Embed Size (px)
Citation preview

196 R. Schopf
MULLER, W.; REYMERS, A.; LUNDERSTADT, J., 1981 : Zur Nahrungsqualitat von Fichtennadeln fur forstliche Schadinsekten. 15. Vergleichende Untersuchungen der Entwicklung und Proteinaus- wertung unterschiedlich alter Larven von Gilpiniu hercyniue (Hym., Diprionidae) an gleichem Nadelmaterial von Fichte (Piceu ubies Karst.) unter kontrollierten und Freilandbedingungen. Z. ang. Ent. 91, 113-123.
OLDIGES, H., 1958: Waldbodendungung und Schadlingsfauna des Kronenraumes. AFZ 13,
OTTO, D., 1970: Zur Bedeutung des Zuckergehaltes der Nahrung fur die Entwicklung nadelfres- sender Kieferninsekten. Arch. Forstwes. 19, 135-150.
SCHOPF, R., 1979: Zur Nahrungsqualitat von Fichtennadeln fur forstliche Schadinsekten. 1 1 . Aufnahme und Ausscheidung I4C-markierter Verbindungen durch Larven von Gilpiniu hercyniue (Hym., Diprionidae) nach Verfutterung radioaktiver Fichtennadeln. 2. ang. Ent. 87,
- 1981a: Untersuchungen zum Aminosaurenstoffwechsel der Fichtennadeln fressenden Blatt- wespe Gilpinia hercyniue Htg. (Hym., Diprionidae). Z. ang. Ent. 92, 84-92.
- 1981b: Speicherung und Mobilisierung von Kohlenhydraten und Lipiden in der Blattwespe Gilpiniu hercyniue. Ent. exp. appl. 29, 218-227.
- 1982: Die Bestimmung der Nahrungsqualitat zur Beurteilung der Eignung von Waldbaumen fur den Befall durch phytophage Insekten. Forstarchiv li3, 205-208.
SCHOPF, R.; MIGNAT, C.; HEDDEN, P., 1982: As to the food quality of spruce needles for forest damaging insects. 18. Resorption of secondary plant metabolites by the sawfly Gilpiniu hercyniue Htg. (Hym., Diprionidae). Z. ang. Ent. 93, 244-257.
SCHWENKE, W., 1960: Uber die Wirkung der Walddungung auf die Massenvermehrung der Kiefernbuschhornblattwespe (Diprion pini L.) 1959 in Mittelfranken und die hieraus ableit- baren gradologischen Folgen. Z. ang. Ent. 46, 371-378.
- 1968: Neue Hinweise auf eine Abhangigkeit der Vermehrung blatt- und nadelfressender Forstinsekten vom Zuckergehalt ihrer Nahrung. Z. ang. Ent. 46, 365-369.
SUMERE, C. F. VAN; ALBRECHT, J.; DEDONDER, A.; POOTER, H.; PE, I., 1975: Plant proteins and phenolics. Chem. and Biochem. of Plant Proteins, ed.: J. B. HARBORNE, C. F. VAN SUMERE, Acad. Press. Chap. 8, 211-264.
138-140.
262-2 76.
SWAIN, T., 1977: Secondary compounds as protective agents. Ann. Rev. Plant Phys. 28,479-501.
Anschrift des Verfassers: Dr. REINHARD SCHOPF, Abt. fur Forstzoologie, Institut fur Standort- lehre und Waldhygiene der Universitat, Busgenweg 3, D-3400 Gottingen-Weende
Aus dem Zoologischen Znstitut und Zoologischen Museum der Universitat Hamburg
Ober das Trockenfallen von Simuliiden-Eiern (Dipt., Simuliidee)
Von W. RUHM',~
Abstract On the desiccation of simuliid eggs (Dipteru, Simuliidae)
Because of the danger of desiccation due to emergence from water, the development of e gs to the point of hatching of first stage larvae depends on the type of deposite produced by femies. With the eggs being laid on a substrate in the vicinity of the water surface the factors which came into play during emergence and which can influence development, can be classified according humid- i t p phase of embryonic development, substrate as well as those of the coincidence of time and p ace.
* Mit Unterstutzung des Niedersachsischen Zahlenlottos und der DFG. * Vortrag Europaischer EntomologenkongreS Kiel, September 1982.
U.S. Copyright Clearance Center Code Statement: 0044-2240/83/9502-0196 $ 02.50/0 2. ang. Ent. 95 (1983), 196-205 0 1983 Verlag Paul Parey, Hamburg und Berlin ISSN 0044-2240 / Intercode: ZANEAE

Uber das Trockenfallen von Simuliiden-Eiern 197
The ability to survive emergence from water shown by the laboratory trials of IMHOF and SMITH (1979) with the eggs of the nearctic species Simulium verecundum Stone & Jamnback and S. vittatum Zett. under controlled conditions for eggdeposits of Boophthora erythrocephala De Geer and Simulium rostvatum Lund. Thus the eggs do not survive any length of time out of water at the beginning of embryonic development, but are relatively resistant at the eye-spot-stage. Towards the end of development they become less resistant again. The importance of the time-related conditioned reflex of the females to deposit eggs at sunset as a means of reducing the danger of desiccation of the eggs at the eye-spot-stage (e.g. Simulium nolleri Fried.) as a way of bridging unfavourable environmental conditions is here expounded. The need to improve analysis through microclimate measurement is indicated.
1 Einleitung
Kriebelmuckenweibchen suchen bestimmte Eiablageplatze auf und zeigen in Anpassung an diese Platze mehr oder minder spezifische Verhaltensweisen (DAVIES und PETERSON 1956; RUHM ab 1968; IMHOF und SMITH 1979). Eigrofle und -form, Lage der Eier auf dem Substrat, Lagerung bzw. Abstande der Eier in den Gelegen, Starke der Eigallerte, Form der haufig ausgebildeten Sammelgelege und deren Altersstruktur sowie die Auswahl einzelner Substrate (Pflanzen, Steine, Sand) und deren Position zum Wasserspiegel konnen artver- schieden sein und bei einer Art variieren. Umweltbedingungen wie u. a. Mange1 an geeignetem Substrat und dessen verschiedene Qualitat (z. B. Ober- flachenstruktur und Benetzbarkeit) konnen das VerhaIten wie die Auswahl der Eiablageplatze im Habitat, Lagerung der Eier, Form und Grofle der Gelege modifizieren (z. B. bei Wilhelmia lineata (Meigen) und Odagmia ornata (Meigen) [RUHM 1971, 19821).
In fruheren Untersuchungen wurde festgestellt, dafl frisch abgelegte Eier gegenuber Austrocknung nach sofortigem und langer anhaltendem Trocken- fallen keine Toleranz besitzen, d. h. stenohydr sind. Die Eier miissen in ein Milieu um 100 % rel. Feuchte bzw. in einen Wasserfilm abgelegt werden, um sich nach Benetzung weiterentwickeln zu konnen und nicht zu Beginn der Embryogenese zu vertrocknen bzw. zu kollabieren (RUHM ab 1968).
Je nach Flieflwassersystem und -abschnitt sowie Bindung an spezielle Eiablageplatze und deren Qualitat (z. B. terrestrische oder aquatische Pflan- Zen) sind die Eigelege der verschiedenen Kriebelmucken unterschiedlich stark durch Trockenfallen mit der Folge des Vertrocknens gefhrdet. Der Ausfall an Eigelegen durch Vertrocknen und die Starke der Eiablage konnen nach unse- ren Analysen bei Boophthora erythrocephala ohne storende Eingriffe quantita- tiv erfaflt werden (RUHM und SCHLEPPER 1979; SCHLEPDER 1981).
2 Typen der Eigelege-Gefahrdung durch Trockenfallen
Drei Typen der Gefahrdung sind grundsatzlich zu unterscheiden: Typ 1: Gelege an bzw. in der Wasserlinie der Flieflgewasser, z. B. auf Pflan-
Zen, die den Pegelschwankungen nicht folgen, sind besonders gefahr- det. Die Gefahrdung ist stark vermindert, wenn ein Angebot aquati- scher Pflanzen (z. B. flottierende Potamo eton-Arten) von den Miik- ken zur Eiablage genutzt werden kann ( B bergangstyp).
Typ 2: Eigelege, die direkt in ein Flieflgewasser unterhalb der Wasserlinie auf Pflanzen abgelegt werden, sind in Abhangigkeit von ihrer Position nur durch relativ starkes und plotzliches Absinken des Pegels kurz nach der Ablage besonders gefahrdet (vgl. Typ 1).

198 W. Ruhm
Typ 3: Gelege oder Eier, die direkt auf den Wasserspiegel oder uber einem Flieflgewasser im Fluge abgelegt werden und auf die Sohle absinken, sind im allgemeinen durch Trockenfallen nicht gefahrdet. Ihre Weiter- entwicklung wird u. a. von der Stromung, der Schleppkraft und der Beschaffenheit der Sohle (Sand, Schlamm) eines Flieflgewassers beein- flui3t. Diese Eier konnen in einen Totwasserbereich verdriftet und von Sedimenten bedeckt werden. Entsprechendes trifft fur Eier zu, die in mehrschichtigen Gelegen (z. B. Odagmia ornatu) von schlupfenden Erstlarven losgelost werden.
Bei einer Art wie beispielsweise Wilhelmia lineata, die sowohl auf Pflanzen als auch auf dem Wasserspiegel und unterhalb der Wasserlinie ablegen kann, gehoren die Gelege je nach Art der Ablage zu einem der drei genannten Typen. Hohe Anforderungen sind an die Oberlebensfahigkeit der Eier in eophemeren Fliei3gewassern oder in einzelnen Abschnitten, die langere Zeit trockenfallen, gestellt (COLBO und MOORHOUSE 1974). Sie konnen nur von Arten auf Dauer besiedelt werden, deren Eier im feuchten Sediment iiberleben konnen. Durch Suchfluge auf mehr oder minder weiten Strecken und/oder durch Verdriftung der Muckenweibchen mit Hilfe des Windes kann in diesen Flieflgewassersyste- men die Wiederbesiedlung eingeleitet werden, wie dies von einigen Arten des Simulium damnorum-Komplexes in Afrika bekannt ist.
3 Beziehungsgefiige Feuchte/Simuliidenei/Substrat/Koinzidenz beim Trockenfallen im Bereich der Wasserlinie
Die Analyse der Populationsdynamik schad- und plageerregender Arten und die Ermittlung der Ursachen der Herdbildung in einem Fliei3gewasser mussen die Umweltbeziehungen des Eistadiums der betreffenden Arten einbeziehen, weil die Abundanz der Populationen uber dieses Stadium entscheidend beein- flui3t werden kann. Im Eistadium uberwiegt nach den bisherigen Befunden der Einflui3 abiotischer Faktoren. Die Analyse von Eigelegen im Bereich der Wasserlinie fordert die Berucksichtigung folgender Fakten bzw. Zusammen- hange: a. Feuchte:
1. Dauer des Trockenfallens und der Oberflutung bzw. Befeuchtung sowie Haufigkeit des Wechsels und dessen zeitliche Verteilung (2. B. Tag/ Nacht).
2. Die jahreszeitlich modifizierten Umweltbedingungen einschliefllich Wit- terung und die dadurch bedingten Temperatur-Feuchte-Verhaltnisse.
3. Die Geschwindigkeit, mit der sich bei absinkendem Pegel in Abhangig- keit von dessen Ausmai3 das Feuchtigkeitsgefalle uber dem Wasserspiegel andert.
b. Ei bzw. Ei opulationen: 1. Zeitpun K t der Eiablage innerhalb der Vegetationsperiode (z. B. fruh-,
schliei3lic K der unterschiedlich potenten bzw. toleranten Embryogenese-
spatschwarmende Arten). 2. Potenz bzw. Toleranz der einzelnen Embryogenesephasen (vgl. 4.2). 3. Artbedin te Entwicklungsgeschwindigkeit (vgl. 4.5) und -dauer ein-
phasen sowie deren zeitlicher Anteil an der gesamten Embryogenese. 4. Individuelle und populare Variabilitat bezuglich Entwicklung bis nach
dem Schlupfen einschliei3lich umweltinduzierter Modifikationen.

Vber das Trockenfallen von Simwliiden-Eiern 199
5. Gesundheitszustand der Eier (taube bzw. nicht entwicklurigsfahige
6 . Beschaffenheit und Starke der Eigallerte, Position und Dichte der Eier in Eier).
der Gallerte. c. Substrat:
1. Beschaffenheit eines Flieflgewassers (z. B. Fehlen oder Vorhandensein einer Beschattung des Uferbereiches bzw. Spritzwasserzone).
2. Qualitat (z. B. Entfernung zum Wasserspiegel) und Quantitat des Sub- strats, einschliefllich jahreszeitlich bedingter Unterschiede bzw. Ver- schiedenheiten.
d. Koinzidenz: 1.
2.
Zeitpunkt des Zusammentreffens von Trockenfallen mit den jeweiligen Embryogenesephasen wie Umweltsituation (z. B. Pflanzenentwicklung, Wasserfiihrung) und dem popularen Anteil dieser Phasen beim Koinzi- d' ieren. Die mogliche zeitliche Beeinflussung der Dauer einzelner Phasen nach dem Koinzidieren mit dem Trockenfallen. So konnte eine Verschiebung der Koinzidenzfristen einschliefllich Veranderung des Schliipfmuste; u. a. die Gefahrdung der Eipopulationen verringern oder vergroflern.
Die komplexen Beziehungen und Wirkungen wahrend des Eistadiums mit moglichen Auswirkungen auf Abundanz und Gesamtdynamik einer Art sind nur durch Annaherung von Versuchen unter konstanten Bedingungen im Labor, u. a. zur Aufklarung von Wirkungsmechanismen und zur Bestimmung der Leistungsgrenzen, und im Freiland unter naturlichen Bedingungen, vor allem zur Erfassung der popularen Wirklichkeit aufzudecken und zu quantifi- zieren. Die mindeste Voraussetzung ist die Altersbestimmung der Gelege, die WELTON und BASS (1980) unterlieflen und daher widerspriichliche Ergebnisse erzielten. Die Exaktheit und Vergleichbarkeit der Untersuchungsergebnisse diirfte mit der morphologischen Differenzierung der Eier in Embryogenese- phasen, die mit bestimmten Stoffwechselprozessen korreliert sind, wachsen.
4 Versuche zur Toleranz gegenuber Austrocknung
IMHOF und SMITH (1979) ermittelten u. a. die Schliipfrate der Eigelege der nearktischen Simulium verecundum Stone & Jamnback und S. vittatum Zett. nach Trockenfallen im Labor unter konstanter Temperatur und Photoperiode. Angaben iiber die Feuchte werden von den Autoren nicht gemacht. Das Alter der Gelege wurde in Stunden angegeben, die Embryogenesephasen wurden nicht erfai3t. Es wurde aber gepriift, welche Phasen die Eier, die sich nicht bis zur Erstlarve entwickelten, erreicht hatten. In diesen Versuchen betrug das maximale Alter der Gelege bei einer Temperatur von 19,5 "C rund 2% Tage. Wir fuhrten noch nicht abgeschlossene, orientierende Versuche im Freiland mit Gelegen von Boophthora erythrocephala und Simulium rostratum (Lund.) (= syn. S. sublacustre Davies nach miindl. Auskunft von Frau Dr. H. ZWICK) in einer Serie vom 8. 7.-24. 7. (A) und einer weiteren Serie in der Zeit vom 31. 7.-11. 8. (B) durch. Im wesentlichen wird uber die mit den Gelegen von Boophthora erythrocephala erzielten Versuchsergebnisse berichtet. Diese Befunde werden, sofern es moglich ist, mit den Angaben von IMHOF und SMITH (1979) iiber S. verecundum verglichen.

200 W. Riihm
4.1 Material und Methode Nach Beendigung der Eiablage am Abend wurden im Freiland die gleichaltrigen Eigelege von Boophthoru erythrocephulu am Substrat individuell durch perforiertes Tesaband markiert. Von jedem der markierten, grogen Sammelgelege Hunderter von Boophthoru-Weibchen wurden einzelne Gelege sofort oder nach einer festgelegten Stundenzahl ungestorter Entwicklung (Abb. 2) in Gruppen von zirka 300-400 Eiern einer Austrocknung von max. 7 Tagen ausgesetzt. Hierzu wurden die Gelege an einem Holzstab mit dem natiirlichen Substrat 30 cm iiber dem Wasserspie- gel eines Teichabflusses iiber dem ursprunglichen Eiablageort befestigt. Die Serie A umfai3te 15, die Serie B 14 Gelege unterschiedlicher GroBe und Anzahl.
Der TeichabfluB war 100-150 cm breit und der Versuchsabschnitt von 200 m Lange, strecken- weise durch Biische und Baume beschattet. An den weniger beschatteten Abschnitten war das Fliengewasser von einem relativ dichtem Bewuchs der steilen Uferboschung eingesaumt. Der Pegel des wahrend der Versuchsdauer etwa 20 cm tiefen Gewasser mit weitgehend beruhigtem AbfluS wurde reguliert, so daS sich der Abstand zu den exponierten Gelegen nicht anderte. Im Versuchszeitraum beider Serien lag die rel. Luftfeuchte bei haufig regnerischem, zum Teil schwiilen Wetter urn 90 %. Die Temperatur erreichte max. Werte von 20-25 "C. Die Gelege lagen im Uferbereich weitgehend windgeschiitzt, sofern in dieser Zeit iiberhaupt starkere Windbewe- gungen auftraten.
Fur Kontrollversuche im Labor wurden jeweils zu gleicher Zeit gravide Weibchen abgefangen, zur Eiablage gebracht und deren Eier bei einer rel. Feuchte von 100 % und einer durchschnittli- chen Temperatur von 15 "C auf FlieBpa ier in Blockschalchen innerhalb groger Petrischalen aufbewahrt. Diese Gelege wurden fortlau/k~d kontrolliert und deren Schliipfrate ermittelt. Nach der festgelegten Expositionszeit wurden die Gelege aus dem Freiland im Labor unter dem Binokular untersucht, ob sie kollabiert waren, anschliefiend bei 100 YO rel. Feuchte auf einen Wasserfilm gelegt und die Schliipfrate bestimmt. Bei den Kontrollen im Labor und Freiland wurden auch die wichtigsten Embryogenesephasen ermittelt. Die Embryogenese war bis zum Schliipfen unter den angegebenen Freilandbedingungen im Durchschnitt nach 6 Tagen abgeschlos- sen. Die ermittelten Schliipfraten sind Durchschnittswerte, die mehrere gleichaltrige, an verschie- denen Stellen des FlieBgewassers exponierte Gelege einschliegen. Sie reprasentieren einen FlieSge- wasserabschnitt und nicht Gelege eines bestimmten Eiablageplatzes. Die Entwicklungsgeschwin- digkeit der Eigelege wurde bei 100 % rel. Feuchte und verschiedenen konstanten Temperaturen in Klimaschrinken bestimmt.
4.2 Toleranz der Embryogenesephasen
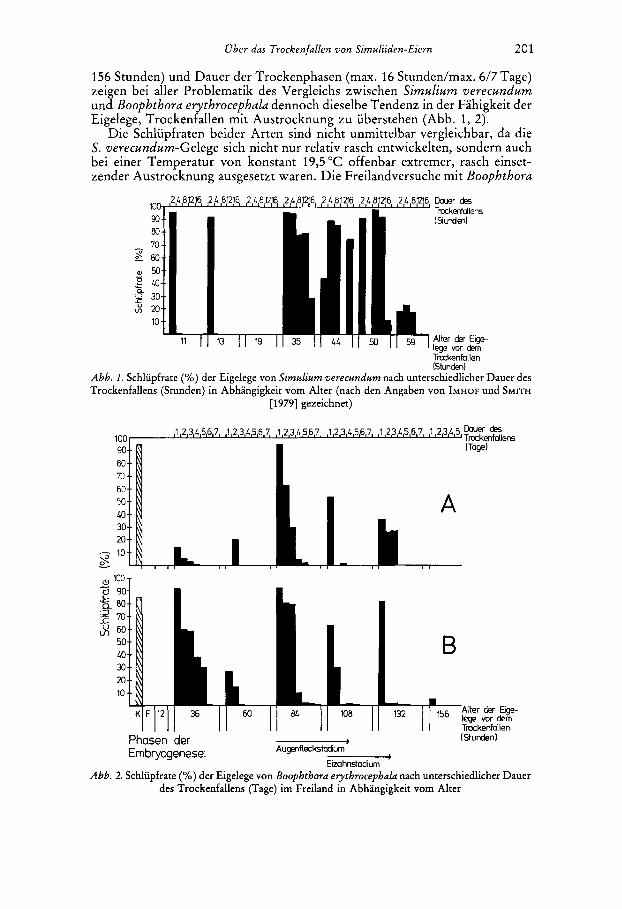
Zu Beginn der Embryogenese iiberstanden die Boophthora-Eier in den Ver- suchsserien (Abb. 2) rnit einer Ausnahme (vgl. 4.4) in einem Alter bis zu 12 Stunden ein eintagiges Trockenfallen nicht. Mit Erreichen der Augenfleck- phase in der Embryogenese (bei 84 Stunden) war die Toleranz der Gelege deutlich erhoht. Vor dem Schliipfen, nach Anlage des Eizahnes (nach 108 Stunden) nahm die Toleranz wieder deutlich ab. In beiden Serien sank, wenn auch unterschiedlich stark, in sht l ichen Embryogenesephasen mit zuneh- mender Dauer des Trockenfallens die Schliipfrate. Insgesamt waren die Schliipfraten beider Serien unterschiedlich hoch. Auffallig sind die Unter- schiede nach 36 Stunden zwischen der Serie A mit einer niedrigen und der Serie B mit einer relativ hohen Schliipfrate. Im Augenfleckstadium wurde in der Serie A nach 3 Tagen Austrocknung eine Schliipfrate von 30 % und in der Serie B von 80 % festgestellt. Auffallige Unterschiede konnten am Ende der Embryogenese zwischen den 132 Stunden alten Gelegen der Serie A und B beziiglich der Schliipfraten wie der Starke ihrer Verminderung nach einem Tag Trockenfallen beobachtet werden (Abb. 2).
4.3 Vergleich von Labor- (Imhof/Smith 1979) und Freilandversuchen
Beide an verschiedenen Simuliidenarten verschieden angelegten Versuche (Labor: konstante Bedingungen, ausgenommen Feuchte; Freiland: variable Bedingungen) mit unterschiedlichem Zeitablauf (max. 59 Stundedmax. 132/
Vber das Trockenfallen won Simuliiden-Eiern 201
90.. 80- 70 60-
aJ 50-
2 30-
- ... p Lo--
x 20-
Q
10
156 Stunden) und Dauer der Trockenphasen (max. 16 Stunden/max. 6/7 Tage) zeigen bei aller Problematik des Vergleichs zwischen Simulium verecundum und Boophthora erythrocephala dennoch dieselbe Tendenz in der Fahigkeit der Eigelege, Trockenfallen mit Austrocknung zu uberstehen (Abb. 1, 2).
Die Schlupfraten beider Arten sind nicht unmittelbar vergleichbar, da die S. verecundum-Gelege sich nicht nur relativ rasch entwickelten, sondern auch bei einer Temperatur von konstant 19,5 "C offenbar extremer, rasch einset- zender Austrocknung ausgesetzt waren. Die Freilandversuche mit Boophthora
B.Undenl
-.
-1
Thenfallen (Stun den1
Abb. 1 . Schluufrate I % ) der Eigelege von Simulium verecundum nach unterschiedlicher Dauer des . , Y Y
Trockenfallens (Stunden) in Abhangigkeit vorn Alter (nach den Angaben von IMHOF und SMITH [1979] gezeichnet)
A
B
K F 12 36 60 ,32 ,56 lege vor dem Alter dw Eige-
Trockenfallen - (Stunden) Augenfleckstcdiurn
Abb. 2. Schliipfrate (YO) der Eigelege von Boophthora erythrocephula nach unterschiedlicher Dauer des Trockenfallens (Tage) im Freiland in Abhangigkeit vom Alter
Phasen der Embrycqenese: -
Eizahnstadium

202 W. Riihrn
6 7
L -
2 -
2 -
I
= Wasserlinie
Abb. 3. Schliipfrate (%) der Eigelege von Boophthora erythrocephakz in Abhangigkeit von der Entfernung
I I I I I I I I
202 W. Riihrn
evythvocephala spiegeln die ,,populare Wirklichkeit" und damit die Potenz der Eier bzw. Gelege unter naturlichen Bedingungen wider. So schlupften nach 4-5 Tagen Trockenfallen aus unterschiedlich alten Gelegen Erstlarven. Die Gelege von S. vevecundum durften unter naturlichen Bedingungen sehr vie1 langer eine Austrocknung uberstehen, als dies in den Versuchen mit max. 16 Stunden der Fall war. In den Versuchen von IMHOF und SMITH (1979) (Abb. I) wird die Abnahme der Schlupfrate mit zunehmender Dauer des Trockenfallens kaum sichtbar, was auf den ,,Alles-oder-Nichts-Versuch" bezuglich der Aus- trocknung zuruckzufuhren sein durfte. Sowohl in den Versuchen mit S. veve- cundum als auch mit B. erythvocephala fallen die grogen Schwankungen in der Schlupfrate auf, die eine statistische Auswertung nicht ermoglichten. In den Versuchen mit B. evythvocephala durfte dies auf die im Versuchszeitraum jeweils unterschiedlichen und wechselnden Feuchtebedingungen im mikrokli- matischen Bereich der verschiedenen Gelege des betreffenden Flieflgewassers zuruckzufuhren sein, die in ihrer Dynamik, insbesondere das Feuchtigkeits- gefalle oberhalb der Wasserlinie, noch nicht genau genug in groi3eren Ver- suchsanordnungen gemessen wurden. Wie rasch oberhalb der Wasserlinie in kurzer Entfernung die Schliipfrate abnehmen kann, zeigt ein Versuch mit unterschiedlicher Position der Eier groger zusammenhangender Sammelgelege zur Wasserlinie, in deren unmittelbarer Nahe die reI. Feuchte um 100% schwankte, im Nahbereich 80 % betrug und in groi3erer Entfernung unter diesem Wert lag. In einem Bereich von 4-6 cm oberhalb der Wasserlinie schliipften nur noch aus 3,5 70 der Boophthora-Gelege Erstlarven aus (Abb. 3). Ein Vergleich dieses Befundes mit den Serien A und B veranschaulicht, wie relativ feucht es wahrend des Versuchszeitraumes noch 30 cm oberhalb der Wasserlinie gewesen sein mufl, um die ermittelten Schlupfraten zu erreichen (Abb. 2).
4.4 Die Augenfleckphase und ihre okologische Bedeutung
Durch den bei zahlreichen Arten von einem bestimmten Helligkeitsabfall induzierten Anflug zur Eiablage (REUTER und RUHM 1976) und daher deren weitgehende Bindung an die Abendstunden, d. h. diese zeitliche Einklinkung, wird die erste Phase starker Gefahrdung durch die Ablage in Stunden relativ hoher Feuchte verkurzt bzw. diese herabgesetzt. In unmittelbarer Nahe des Wasserspiegels durften bei geringem Pegelfall bzw. geringen Schwankungen der Eigelege bessere Oberlebensmoglichkeiten besitzen, als es in dem orientie- renden Freilandversuch der Fall war, in dem ein plotzliches Absinken des Pegels von 30 cm simuliert wurde, wie er unter natiirlichen Bedingungen nicht

Vber das Trockenfallen won Simuliiden-Eiern 203
die Regel ist. In einem der Versuche mit frischen Gelegen bei einer Exposition von einem Tag betrug die Schliipfrate 72,3 %. Schon diese orientierenden Versuche lassen die Bedeutung der zeitlichen Bindung der Eiablage fur das Oberleben von Embryogenesephasen mit hohen Feuchtigkeitsanspriichen erkennen. Wir diirfen vermuten, dafi in einem reich strukturierten Fliefigewas- ser mit vielfaltigem Pflanzenbewuchs im Uferbereich die Gefahrdung der Gelege wegen gunstigerer Feuchtebedingungen geringer ist als in einem Fliefi- gewasser mit verarmter Struktur (z. B. Ufer ohne Beschattung). Sirnufium nofleri, die in der Nahe ihres Aufenthaltes Oberlaufe bzw. Wehre zur Eiablage aufsucht und direkt am Oberlauf oder unter ihn ablegt (RUHM 1975), ist nicht so eng an die Abendstunden bei der Eiablage gebunden. Die Augenfleckphase scheint auch bei anderen Simuliidenarten relativ tolerant bzw. widerstands- fahig gegeniiber ungiinstigen Milieubedingungen, insbesondere abnehmender Feuchte, zu sein. In unserer Fauna uberdauern in dieser Phase nur die diapausierenden Eier von S. nofleri monatelanges Trockenfallen (max. 8 Monate) und Frost. Die Gelege von S. rostratum ruhen uber mehrere Monate im Sediment wie die Eier von S. morsitans (Edwards) nach meinen Beobach- tungen mehrere Monate im feuchten Sediment eines trockengelegten Abflusses einer Teichwirtschaft unbeschadet uberleben konnten. Die Augenfleckphase ist gleichfalls bei den Eiern anderer Insektenarten gegenuber verschiedenen ungiinstigen Umweltbedingungen als widerstandsfahig bekannt. Nach der Augenfleckphase, d. h. nach der Ausbildung des Eizahnes, ist trotz eines hohen Prozentsatzes vollentwickelter Embryonen bzw. Larven die Verminde- rung der Schliipfrate nach Trockenfallen relativ groi3. Nach meinen Feststel- lungen durchbricht zwar ein Teil der Larven das Chorion, kann sich aber nicht vollig befreien (bis zu 80 Yo) und stirbt ab. Kiinftig mufl u. a. in diesem Zusammenhang gepruft werden: a. ob dies auf mechanischen Ursachen beruht (z. B. Verfestigung des Cho-
rions beim Austrocknen), b. ob der Wasserhaushalt der Embryonen bzw. der Erstlarven gestort ist, c. ob die Erstlarven evtl. wegen verzogertem Schlupfen und begrenztem
d. ob die in a-c genannten Fakten zusammenwirken. Es ist sicher, dai3 die Wirkungsmechanismen, die eine erhohte Mortalitat im Eistadium bewirken, in der Anfang- und Endphase verschieden sind. Ohne Ruckkehr des Pegels - dies wurde bereits fruher erortert - sind die Erstlarven bei Fehlen eines Spritzwasserbereiches und eines Flussigkeitsfilmes auf dem Substrat nicht in der Lage, ein Fliefigewasser zu erreichen. Diese Larven sterben ab. Der Anteil abgestorbener Erstlarven ist aus methodischen Grunden quantitativ schwer bzw. nicht zu erfassen. Dies trifft auch fur die nicht entwicklungsfiihigen Eier zu. Sie sind kaum von den unmittelbar durch Vertrocknen abgestorbenen Eier zu unterscheiden.
Hungervermogen geschwacht sind,
4.5 Die Entwicklungsgeschwindigkeit der Eier und deren Gefihrdung durch Trockenfallen
Die arttypische wie umweltinduzierte Entwicklungsgeschwindigkeit ist fur die Oberbruckung ungiinstiger Milieubedingungen sowie zur Minderung der Gefahrdung von Bedeutung. Boophthora erythrocephala, Odagmia ornata und Simulium rostratum konnen in einem Flieflwasserabschnitt nebeneinander vorkommen. Die meist einschichtigen Gelege von B. erythrocephala und S. rostratum, die im Sommer oft dicht nebeneinander liegen, sind sehr ahnlichen

204 W. Riihm
bis identischen Bedingungen der Feuchte wie der Austrocknung auf ein- und demselben Substrat ausgesetzt. Wie die Versuche zeigen (Tab.), erreicht das Eistadium von B. erythrocephulu die Phase erhohter Toleranz schneller als das von S. rostrutum. Eine zeitlich synchrone Entwicklung gleichaltriger Popula- tionen ist in diesem Zusammenhang gunstiger als eine zeitlich weite Streuung. Moglicherweise sind unterschiedliche Dichten ein- um denselben Abschnitt besiedelnder Simuliidenarten auch eine Folge derartiger Wechselwirkungen.
Entwicklungsgeschwindigkeit bis zum Erreichen der rel. roleranten Augenfleckphase (konstant 100 YO rel. Feuchte, Photoperiode: 16/8)
A n Boophthora eyythrocephnln Odagmia ornntd Stmulzum rostmtam
Temperatur (“C) 10 20 10 20 10 20 Entwicklung (h) 100 60 180 84 230 84
4.6 Verdriftung der Eier und Gefahrdung im Totwasserbereich
Gelangen die Eier durch Verdriftung in Totwasserbereiche konnten sie in ihrer Entwicklung gestort sein bzw. absterben. In einem Versuch wurde die Schlupfrate von Boophthoru-Eiern in einer 10 cm Wassersaule ermittelt (n = 5), die wahrend der Beobachtungsdauer von 16 Tagen nicht erneuert wurde. Folgende durchschnittliche Schlupfraten wurden ermittelt :
Eier frisch 6 Yo 37 Yo Eier 2 Tage alt
Eier mit Augenfleck 74 Yo Von den frischen Eiern, die nicht schlupften, hatten nur wenige die Augen- fleckphase erreicht bzw. standen kurz vor dem Schlupfen. Bemerkenswert ist die 6%ige Schlupfrate frischer Eier, deren Zustandekommen noch nicht erklart werden kann. Die Mortalitat wurde bei einem Teil der Eier durch Pilze verursacht, die mit den Hyphen in die Eier gedrungen waren. Die Entwick- lungsdauer war gegenuber den unter normalen Bedingungen (6 Tage) im Bereich des Wasserspiegels befindlichen Eiern um mehrere Tage verlangert. Wie in den vorangehenden Versuchen erwies sich auch hier das Augenfleck- stadium als relativ widerstandsfahig.
5 Ausblick
Zur Verbesserung der Analyse der Abundanzdynamik im Eistadium ein- schlieillich der Vergleichbarkeit und statistischen Absicherung vor allem beziiglich des Einflusses des Feuchtefaktors ist es notwendig: 1. die Grenzen des stenovalenten Faktors Feuchte fur die einzelnen Arten zu
2. mikroklimatische Messungen zur Erfassung unterschiedlicher Feuchtig-
3. den Einflufi der zeitlichen Veranderung dieses Gefalles zu erfassen, 4. den Beginn der Embryonalentwicklung bis zum Augenfleckstatium mor-
phologisch genauer zu differenzieren und den Zeitraster (in Stunden) noch enger zu legen.
Unter diesen Voraussetzungen mui3te es moglich sein, vorhandene, vermutlich geringe, aber fur die Dynamik bedeutsame Toleranzunterschiede gegenuber Austrocknung zwischen verschiedenen Arten festzustellen.
bestimmen,
keitsgefdle im Bereich einzelner Gelege durchzufuhren,

Uber dus Trockenfallen von Simuliiden-Eiern 205
Danksagung Fur die wissenschaftliche Mitarbeit im Freiland und Labor danke ich Herrn U. REUTER (Hanno- ver). Fur technische Unterstiitzung danke ich meiner Frau VERENA.
Zusammenfassung Die Weiterentwicklung der Eigelege bis zum Schliipfen der Erstlarven ist je nach Art der Eiablage beim Trockenfallen unterschiedlich stark gefahrdet. Die Faktoren, die beim Trockenfallen der Gelege, die auf ein Substrat im Bereich des Wassers iegels abgelegt werden, wirksam sind und die Entwicklung beeinflussen, werden nach Feuchtelktor, Embryogenesephasen, Substrat sowie raumlich-zeitlicher Koinzidenz aufgeschlusselt. Wie in den Laborversuchen von IMHOF und SMITH (1979) mit Gelegen der nearktischen Simulium verecundum und S . vittutum unter konstan- ten Bedingungen konnte auch in Freilandversuchen unter natiirlichen Verhaltnissen mit Gelegen von Boo hthoru erythrocephala und Simufium rostrutum dieselbe Tendenz in der Fahigkeir, Trockentllen zu uberstehen, nachgewiesen werden. Zu Beginn der Embryogenese iiberstehen die Eier bei plotzlichem Trockenfallen eine langere Austrocknung. In der Augenfleckphase sind sie relativ widerstandsfahig, gegen Ende der Entwicklung nimmt ihre Widerstandsfahigkeit wieder ab. Die Bedeutung der zeitlichen Einklinkung der Eiablage der Weibchen in die Zeit vor Sonnenunter- gang, durch die vor allem die Gefahrdung der Eier durch Austrocknung gemindert wird, wird ebenso wie die der widerstandsfahigen Augenfleckphase diapausierender Eier (z. B. Simulium nollerz) zur Uberbruckung besonders ungunstiger Milieubedingungen hervorgehoben. Auf die Notwendigkeit u. a. durch mikroklimatische Messungen die Analyse noch zu verbessern, wird hingewiesen.
Literatur
COLBO, M. H.; MOORHOUSE, D. E., 1974: The survival of the eggs of Austrosimuliumpestilens Mack. & Mack. (Diptera, Simuliidae). Bull. ent. Res. 64, 629-632.
DAVIES, D. M.; PETERSON, B. V., 1956: Observations on the mating feeding, ovarian development, and oviposition of adult black flies (Simuliidae, Diptera). Can. J. Zool. 34, 615-655.
DAVIES, CH. C., 1971 : A study of the hatching process in aquatic invertebrates. XXV. Hatching in the blackfly, Simulium (probably venustum) (Diptera, Simuliidae). Can. J. Zool. 49,333-336.
IMHOF, J. E.; SMITH, ST. M., 1979: Oviposition bahaviour, eggmasses and hatching response of the eggs of five Nearctic species of Simulium (Diptera: Simuliidae). Bull. ent. Res. 69, 405-425.
PETERSON, D. G.; WOLFE, L. S., 1960: Diurnal behaviour and biting habits of black flies in the forest of Quebec. Can. J. Zool. 38, 489-498.
REUTER, U.; RUHM, W., 1976: Uber die weitere Verteilun der anfliegenden Weibchen von Boophthoru erythrocephulu De Geer und Simulium suhucustre Davies bei der Eiablage (Simuliidae, Diptera). Z. ang. Zool. 63, 385-391.
RUHM, W., 1968: Zur Autokologie einiger Simuliidenarten. Z. ang. Ent. 61, 466-471. - 1969: Zur Populationsdynamik der Kriebelmucken, insbesondere von Boophthoru erythroce-
phukz De Geer und des Odugmiu ornutu-Komplexes. Z. ang. Ent. 63,212-227. - 1971 : Eiablagen einiger Simuliidenarten. Ang. Parasitol. 22, 68-78. - 1975: Freilandbeobachtungen zum Funktionskreis der Eiablage verschiedener Simuliidenarten
unter besonderer Berucksichtigung von Simulium urgyreutum Meig. (Simuliidae, Diptera). Z. ang. Ent. 78, 321-334.
der mammalophilen Odugmiu ornutu im Bereich der Rur - 1982: Zur Massenvermehrun (Simuliidae, Diptera). 11. J aB restagung des Ges. f. Okologie, Mainz, 28. 9.-3. 10. 81 (i. Druck).
RUHM, W.; SCHLEPPER, R., 1979: Versuche zur quantitativen Erfassung der Eiablageintensitat von Simuliiden (Dipt.). Z. ang. Ent. 88, 204-216.
SCHLEPPER, R., 1981 : Die Eiablageintensitat von Boophthoru erythrocephulu De Geer (Diptera, Simuliidae), ein Mai3 zur Erfassung ihrer Abundanzdynamik im kleinlokalen Bereich sowie innerhalb eines Fliefigewissers. Mitt. dtsch. Ges. allg. angew. Ent. 3, 288-290.
WELTON, J. S.; BASS, A. B., 1980: Quantitative studies on the eggs of Simulium ornuturn Meigen und Simulium (Wilbelmiu) equinum L. in a chalk stream in southern England. Ecol. Ent. 5, 87-96.
Anschrift des Verfussers: Prof. Dr. W. RUHM, Zoologisches Institut und Zoologisches Museum der Universitat Hamburg, Martin-Luther-King-Platz 3, D-2000 Ham- burg 13





























