Embed Size (px)
Citation preview

Planta, Bd. 51, S. 74--83 (1958)
Aus dem Botanischen Institut der Universiti~t Tiibingen
UBEI~ DEN PRIMARVORGANG BEI DER POLARISIERENDEN WIRKUNG DES LICHTES
AUF KEIMENDE EQUISETUM-SPOI~EN*
Von
WOLFGA~G IIAUPT
Mit 11 Textabbildungen
(Eingegangen am 19. ~ovember 1957)
~ber die Polarisierung der keimenden Equisetumspore oder des be- fruchteten Fucaceen-Eies durch Licht liegen zahlreiche Untersuehungen vor (vgl. insbes, die Literaturangaben bei MOSEBAC~ 1943 und HAVPT 1957). Doch ist noch wenig bekannt fiber die primgre Wirkung des Lichtes. Wir wissen, dab bei Equisetum die maBgebliche Lichtabsorption im Cytoplasm~ erfolgt (MOSE~ACI~), dag die Induktionswirkung des Lichtes unabhs yon der Temperatur w~hrend der Belichtung ist, dab das Reizmengengesetz in gewissen Grenzen gfiltig ist, dab bei ge- n/igend hoher Intensit/~t bereits sehr kurze Beliehtungszeiten (Elek- tronenblitz) erheblieh wirksam sein k6nnen (HArlOT), und dab nur Wellen]/ingen unter 500 m# Ifir die Induktion ill Frage kommen mit einem Maximum bei 450 m# (MOSEBACI~, HAUPT; der Bereich unter 300 m# wurde bei Equisetum noch nicht geprfift, ist aber bei Fucus be- sollders wirksam). In der vorliegenden Untersuehung sollen diesen Tat- s~ehen nun einige weitere Ergebnisse zur Char~kterisierung der Prim//r- reM~tion hinzugeffigt werden.
Allgemeine Methodik Die Aussaat und Anzucht der Sporen erfolgte naeh der friiher besehriebenen
Methode (ttAu1,T 1957). Meist wurde mit der Hg-Lampe beliehtet, in vereinzelten Fallen mit der Niedervolt-Lampe bei 6,0 A. Da in vielen Versuchen der UV-Anteil der Hg-Lampe bestm5glich ausgenutzt werden sollte, mul3te in verst~rktem MaBe auf die Aussaat au/start zwischen Agar zurtickgegriffen werden (in den Abbildungs- legenden bezeiehnet: ,,ohne Oeckagar"). Das Lieht der Hg-Lampe wurde durch Verwendung yon 2 Quarzlinsen konzentriert; da in diesen Versuehen nicht mit Interferenzfiltern gearbeitet wurde und die zu bestrahlende ~'li~che klein war, konnte im Interesse mSglichs~er Intensit~tssteigerung eine leiehte Konvergenz des Strahlenganges in Kauf genommen werden. Der Zeitpunkt der Beliehtung naeh der Aussaat wurde ira allgemeinen so gewghlt, daft er m5gliehst in das Maximum der sensiblen Phase fiel. Dieses lag bei den versehiedenen Arten etwas verschieden und hing offensichtlich yon den Bedingungen ab, unter denen die Sporen zur Reife kamen; dariiber soll spi~ter an anderer Stelle berichtet werden.
* Herrn Prof. Dr. Dr. h. e. WIL~.LM I~V~LAND zum 80. Gebur~stag gewidmet.

Prim~rvorgang der Polarit/itsinduktion bei Equisetum-Sporen 75
Sofern Auss~a~en ohne Deck~gar bei niederen Temperaturen betich~et werden sollten, konnte naturgem/~8 der TemPeraturangleieh nieht, wie fffiher beschrieben, im W~sserbad der entsprechenden Temperatur effolgen; in diesem Falle wurden die Glasplatten in der entsprechend temperierten Dunkelkammer (Klimakammer) so lange auf einen glatten Eisblock gelegt (meist etw~ 1 Stunde), bis sieh auf der Oberflaehe des Agars die ersten Anzeichen leic.hter Vereisung zeigten;~ dies wurde yon den Sporen ohne weiteres vertragen.
Auch die Auswertung der Keimungsrichtungen sowie die Berechnung des In- duktionsvektors ,,V" ~us diesen (Prozentsatz der yon der Induktion erfal]ten Sporen im Gegensatz zu den zufallsm/~Big verteilten) effolgte wie in der vorher- gehenden Un~ersuchung. Doeh wurde in vielen F/~llen eine Zwischenauswertung vorgenommen, wenn etwa die HMfte der vorgesehenen Sporenzahl ~usgez/~hlt war; die Abweichung des so bereehneten Induktionsvek~ors veto endgfiltigen gestattet eine t)berprfifung der Genauigkeit der Werte. In Ubereinstimmung mit der ereignis- statistisehen Betrachtung des Induktionsvektors (HAvP~ 1957) betrugen die Diffe- renzen zwisehen End- und Zwischen~uswertung nur in seltenen F/~llen 10 %, meist nieht fiber 5%.
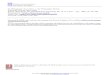
Experimenteller Teil 1. Die S~ittigung der Liehtreaktion. Ein Ansatzpunkt for die weitere
Analyse des Prim/~rvorganges ergab sieh dureh die Feststellung, dab bei starken kurzen Belichtnngen die begrenzende Reaktion sehnell abge- s/ittigt werden kann (HAuPT 1957); wird ngmlieh die Belichtung tiber ein Optimum hinaus gesteigert, so wird die polarits Wir- kung des Liehtes (statistisch gesehen) wieder geringer, verursacht da- dutch, d~B nun aueh auf der SChattenseite der Spore die Reaktion in zu- nehmendem AusmaB erfolgt, w/~hrend auf der Liehtseite keine weitere Steigerung mSglich ist. Dadurch muB der Gradient, der durch die un- gleiehe Beliehtung in der Spore entsteht, naturgem/~l] geringer werden. Abb. 1 zeigt eine solche Wirkungskurve fiir Equ. arvense. Es ist nun zu- n~chst yon Interesse, wie lange dieser S~ttigungszustand anhs bzw. unter welehen Bedingungen die Ursprtingliehe Liehtempfindliehkeit wiederhergestellt wird. Zu diesem Zweck wurden alle Sporen bis zurn Kurvenminimum beliehtet, also 25 see (Punkt B in Abb. 1), so dab die Lichtreaktion sowohl auf der Lichtseite ale auch auf der Sehattenseite weitgehend ges~ttigt ist; eine sofort ansehlie~ende Beliehtung yon 2 (oder 5) sec, die far sich allein maximal pol~risierende Wirkung her- vorrufen wiirde (Abb. 1, Punkt A), ist unter diesen Umst~nden praktisch ohne Wirkung. Wird nun zwischen die erste Beliehtung yon 25 sec und die zweite yon 2- -5 sec eine hinreiehend lange Dunkelphase ein- geschoben, so kann die zweite Belichtung wieder voile Wirkung ent- falten, die Lichtempfindlichkeit der Spore ist offensichtlieh wieder- hergestellt (nachgewiesen far die Lichtseite der Spore, die bei der Karze dieser Beliehtung praktissh allein reagiert). Diese Wiederherstellung der Empfindlichkeit i~Bt sich in ihrem zeitliehen Verlauf quantitati-v ver- folgen, wenn die Zeitspanne der Dunkelphase (die ,,Erholungszeit")
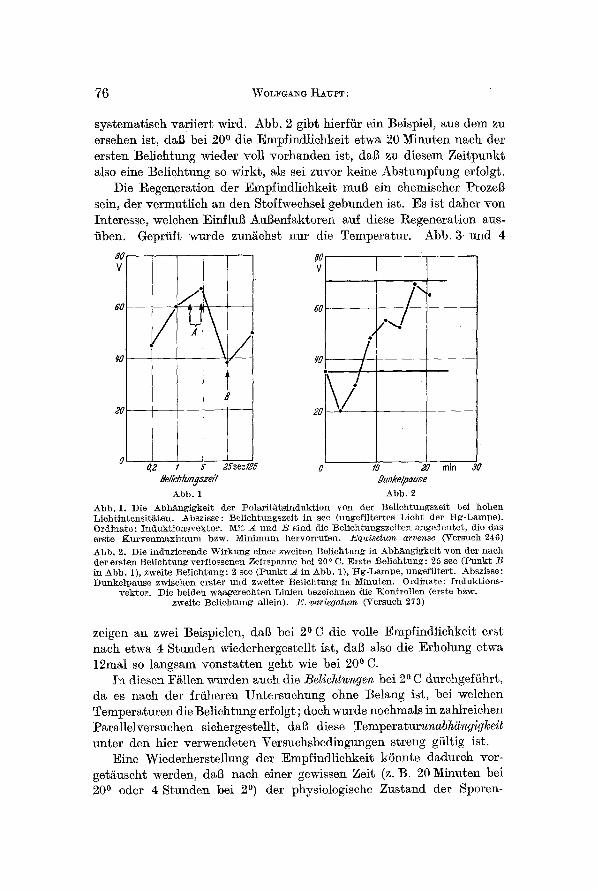
systematisch variiert wird. Abb. 2 gibt hierfiir ein Beispiel, aus dem zu ersehen ist, dab bei 200 die Empfindlichkeit etwa 20 Minuten n~ch der ersten Belichtung wieder roll vorhanden ist, dai~ zu diesem Zeitpunkt also eine Belichtung so wirkt, ~ls sei zuvor keine Abstumpfung erfolgt.
Die l~egeneration der Empfindlichkeit mu2 ein chemischer Proze[~ sein, der vermu~lich an den Stoffwechsel gebnnden ist. Es ist daher yon Interesse, wetchen Ein~luii Aui~enf~ktoren ~ f diese l~egener~tion aus- fiben. Gepriift wurde zunachst nur die Temperatur. Abb. 3 nnd 4
V
qg
20
I i/? lz g
J 0,2 / 5 2gsecf25
deh'ch/un]:~ze//
2Lbb. 1
v
6# , , ,~ . / \ J
2#
0 10 20 rain 30 #unkelpcruse
~_bb. 2
Abb, 1. Die Abh~n~:igkeit tier Polar i t~ ts i~dukt ion yon d er Belichtungszei t bei h o h e n Lich t in tens i ta ten . Abszisse: Bel ichtungszei t in sea (~mgefiltertes Licht der H g - L a m p e ) . Ordina te : Induk t io~svek to r . lV~t A u n d B si~d die Belichtu~gszeitez~ angedente t , die das erste K u r v e n m a x i m ~ m bzw. M i n i m u m hervorrufen . Equ@e~um arvense, (Versuch 246)
Abb, 2. Die i~d~zierende W i rkung einer zweiten Be]icht~mg in Abh~ngigkei t won der a a c h tier ers ten Bel ichtung verf lossenen Zei tspannc bei 200 C. :Erste Bel ichtung: 25 sec ( l~ B in Abb. 1), zweite Bel icht~ng: 2 sec (Punk t A in Abb. 1), I t g -L~mpe , nngefi l ter t . Abszisse: Du~kelpause zwischen erster und zweiter Bel ichtung i a Minuten. Ordinate : Indukt ions-
vek tor . Die beiden wa~gerech tea Limie~ bezeich~en die Koa t ro l l en (erste bzw. zweite Bel ichtung aUein). E. variegatum (Versuch 273)
zeigen ~n zwei Beispielen, dub bei 20 C die volle Empfind]ichkeit erst nach etwa 4 Stunden wiederhergestellt ist, dab ~lso die Erholung etwa 12mal so langsam vonstatten geht wie bei 200 C.
In diesen Fi~lle~ wurden ~neh die Belichtungen bei 20 C durchgefiihrt, da es ngch der Irfiheren Untersuchung ohne ]~e]ang ist, bei welchen Temperaturen die Belichtung erfo]gt; doch wurde nochmals in zahlreichen l%rallelversuchen sichergestellt, dab diese Temperaturunabhgngigkeit unter den hier verwendeten Versuchsbedingungen streng giiItig ist.
Eine Wiederherstellung der Empfindlichkeit kSnnte dadurch vor- getguscht werden, dag nach einer gewissen Zeit (z. B. 20 Minuten bei 20 ~ oder 4 Stunden bei 2 ~ der physiologische Zustand der Sporen-
Prim~rvorgang der Polarit/~tsinduktion bei Equisetum-Sporen 77
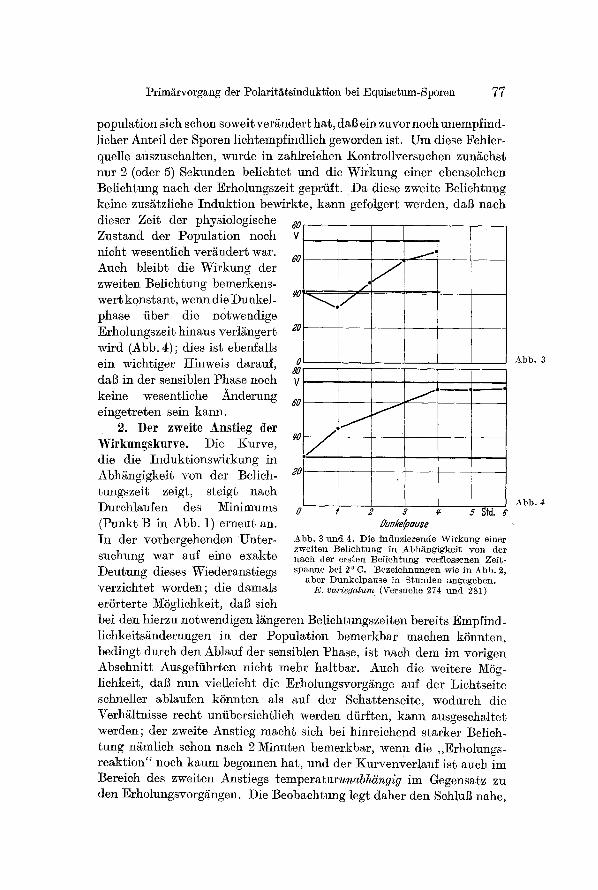
population sieh sehon soweit ver~ndert hat, dal] ein zuvor noeh unempfind- licher Anteil der Sporen lichtempfindlich geworden ist. Um diese Fehler- quelle auszusehalten, wurde in zahlreiehen Kontro]lversuchen zungehst nnr 2 (oder 5) Sekunden belichtet und die Wirkung einer ebensolchen Belichtung nach der Erholungszeit geprfift. Da diese zweite Beliehtung keine zus~tzliche Induktion bewirkte, kann gefolgert werden, dai3 nach dieser Zeit der physiologisehe ~o Zustand der Population noch v nicht wesentlich vergndert war.
6 0 - - Auch bleibt die Wirkung der zweiten Belichtung bemerkens- wert k0nstant, wenn die Dunkel- phase fiber die n0twendige Erholungszeit hinaus verlgngert ;0 - - wird (Abb. 4); dies ist ebenfalls ein wiehtiger ginweis darauf, 0
8O dal] in der sensiblen Phase noeh V keine wesentliche )kndernng ~0 eingetreten sein kann.
2. Der zweite Anstieg der Wirkungskurve. Die Xurve, die die Induktionswirkung in Abh/~ngigkeit ~r der Belieh- 2 o - - tungszeit zeigt, steigt nach Dnrehlaufen des Minimums o (Punkt B in Abb. 1) erneut an. In der vorhergehenden Unter- suchung war auf eine exakte Deutung dieses Wiederanstiegs verziehtet worden; die damals erSrterte MSgliehkeit, da6 sich
Jq j ,
J l
/ J
j ,
2 3 g 5 td. 6 Dunke/pause
Abb . 3 n n d 4, Die incluzierencle ~Virkung einer zwei ten Be l i ch tung in Abh/~ngigkei t y o n der n a c h der e r s t en Be l i ch tung verf lossenen Zei t- s p a n n e bei 20 C. Beze ichnungen wie in Abb . 2,
a b e t D u n k e l p a u s e i a S t n n d e n angegeben . E. variegatum (Versuche 274 uncl 281)
bei den hierzu notwendigen 1/~ngeren Belichtungszeiten bereits Empfind- lichkeits/~nderungen in der Population bemerkbar maehen kSnnten, bedingt dutch den Ablauf der sensiblen Phase, ist nach dem im vorigen Absehnitt Ausgeffihrten nicht mehr haltbar. Auch die weitere MSg- liehkeit, dab nnn vielleicht die Erholungsvorg/~nge auf der Lichtseite schneller ablanfen kSnnten als auf der Schattenseite, wodureh die Verh/~ltnisse recht unfibersichtlich werden diirften, kann ausgeschaltet werden; der zweite Anstieg macht sich bei hinreichend starker Belieh- tung n~mlich schon nach 2 Minuten bemerkbar, wenn die ,,Erholungs- reaktion" noch kanm begonnen hat, and der Kurvenverlauf ist auch im Bereieh des zweiten Anstiegs temperaturunabhiingig im Gegensatz zu den Erholungsvorg/~ngen. Die Beohachtung legt daher den SehluB nahe,
Abb . 3
A b b . 4
78 Wo~A~o HiveT:
dab bei Einstrahlung groBer Energien ein photochemiseher Prim~r- prozeB stattfindet, der sieh yon demjenigen nnterseheidet, der far den ersten Knrvenanstieg verantwortlieh ist. Es erseheint somit lohnend, nach Untersehieden in beiden Prozessen zu suchen.
Ein eindeutiger Nachweis far zwei verschiedene photochemische Primi~rprozesse w~re erbracht, wenn far den ersten and den zweiten Knrvenanstieg versehiedene Aktionsspektren gefunden warden. Da jedoch far den zweiten Anstieg sehr hohe Energien innerhalb nicht zn langer Zeit eingestrahlt werden mfissen, sind der Untersuchung hier zn- ngehst recht enge methodische Grenzen gesetzt. Start eines vollsti~n- 100 . . . . . . . . . . ~_.. digen Aktionsspektrnms 80 % " ~ ~ I . . . . . '-" ~&3 konnte nut gepraft wer-
�9 -" den, ob das Verh~ltnis derWirksamkeit willkiir-
co , / -- lich abgegrenzter Spek- r tralbereiehe far beide
Kurvenanstiege gleich 20 o , / / : : ~ . . ~ . ~ oder verschieden ist. AlS
--. brauehbar erwies sich see coo r :r~176 die Verwendung folgen-
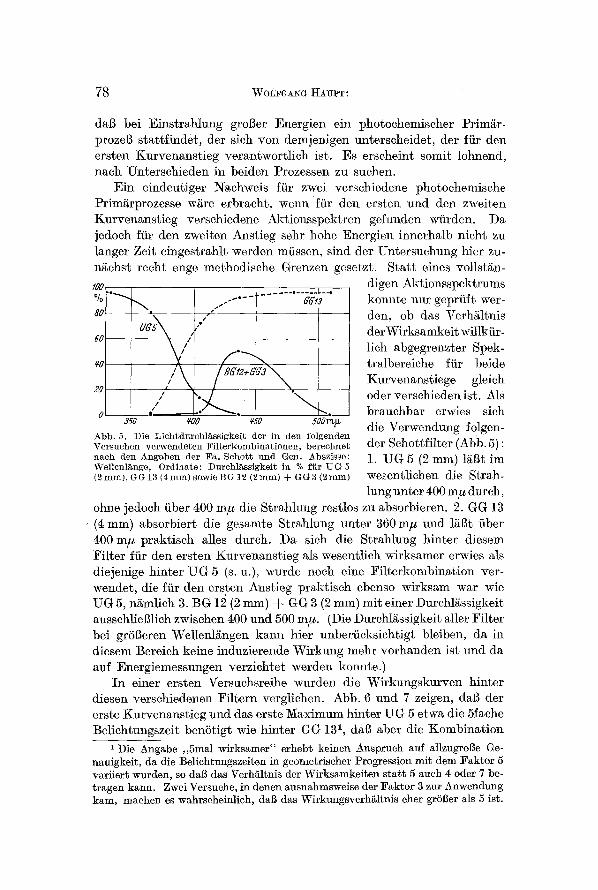
Abb. 5. Die Liehtdurehl~ssig'keit der in den folger~den Versuehen verwendeten Filterkombinationen, bereehnet der Sehottfilter (Abb. 5) : nach den Ang'aben der Fa. Sehott uxld Gem Abszisse: 1. UG 5 (2 mm) l/igt im Wellenlinge, Ordinate: Dnrehli~ssigkeit in % fiir UG 5 (2mm),GG13(4mm)so.wieBG12(2mm)+GG3(2mm) wesentlichen die Strah-
lung nnter 400 m# dutch, ohne jedoch fiber 400 m# die Strahlung restlos zu absorbieren. 2. GG 13 (4 ram) absorbiert die gesamte Strahlung nnter 360 m# and lggt fiber 400 m# 9raktisch alles dureh. Da sieh die Strahlnng hinter diesem Filter far den ersten Kurvenanstieg als wesentlich wirksamer erwies als diejenige hinter UG 5 (s. u.), wurde noeh eine ~ilterkombination ver- wendet, die far den ersten Anstieg praktisch ebenso wirksam war wie UG 5, ngmlich 3. BG 12 (2 ram) -~ GG 3 (2 ram) mit einer Durchliissigkeit aussehlieBlich zwischen 400 und 500 mtt. (Die Durchl~ssigkeit aller l~ilter bei grSBeren Wellenlgngen karm hier unberacksichtigt bleiben, da in diesem Bereich keine induzierende Wirkung mehr vorhanden ist nnd da auf Energiemessungen verzichtet werden konnte.)
In einer ersten Versuchsreihe wurden die Wirkungskurven hinter diesen verschiedenen Filtern verglichen. Abb. 6 and 7 zeigen, dab der erste Kurvenanstieg and das erste l~aximum hinter UG 5 etwa die 5fache Beliehtnngszeit benStigt wie hinter GG 13 l, dab aber die Kombination
i Die Angabe ,,5real wirksamer" erhebt keinen Anspruch auf MlzugroBe Ge- nauigkeit, da die Belichtungszeiten in geometriseher Progression mit dem Faktor 5 variiert warden, so dab das Verhil tnis der Wirksamkeiten start 5 auch 4 oder 7 be- tragen kann. Zwei Versuche, in denen ausnahmsweise der 1%ktor 3 zur Anwendung kam, maehen es wahrscheinlich, dab das Wirkungsverhil tnis eher gr6Ber Ms 5 ist.

Primarvorgang der Polaritatsinduktion bei Equisetum-Sporen 79
GG 3 q- BG 12 bei gleieher Belichtnngszeit den gleichen Effekt hervor- ruft wie UG 5. In Abb. 8 ist ein Versuch dargestellt, der alle 3 Belieh- tungsarten zusammenfgBt und die gleiehe Gesetzm~Bigkeit erkennen ls Diese Abbildung zeigt zugleich, dal~ fiir den zweiten Kurven- anstieg andere Verh/fltnisse vorliegen, indem nun GG 13 nieht mehr wirks~mer ist ~ls UG 5 und die Xombingtion GG 3 -~ BG 12 in ihrer Wirksamkeit welt hinter UG 5 znriickbleibt. DaB hier im iibrigen die Knrve fiir UG 5 insgesamt um einen konstanten Betrag hSher liegt als
v
I 5 25seclZE
- u~'~-'~ ~
I E 25s e c 125 I
8elichl~ngszeil Abb . 6 Abb . 7 Abb . 8
5 25see 2 rain I0
Abb . 6 - - 8 . Die P o l a r i t ~ t s i n d u k t i o n in A b h h u g i g k e i t y o n der Be l i ch tungsze i t h i n t e r ver- seh iedenen F i l t e r k o m b i n a t i o n e n ( t t g - L a m p e ) . Ohne Deekaga r . E inges t r ah l t e Wel len- l~ngen: < 4 0 0 m/~ (UG 5), > 4 0 0 m# (BG 12 + G G 3) bzw. > 3 7 0 m p (GG 13); vgl . Abb . 5). Abszisse: Be l i eh tungsze i t in see n n d ra in . O r d i n a t e : I n d u k t i o n s v e k t o r . E. l imosum (Abb. 6, Ve r such 250), E. variegatum (Abb. 7, Versueh 305) bzw. E. palustre (Abb. 8, u 309)
die zu vergleichenden Kurven, hat methodisohe Griinde und ist ffir unsere ]%lgerungen belanglos.
Der n~heliegende Schlu{~, d~l~ die fiir den zweiten Kurvenanstieg ver~ntwortliche l~eaktion tats~chlich ein anderes Aktionsspektrum be- sitzt als die ffir den ersten Anstieg verantwortliche, ist jedoch noch nieht zwingend, da der Kurvenverlauf oberhalb des ersten Maximums mit- bestimmt wird yon der Durchl~ssigkeit der Spore ffir die betreffende We]lenl~nge; denn yon dieser Durchl~ssigkeit h~ngt des Ausmaf] der Re~ktion auf der Schattenseite ab. Nun konnte bis jetzt nicht nach- gewiesen werden, daI~ die Durchl~ssigkeit oberhalb und unterhalb 400 m F in der gleiehen GrSl]enordnung liegt. Deshalb wurde in einer zweiten Versuehsreihe die Abh/~ngigkeit des zweiten Kurvenanstiegs yon der Lichtqualit~t direkter ~ls in der ersten Versuchsreihe unter- sucht. Mit einer Strahlung, die des Kurvenminimum mSglichst ~usge- pr~gt auftreten l~l]t, wurde einheitlich vorbelichtet (ungefiltertes Licht der Hg-Lampe, 20--25 see oder BG 12 d- GG 3, 2 rain) und sofort
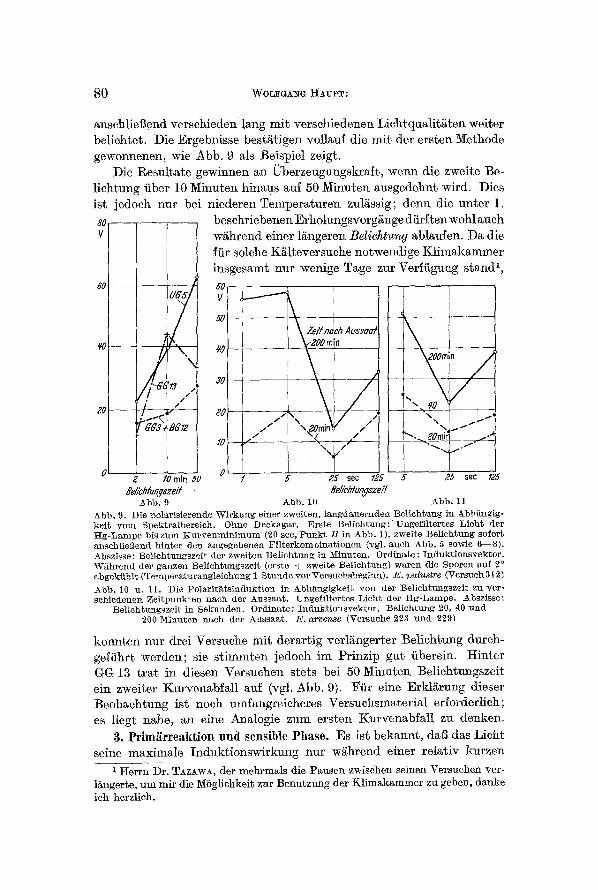
~nschlieI~end verschieden lang mit verschiedenen Lichtqu~lit~ten weiter beliehtet. Die Ergebnisse bestgtigen vollauf die mit der ersten l~Iethode gewonnenen, wie Abb. 9 ~ls Beisloiel zeigt.
Die l~esult~te gewinnen an Uberzeugungskr~ft, wenn die zweite Be- liehtung fiber 10 Minuten hindus ~uf 50 Minuten ausgedehnt wird. Dies ist ]edoeh nur bei niederen Temloeraturen zul~ssig; denn die unter 1. so beschriebenen Erholungsvorg~nge diirften wohl auch V w~hrend einer l~ngeren Belichtung ablaufen. Da die
iiir solche K~lteversuehe notwendige Klimakammer insgesamt nnr wenige Tage zur Verffigung stand 1,
'13 i" 30
f
. 0 ~ 0 z I0 rain 50 I 5 25 sec 125
8ellchluzgszeit . Bel/ehlunflsze# 2~hb. 9 A b b . 10 A b b . 11
A b b . 9. Die DOLUrisierende W i r k u l l g e iner zwei ten , l a n g d a u e r n d e n Be l i ch tung in ~_bhEn~ig- ke i t v e t o Spek t rg lbere ieh . Ohne D e e k a g a r . E r s t e Be l i eh tung : " Un~ef i l te r tes L i e h t tier t t g - L a m o e bis z n m K u r v e n m i n i m n m (20 sec, P u n k t / ~ in 2~bb. 1), zweite Be l i ch tnng sofor t anselfliei~end h i n t e r den a n g e g e b e n e n F i l t e r k o m h i n a t i o n e n (vgl. a n e h Abb . 5 sowie 6 - - 8 ) . Abszisse : Be l i eh tnngsze i t tier zwe i ten B e l i c h t u n g in iV[inuten. O r d i n a t e : I n d u k t i o n s v e k t o r . W ~ h r e n d der g~nzen Be l i ch tungsze i t (erste § zweite Be l i ch tung) w a r e n die Sporen ~uf 20 abgek i ih ] t (TemDera tu ra~g te l chung 1 S tu~de v e t Versuehsheg lnn) . E . 7enquire (Versuch 312)
Abb . 10 n . 11. Die P o l a r i t h t s i n d n k t i o n in A_bh~ngtgkeit y o n tier Bel ieh t~ngsze i t zu ver - seh iedenen Zei t !ounkten n a e h der A u s s a a t . Ungef i l t e r tes L i e h t t ier H g - L a m p e . 2~bszisse:
Be l ieh tu~gsze i t in Seknnden . O r d i n a t e : I n d u k t i o n s v e k t o r . Be l i eh tu~g 20, 40 ~md 200 Minu ten n a c h tier A u s s a a t . E . arvense (Versuche 223 u u d 229)
konnten nut drei Versuche mit der~rtig verl~ngerber Beliehtung dnreh- geifihrt werden; sie stimmten jedoch im Prinzip gut fiberein. Hinter GG 13 trot in diesen Versuchen stets bei 50 Minuten Be]ichtungszeit ein zweiter Kurven~bf~ll ~uf (vgl. Abb: 9). ~fir eine Erkli~rung dieser Beob~ehtung ist noeh umfangreicheres Versuehsm~terial erforderlich; es liegt nahe, an eine Analogie zum ersten Kurven~bf~ll zu denken.
3. Prim~rreaktion und sensible Phase. Es ist bekannt, da~ d~s Licht seine m~xim~le Indtuktionswirkung nur w~hrend einer relativ kurzen
H e r r n D r . TAZAWA, d e r m e h r m ~ l s d i e P ~ u s e n z w i s c h e n s e i n e n V e r s u c h e n v e r -
l ~ n g e r t e , u m m i r d i e M b g l i c h k e i t z u r B e n u t z u n g d e r K l i m ~ k ~ m m e r z u g e b e n , d a n k e
i e h h e r z l i c h .
5 25 sec ;?5

Prirn~rvorgang der Polarit~tsinduktion bei Equisetum-Sporen 81
sensiblen Phase aus/ibt, die im Mi~tel etwa 4 Stunden nach der Aussaat ihren HShepunkt erreicht. Diese sensible Phase ist der Ausdruck eines bestimmten physiologischen Zustandes der Spore und k6nnte entweder yon Einflug auf die Prim~rreaktion der Lichtwirkung sein oder nut ffir den Ablauf der Folgereaktionen yon Bedeutung sein. Im ersten Yalle mfigte die in Abb. 1 dargestellte Wirkungskurve bei weniger empfindliehen Sporen naeh rechts ~rersehoben sein (gleiche Wirk- samkeit bei h6heren Liehtmengen), im zweiten Falle jedoch miigten Maximum und Minimum der Kurve stets bei der gleichen Liehtmenge (bzw. Belichtungszeit) liegen, und die Empfindlichkeit der Sporen wfirde lediglieh fiber die absolute H6he der Ordinaten-Werte bestim- men. Eine Entseheidung konnte dutch die folgenden Versuche herbei geffihrt werden.
In verschiedenen Zeitabst/inden naeh der Aussaat wurde dureh Va- riieren der Belichtungszeit die Wirkungskurve ermittelt. Abb. 10 und 11 zeigen als Beispiele, dal~ tats/ichlieh in alien F/~llen die Kurvenmaxima bzw. Minima mit der gleichen Belichtungszeit erreieht werden, dab es also unm6glieh ist, herabgesetzte Empfindliehkeit der Sporen dutch l~ngere Beliehtung (bzw. grSgere Liehtmengen) zu kompensieren. Der Primiirvorgang is~ somit unabh/~ngig yon der sensiblen Phase.
Diskussion
Die Ermiidung oder S/ittigung der I~eaktion darf wohl so aufgefal3t werden, daft eine fiir die Reaktion wesen~liche Substanz verbraucht und erst nach angemessener Zeit wieder regeneriert oder neugebildet wird. Grunds/itzlich w/ire denkbar, dag es sich bei dieser Substanz entweder um das lichtabsorbierende Pigment handelt oder um den Stoff, der unter Vermittlung dieses Pigments durch das Lieht ver/indert wird (ira folgenden als Stoff ,,X" bezeichnet). Nun zeigt aber der zweite Kurven- anstieg, dab unter geeigneten Bedingungen noch eine weiLere Steigerung der Reaktion m6glich ist (offenbar mit Hilfe eines anderen Pigments, s. u.) ; die Annahme, dal3 die Siittigung auf einen Verbraueh des Stoffes X zur/iekzuffihren ist, erscheint daher unzweckm/il3ig, und wir k6nnen folgern, dal3 die Menge des Pigments zum begrenzenden Faktor werden kann. Die i~egeneration (oder Neubi]dung) dieses Pigments ist tem- peraturabh/~ngig. Zwar wurden keine Versuche speziell zur Ermittlung des Q10 angesetzt, doch lassen die vorliegenden Ergebnisse eine un- gef/~hre Absch/i~zung dieses Wertes zu. Eine Steigerung der tCeaktions- geschwindigkeit yon 2--20 oum das 12faehe wfirde etwa einem Qlo yon 4 entslorechen. Weitergehende Schliisse aus dieser Berechnung zu ziehen scheint jedoeh noeh zu gewagt.
Der zweite Kurvenanstieg zeichnet sich durch eine relativ hShere Wirksamkeit des langwelligen UV gegenfiber dem ersten Kurvenanstieg
Planta . Bd. 5 l 6

aus, so dab hier mit Sieherheit nicht d~s gleiche Pigment ftir die Absorption verantwortlieh sein kann. Ob in diesem FMle ein anderes Pigment die Reaktion ein]eitet oder ob hier (bei etwa 100fucher Struh- lungsenergie; vgl. Abb. 8) der Stoff , ,X" selbst sehon die Liehtabsorption vollzieht 1, kuna vorerst noch nieht entschieden werden. Jedenf~lls haben wit mit zwei versehiedenen photochem. Prim~rprozessen zu reehnen, die sieh in ihrem Eaergiebedarf und in ihrer Wellenl~ngenabh~ngigkeit von- einander unterscheiden, abet sehlieBlich zum gleiehen End-Ergebnis fiihren, g~nz analog zn dem yon Moor (1957) beschriebenen EinfluB der Beliehtung anf die Anthocyunbildung bei Sinapis-Keimlingen. Eine weitere AnMyse, die yon der Aufstellung eines genaueren Aktionsspek- trums der zweiten Reaktion ausgehen muB, erfordert bei Einengung der Spektrulbereiche erheblieh l~ngere Belichtungszeiten; vielleieht lassen sieh derartige Versuche bei Anwendung niederer Temperature~ in gewissem Ausmaft durchftihren.
Nach dem muster der ersten Reuktion wgre zu erwarten, dab such die zweite l~euktion zu einem ~ x i m u m mit anschlieBendem Wieder- ~bfallen der Kurve fiihren mii~te. Mit dem ungefilterten Licht der Hg- Lampe konnte davon jedoeh selbst bei einer Belichtungsdaner yon 4 Stunden (bei 2 o C) nichts bemerkt werden. Das kSnnte einmM daran ]iegen, daft diese zweite l~egktion nicht so schnell abges/~ttigt wird, zum anderen aber auch, dab die Durchl~ssigkeit der Spore fiir die bier wirk- s~men Wellenlgngen viel geringer ist Ms fiir den Spektralbereieh, der die erste l~e~ktion hervorruft. Eine Entscheidung steht noeh aus ; es sei hier nur auf den regelms wiederkehrenden Kurvenabfall hinter GG 13 bei 50miniitiger Belichtung hingewiesen, der bei weiteren Untersuehungen zu beriicksiehtigen sein wird.
Die friiher ges Vermutung ( W m T A ~ 1941, HAUPT 1957), dab die primgre Liehtwirkung mehr oder weniger unabh~ngig vort der sensiblen Phase ist, hut sich in den vorliegenden Versuchen bestgtigt. Damit ist jedoeh noeh nicht der Nachweis erbraeht, dab diese primate Liehtwirkung tats~chlich zu jeder Zeit zu einer l~bilen Pol~risierung ~iihrt, iiber deren Stabilisierung oder vorzeitiges Wiederabklingen der ,,Sensibilit~tszustand" der Sporen entscheidet. Vorl~ufig muB noeh die ~I5gliehkeit o~en bleiben, dab ein Schritt zwischen der prim~ren I~eak- tion und der labilen Polarisierung nur im ~gximum der sensiblen Phuse optimul ~blaufen kann. Eine weitere Diskussion dieser Fr~ge ist ohne neue Versuchsergebnisse nicht sinnvoll.
Ein Modellbeispiel ware etwa die Photolyse der Indolessigsaure, einm~l unter Mitwirkung von Riboflavin, zum anderen (bei geni~gend kurzweliiger Str~hlung) durch eigene Absorption; dab ~ber diesem Modell im Zusammenh~ng mit unserem Versuchsobjekt bis jetzt keine l~ealitgt zugesprochen werden kann, wurde schon fr[iher ausgefiihrt (ItAurT 1957).

Prim/~rvorgang der Polarit~tsinduktion bei Equisetum-Sporen 83
Zasammenfassung Die Lichtreaktion, die schliel~lich zur Polarisierung der Equisetum-
spore Ifihrt, erreicht bei kurzen starken Belichtungen sehnell einen S~tti- gungszust~nd. Eine erneute Reaktion ist bei gleichartiger Belichtung erst nach einer Erholungsphase im Dunkeln m5glieh. Die volle Reak- tionsf~higkei~ is~ bei 200 naeh etwa 20 Minuten, bei 20 nach etwa 4 S~un- den wiederhergestellt. S/it~igung und Wiederherstellung der Lieht- empfindlichkeit beruhen wahrscheinlich au~ Verbraueh und Regenera- tion (oder Neubildung) des die Liehtabsorption vo]lziehenden Pigments.
Aus dem Verlauf der Kurven, die die Abh~ngigkeit der Induktions- wirkung yon der Be]iehtungsd~uer darstellen, 1/~Bt sich ein zweiter photochemischer Prozef] ableiten, der einen um 2--3 GrSl]enordnungen hSheren Energiebed~rf besitzt. Ftir diesen zweiten Prozel] ist nicht das gleiche Pigment verantwortlich wie Ifir den ersten, da die spektrale Empfindlichkeitsverteilung in beiden Fiillen verschieden ist; der lang- wellige UV-Bereieh ist fiir den zweiten Kurvenanstieg relativ wirks~mer als fiir den ersten. Beim zweiten photochemischen Prozel3 (der hShere Strahlungsenergien erfordert ~ls der erste) ist eine Siittigung bis jetzt nicht nachweisbar.
Die prim~ren Liehtreaktionen sind nicht an die sensible Phase der Sporen gebunden; der VerlauI der Wirkungskurven ist unabhiingig vom Zeitpunkt der Belichtung nach der Aussa~t, d .h . das Maximum liegt stets bei der gleiehen Belichtungszeit, und entsprechendes gil~ ftir das Minimum. Nur die absolute HShe der ~ a x i m a und Minima ist am gr51~ten in der Mitre der sensiblen Phase.
Die Untersuchungen wurden durch eine grol]ztigige Forschungs- and Saeh- beihilfe der Deutschen Forschungsgemeinschaft unterstiitzt, wofiir ich auch an dieser Stelle meinen herzlichen Dank aussprechen mSchte, Fiir die zuverlgssige Ausz~hlung der Keimungsrich~ungen yon insgesamt etwa 200000 Sporen bin ich Herrn Dr. W. M~ss zu grol]em Dank verpflichtet, ebenso Frau ~. KV~MEROW- BEECK fiir ihre Hilfe bei der Vorbereitung zahlreicher Versuche.
Literatur HAVPT, W. : Die Induktion der Polariti~t bei der Spore yon Equisetum. Planta
(Berl.) 49, 61--90 (1957). - - Mo~tR, H. : Der Einflul3 monochromatischer S~rahlung auf das L~ngenwachstum des Hypoco~yls und auf die Anthocyanbildung bei Keim- lingen yon Sinapis alb~ L. (~ Brassica alba Boiss.). Planta (Berl.) 49, 389--405 (1957). - - MOSEBACH, G. : 13ber die Polarisierung der Equisetumspore dureh d~s Licht. Planta (Berl.) 33, 340--387 (1943). - - W~ITAKER, D. M. : The effect of uni- lateral ultraviolet light on the development of the Fueus egg. J. gen. Physiol. 24, 263--278 (1941).
Dozen t Dr. WOLFGA~ HAV~T, Botamsches Institut der Universiti~t, Tiibingen, WilhelmstraBe 5
6*