of the unin-hibited rate in the absence of the tryptophanand was somewhat smaller than the residualactivity of80% observed in vitro (Fig. 2A). In theabsence of tryptophan, feedback inhibition wasrelieved in all strains, and it was apparent thatthe extent of anthranilate accumulation is de-pendent on the state of derepression of the cul-tures. The rates of anthranilate accumulation inthe non-derepressible strain RH 21- 375 and theconstitutively derepressed strain RH 21- 558were virtually identical to the activities of an-thranilate synthase measured in the same strinsin vitro (Table 4), whereas in strains RH 21 andRH 21-511, which can derepress their enzymesnormally, we found intermediate rates of anthra-nilate accumulation, reflecting the state of de-repression of the individual cultures.Tryptophan starvation of an auxotroph did
not lead to reproducible derepression of the en-zyme levels, since growth and protein synthesisstopped soon after the removal of the aminoacid. Strains with leaky mutations, however,continued to grow in the absence of the aminoacid, allowing protein synthesis to proceed andderepression ofthe enzymes to take place. StrainRH 570 is a revertant of strain RH 21 thatcarries an external suppressor mutation of un-known nature and grows with 25% of the wild-type rate in the absence of typtophan. As wasdone with strain RH 21, we recombined strainRH 570 with various regulatory mutations andmeasured the rate of anthranilate accumulationin these strains after removing tryptophan fromthe growth medium (Fig. 3). Whereas the rateof accumulation reached a plateau 60 to 90 minafter removal of the amino acid for the regula-tory mutants RH 570-558 and RH 570-375, itcontinued to increase over a period of at least 8h for strain RH 570- 511. This increase is gener-ated by the process of derepression and presum-
ably is a direct reflection ofthe increased activityof anthranilate synthase.Influence of derepression and feedback
inhibition on the endogenous tryptophanpool. Although measurements of the in vivoactivity of anthranilate synthase in auxotrophsunder conditions of tryptophan excess or tryp-tophan limitation may give an idea as to howthe regulatory mechanisms of derepression andfeedback inhibition function under extreme con-ditions, they do not tell to what extent thesemechanisms contribute to the control of the fluxthrough the pathway under normal conditions.Measurements of the tryptophan pool in mu-tants with altered feedback sensitivity of anthra-nilate synthase or with constitutively dere-pressed enzyme levels may provide an answer tothis question, although the interpretation ofpooldata is complicated by the fact that the aminoacid may be rapidly degraded or extensivelycompartmentalized within the cell.As reported previously by Fantes et al. (7) and
illustrated in Table 5, the tryptophan pool in awild-type strain of S. cerevisiae is extremelysmall. In contrast to this, strains with a feed-back-insensitive anthranilate synthase showeda 40-fold increase in pool level and, in addition,excreted large amounts of tryptophan into thegrowth medium. While the pool level was essen-tially the same for both strains RH 511 and RH511-558, the latter strain excreted considerablymore tryptophan into the growth medium, re-flecting its elevated enzyme levels. On the otherhand, strain RH 558 showed a mere twofoldincrease in the level of the tryptophan pool andexcreted no tryptophan into the medium despiteits constitutively derepressed enzyme levels. Itthus appears that the tryptophan pool level isrigorously controlled in strains with intact feed-back sensitivity of anthranilate synthase, whilethe amino acid is overproduced when the feed-back mechanism does not operate. From therates of tryptophan accumulation given in Table5, it is evident that wild-type cells growing inminimal MV medium utilize only a fraction oftheir enzymatic capacity to synthesize trypto-phan, whether their enzymes are derepressed ornot.Tetraploid gene dosage series. A reduction
in the gene dose for a particular enzyme canresult in either a proportional reduction in en-zyme activity, if the enzyme is synthesized con-stitutively, or an adjustment of the enzyme ac-tivity, if the corresponding enzymatic step be-comes limiting and the enzyme is derepressible.While the reduction ofan enzyme that is presentin nonlimiting amounts should have no influenceon growth, the reduction of an enzyme present
53VOL. 134, 1978
on April 27, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
54 MIOZZARI, NIEDERBERGER, AND HOTTER J. BACTERIOL.
i 80 <_
I60X
[0 1
20-
10.6 Y T
L-Trp (M)
B60. -3.0
0 20
400
2001~0.
2 4BX 12 14
246810 12 t(h) 0.1 6 8 10 12t(h)
C 2010~~~~~~~~~~~~~0
*~10-0Ei8- -&,
c c
0 (U
4-
(U~~~~~~~~~~~~~~~~~2
2 4 6 8 10 12 t(h) 2 4 6 8 10 12 t(h)
FIG. 2. Feedback inhibition of anthranilate synthase in vitro and in vivo. (A) Feedback inhibition ofanthranilate synthase by increasing amounts of L-tryptophan in vitro. Symbols: A, strain X2180-1A (wild-typestrain); *, strain RH 511 (feedback-insensitive mutant strain). Arrow indicates 50% inhibition ofthe wild-typeenzyme. (B) Accumulation of anthranilate in a anthranilate phosphoribosyltransferase-negative mutantstrain RH21 in vivo. At the indicated timepoints, samples were removedfrom a culturegrowing exponentiallyin MV medium that had been supplemented with 3 pg of indole per ml at the start of the experiment.
on April 27, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
FLUX IN TRYPTOPHAN PATHWAY OF S. CEREVISIAE
in limiting amounts should result in tryptophanlimitation, growth inhibition, and derepressionof the tryptophan enzymes. To investigate thispoint we constructed eight different tetraploidgene dosage series, one for each enzyme of thetryptophan pathway and additional ones foreach separate mutant class in the genes forindoleglycerolphosphate synthase (trpA,
TABLE 4. Rates of anthranilate accumulation invarious mutants
Rate of anthranilateAnthranilate accumulationbStXaina synthase
sp act -zrypto- +1rypto-in vitro phan phan
RH 21 (trp4) 2.2 1.5-3.6 <0.02RH 21 375 (tip4, Ndr) 1.5 1.5 <0.02RH21 558(ftp4,Cdr) 4.2 4.7 <0.02RH 21-511 (tp4, Fbr) 2.8 1.5-3.6 1.1RH 21-511 558 (trp4, 4.0 4.2 2.9
Fbr, Cdr)a For phenotypes, see Table 3, footnote a.b Cultures were grown exponentially in the presence of
extemal tryptophan or indole. Tryptophan limitation wasimposed by filtering and washing the cells, followed by resus-pension in the absence of tryptophan. The rates of anthrani-late accumulation were recorded as shown in Fig. 2. Thevalues given are the steady states ofanthranilate accumulationafter complete release from feedback inhibition. They repre-sent the average of at least two separate experiments Boththe specific enzyme activity of anthranilate synthase and therates of anthranilate accumulation are given in nanomoles permilligram of protein per minute. -Tryptophan, In the absenceof tryptophan; + Tryptophan, in the presence of tryptophan.
t7p3B, trp3C; see 23) and tryptophan synthase(trp5A, trp5B; see 19). All of the auxotrophsused in this study were nonleaky and had a verylow reversion rate. In addition to the activitiesof the enzyme with reduced gene dose, the activ-ity of at least one additional enzyme of the
6'
5 -
3-
2-.JE ~AA
2 4 6 8 Kh)FIG. 3. Kinetics of anthranilate accumulation in
various mutants. Cultures growing exponentially inMVmedium supplemented with 20,pg ofL-tryptophanper ml were filtered (t = 0), washed, and suspendedin unsupplementedMVmedium. Growth and anthra-nilate accumulation were followed, and the rate ofanthranilate accumulation between the individualsamples was calculated. Symbols: 0, strain RH 570511 (trp4 sup-RH 21, Fbr); , strain RH 570 558 (trp4sup-RH 21, Cdr); A, strain RH 570 375 (trp4 sup-RH21, Ndr).
TABLE 5. Influence offeedback inhibition and derepression on the accumulation of tryptophanSp act (nmol/mg of protein per min) Rate of trypto-
Feedback sensitivity Tryptophan phan accumula-Straina Anthraiflate Indoleglycerol- of anthranilate syn- pool (nmol/mg tion (nmol/mg
synthase phosphate syn- thase (Ki) dry wt) of protein perthase min)
X2180-1A (wild type) 1.4 1.7 2.4 x 10-5 M 0.07 0.001RH 558 (Cdr) 4.9 5.8 2.1 x 10-5M 0.17 0.002RH 511 (Fbr) 1.4 1.8 >>10-3M 2.82 0.17RH 511-558 (Fbr, Cdr) 4.5 4.9 >10-3 M 2.92 0.71
aAll values given were determined in cultures growing exponentially in MV medium. For the nonexcretingstrains X2180-1A and RH 558, the rates of tryptophan accumulation were calculated from the size of thetryptophan pool and the respective growth rates; for strains RH 511 and RH 511 558, which excrete the aminoacid into the medium, the rates of tryptophan accumulation were determined in a bioassay using a tryptophanauxotrophic double mutant as described in the text. For phenotypes, see Table 3, footnote a.
Anthranilate accumulation is given in nanomoles per milliliter of culture at an optical density of 1.0. Arrowindicates the onset ofgrowth lmitation due to exhaustion of the exogenously supplied indole. (C) Feedbackinhibition of anthranilate synthase in vivo. Strain RH 21 (trp4) was grown in MVmedium supplemented with0.25 ug of indole per ml, and accumulation of anthranilate was followed. Five hours after the onset oftryptophan limitation (t = 0), the culture was divided, and indole (15 pg/ml, final concentration) was addedto a portion of the culture. Symbols: 0, unsupplemented control culture; A, culture plus indole. (D) Feedbackinhibition ofa feedback-insensitive anthranilate synthase in vivo. Strain RH 21-511 (trp4, Fbr) was grown inMVmedium supplemented with 2 pg of L-tryptophan per ml, and accumulation of anthranilate was followed.One hour after the onset of tryptophan limitation (reflected by a reduction in growth rate, data not shown),the culture was divided, and indole (50 pg/ml, final concentration) was added to a portion of the culture(arrow). Symbols: 0, unsupplemented control culture; A, culture plus indole (50 ig/ml).
VOL. 134, 1978 55
on April 27, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
56 MIOZZARI, NIEDERBERGER, AND HOTTER
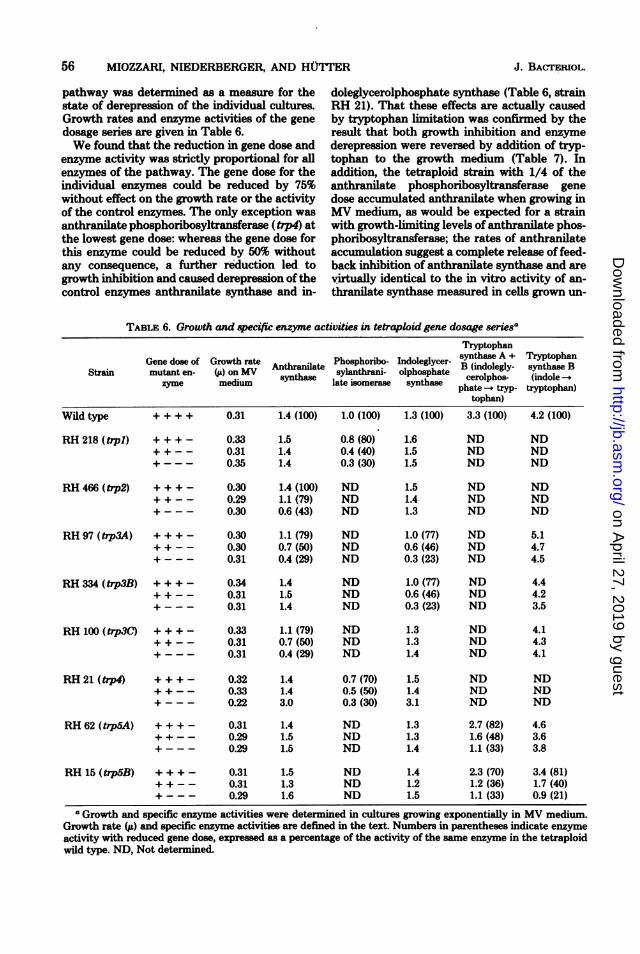
pathway was determined as a measure for thestate of derepression of the individual cultures.Growth rates and enzyme activities of the genedosage series are given in Table 6.We found that the reduction in gene dose and
enzyme activity was strictly proportional for allenzymes of the pathway. The gene dose for theindividual enzymes could be reduced by 75%without effect on the growth rate or the activityof the control enzymes. The only exception wasanthranilate phosphoribosyltransferase (trp4) atthe lowest gene dose: whereas the gene dose forthis enzyme could be reduced by 50% withoutany consequence, a further reduction led togrowth inhibition and caused derepression ofthecontrol enzymes anthranilate synthase and in-
doleglycerolphosphate synthase (Table 6, strainRH 21). That these effects are actually causedby tryptophan limitation was confirmed by theresult that both growth inhibition and enzyme
derepression were reversed by addition of tryp-tophan to the growth medium (Table 7). Inaddition, the tetraploid strain with 1/4 of theanthranilate phosphoribosyltransferase genedose accumulated anthranilate when growing inMV medium, as would be expected for a strainwith growth-limiting levels of anthranilate phos-phoribosyltransferase; the rates of anthranilateaccumulation suggest a complete release of feed-back inhibition of anthranilate synthase and arevirtually identical to the in vitro activity of an-
thranilate synthase measured in cells grown un-
TABLE 6. Growth and specific enzyme activities in tetraploid gene dosage seriesa
TryptophanGene dose of Growth rate . Phosphoribo- Indoleglycer- synthase A + Tryptophan_. . Anthramlate . . . .. . ~~~~~~B(indolegly- synthase B
Strain mutant en- y on MV Anthate sylanthrani- olphosphate ceroiphos- (idole -zyme medium synthase late isomerase synthase phate -. tryp- tryptophan)
tophan)
Wild type ++++ 0.31 1.4 (100) 1.0 (100) 1.3 (100) 3.3 (100) 4.2 (100)
RH218(typl) +++- 0.33 1.5 0.8(80) 1.6 ND ND++-- 0.31 1.4 0.4(40) 1.5 ND ND+--- 0.35 1.4 0.3(30) 1.5 ND ND
RH466(typ2) +++- 0.30 1.4(100) ND 1.5 ND ND++-- 0.29 1.1(79) ND 1.4 ND ND+--- 0.30 0.6(43) ND 1.3 ND ND
RH 97 (t7p3A) + + + - 0.30 1.1 (79) ND 1.0 (77) ND 5.1+ + - - 0.30 0.7 (50) ND 0.6 (46) ND 4.7+ -- - 0.31 0.4 (29) ND 0.3 (23) ND 4.5
RH334 (trp3B) + + + - 0.34 1.4 ND 1.0(77) ND 4.4++-- 0.31 1.5 ND 0.6(46) ND 4.2+ --- 0.31 1.4 ND 0.3 (23) ND 3.5
RH100 (tp3C + + + - 0.33 1.1(79) ND 1.3 ND 4.1++-- 0.31 0.7(50) ND 1.3 ND 4.3+--- 0.31 0.4(29) ND 1.4 ND 4.1
RH 21 (tp4) +++ - 0.32 1.4 0.7 (70) 1.5 ND ND++-- 0.33 1.4 0.5(50) 1.4 ND ND+--- 0.22 3.0 0.3(30) 3.1 ND ND
RH 62 (tp5A) + + + - 0.31 1.4 ND 1.3 2.7 (82) 4.6++-- 0.29 1.5 ND 1.3 1.6(48) 3.6+--- 0.29 1.5 ND 1.4 1.1(33) 3.8
RH 15 (trp5B) + + + - 0.31 1.5 ND 1.4 2.3 (70) 3.4 (81)+ + - - 0.31 1.3 ND 1.2 1.2 (36) 1.7 (40)+ --- 0.29 1.6 ND 1.5 1.1 (33) 0.9 (21)
a Growth and specific enzyme activities were determined in cultures growing exponentially in MV medium.Growth rate (i) and specific enzyme activities are defined in the text. Numbers in parentheses indicate enzymeactivity with reduced gene dose, expressed as a percentage of the activity of the same enzyme in the tetraploidwild type. ND, Not determined.
J. BACTERIOL.
on April 27, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

FLUX IN TRYPTOPHAN PATHWAY OF S. CEREVISIAE
TABLE 7. TIrptophan limitation at reduced gene doses of anthranilate phosphoribosyltransferaseaGrowth rate (u) in Anthranilate syn- Indoleglycerolphos- Rate of anthranilate
medium: thase phate synthase accumulationStrain trp4 gene dose
MV MMV + MV MV+ MV MV+ MV MV +trp trp trp IA
RH 21 (trp4) + + + + 0.31 ND 1.7 ND 1.5 ND 40.02 1.6+ + - 0.32 ND 1.6 ND 1.7 ND t60.02 2.2+ + - - 0.33 0.34 1.6 1.90 1.6 2.2 G0.02 2.6+--- 0.24 0.35 4.0 2.15 3.7 1.9 3.61 3.2
aSpecific enzyme activities and rates of anthranilate accumulation are given in nanomoles per milligram ofprotein per minute. MV + trp, MV medium supplemented with 50 pg of L-tryptophan per ml. MV + IA, MVplus 2.5 x 10-5 M indole acrylic acid. All strains were grown overnight in MV medium supplemented with 50,g of L-tryptophan per ml, filtered at an optical density of 0.4, washed, and suspended in unsupplemented MVmedium. After 30 min of incubation to deplete the tryptophan pool, the analog was added. Samples wereremoved at various time points for the determination of growth and anthranilate accumulation. The valuesgiven represent the rates of anthranilate accumulation 4 h after addition of the analog. ND, Not determined.
der the same conditions in MV medium.While the strains with higher trp4 gene dose
did not accumulate anthranilate during growthin minimal medium, they did so in the presenceof indole acrylic acid (Table 7). The analogcauses a depletion of the tryptophan pool and,as a consequence, release of feedback inhibitionof anthranilate synthase (21a). The rates of an-thranilate accumulation reflected the levels ofanthranilate phosphoribosyltransferase presentat a particular trp4 gene dose; a reduction ofanthranilate phosphoribosyltransferase activityby 25% led to an increase in the rate of accu-mulation of the intermediate by 0.5 to 0.6nmol/mg of protein per min. The uninhibitedrate of anthranilate formation by anthranilatesynthase exceeded the capacity of anthranilatephosphoribosyltransferase to convert the inter-mediate even in the presence of a full trp4 genedose. Since anthranilate phosphoribosyltrans-ferase is present in the smallest excess of alltryptophan enzymes, this is not surprising.
DISCUSSION
The role offeedback inhibition ofanthra-nilate synthase. With 0.07 nmol/mg dryweight, the tryptophan pool of the wild-typestrain X2180-1A of S. cerevisiae (7) is consider-ably smaller than the published values for otheramino acids in yeast (4, 8) and more than 10times smaller than the tryptophan pool in Neu-rospora crassa (21). Because of its small size,the buffering capacity of the tryptophan poolcan be only very limited, and one could expectthat even a small reduction in the flux throughthe pathway should result in tryptophan limi-tation. As shown by the analysis of growth andenzyme activities in a tetraploid gene dosageseries of all five tryptophan enzymes, none ofthe enzymes of the pathway is synthesized in
limiting amounts. With one exception, whichwill be discussed below, gene dose and enzymeactivities could be reduced by as much as 75%without any effect on growth rate or on theactivities of the other enzymes of the pathway.That at least the last four enzymes of the path-way are produced in excess is also evident fromthe fact that mutants with a feedback-insensitiveanthranilate synthase showed a 40-fold increasein the tryptophan pool level and, in addition,excreted the amino acid into the medium. Thebreakdown of regulation of the tryptophan poolin these mutants suggests that feedback inhibi-tion of anthranilate synthase plays a central rolein the control of the flux through the pathway.This conclusion was corroborated by measur-
ing the rate of anthranilate accumulation inmutants lacking anthranilate phosphoribosyl-transferase activity, both in the presence and inthe absence of tryptophan. The amino acid,whether introduced exogenously as tryptophanor synthesized endogenously from indole, causeda complete inhibition of anthranilate synthase,while removal of tryptophan from the growthmedium led to the accumulation of largeamounts of anthranilate. The rates of anthrani-late accumulation in strains carrying variousregulatory mutations were virtually identical tothe corresponding levels of anthranilate syn-thase activity determined in vitro and reflect thestate of derepression of the cultures. For maxi-mally derepressed cultures, they reached valuesabout 10 times higher than the 0.3 nmol oftryptophan per mg of protein per min (7) neededin the wild-type strain to maintain the growthrate of 0.3 in MV medium. The great majorityof tryptophan is probably used for protein syn-thesis, since the amount needed for nicotinicacid biosynthesis is very small, and the degra-dative enzymes presumably need to be inducedby growth in the presence of exogenous trypto-
57VOL. 134, 1978
on April 27, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

58 MIOZZARI, NIEDERBERGER, AND HOTTER
phan (10, 11); or, alternatively, their affinityconstants for tryptophan may require high in-ternal concentrations of the amino acid. Fromthe uninhibited rate of anthranilate accumula-tion by anthranilate synthase in vivo, about 1.5nmol/mg of protein per min, and the calculatedflux through the pathway of 0.31 nmol/mg ofprotein per min, it follows that the tryptophanpool inhibits the activity of anthranilate syn-thase by about 75 to 80% in a wild-type culturegrowing in minimal MV medium. This agreeswell with the estimation of Fantes et al. (7)based on in vitro data only.While the feedback-insensitive strain RH 511
and its constitutively derepressed derivative RH511-558 accumulated tryptophan at 170 and 710times the wild-type rate, respectively, the con-stitutively derepressed strain RH 558 did notaccumulate the amino acid in significantlyhigher amounts than the wild-type strain itself(Table 5). This means that derepression of theenzymes of the pathway does not lead to anincrease of the flux; quite to the contrary, thenet flux through the pathway in strain RH 558was reduced to 0.23 nmol/mg of protein per minbecause of the slower growth rate of this mutantstrain (Ju = 0.22 in MV medium). Thus, whilederepression clearly affects the maximal capac-ity of both anthranilate synthase and the tryp-tophan pathway as a whole, this increased po-tential is expressed only under conditions wherefeedback inhibition of anthranilate synthasedoes not operate, such as during tryptophanlimitation or in feedback-insensitive mutants.Both an increase in anthranilate synthase con-
centration in derepressed cells and a reductionof it by decreasing the gene dose of the enzymeremained without effect on the flux through thepathway. Since the activities of the enzyme inderepressed cells (4.6 nmol/mg of protein permin) and in the tetraploid strain with the small-est gene dose (0.36 nmol/mg of protein per minfor tip3C) differed by more than 10-fold, it isevident that the net flux through the pathway isindependent from the absolute amount of en-zyme present.The flux through the pathway is adjusted to
the growth rate by feedback inhibition of an-thranilate synthase only. The feedback inhibi-tion constant (KJ) of the enzyme and the growthrate also determine the size of the tryptophanpool.
Control of enzyme synthesis. As shownpreviously by Fantes et al. (7), the tryptophanenzymes are not regulated in response to thelevel of the tryptophan pool; neither increasedpool levels in the presence of exogenously sup-plied tryptophan or indole, or as a result of afeedback-insensitive anthranilate synthase, nor
reduced pool levels in mutants with increasedfeedback sensitivity of anthranilate synthase af-fect the activity of the tryptophan enzymes. Inthis study we have shown that the activity of alltryptophan enzymes is proportional to their genedose, indicating that they are transcribed andtranslated constitutively. In addition, the en-zymes of the pathway are synthesized in at leasta threefold excess during growth on minimalmedium. A slight deviation from the strict pro-portionality was observed for anthranilate syn-thase in the trp2 gene dosage series. This maybe due to the fact that the activity of anthrani-late synthase is dependent on the formation ofan intact aggregate between the trp2 and trp3gene products (23).An increase in the specific activity of four of
the five tryptophan enzymes can be elicited bygrowth under conditions where the availabilityoftryptophan becomes growth limiting, as in thepresence of analogs or in bradytrophic mutantswith a leaky mutation in one of the enzymes ofthe pathway. It has been shown (P. Niederber-ger, Ph.D. thesis, Eidgenossische TechnischeHochschule, Zurich, Switzerland, 1977) that thisderepression is not a specific response of the cellto tryptophan limitation, since derepression ofthe tryptophan enzymes also occurs upon limi-tation of unrelated amino acids such as leucine,histidine, and arginine. Similarly, tryptophanlimitation causes derepression of histidine andarginine as well as tryptophan enzymes (22).This general control of amino acid biosynthesishas been reported by several laboratories forboth N. crassa (1, 2) and S. cerevisiae (4, 22, 28)and affects at least the basic, the aromatic, andthe branched-chain amino acid pathways.Capacity of the pathway. Anthranilate
phosphoribosyltransferase, the second enzymeof the pathway, was synthesized in the smallestexcess of all tryptophan enzymes and thus lim-ited the maximal capacity of the pathway underconditions where feedback inhibition of anthra-nilate synthase was eliminated. While a 50%reduction of the gene dose for the enzyme didnot influence growth and enzyme activities, afurther reduction caused the enzyme to becomelimiting and led to reduced growth rate andderepression of the other enzymes of the path-way. From the rates of anthranilate accumula-tion in the strains of the trp4 gene dosage seriesunder the influence of indole acrylic acid (Table6), one can calculate that the anthranilate phos-phoribosyltransferase activity in a strain with anintact trp4 gene dose represents about 65% ofthe uninhibited activity of anthranilate syn-thase. This number is consistent with the ratioof the activities of the two enzymes determinedin vitro. From the uninhibited in vivo activity of
J. BACTrERIOL.
on April 27, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

FLUX IN TRYPTOPHAN PATHWAY OF S. CEREVISIAE
anthranilate synthase, 1.5 nmol/mg of proteinper min in a non-derepressed strain, it followsthat the basic capacity of the pathway is about1 nmol/mg of protein per min, about a threefoldexcess over the value of 0.31 nmol/mg of proteinper min needed to maintain the growth rate ofthe wild-type strain i minimal MV medium.This agrees with the fact that tetraploid cellsneed less than 50% but more than 25% of theanthranilate phosphoribosyltransferase activityfor normal growth.Although derepression of the tryptophan en-
zymes led to a further increase in the capacity ofthe pathway, the basic capacity of the pathwayseemed to be sufficient to support growth underall conditions. In particular, growth in the pres-ence of all amino acids except tryptophan didnot result in tryptophan limitation or derepres-sion despite a 40% increase in growth rate com-pared to unsupplemented minimal MV medium.Because of the large reserve capacity of the non-derepressed pathway, it was virtually impossibleto cause tryptophan limitation of the wild-typestrain without the use of analogs or the intro-duction of mutations that reduce the fluxthrough the pathway. Thus, tryptophan limita-tion is unlikely to be encountered by wild-typecells under natural conditions. Possibly, de-repression of the tryptophan enzymes is onlyone aspect of a general response of the cell toamino acid limitation.
ACKNOWLEDGMETThis work was supported by the Swiss National Foundation
for Scientific Research projects 3.324.70 and 3.053.73.
LrTERATURE CMD1. Carsiods, ML, and R. F. Jones. 1974. Cross-pathway
regulation: tryptophan-mediated control of histidineand arginine biosynthetic enzymes in Neurosporacrassa. J. Bacteriol. 119:889-892.
2. Carsiotls, M., R. F. Jones, and A. C. WeselHng. 1974.Cross-pathwayregulation: histidine-mediated control ofhistidine, tryptophan, and argnine biosynthetic en-zymes in Neuro.pora crassa J. Bacteriol. 119:893898.
3. Creighton, T. E., and C. Yanofaky. 1970. Chorismateto tryptophan (EAcherichia coLi)-anthranilate synthe-tase, PR transferase, PRA isomerase, InGP synthetase,tryptophan synthetase. Methods Enzymol. 17A:365-380.
4. Delforge, J., F. Mesmnguy, and J. M. Wiame. 1975.The regulation of arginine biosynthesis in Saccharo-myces cerevisiae. The specificity of argR- mutationsand the general control of amino-acid biosynthesis. Eur.J. Biochem. 57:231-239.
5. Doy, C. HI, and J. ML Cooper. 1966. Aromatic biosyn-thesis in yeast. L. The synthesis of tryptophan and theregulation of this pathway. Biochim. Biophys. Acta127:302-316.
6. Duntze, W., and T. R. Manney. 1968. Two mechanimsof allelic complementation among tryptophan synthe-tase mutants ofSaccharomyce8 cerevisiae. J. Bacteriol.96:2086-2093.
7. Fantes, P. A.,L K Roberts, and R. Hutter. 1976. Freetryptophan pool and tryptophan biosynthetic enzymesin Saccharomyces cerevisiae. Arch. Microbiol.
107:207-214.8. Gancedo, J. M., and C. Gancedo. 1973. Concentration
of intermediary metabolites in yeast. Biochimie55:205-211.
9. Gibson, F. 1970. Preparation of choriamic acid. MethodsEnzymol. 17A:362-364.
10. Heilmann, H. D., and F. Lingens. 1968. Reinigung undEigenschaften der 3-Hydroxyanthranilat-Oxygenaseaus Saccharomyces cerevisiae. Z. Physiol. Chem.349:223-230.
11. Heilmann, H. D., and F. Lingens. 1968. Zur Regulationder Nicotinsaure-Biosynthese in Saccharomyces cere-visiae. Z. Physiol. Chem. 349:231-236.
12. Herbert, D., P. J. Phipps, and R. E. Strange. 1971.Chemical analysis of microbial cells, p. 209-344. In J.R. Norris and D. W. Ribbons (ed.), Methods in micro-biology, vol. 5B. Academic Press Inc., New York.
13. Hilger, F. 1973. Construction and analysis of tetraploidyeast sets for gene dosage studies. J. Gen. Microbiol.75:23-31.
14. Lester, G. 1971. Regulation of tryptophan biosyntheticenzymes in Neurospora crassa. J. Bacteriol. 107:193-202.
15. Lngens, F. 1968. Die Biosynthese aromatischer Amino-sauren und deren Regulation. Angew. Chem. 80:384-394.
16. Lingens, F., W. Goebel, and H. Uesseler. 1966. Regu-lation der Biosynthese der aromatischen Aminosaurenin Saccharomyces cerevisiae. 1. Hemmung der En-zymaktivititen (Feedback-Wirkung). Biochem. Z.346:357-367.
17. Lingens, F., W. Goebel, and H. Ueseler. 1967. Regu-lation der Biosynthese der aromatischen Aminosaurenin Saccharomyces cereviaiae. 2. Repression, Induktionund Aktivierung. Eur. J. Biochem. 1:363-374.
18. Manney, T. R. 1968. Regulation of factors that influencethe in vitro stability of tryptophan synthetase fromyeast. J. Bacteriol. 96:403-408.
19. Manney, T. R., W. Duntze, N. Janooko, and J. Sala-zar. 1969. Genetic and biochemical studies of partiallyactive tryptophan synthetase mutants of Saccharomy-ces cerevisiae. J. Bacteriol. 9:590-596.
20. Matchett, W. H. 1972. Inhibition of tryptophan synthe-tase by indoleacrylic acid. J. Bacteriol. 110:146-154.
21. Matchett, W. H., J. R. Turner, and W. R. Wiley. 1968.The role oftryptophan in the physiology ofNeurospora.Yale J. Biol. Med. 40:257-283.
21a. Mizzoari, G., P. Niederberger, and R. Hutter. 1977.Action of tryptophan analogues in Saccharomyces cer-evisiae. Arch. Microbiol. 115:307-316.
22. Schurch, A., J. Miozzari, and R. Hutter. 1974. Regu-lation of tryptophan biosynthesis in Saccharomycescerevisiae: mode of action of 5-methyl-tryptophan and5-methyl-tryptophan-sensitive mutants J. Bacteriol.117:1131-1140.
23. Schurch-Rathgeb, Y. 1972. Der trp3-Locus von Saccha-romyces cerevisiae. Arch. Genet. 45:129-192.
24. Smith, O. H, and C. Yanofsky. 1962. Enzymes involvedin the biosynthesis of tryptophan. Methods Enzymol.5:794-806.
25. Turner, J. R., and W. H. Matchett. 1968. Alteration oftryptophan-mediated regulation in Neurospora crassaby indoleglycerol phosphate. J. Bacteriol. 96:160-1614.
26. Wegmnan, J., and J. A. DeMoss. 1965. The enzymicconversion of anthranilate to indolylglycerol phosphatein Neurospora crasa J. Biol. Chem. 240:3781-3788.
27. Weiss, R. L., J. R. Kukova, and J. Adams. 1975. Therelationship between enzyme activity, cell geometry,and fitness in Saccharomyces cerevisiae. Proc. Natl.Acad. Sci. U.S.A. 72:794-798.
28. Wolfner, M., D. Yep, F. Me8senguy, and G. R. Fink.1975. Integration of amino acid biosynthesis into thecell cycle of Saccharomyces cerevisiae. J. Mol. Biol.90:273-290.
59VOL. 134, 1978
on April 27, 2019 by guest
http://jb.asm.org/
Dow
nloaded from





























