Embed Size (px)
Citation preview

Aus der Klinik für Hals-Nasen-Ohrenheilkunde und Kopf- und Halschirurgie
St. Elisabeth-Hospital –Universitätsklinik- der Ruhr-Universität Bochum
Direktor: Professor Dr. med. H. Hildmann
Untersuchung der Expression Osteoklasten-stimulierender und Osteoklasten-differenzierender
Faktoren im Cholesteatom
Inaugural-Dissertation zur
Erlangung des Doktorgrades der Medizin einer
Hohen Medizinschen Fakultät der Ruhr-Universität Bochum
vorgelegt von Michael Sand aus Bochum
2003

Dekan: Prof. Dr. med. G. Muhr
Referent: Priv.-Doz. Dr. med. H. Sudhoff
Koreferent: Prof. Dr. med. Jürgen Krämer
Tag der mündlichen Prüfung: 09.12.2003

Ich widme die vorliegende Arbeit meinen Eltern und meinem Bruder.

Abkürzungen
AP = alkaline phosphatase = Alkalische Phosphatase
bp = Basenpaare
cDNA = complementary desoxyribonucleic acid
DAB =3,3`-Diaminobenzidin
DEPC = Diethylpyrocarbonat
DNA = desoxyribonucleic acid
DNS = Desoxyribonucleinsäure
dNTP-Mix = Desoxy-Nucleosid-Triphosphat-Mix
DTT = Dithiothreitol
g = Erdbeschleunigung = 9,81 N/kg
Il = Interleukin
HRP = horse raddish peroxidase = Meerrettichperoxidase
JNK = c-jun N-terminal kinase
kDa= kilo Dalton
LAB-SA = labeled-streptavidin-biotin
LAMP = lysosomal-associated membrane proteins
LGPs = lysosomal glycosylated proteins
M-CSF = macrophage colony-stimulating-factor
MHC = major histocompatibility complex = Haupthistokompatibilitätskomplex
min= Minute
nm= Nanometer = ( 910
− m)
OD= optische Dichte
OPG = Osteoprotegerin (OCIF/TR1/RANK)
OPGL = Osteoprotegerin Ligand (ODF/TRANCE/RANKL)
PBS = phosphat-gepufferte Natriumchlorid-Lösung
RANK = receptor activator of NF-κB
RNA = ribonucleic acid
RNS = Ribonukleinsäure
RT-PCR = reverse Transkriptase-Polymerase-Kettenreaktion
sec= Sekunde
TBE = Tris-Borat EDTA
TBS = Tris-gepufferte Natriumchlorid-Lösung
TNF = Tumor-Nekrose-Faktor
TRAFs = tumor necrosis factor receptor-associated factors
TRAP = Tatrat resistente alkalische Phosphatase

Inhaltsverzeichnis
Abkürzungen
1. Einleitung ______________________________________________________ 6
1.1 Das Cholesteatom __________________________________________________6
1.2 Osteoklasten im Rahmen des Knochenabbaus beim Cholesteatom_____7
1.2.1 Osteoprotegerin-Ligand (OPGL)____________________________________8
1.2.2 Osteoprotegerin (OPG) ____________________________________________9
1.2.3 CD4 _____________________________________________________________10
1.2.4 CD68 ____________________________________________________________10
1.3 Fragestellung der Arbeit____________________________________________11
2. Material und Methoden _________________________________________ 12
2.1 Experimenteller Teil Molekularbiologie______________________________12
2.1.1 RNA–Isolierung __________________________________________________12
2.1.2 Photometrische Konzentrationsbestimmung der totalen RNA_______14
2.1.3 Reverse Transkription – Reaktion (RT-Reaktion) ___________________15
2.1.4 Polymerase-Kettenreaktion (PCR) _________________________________18
2.1.5 Agarosegelelektrophorese________________________________________22
2.2. Immunhistochemie ________________________________________________24
2.2.1 Präparate________________________________________________________25
2.2.2 Einfachfärbung von CD 4, CD 68, OPG und OPGL __________________25
2.2.3 Doppelfärbung von CD 4 und OPGL _______________________________31
3. Ergebnisse ____________________________________________________ 33
3.1 Molekularbiologischer Nachweis von OPGL und OPG ________________33
3.2 Immunhistochemie CD 4, CD 68, OPG und OPGL sowie_______________43
Doppelfärbung von CD 4 und OPGL ____________________________________43
4. Diskussion ____________________________________________________ 58
5. Zusammenfassung_____________________________________________ 65
6.Anhang ________________________________________________________ 67
7. Literatur_______________________________________________________ 70
8. Danksagung ___________________________________________________ 76
9. Lebenslauf ____________________________________________________ 77

Einleitung 6
1. Einleitung
1.1 Das Cholesteatom
Unter dem Krankheitsbild des Cholesteatoms versteht man einen entzündlichen
Prozess im Mittelohr, der in ein kongenitales und ein erworbenes Cholesteatom
eingeteilt werden kann. Bei der kongenitalen Form ist das Trommelfell
charakteristischerweise intakt und man vermutet, dass sich das Cholesteatom aus
epithelialen Resten eines defekten Neuralrohrschlusses zwischen der dritten und
fünften Schwangerschaftwoche bildet (10). Persistierende epitheliale Reste nach
der 33. Schwangerschaftswoche wurden bereits 1857 von Remak beschrieben und
1986 von Michaels bestätigt (32,40). Nach dieser Theorie ist die Persistenz der
epithelialen Reste, die sich während der Entwicklung des Mittelohres zwischen der
10. und 33. Schwangerschaftswoche bilden, für die mögliche Entstehung eines
kongenitalen Cholesteatoms verantwortlich. Ein eindeutiger Nachweis für diese
Theorie steht jedoch noch aus. Das wesentlich häufigere sekundäre Cholesteatom
ist meist Folge einer chronischen Otitis media, einer Ventilationsstörung der Tuba
eustachii oder einer serösen Otitis media. Im Gegensatz zum kongenitalen
Cholesteatom zeigt sich am Trommelfell meist ein randständiger Defekt. Ursächlich
wurden für das Cholesteatom in der Vergangenheit neben einer infektiösen Genese
auch ein neoplastisches Geschehen diskutiert, da sich im Cholesteatom ein Anstieg
sowohl des im neoplastischen Gewebe erhöhten Tumorsuppressorgens p53 als
auch von MIB(Ki-67) und Proliferationsmarkern (PCNA, AgNOR) im Vergleich zu
gesundem Gewebe beobachten ließ (1,46). Nach neueren Veröffentlichungen ist
jedoch die so genannte Retraktionstheorie für das primär und sekundär erworbene
Cholesteatom weitläufig akzeptiert. Dabei nimmt man an, dass es durch eine
Retraktion der Pars flaccida des Trommelfells (Shrapnell`sche Membran) zur
Ausbildung einer Retraktiontasche kommt. Durch einen gestörten
Selbstreinigungsmechanismus des Gehörgangshautepithels oder Epithels des
Trommelfells in die Retraktionstasche kommt es zu einer Akkumulation von Epithel-
und Keratinmassen. Diese Epithel- und Keratinansammlung bildet den Debris, der
-unter anderem bedingt durch den überforderten Selbstreinigungsmechanismus- zu
einer Entzündungsreaktion führt (50). Auslöser einer derartigen Retraktion des
Trommelfells können eine chronische Otitis media oder ein Tubenverschluss sein.

Einleitung 7
Nach Luftresorption durch die Mittelohrschleimhaut kommt es im Mittelohr durch den
Verschluss der Tuba eustachii zu keinem Druckausgleich.
Es bildet sich ein relativer Unterdruck, womit die Ausbildung einer Retraktionstasche
des Trommelfells begünstigt wird. Hierbei ist oft die Pars flaccida des Trommelfells
betroffen, da hier im Unterschied zur Pars tensa nur wenig organisiertes Kollagen
sowie praktisch keine elastischen Fasern vorhanden sind (5).
Die Entzündung beginnt in der Retraktionstasche des Trommelfells und resultiert in
einem vorranschreitenden Knochenabbau der knöchernen Mittelohrstrukturen sowie
der Gehörknöchelchen. Als Folge kann sich, klinisch eine Schalleitungsstörung und
eine damit verbundene Beeinträchtigung des Hörvermögens bis hin zur Taubheit
entwickeln. Bedingt durch den stark proliferativen und migrativen Charakter des
Cholesteatoms sowie der knochendestruierenden Potenz kann es zu
schwerwiegenden z.T. auch lebensbedrohlichen Komplikationen wie Hirnabzess,
Meningitis, Sinusvenenthrombose, Mastoiditis oder Labyrinthitis kommen. Bei
ausgedehnten Cholesteatomen kann sich zusätzlich eine entzündungsbedingte
Fazialisparese entwickeln. Die Therapie des Cholesteatoms beschränkt sich auf
eine operative Entfernung des Cholesteatoms. Die medikamentöse Inhibition des
Knochenabbaus durch Bisphosphonate (Zoledronat) ist Bestandteil aktueller
Forschung und ein möglicher Ansatz für eine medikamentöse Therapie (57).
1.2 Osteoklasten im Rahmen des Knochenabbaus beim Cholesteatom
Osteoklasten sind spezialisierte Zellen der Monozyten-Makrophagen Zelllinie mit der
Funktion des Knochenabbaus. Ein Knochenabbau beim Cholesteatom wurde
bereits 1864 von Virchow (10) beschrieben. Zu dieser Zeit wurden dem
Knochenabbau jedoch noch keine Osteoklasten zugeordnet. Man macht heute den
entzündlichen Prozess des Cholesteatoms, dessen Grundlage die Epithelinvasion
bildet, für den beim Cholesteatom zu beobachtenden Osteoklasten vermittelten
Knochenabbau verantwortlich. Es kommt bei der verstärkten Aktivierung von
Osteoklasten im Rahmen des Cholesteatoms häufig zum Abbau der
Gehörknöchelchen, insbesondere des langen Fortsatzes des Incus (56) sowie
anderen knöchernen Bestandteilen des Mittelohres. Eine Erklärung für die
beobachtete Knochendestruktion wurde in dem Anstieg des Druckes im Mittelohr
gesucht.

Einleitung 8
Der durch das Cholesteatom ansteigende Druck im Mittelohr wurde zunächst für
den beobachteten Knochenabbau verantwortlich gemacht. Nachdem jedoch die
Aufklärung des Stoffwechsels der Osteoklasten in den vergangenen Jahren
erhebliche Fortschritte erfahren hat, wird ein neues Konzept zum Knochenabbau
des Cholesteatoms entwickelt, welches die Rolle des Osteoklasten und der
entzündungsvermittelten Osteoklasten-Aktivierung stärker hervorhebt und mit
einbezieht:
Durch das Cholesteatom kommt es bedingt durch den entzündlichen Prozess im
Mittelohr lokal zu einem Anstieg von T-Helfer Zellen, Monozyten und Makrophagen
sowie einer erhöhten Freisetzung von entzündungstypischen Cytokinen (Il-1, Il-1l,
Il-6, TNF-α, TNF-β, Interferon-γ). Diese Cytokine führen wie die physiologischen
Regulatoren des Knochenstoffwechsels (Vitamin D3, Parathormon, Prostaglandine)
durch eine direkte Aktivierung von Osteoblasten zu einer erhöhten Expression von
essentiellen Schlüsselfaktoren des Osteoklasten-Stoffwechsels (23). Zu diesen
Faktoren zählt man neben Macrophage-Colony-Stimulating-Factor (M-CSF), das
Osteoprotegerin (OPG) sowie den Osteoprotegerin-Ligand (OPGL). Die
Oberflächenproteine CD4 und CD68 sind charakteristisch für Makrophagen, die als
Vorläuferzellen von Osteoklasten bezeichnet werden können. Im Folgenden sollen
die Funktionen von OPGL, OPG, CD4 und CD68 erläutert werden.
1.2.1 Osteoprotegerin-Ligand (OPGL)
OPGL ist ein membranständiges, 316 Aminosäuren langes (38kDa) von
Osteoblasten und aktivierten T-Zellen exprimiertes Molekül, das einen sehr starken
stimulierenden Effekt sowohl auf die Osteoklastogenese als auch auf die
Knochenresorptionsaktivität von Osteoklasten besitzt. Die Expression von OPGL
wird durch eine Reihe von Faktoren erhöht, welche für eine verstärkte
Knochenresorption verantwortlich zu sein scheinen. Zu diesen Faktoren zählt man
neben Glucokortikoiden auch das 1,25-Hydroxyvitamin D3, Interleukin-1, Interleukin-
6, Interleukin-1l, Interleukin-17, TNF-α (Tumor Nekrose Faktor α), Prostaglandin E2
und das Parathormon (24). OPGL - ein Mitglied der Tumor Nekrose Faktor-Familie
(TNF) - bindet an seinen auf der Osteoklasten- und Osteoblasten-Vorläuferzellen-
Oberfläche exprimierten Rezeptor RANK (receptor activator of NF-κB) (12).

Einleitung 9
Durch die Bindung von OPGL an seinen Rezeptor RANK - einem Mitglied der
Tumor Nekrose Faktor Rezeptor Familie (TNF-R) - führt der durch OPGL aktivierte
Rezeptor zusammen mit verschiedenen TRAFs (tumor necrosis factor receptor-
associated factors) zur Aktivierung einer Reihe von Stoffwechselwegen, die im
Osteoklasten bzw. in seinen Vorläuferzellen zu einer verstärkten Aktivierung und
Differenzierung führen. Zu diesen Stoffwechselwegen zählt die Aktivierung der c-jun
N-terminal Kinase (JNK) sowie des nuclear factor-κB (16). Am Ende der Aktivierung
der Stoffwechselwege von JNK und NF-κB durch den OPGL aktivierten Rezeptor
RANK und den TRAFs steht im Rahmen des Cholesteatoms der Knochenabbau von
Gehörknöchelchen und knöchernen Bestandteilen des Mittelohres.
1.2.2 Osteoprotegerin (OPG)
OPG ist ein nicht membranständiges, 401 Aminosäuren langes Molekül (45), das
zur Familie der TNF-Rezeptoren gehört. OPG bindet sich an das OPGL auf den
Osteoblasten und verhindert damit die Aktivierung oder Differenzierung eines
Osteoklasten bzw. Osteoklastenvorläufers. Es kann zu keiner Aktivierung des RANK
Rezeptors im Osteoklasten und seinen Vorstufen kommen. OPG fungiert damit als
physiologischer Inhibitor der Knochenresorption. Zusätzlich besitzt es eine im
Osteoklasten Apoptose induzierende Potenz. Es kann deswegen als Gegenspieler
von OPGL bezeichnet werden.
Abb. 1 OPG und OPGL (modifiziert nach Greenfield)

Einleitung 10
Wie oben bereits dargestellt, haben der von Osteoblasten und aktivierten T-Zellen
produzierte Osteoklasten-Inhibierungs-Faktor OPG und der auf Osteoblasten
exprimierte Osteoklasten-Differenzierungs-Faktor OPGL eine sehr bedeutende
Position im Rahmen der Osteoklastendifferenzierung und des
Osteoklastenstoffwechsels.
1.2.3 CD4
CD4 ist ein 55 kDa schweres, membranassoziiertes, heterodimeres Glycoprotein,
das vier extrazelluläre dem Immunglobulin ähnliche Domänen enthält (2,4,30). Für
eine effiziente Signalverarbeitung durch den T-Zell-Rezeptor, der sowohl auf
zytotoxischen T-Zellen als auch auf T-Helferzellen exprimiert wird, sind CD3 Ketten
und CD4 oder CD8 nötig (19). Die zytotoxischen T-Zellen exprimieren CD8,
während T-Helfer Zellen das CD4 Protein exprimieren (20). Der T-Zellrezeptor ist
mit CD4 in der Lage, MHC-Klasse-II-Moleküle zu binden, die von Antigen
präsentierenden Zellen präsentiert werden (7,54). Das CD4 Protein erhöht hier die
Bindungsneigung zwischen dem T-Zellrezeptor und dem Antigen-/MHC-Klasse-II-
Molekül Komplex (54). CD4 wird ebenfalls in dendritischen Zellen, Mikroglia-Zellen,
kortikalen Zellen und ausgereiften medullären Thymozyten exprimiert (15).
1.2.4 CD68
CD68 ist ein 110 kDa schweres, glykosyliertes, transmembranes Glykoprotein. Es
gehört zur Gruppe der glykosylierten lysosomalen Glykoproteine (LGPs), zu denen
man auch LAMP-1 und LAMP-2 zählt (17,38). LGPs sind Hauptbestandteile
lysosomaler Membranen. Ihnen wird eine protektive Funktion gegenüber
Hydrolasen zugesprochen (11). CD68 wird vornehmlich in cytoplasmatischen
Vesikeln und auf der Oberfläche von Makrophagen, Monozyten und Lymphozyten
exprimiert (36,39,43).

Einleitung 11
1.3 Fragestellung der Arbeit
Die chronische Entzündung des Mittelohres geht mit einer erhöhten Konzentration
von entzündungsspezifischen Cytokinen einher. Cytokine, wie z.B. das von T-
Helferzellen produzierte Il-1, führen zu einer Aktivierung der Produktion von
Schlüsselfaktoren der Osteoklasten-Aktivierung und -Differenzierung (41,45). Dies
geschieht über die direkte Aktivierung von Osteoblasten, die für die Produktion der
Faktoren des Osteoklastenstoffwechsels verantwortlich sind. Zu diesen
Schlüsselfaktoren gehört neben OPG auch sein Ligand OPGL. Um ihre Rolle im
Cholesteatom induzierten Knochenabbau näher beschreiben zu können, werden im
Rahmen des DFG-geförderten Projekts „Knochenabbau durch das Cholesteatom“
(DFG Su 226/2-1) im ersten experimentellen Teil mit Hilfe der reversen
Transkriptase-Polymerase-Kettenreaktion (RT-PCR) die Expression von OPG und
OPGL in Cholesteatomen im Vergleich zu Gehörgangshäuten untersucht.
Aus dieser Untersuchung sollen Erkenntnisse über eine eventuelle Beteiligung von
OPG und OPGL am destruktiven Knochenabbau des Cholesteatoms gewonnen
werden. Anschließend an den molekularbiologischen Nachweis werden im zweiten
experimentellen Teil die mit Makrophagen assoziierten Antigene CD4 und CD68,
sowie OPG und OPGL, jeweils in Cholesteatomen und in Gehörgangshäuten
immunhistochemisch untersucht. Durch einen Vergleich zwischen
Cholesteatomgewebe und Gehörgangshaut sollen Rückschlüsse auf Verteilung und
die mögliche Expression der Osteoklastenstoffwechsel regulierenden Faktoren OPG
und OPGL, sowie über Vorhandensein und Verteilung von Vorläuferzellen der
Osteoklasten aus der Granulozyten-Makrophagen-Linie im Cholesteatom gezogen
werden. Dies schafft eine Grundlage für weitere Untersuchungen, welche die
Beteiligung der Osteoklasten am Knochenabbau im Cholesteatom untersuchen.
Abschließend wurden in einer Doppelfärbung CD4 und OPGL in einer Präparatserie
(15 Cholesteatome im Vergleich zu fünf Gehörgangshäute und zwei Negativ-
Kontrollen) immunhistochemisch gefärbt, um eine eventuelle Korrelation der
Lokalisation dieser Antigene zu untersuchen.

Material und Methoden 12
2. Material und Methoden
Die vorliegende Arbeit besteht aus zwei experimentellen Teilen:
1. Es wird mit Hilfe der RT-PCR die Expression des Osteoklasten-
Differenzierungsfaktor OPGL und Osteoklasten-Inhibierungsfaktor OPG jeweils im
Cholesteatom und in der Gehörgangshaut untersucht.
2. Es werden Paraffinschnitte von 15 Cholesteatomen und fünf Gehörgangshäuten
immunhistochemisch auf die Existenz und Lokalisation von OPG, OPGL, und der
Makrophagen-assoziierten Antigene CD4 und CD68 hin untersucht. Eine
abschließende immunhistochemische Doppelfärbung untersucht die Expression und
Lokalisation von CD4 und OPGL in einer Präparatserie.
2.1 Experimenteller Teil Molekularbiologie
Ausgangsmaterial der RT-PCR ist totale RNA, dessen Gewinnung im nächsten
Punkt erläutert wird. Das Versuchsprotokoll für Cholesteatomgewebe und
Gehörgangshaut ist identisch. Zum Schutz vor DNAsen, RNAsen und zur
Vermeidung von möglichen Kontaminationen werden bei allen folgenden Versuchen
Einmalhandschuhe, autoklavierte Glas- und Eppendorfgefäße sowie sterile
Einmalpipettenspitzen benutzt.
2.1.1 RNA–Isolierung
Das gewünschte Gewebe (Cholesteatom bzw. Gehörgangshaut) stammt aus OP-
Eingriffen der HNO-Klinik der Ruhr-Universität-Bochum, St. Elisabeth-Hospital. Die
Einwilligung der Ethik-Komission der Medizinischen Fakultät der Ruhr-Universitat
Bochum liegt vor (Registrierungs Nr. 1356). Die totale RNA wird mit Hilfe des
RNeasy Midi Kits (Qiagen, Hilden) anhand den Angaben des Herstellers isoliert.
Das Gewebe wird nach der operativen Entfernung unmittelbar in flüssigem Stickstoff
tiefgefroren, um die Aktivierung von RNA digestiven RNAsen zu verhindern. Das
tiefgefrorene Gewebe wird entweder direkt

Material und Methoden 13
weiterbearbeitet oder bei –80 °C zwischengelagert. Zum Isolieren wird das Gewebe
zuerst gewogen und mit Hilfe eines mit flüssigem Stickstoff vorgekühlten Mörsers
zerkleinert.
Das pulverisierte Gewebe (75-250 mg) wird mit 4 ml RLT-Puffer supplementiert mit
40 µl β-Mercaptoethanol lysiert. Danach wird das Lysat mit Hilfe einer Spritze (20G,
Durchmesser 0,9 mm) homogenisiert und 10 min bei 3.000-5.000xg (8.000 U/min)
zentrifugiert. Der Überstand wird in ein neues 10 ml Röhrchen gegeben, mit einem
Volumen 70%igem Ethanol versetzt und vorsichtig gemixt. Das Ethanol/Lysat-
Gemisch wird dann auf eine Silicasäule (RNeasy Spin midi column,Qiagen,
Hilden) aufgetragen und 5 min bei 3.000-5.000xg abzentrifugiert. Der Durchfluss
wird verworfen. Die Waschschritte, die zum Eluieren der totalen RNA führen,
beginnen mit dem nächsten Schritt.
Hierzu wird 2 ml des RW1-Puffers auf die Silicasäule aufgetragen, 5 min bei 3.000-
5.000xg zentrifugiert und das Zentrifugat verworfen. Um eine eventuelle
Kontamination der Probe mit Fremd-DNA zu vermeiden, wird eine DNase-
Behandlung mit dem RNase-free DNase-Set (Qiagen) anhand den Angaben des
Herstellers durchgeführt. Es wird 140 µl des RDD-Puffers zu 20 µl der DNase I-
Stammlösung gegeben und sehr vorsichtig gemischt, da die DNase I sehr
empfindlich physikalischer Einwirkung gegenüber ist. Die so fertige DNase-I Lösung
(160 µl) wird in die Mitte der Silicasäule pipettiert und 15 min bei Raumtemperatur
inkubiert. Danach wird 2 ml des RW 1-Puffers dazu gegeben und 5 min bei
Raumtemperatur inkubiert. Es folgt eine 5 minütige Zentrifugation bei 3.000-
5.000xg, wobei der Durchfluss verworfen wird. Beim nächsten Waschschritt wird 2,5
ml des RPE-Puffers auf die Säule gegeben und 2 min bei 3.000-5.000xg
zentrifugiert. Es folgt eine erneute Zugabe von 2,5 ml RPE-Puffer und eine
anschliessende Zentrifugation für 5 min bei 3.000-5.000xg. Nach behutsamen
Verwerfen des Durchflusses wird die Säule auf ein neues 15 ml Collection-Tube
gegeben. Zur Eluation werden 150 µl RNase freies Wasser in die Mitte der Säule
gegeben und für 1 min stehen gelassen. Es folgt dann eine Zentrifugation für 3 min
mit 3.000-5.000xg. Um eine höhere RNA Konzentration zu erhalten und das
Ergebnis der Eluation zu optimieren, wird das Eluat nochmals auf die Säule
gegeben und wiederum 3 min bei 3.000-5.000xg abzentrifugiert. Es folgt die
photometrische Konzentrationsbestimmung der totalen RNA.

Material und Methoden 14
2.1.2 Photometrische Konzentrationsbestimmung der totalen RNA
Die optische Dichte wird mit Hilfe eines Photometers (Ultrospec 2000, Pharmacia
Biotech) jeweils bei 260 nm und 280 nm bestimmt. 1000 µl RNase freies Wasser
dient dabei als Referenz. Zur Konzentrationsbestimmung mit Hilfe der gewonnenen
optischen Dichten bei 260/280 nm wird folgende Formel verwendet:
( )
=
l
gVerdünnungnmODC RNA
µ
µ
1000
*40*)260(
Hinweise:
OD= optische Dichte
Verdünnung= 1:500
40= Es ist allgemein bekannt, dass eine RNA-Lösung mit einer optischen Dichte
von 1 eine Konzentration von 40 l
g
µ
µ in einer Zelle mit einer Schichtdicke von 10
mm besitzt.
Für eine rasche Qualitätsermittlung der isolierten RNA wird zusätzlich das
Absorptionsverhältnis A260nm/A280nm (Ratio) bestimmt.
*)280(
)260(
RnmOD
nmODRatio
−=
*R = Referenz = Absorption von Probe (280nm) – Absorption von Wasser (280nm)
Das Optimum liegt bei reiner RNA zwischen 1,8-2,0.

Material und Methoden 15
2.1.3 Reverse Transkription – Reaktion (RT-Reaktion)
Aus der in 2.1.1 gewonnenen totalen RNA muß zunächst in der RT-Reaktion cDNA
gewonnen werden, um an Ausgangsmaterial für eine PCR zu gelangen. Die
reverse-Transkription-Reaktion wird mit Hilfe des SuperScript First-Strand Synthesis
System (GIBCO BRL) durchgeführt. Dabei wird mit Hilfe einer reversen
Transkriptase die komplementäre DNA aus einzelsträngiger RNA nach folgendem
Prinzip synthetisiert:
Die aus Retroviren gewonnene reverse Transkriptase ist eine RNA-abhängige DNA-
Polymerase. In dieser Arbeit wurde als Primer Random Hexamer verwendet, der
sich an das 3´-Ende des Poly(A)-Schwanz der Ausgangs-RNA bindet. Von diesem
Primer aus startet die reverse Transkriptase die Kettenverlängerung, bei der die
Desoxyribonucleosidtriphosphate (dNTP) als Substrate dienen. Die durch die
Ribonuclease-H-Domäne der reversen Transkriptase normalerweise vorhandene
RNase H Aktivität dient der Hydrolysierung des RNA Teils des entstehenden RNA-
DNA-Hybridanteils (44). Diese wurde vom Hersteller inaktiviert, um eine optimale
cDNA Ausbeute zu erhalten. Am Ende der Reaktion wird durch die Zugabe von
RNase H das RNA-DNA-Hybrid abgebaut.

Material und Methoden 16
Abb. 2 RT-Reaktion (modifiziert nach Stryer)
In dieser Arbeit wurde als Ausgangsmaterial zur Synthese der cDNA 1µg totale RNA
eingesetzt. Das Pipettierschema für die RT-Reaktion ist in Tabelle 1
wiedergegeben, wobei +RT1 und +RT2 positive Ansätze mit Zugabe der reversen
Transkriptase und analog –RT1 und –RT2 negative Kontrollen ohne Zugabe des für
die Reaktion benötigten Enzyms, darstellen.
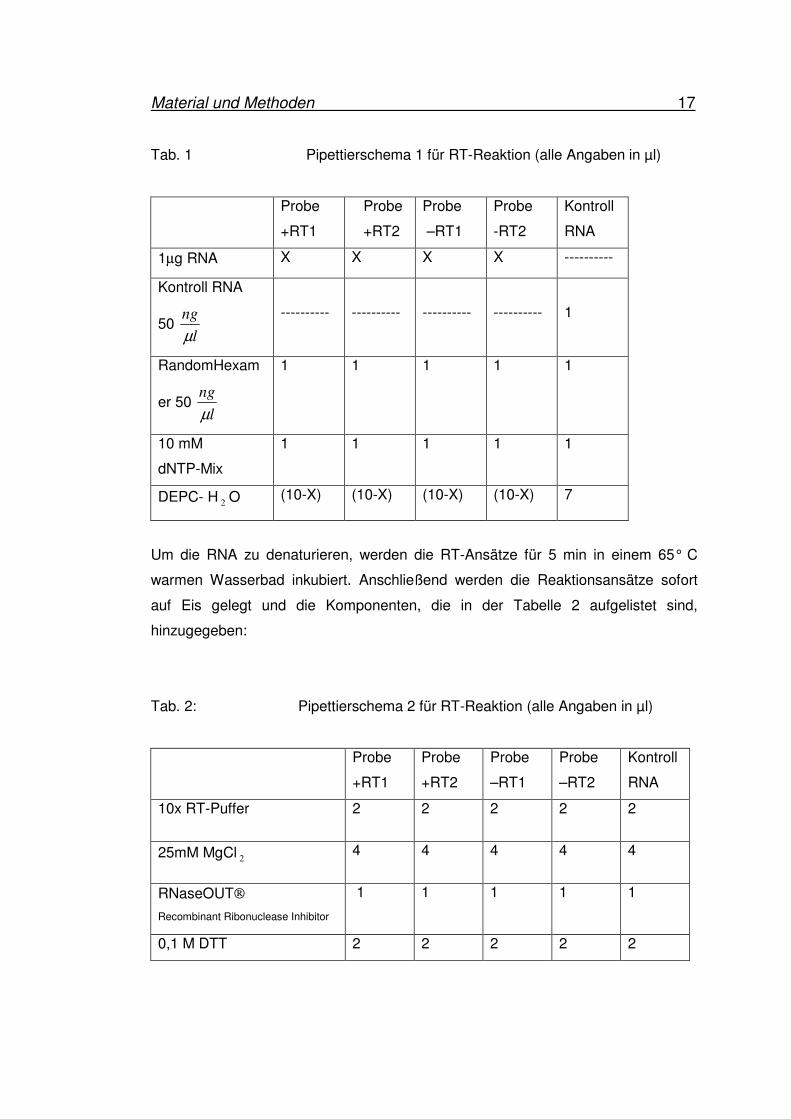
Material und Methoden 17
Tab. 1 Pipettierschema 1 für RT-Reaktion (alle Angaben in µl)
Probe
+RT1
Probe
+RT2
Probe
–RT1
Probe
-RT2
Kontroll
RNA
1µg RNA X X X X ----------
Kontroll RNA
50 l
ng
µ
----------
----------
----------
----------
1
RandomHexam
er 50 l
ng
µ
1 1 1 1 1
10 mM
dNTP-Mix
1 1 1 1 1
DEPC- H2O (10-X) (10-X) (10-X) (10-X) 7
Um die RNA zu denaturieren, werden die RT-Ansätze für 5 min in einem 65° C
warmen Wasserbad inkubiert. Anschließend werden die Reaktionsansätze sofort
auf Eis gelegt und die Komponenten, die in der Tabelle 2 aufgelistet sind,
hinzugegeben:
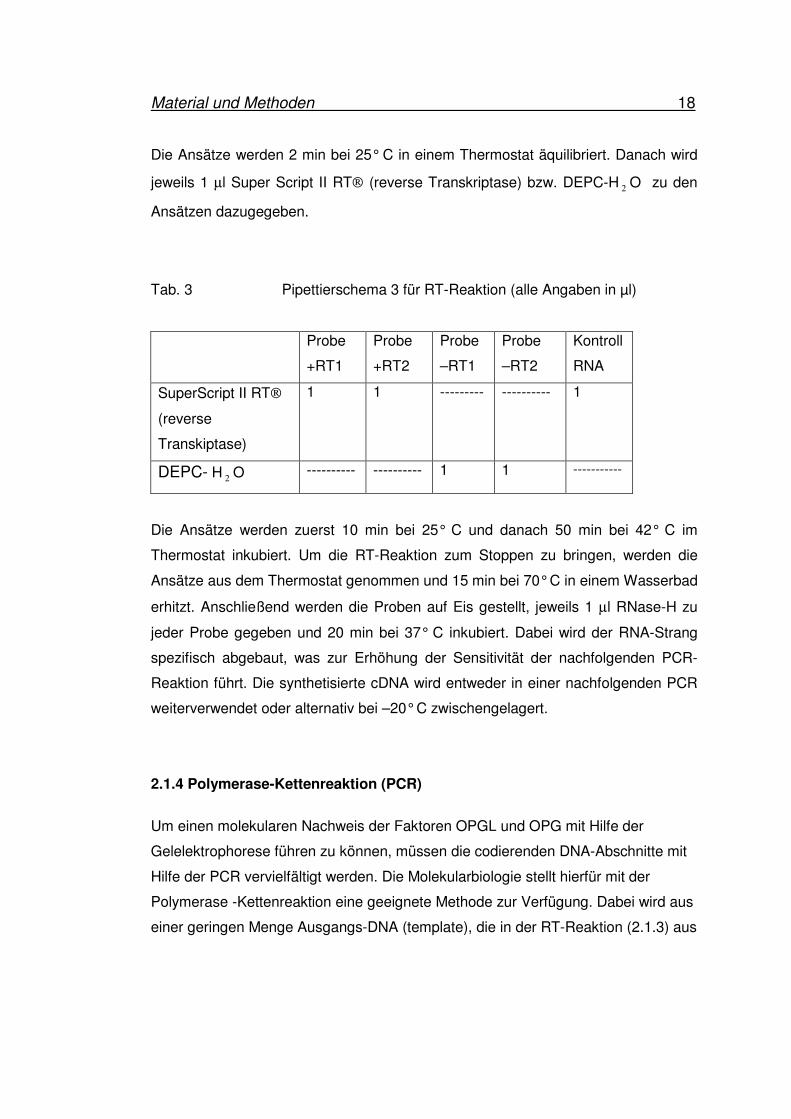
Tab. 2: Pipettierschema 2 für RT-Reaktion (alle Angaben in µl)
Probe
+RT1
Probe
+RT2
Probe
–RT1
Probe
–RT2
Kontroll
RNA
10x RT-Puffer 2 2 2 2 2
25mM MgCl2 4 4 4 4 4
RNaseOUT
Recombinant Ribonuclease Inhibitor
1 1
1 1 1
0,1 M DTT 2 2 2 2 2
Material und Methoden 18
Die Ansätze werden 2 min bei 25° C in einem Thermostat äquilibriert. Danach wird
jeweils 1 µl Super Script II RT (reverse Transkriptase) bzw. DEPC-H2O zu den
Ansätzen dazugegeben.
Tab. 3 Pipettierschema 3 für RT-Reaktion (alle Angaben in µl)
Probe
+RT1
Probe
+RT2
Probe
–RT1
Probe
–RT2
Kontroll
RNA
SuperScript II RT
(reverse
Transkiptase)
1 1 --------- ---------- 1
DEPC- H2O ---------- ---------- 1 1 -----------
Die Ansätze werden zuerst 10 min bei 25° C und danach 50 min bei 42° C im
Thermostat inkubiert. Um die RT-Reaktion zum Stoppen zu bringen, werden die
Ansätze aus dem Thermostat genommen und 15 min bei 70° C in einem Wasserbad
erhitzt. Anschließend werden die Proben auf Eis gestellt, jeweils 1 µl RNase-H zu
jeder Probe gegeben und 20 min bei 37° C inkubiert. Dabei wird der RNA-Strang
spezifisch abgebaut, was zur Erhöhung der Sensitivität der nachfolgenden PCR-
Reaktion führt. Die synthetisierte cDNA wird entweder in einer nachfolgenden PCR
weiterverwendet oder alternativ bei –20° C zwischengelagert.
2.1.4 Polymerase-Kettenreaktion (PCR)
Um einen molekularen Nachweis der Faktoren OPGL und OPG mit Hilfe der
Gelelektrophorese führen zu können, müssen die codierenden DNA-Abschnitte mit
Hilfe der PCR vervielfältigt werden. Die Molekularbiologie stellt hierfür mit der
Polymerase -Kettenreaktion eine geeignete Methode zur Verfügung. Dabei wird aus
einer geringen Menge Ausgangs-DNA (template), die in der RT-Reaktion (2.1.3) aus

Material und Methoden 19
totaler RNA synthetisiert wurde, eine in vitro-Vermehrung durchgeführt, die in einer
großen Menge amplifizierter DNA resultiert. Die folgenden Schritte sind dafür nötig:
Zunächst kommt es durch Wärme zu einer Denaturierung der DNA-Doppelhelix.
Nach dieser Strangtrennung bei 94 °C wird die Temperatur auf 72 °C gesenkt. Da
jetzt Einzelstränge vorliegen, können sich die spezifischen Primer im
Hybridisierungsschritt an die beiden komplementären Sequenzen auf der DNA
binden. Ein Primer bindet sich an den einen Einzelstrang, während sich der andere
Primer an den komplementären Einzelstrang in einer bestimmten Entfernung bindet.
Nach diesem Hybridisierungschritt, der auch Annealing genannt wird, kommt es
unter Zugabe einer DNA-Polymerase und Substraten dNTPs
(Desoxynucleosidtriphosphate) ausgehend von den Primern zu einer
Kettenverlängerung. Die hierfür verwendete DNA-Polymerase ist ein hitzestabiles,
aus dem thermophilen Bakterium Thermus aquaticus gewonnenes Enzym. Diese
Taq-DNA-Polymerase (Taq) bindet sich an einen Primer und synthetisiert einen
komplementären DNA-Strang des zu vervielfältigenden Abschnittes. Wiederholt
man diese Reaktionen mehrfach, so hat man im Endprodukt der PCR-Reaktion eine
große Menge der nachzuweisenden DNA-Sequenz synthetisiert (53). Die in dieser
Arbeit verwendeten spezifischen Primerpaare wurden analog zu schon bekannten
mRNA-Sequenzen konzipiert (Quelle: National Center for Biotechnology Information
- Datenbank) und bei der Firma MWG-Biotech (Ebersberg) hergestellt. Die
Primersequenzen sind im Anhang dargestellt.
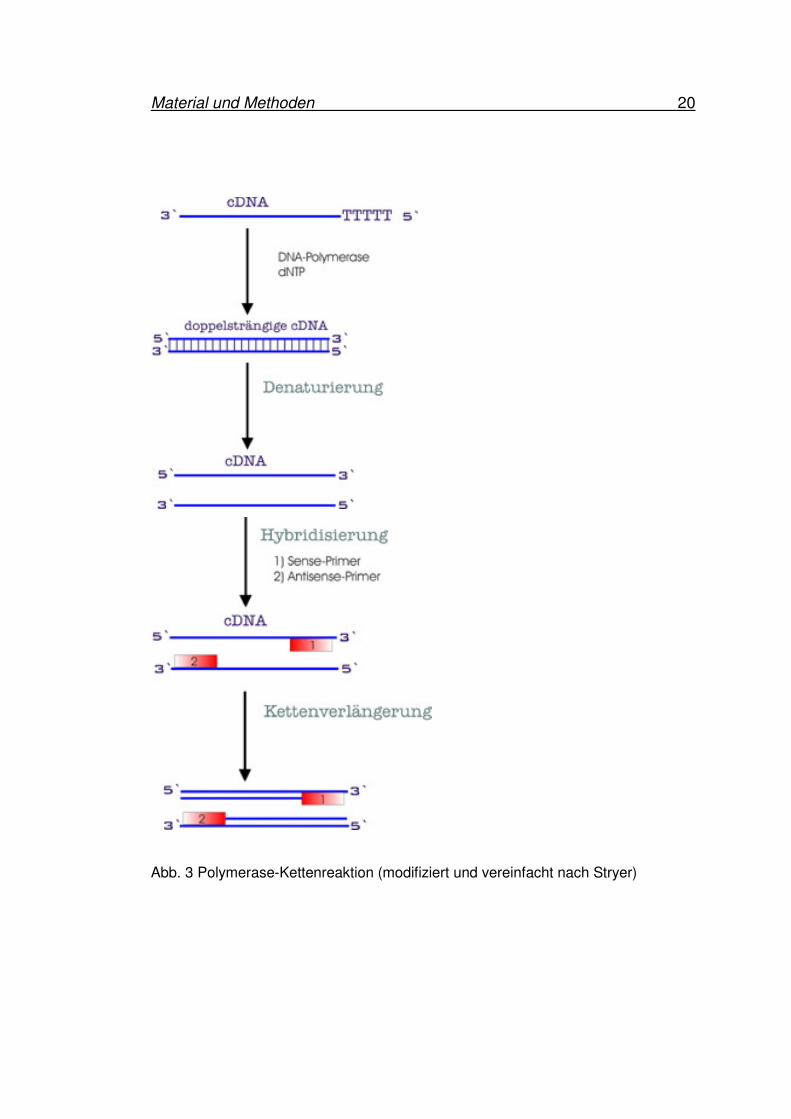
Material und Methoden 20
Abb. 3 Polymerase-Kettenreaktion (modifiziert und vereinfacht nach Stryer)
Material und Methoden 21
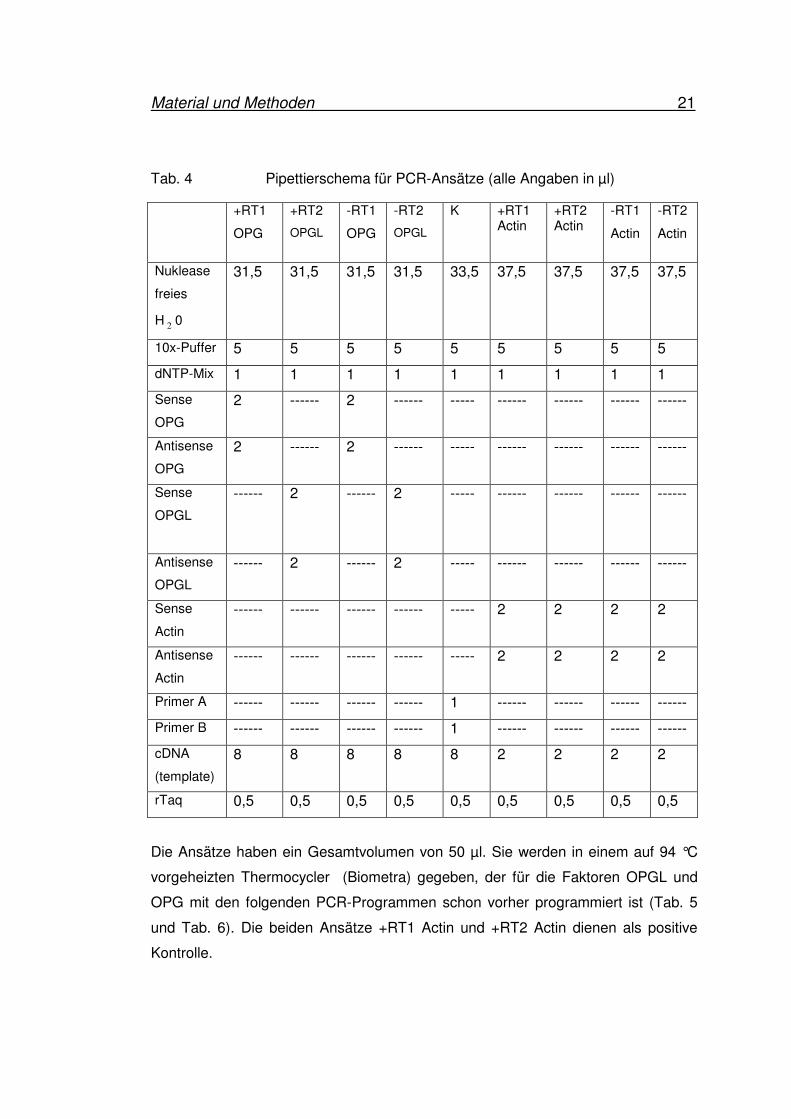
Tab. 4 Pipettierschema für PCR-Ansätze (alle Angaben in µl)
+RT1
OPG
+RT2
OPGL
-RT1
OPG
-RT2
OPGL
K +RT1 Actin
+RT2 Actin
-RT1
Actin
-RT2
Actin
Nuklease
freies
H2
0
31,5 31,5 31,5 31,5 33,5 37,5 37,5
37,5 37,5
10x-Puffer 5 5 5 5 5 5 5 5 5
dNTP-Mix 1 1 1 1 1 1 1 1 1
Sense
OPG
2 ------ 2 ------ ----- ------ ------ ------ ------
Antisense
OPG
2 ------ 2 ------ ----- ------ ------ ------ ------
Sense
OPGL
------ 2 ------ 2 ----- ------ ------ ------ ------
Antisense
OPGL
------ 2 ------ 2 ----- ------ ------ ------ ------
Sense
Actin
------ ------ ------ ------ ----- 2 2 2 2
Antisense
Actin
------ ------ ------ ------ ----- 2 2 2 2
Primer A ------ ------ ------ ------ 1 ------ ------ ------ ------
Primer B ------ ------ ------ ------ 1 ------ ------ ------ ------
cDNA
(template)
8 8 8 8 8 2 2 2 2
rTaq 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5 0,5
Die Ansätze haben ein Gesamtvolumen von 50 µl. Sie werden in einem auf 94 °C
vorgeheizten Thermocycler (Biometra) gegeben, der für die Faktoren OPGL und
OPG mit den folgenden PCR-Programmen schon vorher programmiert ist (Tab. 5
und Tab. 6). Die beiden Ansätze +RT1 Actin und +RT2 Actin dienen als positive
Kontrolle.
Material und Methoden 22
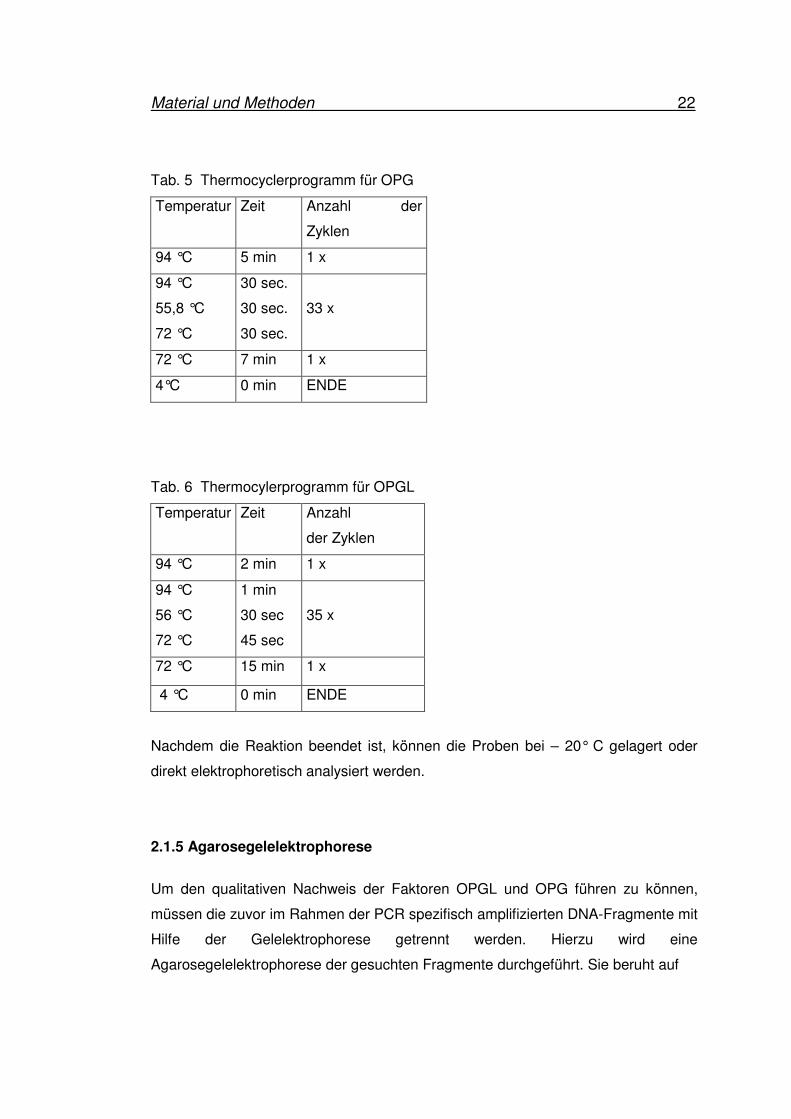
Tab. 5 Thermocyclerprogramm für OPG
Temperatur Zeit Anzahl der
Zyklen
94 °C 5 min 1 x
94 °C
55,8 °C
72 °C
30 sec.
30 sec.
30 sec.
33 x
72 °C 7 min 1 x
4°C 0 min ENDE
Tab. 6 Thermocylerprogramm für OPGL
Temperatur Zeit Anzahl
der Zyklen
94 °C 2 min 1 x
94 °C
56 °C
72 °C
1 min
30 sec
45 sec
35 x
72 °C 15 min 1 x
4 °C 0 min ENDE
Nachdem die Reaktion beendet ist, können die Proben bei – 20° C gelagert oder
direkt elektrophoretisch analysiert werden.
2.1.5 Agarosegelelektrophorese
Um den qualitativen Nachweis der Faktoren OPGL und OPG führen zu können,
müssen die zuvor im Rahmen der PCR spezifisch amplifizierten DNA-Fragmente mit
Hilfe der Gelelektrophorese getrennt werden. Hierzu wird eine
Agarosegelelektrophorese der gesuchten Fragmente durchgeführt. Sie beruht auf

Material und Methoden 23
dem Prinzip, dass Moleküle bedingt durch ihre Eigenladung und Größe in einem
elektrischen Feld unterschiedlich weit wandern. Als Trägermedium kann ein
Polyacrylamidgel oder, wie in dieser Arbeit, ein Agarosegel verwendet werden.
Die Proben werden in einem Agarosegel nach Ihrem Molekulargewicht
elektrophoretisch getrennt und die Fragmentlängen der Proben anhand eines DNA-
Standard bestimmt. Zur Herstellung eines 2%igen Agarosegels werden 1,6 g
Agarose (amresco, Solon/USA) und 80 ml 1 x TBE-Puffer (Merck, Darmstadt) in
einem 250 ml Glasbecher gemischt und 3-4 min in einer Mikrowelle erhitzt.
Nachdem die Agarosekristalle sich vollständig aufgelöst haben, wird der
Flüssigkeitsverlust mit destilliertem Wasser ausgeglichen. Nach wiederholtem
Mischen wird nun 1 µl Ethidiumbromid (10 mg/ml, amresco, Solon/USA) dazu
pipettiert und gemischt. Das Ethidiumbromid, hat die Eigenschaft sich zwischen
einzelne Basenpaare eines doppelsträngigen DNA-Moleküls einzulagern, die dann
unter UV-Licht fluoreszieren, und dient damit der Markierung der DNA (44). Die
Agaroselösung wird möglichst blasenfrei in einem Gelträger mit Kamm
ausgegossen und ist nach dem Abkühlen fertig zum Gebrauch.
Die Proben müssen vor der Elektrophorese mit Probenpuffer (Zentralapotheke der
St. Elisabeth-Stiftung) vorbereitet werden. Dazu werden jeweils 10 µl der zuvor bei
der PCR amplifizierten DNA mit 1,7 µl 6 x Probenpuffer versetzt, gemixt und
herunterzentrifugiert. Um eine Aussage über die Größe der DNA-Fragmente
machen zu können, ist es erforderlich einen DNA-Standard bei jeder
elektrophoretischen Trennung mitlaufen zu lassen. In dieser Arbeit wurde der 50 bp
DNA-Standard (Gibco BRL, Invitrogen life technologies, Carlsbad/USA) eingesetzt.
Das fertige Gel wird in eine mit 1 x TBE-Laufpuffer gefüllte horizontale Gelkammer
(Biometra, Göttingen) gegeben. Hierbei ist darauf zu achten, dass das Gel mit
ausreichend Laufpuffer bedeckt ist. Von den vorbereiteten Proben und dem DNA-
Standard werden jeweils 11 µl in eine Geltasche pipettiert. Bei einer Spannung von
60 V lässt man die Proben für etwa 90 min laufen. Nach 90 min ist die Auftrennung
beendet und das Gel kann zur Dokumentation der DNA-Fragmente mit Hilfe einer
computergesteuerten Digitalkamera unter UV-Licht (Herolab Gel-
Dokumentationssystem, Wiesloch) analysiert und anschließend fotodokumentiert
werden.

Material und Methoden 24
2.2. Immunhistochemie
Die immunhistochemischen Untersuchungen werden an in Formalin fixierten und in
Paraffin eigebetteten histologischen Schnitten von Cholesteatomen und
Gehörgangshäuten durchgeführt. Dabei wird die LSAB-Methode (labeled
streptavidin-biotin) angewendet, welche zu den indirekten immunhistologischen
Techniken gezählt wird. Im Gegensatz zu den direkten immunhistologischen
Techniken sind hierbei die Primärantikörper unmarkiert.
Die LSAB-Methode basiert auf dem folgenden Prinzip: Das anzufärbende Antigen
wird mit einem spezifischen Primärantikörper markiert. Ein biotinylierter
Sekundärantikörper bindet an den Primärantikörper und fungiert als Brücke
zwischen dem Streptavidin-Peroxidase Konjugat (im Falle von CD4 Streptavidin-
alkalische Phosphatase Konjugat), was an den Biotinrest des Sekundärantikörpers
(Brückenantikörper) bindet. Die an das Streptavidin gekoppelte Peroxidase (bzw.
alkalische-Phosphatase) wird durch Zugabe eines Chromogens infolge einer
enzymatischen Spaltungreaktion sichtbar gemacht.
Abb. 4 Testprinzip der LSAB-Methode (modifizierte Abb. Zymed Laboratories Inc.)

Material und Methoden 25
Die Antigene CD4 und CD68, sowie der Osteoklasten-Differenzierungsfaktor OPGL
und der Osteoklasten-Inhibierungsfaktor OPG werden jeweils pro Antigen auf einer
Präparat-Serie einfach gefärbt. Für die Färbung von CD68 wird das Histostain-Plus
Kit (Zymed Laboratories Inc. 85-9643) benutzt. Bei allen anderen angefärbten
Antigenen werden die angegebenen Reagenzien verwendet. In einer Präparat-Serie
wird zusätzlich eine Doppelfärbung von CD4 und OPGL durchgeführt.
2.2.1 Präparate
Eine Präparat-Serie besteht aus 15 verschiedenen Cholesteatomschnitten, fünf
verschiedenen Gehörgangshautschnitten und zwei Negativ-Kontrollen jeweils
unterschiedlicher Patienten.
Die Präparate stammen aus der Hals-Nasen-Ohren-Klinik der Ruhr-Universität-
Bochum, St. Elisabeth-Hospital. Nach der direkten Fixierung in Formalin und
anschließenden Einbettung in Paraffin werden die Präparate mit Hilfe eines
Mikrotoms geschnitten und auf Objektträger aufgezogen. Anschließend werden sie
über Nacht bei 37 °C getrocknet und dann im Kühlschrank bei 6 – 8 °C gelagert.
2.2.2 Einfachfärbung von CD 4, CD 68, OPG und OPGL
Die immunhistologische Einfachfärbung läßt sich in die folgenden sechs Schritte
gliedern:
2.2.2.1 Entparaffinierung und Rehydrierung
2.2.2.2 Demaskierung der Antigene
2.2.2.3 Behandlung mit einem Primärantikörper gegen das nachzuweisende
Antigen
2.2.2.4 Behandlung mit einem Sekundärantikörper gegen den Primärantikörper
2.2.2.5 Behandlung mit einem Enzymkonjugat
2.2.2.6 Sichtbar machen des Primär-/Sekundär-Antikörper Komplexes mit einem
Chromogen

Material und Methoden 26
Zu 2.2.2.1 Entparaffinierung und Rehydrierung
Die Paraffinschnitte werden auf einer Wärmeplatte bei 57-59°C für 10-15 min
getrocknet. Alle nachfolgenden Reagenzien müssen für ein optimales Ergebnis bei
Raumtemperatur verwendet werden. Zur Entparaffinierung werden die
Paraffinschnitte zweimal für 10 min mit Xylol behandelt. Es folgt eine absteigende
Alkoholreihe, die der Rehydrierung des Gewebes dient. Hierzu werden die
Paraffinschnitte
10 Minuten mit 100 Vol% Ethanol,
10 Minuten mit 96 Vol% Ethanol,
5 Minuten mit 70 Vol% Ethanol,
5 Minuten mit 50 Vol% Ethanol und
5 Minuten mit Aqua dest.
behandelt.
Zu 2.2.2.2 Demaskierung der Antigene
Durch die Formalinfixierung kommt es zu einer Eiweißvernetzung im Gewebe. Dies
führt zu einer Maskierung der Antigene, wobei es passieren kann, dass ein
Antikörper ein Antigen nicht erkennt. Daher werden die Präparate zur Demaskierung
der Antigene abhängig vom anzufärbenden Antigen vorbehandelt, um die bei der
Formalinfixierung entstandenen Methylenbrücken, welche durch Quervernetzungen
zu Strukturveränderungen von Proteinen führen können, aufzubrechen (Krull). Bei
CD4, CD68 und OPGL wird die Vorbehandlung der Präparate mit einem
Schnellkochtopf durchgeführt. Bei der Färbung von OPG wird eine proteolytische
Vorbehandlung mit Trypsin vorgenommen.
Vorbehandlung mit Trypsin (für OPG)
Es werden 500 mg des bovinen Trypsins der Firma DAKO, Carpinteria/USA (Code
No. S 20 12 Lot 109) mit 25 ml 0,1%iger Calcium Chlorid (CaCl2) Lösung versetzt.
Danach wird die so gewonnene Lösung in 0,1 ml Portionen alliquotiert und bei

Material und Methoden 27
–20°C gelagert. Zum Herstellen der Gebrauchslösung wird die im Schritt zuvor
gewonnene 0,1 ml Trypsin Stammlösung mit 1,9 ml Calcium Chlorid (0,1%) versetzt.
Diese Lösung ist unmittelbar danach gebrauchsfertig und kann 5 Tage bei 2-8°C
gelagert werden.
Zur proteolytischen Demaskierung der Antigene wird das in der Alkoholreihe
rehydrierte Cholesteatom- / Gehörgangshautpräparat bei 37 °C für 30 min mit 150 µl
Trypsin-Lösung versetzt. Dabei werden Peptidbindungen, die von den
Carboxylgruppen von Arginin und Lysin gebildet werden, hydrolysiert und
ermöglichen somit dem Primärantikörper eine optimale Bindung an das
nachzuweisende Antigen.
Vorbehandlung mit Hitze (für CD4, CD68 und OPGL)
In einem Schnellkochtopf werden 2 l 0,01M Citratpuffer (pH 6) angesetzt. Dazu
werden in 1,9 l Aqua dest. 4,2 g Citronensäuremonohydrat (Merck, Darmstadt
1.00244) aufgelöst. Der pH-Wert wird mit Hilfe von pH-Indikatorstäbchen (Merck
1.09535) und 5 %iger NaOH auf einen pH-Wert von 6 eingestellt. Es wird dann mit
Aqua dest. auf 2 l aufgefüllt. Der so hergestellte Puffer wird bei mittlerer Stufe in
einem Schnellkochtopf erwärmt. Die Paraffinschnitte werden in den Schnellkochtopf
gegeben und 5 min erhitzt.
Zu 2.2.2.3 Behandlung mit einem Primärantikörper
Nach der Hitzebehandlung bzw. der Vorbehandlung mit Trypsin, werden die
Präparate 10 min in TBS Puffer gegeben. Es folgt eine 10 minütige Behandlung mit
3 %iger H2O
2-Lösung, die der Inaktivierung endogener Peroxidasen dient. Danach
wird für 10 min mit TBS Puffer behandelt. Eine lichtundurchlässige
Inkubationskammer aus Plastik wird mit 10 ml TBS Puffer befeuchtet. Die Präparate
werden aus dem TBS genommen und die vorhandene Pufferflüssigkeit auf dem
Objektträger durch seitliches Abklopfen auf Zellstofftüchern entfernt. Mit einem
Fettstift (PAP Pen, Cedarlane, Ontario/Canada) werden die Präparate eingekreist,
um den Reagenzienverbrauch zu reduzieren und eine optimale Inkubationssituation
zu schaffen. Für OPG, OPGL und CD4 werden 100 µl oder 2 Tropfen 10% Normal

Material und Methoden 28
Rabbit Serum (Zymed Laboratories Inc., South San Francisco/USA), für CD68
Blocking Solution A (Zymed Laboratories Inc., South San Francisco/USA) 10 min
auf das Präparat gegeben. Die Blocking Solution bzw. das Normalserum dienen der
Neutralisierung von Ladungen auf der Präparatoberfläche, um unspezifische
Hintergrundfärbung zu vermeiden. Man bereitet die benötigte Menge Primär-
Antikörper vor. Die verwendeten Antikörper und Verdünnungen können aus Tab. 7
entnommen werden.
Tab. 7
Die Blocking Solution wird durch seitliches Abklopfen auf Zellstoffpapier entfernt.
Mit Ausnahme der Negativ-Kontrolle werden auf jedes Präparat 100 µl des mit TBS-
Puffer verdünnten Primär-Antikörpers pipettiert und dann im Kühlschrank bei 4-6 °C
über Nacht inkubiert.
Zu 2.2.2.4 Behandlung mit einem Sekundärantikörper
Am darauffolgenden Tag wird die Inkubationskammer mit den Präparaten bei
Raumtemperatur 10 min aufgewärmt und danach werden die Präparate drei mal
zwei Minuten in TBS-Puffer gewaschen. Die Präparate werden dann seitlich auf
Zellstoffpapier abgeklopft, um den restlichen Puffer zu entfernen. Es werden 2
Tropfen oder 100µl des Biotin-gekoppelten Sekundärantikörpers auf das Präparat
gegeben. Der verwendete Sekundärantikörper ist abhängig vom Primärantikörper,
da der Primärantikörper für den Sekundärantikörper das zu bindende Antigen ist.
Primär-Antikörper Verdünnung mit TBS-Puffer Hersteller des Primär Antikörpers
CD4
CD68
OPG (RANK)
OPGL (RANKL)
1: 50
1: 50
1: 50
1: 50
Santa Cruz Biotech, sc1140
goat polyclonal
DAKO, Code No. M0814
mouse monoclonal
Santa Cruz, sc7625
goat polyclonal
Santa Cruz, sc7627
goat polyclonal
Material und Methoden 29
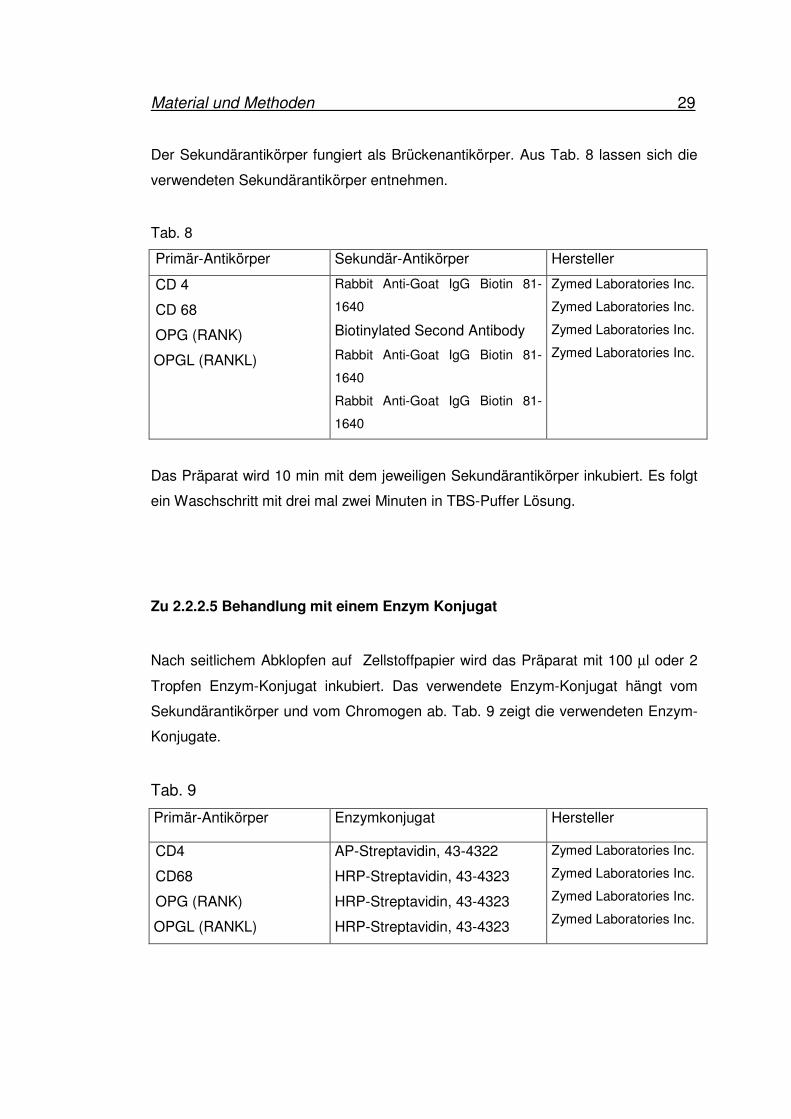
Der Sekundärantikörper fungiert als Brückenantikörper. Aus Tab. 8 lassen sich die
verwendeten Sekundärantikörper entnehmen.
Tab. 8
Primär-Antikörper Sekundär-Antikörper Hersteller
CD 4
CD 68
OPG (RANK)
OPGL (RANKL)
Rabbit Anti-Goat IgG Biotin 81-
1640
Biotinylated Second Antibody
Rabbit Anti-Goat IgG Biotin 81-
1640
Rabbit Anti-Goat IgG Biotin 81-
1640
Zymed Laboratories Inc.
Zymed Laboratories Inc.
Zymed Laboratories Inc.
Zymed Laboratories Inc.
Das Präparat wird 10 min mit dem jeweiligen Sekundärantikörper inkubiert. Es folgt
ein Waschschritt mit drei mal zwei Minuten in TBS-Puffer Lösung.
Zu 2.2.2.5 Behandlung mit einem Enzym Konjugat
Nach seitlichem Abklopfen auf Zellstoffpapier wird das Präparat mit 100 µl oder 2
Tropfen Enzym-Konjugat inkubiert. Das verwendete Enzym-Konjugat hängt vom
Sekundärantikörper und vom Chromogen ab. Tab. 9 zeigt die verwendeten Enzym-
Konjugate.
Tab. 9
Primär-Antikörper Enzymkonjugat Hersteller
CD4
CD68
OPG (RANK)
OPGL (RANKL)
AP-Streptavidin, 43-4322
HRP-Streptavidin, 43-4323
HRP-Streptavidin, 43-4323
HRP-Streptavidin, 43-4323
Zymed Laboratories Inc.
Zymed Laboratories Inc.
Zymed Laboratories Inc.
Zymed Laboratories Inc.

Material und Methoden 30
Nachdem das Präparat 10 min mit dem Enzymkonjugat behandelt wurde, folgt
wieder ein Waschschritt mit drei mal zwei Minuten in TBS-Puffer.
Zu 2.2.2.6 Sichtbar machen des Primär-/Sekundär-Antikörper Komplexes mit
einem Chromogen
Nach seitlichem Abklopfen auf Zellstoff wird das Präparat mit einem Chromogen
behandelt (Tab.10), das den Primär-/ Sekundärantikörper-Komplex sichtbar macht.
Tab. 10 Die verwendeten Chromogene.
Primär-Antikörper Chromogen Hersteller
CD4
CD68
OPG (RANK)
OPGL (RANKL)
Fast Red
DAB
DAB
DAB
Boehringer Mannheim
Zymed Laboratories Inc.
Zymed Laboratories Inc.
Zymed Laboratories Inc.
DAB ist ein lichtempfindliches Chromogen und muß daher in einem lichtgeschützten
Reagenzglas hergestellt und gelagert werden. Nach Herstellung ist es maximal eine
Stunde verwendbar.
Das Chromogen Fast Red (Boehringer, Ingelheim) liegt in Tablettenform vor und
muss in TRIS-HCl Lösung gelöst werden. Je 100 µl der frisch angesetzten
Chromogenlösung werden 3-10 min auf die Präparate gegeben. Danach werden die
Präparate mit TBS gewaschen. Nach seitlichem Abklopfen auf Zellstoff werden zur
Gegenfärbung einige Tropfen Mayer`s Hämalaun-Lösung auf das Präparat
gegeben. Nach einer Minute wird das Präparat mit Aqua dest. gespült. Nach 10 min
in Aqua dest. wird das Präparat eingedeckt. Es werden drei Tropfen des
Eindeckmittels Aquatex (Merck, Darmstadt, 1.08562) auf den Objektträger gegeben
und mit einem Deckglas eingedeckt. Die Farbentwicklung sollte im Mikroskop
kontrolliert werden.

Material und Methoden 31
2.2.3 Doppelfärbung von CD 4 und OPGL
Die Doppelfärbung von CD 4 und OPGL an einer Präparatserie lässt sich ähnlich
wie die immunhistochemische Einfachfärbung (2.2.2) in folgende Punkte gliedern:
2.2.3.1 Demaskierung der Antigene
2.2.3.2 Behandlung mit
- einem Primärantikörper gegen OPGL
- einem Sekundärantikörper gegen den Primärantikörper
- einem Enzymkonjugat
- einem Chromogen
- Double-Staining-Enhancer (Zymed)
2.2.3.3 Behandlung mit
- einem zweiten Primärantikörper gegen CD 4
- einem zweiten Sekundärantikörper gegen den zweiten Primärantikörper
- einem zweiten Enzymkonjugat
- einem Chromogen zum Sichtbarmachen des Primär-/Sekundär-Antikörper
Komplexes
2.2.3.4 Eindecken
2.2.3.1
identisch mit 2.2.2.1
2.2.3.2
Nach der Demaskierung der Antigene durch die Vorbehandlung mit Hitze werden
die Präparate 10 min mit der Blocking Solution (10% Normal Rabbit Serum, Zymed
Laboratories Inc., South San Francisco/USA) behandelt. Es folgt die Inkubation mit
dem Primärantikörper gegen OPGL (Santa Cruz, Heidelberg, sc7627, goat
polyclonal) in einer Verdünnung von 1: 50. Nachdem der erste Primärantikörper
gegen OPGL bei einer Temperatur von ca. 4°C im Kühlschrank über Nacht in einer

Material und Methoden 32
mit TBS befeuchteten, lichtundurchlässigen Inkubationskammer mit dem Gewebe
reagieren konnte, wird der Überschuß mit Hilfe eines 3 x 2 minütigen
Waschvorganges in TBS von dem Gewebe entfernt. Es werden dann im nächsten
Schritt 100 µl Sekundärantikörper (Rabbit Anti-Goat IgG Biotin 81-1640, Zymed
Laboratories Inc., South San Francisco/USA) auf die Präparate gegeben und 15 min
abgedeckt inkubiert. Nach einem anschließenden Waschschritt (3x2 min TBS)
werden 100 µl Enzymkonjugat (HRP-Streptavidin, 43-4323, Zymed Laboratories
Inc., Verdünnung 1:150, South San Francisco/USA) auf das Präparat pipettiert. Das
Enzymkonjugat bindet an das Biotin des Sekundärantikörpers, und die daran
gekoppelte Peroxidase führt durch eine Reaktion mit dem Chromogen zur
Farbentwicklung. Nach dem Waschschritt (3 x 2 min TBS) werden als Chromogen
100 µl DAB, 3 min auf das Präparat gegeben. Nach erfolgter Farbentwicklung wird
das DAB mit TBS abgespült. Es folgt eine 30 minütige Inkubation mit einem Double-
Staining-Enhacer der Firma Zymed Laboratories Inc.
2.2.3.3
Der Double-Staining-Enhancer wird mit Aqua dest. abgespült und das Präparat
3x2 min in TBS gewaschen. Nach der 10 minütigen Behandlung mit Blocking
Solution wird im nächsten Schritt 100 µl des zweiten Primärantikörpers gegen CD 4
(Santa Cruz Biotech, Heidelberg, sc1440, goat polyclonal) in einer Verdünnung von
1:50 auf das Präparat gegeben und über Nacht bei 4°C im Kühlschrank inkubiert.
Am nächsten Tag erfolgt nach dem Waschschritt (3x2 min TBS) die Zugabe von 100
µl des Sekundärantikörpers Rabbit Anti-Goat IgG Biotin 81-1640 der Firma Zymed
Laboratories Inc., South San Francisco/USA. Das Präparat wird nach 15 min
Inkubation mit dem Sekundärantikörper und einem Waschschritt (3x2 min TBS) für
15 min mit 100µl des Enzymkonjugates (AP-Streptavidin, 43-4322, Zymed
Laboratories Inc., South San Francisco/USA) inkubiert. Es folgt ein Waschschritt
(3x2 min) und die Behandlung mit 100µl des Chromogens Fast Red (Anleitung
identisch mit 2.2.2.5).
2.2.3.4 siehe 2.2.2.5
Die Beschreibung der computergestützten, lichtmikroskopisch gewonnen
Ergebnisse des immunhistochemischen Teils der Arbeit erfolgt unter 3.2.
Ergebnisse 33
3. Ergebnisse
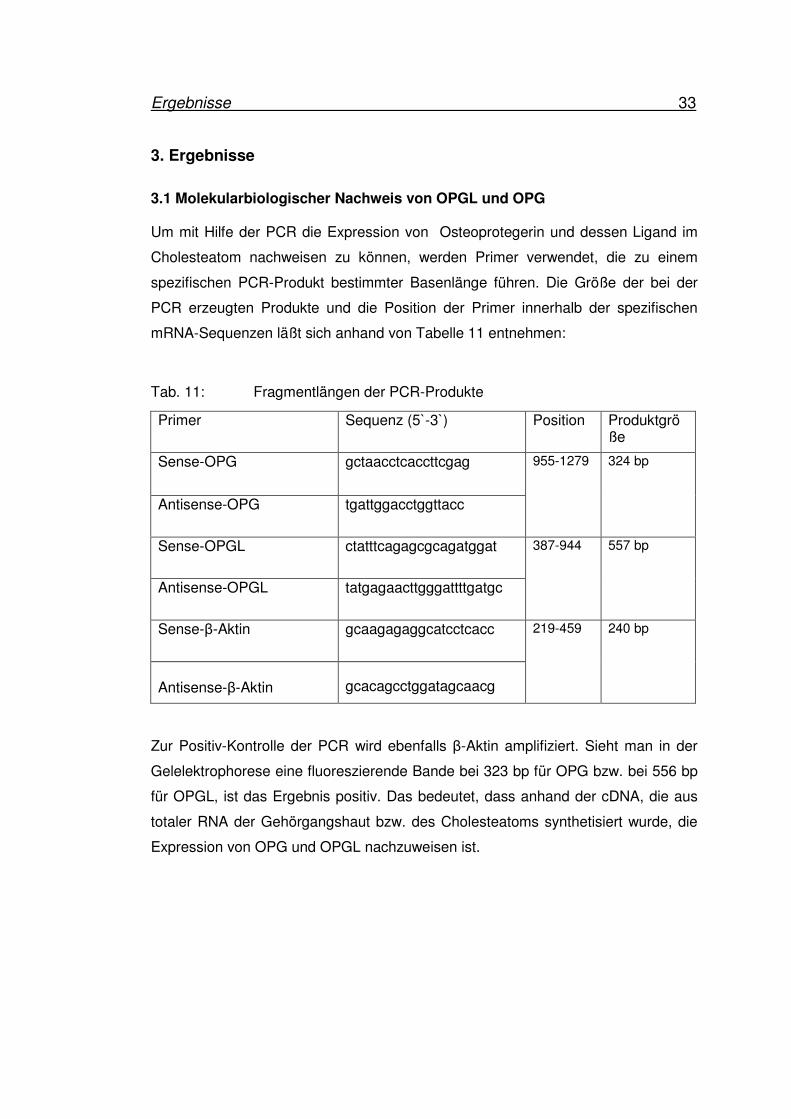
3.1 Molekularbiologischer Nachweis von OPGL und OPG Um mit Hilfe der PCR die Expression von Osteoprotegerin und dessen Ligand im
Cholesteatom nachweisen zu können, werden Primer verwendet, die zu einem
spezifischen PCR-Produkt bestimmter Basenlänge führen. Die Größe der bei der
PCR erzeugten Produkte und die Position der Primer innerhalb der spezifischen
mRNA-Sequenzen läßt sich anhand von Tabelle 11 entnehmen:
Tab. 11: Fragmentlängen der PCR-Produkte
Zur Positiv-Kontrolle der PCR wird ebenfalls β-Aktin amplifiziert. Sieht man in der
Gelelektrophorese eine fluoreszierende Bande bei 323 bp für OPG bzw. bei 556 bp
für OPGL, ist das Ergebnis positiv. Das bedeutet, dass anhand der cDNA, die aus
totaler RNA der Gehörgangshaut bzw. des Cholesteatoms synthetisiert wurde, die
Expression von OPG und OPGL nachzuweisen ist.
Primer Sequenz (5`-3`) Position Produktgröße
Sense-OPG
gctaacctcaccttcgag
Antisense-OPG tgattggacctggttacc
955-1279 324 bp
Sense-OPGL ctatttcagagcgcagatggat
Antisense-OPGL tatgagaacttgggattttgatgc
387-944 557 bp
Sense-β-Aktin
gcaagagaggcatcctcacc
Antisense-β-Aktin
gcacagcctggatagcaacg
219-459 240 bp

Ergebnisse 34
OPG-Nachweis im Cholesteatom
Die molekularbiologische Untersuchung zeigte bei allen acht untersuchten
Cholesteatomen verschiedener Patienten eine spezifische Bande bei 324
Basenpaaren. Damit wurde der codierende DNA-Abschnitt für OPG in allen acht
untersuchten Cholesteatomen nachgewiesen. Die Negativkontrollen waren
ausnahmslos negativ. Im folgenden sind die relevanten Gelabschnitte aus den
dokumentierten Bildern dargestellt:
Abb. 5 Cholesteatom 1 OPG Abb. 6 Cholesteatom 2 OPG
Banden: Banden: siehe links
1= DNA-Standard
2= β-Aktin
3= 324bp Produkt (OPG-Nachweis)

Ergebnisse 35
Abb. 7 Cholesteatom 3 OPG Abb. 8 Cholesteatom 4 OPG
Banden: Banden: siehe links
1= DNA-Standard
2= 324bp Produkt
Abb. 9 Cholesteatom 5 OPG Abb. 10 Cholesteatom 6 OPG
Banden: Banden: siehe links
1= DNA-Standard
2= 324bp Produkt
Ergebnisse 36
Abb. 11 Cholesteatom 7 OPG Abb. 12 Cholesteatom 8 OPG
Banden: Abb. 9 Banden: siehe Abb. 9

Ergebnisse 37
OPG Nachweis in Gehörgangshaut
In den fünf untersuchten Gehörgangshäuten ließ sich eine Bande bei ca. 324 bp
nachweisen. Damit wurde auch hier in allen untersuchten Proben ein Nachweis für
die Expression von OPG erbracht. In drei Fällen erscheinen die Banden relativ
schwach, sind aber dennoch als positiv zu werten. Die Negativkontrollen waren alle
negativ.
Abb. 13 Gehörgangshaut OPG
Banden:
1= DNA-Standard
2= 324bp Produkt
OPGL Nachweis im Cholesteatom
Das spezifische Amplikon, das für OPGL codiert, wurde in allen acht
Cholesteatomen nachgewiesen. Die folgenden Gelabschnitte zeigen in Tasche Nr. 1
den DNA-Standard und in Tasche Nr. 2 das 557bp lange PCR-Produkt für den
OPGL Nachweis.

Ergebnisse 38
Abb. 14 Cholesteatom 1 OPGL Abb. 15 Cholesteatom 2 OPGL
Banden: Banden: siehe Abb. 14
1= DNA-Standard
2= 557bp Produkt
Abb. 16 Cholesteatom 3 OPGL Abb. 17 Cholesteatom 4 OPGL
Banden: siehe Abb. 14 Banden: siehe Abb. 14
Ergebnisse 39
Abb. 18 Cholesteatom 5 OPGL Abb. 19 Cholesteatom 6 OPGL
Banden: Banden: siehe Abb. 18
1= DNA-Standard
2= 557bp Produkt
Abb. 20 Cholesteatom 7 OPGL Abb. 21 Cholesteatom 8 OPGL
Banden: siehe Abb. 18 Banden: siehe Abb. 18

Ergebnisse 40
OPGL Nachweis in Gehörgangshaut
In der Gehörgangshaut konnte in einem Fall eine stark positive Bande
(Gehörgangshaut 2), und in einem anderen Fall eine sehr schwach positive Bande
(Gehörgangshaut 3) detektiert werden. In drei weiteren Fällen war der Nachweis für
OPGL negativ. Auch hier zeigt die erste Tasche wie zuvor den DNA-Standard und
die zweite Tasche eine zu erwartende Bande bei 557bp. Bei negativen Proben ist in
der zweiten Tasche kein spezifisches PCR-Produkt nachzuweisen.
Abb. 22 Gehörgangshaut 1 OPGL Abb. 23 Gehörgangshaut 2
OPGL
Nachweis negativ Banden:
1= DNA-Standard
2= 557bp Produkt

Ergebnisse 41
Abb. 24 Gehörgangshaut 3 OPGL Abb. 25 Gehörgangshaut 4 OPGL
Banden: Nachweis negativ
1= DNA-Standard
2= 557bp Produkt
Abb. 26 Gehörgangshaut 5 OPGL
Nachweis negativ
Ergebnisse 42
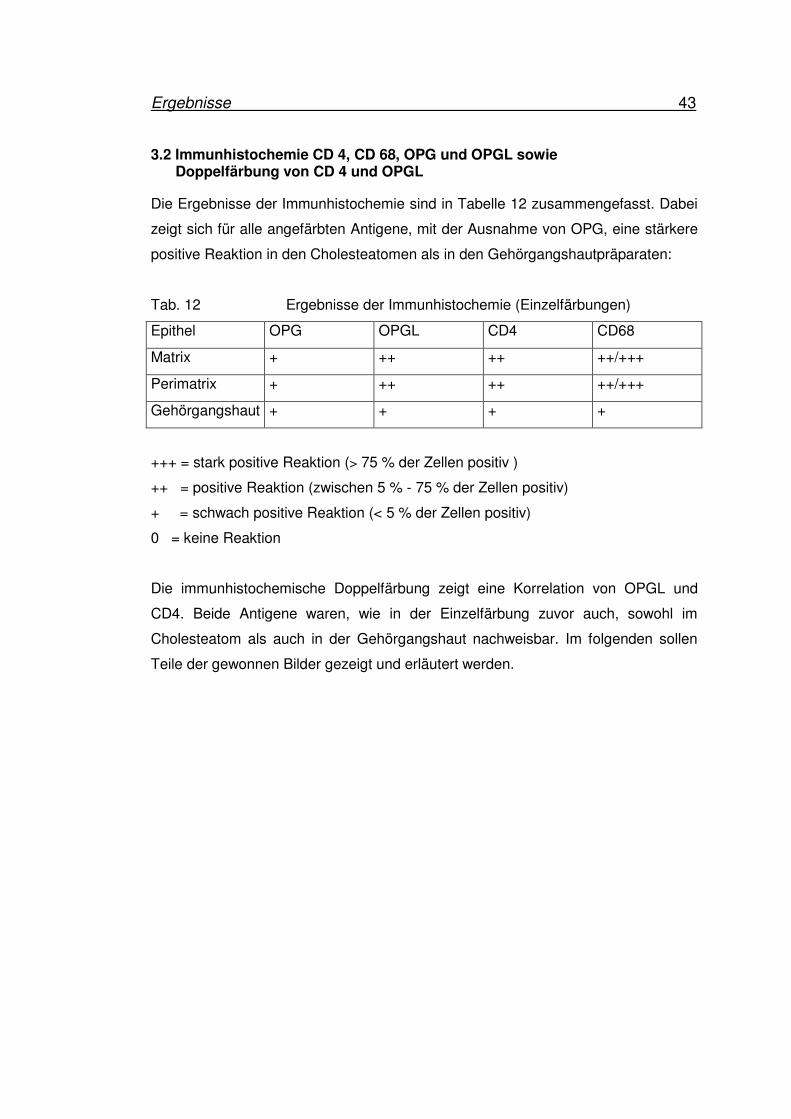
Zusammenfassung der Ergebnisse
Die Abbildungen 27 und 28 zeigen einen Überblick über die Ergebnisse des
molekularbiologischen Teils:
0
1
2
3
4
5
6
7
8
positive
Proben
1 2
OPGL OPG
Abb. 27 Expression von OPGL (1) und OPG (2) im Cholesteatom (n=8)
0
1
2
3
4
5
positive
Proben
1 2
OPGL OPG
Abb. 28 Expression von OPGL (1) und OPG (2) in Gehörgangshaut (n=5)
Ergebnisse 43
3.2 Immunhistochemie CD 4, CD 68, OPG und OPGL sowie Doppelfärbung von CD 4 und OPGL
Die Ergebnisse der Immunhistochemie sind in Tabelle 12 zusammengefasst. Dabei
zeigt sich für alle angefärbten Antigene, mit der Ausnahme von OPG, eine stärkere
positive Reaktion in den Cholesteatomen als in den Gehörgangshautpräparaten:
Tab. 12 Ergebnisse der Immunhistochemie (Einzelfärbungen)
Epithel OPG OPGL CD4 CD68
Matrix + ++ ++ ++/+++
Perimatrix + ++ ++ ++/+++
Gehörgangshaut + + + +
+++ = stark positive Reaktion (> 75 % der Zellen positiv )
++ = positive Reaktion (zwischen 5 % - 75 % der Zellen positiv)
+ = schwach positive Reaktion (< 5 % der Zellen positiv)
0 = keine Reaktion
Die immunhistochemische Doppelfärbung zeigt eine Korrelation von OPGL und
CD4. Beide Antigene waren, wie in der Einzelfärbung zuvor auch, sowohl im
Cholesteatom als auch in der Gehörgangshaut nachweisbar. Im folgenden sollen
Teile der gewonnen Bilder gezeigt und erläutert werden.
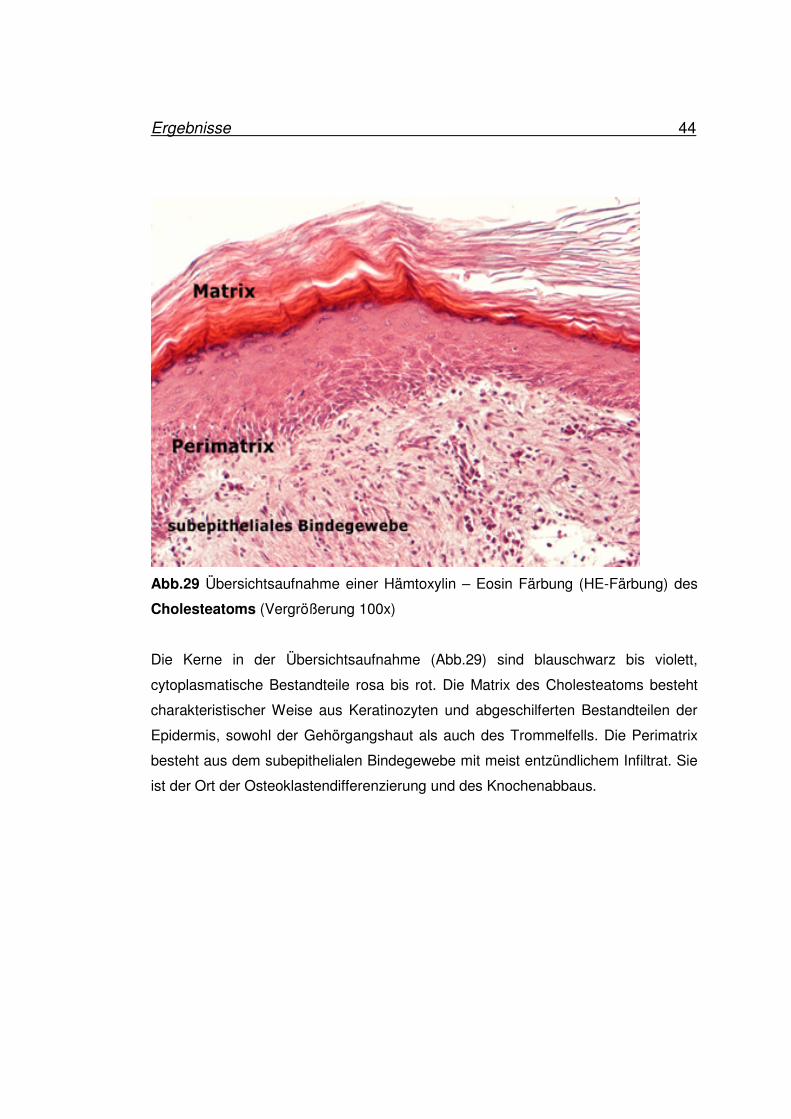
Ergebnisse 44
Abb.29 Übersichtsaufnahme einer Hämtoxylin – Eosin Färbung (HE-Färbung) des
Cholesteatoms (Vergrößerung 100x)
Die Kerne in der Übersichtsaufnahme (Abb.29) sind blauschwarz bis violett,
cytoplasmatische Bestandteile rosa bis rot. Die Matrix des Cholesteatoms besteht
charakteristischer Weise aus Keratinozyten und abgeschilferten Bestandteilen der
Epidermis, sowohl der Gehörgangshaut als auch des Trommelfells. Die Perimatrix
besteht aus dem subepithelialen Bindegewebe mit meist entzündlichem Infiltrat. Sie
ist der Ort der Osteoklastendifferenzierung und des Knochenabbaus.
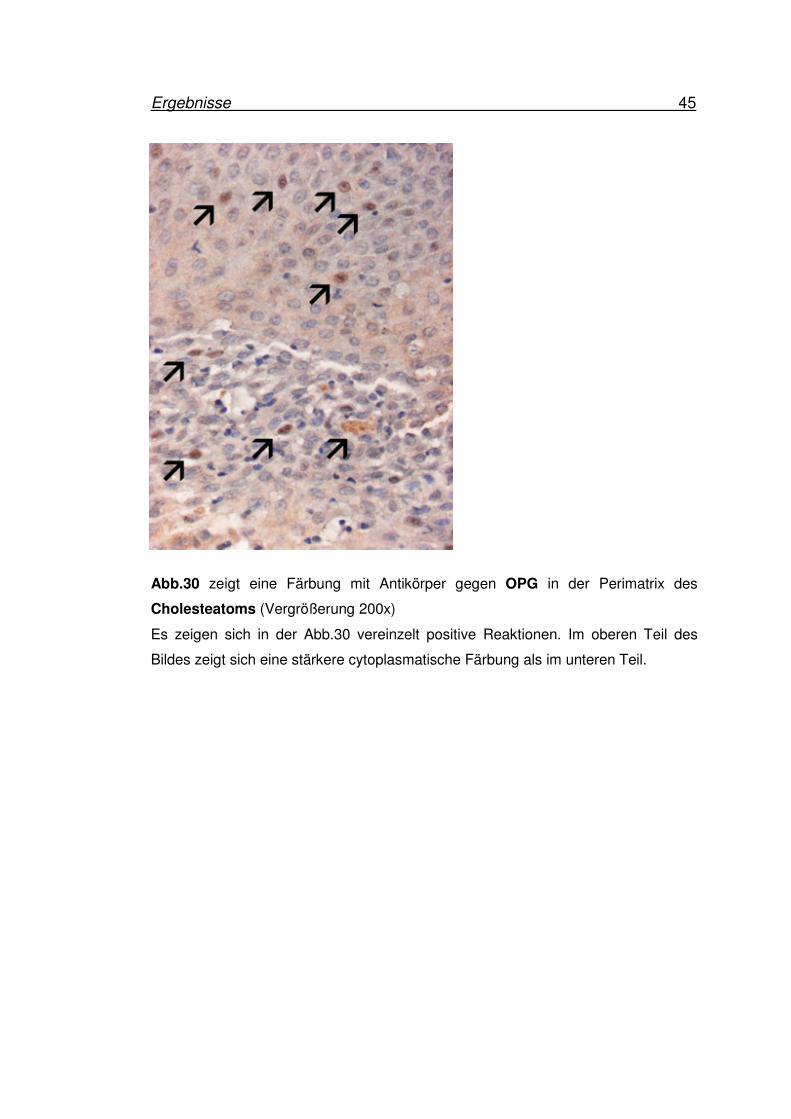
Ergebnisse 45
Abb.30 zeigt eine Färbung mit Antikörper gegen OPG in der Perimatrix des
Cholesteatoms (Vergrößerung 200x)
Es zeigen sich in der Abb.30 vereinzelt positive Reaktionen. Im oberen Teil des
Bildes zeigt sich eine stärkere cytoplasmatische Färbung als im unteren Teil.
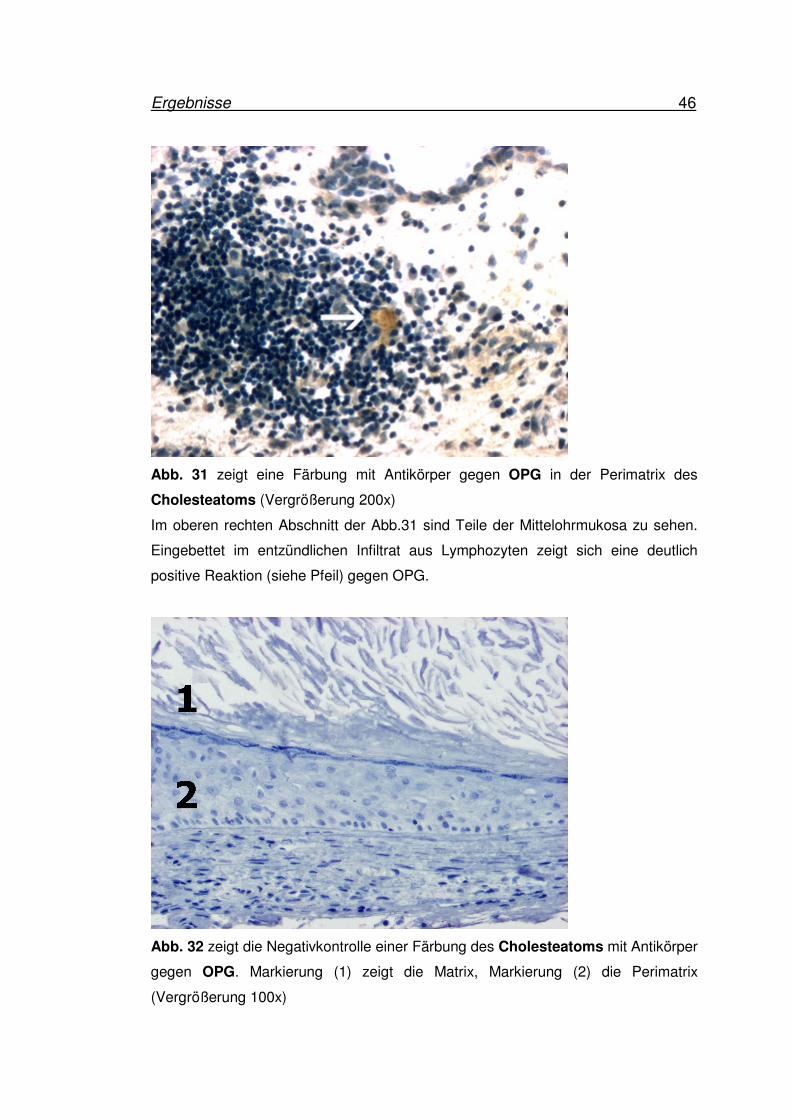
Ergebnisse 46
Abb. 31 zeigt eine Färbung mit Antikörper gegen OPG in der Perimatrix des
Cholesteatoms (Vergrößerung 200x)
Im oberen rechten Abschnitt der Abb.31 sind Teile der Mittelohrmukosa zu sehen.
Eingebettet im entzündlichen Infiltrat aus Lymphozyten zeigt sich eine deutlich
positive Reaktion (siehe Pfeil) gegen OPG.
Abb. 32 zeigt die Negativkontrolle einer Färbung des Cholesteatoms mit Antikörper
gegen OPG. Markierung (1) zeigt die Matrix, Markierung (2) die Perimatrix
(Vergrößerung 100x)
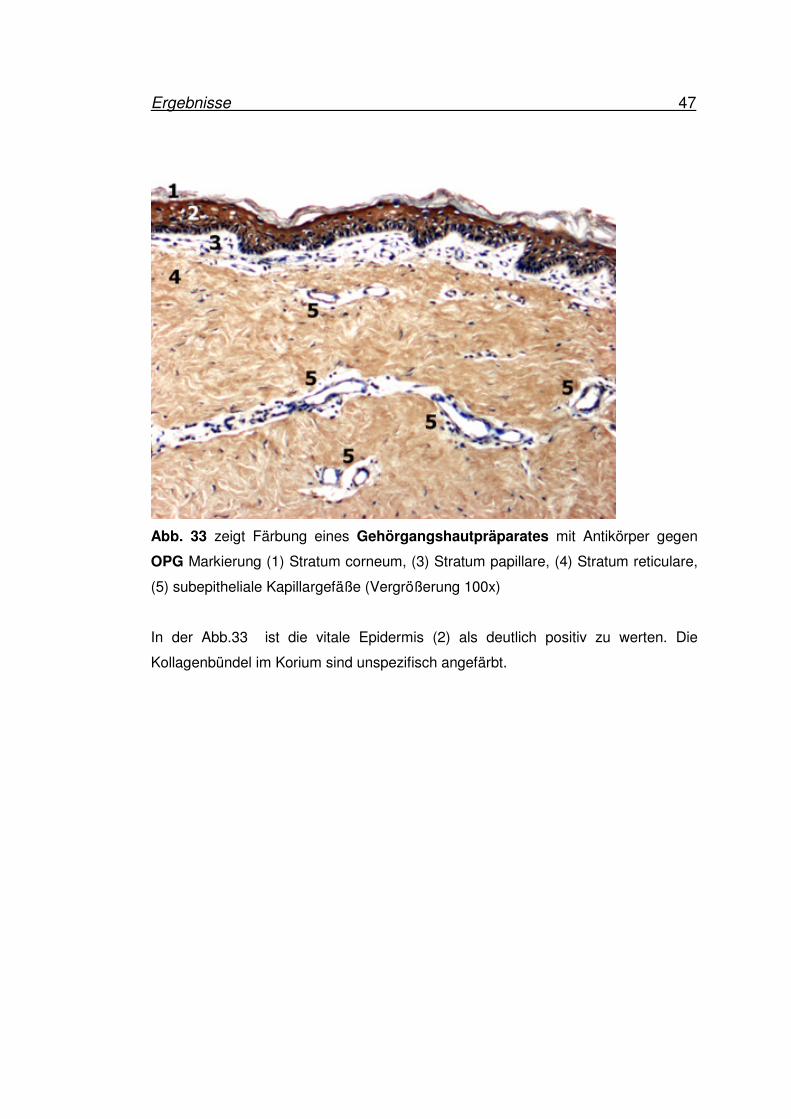
Ergebnisse 47
Abb. 33 zeigt Färbung eines Gehörgangshautpräparates mit Antikörper gegen
OPG Markierung (1) Stratum corneum, (3) Stratum papillare, (4) Stratum reticulare,
(5) subepitheliale Kapillargefäße (Vergrößerung 100x)
In der Abb.33 ist die vitale Epidermis (2) als deutlich positiv zu werten. Die
Kollagenbündel im Korium sind unspezifisch angefärbt.
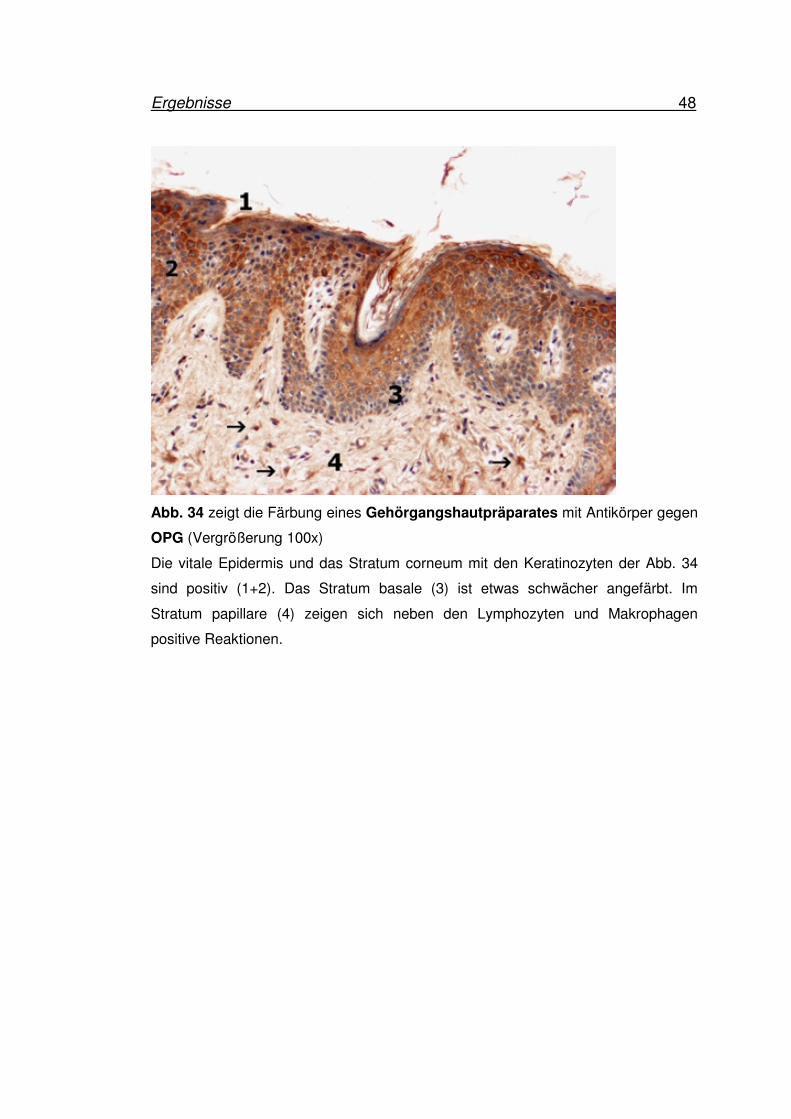
Ergebnisse 48
Abb. 34 zeigt die Färbung eines Gehörgangshautpräparates mit Antikörper gegen
OPG (Vergrößerung 100x)
Die vitale Epidermis und das Stratum corneum mit den Keratinozyten der Abb. 34
sind positiv (1+2). Das Stratum basale (3) ist etwas schwächer angefärbt. Im
Stratum papillare (4) zeigen sich neben den Lymphozyten und Makrophagen
positive Reaktionen.
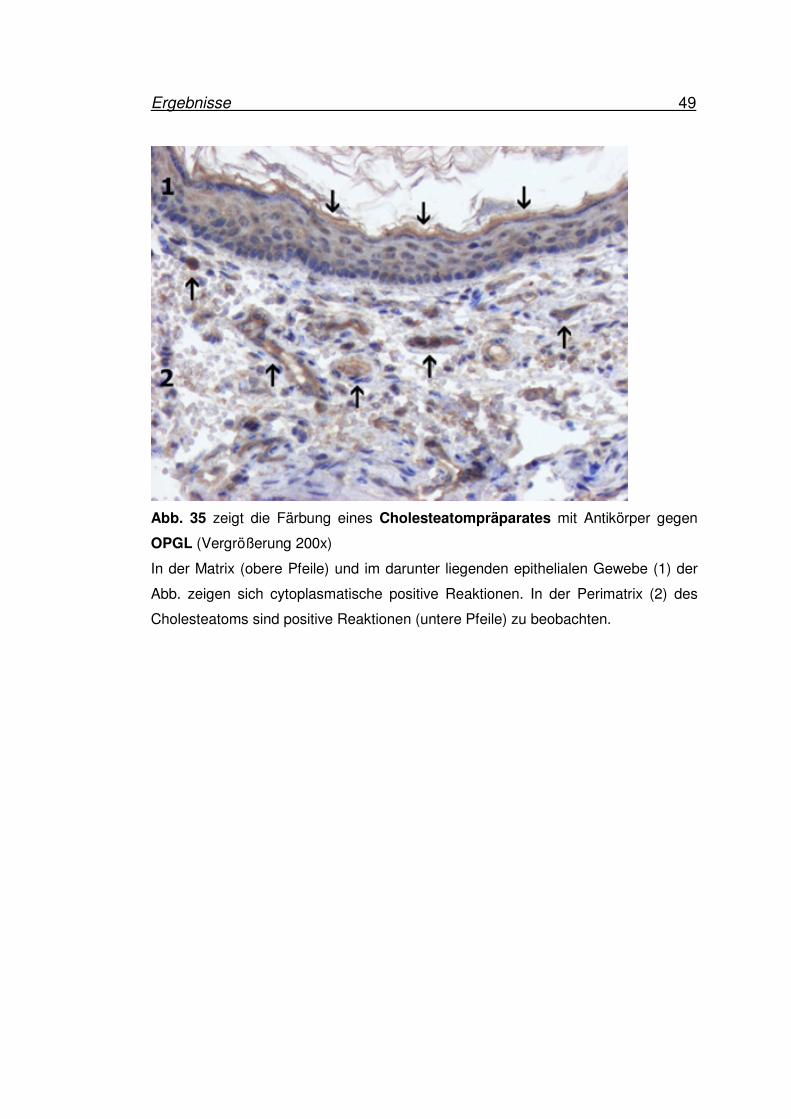
Ergebnisse 49
Abb. 35 zeigt die Färbung eines Cholesteatompräparates mit Antikörper gegen
OPGL (Vergrößerung 200x)
In der Matrix (obere Pfeile) und im darunter liegenden epithelialen Gewebe (1) der
Abb. zeigen sich cytoplasmatische positive Reaktionen. In der Perimatrix (2) des
Cholesteatoms sind positive Reaktionen (untere Pfeile) zu beobachten.
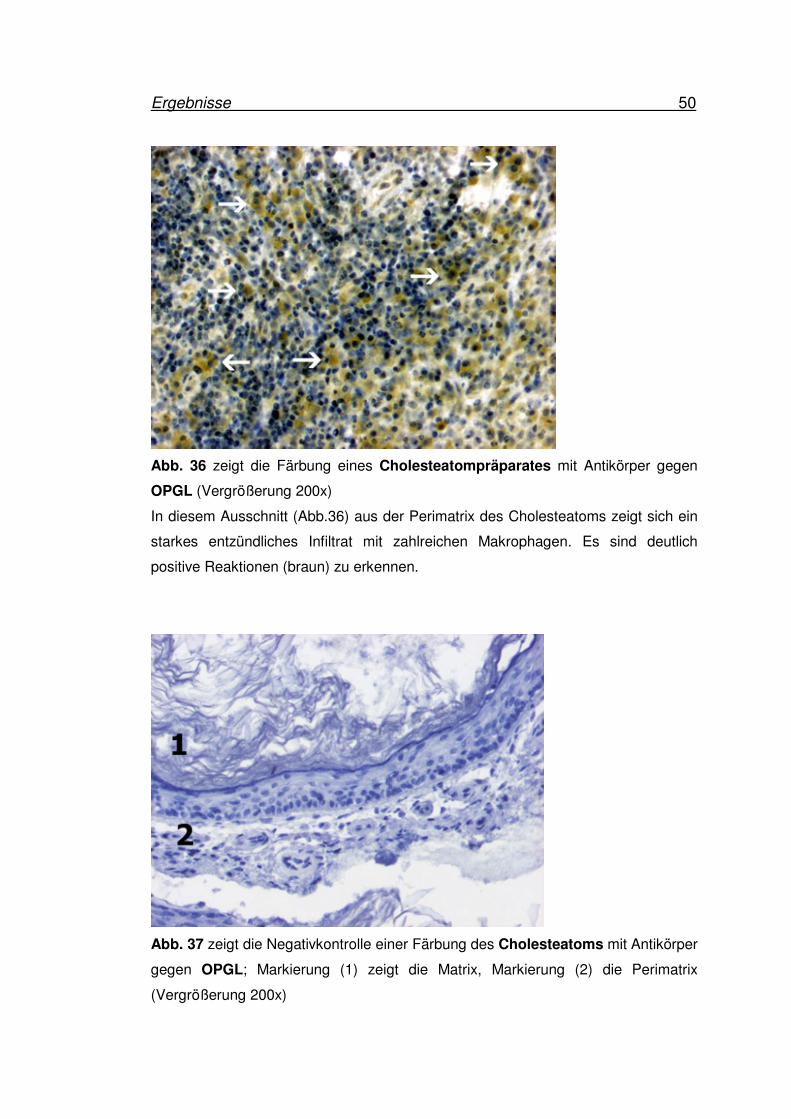
Ergebnisse 50
Abb. 36 zeigt die Färbung eines Cholesteatompräparates mit Antikörper gegen
OPGL (Vergrößerung 200x)
In diesem Ausschnitt (Abb.36) aus der Perimatrix des Cholesteatoms zeigt sich ein
starkes entzündliches Infiltrat mit zahlreichen Makrophagen. Es sind deutlich
positive Reaktionen (braun) zu erkennen.
Abb. 37 zeigt die Negativkontrolle einer Färbung des Cholesteatoms mit Antikörper
gegen OPGL; Markierung (1) zeigt die Matrix, Markierung (2) die Perimatrix
(Vergrößerung 200x)
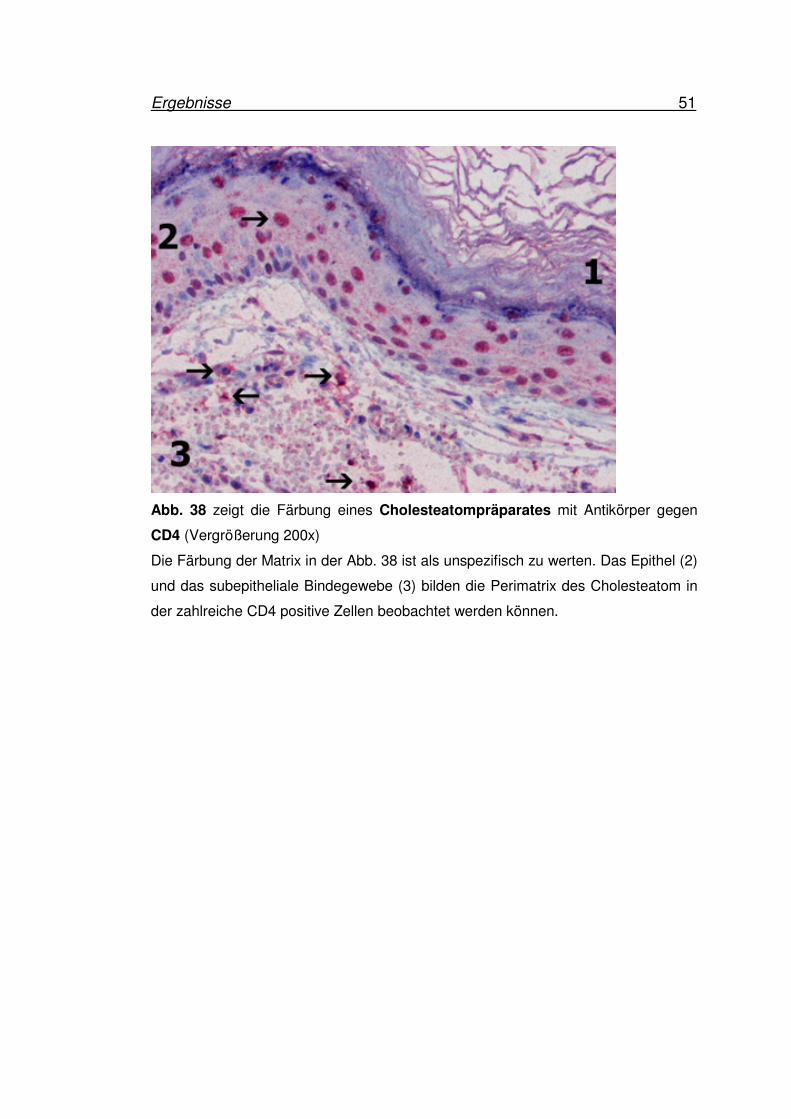
Ergebnisse 51
Abb. 38 zeigt die Färbung eines Cholesteatompräparates mit Antikörper gegen
CD4 (Vergrößerung 200x)
Die Färbung der Matrix in der Abb. 38 ist als unspezifisch zu werten. Das Epithel (2)
und das subepitheliale Bindegewebe (3) bilden die Perimatrix des Cholesteatom in
der zahlreiche CD4 positive Zellen beobachtet werden können.
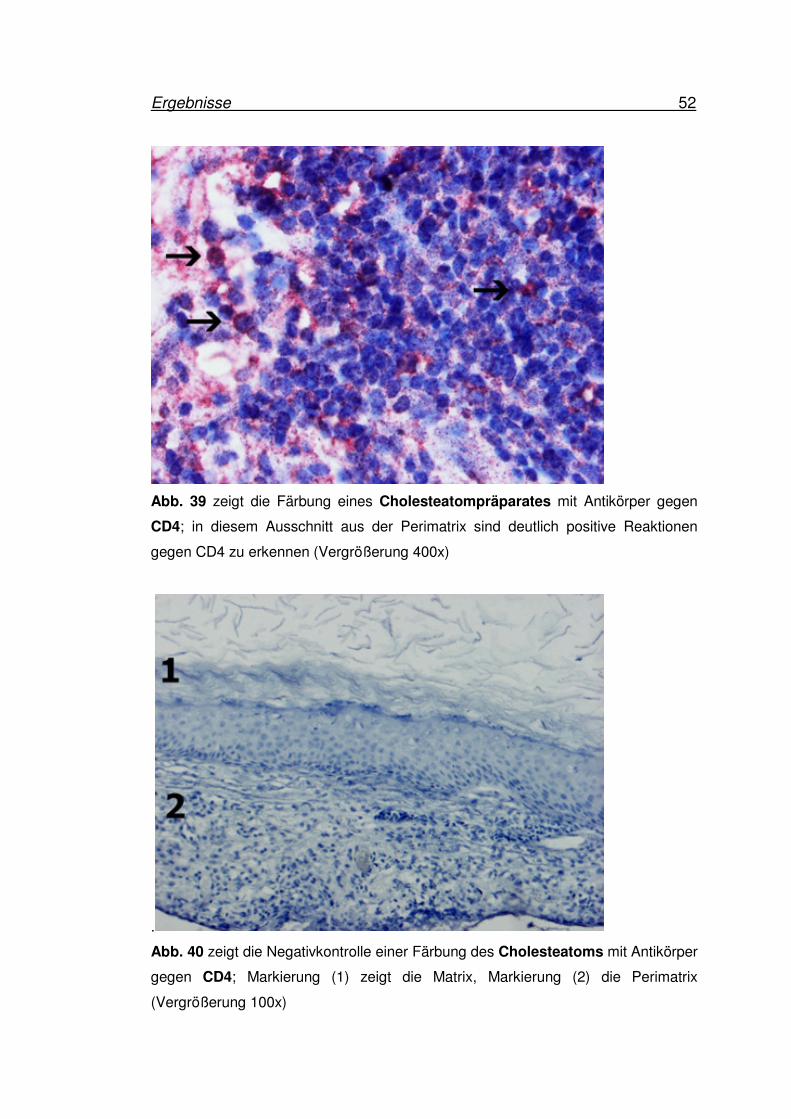
Ergebnisse 52
Abb. 39 zeigt die Färbung eines Cholesteatompräparates mit Antikörper gegen
CD4; in diesem Ausschnitt aus der Perimatrix sind deutlich positive Reaktionen
gegen CD4 zu erkennen (Vergrößerung 400x)
.
Abb. 40 zeigt die Negativkontrolle einer Färbung des Cholesteatoms mit Antikörper
gegen CD4; Markierung (1) zeigt die Matrix, Markierung (2) die Perimatrix
(Vergrößerung 100x)
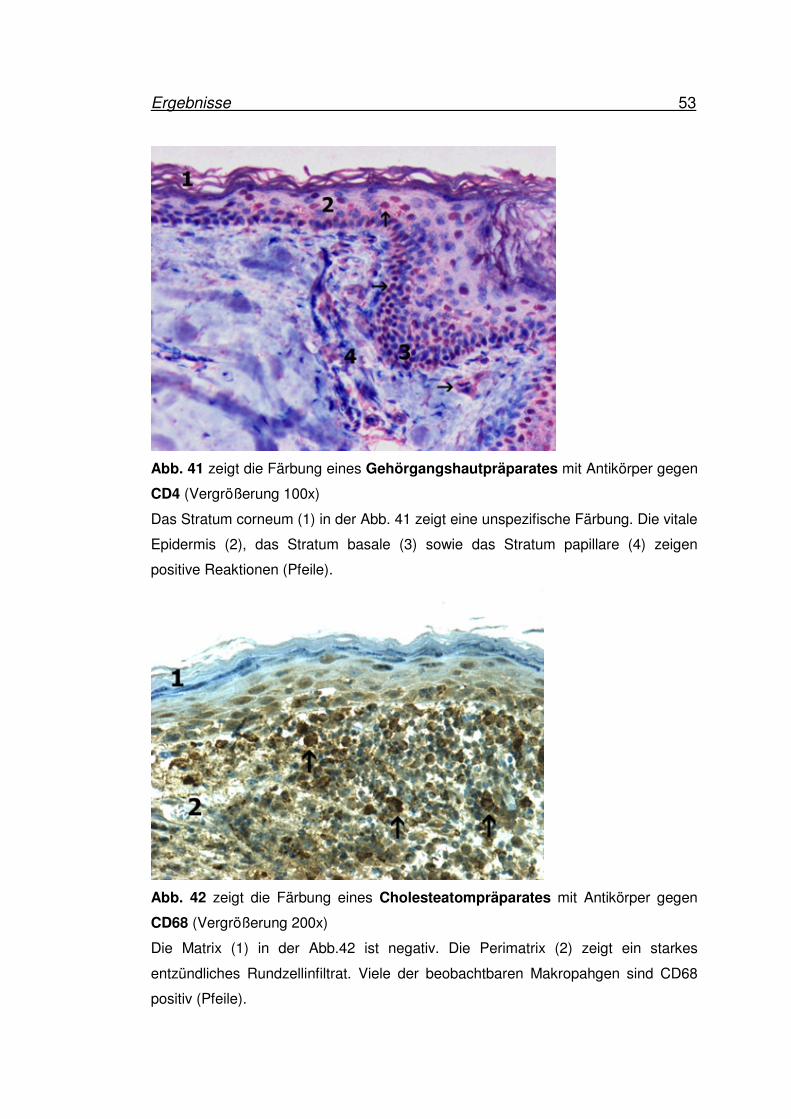
Ergebnisse 53
Abb. 41 zeigt die Färbung eines Gehörgangshautpräparates mit Antikörper gegen
CD4 (Vergrößerung 100x)
Das Stratum corneum (1) in der Abb. 41 zeigt eine unspezifische Färbung. Die vitale
Epidermis (2), das Stratum basale (3) sowie das Stratum papillare (4) zeigen
positive Reaktionen (Pfeile).
Abb. 42 zeigt die Färbung eines Cholesteatompräparates mit Antikörper gegen
CD68 (Vergrößerung 200x)
Die Matrix (1) in der Abb.42 ist negativ. Die Perimatrix (2) zeigt ein starkes
entzündliches Rundzellinfiltrat. Viele der beobachtbaren Makropahgen sind CD68
positiv (Pfeile).
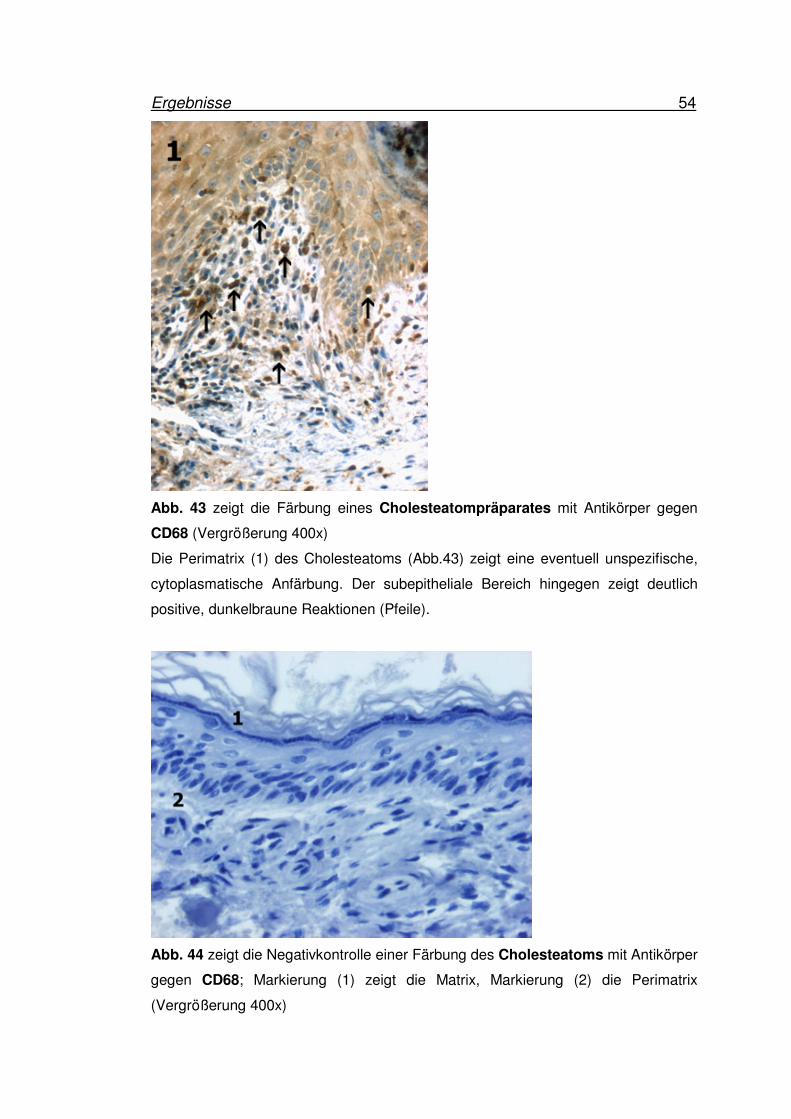
Ergebnisse 54
Abb. 43 zeigt die Färbung eines Cholesteatompräparates mit Antikörper gegen
CD68 (Vergrößerung 400x)
Die Perimatrix (1) des Cholesteatoms (Abb.43) zeigt eine eventuell unspezifische,
cytoplasmatische Anfärbung. Der subepitheliale Bereich hingegen zeigt deutlich
positive, dunkelbraune Reaktionen (Pfeile).
Abb. 44 zeigt die Negativkontrolle einer Färbung des Cholesteatoms mit Antikörper
gegen CD68; Markierung (1) zeigt die Matrix, Markierung (2) die Perimatrix
(Vergrößerung 400x)
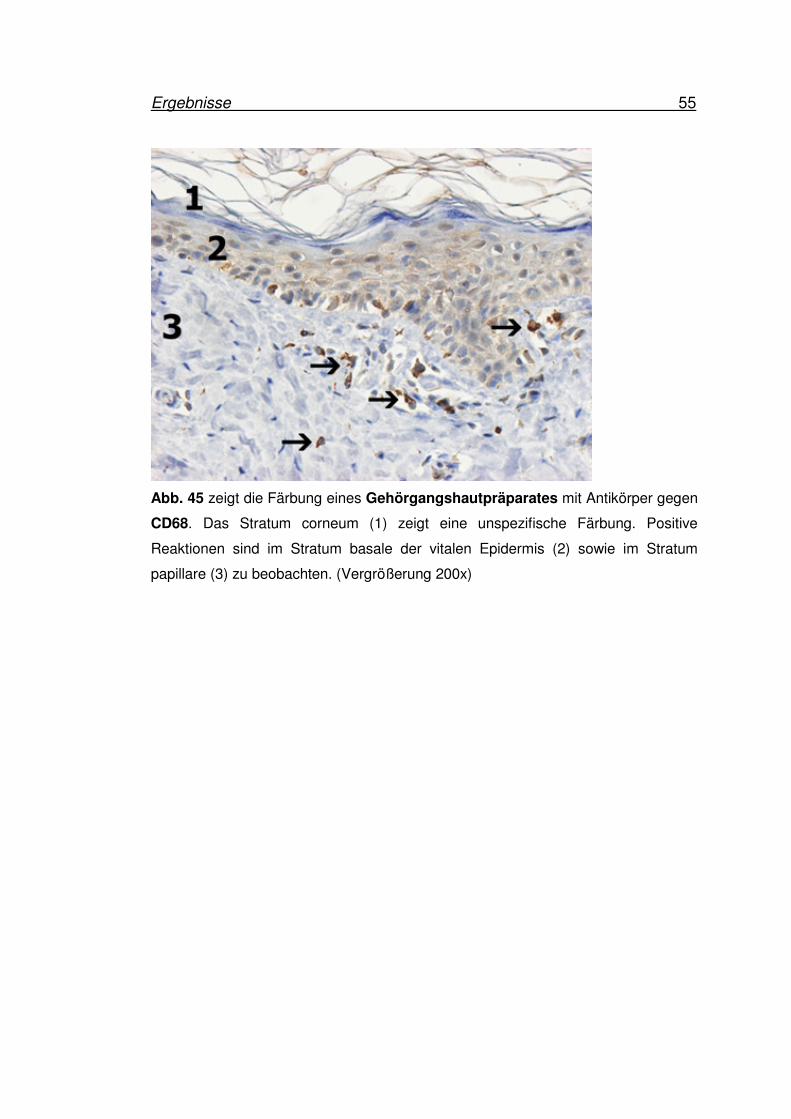
Ergebnisse 55
Abb. 45 zeigt die Färbung eines Gehörgangshautpräparates mit Antikörper gegen
CD68. Das Stratum corneum (1) zeigt eine unspezifische Färbung. Positive
Reaktionen sind im Stratum basale der vitalen Epidermis (2) sowie im Stratum
papillare (3) zu beobachten. (Vergrößerung 200x)
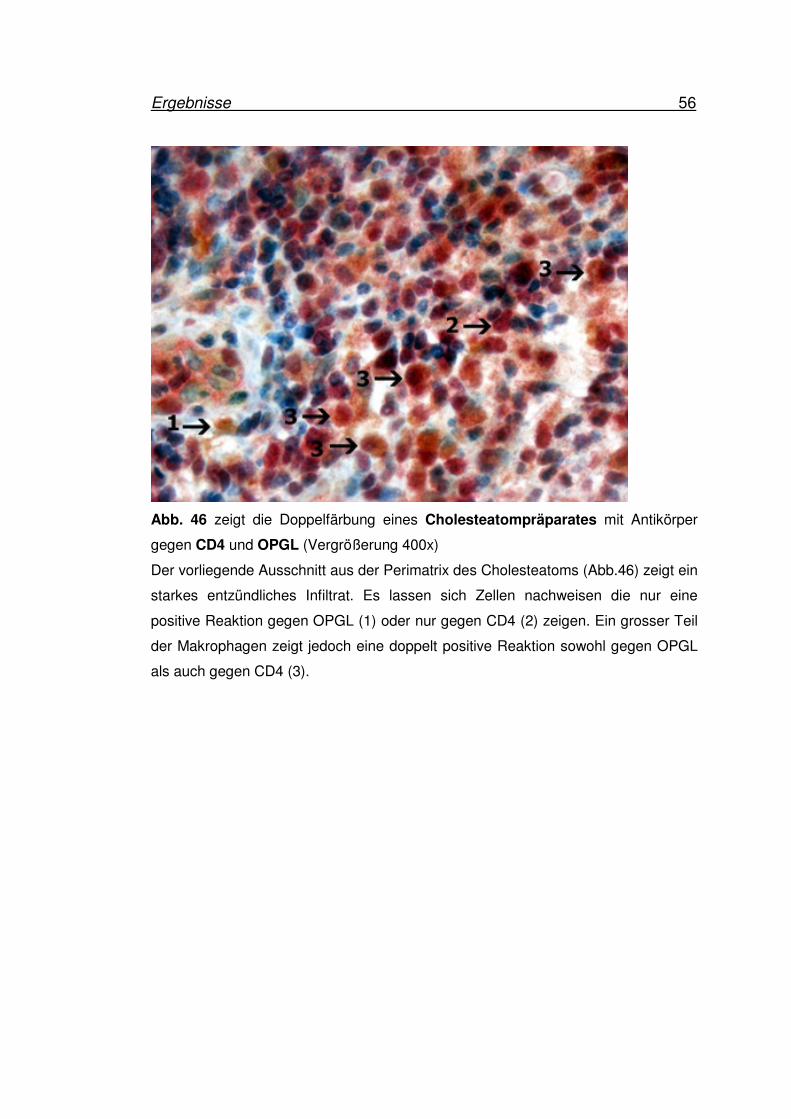
Ergebnisse 56
Abb. 46 zeigt die Doppelfärbung eines Cholesteatompräparates mit Antikörper
gegen CD4 und OPGL (Vergrößerung 400x)
Der vorliegende Ausschnitt aus der Perimatrix des Cholesteatoms (Abb.46) zeigt ein
starkes entzündliches Infiltrat. Es lassen sich Zellen nachweisen die nur eine
positive Reaktion gegen OPGL (1) oder nur gegen CD4 (2) zeigen. Ein grosser Teil
der Makrophagen zeigt jedoch eine doppelt positive Reaktion sowohl gegen OPGL
als auch gegen CD4 (3).
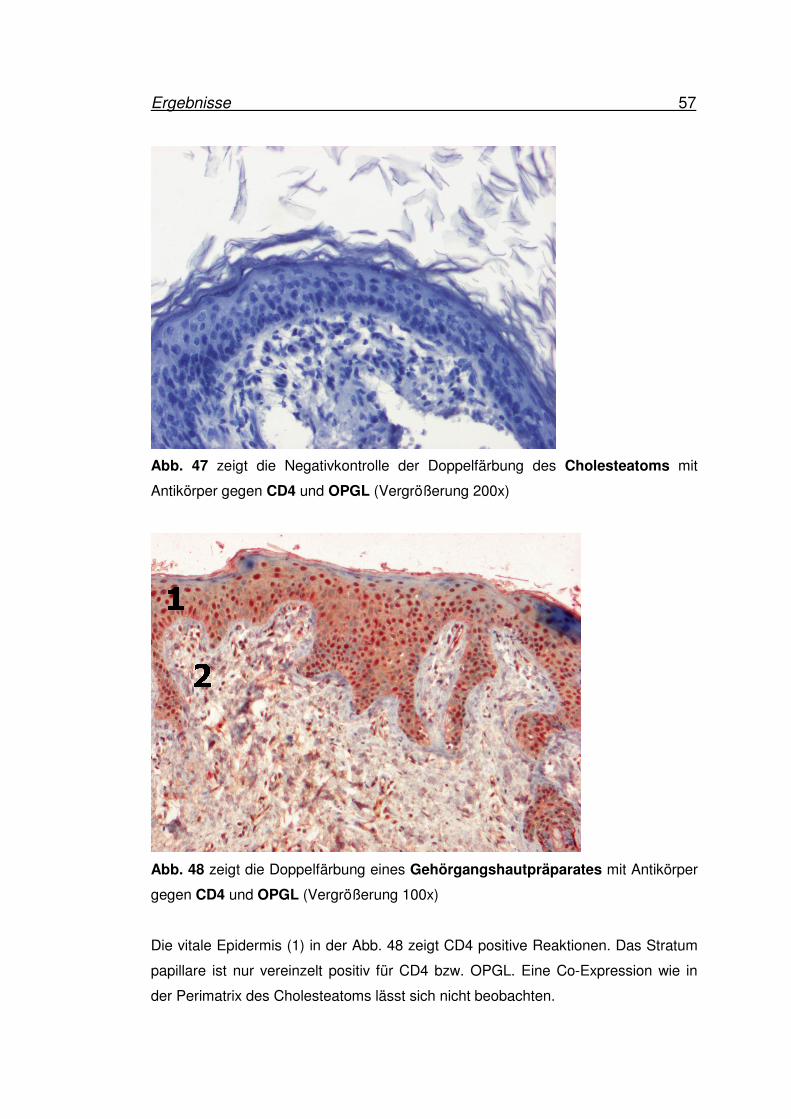
Ergebnisse 57
Abb. 47 zeigt die Negativkontrolle der Doppelfärbung des Cholesteatoms mit
Antikörper gegen CD4 und OPGL (Vergrößerung 200x)
Abb. 48 zeigt die Doppelfärbung eines Gehörgangshautpräparates mit Antikörper
gegen CD4 und OPGL (Vergrößerung 100x)
Die vitale Epidermis (1) in der Abb. 48 zeigt CD4 positive Reaktionen. Das Stratum
papillare ist nur vereinzelt positiv für CD4 bzw. OPGL. Eine Co-Expression wie in
der Perimatrix des Cholesteatoms lässt sich nicht beobachten.

Diskussion 58
4. Diskussion
Die Pathogenese des Cholesteatoms ist seit mehreren Jahren Gegenstand
kontroverser Diskussionen und kann zur Zeit noch nicht als eindeutig aufgeklärt
bezeichnet werden. In Anbetracht der Tatsache, dass ein großer Teil der
chronischen Mittelohrentzündungen Cholesteatome sind oder sich zu solchen
enwickeln, ist es von epidemiologischer Bedeutung durch das Entschlüsseln der
Pathogenese des Cholesteatoms, einschliesslich seiner Komplikationen, mögliche,
die Operation unterstützende Therapieformen zu entwickeln.
Als eine der bekannten Hauptkomplikationen des voranschreitenden Cholesteatoms
kann es zu einer knöchernen Destruktion des Mittel- und Innenohres mit potentiell
lebensbedrohlichen, intracraniellen Komplikationen kommen. Dabei stellt sich die
Frage, wie das Cholesteatom zu einem Knochenabbau führen kann. Der direkte
Mechanismus und seine Steuerung sind ebenfalls Bestandteil aktueller Diskussion.
Hierzu einige Vorbemerkungen:
Knochen Auf-, Ab- und Umbau sind Prozesse, die je nach Ausprägung sowohl
physiologischen als auch pathologischen Charakter annehmen können. Ein
physiologischer Knochen Auf- und Umbau beinhaltet die Synthese der
Knochenmatrix sowohl durch Osteoblasten als auch die Resorption von Knochen
durch Osteoklasten. Kommt es zu einem pathologischen Knochenabbau, ist das
Verhältnis von Synthese der Knochenmatrix durch Osteoblasten zur Resorption des
Knochens durch Osteoklasten zugunsten der Resorptionsvorgänge verschoben. Im
Rahmen des Cholesteatoms lässt sich dann eine knöcherne Destruktion
morphologisch beschreiben, welche die Gehörknöchelchenkette und andere
knöcherner Bestandteile des Mittel- und Innenohres betreffen kann.
Fest steht, dass als direkte nachgewiesene Ursache für die knöchernen
Destruktionen nur eine einzige Zelle im menschlichen Körper verantwortlich
gemacht werden kann: der Osteoklast. Sein genauer Aktivierungs- und
Kontrollmechanismus ist im Rahmen des Cholesteatoms jedoch auch noch nicht
eindeutig geklärt. Neben einer cytokinbedingten Aktivierung von Osteoklasten
wurden in der Vergangenheit mehrere verschiedene Faktoren als Auslöser für die
Aktivierung von Osteoklasten und dem damit verbundenen Knochenabbau diskutiert
(35,47), die im folgenden in groben Zügen dargestellt werden sollen.

Diskussion 59
Bedingt durch den Verlust der Kontaktinhibierung der Keratinozyten des
Trommelfells und des Gehörgangsepithels kommt es zu einer fortschreitenden
Akkumulation von Keratinozyten in eine Retraktionstasche des Trommelfells. Diese
progrediente Ansammlung von Keratinozyten führt zu einer lokalen Expansion des
Cholesteatoms und damit verbunden zu einer Druckerhöhung. Dies führte zu der
Annahme, dass der erhöhte Druck als Auslöser für die Osteoklastenaktivierung
verantwortlich zu machen sei (21,55). Orisek und Chole zeigten, dass der durch das
Cholesteatom bedingte Druckanstieg zwischen 1.3 und 11,9 mm Hg und damit unter
dem kapillären Perfusionsdruck von 25mm Hg liegt (35). Damit stellte sich die
druckbedingte Aktivierung von Osteoklasten als unwahrscheinlich heraus. Durch
eine sekundäre Infektion des Cholesteatoms und des umliegenden Gewebes kann
es zu einer Produktion sowohl bakterieller Endotoxine als auch zu einer Erhöhung
von Enzymen wie z.B. Kollagenasen und sauren Hydrolasen durch das umliegende
entzündliche Gewebe kommen (33). Zelluläre Faktoren wie Cytokine und
Wachstumsfaktoren sind demnach nicht als einziger Auslöser der
Osteoklastenaktivierung zu sehen, scheinen jedoch eine der wichtigsten Rollen zu
übernehmen. Dabei ist der folgende grundsätzliche Ansatz weitläufig akzeptiert:
Durch die Migration von Keratinozyten in eine Retraktionstasche des Trommelfells
kommt es zu einer, das Cholesteatom begleitenden, entzündlichen Reaktion des
subepithelialen Bindegewebes. In der sich bildenden Perimatrix führt dies zu einer
erhöhten Konzentration von Cytokinen, welche die Osteoklasten-Aktivierung und -
Differenzierung entscheidend beeinflussen.
Dies ist insofern von herausragender Bedeutung, da Osteoklasten die einzigen
Zellen im menschlichen Körper sind, welche die Fähigkeit besitzen, Knochen
abzubauen, und es sich hierbei um eine direkten Aktivierungsmechanismus gerade
dieser Zellen handelt. Sie werden direkt für den Knochenabbau im Rahmen des
Cholesteatoms verantwortlich gemacht und stellen ein morphologisches Substrat
dar, dass den beobachteten Knochendestruktionen zugeordnet werden kann.
Es wurde in dieser Arbeit daher ein Augenmerk auf das Vorhandensein der
Osteoklasten- Vorläuferzellen, die aus der Monozytenmakrophagenzelllinie
stammen, sowie auf den Aktivierungsprozeß adulter Osteoklasten gelegt. Beides
sind essentielle Vorraussetzungen bzw. Bestandteile der
Osteoklastendifferenzierung und -steuerung. Hierzu wurden zur genaueren
Beschreibung des Vorhandenseins und der Verteilung von Osteoklasten-

Diskussion 60
Vorläuferzellen, die mit Makrophagen assoziierten Antigene CD4 und CD68
immunhistochemisch untersucht. Es zeigte sich dabei, dass sich beim Cholesteatom
in den untersuchten 15 Präparaten CD4 positive Reaktionen nachweisen ließen
(5% - 75% der Zellen positiv). Die Gehörgangshäute wiesen im Vergleich nur eine
schwach positive Reaktion (unter 5% der Zellen positiv) auf.
Bei dem immunhistochemischen Nachweis von CD68 war der Unterschied zwischen
den Cholesteatom- und den Gehörgangshautpräparaten noch eindeutiger. Im
Vergleich zur Gehörgangshaut ließen sich hier Cholesteatom-Präparate mit stark
positiven Reaktionen (über 75% positive Zellen) beobachten (siehe Abb.42). Damit
konnte das Vorhandensein von Osteoklastenvorläuferzellen eindeutig
nachgewiesen werden. Das Epithel zeigt zwar vereinzelt positive Reaktionen, in der
Mehrheit sind diese jedoch im subepithelialen Bindegewebe zu beobachten (siehe
Abb.43). Dieses Ergebnis war zu erwarten, da das subepitheliale Bindegewebe Ort
des entzündlichen Geschehens ist.
In den untersuchten immunhistochemischen Cholesteatomschnitten ist das
subepitheliale Bindegewebe also der Ort mit dem stärksten entzündlichen Infiltrat.
Dies deckt sich mit den Beobachtungen von Negri et al., die eine verstärkte
Konzentration von aktivierten Makrophagen und T- Zellen im Cholesteatom
beschrieben haben (34).
Makrophagen und aktivierte T-Zellen spielen bekanntlich eine bedeutende Rolle im
Immunsystem des Menschen. Neben der eigenen Körperabwehr nehmen sie auch
eine Schlüsselposition in Entzündungsprozessen ein. Makrophagen haben dabei
vielfältige Funktionen, wie z.B. einen großen Teil der immunologischen Antwort auf
fremde Substanzen, die Produktion von Cytokinen im Rahmen der Entzündung,
sowie die Phagozytose und die anschließende Präsentation von Antigenen.
Dadurch werden sie zu zentralen Zellen der immunologischen Antwort auf
eingewanderte Keratinozyten im Mittelohr (52).
Die immunhistochemische Untersuchung zeigt, dass es sich aufgrund der stärkeren
Anfärbung der makrophagenassoziierten Antigene CD4 und CD68 beim
Cholesteatom um einen Entzündungsprozess mit einer eindrucksvollen Erhöhung
der Makrophagendichte handelt. Da Makrophagen die Osteoklasten-Vorläuferzellen
darstellen, lässt sich aufgrund der Ergebnisse im Cholesteatom eine erhöhte Anzahl
von Osteoklasten erwarten. Darüber lässt sich jedoch, bedingt durch die

Diskussion 61
verwendeten Antikörper, keine Aussage treffen, da CD4 und CD68 keine
spezifischen Osteoklasten-Marker sind.
Sowohl unter physiologischen, als auch unter pathologischen Bedingungen, sind für
die Osteoklastogenese einige wichtige Voraussetzungen zu erfüllen. Eine dieser
wichtigen Voraussetzungen ist die Zell-Zell-Interaktion zwischen den
Bindegewebszellen beziehungsweise Osteoblasten und den Osteoklasten-
Vorläuferzellen, die zu der sogenannten parakrinen Stimulation führt (12). Die Liste
der Osteoklasten-steuernden Faktoren ist lang. Man zählt neben Prostaglandinen,
Glucokortikoiden, Parathormon, Vitamin D, Endothelinen, Il-1, Il-6, Hepatocyte
Growth Factor (HGF), Tumor Nekrose Faktor (TNF), Transformig Growth Factor β
(TGFβ) auch das in dieser Arbeit untersuchte Osteoprotegerinligand (OPGL) und
sein Rezeptor Osteoprotegerin (OPG) dazu (12).
Osteoblasten und T-Zellen antworten auf die entzündungsspezifischen Zytokine (Il-
1, Il-1l, Il-6, TNF-α, TNF-β, Interferon-γ) mit einer verstärkten Expression von
Osteoprotegerinligand (OPGL), eines membranständigen Proteins aus der Tumor-
Nekrose- Faktor-Familie (TNF), dass im Rahmen des Stoffwechsels von
Osteoklasten zu den Faktoren mit dem stärksten aktivierenden Potential zählt und
als essentiell für die Osteoklastogenese einzustufen ist (24). OPGL ist sowohl ein
Schlüsselmolekül im Rahmen der Differenzierung von Osteoklasten-Vorläuferzellen
(Makrophagen) zu Osteoklasten, als auch in der Aktivierung von adulten
Osteoklasten. Um den Aktivierungsprozeß adulter Osteoklasten zu untersuchen,
wurde hierzu der immunhistochemische Nachweis dieses Schlüsselmoleküls der
Osteoklastogenese erbracht.
Es ließ sich im Cholesteatom eine stark positive Reaktion von OPGL sowohl in der
Matrix, als auch in der entzündlichen Perimatrix beobachten. In der Gehörgangshaut
zeigte sich hingegen nur eine schwach positive Reaktion. Diese Ergebnisse der
Immunhistochemie decken sich mit den molekularbiologischen Nachweisen von
OPGL. Der molekularbiologische Nachweis der Expression von OPGL wurde
ebenfalls sowohl im Cholesteatom, als auch in der Gehörgangshaut vorgenommen.
In allen acht untersuchten Cholesteatomen konnte dabei ein eindeutiger Nachweis
der Expression von OPGL geführt werden. In Anbetracht der Tatsache, dass es sich
beim Cholesteatom um ein stark entzündliches Gewebe handelt, wurde dieses
Ergebnis aufgrund der damit verbundenen erhöhten T-Zell Konzentration auch
erwartet.

Diskussion 62
Anders hingegen sieht es in den untersuchten Proben der Gehörgangshaut aus.
Hier konnte nur bei einer von fünf untersuchten Gehörgangshäuten eine eindeutig
positive (Gehörgangshaut 2) bzw. in einer weiteren Probe nur ein schwach positives
Ergebnis (Gehörgangshaut 3) des Nachweises der Expression von OPGL erzielt
werden.
Der sich immunhistochemisch als schwach und molekularbiologisch als schwierig
herausgestellte Nachweis von OPGL in der Gehörgangshaut ist mit dem Fehlen
einer entzündungsbedingten Aktivierung der Expression von OPGL in Osteoblasten
und in aktivierten T-Zellen zu erklären. Unter regelrechten physiologischen
Bedingungen kommt es daher nur zur einer geringen Freisetzung von OPGL in der
Gehörgangshaut.
Die Ergebnisse der Doppelfärbung von CD4 und OPGL bestätigen, dass OPGL von
den CD4 positiven T-Helfer Zellen in der Perimatrix des Cholesteatoms produziert
wird. Damit sind in der Perimatrix des Cholesteatoms gute Vorraussetzungen für die
entzündungsbedingte verstärkte Aktivierung von adulten Osteoklasten bzw. für die
Differenzierung von Makrophagen zu multinukleären Osteoklasten gegeben. Hiermit
ist ein direkter Zusammenhang zwischen entzündungsbedingter Aktivierung und
potentieller Knochendestruktion durch Osteoklasten gegeben. Es wurde damit
erstmalig eine Co-Expression von CD4 und OPGL in der Perimatrix des
Cholesteatoms beschrieben. Es ist aufgrund dieser Beobachtungen nachgewiesen,
dass es im Cholesteatom zu einer verstärkten OPGL Expression durch aktivierte T-
Zellen kommt. Das Konzept einer tumor- oder entzündungsbedingten Aktivierung
von adulten Osteoklasten bzw. Differenzierung von Makrophagen zu Osteoklasten
wurde bereits für andere Erkrankungen, die zu Knochenabbau führen können,
entwickelt und ist hiermit in Einklang zu bringen (37). Zusammenfassend lässt sich
zu OPGL sagen, dass das Vorhandensein im Cholesteatom einen wichtigen
Hinweis für die Osteoklasten-Aktivierung und der damit einhergehenden
knochendestruierenden Potenz des Cholesteatoms liefert. Dies kann ein wichtiger
Ansatzpunkt weiterer Untersuchungen zum Knochenabbau im Cholesteatom sein.
Osteoprotegerin (OPG) ist ein löslicher TNF-Rezeptor und physiologischer Inhibitor
der Knochenresoprtion und wird auch als „osteoclastogenesis inhibitory factor„
(OCIF) bezeichnet.

Diskussion 63
Die Untersuchungen des Osteoprotegerin (OPG), dem Gegenspieler von
Osteoprotegerinligand (OPGL), kommen zu folgenden Ergebnissen:
Der Nachweis der Expression von OPG mit Hilfe der RT-PCR war in allen
untersuchten Cholesteatomen und Gehörgangshäuten positiv.
Diese gewonnenen Ergebnisse decken sich mit den Ergebnissen der
immmunhistochemischen Untersuchung. OPG kann sowohl im Cholesteatom als
auch in der Gehörgangshaut eindeutig nachgewiesen werden. OPG lässt sich
immunhistochemisch in der Gehörgangshaut jedoch stärker anfärben als im
Cholesteatom. Das Osteoklasten aktivierende OPGL hingegen zeigte sich im
Cholesteatom stärker positiv als in den untersuchten Gehörgangshäuten. Dies
untersützt die These, dass das entzündliche Infiltrat in der Perimatrix des
Cholesteatoms zu einer verstärkten Produktion von OPGL führt, und es gleichzeitig
zu einer verringerten Expression von OPG kommt.
Durch das Verwenden einer hochsensitiven und spezifischen Methode, der
Polymerase-Ketten-Reaktion, kann den Ergebnissen der Untersuchung der
Expression von OPG und OPGL eine hohe Validität zugesprochen werden. Ein
falsch positives Ergebnis durch eine Kontamination mit Fremd-DNA ist prinzipiell
möglich, aber als sehr unwahrscheinlich einzuschätzen. Dieselbe Aussage gilt auch
für falsch negative Ergebnisse, da es mit Hilfe der PCR möglich ist, bereits aus einer
sehr geringen Menge Ausgangs-DNA eine Aussage über die Expression eines
Faktors zu machen. Nach den erfolgten qualitativen Nachweisen sowohl im
Cholesteatom als auch in der Gehörgangshaut kann man in einem quantitativen
Verfahren z.B. durch eine quantitative PCR oder Northern-Blot-Experiment die
Stärke der Expression von OPG und OPGL vergleichen.
Letztendlich ist das progressive Wachstumsverhalten des Cholesteatoms mit den
beschriebenen Knochenarrosionen sicherlich ein multifaktorielles Geschehen und
nicht einzig und allein mit dem Vorhandensein der beobachteten erhöhten
Makrophagen-Dichte oder Osteoklasten-differenzierender Zytokine zu erklären.
Dennoch zeigen sich hier wichtige Hinweise auf ein von Kong enwickeltes Konzept
der Knochendestruktion, dass uns von der entzündlichen Polyarthritis bekannt ist
(23) :

Diskussion 64
Eine das Cholesteatom begleitende, entzündliche Reaktion führt zu einer
verstärkten Produktion von OPGL durch entzündungsbedingt aktivierte T-Zellen.
Diese Reaktion ist mitverantwortlich für eine verstärkte Osteoklastogenese und
einem Knochenabbau durch Osteoklasten. Kong et al. haben weiterhin gezeigt,
dass Mäuse mit einem ausgeschalteten OPGL-Gen eine starke Osteopetrose sowie
Störungen der Odontogenese zeigen. Daher kann eine weitere Erforschung des
Osteoklasten aktivierenden Systems zur Entwicklung einer unterstützenden
medikamentösen Therapie in Form von Osteoklasten hemmenden Substanzen z.B.
in Form eines OPGL Antagonisten führen. Neben der Operation würde dies zu einer
ergänzenden möglichen Therapieform führen, welche eventuell zu einer Hemmung
der Osteoklastenproliferation und damit des Knochenabbaus im Rahmen des
Cholesteatoms führt.

Zusammenfassung 65
5. Zusammenfassung
Das Cholesteatom ist ein chronisch entzündlicher Prozeß im Mittelohr. Eine
begleitende, entzündungsbedingte Aktivierung von Osteoklasten kann zu einer
Resorption von Gehörknöchelchen und knöchernen Bestandteilen des Mittelohres
mit zum Teil lebensbedrohlichen Komplikationen führen. Der Prozess der
Knochenresorption ist noch nicht eindeutig geklärt. Ein Hauptdifferenzierungs- und
Aktivierungsfaktor der Osteoklasten, die für den Knochenabbau verantwortlich sind,
ist das Osteprotegerin-Ligand (OPGL). OPGL ist ein von aktivierten T-Zellen und
Osteoblasten exprimiertes, membranständiges Molekül, was sowohl eine
stimulierende Wirkung auf die Osteoklastogenese als auch auf die
Knochenresorptionsaktivität von Osteoklasten besitzt. Sein Gegenspieler ist das
Osteoprotegerin (OPG), ein nicht membranständiges, ebenfalls von Osteoblasten
und aktivierten T-Zellen produziertes Molekül. Es bindet sich an OPGL und hemmt
damit die Aktivierung der Osteoklasten. Sowohl OPG als auch OPGL konnten in der
Gehörgangshaut und im Cholesteatomgewebe mit Hilfe einer RT-PCR
nachgewiesen werden. Damit ist nach dem erfolgten qualitativen Nachweis dieser
beiden wichtigen Faktoren die Grundlage für weitere quantitative
Nachweisverfahren gegeben.
In den immunhistochemischen Untersuchungen zeigte sich in der Perimatrix des
Cholesteatoms eine erhöhte Anfärbbarkeit sowohl für OPGL als auch für die
makrophagenassoziierten Antigene CD4 und CD68. Die Makrophagendichte im
Cholesteatom ist im Vergleich zur Gehörgangshaut deutlich erhöht. OPG hingegen
war im Cholesteatom schwächer anfärbbar als in der Gehörgangshaut.
In einer immunhistochemischen Doppelfärbung zeigt sich sowohl in der Matrix als
auch in der Perimatrix der Cholesteatomproben ein Vorhandensein von gleichzeitig
CD4 und OPGL positiven Zellen. Dies deutet auf eine Co-Expression von CD4 und
OPGL hin. Verantwortlich dafür sind durch die Entzündung aktivierte T-Zellen, die
mit einer verstärkten Produktion von OPGL auf die Entzündung reagieren. Eine
erhöhte OPGL-Konzentration kann dann in einer ausgeprägten Aktivierung von
Osteoklasten resultieren, was einen verstärkten Knochenabbau zur Folge haben
kann. Die Ergebnisse der Arbeit unterstützen ein Konzept, dass bereits für die
entzündliche Arthritis formuliert wurde:

Zusammenfassung 66
Die das Cholesteatom begleitende Entzündung führt zu einer erhöhten Freisetzung
von OPGL durch aktivierte T-Zellen. Dies ist ein wichtiger Faktor in der Aktivierung
und Differenzierung von Osteoklasten. Daher könnte neben der Operation die
medikamentöse Inhibierung von OPGL zu einer möglichen begleitenden
Therapieform führen.

Anhang 67
6.Anhang Im folgenden sind die mRNA-Sequenzen von humanem OPGL und OPG
dargestellt. Die für die PCR verwendeten Primersequenzen sind markiert.
Quelle : National Center for Biotechnology Information (NCBI)
humanes Osteoprotegerin Ligand (OPGL):
-Anfang-
1 aagcttggta ccgagctcgg atccactact cgacccacgc gtccgcgcgc cccaggagcc
61 aaagccgggc tccaagtcgg cgccccacgt cgaggctccg ccgcagcctc cggagttggc
121 cgcagacaag aaggggaggg agcgggagag ggaggagagc tccgaagcga gagggccgag
181 cgccatgcgc cgcgccagca gagactacac caagtacctg cgtggctcgg aggagatggg
241 cggcggcccc ggagccccgc acgagggccc cctgcacgcc ccgccgccgc ctgcgccgca
301 ccagcccccc gccgcctccc gctccatgtt cgtggccctc ctggggctgg ggctgggcca
361 ggttgtctgc agcgtcgccc tgttcttcta tttcagagcg cagatggatc ctaatagaat
421 atcagaagat ggcactcact gcatttatag aattttgaga ctccatgaaa atgcagattt
481 tcaagacaca actctggaga gtcaagatac aaaattaata cctgattcat gtaggagaat
541 taaacaggcc tttcaaggag ctgtgcaaaa ggaattacaa catatcgttg gatcacagca
601 catcagagca gagaaagcga tggtggatgg ctcatggtta gatctggcca agaggagcaa
661 gcttgaagct cagccttttg ctcatctcac tattaatgcc accgacatcc catctggttc
721 ccataaagtg agtctgtcct cttggtacca tgatcggggt tgggccaaga tctccaacat
781 gacttttagc aatggaaaac taatagttaa tcaggatggc ttttattacc tgtatgccaa
841 catttgcttt cgacatcatg aaacttcagg agacctagct acagagtatc ttcaactaat
901 ggtgtacgtc actaaaacca gcatcaaaat cccaagttct cataccctga tgaaaggagg
961 aagcaccaag tattggtcag ggaattctga attccatttt tattccataa acgttggtgg
1021 attttttaag ttacggtctg gagaggaaat cagcatcgag gtctccaacc cctccttact
1081 ggatccggat caggatgcaa catactttgg ggcttttaaa gttcgagata tagattgagc
1141 cccagttttt ggagtgttat gtatttcctg gatgtttgga aacatttttt aaaacaagcc
1201 aagaaagatg tatataggtg tgtgagacta ctaagaggca tggccccaac ggtacacgac
1261 tcagtatcca tgctcttgac cttgtagaga acacgcgtat ttacagccag tgggagatgt
1321 tagactcatg gtgtgttaca caatggtttt taaattttgt aatgaattcc tagaattaaa
1381 ccagattgga gcaattacgg gttgacctta tgagaaactg catgtgggct atgggagggg
1441 ttggtccctg gtcatgtgcc ccttcgcagc tgaagtggag agggtgtcat ctagcgcaat
1501 tgaaggatca tctgaagggg caaattcttt tgaattgtta catcatgctg gaacctgcaa
1561 aaaatacttt ttctaatgag gagagaaaat atatgtattt ttatataata tctaaagtta

Anhang 68
1621 tatttcagat gtaatgtttt ctttgcaaag tattgtaaat tatatttgtg ctatagtatt
1681 tgattcaaaa tatttaaaaa tgtcttgctg ttgacatatt taatgtttta aatgtacaga
1741 catatttaac tggtgcactt tgtaaattcc ctggggaaaa cttgcagcta aggaggggaa
1801 aaaaatgttg tttcctaata tcaaatgcag tatatttctt cgttcttttt aagttaatag
1861 attttttcag acttgtcaag cctgtgcaaa aaaattaaaa tggatgcctt gaataataag
1921 caggatgttg gccaccaggt gcctttcaaa tttagaaact aattgacttt agaaagctga
1981 cattgccaaa aaggatacat aatgggccac tgaaatctgt caagagtagt tatataattg
2041 ttgaacaggt gtttttccac aagtgccgca aattgtacct tttttttttt ttcaaaatag
2101 aaaagttatt agtggtttat cagcaaaaaa gtccaatttt aatttagtaa atgttatctt
2161 atactgtaca ataaaaacat tgcctttgaa tgttaatttt ttggtacaaa aataaattta
2221 tatgaaaaaa aaaaaaaaag ggcggccgct ctagagggcc ctattctata g
-Ende-
humanes Osteoprotegerin (OPG) :
-Anfang-
1 gtatatataa cgtgatgagc gtacgggtgc ggagacgcac cggagcgctc gcccagccgc
61 cgyctccaag cccctgaggt ttccggggac cacaatgaac aagttgctgt gctgcgcgct
121 cgtgtttctg gacatctcca ttaagtggac cacccaggaa acgtttcctc caaagtacct
181 tcattatgac gaagaaacct ctcatcagct gttgtgtgac aaatgtcctc ctggtaccta
241 cctaaaacaa cactgtacag caaagtggaa gaccgtgtgc gccccttgcc ctgaccacta
301 ctacacagac agctggcaca ccagtgacga gtgtctatac tgcagccccg tgtgcaagga
361 gctgcagtac gtcaagcagg agtgcaatcg cacccacaac cgcgtgtgcg aatgcaagga
421 agggcgctac cttgagatag agttctgctt gaaacatagg agctgccctc ctggatttgg
481 agtggtgcaa gctggaaccc cagagcgaaa tacagtttgc aaaagatgtc cagatgggtt
541 cttctcaaat gagacgtcat ctaaagcacc ctgtagaaaa cacacaaatt gcagtgtctt
601 tggtctcctg ctaactcaga aaggaaatgc aacacacgac aacatatgtt ccggaaacag
661 tgaatcaact caaaaatgtg gaatagatgt taccctgtgt gaggaggcat tcttcaggtt
721 tgctgttcct acaaagttta cgcctaactg gcttagtgtc ttggtagaca atttgcctgg
781 caccaaagta aacgcagaga gtgtagagag gataaaacgg caacacagct cacaagaaca
841 gactttccag ctgctgaagt tatggaaaca tcaaaacaaa gcccaagata tagtcaagaa
901 gatcatccaa gatattgacc tctgtgaaaa cagcgtgcag cggcacattg gacatgctaa
961 cctcaccttc gagcagcttc gtagcttgat ggaaagctta ccgggaaaga aagtgggagc
1021 agaagacatt gaaaaaacaa taaaggcatg caaacccagt gaccagatcc tgaagctgct
1081 cagtttgtgg cgaataaaaa atggcgacca agacaccttg aagggcctaa tgcacgcact

Anhang 69
1141 aaagcactca aagacgtacc actttcccaa aactgtcact cagagtctaa agaagaccat
1201 caggttcctt cacagcttca caatgtacaa attgtatcag aagttatttt tagaaatgat
1261 aggtaaccag gtccaatcag taaaaataag ctgcttataa ctggaaatgg ccattgagct
1321 gtttcctcac aattggcgag atcccatgga tgataa -Ende-

Literatur 70
7. Literatur 1) Albino, AP, Kimmelman CP, Parisier SC. Cholesteatoma: a molecular and cellular puzzle. Am J Otol 1998, 19(1), 7-19. 2) Allison, JP, Havran WL. The immunbiology of T cells with invariant gamma delta antigen receptors. Ann Rev Immunol 1991, 9, 679-705. 3) Amling, M, Delling G. Osteoklastenbiologie Bedeutung im Rahmen von Knochenmetastasen. Orthopäde 1998, 27, 214-223. 4) Arthos, J, Deen KC, Chaikin MA, Fornwald JA, Sathe G, Sattentau QJ, Clapham PR, Weiss RA, McDougal JS, Pietropaolo C. Identification of the residues in human CD4 critical for the binding of HIV. Cell 1989, 57, 469-481. 5) Canalis, RF, Lambert PR. The Ear Comprehensive Otology. Philadelphia, Lippincott Wiliams & Wilkins, 1999. 6) Chambers, TJ. Regulation of the differentiation and function of osteoclasts. J Pathol 2000, 192(1), 4-13. 7) Ehrich, EW, Devaux B, Rock EP, Jorgensen JL, Davis MN, Chien YH. T cell receptor interaction with peptide/major histocompatibility complex (MHC) and superantigen/MHC ligands is dominated by antigen. J Exp Med 1993, 178, 713-722. 8) Fixe, P, Praloran V. M-CSF: haematopoietic growth factor or inflammatory cytokine? Cytokin 1998, 10(1), 32-7. 9) Flanagan, AM, Lader CS. Update on the biologic effects of macrophage colony-stimulating factor. Curr Opin Hematol 1998, 5(3), 181-185. 10) Friedmann, I, Arnold W. Pathology of the Ear. London, Churchill & Livingstone, 1993. 11) Fukuda, M. Lysosomal membrane glycoproteins. Structure, biosynthesis, and intracellular trafficking. J Biol Chem 1991, 266, 21327-21330.

Literatur 71
12) Greenfield, EM, Bi Y, Miyauchi A. Regulation of osteoclast activity. Life sci 1999, 65(11), 1087-1102. 13) Hall, TJ, Chambers TJ. Molecular aspects of osteoclast function. Inflamm Res 1996, 45(1), 1-9. 14) Hayashi, S, Yamane T, Miyamoto A, Hemmi H, Tagaya H, Tanio Y, Kanda H, Yamazaki H, Kunisada T. Commitment and differentiation of stem cells to the osteoclast lineage. Biochem Cell Biol 1998, 76(6), 911-22. 15) Healy, D, Dianda L, Moore JP, McDougal JS, Moore MJ, Estess P, Buck D, Kwong PD, Beverley PC, Sattentau QJ. Novel anti-CD4 monoclonal antibodies separate human immunodeficiency virus infection and fusion of CD4+ cells from virus binding. J Exp Med 1990, 172, 1233-1242. 16) Hofbauer, LC, Heufelder AE. Role of receptor activator of nuclear factor-kappaB ligand and osteoprotegerin in bone cell biology. J Mol Med 2001, 79(5/6), 243-253. 17) Holness, CL, Simmons DL. Molecular cloning of CD68, a human macrophage marker related to lysosomal glycoproteins. Blood 1993, 81, 1607-1613. 18) Ingalls, RR, Heine H, Lien E, Yoshimura A, Golenbock D. Lipopolysaccharide Recognition, CD14, and lipopolysaccharide receptors. Infect Dis Clin North Am 1999, 13(2), 341-53. 19) Janeway, CA. The T cell receptor as a multicomponent signaling machine: CD4/CD8 coreceptors and CD45 in T cell activation. Ann Rev Immunol 1992, 10, 645-674. 20) Julius, M, Maroun CR, Haughn L. Distinct roles for CD4 and CD68 as co-receptors in antigen receptor signaling. Immunol Today 1993, 14, 177-183. 21) Kirchner The pathology in cholesteatoma. Proc R Soc Med 1930, 2, 332. 22) Kojima, H, Miyazaki H, Shiwa M, Tanaka Y, Moriyama H. Molecular biological diagnosis of congenital and acquired cholesteatoma on the basis of differences in telomere length. Laryngoscope 2001, 111(5), 867-73.

Literatur 72
23) Kong, YY, Penninger JM. Molecular control of bone remodeling and osteoporosis. Exp Gerontol 2000, 35(8), 947-56. 24) Kong, YY, Yoshida H, Sarosi I, Tan HL, Timms E, Capparelli C, Morony S, Oliveira-dos-Santos AJ, Van G, Itie A, Khoo W, Wakeham A, Dunstan CR, Lacey DL, Mak TW, Boyle WJ, Penninger JM. OPGL is a key regulator of osteoclastogenesis, lymphocyte development and lymph-node organogenesis. Nature 1999, 397(6717), 315-23. 25) Kostenuik, PJ, Shalhoub V. Osteoprotegerin: a physiological and pharmacological inhibitor of bone resorption Curr Pharm Des 2001, 7(8), 613-35. 26) Krull, G. Vorbehandlung von Paraffinschnitten in der Immunhistologie. Gerhard-Domagk-Institut für Pathologie, Domagkstr. 17, 41849 Münster. 27) Lacey, DL, Timms E, Tan HL, Kelley MJ, Dunstan CR, Burgess T, Elliott R, Colombero A, Elliott G, Scully S, Hsu H, Sullivan J, Hawkins N, Davy E, Capparelli C, Eli A, Qian YX, Kaufman S, Sarosi I, Shalhoub V, Senaldi G, Guo J, Delaney J, Boyle WJ. Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell 1998, 93(2), 165-76. 28) Lees, RL, Heersche JN. Macrophage colony stimulating factor increases bone resorption in dispersed osteoclast cultures by increasing osteoclast size. J Bone Miner Res 1999, 14(6), 937-45. 29) Lerner, UH. Osteoclast formation and resorption. Matrix Biol 2000, 19(2), 107-20. 30) Maddon, PJ, Molineaux SM, Maddon DE, Zimmermann KA, Godfrey M, Alt FW, Chess L, Axel R. Structure and expression of human and mouse T4 genes. Proc Natl Acad Sci USA 1987, 84, 9155-9159 31) Martinez-Pomares, L, Platt N, McKnight AJ, da Silva RP, Gordon S. Macrophage membrane molecules: markers of tissue differentiation and heterogenity. Immunobiology 1996, 195(4-5), 407-16. 32) Michaels, L. An epidermoid formation in the developing middle ear: possible source of cholesteatoma. J Otolaryngolog 1986, 15, 169-74.

Literatur 73
33) Moriyama, H. Bone resorption in choilesteatoma: epithelial mesenchymal cell interaction and collagenase production. Laryngoscope 1987, 97, 854. 34) Negri, B, Schilling V, Bujia J, Schulz P, Kastenbauer E. Immunotype findings in macrophages and aural cholesteatomas. Eur Arch Otorhinolaryngol 1992, 249, 87-90. 35) Orisek, B, Chole RA. Pressures exerted by experimental cholesteatomas. Arch Otolaryngol 1987, 113, 386. 36) Pulford, KA, Sipos A, Cordell JL, Stross WP, Mason DY. Distribution of the CD68 macrophage/myeloid associated antigen. Int Immunol 1990, 2, 973-980. 37) Quinn, J, McGee J, Athanasou N. Human tumour-associated macrophages differentiate into osteoclastic bone-resorbing cells. J Pathol 1998, 184, 31-36. 38) Ramprasad, MP, Fischer W, Witztum JL, Sambrano GR, Quehenberger O, Steinberg D. The 94- to 97-kDa mouse macrophage membrane protein that recognizes oxidized low density lipoprotein and phosphatidylserine-rich liposomes is identical to macrosialin, the mouse homologue of human CD68. Proc Natl Acad Sci USA 1995, 92, 9580-9584. 39) Ramprasad, MP, Terpstra V, Kondratenko N, Quehenberger O, Steinberg D. Cell surface expression of mouse macrosialin and human CD68 and their role as macrophage receptors for oxidized low density lipoprotein. Proc Natl Acad Sci USA 1996, 93, 14833-14838. 40) Remak, R. Ein Beitrag zur Entwicklungsgeschichte der Krebshaften Geschwülste. Dtsch Klin 1854, 6, 170-4. 41) Roodman, GD. Cell biology of the osteoclast. Exp Hematol 1999, 27(8), 1229-41. 42) Stanley, ER, Berg KL, Einstein DB, Lee PS, Pixley FJ, Wang Y, Yeung YG. Biology and action of colony--stimulating factor-1. Mol Reprod Dev 1997, 46(1), 4-10. 43) Strobl, H, Scheinecker C, Csmarits B, Majdic O, Knapp W. Flow cytometric analysis of intracellular CD68 molecule expression in normal and malignant haemopoiesis. Brit J Haematol 1995, 90, 774-782.

Literatur 74
44) Stryer, L. Biochemie, 4.Aufl., Heidelberg, Spektrum Akademischer Verlag, 1996. 45) Suda, T, Takahashi N, Udagawa N, Jimi E, Gillespie MT, Martin TJ. Modulation of osteoclast differentiation and function by the new members of the tumor necrosis factor receptor and ligand families. Endocr Rev 1999, 20(3), 345-57. 46) Sudhoff, H, Bujia J, Fisseler-Eckhoff A. Expression of a cell-cycle associated nuclear antigen (MIB1) in cholesteatoma and auditory meatal skin. Laryngoscope 1995, 105,1227-1231. 47) Sudhoff, H, Hildmann H, Michaels L. Pathogenesis in Cholesteatoma.The Hague, The Netherlands, Kugler Publications, 1999. 48) Takahashi, N, Udagawa N, Suda T. A new member of tumor necrosis factor ligand family, ODF/OPGL/TRANCE/RANKL, regulates osteoclast differentiation and function. Biochem Biophys Res Commun 1999; 256(3), 449-55. 49) Teitelbaum, SL. Bone resorption by osteoclasts. Science 2000, 289(5484), 1504-8. 50) Tos, M. A new pathogenesis of mesotympanic (congenital) cholesteatoma. Laryngoscope 2000, 110(11), 1890-7. 51) Vaananen, HK, Zhao H, Mulari M, Halleen JM. The cell biology of osteoclast function. J Cell Sci 2000 Feb, 113(Pt 3), 377-81. 52) van Dijk, CM, Visser CE, Veldmann JE. Spatial distribution of Langerhans` cells and T-lymphocyte subpopulations in human tympanic membrane and aural cholesteatoma. Virchows Arch B 1986, 52, 143-152. 53) Varsányi, M. Biochemisch-Molekularbiologisches Praktikum für Mediziner. Bochum, Universitätsverlag Brockmeyer, 1996. 54) Vignali, DA. The interaction between CD4 and MHC class II molecules and its effect on T cell function. Behring Institute Mitteilungen 1994, 133-147. 55) Walsh, TE, Covell WP, Ogura JH. The effect of cholesteatosis on bone. Ann Otol Rhinol Laryngol 1951, 60, 1100.

Literatur 75
56) Wetmore, RF, Muntz HR, McGill TJ. Pediatric Otolaryngology: Principles And Practic Pathways. New York, Thieme Medical Publishers Inc., 2000. 57) Yoon, TH, Lee SH, Park MH, Chang HJ, Chung JW, Kim HJ. Inhibition of Cholesteatomatous Bone Resorption with Pamidronate Disodium. Acta Otolaryngol 2001, 121, 178-181.

Danksagung 76
8. Danksagung Das Zustandekommen dieser Arbeit wurde durch viele Personen unterstützt. Ich möchte mich an erster Stelle bei dem Direktor der Universitätsklinik für Hals-Nasen-Ohrenheilkunde, Kopf- und Halschirurgie der Ruhr-Universität Bochum Herrn Professor Dr. med. H. Hildmann für die Überlassung der Arbeit bedanken. Bei Herrn Priv.-Doz. Dr. med. H. Sudhoff bedanke ich mich für die hervorragende Betreuung während der gesamten Arbeit. Seine kompetenten Ratschläge haben einen wesentlichen Beitrag zur Vollendung dieser Arbeit geleistet. Frau Dipl.-Biol. M. Hagnia, Frau Dipl.-Biol. A. Antonopoulos und Frau S. Kanabey danke ich für die exzellente Betreuung und Durchsicht des experimentellen Teils. Neben der Deutschen-Forschungs-Gemeinschaft die das Projekt „Knochenabbau durch das Cholesteatom“ (DFG Su 226/2-1) gefördert und das Zustandekommen dieser Arbeit ermöglicht hat, bedanke ich mich bei Professor Dr. S. Merchant, Massachusetts Eye and Ear Infirmary, Harvard Medical School, Boston für den Zugang zur Felsenbeinsammlung und hilfreiche Diskussionen zur Pathogenese des Cholesteatoms. Für die Durchsicht der Arbeit möchte ich mich bei Frau cand. med. C. Temp, Recklinghausen sowie Herrn Dr. rer. nat. K. Uludag, San Diego bedanken. Meinen Gymnasiallehrern Dr. phil. A. Vogt und Dr. rer. nat. W. Rücker bin ich ebenfalls zu großem Dank verpflichtet, da sie in mir frühzeitig das Interesse an der Komplexität der Perzeption und den damit verbundenen Organen geweckt haben. Meiner Familie und meinen Freunden sei für Ihre großzügige Unterstützung gedankt.