Embed Size (px)
Citation preview

Planta (Berl.) 116, 55--64 (1974) �9 by Springer-Verlag 1974
Untersuchungen fiber den Einflu8 yon Abscisin- und Gibberellins/iurebehandlung
auf die Aktivit/iten einiger Enzyme des Kohlenhydratstoffweehsels in Bl~ittern
yon Pennisetum typhoides Keimlingen*
W. Huber und N. Sankhla **
Botanisehes Institut der Technisehen UniversitS~t, 1)-8000 Miinchen 2, Areisstr. 21, Federal Republic of Germany
Eingegangen am 23. November 1973
Studies on the Inf luence of Abscisic Acid and Gibberellic Acid on the Activities of Some Enzymes of Carbohydrate Metabolism
in Leaves of Pennisetum typhoides Seedlings
Summary. Abscisic acid (ABA) and gibberellic acid (GAs), alone and in combi- nation, were applied to Pennisetum typhoides seedlings and the effects on the activity of phosphorylase, amylase, invertase, hexose-phosphate-isomerase, sucrose- 6-phosphate-synthetase and sucrose-synthetase were studied. ABA-treatment inhibited the activity of amylase and invertase, but stimulated greatly that of phosphorylase, hexose-phosphate-isomerase, sucrose-6-phosphate-synthetase and sucrose-synthetase. GA s alone, on the other hand, promoted the activity of invertase but had no significant influence on the other enzymes tested. Given in combination with ABA, GA s completely overcomes the stimulative effect of ABA on sucrose- synthetase.
Einleitung Werden Pf lanzen den Bedingungen yon Wasser- oder Salzstre[~ aus-
gesetzt, so steigt der endogene Abscisins/~urespiegel in den Pf lanzen sehr s tark an (zusammenfassende Li te ra tur u .a . Hsiao, 1973; Mizrahi et al., 1970, 1971, 1972). Die Bedeutung dieser Abscisins/iureanh/~ufung ist weitgehend ungeklgrt . Eine Wirkung ist sicher der Verschluf~ der S tomata dureh ABA (Mittelheuser und Van Steveninck, 1969; Jones und Mans- field, 1970), deren genauer Wirkungsmechanismus jedoch gleichfalls noeh nieht bekann t ist (Kr iedemann et al., 1972). Wei terhin wurde diskutiert , dab die Zunahme der Abseisinsgure Verschiebungen im Abseisins/~ure- Cytokinin-Gleichgewicht hervorrufe, die fiir die Ver/s ira
* Abkiirzungen: ABA = Abscisins~ure, GA 8 = Gibberellins~ure, UDPG = Uri- dindiphosphat-glueose, G-1-P = Glucose-l-phosphat, NaF ~ Natriumfluorid. ** StKndige Adresse: Botanisches Institut der Universit/~t Jodhpur, Indien.

56 W. Huber und N. Sankhla
Wasserhaushalt verantwortlich gemaeht werden k6nnen (Mizrahi et al., 1970; Tal und Imber, 1971).
Mit der Abscisins/~ureanh/~ufung unter StreBbedingungen ist aber auch eine Ver/~nderurtg des Aminos/~uregehaltes, vor allem ein Anstieg an freiem Prolin (Barnett and Naylor, 1966; Singh etal., 1972 u.a.), eine Reduktion der Photosynthese (UbGrsicht Hsiao, 1973) und eine Zunahme der 16slichen Kohlenhydrate in den Pflanzen verbunden (Stewart, 1971). Eine direkte Wirkung der ABA auf diese Prozesse kann daher sieher nieht ausgeschlossen werden. Da es m6glieh ist, den Anstieg des Prolins (Aspinall et al., 1973) und die Hemmung der Photosynthese (Mittelheuser und Van Steveninek, 1969; WGllburn et aI., 1973; Sankhla und Huber, 1974a) aueh dutch exogene Applikation yon ABA auf Pflanzen zu erzielen, scheint es uns gerechtfertigt, die Zunahme des ABA-Gehaltes in den Pflanzen untGr StreBbedi~gungen durch Anzucht dGr Pflanzen in Absei- sins/turel6sungen zu simulieren, um damit Gin fibersehaubares System ffir die Untersuchung der ABA-Wirkungen auf Enzymaktivit/~ten zu haben.
W~hrend fiber den Einflul] yon ABA auf die Enzyme des Amino- s/~urestoffweehsels and auf die Photosynthese sehon beriehtet wurde (Sankhla und Huber, 1974; Sankhla und Huber, 1974a; Huber und Sankhla, 1974), soll hier geprfift werden, ob Enzyme des Kohlenhydrat- stoffweehsels beeinflugt werden k6nnen, da auch der Kohlenhydrat- umsatz unter Stregbedingungen ver/~ndert wird (Stewart, 1971). Parallel dazu wurdGn auch die Wirkungen yon Gibberellins/~ure, allein and in Kombination mit ABA, auf die Enzyme gemessen. GAa hat zwar - - soweit bekannt - - keinen direkten EinfluB auf den Wasserhaushalt, macht jedoeh z.B. die mit der Erh6hung des ABA-Spiegels verbundene Zunahme des freien Prolins wieder rfickg/~ngig (Singh et al., 1973).
Material und Methoden Pflanzenmaterial. Als Material wurde Penni~etum typhoide.s (Burm. f.) Stapf
& Hubbard verwendet. Die Keimlinge wurden in Petrisch~len angezogen, die mit Filterpapier ausgelegt waren, welches mit 5 ml dest. Wasser oder mit der Testl6sung angefeuehtet wurde. Die Testl6sungen enthielten 3,8-10 6 ~, 7,4.10-6 M oder 1,48.10 -5 M ABA oder 2,89.10 -5 M bzw. 2,89.10 -~ M GA~. Die Keimlinge wurden 4 Tage bei 28~-2~ und ca.. 3000 lx (Osram-L Fluora-Lampen) aufgezogen und dann die BI~itter und Koleoptilen zum Test verwendet.
Enzymextraktion. Etw~ 2 g Pflanzenmaterial, bestehend aus den ersten Bl~ttern und den Koleoptilen, wurden mit 8 ml Isolierungsmedium (0,05 M Tris-HC1-Puffer pH 7,0; 0,001 M EDTA; 0,003 M ~gC12 und 0,02% v/v Mercapto~thanol) in einem Potter-Elvehjem Homogenisator (Potter S, Braun, Melsungen, 1200 Umin) etwa 2 min unter Eiskiihlung m6glichst gleichmi~l~ig homogenisiert. Der Rohextrakt wurde sofort in einer Sorvall SS 1 Zentrifuge bei 4~ 15min bei 20000• g ah- zentrifugiert. Aus der fiberstehenden L6sung der Zentrifugation wurden die Proteine mit festem Ammoniumsulfat (0--80% S~ttigung) ausgef~llt. Der Niederschlag wurde durch erneutes Zentrifugieren gesammelt (15 rain bei 20000• und mit

Einflul3 yon ABA und GAa aug Enzyme 57
2 ml Medium (jedoch ohne Mercaptoi~thanol) wieder aufgenommen. Diese L6sung wurde als 16sliche Enzymfraktion verwendet.
Bestimmung der Enzymalctivitiiten. Die Messung der Enzymaktivit~ten effolgte nach tIuber et al. (1969, 1973), Fekete et al. (1967) und Kennecke et al. (1971). Zu den Enzymbestimmungen wurden folgende Inkubationsgemische verwendet:
1. Phosphorylase (~-l,4-glucan-orthophosphat- glucosyltrans]erase, EC 2.4.1.1)
0,25 fzmol G-1-P14C (16000 imp), 35 fzmol Citratpuffer pH 6,1, 10 tzl Amylo- pektin (~ 55 ~zmol Glykogen), 5 fzmol I~aF, 50 ~l 15sliches Enzym in einem Ge- samtvolumen yon 140 ~zl.
2. Amylase (~-l,4-glucan-4-glucanohydrolase, EC 3.2.11 bzw. ~-l,4-glucanmaltohydrolase)
0,18 mg St~rke, 10 ~mol Citratpuffer pI-I 6,1, 10 fzl 16sliches Enzym in einem Gesamtvolumen yon 60 fzl.
3. Saccharose-Synthetase ( UD PG-D-/ruetose-2-glucosyltrans/erase, EC 2.4.1.13)
Saccharose.6-phosphat-Synthetase ( U D PG-D-/ructose-6-phosphat-2-glueosyltrans/erase, EC 2.4.1.14)
Es wurde in Richtung der Saccharosesynthese gemessen. Die Bestimmung der gebildeten Saccharose bzw. des Saccharose-6-phosphats erfolgte nach Roe (1934). 0,2 ~zmol UDPG, 20 ~zmol Glycyl-Glycinpuffer pH 7,8 0,6 tzmol EDTA, 1 tzmol Mg 2+, 0,5 fzmol Fructose bzw. Fructose-6-phosphat, 5 Izmol NaF, 50 ~zl 16sliches Enzym in einem Gesamtvolumen yon 120 ~zl.
4. Invertase (fi-D-/ructo/uranosid-/ructohydrolase, EC 3.2.1.26) oder alkalisehe Saeeharase
Die nichtabgebaute, alkalistabile Saccharose wurde nach Beendigung der Enzymeinwirkung bestimmt.
0,2 ~zmol Saccharose, 4 {zmol Glycyl-Glycinpuffer pH 7,8, 0,12 fzmol EDTA, 0,2 fzmol Mg ~+, 20 {z116sliches Enzym in einem Gesamtvolumen yon 50 ~l.
5. Phosphohexoseisomerase (D-Glucose-6-phosphat-ketol-isomerase, EC 5.3.1.9)
0,4 fzmol Glucose-6-phosphat, 4 ~zmol Glycyl-Glycinpuffer pH 7,3, 0,12 ~zmol EDTA, 0,2 ~zmol Mg 2+ und 20 tzl 15sliches Enzym in einem Gesamtvolumen yon 50 ~I.
Bei allen Enzymbestimmungen wurde 1 h bei 37~C inkubiert. Proteinbestimmung. Die Proteinbestimmungen wurden nach Lowry et al. (1951)
durchgefiihrt. Dabei wurde Rinderserumalbumin als Standard verwendet. Berechnung dcr Ergebnisse. Die Enzymaktivit~ten wurden in tzmol Substrat-
umsatz/mg Protein errechnet und dann in (%) des Kontrollwertes angegeben. Die Experimente wurden mindestens siebenmal wiederholt. Die Ergebnisse wurden statistisch mit dem t-Test abgesicbert. Bei der Diskussion wurden nur solche Werte beriicksichtigt, bei denen die Nullbypothese mit 95 % Wahrscheinlichkeit verwoffen werden konnte.
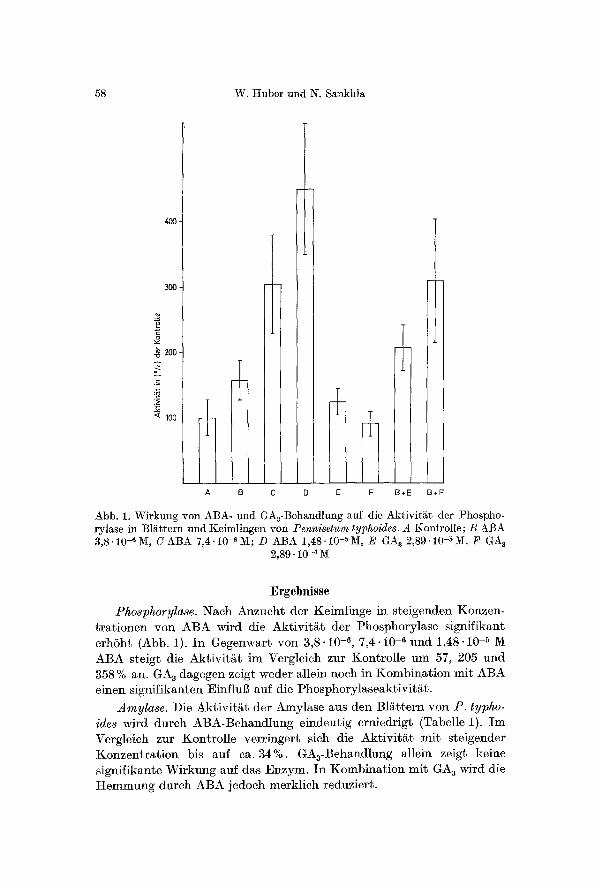
58 W. Huberund N. Sankhla
t, O0-
300-
o
200-
.~_
< 100
A D B+E B+F E F
Abb. 1. Wirkung yon ABA- und GA3-Behandlung auf die Aktivit~t der Phospho- rylase in Bliittem und Keimlingen yon Pennisetum typhoides. A Kontrolle; B ABA 3,8.10-e~, CABA 7,4.10-6M; D ABA 1,48.10-aM, E GA 3 2,89 .10-5 M, F GA~
2,89.10 -a
Ergebnisse Phosphorylase. Nach Anzucht der KeimlirLge in steigenden Koazen-
trationen yon ABA wird die Aktivit&t der Phosphorylase signifik~nt erhSht (Abb. 1). In Gegenwart yon 3,8-10 -6, 7,4-10 -6 und 1,48.10 -5 M ABA steigt die Aktivits im Vergleich zur Kontrolle um 57, 205 und 358% an. GA~ dagegen zeigt weder allein noch in Kombination mit ABA einen sigr~ifikanten Einflu[~ auf die 1)hosphorylaseaktiviti~t.
Amylase. Die Aktivit~t der Amylase aus den Bl~ttern yon P. typho- ides wird durch ABA-Behandlung eir~deutig erniedrigt (T~belle 1). Im Vergleich zur Kontrolle verringert sich die Aktiviti~ mit steigender Konzentration bis auf ca. 34%. GA3-Beh~ndlung allein zeigt keine signifikante Wirkung auf das Enzym. In Kombinatior~ mit GA~ wird die Hemmung durch ABA jedoch merklich reduziert.
Einflul~ yon ABA und GA sauf Enzyme 59
Tabelle 1. Wirkung yon ABA- und GAs-Behandlung auf die Aktivit~it der Amylase in Bli~ttern und Koleoptilen yon Pennisetum typhoides
Behandlung Aktivi t~t in ( % ) der Kontrolle
Kontrolle 100 • 18,75
ABA 3,8 - 10 -6 M 106,1 ~_ 26,1
ABA 7,4 �9 10 -6 M 71,6 • 18,0
ABA 1,48-10 -~ iV[ 33,6 • 13,75
GA s 2,89.10 -~ M 96,5 • 21,7
GA a 2,89.10 -~ ~r 112,8 ~- 20,05
ABA ~- GA s 3,8 �9 l0 -6 M 74,7 ~- 19,85 2,89.10 -s M
ABA ~- GA s 7,4 - 10 -6 M 61,0 • 11,82 2,89.10 -s M
Tabelle 2. Wirkung von ABA- und GAs-Behandlung auf die Aktivi t~t der Invertase in BlOtter und Koleopti]en yon Pennisetum typhoides
Behandlung Aktivi t~t in ( % ) der Kontrolle
Kontrolle 100 ~ 21,8
ABA 3,8 �9 10 -6 M 78,4 • 18,8
ABA 7,4 �9 10 -G ~ 78,5 • 18,1
ABA 1,48' 10 -5 M 83,3 • 13,75
GA a 2,89" 10 ~ M 146,4 • 26,54
GA s 2,89" 10 -4 M 145,9 • 26,1
ABA + GA s 3,8 " 10 -6 M 130,3 • 24,58 2,89" 10 -5 M
ABA ~- GA 3 7,4 " 10 -6 M 104,2 • 24,83 2,89" 10 -5 Yi
Invertase. A u c h die I n v e r t a s e w i r d d u r c h d ie A B A - B e h a n d l u n g ge-
h e m m t , j e d o c h n i c h t in g l e i c h e m Ausmal3 wie die A m y l a s e (Tabe l l e 2).
I m V e r g l e i c h z u r K o n t r o l l e e r n i e d r i g t s ich die A k t i v i t s u m 2 0 % , w o b e i
d ie A k t i v i t ~ t s a b n a h m e n i c h t m i t e i n e r Z u n a h m e d e r A B A - K o n z e n t r a t i o n
k o r r e l i e r t is t . G A 3 d a g e g e n vergr613ert d ie E n z y m a k t i v i t s s i g n i f i k a n t
u n d b e w i r k t in K o m b i n a t i o n m i t A B A e ine A u f h e b u n g d e r d u r c h A B A
h e r v o r g e r u f e n e n H e m m u n g .
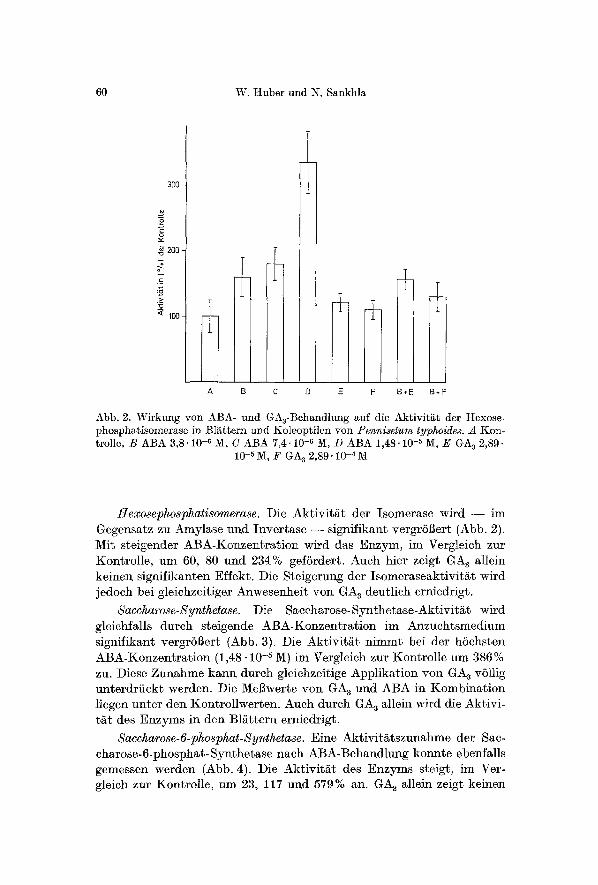
60 W. Huber und N. Sankhla
300 -
200-
,s
< 100-
A E F:
-[-
B C D B * E B * F
Abb. 2. Wirkung yon ABA- und GA3-Behandlung auf die Aktiviti~t der Hexose- phosphafisomerase in Illi~ttern und Koleop%iien yon Pennisetum typhoides. A t{on- trolle, B ABA 3,8.10 -6 M, C ABA 7,4-10 -6 M, D ABA 1,48.10 -s M, E GA a 2,89.
10 -5 IV[, F GA s 2,89.10 -a M
Hexosephosphatisomerase. Die Akt iv i t / t t der I somerase wird - - im Gegensatz zu Amylase und In~er t a se - - s ignif ikant ve rg r6ger t (Abb. 2). Mit sgeigender ABA-Konzen t r a$ ion wird das Enzym, im Vergleich zur KontroI le , um 60, 80 und 234% gef6rdert . Auch hier zeigt GA s al lein ke inen s igni f ikanten Effekt . Die Ste igerung der Isomeraseakt ivi t i~t wird jedoch bei gleichzei t iger Anwesenhe i t yon GA 8 deut l ich erniedrigt .
Sacchctrose-Synthetase. Die Saccharose-Synthe tase-Akt iv i t /~ t wird gleichfal ls du tch s te igende A B A - K o n z e n t r a t i o n im Anzuch t smed ium s ignif ikant vergr66er t (Abb. 3). Die Akt iv i t / i t n i m m t bei der h6chs ten A B A - K o n z e n t r a t i o n (1,48.10 -s IV[) im Vergleich zur Kon t ro l l e urn 386 % zu. Diese Zunahme k a n n dm-ch gleichzei t ige App l ika t ion yon GA s v6llig un te rdr f i ck t werden. Die Mel3werte von GA a und A B A in K o m b i n a t i o n ]iegen un te r den Kont ro l lwer ten . Auch durch GA s al lein wird die Akt iv i - ti~t des E n z y m s in den Bl&ttern erniedr igt .
Saccharose-6-phosphat-Synthetase. Eine Akt iv i t&tszunahme der Sac- cha rose -6 -phospha t -Sya the t a se nach A B A - B e h a n d l u n g kormte ebenfal ls gemessen werden (Abb. 4). Die Aktivit /~t des E n z y m s steigt , im Ver- gleich zur Kont ro l le , um 23, 117 und 579% an. GA~ al lein zeigt ke inen
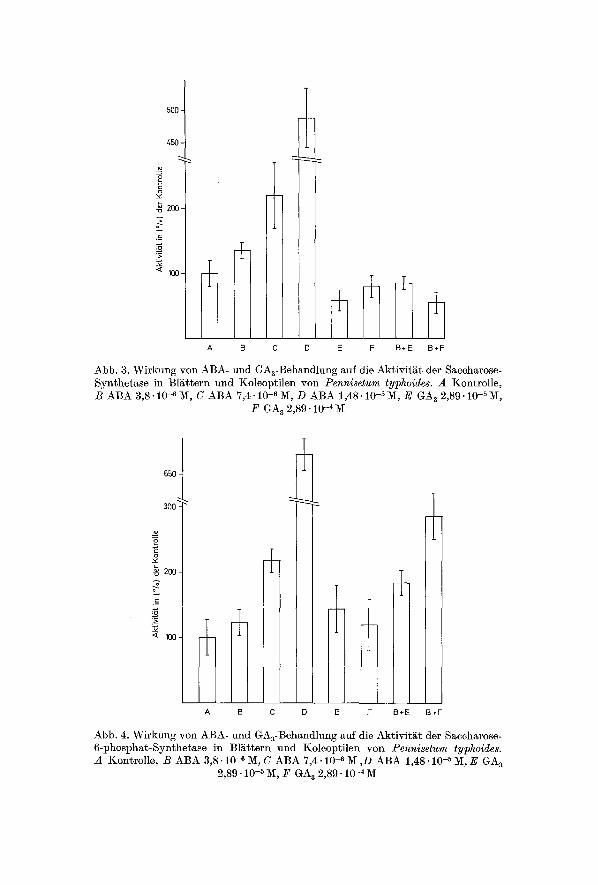
500 -
4 5 0 -
#, o
~c
-~ 200 -
._c
< 100
--I-
A B
t C D E F B§ B+F
Abb. 3. Wirkung yon ABA- und GAa-Behandlung auf die Aktivit~it der Saccharose- Synthetase in BIEttern und Koleoptilen yon Pennisetum typhoides. A Kontrolle, B ABA 3,8.10 _6 M, C ABA 7,4.10 -6 M, D ABA 1,48.10-5 M, E GA s 2,89.10-5 M,
F GA~ 2,89.10 -~
65O
300"
A
2oo
.E
< 100
B
+
C D
1 F B+E B*F
Abb. 4. Wirkung von ABA- und GAa-Behandlung auf die Aktivit/it der Saccharose- 6-phosphat-Synthetase in Bl~ttern und Koleoptilen yon Pennisetum typhoides. A Kontrolle, B ABA 3,8.10 -~ iK, C ABA 7,4- 10 -~ 1~ ,D ABA 1,48.10 -5 M, E GA a
2,89.10 -5 M, F GA a 2,89.10-aM

62 W. Huber und N. Sankhla
signifikanten Ein~luB, aueh in Kombination mit ABA wird keine Wir- kmlg siehtbar.
Diskussion
Die Untersuchungen fiber den Ei~luB yon exogener ABA- und GAs-Zugabe auf die Enzyme des Kohlenhydratstoffwechsels in Keim- lingen yon Pennisetum typhoides sollten AufschluB darfiber geben, wie der Kohlenhydrats~offwechsel durch diese I-Iormone beeinitul3t wird.
Wie bei anderen Objekten (zusammen~assende (~bersicht, Addicott, 1972) wurde auch in Bl~tttern yon P. typhoides die Amylase und die Invertase durch ABA-Behandlung gehemmt. Bei allen anderen unter- suchten Enzymen dagegen fanden wir eine deutliche Steigerung der Enzymaktivit~ten. ~'ber die Wirkung yon ABA guf diese Enzyme, die wichtige Funktionen bei der Phosphorolyse der St~rke, ir~ der Glykolyse oder bei der Saccharose-Sy-nthese haben, wurde bisher noch nicht berichtet.
Betrachtet man die geschilderten Ergebnisse in Verbindung mit den Vers des Kohlenhydratgehaltes in den Pflanzen unter Wasser- oder SalzstreB, so stellt man eine weitgehende ~Jbereil~stimmung lest. Die mit dem Anstieg der ABA-Konzentration verbundene Zunahme der 16slichen Zucker - - vor allem der Saccharose - - kann durch die Akti- vierung der Phosphorylase, I-Iexosephosphatisomerase, Saccharose- und Saccharose-6-phosphat-Synthetase dutch ABA kausal hervorgerufen werden. Die F6rderung beso~tders der Saccharose-Synthese ist wichtig, da nur bei Anwesenheit yon Saccharose der Spiegel des freiea Prolins aufrecht erhalten wird, der sich unter StreBbedingungen oder ABA-Be- handlung angesammelt hat (Stewart, 1971, 1972). Diese Saccharose- anh~ufung wird welter durch die Hemmung der Invertase durch ABA begfinstigt. ])as Prolin hat offer~bar eine wichtige Funktion beim lJber- dauern yon StreBeinwirkungen. Es wird aus Glutaminsaure gebildet (Barnett nnd Naylor, 1966), die wiederum mit den yon uns schon fr~her diskutierten Mechanismen bei ABA-Behandlung oder unter SalzstreB vermehrt synthetisiert werden karm (Sankhla und I-Iuber, 1974; Huber und Sankhla, 1973). Der Stickstoff bleibt duher den Pflanzen in Form des Prolins auch unter StreBbedingungea erhalten und kann bei Eintri t t gfinstiger Bedingungen wieder zur Neusynthese von Protein nutzbar gemacht werden.
GA 8 verhi~lt sich auch hier, wie bei einigen anderen Vorgitngen in der Pflanze (Addicott, 1972), antagolfistisch zur ABA. Vor allem durch die F6rderung der Invertase und durch die Hemmung oderNichtbeeinflussuag der anderen Enzyme kann der Kohlenhydratgehalt unter GA 3 Ein- wirkung abgebaut werden, oder er wird bei hohem GAa-Spiegel erst gar

Einflul3 yon ABA und GA s auf Enzyme 63
nicht angehoben. Dami t ist auch die in der Ein le i tung erw~hnte schaelle
Oxidatiorr des Prolins un te r GAs-EinftuB zu erkl/iren. Ob diese Befurrde aueh direkt auf strel3behandelte Pf lanzen zutreffen,
wird yon uns gerade gepriift. Es ist jedoch anzunehmen, dal3 ABA u n d GA s un te r Stref3bedingungen prim/~r auf dem enzymat isehen Niveau eingreifen und dal3 eventuel l aueh die Regelung des Wasserhaushal tes fiber die S tomatabewegung auf diese Effekte zuriiekzuffihren ist.
Wir danken Prof. H. Ziegler fiir fruchtbare Diskussionen und Anregungen und Frl. S. Ballzus far sorgfEltige technische Assistenz. Die Untersuehung wurde von der Deutschen Forschungsgemeinschaft und yon der Alexander-von-Humboldt- Stiftung durch eine Sachbeihilfe unterst~itzt.
Literatur Addicott, F. T. : Biochemical aspects of abscisic acid. In: Plant growth substances
1970, p. 272--280, Carr, D. J., ed. Berlin-Heidelberg-New York: Springer 1972 Aspinall, D., Singh, N. T., Paleg, L. G.: Stress metabolism. V. Abscisic acid and
nitrogen metabolism in barley and Lolium temulentum L. Aust. J. biol. Sci. 26, 3 1 9 ~ 2 7 (1973)
Barnett, N. M., Naylor, A. W.: Amino acid and protein metabolism in Bermuda grass during water stress. Plant Physiol. 41, 1222--1230 (1966)
Fekete, M. A. R. de, Ziegler, H., Wolf, R. : Enzyme des Kohlenhydratstoffwechsels in Nektarien. Planta (Berl.) 75, 125--138 (1967)
Hsiao, T. C. : Plant responses to water stress. Arm. Rev. Plant Physiol. 24,519--570 (1973)
Huber, W., Fekete, M. A. R. de, Ziegler, H.: Enzyme des St~rkeumsatzes in Biindelscheiden- und Palisadenzellchloroplasten von Zea mays. Planta (Berl.) 87, 360--364 (1969)
Huber, W., Fekete, M. A. R. de, Ziegler, H. : Enzyme des St~rkeumsatzes in den Wurzelhaubenzellen yon Zea mays L. Planta (BEE.) l l2,343--356 (1973)
Huber, W., Sankhla, N.: Eco-physiological studies on Indian arid zone plants. II. Effect of salinity and gibberellin on the activity of the enzymes of amino-acid metabolism in leaves of Pennisetum typhoides. Oecologia (Berl.) 13, 271--277 (1973)
Huber, W., Sankhla, N. : Effect of gibberellic acid on the activities of photosyn- thetic enzymes and laCO~ fixation products in leaves of Pennisetum typhoides seedlings. Z. Pflanzenphysiol. 71, (1974)
Jones, R. J., Mansfield, T. A. : Suppression of stomatal opening in leaves treated with abscisic acid. J. exp. Bot. 21, 714--719 (1970)
Kennecke, M., Ziegler, H., Fekete, M. A. R. de: Enzymaktivit/iten im SiebrShren- saft yon Robinia pseudoacacia L. und anderen Baumarten. Planta (Berl.) 98, 330--356 (1971)
Kriedemann, P. E., Loveys, B. R., Fuller, G. L., Leopold, A. C.: Abscisic acid and stomatal regulation. Plant Physiol. 49, 842--847 (1972)
Lowry, O. H., Rosebrough, N. J., Lewis, A., Randall, R. J. : Protein measurement with the folin-phenol reagent. J. biol. Chem. 193, 265--275 (1951)
Mittelheuser, C. J., van Steveninck, R. F. H. : Stomatal closure and inhibition of transpiration by (RS)-abscisic acid. Nature (Lond.) 221, 281--282 (1969)
5 Planta (Berl.), Vol. 116

64 W. Huber und N. Sankhla
Mizrahi, Y., Blumenfeld, A., Bittner, S., Richmond, A. E. : Abseisic acid and cyto- kinin contents of leaves in relation to salinity and relative humidity. Plant Physiol. 48, 752--755 (1971)
Mizrahi, Y., Blumenfeld, A., Richmond, A. E. : Abscisic acid and transpiration in leaves in relation to osmotic root stress. Plant Physiol. 46, 169--171 (1970)
Mizrahi, Y., Blumenfeld, A., Richmond, A. E. : The role of abscisic acid and salini- ration in the adaptive response of plants to reduced root aeration. Plant & Cell Physiol. 1], 15--21 (1972)
Roe, J. H. : A colorimetric method for the determination of fructose in blood and urine. J. biol. Chem. 107, 15--22 (1934)
Sankhla, N., Huber, W. : Enzyme activities in Pennisetum seedlings germinated in the presence of abscisie and gibberellic acids. Phytochemistry 1] (1974)
Sankhla, N., Huber, W." Effect of abscisic acid on the activities of photosynthetic enzymes and ltCO~-fixation products in leaves of Pennisetum typhoides seedlings. Physiol. Plant., in press (1974a)
Singh, T. N., Aspinall, D., Paleg, L. G. : Proline accumulation and adaptability to drought in barley: a potential metabolic measure of drought resistance. Nature (Lond.) New Biol. 286, 188--190 (1972)
Singh, T. N., Aspinall, D., Paleg, L. G.: Stress metabolism IV. The influence of (2-chloroethyl)trimethylammonium chloride and gibberellic acid on the growth and proline accumulation of wheat plants during water stress. Aust. J. biol. Sci. 26, 77--86 (1973)
Stewart, C. R. : Effect of wilting on carbohydrates during incubation of excised Bean leaves in the dark. Plant Physiol. 48, 792--794 (1971)
Stewart, C. R.: Proline content and metabolism during rehydration of wilted excised leaves in the dark. Plant Physiol. 50, 679--681 (1972)
Tal, M., Imber, D.: Abnormal stomatal behaviour and hormonal imbalance in flacca, a wilty mutant of tomato I I I . Hormonal effects on the water balance in the plant. Plant Physiol. 47, 849--850 (1971)
Wellburn, F. A. M., Wellburn, A. R., Stoddart, J . L., Treharne, K. J. : Influence of gibberellic and abscisic acids and the growth retardant CCC upon plastid devel- opment. Planta (Berl.) l l l , 337--346 (1973)





























