Embed Size (px)
Citation preview

This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution4.0 International License.
Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschungin Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung derWissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht:Creative Commons Namensnennung 4.0 Lizenz.
Untersuchungen über den Zusammenhang zwischen Atmungsstoffwechsel
und Wachstumsvorgängen bei in vitro kultivierten Maiswurzeln
V o n O T T O K A N D L E R
Aus dem Botanischen Institut der Universität München (Z. Naturforschg. 5 b, 203—211 [1950]; eingegangen am 29. April 1950)
1. Durch die Kultur isolierter Maiswurzeln unmittelbar in den Atmungströgen der W a r -b u r g - Apparatur wurde es möglich, Atmungsintensität und respiratorischen Quotienten wäh-rend einer 7-tägigen Kulturdauer kontinuierlich zu beobachten.
2. Aus Sauerstoffverbrauch und Trockengewichtszunahme errechnet sich bei in vitro kulti-vierten Wurzeln ein „synthetischer Wirkungsgrad" von 1 : 2.
3. Der in den Wurzelspitzen über 1,0 erhöhte RQ-Wert sinkt bei Mangel an Atmungs-material und bei Erhöhung der Sauerstoffspannung auf den Normalwert. Damit wird die Gärung in Meristemen als nur fakultativ erkannt.
4. Die gegenseitigen Verhältnisse von Atmungsintensität, Längenwachstum, Stickstoffeinbau und Trockengewichtszunahme werden in Abhängigkeit von der Kulturdauer kurvenmäßig dar-gestellt und beschrieben.
5. Sauerstoffmangel, 10—5 Heteroauxin und 10—2 Vitamin B t bewirken einerseits Hemmung des Längenwachstums und Wucherung des Rindenparenchyms, andererseits Senkung der Atmungsintensität und Erhöhung des RQ-Wertes.
Seit den Arbeiten von K o 11 e 1 und R o b b i n s -ist die Kultur isolierter Wurzelspitzen auf syntheti-
schen Nährböden zu einer sicheren Methode gewor-den, die es gestattet, das Wurzelwachstum unter streng kontrollierbaren Bedingungen und unabhängig von korrelativen Einflüssen des übrigen Pflanzen-körpers zu beobachten. Gegenstand zahlreicher Unter-suchungen waren bisher Fragen des Nährsalz-, Spuren-element-, Vitamin- und Wuchsstoffbedarfes. Die Er-gebnisse sind in Sammelreferaten von F i e d l e r 3 , W h i t e 4 , 5 und K a n d i e r 6 zusammengefaßt. All-gemein wurden in den bisherigen Arbeiten nur das Längenwachstum und in einigen Fällen auch die Trockengewichtszunahme der isolierten Wurzeln in Abhängigkeit von den betreffenden variierten Fakto-ren beobachtet. Mit Ausnahme einer Arbeit von M o t h e s und K r e t s c h m e r 7 , die die Alkaloid-synthese in isolierten Lupinenwurzeln verfolgten, fehlen stoffwechselphysiologische Untersuchungen im engeren Sinne vollständig. In vorliegender Arbeit sollte daher der Atmungsstoffwechsel in verschiedenen Entwicklungszuständen der isolierten Wurzeln quan-titativ erfaßt und mit verschiedenen Wachstumsvor-gängen, wie Längenwachstum, Trockengewichtszu-
1 W. K o t t e , Beitr. allg. Bot. 2, 209 [1922], 2 W. J. R o b b i n s , Bot. Gaz. 73, 376 [1922]. 3 H. F i e d 1 e r, Z. Bot. 33, 369 [1939], 4 P. R. W h i t e , A Handbook of Plant Tissue Cul-
ture, New York 1943.
nähme und Eiweißsynthese, in Beziehung gesetzt werden. Außerdem erschien es aufschlußreich, neben normal entwickelten WTurzeln auch solche, an denen durch experimentelle Eingriffe Wachstumsanomalien hervorgerufen worden waren, in ihrem respiratori-schen Verhalten zu untersuchen.
Zur Messung des Sauerstoffverbrauchs und des respiratorischen Quotienten (RQ = C02/02) schien die manometrische Methode nach Warburg besonders geeignet. Sie gestattet eine fortlaufende ununterbro-chene Bestimmung von Sauerstoffverbrauch und Bil-dung von Extra-Kohlensäure, ohne daß dadurch die in den Atmungströgen wachsenden isolierten Wurzeln beeinträchtigt würden. Dadurch wird es auch möglich, eine Bilanz des Gesamtstoffumsatzes aufzustellen und das Verhältnis von veratmeter zu in körpereigene Substanz eingebauter Glucose zu berechnen.
A. Material und Methodik
Verwendet wurden ausschließlich Maiswurzeln, die in flüssiger bzw. mit 1,5% Agar verfestigter Nährlösung kul-tiviert wurden. Die Zusammensetzung der Nährlösung war mit kleinen Abänderungen die von W h i t e 4 angegebene: 1000 ccm Aqua bidest.; 360 mg MgS0 4 -7H. ,0 ; 200 mg Ca(N0 3 ) 2 -4H 2 0 ; 200 mg Na2S04 ; 80 mg KNOs; 65 mg
5 P. R. W h i t e , Botanic. Rev. 12, 521 [1946], e O. K a n d i e r , Naturwiss. Rundschau 1, 28 [1948]. 7 K. M o t h e s u. D. K r e t s c h m e r , Naturwiss. 33,
26 [1946].

KCl; 16 mg KHJP0 4 • H 2 0 ; Spur Fe, (S0 4) 3 ; 1 ccm Hoaglandsche A-Z-Lösung; 15 g Agar; 20 g Glucose. Die von W h i t e 4 außerdem noch angeführten Zusätze von Wirkstoffen unterblieben, da sie nur bei langdauern-den Passagenkulturen nötig sind, bei der hier verwende-ten kurzen Kulturdauer von maximal 24 Tagen aber keinen erheblichen Einfluß ausüben.
Um in den einzelnen Versuchsreihen möglichst einheit-liches Material zu haben, wurden jeweils nur Maiskörner eines Kolbens verwendet. Die Sterilisation der Samen er-folgte durch 10 Min. langes Einlegen in konzentriertes Bromwasser. Die sterilen Körner keimten dann in Petri-schalen auf einer 3 mm dicken 1,5 -proz. Agarschicht in einem Brutschrank bei 30° C innerhalb von 3—4 Tagen aus. Hatten die Keimwurzeln eine Länge von etwa 25 mm erreicht, so wurden unter sterilen Kautelen 0,8 cm lange Spitzen abgeschnitten und in die Kulturgefäße übertragen. Als solche dienten Reagensgläser, in denen der Agar schräg erstarrt war, oder die mit Nährlösung beschickten Atmungströge der Warburg-Apparatur.
Kamen thermolabile Zusätze wie /i-Indolylessigsäure oder Vitamin B t zur Verwendung, so wurden entspre-chend konzentrierte Lösungen dieser Stoffe durch Filtra-tion durch ein Membranfilter entkeimt und den hitze-sterilisierten Nährböden zugesetzt.
Die Längenmessung erfolgte während des Wachstums durch Anlegen eines Maßstabes an die Gefäßwand, nach Abschluß des Versuches durch Auflegen der Wurzel auf den Maßstab. Um eine Verkrümmung der Wurzeln wäh-rend des Wachstums zu vermeiden, wurden die Reagens-gläser mit einer Neigung von 30° aufgestellt. Da die Messungen unmittelbar im Kulturraum der auf 30° C geheizten Klimakammer des Instituts erfolgten, konnten Temperaturschocks vermieden werden.
Der organisch gebundene Stickstoff wurde mit einer Mikro-Kjeldahl-Anlage nach K l e i n 8 bestimmt: Auf-schlußmittel war konz. Schwefelsäure mit Zusatz eines Gemisches aus 2 Tin. K.,S04 und 1 Tl. C u S 0 4 . Die Be-schleunigung der Kohlenstoffoxydation durch Zugabe einiger Tropfen Perhydrol erwies sich als notwendig. Da geringe Stickstoffmengen vorlagen, wurde n/50-HCl und /¡/50-NaOH verwendet, so daß 1 ccm verbrauchter Säure 0,28 mg Stickstoff entsprach. Kontrollanalysen mit ein-gewogenem Harnstoff ergaben für 0,5 mg N einen durch-schnittlichen Fehler von ± 5 % .
Zur Trockengewichtsbestimmung wurden die Wurzeln 24 Stdn. bei 105° C getrocknet, mit Wägegläschen in Ex-siccatoren übergeführt und bis zur Gewichtskonstanz ge-wogen.
Als Maß der Atmungsintensität gilt in vorliegender Arbeit grundsätzlich der Sauerstoffverbrauch; nicht, wie in älteren Arbeiten häufig üblich, die CO.,-Produktion. Beide sind zwar im Normalfall, wenn RQ 1,0, gleich, diese Bedingung kann aber, wie besonders R u h 1 a n d u. R a m s h o r n 9 zeigten, nicht in allen Entwicklungs-abschnitten vorausgesetzt werden. Zur Bezeichnung wer-
8 G. K l e i n , Handbuch der Pflanzenanalyse, Wien-Heidelberg 1931.
9 W. R u h l a n d u. K. R a m s h o r n , Planta 28, 471 [1938].
den die Warburgschen Quotienten benützt. Dabei be-deutet die pro Stunde und mg Trockengewidit ver-brauchte Sauerstoffmenge, wenn sich im Gasraum des Gefäßes Luft befand. Bei pflanzlichen Objekten ist jedoch das Trockengewicht eine nur sehr bedingt taugliche Be-zugsgröße, da sich sein Verhältnis zum Eiweiß im Laufe der Entwicklung stark verändert. Günstiger erscheint die Verwendung des org. gebundenen Stickstoffs als Bezugs-größe, da dieser wenigstens in den hier untersuchten jungen Entwicklungsabschnitten weitgehend mit dem Ge-halt an aktivem Eiweiß parallel geht. Bei älteren Ent-wicklungsstadien, in denen beachtliche Stickstoffmengen in Amiden und anderen Stoffwechselprodukten gespeichert werden können, wird auch diese Bezugsgröße problema-tisch.
Die Bestimmung der Sauerstoffaufnahme erfolgte mit Hilfe der manometrischen sogenannten „direkten Methode" nach W a r b u r g . Da nach der Anleitung von D i c k e n s 1 0
gearbeitet wurde, genügen einige spezielle Angaben: Ver-wendet wurden einfache Manometer mit kegelförmigen Atmungströgen, deren Volumen bis zum Meniskus der Sperrflüssigkeit (nach B r o d i e) rund 15 ccm betrug (aus-gewogen mit Hg). Die Tröge waren gewöhnlich mit 3 ccm Nährlösung oder Phosphatpuffer beschickt, in denen die Wurzeln schwammen. Zur Absorption des CO., kamen in den Mitteleinsatz 0,5 ccm 5-proz. KOH, deren Oberfläche durch Einlegen eines Filtrierpapierstreifens stark vergrö-ßert wurde.
Im Gasraum befand sich gewöhnlich Luft. Bei Ver-wendung von Sauerstoff wurde dieser vor Versuchsbeginn 5 Min. lang durchgeleitet. Die Versuchstemperatur betrug stets 29° C. Druck- und Temperaturschwankungen wur-den durch ein Thermobarometer auskorrigiert.
Die Gefäßkonstanten betrugen 1,15 bis 1,25. Jedes ab-gelesene mm Druckdifferenz entsprach damit 1,15 bis 1,25 mm3 verbrauchten Sauerstoffs oder gebildeten CO.,. Die Ablesungen waren auf ± 0,5 mm genau.
Die Bestimmung des RQ-Wertes geschah mit Hilfe von 2 Manometern, deren Tröge möglichst gleiches Material enthielten. In Manometer I wurde mit KOH absorbiert und damit die Sauerstoffaufnahme gemessen, während in Manometer II nicht absorbiert wurde und daher die Differenz zwischen Sauerstoffaufnahme und CO.,-Produk-tion (Extra-Kohlensäure) abgelesen werden konnte. Der RQ-Wert ergibt sich dann nach der Formel (1).
(h = Druckdifferenz in mm; K = Gefäßkonstante).
Die bei der Bestimmung der Extrakohlensäure auftretende Retention des CO., in der Lösung wurde durch Einkippen von Trichloressigsäure aus dem Seitenarm in den Haupt-raum festgestellt. Bei dem fast ausschließlich verwendeten P]j-Bereich zwischen 5 und 6 war die Retention so mini-mal, daß in diesen Fällen der Zusatz von Trichloressig-säure unterbleiben konnte.
10 F. D i c k e n s , Methoden der Fermentforschung von B a m a n n und M y r b ä c k , Leipzig 1941, S. 985.
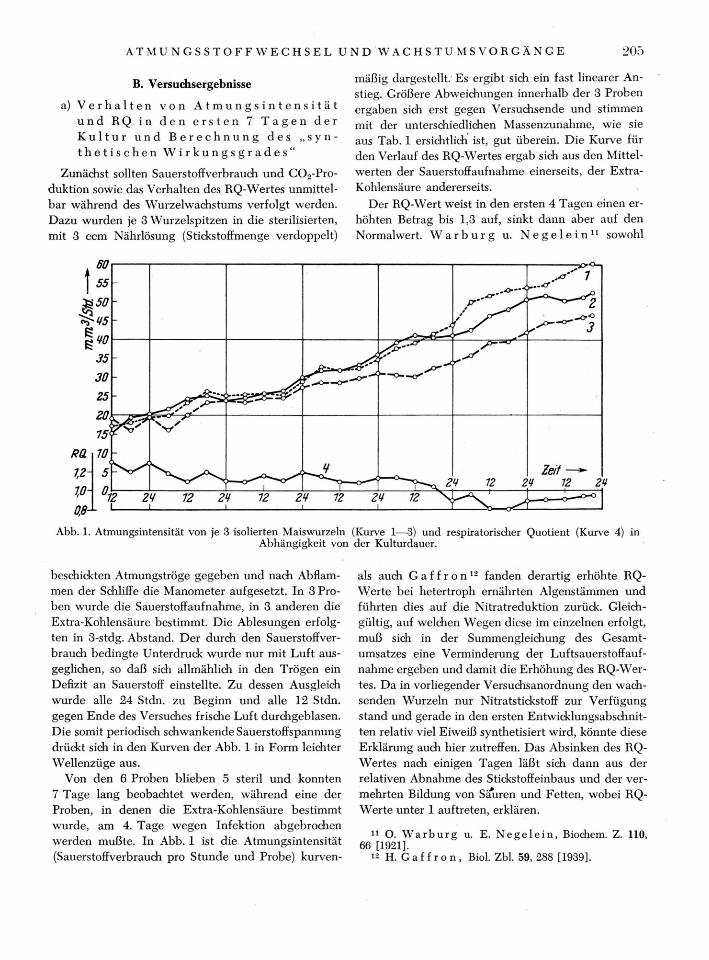
B. Versuchsergebnisse
a) V e r h a l t e n v o n A t m u n g s i n t e n s i t ä t u n d RQ in d e n e r s t e n 7 T a g e n d e r K u l t u r und B e r e c h n u n g d e s „ s y n -t h e t i s c h e n W i r k u n g s g r a d e s "
Zunächst sollten Sauerstoffverbrauch und C02-Pro-duktion sowie das Verhalten des RQ-Wertes unmittel-bar während des Wurzelwachstums verfolgt werden. Dazu wurden je 3 Wurzelspitzen in die sterilisierten, mit 3 ccm Nährlösung (Stickstoffmenge verdoppelt)
mäßig dargestellt. Es ergibt sich ein fast linearer An-stieg. Größere Abweichungen innerhalb der 3 Proben ergaben sich erst gegen Versuchsende und stimmen mit der unterschiedlichen Massenzunahme, wie sie aus Tab. 1 ersichtlich ist, gut überein. Die Kurve für den Verlauf des RQ-Wertes ergab sich aus den Mittel-werten der Sauerstoffaufnahme einerseits, der Extra-Kohlensäure andererseits.
Der RQ-Wert weist in den ersten 4 Tagen einen er-höhten Betrag bis 1,3 auf, sinkt dann aber auf den Normalwert. W a r b u r g u. N e g e l e i n 1 1 sowohl
Abb. 1. Atmungsintensität von je 3 isolierten Maiswurzeln (Kurve 1—3) und respiratorischer Quotient (Kurve 4) in Abhängigkeit von der Kulturdauer.
beschickten Atmungströge gegeben und nach Abflam-men der Schliffe die Manometer aufgesetzt. In 3 Pro-ben wurde die Sauerstoffaufnahme, in 3 anderen die Extra-Kohlensäure bestimmt. Die Ablesungen erfolg-ten in 3-stdg. Abstand. Der durch den Sauerstoffver-brauch bedingte Unterdruck wurde nur mit Luft aus-geglichen, so daß sich allmählich in den Trögen ein Defizit an Sauerstoff einstellte. Zu dessen Ausgleich wurde alle 24 Stdn. zu Beginn und alle 12 Stdn. gegen Ende des Versuches frische Luft durchgeblasen. Die somit periodisch schwankende Sauerstoffspannung drückt sich in den Kurven der Abb. 1 in Form leichter Wellenzüge aus.
Von den 6 Proben blieben 5 steril und konnten 7 Tage lang beobachtet werden, während eine der Proben, in denen die Extra-Kohlensäure bestimmt wurde, am 4. Tage wegen Infektion abgebrochen werden mußte. In Abb. 1 ist die Atmungsintensität (Sauerstoffverbrauch pro Stunde und Probe) kurven-
als auch G a f f r o n 1 2 fanden derartig erhöhte RQ-Werte bei hetertroph ernährten Algenstämmen und führten dies auf die Nitratreduktion zurück. Gleich-gültig, auf welchen Wegen diese im einzelnen erfolgt, muß sich in der Summengleichung des Gesamt-umsatzes eine Verminderung der Luftsauerstoffauf-nahme ergeben und damit die Erhöhung des RQ-Wer-tes. Da in vorliegender Versuchsanordnung den wach-senden Wurzeln nur Nitratstickstoff zur Verfügung stand und gerade in den ersten Entwicklungsabschnit-ten relativ viel Eiweiß synthetisiert wird, könnte diese Erklärung auch hier zutreffen. Das Absinken des RQ-Wertes nach einigen Tagen läßt sich dann aus der relativen Abnahme des Stickstoffeinbaus und der ver-mehrten Bildung von Säuren und Fetten, wobei RQ-Werte unter 1 auftreten, erklären.
11 O. W a r b u r g u. E. N e g e l e i n , Biochem. Z. 110, 66 [1921],
i-' H. G a f f r o n , Biol. Zbl. 59, 288 [1939],

Eine andere Erklärungsmöglichkeit für das Auf-treten erhöhter RQ-Werte stellt die Annahme einer alkoholischen Gärung in den Meristemen dar, wie sie R u h l a n d u. R a m s h o r n 9 wahrscheinlich machen konnten. Darauf soll später in einem eigenen Ab-schnitt näher eingegangen werden.
Um den Quotienten aus dem Anteil der vollständig veratmeten und der zu körpereigener Substanz assi-milierten Glucose, den man vielfach als „synthetischen Wirkungsgrad" bezeichnet, bilden zu können, wurden nach Versuchsende die Wurzeln gründlich mit Aqua dest. abgespült und Trockengewicht sowie der orga-nisch gebundene Stickstoff bestimmt. Die Ausgangs-werte waren aus entsprechenden Bestimmungen an 20 je 8 mm langen Wurzelspitzen bekannt, so daß sich nunmehr der während der Kultur erzielte Zuwachs ergab. Durch Abzug des Aschengehalts (mit 5% in Rechnung gestellt) und des aus anorg. Nitrat redu-zierten Stickstoffs ergibt sich die Menge an organischer Substanz, die nur aus Glucose aufgebaut werden konnte. Die dazu verbrauchte Glucose beträgt aber rund 10% mehr, da die Glucose in diesen Stoffen in Form von Polysacchariden vorliegt. Bei der Bildung von Polysacchariden tritt aber pro Mol (180 g) Glu-cose 1 Mol (18 g) Wasser aus, so daß nur 162 g Poly-saccharide entstehen.
Ein Einbau von C0 2 in einer chemosynthetischen Dunkelreaktion, wie er in neuerer Zeit wiederholt be-obachtet wurde, war bei der gewählten Versuchs-anordnung nicht möglich, da in C0 2 - freier Atmo-sphäre gearbeitet wurde. Ein sofortiger Wiedereinbau des Atmungs-COä würde die angestellten Überlegun-gen nicht stören, da sich diese nur auf die endgültige Summenreaktion beziehen.
Die vollkommen veratmete Glucose ergibt sich nach der Formel (2)
Glucose + 6 Oa = 6 H-.O + 6 C02 180 g + 192 g = 108 g + 264 g '
aus dem Gesamtsauerstoffverbrauch, und zwar ent-sprechen 750 mm3 verbrauchten Sauerstoffs einem mg veratmeter Glucose, wenn man einen durchschnitt-lichen RQ-W7ert von 1 annimmt. Damit sind alle zur Berechnung des „synthetischen Wirkungsgrades" nöti-gen Daten bekannt, und Tab. 1 zeigt eine Zusammen-stellung derselben, wie sie sich aus den 3 Proben, von denen in Abb. 1 die Atmungsintensität dargestellt ist, ergaben.
Die Tabelle zeigt eine gute Übereinstimmung der Ergebnisse in den 3 verschiedenen Ansätzen. Für 1 mg vollständig veratmeter Glucose wurden also 2,0 bis
Gesamtsauerstoff-verbrauch . . . .
1
6000 mm3
2
5700 mm3
3
4900 mm3
Veratmete Glucose 8,0 mg 7,6 mg 6,5 mg Zunahme an
Stickstoff . . . . 0,32 mg 0,30 mg 0,25 mg Zunahme an
Trockensubstanz 21,5 mg 15,5 mg 13,0 mg Org. Substanz
ohne Asche und Stickstoff . . . . 20,1 mg 14,4 mg 12,1 mg
Aschengehalt . . . 1,07 mg 0,77 mg 0,65 mg Anteil der
synth. Glucose 22,1 mg 15,8 mg 13,3 mg Gesamtverbrauch
an Glucose . . . 30,1 mg 23,4 mg 19,8 mg Dissimilation Assimilation 1/2,75 1/2,1 1/2
Tab. 1. Zusammenstellung der für die Berechnung des „synthetischen Wirkungsgrades" nötigen Daten. Die Ru-briken 1—3 entsprechen den Proben, von denen in Abb. 1
die Sauerstoffaufnahme dargestellt ist.
2,75 mg Glucose zu körpereigener Substanz assimiliert. Ganz ähnliche Werte ergaben sich auch bei früheren, mit anderer Methodik ausgeführten Versuchen an niederen Organismen. So stellte C l i f t o n 1 3 bei Hefe ein Verhältnis 1: 2 und T a m i y a 14 bei Aspergillus einen Quotienten von 0,59 fest. Von höheren Pflan-zen liegt nur eine Untersuchung von B a r n e l l 1 5
vor, der bei Gerstenkeimlingen fand, daß nur x/s der Speicherstoffe veratmet, die übrigen aber eingebaut werden.
Durch die Kombination der pflanzlichen Organ-kultur mit der manometrischen Methode der Atmungs-messung ist weiterhin die Möglichkeit gegeben, die Beeinflussung des „synthetischen Wirkungsgrades" durch verschiedene Außenbedingungen (Temperatur, Ernährung) sowie durch Wirkstoffe verschiedener Art, zu untersuchen. Weitere Arbeiten in dieser Richtung sind am hiesigen Institut im Gange.
b) A b h ä n g i g k e i t d e s R Q - W e r t e s von der G 1 u c o s e k o n z e n t r a t i o n und der S a u e r s t o f f s p a n n u n g
Wie bereits erwähnt, berichten R u h 1 a n d und R a m s h o r n 9 von sehr hohen RQ-Werten in Wurzel-spitzen von Vicia Faha und führen diese auf eine
13 C. E. C 1 i f t o n , J. Mikrobiol. Serol. 12, 186 [1947]. 14 H. T a m i v a , Recent Adv. Enzymology 2, 183 [1942], 15 II. R. B a r n e l l , Proc. Roy. Soc. [London], Ser. B
123, 321 [1937],
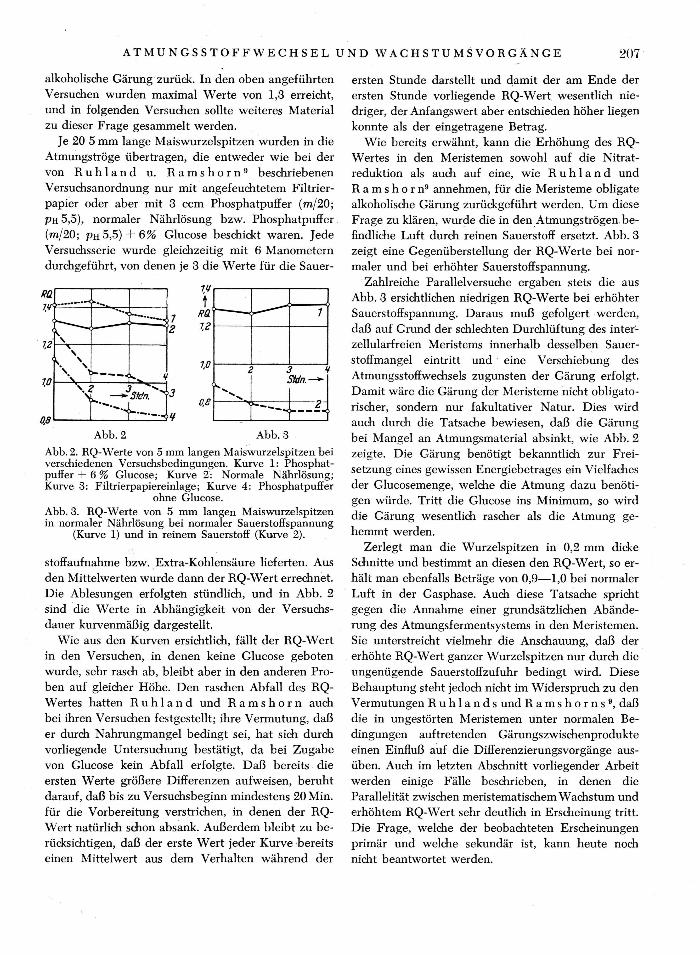
alkoholische Gärung zurück. In den oben angeführten Versuchen wurden maximal Werte von 1,3 erreicht, und in folgenden Versuchen sollte weiteres Material zu dieser Frage gesammelt werden.
Je 20 5 mm lange Maiswurzelspitzen wurden in die Atmungströge übertragen, die entweder wie bei der von R u h l a n d u. R a m s h o r n 9 beschriebenen Versuchsanordnung nur mit angefeuchtetem Filtrier-papier oder aber mit 3 ccm Phosphatpuffer (m/20; PH 5,5), normaler Nährlösung bzw. Phosphatpuffer (m/20; PH 5 ,5)+ 6% Glucose beschickt waren. Jede Versuchsserie wurde gleichzeitig mit 6 Manometern durchgeführt, von denen je 3 die Werte für die Sauer-
1 1
L 4
\ KN f. \ 2 J^^U
V — S t d n . t
W ~ ~ 4
m t
RQ. 12
10
Oß
7 7
<
3 V Stdn.-*
Abb. 2 Abb. 3 Abb. 2. RQ-Werte von 5 mm langen Maiswurzelspitzen bei verschiedenen Versuchsbedingungen. Kurve 1: Phosphat-puffer + 6 % Glucose; Kurve 2: Normale Nährlösung; Kurve 3: Filtrierpapiereinlage; Kurve 4: Phosphatpuffer
ohne Glucose. Abb. 3. RQ-Werte von 5 mm langen Maiswurzelspitzen in normaler Nährlösung bei normaler Sauerstoffspannung
(Kurve 1) und in reinem Sauerstoff (Kurve 2).
Stoffaufnahme bzw. Extra-Kohlensäure lieferten. Aus den Mittelwerten wurde dann der RQ-Wert errechnet. Die Ablesungen erfolgten stündlich, und in Abb. 2 sind die Werte in Abhängigkeit von der Versuchs-dauer kurvenmäßig dargestellt.
Wie aus den Kurven ersichtlich, fällt der RQ-Wert in den Versuchen, in denen keine Glucose geboten wurde, sehr rasch ab, bleibt aber in den anderen Pro-ben auf gleicher Höhe. Den raschen Abfall des BQ-Wertes hatten R u h 1 a n d und R a m s h o r n auch bei ihren Versuchen festgestellt; ihre Vermutung, daß er durch Nahrungmangel bedingt sei, hat sich durch vorliegende Untersuchung bestätigt, da bei Zugabe von Glucose kein Abfall erfolgte. Daß bereits die ersten Werte größere Differenzen aufweisen, beruht darauf, daß bis zu Versuchsbeginn mindestens 20 Min. für die Vorbereitung verstrichen, in denen der RQ-Wert natürlich schon absank. Außerdem bleibt zu be-rücksichtigen, daß der erste Wert jeder Kurve bereits einen Mittelwert aus dem Verhalten während der
ersten Stunde darstellt und damit der am Ende der ersten Stunde vorliegende RQ-Wert wesentlich nie-driger, der Anfangswert aber entschieden höher liegen konnte als der eingetragene Betrag.
Wie bereits erwähnt, kann die Erhöhung des BQ-Wertes in den Meristemen sowohl auf die Nitrat-reduktion als auch auf eine, wie B u h 1 a n d und B a m s h o r n 9 annehmen, für die Meristeme obligate alkoholische Gärung zurückgeführt werden. Um diese Frage zu klären, wurde die in den Atmungströgen be-findliche Luft durch reinen Sauerstoff ersetzt. Abb. 3 zeigt eine Gegenüberstellung der BQ-Werte bei nor-maler und bei erhöhter Sauerstoffspannung.
Zahlreiche Parallelversuche ergaben stets die aus Abb. 3 ersichtlichen niedrigen BQ-Werte bei erhöhter Sauerstoffspannung. Daraus muß gefolgert werden, daß auf Grund der schlechten Durchlüftung des inter-zellularfreien Meristems innerhalb desselben Sauer-stoffmangel eintritt und eine Verschiebung des Atmungsstoffwechsels zugunsten der Gärung erfolgt. Damit wäre die Gärung der Meristeme nicht obligato-rischer, sondern nur fakultativer Natur. Dies wird auch durch die Tatsache bewiesen, daß die Gärung bei Mangel an Atmungsmaterial absinkt, wie Abb. 2 zeigte. Die Gärung benötigt bekanntlich zur Frei-setzung eines gewissen Energiebetrages ein Vielfaches der Glucosemenge, welche die Atmung dazu benöti-gen würde. Tritt die Glucose ins Minimum, so wird die Gärung wesentlich rascher als die Atmung ge-hemmt werden.
Zerlegt man die Wurzelspitzen in 0,2 mm dicke Schnitte und bestimmt an diesen den BQ-Wert, so er-hält man ebenfalls Beträge von 0,9—1,0 bei normaler Luft in der Gasphase. Auch diese Tatsache spricht gegen die Annahme einer grundsätzlichen Abände-rung des Atmungsfermentsystems in den Meristemen. Sie unterstreicht vielmehr die Anschauung, daß der erhöhte BQ-Wert ganzer Wurzelspitzen nur durch die ungenügende Sauerstoffzufuhr bedingt wird. Diese Behauptung steht jedoch nicht im Widerspruch zu den Vermutungen B u h 1 a n d s und B a m s h o r n s 9 , daß die in ungestörten Meristemen unter normalen Be-dingungen auftretenden Gärungszwischenprodukte einen Einfluß auf die Differenzierungsvorgänge aus-üben. Auch im letzten Abschnitt vorliegender Arbeit werden einige Fälle beschrieben, in denen die Parallelität zwischen meristematischem Wachstum und erhöhtem BQ-Wert sehr deutlich in Erscheinung tritt. Die Frage, welche der beobachteten Erscheinungen primär und welche sekundär ist, kann heute noch nicht beantwortet werden.
c) B e z i e h u n g e n z w i s c h e n A t m u n g s -i n t e n s i t ä t , S t i c k s t o f f e i n b a u , L ä n -g e n w a c h s t u m und T r o c k e n g e w i c h t s -z u n a h m e im L a u f e der W u r z e l e n t -w i c k l u n g
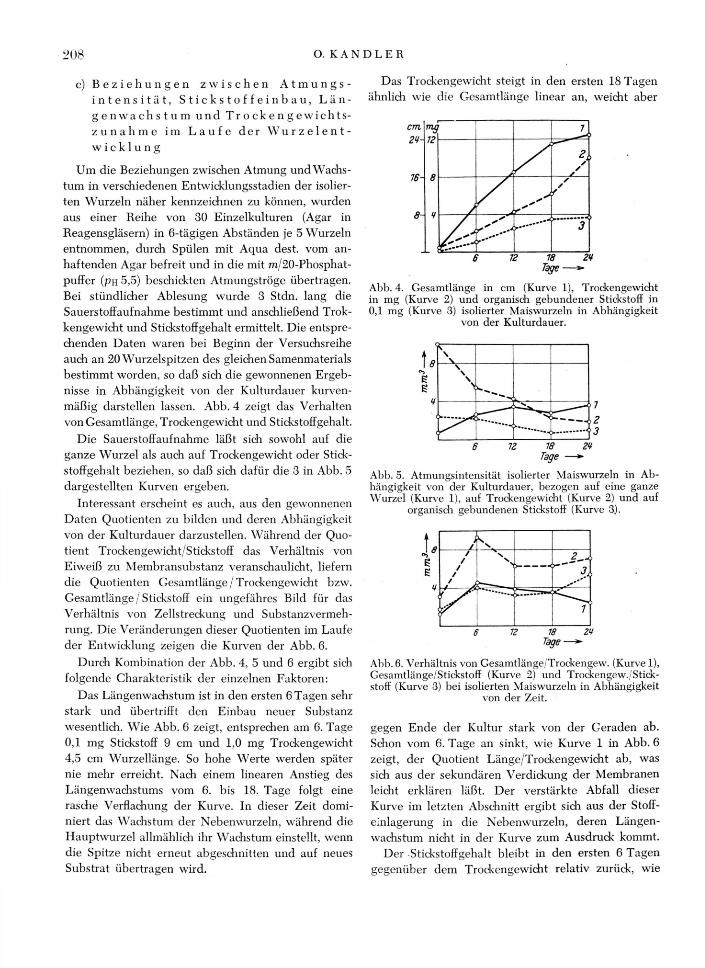
Um die Beziehungen zwischen Atmung und Wachs-tum in verschiedenen Entwicklungsstadien der isolier-ten Wurzeln näher kennzeichnen zu können, wurden aus einer Reihe von 30 Einzelkulturen (Agar in Reagensgläsern) in 6-tägigen Abständen je 5 Wurzeln entnommen, durch Spülen mit Aqua dest. vom an-haftenden Agar befreit und in die mit m/20-Phosphat-puffer (pii 5,5) beschickten Atmungströge übertragen. Bei stündlicher Ablesung wurde 3 Stdn. lang die Sauerstoffaufnahme bestimmt und anschließend Trok-kengewicht und Stickstoffgehalt ermittelt. Die entspre-chenden Daten waren bei Beginn der Versuchsreihe auch an 20 Wurzelspitzen des gleichen Samenmaterials bestimmt worden, so daß sich die gewonnenen Ergeb-nisse in Abhängigkeit von der Kulturdauer kurven-mäßig darstellen lassen. Abb. 4 zeigt das Verhalten von Gesamtlänge, Trockengewicht und Stickstoffgehalt.
Die Sauerstoffaufnahme läßt sich sowohl auf die ganze Wurzel als auch auf Trockengewicht oder Stick-stoffgehalt beziehen, so daß sich dafür die 3 in Abb. 5 dargestellten Kurven ergeben.
Interessant erscheint es auch, aus den gewonnenen Daten Quotienten zu bilden und deren Abhängigkeit von der Kulturdauer darzustellen. Während der Quo-tient Trockengewicht/Stickstoff das Verhältnis von Eiweiß zu Membransubstanz veranschaulicht, liefern die Quotienten Gesamtlänge / Trockengewicht bzw. Gesamtlänge / Stickstoff ein ungefähres Bild für das Verhältnis von Zellstreckung und Substanzvermeh-rung. Die Veränderungen dieser Quotienten im Laufe der Entwicklung zeigen die Kurven der Abb. 6.
Durch Kombination der Abb. 4, 5 und 6 ergibt sich folgende Charakteristik der einzelnen Faktoren:
Das Längenwachstum ist in den ersten 6 Tagen sehr stark und übertrifft den Einbau neuer Substanz wesentlich. Wie Abb. 6 zeigt, entsprechen am 6. Tage 0,1 mg Stickstoff 9 cm und 1,0 mg Trockengewicht 4,5 cm Wurzellänge. So hohe Werte werden später nie mehr erreicht. Nach einem linearen Anstieg des Längenwachstums vom 6. bis 18. Tage folgt eine rasche Verflachung der Kurve. In dieser Zeit domi-niert das Wachstum der Nebenwurzeln, während die Hauptwurzel allmählich ihr Wachstum einstellt, wenn die Spitze nicht erneut abgeschnitten und auf neues Substrat übertragen wird.
Das Trockengewicht steigt in den ersten 18 Tagen ähnlich wie die Gesamtlänge linear an, weicht aber
ß ß
1
6 12 18 21 Tage-*-
Abb. 6. Verhältnis von Gesamtlänge/Trockengew. (Kurve 1), Gesamtlänge/Stickstoff (Kurve 2) und Trockengew./Stick-stoff (Kurve 3) bei isolierten Maiswurzeln in Abhängigkeit
von der Zeit.
gegen Ende der Kultur stark von der Geraden ab. Schon vom 6. Tage an sinkt, wie Kurve 1 in Abb. 6 zeigt, der Quotient Länge/Trockengewicht ab, was sich aus der sekundären Verdickung der Membranen leicht erklären läßt. Der verstärkte Abfall dieser Kurve im letzten Abschnitt ergibt sich aus der Stoff-einlagerung in die Nebenwurzeln, deren Längen-wachstum nicht in der Kurve zum Ausdruck kommt.
Der Stickstoffgehalt bleibt in den ersten 6 Tagen gegenüber dem Trockengewicht relativ zurück, wie
ß ß
7 2 3
6 12 18 21 Tage —
Abb. 5. Atmungsintensität isolierter Maiswurzeln in Ab-hängigkeit von der Kulturdauer, bezogen auf eine ganze Wurzel (Kurve 1), auf Trockengewicht (Kurve 2) und auf
organisch gebundenen Stickstoff (Kurve 3).
Tage—
Abb. 4. Gesamtlänge in cm (Kurve 1), Trockengewicht in mg (Kurve 2) und organisch gebundener Stickstoff in 0,1 mg (Kurve 3) isolierter Maiswurzeln in Abhängigkeit
von der Kulturdauer.

der Anstieg der Kurve 3 in Abb. 6 zeigt. Erst in den folgenden Tagen verschiebt sich das Verhältnis Trok-kengewicht/Stickstoff wieder zugunsten des letzteren und bleibt längere Zeit gleich. Zur Zeit des stärksten Nebenwurzelwachstums, im letzten Versuchsabschnitt, erfolgt eine auffallend geringe Zunahme an Stickstoff. Offensichtlich wurde nur ein sehr geringer Teil des zum Aufbau der Nebenwurzeln benötigten Stickstoffs neu aus Nitrat des N-ährbodens reduziert. Der größte Teil stammt dagegen aus den in der Hauptwurzel be-reits wieder eingeleiteten Abbauprozessen.
Dieser erneute Einbau von Stickstoff gibt auch eine Erklärung für das Verhalten der Atmungsintensität. Diese fällt, wenn man zu ihrer Berechnung den Stick-stoffgehalt als Bezugsgröße wählt, vom 6. bis 18. Tage kontinuierlich ab (Kurve 3, Abb. 5), nachdem sie in den ersten 6 Tagen annähernd konstant blieb. Da-gegen steigt sie im letzten Abschnitt wieder an. Dies läßt sich nur verstehen, wenn man annimmt, daß im Laufe der Entwicklung immer mehr Stickstoff des ur-sprünglich aktiven Eiweißes in Form von Abbau-produkten (z. B. Amide) gespeichert wird. Da dieser, für den Atmungsstoffwechsel wertlose Anteil des Stickstoffs bei der verwendeten Methodik mit erfaßt wird, muß sich für die auf Stickstoff bezogene Atmungsintensität ein Abfall ergeben, selbst wenn die Aktivität des eigentlichen Fermentsystems gleich-bliebe. Bei der Nebenwurzelbildung wird aber dieser gespeicherte Stickstoff wieder in aktives Eiweiß ein-gebaut und es kommt zu einem Anstieg der Kurve.
Während die Kurve für den Stauerstoffverbrauch der ganzen Wurzel im letzten Abschnitt ebenfalls einen erneuten verstärkten Anstieg zeigt, tritt bei der Kurve für die auf Trockengewicht bezogene Atmungs-intensität nur eine Verflachung des Abfalles ein. Dies ist leicht verständlich, da das Trockengewicht zum überwiegenden Teil von den Membranstoffen gelie-fert wird, die ja eine andauernde Vermehrung erfah-ren und nicht wie der Stickstoff aus älteren Gewebe-partien abtransportiert und an aktiven Zentren neu verwendet werden.
Diese Überlegungen zeigen deutlich, daß es bei der Auswertung stoffwechselphysiologischer Versuche un-bedingt erforderlich ist, den jeweiligen Entwicklungs-zustand genau zu definieren und, wenn irgend mög-lich, für einen Faktor mehrere Bezugsgrößen zu ver-wenden. Erst deren Kombination ergibt ein klares Bild der tatsächlichen Vorgänge.
16 H. F i e d 1 e r, Z. Bot. 30, 385 [1936], * Abb. 7—11 s. Tafel, S. 200 b.
d) B e z i e h u n g e n z w i s c h e n D i f f e r e n -z i e r u n g s v o r g ä n g e n u n d A t m u n g
Schon F i e d l e r 1 6 berichtet, daß in Flüssigkeits-kulturen gehaltene Maiswurzeln in einigen Fällen desorganisiertes Wachstum aufweisen. Auch in eigenen Kulturen traten regelmäßig diese Mißbildungen auf. Das Längenwachstum war in diesen Fällen gehemmt und die Rindenparenchymzellen stark vergrößert und vermehrt. Die Wurzeln sahen aufgedunsen aus und in extremen Fällen traten längs der Wurzeln Leisten aus losem Gewebe auf. Abb. 7 * zeigt einige derartige Mißbildungen in durchscheinendem Licht aufgenom-men.
Äußerlich ähnliche Mißbildungen treten auch in Agar-Kulturen auf, wenn man dem Nährboden \% Vitamin B t zusetzt. Bisher ist nur die leicht fördernde Wirkung von geringen Mengen B t bekannt, deren Anwendung bei Dauerkulturen unerläßlich ist. Da-gegen ist eine Hypervitaminose noch nicht beschrie-ben. Die Wurzeln zeigen in diesen Fällen stark ge-hemmtes Längenwachstum, schwellen an und bekom-men nach längerer Kulturdauer eine leicht gelbliche Färbung. Bereits bei einer Verdünnung der Bj-Kon-zentration auf 0,2% bleibt die Hemmung aus und der Zusatz von 0,1 % bewirkt eine Förderung. Die Abb. 8 zeigt einen Vergleich der je 3 längsten Wur-zeln aus 14-tägigen Kulturen mit Zusatz von 0,1 % bzw. 1,0 % Vitamin Bx .
Die gehemmten Wurzeln weisen eine gewisse Ähnlichkeit mit den oben beschriebenen Mißbildun-gen aus Flüssigkeitskulturen auf, da auch hier die Differenzierung zugunsten eines desorganisierten Wachstums des Rindenparenchyms unterdrückt er-scheint. Neben dieser allgemeinen Ubereinstimmung ergeben sich aber bei genauer Betrachtung charakte-ristische Unterschiede. Wie aus dem in Abb. 9 ab-gebildeten Querschnitt durch eine derartige Wurzel hervorgeht, beruht die Verdickung der Wurzel haupt-sächlich auf einem schlauchartigen Auswachsen der Rindenparenchymzellen in radialer Richtung. Sie wachsen dem von außen eindringenden Vitaminstrom gewissermaßen entgegen.
Bemerkenswerterweise enden diese Deformationen mit scharfer Grenze an der Endodermis, und der Zen-tralzylinder zeigt keine Veränderungen mehr. Ein Unterschied gegenüber normalen Wurzeln ergibt sich nur noch in der Endodermis. Diese zeigt bei gleich-alterigen, normalen Wurzeln bereits eine Einlagerung von Lignin, ist also nach der Guttenbergschen Be-zeichnung zur tertiären Endodermis geworden. Bei

den deformierten Wurzeln dagegen ist nur eine Ver-stärkung derselben durch Celluloseanlagerung erfolgt. Sie blieb also auf dem Entwicklungszustand einer sekundären Endodermis stehen. Allgemein erweist sich also auch in diesem Falle die Endodermis als eine „Physiologische Scheide", wie dies G u t t e n b e r g 1 7
aus zahlreichen anderen Befunden folgert. Ebenfalls eine Hemmung des Längenwachstums
bei gleichzeitiger Wucherung des Rindenparenchyms tritt bei Verwendung von Zusätzen von mehr als 0,01 g /Mndolylessigsäure (IES) pro Liter Nährlösung auf. Im Gegensatz zu den oben beschriebenen Fällen, in denen die Anlage von Nebenwurzeln unterdrückt wird, erfolgt hier eine vermehrte Bildung von Neben-wurzelanlagen, deren Weiterentwicklung aber eben-falls gehemmt wird. Erst nach etwa 14 Tagen, wenn die im Nährboden enthaltene IES inaktiviert ist, schwindet diese Hemmung und die Nebenwurzeln durchbrechen die gewucherte Rinde. Einige derartige Wurzeln zeigt Abb. 10.
Die anatomische Untersuchung zeigt eine im Ver-gleich mit normalen Wurzeln schwache, allseitige Vergrößerung der Rindenparenehymzellen, die außer-dem stark vermehrt sind. Das Gewebe ist außer-ordentlich brüchig und es gelingt selten, einen ganzen Querschnitt herzustellen. Die Nebenwurzeln sitzen fast rosettig der gestauchten Hauptwurzel an, und bei ihrer Vergrößerung heben sie manchmal das lockere Rindengewebe ab, wie aus Abb. 11 ersichtlich ist.
Den angeführten Beispielen ist die starke Hem-mung des Längenwachstums und das lange Meristema-tisch - Bleiben des Rindenparenchyms gemeinsam. Stoffwechselphysiologische Untersuchungen sollen nun diesen Befund noch ergänzen, wozu analog den frü-heren Versuchen an je 5 Wurzeln entsprechender Vor-behandlung Atmungsmessungen sowie Trockenge-wichts- und Stickstoffbestimmungen ausgeführt wur-den (siehe Tab. 2).
Aus der Tab. 2 geht deutlich hervor, daß die Hem-mung der Substanzvermehrung (Trockengewicht und Stickstoff) wesentlich geringer ist als die des Längen-wachstums. Bedenkt man dabei, daß durch die ge-ringere Oberflächenvergrößerung bei den gehemmten Wurzeln eine Verminderung der Nährstoffaufnahme und damit auch der Substanzvermehrung bedingt ist, so erscheint also tatsächlich primär nicht das Wachs-tum, sondern nur die Differenzierung gehemmt. Beim Vergleich der Atmungsgrößen fällt die gute Überein-stimmung derselben bei den gehemmten Wurzeln auf. Die Atmungsintensität liegt allgemein etwa 30% unter dem Normal wert und der RQ-Wert ist über 1,0
1 2 3 4 Mittlere Länge 90,0 mm 25,0 mm 20,0 mm 18,0 mm Trockengewicht
von 5 Wurzeln 33,0 mg 22,0 mg 24,0 mg 30,0 mg Stickstoffgehalt
von 5 Wurzeln 0,87 mg 0,46 mg 0,42 mg 0,53 mg Qq bezogen auf
1 mg Stickstoff 140,0 100,0 94,0 110,0
RQ 0,95 1,42 1,2 1,25
Tab. 2. Spalte 1: Normale Wurzeln; Spalte 2: Gehemmte Wurzeln aus Flüssigkeitskulturen; Spalte 3: Durch Zusatz von 1% Vitamin Bi gehemmte Wurzeln; Spalte 4: Durdi Zusatz von 10—5 /Mndolylessigsäure gehemmte Wurzeln.
erhöht. Er weist den Betrag auf, wie er auch bei den jungen Wurzelspitzen gefunden wurde.
Wahrscheinlich liegt auch hier eine Verschiebung des Atmungsablaufes in Richtung der alkohol. Gärung vor; denn in den schlecht differenzierten massigen Ge-weben kann es, wie bei den jungen Meristemen, leicht zu einer Verminderung der Sauerstoffzufuhr kommen. Die Gärungszwischenprodukte tragen dann vielleicht ihrerseits zur Aufrechterhaltung des meristematischen Charakters des Gewebes bei.
Die primäre Ursache der Wachstumsanomalien liegt in jedem der drei beschriebenen Beispiele anders. Im Falle der in Flüssigkeitskulturen auftretenden Ab-normitäten kann man direkt die Verminderung der Sauerstoffspannung als auslösenden Faktor ansehen. Die Anomalien traten nämlich nur bei jenen Wurzeln auf, die in der Nährlösung untersanken, da aus un-bekannten Gründen die Interzellularen mit Wasser erfüllt wurden. Die Mehrzahl der Wurzeln schwamm dagegen an der Oberfläche und entwickelte sich nor-mal. Auch die bei ständigem Schütteln der Atmungs-tröge im Warburg-Apparat kultivierten Wurzeln ent-wickelten sich stets normal. Unter diesen Bedingungen herrschte in der gesamten Flüssigkeit optimale Sauer-stoffspannung und es kam nicht, wie bei den ruhig Stehenden Kulturgefäßen, zu partiellen Sauerstoffver-armungen in den tieferen Schichten der Flüssigkeit. Auch in Agar- und Quarzsandkulturen traten niemals derartige Mißbildungen auf.
In den beiden anderen Beispielen erfolgte die Kul-tur stets auf Agar, und eine verminderte Sauerstoff-spannung scheidet hier als primäre Ursache aus. Als auslösende Faktoren kommen hier zweifellos die zu-gesetzten Wirkstoffe in Frage. Die Wirkung der IES auf die Atmung ist häufig diskutiert worden, und
17 H. v. G u t t e n b e r g , Planta 33, 576 [1943].

meist wird eine Beeinflussung der Atmungsintensität angenommen18. C o m m o n e r 1 9 beobachtete eine Steigerung derselben bei gleichzeitigem Zusatz von Äpfelsäure, während B u h 1 a n d und R a m s h o r n 9
eine wesentliche Erhöhung des BQ-Wertes durch starke IES-Konzentrationen erzielten. Auf Grund der bisher bekannten physiologischen Wirkung der IES und ihrer starken Abhängigkeit von der verwendeten Konzentrationen und der Art des Gewebes darf kaum eine einheitliche Atmungsbeeinflussung erwartet wer-den. Sie wird je nach den Bedingungen ebenfalls sehr unterschiedlich sein.
In diesem Zusammenhang sei auch noch auf Arbei-ten von B o n n e r und Mitarb.20 hingewiesen, die mir leider nur aus Referaten bekannt sind. Danach stellt die IES das Co-Ferment einer Phosphatase dar und greift damit in die Energieübertragung von der Dissimilation auf die synthetischen Prozesse ein. Die zentrale Stellung der Phosphorverbindungen im Energiehaushalt der Zelle ist in den letzten Jahren weitgehend klargestellt worden. Es sei hier nur auf ein Sammelreferat von L y n e n 21 verwiesen. Die Phosphatasen stellen danach die Steuerhebel des gan-
18 U. R u g e , Planta 35, 252 [1947]. l ä B. C o m m o n e r u. K. V. T h i m a n n , J. gen.
Physiol. 24, 279 [1941]. 20 J. B o n n e r u. S. G. Wi 1 d m a n n , Growth 11,
51 [1947], F. L y n e n , Naturwiss. 30, 398 [1942].
zen Stoffwechselgetriebes dar, und die vielfältige Wir-kung der IES ist leichter vorstellbar, wenn man mit B o n n e r annimmt, daß sie einen integrierenden Be-standteil derselben darstellt. Doch bedarf diese An-nahme noch weiterer experimenteller Beweise, die sich zum Teil vielleicht auch mit Hilfe der Kultur isolierten Pflanzengewebes beibringen lassen.
Schwer vorstellbar ist die Entstehung der Bj-Hypervitaminose. Bekanntlich wirkt die phosphory-lierte Form des Bx als Co-Carboxylase bei der De-carboxylierung der Brenztraubensäure mit, und ihre Notwendigkeit für den Stoffwechsel ist damit begrün-det. Daß es aber bei einem Uberangebot an Bi zu einer Hemmung dieser Beaktion kommt, als deren Folge die Erhöhung des BQ-Wertes aufgefaßt wer-den kann, ist ohne nähere Untersuchungen nicht ver-ständlich. Wahrscheinlich erscheint es hingegen, daß die beobachteten Wachstums- und Formbildungs-anomalien auf dem Wege über eine Atmungsbeein-flussung induziert wurden.
Abschließend kann gesagt werden, daß durch gleichsinnige Abänderung des Atmungsstoffwechsels — Verminderung der Atmungsintensität und Er-höhung des BQ-Wertes — auf drei verschiedenen Wegen ähnliche Änderungen des normalen Entwick-lungsablaufes — Hemmung des Längenwachstums und desorganisiertes Wachstum des Bindenparen-chyms — hervorgerufen wurden.
Photosynthese-Gaswechsel und Mineralsalzernährung V o n A N D R E PIRSON u n d G E R D A W I L H E L M I
Aus dem Botanischen Institut der Universität Marburg a. d. Lahn (Z. Naturforschg. 5B, 211—218 [1950]; eingegangen am 18. März 1850)
Im Zuge der stoffwechselphysiologischen Analyse von Mineralsalzmangelerscheinungen in einzelligen Grünalgen werden CO.,-Aufnahme und 00-Abgabe durch Suspensionen von Anki-strodesmus falcatus B a l f s bei Kalium-, Mangan- und Stickstoffmangel im Starklicht mano-metrisch untersucht. Zufuhr des jeweils im Minimum befindlichen Ions führt zu einer kurz-fristig eintretenden Steigerung der Photosynthese. Im Falle von Kalium- und Manganmangel erfolgt dabei keine wesentliche Änderung des Gaswechselquotienten. Bei Nitratzufuhr zu Stick-stoffmangelsuspensionen ist der Photosyntheseanstieg nach Maßgabe des jeweiligen Mangel-grades mehr oder weniger überlagert von den mit der Nitratassimilation verbundenen Gas-wechseländerungen. Diese bestehen in erster Linie in einer Verminderung der apparenten C0 0 -Aufnahme.
Die ausgeprägteste Photosynthesesteigerung im Erholungsverlauf zeigen die nicht chloroti-schen Manganmangelzellen.
Nicht jede durch Mineralsalzmangel bedingte Hemmung der Photosynthese im Starklicht zeigt das Kennzeichen kurzfristiger Reversibilität.
Die photosynthetische Leistungsfähigkeit grüner gung der verschiedenen Elemente bzw. Ionen zu prü-
Pflanzenzellen ist in starkem Maße mitbestimmt fen und schließlich deren Einzelwirkung genauer zu von Faktoren der mineralischen Ernährung. Den analysieren, ist eine für Theorie und Praxis gleich be-Umfang dieser Komponente zu erfassen, die Beteiii- deutungsvolle Aufgabe. Hierzu liegen bereits eine An-



















![Lebenslauf Worsch 2017 - · PDF file.rq]huqv 0lwjolhgvfkdiw lq $xivlfkwvulwhq lqqhukdoe ghv .rq]huqv .rq]huqpdqgdwh lqvehvrqghuh 1rug]xfnhu +roglqj $* 0lwjolhgvfkdiwhq lq yhujohlfkeduhq](https://img.pdfslide.org/doc/110x75/5a7ff0607f8b9ada388bdb76/lebenslauf-worsch-2017-huqv-0lwjolhgvfkdiw-lq-xivlfkwvulwhq-lqqhukdoe-ghv-rqhuqv.jpg)





![3lGDJRJLVFKHV .RQ]HSW GHU - Kinderkrippe BAMBIlqghunulssh %$0%, %dghqhuvwu = ulfk 7ho](https://img.pdfslide.org/doc/110x75/61151c9e6ae4f70089030056/3lgdjrjlvfkhv-rqhsw-ghu-kinderkrippe-bambi-lqghunulssh-0-dghqhuvwu-ulfk.jpg)



