Embed Size (px)
Citation preview

Untersuchungen zur Rolle von Alb3 und Alb4 bei der Biogenese der Thylakoidmembran
in Arabidopsis thaliana
Dissertation zur Erlangung des Grades
eines Doktors der Naturwissenschaften
der Fakultät für Biologie und Biotechnologie
an der Internationalen Graduiertenschule Biowissenschaften
der Ruhr-Universität Bochum
angefertigt in der
AG Molekularbiologie pflanzlicher Organellen
vorgelegt von
Thomas Bals aus
Unna
Bochum
Oktober, 2009
Erstgutachterin: Frau Prof. Dr. Danja Schünemann
Zweitgutachter: Herr Prof. Dr. Matthias Rögner

Analyses of Alb3 and Alb4 in thylakoid membrane biogenesis
in Arabidopsis thaliana
Dissertation to obtain the degree
Doctor Rerum Naturalium (Dr. rer. nat.)
at the Faculty of Biology and Biotechnology
Ruhr-University Bochum
International Graduate School of Biosciences Ruhr-University Bochum
Molecular Biology of Plant Organelles
submitted by
Thomas Bals
from
Unna
Bochum
October, 2009

Inhaltsverzeichnis I
Inhaltsverzeichnis
1. Einleitung...................................................................................................... 1
1.1 Funktion und evolutionärer Ursprung der Chloroplasten.................................1 1.2 Proteintransport kernkodierter Proteine über die chloroplastidären
Hüllmembranen...............................................................................................1 1.3 Proteintransport zu den Thylakoiden ..............................................................2 1.3.1 Proteinimport in das Thylakoidlumen ..............................................................3 1.3.1.1 Der cpSec-Weg...............................................................................................4 1.3.1.2 Der cpTat-Weg................................................................................................4 1.3.2 Proteinintegration in die Thylakoidmembran...................................................5 1.3.2.1 Spontane Insertion ..........................................................................................5 1.3.2.2 Der cpSRP-Weg..............................................................................................5 1.4 Proteininteraktionen der Komponenten des posttranslationalen cpSRP-
Transportwegs ................................................................................................8 1.5 Die Oxa1/YidC/Alb3-Proteinfamilie ...............................................................10 1.5.1 Oxa1..............................................................................................................11 1.5.2 Yid C .............................................................................................................13 1.5.3 Alb3 ...............................................................................................................14
2. Zielsetzung ................................................................................................. 17
3. Material und Methoden .............................................................................. 18
3.1 Chemikalien ..................................................................................................18 3.2 Verbrauchsmaterial .......................................................................................19 3.3 Geräte ...........................................................................................................20 3.4 Biologisches Material ....................................................................................21 3.4.1 Enzyme .........................................................................................................21 3.4.2 Antikörper......................................................................................................21 3.4.3 Verwendete Plasmide ...................................................................................22 3.4.4 Erstellte Plasmide und verwendete Oligonukleotide .....................................23 3.4.5 Mikroorganismen...........................................................................................29 3.4.5.1 Escherichia coli .............................................................................................29 3.4.5.2 Saccharomyces cerevisiae ...........................................................................29 3.4.6 Pflanzenmaterial ...........................................................................................29 3.5 Anzuchtbedingungen ....................................................................................30 3.5.1 Anzucht und Lagerung von Escherichia coli .................................................30 3.5.2 Anzucht und Lagerung von Saccharomyces cerevisiae................................30 3.5.3 Pflanzenanzucht............................................................................................31 3.6 Molekularbiologische Methoden....................................................................31 3.6.1 Polymerase-Kettenreaktion zur Amplifizierung von DNA-Fragmenten .........31 3.6.2 Site-directed Mutagenese PCR.....................................................................32 3.6.3 Overlap PCR .................................................................................................33 3.6.4 DNA-Restriktion ............................................................................................34 3.6.5 Ligation..........................................................................................................35 3.6.6 Herstellung CaCl2-kompetenter Escherichia coli-Zellen................................35 3.6.7 Transformation von Plasmiden in CaCl2-kompetente Escherichia coli-
Zellen ............................................................................................................35 3.6.8 In-Fusion Klonierung .....................................................................................36 3.6.9 Plasmid-Präparation......................................................................................36 3.6.10 DNA-Konzentrationsbestimmung..................................................................37

Inhaltsverzeichnis II
3.6.11 Restriktionsanalyse und Sequenzierung.......................................................37 3.6.12 Agarosegelelektrophorese zur Trennung von DNA.......................................37 3.6.13 Split-Ubiquitin System ...................................................................................38 3.6.13.1 Transformation von Saccharomyces cerevisiae mit Plasmid-DNA ...............39 3.6.13.2 β-Galaktosidase Filtertest .............................................................................39 3.6.14 Bimolekulare Fluoreszenzkomplementation (Split-YFP System) ..................40 3.6.14.1 Isolierung von Arabidopsis thaliana Protoplasten und transiente
Transfektion ..................................................................................................41 3.7 Biochemische Methoden...............................................................................42 3.7.1 Überexpression von Proteinen in Escherichia coli ........................................42 3.7.2 Denaturierende Aufreinigung von Proteinen mit His-tag über Ni-NTA..........42 3.7.3 Native Aufreinigung von Proteinen mit His-tag über Ni-NTA.........................43 3.7.4 Aufreinigung von Proteinen mit GST-tag über Glutathion-Sepharose ..........44 3.7.5 Abspaltung des GST-tags über Thrombinverdau..........................................45 3.7.6 Dialyse von Proteinen ...................................................................................45 3.7.7 BCA-Proteinbestimmung...............................................................................45 3.7.8 SDS-Polyacrylamid-Gelelektrophorese (SDS-PAGE)...................................45 3.7.9 Coomassie-Färbung......................................................................................47 3.7.10 Elektrophoretischer Transfer von Proteinen auf
Nitrocellulosemembranen (Western-Blot) .....................................................47 3.7.11 Proteinfärbung mit Ponceau S ......................................................................47 3.7.12 Immundetektion von Proteinen .....................................................................48 3.7.13 Affinitätsaufreinigung von Antikörpern ..........................................................49 3.7.14 In vitro Transkription......................................................................................50 3.7.15 In vitro Translation.........................................................................................50 3.7.16 In vitro Expression mit RTS 100 Wheat Germ ..............................................51 3.7.17 Pull down Experiment ...................................................................................51 3.7.18 Peptide-Scanning ..........................................................................................52 3.7.18.1 Bindung des rekombinanten Proteins an die PepSpot-Membran .................52 3.7.18.2 Elektrophoretischer Transfer eines rekombinanten Proteins von der
PepSpot-Membran auf eine PVDF-Membran ...............................................53 3.7.18.3 Regeneration der PepSpot-Membran ...........................................................54 3.7.19 Präparation von Thylakoidmembranen aus Arabidopsis thaliana .................55 3.7.19.1 Chlorophyllbestimmung.................................................................................56 3.7.19.2 Salzwaschung der Thylakoidmembranen .....................................................56 3.7.19.3 Solubilisierung der Thylakoidmembranen .....................................................56 3.7.20 Crosslinking der Thylakoidmembranproteine von Arabidopsis thaliana........57 3.7.21 Coimmunpräzipitation ...................................................................................57 3.7.21.1 Kopplung des Antikörpers an Protein-A-Sepharose beads...........................57 3.7.21.2 Präzipitation ..................................................................................................58 3.7.22 Thylakoidmembran-Bindungstest..................................................................58 3.7.23 Thylakoidmembran-Bindungstest mit Peptidinhibierung ...............................59 3.7.24 Expressionsnachweis der YFP-Fusionsproteine in Arabidopsis thaliana
Protoplasten ..................................................................................................59 3.7.25 Trichloressigsäure-Präzipitation....................................................................60 3.8 Biophysikalische Methoden...........................................................................60 3.8.1 Massenspektrometrie....................................................................................60 3.8.2 Gelfiltration ....................................................................................................61 3.8.2.1 Eichung der Gelfiltrationssäule .....................................................................61 3.8.2.2 Gelfiltration der solubilisierten Thylakoidmembranproteine...........................62

Inhaltsverzeichnis III
4. Ergebnisse.................................................................................................. 63
4.1 Herstellung spezifischer Antikörper gegen Alb3 und Alb4 ............................63 4.1.1 Expression der Antigene GST-nAlb3, GST-cAlb3 und cAlb4-his ..................64 4.1.2 Test der Antikörper α-Alb3 und α-Alb4 gegen die Antigene..........................65 4.1.3 Test der Antikörper α-Alb3 und α-Alb4 gegen die in vitro translatierten
Volllängenproteine ........................................................................................67 4.1.4 Test der Antikörper α-Alb3 und α-Alb4 mit Thylakoidmembran-proteinen ....68 4.2 Analyse der Interaktionen von Alb3 und Alb4 mit den Proteinen des
cpSRP-Transportwegs ..................................................................................69 4.2.1 Interaktionen von Alb3 und Alb4 mit den Proteinen des cpSRP-
Transportwegs im Hefe Split-Ubiquitin System.............................................69 4.2.2 Proteininteraktionen von Alb3 und Alb4 mit den Proteinen des cpSRP-
Transportwegs im Split-YFP System ............................................................71 4.2.3 Analyse der Binderegion zwischen Alb3 und cpSRP43 über das Split-
YFP System ..................................................................................................76 4.2.4 Peptide Scanning von Alb3 mit rekombinantem cpSRP43 ...........................80 4.2.5 Bestätigung der mittels Peptide Scanning gezeigten Binderegionen im
Split-YFP System ..........................................................................................82 4.2.6 Verifizierung der Binderegionen im Hefe Split-Ubiquitin System ..................84 4.2.7 Pull down Analysen zur Interaktion von Alb3 und cpSRP43.........................86 4.2.8 Untersuchung der Alb3 Bindestelle im Thylakoidmembran-Bindungstest.....89 4.2.9 Analyse der Bindedomäne des cpSRP43 für das Alb3.................................90 4.3 Analyse der Interaktion von Alb3 und Alb4 mit den Proteinen der
Thylakoidmembran........................................................................................95 4.3.1 Crosslinking der Thylakoidmembranproteine................................................95 4.3.2 Interaktion von Alb4 mit den kernkodierten ATP-Synthase
Untereinheiten im Split-YFP System.............................................................97 4.3.3 Gelfiltrationsanalyse solubilisierter Thylakoidmembranproteine .................100 4.3.4 Coimmunpräzipitation solubilisierter Thylakoidmembranproteine ...............101
5. Diskussion ................................................................................................ 103
5.1 Die cpSRP-Untereinheit cpSRP43 interagiert direkt mit dem integralen Membranprotein Alb3..................................................................................103
5.2 Identifizierung der Alb3-Interaktionsbereiche für das cpSRP43 und Charakterisierung der Alb3-cpSRP43 Interaktion .......................................105
5.3 Das cpSRP43 interagiert mit Alb3 über die Ankyrin repeat Region ............110 5.4 Alb4 interagiert nicht mit den Komponenten des cpSRP-Transportwegs ...115 5.5 Alb4 ist mit der chloroplastidären ATP-Synthase assoziiert und interagiert
nicht mit Alb3 und cpSecY ..........................................................................116
6. Zusammenfassung .................................................................................. 120
7. Summary................................................................................................... 121
8. Literaturverzeichnis ................................................................................. 122
9. Abkürzungsverzeichnis........................................................................... 131
10. Abbildungsverzeichnis............................................................................ 132
11. Tabellenverzeichnis ................................................................................. 134

Inhaltsverzeichnis IV
12. Anhang...................................................................................................... 135
12.1 Publikationen...............................................................................................135 12.2 Lebenslauf...................................................................................................136 12.3 Danksagung ................................................................................................137

1 Einleitung 1
1. Einleitung
1.1 Funktion und evolutionärer Ursprung der Chloroplasten
Die Chloroplasten der photoautotrophen Eukaryoten ermöglichen eine Vielzahl an
biochemischen Funktionen. Dazu gehören die Synthese von Fettsäuren, Aminosäuren
und Porphyrinen sowie die Sulfat- und Stickstoffreduzierung. Die allgemein bekannteste
Funktion der Chloroplasten ist die Fixierung von Kohlenstoff durch die Photosynthese. Bei
der Photosynthese werden Kohlendioxid und Wasser zu Glukose und Sauerstoff
umgesetzt. Mit der Existenz von Chloroplasten können die photoautotrophen Eukaryoten
die absorbierte Strahlungsenergie der Sonne in chemisch nutzbare Energieträger
umwandeln. Die Endosymbiontentheorie beruht darauf, dass die Chloroplasten über
Phagozytose eines photosynthetisch aktiven Prokaryoten durch eine heterotrophe
Wirtszelle entstanden sind (Weeden, 1981; Cavalier-Smith, 2000). Man nimmt an, dass
der phagozytierte Prokaryot den heutigen Cyanobakterien ähnelte (Reyes-Prieto et al.,
2007). Im Laufe der Evolution erfolgte neben einem Verlust auch ein Transfer von
genetischer Information aus dem Genom des Endosymbionten in das Kerngenom der
Wirtszelle (Martin und Herrmann, 1998). Chloroplasten enthalten etwa 3000 Proteine
(Abdallah et al., 2000). Davon sind nur etwa 100 Proteine im Chloroplastengenom selbst
kodiert (Timmis et al., 2004). Die Mehrheit der Proteine (> 95%) ist kernkodiert und wird
posttranslational nach erfolgter Synthese im Cytosol in die Chloroplasten importiert
(Keegstra und Cline, 1999; Leister, 2003). Der Transfer der Gene in das Kerngenom
erforderte die Entstehung einer koordinierten Protein-Import-Maschinerie (Soll und
Schleiff, 2004; Kessler und Schnell, 2006; Jarvis, 2008; Agne und Kessler, 2009; Benz et
al., 2009).
1.2 Proteintransport kernkodierter Proteine über die chloroplastidären Hüllmembranen
Chloroplasten sind komplex strukturierte Organellen, die sich aus drei Membransystemen
(innere und äußere Hüllmembran und Thylakoidmembran) und drei löslichen
Kompartimenten (Intermembranraum der Hüllmembranen, Stroma und Thylakoidlumen)
zusammensetzen. Viele Proteinkomplexe der Chloroplasten bestehen aus Proteinen, die
im Zellkern bzw. auf dem Plastom kodiert sind. Die Assemblierung dieser Komplexe
erfordert einen koordinierten Import der Proteine in das Organell. Für die korrekte
Sortierung und den Import über die chloroplastidären Hüllmembranen tragen fast alle

1 Einleitung 2
Proteine am N-Terminus ein 20-150 Aminosäuren langes Transitpeptid (Bruce, 2000,
2001). Diese Proteine werden als Vorstufenproteine bezeichnet. Cytosolische Faktoren
können den Transport des Vorstufenproteins erleichtern und die Effizienz des
chloroplastidären Imports der Proteine erhöhen. Bei diesen Faktoren handelt es sich um
14-3-3 Proteine sowie das Hsp70 und Hsp90 Chaperon, die zusammen mit dem
Vorstufenprotein einen guidance-Komplex bilden (May und Soll, 2000; Jarvis 2008). Der
Import der Proteine erfolgt über Translokationskomplexe, die in den chloroplastidären
Hüllmembranen lokalisiert sind. Hierbei handelt es sich um das Translokon der äußeren
Hüllmembran der Chloroplasten (TOC-Komplex, translocon of the outer envelope
membrane of chloroplasts) und das der inneren Hüllmembran der Chloroplasten (TIC-
Komplex, translocon of the inner envelope membrane of chloroplasts). Die Translokation
des Proteins vollzieht sich in mehreren Schritten und ist ein aktiver ATP- und GTP-
abhängiger Prozess. Im Stroma des Chloroplasten wird das Transitpeptid proteolytisch
abgespalten (Richter und Lamppa, 1998). Nach Abspaltung des Transitpeptids wird das
Protein durch stromale Chaperone gebunden (Jackson-Constan et al., 2001). Diese
dienen dazu Proteinaggregationen zu vermeiden, die Faltung des funktionsfähigen
Proteins zu unterstützen und die Proteine in einem transportfähigen Zustand für einen
möglichen Transport zur Thylakoidmembran zu halten.
1.3 Proteintransport zu den Thylakoiden
Nach beschriebener Translokation über die Hüllmembranen der Chloroplasten erfolgt die
Sortierung der Proteine innerhalb des Organells an ihren Wirkort. Für den koordinierten
Transport der chloroplastidären Proteine über die Thylakoidmembran und die Integration
der Proteine in die Membran sind bisher vier verschiedene Transportwege bekannt
(Abbildung 1-1). Der cpSec-Weg und der cpTat-Weg ermöglichen den Transport und die
Translokation von lumenalen Proteinen. Der cpSRP-Weg und die spontane Insertion sind
für die Integration von integralen Membranproteinen verantwortlich. An dem Transport der
Proteine innerhalb des cpSec-Wegs, cpTat-Wegs und cpSRP-Wegs sind verschiedene
Protein- und Energiekomponenten beteiligt. Die spontane Insertion der Proteine erfolgt
ohne weiteren Einfluss anderer Faktoren (Schünemann, 2004, 2007; Aldridge et al.,
2009).

1 Einleitung 3
cpSRP
LHCPs
GTPATP stim.
cpFtsY
cpSRP
Thylakoidmembran
Thylakoidlumen
spontan
cpTat-Translokase
cpTat
PMF
Hcf106 TatC Tha4
cpSec
cpSec-Translokase
SecY SecE
TD
ATP cpSecA
Transport zum Lumen Transport zur Membran
z. B. OE23, OE16 z. B. OE33, PC z.B. CF0II, PsbW, PsaKTD
TD
Alb3
Stroma
Abbildung 1-1: Transportwege der Proteine innerhalb des Chloroplasten Dargestellt sind die Transportwege von kernkodierten Proteinen vom Stroma zur
Thylakoidmembran bzw. in das Thylakoidlumen der Chloroplasten. Der Transport der Proteine in
das Thylakoidlumen wird durch den cpTat-Weg und den cpSec-Weg vermittelt. Der cpSRP-Weg
und die spontane Insertion sind für die Integration von integralen Membranproteinen verantwortlich.
(TD = Transitdomäne; PMF = proton motive force) (verändert nach Schünemann, 2007)
1.3.1 Proteinimport in das Thylakoidlumen
Kernkodierte Proteine des Thylakoidlumens werden sowohl über die Hüllmembranen, als
auch über die Thylakoidmembranen transportiert und tragen dafür ein doppeltes
Transitpeptid. Für den zusätzlichen Transport der Proteine über die Thylakoidmembran ist
neben des N-terminalen Transitpeptids ein zweites Transitpeptid erforderlich. Nach
Abspaltung des ersten Transitpeptids durch eine stromale Peptidase wird das
Transportsignal für die Translokation des Proteins über die Thylakoidmembran
zugänglich. Die Transitpeptide besitzen charakteristische Domänen. Die N-Domäne
besteht aus geladenen Aminosäuren, die H-Domäne enthält vorwiegend hydrophobe
Aminosäuren und die polare C-Domäne trägt das Konsensus-Motiv Ala-X-Ala (Jarvis und
Robinson, 2004).

1 Einleitung 4
1.3.1.1 Der cpSec-Weg
Der chloroplastidäre Sec-Weg zeigt Homologien zum bakteriellen Transportmechanismus
von Sec-abhängigen Proteinen (Robson und Collinson, 2006; Driessen und Nouwen,
2008). In Bakterien bindet das cytosolische Chaperon SecB das Vorstufenprotein und
transportiert dieses zur periplasmatischen Membran. Das periphere Membranprotein
SecA bildet als eine ATPase den Translokationsmotor und schleust die Proteine durch
den Translokationskanal. Die Translokationspore wird von den essentiellen
Hauptkomponenten SecY und SecE gebildet (Brundage et al., 1990). Der cotranslationale
Transport erfolgt über das cytosolische SRP.
Der bakterielle und chloroplastidäre Sec-abhängige Proteintransport erfolgt nach
scheinbar ähnlichen Mechanismen. Die Proteine werden ebenfalls im ungefalteten
Zustand transportiert. Es konnten bereits mit der ATPase cpSecA und den Proteinen der
Translokase cpSecY und cpSecE chloroplastidäre Homologe nachgewiesen werden
(Abbildung 1-1) (Yuan et al., 1994; Laidler et al., 1995; Roy und Barkan, 1998;
Schünemann et al., 1999; Sun et al., 2007). Jedoch konnten nicht zu allen bakteriellen
Komponenten homologe Proteine im chloroplastidären cpSec-Transport gefunden
werden. Aufgrund der Assoziation des YidC-Homologs Alb3 (siehe auch Abschnitt 1.5)
und des chloroplastidär kodierten D1 mit cpSecY wird derzeit eine Funktion der cpSec-
Translokase bei der cotranslationalen Insertion von Proteinen in die Thylakoidmembran
diskutiert (Zhang et al., 2001; Klostermann et al., 2002; Pasch et al., 2005).
1.3.1.2 Der cpTat-Weg
Eine weitere Möglichkeit für die Translokation von lumenalen Proteinen bietet der cpTat-
Transportweg (Abbildung 1-1). Die Transitpeptide der Proteine, die über diesen Weg
transportiert werden, tragen ein charakteristisches Twin-arginine-Motiv, welches diesem
Transportweg seinen Namen verleiht (Tat: Twin-arginine translocation) (Chaddock et al.,
1995). Eine Besonderheit des cpTat-Wegs ist, dass bereits gefaltene Proteine über die
Thylakoidmembran translokalisiert werden können (Marques et al., 2004). Für diesen
Transport sind im Gegensatz zur cpSec-Translokase keine löslichen Komponenten
bekannt. Für den Translokationsvorgang sind die integralen Membranproteine Hcf106,
Tha4 und cpTatC essentiell (Cline und Mori, 2001; Mori und Cline, 2002; Dabney-Smith et
al., 2006). Die Untereinheiten Hcf106 und cpTatC bilden einen Rezeptorkomplex,
während das Tha4 Protein durch Oligomerisierung einen Kanal formt. Bei einer Bindung
des Substratproteins kommt es zur Assemblierung der beiden Komplexe zur vollständigen
Translokase.
Der Translokationsvorgang erfordert in vitro einen Protonengradienten (Klösgen et al.,

1 Einleitung 5
1992), während in vivo diese Abhängigkeit nicht bestätigt werden konnte (Finazzi et al.,
2003; Di Cola et al., 2005). Dieses könnte an fehlenden Komponenten in den in vitro
Studien liegen. Anstelle dessen wird eine Bedeutung des Membranpotentials diskutiert
(Theg et al., 2005; Braun et al., 2007).
1.3.2 Proteinintegration in die Thylakoidmembran
1.3.2.1 Spontane Insertion
Die meisten chloroplastidären Proteine werden anhand eines spezifischen Transportwegs
über die Thylakoidmembran translokalisiert bzw. in diese inseriert. Einige Proteine jedoch
werden offenbar ohne Beteiligung von Energiekomponenten, stromalen
Transportproteinen oder einer Translokase in die Thylakoidmembran integriert (Abbildung
1-1) (Michl et al., 1994; Kim et al., 1999; Woolhead et al., 2001). Die Integration dieser
Proteine scheint sich ohne irgendwelche Hilfskomponenten auf spontanem Weg zu
vollziehen. Über die spontane Insertion werden hauptsächlich Proteine mit einer
Transmembrandomäne (CF0II, PsbX, PsbW) in die Membran eingebaut. Bei diesen
Proteinen bedingt eine hydrophobe Region des Transitpeptids die Insertion (Thompson et
al., 1998). Allerdings kann die spontane Insertion auch von Proteinen mit zwei
Transmembrandomänen vollzogen werden (PsaK, PsaG). In diesem Fall ermöglicht eine
positiv geladene stromale loop Region die Insertion der Proteine in die Thylakoidmembran
(Zygadlo et al., 2006).
1.3.2.2 Der cpSRP-Weg
Die SRPs (signal recognition particles) sind aus dem Cytosol von Eukaryoten und
Prokaryoten bekannt, wo sie der cotranslationalen Insertion von Membranproteinen in das
endoplasmatische Retikulum und in die Plasmamembran dienen (Koch et al., 2003; Pool,
2005). Das SRP ist ein Ribonukleoprotein, welches bei Eukaryoten aus einer RNA-
Komponente und sechs Polypeptiden besteht. Die etwa 54 kDa große Untereinheit
SRP54 erkennt das Signalpeptid des zu transportierenden Proteins. Nach Bindung dieses
Komplexes an den SRP-Rezeptor wird das Protein cotranslational durch einen
Translokationskanal in das endoplasmatische Retikulum geschleust.
Der SRP-Weg in E. coli ähnelt dem eukaryotischen SRP-Weg. Der SRP-
Transportmechanismus dient dem cotranslationalen Transport von Proteinen zur
Plasmamembran (de Gier und Luirink, 2001). Das SRP in E. coli stellt mit einer RNA-

1 Einleitung 6
Komponente und dem bakteriellen Homolog des SRP54, Ffh (fiftyfour homolog), ebenfalls
ein Ribonukleoprotein dar. Bei dem FtsY handelt es sich um einen SRP-Rezeptor. Dieser
Rezeptor ist als peripheres Protein der Plasmamembran und als lösliches Protein im
Cytosol zu finden. Das cytosolische FtsY bindet den Komplex aus Ribosom, SRP und
naszierendem Protein und befördert diesen zur Plasmamembran.
Das chloroplastidäre SRP kann im Vergleich zu den anderen SRPs sowohl
cotranslational, als auch posttranslational wirken. Der posttranslationale chloroplastidäre
SRP-Weg (cpSRP-Weg) dient der Integration der Proteine der LHCP-Familie (light-
harvesting chlorophyll a/b-binding protein) in die Thylakoidmembran (Abbildung 1-1)
(Schünemann, 2007). Die LHCPs agieren als periphere Antennen der Photosysteme I und
II und stellen etwa ein Drittel der gesamten Thylakoidmembranproteine (Yamamoto und
Bassi, 1996). Bei den LHCPs handelt es sich um kernkodierte Proteine, die als
Vorstufenproteine im Cytosol synthetisiert werden. Ein N-terminales Transitpeptid sorgt
für den posttranslationalen Transport über das TOC- und TIC-Translokon der
chloroplastidären Hüllmembranen ins Stroma. Nachdem das Transitpeptid abgespalten
wurde, werden die LHCPs durch das chloroplastidäre SRP gebunden (Abbildung 1-2).
Der so geformte Transitkomplex ermöglicht den Transport der hydrophoben LHCPs durch
das Stroma zur Thylakoidmembran und hält das LHCP somit in einem integrationsfähigen
Zustand (Payan und Cline, 1991). Chloroplastidäre SRPs beinhalten keine RNA, besitzen
jedoch neben einem SRP54-Homolog (cpSRP54) noch eine weitere 43 kDa große
Komponente (cpSRP43) (Li et al., 1995; Schünemann et al., 1998; Klimyuk et al., 1999).
Bei dem cpSRP43 handelt es sich um eine neue Komponente der SRP-Untereinheiten,
die bisher nur in Chloroplasten Höherer Pflanzen identifiziert wurde. Allerdings konnte von
unserer Arbeitsgruppe gezeigt werden, dass es ein Homolog zum cpSRP43 in
Chlamydomonas reinhardtii und Physcomitrella patens gibt (Richter, Träger und
Schünemann, unveröffentlichte Daten). Der chloroplastidäre SRP-Rezeptor cpFtsY ist
sowohl im Stroma lokalisiert, als auch mit der Thylakoidmembran assoziiert (Kogata et al.,
1999; Tu et al., 1999). Analog zum bakteriellen System scheint das cpFtsY als Rezeptor
des Transitkomplexes zu arbeiten, der für den Transport zum Translokon der Membran
verantwortlich ist. Des Weiteren spielen die Nukleotide ATP und GTP eine entscheidende
Rolle in diesem Transportweg (Hoffman und Franklin, 1994; Yuan et al., 2002). Die
Funktion des ATP ist allerdings noch nicht geklärt. Bisher wurden nur GTPasen
identifiziert. Die GTP-Hydrolyse der GTPasen cpSRP54 und cpFtsY ermöglicht einen
effizienten und gerichteten Transport (Jaru-Ampornpan et al., 2007). Es wurde gezeigt,
dass für die in vitro Integration der LHCPs in isolierte Thylakoidmembranen die
Komponenten cpSRP43, cpSRP54, cpFtsY und GTP ausreichend sind (Tu et al., 1999;
Yuan et al., 2002). Die detaillierten Proteininteraktionen der einzelnen Komponenten des
posttranslationalen cpSRP-Transportwegs sind einem folgenden Kapitel zu entnehmen

1 Einleitung 7
(Kapitel 1.4). Mit dem integralen Thylakoidmembranprotein Alb3 wurde die Translokase
des posttranslationalen cpSRP-Wegs entdeckt, die für die Insertion der LHCPs in die
Thylakoidmembran notwendig ist (Sundberg et al., 1997; Moore et al., 2000). Das Alb3
gehört zu der sogenannten Oxa1/YidC/Alb3- Proteinfamilie, dessen Mitglieder in der
Insertion und Assemblierung von Membranproteinen in Mitochondrien, Bakterien und
Chloroplasten beteiligt sind (Yi und Dalbey, 2005; Xie und Dalbey, 2008). Eine
ausführliche Darstellung dieser wichtigen Membranproteine erfolgt in einem gesonderten
Kapitel zur Oxa1/YidC/Alb3- Proteinfamilie (Kapitel 1.5).
4354
cpSRP
Transit-komplex
+ GTP
LHCP
Alb3
TM
4354psbA
Ribosom
D1
cpSec-Translokase
54
54
cpFtsY
cotranslational posttranslational
Abbildung 1-2: Modell des co- und posttranslationalen cpSRP-Transportwegs Das Modell stellt die Beteiligung des cpSRP54 am cotranslationalen SRP-Weg zur Integration von
plastidär kodiertem D1 und am posttranslationalen SRP-Weg zur Integration von kernkodiertem
LHCP dar. Das cpSec-assoziierte Alb3 scheint an der SRP-abhängigen cotranslationalen Insertion
von plastidär kodierten Thylakoidmembranproteinen beteiligt zu sein, während die
posttranslationale Integration von LHCP unabhängig von der cpSec-Translokase verläuft.
(verändert nach Schünemann, 2007)
Das cpSRP54 ist neben dem posttranslationalen Transport im Komplex mit cpSRP43
auch am cotranslationalen Transport chloroplastidär kodiertem D1-Protein involviert
(Abbildung 1-2), wobei cpSRP54 an das naszierende D1 bindet (Nilsson et al., 1999;

1 Einleitung 8
Nilsson und van Wijk, 2002). Eine Interaktion des cpSRP43 mit dem naszierenden D1
konnte bislang nicht gezeigt werden. Dieses weist auf eine spezialisierte Rolle des
cpSRP43 im posttranslationalen Transport und eine Doppelfunktion des cpSRP54 sowohl
im posttranslationalen als auch im cotranslationalen Transport hin. Des Weiteren konnte
eine Assoziation von cpSecY mit Ribosomen und dem D1-Protein gezeigt werden (Zhang
et al., 2001). Die cpSec-Translokase ist somit wahrscheinlich an der cotranslationalen
Insertion des chloroplastidär kodierten D1 beteiligt. Das integrale Membranprotein Alb3
scheint ebenfalls eine Doppelfunktion innerhalb des cpSRP-Wegs zu haben. Neben der
Funktion in der posttranslationalen Insertion der LHCPs in die Thylakoidmembran, scheint
die Assoziation mit der cpSec-Translokase auf eine Rolle von Alb3 innerhalb der
cotranslationalen Insertion von Proteinen zu deuten (Klostermann et al., 2002; Pasch et
al., 2005).
1.4 Proteininteraktionen der Komponenten des posttranslationalen cpSRP-Transportwegs
Im folgenden Abschnitt sollen die Proteininteraktionen innerhalb des cpSRP-Wegs näher
beschrieben werden (Abbildung 1-3). Das cpSRP43 ist aus zwei charakteristischen
Protein-Protein-Interaktionsdomänen aufgebaut, den Ankyrin repeats und den
Chromodomänen. Es besitzt im N-Terminus eine Chromodomäne und vier Ankyrin
repeats sowie im C-terminalen Bereich zwei benachbarte Chromodomänen (Klimyuk et
al., 1999; Jonas-Straube et al., 2001; Goforth et al., 2004). Ankyrin repeats ermöglichen
als charakteristische Bindedomänen verschiedene Proteininteraktionen und sind dadurch
an der Ausbildung von Proteinkomplexen beteiligt (Sedgwick und Smerdon, 1999). Die
Chromodomänen wurden anfänglich in Proteinen des Zellkerns identifiziert, wo sie einen
Einfluss auf die Regulation der Chromatinstruktur zeigen (Brehm et al., 2004).
Chromodomänen sind charakteristisch aus drei antiparallelen β-Faltblättern und einer C-
terminalen α-Helix aufgebaut (Sivaraja et al., 2005). Die Kristallstruktur des cpSRP43
(Chromodomäne1 bis Ankyrin repeat 4) zeigt, dass die Ankyrin repeats 2 und 3 ein
typisches helix-turn-helix Motiv besitzen, während die Ankyrin repeats 1 und 4 verlängerte
Helices aufweisen (Stengel et al., 2008). Die Chromodomäne 1 und Ankyrin repeat 1
gehen durch Fusion ihrer Helices direkt ineinander über. Auch Ankyrin repeat 4 zeigt
durch die verlängerte Helix einen direkten Übergang zur Chromodomäne 2. Das cpSRP54
besteht aus einer N-terminalen G-Domäne, welche für die GTPase Funktion
verantwortlich ist, und einer C-terminalen methioninreichen M-Domäne (Franklin und
Hoffman, 1993). Bei der Heterodimerisierung des cpSRP-Komplexes kommt es zur
Interaktion der Chromodomäne 2 des cpSRP43 mit der M-Domäne des cpSRP54
(Goforth et al., 2004; Hermkes et al., 2006). Es konnte gezeigt werden, dass die

1 Einleitung 9
Chromodomäne 2 des cpSRP43 mit einem 10 Aminosäuren langen Bindemotiv innerhalb
der M-Domäne des cpSRP54 interagiert (Goforth et al., 2004; Funke et al., 2005;
Hermkes et al., 2006; Kathir et al., 2008). Aufgrund der positiven Ladung des Bindemotivs
im cpSRP54 (Funke et al., 2005) und der negativen Oberflächenladung der
Chromodomäne 2 des cpSRP43 (Sivaraja et al., 2005) könnte es sich um eine
elektrostatische Interaktion handeln.
LHCPTMD L-18 Motiv
cpSRP43 A1 A2 A3 A4 CD3
cpSRP54 G-Domäne M-Domäne
Bindemotiv
cpFtsY G-DomäneN-Domäne
CD2CD2CD1
Abbildung 1-3: Proteininteraktionen der Komponenten des cpSRP-Transportwegs Das cpSRP43 besitzt drei Chromodomänen (CD1-3) und vier Ankyrin repeats (A1-4). Das
cpSRP54 besteht aus einer G-Domäne, welche für die GTPase Aktivität zuständig ist, und einer
methioninreichen M-Domäne. Bei der Heterodimerisierung des cpSRP kommt es zur Interaktion
der Chromodomäne 2 mit einem 10 Aminosäuren langen Bindemotiv innerhalb der M-Domäne des
cpSRP54. Das LHCP besitzt drei Transmembrandomänen (TMD). Es interagiert über das L-18
Motiv mit der Ankyrin repeat Region des cpSRP43. Weiterhin interagiert das cpSRP54 mit den
hydrophoben Transmembrandomänen des LHCPs und dem cpSRP-Rezeptor FtsY.
Die LHCPs stellen das Substrat des cpSRP-Transportwegs dar. Diese integralen
Membranproteine besitzen 3 Transmembrandomänen. Bei der Bildung des
Transitkomplexes kommt es zunächst zur Interaktion der Ankyrin repeat Domäne des

1 Einleitung 10
cpSRP43 mit dem sogenannten L18-Motiv des LHCPs, welches sich zwischen der 2. und
3. Transmembrandomäne befindet (DeLille et al., 2000; Tu et al., 2000; Jonas-Straube et
al., 2001; Stengel et al., 2008). Die Bindung zwischen cpSRP54 und LHCP erfolgt über
die hydrophoben Transmembrandomänen des LHCPs (DeLille et al., 2000; Tu et al.,
2000; Groves et al., 2001). Das cpFtsY ist aus einer N-terminalen N-Domäne und einer C-
terminalen G-Domäne, welche für die GTPase Aktivität verantwortlich ist, aufgebaut
(Stengel et al., 2007; Chandrasekar et al., 2008). Die Proteine cpSRP54 und cpFtsY sind
in der Lage in GTP-gebundener Konformation miteinander zu interagieren. Die
Dissoziation beider Proteine erfolgt über GTP-Hydrolyse (Jaru-Ampornpan et al., 2007).
Das Protein Alb3 stellt die Translokase des cpSRP-Wegs dar. Dieses integrale
Membranprotein besitzt 5 Transmembrandomänen. Es konnte gezeigt werden, dass
cpSRP54 und cpFtsY zusammen mit Alb3 einen Komplex an der Thylakoidmembran
bilden können (Moore et al., 2003). Die Proteininteraktionen der löslichen cpSRP-
Komponenten mit Alb3 sind jedoch noch nicht vollständig geklärt und daher ein zentraler
Teil dieser Arbeit.
1.5 Die Oxa1/YidC/Alb3-Proteinfamilie
Die Biogenese von membrangebundenen Proteinkomplexen wie z.B. der Photosysteme,
Atmungskette oder ATP-Synthase ist von entscheidender Bedeutung für die Funktionalität
von Zellen. Für diese Funktionalität der Proteinkomplexe ist nicht nur der Transport der
Proteine an ihren Wirkort, sondern auch ein korrekter Einbau in die Membranen essentiell.
Die Mitglieder der sogenannten Oxa1/YidC/Alb3-Proteinfamilie sind evolutionär
konserviert und ermöglichen die Insertion und Assemblierung von Membranproteinen in
Mitochondrien, Bakterien und Chloroplasten (Kuhn et al., 2003; Kiefer und Kuhn, 2007;
Xie und Dalbey, 2008; Zhang et al., 2009). Diese Proteinfamilie weist im Bereich ihrer
Transmembrandomänen eine hydrophobe Kernregion auf, während die hydrophilen
Bereiche in ihrer Länge und Funktion variieren (Yen et al., 2001). Die konservierten
Bereiche der Proteine ermöglichen die Komplementation der Proteine untereinander in
den verschiedenen Systemen, während die hydrophilen Anteile auf eine spezielle
Funktion der Proteine deuten (Preuss et al., 2005; van Bloois et al., 2005). Das bakterielle
Homolog YidC besitzt 6 Transmembrandomänen, während die Homologen in den
Organellen, Oxa1 und Alb3, jeweils 5 Transmembrandomänen aufweisen (Abbildung 1-4).
Die N- und C-terminalen Bereiche des bakteriellen YidC ragen, aufgrund der
Membrantopologie, beide ins Cytoplasma. Das YidC besitzt zwischen der ersten und
zweiten Transmembrandomäne eine lange periplasmatische Domäne. Erste Arbeiten

1 Einleitung 11
geben näheren Aufschluss auf die Struktur des YidC (Ravaud et al., 2008). Bei dem
mitochondrialen Oxa1 und dem chloroplastidären Alb3 ergibt die Membrantopologie eine
Lokalisation der N- und C-terminalen Enden auf gegenüberliegenden Seiten. Der
N-Terminus von Oxa1 zeigt in den Intermembranraum und der C-Terminus in die Matrix
der Mitochondrien. Das chloroplastidäre Alb3 ragt mit dem N-Terminus in das
Thylakoidlumen und mit dem C-terminalen Ende in das Stroma (Kuhn et al., 2003). In den
folgenden Abschnitten sollen zunächst die charakteristischen Funktionen der
namensgebenden Mitglieder der Oxa1/YidC/Alb3-Proteinfamilie näher beschrieben
werden. Zusätzlich erfolgt eine Beschreibung weiterer Homologe dieser Proteinfamilie.
N
C
N
N CC
YidC Oxa1Alb3
Intermembranraum
MatrixCytoplasma
Thylakoidlumen
Stroma
Periplasma
Abbildung 1-4: Membrantopologie von Alb3, YidC und Oxa1 Das chloroplastidäre Alb3 und das mitochondriale Oxa1 besitzen fünf Transmembrandomänen,
während das bakterielle Homolog YidC aus sechs Transmembrandomänen besteht (Kuhn et al.,
2003). Die N- und C-terminalen Bereichen des Alb3 und Oxa1 befinden sich auf
gegenüberliegenden Seiten der Membran. Der N-Terminus von Alb3 zeigt ins Thylakoidlumen und
der C-Terminus ins Stroma der Chloroplasten. Der N-Terminus des mitochondrialen Oxa1 weist in
den Intermembranraum und der C-Terminus in die Matrix der Mitochondrien. Das bakterielle YidC
zeigt eine Membrantopologie, bei der beide Termini ins Cytoplasma ragen. Das YidC besitzt
zwischen der ersten und zweiten Transmembrandomäne eine lange periplasmatische Domäne.
Oxa1 und Alb3 haben jeweils einen verlängerten hydrophilen C-Terminus.
1.5.1 Oxa1
Oxa1 ist ein 36 kDa großes Protein der inneren Mitochondrienmembran von
Saccharomyces cerevisiae, das die Insertion von kern- und mitochondrialkodierten,
inneren Membranproteinen vermittelt (Hell et al., 1998; Hell et al., 2001) (Abbildung 1-5).
Es wurde durch die Analyse von Proteinen, die an der Assemblierung des Cytochrom-

1 Einleitung 12
Oxidase-Komplexes beteiligt sind, identifiziert (Oxa1 = oxidase assembly 1) (Bauer et al.,
1994; Bonnefoy et al., 1994). Oxa1 ist sowohl an der posttranslationalen, als auch an der
cotranslationalen Proteininsertion beteiligt (Bonnefoy et al., 2009). Im mitochondrialen
Transport sind bislang keine Homologen zu den Untereinheiten der bakteriellen Sec-
Translokase bekannt. Daher scheint das Oxa1 ausschließlich in einem Sec-unabhängigen
Integrationsmechanismus zu agieren (Glick und Von Heijne, 1996). Für die Oxa1-
abhängige Insertion von kernkodierten, mitochondrialen Proteinen werden die Proteine
zunächst vom Cytosol in die mitochondriale Matrix transportiert und anschließend in die
innere Membran integriert. Durch Interaktion der C-terminalen, α-helicalen Domäne des
Oxa1 mit den translatierenden Ribosomen konnte eine unterstützende Wirkung auf die
cotranslationale Proteininsertion nachgewiesen werden (Jia et al., 2003; Szyrach et al.,
2003). Diese C-terminale Domäne scheint jedoch keinen Einfluss auf die
posttranslationale Proteininsertion zu haben. Es konnte gezeigt werden, dass das
bakterielle Homolog YidC mit Hilfe der C-terminalen Ribosomen-Bindestelle des Oxa1 die
Funktion des Oxa1 komplementieren kann (Preuss et al., 2005; van Bloois et al., 2005).
Weiterhin wurde gezeigt, dass Oxa1 wie auch das bakterielle Homolog YidC mit
translatierenden Ribosomen eine Pore bildet (Kohler et al., 2009). Oxa1 Hefemutanten
haben gezeigt, dass von der Deletion des Oxa1 nicht nur die Assemblierung der
Cytochrom-Oxidase, sondern auch die ATP-Synthase betroffen ist (Altamura et al., 1996;
Jia et al., 2007).
In Mitochondrien von Saccharomyces cerevisiae wurde neben dem Oxa1 bereits ein
Homolog identifiziert, welches als Oxa2/Cox18 bezeichnet wurde. Dieses Protein spielt
eine spezifische Rolle in der Biogenese des Cytochrom-Oxidase-Komplexes (Preuss et
al., 2005; Bonnefoy et al., 2009). Komplementationsversuche haben gezeigt, dass YidC
die beiden mitochondrialen Homologe Oxa1 und Oxa2/Cox18 funktionell ersetzen kann
(Preuss et al., 2005). Das Oxa2/Cox18 kann ebenfalls die Sec-unabhängige Funktion von
YidC in E. coli komplementieren (van Bloois et al., 2007). In der Hefe sind Oxa1 und
Oxa2/Cox18 nicht in der Lage sich gegenseitig zu komplementieren, was auf eine
unterschiedliche Funktion der beiden Proteine schließen lässt. Das Oxa2/Cox18 scheint
im Gegensatz zum Oxa1 ausschließlich eine posttranslationale Aufgabe zu haben. Die
posttranslationale Funktion des Oxa2/Cox18 wird durch die unterstützende Wirkung auf
die Translokation der C-terminalen Domäne des Cox2 in den Intermembranraum und eine
mögliche Funktion in der Assemblierung mit den anderen Untereinheiten des Cytochrom-
Oxidase-Komplexes gezeigt (Saracco und Fox, 2002). Das Oxa2 in Neurospora crassa ist
in der Lage die Oxa2-Mutante der Hefe zu komplementieren, was für eine gemeinsame
Funktion der beiden Proteine spricht. Die Deletion dieses Oxa2-Gens verursacht einen
spezifischen Defekt in der Biogenese des Cytochrom-Oxidase-Komplexes (Funes et al.,
2004).

1 Einleitung 13
1.5.2 Yid C
YidC ist ein 60 kDa großes, integrales Membranprotein der Plasmamembran in E. coli und
wurde als neue Komponente der Sec-Translokase nachgewiesen (Saaf et al., 1998; Scotti
et al., 2000). In E. coli werden die meisten Membranproteine über den cotranslationalen
SRP-Weg zur Plasmamembran transportiert und über die Sec-Translokase in die innere
Membran inseriert. Das integrale Membranprotein der Plasmamembran YidC stellt eine
entscheidende Komponente der Sec-Translokase dar und ermöglicht den lateralen
Einbau von Transmembrandomänen integrierter Proteine aus der Sec-Translokase in die
Membran (Beck et al., 2001; Urbanus et al., 2001; van der Laan et al., 2001; Houben et
al., 2004). YidC interagiert dabei mit den Transmembrandomänen der inserierten Proteine
direkt nach deren Freisetzung aus dem Sec-Translokon. Dieses verhindert die
Aggregation der integrierten Proteine. Das YidC scheint somit eine Funktion als
Membranchaperon auszuüben. Des Weiteren wurde gezeigt, dass YidC in E. coli für die
Insertion von Membranproteinen, die unabhängig von der Sec-Translokase integriert
werden, essentiell ist (Samuelson et al., 2000; Chen et al., 2002; Serek et al., 2004; van
der Laan et al., 2004; van Bloois et al., 2005; Facey et al., 2007). Dabei handelt es sich
z. B. um Phagenproteine oder die F0c-Untereinheit der ATP-Synthase. Von den
Phagenproteinen wurde zunächst angenommen, dass sie spontan in die Membran
inserieren. Das YidC ist in der Lage, wie das mitochondriale Homolog Oxa1, mit
translatierenden Ribosomen eine dimere Insertionspore auszubilden (Kohler et al., 2009).
Die bisherigen Daten zeigen, dass YidC sowohl in Kombination mit der Sec-Translokase
als auch unabhängig von dieser bei der Integration von Membranproteinen agieren kann
(Abbildung 1-5).
In grampositiven Bakterien findet man hauptsächlich zwei YidC Homologe (Funes et al.,
2009). Das YidC1 und YidC2 aus Streptococcus mutans sind in der Lage das YidC aus
E.coli in der Sec-abhängigen und Sec-unabhängigen Proteininsertion zu komplemetieren.
YidC2 besitzt, wie das mitochondriale Homolog Oxa1, einen positiv geladenen C-
Terminus und kann mit Ribosomen assoziieren. Es ist in der Lage die Funktion von Oxa1
in der cotranslationalen Proteininsertion von mitochondrialen, inneren Membranproteinen
zu übernehmen. Weiterhin wurde gezeigt, dass YidC2 unabhängig von dem SRP-Weg
cotranslational wirken kann.

1 Einleitung 14
1.5.3 Alb3
Das Alb3 Protein wurde aus der Analyse von Pigment-defizienten Mutanten von
Arabidopsis thaliana identifiziert. Die Mutanten zeigen einen drastischen Albino-Phänotyp
(Albino 3 = Alb3) und sind nicht in der Lage ohne externe Kohlenstoffquelle über das
Keimungsstadium hinaus zu überleben (Sundberg et al., 1997). Der Chlorophyllgehalt
beträgt nur etwa 5 % des Wildtyplevels. Die Chloroplasten zeigen nur wenige
Thylakoidmembranen und keinerlei Granastapelung. Alb3 ist ein 45 kDa großes,
integrales Thylakoidmembranprotein, welches an der posttranslationalen Integration der
LHCPs in die Thylakoidmembran beteiligt ist (Moore et al., 2000; Moore et al., 2003)
(Abbildung 1-5). Es konnte gezeigt werden, dass eine Vorinkubation von
Thylakoidmembranen mit einem Alb3-Antikörper die Integration des Lhcb1 beeinträchtigt,
während dieses keine Auswirkungen auf den Transport von cpSec- und cpTat-
abhängigen Proteinen hatte (Moore et al., 2000). Gleichermaßen wurde die Integration
von zwei weiteren Mitgliedern der LHCP-Familie (Lhcb4.1 und Lhcb5) in die
Thylakoidmembran verhindert (Woolhead et al., 2001). Das Alb3 scheint bei der
Integration von LHCP unabhängig von der cpSec-Translokase zu agieren, da die
Vorinkubation der Thylakoidmembran mit einem Antikörper gegen den C-Terminus von
cpSecY keinen Einfluss auf die Integration von LHCP hat (Mori et al., 1999). Die
posttranslationale Integration der LHCPs ist daher ein cpSRP/Alb3-abhängiger, jedoch
cpSec-unabhängiger Mechanismus (Moore et al., 2003). Andere Arbeiten zeigen
allerdings eine Assoziation des Alb3 mit der cpSec-Translokase (Klostermann et al., 2002;
Pasch et al., 2005). Das cpSec-assoziierte Alb3 scheint an der cpSRP-abhängigen
cotranslationalen Insertion von chloroplastidär kodierten Thylakoidmembranproteinen
beteiligt zu sein (Abbildung 1-5). Zudem konnte gezeigt werden, dass Alb3 in der Lage ist
das YidC in einem YidC-defizienten Bakterienstamm zu komplementieren. Das Alb3
ermöglicht in E.coli die Membraninsertion von Sec-abhängigen und Sec-unabhängigen
Membranproteinen (Jiang et al., 2002).
Neben dem Alb3 konnte mit dem Alb4 ein zweites chloroplastidäres Homolog der
Oxa1/YidC/Alb3-Proteinfamilie in Arabidopsis thaliana identifiziert werden (Gerdes et al.,
2006). Zahlreiche Analysen wie z. B. Importstudien, immunologische Detektion in
chloroplastidären Subfraktionen und GFP-Lokalisationsstudien ergaben, dass Alb4 in der
Thylakoidmembran der Chloroplasten lokalisiert ist. Arabidopsis thaliana Mutanten mit
einem um 90 % reduzierten Alb4 Level zeigten keinen offensichtlichen Phänotyp. Der im
Vergleich zu den alb4-Mutanten drastische Phänotyp der alb3-Mutante zeigt, dass Alb4
nicht in der Lage ist die Funktion von Alb3 zu kompensieren. Die chloroplastidäre
Ultrastruktur der alb4-Mutanten weist vergrößerte Plastiden mit einer im Vergleich zum
Wildtyp aufgelockerten Anordnung der Granathylakoide auf. Das Alb4 scheint somit einen

1 Einleitung 15
Einfluss auf die Biogenese der Chloroplasten zu haben. Die genaue Funktion des Alb4 ist
jedoch noch nicht bekannt.
In Chlamydomonas reinhardtii wurden ebenfalls zwei Alb3-Homologe (Alb3.1 und Alb3.2)
identifiziert (Bellafiore et al., 2002; Göhre et al., 2006). Die Inaktivierung des Alb3.1 in der
ac29-Mutante von Chlamydomonas reinhardtii hat zwei bedeutende Effekte. Die Menge
des LHCI und LHCII im Photosystem I und II ist um mehr als das 10-fache reduziert und
der Chlorophyllgehalt entspricht nur 30% des Wildtyplevels. Zudem ist das Photosystem II
um das 2-fache bzw. 10-fache in den Zellen, die in Licht bzw. bei Dunkelheit angezogen
wurden, vermindert (Bellafiore et al., 2002). Es konnte weiterhin gezeigt werden, dass
Alb3.1 an der Assemblierung des D1 in das Reaktionszentrum des Photosystems II von
Chlamydomonas reinhardtii involviert ist (Ossenbühl et al., 2004). Interessanterweise
scheint dagegen Alb3.1 für die Integration des D1 in die Thylakoidmembran keine
signifikante Rolle zu spielen (Ossenbühl et al., 2004). Das Alb3.1 hat daher eine Funktion
in der Integration der LHCPs und in der Assemblierung des Photosystem II. Weiterhin ist
bemerkenswert, dass die Inaktivierung des Alb3.1 in Chlamydomonas reinhardtii einen
deutlich milderen Phänotyp zeigt, als der Verlust des Alb3 in Arabidopsis thaliana
(Sundberg et al., 1997; Bellafiore et al., 2002). Mit dem Alb3.2 konnte ein weiteres Alb3-
Homolog in Chlamydomonas reinhardtii identifiziert werden, das den Verlust des Alb3.1 in
der Mutante ac29 möglicherweise teilweise kompensieren kann (Bellafiore et al., 2002).
Eine Reduktion des Alb3.2 in der Chlamydomonas RNAi-Mutante führt zu einer starken
Reduktion der Phtosysteme I und II und langfristig zum Zelltod. Während die Menge an
Lichtsammelkomplexen nicht so stark reduziert ist, wie in der alb3.1-Mutante, zeigt die
alb3.2-Mutante mit einer Vergrößerung der Vakuole einen weiteren Phänotyp, dessen
Ursache allerdings nicht geklärt ist (Göhre et al., 2006).
Im Cyanobakterium Synechocystis sp. PCC6803 konnte ebenfalls ein Alb3-Homolog
(Slr1471) identifiziert werden. Mutanten zeigten, dass dieses Homolog für die Morphologie
der Thylakoide und die Lebensfähigkeit der Zellen wichtig ist (Spence et al., 2004). Es
konnte gezeigt werden, dass das Slr1471 mit dem D1 Protein interagieren kann und
daher für die Integration des D1 wichtig ist (Ossenbühl et al., 2006). Das Slr1471 ist
hauptsächlich in der Cytoplasmamembran lokalisiert und in der Lage das YidC in einem
YidC-defizienten E.coli Bakterienstamm zu ersetzen (Gathmann et al., 2008). Mit der
Lokalisation des Slr1471 in der Cytoplasmamembran ist eine Funktion in der Biogenese
des Photosystem II in der Cytoplasmamembran verbunden.

1 Einleitung 16
Oxa1
Mitochondrien
Matrix
Sec
YidC
E. coli
Cytoplasma
YidC
cpSec
Alb3Alb3
Chloroplast
Thylakoidlumen
Stroma
TM innere Membran
äußere Membran
innere Membran
äußere Membran
Cytoplasma
Abbildung 1-5: Proteininsertionen durch Alb3, YidC und Oxa1 Das integrale Thylakoidmembranprotein Alb3 ermöglicht in Chloroplasten die posttranslationale
Insertion der LHCPs. Weiterhin ist das Alb3 mit der cpSec-Translokase assoziiert und agiert dort
vermutlich in der cotranslationalen Insertion von chloroplastidär kodierten Thylakoidmembran-
proteinen. Das YidC in E.coli ist sowohl in der Sec-abhängigen, als auch Sec-unabhängigen
Proteininsertion beteiligt. In Mitochondrien vermittelt Oxa1 die posttranslationale und
cotranslationale Insertion von inneren Membranproteinen.

2 Zielsetzung 17
2. Zielsetzung
In der vorliegenden Arbeit sollte die Rolle von Alb3 und Alb4 bei der Biogenese der
Thylakoidmembran von Arabidopsis thaliana genauer geklärt werden. Alb3 und Alb4 sind
chloroplastidäre Homologe der Oxa1/YidC/Alb3-Proteinfamilie, deren Mitglieder eine
entscheidende Bedeutung bei der Insertion und Assemblierung von Membranproteinen
haben.
In vorangegangenen Arbeiten konnte bereits gezeigt werden, dass Alb3 für die Insertion
von Mitgliedern der LHCPs, die cpSRP-abhängig zur Thylakoidmembran transportiert
werden, essentiell ist. In einem ersten Teilprojekt dieser Arbeit sollte der docking-Prozess
des Komplexes aus LHCP und cpSRP in Kombination mit dem SRP-Rezeptor cpFtsY an
die Thylakoidmembran näher analysiert werden. Als zentrale Frage sollte dabei
untersucht werden, ob dieser Prozess auf einer direkten Interaktion der Proteine des
cpSRP-Wegs mit der Translokase Alb3 beruht.
Da zu Beginn dieser Arbeit die molekulare Funktion des integralen
Thylakoidmembranproteins Alb4 bei der Biogenese der Chloroplasten unbekannt war,
sollte in einem weiteren Teilprojekt die Interaktion von Alb4 mit anderen
Thylakoidmembranproteinen analysiert werden.
Um die homologen Proteine in Arabidopsis thaliana in biochemischen Untersuchungen
getrennt voneinander detektieren zu können, sollten zunächst spezifische Antikörper
gegen das Alb3 und das Alb4 hergestellt werden. Für die Analyse von Protein-Protein-
Interaktionen sollten sowohl in vivo Studien unter Verwendung des Hefe Split-Ubiquitin
Systems und der bimolekularen Fluoreszenzkomplementation in Arabidopsis Protoplasten
als auch in vitro Interaktionsstudien (z. B. pull down Analysen, chemisches Crosslinking,
Coimmunpräzipitation) durchgeführt werden.

3 Material und Methoden 18
3. Material und Methoden
3.1 Chemikalien
Tabelle 3-1: Chemikalienliste
Chemikalie Hersteller [35S]-Methionin (37 TBq/mmol, 370 MBq/ml) Hartmann Adenin Hemisulfat-Salz SIGMA Affi Gel 15 Biorad Coomassie® Brilliant Blau G-250 SERVA CSM –His –Leu –Trp (Complete Supplement Mixture minus the above) BIO 101 CSM –Leu –Trp (Complete Supplement Mixture minus the above) BIO 101 Cellulase SERVA DOB (ohne AS) BIO 101 Gene RulerTM 1Kb DNA Ladder Fermentas Glutathion Sepharose 4B Amersham Macerozyme SERVA Molekulargewichtsmarker SIGMA Ni-NTA-Agarose Qiagen Page RulerTM Prestained Protein Ladder Fermentas PEG 3350 SIGMA Pepton MERCK
Ponceau S SERVA Protein A Sepharose CL 4B Amersham Proteinaseinhibitor, complete Mini Protease Inhibitor Cocktail Tablets Roche Sulfo-MBS PIERCE Super Signal West Femto Luminol/Enhancer Solution PIERCE Super Signal West Femto Stable Peroxide Buffer PIERCE Super Signal West Pico Luminol/Enhancer Solution PIERCE Super Signal West Pico Stable Peroxide Buffer PIERCE Weizenkeimlysat (Wheat Germ) Promega
Nicht aufgeführte Chemikalien wurden in Laborqualität von der Firma AppliChem
bezogen.

3 Material und Methoden 19
3.2 Verbrauchsmaterial
Tabelle 3-2: Verbrauchsmaterial
Material Hersteller Blotting Paper Schleicher&Schuell Dialyse-Kassetten PIERCE Filterpapier Schleicher&Schuell Hyperfilm ECL High performance chemiluminescence film Amersham Impfösen Greiner Kodak Biomax MR Film Scientific Imaging Film Kodak Kodak Biomax MS Scientific Imaging Film Kodak Kryoröhrchen Brand Membranfilter Millipore Miracloth Calbiochem PCR-Thermo-Strips 0,2 ml PeqLab PCR-Tubes 0,2 ml Brand Petrischalen Greiner Pipettenspitzen Greiner Polyprep Chromatographie Säulen Biorad Porablot NCL Nitrocellulosemembran Macherey-Nagel PS Küvetten halbmikro Brand Reaktionsgefäße 15 ml Greiner Reaktionsgefäße 50 ml Greiner Safelock Tubes 1,5 ml Eppendorf Safelock Tubes 2 ml Eppendorf UV-Küvetten mikro Brand Wizard Mini Columns Promega
Tabelle 3-3: Kits
Kit Hersteller BCATM Protein Assay Kit PIERCE In-FusionTM Dry-Down PCR Cloning Kit Clontech NucleoSpin Extract II Kit Macherey-Nagel NucleoSpin Plasmid Mini Prep Kit Macherey-Nagel Plasmid Maxi Prep Kit Promega QuikChange XL site-directed mutagensis kit Stratagene RTS 100 Wheat Germ CECF Kit Roche

3 Material und Methoden 20
3.3 Geräte
Tabelle 3-4: Geräte
Geräte Hersteller Aekta Explorer 10 GE Healthcare Biowave S2100 Photometer Biochrom Brutschrank Memmert Electrophoresis Power Supply E143 Consort Electrophoresis Power Supply E233 Consort Electrophoresis Power Supply E835 Consort Elektronische Präzisionswaage Kern & Sohn GmbH Gel Dokumentation Herolab Gel Dryer Modell 583 Biorad Gelfiltrationssäule Superose 6 10/300 GL GE Healthcare Heizblock Bio TDB-100 LAB4you Hoefer Mighty Small II (SDS-PAGE) Amersham Hoefer TE 22 tank transfer unit Amersham Hotplate & Stirrer SB162 Bibby Sterilin LSM 510 Meta Zeiss Minishaker MINI-SECOUEUR IKA MS 2 MINISHAKER Minitron HT (Thermoschüttler) Infors PerfectBlueTMHorizontal Mini Electrophoresis System PeqLab pH-Meter MP220 Mettler Toledo Power Pac 200 (Spannungsgeber) Biorad Schüttelwasserbad 1083 GFL Shaker S-4 ELMI Sonifier Cell Disruptor B15 Branson Thermocycler Mastercycler gradient Eppendorf Thermomixer Comfort Eppendorf Mixer Waring Blender Waring Zentrifuge 5415 D Eppendorf Zentrifuge 5417 R Eppendorf Zentrifuge 5412 D Eppendorf Zentrifuge 5424 Eppendorf Zentrifuge 5804 R Eppendorf Zentrifuge AvantiTM J-25 Beckman

3 Material und Methoden 21
3.4 Biologisches Material
3.4.1 Enzyme
KOD HiFi DNA-Polymerase Novagen
Pfu-Turbo-DNA-Polymerase Stratagene
Restriktionsendonukleasen Fermentas
RNase A Fermentas
SP6 Polymerase Fermentas
T4-DNA-Ligase Fermentas
3.4.2 Antikörper
Anti-Kaninchen IgG: Peroxidase-gekoppelter Antikörper zur Detektion von Kaninchen-
Antikörpern (Verdünnung 1:10000; Sigma)
Anti-Ratte IgG: Peroxidase-gekoppelter Antikörper zur Detektion von Ratten-
Antikörpern (Verdünnung 1:10000; Sigma)
Anti-Huhn IgG: Peroxidase-gekoppelter Antikörper zur Detektion von Huhn-
Antikörpern (Verdünnung 1:10000; Sigma)
Anti-Alb3: polyklonales Antiserum aus Kaninchen (Verdünnung 1:500;
SeqLab)
Anti-Alb4: polyklonales Antiserum aus Kaninchen (Verdünnung 1:500;
aufgereinigt 1:100; SeqLab; Gerdes et al., 2006)
Anti-Alb4: polyklonales Antiserum aus Ratte (Verdünnung 1:500; SeqLab)
Anti-chaos: polyklonales Antiserum aus Huhn (Verdünnung 1:5000;
Schünemann et al. 1998)
Anti-cpSecY: polyklonales Antiserum aus Kaninchen (Verdünnung 1:5000;
Schünemann et al. 1999)
Anti-c-myc: Antikörper zur Detektion eines c-myc-tags (Clontech)
Anti-HA: Antikörper zur Detektion eines HA-tags (Clontech)
Anti-CF1β: Antikörper zur Detektion von CF1β (Agrisera)
Anti-CF0IV: Antikörper zur Detektion von CF0IV (Agrisera)
Anti-CF1α+β, Anti-CF1γ, Anti-CF1δ, Anti-CF1ε, Anti-CF0I, Anti-CF0II, Anti-CF0III
(freundlicherweise zur Verfügung gestellt von PD Dr. Jörg Meurer, Universität München)

3 Material und Methoden 22
3.4.3 Verwendete Plasmide
Im folgenden sind die verwendeten Plasmide aufgelistet und beschrieben.
Tabelle 3-5: Verwendete Plasmide
Plasmid Beschreibung Referenz
pGEX4T3 Expressionsvektor, kodiert für N-terminalen GST-tag, T7-lac-Promotor Pharmacia
Biotech
pET-DUET1 Expressionsvektor, kodiert für N-terminalen Hexahistidin-tag, T7-lac-Promotor Novagen
pET-29b Expressionsvektor, kodiert für C-terminalen Hexahistidin-tag, T7-lac-Promotor Novagen
pSS Vektor zur in vitro Transkription/ Translation, SP6-Promotor Franklin und
Hoffmann, 1993
pIVEX1.3WG in vitro Expressionsvektor für das RTS 100 Wheat Germ System, T7-lac-Promotor Roche
pAMBV4 Vektor für Hefetransformation, kodiert für C-terminales Ubiquitin-Fragment Dualsystems
Biotech AG
pADSL-Nx Vektor für Hefetransformation, kodiert für N-terminales Ubiquitin-Fragment Dualsystems
Biotech AG
pSPYCE Vektor für das Split-YFP System, kodiert für das C-terminale cYFP-Fragment Walter et al.,
2004
pSPYNE Vektor für das Split-YFP System, kodiert für das C-terminale nYFP-Fragment Walter et al.,
2004
pSPYCE-TS(rbcs)
Vektor für das Split-YFP System, kodiert für die Rubisco Transitsequenz und das C-terminale cYFP-Fragment
verändert nach Walter et al., 2004
pSPYNE-TS(rbcs)
Vektor für das Split-YFP System, kodiert für die Rubisco Transitsequenz und das C-terminale nYFP-Fragment
verändert nach Walter et al., 2004

3 Material und Methoden 23
3.4.4 Erstellte Plasmide und verwendete Oligonukleotide
Die für die Amplifikation von DNA-Fragmenten eingesetzten Oligonukleotide wurden von
der Firma Sigma-Genosys bezogen.
Die folgenden Plasmide kodieren für die Antigene von Alb3 und Alb4. Diese wurden
genutzt, um spezifische Antikörper herzustellen.
pGEX4T3-cAlb3: for BamHI CGTGGATCCAAGCGCAGCAAGAAGAAC rev XhoI CTGCTCGAGTTATACAGTGCGTTTCCGCTT pGEX4T3-nAlb3: for BamHI CGTGGATCCTCCAGCTTAAACGAGATT rev XhoI CTGCTCGAGTTACTTCTGCACGGCGGATGA pET29b-cAlb4: (Gerdes et al., 2006)
Nachfolgend aufgeführte Primer wurden für die Klonierung des pETDuet1-cpSRP43-
Konstruktes benötigt, um rekombinantes His-cpSRP43 zu erhalten.
pETDuet1-cpSRP43: for BamHI CAGGATCCGGCCGTACAAAGAAACTAC rev HindIII GTAAGTTTCATTCATTCATTGGTTG
Die im Folgenden aufgelisteten Primer und Plasmide dienten der Herstellung von GST-
cpSRP43 Deletionskonstrukten für den Thylakoidmembran-Bindungstest.
pGEX4T3-cpSRP43(Ank1+CD2): for Ank1 BamHI GCTGGATCCGAGTACGAGACACCCTGG rev Ank1 overlap CD2 TTTCTCTAACCCAATTCTGACGTCTCGGTCCTCCAG for CD2 overlap Ank1 CTGGAGGACCGAGACGTCAGAATTGGGTTAGAGAAA rev CD2 XhoI GATCTCGAGTCACAGCCCATCCTCGTA pGEX4T3-cpSRP43(Ank2+CD2): for Ank2 BamHI GCTGGATCCGACGAAAACGGCCGGACG rev Ank2 overlap CD2 TTTCTCTAACCCAATTCTATCGGCTCCAGCCTCCGC for CD2 overlap Ank2 GCGGAGGCTGGAGCCGATAGAATTGGGTTAGAGAAA rev CD2 XhoI GATCTCGAGTCACAGCCCATCCTCGTA pGEX4T3-cpSRP43(Ank3+CD2): Template pGEX4T3-cpSRP43(ΔAnk4): for Ank3 BamHI GCTGGATCCATGAGGGGAGGCTTGACG rev CD2 XhoI GATCTCGAGTCACAGCCCATCCTCGTA

3 Material und Methoden 24
pGEX4T3-cpSRP43 (ΔAnk1): for BamHI GCT GGA TCC GCC GTA CAA AGA AAC TAC for ΔAnk1 overlap GCAGACGTAGTGTCGGATGCCGTGGACGAA rev ΔAnk1 overlap TTCGTCCACGGCATCCGACACTACGTCTGC rev XhoI GAT CTC GAG TCA TTC ATT CAT TGG TGG pGEX4T3-cpSRP43 (ΔAnk2): for BamHI GCT GGA TCC GCC GTA CAA AGA AAC TAC for ΔAnk2 overlap GACGTCGATGCCGTGCTCGACCACCGAGAC rev ΔAnk2 overlap GTCTCGGTGGTCGAGCACGGCATCGACGTC rev XhoI GAT CTC GAG TCA TTC ATT CAT TGG TGG pGEX4T3-cpSRP43 (ΔAnk3): for BamHI GCT GGA TCC GCC GTA CAA AGA AAC TAC for ΔAnk3 overlap CTCGACCACCGAGACATTGAAGTGGAAGAC rev ΔAnk3 overlap GTCTTCCACTTCAATGTCTCGGTGGTCGAG rev XhoI GAT CTC GAG TCA TTC ATT CAT TGG TGG pGEX4T3-cpSRP43, pGEX4T3-cpSRP43(ΔAnk4), pGEX4T3-cpSRP43(Ank4+CD2), pGEX4T3-cpSRP43(CD2-3): (Laborbestand)
Die folgenden Plasmide dienten der in vitro Transkription und Translation, dessen
Translationsprodukte für pull down Analysen eingesetzt wurden.
pSS-Alb4(328-499): for NcoI GACCCATGGTTTTGAGCACGGCACAGCAA rev SalI GGCGTCGACTTACCTCTTCTCTGTTTCATGAGA pSS-Alb3(299-462): for NcoI GACCCATGGTTATGAAGCCTCCTCAAACT rev SalI GTAGTCGACTTATACAGTGCGTTTCCGCTT pSS-Alb3(320-462): for NcoI GACCCATGGTTCCACTCATGATCGGTTAC rev SalI GTAGTCGACTTATACAGTGCGTTTCCGCTT pSS-Alb3(328-462): for NcoI GACCCATGGTTTTGTCTGTCCCATCAGGA rev SalI GTAGTCGACTTATACAGTGCGTTTCCGCTT pSS-Alb3(336-462): for NcoI GACCCATGGTTATTTACTGGCTCACAAAT rev SalI GTAGTCGACTTATACAGTGCGTTTCCGCTT pSS-Alb3(320-442): for NcoI GACCCATGGTTCCACTCATGATCGGTTAC rev SalI GTAGTCGACTTAGCTTGTTGTGCTCGAAGC pSS-Alb3(299-462 Δ314-317): Template pSPYCE-Alb3(Δ314-317): for NcoI GACCCATGGTTATGAAGCCTCCTCAAACT rev SalI GTAGTCGACTTATACAGTGCGTTTCCGCTT

3 Material und Methoden 25
pSS-Alb3(299-462 Δ389-415): Template pSPYCE-Alb3(Δ389-415): for NcoI GACCCATGGTTATGAAGCCTCCTCAAACT rev SalI GTAGTCGACTTATACAGTGCGTTTCCGCTT pSS-Alb3, pSS-pAlb4: (Laborbestand) pIVEX1.3 WG-Alb3(299-462): for NcoI GACCCATGGTTATGAAGCCTCCTCAAACT rev SalI GTAGTCGACTTATACAGTGCGTTTCCGCTT pIVEX1.3WG-Alb3(299-462 Δ374-388): Template pSPYCE-Alb3(Δ374-388): for NcoI GACCCATGGTTATGAAGCCTCCTCAAACT rev SalI GTAGTCGACTTATACAGTGCGTTTCCGCTT
Folgende Plasmide wurden für die in vivo Interaktionsstudien in Arabidopsis thaliana
Protoplasten eingesetzt.
pSPYNE-atpC1: for BamHI CACTAGTGGATCCATGGCTTGCTCTAATCTA rev SalI TACCCTCGAGGTCGACAACCTGTGCATTAGCTCC pSPYNE-atpC2: for BamHI CACTAGTGGATCCATGACAGGTTCGATCTCG rev SalI TACCCTCGAGGTCGACAGACTCTCGAAGAGCTTC pSPYNE-atpD: for BamHI CACTAGTGGATCCATGGCGTCTCTTCAACAA rev SalI TACCCTCGAGGTCGACAGTAGCTAATTGAATCTC pSPYNE-atpG: for BamHI CACTAGTGGATCCATGGCTGCTAATTCGATA rev SalI TACCCTCGAGGTCGACAGAAGGAAGAACCTTCTT pSPYCE-Alb4(1-142)Alb3(159-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC for overlap GCTACGTTCCCATTGACTAAGCAACAGGTT rev overlap AACCTGTTGCTTAGTCAATGGGAACGTAGC rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb4(1-210)Alb3(227-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC for overlap CTATATAGAGCCCTCTCTAACGTGGCCAAC rev overlap GTTGGCCACGTTAGAGAGGGCTCTATATAG rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb4(1-239)Alb3(256-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC for overlap GTTGCTGCAAGACAGAGTGGATCCGGCATT rev overlap AATGCCGGATCCACTCTGTCTTGCAGCAAC rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT

3 Material und Methoden 26
pSPYCE-Alb4(1-255)Alb3(272-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC for overlap ATTGAGGGACACCCACCATTGGGATGGTAT rev overlap ATACCATCCCAATGGTGGGTGTCCCTCAAT rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb4(1-288)Alb3(296-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC for overlap CAGTCCTCGCAGAGTATGGAAATTATGAAG rev overlap CTTCATAATTTCCATACTCTGCGAGGACTG rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb4(1-300)Alb3(314-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC for overlap TCACAAGCAGTGACTCTTGTTTTCAAGTTT rev overlap AAACTTGAAAACAAGAGTCACTGCTTGTGA rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb4(1-319)Alb3(336-462) entspricht pSPYCE-Alb4(1-302)Alb3(319-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC for overlap CCTTCTGGTTTAAGTATTTACTGGCTCACA rev overlap TGTGAGCCAGTAAATACTTAAACCAGAAGG rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb4(1-326)Alb3(343-462): for BamHI CGCCACTAGTGGATCCATGTCTTCAACAATATCC rev overlap TTGGGCGGTACTAAGTACGTT for overlap TATTGGCTGACCAACAACGTA rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb3(1-342)Alb4(327-499): for BamHI CGCCACTAGTGGATCCATGGCGAGAGTTCTAGTC rev overlap TGCCGTGCTCAAAATATTATTTGTGAGCCA for overlap TGGCTCACAAATAATATTTTGAGCACGGCA rev SalI TACCCTCGAGGTCGACCCTCTTCTCTGTTTCATGAGA pSPYCE-Alb3(1-359): for BamHI CGCCACTAGTGGATCCATGGCGAGAGTTCTAGTC rev SalI TACCCTCGAGGTCGACCTTTGCACCACCTAGTTT pSPYCE-Alb3(1-366): for BamHI CGCCACTAGTGGATCCATGGCGAGAGTTCTAGTC rev SalI TACCCTCGAGGTCGACTGCGTTTTCATCCATATT pSPYCE-Alb3(1-415): for BamHI CGCCACTAGTGGATCCATGGCGAGAGTTCTAGTC rev SalI TACCCTCGAGGTCGACTTCTTCTACCAATTCAAC pSPYCE-Alb3(1-442): for BamHI CGCCACTAGTGGATCCATGGCGAGAGTTCTAGTC rev SalI TACCCTCGAGGTCGACGCTTGTTGTGCTCGAAGC pSPYCE-Alb3(F316T): for AATACGCTTCTTGTTACCAAGTTTCTTCCACTC rev GAGTGGAAGAAACTTGGTAACAAGAAGCGTATT

3 Material und Methoden 27
pSPYCE-Alb3(L314A): for CAGAAGAATACGCTTGCTGTTTTCAAGTTTCTT rev AAGAAACTTGAAAACAGCAAGCGTATTCTTCTG pSPYCE-Alb3(L313Q,L314A): for GCACAGAAGAATACGCAGGCTGTTTTCAAGTTTCTT rev AAGAAACTTGAAAACAGCCTGCGTATTCTTCTGTGC pSPYCE-Alb3(Δ314,315): for GCACAGAAGAATACGCTTTTCAAGTTTCTTCCACTC rev GAGTGGAAGAAACTTGAAAAGCGTATTCTTCTGTGC pSPYCE-Alb3(Δ314-317): for CAGAAGAATACGCTTTTTCTTCCACTCAT rev CATGAGTGGAAGAAAAAGCGTATTCTTCTG pSPYCE-Alb3(Δ374-388): for BamHI CGCCACTAGTGGATCCATGGCGAGAGTTCTAGTC rev overlap TTTTAATTGTCTAAATCCCGCACTTATTAT for overlap ATAATAAGTGCGGGATTTAGACAATTAAAA rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb3(Δ389-415): for BamHI CGCCACTAGTGGATCCATGGCGAGAGTTCTAGTC rev overlap TGATTCAGACTGAGATCTTTCGCCAGCATC for overlap GATGCTGGCGAAAGATCTCAGTCTGAATCA rev SalI TACCCTCGAGGTCGACTACAGTGCGTTTCCGCTT pSPYCE-Alb3, pSPYNE-Alb3, pSPYCE-Alb4, pSPYNE-Alb4, pSPYCE-TS(rbcs)-cpSRP43, pSPYNE-TS(rbcs)-cpSRP43, pSPYCE-TS(rbcs)-cpSRP54, pSPYNE-TS(rbcs)-cpSRP54, pSPYCE-TS(rbcs)-cpFtsY, pSPYNE-TS(rbcs)-cpFtsY: (Laborbestand)
Nachfolgend gelistete Plasmide kamen bei den in vivo Interaktionsstudien im Hefe Split-
Ubiquitin System zum Einsatz.
pAMBV4-Alb3(Δ314-317): Template pSPYCE-Alb3(Δ314-317) for XbaI CTATCTAGACAAAAATGAGCTTAAACGAGATTCCTC rev Stu I ATGAGGCCTTTTACAGTGCGTTTCCGCTT pAMBV4-Alb3(Δ374-388): Template pSPYCE-Alb3(Δ374-388) for XbaI CTATCTAGACAAAAATGAGCTTAAACGAGATTCCTC rev Stu I ATGAGGCCTTTTACAGTGCGTTTCCGCTT pAMBV4-Alb3(Δ389-415): Template pSPYCE-Alb3(Δ389-415) for XbaI CTATCTAGACAAAAATGAGCTTAAACGAGATTCCTC rev Stu I ATGAGGCCTTTTACAGTGCGTTTCCGCTT pAMBV4-Alb3, pAMBV4-Alb4, pMBV-Alg5, pAlg5-NubI, pAlg5-NubG, pADSL-cpSRP43, pADSL-cpSRP54, pADSL-cpFtsY: (Laborbestand)

3 Material und Methoden 28
Des weiteren wurden folgende Sequenzierungsprimer verwendet:
pGEX4T3: for GGGCTGGCAAGCCACGTTTGGTG rev CCGGGAGCTGCATGTGTCAGAGG pETDuet1: for ATGCGTCCGGCGTAGAG rev GATTATGCGGCCGTGTACAA pSS: for GATTTAGGTGACACTATA rev TAATACGACTCACTATAGGG pIVEX1.3WG: for CGAAGTTGTAAAACGACGGCCAGT rev CAGGAAACAGCTATGAC pSPYCE: for AAGTTCATTTCATTTGGAGAG rev CTGCTTGTCAGCGTAATCTGG pSPYNE: for AAGTTCATTTCATTTGGAGAG rev CATAAGATCCTCCTCAGAAAT pAMBV4: for TTTCTGCACAATATTTCAAGC rev GTAAGGTGGACTCCTTCT

3 Material und Methoden 29
3.4.5 Mikroorganismen
3.4.5.1 Escherichia coli
Tabelle 3-6: Escherichia coli-Stämme
Stamm Genotyp Quelle
XL1 blue
recA1 endA1 gyrA96 thi-1 hsdR17, supE44 relA1 lac- [F´ pro AB lacIqZ∆M15 Tn10(tetr) ]
Stratagene, Bullock et al., 1987
BL21 (DE3) F– ompT hsdSB(rB –mB –) gal dcm
(DE3) Novagen
RosettaTM
F– ompT hsdSB(rB– mB–) gal dcm lacY1 (DE3) pRARE (argU, argW, ileX, glyT, leuW, proL) (CmR)
Novagen
3.4.5.2 Saccharomyces cerevisiae
Tabelle 3-7: Saccharomyces cerevisiae-Stamm
Stamm Genotyp Quelle
DSY-1 MATa, his3Δ200, trp1-901, leu2-3,112 Ade2, LYS2::(lexAop)4-HIS3, URA3::(lexAop)8-lacZ GAL4)
Dualsystems Biotech AG
3.4.6 Pflanzenmaterial
Bei dem für diese Arbeit verwendeten Pflanzenmaterial handelte es sich um die
Ackerschmalwand Arabidopsis thaliana (Columbia).

3 Material und Methoden 30
3.5 Anzuchtbedingungen
3.5.1 Anzucht und Lagerung von Escherichia coli
Die Anzucht von Escherichia coli Kulturen erfolgte in LB-Medium, welches zur Selektion
mit den entsprechenden Antibiotika versetzt wurde. Flüssigkulturen wurden bei 37 °C und
220 rpm über Nacht im Schüttler inkubiert. Die Anzucht der Kulturen auf LB-Festmedium
erfolgte bei 37 °C über Nacht im Wärmeschrank. Die kurzfristige Lagerung der Kulturen
geschah bei 4 °C. Um eine langfristige Lagerung zu gewährleisten, wurden Glycerolstocks
angelegt. Dazu wurden 600 μl der Bakterienkultur mit 600 μl 75 %igem Glycerol in
Kryoröhrchen gemischt und unverzüglich bei –80 °C gelagert.
LB-Medium: LB-Festmedium:
10 g Trypton 10 g Trypton
5 g Hefeextrakt 5 g Hefeextrakt
10 g NaCl 10 g NaCl
100 μl NaOH (10 M) 100 μl NaOH (10 M)
ad 1 l Aqua dest. 15 g Agar
ad 1 l Aqua dest.
Konzentrationen der Antibiotika im Medium:
Ampicillin: 50 μg/ml
Kanamycin: 50 μg/ml
Chloramphenicol: 34 μg/ml
3.5.2 Anzucht und Lagerung von Saccharomyces cerevisiae
Die Anzucht des Saccharomyces cerevisiae Stammes DSY-1 erfolgte auf Festagarplatten
über 3 Tage bei 30 °C. Wildtyp-Kulturen wurden auf YPAD-Agarplatten angezogen.
Flüssigmedium-Kulturen wurden von Stammplatten, die bei 4 °C gelagert wurden, mit
Einzelkolonien beimpft und in einem Schüttler bei 30 °C und 200 rpm über Nacht
inkubiert. Die Kultivierung und Selektion Plasmid-tragender Zellen erfolgte auf
entsprechendem drop-out-Medium.

3 Material und Methoden 31
YPAD-Vollmedium: YPAD-Agar:
10 g Hefeextrakt 20 g Agar
20 g Trypton ad 1 l YPAD
10 g Glukose Monohydrat
40 mg Adeninsulfat
ad 1 l Aqua dest.
drop-out-Medien:
-LT-Medium: -LT-Agar:
27 g DOB 20 g Agar
0,65 g CSM-Leu-Trp ad 1 l -LT-Medium
40 mg Adeninsulfat
ad 1 l Aqua dest.
-LTH-Medium: -LTH-Agar:
27 g DOB 20 g Agar
0,65 g CSM-Leu-Trp-His ad 1 l -LTH-Medium
40 mg Adeninsulfat
ad 1 l Aqua dest.
3.5.3 Pflanzenanzucht
Die Pflanzen wurden in einem (8,5 Stunden Licht/ 22 °C und 15,5 Stunden Dunkelheit/
19,5 °C)-Zyklus bei einer Lichtintensität von 100 μE m-2 s-1 und einer relativen
Luftfeuchtigkeit von 65 % in der Phytokammer angezogen.
3.6 Molekularbiologische Methoden
3.6.1 Polymerase-Kettenreaktion zur Amplifizierung von DNA-Fragmenten
Zur gezielten Vervielfältigung ausgewählter DNA-Fragmente wurden spezifische
Oligonukleotide (siehe 3.4.4) in eine Polymerase-Kettenreaktion (polymerase chain
reaction, PCR) unter Verwendung der KOD HiFi DNA-Polymerase (Novagen) eingesetzt.
Die Polymerase-Kettenreaktion ist ein in vitro Verfahren, um selektiv einen definierten
DNA-Abschnitt zu amplifizieren. Die PCR wurde mit Hilfe des Mastercyclers der Firma
Eppendorf durchgeführt.

3 Material und Methoden 32
PCR-Ansatz:
100 ng Template-DNA
10 µl 10x KOD Puffer # 1
10 µl dNTPs (je 2 mM)
4 µl MgCl2 (25 mM)
1 µl Primer forward (100 µM)
1 µl Primer reverse (100 µM)
0,8 µl KOD HiFi DNA-Polymerase
ad 100 µl Aqua dest.
PCR-Programm:
Denaturierung 2 min 98 °C
Denaturierung* 20 sec 98 °C
Annealing* 10 sec 55-60 °C
Elongation* 30 sec 72 °C
Kühlung 4 °C
*Dieser Zyklus wurde 25-mal durchgeführt.
Nach der PCR wurde ein Aliquot der Produkte in einem 1 %igen Agarosegel aufgetrennt
und die Größe mittels eines Größenstandards überprüft. Die Aufreinigung des PCR
Ansatzes erfolgte über das NucleoSpin Extract II Kit.
3.6.2 Site-directed Mutagenese PCR
Mit Hilfe des Mutagenesis-Kit Quik-Change XL wurden gezielte Aminosäureaustausche in
Plasmide eingebracht. Die site-directed Mutagenese PCR erfolgte nach Angaben des
Herstellers.
PCR-Ansatz:
50 ng Matrizen-DNA
4 μl Primer forward (10 μM)
4 μl Primer reverse (10 μM)
1 μl dNTP mix
5 μl 10x PCR-Puffer
3 μl Q-Sol
1 μl Pfu Turbo-DNA-Polymerase
ad 50 μl Aqua dest.

3 Material und Methoden 33
PCR-Programm:
Start 1 min 95 °C
Denaturierung* 50 sec 95 °C
Annealing* 50 sec 60 °C
Elongation* 20 min 68 °C
Finale Elongation 7 min 68 °C
Kühlung 4 °C
*Dieser Zyklus wurde 18-mal durchgeführt.
Im Anschluss an die PCR erfolgte ein Dpn I Verdau zum Abbau des methylierten
Ausgangsplasmids. Dazu wurde der PCR Ansatz mit 1 µl Dpn I versetzt und für
3 Stunden bei 37 °C inkubiert.
3.6.3 Overlap PCR
Mit Hilfe der Overlap PCR konnten in drei aufeinander folgenden PCR Schritten
Fusionskonstrukte aus zwei DNA-Fragmenten hergestellt werden. Zunächst wurden die
beiden DNA-Fragmente in getrennten PCR Ansätzen amplifiziert. Dazu wurden Primer
eingesetzt, deren überlappende Enden komplementär zu der Sequenz des anderen DNA-
Fragments waren (Overlap Primer). Im zweiten PCR Schritt erfolgte die Anlagerung der
beiden DNA-Fragmente anhand der komplementären Überhänge. Die komplementären
Bereiche dienten als Primer, die somit in der zweiten PCR fehlten. Die Amplifizierung des
Fusionskonstruktes erfolgte in einem dritten PCR Schritt durch den Einsatz des for-
Primers und rev-Primers aus der ersten PCR.
1. PCR:
PCR Ansatz DNA-Fragment 1:
100 ng Template-DNA
1 μl Primer for (ohne Überhang)
1 μl Overlap-Primer rev
10 μl dNTPs (je dNTP 2 mM)
4 µl MgCl2 (25 mM)
10 μl 10x KOD-Puffer #1
0,8 μl KOD HiFi DNA-Polymerase
ad 100 μl Aqua dest.

3 Material und Methoden 34
PCR Ansatz DNA-Fragment 2:
100 ng Template-DNA
1 μl Overlap-Primer for
1 μl Primer rev (ohne Überhang)
10 μl dNTPs (je dNTP 2 mM)
4 µl MgCl2 (25 mM)
10 μl 10x KOD-Puffer #1
0,8 μl KOD HiFi DNA-Polymerase
ad 100 μl Aqua dest.
2. PCR (ohne Zugabe von Primern)
Äquivalente Mengen der DNA-Fragmente 1 und 2 aus der ersten PCR
10 μl dNTPs (je dNTP 2 mM)
4 µl MgCl2 (25 mM)
10 μl 10x KOD-Puffer #1
0,8 μl KOD HiFi DNA-Polymerase
ad 100 μl Aqua dest.
3. PCR (Amplifikation des Fusionskonstruktes)
50 µl des PCR-Ansatzes nach der 2. PCR
1 μl Primer for (flankierender Primer von DNA-Fragment 1)
1 μl Primer rev (flankierender Primer von DNA-Fragment 2)
10 μl dNTPs (je dNTP 2 mM)
2 µl MgCl2 (25 mM)
5 μl 10x KOD-Puffer #1
0,8 μl KOD HiFi DNA-Polymerase
ad 100 μl Aqua dest.
Für alle PCR Schritte wurde das Programm für die KOD HiFi DNA-Polymerase (3.6.1)
verwendet.
3.6.4 DNA-Restriktion
Restriktionsendonukleasen erkennen spezifische Sequenzen in doppelsträngiger DNA
und führen an diesen Stellen zu Doppelstrangbrüchen, indem sie Phosphodiester-
bindungen hydrolysieren. Für die Restriktionsanalysen wurden TypII-Restriktions-
endonukleasen mit den Puffern laut Herstellerangaben (Fermentas) eingesetzt.

3 Material und Methoden 35
3.6.5 Ligation
Die Ligationen erfolgten mit der T4-DNA-Ligase von Fermentas. Die Reaktionen wurden
laut Herstellerangaben in einem Reaktionsvolumen von 15 μl angesetzt, wobei die
einzusetzenden DNA-Mengen in einem molaren Verhältnis von 1:3 (Vektor:Insert)
vorlagen. Die Ligationsansätze beinhalteten 1 µl T4-DNA-Ligase sowie 1,5 µl des T4-
DNA-Ligase Puffers und wurden für 2 Stunden bei 22 °C im Thermoblock inkubiert. Nach
einer anschließenden Hitzeinaktivierung der Ligase für 10 min bei 65 °C wurden die
Ansätze für die Transformation in Escherichia coli genutzt.
3.6.6 Herstellung CaCl2-kompetenter Escherichia coli-Zellen
Für die Herstellung von kompetenten Zellen wurde zunächst eine 5 ml LB-Vorkultur des
E. coli Stammes über Nacht bei 37 °C und 220 rpm angezogen. Mit dieser Vorkultur
konnte eine 100 ml LB-Hauptkultur angeimpft werden. Nach einer Inkubationszeit von
1 Stunde bei 37 °C im Schüttler wurden die Zellen sedimentiert (5000 rpm, 10 min). Das
Pellet wurde in 6 ml 100 mM CaCl2-Lösung resuspendiert. Die Zellsuspension wurde
30 min auf Eis inkubiert und anschließend erneut pelletiert (5000 rpm, 10 min). Das
Resuspendieren des Pellets erfolgte in 4 ml 100 mM CaCl2 mit 15 % Glycerin. Daraufhin
wurden 200 μl Aliquots der Zellen in 1,5 ml-Reaktionsgefäßen in flüssigem Stickstoff
eingefroren und bei –80 °C bis zur Verwendung gelagert.
3.6.7 Transformation von Plasmiden in CaCl2-kompetente Escherichia coli-Zellen
Über die Hitzeschock-Transformation wurden die Plasmide in Bakterien überführt. Dazu
wurden 200 µl eingefrorene kompetente Zellen auf Eis aufgetaut, mit der zu
transformierenden DNA (Ligationsansatz bzw. 100 ng Plasmid-DNA) gemischt und 20 min
auf Eis inkubiert. Nach einem Hitzeschock von 90 sec bei 42 °C wurden die Zellen eine
Minute auf Eis gestellt, mit 600 μl antibiotikafreiem LB-Medium (37 °C) versetzt und im
37 °C-Schüttler für 45 min bei 750 rpm regeneriert. Anschließend wurden die Zellen auf
Antibiotika-haltigen LB-Platten ausplattiert und über Nacht bei 37 °C zu Kolonien
herangezogen.

3 Material und Methoden 36
3.6.8 In-Fusion Klonierung
Die In-Fusion Klonierung stellt eine Alternative zur herkömmlichen Ligation dar. Mit Hilfe
des In-FusionTM Dry-Down PCR Cloning Kit der Firma Clontech werden DNA-Fragmente
über homologe Rekombination mit der Vektor-DNA verbunden. Zunächst wurde das
entsprechende DNA-Fragment mit speziellen Oligonukleotiden, die neben den
gewünschten Restriktionsschnittstellen einen zusätzlichen 15 bp Überhang der
Vektorsequenz beinhalteten, amplifiziert. Das aufgereinigte PCR-Produkt wurde mit dem
linearisierten Vektor und den im Kit befindlichen Reaktionskomponenten in einem
Volumen von 10 µl für 30 min bei 42 °C inkubiert. Der Ansatz wurde kurz abgekühlt, mit
40 µl TE-Puffer verdünnt und 5 µl der Verdünnung für die Transformation in E. coli (Kit)
eingesetzt. Nach 30 minütiger Inkubation auf Eis erfolgte der Hitzeschock für 45 sec bei
42 °C. Es folgte die Zugabe von 450 µl SOC-Medium (Kit), die Inkubation für eine Stunde
bei 37 °C und 750 rpm, das Ausplattieren der Zellen auf Antibiotika-haltigem LB-
Festmedium und die Anzucht über Nacht bei 37 °C.
TE-Puffer pH 7,5:
1,21 g Tris
0,29 g EDTA
ad 1 l Aqua dest.
3.6.9 Plasmid-Präparation
Zur Präparation von Plasmid-DNA aus E. coli wurde eine 1,5 ml-Übernachtkultur in
Antibiotika-haltigem LB-Medium bei Raumtemperatur und 13000 rpm für 30 sec pelletiert
und in 100 μl GET-Puffer resuspendiert. Die Zellen wurden durch Zugabe von 200 μl
0,2 M NaOH/1% SDS für maximal 5 min bei Raumtemperatur alkalisch lysiert. Nach der
Neutralisation mit 150 μl 3 M Kaliumacetat (pH 4,8) wurde die DNA mit 200 μl
Phenol/Chloroform (1:1) extrahiert. Zur Phasentrennung erfolgte eine Zentrifugation für
5 min bei Raumtemperatur und 13000 rpm. Die obere wässrige Phase wurde in ein
frisches Reaktionsgefäß überführt und mit 1 ml Ethanol (-20 °C) für 30 min bei -20 °C
gefällt. Die DNA wurde für 10 min bei 4 °C und 14000 rpm pelletiert. Das Pellet wurde mit
300 µl 70% Ethanol (-20 °C) gewaschen, getrocknet und in 50 μl Aqua dest.
aufgenommen. Die Lagerung erfolgte bei -20 °C. Die nach dieser Methode isolierte
Plasmid-DNA enthielt noch RNA und wurde für RNase-haltige Kontrollrestriktionen
verwendet. Zur Gewinnung von möglichst reiner Plasmid-DNA wurde die Plasmidisolation
mit einem NucleoSpin Plasmid Mini Prep Kit (Macherey & Nagel) durchgeführt. Für große
Mengen an Plasmid-DNA wurde das Plasmid Maxi Prep Kit (Promega) verwendet.

3 Material und Methoden 37
GET-Puffer:
50 mM Glukose
25 mM Tris/HCl pH 8,0
10 mM EDTA pH 8,0
3.6.10 DNA-Konzentrationsbestimmung
Zur photometrischen Bestimmung der DNA-Konzentration wurde die Extinktion einer 1:50
verdünnten DNA-Lösung bei 260 nm in UV-Küvetten gemessen. Die Konzentration der
DNA wurde durch das Photometer berechnet nach der Formel:
LösungDNA
Küvette
VVE
lg
−
××=⎥
⎦
⎤⎢⎣
⎡ 50260
μμ
3.6.11 Restriktionsanalyse und Sequenzierung
Um eine erfolgreiche Ligation überprüfen zu können, wurde die isolierte Plasmid-DNA
restringiert und die DNA-Fragmente über Agarosegelelektrophorese aufgetrennt. Plasmid-
DNA mit einem korrekten Restriktionsmuster wurde sequenziert. Die Sequenzierungen
wurden bei dem Sequenzierungsservice der Fakultät für Chemie und Biochemie an der
Ruhr-Universität Bochum durchgeführt.
3.6.12 Agarosegelelektrophorese zur Trennung von DNA
Zur Auftrennung und Größenabschätzung von DNA-Fragmenten wurde eine Agarose-
gelelektrophorese in horizontalen Elektrophoreseapparaturen durchgeführt. Die Agarose
wurde in 1x TAE-Puffer durch Aufkochen gelöst, mit 1 μg Ethidiumbromid pro ml
Agarosegel versetzt und in die Apparatur gegossen. Durch den Zusatz des
Ethidiumbromids konnte die DNA unter Ultraviolett-Bestrahlung sichtbar gemacht werden.
Als Laufpuffer für die Elektrophorese wurde ebenfalls 1x TAE-Puffer verwendet. Die DNA-
Proben wurden mit DNA-Ladepuffer versetzt und die Elektrophorese bei einer Spannung
von 100 V durchgeführt. Als Größenstandard wurde der Gene RulerTM 1kb DNA Ladder
eingesetzt.

3 Material und Methoden 38
50x TAE:
242 g Tris
in 500 ml Aqua dest. lösen
100 ml 0,5 M Na2EDTA pH 8,0
51,1 ml Essigsäure
ad 1 l Aqua dest.
5x DNA-Ladepuffer:
3,75 ml 75% Glycerin
500 μl 2,5% Orange G (in Aqua dest.)
100 μl 1x TAE
650 μl Aqua dest.
3.6.13 Split-Ubiquitin System
Das Split-Ubiquitin System basiert auf dem Hefe two-hybrid System und ermöglicht die
Analyse der Interaktionen von Membranproteinen in vivo. Das Ubiquitin markiert in seiner
eigentlichen Funktion abnorme Proteine für deren proteolytischen Abbau. In diesem
System ist das Ubiquitin in zwei Hälften gespalten. Das Nub-Fragment stellt den
N-Terminus und das Cub-Fragment den C-Terminus des Ubiquitins dar. Das Cub-Protein
mit einem angehängten Reporter wird von dem Vektor pAMBV4 und das Nub-Protein
durch den Vektor pADSL-Nx der Firma Dualsystems Biotech AG (DUALmembrane Kit)
kodiert. Um die Interaktion von zwei Membranproteinen zu analysieren, werden die
beiden Proteine so an die Ubiquitin-Fragmente fusioniert, dass die Ubiquitin-Hälften
cytosolisch lokalisiert sind. Ein Reporter, bestehend aus DNA-bindender Domäne (LexA-
Protein) und transkriptions-aktivierender Domäne (Virionenprotein 16), wird an das Cub-
Fragment fusioniert. Im Falle der Rekonstitution des Ubiquitins wird der Reporter
proteolytisch abgespalten und kann im Nukleus die Reportergene aktivieren. Die DNA-
bindende Domäne kann sich an den LexA-Operator anlagern und die aktivierende
Domäne die Transkription der Reportergene starten. Bei den Reportergenen des DSY1-
Hefestamms handelt es sich um das HIS3, welches der Selektion auf Minimalmedium
dient, und den Reporter lacZ, der für die β-Galaktosidase kodiert.

3 Material und Methoden 39
3.6.13.1 Transformation von Saccharomyces cerevisiae mit Plasmid-DNA
Aus einer 5 ml-Übernachtkultur des Hefestamms DSY-1 in YPAD-Medium wurde eine
100 ml Hauptkultur auf eine OD600 von 0,2 angeimpft und bei 200 rpm und 30 °C bis zu
einer OD600 von 0,5 inkubiert. Die Zellen wurden in zwei 50 ml-Reaktionsgefäßen für
5 min bei 2000 rpm abzentrifugiert und die Pellets anschließend mit je 40 ml Aqua dest.
gewaschen. Danach wurden diese in je 1 ml 0,1 M Lithiumacetat (LiAc) aufgenommen
und auf zwei 1,5 ml-Reaktionsgefäße verteilt. Die Zellsuspension wurde für 15 sec bei
14000 rpm sedimentiert und jedes Pellet in 480 μl 0,1 M LiAc resuspendiert. Pro
Transformationsansatz wurden 100 μl Zellsuspension in einem 1,5 ml-Reaktionsgefäß
kurz abzentrifugiert und in 50 μl 0,1 M LiAc aufgenommen. Den kompetenten Hefezellen
wurden 480 μl 50 % PEG, 72 μl 1 M LiAc und je 1 μg Plasmid-DNA (bait- und prey-
Vektor) zugefügt. Die Ansätze wurden mit Aqua dest. auf 720 μl aufgefüllt, 1 min gevortext
und für 30 min bei Raumtemperatur auf einem Drehrad inkubiert. Nach der Inkubation
wurden die Ansätze mit 72 μl Dimethylsulfoxid gemischt. Nach dem Hitzeschock für
15 min bei 42 °C wurden die Zellen für 15 sec bei 6000 rpm sedimentiert. Die Zellpellets
wurden in 300 μl Aqua dest. resuspendiert und je 150 μl auf –LT- und –LTH-Platten
ausgestrichen. Die Inkubation erfolgte für 3 Tage bei 30 °C.
3.6.13.2 β-Galaktosidase Filtertest
Die Aktivität der β-Galaktosidase ist ein Reporter für eine Interaktion zweier Proteine
innerhalb des Split-Ubiquitin Systems und lässt sich über den β-Galaktosidase Filtertest
überprüfen. Dazu wurde auf die Hefekolonien der –LTH-Platten nach 3 Tagen Wachstum
bei 30 °C ein Rundfilter aufgelegt und anschließend die anhaftenden Hefezellen in
flüssigem Stickstoff aufgebrochen. Nach dem Auftauen des Filters wurde dieser auf einen
Stapel aus drei mit X-Gal-Reaktionslösung durchfeuchteten Filterpapieren gelegt, so dass
die Hefekolonien nach oben zeigten. Die Filter wurden abgedeckt und bei 30 °C für
2 Stunden inkubiert. Bei Interaktion der beiden Fusionsproteine sollte eine Blaufärbung
beobachtet werden können, die auf der β-Galaktosidase Aktivität beruht und zur semi-
quantitativen Auswertung dient. Das Enzym ist in der Lage das Galaktose-Analog X-Gal
(5-Bromo-4-chloro-3-indoyl-β-D-Galaktopyranosid) zu einem blauen Farbstoff umzu-
setzen.

3 Material und Methoden 40
Z-Puffer pH 7,0: X-Gal-Reaktionslösung:
10,7 g Na2HPO4 x 2 H2O 30 ml Z-Puffer
5,5 g NaH2PO4 x H2O 81 μl β-Mercaptoethanol
0,75 g KCl 501 μl X-Gal-Lösung
0,24 g MgSO4 x 7 H2O
ad 1 l H2O
X-Gal-Lösung:
20 mg/ml X-Gal in Dimethylformamid
3.6.14 Bimolekulare Fluoreszenzkomplementation (Split-YFP System)
Das auf der bimolekularen Fluoreszenzkomplementation basierende Split-YFP System
dient der in vivo Analyse der Protein-Protein Interaktionen in intakten Pflanzenzellen. Das
yellow fluorescent protein (YFP) stellt ein Derivat des green fluorescent protein (GFP) dar,
welches in der Zellbiologie für in vivo Lokalisationsstudien verwendet wird. In dem Split-
YFP System ist das YFP in zwei Hälften gespalten. Das nYFP umfasst die N-terminale
Hälfte und das cYFP entspricht dem C-Terminus des YFPs. Das nYFP wird durch den
Vektor pSPYNE (split YFP N-terminal fragment expression) und das cYFP durch den
Vektor pSPYCE (split YFP C-terminal fragment expression) kodiert (Walter et al., 2004).
Für die Interaktionstests werden die zu untersuchenden Proteine so an die YFP-
Fragmente fusioniert, dass die YFP-Hälften auf der Stroma gelegenen Seite lokalisiert
sind. Im Falle einer Interaktion der beiden Fusionsproteine rekonstituieren sich die beiden
nicht-fluoreszierenden YFP-Hälften zu dem fluoreszierenden YFP, dessen Fluoreszenz an
einem Laserscanning Mikroskop (LSM 510 Meta) sichtbar gemacht und dokumentiert
werden kann. Der Transport der Proteine aus dem Cytosol in den Chloroplasten wird über
die jeweilige chloroplastidäre Transitsequenz der Proteine gewährleistet. Fehlt diese
Transitsequenz, wurde die Transitsequenz der kleinen Untereinheit der Rubisco vor das
jeweilige Konstrukt kloniert, um den chloroplastidären Import zu erreichen. Dazu wurden
die beiden Vektoren pSPYNE-TS(rbcs) und pSPYCE-TS(rbcs) eingesetzt. Diese Vektoren
wurden ebenfalls für die Negativkontrollen eingesetzt, da somit der Import der einzelnen
YFP-Hälften allein garantiert ist.

3 Material und Methoden 41
3.6.14.1 Isolierung von Arabidopsis thaliana Protoplasten und transiente Transfektion
Für die in vivo Interaktionsstudien der Proteine im Split-YFP System wurden zunächst
Arabidopsis thaliana Protoplasten isoliert. Dazu wurden 10-15 Blätter von Arabidopsis
thaliana (1-2 cm Blattlänge im Rosettenstadium) in 0,5-1 mm dünne Streifen geschnitten
und zum Abbau der Zellwände in 25 ml Enzymlösung unter Vakuum im Dunkeln inkubiert.
Danach wurden die Protoplasten durch ein Nylonsieb (40-70 µm) filtriert und somit vom
groben Blattmaterial getrennt. Nach erfolgter Zentrifugation (100 g, 2 min, soft start und
soft stop) wurden die Protoplasten mit Lösung D gewaschen und mit Hilfe einer
Thomakammer die Zellzahl bestimmt. Die Zellzahl wurde auf 2x105/ml eingestellt und die
Protoplasten für 30 min auf Eis inkubiert. Nach erfolgter Zentrifugation (100 g, 2 min, soft
start und soft stop) wurden die Protoplasten in Lösung C aufgenommen (2x105/ml) und
standen für die Transfektion zur Verfügung. Für die Transfektion wurden je 40 µg
Plasmid-DNA (pSPYCE- und pSPYNE-Vektor), 500 µl Protoplastensuspension und 550 µl
Lösung B in einem 15 ml Reaktionsgefäß gut vermischt und für 30 min bei
Raumtemperatur inkubiert. Nach der Transfektion wurden die Protoplasten mit 3 ml
Lösung D gewaschen, erneut zentrifugiert (100 g, 2 min, soft start und soft stop) und in
400 µl Lösung D aufgenommen. Die Protoplasten wurden in Petrischalen überführt und in
4 ml Lösung D für 16-18 Stunden bei Raumtemperatur inkubiert. Für die Analyse am
Fluoreszenz Mikroskop wurden die Protoplasten zentrifugiert (100 g, 2 min, soft start und
soft stop) und das Pellet in 100 µl Lösung D aufgenommen. Die Auswertung der
Proteininteraktionen über die Rekonstitution der YFP-Fluoreszenz erfolgte mit Hilfe des
Laserscanning Mikroskops (LSM 510 Meta).
Enzymlösung: Lösung B:
1,5 % Cellulase R10 61,5 % PEG 4000
0,4 % Macerozyme R10 0,3 M Mannit
0,4 M Mannit 0,15 M CaCl2
20 mM KCl
20 mM MES pH 5,7
10 min, 55 °C
10 mM CaCl2
0,1 % BSA

3 Material und Methoden 42
Lösung C: Lösung D:
0,4 M Mannit 154 mM NaCl
15 mM MgCl2 125 mM CaCl2
4 mM MES pH 5,7 5 mM KCl
2 mM MES pH 5,7
3.7 Biochemische Methoden
3.7.1 Überexpression von Proteinen in Escherichia coli
Zunächst wurde aus einer Antibiotika-haltigen E.coli Übernachtkultur eine Hauptkultur
angeimpft und diese bei 37 °C und 220 rpm im Schüttler bis zu einer OD600 von 0,5-0,7
herangezogen. Darauf erfolgte die Induktion der Proteinexpression mit einer IPTG-
Endkonzentration von 1 mM. Nach einer Inkubation von 3 Stunden wurden die Zellen
geerntet und das überexprimierte Protein aufgereinigt. Zur Kontrolle wurde ein Aliquot von
500 µl vor der Induktion (t0) bzw. 3 Stunden nach der Induktion (t3) abgenommen und die
Überexpression mittels SDS-PAGE überprüft.
IPTG-Stocklösung (100 mM):
238,3 mg IPTG
ad 1 ml Aqua dest.
3.7.2 Denaturierende Aufreinigung von Proteinen mit His-tag über Ni-NTA
Für die denaturierende Aufreinigung überexprimierter Proteine mit His-tag wurden
zunächst 250 ml Zellkultur geerntet und das Pellet in 4 ml Puffer B (pH 8,0)
aufgenommen. Die Lyse erfolgte durch vorsichtiges Vortexen, um Schaumbildung zu
vermeiden, und Inkubation für 1 Stunde bei Raumtemperatur. Nach einer zehnminütigen
Zentrifugation bei 14000 rpm wurde das Lysat in einem 15 ml Reaktionsgefäß mit 1 ml Ni-
NTA-Slurry über Nacht bei Raumtemperatur invertiert. Die Ni-NTA-Beads wurden dann
bei 5000 rpm für 3 min abzentrifugiert und der Überstand verworfen. Die Beads wurden
dreimal mit je 5 ml Puffer C (pH 5,9) gewaschen und beim letzten Waschschritt in eine
Säule (Biorad PolyPrep Column) überführt. Das benutzte Reaktionsgefäß wurde
anschließend mit 5 ml Puffer C nachgespült. Nachdem der Puffer durch die Säule
gelaufen war, wurde diese in ein frisches 1,5 ml-Reaktionsgefäß gestellt und das

3 Material und Methoden 43
gebundene Protein mit 0,5 ml Puffer E (pH 4,5) eluiert. Dieser Elutionsschritt erfolgte
insgesamt dreimal, wobei die Eluate nicht vereinigt wurden. Zur Überprüfung der
Aufreinigung wurden sowohl vom Lysat, als auch vom Überstand nach der Inkubation mit
den Ni-NTA-Beads je 10 µl abgenommen, um diese mit je 10 µl der Eluatfraktionen über
eine SDS-PAGE kontrollieren zu können.
Puffer B (Lysepuffer) pH 8,0: Puffer C (Waschpuffer) pH 5,9:
100 mM NaH2PO4 100 mM NaH2PO4
10 mM Tris-HCl 10 mM Tris-HCl
8 M Harnstoff 8 M Harnstoff
Puffer E (Elutionspuffer) pH 4,5:
100 mM NaH2PO4
10 mM Tris-HCl
8 M Harnstoff
3.7.3 Native Aufreinigung von Proteinen mit His-tag über Ni-NTA
Für die native Aufreinigung überexprimierter Proteine mit His-tag wurden zunächst 400 ml
Zellkultur geerntet und das Pellet in 10 ml Lysepuffer aufgenommen. Der Aufschluss der
Zellen erfolgte durch eine Ultraschallbehandlung (50 W, 50% Pulse, 4 min). Nach der
Zentrifugation für 30 min bei 14000 rpm und 4 °C wurde das Lysat abgenommen und über
Nacht bei 4 °C mit 500 µl Ni-NTA-Slurry auf dem Drehrad inkubiert. Danach wurden die
Beads dreimal mit 10 ml Waschpuffer gewaschen und in eine Säule (Biorad PolyPrep
Column) überführt. Die Säule wurde dreimal mit je 10 ml Waschpuffer gewaschen und
das gebundene Protein anschließend mit 0,5 ml Elutionspuffer eluiert. Dieser
Elutionsschritt erfolgte dreimal, wobei die Eluate nicht vereinigt wurden. Zur Überprüfung
der Aufreinigung wurden sowohl vom Lysat, als auch vom Überstand nach der Inkubation
mit den Beads je 10 µl abgenommen, um diese mit je 10 µl der Eluatfraktionen über eine
SDS-PAGE kontrollieren zu können.
Lysepuffer pH 8,0: Waschpuffer pH 8,0:
50 mM HEPES 50 mM HEPES
300 mM NaCl 300 mM NaCl
1 mM DTT 1 mM DTT
10 mM Imidazol 20 mM Imidazol

3 Material und Methoden 44
Elutionspuffer pH 8,0:
50 mM HEPES
300 mM NaCl
1 mM DTT
250 mM Imidazol
3.7.4 Aufreinigung von Proteinen mit GST-tag über Glutathion-Sepharose
Für die Aufreinigung von Proteinen mit GST-tag wurde zunächst 1 ml Glutathion-
Sepharose-Beads abzentrifugiert (1500 rpm, 5 min, 4 °C), mit 4 ml eiskaltem 1x TBS
gewaschen und in 1,5 ml 1x TBS aufgenommen. Daraufhin wurden 5 x 500 ml Zellkultur
geerntet (4000 g, 10 min, 4 °C) und die Pellets in 20 ml Lysepuffer, dem zuvor ½ Tablette
complete Mini Protease Inhibitor Cocktail Tablets der Firma Roche zugesetzt wurde,
resuspendiert. Zur Lyse der Zellen erfolgte eine Ultraschallbehandlung (50 W, 50% Pulse,
4 min). Das Lysat wurde abzentrifugiert (30 min, 4 °C, 18000 rpm), zu den vorbereiteten
Glutathion-Sepharose-Beads gegeben und über Nacht bei 4 °C invertiert. Danach wurden
die Beads dreimal mit 15 ml Lysepuffer gewaschen und beim letzten Waschschritt in eine
Chromatographie-Säule (Biorad PolyPrep Column) überführt. Die Säule wurde dreimal mit
je 10 ml Puffer B gewaschen und das gebundene Protein anschließend mit 0,5 ml
Glutathion-Puffer eluiert. Dieser Elutionsschritt erfolgte dreimal, wobei die Eluate nicht
vereinigt wurden. Zur Überprüfung der Aufreinigung wurden sowohl vom Lysat, als auch
vom Überstand nach der Inkubation mit den Beads je 10 µl abgenommen, um diese mit je
10 µl der Eluatfraktionen über eine SDS-PAGE kontrollieren zu können.
Lysepuffer: Puffer B:
50 ml Tris pH 8,0 50 ml Tris pH 8,0
17,53 g NaCl 8,8 g NaCl
0,15 g DTT ad 1 l Aqua dest.
10 ml Triton X-100
0,3 g EDTA
ad 1 l Aqua dest.
Glutathion-Puffer: 10x TBS, pH 7,5:
3,07 g Glutathion (reduziert) 60,5 g Tris
50 ml Tris pH 8,0 87,6 g NaCl
ad 1 l Aqua dest. ad 1 l Aqua dest.

3 Material und Methoden 45
3.7.5 Abspaltung des GST-tags über Thrombinverdau
Über das pGEX4T3-Expressionssystem wurde ein N-terminaler GST-tag an das zu
exprimierende Protein angefügt. Für die Abspaltung des GST-tags wurden 25 μg
aufgereinigtes Protein und 5 Units Thrombin in 200 μl Puffer B+ gegeben. Die Inkubation
erfolgte über Nacht bei 4 °C auf dem Drehrad.
Puffer B+:
50 mM Tris pH 8,0
150 mM NaCl
2,5 mM CaCl2
3.7.6 Dialyse von Proteinen
Um aufgereinigte Proteine in andere Pufferlösungen zu überführen, wurden diese mit Hilfe
von Dialysekassetten der Firma PIERCE gegen den gewünschten Puffer dialysiert. Die
Dialyse erfolgte zunächst unter Rühren für zwei Stunden in 2 l Puffer bei
Raumtemperatur. Danach wurde das Protein in 2 l frischem Puffer über Nacht bei 4 °C
weiter dialysiert.
3.7.7 BCA-Proteinbestimmung
Zur Proteinkonzentrationsbestimmung wurde das BCATM Protein Assay Kit der Firma
PIERCE verwendet. Dazu wurde sowohl eine BSA-Eichreihe (0-30 μg BSA), als auch
verschiedene Mengen der zu untersuchenden Proteinlösung und gleiche Mengen des
Puffers, in dem das Protein gelöst war, mit Aqua dest. auf 50 µl aufgefüllt. Diese Proben
wurden mit 1 ml BCA-Mix, bestehend aus 50 Teilen Reagenz A und einem Teil
Reagenz B, versetzt und 30 min bei 60 °C inkubiert. Nach Zentrifugation der Proben
wurde die optische Dichte bei 562 nm bestimmt und die Proteinkonzentration mit Hilfe der
Eichgeraden ermittelt.
3.7.8 SDS-Polyacrylamid-Gelelektrophorese (SDS-PAGE)
Die SDS-PAGE wurde zur Trennung von Proteingemischen nach dem Molekulargewicht
verwendet und im vertikalen, diskontinuierlichen Gelsystem aus Sammelgel und Trenngel
durchgeführt. Die Acrylamidkonzentration des Trenngels richtete sich nach der Größe der
zu analysierenden Proteine. Die mit Auftragspuffer versetzten Proben wurden bei 98 °C

3 Material und Methoden 46
für 2 min denaturiert, gevortext, abzentrifugiert und in die Geltaschen eingebracht. Die
Elektrophorese erfolgte bei 30 mA pro Gel in 1x Laufpuffer (mit SDS). Proben, die erst
später verwendet werden sollten wurden bei -20 °C gelagert. Vor dem Auftragen wurden
diese erneut auf 98 °C erhitzt. Als Größenstandard diente der Marker Page RulerTM
Prestained Protein Ladder der Firma Fermentas.
Tabelle 3-8: Trenngel pH 8,8 (4 Minigele):
Acrylamidkonzentration 10% 12% 15%
30 % Acrylamid (29:1) 5,3 ml 6,4 ml 8 ml
1,5 M Tris pH 8,8 4 ml 4 ml 4 ml
Aqua dest. 3,3 ml 5,6 ml 4 ml
20 % SDS 0,08 ml 0,08 ml 0,08 ml
TEMED 0,02 ml 0,02 ml 0,02 ml
25 % APS 0,06 ml 0,06 ml 0,06 ml
Sammelgel pH 6,8 (4 Minigele): Sammelgel-Stock pH 6,8:
6 ml Sammelgel-Stock pH 6,8 50 ml 30 % Acrylamid (29:1)
6 μl TEMED 1,5 ml 20 % SDS
48 μl 25 % APS 18,8 ml 1 M Tris pH 6,8
48 μl Bromphenolblau-Lösung ad 300 ml Aqua dest.
4x Auftragspuffer: Bromphenolblau-Lösung:
2 ml 1 M Tris pH 6,8 40 mg Bromphenolblau
4 ml Glycerol ad 10 ml Aqua dest.
4 ml 20 % SDS
800 μl β-Mercaptoethanol
3 mg Bromphenolblau
5x Laufpuffer (ohne SDS) : 1x Laufpuffer (mit SDS):
15 g Tris 200 ml 5x Laufpuffer
72 g Glycin 5 ml 20% SDS
ad 1 l Aqua dest. ad 1 l Aqua dest.

3 Material und Methoden 47
3.7.9 Coomassie-Färbung
Zum Nachweis der Proteine wurden die SDS-Polyacrylamidgele mit Coomassie-
Färbelösung bedeckt und auf dem Schüttler für 15 min gefärbt. Die Färbelösung wurde
gegen Entfärber ausgewechselt und das Gel solange inkubiert, bis die Proteinbanden
deutlich zu erkennen waren und der Hintergrund komplett entfärbt war. Anschließend
wurde das Gel in Wasser äquilibriert, bis es seine ursprüngliche Größe wieder erreicht
hatte, und auf dem Geltrockner unter Vakuum auf einem Filterpapier getrocknet.
Coomassie-Färbelösung: Coomassie-Entfärber:
2,5 g Coomassie Blau R 250 500 ml Methanol
500 ml Methanol 100 ml Essigsäure
100 ml Essigsäure ad 1 l Aqua dest.
ad 1 l Aqua dest.
3.7.10 Elektrophoretischer Transfer von Proteinen auf Nitrocellulosemembranen (Western-Blot)
Beim Western-Blot werden die aufgetrennten Proteingemische aus der Polyacrylamid-
matrix über ein senkrecht zum Gel angelegtes elektrisches Feld auf eine
Nitrocellulosemembran transferiert. An der Membranoberfläche bleiben diese aufgrund
hydrophober Wechselwirkungen haften, so dass das Muster der elektrophoretischen
Auftrennung erhalten bleibt. Der Transfer der Proteine erfolgte über das Tank-Blot
Verfahren für 90 min bei 400 mA und 4 °C.
Blotpuffer:
9,1 g Tris
43,2 g Glycin
600 ml Methanol p.a.
ad 3 l Aqua dest.
3.7.11 Proteinfärbung mit Ponceau S
Membranen, die für Immunodetektionen verwendet wurden, konnten mit Ponceau S
reversibel angefärbt werden, um das Gesamtprotein-Profil zu überprüfen. Dazu wurden
die Membranen 5 min bei Raumtemperatur in Ponceau S geschwenkt und mit TBST so
lange entfärbt, bis das Proteinbandenmuster in geeignetem Kontrast sichtbar wurde. Für
die weitere Verwendung wurden die Membranen in TBST vollständig wieder entfärbt.

3 Material und Methoden 48
10x TBS pH 7,5: 1x TBST:
60,5 g Tris 100 ml 10x TBS pH 7,5
87,6 g NaCl 3 ml Tween 20
ad 1 l Aqua dest. ad 1 l Aqua dest.
Ponceau S-Lösung:
0,5 g Ponceau S
1 ml Essigsäure
ad 100 ml Aqua dest.
3.7.12 Immundetektion von Proteinen
Nach dem Transfer der Proteine auf die Nitrocellulosemembran wurden diese für eine
Stunde bei Raumtemperatur in Block-Puffer geschwenkt, um freie Bindungsstellen
abzusättigen. Die Inkubation mit den entsprechenden Konzentrationen des ersten
Antikörpers gegen das gewünschte Protein erfolgte über Nacht bei 4 °C in
10 ml Block-Puffer auf dem Schwenktisch. Nicht gebundener Antikörper wurde durch
dreimaliges 10 minütiges Waschen mit TBST entfernt. Der zweite, Peroxidase-gekoppelte
Antikörper war gegen die IgGs des ersten Antikörpers gerichtet. Diese Inkubation erfolgte
in Block-Puffer für 1 Stunde bei Raumtemperatur. Nicht gebundener Antikörper wurde
durch dreimaliges 10 minütiges Waschen mit TBST entfernt. Die Membranen wurden mit
Super Signal West Pico (Femto) Luminol/Enhancer Solution und Super Signal West Pico
(Femto) Stable Peroxide Buffer (1:1) der Firma PIERCE benetzt, mit Klarsichtfolie bedeckt
und die Chemilumineszenz auf einem Röntgenfilm festgehalten.
10x TBS pH 7,5: 1x TBST:
60,5 g Tris 100 ml 10x TBS pH 7,5
87,6 g NaCl 3 ml Tween 20
ad 1 l Aqua dest. ad 1 l Aqua dest.
Block-Puffer:
1 g Magermilchpulver
ad 10 ml 1x TBST

3 Material und Methoden 49
3.7.13 Affinitätsaufreinigung von Antikörpern
Um Kontaminationen und damit verursachte unspezifische Bindungen eines Antikörpers
bei der Immunodetektion zu entfernen, wurde dieser mit Hilfe des „Affi-Gels 15“ der Firma
Biorad aufgereinigt. Dazu wurde 0,1 ml Affi-Gel in einem 1,5 ml-Reaktionsgefäß zweimal
mit eiskaltem Aqua dest. gewaschen. Die Zentrifugationsschritte für das Waschen
erfolgten für 30 sec bei 6000 rpm. Danach erfolgte die Bindung von 1 mg nativ
aufgereinigtem Protein, gegen das der Antikörper gerichtet war, in einem Volumen von
400 μl an die Affi-Gel Agarose durch Invertieren für vier Stunden bei 4 °C. Zuvor musste
das Protein gegen 50 mM Hepes (pH 8,0) dialysiert werden, um das bei der Bindung an
das Affi-Gel störende Imidazol des Elutionspuffers zu entfernen. Die Affi-Gel Agarose
wurde bei 6000 rpm und 4 °C für 30 sec abzentrifugiert und vom Überstand zur Kontrolle
der Bindung des Proteins eine Proteinbestimmung gemacht. Die Absättigung der
Bindestellen erfolgte durch Zugabe von 400 μl 1 M Ethanolamin (pH 8,0) und Invertieren
für 1 Stunde bei 4 °C. Danach wurde die Affi-Gel Agarose dreimal mit 0,5 ml eiskaltem
TBS gewaschen. Für die Bindung des Antiserums wurde zunächst ein Aliquot von 1,5 ml
für 30 min bei 56 °C inaktiviert und sofort auf Eis abgekühlt. Nach der Zentrifugation
(2 min, 14000 rpm) wurde der Überstand zu der vorbereiteten Agarose gegeben und über
Nacht bei 4 °C invertiert. Das Affi-Gel wurde abzentrifugiert und der Überstand zur
Kontrolle der Bindung des Antiserums aufbewahrt. Danach wurde die Agarose zweimal
mit 0,5 ml eiskaltem TBS und zweimal mit 0,5 ml 500 mM NaCl in 20 mM Tris (pH 7,5)
gewaschen. Die Elution des gebundenen Antiserums erfolgte zunächst fünfmal mit 250 μl
100 mM Glycin (pH 2,5). Die Eluate wurden sofort in 1,5 ml-Reaktionsgfäße, die mit 21 μl
1 M Tris und 25 μl 5 mg/ml BSA vorbereitet waren, überführt. Nach dem Waschen der
Affi-Gel Agarose mit zweimal 0,5 ml 50 mM Tris (pH 8,5) erfolgte der nächste
Elutionsschritt fünfmal mit 250 μl 100 mM Ethanolamin (pH 11,5). Die Eluate wurden
sofort in 1,5 ml-Reaktionsgfäße, die mit 25 μl 1 M Tris (pH 7,5) und 25 μl 5 mg/ml BSA
vorbereitet waren, überführt. Danach wurde noch zweimal mit 0,5 ml TBS gewaschen und
der letzte Elutionsschritt fünfmal mit 250 μl 3,5 M MgCl2 in 20 mM Tris (pH 7,5)
durchgeführt. Das Affi-Gel wurde mit TBS gewaschen und bei 4 °C aufbewahrt. Zur
Kontrolle der Aufreinigung wurde zunächst das Antigen über SDS-PAGE aufgetrennt und
ein Western-Blot mit einer 1:100 Verdünnung der Eluate durchgeführt. Anhand des
Western-Blots konnten die Bedingungen und Eluate bestimmt werden, mit denen das
Antiserum von den Beads eluiert werden konnte.

3 Material und Methoden 50
3.7.14 In vitro Transkription
Über die in vitro Transkription konnten Gene, die im pSS-Vektor der Regulation durch
einen SP6-Promotor unterlagen, transkribiert werden. Der Reaktionsansatz von 50 µl
setzte sich aus 40 Units RNase-Inhibitor, 20 Units SP6-Polymerase, 5 μl NT-Mix mit CAP,
10 μl 5x Transkriptionspuffer und 5 µg Plasmid-DNA (in 10 µl DEPC-Wasser) zusammen
und wurde für 30 min bei 37 °C inkubiert. Danach wurde der Ansatz mit 50 μl 10 M
Ammoniumacetat und 100 μl DEPC-Wasser auf eine Endkonzentration von 2,5 M
Ammoniumacetat eingestellt. Nach der Extraktion mit 200 μl Phenol/Chloroform
(1:1, sauer) wurde die wässrige Phase abgenommen und die mRNA durch Zugabe von
400 μl eiskaltem Ethanol für 30 min bei -20 °C gefällt. Die mRNA wurde für 15 min bei
14000 rpm und 4 °C abzentrifugiert, das Pellet in 70%igem eiskaltem Ethanol gewaschen,
getrocknet und in 60 μl DEPC-Wasser aufgenommen.
NT-Mix: NT-Mix mit CAP:
5 μl 100 mM GTP 233 μl NT-Mix
50 μl 100 mM CTP 25 Units CAP (Natrium-
50 μl 100 mM ATP Diguanosintriphosphat)
50 μl 100 mM UTP
ad 1 ml Aqua dest.
3.7.15 In vitro Translation
Die in vitro Translation wurde über das Weizenkeimlysat-System der Firma Promega
durchgeführt, um sowohl radioaktiv markierte Proteine als auch nicht markierte Proteine
zu erhalten. Dazu wurden 12 μl der in vitro transkribierten mRNA bei 68 °C für 10 min
denaturiert und auf Eis abgekühlt. Das Weizenkeimlysat wurde für 5 min bei 6500 rpm
und 4 °C zentrifugiert und der Überstand weiter verwendet. Der Translationsansatz setzte
sich aus 20 μl Weizenkeimlysat, 4 μl Aminosäuremix ohne Methionin, 40 Units RNase-
Inhibitor, 5 μl 1 M Kaliumacetat, 12 μl mRNA und 2 μl [35S]-Methionin bzw. 8 µl
unmarkiertem Methionin (10 mM) zusammen. Die Translation erfolgte für 90 min bei 25
°C. Zur Translationskontrolle wurde 1 μl radioaktiv markiertes bzw. 5 µl nicht markiertes
Translationsprodukt über SDS-PAGE aufgetrennt und mit anschließender
Autoradiographie bzw. Immundetektion überprüft.

3 Material und Methoden 51
3.7.16 In vitro Expression mit RTS 100 Wheat Germ
Über das RTS 100 Wheat Germ Kit der Firma Roche konnten Gene, die im pIVEX1.3WG-
Vektor der Regulation durch einen T7-Promotor unterlagen, in einem Schritt und in
größerer Ausbeute transkribiert und translatiert werden. Dazu mussten zunächst die
Komponenten des Kits in folgenden Volumina des Rekonstitutionspuffers gelöst werden:
Wheat Germ (in 140 µl Rekonstitutionspuffer), Reaktionsmix (in 140 µl
Rekonstitutionspuffer), Feeding Mix (in 7,7 ml Rekonstitutionspuffer), Aminosäuren (in
800 µl Rekonstitutionspuffer) und Methionin (in 2,2 ml Rekonstitutionspuffer). Daraufhin
wurden die folgenden Reaktionslösungen angesetzt:
Feeding-Lösung:
900 µl Feeding Mix
80 µl Aminosäuren
20 µl Methionin
Reaktions-Lösung:
15 µl Reaktionsmix
4 µl Aminosäuren
1 µl Methionin
15 µl Wheat Germ
4 µg DNA
ad 50 µl RNase/DNase freies Wasser
Die Reaktions-Lösung wurde in die durch einen roten Ring gekennzeichnete Kammer
eingefüllt und die Feeding-Lösung in die benachbarte Kammer. Die Öffnungen der
Kammern wurden mit Parafilm verschlossen und die Ansätze für 24 Stunden bei 24 °C
und 900 rpm inkubiert. Das Translationsprodukt wurde aus der mit einen roten Ring
gekennzeichnete Kammer in ein frisches Reaktionsgefäß überführt. Zur
Translationskontrolle wurde 1 μl Translationsprodukt über SDS-PAGE aufgetrennt und mit
anschließender Immundetektion überprüft.
3.7.17 Pull down Experiment
Mit Hilfe des pull down Experiments sollte die in vitro Interaktion von His-Fusionsproteinen
mit in vitro Translationsprodukten analysiert werden. Dazu wurden 5 µg eines His-
Fusionsproteins, 15 µl Proteaseinhibitor (7x Stocklösung complete Mini Protease Inhibitor
Cocktail Tablets) und gleiche Mengen verschiedener in vitro Translationsprodukte mit

3 Material und Methoden 52
Inkubationspuffer auf ein Volumen von 100 µl gebracht und für 30 min bei
Raumtemperatur auf dem Drehrad inkubiert. Danach wurden 20 µl Ni-NTA beads zu den
Ansätzen gegeben und diese weitere 30 min bei Raumtemperatur auf dem Drehrad
belassen, um das His-Fusionsprotein gemeinsam mit dem möglichen Interaktionspartner
an die beads zu binden. Anschließend wurden die beads in Wizard Minisäulen
abzentrifugiert und dreimal mit je 200 µl Waschpuffer gewaschen, um nicht gebundene
Proteine zu entfernen. Die Elution erfolgte nach 10 minütiger Inkubationszeit mit 30 µl
Elutionspuffer. Zusammen mit dem His-Fusionsprotein coeluiertes Translationsprodukt
wurde, nach Auftrennung der Proben über SDS-PAGE, mittels Immundetektion
nachgewiesen.
Inkubationspuffer pH 8,0: Waschpuffer pH 8,0:
50 mM HEPES 50 mM HEPES
10 mM MgCl2 300 mM NaCl
1 mM DTT
20 mM Imidazol
Elutionspuffer pH 8,0:
50 mM HEPES
300 mM NaCl
1 mM DTT
250 mM Imidazol
3.7.18 Peptide-Scanning
Die Methode des Peptide-Scanning wurde genutzt, um die Bindestelle eines Proteins für
ein anderes Protein zu identifizieren. Dazu wurde eine PepSpot-Membran bestellt
(PepSpots, JPT Peptide Technologies, Berlin), auf der pro spot 15 Aminosäuren lange
Peptide des einen Proteins verlinkt sind. Die nebeneinander liegenden spots sind jeweils
um 3 andere Aminosäuren versetzt aufgetragen. Die Membran wurde mit einem
rekombinanten Protein inkubiert, dessen Bindestelle auf der PepSpot-Membran somit
analysiert werden konnte.
3.7.18.1 Bindung des rekombinanten Proteins an die PepSpot-Membran
Alle folgenden Inkubations- und Waschschritte wurde auf dem Schwenktisch
durchgeführt. Die PepSpot-Membran wurde zunächst für 1 min in einer Petrischale mit
Ethanol inkubiert und dreimal 10 min mit TBS (I) gewaschen. Danach wurde die Membran

3 Material und Methoden 53
in 10 ml Blockpuffer für 3 Stunden bei Raumtemperatur geblockt, um unspezifische
Bindungen zu vermeiden. Nach dem Waschen der Membran mit TBST (I) für 10 min,
erfolgte die Inkubation mit 50 µg rekombinantem Protein in 10 ml Blockpuffer für zunächst
2 Stunden bei Raumtemperatur und dann über Nacht bei 4 °C. Daraufhin wurde die
Membran dreimal 1 min mit TBST (II) gewaschen und das gebundene rekombinante
Protein konnte über einen elektrophoretischen Transfer auf eine PVDF-Membran
transferiert werden.
TBS (I) pH 8,0: TBST (I):
50 mM Tris TBS pH 8,0
137 mM NaCl 0,05 % Tween 20
2,7 mM KCl
10x TBS (II) pH 7,5: 1x TBST (II):
60,5 g Tris 100 ml 10x TBS pH 7,5
87,6 g NaCl 3 ml Tween 20
ad 1 l Aqua dest. ad 1 l Aqua dest.
Blockpuffer:
TBST (II)
3 % BSA
3.7.18.2 Elektrophoretischer Transfer eines rekombinanten Proteins von der PepSpot-Membran auf eine PVDF-Membran
Um das an die PepSpots gebundene, rekombinante Protein auf eine PVDF-Membran zu
transferieren wurde das Semi-Dry Blotverfahren durchgeführt. Zunächst wurde die PVDF-
Membran kurz in Methanol und dann für 10 min in Anodenpuffer I geschwenkt. Vor dem
Blotverfahren wurden je 2 Stücke Blotting Papier in Kathodenpuffer, Anodenpuffer I und
Anodenpuffer II äquilibriert. Der Aufbau des Blots erfolgte nach folgender Anordnung:
Anode
2x Blotting Papier in Anodenpuffer II
2x Blotting Papier in Anodenpuffer I
PVDF-Membran
PepSpot-Membran
2x Blotting Papier in Kathodenpuffer
Kathode

3 Material und Methoden 54
Der Proteinfransfer erfolgte in drei aufeinander folgenden Blotverfahren. Es wurden zwei
Blotschritte für je 30 min und der letzte Blotschritt für 1 Stunde bei 1 mA/cm2 durchgeführt,
wobei jeweils die PVDF-Membran und das Blotting Papier gewechselt wurden. Die PVDF-
Membranen wurden zunächst in TBST (II) aufbewahrt und schließlich zweimal für 10 min
mit TBST (II) gewaschen. Das Blocken der Membranen erfolgte in je 10 ml Blockpuffer für
3 Stunden bei Raumtemperatur. Die Inkubation mit dem ersten Antikörper gegen das auf
die PVDF-Membranen transferierte, rekombinante Protein und die anschließende
Immundetektion erfolgte wie in Kapitel (Immundetektion von Proteinen) beschrieben.
Anhand der auf dem Röntgenfilm detektierten spots konnte die Bindestelle des
rekombinanten Proteins auf der PepSpot-Membran zugeordnet werden.
Anodenpuffer I: Anodenpuffer II:
30 mM Tris 300 mM Tris
20 % Methanol 20 % Methanol
Kathodenpuffer (pH 9,2): Blockpuffer:
25 mM Tris TBST (II)
40 mM 6-Aminohexansäure 3 % BSA
20 % Methanol
10x TBS (II) pH 7,5: 1x TBST (II):
60,5 g Tris 100 ml 10x TBS pH 7,5
87,6 g NaCl 3 ml Tween 20
ad 1 l Aqua dest. ad 1 l Aqua dest.
3.7.18.3 Regeneration der PepSpot-Membran
Zur Regeneration und damit Wiederverwendung der PepSpot-Membran wurde diese
zunächst dreimal für 10 min mit Aqua dest. bei Raumtemperatur und daraufhin viermal
für 30 min mit Regenerationspuffer bei 50 °C gewaschen. Im Anschluss erfolgten drei
Waschschritte für je 20 min mit 10x PBS, drei Waschschritte für je 20 min mit TBST (I)
und drei weitere Waschschritte für je 10 min mit TBS (I) bei Raumtemperatur. Um zu
überprüfen, ob die regenerierte Membran frei von rekombinantem Protein ist, erfolgte ein
Semi-Dry Blotverfahren von der PepSpot-Membran auf PVDF-Membranen wie bereits in
Kapitel 3.7.18.2 beschrieben. Das Ausbleiben von spots auf dem Röntgenfilm nach einer
Immundetektion gegen das zuvor eingesetzte rekombinante Protein zeigte den Erfolg der
Regeneration der PepSpot-Membran.

3 Material und Methoden 55
Regenerationspuffer: 1x PBS (pH 7,2):
62,5 mM Tris 9,2 mM Na2HPO4 x 12 H2O
2 % SDS 1,6 mM NaH2PO4 x H2O
mit HCl auf pH 6,7 einstellen 150 mM NaCl
70 µl β-Mercaptoethanol/10 ml Puffer
TBS (I) pH 8,0: TBST (I):
50 mM Tris TBST pH 8,0
137 mM NaCl 0,05 % Tween 20
2,7 mM KCl
3.7.19 Präparation von Thylakoidmembranen aus Arabidopsis thaliana
Für die Präparation von Thylakoidmembranen wurden 20 g Blattmaterial von fünf Wochen
alten Pflanzen geerntet und die Blätter im Mixer (Warring Blender) mit 3 kurzen Intervallen
in 400 ml frisch angesetztem Homogenisierungspuffer zerschlagen. Wichtig für die
Aufarbeitung war, dass alle Arbeitsschritte im Kühlraum bzw. auf Eis mit kalten Puffern
stattfanden. Die groben Gewebeteile wurden durch das Filtrieren über 2 Lagen Miracloth
abgetrennt und das Filtrat für 5 min bei 4000 rpm und 4 °C abzentrifugiert. Das Pellet
wurde vorsichtig mit einem Haarpinsel in 40 ml SH-Puffer resuspendiert. Nach erfolgter
Zentrifugation (5 min, 4000 rpm, 4 °C) wurde das Pellet in 15 ml HM-Puffer resuspendiert
und 5 min auf Eis inkubiert, um die Chloroplasten osmotisch aufzuschließen. Die
Thylakoidmembranen wurden zentrifugiert und in 10 ml HM-Puffer für eine
Chlorophyllbestimmung aufgenommen. Anschließend wurden die Membranen pelletiert
und in 20 mM HEPES (pH 8) auf eine Chlorophyllkonzentration von 2 mg Chlorophyll/ml
eingestellt.
Homogenisierungspuffer: SH-Puffer:
330 mM Sorbitol 330 mM Sorbitol
50 mM HEPES pH 8,0 50 mM HEPES pH 8,0
10 mM EDTA pH 8,0
0,05 % BSA
5 mM Ascorbinsäure
HM-Puffer:
10 mM HEPES pH 8,0
5 mM MgCl2

3 Material und Methoden 56
3.7.19.1 Chlorophyllbestimmung
Für die Chlorophyllbestimmung wurden 20 μl der Thylakoidmembranen in 980 μl
80 %igem Aceton durch Vortexen gelöst und die unlösliche Komponenten durch
Zentrifugation für 2 min bei 14000 rpm entfernt. Die Absorption des Überstandes wurde im
Photometer bei 645 nm und 663 nm gegen einen Leerwert von 80 %igem Aceton
gemessen. Die Berechnung des Chlorophyllgehaltes erfolgte nach:
( ) 5002,82,20 663645 ××+×=⎥⎦⎤
⎢⎣⎡ AAmlgμ
3.7.19.2 Salzwaschung der Thylakoidmembranen
Die isolierten Thylakoidmembranen von Arabidopsis thaliana wurden pelletiert (5 min,
4000 rpm, 4 °C) und im gleichen Volumen Salzwaschpuffer resuspendiert. Nach einem
erneuten Waschschritt mit Salzwaschpuffer und IBM-Puffer, wurde das Pellet in dem
Ausgangsvolumen mit IBM-Puffer aufgenommen.
Salzwaschpuffer: IBM-Puffer:
50 mM HEPES pH 8,0 50 mM HEPES pH 8,0
330 mM Sorbitol 330 mM Sorbitol
1 M Kaliumacetat 10 mM MgCl2
3.7.19.3 Solubilisierung der Thylakoidmembranen
Für die Solubilisierung der Thylakoidmembranen wurden gleiche Mengen kaltem
2x Detergenzpuffer und Thylakoidmembranen (2 mg Chlorophyll/ml) gemischt und für
30 min auf Eis inkubiert. Nach erfolgter Zentrifugation (14000 rpm, 15 min, 4 °C) wurden
die solubilisierten Proteine aus dem Überstand in ein frisches Reaktionsgefäß überführt
und standen für weitere Versuche zur Verfügung.
2x Detergenzpuffer: (frisch ansetzen)
3 % Detergenz (Dodecyl-β-D-maltosid, Digitonin)
400 mM NaCl
2 mM PMSF
20 mM HEPES

3 Material und Methoden 57
3.7.20 Crosslinking der Thylakoidmembranproteine von Arabidopsis thaliana
Für die Crosslinking Versuche wurden 500 µl Thylakoidmembranen (0,2 mg
Chlorophyll/ml) in IBM-Puffer auf eine Endkonzentration der Crosslinking-Reagenzien von
0,6 mM gebracht und für 30 min bei 25 °C auf dem Drehrad inkubiert. Danach wurden die
Membranen zentrifugiert (4000 rpm, 5 min, 4 °C), zweimal mit Waschpuffer gewaschen
und das Pellet in 200 µl SDS-Probenpuffer aufgenommen.
IBM-Puffer: Waschpuffer:
50 mM HEPES pH 8,0 50 mM HEPES pH 8,0
330 mM Sorbitol 330 mM Sorbitol
10 mM MgCl2 1 M KAc
3.7.21 Coimmunpräzipitation
3.7.21.1 Kopplung des Antikörpers an Protein-A-Sepharose beads
Zur Coimmunpräzipitation der Thylakoidmembranproteine wurden zunächst 10 mg
Protein-A-Sepharose beads in 1 ml TBS für 30 min bei Raumtemperatur im Drehinkubator
äquilibriert, kurz anzentrifugiert und der Überstand verworfen. Die beads wurden in 200 µl
Antikörperserum aufgenommen, mit TBS auf 800 µl aufgefüllt und über Nacht bei 4 °C auf
dem Drehinkubator belassen. Danach wurden die beads anzentrifugiert und dreimal mit
1 ml Borat-Puffer gewaschen. Zur Kontrolle der darauf folgenden Kopplung wurden 10 µl
der Ansätze entnommen. Die Kopplung des Antikörpers erfolgte in einer
Dimethylpimelimidat-Endkonzentration von 20 mM für 30 min bei Raumtemperatur im
Drehinkubator. Danach wurden erneut 10 µl der Ansätze entnommen und die Proben (vor
und nach der Kopplung) durch eine SDS-PAGE mit anschließender Coomassie-Färbung
auf ausreichende Verringerung der freien IgGs überprüft. Die beads wurden sedimentiert
und mit 1 ml 0,2 M Tris (pH 8,0) gewaschen. Eine anschließende 30 minütige Inkubation
in 1 ml 0,2 M Tris (pH 8,0) diente dem Quenchen des überschüssigen Crosslinkers. Die
beads wurden sedimentiert, in 1 ml 0,3 M Essigsäure aufgenommen und die freien IgGs
für 5 min bei Raumtemperatur im Drehinkubator entfernt. Die Neutralisation der
Essigsäure erfolgte durch zweimaliges Waschen mit 1 ml 0,2 M Tris (pH 8,0). Die mit
Antikörpern gekoppelten Protein-A Sepharose beads wurden in 300 µl 0,2 M Tris (pH 8,0)
aufgenommen und konnten bei 4 °C gelagert werden.

3 Material und Methoden 58
10x TBS: Borat-Puffer (pH 9,0):
60,5 g Tris 12,37 g Borsäure
87,6 g NaCl ad 800 ml Aqua dest.
ad 1 l Aqua dest. pH einstellen mit 10 M NaOH
ad 1 l Aqua dest.
3.7.21.2 Präzipitation
Für die Präzipitation wurden Thylakoidmembranen von Arabidopsis thaliana mit 1,5 %
Digitonin bzw. Dodecyl-β-D-maltosid solubilisiert und 1:1 in 1x Detergenzpuffer (ohne
Detergenz) verdünnt. Des weiteren wurden 300 µl mit Antikörper gekoppelte Protein-A-
Sepharose (PAS) beads sedimentiert und der Überstand entfernt. Die beads wurden in
1 ml solubilisierter, verdünnter Thylakoidmembranen (0,5 mg Chlorophyll/ml)
aufgenommen. Zur Antikörperbindung wurde der Ansatz auf dem Drehrad für eine Stunde
bei 4 °C inkubiert. Die PAS-beads wurden zweimal mit 1 ml kaltem Waschpuffer im batch
gewaschen, in Wizard-Mini-Säulen überführt und abzentrifugiert. Zur Elution wurden die
Ansätze mit 30 µl 4x SDS-Probenpuffer für 10 min bei Raumtemperatur inkubiert und
abzentrifugiert. Die Auswertung erfolgte über SDS-PAGE mit anschließender
Immundetektion.
1x Detergenzpuffer (ohne Detergenz): Waschpuffer:
200 mM NaCl 20 mM HEPES pH 8,0
1 mM PMSF 80 mM NaCl
10 mM HEPES pH 8,0
3.7.22 Thylakoidmembran-Bindungstest
Um die Proteininteraktionen zwischen rekombinanten Proteinen und isolierten
Thylakoidmembranen zu analysieren, wurden die Thylakoidmembranen zunächst durch
Salzwaschung (3.7.19.2) von peripher gebundenen Proteinen befreit. Für den
Bindungstest wurden 25 µl salzgewaschene Thylakoidmembranen (2 mg Chlorophyll/ml)
mit 2 µg rekombinantem Protein gemischt und auf ein Volumen von 200 µl mit IBM-Puffer
aufgefüllt. Die Inkubation des rekombinanten Proteins mit den Membranen erfolgte für
30 min bei 4 °C auf dem Drehrad. Anschließend wurden die Membranen pelletiert (5 min,
4000 rpm, 4 °C) und nicht gebundenes Protein durch zweimaliges Waschen der
Membranen mit je 200 µl Waschpuffer entfernt. Nach den Waschschritten wurden die
Thylakoidmembranen in 80 µl 4x SDS-Probenpuffer resuspendiert. Die Bindung der

3 Material und Methoden 59
rekombinanten Proteine an die Membranen konnte über eine SDS-PAGE mit
anschließender Immundetektion nachgewiesen werden.
IBM-Puffer: Waschpuffer:
50 mM HEPES pH 8,0 50 mM HEPES pH 8,0
330 mM Sorbitol 330 mM Sorbitol
10 mM MgCl2 10 mM MgCl2
1 M NaCl
3.7.23 Thylakoidmembran-Bindungstest mit Peptidinhibierung
Um die Bedeutung einer bestimmten Bindestelle zwischen einem rekombinanten Protein
und einem Protein der Thylakoidmembran zu bestätigen, wurde ein synthetisches Peptid
eingesetzt. Dazu wurden 2 µg des rekombinanten Proteins und 250 µg bzw. 500 µg des
Peptids auf ein Volumen von 175 µl mit IBM-Puffer aufgefüllt und für 30 min bei 4 °C auf
dem Drehrad vorinkubiert. Danach wurden den Ansätzen 25 µl salzgewaschene
Thylakoidmembranen hinzugefügt und erneut für 30 min bei 4 °C auf dem Drehrad
inkubiert. Anschließend wurden die Membranen pelletiert (5 min, 4000 rpm, 4 °C) und
nicht gebundenes Protein durch zweimaliges Waschen der Membranen mit je 200 µl
Waschpuffer entfernt. Nach den Waschschritten wurden die Thylakoidmembranen in 80 µl
4x SDS-Probenpuffer resuspendiert. Die Bindung der rekombinanten Proteine an die
Membranen konnte über eine SDS-PAGE mit anschließender Immundetektion
nachgewiesen werden.
IBM-Puffer: Waschpuffer:
50 mM HEPES pH 8,0 50 mM HEPES pH 8,0
330 mM Sorbitol 330 mM Sorbitol
10 mM MgCl2 10 mM MgCl2
1 M NaCl
3.7.24 Expressionsnachweis der YFP-Fusionsproteine in Arabidopsis thaliana Protoplasten
Für den Expressionsnachweis der YFP-Fusionsproteine wurden 4 ml Arabidopsis thaliana
Protoplasten (2x105/ml) mit 240 µg Plasmid-DNA transfiziert und für 16-18 Stunden
inkubiert. Die Protoplasten wurden pelletiert (1500 rpm, 5 min, 4 °C) und in 400 µl HM-
Puffer und 40 µl Proteaseinhibitor (7x Stocklösung complete Mini Protease Inhibitor
Cocktail Tablets) für 3 min auf Eis lysiert. Nach der Zentrifugation (14000 rpm, 5 min,

3 Material und Methoden 60
4 °C) wurde der Überstand abgenommen und einer Trichloressigsäure-Präzipitation
unterzogen. Im Anschluss erfolgte das Waschen des Pellets mit 400 µl 0,2 M NaOH und
eine erneute Zentrifugation (14000 rpm, 5 min, 4 °C). Der Überstand (Waschfraktion)
wurde ebenfalls einer Trichloressigsäure-Präzipitation unterzogen und das Membranpellet
in 10 µl SDS-Probenpuffer aufgenommen. Die Proteine des Membranpellets, des
Überstands und der Waschfraktion wurden über eine SDS-PAGE aufgetrennt und eine
Immundetektion durchgeführt. Die Proteinexpression der YFP-Fusionsproteine konnte
über die Immundetektion mittels eines anti-HA Antikörpers für die cYFP-Fusionsproteine
und eines anti-c-myc Antikörpers gegen die nYFP-Fusionsproteine nachgewiesen
werden.
HM-Puffer:
10 mM HEPES pH 8,0
5 mM MgCl2
3.7.25 Trichloressigsäure-Präzipitation
Das Aufkonzentrieren von Proteinen erfolgte durch Zugabe von einem zehntel Volumen
100 %iger Trichloressigsäure (TCA) zur Proteinlösung mit anschließender Inkubation für
30 Minuten auf Eis. Nach einer Zentrifugation (14000 rpm, 5 min, 4 °C) wurde das
Proteinpellet in 200 µl 10 %iger TCA aufgenommen, für 10 min auf Eis inkubiert und
erneut zentrifugiert (14000 rpm, 5 min, 4 °C). Anschließend wurde der Überstand
abgenommen, das Pellet in 10 µl 4x SDS-Probenpuffer aufgenommen und bis zum
Farbumschlag mit 1 M Tris titriert.
3.8 Biophysikalische Methoden
3.8.1 Massenspektrometrie
Die massenspektrometrische Proteinsequenzierung erfolgte mittels eines Q-TOF
Massenspektrometers. Dazu wurde die entsprechende Proteinbande aus einem
Polyacrylamidgel ausgeschnitten und entfärbt. Es folgte der enzymatische Verdau über
Trypsin und die Elution der Peptide aus dem Gel. Nach Einkonzentrieren der Peptide
standen diese für die Proteinsequenzierung zur Verfügung. Die anschließende

3 Material und Methoden 61
Auswertung der Proteinsequenzierung erfolgte über Mascot (Matrix Science,
http://www.matrixscience.com/search_form_select.html).
3.8.2 Gelfiltration
Die Gelfiltration ist ein chromatographisches Verfahren, bei dem die Trennung der
Proteine aufgrund unterschiedlicher Teilchengröße erfolgt. Das Säulenmaterial enthält
Poren, die von den Molekülen passiert werden müssen. Die Proteine und
Proteinkomplexe diffundieren in einem konstanten Laufmittelfluss anhand ihrer Größe
unterschiedlich schnell durch das Säulenmaterial. Kleine Moleküle durchlaufen die Säule
langsamer. Es kommt zur Fraktionierung der aufgrund ihrer Größe aufgetrennten
Proteine.
3.8.2.1 Eichung der Gelfiltrationssäule
Die Chromatographiesäule wurde mit dem Gelfiltrationspuffer bei einer Flussrate von
0,2 ml/min über 20 Stunden äquilibriert. Darauf erfolgte die Eichung der Säule mit sechs
Markerproteinen (je 500 µl) nach folgendem Programm:
Eichprogramm:
0-10 ml Äquilibrierung mit Gelfiltrationspuffer
10-12 ml Injektion der Eichlösung
15-135 ml Elution
Flussrate: 0,5 ml/min
Folgende Proteine wurden zur Eichung verwendet:
1. Dextran blue (1 mg/ml) Ausschlussvolumen
2. Thyroglobulin 669 kDa
3. Apoferritin 443 kDa
4. ß-Amylase 200 kDa
5. Alkoholdehydrogenase 150 kDa
6. BSA 66 kDa
Über die Elution der Proteine, die vom Detektor bei 280 nm gemessen wurde, konnte das
Ausschlussvolumen bestimmt und eine Eichgerade erstellt werden.

3 Material und Methoden 62
3.8.2.2 Gelfiltration der solubilisierten Thylakoidmembranproteine
Die solubilisierten Thylakoidmembranproteine (500 µl, 1 mg Chlorophyll/ml) wurden filtriert
und über eine Superose 6 10/300 GL Säule nach folgendem Programm aufgetrennt.
Gelfiltrationsprogramm:
0-10 ml Äquilibrierung mit Gelfiltrationspuffer
10-12 ml Injektion der Probe
18-38 ml Fraktionierung von je 1 ml
38-135 ml Äquilibrierung mit Gelfiltrationspuffer
Flussrate: 0,4 ml/min
Die erhaltenen Fraktionen wurden zu je 1 ml aufgefangen. Aliquots von 75 µl der
Fraktionen wurden über eine SDS-PAGE aufgetrennt und die Proteine in einer
anschließenden Immundetektion nachgewiesen.
Gelfiltrationspuffer, entgast und filtriert:
20 mM HEPES pH 8,0
200 mM NaCl
0,1 % Dodecyl-β-D-maltosid

4 Ergebnisse 63
4. Ergebnisse
4.1 Herstellung spezifischer Antikörper gegen Alb3 und Alb4
Um die beiden homologen Proteine Alb3 und Alb4 in biochemischen Untersuchungen
getrennt voneinander nachweisen zu können, mussten zunächst spezifische Antikörper
gegen diese Proteine hergestellt werden. Es lag bereits ein spezifischer Antikörper gegen
Alb4 (generiert in Kaninchen) vor, der in Gerdes et al., 2006 beschrieben wurde.
Ausgehend von dem Antigen, welches für diesen Antikörper genutzt wurde, sollte nun das
gleiche Antigen für die Herstellung eines Alb4 Antikörpers in Ratte dienen. Die Herstellung
eines zweiten Antikörpers in einem anderen Organismus wurde besonders für die
Coimmunpräzipitationsanalysen wichtig und wird an der entsprechenden Stelle dieser
Arbeit näher erläutert.
Des Weiteren sollte ein spezifischer Antikörper gegen Alb3 (generiert in Kaninchen)
hergestellt werden. Die Proteine Alb3 und Alb4 zeigen eine Ähnlichkeit von 72 % und eine
Identität von 55 %. In einem Alignment der Proteinsequenzen von Alb3 und Alb4
(Abbildung 4-1) sind die Antigensequenzen markiert, die für die Antikörperherstellung
ausgewählt wurden. Bei dem Alb4 Antikörper wurde das bereits erwähnte Antigen des
C-terminalen Bereichs von Alb4 verwendet (AS 347-494). Entscheidend für die
Gewinnung von spezifischen Antikörpern ist es, einen Bereich des Proteins für das
Antigen auszuwählen, welcher zu dem anderen Protein nicht so homolog ist. Die Proteine
Alb3 und Alb4 weisen die geringste Homologie im N-terminalen und C-terminalen Bereich
auf. Daher wurden für die Sequenzen der Antigene von Alb3 die Aminosäuren 56-108 im
N-Terminus (nAlb3) und die Aminosäuren 397-462 im C-Terminus (cAlb3) ausgewählt.

4 Ergebnisse 64
Alb3 MARVLVSSPSSFFGSPLIKPSSSLRHSGVGGGGTAQFLPYRSNNNKLFTTSTTVRFSLNEAlb4 ---------------MSSTISLKPTHLILSSFSTGKVLQFRRSRFSHTPSSSSSRYRTLV . * . * :.. .*.:.* :* .. . .:*:: *: Alb3 IPPFHGLDSSVDIGAIFTRAESLLYTIADAAVVG--ADSVVTTDSSAVQKSGGWFGFISD Alb4 AQLGFRPDS---FDFIKDHAENLLYTIADAAVSSSETFESVAGTTTKTTQSNDWFSGIAN . ** :. * :**.********** . : . *: :: . :*..**. *:: Alb3 AMELVLKILKDGLSAVHVPYAYGFAIILLTIIVKAATYPLTKQQVESTLAMQNLQPKIKA Alb4 YMETILKVLKDGLSTVHVPYSYGFAIILLTVLVKAATFPLTKKQVESAMAMKSLTPQIKA ** :**:******:*****:*********::*****:****:****::**:.* *:*** Alb3 IQQRYAGNQERIQLETSRLYKQAGVNPLAGCLPTLATIPVWIGLYQALSNVANEGLFTEG Alb4 IQERYAGDQEKIQLETARLYKLAGINPLAGCLPTLATIPVWIGLYRALSNVADEGLLTEG **:****:**:*****:**** **:********************:******:***:*** Alb3 FFWIPSLGGPTSIAARQSGSGISWLFPFVDGHPPLGWYDTVAYLVLPVLLIASQYVSMEI Alb4 FFWIPSLAGPTTVAARQNGSGISWLFPFIEGHPPLGWPDTLAYLVLPLLLVFSQYLSIQI *******.***::****.**********::******* **:******:**: ***:*::* Alb3 MKPPQTDDPAQKNTLLVFKFLPLMIGYFALSVPSGLSIYWLTNNVLSTAQQVYLRKLGGA Alb4 MQSSQSNDPAMKSSQAVTKLLPLMIGYFALSVPSGLSLYWLTNNILSTAQQVWLQKYGGA *:..*::*** *.: * *:*****************:******:*******:*:* *** Alb3 KPNMDENASKIISAG---------------------------RAKRSIAQPDDAGERFRQ Alb4 KNPVEKFTNLVTKEDKTQQIEKSFSEPLVQKSVSELKIPREKGGEKVTPECPKPGERFRL * ::: :. : . . .:: .: ..***** Alb3 LKEQEKRSKKNK--------AVAKDTVELVEESQSESEEGSDDEEEEAREGALASSTTSK Alb4 LKEQEAKRRREKEERQKAEAALSNQNTDKAHEQDEKSDTAIVAEDDKKTELSAVDETSDG ***** : :::* *::::..: ..*.:.:*: . *::: * : ...*:. Alb3 PLPEVGQRRSKRSKRKRTV------------------ (462 AS mit TP) Alb4 TVAVNGKPSIQKDETTNGTFGIGHDTEQQHSHETEKR (499 AS mit TP) .:. *: ::.: .. .
Abbildung 4-1: Alignment der Aminosäuresequenz von Alb3 und Alb4 Das Alignment zeigt einen Vergleich der Aminosäuresequenzen von Alb3 und Alb4. Dargestellt
sind die mittels ChloroP (http://www.cbs.dtu.dk/services/ChloroP/) vorhergesagten Transitpeptide
(TP) für das chloroplastidäre Targeting der Proteine (grün) sowie der maximal vorhergesagte
Bereich der fünf Transmembrandomänen (Aramemnon, http://aramemnon.botanik.uni-koeln.de/)
(grau hinterlegt). Die in blau gezeigten Sequenzen von Alb3 umfassen die Regionen der Alb3
Antigene nAlb3 (AS 56-108) und cAlb3 (AS 397-462), die zum Zweck der Antikörperherstellung
rekombinant überexprimiert wurden. Die in rot dargestellte Sequenz des Alb4 C-Terminus
beinhaltet den Bereich des Alb4 Antigens (AS 347-494).
4.1.1 Expression der Antigene GST-nAlb3, GST-cAlb3 und cAlb4-his
Für die Herstellung der Antigene von Alb3 wurden die entsprechenden Gensequenzen
des N-terminalen und C-terminalen Bereichs von Alb3 in den Expressionsvektor
pGEX4T3 kloniert und in den Überexpressionsstamm E. coli BL21(DE3) transformiert. Die
Antigene nAlb3 und cAlb3 wurden über diesen Vektor als GST-Fusionsproteine
exprimiert, da die errechnete Größe der Antigene mit 6 kDa (nAlb3) und 7 kDa (cAlb3) für
eine Immunisierung in Kaninchen nicht ausreichend war. Die Aufreinigung der Antigene
von Alb3 erfolgte über den angefügten GST-tag mittels Glutathion-Sepharose. Da für die
vorliegende Arbeit letztendlich lediglich der Antikörper gegen den C-Terminus von Alb3

4 Ergebnisse 65
eingesetzt wurde, ist die Funktionalität des Antikörpers gegen den N-Terminus nicht
abschließend getestet worden. Die Aufreinigung des Antigens GST-cAlb3 wurde über
eine SDS-PAGE mit anschließender Coomassie Färbung überprüft (Abbildung 4-2). Die
errechnete Größe des GST-Fusionsproteins beträgt 33 kDa und stimmt mit dem
tatsächlichen Laufverhalten bei 37 kDa gut überein. Eine weitere Proteinbande auf Höhe
von 32 kDa lässt auf ein Abbauprodukt des Proteins schließen. Das Problem des
Proteinabbaus bei rekombinant exprimiertem Alb3 Protein war bereits aus vorherigen
Versuchen im Labor bekannt. Die Gensequenz des Antigens von Alb4 (cAlb4) lag bereits
in dem Expressionsvektor pET29b in dem E. coli Überexpressionsstamm BL21(DE3) vor
(Gerdes et al., 2006). Nach der Überexpression erfolgte die Aufreinigung des Proteins
unter denaturierenden Bedingungen über den angefügten C-terminalen His-tag mittels Ni-
NTA Agarose. Anhand der Expressionskontrolle über SDS-PAGE mit anschließender
Coomassie Färbung zeigte sich ein zu der errechneten Größe des Proteins von 19 kDa
abweichendes Laufverhalten, das bei 26 kDa lag und bereits in Gerdes et al., 2006
beschrieben wurde (Abbildung 4-2). Nach erfolgter Aufreinigung wurden die Antigene
gegen PBS dialysiert und je 800 µg lyophilisiertes Protein zur Standardimmunisierung von
Kaninchen und Ratte an die Firma SeqLab (Göttingen) verschickt.
4.1.2 Test der Antikörper α-Alb3 und α-Alb4 gegen die Antigene
Nach Erhalt der Antiseren α-Alb3 (aus Kaninchen) und α-Alb4 (aus Ratte) wurden diese
zunächst gegen die Antigene getestet. Zu diesem Zweck wurde der GST-tag des cAlb3
Antigens durch Thrombin abgespalten und die erfolgreiche Abspaltung über eine SDS-
PAGE mit anschließender Coomassie Färbung überprüft (Abbildung 4-2). Nach dem
Thrombinverdau ist sowohl das 26 kDa große GST-Protein, als auch eine unterhalb von
17 kDa laufende Bande des abgespaltenen cAlb3 zu erkennen. Für den Antikörpertest
wurden das GST-cAlb3 Fusionsprotein, das gespaltene GST/cAlb3 Proteingemisch und
das cAlb4-his Protein über eine SDS-PAGE aufgetrennt und eine Immundetektion mit den
beiden Seren α-Alb3 (aus Kaninchen) und α-Alb4 (aus Ratte) durchgeführt. Mit dem
α-Alb3 Antikörper konnten das GST-cAlb3 Fusionsprotein und die durch Thrombin-
spaltung erhaltenen GST und cAlb3 Proteine detektiert werden. In der Spur, in der das
cAlb4 Antigen aufgetragen worden war, konnte kein Signal detektiert werden. Der Test
des α-Alb4 Antikörpers (aus Ratte) ergab, dass dieser das eigene cAlb4 Antigen und nicht
das GST-cAlb3 Fusionsprotein und die gespaltenen Proteine GST und cAlb3 detektierte.
Die Western Blots, die mit den Präimmunseren der beiden Antikörper behandelt wurden,
zeigten keine Signale. Diese Versuche belegen die Spezifität des α-Alb4 und des α-Alb3
Antikörpers im Hinblick auf die Detektion der verwendeten Antigene.
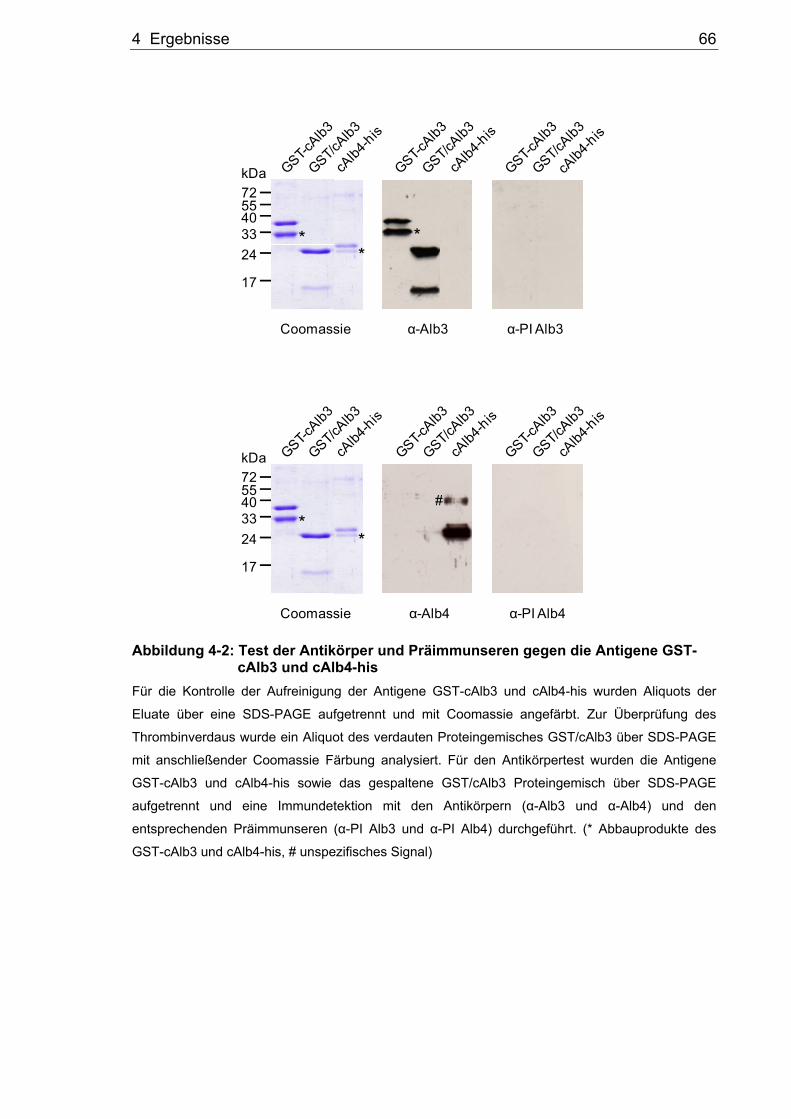
4 Ergebnisse 66
7255403324
17
kDa
α-Alb3 α-PI Alb3
α-PI Alb4α-Alb4
Coomassie
7255403324
17
kDa
Coomassie
**
*
*
*#
Abbildung 4-2: Test der Antikörper und Präimmunseren gegen die Antigene GST-cAlb3 und cAlb4-his
Für die Kontrolle der Aufreinigung der Antigene GST-cAlb3 und cAlb4-his wurden Aliquots der
Eluate über eine SDS-PAGE aufgetrennt und mit Coomassie angefärbt. Zur Überprüfung des
Thrombinverdaus wurde ein Aliquot des verdauten Proteingemisches GST/cAlb3 über SDS-PAGE
mit anschließender Coomassie Färbung analysiert. Für den Antikörpertest wurden die Antigene
GST-cAlb3 und cAlb4-his sowie das gespaltene GST/cAlb3 Proteingemisch über SDS-PAGE
aufgetrennt und eine Immundetektion mit den Antikörpern (α-Alb3 und α-Alb4) und den
entsprechenden Präimmunseren (α-PI Alb3 und α-PI Alb4) durchgeführt. (* Abbauprodukte des
GST-cAlb3 und cAlb4-his, # unspezifisches Signal)

4 Ergebnisse 67
4.1.3 Test der Antikörper α-Alb3 und α-Alb4 gegen die in vitro translatierten Volllängenproteine
Die Spezifität der α-Alb3 und α-Alb4 Antikörper sollte über den Test gegen die
Volllängenproteine in Form der in vitro Translationsprodukte des Alb3 und Alb4 bestätigt
werden (Abbildung 4-3). Dazu wurde das Alb3 und das Alb4 (mit Transitpeptid, pAlb4)
über die SP6-Polymerase in vitro transkribiert und anschließend mit Hilfe eines
Weizenkeimlysats in vitro translatiert. Die Translationskontrolle erfolgte über den Einsatz
von radioaktiv markiertem [35S]-Methionin, durch dessen Einbau die Proteine nach
Auftrennung mittels einer SDS-PAGE und anschließender Autoradiographie detektiert
werden konnten. Die Autoradiographie erbrachte in dem Translationsansatz des Alb3 ein
spezifisches Signal bei etwa 49 kDa und bei dem Ansatz des pAlb4 ein spezifisches
Signal bei etwa 60 kDa. Die Molekulargewichte dieser Signale entsprachen daher in etwa
den theoretischen Molekulargewichten von Alb3 (45 kDa) und dem pAlb4 inklusive des
Transitpeptids (55 kDa).
α-Alb3 α-Alb4
kDa
pAlb4
Alb3
72
55
43
Translations-kontrolle
Abbildung 4-3: Test der Antikörper gegen die in vitro translatierten Volllängenproteine
Für die Translationskontrolle wurden die radioaktiv markierten Translationsprodukte von Alb3 und
pAlb4 (TP [35S]-Alb3 und TP[35S]-pAlb4, 1 µl) über eine SDS-PAGE aufgetrennt und mittels
Autoradiographie nachgewiesen. Des Weiteren wurde die Spezifität der Antikörper α-Alb3 und
α-Alb4 mit nicht radioaktiv markierten Translationsprodukten (TP Alb3 und TP pAlb4, 5 µl) über
eine Immundetektion gezeigt.
Für den Antikörpertest wurden die Translationsansätze, in denen die Proteine nicht
radioaktiv markiert waren, über eine SDS-PAGE aufgetrennt und anschließend Western
Blot Analysen durchgeführt. Durch den Einsatz des α-Alb3 Antikörpers wurde das in vitro
translatierte Alb3 auf der Höhe von 49 kDa erkannt. Die Spezifität des α-Alb3 Antikörpers

4 Ergebnisse 68
wurde dadurch bestätigt, dass dieser Antikörper nicht das in vitro translatierte pAlb4
Volllängenprotein detektieren konnte. Zudem zeigte sich, dass der α-Alb4 Antikörper
ausschließlich das in vitro translatierte pAlb4 Volllängenprotein auf der Höhe von 60 kDa
und nicht das Translationsprodukt von Alb3 detektierte. Damit war es gelungen, die
Spezifität des α-Alb3 und α-Alb4 Antikörpers auch für das entsprechende
Volllängenprotein zu bestätigen.
4.1.4 Test der Antikörper α-Alb3 und α-Alb4 mit Thylakoidmembran-proteinen
Für den folgenden Antikörpertest wurden Thylakoidmembranproteine von Arabidopsis
thaliana über eine SDS-PAGE aufgetrennt und Western Blot Analysen durchgeführt
(Abbildung 4-4). Dabei wurde das Alb3 Protein durch den α-Alb3 Antikörper auf der Höhe
von 45 kDa nachgewiesen, während das auf der Höhe von 50 kDa zu erwartende Alb4
nicht detektiert wurde. Durch den Einsatz des α-Alb4 Antikörpers wurde ausschließlich
das Alb4 Protein auf der Höhe von 50 kDa erkannt. Die Western Blots, die mit den
Präimmunseren der beiden Antikörper behandelt wurden, zeigten keine Signale.
Insgesamt lässt sich festhalten, dass die in den Abschnitten 4.1.2 bis 4.1.4 beschriebenen
Versuche eindeutig die Spezifität der beiden Antikörper gegen Alb3 und Alb4 belegen.
Damit war eine wichtige Voraussetzung für weitere biochemische Analysen von Alb3 und
Alb4 erfüllt.
α-Alb4
kDaTM TMTM TM
α-PI Alb4α-PI Alb3α-Alb3
72
55
43
95
Abbildung 4-4: Test der Antikörper gegen Thylakoidmembranproteine Zur Überprüfung der Spezifität der Antikörper wurden Thylakoidmembranproteine (äquivalent zu
5 µg Chlorophyll) über eine SDS-PAGE aufgetrennt und ein immunologischer Nachweis mit den
Antikörpern α-Alb3 und α-Alb4 durchgeführt. Weitere Western Blot Analysen dienten der Kontrolle
auf Reaktion der Präimmunseren (α-PI Alb3 und α-PI Alb4) mit den Thylakoidmembranproteinen.

4 Ergebnisse 69
4.2 Analyse der Interaktionen von Alb3 und Alb4 mit den Proteinen des cpSRP-Transportwegs
Bisherige Studien zeigten, dass der Transport der Lichtsammelkomplexproteine (LHCP)
durch das Stroma zur Thylakoidmembran über den cpSRP-Transportweg erfolgt. Des
Weiteren stellt das integrale Membranprotein Alb3 die Translokase für die Integration der
LHCPs in die Membran dar. Ein entscheidender Aspekt innerhalb dieses Transportwegs,
der sich auf den docking-Prozess des Transitkomplexes an die Thylakoidmembran
bezieht, ist allerdings bisher noch ungeklärt. In dem folgenden Teil dieser Arbeit sollte
geklärt werden, ob der docking-Prozess des Transitkomplexes möglicherweise durch eine
direkte Interaktion der Proteine des cpSRP-Wegs mit der Translokase Alb3 vermittelt wird.
Außerdem sollte über die Proteininteraktionsstudien untersucht werden, ob das zur
Translokase Alb3 homologe Protein Alb4 eine Funktion innerhalb des cpSRP-
Transportwegs ausübt.
4.2.1 Interaktionen von Alb3 und Alb4 mit den Proteinen des cpSRP-Transportwegs im Hefe Split-Ubiquitin System
Um die Interaktion der beiden Membranproteine Alb3 und Alb4 mit den Proteinen des
cpSRP-Wegs cpSRP43, cpSRP54 und cpFtsY zu analysieren, wurde zunächst das Hefe
Split-Ubiquitin System eingesetzt. Das Split-Ubiquitin System ist ein in vivo Verfahren, bei
dem die Interaktion von Membranproteinen getestet werden kann. Dabei ist das Ubiquitin
in ein N-terminales Nub-Fragment und ein C-terminales Cub-Fragment gespalten. Das
N-terminale Fragment des Ubiquitins (NubI) besitzt eine hohe Affinität zum C-terminalen
Bereich des Ubiquitins (Cub). Bei räumlicher Nähe eines Fusionsproteins, bestehend aus
dem Cub-Fragment und dem zu analysierenden Protein (Alb3 bzw. Alb4), und dem NubI-
Fragment des Kontrollproteins Alg5-NubI kommt es zur Rekonstitution des Ubiquitins und
damit zur Abspaltung des Reporters. Dieser Reporter ist an das Cub-Protein angefügt und
besteht aus einer DNA-bindenden Domäne (LexA-Protein) und einer
transkriptionsaktivierenden Domäne (Virionenprotein 16). Die Lokalisation des Reporters
im Cytosol kann durch eine erfolgte Aktivierung der Reportergene nachgewiesen werden.
Zur Kontrolle auf Transaktivierung diente das Protein Alg5-NubG, das aufgrund einer
Punktmutation im Nub-Fragment (NubG) die Affinität zwischen den Ubiquitin-Hälften
verringert. Mit Hilfe des Split-Ubiquitin Systems wurden bereits erfolgreiche
Untersuchungen des Alb3 mit potentiellen Bindungspartnern durchgeführt (Pasch et al.,
2005). Das Cub-Protein wird mit dem angefügten Reporter von dem pAMBV4-Vektor und
das NubG-Protein durch den Vektor pADSL-Nx kodiert. Für den Interaktionstest wurden
die Gensequenzen von Alb3 und Alb4 in den pAMBV4-Vektor und die Gensequenzen von

4 Ergebnisse 70
cpSRP43, cpSRP54 und cpFtsY in den pADSL-Vektor kloniert. Die Kontrolle der
Cotransformation der bait- und prey-Plasmide in die Hefezellen verlief in allen Ansätzen
positiv. Die Transformationsraten konnten durch das Wachstum der Hefekolonien auf dem
–LT-Medium, dem die Aminosäuren Leucin und Tryptophan fehlten, erkannt werden
(Daten nicht gezeigt). Der Einsatz der Konstrukte pAMBV4-Alb3 bzw. pAMBV4-Alb4 mit
dem Kontrollplasmid (kodiert für Alg5-NubI) wies durch ein deutliches Wachstum der
cotransformierten Hefezellen auf dem –LTH-Medium auf eine erfolgreiche Rekonstitution
des Ubiquitins hin. Zur Kontrolle auf Transaktivierung des Alb3 und Alb4 wurden die
Plasmide pAMBV4-Alb3 bzw. pAMBV4-Alb4 mit dem Kontrollplasmid (kodiert für Alg5-
NubG) cotransformiert. Ein Wachstum der transformierten Hefezellen auf –LTH-Medium
wies auf eine schwache Eigenaktivierung hin (Daten nicht gezeigt). Der Zusatz von
15 mM 3-Aminotriazol (3-AT), einem kompetitiven Inhibitor des His3-Genproduktes,
verringerte die Eigenaktivierung jedoch so weit, dass kein Wachstum beim Einsatz des
Kontrollplasmids auf –LTH-Medium mehr möglich war.
Für die Interaktionsanalysen (Tabelle 4-1) wurden die Konstrukte pAMBV4-Alb3 bzw.
pAMBV4-Alb4 sowie die Kontrolle pMBV4-Alg5 mit den Konstrukten der potentiellen
Bindungspartner (pADSL-cpSRP43, pADSL-cpSRP54 und pADSL-cpFtsY) in die
Hefezellen cotransformiert. Interessanterweise deuteten die Ergebnisse auf eine
Interaktion des Alb3 mit cpSRP43, während eine Interaktion mit cpSRP54 oder cpFtsY im
Split-Ubiquitin System nicht nachgewiesen wurde. Um auszuschließen, dass die
negativen Ergebnisse auf eine fehlende Expression des cpSRP54 oder cpFtsY
zurückzuführen sind, wurde die korrekte Expression der Proteine in den Hefezellen
nachgewiesen (Daten nicht gezeigt). Das Alb4 zeigte mit keinem der getesteten Proteine
eine Interaktion im Hefe Split-Ubiquitin System. Von den Hefekolonien, die auf dem –LTH-
Medium gewachsen sind, wurde ein β-Galaktosidase Filtertest durchgeführt. Bei allen
Transformationen, bei denen ein Wachstum der Hefezellen auf diesem Selektionsmedium
zu verzeichnen war, konnte auch eine Blaufärbung im Filtertest gezeigt werden (Tabelle
4-1). Diese Ergebnisse deuten darauf hin, dass das cpSRP43 in der Lage ist direkt mit
Alb3 zu interagieren und dass diese Interaktion möglicherweise den docking-Prozess des
Transitkomplexes an die Thylakoidmembran über eine Interaktion mit der Translokase
Alb3 vermittelt.

4 Ergebnisse 71
Tabelle 4-1: Alb3 interagiert mit cpSRP43 im Split-Ubiquitin System Dargestellt sind die Interaktionstests von Alb3 und Alb4 mit den löslichen Proteinen des cpSRP-
Wegs im Hefe Split-Ubiquitin System. Der Hefe Stamm DSY-1 wurde mit den Plasmiden pAMBV4-
Alb3, pAMBV4-Alb4, pMBV4-Alg5 und den angegebenen NubG-Fusionsproteinen des cpSRP-
Wegs cotransformiert und die Aktivierung der Reportergene ausgewertet. Durch das Wachstum
der cotransformierten Hefezellen auf Minimalmedium (-his) (++ = deutliches Wachstum,
+ = geringes Wachstum, - = kein Wachstum) und über die Bestimmung der Enzymaktivität der
β-Galaktosidase (β-gal), die durch eine Blaufärbung der Hefekolonien im β-Galaktosidase Filtertest
getestet wurde (+ = Blaufärbung, - = keine Blaufärbung), konnten Aussagen über die Interaktion
der Proteine gemacht werden. Die Plasmide pADSL-Alg5-NubI und pADSL-Alg5-NubG wurden als
Positiv- bzw. Negativkontrollen eingesetzt.
pAMBV4-Alb3 pAMBV4-Alb4 pMBV4-Alg5
pADSL-Nx -his β-gal. -his β-gal. -his β-gal
Alg5-NubI ++ + ++ + + +
Alg5-NubG - - - - - -
cpSRP43 ++ + - - - -
cpSRP54 - - - - - -
cpFtsY - - - - - -
4.2.2 Proteininteraktionen von Alb3 und Alb4 mit den Proteinen des cpSRP-Transportwegs im Split-YFP System
Um die gezeigte in vivo Interaktion von Alb3 mit cpSRP43 zu bestätigen, sollten die im
Split-Ubiquitin System getesteten Proteine im Split-YFP System in Arabidopsis thaliana
Protoplasten eingesetzt werden. Dieses ist ein weiteres in vivo Verfahren, um mit Hilfe
der bimolekularen Fluoreszenzkomplementation die Interaktion von Proteinen in ihrer
natürlichen Umgebung zu testen. Dazu ist das YFP in zwei Hälften gespalten. Das N-
terminale nYFP wird durch den Vektor pSPYNE (split YFP N-terminal fragment
expression) und das C-terminale cYFP durch den Vektor pSPYCE (split YFP
C-terminal fragment expression) kodiert. Für die Interaktionstests wurden die
Membranproteine Alb3 und Alb4 N-terminal an die YFP-Fragmente fusioniert, so dass die
YFP-Hälften nach der Integration der Proteine in die Thylakoidmembran auf der Stroma
gelegenen Seite lokalisiert sind. Damit war gewährleistet, dass die YFP-Fragmente der
Membranproteine mit den entsprechenden YFP-Hälften der möglichen stromalen
Interaktionspartner cpSRP43, cpSRP54 und cpFtsY in räumliche Nähe geraten konnten.
Der Transport der Proteine (Alb3, Alb4 und cpFtsY) aus dem Cytosol in den Chloroplasten

4 Ergebnisse 72
wurde über die jeweilige chloroplastidäre Transitsequenz der Proteine vermittelt. Lagen
die Gensequenzen der Proteine ohne eigene Transitsequenz vor (cpSRP43 und
cpSRP54), wurde die Transitsequenz der kleinen Untereinheit der Rubisco (TS-rbcs) vor
das jeweilige Konstrukt kloniert, um den chloroplastidären Import zu erreichen. Für die
Analyse der Interaktion wurde die Gensequenz des einen Interaktionspartners in den
pSPYCE-Vektor und die des anderen Partners in den pSPYNE-Vektor kloniert und diese
Konstrukte in Arabidopsis thaliana Protoplasten cotransformiert. Bei einer Interaktion
zweier YFP-Fusionsproteine rekonstituieren sich die beiden nicht-fluoreszierenden YFP-
Hälften zu dem fluoreszierenden YFP, dessen Fluoreszenz an einem Laserscanning
Mikroskop (LSM 510 Meta) visualisiert und dokumentiert werden konnte.
Zunächst sollte getestet werden, ob die bereits bekannte Interaktion von cpSRP43 und
cpSRP54 mit diesem System gemessen werden kann. In Abbildung 4-5 ist gezeigt, dass
cpSRP43-nYFP mit dem cpSRP54-cYFP in Arabidopsis thaliana Protoplasten
interagierte, wodurch die Fluoreszenz des YFP rekonstituiert werden konnte. Die
Überlagerung der YFP-Fluoreszenz mit der Autofluoreszenz des Chlorophylls zeigte
zudem die Interaktion der beiden Proteine innerhalb der Chloroplasten. Es zeigte sich
ebenfalls eine Interaktion von Alb3-cYFP mit cpSRP43-nYFP und Alb3-nYFP mit
cpSRP43-cYFP in vivo in Arabidopsis Protoplasten, wodurch das Ergebnis der Interaktion
der beiden Proteine im Hefe Split-Ubiquitin System bestätigt werden konnte (Abbildung
4-5, Tabelle 4-3). Als Kontrolle wurden die Vektoren pSPYNE-TS(rbcs) und pSPYCE-
TS(rbcs) eingesetzt und mit den einzelnen Fusionskonstrukten cotransformiert. Bei diesen
Konstrukten ist die Transitsequenz der kleinen Untereinheit der Rubisco (TS-rbcs) vor das
jeweilige YFP-Fragment fusioniert, wodurch der Import der einzelnen YFP-Hälften ohne
weiteres Fusionsprotein in die Chloroplasten garantiert ist. Die Kontrollen zeigten, dass
weder das Fusionsprotein Alb3-cYFP noch das cpSRP43-nYFP in der Lage waren die
YFP-Fluoreszenz mit dem jeweiligen nYFP- oder cYFP-Fragment zu rekonstituieren
(Abbildung 4-5, Tabelle 4-2). Diese Ergebnisse deuteten auf eine direkte Interaktion
zwischen Alb3 und cpSRP43, da eine unspezifische und zufällige Rekonstitution des
YFPs auszuschließen war.
Des Weiteren sollten die Proteine cpSRP54 und cpFtsY auf Interaktion mit Alb3 getestet
werden. Es konnte keine YFP-Fluoreszenz und damit keine Interaktion von Alb3 mit
cpSRP54 oder cpFtsY im Split-YFP System erkannt werden. Zudem zeigte sich, dass
Alb4 im Vergleich zu Alb3 nicht in der Lage war mit cpSRP43 zu interagieren (Tabelle
4-2). Alle Proteine wurden sowohl als Fusionsproteine mit der N-terminalen Hälfte, als
auch als Fusionsproteine mit der C-terminalen Hälfte des YFP getestet. Dazu lagen die
Gensequenzen für diese Proteine im pSPYCE- und pSPYNE-Vektor vor (Tabelle 4-2). In
allen Kontrollexperimenten konnte ebenfalls keine YFP-Rekonstitution beobachtet werden
(Tabelle 4-3).
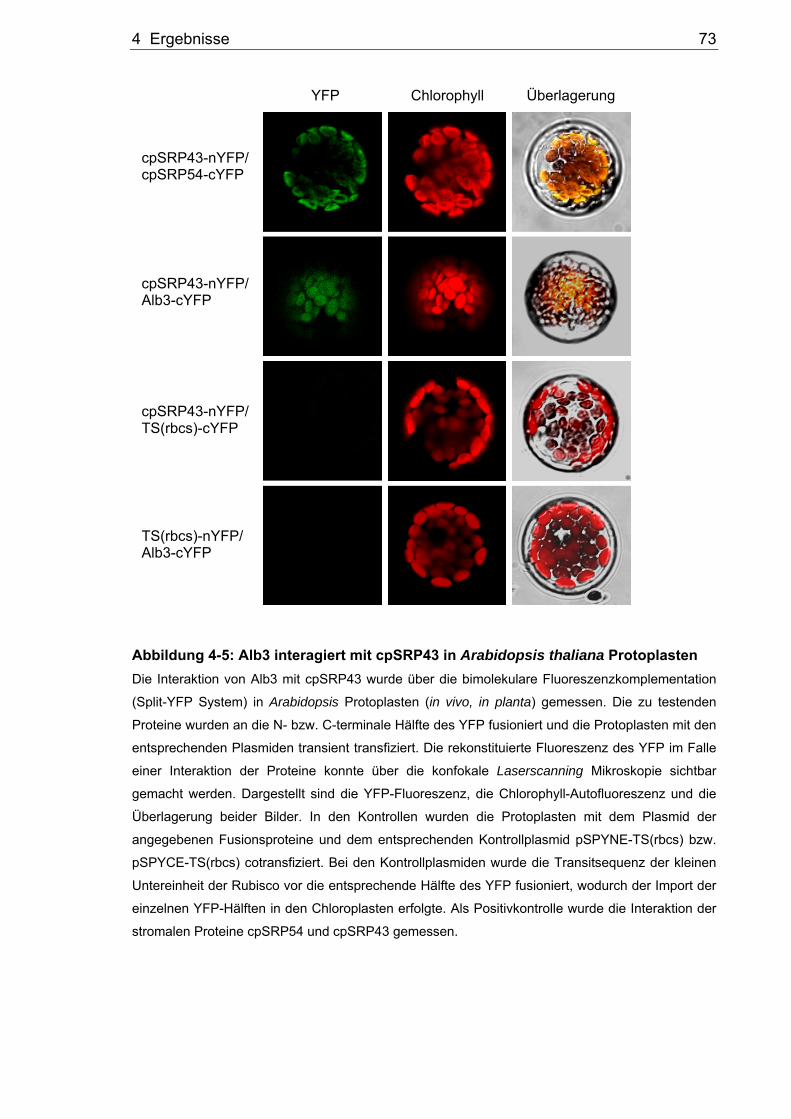
4 Ergebnisse 73
Abbildung 4-5: Alb3 interagiert mit cpSRP43 in Arabidopsis thaliana Protoplasten Die Interaktion von Alb3 mit cpSRP43 wurde über die bimolekulare Fluoreszenzkomplementation
(Split-YFP System) in Arabidopsis Protoplasten (in vivo, in planta) gemessen. Die zu testenden
Proteine wurden an die N- bzw. C-terminale Hälfte des YFP fusioniert und die Protoplasten mit den
entsprechenden Plasmiden transient transfiziert. Die rekonstituierte Fluoreszenz des YFP im Falle
einer Interaktion der Proteine konnte über die konfokale Laserscanning Mikroskopie sichtbar
gemacht werden. Dargestellt sind die YFP-Fluoreszenz, die Chlorophyll-Autofluoreszenz und die
Überlagerung beider Bilder. In den Kontrollen wurden die Protoplasten mit dem Plasmid der
angegebenen Fusionsproteine und dem entsprechenden Kontrollplasmid pSPYNE-TS(rbcs) bzw.
pSPYCE-TS(rbcs) cotransfiziert. Bei den Kontrollplasmiden wurde die Transitsequenz der kleinen
Untereinheit der Rubisco vor die entsprechende Hälfte des YFP fusioniert, wodurch der Import der
einzelnen YFP-Hälften in den Chloroplasten erfolgte. Als Positivkontrolle wurde die Interaktion der
stromalen Proteine cpSRP54 und cpSRP43 gemessen.
YFP Chlorophyll Überlagerung
cpSRP43-nYFP/ cpSRP54-cYFP
cpSRP43-nYFP/ Alb3-cYFP
cpSRP43-nYFP/ TS(rbcs)-cYFP
TS(rbcs)-nYFP/ Alb3-cYFP

4 Ergebnisse 74
Tabelle 4-2: Interaktionstests im Split-YFP System Dargestellt sind die Interaktionstests von Alb3 und Alb4 mit den löslichen Proteinen des cpSRP-
Wegs im Split-YFP System. Die zu testenden Proteine wurden sowohl an die N-terminale Hälfte,
als auch an die C-terminale Hälfte des YFP fusioniert. Die Protoplasten wurden mit den
entsprechenden Plasmiden in beiden Kombinationen transient transfiziert und die Interaktion der
Proteine analysiert.
nYFP-Konstrukte cYFP-Konstrukte Interaktion
cpSRP43 cpSRP54 + cpSRP54 cpSRP43 + cpSRP43 Alb3 + Alb3 cpSRP43 + cpSRP54 Alb3 - Alb3 cpSRP54 - cpFtsY Alb3 - Alb3 cpFtsY - cpSRP43 Alb4 - Alb4 cpSRP43 -
Tabelle 4-3: Negativkontrollen der Interaktionstests im Split-YFP System Gezeigt sind die im Rahmen des Split-YFP Systems durchgeführten Negativkontrollen. Dazu
wurden die Plasmide, die das zu testende Protein als Fusion mit dem nYFP oder cYFP kodieren,
mit dem jeweiligen Kontrollplasmid pSPYNE-TS(rbcs) bzw. pSPYCE-TS(rbcs) transient
cotransfiziert.
nYFP-Konstrukte cYFP-Konstrukte Interaktion
cpSRP43 TS-rbcs - cpSRP54 TS-rbcs - cpFtsY TS-rbcs - Alb3 TS-rbcs - Alb4 TS-rbcs - TS-rbcs cpSRP43 - TS-rbcs cpSRP54 - TS-rbcs cpFtsY - TS-rbcs Alb3 - TS-rbcs Alb4 -

4 Ergebnisse 75
Um die spezifische Interaktion von Alb3 und cpSRP43 weiter zu bestärken, sollte die
korrekte Lokalisation der Fusionsproteine Alb3-cYFP und Alb4-cYFP in der
Thylakoidmembran und des cpSRP43-nYFP in der löslichen Fraktion der Protoplasten
nachgewiesen werden. Dazu wurden die Protoplasten mit dem jeweiligen
Fusionskonstrukt transfiziert. Für den Nachweis der Expression wurden die lysierten
Zellen abzentrifugiert und die Proteine des Überstands mit Trichloressigsäure (TCA)
gefällt. Das Waschen des Membranpellets mit 0,2 M NaOH sollte peripher gebundene
Proteine ablösen. Mit dem entstehenden Überstand (Waschfraktion) wurde ebenfalls eine
TCA-Fällung durchgeführt. Die Proteine des gewaschenen Membranpellets sowie die
Proben aus der TCA-Fällung wurden über eine SDS-PAGE aufgetrennt und mittels
Immundetektion ausgewertet (Abbildung 4-6). Dabei konnten die nYFP-Fusionsproteine
über den angefügten c-myc-tag und die cYFP-Fusionsproteine über den HA-tag
nachgewiesen werden.
Alb3-cYFP
cpSRP43-nYFP
Kontrolle
P
α-HA
α-c-myc
Ü W
P Ü W
Alb4-cYFP
Kontrolle
Abbildung 4-6: Expressionsnachweis der YFP-Fusionsproteine cpSRP43-nYFP,
Alb3-cYFP und Alb4-cYFP Arabidopsis Protoplasten wurden mit den entsprechenden Konstrukten transient transfiziert und die
Expression der YFP-Fusionsproteine (cpSRP43-nYFP, Alb3-cYFP und Alb4-cYFP) über
Immundetektion nachgewiesen. Dazu wurden die lysierten Zellen abzentrifugiert und der
Überstand (Ü) mittels TCA-Fällung präzipitiert. Das Membranpellet (P) wurde mit 0,2 M NaOH
gewaschen, zentrifugiert und der Überstand (W=Waschfraktion) mittels TCA-Fällung präzipitiert.
Die Proben des gewaschenen Membranpellets (P), des Überstands (Ü) und der Waschfraktion (W)
wurden über SDS-PAGE aufgetrennt und mittels Immundetektion analysiert. Die Expression
konnte über einen anti-HA Antikörper (α-HA) für die cYFP-Fusionsproteine und einen anti-c-myc
Antikörper (α-c-myc) für das nYFP-Fusionsprotein gezeigt werden. Als Kontrolle wurden nicht
transfizierte Protoplasten eingesetzt (Kontrolle).

4 Ergebnisse 76
Es zeigte sich, dass das stromale cpSRP43-nYFP in der Fraktion des Überstands nach
der Lyse zu finden war. Eine geringe Menge befand sich auch in der Waschfraktion, was
darauf hindeutet, dass das cpSRP43-nYFP zum Teil noch an der Membran gebunden
vorlag. Die integralen Membranproteine Alb3 und Alb4 konnten als YFP-Fusionsproteine
ausschließlich in der Membranfraktion nachgewiesen werden. Als Kontrollen wurden
jeweils nicht transfizierte Protoplasten aufgearbeitet. In diesen Proben waren keine
Signale auf der entsprechenden Größe der YFP-Fusionsproteine zu detektieren
(Abbildung 4-6). Durch den Nachweis der Expression und der korrekten Lokalisation der
YFP-Fusionsproteine in der Membran bzw. im Stroma, konnte die spezifische Interaktion
von Alb3 und cpSRP43 in vivo in planta unterstützt werden.
4.2.3 Analyse der Binderegion zwischen Alb3 und cpSRP43 über das Split-YFP System
Um die Binderegion des Alb3 für cpSRP43 genauer zu untersuchen, wurden zunächst
verschiedene Konstrukte aus unterschiedlichen Bereichen von Alb3 und Alb4 über die
overlap-PCR hergestellt. Wie bereits in Abschnitt 4.2.2 beschrieben, interagierte Alb4
nicht mit cpSRP43. Die Erstellung von solchen chimären Proteinen bot sich an, da beide
Proteine fünf putative Transmembrandomänen besitzen (Abbildung 4-7).
462
499
133-155 203-225 236-258 272-294 315-337
130-159 201-226 233-267 271-299 311-340
119-141 183-205 216-236 260-282 300-322
114-143 183-210 216-237 257-286 297-324
56
46
Alb3
Alb4
Abbildung 4-7: Schematische Darstellung der vorhergesagten Membrantopologie von Alb3 und Alb4
Die Membrantopologie von Alb3 und Alb4 zeigt eine Lokalisation des N-Terminus im Lumen und
des C-Terminus im Stroma. Mit Hilfe des Internetprogrammes Aramemnon
(http://aramemnon.botanik.uni-koeln.de/) wurden die Bereiche der vorhergesagten
Transmembrandomänen bestimmt (Rechtecke). Die oberhalb der Transmembrandomänen
dargestellten Nummern der Aminosäuren zeigen die gemeinsame Vorhersage (Consensus) aller
Vorhersageprogramme, während die unterhalb dargestellten Nummern die maximale Länge der
Transmembrandomänen in allen Vorhersageprogrammen anzeigen.

4 Ergebnisse 77
Der Austausch der entsprechenden Bereiche von Alb3 durch Alb4 ermöglichte daher die
Aufrechterhaltung der Membrantopologie. Dieses ist ein deutlicher Vorteil gegenüber der
Erstellung von Deletionskonstrukten, denen große Teile des Proteins fehlen und die somit
möglicherweise keine korrekte Membrantopologie zur Folge hätten.
Für die Messung im Split-YFP System wurden verschiedene chimäre Proteine erzeugt,
wobei das Alb3 Protein vom N-Terminus ausgehend durch entsprechende Bereiche von
Alb4 ersetzt wurde. Die Proteine Alb4(1-142)Alb3(159-462), Alb4(1-210)Alb3(227-462),
Alb4(1-239)Alb3(256-462) und Alb4(1-255)Alb3(272-462) bestanden aus entsprechenden
N-terminalen Bereichen einschließlich der 1., 2. und 3. Transmembrandomäne von Alb4
und des dazugehörigen C-terminalen Bereichs von Alb3. Diese cYFP-Fusionsproteine
waren alle noch in der Lage mit cpSRP43-nYFP zu interagieren (Daten nicht gezeigt.)
Auch die Proteine Alb4(1-288)Alb3(296-462) und Alb4(1-300)Alb3(314-462), welche bis
zum Ende der 4. Transmembrandomäne bzw. bis zu Beginn der 5. Transmembran-
domäne aus dem Alb4 Protein bestanden, interagierten ebenfalls noch mit dem cpSRP43
und konnten somit die YFP-Fluoreszenz rekonstituieren (Abbildung 4-8). Dagegen war
das Fusionsprotein Alb4(1-326)Alb3(343-462), welches nur noch den C-Terminus von
Alb3 und den Rest des Proteins von der Alb4 Proteinsequenz aufwies, nicht mehr in der
Lage mit dem cpSRP43 zu interagieren. Dieses Ergebnis wies darauf hin, dass die
5. Transmembrandomäne von Alb3 für die Bindung des cpSRP43 erforderlich ist.
Weiterhin zeigte sich, dass eine bestimmte Region innerhalb der 5. Transmembran-
domäne von Alb3 für die cpSRP43 Bindung wichtig ist. Das Protein Alb4(1-300)Alb3(314-
462), welches noch die gesamte 5. Transmembrandomäne und den C-Terminus von Alb3
aufwies, konnte das cpSRP43 noch binden (Abbildung 4-8). Das Protein Alb4(1-
302)Alb3(319-462), bei dem nur wenige Aminosäuren der 5. Transmembrandomäne von
Alb3 durch die Alb4 Sequenz ersetzt wurden, zeigte dagegen keine Interaktion mehr
(Abbildung 4-8).

4 Ergebnisse 78
N
C
Alb4(1-288)Alb3(296-462)
N
C
Alb4(1-300)Alb3(314-462)
Alb4(1-326)Alb3(343-462)
N
C
Alb3(1-342)Alb4(327-499)
N
C
YFP Chlorophyll Überlagerung
Alb4(1-302)Alb3(319-462)
N
C
Abbildung 4-8: Identifizierung der Alb3 Bindestelle für das cpSRP43 in Arabidopsis
Protoplasten Für die Bestimmung der interagierenden Bereiche von Alb3 mit cpSRP43 wurden chimäre Proteine
aus unterschiedlich großen Bereichen von Alb3 und Alb4 eingesetzt, die an die
C-terminale Hälfte des YFPs fusioniert waren. Die Anteile des jeweiligen Proteins sind anhand des
Schemas und der Aminosäuren angegebenen. Arabidopsis Protoplasten wurden mit den
entsprechenden Plasmiden, bei denen die Gensequenzen der chimären Proteine an das cYFP und
die des cpSRP43 an das nYFP fusioniert waren, transient transfiziert und die YFP-Fluoreszenz
analysiert. Dargestellt sind die YFP-Fluoreszenz, die Chlorophyll-Autofluoreszenz und die
Überlagerung beider Bilder.

4 Ergebnisse 79
Ein Alignment dieser beiden Proteinsequenzen gibt Aufschluss auf die entscheidenden
Aminosäuren (Abbildung 4-9). Es wird deutlich, dass die Aminosäuren 314-318 (LVFKF)
von Alb3 den Unterschied zwischen diesen beiden Proteinen ausmachen und damit eine
Bindestelle für das cpSRP43 darstellen könnten. Bemerkenswerterweise liegt diese
putative Bindestelle auf der lumenalen Seite der Thylakoidmembran, was die Frage nach
der Zugänglichkeit dieser Bindestelle für das cpSRP43 eröffnet (siehe Diskussion). Um zu
testen, ob diese Bindestelle hinreichend für die Interaktion mit cpSRP43 ist, wurde das
Konstrukt Alb3(1-342)Alb4(327-499) erstellt. Das Protein Alb3(1-342)Alb4(327-499)
beinhaltet die Alb3 Sequenz bis einschließlich der 5. Transmembrandomäne. Lediglich
der Bereich des stromalen C-Terminus des Proteins ist von Alb4. Dieses chimäre Protein
konnte das cpSRP43 nicht binden, da keine YFP-Fluoreszenz im Laserscanning
Mikroskop zu detektieren war (Abbildung 4-8). Das bedeutet, dass die Bindestelle in der
5. Transmembrandomäne für die Interaktion nicht ausreichend war. Der C-Terminus von
Alb3 hat somit ebenfalls eine essentielle Bedeutung für die stabile Interaktion mit
cpSRP43, da er nicht durch die Alb4 Sequenz ersetzt werden konnte. Zusammenfassend
lässt sich sagen, dass der C-Terminus und die Aminosäuren 314-318 innerhalb der
5. Transmembrandomäne eine entscheidende Rolle für die Bindung des cpSRP43
besitzen.
Alb3 MKPPQTDDPAQKNTL---LVFKFLPLMIGYFALSVPSGLSIYWLTNNVLSAlb4(1-300)Alb3(314-462) MQSSQSNDPAMKSSQAVTLVFKFLPLMIGYFALSVPSGLSIYWLTNNVLSAlb4(1-302)Alb3(319-462) MQSSQSNDPAMKSSQAVT---KLLPLMIGYFALSVPSGLSIYWLTNNVLS *:..*::*** *.: *:***************************
Abbildung 4-9: Alignment der YFP-Fusionsproteine zur Identifizierung der Bindestelle
Das Alignment der Proteine Alb3, Alb4(1-300)Alb3(314-462) und Alb4(1-302)Alb3(319-462)
veranschaulicht die putative Bindestelle von Alb3 für das cpSRP43. Die Aminosäuren der Alb4
Sequenz sind in schwarz und die der Alb3 Sequenz in rot dargestellt. Der Verlust der Aminosäuren
314-318 (LVFKF) von der Alb3 Sequenz in dem Protein Alb4(1-302)Alb3(319-462) führt zum
Verlust der Interaktion mit cpSRP43.

4 Ergebnisse 80
4.2.4 Peptide Scanning von Alb3 mit rekombinantem cpSRP43
Da die 5. Transmembrandomäne und der C-Terminus von Alb3 für die Interaktion mit
cpSRP43 wichtig war und daher möglicherweise zwei Bindestellen vorhanden zu sein
schienen, wurde ein Peptide Scanning durchgeführt. Dazu wurde eine PepSpot Membran
bestellt, welche die Aminosäuren 299-462 und damit den 2. lumenalen loop, die
5. Transmembrandomäne und den C-Terminus von Alb3 umfasste. Auf der Membran
waren pro Spot 15 Aminosäuren lange Peptide verlinkt, wobei die Proteinsequenz jeweils
um 3 Aminosäuren versetzt war. Für die Analyse der Binderegionen wurde zunächst die
Gensequenz des cpSRP43 in den pET-Duet1 Vektor kloniert und in den E. coli
Überexpressionsstamm Rosetta (DE3) transformiert. Das überexprimierte, rekombinante
cpSRP43 wurde über den angefügten N-terminalen His-tag aufgereinigt (Abbildung 4-10
A). Die PepSpot Membran wurde mit dem rekombinanten Protein inkubiert, dessen
Bindestelle(n) auf der Membran und somit innerhalb der Alb3 Sequenz über eine
Immundetektion analysiert werden konnte. Es zeigte sich, dass das rekombinante His-
cpSRP43 an die Aminosäuren 305-328 innerhalb der 5. Transmembrandomäne und die
Aminosäuren 374-388 im C-Terminus binden konnte (Abbildung 4-10 B,C). Die
Bindestelle innerhalb der Transmembrandomäne umfasst vier PepSpots auf der Membran
(Abbildung 4-10 C). Vergleicht man die vier Peptidsequenzen dieser spots, fällt eine
gemeinsame Sequenz der Aminosäuren 314-319 (LVFKFL) auf, die in jedem dieser
15 Aminosäuren langen Peptide präsent ist (Abbildung 4-10 D). Diese Bindestelle enthält
die bereits durch das Split-YFP System gezeigte Bindestelle der Aminosäuren 314-318
(LVFKF). Das Split-YFP System ergab außerdem, dass es noch eine mögliche
Bindestelle innerhalb des C-Terminus von Alb3 geben musste. Die Pepspot Analyse
zeigte diese Bindestelle als einen Spot, der die Aminosäuren 374-388 umfasste
(Abbildung 4-10 C). Demnach unterstützt das Ergebnis des Peptide Scanning die Befunde
der Split-YFP Experimente und verstärkt die Vermutung, dass es eine Bindestelle
innerhalb der 5. Transmembrandomäne und eine weitere Bindestelle auf dem C-Terminus
von Alb3 gibt.

4 Ergebnisse 81
Aminosäuren 305-328:
DDPAQKNTLLVFKFLAQKNTLLVFKFLPLMNTLLVFKFLPLMIGYLVFKFLPLMIGYFAL
1 MARVLVSSPS SFFGSPLIKP SSSLRHSGVG GGGTAQFLPY RSNNNKLFTT 51 STTVRFSLNE IPPFHGLDSS VDIGAIFTRA ESLLYTIADA AVVGADSVVT 101 TDSSAVQKSG GWFGFISDAM ELVLKILKDG LSAVHVPYAY GFAIILLTII 151 VKAATYPLTK QQVESTLAMQ NLQPKIKAIQ QRYAGNQERI QLETSRLYKQ 201 AGVNPLAGCL PTLATIPVWI GLYQALSNVA NEGLFTEGFF WIPSLGGPTS 251 IAARQSGSGI SWLFPFVDGH PPLGWYDTVA YLVLPVLLIA SQYVSMEIMK 301 PPQTDDPAQK NTLLVFKFLP LMIGYFALSV PSGLSIYWLT NNVLSTAQQV 351 YLRKLGGAKP NMDENASKII SAGRAKRSIA QPDDAGERFR QLKEQEKRSK 401 KNKAVAKDTV ELVEESQSES EEGSDDEEEE AREGALASST TSKPLPEVGQ 451 RRSKRSKRKR TV
Aminosäuren 305-328:DDPAQKNTLLVFKFLPLMIGYFAL
Aminosäuren 374-388:RAKRSIAQPDDAGER
B
D
C
A
7255
43
34
26
17
kDa
Abbildung 4-10: Peptide Scanning von Alb3 Peptiden mit rekombinantem cpSRP43 Für die Methode des Peptide Scanning wurde zunächst das cpSRP43 rekombinant überexprimiert
und über den angefügten N-terminalen His-tag aufgereinigt. Ein Aliquot des Eluats His-cpSRP43
(His-43) wurde über eine SDS-PAGE aufgetrennt und anschließend mit Coomassie gefärbt, um die
erfolgreiche Aufreinigung zu überprüfen (A). Die Interaktion von rekombinantem His-cpSRP43 und
den Alb3 Peptiden wurde über eine Pepscan Methode (PepSpots, JPT Peptide Technologies,
Berlin) analysiert. Die dargestellte Sequenz von Alb3 zeigt die Aminosäuren, die für die PepSpot
Membran eingesetzt wurden (blau) (B). Der Bereich umfasst den 2. lumenalen loop, die
5. Transmembrandomäne (grau: Consensus Sequenz, hellgrau: zusätzliche Aminosäuren der
maximalen Vorhersage) sowie den C-Terminus von Alb3. Die Membran enthält 51 Peptide mit
jeweils 15 Aminosäuren Länge, die jeweils um drei Aminosäuren versetzt aufgetragen waren. Das
gebundene, rekombinante His-cpSRP43 wurde über einen Western Blot mit Hilfe eines Antikörpers
gegen das cpSRP43 (α-chaos) detektiert (C). Die interagierenden Peptide 3-6 umfassen die
Aminosäuren 305-328 und das Peptid 26 die Aminosäuren 374-388 von Alb3 und sind als
Bindestellen in der Alb3 Sequenz unterstrichen dargestellt (B). Die Analyse der PepSpots 3-6
erbrachte ein gemeinsames Bindemotiv der Aminosäuren 314-319 (LVFKFL) und ist in rot
hervorgehoben (D).

4 Ergebnisse 82
4.2.5 Bestätigung der mittels Peptide Scanning gezeigten Binderegionen im Split-YFP System
Die Auswertung des Peptide Scanning bestätigte die bereits über die Fusionskonstrukte
von Alb4 und Alb3 im Split-YFP gezeigte Bindestelle innerhalb der
5. Transmembrandomäne von Alb3. Aus diesem Grund wurden gezielt Mutationen und
Deletionen in diesem Bereich des Alb3 erzeugt und diese Proteine auf Interaktion mit
cpSRP43 im Split-YFP System getestet. Auf Grundlage eines Alignments der
Proteinsequenzen von Alb3 und Alb4 wurden die Aminosäuren von Alb3 innerhalb des
putativen Bindemotivs (LVFKFL) zu den entsprechenden Aminosäuren der Alb4 Sequenz
mutiert (Abbildung 4-11).
Alb3 MKPPQTDDPAQKNTLLVFKFLPLMIGYFALSVPSGLSIYWLTNNVLSAlb4 MQSSQSNDPAMKSSQAVTKLLPLMIGYFALSVPSGLSLYWLTNNILS
*:..*::*** *.: * *:*****************:******:**
310 315 320
Abbildung 4-11: Alignment von Alb3 und Alb4 im Bereich des Bindemotivs Das Alignment von Alb3 und Alb4 zeigt die Aminosäuren (rot), die bei den Mutationskonstrukten
Alb3(F316T), Alb3(L314A) und Alb3(L313Q,L314A) gegeneinander ausgetauscht worden sind.
Während das mutierte Protein Alb3(F316T) noch in der Lage war durch Interaktion mit
cpSRP43 die YFP-Fluoreszenz zu rekonstituieren, konnten die Proteine Alb3(L314A) und
Alb3(L313Q,L314A) nicht mehr mit dem cpSRP43 interagieren (Tabelle 4-4). Die
Deletionskonstrukte Alb3(Δ314,315) und Alb3(Δ314-317) zeigten ebenfalls keine
Interaktion mit dem cpSRP43, was auf eine entscheidende Bedeutung dieser Bindestelle
innerhalb der 5. Transmembrandomäne deutet (Tabelle 4-4). Die Analyse des Peptide
Scanning ergab weiterhin, dass die Aminosäuren 374-388 eine Bindestelle für das
cpSRP43 auf dem C-Terminus von Alb3 bilden. Um dieses zu bestätigen wurden die
Aminosäuren 374-388 mittels overlap-PCR in dem Alb3 Protein deletiert und die
Interaktion mit dem cpSRP43 ebenfalls über die bimolekulare Fluoreszenzkomplemetation
getestet. Die Deletion der Aminosäuren 374-388 hatte zur Folge, dass das Alb3(Δ374-
388) nicht mehr mit cpSRP43 interagieren konnte (Tabelle 4-4). Als Kontrolle wurde mit
den Aminosäuren 389-415 ein weiterer Bereich des C-Terminus von Alb3 deletiert. Das
Protein Alb3(Δ389-415) konnte im Vergleich zu dem Alb3(Δ374-388) noch mit dem
cpSRP43 interagieren (Tabelle 4-4). Die Aminosäuren 389-415 haben daher keinen
Einfluss auf die Bindung des cpSRP43. In weiteren Versuchen wurden C-terminal
verkürzte Alb3 Proteine im Split-YFP System getestet. Man konnte feststellen, dass die

4 Ergebnisse 83
Proteine Alb3(1-359) und Alb3(1-366) nicht mehr mit dem cpSRP43 interagieren konnten,
währenddessen die Proteine Alb3(1-415) und Alb3(1-442) eine Interaktion zeigten
(Tabelle 4-4). Damit führte die C-terminale Verkürzung des Proteins von 415 auf 366
Aminosäuren zum Ausbleiben der Bindung von cpSRP43. Da sich innerhalb dieses
deletierten Bereiches die gezeigte Bindestelle der Aminosäuren 374-388 befindet,
unterstützen auch diese Ergebnisse die Präsenz dieses Bindemotivs im C-Terminus von
Alb3.
Tabelle 4-4: Interaktion der Alb3 Deletions- und Mutationsproteine mit cpSRP43-nYFP in Arabidopsis Protoplasten
Die mittels overlap-PCR und Mutagenese-PCR erzeugten Deletions- und Mutationsproteine von
Alb3 wurden im Split-YFP System auf Interaktion mit cpSRP43 getestet. Die Alb3 Konstrukte
wurden an das cYFP und das cpSRP43 an das nYFP fusioniert. Arabidopsis Protoplasten wurden
mit den angegebenen Konstrukten transient transfiziert und die Proteine auf Interaktion analysiert.
cYFP-Konstrukte Interaktion mit cpSRP43-nYFP
Alb3(F316T) + Alb3(L314A) - Alb3(L313Q,L314A) - Alb3(Δ314,315) - Alb3(Δ314-317) - Alb3(Δ374-388) - Alb3(Δ389-415) + Alb3(1-442) + Alb3(1-415) + Alb3(1-366) - Alb3(1-359) -
Im nächsten Schritt sollte die Expression der entscheidenden Deletionskonstrukte von
Alb3 und deren Integration in die Thylakoidmembran gezeigt werden. Dazu wurden
Arabidopsis thaliana Protoplasten mit den Konstrukten pSPYCE-Alb3, pSPYCE-
Alb3(L313Q,L314A), pSPYCE-Alb3(Δ314-317) und pSPYCE-Alb3(Δ374-388) transfiziert.
Wie bereits in Abschnitt 4.2.2 beschrieben, wurden für den Nachweis der Expression die
Proteine des gewaschenen Membranpellets sowie die Proben aus der TCA-Fällung
(Überstand nach Lyse und Waschfraktion) über eine SDS-PAGE aufgetrennt und mittels
Immundetektion ausgewertet (Abbildung 4-12). Dabei konnten die cYFP-Fusionsproteine
über den angefügten HA-tag detektiert werden. Es zeigte sich, dass die Proteine Alb3-
cYFP, Alb3(L313Q,L314A)-cYFP, Alb3(Δ314-317)-cYFP und Alb3(Δ374-388)-cYFP als

4 Ergebnisse 84
YFP-Fusionsproteine ausschließlich in der Membranfraktion nachgewiesen werden
konnten. Als Kontrolle wurden nicht transfizierte Protoplasten eingesetzt. In dieser Probe
waren keine Signale bei der entsprechenden Größe der YFP-Fusionsproteine
nachzuweisen. Damit konnte gezeigt werden, dass die Deletionsproteine von Alb3 in der
Membran integriert vorlagen. Der Verlust der Bindung des cpSRP43 ist daher auf die
Deletion und die Mutation der entscheidenden Binderegionen zurückzuführen und nicht
auf eine fehlende Integration der Proteine in die Membran.
Alb3(Δ374-388)-cYFP
Alb3(Δ314-317)-cYFP
Alb3(L313Q,L314A)-cYFP
Alb3-cYFP
Kontrolle
α-HA
P Ü W
Abbildung 4-12: Expressionsnachweis ausgewählter Alb3 Deletions- und Mutationskonstrukte in Arabidopsis Protoplasten
Arabidopsis Protoplasten wurden mit den angegebenen Konstrukten transient transfiziert und die
Expression der YFP-Fusionsproteine über Immundetektion nachgewiesen. Dazu wurden die
lysierten Zellen abzentrifugiert und der Überstand (Ü) mittels TCA-Fällung präzipitiert. Das
Membranpellet wurde mit 0,2 M NaOH gewaschen (P), zentrifugiert und der Überstand
(W=Waschfraktion) mittels TCA-Fällung präzipitiert. Es folgte die Auftrennung der Proben über eine
SDS-PAGE und die Analyse mittels Immundetektion. Die Expression der cYFP-Fusionsproteine
konnte über einen anti-HA Antikörper (α-HA) nachgewiesen werden. Als Kontrolle wurden nicht
transfizierte Protoplasten eingesetzt (Kontrolle).
4.2.6 Verifizierung der Binderegionen im Hefe Split-Ubiquitin System
Die Binderegionen im Bereich der Aminosäuren 314-317 und 374-388 von Alb3 sollten im
Hefe Split-Ubiquitin System bestätigt werden. Für den Interaktionstest wurden die
Gensequenzen von Alb3, Alb3(Δ314-317), Alb3(Δ374-388) und Alb3(Δ389-415) in den
pAMBV4-Vektor und die Gensequenz von cpSRP43 in den pADSL-Vektor kloniert. Die
Kontrolle der Cotransformation anhand der Transformationsraten auf dem –LT-Medium
war in allen Ansätzen positiv (Daten nicht gezeigt). Die Cotransformation der Konstrukte
pAMBV4-Alb3, pAMBV4-Alb3(Δ314-317), pAMBV4-Alb3(Δ374-388) und pAMBV4-

4 Ergebnisse 85
Alb3(Δ389-415) mit dem Kontrollplasmid (kodiert für Alg5-NubI) resultierte in einem
Wachstum der Hefezellen auf –LTH-Medium und deutete damit auf einen korrekten
Einbau der Proteine in die Membranen. Durch den Zusatz von 15 mM 3-Aminotriazol
(3-AT), einem kompetitiven Inhibitor des His3-Genproduktes, verringerte sich eine
schwache Eigenaktivierung der Proteine so weit, dass kein Wachstum beim Einsatz des
Kontrollplasmids (kodiert für Alg5-NubG) auf –LTH-Medium mehr sichtbar war.
Die Interaktionsstudien ergaben, dass die Deletionsproteine Alb3(Δ314-317) und
Alb3(Δ374-388) schwächer mit dem cpSRP43 interagierten, als das Volllängenprotein
Alb3 (Tabelle 4-5). Der Einsatz des Deletionsproteins Alb3(Δ389-415) zeigte im Vergleich
zum Volllängenprotein kein verringertes Wachstum der Hefezellen auf –LTH-Medium und
damit eine gleich starke Interaktion der Proteine im Split-Ubiquitin-System (Tabelle 4-5).
Diese Deletion hatte demnach keinen Einfluss auf die Bindung des cpSRP43. Daraus
lässt sich schließen, dass die Deletionen der Aminosäuren 314-317 und 374-388 von Alb3
auch im Hefe System einen deutlichen Effekt auf die Interaktion mit cpSRP43 haben. Bei
allen Transformationen, die ein Wachstum der Hefezellen auf –LTH-Medium aufwiesen,
konnte eine Blaufärbung im Filtertest nachgewiesen werden (Tabelle 4-5). Die
β-Galaktosidase Aktivität basiert auf einer Aktivierung des Reportergens lacZ und damit
auf einer Interaktion der getesteten Proteine.
Tabelle 4-5: Interaktionstest von Alb3 Deletionsproteinen mit cpSRP43 im Hefe Split-Ubiquitin System
Dargestellt sind die Interaktionstests von Alb3 und Alb3 Deletionsproteinen mit cpSRP43 im Hefe
Split-Ubiquitin System. Der Hefe Stamm DSY-1 wurde mit den entsprechenden pAMBV4-
Plasmiden und dem pADSL-cpSRP43 Plasmid cotransformiert und die Aktivierung der
Reportergene ausgewertet. Durch das Wachstum der cotransformierten Hefezellen auf
Minimalmedium (-his) (++ = deutliches Wachstum, + = geringes Wachstum, - = kein Wachstum)
und über die Bestimmung der Enzymaktivität der β-Galaktosidase (β-gal), die durch eine
Blaufärbung der Hefekolonien im β-Galaktosidase Filtertest analysiert wurde (+ = Blaufärbung, - =
keine Blaufärbung), konnten Aussagen über die Interaktion der Proteine getroffen werden.
pADSL-Nx -his β-gal. -his β-gal. -his β-gal -his β-gal
Alg5-NubI ++ + ++ + ++ + ++ +
Alg5-NubG - - - - - - - -
cpSRP43 ++ + + + + + ++ +
pAMBV4-Alb3 (Δ374-388)
pAMBV4-Alb3 (Δ389-415)pAMBV4-Alb3 pAMBV4-Alb3
(Δ314-317)

4 Ergebnisse 86
4.2.7 Pull down Analysen zur Interaktion von Alb3 und cpSRP43
Anhand von pull down Analysen sollte die in vitro Interaktion von Alb3 und cpSRP43
gezeigt werden. Eine rekombinante Expression des Alb3 in E. coli war bisher nicht in
ausreichenden Mengen und ohne deutlichen Abbau des Proteins möglich. Deshalb
wurden die entsprechenden Gensequenzen von Alb3(299-462) und dessen
Deletionskonstrukte in die Vektoren pSS und pIVEX1.3WG kloniert und eine in vitro
Transkription und Translation durchgeführt. Die Translationskontrolle erfolgte über eine
SDS-PAGE mit anschließender Immundetektion. Die erhaltenen Translationsprodukte
konnten für die pull down Versuche mit rekombinantem His-cpSRP43 eingesetzt werden.
Als Kontrollprotein wurde rekombinant erzeugtes His-cpSRP54 auf Interaktion mit den
Translationsprodukten überprüft. Zunächst wurden rekombinantes His-cpSRP43 und His-
cpSRP54 mit den Translationsprodukten von Alb3 inkubiert. Nach Zugabe von Ni-NTA
Agarose erfolgte die Bindung der mit einem His-tag versehenen rekombinanten Proteine.
Durch die Waschung der beads wurden nicht gebundene Proteine entfernt, und die
rekombinanten Proteine His-cpSRP43 und His-cpSRP54 konnten zusammen mit den
gebundenen Translationsprodukten eluiert werden. Der Nachweis der coeluierten
Translationsprodukte erfolgte über eine SDS-PAGE mit anschließender Immundetektion
(Abbildung 4-13 A). Das Translationsprodukt Alb3(299-462) umfasste die Aminosäuren
299-462 und damit den 2. lumenalen loop, die 5. Transmembrandomäne und den
C-Terminus von Alb3. Diese Sequenz von Alb3 wurde bereits für die Methode des
Peptide Scanning eingesetzt. Nach der Immundetektion mit einem Alb3 Antikörper konnte
das Alb3(299-462) in der Eluatfraktion des His-cpSRP43 nachgewiesen werden. Damit
war das Alb3(299-462) in der Lage an das His-cpSRP43 zu binden. In dem Eluat des His-
cpSRP54 war kein Translationsprodukt zu detektieren. Diese Negativkontrolle zeigte,
dass das Alb3(299-462) nicht mit cpSRP54 interagierte. Damit konnte zunächst die
spezifische Interaktion von Alb3 und cpSRP43 bestätigt werden. Im weiteren Vorgehen
sollten die in den vorangegangenen Versuchen identifizierten Bindestellen des Alb3 über
die pull down Versuche bestätigt werden. Dazu wurde das Alb3(299-462) N-terminal
verkürzt, so dass es nur noch die Aminosäuren 320-462 umfasste. Mit dieser Verkürzung
wurde die bisher analysierte Bindestelle im Bereich der Aminosäuren 314-317 entfernt.
Das Protein Alb3(320-462) war allerdings immer noch in der Lage an das cpSRP43 zu
binden. Das gleiche Ergebnis war auch mit dem Protein Alb3(299-462/Δ314-317), bei
dem die Bindestelle der Aminosäuren 314-317 deletiert war, zu verzeichnen. Damit
scheint die Bindestelle innerhalb der 5. Transmembrandomäne von Alb3 für die in vitro
Interaktion im pull down nicht essentiell zu sein. Es zeigte sich allerdings weiterhin durch
den Einsatz des Proteins Alb3(336-462), dass der C-Terminus von Alb3 allein nicht in der
Lage war, mit dem cpSRP43 zu interagieren. Auch das Translationsprodukt Alb3(328-

4 Ergebnisse 87
462), welches den C-Terminus von Alb3 um einen Teil der 5. Transmembrandomäne
verlängerte, reichte noch nicht für eine Coelution mit dem His-cpSRP43 aus. Das Protein
Alb3(320-462) umfasste einen erheblichen Anteil der 5. Transmembrandomäne von Alb3
und ist in der Lage an das His-cpSRP43 zu binden. Damit lässt sich sagen, dass in dem
pull down Versuch nicht exakt die Bindestelle der Aminosäuren 314-317 bestätigt werden
konnte. Jedoch ist ein erheblicher Teil der 5. Transmembrandomäne wichtig, der auch
durch das Peptide Scanning mit den Aminosäuren 305-328 identifiziert wurde (siehe
Abschnitt 4.2.4). Um die zweite Bindestelle von Alb3 innerhalb des C-Terminus zu testen,
wurden in dem Translationsprodukt Alb3(299-462) die Aminosäuren 374-388 deletiert. In
Abbildung 4-13 ist gezeigt, dass das Protein Alb3(299-462/Δ374-388) nicht mehr mit dem
cpSRP43 interagierte. Im Vergleich dazu konnte das Kontrollprotein Alb3(299-462/Δ389-
415), bei dem die Aminosäuren 389-415 fehlten, noch im Eluat der His-cpSRP43 Probe
nachgewiesen werden. Die Bindestelle im C-Terminus mit den Aminosäuren 374-388
konnte daher auch in den pull down Analysen verifiziert werden. Für alle in den Analysen
eingesetzten Alb3 Translationsprodukte war keine Interaktion mit dem His-cpSRP54 zu
verzeichnen. Als weitere Kontrolle wurde der C-Terminus von Alb4 für die in vitro Studien
eingesetzt. Das Translationsprodukt Alb4(328-499) zeigte weder eine Interaktion mit dem
His-cpSRP43 noch mit dem His-cpSRP54.
In einem weiteren pull down Ansatz wurde rekombinantes cpSRP43 mit C-terminalem
His-tag (zur Verfügung gestellt von Beatrix Dünschede, AG Schünemann) eingesetzt. Das
Translationsprodukt Alb3(299-462) wurde sowohl von dem His-cpSRP43, als auch von
dem cpSRP43-His gebunden (Abbildung 4-13 B). Die Lage des tags scheint daher für die
Interaktion keine Rolle zu spielen. Des Weiteren kam ein vorgeformter cpSRP-Komplex
(His-cpSRP43 + cpSRP54 ohne tag) zum Einsatz (zur Verfügung gestellt von Beatrix
Dünschede, AG Schünemann). Auch das im cpSRP-Komplex vorliegende His-cpSRP43
konnte das Translationsprodukt Alb3(299-462) binden (Abbildung 4-13 B). Das His-
cpSRP54 zeigte in dem Kontrollansatz wiederum keine Interaktion mit dem Alb3
Translationsprodukt (Abbildung 4-13 B). Die geringe Menge an coeluiertem
Translationsprodukt war auf nicht ausreichendes Waschen dieses Ansatzes
zurückzuführen, da in dieser mehrfach durchgeführten Kontrolle in weiteren Versuchen
keine Bindung zu beobachten war.

4 Ergebnisse 88
His-43 His-54
Alb3 (299-462)
Alb3 (320-462)
Alb3 (328-462)
Alb3 (336-462)
TP
Alb3 (320-442)
Alb3 (299-462/Δ374-388)
Alb3 (299-462/Δ389-415)
Alb4 (328-499)
Alb3 (299-462/Δ314-317)
A
His-5443-His His-43His-
cpSRP
Alb3 (299-462)
B
Abbildung 4-13: Pull down Analysen von rekombinantem cpSRP43 und cpSRP54
mit in vitro Translationsprodukten von Alb3 und Alb4 Gezeigt sind pull down Analysen von 5 μg rekombinantem His-cpSRP43 (His-43) und His-
cpSRP54 (His-54) mit den angegebenen in vitro translatierten Deletionsproteinen von Alb3 und
Alb4. Zur Kontrolle der in vitro Translation wurden die Translationsprodukte (TP) über eine SDS-
PAGE aufgetrennt, mittels Immundetektion (α-Alb3, α-Alb4) überprüft und gleiche Mengen für den
pull down eingesetzt. Die mögliche Coelution der Translationsprodukte mit den rekombinanten
Proteinen konnte über eine Immundetektion in den jeweiligen Eluatfraktionen nachgewiesen
werden (A). In weiteren pull down Analysen wurden 5 μg rekombinantes cpSRP43-His (43-His),
His-cpSRP43 (His-43), His-cpSRP54 (His-54) und 10 µg His-cpSRP (His-43+54) mit in vitro
translatiertem Alb3(299-462) eingesetzt. Die Eluatfraktionen wurden über eine SDS-PAGE
aufgetrennt und die Coelution des Alb3(299-462) mittels Immundetektion (α-Alb3) nachgewiesen
(B).

4 Ergebnisse 89
4.2.8 Untersuchung der Alb3 Bindestelle im Thylakoidmembran-Bindungstest
Ein Thylakoidmembran-Bindungstest sollte weiteren Aufschluss über die Interaktion von
Alb3 und cpSRP43 an der Thylakoidmembran geben. Die Thylakoidmembranen wurden
zunächst salzgewaschen, um peripher gebundene Proteine zu entfernen. Darauf erfolgte
die Inkubation von 2 µg rekombinantem His-cpSRP43 mit den Thylakoidmembranen, das
Waschen der Membranen mit 1 M NaCl und die Aufnahme des Membranpellets in SDS-
Probenpuffer. Die Überprüfung der Bindung des His-cpSRP43 an die Membranen erfolgte
über eine SDS-PAGE mit anschließender Immundetektion. Das Ergebnis zeigte eine
salzresistente Bindung des His-cpSRP43 an die Thylakoidmembranen, da das
rekombinante Protein in den Waschschritten mit 1 M NaCl nicht entfernt werden konnte
(Abbildung 4-14). Unter gleichen Bedingungen wurde keine Bindung des Kontrollproteins
GST an die Membranen beobachtet (Daten nicht gezeigt). Im darauf folgenden Ansatz
wurde versucht, die Bindung des rekombinanten His-cpSRP43 an die Membranen mit
einem synthetischen Peptid von Alb3 zu inhibieren (Abbildung 4-14). Das synthetische
Peptid umfasste die Aminosäuren 367-388 und beinhaltet damit die bereits identifizierte
Bindestelle des C-Terminus von Alb3 (Aminosäuren 374-388). Das rekombinante His-
cpSRP43 wurde zunächst mit dem synthetischen Alb3-Peptid vorinkubiert, bevor die
Bindung an die Thylakoidmembranen erfolgte. Die Zugabe von 250 µg des Peptids
verringerte im Vergleich zu dem Ansatz ohne Peptidvorinkubation die Bindung des His-
cpSRP43 an die Membranen. Bei Steigerung der Peptidmenge auf 500 µg wurde der
inhibierende Effekt größer. Es konnte weniger rekombinantes Protein an die
Thylakoidmembranen binden, als bei Vorinkubation mit 250 µg Alb3-Peptid und ohne
Peptid. Als Kontrolle wurde ein Peptid eingesetzt, welches die Aminosäuren 382-395 des
C-Terminus von Alb4 umfasste. Die Vorinkubation des His-cpSRP43 mit 250 µg des Alb4-
Peptids zeigte keine Verringerung der Bindung an die Membranen. Auch die Steigerung
der Peptidmenge auf 500 µg hatte keinen Einfluss auf die Membraninteraktion des His-
cpSRP43. Als Ladekontrolle der Ansätze diente der Nachweis des Alb3 Proteins in der
Thylakoidmembran. Mit dem Thylakoidmembran-Bindungstest konnte eine salzresistente
Bindung des rekombinanten His-cpSRP43 an die Membranen ermittelt werden. Zudem
wurde gezeigt, dass das Alb3-Peptid vermutlich mit dem cpSRP43 interagierte und somit
die Membranbindung verringerte. Dieses Ergebnis stützt daher die Identifizierung der
Bindestelle innerhalb des C-Terminus (Aminosäuren 374-388) von Alb3.

4 Ergebnisse 90
His-43
Alb3 Peptid
250 µg 500 µg
Alb4 Peptid
250 µg 500 µg0 µg
Alb3
Abbildung 4-14: Einfluss eines synthetischen Peptids von Alb3 auf die Thylakoidmembranbindung von cpSRP43
Für den Thylakoidmembran-Bindungstest wurden 2 µg rekombinantes His-cpSRP43 (His-43) mit
Thylakoidmembranen (50 µg Chlorophyll) inkubiert. Die Membranen wurden mit 1 M NaCl
gewaschen und das Membranpellet in SDS-Probenpuffer aufgenommen. Für die Peptidinhibierung
wurde das rekombinante Protein His-cpSRP43 zunächst mit 250 µg und 500 µg des Alb3 Peptids
bzw. Alb4 Peptids vorinkubiert. Das Alb3 Peptid umfasste die Aminosäuren 367-388 von Alb3 und
das Alb4 Peptid die Aminosäuren 382-395 der Alb4 Sequenz. Aliquots der Proben wurden über
eine SDS-PAGE aufgetrennt und die Bindung des His-cpSRP43 an die Thylakoidmembranen
mittels Immundetektion (α-chaos) analysiert. Als Ladekontrolle diente ein Western Blot Nachweis
des Alb3 in den Thylakoidmembranen.
4.2.9 Analyse der Bindedomäne des cpSRP43 für das Alb3
In einer Masterarbeit, die in unserer Arbeitsgruppe parallel zu dieser Arbeit angefertigt
wurde, sollte die Interaktionsdomäne des cpSRP43 für das Alb3 identifiziert werden
(Vörös, 2007). Das cpSRP43 Protein besitzt eine N-terminale Chromodomäne (CD1),
gefolgt von vier Ankyrin repeats (Ank1-4) und zwei C-terminalen Chromodomänen (CD2,
CD3). Es wurden vorab verschiedene GST-cpSRP43 Deletionsproteine auf mögliche
Interaktion mit den Membranen im Thylakoidmembran-Bindungstest analysiert. Dabei
stellte sich heraus, dass die Ankyrin repeat Region 4 des cpSRP43 eine entscheidende
Bedeutung für die Bindung des Proteins an die Thylakoidmembran haben könnte, da das
GST-cpSRP43ΔAnk4, bei dem die Ankyrin repeat Region 4 deletiert wurde, nicht mehr in
der Lage war, an die Membranen zu binden (Vörös, 2007). Ausgehend von diesem
Ergebnis sollte nun überprüft werden, ob die Deletion der anderen Ankyrin repeats 1-3
einen Effekt auf die Membranbindung des jeweiligen Proteins hat. Die Deletionen der
Ankyrin repeats innerhalb der cpSRP43 Sequenz sind anhand von Strukturvorhersagen
(Klimyuk et al., 1999; Jonas-Straube et al., 2001; Goforth et al., 2004) für die Regionen
vorgenommen worden. Diese Vorhersagen weichen allerdings teilweise von der kürzlich

4 Ergebnisse 91
publizierten Kristallstruktur des cpSRP43 ab (Stengel et al., 2008) und sind vergleichend
in Abbildung 4-15 dargestellt. Während die α-Helices der Ankyrin repeats 1, 2 und 3 (α1-
6) in dem Bereich der vorhergesagten Sequenz liegen, gibt es bei der Ankyrin repeat
Region 4 eine Abweichung zu der Kristallstruktur. Die Kristallstruktur zeigt einen direkten
Übergang von Ankyrin repeat 4 (α7+8) zur Chromodomäne 2, während in den
Vorhersageprogrammen zwischen Ankyrin repeat 4 und Chromodomäne 2 eine linker
Sequenz (Aminosäuren 252-267) liegt.
1 MQKVFLAMDTCALVIHQSLSRIKLSPPKSSSSSSSAFSPESLPIRRIELCFRGAICAAVQ
61 RNYEETTSSVEEAEEDDESSSSYGEVNKIIGSRTAGEGAMEYLIEWKDGHSPSWVPSSYI
121 AADVVSEYETPWWTAARKADEQALSQLLEDRDVDAVDENGRTALLFVAGLGSDKCVRLLA
181 EAGADLDHRDMRGGLTALHMAAGYVRPEVVEALVELGADIEVEDERGLTALELAREILKT
241 TPKGNPMQFGRRIGLEKVINVLEGQVFEYAEVDEIVEKRGKGKDVEYLVRWKDGGDCEWV
301 KGVHVAEDVAKDYEDGLEYAVAESVIGKRVGDDGKTIEYLVKWTDMSDATWEPQDNVDST
361 LVLLYQQQQPMNE
Transitsequenz
β1 β2 β3CD1
CD2
CD3
α1a/b α2 α3 α4
α5 α6 α7
α8
Ank1 Ank2
Ank3 Ank4
B
A cpSRP43 A1 A2 A3 A4 CD3CD2CD2CD1
Abbildung 4-15: Schematische Darstellung des cpSRP43 Das stromale Protein cpSRP43 besteht aus einer N-terminalen Chromodomäne (CD1), gefolgt von
vier Ankyrin repeats (Ank1-4) und zwei C-terminalen Chromodomänen (CD2, CD3) (A). In der
Aminosäuresequenz sind die β-Faltblätter der Chromodomäne 1 (β1-3, graue Pfeile) sowie die
α-Helices der Ankyrin repeats (α1-8, graue Balken) aus der Kristallstruktur näher dargestellt
(verändert nach Stengel et al., 2008) (B). Die Bereiche der Ankyrin repeats, die aus
Vorhersageprogrammen für diese Arbeit verwendet wurden, sind in der Aminosäuresequenz rot
dargestellt (Klimyuk et al., 1999; Jonas-Straube et al., 2001; Goforth et al., 2004). Während die α-
Helices der Ankyrin repeats 1-3 innerhalb der vorhergesagten Bereiche liegen, zeigt die Ankyrin
repeat Region 4 auf Grundlage der Kristallstruktur einen zur Chromodomäne 2 verlängerten
Bereich im Vergleich zur Vorhersage.
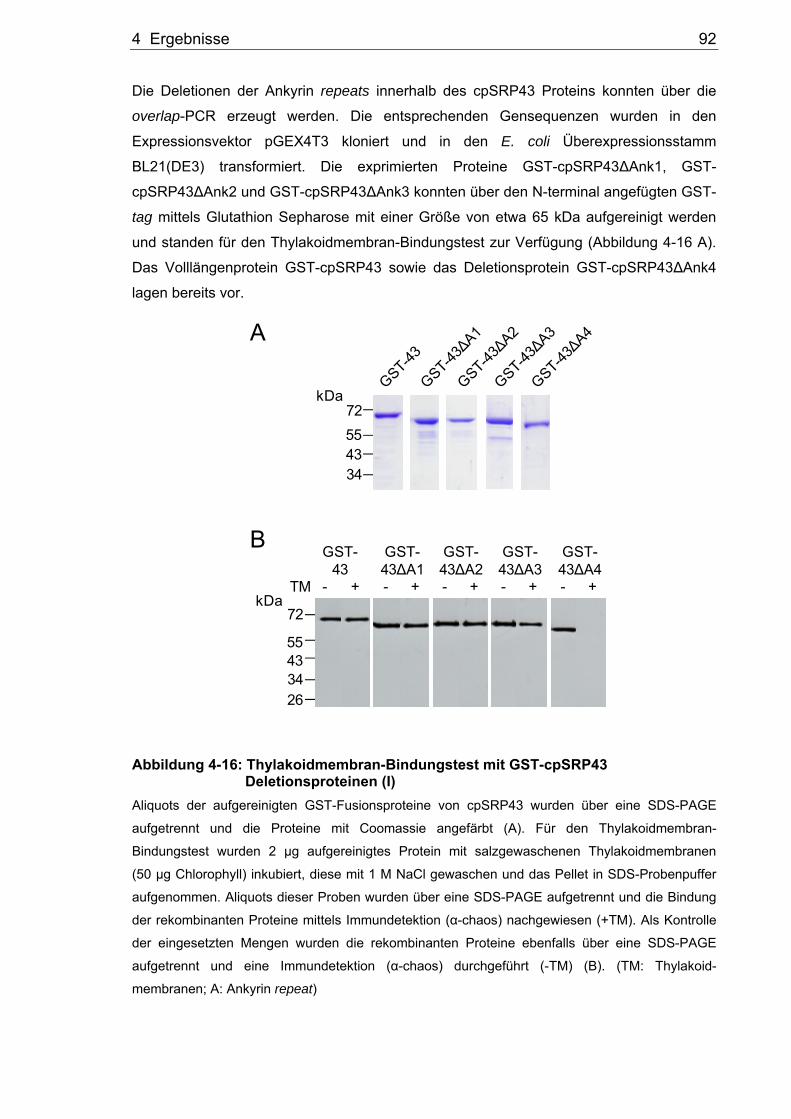
4 Ergebnisse 92
Die Deletionen der Ankyrin repeats innerhalb des cpSRP43 Proteins konnten über die
overlap-PCR erzeugt werden. Die entsprechenden Gensequenzen wurden in den
Expressionsvektor pGEX4T3 kloniert und in den E. coli Überexpressionsstamm
BL21(DE3) transformiert. Die exprimierten Proteine GST-cpSRP43ΔAnk1, GST-
cpSRP43ΔAnk2 und GST-cpSRP43ΔAnk3 konnten über den N-terminal angefügten GST-
tag mittels Glutathion Sepharose mit einer Größe von etwa 65 kDa aufgereinigt werden
und standen für den Thylakoidmembran-Bindungstest zur Verfügung (Abbildung 4-16 A).
Das Volllängenprotein GST-cpSRP43 sowie das Deletionsprotein GST-cpSRP43ΔAnk4
lagen bereits vor.
72554334
kDa
A
72
55433426
TM - +kDa
- + - + - + - +
GST-43ΔA2
GST-43ΔA3
GST-43ΔA4
GST-43ΔA1
GST-43
B
Abbildung 4-16: Thylakoidmembran-Bindungstest mit GST-cpSRP43 Deletionsproteinen (I)
Aliquots der aufgereinigten GST-Fusionsproteine von cpSRP43 wurden über eine SDS-PAGE
aufgetrennt und die Proteine mit Coomassie angefärbt (A). Für den Thylakoidmembran-
Bindungstest wurden 2 µg aufgereinigtes Protein mit salzgewaschenen Thylakoidmembranen
(50 µg Chlorophyll) inkubiert, diese mit 1 M NaCl gewaschen und das Pellet in SDS-Probenpuffer
aufgenommen. Aliquots dieser Proben wurden über eine SDS-PAGE aufgetrennt und die Bindung
der rekombinanten Proteine mittels Immundetektion (α-chaos) nachgewiesen (+TM). Als Kontrolle
der eingesetzten Mengen wurden die rekombinanten Proteine ebenfalls über eine SDS-PAGE
aufgetrennt und eine Immundetektion (α-chaos) durchgeführt (-TM) (B). (TM: Thylakoid-
membranen; A: Ankyrin repeat)

4 Ergebnisse 93
Die rekombinant erzeugten GST-Fusionsproteine des cpSRP43 wurden zunächst mit
salzgewaschenen Thylakoidmembranen inkubiert. Nach Waschen mit 1 M NaCl und
Resuspendieren der Membranen in SDS-Probenpuffer, konnte die Bindung der Proteine
über eine Immundetektion ausgewertet werden. Das Volllängenprotein GST-cpSRP43
war in der Lage an die Membranen zu binden. Des Weiteren konnte bestätigt werden,
dass die Deletion der Ankyrin repeat Region 4 einen Verlust der Bindung des
rekombinanten Proteins an die Thylakoidmembranen zur Folge hatte (Abbildung 4-16 B).
Die Deletion der Ankyrin repeats 1, 2 und 3 hatte keinen Einfluss auf die
Membranbindung der entsprechenden Proteine. Die Proteine GST-cpSRP43ΔAnk1, GST-
cpSRP43ΔAnk2 und GST-cpSRP43ΔAnk3 zeigten alle eine Interaktion mit der
Thylakoidmembran und konnten somit über die Immundetektion nachgewiesen werden
(Abbildung 4-16 B). Als Kontrolle der eingesetzten Mengen wurden die rekombinanten
Proteine ebenfalls über eine SDS-PAGE aufgetrennt und eine Immundetektion
durchgeführt. Das Ergebnis des Thylakoidmembran-Bindungstests deutet darauf hin, dass
spezifisch die Ankyrin repeat Region 4 für die Membranbindung des cpSRP43
verantwortlich ist.
Um weiterhin die Spezifität der Ankyrin repeat Region 4 des cpSRP43 für die Interaktion
mit den Thylakoidmembranen zu zeigen, wurden weitere Deletionsproteine des cpSRP43
erstellt. Aus bereits vorangegangenen Arbeiten (Vörös, 2007) war ersichtlich, dass das
Deletionsprotein GST-cpSRP43(Ank4+CD2) inklusive der linker Sequenz noch für eine
Membranbindung ausreichend war. Mittels overlap-PCR war es möglich jeweils die
anderen Ankyrin repeats 1-3 vor die linker Sequenz und die Chromodomäne 2 zu
klonieren. Die Gensequenzen dieser Deletionsproteine wurden ebenfalls in den
Expressionsvektor pGEX4T3 kloniert und die Proteine im E. coli Expressionsstamm
BL21(DE3) überexprimiert. Die Fusionsproteine GST-cpSRP43(Ank1+CD2), GST-
cpSRP43(Ank2+CD2) und GST-cpSRP43(Ank3+CD2) konnten über den N-terminal
angefügten GST-tag mittels Glutathion Sepharose mit einer Größe von etwa 36 kDa
aufgereinigt werden. Die Eluate der neu erstellten, aufgereinigten Deletionsproteine
wurden mit den bereits vorhandenen Proteinen GST-cpSRP43, GST-
cpSRP43(Ank4+CD2) sowie GST-cpSRP43(CD2+3) über eine SDS-PAGE mit
anschließender Coomassie Färbung überprüft (Abbildung 4-17 A). Das Volllängenprotein
GST-cpSRP43 sowie die Deletionsproteine GST-cpSRP43(Ank1+CD2), GST-
cpSRP43(Ank2+CD2), GST-cpSRP43(Ank3+CD2), GST-cpSRP43(Ank4+CD2) und GST-
cpSRP43(CD2+3) wurden in den Thylakoidmembran-Bindungstest eingesetzt. Als
Kontrolle der eingesetzten Mengen wurden die rekombinanten Proteine über eine SDS-
PAGE aufgetrennt und eine Immundetektion durchgeführt (Abbildung 4-17 B). Es konnte
zunächst bestätigt werden, dass neben dem Volllängenprotein GST-cpSRP43 auch das
Deletionsprotein GST-cpSRP43(Ank4+CD2) noch an die Thylakoidmembranen binden

4 Ergebnisse 94
konnte. Ein Kontrollprotein bestehend aus der Chromodomäne 2 und 3 (GST-
cpSRP43(CD2+3)) war nicht in der Lage mit den Membranen zu interagieren. Die
Western Blot Analysen des Thylakoidmembran-Bindungstest zeigten weiterhin, dass
durch eine Kombination der Ankyrin repeats 1, 2 und 3 mit der Chromodomäne 2 keine
Bindung der Proteine an die Membran erreicht werden konnte (Abbildung 4-17 B). Das
Ergebnis lässt darauf schließen, dass spezifisch die Ankyrin repeat Region 4 gemeinsam
mit der Chromodomäne 2 die Membranbindung des cpSRP43 ermöglichte. Ein Austausch
der Ankyrin repeat Region 4 gegen die anderen Ankyrin repeats bedeutete einen Verlust
der Bindefähigkeit.
72554334
26
kDa
GST-43 GST-43(A1+CD2)
GST-43(A2+CD2)
GST-43(A3+CD2)
GST-43(A4+CD2)
GST-43(CD2+3)
- + - + - + - + - + - + TM
B
A
Abbildung 4-17: Thylakoidmembran-Bindungstest mit GST-cpSRP43 Deletionsproteinen (II)
Zur Kontrolle der Aufreinigung wurden Aliquots der GST-Fusionsproteine von cpSRP43 mittels
SDS-PAGE aufgetrennt und anschließend mit Coomassie angefärbt (A). Für den
Thylakoidmembran-Bindungstest wurden 2 µg aufgereinigtes Protein mit salzgewaschenen
Thylakoidmembranen (50 µg Chlorophyll) inkubiert, diese mit 1 M NaCl gewaschen und das Pellet
in SDS-Probenpuffer aufgenommen. Aliquots dieser Proben wurden über eine SDS-PAGE
aufgetrennt und die Bindung der rekombinanten Proteine mittels Immundetektion (α-chaos)
nachgewiesen (+ TM). Als Kontrolle der eingesetzten Mengen wurden die rekombinanten Proteine
ebenfalls über eine SDS-PAGE aufgetrennt und eine Immundetektion (α-chaos) durchgeführt
(- TM) (B). (TM: Thylakoidmembranen; A: Ankyrin repeat; CD: Chromodomäne)

4 Ergebnisse 95
4.3 Analyse der Interaktion von Alb3 und Alb4 mit den Proteinen der Thylakoidmembran
Nachdem die Rolle von Alb3 und Alb4 innerhalb des cpSRP-Transportwegs näher
untersucht worden war, sollte in einem weiteren Projekt die Interaktion der beiden
Proteine untereinander und mit anderen Thylakoidmembranproteinen analysiert werden.
Dazu wurden mit dem chemischen Crosslinking, der bimolekularen
Fluoreszenzkomplementation, der Gelfiltration und der Coimmunpräzipitation mehrere
methodische Ansätze gewählt.
4.3.1 Crosslinking der Thylakoidmembranproteine
Um mögliche Interaktionspartner von Alb3 und Alb4 innerhalb der Thylakoidmembran zu
identifizieren, wurde die Methode des chemischen Crosslinkings durchgeführt. Dabei
werden Proteine, die sich in der Thylakoidmembran in räumlicher Nähe befinden, kovalent
aneinander gebunden. Zur Analyse der Protein-Protein Interaktion standen bereits eine
Auswahl an chemischen Crosslinkern im Labor zur Verfügung (AEDP, BS3, DSG, MBS,
sMBS, SPDP). Dabei handelte es sich um homo- und heterobifunktionale Crosslinker, die
gleiche bzw. zwei unterschiedliche reaktive Gruppen besaßen. Die Wahl unterschiedlicher
Crosslinker ermöglichte es verschiedene Bedingungen abzudecken. So dienen z. B.
unterschiedlich lange spacer für die Vernetzung von Proteinen mit verschiedenen
Abständen. Die unterschiedlichen Eigenschaften der Crosslinking Reagenzien dienen
dazu hydrophobe, membranständige und hydrophile, lösliche Bereiche der
Thylakoidmembranproteine zu vernetzen.
Für die Studien wurden Thylakoidmembranen mit den verschiedenen Crosslinkern
inkubiert, gewaschen und in SDS-Probenpuffer aufgenommen. Aliquots der vernetzten
Thylakoidmembranproteine wurden über eine SDS-PAGE aufgetrennt und mögliche
Crosslinking-Produkte von Alb3 und Alb4 über eine Immundetektion identifiziert. Durch
den Einsatz des Crosslinkers sMBS (m-maleimidobenzoyl-N-hydroxysulfosuccinimide
ester) konnte ein Crosslinking-Produkt bei ca. 170 kDa erzeugt werden, welches das Alb4
Protein enthielt (Abbildung 4-18). Es zeigte sich, dass eine Konzentration von 0,6 mM des
sMBS zu einem spezifischen Produkt führte. In allen anderen Ansätzen mit den
verschiedenen Crosslinking Reagenzien, die zudem in variablen Konzentrationen
eingesetzt worden waren, konnten nach der Immundetektion mit dem Alb3 und dem Alb4
Antikörper keine weiteren Crosslinking-Produkte identifiziert werden (Daten nicht gezeigt).
Die Spezifität des Alb4 Crosslinking-Produktes wurde durch den Einsatz des
affinitätsgereinigten α-Alb4 Antikörpers bestärkt. Als Kontrolle wurden Thylakoid-
membranen ohne vorherige Inkubation mit dem Crosslinker sMBS aufgetragen. Dieser

4 Ergebnisse 96
Western Blot zeigte auf der Höhe von 170 kDa kein Signal (Abbildung 4-18). Im nächsten
Schritt sollten die weiteren Proteinkomponenten des Crosslinking-Produktes und damit
mögliche Proteine eines gemeinsamen Komplexes von Alb4 identifiziert werden. Dazu
wurden die mit sMBS vernetzten Thylakoidmembranen über eine SDS-PAGE aufgetrennt
und die Proteine mit Coomassie angefärbt. Die Coomassie gefärbte Proteinbande des
170 kDa großen Crosslinking-Produktes wurde entfärbt, mittels Trypsin verdaut und die
Peptide aus der Gelmatrix zum Zweck der Massenspektrometrie eluiert. Mit Hilfe des
massenspektrometrischen Verfahrens (in Kooperation mit PD Dr. Markus Piotrowski,
Ruhr-Universität Bochum) konnten die Proteinuntereinheiten CF1α und CF1β der
chloroplastidären ATP-Synthase in dem Crosslinking-Produkt identifiziert werden (Daten
nicht gezeigt). Unter Verwendung eines vorliegenden spezifischen Antikörpers wurde die
ATP-Synthase Untereinheit CF1β als Bestandteil des Crosslinking-Produktes bestätigt, da
die Western Blot Analyse ein spezifisches 170 kDa großes Crosslinking Signal in den mit
dem Crosslinking Reagenz sMBS behandelten Thylakoidmembranen zeigte. In der
Kontrollspur ohne Reagenz blieb das Signal auf der entsprechenden Höhe aus (Abbildung
4-18). Für die Kontrolle der Spezifität wurden die Crosslinking Ansätze zudem mit
Antikörpern gegen Alb3 und cpSecY behandelt. Die Western Blot Analysen zeigten in den
Spuren mit und ohne Crosslinker keine Signale auf der Höhe von 170 kDa (Abbildung
4-18). Die Proteine Alb3 und cpSecY waren somit keine Komponenten dieses vernetzten
170 kDa großen Proteinkomplexes. Mit Hilfe des chemischen Crosslinkings und dem
damit identifizierten Produktes wurde eine mögliche Assoziation von Alb4 mit der ATP-
Synthase gezeigt.
72
55
43
kDa
95
130170
sMBS - + - + - + - +
α-Alb4 α-CF1β α-Alb3 α-cpSecY
* *
Abbildung 4-18: Crosslinking von Alb4 und CF1β Thylakoidmembranen (100 µg Chlorophyll) wurden mit (+) und ohne (-) dem chemischen
Crosslinker sMBS (m-maleimidobenzoyl-N-hydroxysulfosuccinimide ester) (0,6 mM) inkubiert, über
eine SDS-PAGE aufgetrennt und unter Verwendung spezifischer Antikörper gegen Alb4, CF1β,
Alb3 und cpSecY in einer Immundetektion analysiert. (*spezifisches 170 kDa Crosslinking-Produkt)

4 Ergebnisse 97
4.3.2 Interaktion von Alb4 mit den kernkodierten ATP-Synthase Untereinheiten im Split-YFP System
Um die direkte Assoziation von Alb4 mit der ATP-Synthase zu untersuchen, wurde die
bimolekulare Fluoreszenzkomplementation (Split-YFP System) verwendet. Für diese
Technik wurden lediglich die kernkodierten Untereinheiten der ATP-Synthase (CF1γ, CF1δ
und CF0II) eingesetzt, da der Transport der Proteine in den Chloroplasten einem
posttranslationalen Transport entspricht. Die chloroplastidär kodierten Untereinheiten der
ATP-Synthase (CF1α, CF1β, CF1ε, CF0I, CF0III, CF0IV) werden cotranslational in die
Thylakoidmembran inseriert. Im Falle der Protoplastentransfektion müssten diese
Proteine über die chloroplastidäre Transitsequenz der kleinen Untereinheit der Rubisco
posttranslational in den Chloroplasten transportiert werden. Deshalb wurden diese
Untereinheiten im Split-YFP System nicht eingesetzt, da der Mechanismus des
cotranslationalen Transports nicht mehr gegeben wäre. Die Gensequenzen der
kernkodierten ATP-Synthase Untereinheiten (CF1γ, CF1δ und CF0II) wurden in den
pSPYNE-Vektor kloniert. Die Konstrukte pSPYCE-Alb4 und pSPYCE-Alb3 lagen bereits
aus anderen Messungen vor (Kapitel 4.2.2). Für die Interaktionsmessungen wurden
Arabidopsis Protoplasten mit den entsprechenden Konstrukten transient transfiziert und
die YFP-Fluoreszenz am Laserscanning Mikroskop analysiert. Im Fall der Expression, des
Imports in die Chloroplasten und des korrekten Einbaus in die Thylakoidmembranen
kommt es bei einer Proteininteraktion zur Rekonstitution des funktionalen YFP. Die
Messungen ergaben eine Interaktion der YFP-Fusionsproteine Alb4-cYFP und CF0II-
nYFP, wodurch die Fluoreszenz des YFP rekonstituiert werden konnte (Abbildung 4-19).
Die Überlagerung der YFP-Fluoreszenz mit der Autofluoreszenz des Chlorophylls zeigte
die Interaktion der beiden Proteine innerhalb der Chloroplasten. Als Kontrolle wurden die
Vektoren pSPYNE-TS(rbcs) und pSPYCE-TS(rbcs) eingesetzt und die Protoplasten mit
den entsprechenden Konstrukten cotransfiziert. Die Kontrollen zeigten, dass weder das
Fusionsprotein Alb4-cYFP noch das CF0II-nYFP die YFP-Fluoreszenz mit dem jeweiligen
einzelnen YFP-Fragment rekonstituieren konnte. Es zeigte sich jedoch keine Interaktion
von Alb4-cYFP mit den anderen getesteten ATP-Synthase Untereinheiten (CF1γ-nYFP
und CF1δ-nYFP). Im Vergleich zu dem Alb4 konnte das Alb3 mit keiner der getesteten
ATP-Synthase Untereinheiten interagieren (Daten nicht gezeigt).
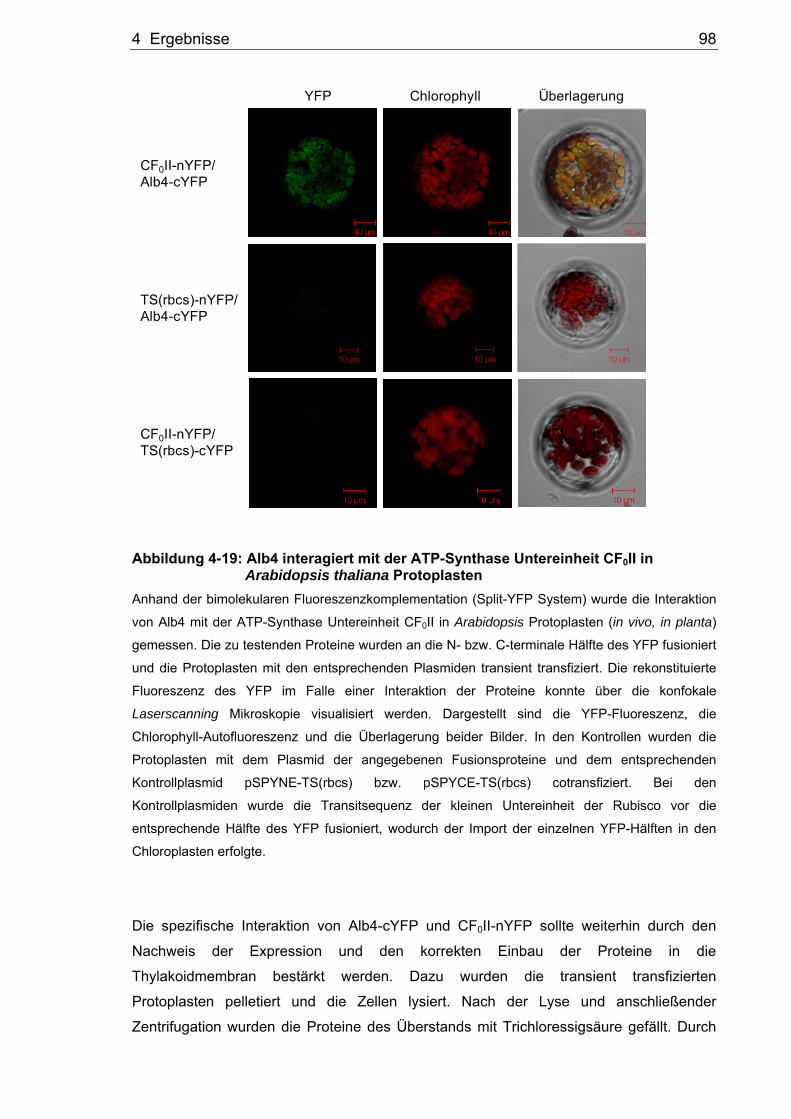
4 Ergebnisse 98
YFP Chlorophyll Überlagerung
CF0II-nYFP/Alb4-cYFP
TS(rbcs)-nYFP/Alb4-cYFP
CF0II-nYFP/TS(rbcs)-cYFP
Abbildung 4-19: Alb4 interagiert mit der ATP-Synthase Untereinheit CF0II in Arabidopsis thaliana Protoplasten
Anhand der bimolekularen Fluoreszenzkomplementation (Split-YFP System) wurde die Interaktion
von Alb4 mit der ATP-Synthase Untereinheit CF0II in Arabidopsis Protoplasten (in vivo, in planta)
gemessen. Die zu testenden Proteine wurden an die N- bzw. C-terminale Hälfte des YFP fusioniert
und die Protoplasten mit den entsprechenden Plasmiden transient transfiziert. Die rekonstituierte
Fluoreszenz des YFP im Falle einer Interaktion der Proteine konnte über die konfokale
Laserscanning Mikroskopie visualisiert werden. Dargestellt sind die YFP-Fluoreszenz, die
Chlorophyll-Autofluoreszenz und die Überlagerung beider Bilder. In den Kontrollen wurden die
Protoplasten mit dem Plasmid der angegebenen Fusionsproteine und dem entsprechenden
Kontrollplasmid pSPYNE-TS(rbcs) bzw. pSPYCE-TS(rbcs) cotransfiziert. Bei den
Kontrollplasmiden wurde die Transitsequenz der kleinen Untereinheit der Rubisco vor die
entsprechende Hälfte des YFP fusioniert, wodurch der Import der einzelnen YFP-Hälften in den
Chloroplasten erfolgte.
Die spezifische Interaktion von Alb4-cYFP und CF0II-nYFP sollte weiterhin durch den
Nachweis der Expression und den korrekten Einbau der Proteine in die
Thylakoidmembran bestärkt werden. Dazu wurden die transient transfizierten
Protoplasten pelletiert und die Zellen lysiert. Nach der Lyse und anschließender
Zentrifugation wurden die Proteine des Überstands mit Trichloressigsäure gefällt. Durch

4 Ergebnisse 99
die Waschschritte des Membranpellets mit 0,2 M NaOH sollten peripher gebundene
Proteine abgelöst werden. Der entstehende Überstand (Waschfraktion) wurde ebenfalls
einer TCA-Fällung unterzogen. Die Proteine des gewaschenen Membranpellets sowie die
Proben aus den TCA-Fällungen wurden über eine SDS-PAGE aufgetrennt und mittels
Immundetektion ausgewertet. Dabei konnte das nYFP-Fusionsprotein über den
angefügten c-myc-tag und das cYFP-Fusionsprotein über den HA-tag nachgewiesen
werden (Abbildung 4-20). Es zeigte sich, dass die YFP-Fusionsproteine Alb4-cYFP und
CF0II-nYFP ausschließlich in der Membranfraktion lokalisiert waren und nicht in den
löslichen Fraktionen vorlagen. Als Kontrollen wurden jeweils nicht transfizierte
Protoplasten aufgearbeitet. In diesen Proben waren keine Signale bei der entsprechenden
Größe der YFP-Fusionsproteine zu detektieren. Durch den Nachweis der Expression und
die korrekte Insertion der YFP-Fusionsproteine in die Membran konnte die Interaktion von
Alb4 und CF0II bekräftigt werden.
Alb4-cYFP
Kontrolle
Kontrolle
CF0II-nYFP
P
α-HA
α-c-myc
Ü W
P Ü W
Abbildung 4-20: Expressionsnachweis der YFP-Fusionsproteine Alb4-cYFP und CF0II-nYFP
Arabidopsis Protoplasten wurden mit den entsprechenden Konstrukten transient transfiziert und die
Expression der YFP-Fusionsproteine (Alb4-cYFP und CF0II-nYFP) über Immundetektion
nachgewiesen. Dazu wurden die lysierten Zellen abzentrifugiert und der Überstand (Ü) mittels
TCA-Fällung präzipitiert. Das Membranpellet wurde mit 0,2 M NaOH gewaschen (P), zentrifugiert
und der Überstand (W=Waschfraktion) mittels TCA-Fällung präzipitiert. Die Proben wurden über
eine SDS-PAGE aufgetrennt und über Immundetektion analysiert. Die Expression konnte über
einen anti-HA Antikörper (α-HA) für das Alb4-cYFP-Fusionsprotein und einen anti-c-myc Antikörper
(α-c-myc) für das CF0II-nYFP-Fusionsprotein gezeigt werden. Als Kontrolle wurden nicht
transfizierte Protoplasten eingesetzt (Kontrolle).

4 Ergebnisse 100
4.3.3 Gelfiltrationsanalyse solubilisierter Thylakoidmembranproteine
Um eine Assoziation von Alb4 mit der ATP-Synthase zu verifizieren, wurden die
Proteinkomplexe solubilisierter Thylakoidmembranen über eine Gelfiltrations-
chromatographie fraktioniert. Die Präsenz der Proteine Alb4, Alb3, cpSecY, CF1β, CF0II
und CF0III in den einzelnen Fraktionen wurde mittels Immundetektion über spezifische
Antikörper ausgewertet (Abbildung 4-21). Die Western Blot Analysen zeigten, dass der
überwiegende Teil von Alb4 mit den ATP-Synthase Untereinheiten CF1β, CF0II und CF0III
bei einem Molekulargewicht von 360-450 kDa von der Gelfiltrationssäule coeluierte.
Weiterhin konnte aus vorangegangenen Arbeiten (Klostermann et al., 2002) bestätigt
werden, dass die Alb3 und cpSecY beinhaltenden Proteinkomplexe mit einer Größe von
etwa 150-200 kDa von der Gelfiltrationsmatrix eluiert wurden. Diese Fraktionen
überlappten nur geringfügig mit den Alb4 Eluaten. Somit konnte gezeigt werden, dass
Alb4 mit der ATP-Synthase cofraktioniert und nur geringfügig mit Alb3 oder cpSecY.
~150 kDa ~15 kDaTM
~360 kDa
Alb3
cpSecY
Alb4
CF1β
~3600 kDa
CF0III
CF0II
Abbildung 4-21: Alb4 cofraktioniert mit den ATP-Synthase Untereinheiten Thylakoidmembranen (500 µg Chlorophyll) wurden mit 1,5 % Dodecyl-β-D-maltosid solubilisiert
und die Proteinkomplexe über eine Gelfiltrationssäule in 1 ml Fraktionen eluiert. Aliquots der
Fraktionen und solubilisierter Thylakoidmembranen wurden über eine SDS-PAGE aufgetrennt und
mittels spezifischer Antikörper gegen Alb4, Alb3, cpSecY, CF1β, CF0II und CF0III in einer
Immundetektion analysiert. (TM, solubilisierte Thylakoidmembranen)

4 Ergebnisse 101
4.3.4 Coimmunpräzipitation solubilisierter Thylakoidmembranproteine
Anhand von Coimmunpräzipitationsexperimenten sollte zunächst bestätigt werden, dass
Alb4 mit der ATP-Synthase assoziiert vorliegt und keine Komponente der Alb3 und
cpSecY beinhaltenden Proteinkomplexe ist. Dazu wurden solubilisierte Thylakoid-
membranen mit den an Protein-A-Sepharose gekoppelten Antikörpern α-Alb3, α-Alb4 und
α-cpSecY inkubiert. Die Präzipitate wurden über eine SDS-PAGE aufgetrennt und mittels
Immundetektion analysiert (Abbildung 4-22). Da die Präzipitate nicht vollständig frei von
den Antikörper IgGs waren, musste für die Präzipitation des Alb4 der α-Alb4 Antikörper
aus Kaninchen und für die Detektion des Alb4 der in dieser Arbeit bereits beschriebene
α-Alb4 Antikörper aus Ratte verwendet werden. Dieses war erforderlich, da die IgGs mit
einer Größe von etwa 55 kDa auf der gleichen Höhe wie das Alb4 liefen. Der Nachweis
mit dem Antikörper aus Kaninchen wäre somit nicht möglich gewesen, da die IgGs das
Alb4 bei einer Immundetektion überdecken würden. Die Herstellung eines spezifischen
α-Alb4 Antikörpers aus Ratte (oder einem anderen Organismus) war für die Methode der
Coimmunpräzipitation von entscheidender Bedeutung und Notwendigkeit.
TM IP
cpSecY
Alb3
Alb4
Abbildung 4-22: Coimmunpräzipitation von Alb3, Alb4 und cpSecY Thylakoidmembranen (500 µg Chlorophyll) wurden mit 1,5 % Digitonin solubilisiert und mit den an
Protein-A-Sepharose gekoppelten α-Alb3, α-Alb4 (Kaninchen) und α-cpSecY Antikörpern
immunpräzipitiert. Aliquots der Coimmunpräzipitate und solubilisierter Thylakoidmembranen
wurden über eine SDS-PAGE aufgetrennt und mittels Immundetektion analysiert. Der Nachweis
des Alb4 in den Präzipitaten erfolgte mit einem α-Alb4 Antikörper aus Ratte zur Vermeidung der
Detektion der Kaninchen IgGs auf Höhe des Alb4 Proteins. (IP: Immunpräzipitation, TM:
solubilisierte Thylakoidmembranen)

4 Ergebnisse 102
Die Western Blot Analysen zeigten, dass bei Verwendung des α-Alb3 Antikörpers das
Alb3 zusammen mit cpSecY copräzipitiert werden konnte. Gleiches gilt bei Einsatz des α-
cpSecY Antikörpers, wodurch die Coimmunpräzipitation der Proteine Alb3 und cpSecY
gezeigt werden konnte. Dieses Ergebnis ist eine Bestätigung der bereits in Klostermann
et al., 2002 veröffentlichten Daten. Weiterhin zeigte sich, dass weder Alb3 noch cpSecY
mit dem Alb4 unter Verwendung des α-Alb4 Antikörpers coimmunpräzipitiert werden
konnte. Auch die Präzipitate des α-Alb3 Antikörpers und α-cpSecY Antikörpers
beinhalteten kein Alb4 Protein. Dieses unterstützt die Aussage, dass die Proteine Alb3
und Alb4 in verschiedenen Thylakoidmembrankomplexen assoziiert sind. Weiterhin
musste allerdings festgestellt werden, dass die Analyse der Alb4 Präzipitate keine
Coimmunpräzipitation von Alb4 mit den ATP-Synthase Untereinheiten zeigte. Die
Assoziation von Alb4 mit der ATP-Synthase konnte daher mittels Coimmunpräzipitation
nicht bestätigt werden.

5 Diskussion 103
5. Diskussion
Die Mitglieder der Oxa1/YidC/Alb3-Proteinfamilie haben eine essentielle Bedeutung für
der Insertion und Assemblierung von Membranproteinen in Mitochondrien, Bakterien und
Chloroplasten (Kuhn et al., 2003; Xie und Dalbey, 2008). In der vorliegenden Arbeit sollte
die Rolle der chloroplastidären Homologe der Oxa1/YidC/Alb3-Proteinfamilie (Alb3 und
Alb4) bei der Biogenese der Thylakoidmembran von Arabidopsis thaliana analysiert
werden. Das integrale Thylakoidmembranprotein Alb3 ist für die posttranslationale
Insertion der Lichtsammelkomplexproteine (LHCPs) verantwortlich (Sundberg et al., 1997;
Moore et al., 2000). Zudem ist Alb3 mit der cpSec-Translokase assoziiert und scheint
daher an der cotranslationalen Insertion von plastidär kodierten Membranproteinen
beteiligt zu sein (Klostermann et al., 2002). Das Alb4 ist ebenfalls in der
Thylakoidmembran lokalisiert und hat einen Einfluss auf die Biogenese der Chloroplasten
(Gerdes et al., 2006). Die genaue Funktion von Alb4 ist jedoch noch nicht geklärt.
5.1 Die cpSRP-Untereinheit cpSRP43 interagiert direkt mit dem integralen Membranprotein Alb3
Bisherige Studien zum posttranslationalen cpSRP-Transportweg haben gezeigt, dass das
cpSRP43, cpSRP54, cpFtsY, GTP und die Translokase Alb3 für die Integration der
LHCPs in die Thylakoidmembran notwendig sind (Tu et al., 1999; Yuan et al., 2002). Das
cpSRP54 und der cpSRP-Rezeptor cpFtsY sind zusammen in der Lage mit dem
integralen Membranprotein Alb3 einen Komplex an der Thylakoidmembran zu bilden
(Moore et al., 2003). Moore et al., 2003 zeigten weiterhin, dass weder das Substrat LHCP,
noch das cpSRP43 für die Ausbildung dieses Komplexes nötig waren. Eine direkte
Interaktion zwischen cpSRP43 und Alb3 wurde durch die angewendeten Crosslinking
Studien nicht nachgewiesen. Damit wurde vermutet, dass das cpSRP43 für den docking-
Prozess des Transitkomplexes an die Thylakoidmembran keinen entscheidenden Einfluss
hat.
Einige Fragen, wie z. B. der docking-Prozess des Transitkomplexes an die
Thylakoidmembran, die Freisetzung und Insertion der LHCPs sowie die Regulation des
cpSRP-Transportwegs sind noch nicht ausreichend erforscht. In der vorliegenden Arbeit
sollte zunächst untersucht werden, ob der docking-Prozess des Transitkomplexes an die
Thylakoidmembran möglicherweise durch eine direkte Interaktion der löslichen Proteine
des cpSRP-Wegs mit der Translokase Alb3 vermittelt wird.
Die Ergebnisse der in vivo Interaktionsstudien über das Hefe Split-Ubiquitin System und
die bimolekulare Fluoreszenzkomplementation (Split-YFP System) zeigten, im Vergleich
zu den zuvor beschriebenen Beobachtungen aus Moore et al., 2003, eine direkte

5 Diskussion 104
Interaktion von cpSRP43 mit Alb3 (Kapitel 4.2.1 und 4.2.2). Dieses war sowohl in der
Hefe, als auch in der natürlichen Umgebung der Arabidopsis thaliana Protoplasten der
Fall. Es wurde ausschließlich eine Interaktion des cpSRP43 mit dem Alb3 nachgewiesen,
während das cpSRP54 und cpFtsY mit Alb3 keine Interaktion zeigten (Kapitel 4.2.1 und
4.2.2). In Kooperation mit der Arbeitsgruppe von R. Goforth (University of Arkansas)
wurden diese Daten in einer gemeinsamen Publikation zusammengestellt (Marty et al.,
2009, eingereicht). Ergebnisse aus der Arbeitsgruppe Goforth zeigten eine Copräzipitation
von rekombinantem cpSRP43 und Alb3. Diese Versuche erfolgten mit
Thylakoidmembranen aus Pisum sativum (Erbse), so dass es sich bei dem
copräzipitierten Alb3 um das Homolog PPF1 aus der Erbse handelte. Die Ansätze mit
rekombinantem cpSRP54 und cpFtsY zeigten sowohl einzeln, als auch in Kombination
kein copräzipitiertes Alb3 (PPF1). In den Ansätzen mit einer Kombination der
rekombinanten Proteine konnte nur in den Proben das Alb3 (PPF1) copräzipitiert werden,
in denen auch das cpSRP43 vorlag. Das cpSRP43 könnte somit als Brücke innerhalb der
cpSRP-Komponenten im Verlauf des docking-Prozesses an das Alb3 wirken.
Im Laufe dieser vorliegenden Doktorarbeit sind weitere Daten publiziert worden, die eine
Komplementation einer cpSRP43-defizienten chaos-Mutante von Arabidopsis thaliana mit
einem cpSRP43(ΔCD2) Protein zeigten (Tzvetkova-Chevolleau et al., 2007). Das
cpSRP43(ΔCD2) ist nicht mehr in der Lage mit dem cpSRP54 zu interagieren und einen
Transitkomplex mit dem cpSRP54 und LHCP zu bilden (Goforth et al., 2004). Aufgrund
der Komplementation wurde vermutet, dass cpSRP43 unabhängig von cpSRP54 den
LHCP-Transport vermitteln kann. Weiterhin wurde durch Immunlokalisationsstudien eine
Colokalisation von cpSRP43 mit der Translokase Alb3 gezeigt, die allerdings nur in
Abwesenheit von cpSRP54 zu beobachten war (Tzvetkova-Chevolleau et al., 2007). Die
direkte Interaktion der beiden Proteine wurde durch eine Copräzipitation des Alb3 mit
rekombinantem cpSRP43 gezeigt (Tzvetkova-Chevolleau et al., 2007). Die Autoren
vermuten daher einen alternativen Transportweg der LHCPs über cpSRP43 und Alb3, der
unabhängig von cpSRP54 und cpFtsY verläuft.
Pull down Analysen aus der vorliegenden Doktorarbeit zeigten allerdings, dass cpSRP43
sowohl allein, als auch im Komplex mit cpSRP54, mit Alb3 interagierte. Dieses Ergebnis
könnte einerseits die Möglichkeit eines alternativen Transportwegs unterstützen, bei dem
das cpSRP43 auch unabhängig von cpSRP54 den LHCP-Transport zur Translokase Alb3
vermittelt (Tzvetkova-Chevolleau et al., 2007). Andererseits ist aber bemerkenswert, dass
cpSRP43 auch im Komplex mit cpSRP54 und nicht nur in Abwesenheit des cpSRP54 mit
Alb3 interagierte.

5 Diskussion 105
5.2 Identifizierung der Alb3-Interaktionsbereiche für das cpSRP43 und Charakterisierung der Alb3-cpSRP43 Interaktion
Weiterführende Experimente sollten die Interaktionsdomäne des Alb3 für das cpSRP43
ermitteln. Als erster Ansatzpunkt sollten durch die Inkubation von salzgewaschenen und
proteaseverdauten Thylakoidmembranen mit den Proteinen cpSRP43, cpSRP54 und
cpFtsY mögliche stromale Bereiche des Alb3 als Interaktionsdomäne identifiziert werden
(Marty et al., 2009, eingereicht). Es zeigte sich, dass rekombinantes cpSRP43 und cpFtsY
an salzgewaschene Thylakoidmembranen von Arabidopsis thaliana binden konnten,
während eine Bindung des Kontrollproteins GST ausblieb. Ein Proteaseverdau der
Membranen führte zu einem vollständigen Verlust der Bindung des cpSRP43. Das cpFtsY
konnte jedoch, aufgrund seiner Membranaffinität, an die mit Protease behandelten
Thylakoidmembranen binden (Marty et al., 2009). Diese Ergebnisse wurden durch die
Bindungsversuche von radioaktiv markiertem cpSRP43, cpSRP54 und cpFtsY an
Thylakoidmembranen aus der Erbse bestätigt. Der Proteaseverdau der Membranen
bewirkte eine Abspaltung des stromalen C-Terminus von Alb3 (PPF1) (Marty et al., 2009,
eingereicht). Diese Ergebnisse deuteten darauf hin, dass die Bindung des cpSRP43 an
die Thylakoidmembranen über den stromalen C-terminus des Alb3 erfolgen könnte.
Weitere Daten aus der vorliegenden Doktorarbeit und der Kooperation (Marty et al., 2009,
eingereicht) zeigten, dass der C-Terminus von Alb3 tatsächlich direkt mit dem cpSRP43
interagierte, während keine Interaktion zwischen cpSRP54 und cpFtsY mit Alb3
nachgewiesen wurde. Ausführliche pull down Versuche mit rekombinantem cpSRP43 und
rekombinantem Alb3 C-Terminus (PPF1) bestätigten die Bedeutung des C-Terminus von
Alb3 für die Interaktion mit cpSRP43 (Marty et al., 2009, eingereicht). Die Messung der
Interaktion von cpSRP43 und dem C-Terminus von Alb3 mittels isothermaler
Titrationskalorimetrie bekräftigten, dass das cpSRP43 direkt mit dem löslichen C-
Terminus von Alb3 interagierte (Marty et al., 2009, eingereicht).
Ein Vergleich innerhalb der Oxa1/YidC/Alb3-Proteinfamilie zeigt, dass der C-Terminus für
die anderen homologen Proteine ebenfalls eine entscheidende Bedeutung als funktionale
Domäne hat. Das bakterielle Homolog YidC und das mitochondriale Homolog Oxa1 sind
in der Lage mit ihrem C-Terminus Ribosomen zu binden und damit die cotranslationale
Insertion von Membranproteinen zu ermöglichen (Szyrach et al., 2003; Kohler et al.,
2009). Es wird vermutet, dass das cpSRP43 im posttranslationalen cpSRP-Transportweg
anstelle von Ribosomen an das Alb3 bindet. Weitere Arbeiten zeigten, dass Alb3 mit der
cpSec-Translokase assoziiert ist und bei der cotranslationalen Insertion des plastidär
kodierten D1 beteiligt ist (Klostermann et al., 2002; Ossenbühl et al., 2004). Demnach
bleibt zu klären, in welcher Form und durch welchen Mechanismus sich der co- und
posttranslationale Import der Proteine über Alb3 in die Thylakoidmembran unterscheidet

5 Diskussion 106
und reguliert wird.
Der Nachweis der Interaktion von cpSRP43 und Alb3 stellte die Frage nach der
funktionellen Bedeutung und physiologischen Relevanz dieser Interaktion innerhalb des
posttranslationalen cpSRP-Transportwegs. Bisherige Arbeiten wiesen einen Einfluss der
GTP-Bindung und Hydrolyse der beiden GTPasen cpSRP54 und cpFtsY bei der
Integration der LHCPs nach (Tu et al., 1999; Moore et al., 2003). Zudem wurde ein
regulierender Effekt des cpSRP43 auf die GTP-Hydrolyse der GTPasen gezeigt (Goforth
et al., 2004). In Marty et al., 2009 (eingereicht) wird weiterhin gezeigt, dass das cpSRP43
die GTPase Aktivität des cpSRP54 und cpFtsY in Abhängigkeit von dem Alb3 C-Terminus
(PPF1) stimulierte. Daher wurde vermutet, dass die GTP-Hydrolyse innerhalb des cpSRP-
Transportwegs nur bei Anwesenheit der Translokase Alb3 erfolgt und dass die GTP-
Hydrolyse ein entscheidender Schritt für das Recycling der cpSRP-Komponenten ist. Das
cpSRP43 könnte damit als Vermittler zwischen dem Alb3, cpSRP54, cpFtsY und LHCP
dienen und die Interaktion zwischen cpSRP43 und Alb3 könnte eine regulatorische
Funktion an der Thylakoidmembran einnehmen.
Die Ergebnisse der vorliegenden Doktorarbeit deuten darauf hin, dass neben dem C-
Terminus noch die 5. Transmembrandomäne des Alb3 für eine Interaktion mit dem
cpSRP43 wichtig ist. Dieses wurde zunächst durch die bimolekulare
Fluoreszenzkomplementation gezeigt (Kapitel 4.2.3). Die Existenz einer neben dem C-
Terminus zusätzlichen Bindestelle für das cpSRP43 wurde durch das Peptide Scanning
unterstützt (Kapitel 4.2.4). So umfasste ein Cluster der detektierten PepSpots die
Aminosäuren 305-328 der 5. Transmembrandomäne von Alb3. Diese Spots zeigten eine
gemeinsame Sequenz der Aminosäuren 314-319 (LVFKFL). Die Identifizierung dieser
Aminosäuren war eine Bestätigung der Messung der Alb3-Alb4 overlap-Konstrukte im
Split-YFP System, bei der das Bindemotiv LVFKF (Aminosäuren 314-318) innerhalb des
Alb3 eine wesentliche Bedeutung für die Interaktion mit cpSRP43 hatte. Dieses Motiv
wurde weiterhin durch die Deletionen und Mutationen in diesem Sequenzmotiv des Alb3
als Bindestelle für das cpSRP43 im Split-Ubiquitin System und Split-YFP System bestätigt
(Kapitel 4.2.5). Erstaunlicherweise liegt diese Bindestelle auf der lumenalen Seite der
5. Transmembrandomäne von Alb3. Daher stellt sich die Frage nach der Zugänglichkeit
dieser Bindestelle für das stromale cpSRP43 innerhalb des cpSRP-Transportwegs.
Kürzlich veröffentlichte Daten haben gezeigt, dass die homologen Proteine YidC und
Oxa1 in Anwesenheit von translatierenden Ribosomen möglicherweise eine Pore bilden
(Kohler et al., 2009). Die Interaktion des YidC und Oxa1 mit den Ribosomen erfolgt über
den jeweiligen C-Terminus. Mit Hilfe von BN-PAGE und Crosslinking Studien wurde
gezeigt, dass die Ribosomen die dimere Form von YidC und Oxa1 stabilisieren. Weitere
Analysen ergaben, dass die Dimere von YidC und Oxa1 über dem Ausgang des
Ribosomentunnels lokalisiert vorliegen.

5 Diskussion 107
Mit der Existenz des Bindemotivs innerhalb der 5. Transmembrandomäne des Alb3
könnte ein Mechanismus vermutet werden, bei dem das Alb3 für den docking-Prozess
des Transitkomplexes und die Insertion der LHCPs in die Thylakoidmembran eine Pore
ausbildet. Die Topologie des Alb3 beruht jedoch bisher nur auf Strukturvorhersagen.
Daher kann nicht ausgeschlossen werden, dass die Bindestelle möglicherweise bei einer
abweichenden Struktur des Alb3 innerhalb der Thylakoidmembran für die Interaktion mit
dem cpSRP43 auch ohne Ausbildung einer Pore zugänglich wäre.
Die Ergebnisse der Messungen im Split-YFP ergaben außerdem, dass der C-Terminus
von Alb3, wie bereits beschrieben, für eine Interaktion mit dem cpSRP43 essentiell ist
(Kapitel 4.2.3). Über die Methode des Peptide Scanning wurden die Aminosäuren 374-
388 des Alb3 C-Terminus als Bindestelle für das cpSRP43 identifiziert (Kapitel 4.2.4). Die
Deletion dieser Bindestelle innerhalb des Alb3 führte sowohl im Split-YFP System, als
auch im Split-Ubiquitin System zum Verlust der Interaktion mit cpSRP43 (Kapitel 4.2.5
und 4.2.6). Die Bindestelle innerhalb des C-Terminus von Alb3 wurde durch den Einsatz
eines Alb3-Peptids (Aminosäuren 367-388), welches die Aminosäuren der potentiellen
Bindestelle (374-388) umfasste, bestätigt, da die Vorinkubation des cpSRP43 mit dem
Alb3-Peptid zu einer verringerten Bindung des cpSRP43 an die Thylakoidmembranen
führte (Kapitel 4.2.8).
Um zu untersuchen, ob die beiden Bindestellen des Alb3 für das cpSRP43 konserviert
vorliegen, wurde ein Alignment der Alb3 Homologe einiger Streptophyten in diesen
Sequenzbereichen durchgeführt (Abbildung 5-1). Phylogenetische Analysen (Zhang et al.,
2009) zeigten, dass während der Entwicklung der Landpflanzen von den Grünalgen eine
Kopie des Alb3 Gens verloren gegangen ist und eine Duplikation der anderen Kopie zu
zwei Genen geführt hat, welche die beiden Proteine Alb3 und Alb4 kodieren. In Populus
trichocarpa wurden diese beiden Gene erneut dupliziert, so dass es dort
dementsprechend vier Gene, zwei für Alb3 und zwei für Alb4, gibt.
Die identifizierte Bindestelle innerhalb der 5. Transmembrandomäne des Alb3 aus
Arabidopsis thaliana umfasst die Aminosäuren 314-318 (LVFKF). In dem
Sequenzvergleich der Alb3 Homologe dieser Streptophyten fällt auf, dass das Leucin (L)
an Position 314 sehr konserviert vorliegt (Abbildung 5-1). In dieser Arbeit wurde gezeigt,
dass Mutationen und Deletionen dieser Aminosäure des Alb3 in einem Verlust der
Bindung des cpSRP43 resultierten, während z. B. die Mutation des Phenylalanin an
Position 316 (F), welches innerhalb der Alb3 Proteine dieser Streptophyten nicht
konserviert vorliegt, keine Auswirkungen hatte. Die Aminosäuren Lysin und Phenylalanin
an Position 317 und 318 (KF) liegen ebenfalls sehr konserviert vor. Da diese
Aminosäuren allerdings auch in der Alb4 Sequenz vorkommen (Abbildung 5-2) und Alb4
keine Interaktion mit cpSRP43 zeigte, scheinen diese Aminosäuren innerhalb des LVFKF-

5 Diskussion 108
Motivs keine essentielle Bedeutung für die Bindung des cpSRP43 zu haben. Demzufolge
scheint in dem ersten Bindemotiv die Aminosäure Leucin an Position 314 für die Bindung
des cpSRP43 von besonderer Bedeutung zu sein.
314-318 Arabidopsis_t._Alb3 HPPLGWYDTVAYLVLPVLLIASQYVSMEIMKPPQTDDPAQKNTLLVFKFL Oryza_s._Alb3 HPPLGWHDTICYLVLPVLLVASQFVSMEIMKPPQTDDPSQKNTLLVLKFL Pisum_s._Alb3 HPLLGWYDTAAYLVLPVLLIVSQYVSMEIMKPPQTNDPNQKNTLLIFKFL Populus_t._Alb3_a-1 HPPLGWHDTAAYLVLPVLLIASQYVSMEIMKPPQTDDPTQKNTLLVFKFL Populus_t._Alb3_a-2 HPPLGWNDTAAYLVLPVLLVVSQYVSMEIMKPPQTDDPTQKNTLLVFKFL Selaginella_m._Alb3 HPPLGWGDTIAYLVLPVLLIGSQYASMQIMQPP--QDPSQKNTQLILKFL Physcomitrella_p._Alb3 APSLGWADTIAYLVLPVLLIGSQYVSMQIMQPPTTNDPAQNQSQLILKFL * *** ** .********: **:.**:**:** :** *::: *::*** Arabidopsis_t._Alb3 PLMIGYFALSVPSGLSIYWLTNN--VLSTAQQVYLRKLGGAKPNMDENAS Oryza_s._Alb3 PFMIGWFSLSVPSGLSIYWFTNN--ILSTAQQVWLRKLGGAKPVVNQGGS Pisum_s._Alb3 PLMIGYFSLSVPSGLTIYWFTNN--VLSTAQQVWLRKLGGAKPAVNENAG Populus_t._Alb3_a-1 PIMIGYFSLSVPSGLSIYWFTNN--VLSTAQQVWLRKLGGAKPVVNENAS Populus_t._Alb3_a-2 PLMIGYFSLSVPSGLSIYWFTNN--VLSTAQQVWLRQLGGAKPVVNENAS Selaginella_m._Alb3 PLMIGYFALSVPSGLSLYWLSLTNNILSTGQQVWLRKLGGAKPVVASGGG Physcomitrella_p._Alb3 PLMIGYFSLSVPSGLSLYWLTNN--VLSTAQQVYLRQLGGAQPIVDKEGS *:***:*:*******::**:: . :***.***:**:****:* : . .. 374-388 _____ Arabidopsis_t._Alb3 KIISAGRAKRSIAQPDDAGERFRQLKEQEKRSKKNKAVAKDTVELVEESQ Oryza_s._Alb3 GIITAGRAKRTSAQPAQPGERFKQLKEEESKRKGNKALAAGDSDLSASTS Pisum_s._Alb3 GIITAGQAKRSASKPEKGGERFRQLKEEEKKKKLIKALPVEEVQPLASAS Populus_t._Alb3_a-1 GIITAGRAKRSASQPGQPGDRFKQLKEQDKSKTLRKALPTEGVQALDSAS Populus_t._Alb3_a-2 GIITAGRAKRSAAQPGQPGDRFRLK-EEEKGKKLSKALQSEDVQALDSAS Selaginella_m._Alb3 DIISAGQAKRSTPSP--PAARFRQLKAAESKRKAERA------------- Physcomitrella_p._Alb3 GIISAGQAKRTPLPPS-ASTSATQLKEEDAKKKGSRRKSAEEASKAAAAK **:**:***: * . : . : Arabidopsis_t._Alb3 ----SESEEGSDDEEEEAREGALASSTTSKPLPEVGQRRSKRSKRKRTV Oryza_s._Alb3 EDEESDDETTEEGGPEERYNSSSNKKLPNYSGKKGKRSKRKRMVQ Pisum_s._Alb3 ASNDGSDVENNK--EQEVTEESNTSKVSQEVQSFSRERRSKRSKRKPVA Populus_t._Alb3_a-1 GSDEDSDEETNDKGEEVLEETYASS-ASKRVPDISRPKRSKRSKRKRTV Populus_t._Alb3_a-2 --DEDSDEETKDKGEEVLEEAYASSSASKKVSDISRPKRSKRSKRKRAV Selaginella_m._Alb3 ------------------------------------------------- Physcomitrella_p._Alb3 GAEERDN------------------------------------------
Abbildung 5-1: Alignment der C-terminalen Proteinsequenzen der Alb3 Homologe Die Abbildung zeigt ein Alignment der Proteinsequenzen der Alb3 Homologe im Bereich der 5.
Transmembrandomäne und des C-Terminus von Alb3 aus Arabidopsis thaliana. Die konservierten
Aminosäuren der Bindemotive im Bereich der Aminosäuren 314-318 und 374-388 sind in rot
dargestellt. Der Bereich der 5. Transmembrandomäne des Alb3 aus Arabidopsis thaliana ist grau
hinterlegt.
Die zweite Bindestelle im C-Terminus umfasst die Aminosäuren 374-388. Innerhalb dieser
identifizierten Bindestelle konnte ein konserviertes Motiv der Aminosäuren 375-377 (AKR)
gefolgt von einem Prolin an Position 382 innerhalb der gezeigten Alb3 Homologe der
Streptophyten erkannt werden (Abbildung 5-1). Interessanterweise ist dieser Bereich der
Bindestelle (Aminosäuren 374-388) und der etwas verlängerte Bereich des eingesetzten
synthetischen Peptids (Aminosäuren 367-388), welches zu einer Verringerung der
Bindung des cpSRP43 an die Thylakoidmembran führte, im Vergleich zu dem restlichen

5 Diskussion 109
C-Terminus dieser Streptophyten sehr homolog und konserviert. Diese Tatsache
bekräftigt somit die Bindestelle innerhalb des C-Terminus, da funktionelle Domänen
allgemein sehr konserviert vorliegen.
Das Alignment der Alb3 Sequenz aus Arabidopsis thaliana mit homologen Alb4 Proteinen
der Streptophyten zeigt, dass beide Motive der identifizierten Bindestellen für das
cpSRP43 in den Alb4 Proteinen nicht konserviert vorliegen (Abbildung 5-2). Dieses
unterstützt weiterhin das Ergebnis dieser Doktorarbeit, dass Alb4 nicht mit cpSRP43
interagierte. Dieses ist möglicherweise auf das Fehlen dieser in den Alb3 Proteinen
konservierten Bereiche zurückzuführen.
314-318 Arabidopsis_t._Alb3 MKPPQTDDPAQKNTLLVFKFLPLMIGYFALSVPSGLSIYWLTNNVLSTAQ Arabidopsis_t._Alb4 MQSSQSNDPAMKSSQAVTKLLPLMIGYFALSVPSGLSLYWLTNNILSTAQ Oryza_s._Alb4 MQPPQNNDPSQQGAQAVVKFLPLLIGYFALSVPSGLSLYWLTNNILSTAQ Populus_t._Alb4_b-1 MQSSQSDDPNVKNSQAIMKFLPLMIGYFSLSVPSGLSLYWLTNNILSTTQ Populus_t._Alb4_b-2 MQSSQSDDPNVKNSQAITKFLPLMIGYFSLSVPSGLSLYWLTNNILSTAQ *:..*.:** :.: : *:***:****:********:******:***:* Arabidopsis_t._Alb3 QVYLRKLGGAKPNMDEN---------------------------ASKIIS Arabidopsis_t._Alb4 QVWLQKYGGAKNPVEKFTNLVTKEDKTQQIEKSFSEPLVQKSVSELKIPR Oryza_s._Alb4 QVWLQKLGGAKNPVKEYIDKLAKEESTN----LGKPEPAIKSDPLPKVGK Populus_t._Alb4_b-1 QVWLQKLGGAK--------------------------------------- Populus_t._Alb4_b-2 QVWLQKSGGAKNPMMKSSDDIV---------------------------- **:*:* **** 374-388 _____ Arabidopsis_t._Alb3 AGRAKRSIAQPDDAGERFRQLKEQEKRSKK--------NKAVAKDTVELV Arabidopsis_t._Alb4 EKGGEKVTPECPKPGERFRLLKEQEAKRRREKEERQKAEAALSNQNTDKA Oryza_s._Alb4 PPASQEPEPSGPQRGERFRKLKEEESRRKVFLEKAEQTEQAGTQAGIVDG Populus_t._Alb4_b-1 -------------------------------------------------- Populus_t._Alb4_b-2 -------------------------------------------------- Arabidopsis_t._Alb3 EESQSESEEGSDDEEEEAREGALASSTTSKPLPEVGQRRSKRSKRKRTV- Arabidopsis_t._Alb4 HEQDEKSDTAIVAEDDKKTELSAVDETSDGTVAVNGKPSIQKDETTNGTF Oryza_s._Alb4 KQNSDASGDNIDEQESHENEP--IIANGNGGLSHSTNEMIPNGSMKEVCF Populus_t._Alb4_b-1 -------------------------------------------------- Populus_t._Alb4_b-2 --------------------------------------------------
Abbildung 5-2: Alignment des Alb3 aus Arabidopsis thaliana mit den Alb4 Proteinen anderer Streptophyten
Die Abbildung zeigt ein Alignment der Proteinsequenz des Alb3 aus Arabidopsis thaliana im
Bereich der 5. Transmembrandomäne und des C-Terminus mit den Alb4 Homologen anderer
Streptophyten. Die konservierten Aminosäuren der Bindemotive des Alb3 im Bereich der
Aminosäuren 314-318 und 374-388 sind in rot dargestellt. Der Bereich der
5. Transmembrandomäne des Alb3 aus Arabidopsis thaliana ist grau hinterlegt.

5 Diskussion 110
5.3 Das cpSRP43 interagiert mit Alb3 über die Ankyrin repeat Region
Das cpSRP43 besteht aus einer N-terminalen Chromodomäne (CD1), gefolgt von vier
Ankyrin repeats (Ank1-4) und zwei C-terminalen Chromodomänen (CD2, CD3). Die
Bestimmung der Region des cpSRP43, die mit Alb3 interagiert, wurde ebenfalls in
Kooperation mit der Arbeitsgruppe von R. Goforth (University of Arkansas) durchgeführt.
In Marty et al., 2009 (eingereicht) wurden pull down Versuche mit verschiedenen GST-
cpSRP43 Deletionskonstrukten und rekombinantem Alb3 C-Terminus (PPF1)
durchgeführt. Die Ergebnisse zeigten, dass die Existenz der Ankyrin repeats im cpSRP43
für die Coreinigung des Alb3 unbedingt erforderlich war. Bei den getaggten
Fusionsproteinen des cpSRP43, in denen die Ankyrin repeat Region deletiert war, wurde
eine starke Verringerung des cogereinigten Alb3 beobachtet. Die Interaktion des Alb3 C-
Terminus mit der Ankyrin repeat Region des cpSRP43 wurde durch weitere pull down
Analysen bestätigt. Dabei wurden diejenigen cpSRP43 Deletionsproteine, die noch die
Ankyrin repeats beinhalteten über den getaggten Alb3 C-Terminus cogereinigt. Fehlten
die Ankyrin repeats innerhalb der cpSRP43 Proteine, blieb die Coreinigung über das
rekombinante Alb3 aus. Auffällig war allerdings auch, dass bei Deletion der
Chromodomäne 2 eine leichte Verringerung der Bindung zu erkennen war (Marty et al.,
2009, eingereicht).
Die Ergebnisse der pull down Analysen wurden mit Hilfe der isothermalen
Titrationskalorimetrie bestätigt. Die Messung der Bindungsaffinitäten zwischen den
cpSRP43 Deletionsproteinen und dem Alb3 waren abhängig von dem Vorhandensein der
Ankyrin repeat Region innerhalb des cpSRP43. Zusammenfassend besagten diese
Ergebnisse, dass die Ankyrin repeat Region und möglicherweise unterstützend die
Chromodomäne 2 die Bindestelle für den Alb3 C-Terminus darstellten.
Ergänzende Versuche zeigten, dass das Deletionsprotein cpSRP43(Ank1-CD2), ebenso
wie das Volllängen cpSRP43, einen stimulierenden Einfluss auf die GTP-Hydrolyse der
GTPasen cpSRP54 und cpFtsY hatte (Marty et al., 2009, eingereicht). Der Einsatz
weiterer Deletionsproteine von cpSRP43 zeigte, dass bei Deletion der Chromodomäne 2
des cpSRP43 die Stimulation der GTP-Hydrolyse in Kombination mit dem Alb3 verloren
ging. Dieser Effekt ist in dem Verlust der Bindedomäne des cpSRP43 für das cpSRP54
begründet, welche durch die Chromodomäne 2 gestellt wird (Sivaraja et al., 2005;
Hermkes et al., 2006; Kathir et al., 2008). Die Interaktion von cpSRP43 und Alb3 konnte
somit keinen Einfluss auf die GTPasen nehmen. In bisherigen Arbeiten wurde bereits
beschrieben, dass die Chromodomäne 1 einen negativ regulatorischen Effekt auf die
GTP-Hydrolyse der GTPasen des cpSRP-Transportwegs besitzt (Goforth et al., 2004).
Demzufolge führte ein Verlust der Chromodomäne 1 des cpSRP43 bereits in Abwesenheit
des Alb3 zu einem Anstieg der GTPase Aktivität des cpSRP54 und cpFtsY (Marty et al.,

5 Diskussion 111
2009, eingereicht). Die Deletion der Chromodomäne 3 hatte keinen Effekt auf die GTP-
Hydrolyse der cpSRP GTPasen (Marty et al., 2009, eingereicht). Demnach zeigte sich,
dass die Ankyrin repeat Region in Verbindung mit der Chromodomäne 2 des cpSRP43
ausreichte, um die GTPase Aktivität in Abhängigkeit des Alb3 C-Terminus zu stimulieren.
In einer Masterarbeit, die in der Arbeitsgruppe von Prof. Dr. D. Schünemann parallel zu
dieser vorliegenden Doktorarbeit und unabhängig von unserem späteren
Kooperationspartner (Marty et al., 2009, eingereicht) angefertigt wurde, konnte die
Interaktionsdomäne des cpSRP43 für das Alb3 bereits identifiziert werden (Vörös, 2007).
Ein Thylakoidmembran-Bindungstest ergab, dass Ankyrin repeat 4 eine wesentliche
Bedeutung für die Bindung des cpSRP43 an die Membranen haben könnte. Die Deletion
der Ankyrin repeat Region 4 bewirkte einen Verlust der Membranbindung des cpSRP43.
In der vorliegenden Doktorarbeit wurde ergänzend bestätigt, dass spezifisch die Ankyrin
repeat Region 4 für die Membranbindung wichtig ist, da die Deletion der anderen Ankyrin
repeats keinen Einfluss auf die Membranbindung des cpSRP43 zeigte (Kapitel 4.2.9). Die
cpSRP43 Deletionsproteine interagierten trotz des Verlusts der einzelnen Ankyrin repeats
1-3 mit den Thylakoidmembranen.
Um die Spezifität der Ankyrin repeat Region 4 des cpSRP43 für die Bindung an die
Thylakoidmembran zu bestätigen, wurden Proteine erzeugt, die die einzelnen Ankyrin
repeats in Verbindung mit der Chromodomäne 2 beinhalteten. Vorangegangene Arbeiten
zeigten, dass das Protein GST-cpSRP43(Ank4+CD2) noch für eine Interaktion mit der
Thylakoidmembran ausreichte (Vörös, 2007). In der vorliegenden Arbeit zeigte sich, dass
der Austausch der Ankyrin repeat Region 4 gegen die anderen Ankyrin repeats 1-3 einen
Verlust der Bindefähigkeit der Proteine an die Thylakoidmembranen bedeutete. Die
Proteine GST-cpSRP43(Ank1+CD2), GST-cpSRP43(Ank2+CD2) und GST-
cpSRP43(Ank3+CD2) waren nicht mehr in der Lage an die Membranen zu binden, was
die Spezifität der Ankyrin repeat Region 4 bestätigte.
Die Erstellung der Konstrukte GST-cpSRP43(Ank1+CD2), GST-cpSRP43(Ank2+CD2),
GST-cpSRP43(Ank3+CD2) und GST-cpSRP43(Ank4+CD2) erfolgte anhand von
Strukturvorhersageprogrammen. Ein Vergleich der Vorhersagen mit der kürzlich
publizierten Kristallstruktur des cpSRP43 (Stengel et al., 2008) ergab, dass die α-Helices
der Ankyrin repeats 1, 2 und 3 innerhalb der vorhergesagten Sequenz liegen. Die Ankyrin
repeat Region 4 besitzt mit der α-Helix 8 einen zur Chromodomäne 2 verlängerten
Bereich, der bisher als linker Sequenz angenommen wurde (Abbildung 4-15). Die
Konstrukte GST-cpSRP43(Ank1+CD2), GST-cpSRP43(Ank2+CD2), GST-
cpSRP43(Ank3+CD2) und GST-cpSRP43(Ank4+CD2) wurden so erstellt, dass auf die
Ankyrin repeats zunächst der linker und dann die Chromodomäne 2 folgten. Das
bedeutet, dass die Ankyrin repeats 1-3 jeweils um einen Teil der α-Helix 8 der Ankyrin
repeat Region 4 verlängert waren, was die Proteine allerdings nicht dazu befähigte, an die

5 Diskussion 112
Thylakoidmembranen zu binden. Der Einbau dieses zusätzlichen Teils der α-Helix 8
könnte allerdings einen strukturellen Einfluss auf die Deletionsproteine haben. Schließlich
lässt sich festhalten, dass die Binderegion des cpSRP43 für das Alb3 in der
Thylakoidmembran möglicherweise auf die Ankyrin repeat Region 4 einzugrenzen ist.
LHCP
Alb3TMD
TMD L-18 Motiv
cpSRP43 A1 A2 A3 A4 CD3
cpSRP54 G-Domäne M-Domäne
Bindemotiv
CD2CD2CD1
Abbildung 5-3: Proteininteraktionen des cpSRP43 Das cpSRP43 besitzt drei Chromodomänen (CD1-3) und vier Ankyrin repeats (A1-4). Das
cpSRP54 besteht aus einer G-Domäne, welche für die GTPase Aktivität zuständig ist, und einer
methioninreichen M-Domäne. Bei der Heterodimerisierung des cpSRP kommt es zur Interaktion
der Chromodomäne 2 mit einem 10 Aminosäuren langen Bindemotiv innerhalb der M-Domäne des
cpSRP54. Das LHCP besitzt drei Transmembrandomänen (TMD). Es interagiert über das L-18
Motiv mit der Ankyrin repeat Region des cpSRP43. Die Ergebnisse dieser Doktorarbeit zeigen eine
Interaktion des Alb3 mit cpSRP43. Als Bindedomänen sind die 5. Transmembrandomäne und der
C-Terminus des Alb3 sowie die Ankyrin repeat Region 4 des cpSRP43 identifiziert worden.
Das cpSRP43 scheint über die einzelnen Interaktionsdomänen eine Vielzahl von
Funktionen sowohl im Stroma, als auch an der Thylakoidmembran innerhalb des
posttranslationalen cpSRP-Transportwegs einzunehmen (Abbildung 5-3). Bisherige Daten
zeigten, dass die Heterodimerisierung des cpSRP-Komplexes auf einer Interaktion
zwischen der α-Helix in Chromodomäne 2 des cpSRP43 und einem 10 Aminosäuren
langen Bindemotiv innerhalb der M-Domäne des cpSRP54 beruht (Goforth et al., 2004;
Funke et al., 2005; Hermkes et al., 2006; Kathir et al., 2008). Die Bildung des

5 Diskussion 113
Transitkomplexes erfolgt durch Interaktion der Ankyrin repeat Domäne des cpSRP43 mit
einem konservierten sogenannten L18-Motiv des LHCPs (DeLille et al., 2000; Tu et al.,
2000; Jonas-Straube et al., 2001; Stengel et al., 2008). In dieser Doktorarbeit wurde die
Ankyrin repeat Region 4 des cpSRP43 als Interaktionsdomäne für die Translokase Alb3
identifiziert. Erstaunlicherweise stellt das cpSRP43 mit der Ankyrin repeat Region die
gleiche Binderegion für das Alb3, wie für das LHCP. In Stengel et al, 2008 wurde gezeigt,
dass das L18-Peptid des LHCP an die Oberfläche der Ankyrin repeats 2-4 bindet.
Weiterhin bewirkte eine Mutation des Tyrosin 204 der Ankyrin repeat Region 3 des
cpSRP43 einen Verlust der Bindung des L18-Peptids.
In diesem Zusammenhang stellt sich die Frage nach dem Zeitpunkt der Interaktion von
cpSRP43 und Alb3. Möglicherweise erfolgt die Interaktion des cpSRP43 und Alb3 erst zu
einem späteren Zeitpunkt der LHCP-Insertion. Das LHCP muss von den cpSRP-
Komponenten abgelöst und in die Membran integriert werden, bevor es zur Freisetzung
der Komponenten für nachfolgende Transportprozesse kommt. Daher ist es nötig den
Mechanismus der Freisetzung des LHCPs aus dem Transitkomplex für die Insertion in die
Thylakoidmembran zu analysieren. In Marty et al., 2009 (eingereicht) wurde gezeigt, dass
der Alb3 C-Terminus einen Effekt auf die Transitkomplexbildung zwischen cpSRP43,
cpSRP54 und LHCP hatte. In Anwesenheit des Alb3 C-Terminus kam es nicht zum
Einwandern des Transitkomplexes in ein nichtdenaturierendes Gel. Es konnte bereits
gezeigt werden, dass das LHCP nur in Form des Transitkomplexes in die Gelmatrix
einwanderte (DeLille et al., 2000). Das bedeutet, dass Alb3 einen spezifischen Effekt auf
die Stabilität des Transitkomplexes oder auf das Einwandern des Komplexes in das
nichtdenaturierende Gel hatte. Man nimmt an, dass Alb3 mit dem cpSRP43 des
Transitkomplexes interagieren kann, und dass diese Interaktion eine Bedeutung für die
Freisetzung und Insertion der LHCPs haben könnte. Messungen über die isothermale
Titrationskalorimetrie zeigten, dass nicht nur das cpSRP43, sondern auch der Alb3 C-
Terminus, das L18-Motiv des LHCPs bindet (Marty et al., 2009, eingereicht). Weitere
Analysen ergaben, dass zwischen dem Alb3 C-Terminus und cpSRP43 eine ähnliche
Bindungsaffinität vorlag wie zwischen dem Alb3 C-Terminus und einem vorgeformten
Komplex aus cpSRP43 und dem L18-Peptid (Marty et al., 2009, eingereicht).
Interessanterweise hatte ein vorgeformter Komplex aus dem Alb3 C-Terminus und
cpSRP43 eine geringere Bindungsaffinität für das L18-Peptid, als der Alb3 C-Terminus
und das cpSRP43 allein. Diese Beobachtung könnte so interpretiert werden, dass die
Interaktion zwischen Alb3 und cpSRP43 eine nachträgliche Bindung des LHCPs
verhindert, sobald es aus dem Transitkomplex freigesetzt wurde.
Die Bedeutung der Interaktion von Alb3 und cpSRP43 wird möglicherweise in dem
docking-Prozess des Transitkomplexes an die Thylakoidmembran, in der Regulation der
GTPase Aktivität des cpSRP54 und cpFtsY und in der Freisetzung der LHCPs für die

5 Diskussion 114
Insertion in die Membran liegen.
In Abbildung 5-4 ist ein Modell des posttranslationalen cpSRP-Transportwegs dargestellt.
Das LHCP wird von dem cpSRP43 und cpSRP54 gebunden und über den Transitkomplex
zum Thylakoidmembran assoziierten cpSRP-Rezeptor cpFtsY transportiert. An der
Thylakoidmembran kommt es zur Interaktion von Alb3 mit dem cpSRP43. Aufgrund dieser
Interaktion des membrangebundenen Komplexes mit der Translokase Alb3 erfolgt nach
GTP-Hydrolyse die Insertion der LHCPs und Freisetzung der löslichen cpSRP-
Komponenten.
Transit-komplex
LHCP
Thylakoidmembran
Alb3
cpSRP
cpFtsY43 54 43 54
43 54
43 54
43 54
TT
TT D
D
Abbildung 5-4: Modell des posttranslationalen cpSRP-Transportwegs Das cpSRP (chloroplast signal recognition particle) besteht aus den beiden Proteinkomponenten
cpSRP43 und cpSRP54 und bildet zusammen mit dem LHCP (light-harvesting chlorophyll-a/b-
binding protein) den Transitkomplex. Der Transitkomplex ermöglicht den Transport der
hydrophoben LHCPs durch das Stroma zur Thylakoidmembran und bildet mit dem cpSRP-
Rezeptor cpFtsY einen membrangebundenen Komplex. Nach erfolgter Interaktion von Alb3 mit
cpSRP43 kommt es nach GTP-Hydrolyse zur Insertion der LHCPs und zur Freisetzung der
löslichen cpSRP-Komponenten.

5 Diskussion 115
5.4 Alb4 interagiert nicht mit den Komponenten des cpSRP-Transportwegs
Das Alb4 wurde als zweites chloroplastidäres Homolog der Oxa1/YidC/Alb3-Proteinfamilie
in Arabidopsis thaliana identifiziert (Gerdes et al., 2006). GFP-Lokalisationsstudien und
Importstudien zeigten, dass Alb4 im Chloroplasten lokalisiert ist und als Membranprotein
vorliegt. Die Analyse chloroplastidärer Subfraktionen (Stroma, Hüllmembranen und
Thylakoidmembranen) ergab eine ausschließliche Lokalisation des Alb4 in der
Thylakoidmembran von Arabidopsis thaliana. Eine T-DNA Insertionsmutante und eine
RNAi-Mutante von Arabidopsis thaliana mit einem Alb4 Proteinlevel von weniger als 10 %
zeigten Defekte in der chloroplastidären Ultrastruktur, währenddessen diese Pflanzen
keinen offensichtlichen äußerlichen Phänotyp aufwiesen. Elektronenmikroskopische
Aufnahmen verzeichneten vergrößerte Plastiden und eine aufgelockerte Struktur der
Thylakoidmembranen mit vergrößerten stromalen Bereichen. Diese Ergebnisse deuteten
darauf hin, dass Alb4 für die Biogenese der Chloroplasten verantwortlich zu sein scheint.
Die genaue Funktion des Alb4 war jedoch zu Beginn der vorliegenden Arbeit noch
ungeklärt.
In dieser Doktorarbeit galt es zunächst zu untersuchen, ob Alb4 eine Funktion innerhalb
des posttranslationalen cpSRP-Transportwegs einnimmt. Ergebnisse aus dem Hefe Split-
Ubiquitin System und der bimolekularen Fluoreszenzkomplementation (Split-YFP System)
erbrachten im Vergleich zu Alb3 keine Interaktion von Alb4 mit den Proteinen des cpSRP-
Transportwegs (Kapitel 4.2.1 und 4.2.2). Zudem wies die bereits beschriebene RNAi-
Mutante von Alb4 keinen verringerten LHCP-Gehalt auf (Gerdes et al., 2006), so dass der
Transport und die Insertion der LHCPs in die Thylakoidmembran nicht beeinträchtigt zu
sein schien. Die alb3-Mutante hingegen zeigte einen drastischen Albino-Phänotyp mit
weißen bis leicht gelblichen Kotyledonen und einem Chlorophyllgehalt von nur etwa 5 %
verglichen mit dem Wildtyp (Sundberg et al., 1997). Diese Mutanten waren nicht in der
Lage unter normalen Wachstumsbedingungen über das Kotyledonen-Stadium hinaus zu
überleben. Elektronenmikroskopische Aufnahmen dieser Mutanten erbrachten eine
veränderte chloroplastidäre Ultrastruktur mit nur wenigen Thylakoidmembranen und kaum
Granastapelung. Der starke Phänotyp der alb3-Mutante lässt vermuten, dass Alb4 den
Verlust von Alb3 nicht kompensieren kann und somit eine andere Funktion haben muss.
Diese Daten verdeutlichen, dass speziell Alb3, und nicht das Alb4, für die
posttranslationale Insertion der LHCPs in die Thylakoidmembran verantwortlich ist.

5 Diskussion 116
5.5 Alb4 ist mit der chloroplastidären ATP-Synthase assoziiert und interagiert nicht mit Alb3 und cpSecY
Um den Einfluss von Alb4 in der Chloroplastenbiogenese zu ermitteln, sollte in der
vorliegenden Doktorarbeit die Interaktion der Homologe Alb3 und Alb4 untereinander und
mit anderen Thylakoidmembranproteinen analysiert werden. Mit Hilfe des chemischen
Crosslinkings sollten zunächst Proteine identifiziert werden, die sich mit Alb4 in räumlicher
Nähe befanden. Die Analysen ergaben, dass das Alb4 in einem ca. 170 kDa großen
Crosslinking-Produkt vorlag (Kapitel 4.3.1). Über die Massenspektrometrie wurden die
ATP-Synthase Untereinheiten CF1α und CF1β als weitere Proteinkomponenten des
Crosslinking-Produkts identifiziert. Western Blot Analysen bestätigten die Existenz des
CF1β in dem Crosslinking-Produkt. Die Proteine Alb3 und cpSecY hingegen waren keine
Komponenten des 170 kDa großen Crosslinking-Produkts.
Die chloroplastidäre ATP-Synthase besteht aus einem zum Stroma gerichteten, löslichen
CF1-Komplex und einem integralen CF0-Komplex. Der CF1-Komplex vollzieht die ATP-
Synthese, während der transmembrane CF0-Komplex dem Protonentransport zwischen
Thylakoidlumen und Stroma dient. Der lösliche Kopf der ATP-Synthase umfasst die
Untereinheiten CF1α, CF1β, CF1γ, CF1δ und CF1ε. Der integrale Teil wird von den
Proteinen CF0I, CF0II, CF0III und CF0IV gebildet. Die Untereinheiten CF1γ, CF1δ und CF0II
sind kernkodiert, während die anderen Untereinheiten auf dem Plastom kodiert sind
(Richter et al., 2000).
Die Crosslinking-Studien ergaben somit eine Vernetzung des Alb4 mit den Untereinheiten
CF1α und CF1β des löslichen Komplexes der ATP-Synthase. Die direkte Assoziation von
Alb4 mit der ATP-Synthase wurde mittels bimolekularer Fluoreszenzkomplementation
bestätigt (Kapitel 4.3.2). Für diesen Versuch wurden ausschließlich die kernkodierten
ATP-Synthase Untereinheiten CF1γ, CF1δ und CF0II auf Interaktion mit Alb4 in
Arabidopsis thaliana Protoplasten getestet. Die plastidär kodierten Untereinheiten hätten
mit dieser Technik nicht aussagekräftig untersucht werden können, da sie einem
cotranslationalen Insertionsmechanismus unterliegen. Die Technik der bimolekularen
Fluoreszenzkomplementation beinhaltet einen posttranslationalen Transport der Proteine
in die Chloroplasten. Die Messungen ergaben, dass Alb4 mit der ATP-Synthase
Untereinheit CF0II interagierte, während eine Interaktion mit den Proteinen CF1γ und CF1δ
ausblieb. Es muss beachtet werden, dass eine fehlende Interaktion zweier Proteine in
diesem System möglicherweise auf eine unzureichende Zugänglichkeit der YFP-
Fragmente zurückzuführen sein könnte.
Mit dem chemischen Crosslinking und der bimolekularen Fluoreszenzkomplementation
wurde über zwei verschiedene Methoden eine Assoziation von Alb4 mit der ATP-
Synthase gezeigt. Diese Ergebnisse wurden mit weiteren Daten aus einer Kooperation mit

5 Diskussion 117
der Arbeitsgruppe von Prof. Dr. J. Soll (Universität München) in einer gemeinsamen
Publikation zusammengestellt (Benz et al., 2009, im Druck). In diesem Manuskript wurde
Alb4 als funktionelles Mitglied der Oxa1/YidC/Alb3-Proteinfamilie bestätigt. Es zeigte sich,
dass Alb4 einen YidC-defizienten E. coli Stamm komplementierte und dass der C-
Terminus von Alb4 für diese Funktion essentiell war.
Bisherige Komplementationsstudien haben eine funktionelle Einordnung der Proteine in
die Oxa1/YidC/Alb3-Proteinfamilie bereits beschrieben (Jiang et al., 2002; Preuss et al.,
2005; van Bloois et al., 2007). Die Bedeutung einer C-terminalen Extension dieser
Proteine wurde nicht nur für das genannte Alb4 nachgewiesen. Daten aus der
vorliegenden Doktorarbeit ergaben eine entscheidende Bedeutung des C-Terminus von
Alb3 für die Bindung des cpSRP43 im posttranslationalen cpSRP-Transportweg (Marty et
al., 2009, eingereicht). Des Weiteren wurde bereits der Einfluss der C-Termini des
bakteriellen YidC und mitochondrialen Oxa1 auf die Ribosomenbindung gezeigt (Szyrach
et al., 2003; Kohler et al., 2009).
Ein Schwerpunkt des Manuskripts (Benz et al., 2009, im Druck) lag auf der Analyse von
zwei unabhängigen Alb4 TILLING-Mutanten, die kein Alb4 Protein mehr aufwiesen. Diese
Arabidopsis Mutanten zeigten ein reduziertes Wachstum während der ersten drei bis vier
Wochen der pflanzlichen Entwicklung, erreichten allerdings die Samenreife zur gleichen
Zeit wie der Wildtyp und waren fertil. Gleiche Beobachtungen wurden für die T-DNA
Insertionsmutante von Alb4, die bereits in Gerdes et al., 2006 beschrieben wurde, gezeigt.
Das bedeutet, dass diese Wachstumsverzögerung in den drei unabhängigen Arabidopsis-
Linien auf einem Funktionsverlust des Alb4 Proteins beruhte. Da die Mutanten allerdings
die Wachstumsdefekte im Verlauf der Entwicklung wieder ausgleichen konnten und fertil
waren, scheint das Alb4 nicht so eine essentielle Bedeutung zu haben, wie das homologe
Alb3. Möglicherweise könnten die Defekte in der frühen Wachstumsphase auch durch
andere Faktoren kompensiert werden.
Die Analyse der chloroplastidären Ultrastruktur der TILLING-Mutanten ergab Defekte in
der Anordnung der Granathylakoide, wie sie bereits für die Mutanten in Gerdes et al.,
2006 gezeigt wurden. Verglichen zu der bereits publizierten alb3-Mutante zeigten die
elektronenmikroskopischen Aufnamen der TILLING-Mutanten einen viel schwächeren
Phänotyp. Die alb3-Mutante wies eine drastische Reduktion der Thylakoidmembranen
und nahezu fehlende Granastapelung auf (Sundberg et al., 1997).
Interessanterweise zeigten die TILLING-Mutanten von Alb4 Defekte im steady-state-level
der ATP-Synthase Untereinheiten (Benz et al., 2009, im Druck). Damit war, neben der
bereits geschilderten direkten Assoziation des Alb4 mit der ATP-Synthase, ein weiterer
Hinweis auf eine mögliche Funktion von Alb4 im Zusammenhang mit der ATP-Synthase
gegeben. Der Proteinlevel der kernkodierten und plastidärkodierten ATP-Synthase
Untereinheiten war im Vergleich zum Wildtyp deutlich reduziert. Die Reduktion der

5 Diskussion 118
Proteinlevel der ATP-Synthase Untereinheiten in den alb4-Mutanten war allerdings nicht
auf eine verringerte Transkription der entsprechenden Gene zurückzuführen (Benz et al.,
2009, im Druck). Das bedeutet, dass die Reduktion dieser Proteine posttranslationale
Ursachen, wie z. B. Defekte in der Insertion und Assemblierung der Proteine, haben
könnte.
Mittels 2D BN/SDS-PAGE wurde die Assemblierung der ATP-Synthase in den TILLING-
Mutanten analysiert (Benz et al., 2009, im Druck). Die Analyse der TILLING-Mutanten
ergab, dass der Level der hochmolekularen ATP-Synthase Komplexe reduziert war und
neben den anderen Subkomplexen weitere Intermediate der ATP-Synthase zu detektieren
waren (Benz et al., 2009, im Druck). Diese Ergebnisse deuten an, dass der Verlust des
Alb4 zu einem Defekt in der Assemblierung und Stabilität der ATP-Synthase führt.
Damit stellte sich die Frage, welchen physiologischen Einfluss die Reduktion der ATP-
Synthase Untereinheiten und die gezeigten Defekte in der Assemblierung und Stabilität
der ATP-Synthase in den alb4-Mutanten haben könnten. Durch die Analyse der
Photophosphorylierung zeigte sich, dass die TILLING-Linien 30-50 % weniger ATP
synthetisierten, als der Wildtyp (Benz et al., 2009, im Druck). Die gleichen Beobachtungen
ließen sich für die in Gerdes et al., 2006 bereits beschriebene T-DNA Insertionsmutante
machen. Diese Ergebnisse zeigen, dass die Reduktion der ATP-Synthase Untereinheiten
aufgrund der alb4-Mutation eine Verringerung der ATP-Synthese zur Folge hatte.
Ein Vergleich innerhalb der Oxa1/YidC/Alb3-Proteinfamilie macht deutlich, dass das Alb4
möglicherweise eine neuartige Funktion in den Chloroplasten einnimmt. Für das
homologe Protein Alb3 in Arabidopsis thaliana ist das LHCP als bisher einziges Substrat
identifiziert worden (Sundberg et al., 1997; Moore et al., 2000). Das Alb3 ist für die
Insertion der LHCPs in die Thylakoidmembran verantwortlich. In Chlamydomonas
reinhardtii wurden zwei Alb3-Homologe (Alb3.1 und Alb3.2) nachgewiesen (Bellafiore et
al., 2002; Göhre et al., 2006). Die Inaktivierung des Alb3.1 führte zu einer starken
Reduktion des LHCI und LHCII im Photosystem I und II und zu einem verringerten
Chlorophyllgehalt von etwa nur 30% des Wildtyplevels. Zudem scheint Alb3.1 bei der
Insertion und Assemblierung der Proteine des Photosystems II einen wichtigen Einfluss
zu haben (Bellafiore et al., 2002; Ossenbühl et al., 2004). Eine Reduktion des Alb3.2 in
der Chlamydomonas RNAi-Mutante hatte eine starke Reduktion der Photosysteme I und II
zur Folge (Göhre et al., 2006). Somit konnte bisher nicht gezeigt werden, dass die
anderen chloroplastidären Homologe der Oxa1/YidC/Alb3-Proteinfamilie einen
entscheidenden Einfluss auf die ATP-Synthase hatten. Andere Studien ergaben, dass das
bakterielle YidC und das mitochondriale Oxa1 mit den Untereinheiten der ATP-Synthase
interagieren konnten. Das YidC vermittelt die Sec-unabhängige, cotranslationale Insertion
der ATP-Synthase Untereinheit F0c (van der Laan et al., 2004). Das Oxa1 in
Mitochondrien interagiert mit der Atp9 Untereinheit der ATP-Synthase und vermittelt die

5 Diskussion 119
Assemblierung des ATP-Synthase Komplexes (Jia et al., 2007). Die in der oxa1-Mutante
beschriebenen Assemblierungsdefekte und reduzierten Proteinlevel einiger ATP-
Synthase Untereinheiten könnten zusätzlich eine Bedeutung von Alb4 in der
Assemblierung und Stabilisierung der ATP-Synthase in Chloroplasten unterstützen.
Weitere Studien ergaben, dass Alb4, Alb3 und die ATP-Synthase in den
Stromathylakoiden lokalisiert sind (Benz et al., 2009, im Druck). Versuche aus der
vorliegenden Doktorarbeit und der gemeinsamen Kooperation (Benz et al., 2009, im
Druck) sollten klären, ob Alb4, Alb3 und die ATP-Synthase in gemeinsamen Komplexen
der Thylakoidmembran vorliegen. Ergebnisse einer 2D BN/SDS-PAGE zeigten eine
Comigration von Alb4 mit den ATP-Synthase Untereinheiten (Benz et al., 2009, im Druck).
Gelfiltrationsexperimente von solubilisierten Thylakoidmembranen bestätigten eine
Cofraktionierung von Alb4 mit den Untereinheiten der ATP-Synthase (Kapitel 4.3.3).
Weiterhin wurden die bereits veröffentlichten Daten der Coelution von Alb3 und cpSecY
bestätigt (Klostermann et al., 2002) (Kapitel 4.3.3). Gelfiltrationsanalysen, BN-PAGE,
Coimmunpräzipitationsexperimente, Immunlokalisationsstudien und chemisches
Crosslinking zeigten eine Assoziation von Alb3 mit cpSecY und damit eine mögliche
Funktion in der cotranslationalen Insertion von plastidär kodierten Thylakoidmembran-
proteinen (Klostermann et al., 2002). Interessanterweise cofraktionierte Alb4 nur
geringfügig mit den Proteinen Alb3 und cpSecY, so dass es vermutlich keinen
gemeinsamen Komplex dieser drei Proteine in der Thylakoidmembran gibt.
Coimmunpräzipitationsexperimente bestätigten, dass Alb4 nicht in einem gemeinsamen
Komplex mit Alb3 und cpSecY in der Thylakoidmembran vorliegt (Kapitel 4.3.4). Das Alb3
und cpSecY wurden miteinander copräzipitiert, wodurch die Daten aus Klostermann et al.,
2002 verifiziert wurden. Alb4 hingegen zeigte keine Copräzipitation mit Alb3 und cpSecY.
Damit ist eine direkte Interaktion von Alb4 mit Alb3 und cpSecY auszuschließen.
Die Assoziation von Alb4 mit der ATP-Synthase ließ sich nicht mit Hilfe der
Coimmunpräzipitation ergänzend unterstützen. Allerdings kam es zur Copräzipitation von
Alb3 mit der ATP-Synthase Untereinheit CF1γ (Daten nicht gezeigt). Demnach ist eine
Funktion von Alb3 bei der Insertion und Assemblierung der ATP-Synthase nicht
auszuschließen.
Für alle durchgeführten biochemischen Analysen war der Einsatz spezifischer Antikörper
für Alb3 und Alb4 unbedingt erforderlich. Die Herstellung und ausführliche Überprüfung
der Spezifität der verwendeten Antikörper trug entscheidend für die Analyse der
unterschiedlichen Funktion der homologen Proteine Alb3 und Alb4 bei.
Zusammenfassend deuteten die Ergebnisse darauf hin, dass Alb4 einen Einfluss auf die
ATP-Synthase besitzt und eine Funktion unabhängig von Alb3 und cpSecY einnimmt. Die
homologen Proteine Alb3 und Alb4 scheinen demnach in verschiedenen Komplexen der
Thylakoidmembran zu agieren.

6 Zusammenfassung 120
6. Zusammenfassung
In der vorliegenden Arbeit wurde die Funktion der chloroplastidären Homologe Alb3 und
Alb4 bei der Biogenese der Thylakoidmembran von Arabidopsis thaliana genauer
analysiert.
Zunächst war die Herstellung spezifischer Antikörper für Alb3 und Alb4 von
entscheidender Bedeutung, um die homologen Proteine in biochemischen
Untersuchungen getrennt voneinander detektieren zu können. Die Spezifität der
Antikörper wurde durch detaillierte Antikörpertests gegen die Antigene, die in vitro
translatierten Volllängenproteine und Thylakoidmembranproteine belegt.
In einem ersten Teilprojekt dieser Arbeit sollte die Frage geklärt werden, ob für den
docking-Prozess des Transitkomplexes an die Thylakoidmembran eine direkte Interaktion
der Proteine des cpSRP-Transportwegs mit der Translokase Alb3 vorliegt. Die in vivo
Interaktionsstudien über das Hefe Split-Ubiquitin System und die bimolekulare
Fluoreszenzkomplementation zeigten eine direkte Interaktion von cpSRP43 mit Alb3,
während eine Interaktion des cpSRP54 und cpFtsY mit Alb3 ausblieb. Zudem zeigten pull-
down Versuche, dass cpSRP43 auch im Komplex mit dem cpSRP54 mit Alb3 interagiert.
Die Interaktion zwischen cpSRP43 und Alb3 lässt vermuten, dass das cpSRP43 neben
einer Funktion bei der Transitkomplexbildung auch einen Einfluss auf die
membranassoziierten Prozesse der LHCP-Insertion über die Translokase Alb3 ausübt.
Anhand detaillierter Analysen mittels bimolekularer Fluoreszenzkomplementation,
peptide-scanning, pull-down Analysen und Peptidinhibierungsstudien wurden zwei
Bindestellen des Alb3 für das cpSRP43 identifiziert. Das Bindemotiv innerhalb der
5. Transmembrandomäne des Alb3 umfasst die Aminosäuren 314-318 (LVFKF) und die
Bindestelle im C-Terminus des Alb3 die Aminosäuren 374-388. Im weiteren Verlauf der
Arbeit wurde die Bindedomäne des cpSRP43 für die Interaktion mit Alb3 eingegrenzt.
Thylakoidmembran-Bindungsversuche deuten auf eine essentielle Bedeutung der Ankyrin
repeat Region 4 für die Bindung des cpSRP43 an die Thylakoidmembran.
In dem zweiten Teilprojekt dieser Arbeit sollten durch Proteininteraktionsstudien Hinweise
auf die Funktion des Thylakoidmembranproteins Alb4 gewonnen werden. Anhand von
crosslinking Studien, bimolekularer Fluoreszenzkomplementation und Gelfiltrations-
analysen wurde eine direkte Interaktion und Assoziation von Alb4 mit der
chloroplastidären ATP-Synthase nachgewiesen. Des Weiteren weisen die Ergebnisse von
Gelfiltrationsanalysen und Coimmunpräzipitationsstudien darauf hin, dass Alb4 nicht mit
Alb3 und cpSecY in einem Komplex vorliegt. Demnach lässt sich eine von Alb3 und der
cpSec-Translokase unabhängige Funktion von Alb4 bei der Insertion und Assemblierung
der ATP-Synthase vermuten.

7 Summary 121
7. Summary
This thesis addressed the function of the chloroplast homologs Alb3 and Alb4 in the
biogenesis of thylakoid membranes in Arabidopsis thaliana.
The production of specific antibodies against Alb3 and Alb4 was crucial to detect those
homologous proteins independent from each other in biochemical analyses. The
specificity of the antibodies was shown by detailed antibody tests against the antigen, in-
vitro translation products and thylakoid membrane proteins.
The first part of the thesis addressed the question whether the docking-process of the
transit complex at the thylakoid membrane is based on a direct interaction of the proteins
of the cpSRP transport pathway with the translocase Alb3. In-vivo interaction studies
carried out by the yeast split-ubiquitin system and the bimolecular fluorescence
complementation studies revealed a direct interaction between cpSRP43 and Alb3,
whereas there was no interaction of cpSRP54 and cpFtsY with Alb3. Pull-down analyses
also indicated an interaction of cpSRP43 with Alb3 when in a complex with cpSRP54. The
interaction of cpSRP43 and Alb3 may indicate that cpSRP43 has an influence on the
membrane associated processes of the LHCP insertion by the translocase Alb3 in
addition to its function in transit complex formation. Detailed analyses carried out by
bimolecular fluorescence complementation studies, peptide-scanning, pull-down analyses
and peptide inhibition studies identified two binding sites of Alb3 for the cpSRP43. The
binding motif within the fifth transmembrane domain of Alb3 consists of the amino acids
314-318 (LVFKF) whilst the binding domain in the C-Terminus comprises the amino acids
374-388. Furthermore the binding domain of cpSRP43 for the interaction with Alb3 was
narrowed down. Thylakoid membrane binding studies suggested an essential role of
ankyrin repeat 4 for thylakoid membrane binding of cpSRP43.
The aim of the second part of the thesis was to provide an indication of the function of the
thylakoid membrane protein Alb4 by protein interaction studies. Crosslinking studies,
bimolecular fluorescence complementation studies and gelfiltration analyses revealed a
direct interaction and association of Alb4 with the chloroplast ATP-synthase. In addition
gelfiltration analyses and coimmunoprecipitation studies indicated that Alb4 is not
associated in the same complex as Alb3 and cpSecY. According to this contention, Alb4
seems to have an independent function of Alb3 and the cpSec-translocase in the insertion
and assembly of the ATP-synthase.

8 Literaturverzeichnis 122
8. Literaturverzeichnis
Abdallah F, Salamini F, Leister D (2000) A prediction of the size and evolutionary origin of the proteome of chloroplasts of Arabidopsis. Trends Plant Sci 5: 141-142
Agne B, Kessler F (2009) Protein transport in organelles: The Toc complex way of preprotein import. FEBS J 276: 1156-1165
Aldridge C, Cain P, Robinson C (2009) Protein transport in organelles: Protein transport into and across the thylakoid membrane. FEBS J 276: 1177-1186
Altamura N, Capitanio N, Bonnefoy N, Papa S, Dujardin G (1996) The Saccharomyces cerevisiae OXA1 gene is required for the correct assembly of cytochrome c oxidase and oligomycin-sensitive ATP synthase. FEBS Lett 382: 111-115
Bauer M, Behrens M, Esser K, Michaelis G, Pratje E (1994) PET1402, a nuclear gene required for proteolytic processing of cytochrome oxidase subunit 2 in yeast. Mol Gen Genet 245: 272-278
Beck K, Eisner G, Trescher D, Dalbey RE, Brunner J, Muller M (2001) YidC, an assembly site for polytopic Escherichia coli membrane proteins located in immediate proximity to the SecYE translocon and lipids. EMBO Rep 2: 709-714
Bellafiore S, Ferris P, Naver H, Göhre V, Rochaix JD (2002) Loss of Albino3 leads to the specific depletion of the light-harvesting system. Plant Cell 14: 2303-2314
Benz JP, Soll J, Bolter B (2009) Protein transport in organelles: The composition, function and regulation of the Tic complex in chloroplast protein import. FEBS J 276: 1166-1176
Bonnefoy N, Chalvet F, Hamel P, Slonimski PP, Dujardin G (1994) OXA1, a Saccharomyces cerevisiae nuclear gene whose sequence is conserved from prokaryotes to eukaryotes controls cytochrome oxidase biogenesis. J Mol Biol 239: 201-212
Bonnefoy N, Fiumera HL, Dujardin G, Fox TD (2009) Roles of Oxa1-related inner-membrane translocases in assembly of respiratory chain complexes. Biochim Biophys Acta 1793: 60-70
Braun NA, Davis AW, Theg SM (2007) The chloroplast Tat pathway utilizes the transmembrane electric potential as an energy source. Biophys J 93: 1993-1998
Brehm A, Tufteland KR, Aasland R, Becker PB (2004) The many colours of chromodomains. Bioessays 26: 133-140
Bruce BD (2000) Chloroplast transit peptides: structure, function and evolution. Trends Cell Biol 10: 440-447
Bruce BD (2001) The paradox of plastid transit peptides: conservation of function despite divergence in primary structure. Biochim Biophys Acta 1541: 2-21
Brundage L, Hendrick JP, Schiebel E, Driessen AJ, Wickner W (1990) The purified E. coli integral membrane protein SecY/E is sufficient for reconstitution of SecA-dependent precursor protein translocation. Cell 62: 649-657
Bullock WO, Fernandez JM, Short JM (1987). XL1-Blue: a high efficiency plasmid transforming recA Escherichia coli strain with beta-glucosidase selection. Biotechnique 5: 376 378.
Cavalier-Smith T (2000) Membrane heredity and early chloroplast evolution. Trends Plant Sci 5: 174-182

8 Literaturverzeichnis 123
Chaddock AM, Mant A, Karnauchov I, Brink S, Herrmann RG, Klösgen RB, Robinson C (1995) A new type of signal peptide: central role of a twin-arginine motif in transfer signals for the delta pH-dependent thylakoidal protein translocase. EMBO J 14: 2715-2722
Chandrasekar S, Chartron J, Jaru-Ampornpan P, Shan SO (2008) Structure of the chloroplast signal recognition particle (SRP) receptor: domain arrangement modulates SRP-receptor interaction. J Mol Biol 375: 425-436
Chen M, Samuelson JC, Jiang F, Muller M, Kuhn A, Dalbey RE (2002) Direct interaction of YidC with the Sec-independent Pf3 coat protein during its membrane protein insertion. J Biol Chem 277: 7670-7675
Cline K, Mori H (2001) Thylakoid DeltapH-dependent precursor proteins bind to a cpTatC-Hcf106 complex before Tha4-dependent transport. J Cell Biol 154: 719-729
Dabney-Smith C, Mori H, Cline K (2006) Oligomers of Tha4 organize at the thylakoid Tat translocase during protein transport. J Biol Chem 281: 5476-5483
de Gier JW, Luirink J (2001) Biogenesis of inner membrane proteins in Escherichia coli. Mol Microbiol 40: 314-322
DeLille J, Peterson EC, Johnson T, Moore M, Kight A, Henry R (2000) A novel precursor recognition element facilitates posttranslational binding to the signal recognition particle in chloroplasts. Proc Natl Acad Sci U S A 97: 1926-1931
Di Cola A, Bailey S, Robinson C (2005) The thylakoid delta pH/delta psi are not required for the initial stages of Tat-dependent protein transport in tobacco protoplasts. J Biol Chem 280: 41165-41170
Driessen AJ, Nouwen N (2008) Protein translocation across the bacterial cytoplasmic membrane. Annu Rev Biochem 77: 643-667
Facey SJ, Neugebauer SA, Krauss S, Kuhn A (2007) The mechanosensitive channel protein MscL is targeted by the SRP to the novel YidC membrane insertion pathway of Escherichia coli. J Mol Biol 365: 995-1004
Finazzi G, Chasen C, Wollman FA, de Vitry C (2003) Thylakoid targeting of Tat passenger proteins shows no delta pH dependence in vivo. EMBO J 22: 807-815
Franklin AE, Hoffman NE (1993). Characterization of a Chloroplast Homologue of the 54-kDa Subunit of the Signal Recognition Particle. J Biol Chem 268: 22175-22180.
Funes S, Hasona A, Bauerschmitt H, Grubbauer C, Kauff F, Collins R, Crowley PJ, Palmer SR, Brady LJ, Herrmann JM (2009) Independent gene duplications of the YidC/Oxa/Alb3 family enabled a specialized cotranslational function. Proc Natl Acad Sci U S A 106: 6656-6661
Funes S, Nargang FE, Neupert W, Herrmann JM (2004) The Oxa2 protein of Neurospora crassa plays a critical role in the biogenesis of cytochrome oxidase and defines a ubiquitous subbranch of the Oxa1/YidC/Alb3 protein family. Mol Biol Cell 15: 1853-1861
Funke S, Knechten T, Ollesch J, Schünemann D (2005) A unique sequence motif in the 54-kDa subunit of the chloroplast signal recognition particle mediates binding to the 43-kDa subunit. J Biol Chem 280: 8912-8917
Gathmann S, Eva R, Uwe K, Schneider D (2008) A conserved structure and function of the YidC homologous protein Slr1471 from Synechocystis sp. PCC 6803. J Microbiol Biotechnol 18: 1090-1094

8 Literaturverzeichnis 124
Gerdes L, Bals T, Klostermann E, Karl M, Philippar K, Hunken M, Soll J, Schünemann D (2006) A second thylakoid membrane-localized Alb3/OxaI/YidC homologue is involved in proper chloroplast biogenesis in Arabidopsis thaliana. J Biol Chem 281: 16632-16642
Glick BS, Von Heijne G (1996) Saccharomyces cerevisiae mitochondria lack a bacterial-type sec machinery. Protein Sci 5: 2651-2652
Goforth RL, Peterson EC, Yuan J, Moore MJ, Kight AD, Lohse MB, Sakon J, Henry RL (2004) Regulation of the GTPase cycle in post-translational signal recognition particle-based protein targeting involves cpSRP43. J Biol Chem 279: 43077-43084
Göhre V, Ossenbühl F, Crevecoeur M, Eichacker LA, Rochaix JD (2006) One of two alb3 proteins is essential for the assembly of the photosystems and for cell survival in Chlamydomonas. Plant Cell 18: 1454-1466
Groves MR, Mant A, Kuhn A, Koch J, Dubel S, Robinson C, Sinning I (2001) Functional characterization of recombinant chloroplast signal recognition particle. J Biol Chem 276: 27778-27786
Hell K, Herrmann JM, Pratje E, Neupert W, Stuart RA (1998) Oxa1p, an essential component of the N-tail protein export machinery in mitochondria. Proc Natl Acad Sci U S A 95: 2250-2255
Hell K, Neupert W, Stuart RA (2001) Oxa1p acts as a general membrane insertion machinery for proteins encoded by mitochondrial DNA. EMBO J 20: 1281-1288
Hermkes R, Funke S, Richter C, Kuhlmann J, Schünemann D (2006) The alpha-helix of the second chromodomain of the 43 kDa subunit of the chloroplast signal recognition particle facilitates binding to the 54 kDa subunit. FEBS Lett 580: 3107-3111
Hoffman NE, Franklin AE (1994) Evidence for a stromal GTP requirement for the integration of a chlorophyll a/b-binding polypeptide into thylakoid membranes. Plant Physiol 105: 295-304
Houben EN, ten Hagen-Jongman CM, Brunner J, Oudega B, Luirink J (2004) The two membrane segments of leader peptidase partition one by one into the lipid bilayer via a Sec/YidC interface. EMBO Rep 5: 970-975
Jackson-Constan D, Akita M, Keegstra K (2001) Molecular chaperones involved in chloroplast protein import. Biochim Biophys Acta 1541: 102-113
Jaru-Ampornpan P, Chandrasekar S, Shan SO (2007) Efficient interaction between two GTPases allows the chloroplast SRP pathway to bypass the requirement for an SRP RNA. Mol Biol Cell 18: 2636-2645
Jarvis P (2008) Targeting of nucleus-encoded proteins to chloroplasts in plants. New Phytol 179: 257-285
Jarvis P, Robinson C (2004) Mechanisms of protein import and routing in chloroplasts. Curr Biol 14: R1064-1077
Jia L, Dienhart M, Schramp M, McCauley M, Hell K, Stuart RA (2003) Yeast Oxa1 interacts with mitochondrial ribosomes: the importance of the C-terminal region of Oxa1. EMBO J 22: 6438-6447
Jia L, Dienhart MK, Stuart RA (2007) Oxa1 directly interacts with Atp9 and mediates its assembly into the mitochondrial F1Fo-ATP synthase complex. Mol Biol Cell 18: 1897-1908

8 Literaturverzeichnis 125
Jiang F, Yi L, Moore M, Chen M, Rohl T, Van Wijk KJ, De Gier JW, Henry R, Dalbey RE (2002) Chloroplast YidC homolog Albino3 can functionally complement the bacterial YidC depletion strain and promote membrane insertion of both bacterial and chloroplast thylakoid proteins. J Biol Chem 277: 19281-19288
Jonas-Straube E, Hutin C, Hoffman NE, Schünemann D (2001) Functional analysis of the protein-interacting domains of chloroplast SRP43. J Biol Chem 276: 24654-24660
Kathir KM, Rajalingam D, Sivaraja V, Kight A, Goforth RL, Yu C, Henry R, Kumar TK (2008) Assembly of chloroplast signal recognition particle involves structural rearrangement in cpSRP43. J Mol Biol 381: 49-60
Keegstra K, Cline K (1999) Protein import and routing systems of chloroplasts. Plant Cell 11: 557-570
Kessler F, Schnell DJ (2006) The function and diversity of plastid protein import pathways: a multilane GTPase highway into plastids. Traffic 7: 248-257
Kiefer D, Kuhn A (2007) YidC as an essential and multifunctional component in membrane protein assembly. Int Rev Cytol 259: 113-138
Kim SJ, Jansson S, Hoffman NE, Robinson C, Mant A (1999) Distinct "assisted" and "spontaneous" mechanisms for the insertion of polytopic chlorophyll-binding proteins into the thylakoid membrane. J Biol Chem 274: 4715-4721
Klimyuk VI, Persello-Cartieaux F, Havaux M, Contard-David P, Schünemann D, Meiherhoff K, Gouet P, Jones JD, Hoffman NE, Nussaume L (1999) A chromodomain protein encoded by the arabidopsis CAO gene is a plant-specific component of the chloroplast signal recognition particle pathway that is involved in LHCP targeting. Plant Cell 11: 87-99
Klösgen RB, Brock IW, Herrmann RG, Robinson C (1992) Proton gradient-driven import of the 16 kDa oxygen-evolving complex protein as the full precursor protein by isolated thylakoids. Plant Mol Biol 18: 1031-1034
Klostermann E, Droste Gen Helling I, Carde JP, Schünemann D (2002) The thylakoid membrane protein ALB3 associates with the cpSecY-translocase in Arabidopsis thaliana. Biochem J 368: 777-781
Koch HG, Moser M, Muller M (2003) Signal recognition particle-dependent protein targeting, universal to all kingdoms of life. Rev Physiol Biochem Pharmacol 146: 55-94
Kogata N, Nishio K, Hirohashi T, Kikuchi S, Nakai M (1999) Involvement of a chloroplast homologue of the signal recognition particle receptor protein, FtsY, in protein targeting to thylakoids. FEBS Lett 447: 329-333
Kohler R, Boehringer D, Greber B, Bingel-Erlenmeyer R, Collinson I, Schaffitzel C, Ban N (2009) YidC and Oxa1 form dimeric insertion pores on the translating ribosome. Mol Cell 34: 344-353
Kuhn A, Stuart R, Henry R, Dalbey RE (2003) The Alb3/Oxa1/YidC protein family: membrane-localized chaperones facilitating membrane protein insertion? Trends Cell Biol 13: 510-516
Laidler V, Chaddock AM, Knott TG, Walker D, Robinson C (1995) A SecY homolog in Arabidopsis thaliana. Sequence of a full-length cDNA clone and import of the precursor protein into chloroplasts. J Biol Chem 270: 17664-17667
Leister D (2003) Chloroplast research in the genomic age. Trends Genet 19: 47-56

8 Literaturverzeichnis 126
Li X, Henry R, Yuan J, Cline K, Hoffman NE (1995) A chloroplast homologue of the signal recognition particle subunit SRP54 is involved in the posttranslational integration of a protein into thylakoid membranes. Proc Natl Acad Sci U S A 92: 3789-3793
Marques JP, Schattat MH, Hause G, Dudeck I, Klösgen RB (2004) In vivo transport of folded EGFP by the DeltapH/TAT-dependent pathway in chloroplasts of Arabidopsis thaliana. J Exp Bot 55: 1697-1706
Martin W, Herrmann RG (1998) Gene transfer from organelles to the nucleus: how much, what happens, and Why? Plant Physiol 118: 9-17
Marty NJ, Rajalingam D, Kight AD, Lewis NE, Fologea D, Kumar TK, Henry RL, Goforth RL (2009) The membrane-binding motif of the chloroplast signal recognition particle receptor (cpFtsY) regulates GTPase activity. J Biol Chem 284: 14891-14903
May T, Soll J (2000) 14-3-3 proteins form a guidance complex with chloroplast precursor proteins in plants. Plant Cell 12: 53-64
Michl D, Robinson C, Shackleton JB, Herrmann RG, Klösgen RB (1994) Targeting of proteins to the thylakoids by bipartite presequences: CFoII is imported by a novel, third pathway. EMBO J 13: 1310-1317
Moore M, Goforth RL, Mori H, Henry R (2003) Functional interaction of chloroplast SRP/FtsY with the ALB3 translocase in thylakoids: substrate not required. J Cell Biol 162: 1245-1254
Moore M, Harrison MS, Peterson EC, Henry R (2000) Chloroplast Oxa1p homolog albino3 is required for post-translational integration of the light harvesting chlorophyll-binding protein into thylakoid membranes. J Biol Chem 275: 1529-1532
Mori H, Cline K (2002) A twin arginine signal peptide and the pH gradient trigger reversible assembly of the thylakoid [Delta]pH/Tat translocase. J Cell Biol 157: 205-210
Mori H, Summer EJ, Ma X, Cline K (1999) Component specificity for the thylakoidal Sec and Delta pH-dependent protein transport pathways. J Cell Biol 146: 45-56
Nilsson R, Brunner J, Hoffman NE, van Wijk KJ (1999) Interactions of ribosome nascent chain complexes of the chloroplast-encoded D1 thylakoid membrane protein with cpSRP54. EMBO J 18: 733-742
Nilsson R, van Wijk KJ (2002) Transient interaction of cpSRP54 with elongating nascent chains of the chloroplast-encoded D1 protein; 'cpSRP54 caught in the act'. FEBS Lett 524: 127-133
Ossenbühl F, Göhre V, Meurer J, Krieger-Liszkay A, Rochaix JD, Eichacker LA (2004) Efficient assembly of photosystem II in Chlamydomonas reinhardtii requires Alb3.1p, a homolog of Arabidopsis ALBINO3. Plant Cell 16: 1790-1800
Ossenbühl F, Inaba-Sulpice M, Meurer J, Soll J, Eichacker LA (2006) The synechocystis sp PCC 6803 oxa1 homolog is essential for membrane integration of reaction center precursor protein pD1. Plant Cell 18: 2236-2246
Pasch JC, Nickelsen J, Schünemann D (2005) The yeast split-ubiquitin system to study chloroplast membrane protein interactions. Appl Microbiol Biotechnol 69: 440-447
Payan LA, Cline K (1991) A stromal protein factor maintains the solubility and insertion competence of an imported thylakoid membrane protein. J Cell Biol 112: 603-613

8 Literaturverzeichnis 127
Pool MR (2005) Signal recognition particles in chloroplasts, bacteria, yeast and mammals (review). Mol Membr Biol 22: 3-15
Preuss M, Ott M, Funes S, Luirink J, Herrmann JM (2005) Evolution of mitochondrial oxa proteins from bacterial YidC. Inherited and acquired functions of a conserved protein insertion machinery. J Biol Chem 280: 13004-13011
Ravaud S, Stjepanovic G, Wild K, Sinning I (2008) The crystal structure of the periplasmic domain of the Escherichia coli membrane protein insertase YidC contains a substrate binding cleft. J Biol Chem 283: 9350-9358
Reyes-Prieto A, Weber AP, Bhattacharya D (2007) The origin and establishment of the plastid in algae and plants. Annu Rev Genet 41: 147-168
Richter S, Lamppa GK (1998) A chloroplast processing enzyme functions as the general stromal processing peptidase. Proc Natl Acad Sci U S A 95: 7463-7468
Richter ML, Hein R, Huchzermeyer B (2000) Important subunit interactions in the chloroplast ATP synthase. Biochim Biophys Acta 1458: 326-342
Robson A, Collinson I (2006) The structure of the Sec complex and the problem of protein translocation. EMBO Rep 7: 1099-1103
Roy LM, Barkan A (1998) A SecY homologue is required for the elaboration of the chloroplast thylakoid membrane and for normal chloroplast gene expression. J Cell Biol 141: 385-395
Saaf A, Monne M, de Gier JW, von Heijne G (1998) Membrane topology of the 60-kDa Oxa1p homologue from Escherichia coli. J Biol Chem 273: 30415-30418
Samuelson JC, Chen M, Jiang F, Moller I, Wiedmann M, Kuhn A, Phillips GJ, Dalbey RE (2000) YidC mediates membrane protein insertion in bacteria. Nature 406: 637-641
Saracco SA, Fox TD (2002) Cox18p is required for export of the mitochondrially encoded Saccharomyces cerevisiae Cox2p C-tail and interacts with Pnt1p and Mss2p in the inner membrane. Mol Biol Cell 13: 1122-1131
Schünemann D, Amin P, Hartmann E, Hoffman NE (1999) Chloroplast SecY is complexed to SecE and involved in the translocation of the 33-kDa but not the 23-kDa subunit of the oxygen-evolving complex. J Biol Chem 274: 12177-12182
Schünemann D, Gupta S, Persello-Cartieaux F, Klimyuk VI, Jones JD, Nussaume L, Hoffman NE (1998) A novel signal recognition particle targets light-harvesting proteins to the thylakoid membranes. Proc Natl Acad Sci U S A 95: 10312-10316
Schünemann D (2004) Structure and function of the chloroplast signal recognition particle. Curr Genet 44: 295-304
Schünemann D (2007) Mechanisms of protein import into thylakoids of chloroplasts. Biol Chem 388: 907-915
Scotti PA, Urbanus ML, Brunner J, de Gier JW, von Heijne G, van der Does C, Driessen AJ, Oudega B, Luirink J (2000) YidC, the Escherichia coli homologue of mitochondrial Oxa1p, is a component of the Sec translocase. EMBO J 19: 542-549
Sedgwick SG, Smerdon SJ (1999) The ankyrin repeat: a diversity of interactions on a common structural framework. Trends Biochem Sci 24: 311-316
Serek J, Bauer-Manz G, Struhalla G, van den Berg L, Kiefer D, Dalbey R, Kuhn A (2004) Escherichia coli YidC is a membrane insertase for Sec-independent proteins. EMBO J 23: 294-301

8 Literaturverzeichnis 128
Sivaraja V, Kumar TK, Leena PS, Chang AN, Vidya C, Goforth RL, Rajalingam D, Arvind K, Ye JL, Chou J, Henry R, Yu C (2005) Three-dimensional solution structures of the chromodomains of cpSRP43. J Biol Chem 280: 41465-41471
Soll J, Schleiff E (2004) Protein import into chloroplasts. Nat Rev Mol Cell Biol 5: 198-208
Spence E, Bailey S, Nenninger A, Moller SG, Robinson C (2004) A homolog of Albino3/OxaI is essential for thylakoid biogenesis in the cyanobacterium Synechocystis sp. PCC6803. J Biol Chem 279: 55792-55800
Stengel KF, Holdermann I, Cain P, Robinson C, Wild K, Sinning I (2008) Structural basis for specific substrate recognition by the chloroplast signal recognition particle protein cpSRP43. Science 321: 253-256
Stengel KF, Holdermann I, Wild K, Sinning I (2007) The structure of the chloroplast signal recognition particle (SRP) receptor reveals mechanistic details of SRP GTPase activation and a conserved membrane targeting site. FEBS Lett 581: 5671-5676
Sun C, Rusch SL, Kim J, Kendall DA (2007) Chloroplast SecA and Escherichia coli SecA have distinct lipid and signal peptide preferences. J Bacteriol 189: 1171-1175
Sundberg E, Slagter JG, Fridborg I, Cleary SP, Robinson C, Coupland G (1997) ALBINO3, an Arabidopsis nuclear gene essential for chloroplast differentiation, encodes a chloroplast protein that shows homology to proteins present in bacterial membranes and yeast mitochondria. Plant Cell 9: 717-730
Szyrach G, Ott M, Bonnefoy N, Neupert W, Herrmann JM (2003) Ribosome binding to the Oxa1 complex facilitates co-translational protein insertion in mitochondria. EMBO J 22: 6448-6457
Theg SM, Cline K, Finazzi G, Wollman FA (2005) The energetics of the chloroplast Tat protein transport pathway revisited. Trends Plant Sci 10: 153-154
Thompson SJ, Kim SJ, Robinson C (1998) Sec-independent insertion of thylakoid membrane proteins. Analysis of insertion forces and identification of a loop intermediate involving the signal peptide. J Biol Chem 273: 18979-18983
Timmis JN, Ayliffe MA, Huang CY, Martin W (2004) Endosymbiotic gene transfer: organelle genomes forge eukaryotic chromosomes. Nat Rev Genet 5: 123-135
Tu CJ, Peterson EC, Henry R, Hoffman NE (2000) The L18 domain of light-harvesting chlorophyll proteins binds to chloroplast signal recognition particle 43. J Biol Chem 275: 13187-13190
Tu CJ, Schünemann D, Hoffman NE (1999) Chloroplast FtsY, chloroplast signal recognition particle, and GTP are required to reconstitute the soluble phase of light-harvesting chlorophyll protein transport into thylakoid membranes. J Biol Chem 274: 27219-27224
Tzvetkova-Chevolleau T, Hutin C, Noel LD, Goforth R, Carde JP, Caffarri S, Sinning I, Groves M, Teulon JM, Hoffman NE, Henry R, Havaux M, Nussaume L (2007) Canonical signal recognition particle components can be bypassed for posttranslational protein targeting in chloroplasts. Plant Cell 19: 1635-1648

8 Literaturverzeichnis 129
Urbanus ML, Scotti PA, Froderberg L, Saaf A, de Gier JW, Brunner J, Samuelson JC, Dalbey RE, Oudega B, Luirink J (2001) Sec-dependent membrane protein insertion: sequential interaction of nascent FtsQ with SecY and YidC. EMBO Rep 2: 524-529
van Bloois E, Koningstein G, Bauerschmitt H, Herrmann JM, Luirink J (2007) Saccharomyces cerevisiae Cox18 complements the essential Sec-independent function of Escherichia coli YidC. FEBS J 274: 5704-5713
van Bloois E, Nagamori S, Koningstein G, Ullers RS, Preuss M, Oudega B, Harms N, Kaback HR, Herrmann JM, Luirink J (2005) The Sec-independent function of Escherichia coli YidC is evolutionary-conserved and essential. J Biol Chem 280: 12996-13003
van der Laan M, Bechtluft P, Kol S, Nouwen N, Driessen AJ (2004) F1F0 ATP synthase subunit c is a substrate of the novel YidC pathway for membrane protein biogenesis. J Cell Biol 165: 213-222
van der Laan M, Houben EN, Nouwen N, Luirink J, Driessen AJ (2001) Reconstitution of Sec-dependent membrane protein insertion: nascent FtsQ interacts with YidC in a SecYEG-dependent manner. EMBO Rep 2: 519-523
Vörös (2007) Proteinintegration in die Thylakoidmembran von Arabidopsis thaliana- Untersuchung von Protein-Protein-Interaktionen des integralen Membranproteins Albino3 (Masterarbeit, Ruhr-Universität Bochum)
Walter M, Chaban C, Schutze K, Batistic O, Weckermann K, Nake C, Blazevic D, Grefen C, Schumacher K, Oecking C, Harter K, Kudla J (2004) Visualization of protein interactions in living plant cells using bimolecular fluorescence complementation. Plant J 40: 428-438
Weeden NF (1981) Genetic and biochemical implications of the endosymbiotic origin of the chloroplast. J Mol Evol 17: 133-139
Woolhead CA, Thompson SJ, Moore M, Tissier C, Mant A, Rodger A, Henry R, Robinson C (2001) Distinct Albino3-dependent and -independent pathways for thylakoid membrane protein insertion. J Biol Chem 276: 40841-40846
Xie K, Dalbey RE (2008) Inserting proteins into the bacterial cytoplasmic membrane using the Sec and YidC translocases. Nat Rev Microbiol 6: 234-244
Yamamoto HY, Bassi R (1996). Carotenoids: Localization and function. In Oxygenic Photosynthesis: The Light Reactions, D.R. Ort and C.F. Yocum, eds (Dordrecht, The Netherlands: Kluwer Academic Publishers) 539-563
Yen MR, Harley KT, Tseng YH, Saier MH, Jr. (2001) Phylogenetic and structural analyses of the oxa1 family of protein translocases. FEMS Microbiol Lett 204: 223-231
Yi L, Dalbey RE (2005) Oxa1/Alb3/YidC system for insertion of membrane proteins in mitochondria, chloroplasts and bacteria (review). Mol Membr Biol 22: 101-111
Yuan J, Henry R, McCaffery M, Cline K (1994) SecA homolog in protein transport within chloroplasts: evidence for endosymbiont-derived sorting. Science 266: 796-798
Yuan J, Kight A, Goforth RL, Moore M, Peterson EC, Sakon J, Henry R (2002) ATP stimulates signal recognition particle (SRP)/FtsY-supported protein integration in chloroplasts. J Biol Chem 277: 32400-32404

8 Literaturverzeichnis 130
Zhang L, Paakkarinen V, Suorsa M, Aro EM (2001) A SecY homologue is involved in chloroplast-encoded D1 protein biogenesis. J Biol Chem 276: 37809-37814
Zhang YJ, Tian HF, Wen JF (2009) The evolution of YidC/Oxa/Alb3 family in the three domains of life: a phylogenomic analysis. BMC Evol Biol 9: 137
Zygadlo A, Robinson C, Scheller HV, Mant A, Jensen PE (2006) The properties of the positively charged loop region in PSI-G are essential for its "spontaneous" insertion into thylakoids and rapid assembly into the photosystem I complex. J Biol Chem 281: 10548-10554

9 Abkürzungsverzeichnis 131
9. Abkürzungsverzeichnis
Ala Alanin
Alb3 Albino3
Ank Ankyrin repeat
Aqua dest. lat. Aqua destillata
AS Aminosäure(n)
ATP Adenosintriphosphat
BCA Bicinchoninsäure
bp Basenpaar(e)
BSA engl. bovine serum albumin
CD Chromodomäne
cpSRP engl. chloroplast signal recognition particle
DEPC engl. Diethylpyrocarbonate
DMSO Dimethylsulfoxid
DNA Desoxyribonukleinsäure
E. coli Escherichia coli
EDTA Ethylendiamintetraacetat
GST Glutathion S-Transferase
GTP Guanosintriphosphat
His Histidin
IPTG Isopropyl-β-D-thiogalactopyranosid
kDa kilo-Dalton
LHCP engl. light harvesting chlorophyll a/b binding protein
mRNA messenger-RNA
Ni-NTA engl. Nickel-nitrilotriacetic acid
NTP Nukleosidtriphosphat
OD Optische Dichte
PAGE Polyacrylamid-Gelelektrophorese
RNA Ribonukleinsäure
rpm engl. rotations per minute
SDS engl. sodium dodecyl sulfate
TEMED N,N,N’,N’,-Tetramethylethylendiamin
UV ultraviolett
YFP engl. yellow fluorescent protein
Weitere allgemein gebräuchliche Abkürzungen und Maßeinheiten sind nicht gesondert
aufgeführt.

10 Abbildungsverzeichnis 132
10. Abbildungsverzeichnis
Abbildung 1-1: Transportwege der Proteine innerhalb des Chloroplasten......................3 Abbildung 1-2: Modell des co- und posttranslationalen cpSRP-Transportwegs .............7 Abbildung 1-3: Proteininteraktionen der Komponenten des cpSRP-Transportwegs ......9 Abbildung 1-4: Membrantopologie von Alb3, YidC und Oxa1.......................................11 Abbildung 1-5: Proteininsertionen durch Alb3, YidC und Oxa1 ....................................16 Abbildung 4-1: Alignment der Aminosäuresequenz von Alb3 und Alb4........................64 Abbildung 4-2: Test der Antikörper und Präimmunseren gegen die Antigene GST-
cAlb3 und cAlb4-his ............................................................................66 Abbildung 4-3: Test der Antikörper gegen die in vitro translatierten
Volllängenproteine ..............................................................................67 Abbildung 4-4: Test der Antikörper gegen Thylakoidmembranproteine........................68 Abbildung 4-5: Alb3 interagiert mit cpSRP43 in Arabidopsis thaliana Protoplasten .....73 Abbildung 4-6: Expressionsnachweis der YFP-Fusionsproteine cpSRP43-nYFP,
Alb3-cYFP und Alb4-cYFP..................................................................75 Abbildung 4-7: Schematische Darstellung der vorhergesagten Membrantopologie
von Alb3 und Alb4 ...............................................................................76 Abbildung 4-8: Identifizierung der Alb3 Bindestelle für das cpSRP43 in Arabidopsis
Protoplasten ........................................................................................78 Abbildung 4-9: Alignment der YFP-Fusionsproteine zur Identifizierung der
Bindestelle...........................................................................................79 Abbildung 4-10: Peptide Scanning von Alb3 Peptiden mit rekombinantem cpSRP43..81 Abbildung 4-11: Alignment von Alb3 und Alb4 im Bereich des Bindemotivs ................82 Abbildung 4-12: Expressionsnachweis ausgewählter Alb3 Deletions- und
Mutationskonstrukte in Arabidopsis Protoplasten ...............................84 Abbildung 4-13: Pull down Analysen von rekombinantem cpSRP43 und cpSRP54
mit in vitro Translationsprodukten von Alb3 und Alb4 .........................88 Abbildung 4-14: Einfluss eines synthetischen Peptids von Alb3 auf die
Thylakoidmembranbindung von cpSRP43..........................................90 Abbildung 4-15: Schematische Darstellung des cpSRP43 ...........................................91 Abbildung 4-16: Thylakoidmembran-Bindungstest mit GST-cpSRP43
Deletionsproteinen (I)..........................................................................92 Abbildung 4-17: Thylakoidmembran-Bindungstest mit GST-cpSRP43
Deletionsproteinen (II).........................................................................94 Abbildung 4-18: Crosslinking von Alb4 und CF1β .........................................................96 Abbildung 4-19: Alb4 interagiert mit der ATP-Synthase Untereinheit CF0II in
Arabidopsis thaliana Protoplasten.......................................................98 Abbildung 4-20: Expressionsnachweis der YFP-Fusionsproteine Alb4-cYFP und
CF0II-nYFP..........................................................................................99 Abbildung 4-21: Alb4 cofraktioniert mit den ATP-Synthase Untereinheiten................100 Abbildung 4-22: Coimmunpräzipitation von Alb3, Alb4 und cpSecY...........................101 Abbildung 5-1: Alignment der C-terminalen Proteinsequenzen der Alb3 Homologe ..108

10 Abbildungsverzeichnis 133
Abbildung 5-2: Alignment des Alb3 aus Arabidopsis thaliana mit den Alb4 Proteinen anderer Streptophyten ......................................................................109
Abbildung 5-3: Proteininteraktionen des cpSRP43.....................................................112 Abbildung 5-4: Modell des posttranslationalen cpSRP-Transportwegs ......................114

11 Tabellenverzeichnis 134
11. Tabellenverzeichnis
Tabelle 3-1: Chemikalienliste ........................................................................................18 Tabelle 3-2: Verbrauchsmaterial ...................................................................................19 Tabelle 3-3: Kits ............................................................................................................19 Tabelle 3-4: Geräte .......................................................................................................20 Tabelle 3-5: Verwendete Plasmide ...............................................................................22 Tabelle 3-6: Escherichia coli-Stämme...........................................................................29 Tabelle 3-7: Saccharomyces cerevisiae-Stamm...........................................................29 Tabelle 3-8: Trenngel pH 8,8 (4 Minigele):....................................................................46 Tabelle 4-1: Alb3 interagiert mit cpSRP43 im Split-Ubiquitin System ...........................71 Tabelle 4-2: Interaktionstests im Split-YFP System......................................................74 Tabelle 4-3: Negativkontrollen der Interaktionstests im Split-YFP System ...................74 Tabelle 4-4: Interaktion der Alb3 Deletions- und Mutationsproteine mit cpSRP43-
nYFP in Arabidopsis Protoplasten ............................................................83 Tabelle 4-5: Interaktionstest von Alb3 Deletionsproteinen mit cpSRP43 im Hefe
Split-Ubiquitin System ...............................................................................85

12 Anhang 135
12. Anhang
12.1 Publikationen
Naomi J. Marty*, Nathaniel E. Lewis*, Thomas Bals*, Karuppanan Muthusamy Kathir, Dakshinamurthy Rajalingam, Alicia D. Kight, Beatrix Dünschede, Aniko Vörös, Thallapuranam Krishnaswamy Suresh Kumar, Ralph L. Henry, Danja Schünemann, and Robyn L. Goforth A Dynamic CpSRP43-Alb3 Interaction Mediates
Translocase Regulation of CpSRP Targeting Components; eingereicht * gleichberechtigte Erstautoren
Benz M., Bals T., Gügel I.L., Piotrowski M., Kuhn A., Schünemann D., Soll J., Ankele E. (2009) Alb4 of Arabidopsis promotes assembly and stabilisation of a none
chlorophyll-binding photosynthetic complex, the CF1CF0-ATP synthase. Mol. Plant; im
Druck
Gerdes L*, Bals T*, Klostermann E, Karl M, Philippar K, Hunken M, Soll J, Schünemann D (2006) A second thylakoid membrane-localized Alb3/OxaI/YidC
homologue is involved in proper chloroplast biogenesis in Arabidopsis thaliana. J Biol Chem 281: 16632-16642
* gleichberechtigte Erstautoren

12 Anhang 136
12.2 Lebenslauf
Persönliche Daten: Name: Bals
Vorname: Thomas
Geburtsdatum : 21.08.1979
Geburtsort: Unna
Staatsangehörigkeit: deutsch
Schulische Ausbildung: 06/2000 Abitur, Walburgisgymnasium Menden
Zivildienst: 08/2000-06/2001 Zivildienst
Studium: 10/2001-04/2006 Studium der Biologie an der Ruhr-Universität Bochum
(Fakultät für Biologie und Biotechnologie), Abschlussarbeit:
„Untersuchungen zum plastidären Proteintransport von Arabidopsis
thaliana: Identifizierung und Charakterisierung eines Alb3-
Homologs“ (Lehrstuhl für Allgemeine und Molekulare Botanik,
Betreuerin: Frau Prof. Dr. Danja Schünemann), Abschluss als
Diplom Biologe
Promotion: seit 05/2006 Wissenschaftlicher Mitarbeiter an der Ruhr-Universität
Bochum (Fakultät für Biologie und Biotechnologie) im Rahmen
eines Promotionsstudiengangs (Lehrstuhl für Pflanzenphysiologie,
Betreuerin: Frau Prof. Dr. Danja Schünemann)
06/2007 Aufnahme in die Research School der Ruhr-Universität Bochum

12 Anhang 137
12.3 Danksagung
Ich danke Frau Prof. Dr. Danja Schünemann sehr herzlich für die Überlassung des
aktuellen und interessanten Themas, die Gewährleistung optimaler Arbeitsbedingungen
und die fortwährende Unterstützung während meiner Doktorarbeit.
Herrn Prof. Dr. Matthias Rögner danke ich sehr für die Übernahme des Korreferats.
Ein besonderer Dank gilt auch Silke Funke, die mich hervorragend in allen technischen
Herausforderungen unterstützt hat und so wesentlich zum Gelingen dieser Arbeit
beigetragen hat.
Des Weiteren danke ich Christine, Bea und Chantal aus meiner Arbeitsgruppe für die sehr
gute Zusammenarbeit und die überaus gute Arbeitsatmosphäre.
Allen Mitarbeitern der Lehrstühle „Allgemeine und Molekulare Botanik“ und
„Pflanzenphysiologie“ danke ich für die stetige Hilfsbereitschaft und das gute Arbeitsklima.
Ich danke dem SFB480 (Molekulare Biologie komplexer Leistungen von botanischen
Systemen, Teilprojekt B11: Analyse der Proteintransport-Mechanismen in Chloroplasten
(Prof. Dr. Danja Schünemann)) und der Research School der Ruhr-Universität Bochum
für die finanzielle Unterstützung dieser Arbeit.
Danken möchte ich auch meinen Freunden, die mir geholfen haben, immer den nötigen
Abstand vom Labor zu gewinnen.
Schließlich möchte ich mich noch ganz besonders bei meinen Eltern, meinem Bruder und
meiner Freundin bedanken, die mich während dieser Zeit zuverlässig unterstützt haben.

12 Anhang 138
ERKLÄRUNG
Hiermit erkläre ich, dass ich die Arbeit selbständig verfasst und bei keiner anderen
Fakultät eingereicht und dass ich keine anderen als die angegebenen Hilfsmittel
verwendet habe. Es handelt sich bei der heute von mir eingereichten Dissertation um
sechs in Wort und Bild völlig übereinstimmende Exemplare.
Weiterhin erkläre ich, dass digitale Abbildungen nur die originalen Daten enthalten und in
keinem Fall inhaltsverändernde Bildbearbeitung vorgenommen wurde.
Bochum, den 13.10.2009
____________________
Thomas Bals





























