Embed Size (px)
Citation preview

Zeitschrift fur Allgemeine Mikrobiologie 1 7
(Instit u t fur Physiologische und Biologische Chemie dcr Humboldt-Universitat Berlin und Insti- tu t fur Pflanzenschutzforschung der Akademie der Landwirtschaftswissenschaften der DDK,
Kleinmachnow)
Wirkmechanismus der Atmungshemmung durch die System- fungicide der Carboxingruppe. Wirkung von Oxathiinderivaten und -analoga auf nichtphosphorylierende submitochondriale
Partikeln aus Rinderherz sowie Trametes versicolor und Trichoderma viridel )
M’. MULLER, T. SCHEWE, H. L Y R und D. ZANKE
5 1977 359-372
(Eingegangen am 26. 3.1976)
23 Derivate und strukturanaloge Verbindungen des Carboxins (5,6-Dihydro-2-methyl-1,4- oxathiin-3-carboxanilid) wurden im Succinat-Cytochrom c-Oxydoreduktasesystem und im NADH- Oxydasesystem von nichtphosphorylierenden Elektronentransportpartikeln ( ETP)2) aus Rinder- herzmitochondrien getestet. Einige dieser Verbindungen wurden dariiber hinaus an partikuliirer Succinatdehydrogenase aus dem carboxinempfindlichen Pilz Trarnetes versicolor und der carboxin- unempfindlichen Art Trichoderma viride gepriift. Aus den Werten fur die Hemmaktivitiiten er- geben sich folgende SchluRfolgerungen:
1. Die Wirksamkciten der Oxathiinderivate und -analogs im Succ.-Cyt.c-OR-System vonRinder- herz-ETP korrclieren hoch signifikant (Irrtumswahrscheinlichkeit <0,1%) mit ihren Wirkungen auf partikuliire Succ.-DH aus den carboxinempfindlichen Pilzarten Trametes versicolor sowie Usti- lago maydis und Cryptococcus laurentii (WHITE u. THORN 1975), nicht aber mit dem cntsprechenden System aus Trichoderma viride. Rinderherz-ETP sind somit ein geeignetes Modellobjekt fur den Carboxinrezeptor in carboxinempfindlichen Pilzen.
2. Die Wirksamkeiten auf die Succ.-DH-Systeme von Rinderherz-ETP und Trametes versicolor korrelieren grob mit dem Hydrophobizitatsterm Ig P, jedoch sind auch elektronische und sterische Einfliisse crkennbar.
3. Fur die Bindung des Carboxins am Rezeptor in der Succ.-DH-Region des Elektronentrans- portsystems wird ein Mehrzentrenmechanismus vorgeschlagen, a n dem folgende Zentren beteiligt sind: 1 elektrophiles Zentrum am C,-Atom des Oxathiinringes, je 1 nucleophiles Zentrum am 0- Atom und am N-Atom der Siiureamidgruppierung sowie 2 hydrophobe Zentren (Phenylrest der Anilidgruppierung und CH,-Gruppierung am Oxathiinring). Das S-Atom iibt eine stabilisierende Wirkung auf den Wirkstoff-Rezeptor-Komplex &us.
4. Die Ursache fur die Carboxinunempfindlichkeit mancher Pilzarten liegt wahrscheinlich in einer abweichenden Struktur des Succ.-DH-Komplexes; die dort ebenfalls auftretende Hemm- wirkung erfolgt vermutlich nach einem anderen Mechanismus.
Die Oxathiine, deren Hauptvertreter das Carboxin (= Vitavax = DCMO = F 480 = 5,6-Dihydro-2-inethy1-1,4-oxathiin-3-carboxanilid) ist, wurden erstmalig durch VON SCHMELING et al. (1966) als systemische Fungicide beschrieben, die eine besondere Selektivitat gegeniiber Basidiomyceten zeigen ( EDGINQTON u. BARRON 1967, LYR et al. 1971). Die Untersuchungen von LYR et al. (1971, 1972) MATHRE (1971) und WHITE
I ) Ein vorliiufigcr Bericht dieser Ergebnisse wurde bereits an anderer Stelle gegeben (LYR et al. 1975a).
*) Abkurzungen: ETP = Elektronentransportpartikeln; Succ.-DH = Succinatdehydrogenase; FeS-Protein = Nichthiimeisenschwefelprotein; lg P = Logarithmus des Octanol-Wasser-Vertei- lungskoeffizienten der Verbindung; ED,, = fur eine 50%ige Hemmung erforderliche Hemmstoff- konzentration; DCPIP = 2,6-Dichlorphenolindophenol; Succ.-DCPIP-OR = Succinat-DCPIP- Oxydoreduktase; Succ.-Cyt.c-OR = Succinat-Cytochromc-Oxydoreduktase; NADH-Ox. = NADH- Oxydase

360 W. MULLER, T. SCHEWE:, H. LYR und D. ZANKE
(1971) an empfindlichen und uneinpfindlichen Pilzen fuhrten zu den1 Ergebnis, daB Carboxin in sehr spezifischer Weise eine Reaktion in der Atmungskette hemmt, die unmittelbar der Succ.-DH nachgelagert ist. Diese Atmungshemmung konnte als primarer Angriffspunkt fur die fungicide Wirkung wahrscheinlich gemacht werden (WHITE 1971). Sie wurde an Mitochondrien aus Pilzen (LYR et al. 1973, MATHRE 1971a, WHITE 1971), hoherenpflanzen (MATHRE 1971a, SCHEWE et al. 1974) und Siiuge- tieren (MATHRE 1971a, SCHEWE et al. 1973) sowie an atmenden Menibranen des Bak- teriums Micrococcus denitrificans (TUCKER u. LILLICH 1974) nachgewiesen. Hinsicht- lich des exakten Wirkortes gelangten ~JLRICH u. MATHRE (1971) und kurzlich auch SINGER et al. (1975) zu der SchluBfolgerung, daB der Wirkort von Carboxin iriit den1 des TTEA (2-Thenoyltrifluoraceton) identisch sei. SCHEWE et al. (1973) fanden jedoch Unterschiede in der Wirkung beider Hemmstoffe, die auf einen im Vergleich Zuni TTFA substratferneren Wirkort fur Carboxin schlieflen lieBen. Auf Grund dieser Ergebnisse wurde postuliert, daB T T F A als Fe'II-Koniplexbildner am oxydierten Eisen des FeS-Proteins im Komplex I1 der Atmungskette nach GREEN angreift, wahrend Carboxin auf die reduzierte Form eines FeS-Proteins dieses Komplexes wirkt. Unter Beriicksichtigung der neueren Ergebnisse von Untersuchungen der FeS-Zen- tren des Succinat-Ubichinon-Reduktasekomplexes mittels der paramagnetischen Elek- tronenresonanzspektroskopie (SINGER et ul. 1975, OHNISHI et al. 1974) liegt die Ver- mutung nahe, dafl der Wirkort des Carboxins mit dein sog. HIPIP (high-potential iron- sulphur protein) identisch ist. Weiterhin schlossen LYR u. SCHEWE (1975) aus Unter- suchungen an einer carboxinenipfindlichen I'ilzart, die iiber den alternativen niito- chondrialen Atmungsweg verfiigt, daB dan rarboxinenipfindliche FeS-Protein bei den betreffenden Pilzen und hoheren Pflanzen gleichzeitig terminale Oxydase ist.
Ziel der vorliegenden Mitteilung ist es, aus deni Vergleich der Atmungskettenwirk- sarnkeit, einer Reihe von Oxathiinderivaten und -analoga weitere Aussagen iiber den Wirkmechanismus dieser Verbindungsklasse zu treffen. Zu diesem Zweck wurden als Testobjekt nichtphosphorylierende Elektronentransportpartikeln (ETP) aus Rinder- herzmitochondrien verwendet'.
Auf Grund ihrer Natur als Umstiilpungen der Mitochondrieninnenmembran treten bei ihnen keine Penetrationsharrieren auf, so dafl eine direkte Zugiinglichkeit der Wirkstoffe an den Rezeptor gewahrleistet ist. Weiterhin lassen sich die Atmungs- kette und ihre Teilreaktionen an diesem Objekt niethodisch einfacher niessen. Da die ETP nur die Enzymsysteme der Atmungskette enthalten, spielen Entgiftungs- oder Giftungsniechanismen sowie sekundare Effekte des Wirkstoffes auflerhalb der Atniungs- kette kauni eine Rolle. So wiirde z. B. das Salicylsaureanilid durch seine Wirkung als Entkoppler der oxydativen Phosphorylierung bei koniplexeren Testobjekten sto- rende Effekte verursachen. AuBerdeiii verfiigen tierische Atniungskettensystenie nicht iiber den methodisch storenden alternativen Oxydationsweg, wahrend andererseits die Hauptwege bei Pilzen und Tieren im wcsentlichen identisch sind. Untersuchungen iiber die Struktur-Wirkungsbeziehungen der Oxathiine wurden bisher an Pilzen auf zellularer Ebene (MATHRE 197lb, TEX HAKEN u. DUNN 1971) sowie an partiku- liirer Succ.-DH (WHITE 1971, WHITE u. THORN 1975) durchgefiihrt. Diese Systeme weisen jedoch die genannten Nachteile auf. I n der vorliegenden Mitteilung wird da- gegen erstnials ein standardisiertes tierisches Atniungskettensystenl benutzt und mit den fungalen Systemen verglichen.
Urn Riickschlusse auf die Ursache der Selektivitat gegeniiber verschiedenen Pilz- arten ziehen zu konnen, wurden die Wirkungen einiger Oxathiinderivate und -analoga auf die Succ.-DCPIP-OR-Aktivitiit von subniitochondrialen partikularen Prapara- tionen aus einer carboxinempfindlichen l'ilzart (Trametes versicolor) sowie einer un- empfindlichen Art (Trichoderrna viride) getestet. Die vorliegenden Ergebnisse wurden

Wirkmechanismus von Carboxin 36 I
untereinander sowie niit den Octanol-Wasser-Verteilungskoeffizienten (lg P) als Hydrophobizitiitsterm verglichen.
Material und Methoden E T P wurden aus Rinderherzmitochondrien mittels alkalischer Schwellung und Messerhomogeni-
sation nach einem modifizierten Verfahren (MULLER et al. 1977) nach CRANE et al. (1956) prapa- riert. Die Versuchspilze Trametes versicolor (Stamm 67a) und Trichoderma v i d e (Stamm I 15a) aus der Stammkulturensammlung des Institutes fur Pflanzenschutzforschung wurden auf Rota- tionsschiittlern submers auf 3% Malzmedium angezogen. Die Mitochondrienpraparation erfolgte in Anlehnung an MAY u. JACOB (1970). Die Mycelien.wurden abgesaugt, 3mal mit Eiswasser und lmalmit Isolationsmedium (Tris/HCl-Puffer 2,4 g/l, ADTA 24 mg/l, Saccharose 137 g/l) gewaschen und im Buhler-Homogenisator (30 sec) im Eisbad homogenisiert. Nach Verwerfen des 1000 x g (10 min) Sedimentes wurde die Mitochondrienfraktion bei 10000 x g (10 min) gewonnen, das Se- diment in Isolationsmedium resuspendiert und wieder sedimentiert, das Sediment in Isolations- medium aufgenommen und in einem POTTER-Homogenisator resuspendiert. Fur die Herstellung der submitochondrialen partikularen Praparation erwies sich 2maliges Einfrieren und Auftauen als am giinstigsten. Durch Zentrifugation bei 10000 x g wurden die unzerstorten Mitochondrien ent- fernt.
Die Oxathiinderivate und -analogs wurden in Dimethylformamid (vakuumdestilliert) gelost. Die Losungsmittelkonzentration betrug in den MeDansatzen maximal 5% bei den Rinderherz- ETP-Untersuchungen und 1% bei den Pilzuntersuchungen.
Die 2-Methyl-5,6-dihydro-1,4-oxathiincarbonsaure-(3) (Verb. 2) wurde nach VON SCHMELINQ et al. (1966) dargestellt. Die N-monosubstituierten 3-Carbonsaureamide des 2-Methyl-5,6-dihydro- 1,4-oxathiins (Verb. 1,3-10,15-20) wurden nach den bekannten Methoden (VON SCHMELINC et al. (1966) ; United States Rubber Company (1967)) aus 2-Methyl-5,6-dihydro-1,4-oxathiincarbon- slure-(3)-chlorid und dem entsprechenden primaren Amin bzw. Ammoniak synthetisiert. 2-Methyl- 5,6-dihydro-1,4-oxathiin-carbons~ure-(3)-phenylester (Verb. 14) wurde folgendermaDen prapa- riert: 3 3 7 g (0,02 Mol) 2-Methyl-5,6-dihydro-l,4-oxathiin-carbonsaure-(3)-chlorid wurden in 10 ml Chloroform gelost, rnit 1,88 g (0,02 Mol) Phenol sowie 2 ml Pyridin (uber KOH getrocknet und destilliert) versetzt und 2 Std. unter RfickfluB und FeuchtigkeitsausschluD erhitzt. Nach dem Abkuhlen wurde mit verdunnter Salzsaure und anschlieBend mit Wasser gewaschen. Das Chloro- form wurde i. V. entfernt. Es schieden sich weiDe Kristalle des 2-Methyl-5,6-dihydro-1,4-oxathiin- carbonsaure-(3)-phenylesters ab. Umkristallisation aus 50yoigem i-Propanol. Das 2-Methyl-5,6- dihydro-l,4-oxathiin-3-carboxanilid-4,4-dioxid (Verb. 13) wurde durch Oxydation von Carboxin mit H,O, erhalten (KULKA et al. 1968). 2-Methyl-5,6-dihydro-(4H)-pyran-3-carboxanilid (Verb. 12) wurde nach TEN HAKEN u. BAYR (1969) dargestellt. Salicylsaureanilid (Verb. 21) und 2-Jodbenz- anilid (Verb. 23) wurden nach den gebrauchlichen Methoden (GRIMMEL et al. 1946, RAPSON u. SHUTTLEWORTH 1941) synthetisiert. o-Toluylsaureanilid (Verb. 22) wurde analog uber o-Toluyl- saure (ZAUGQ u. RAPALIA 1947), Uberfiihriing rnit SOCI, in das Saurechlorid und Umsetzung rnit Anilin erhalten. Trans-Crotonsaureanilid (Verb. 11) erhielten wir von Prof. Dr. G. WESTPHAL~} (Humholdt-Universitat Berlin, Sektion Chemie). Von allen bisher in der Literatur nicht beschrie- benen Verbindungen wurden C-, H- und N-Elementaranalysen angefertigt. Sie stimmen rnit den berechneten Werten innerhalb der Fehlergrenzen iiberein. Die Schmelzpunkte aller Verbindungen sind in Tabelle 1 abgegeben.
Die Octanol-Wasser-Verteilungskoeffizienten der Oxathiinderivate und -analogs wurden aus der Literatur (MATHRE 1971) entnommen oder nach den gebrauchlichen Methoden (LEO et al. 1971) berechnet.
Die iibrigen Chemikalien waren folgender Herkunft : NADH, VEB Arzneimittelwerk Dresden; Cytochrom c, BOEHRINCER Mannheim GmbH; Bernsteinsaure, VEB Laborchemie Apolda (als Ka- liumsalz verwendet) ; Dinatriumsuccinat, Feinchemie K.-H. KALLIES KG ; KCN, CHEMAPOL, C'SSR (mit HCI neutralisiert) ; Dichlorphenolindophenol (DCPIP), G. SCHONERT K.-G., Leipzig.
Die Enzymaktivitaten in den Systemen der Rinderherz-ETP wurden durch kinetische photo- metrische Untersuchungen am registrierenden Spektralphotometer UNICAM SP. 800 A ermittelt. Dazu wurden die E T P im MeBmedium mit variierenden Konzentrationen an Hemmstoff 15 min bei 37 "C vorinkubiert. Alle Messungen erfolgten bei 37 "C (Kiivettenvolumen 1 ml, Schichtdicke 1 cm). Die Enzymaktivitaten der Kontrollansiitze betrugen in beiden MeDsystemen 0,100 f 0,020 AEa0 bzw. A E,,,/min bei einer ETP-Konzentration von etwa 90 pg Protein/Ansatz. Die Bestim- mung der Enzymaktivitaten wurde fur die NADH-Ox. nach SANADI et al. (1967) und fur die Succ.- Cyt.c-OR nach TISDALE (1967) (hierbei wurde Natriumazid durch Kaliumcyanid ersetzt sowie ADTA und Rinderserumalbumin weggelassen) durchgefuhrt.
Fur die Uberlassung von trans-Crotonsaureanilid mochten wir an dieser Stelle unseren herz- lichen Dank aussprechen.

362 W. MULLER, T. SCHEWE, H. LYR und D. ZANKE
Die Messung der Succinatdehydrogenase-Aktivitiit (Succ.-DCPIP-OR) in den fungalen sub- mitochondrialen partikuliiren Praparationen erfolgte kolorimetrisch mit Dichlorphenolindophenol (DCPIP) nach BERNATHU. SINQER (1962) in folgendem Ansatz: 1 ml Phosphatpuffer (0,3 M) pH 7,2; 0,3 ml KCN (0,l M); 1,0 ml Wirkstoff bzw. Losungsmittel, 0,2 ml submitochondriale partikuliire Priiparation; 0,3 ml DCPIP (0,001 M); O,2 ml Dinatriumsuccinat (0,2 M). Reaktionszeit 15 min bei 20 "C, Messung im SPEKOL oder UNIcm-Spektrophotometer bei 580 nm (l-cm-Kuvette).
Die Ermittlung der Halbhemmungskonzentrationen erfolgte durch Auftragung der prozentua- len Hemmung gegen die Konzentration im halblogarithmischen MaBstab.
Die Proteinbestimmung erfolgte nach Lomy et al. (1951). Die linearen Korrelationen und Re- gressionen erfolgten auf dem Kleinrechner Typ 9810 A Calculator Hewlett-Packard.
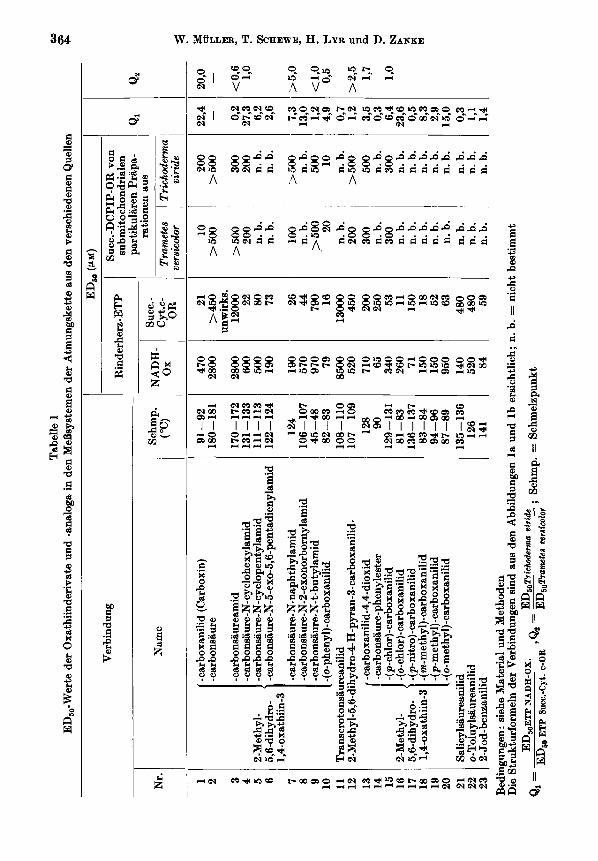
Ergehnisse Die Halbheinmungskonzentrationen der untersuchten Oxathiinderivate und -am-
loga in den vier MeBsystemen: das NADH-Ox.- und das Succ.-Cyt. c-OR-System der Rinderherz-ETP sowie die Succ.-DCPlP-OR-Systeme der submitochondrialen parti- kularen Praparationen aus Trametes versicolor und Trichoderma viride sind in Tabelle 1 aufgefuhrt.
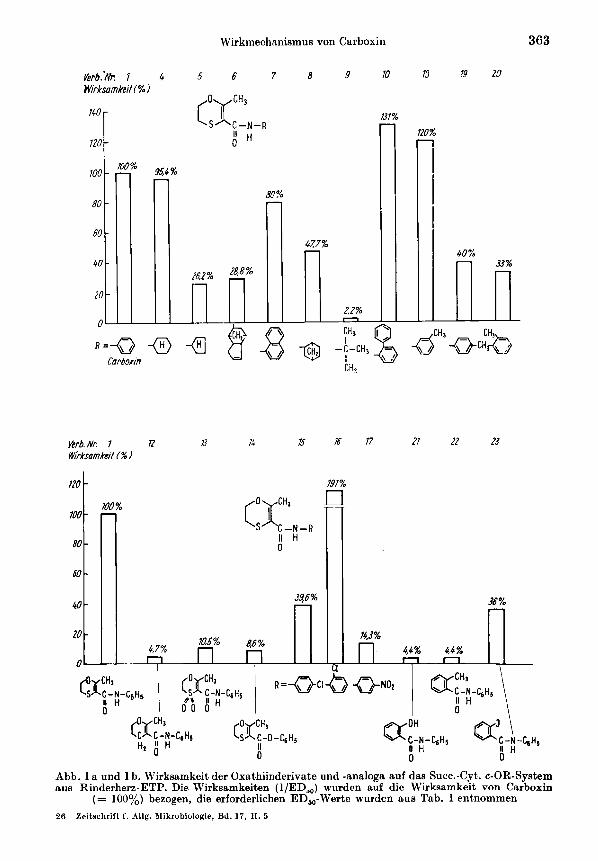
Zur Analyse des Wirkmechanismus der Oxathiine seien zuniichst die Hemmaktivi- tiiten im Succ-Cyt.c.-OR-System in den Rinderherz-ETP-betrachtet. Die Oxathiin- saure (Verb. 2) und das Oxathiinsaureamid (Verb. 3) sind praktisch unwirksani. Das ist ein qualitstiver Unterschied zu allen anderen Oxathiinderivaten, die durchweg wirksam waren - wenn auch teilweise erst bei hoherer Konzentration. Die strukturelle Besonderheit dieser unwirksamen Verbindungen besteht im Fehlen eines hydrophoben Restes, der somit als essentiell anzusehen ist. Abbildung l a zeigt, daB die Anwesen- heit einer beliebigen hydrophoben Gruppierung an dieser Stelle eine Wirkung bedingt. Die unterschiedlichen Aktivitaten der in Abbildung 1 a aufgefuhrten Verbindungen lassen sich jedoch nicht allein mit der differierenden Hydrophobizitat erklaren, son- dern es spielen wahrscheinlich auch sterische und elektronische Einflusse eine Rolle. Dafur sprechen die unterschiedlichen Aktivitaten der Verb. 1 und 4 einerseits und 5 und 8 andererseits sowie die dreifach hiihere Aktivitat der Verb. 18 gegenuber den Verb. 19 und 20 trotz jeweils etwa vergleichbarer Hydrophobizitat. Weiterhin zeigen die Verb. 5 und 6 etwa die gleiche Wirkung, obwohl sie eine unterschiedliche Hydrophobizitat aufweisen. Eine Erhohung der Hydrophobizitat fuhrt nicht zwangs- laufig zu einer Wirkungssteigerung (vgl. Verb. 6 und 7 mit 1).
Die zwar schwachen aber doch eindeutigen Wirkungen der Verb. 12 und 22 zeigen, daB der Oxathiinring nicht unbedingt erforderlich ist, sondern durch ahnliche Ring- systeme ersetzbar ist (Abb. lb). Die Ersetzbarkeit des Oxathiinringes durch Thiazol wurde von WHITE (1971, 1975) gezeigt. Von entscheidender Bedeutung ist offenbar das Vorliegen einer Acrylaniidgruppieruna. Das geht bereits au8 den Untersuchungen von TEN HAKEN 11. DUNN (1871) hervor, die die Wirkungen verschiedener Oxathiin- derivate und -analoga auf intakte Zellen von Uromyces fabae und Puccinia recondita halbquantitativ abgeschatzt hatten ; danach fuhrt eine Hydrierung dieser Doppel- bindungen sowie der Ersatz der Carbonyl- durch eine Thiocarbonylgruppierung zum Verlust bzw. zur Abschwachung der biologischen Wirkung. Bemerkenswert ist die Ersetzbarkeit des Oxathiins durch den Benzolring, wobei jedoch die Aktivitat stark abnimmt (Abb. lb, Verb. 22 und 23 gegenuber Verb. 1). Transcrotonsiiureanilid (Verb. 11) war in unseren Experimenten unwirksam, wahrend das cis-lsomere bei WHITE u. THORN (1975) schwach wirksam war. TEN HAKEN u. DUNN (1971) stellten auBerdem fest, daB die Methylgruppe in 2-Stellung der Verb. 12 bedingt durch eine Athylgruppe, nicht aber durch Wasserstoff, eine Phenyl- oder eine Hydroxymethylgmppe ersetzbar ist. Das zeigt, daB auch dieser Methylgruppe eine essentielle Bedeutung fur die Wir- kung zukoninit. Aus der gegenuber dem Carboxin deut,lich schwacheren Wirkung der Verb. 12, 13 und 22 (Abb. lb) ergibt sich, daB auch das Schwefelatom im Oxathiirlring eine Rolle beim Wirkmechanismus spielt, jedoch nicht essentiell ist.
Wirkmechanismus von Carboxin 363
Verb.’@. T 4 Wlrksarnkeif (% )
“4 120
?3 ?9 2.9 5 6 7 8 9 10
/ V H 3
R=a -Q -Q Carbgxin
Yerb.Nr. 7 I? 73 bvkksarnkeif I% I
700 %
40 in 20 0 47% n
J4 73
(;IcH3 C-N- I1 H 0
396%
76 77 27 22 23
3%
Abb. 1 a und 1 b. Wirksamkeit der Oxathiinderivate und -analogs auf das Succ.-Cyt. c-OR-System aus Rinderhcrz-ETP. Die Wirksamkeiten (l/ED,,) wurden auf die Wirksamkeit von Carboxin
(= 100%) bezogen, die erforderlichen ED,,-Werte wurden aus Tab. 1 entnommen 26 Zeitschrift f . Allg. Nikrobiologie, Bd. 17, 11. 5
Tab
elle
1
ED,,-
Wer
te
der
Oxa
thiin
deri
vate
und
-an
alog
s in
den
MeD
syst
emen
der
Atm
ungs
kette
aus
den
ver
schi
eden
en Q
uelle
n
2-M
ethy
l-
5,6-dihydro- '
1,4-oxathiin-3
Nr.
' -ca
rbox
anili
d (C
arbo
xin)
-c
arbo
ns&
ure
-car
bons
iiure
amid
-carbonsiiure-N-cyclohexylamid
-carbonsiiure-N-cyclopentylamid
-carbonsiiure-N-5-exo-5,6-pentadienylamid
-carbonsiiure-N-naphthylamid
-carbonsaure-N-2-exonorbornylamid
-carbonsiiure-N-t-butylamid
, -(o-pheny1)-carboxanilid
1 2 3 4 5 6 7 8 9 10
11
12
13
14
15
16
17
18
19
20
21
22
23
Ver
bind
ung
Nam
e
I E
Dl
I Rin
derh
erz-
ET
P
91 -92
180-181
170-172
131 -133
111-113
122-124
124
106-107
45 -48
82 -83
108-110
107
109
128 90
129 - 131
81-83
136-137
83-84
94-96
87-89
135-136
126
141
470
2800
2800
600
500
190
190
570
970 79
8500
520
710 65
340
260 71
150
150
950
140
520 84
succ
.-
OR
C
yt.e
- 21
> 450
unw
irks
. 12000 22
80
73
26
44
790 16
13000
450
200
250 53
11
150 18
52
63
480
480 59
Succ
.-DC
PIP-
OR
von
su
bmito
chon
dria
len
part
ikul
iiren
Pra
pa-
ratio
nen
aus
Trar
nete
a ve
rsie
olor
10
>50
0
>50
0 200
n. b
. n. b
.
100
n. b
. 20
n. b
. 200
300
n. b
. 300
n. b
. n.
b.
n. b
. n.
b.
n. b.
n. b
. n.
b.
n. b
.
-~
,> 500
Bed
ingu
ngen
: si
ehe
Mat
eria
l und
Met
hode
n D
ie S
truk
turf
orm
eln
der
Ver
bind
unge
n si
nd a
us d
en A
bbild
unge
n la u
nd l
b er
sich
tlich
; n.
b. =
nic
ht b
estim
mt
Fri
chal
erm
vi
ride
200
> 500
300
200
n. b
. n.
b.
>Boo
n.
b.
500 10
n. b
. >500
500
n. b
. 300
n. b
. n.
b.
n. b
. n.
b.
n. b
. n.
b.
n. b
. n.
b.
ED
So
~~
p
NA
DH
-OX
. ED
SoTr
ichod
srma
viri
dc ; S
chm
p. =
Sch
mel
zpun
kt
Q1 =
ED
,, E
TP
succ
.-cyt
. c-
OX '" =
ED
soTr
umG
tea w
emico
lor

Wirkmechanismus von Carboxin 365
Die uni etwa eine GroBenordnung geringere Wirksamkeit des Phenylesters der Oxa- thiinsiiure (Verb. 14) gegeniiber Carboxin (Abb. lb ) zeigt, daB vermutlich auch der Amid-Stickstoff am Wirkmechanismus beteiligt ist. Dafiir sprechen auch die elektro- nischen Substituenteneinfliisse am Phenylring. Elektronenziehende Substituenten (p-C1, p-NO,) senken die Hemmwirkung (Abb. lb , Verb. 15 und 17). Den gleichen Ef- fekt verursachen elektronendriickende Substituenten (p-CH,, o-CH,) ; wahrend die m- CH,-Substitution, die die Elektronendichte am N-Atom nicht wesentlich beeinflufit, sogar zu einer geringen Wirkungsverstiirkung fiihrt. Ini letzteren Falle konnte je- doch die Erhohung der Hydrophobizitiit ausschlaggebend sein (Abb. la, Verb. 18, 19 und 20). Die unterschiedlichen Aktivitiiten des 0-C1- und des p-C1-Derivates lassen sich dagegen offenbar nur durch sterische Einfliisse erkliiren. hfolge des gleichen Effektes von elektronisch verschieden wirkenden Substituenten konnte man schluflfolgern, daB ein gewisses Optimum der Elektronendichte am Phenylring fur die Wirkung notwen- dig ist, das gerade beim Carboxin erreicht wird.
Die Ergebnisse der Vergleiche der Wirksamkeiten der Oxathiinderivate und -analoga auf die einzelnen MeBsysteme mit den1 Hydrophobizitatsterm lg P und untereinander sind in Tabelle 2 dargestellt. Die Wirksamkeit im Succ.-Cyt. c-OR-System der Rinder- herz-ETP korreliert maBig mit der Hydrophobizitat (a). Ohne die Verbindungen 21, 22 und 23, bei denen der Oxathiinring durch einen Benzolring ersetzt ist, verbessert sich die Korrelation signifikant (b), es wird jedoch auch dadurch keine sehr gute Kor- relation erreicht. Diese Verbesserung und der relativ niedrige Korrelationskoeffizient bestiitigen, daB die Wirkung auBer von der Hydrophobizitiit auch noch von elektro- nischen und sterischen Parametern abhangen mul3.
Die Henimung des NADH-Ox.-Systems hat sich als geeignete MeBgroBe fur die uni- verselle Neigung einer Verbindung, hydrophobe Wechselwirkungen mit Protein- rezeptoren einzugehen, herausgestellt (DOVE 1974, FRANKE u. DOVE 1975). Folglich
Tabelle 2 Lineare Korrelationen und Regressionen der Logarithmen der Wirksamkeiten (Ig l/ED,,) der Oxa- thiinderivate und -analogs in den untersuchten Systemen gegen den Hydrophobizitiitsterm Ig P
sowie untereinander
I n ! Verglichene Parameter Signifikanz
von r bei P = 1%
a) ETPRH, sz - Ig P b) ETPRH, szl) - Ig P c) ETPRH, NA - Ig P d) ETPRH.NA') - la P
22 19 22 19 10 10 10 , ::
0,60 0,80 0,74 0,77 0,78 0,51 0,94 0,73 0,75
+ + + + + + - - -
20 15 9 9
23 27 12 21 24
Anstieg*) I Abschnitt der Regressionsgeraden
I
0,58 f 0,17 0,93 f 0,17 0,36 f 0,07 0,42 f 0,08 0,84 f 0,24 0,46 f 0,27 0,86 f. 0,11 0,56 f 0,18 0,90 f 0,28
2,72 f 0,38 2,26 f 0,33 2,77 f 0,16 2,68 f 0,17 2,12 f 0,44 2,60 f 0,51 0,30 f 0,42 1,28 f 0,70 0,43 f 0,96
Bei allen Rechnungen wurde Transcrotonsiiureanilid (Verb. 11) nicht mit einbezogen.
n = Anzahl der fur die Rechnung benutzten Wertepaare A Verbindungen; r = Korrelations- koeffizient ; Signifikanz + bedeutet, daD der Korrelationskoeffizient groBer als der Zufallshochst- korrelationskoeffizient ist; p = Irrtumswahrscheinlichkeit; S,,(%) = Durchschnittliche Stan- dardabweichung jedes einzelnen Punktes von der Regressionsgeraden in yo bezogen auf den Mittel- wert der Ordinatenwerte (1. Parameter) ?) Anstieg = Regressionskoeffizient; f Werte bei Anstieg und Abschnitt sind die Standardabwei- chungen dieser GroDen ; RH = Rinderherz; SZ = Succinat-Cytochrom c-Oxydoreduktasesystm; NA = NADH-Oxydasesystem ; SPP = submitochondriale partikuliire Priiparation des betreffenden Pilzes, Succinat-DCPIP-Oxydoreduktasesystem ; Tra = Trametea versicolor (empfind- licher Pilz) ; Tri = Trichoderma viride (unempfindlicher Pilz)
I ) Bei diesen Rechnungen wurden die Verb. 21,22 und 23 nicht rnit einbezogen.
26'

366 W. MULLER, T. SCIIEWE, H. LYR und D. ZANKE
ist erkliirlich, daB alle Verbindungen, die iin Succ.-Cyt.c-OR-System wirksani sind, auch das NADH-Ox.-System hemmen. Die Wirkungen der Oxathiinderivate und -ana- loga auf die NADH-Ox.-Aktivitat der Rinderherz-ETP sind jedoch bei den meisten Verbindungen deutlich schwacher, so daB eine spezifische Wirkung auf die Succinat- oxydation innerhalb der Atmungskette in tfbereinstininiung mit der Literatur (LYR et al. 1972, MATHRE 1971a, WHITE 1971, SCHEWE et aZ. 1973) vorliegt. Allerdings be- stehen hinsichtlich der Wirkortspezifitat betriichtliche Unterschiede zwischen den einzelnen Verbindungen, wie aus den Quotienten der Wirksamkeiten ini Succ.-Cyt.c- OR-System und im NADH-0x.system (Q1, Tab. 1) hervorgeht. Hohe Werte, wie sie fur die spezifisch wirkenden Vertreter der Oxathiingruppe gefordert werden niiissen, er- geben sich bei den Verb. 4,16,1,20 und 8. Dagegen weist die im Succ.-Cyt.c-OR-System recht gut wirksame Verb. 23 eine geringe Wirkortspezifitat auf ; die hohe Wirksam- keit im NADH-Ox.system laBt eine Reihe weiterer unspezifischer Wirkungen auf Mem- branfunktionen vermuten. Bei den Verbindungen, die im NADH-0x.system starker hemmen als im Succ.-Cyt.c-OR-System (Verb. 3, 11, 14, 17 und 21) ist zu beachten, daB mogliche biologische Effekte auf zellullirer Ebene nicht primar auf eine Carboxin- wirkung zuruckzufuhren sein mussen ; z. B. ist Verb. 21 (Salicylsaureanilid) in1 Gegen- satz zum Carboxin (MATHRE 1971a) als Entkoppler der oxydativen Phosphorylierung bekannt (WILLIAMSON u. METCALF 1967). Andererseits zeigen die Ergebnisse in Ta- belle 1 eindeutig, daB Salicylsaureanilid eine echte, wenn auch schwache Carboxin- wirkung zeigt (Abb. lb).
Die Wirksamkeiten im NADH-Ox.-System korrelieren ebenfalls mit der Hydropho- bizitat, allerdings auch nur mit eineni relativ geringen Korrelationskoeffizienten (c). Das Weglassen der Verbindungen 21,22 und 23 fuhrt hier nicht zu einer Verbesserung der Korrelation (d). Die Korrelation ist wesentlich schlechter als bei vergleichbaren Untersuchungen anderer Homologserien hydrophober Verbindungen (DOVE 1974, FF~ANKE u. DOVE 1975). M'ir erkliiren die relstiv schlechte Korrelation durch das Vor- handensein des zweiten, spezifischen Bindungsortes, den1 eigentlichen Wirkort von Carboxin in der Succ.-DH-Region, die vermutlich die Wechselwirkung zwischen Wirk- stoff und Rezeptorregion im NADH-Ox.-System beeinfluot.
Die Hemmwirkungen der Oxathiinderivate und -analoga auf das Succ.-DCPIP- OR-System der submitochondrialen partikularen Praparation aus Trametes versicolor, dem carboxinempfindlichen Pilz, korrelieren ebenfalls ma big mit der Hydrophobizitat (e). Der geringe Korrelationskoeffizient zeigt auch hier, daB die Hydrophobizitiit fur die Wirkung nicht allein entscheidend ist. Die Wirkungen auf das analoge System aus dem carboxinunenipfindlichen Pilz Trichoderma viride zeigen keine signifikante Kor- relation mit der Hydrophobizitat (f), allerdings ist die Differenz der Korrelations- koeffizienten beider Systeme zu lg P nicht sehr grog.
FRANKE u. OEHME (1973) stellten fest, daW bei hydrophoben Wechselwirkungen von Verbindungen kongener Serien mit Proteinen der Anstieg der ltegressionsgeraden (Re- gressionskoeffizient) bei der Auftragung der Logarithmen der biologischen Wirkung gegen lg P entweder 0,5 f 0,2 bei einer Bindung des Wirkstoffmolekiils an die Ober- flache des Rezeptorproteins oder 1,0 f 0,2 bei der vollstandigen Uberfiihrung in das Innere einer hydrophoben Tasrhe betragt. J)ie Regressionskoeffizienten &us dem Ver- gleich der Systeme Succ.-Cyt.c-OR aus Rinderherz-ETP und Succ.-DCPlP-OR aus Trametee versicolor mit lg P fallen in den Bereich von 1.0 f. 0,2 (b, e) wiihrend die der Systeme NADH-Ox. aus Rinderherz-ETP und Succ.-DCPIP-Olt aus Trichoderma viride mit lg P in den Bereich 0,5 f 0,2 (d, f ) fallen. Wir schluBfolgern daraus, daB es sich bei den beiden erstgenannten Systemen um eine spezifische Wechselwirkung der Oxathiine mit der Rezeptorregion, d. h. den FeS-Proteinen der Succ.-DH, handelt, wiihrend bei den anderen beiden Systemen eine unspezifische Bindung an die Ober- fliiche von Proteinkomponenten des Elektronentransportsystems vorliegt.
Wirkmechanismus von Carboxin 367
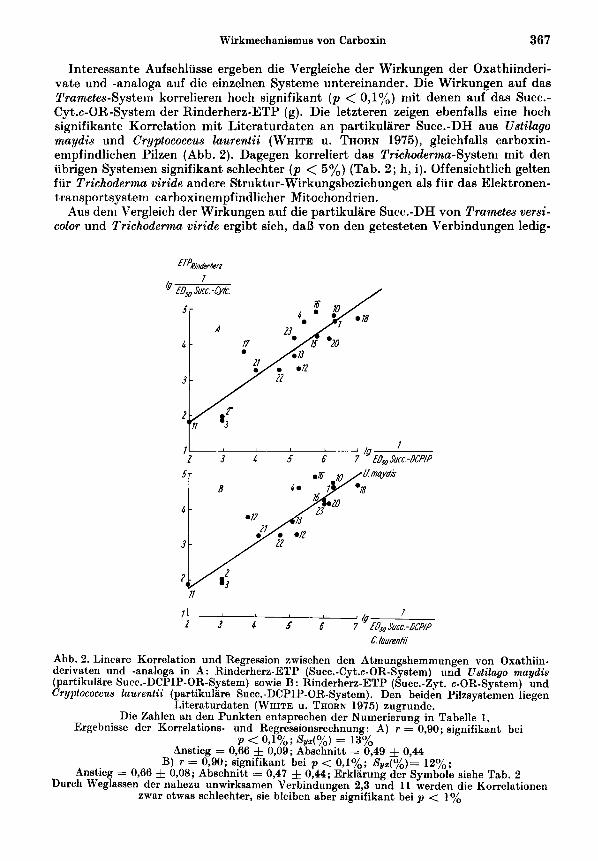
lnteressante Aufschliisse ergeben die Vergleiche der Wirkungen der Oxathiinderi- vate und -analoga auf die einzelnen Systeme untereinander. Die Wirkungen auf das Trametes-System korrelieren hoch signifikant ( p <: 0,1%) niit denen auf das Succ.- Cyt.c-OR-System der Rinderherz-ETP (g). Die letzteren zeigen ebenfalls eine hoch signifikante Korrelation mit Literaturdaten an partikularer Succ.-DH aus Ustilago maydis und Cryptococcus Zaurentii (WHITE u. THORN 1975), gleichfalls carboxin- empfindlichen Pilzen (Abb. 2). Dagegen korreliert das Trichoderma-System mit den ubrigen Systeinen signifikant schlechter ( p < 5 % ) (Tab. 2; h, i). Offensichtlich gelten fur Trichoderma viride andere Struktur-Wirkungsbeziehungen als fur das Elektronen- transportsystem carhoxinempfindlicher Mitochondrien.
Aus dem Vergleich der Wirkungen auf die partikulare Succ.-DH von Trametes versi- color und Trichoderma viride ergibt sich, da13 von den getesteten Verbindungen ledig-
7 ' FDN Succ. -oh
7-Q 7 2 3 4 5 6 7 mnsucc.-Dcp/p
6 Iaumtfii
Abb. 2. Lineare Korrelation und Regression zwischen den Atmungshemmungen von Oxathiin- derivaten und -analogs in A: Rinderherz-ETP (Succ.-Cyt.c-OR-System) und Ustilago maydis (partikuliire Sum-DCPIP-OR-System) sowie B: Rinderherz-ETP (Succ.-Zyt. c-OR-System) und Cryptococcus Zaurentii (partikuliire Succ.-DCPIP-OR-System). Den beiden Pilzsystemen liegen
Literaturdaten (WHITE u. THORN 1975) zugrunde. Die Zahlen an den Punkten entsprechen der Numerierung in Tabelle 1.
Ergebnisse der Korrelations- und Regressionsrechnung : A) r = 0,90; signifikant bei p < O J % ; %/z(%) = 13%
Anstieg = 0,66 0,09; Abschnitt = 0,49 -j= 0,44
0,08; Abschnitt = 0.47 & 0 , U ; Erklarung der Symbole siehe Tab. 2 B) r = 0,90; signifikant bei p < O,lyo; S V Z ( ~ ~ ) = 12%;
Anstieg = 0,66 Durch Weglassen der nahezu unwirksamen Verbindungen 2,3 und 11 nerden die Korrelationen
zwar etwas schlechter, sie bleiben aber signifikant bei p < 1%
368 W. MULLER, T. SCHEWE, H. LYR und D. ZANKE
lich das Carboxin eine ausgepragte Selektivitat zeigt. Die Verb. 3, 9 und 10 waren im Trichoderma-System sogar wirksamer als im Trametee-System. Bemerkenswert ist die hohe Wirksamkeit der Verb. 10 bei beiden Species. Eine solche geringe Speciesselekti- vitat konnte von Vorteil fur die Auswahl von Fungiciden mit breitem Wirkungs- spektrum sein, jedoch weist diese Verbindung eine nur maDige Wirkortspezifitat auf, so daS mit Nebenwirkungen zu rechnen ist.
Diskuseion
Die dargelegten Resultete zeigen, da13 folgende Molekulteile des Carboxins an der Wirkung beteiligt sind : Die hydrophobe Gruppe am Aniidstickstoff und die Methyl- gruppe in 2-Stellung des Oxathiinringes sowie die vinyloge Carbonylgruppe scheinen essentiell fur die Wirkung zu sein, und der Amidstickstoff und der Schwefel bedingen eine erhebliche Wirkungsverstarkung. Zu identischen SchluSfolgerungen gelangten TEN KAREN u. DUNN (1971) bei ihren Untersuchungen an intakten Pilzzellen, die je- doch auf Grund der Komplexizitat der Testsysteme mit einigen Unsicherheiten be- haftet waren. Aus dem Vergleich dieser und unserer Untersuchungen geht hervor, daD die bei Pilzen zu beobachtenden Struktur-Wirkungsbeziehungen auf die unterschied- lichen Affinitaten der Oxathiinderivate zu ihrem Rezeptor in der Atmungskette zu- riickzufuhren sind, wkhrend zellulare Entgiftung, Permeationsunterschiede und ahn- liche Faktoren von untergeordneter Bedeutung zu sein scheinen.
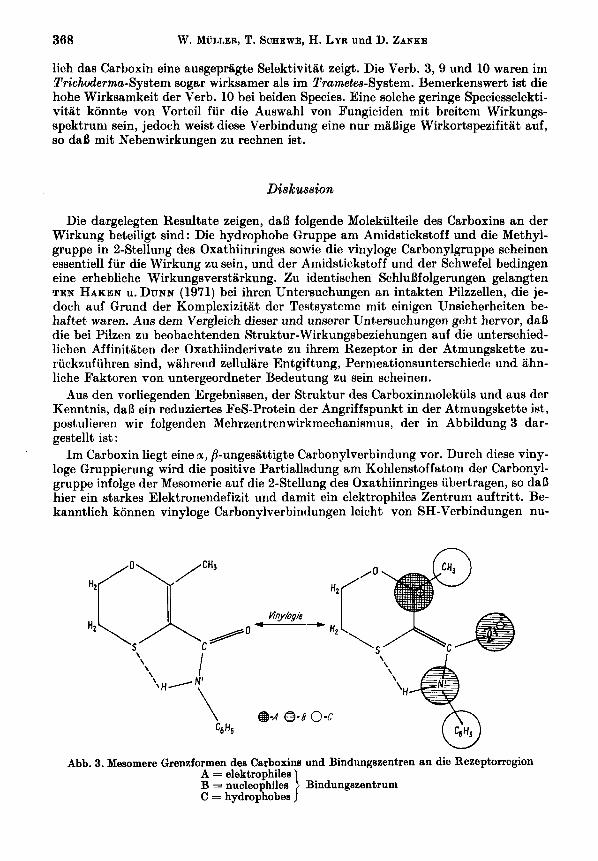
Bus den vorliegenden Ergebnissen, der Struktur des Carboxinniolekuls und aus der Kenntnis, daD ein reduziertes FeS-Protein der Angriffspunkt in der Atmungskette ist, postulieren wir folgenden Mehrzentrenwirkmechanismus, der in Abbildung 3 dar- gestellt ist :
Im Carboxin liegt eine a, /3-ungesattigte Carbonylverbindung vor. Durch diese viny- loge Gruppierung wird die positive Partialladung am Kohlenstoffatom der Carbonyl- gruppe infolge der Mesomerie auf die $-Stellung des Oxathiinringes iibertragen, so daD hier ein starkes Elektronendefizit und damit ein elektrophiles Zentrum auftritt. Be- kanntlich konnen vinyloge Carbonylverbindungen leicht von SH-Verbindungen nu-
\ \ . '\H/"
\ @ad e*B 0 - C '6H5
Abb. 3. Mesomere Grenzformen des Carboxine und Bindungszentren an die Rezeptorregion
Bindungezentrum C = hydrophobee

t
cleophil angegriffen werden, so geht z. B. die Addition von Methylmercaptan an Acro- lein bereits unter physiologischen Bedingungen (37 "C, ohne Katalysator) vor sich (PIERSON et al. 1948). In ahnlicher Weise konnten auch bestimmte S-Atome des FeS- Proteins, die durch den EinfluB des reduzierten Eisen-Zentralatoms eine hohe nucleo- phile Kraft besitzen sollten, mit dem elektrophilen C,-Atom des Oxathiinringes re- agieren. Dadurch wird die Carbonylgruppe am Sauerstoff stark negativ, und es konnte sich zusatzlich eine koordinative Bindun zwischen dem Sauerstoff und bestimmten Fe-Atomen der FeS-Proteine ausbilden. 7 7 ber Carbonyl-Sauerstoff-Atone wird z. B. auch TTFA am Eisen gebunden (REDFEARN et al. 1965). Der Amidstickstoff verstarkt verniutlich die Wirkung iiber sein freies Elektronenpaar durch eine weitere koordi- native Bindung an1 Eisen des FeS-Proteins. Die hydrophobe Phenylgruppe am Amid- stickstoff und die 2-Methylgruppe sind offenbar fur die Verankerung der Verbindung in der hydrophoben Rezeptorregion verantwortlich. Die Lokalisation der prostetischen Gruppe der FeS-Proteine in einer unpolaren Region geht auch daraus hervor, daB hydrophobe Eisenkoniplexbildner wie z. B. TTFA oder 4,7-Diphenyl-2,9-dimethyl- 1,lO-phenanthrolin, nicht aber hydrophile Verbindungen, wie z. B. ADTA, das Elek- tronentransportsysteni henimen (TAPPEL 1960). Die Funktion des Oxathiinringes konnte darin bestehen, die vinyloge Carbonylgruppe raumlich zu stabilisieren und die Methylgruppe in cis-Stellung zu fixieren. Der Schwefel konnte dabei iiber eine Wasser- stoffbriickenbindung (vgl. Abb. 3), deren Ausbildung in einem unpolaren Milieu ener- getisch begiinstigt ist, die Saureaniidgruppierung in ihrer raumlichen Lage noch zu- satzlich fixieren.
Die Bindung des Wirkstoffes am Rezeptor wird somit durch insgesanit funf Wirk- zentren - ein elektrophiles, zwei nucleophile und zwei hydrophobe - vermittelt. *Jedes der einzelnen Bindungszentren scheint einen wesentlichen Beitrag fur die Bin- dungsaffinitat zu liefern, denn bei Verlust eines von ihnen sinkt die Heminaktivitat uin eine GroBenordnung. So wird z. B. nur noch eine schwache Wirkung erreicht, wenn die Vinylogie nicht mehr gewahrleistet ist (vgl. Verb. 22). Einfiihrung eines elektro- nenziehenden Substituenten an 2-Stellung des Benzolringes (Verb. 23) vergroBert jedoch wiederuni die elektrophile Kraft an dieser Stelle und damit auch die biologische Aktivitat. &Tit diesem Mehrzentrenniechanismus la fit sich auch das anscheinende Op- timum der Elektronendichte ani Phenylring fiir die Wirkung erkliiren. Alle drei elek- tronisch wirkenden Bindungszentren, der Amidstickstoff, der Carbonylsauerstoff und das C,-Atom des Oxathiinringes sind durch die Doppelbindungen und freien Elek- tronenpaare konjugativ niiteinander verbunden, so da13 sich elektronische Einflusse aus den1 Phenylring auf alle drei Zentren auswirken miissen. Dnrch ein Elektronen- defizit wird die Nucleophilie des Amidstickstoffes und des Carbonylsauerstoffes ver- kleinert, die elektrophile Kraft des C,-Atoms aber erhoht, und durch einen Elektronen- iiberschuB werden die entgegengesetzten Verhaltnisse erreicht. Bei einer bestimmten optimalen Elektronendichte mussen alle drei Zentren iiber eine ausreichende Reakti- vitat verfiigen. Der Mehrzentrenwirkniechanisnius ist vermutlich die Grundlage fur die hohe Spezifitat der Wirkung, da nur die FeS-Proteine des Komplexes I1 und nicht die des Komplexes I angegriffen werden. denn die Spezifitat eines Angriffs ist um so groBer, je mehr Parameter an Wirkort und Reagens iibereinstimmen mussen. Anhand des hier vorgeschlagenen Wirkmechanismus des Carboxins wurde ein Wirkstoff-Re- zeptor-Model1 entwickelt, das Gegenstand einer spateren Puhlikation sein wird.
Die gute ubereinstimniung zwischen den Ergebnissen aus den Rinderherz-ETP Untersuchungen mit denen an den empfindlichen Pilzen Trametes versicolor sowie Ustilago ma& und Cryptococcus laurentii spricht fur einen prinzipiell gleichen Auf- bau der Carboxinrezeptorregion und damit gleichen Struktur-Wirkungsbeziehungen bei carboxinempfindlichen Pilzen und bei Saugetieren. Da eine gute Korrelation fun-

370 W. MULLER, T. SCHEWE, H. LYR und D. ZANRE
galer Atniungskettenwirksanikeit und fungistatischer Wirkung gefunden wurde (WHITE 1971, WHITE u. TIIORN 1975), korrelieren zwangslaufig die Wirksanikeiten auf das ETP-System aus Rinderherz mit der fungistatischen Wirkung. Das bedeutet, daS die Ergebnisse, die an dem Rinderherz-ETY-System erhalten wurden, auch fur das fungale System zutreffen sollten. Das im Gegensatz zu den fungalen Systemen wesent- lich einfacher handhabbare ETP-System ist somit ein geeignetes Modellobjekt fur den Carboxinrezeptor in carboxinempfindlichen Pilzen.
Trotz der vielfaltigen Ubereinstimmung des Verhaltens der Atmungskettensysteme aus Rinderherz-ETP und carboxinempfindlichen Pilzen gibt es auch Unterschiede. So reagieren Ustilago maydis und Cryptococcus laurentii (WHITE u. THORN 1975) gene- re11 etwa 30mal empfindlicher ; allerdings fanden wir keinen wesentlichen Unterschied in der Empfindlichkeit zwischen ETP und l'rametes versicolor. Weiterhin ergeben sich fur einzelne Vertreter der Oxathiine Unterschiede hinsichtlich der relativen Wirksam- keit bezogen auf das Carboxin. So ist die Verb. 4 sowohl bei C . lairrentii, U . maydis (WHITE u. THORN 1975) als auch T . versicolor wesentlich schwacher wirksam als Carb- oxin, wahrend sich beide Verbindungen im Rinderherz-ETP-System nicht unter- scheiden. Derartige Differenzen treten auch bei den Verb. 16, 18 und 21 (WHITE u. THORN 1975) auf. Daraus 1a13t sich ableiten, daS trotz identischem Wirkmechanismus subtile Unterschiede in der Rezeptorregion bei Rinderherz-ETP und entsprechenden Systenien aus carboxinempfindlichen Pilzen vorhanden sind. Ein solcher Unterschied besteht z. B. in der Anwesenheit des alternativen Oxydationsweges bei vielen carboxin- empfindlichen Pilzen (WHITE 1971). Die Beziehung zwischen Carboxinempfindlichkeit und alternativem Oxydationsweg ist verstandlich, da der Carboxinrezeptor und die alternative Oxydase vermutlich identisch sind (siehe Einleitung) (LYR u. SCHEWE 1975).
Auf Grund der Universalitat der Atniungskette ist fur ein systemisches Pflanzen- schutzmittel, das auf der Basis einer Atniungshemmung wirkt, die Selektivitat eine notwendige Forderung. Diese ist bei den Oxathiinen erfiillt, jedoch sind die Ursachen hierfur noch nicht endgultig geklart. Folgmde Faktoren konnen prinzipiell eine Selek- tivitat bedingen: a ) unterschiedliche Aufnahme durch die Zellen, b) Penetrations- unterschiede der Mitochondrieninnenmem bran, c) speciesabhangige Entgiftungs- und Abbaumechanismen oder Giftungsmechariismen, d) quantitative Unterschiede in der Empfindlichkeit des Rezeptors sowie e) die selektive Fahigkeit, den Atniungsketten- block durch Nebenwege der Atniungskette zu umgehen, wie sie bei pflanzlichen und fungalen Systemen bekannt sind. LYR et al. (1972) zeigten anhand eines Vergleiches von empfindlichen und unempfindlichen Hefearten, daB die selektive Wirkung des Carboxins nicht durch eine unterschiedliche Aufnahmefiihigkeit der Zellen bedingt ist. Eine Entgiftung in den Mitochondrien durch Oxydation des Carboxins zurn Sulf- oxid findet sowohl bci enipfindlichen als auch unempfindlichen Pilzarten in ahnlichem AusmaS statt (LYR et al. 1974) und kann ebenfalls nicht die Ursache fur die Selektivi- t a t sein. Ein weiterer Abbau aul3er der Oxydation Zuni Sulfoxid und Sulfon findet innerhalb von 24 Stunden in beiden Pilzarten nicht statt (LYR et al. 1974). Eine Gif- tung scheidet durch die gezeigte Wirkung auf ETY (SCHEWE et al. 1973) von vorn- herein aus. MATHRE (19'7la) diskutiert auf Grund unterschiedlicher Empfindlichkeiten von Mitochondrien verschiedener Quellen gegeniiber Carboxin und eineni Analogen, daB die Permeabilitat der Mitochondrierinienibranen eine wesentliche Rolle fur die Selektivitat spielen konnte. Da jedoch in unseren Untersuchungen mit subniitochon- drialen Praparationen gearbeitet wurde, trotzdem aber die Speciesselektivitat des Carboxins nachweisbar war, scheiden Pernieabilitatsunterschiede als Ursache prak- tisch aus. WHITE (1971) vermutet, daS die Carboxin-Empfindlichkeit der betreffenden Pilze auf eine veranderte Struktur der Succ.-DH zuriickzufiihren ist. Dem widerspricht jedoch die Tatsache, daS auch Mitochondrien aus Bohnen (MATHRE 1971a), Blumen-

Wirkmechanismus von Carboxin 371
kohl (SCHEWE et al. 1974) und Rattenleber (MATHRE 1971a, SCHEWE et al. 1973) sowie ETP aus Rinderherz (SCHEWE et al. 1973) in ihrer Atmungsaktivitiit gehemmt werden, so da13 die Carboxin-Enipfindlichkeit als eine universelle Eigenschaft des nornialen Elektronentransportes angesehen werden niu13.
Verniutlich ist unigekehrt eine abweichende Struktur der Succ.-DH-Region die Voraussetzung fiir eine Carboxin-Uriempfindlichkeit. Fur eine solche Annahnie spre- chen die schlechte Korrelation der Struktur-Wirkungsbeziehungen zwischen den bei- den hier untersuchten Pilzarten sowie der unterschiedliche Anstieg der Regressions- geraden gegen lg P. Ein weiteres Argument liefert die Untersuchung von GEORGO- POULOS et al. (1972). Die Autoren erhielten eine carboxinresistente Mutante von Usfi- lago naaydis, deren Succ.-DH deutlich veranderte Eigenschaften, wie z. B. extreme Labilitat, aufwies ; die Mutation betraf folglich eine Komponente des Succ.-DH-Kom- plexes, offenbar den Carboxinrezeptor.
Herrn Professor RAPOPORT sind wir fur wertvolle Hinweise und Anregungen zu grol3em Dank
Fraulein S. K ~ H L E R danken wir fur die sorgfiiltigen Untersuchungen an den submitochondrialen verpflichtet.
Priiparationen von Trametes und Trdchoderma.
L i t e r a t u r
BERNATH, P. and SINGER, T. P., 1962. Succinic dehydrogenase. In: Methods in Enzymology, 6 ,
CRANE, F. L., GLENN, I. L. and GREEN, D. E., 1956. Studies on the electron transfer system. IV.
DOVE, R., 1974. Zur Moglichkeit der Anwendung des ETP-Testes fur pharmakologische Zwecke.
EDGINGTON. L. V. and BARRON. G. L.. 1967. Funnitoxic sDectrum of oxathiin comDound. Phvto-
597.
The electron transfer particle. Biochim. biophysica Acta, 22,475.
Diplomarbeit, Martin-Luther-Universitlt Halle- Wittenberg, 27. 9.
pathology, 57, 1256. -
FRANHE. R. und OEHME, P.. 1973. Aktuelle Probleme der Wirkstofforschunn. 1. Ermittlung uuan- titativer Struktur- Wirkungs-Beziehungen bei Biowirkstoffen : TheoretiscGe Grundlagen,-D;rch- fiihrung, Voraussetzungen und gegenwlrtiger Stand. Pharmazie, 28,489.
FRANRE, R. und DOVE, S., 1975. Unveroffentlichte Ergebnisse. GEORGOPOULOS, S. G., ALEXANDRI, E. and CHRYSAYI, M., 1972. Genetic evidence for the action of
oxathiin and thiazole derivatives on the succinic dehydrogenase system of Ustilago maydis mito- chondria. J. Bacteriol., 110, 809.
GRIMMEL, H. W., GUENTHER, A. and MORGAN, J. F., 1946. Phosphazo compounds and their use in preparing amides. J. Amer. chem. SOC., 68, 539.
TEN HAHEN. P. and BAYR. H.. 1970. BRDPat. 1.914.954 ICIAOln. 9/10) vom 23. 9. 1969. Chem. , I I I ,
Abstr., 7'& 31 613t. TEN HAKEN. P. and DUNN. c. L.. 1971. Structure-activitv re1ationshiDs in a mouD of carboxanilides
systemically active against broad bean rust ( UromGes fubae) a i d w h e 2 rust (Puccinia recon- dita). Proc. 6th Br. Insectic. Fungic. Conf.
KULKA, A., THIARA, D. S. and HARRISON, W. A., 1968. U.S. Pat. 3.339.214 (C1260-327) vom 27.8. LEO, A., HANSCH, C. and ELKINS, D., 1971. Partition coefficients and their uses. Chem. Rev.,
LOWRY, 0. H., ROSEBROUGH, N. J., FARR, A. L. and RANDALL, R. J., 1951. Protein measurement
LYR, H., LUTHARDT, W. und RITTER, G., 1971. Wirkungsweise von Oxathiin-Derivaten auf die
LYR, H. RITTEH, G. und CaSPERSON, G., 1972. Wirkungsweise des systemischen Fungicides Car-
LYR, H., RITTER, G. and BANASIAK, L., 1974. Detoxicationof carb0xin.Z. Allg. Mikrobiol., 14,313. LYR, H., SCHEWE, T., MULLER, W. und ZANKE, D., 1975. Zum Problem der Selektivitiit sowie
der Struktur-Rezeptorbeziehungen von Carboxin und seinen Analogen. Internationales Sympo- sium ,,Systemfungizide" Reinhardsbrunn, Mai 1974, Akademie-Verlag Berlin S. 153.
LYR, H. and SCHEWE. T., 1975. On the mechanism of the cyanide-insensitive alternative pathway of respiration in fungi and higher plants and the nature of the alternative terminal oxidase. Acta biol. med. german., 34, 1631.
71, 525.
with the Folin phenol reagent. J. biol. Chemistry, 193,265.
Physiologie sensitiver und insensitiver Hefearten. Z. Allg. Mikrobiol., 11, 373.
hoxin. Z. Allg. Mikrobiol., 12, 271.

372 W. MULLER, T. SCHEWE, H. LYR und D. ZANKE
MATERE, D. E., 1971 a. Mode of action of oxathiin systemic fungicides. 111. Effects on mitochon- drial activities. Pestic. Biochem. Physiol., 1, 216.
MATHRE, D. E., 1971 b. Mode of action of oxathiin systemic fungicides. Structure-activity rela- tionships. Agric. and Food Chem., 19,872.
MAY, R. und JACOB, H.-E., 1970. Vergleich zwiwchen Atmungsaktivitiit und morphologischem Zustand isolierter Basidiomyceten-Mitochondrien. Z. Allg. Mikrobiol., 10, 275.
MULLER, W., SCHEWE, T. und RAPOPORT, S., 1977. Elektronentransportpartikeln aus Rinderherz als Testsystem auf Hemmstoffe der Atmungekette. Acta biol. med. german., 36, Heft 5-6.
OHNISHI, T., WINTER, D. B., LIM, J. and KING, T. E., 1974. Studies on a Hipip type iron-sulfur center in the succinate dehydrogenase segment of the respiratory chain. Biochem. biophysic. Res. Commun., 61, 1017.
~ E R S O N , E., GIELLA, M. and TISBLER, M., 1948. Synthesis of DL-methionine. J. Amer. chem. SOC., 70, 1450.
RAPSON, W. S. and SHUTTLEWORTH, R. G., 1941. Benzcyclooctatetraenes, I. J. chem. SOC., 1941, 487.
REDFEARN, E. R., WHITTAKER, P. A. and BURQOS, J., 1965. The Interaction of Electron Carriers the Mitochondria1 NADH, and Succinate Oxidase Systems. Symposium on Oxidases and Re- lated Redox Systems (KINQ, T. E., MASON, H. S. and MORRISON, M., Editors). Wiley New York, 11, p. 942.
SANADI, D. R., PHARO, R. L. and SORDAHL, L. A,, 1967. NADH-CoQ reductase-assay and puri- fication. In: Methods in Enzymology, 10, 297.
SCHEWE, T., RAPOPORT, S., BOFIME, G. und KUNZ, W., 1973. Zum Angriffspunkt des Systemfungi- zids Carboxin in der Atmungskette. Acta biol. med. german., 31, 73.
SCHEWE, T., HIEBSCH, CH., GARCIA PARRA, M. und RAPOPORT, S., 1974. Zur Wirkung von At- mungshemmstoffen auf die Atmungsenzyme der Mitochondrien aus Blumenkohl (Brassica olera- cea L.) . Acta biol. med. german., 32,419.
VON SCHMELING, B., KULKA, M. TEIARA, D. S. and HARRISON, W. A., 1966. U.S. Pat. 3.249.499 (Cl 167-33) vom 3.5. 1966; Chem. Abstr. 66,7190g.
SINQER, T. P., BEINERT, H., ACKRELL, B. A. C. and KEARNEY, E. B., 1975. Electron carriers in the succinate dehydrogenase region of the respiratory chain, their reactions and relationships. Abstracts of the “Tenth Meeting of the Federation of European Biochemical Societies”, Paris, 20-25 July 1975; published by the SocietB de Chimie Biologique, Paris 1975, Abstract- No. 95.
TAPPEL, A. L., 1960. Inhibition of electron transport by antimycin A, alkyl hydroxy naphtho- quinones and metal coordination compounds. Biochem. Pharmacol., 3,289.
TISDALE, H. D., 1967. Preparation and properties of succinic cytochrome c reductase (complex 11-111). In: Methods in Enzymology, 10,213.
TUCKER, A. N. and LILLICH, T. T., 1974. Effect of the systemic fungicide carboxin on electron trans- port function in membranes of Micrococcus denitrificans. Antimicrob. Agents and Chemotherapy, 6,572.
ULRICH, T. J. and MATISRE, D. E., 1971. Mode of action of oxathiin systemic fungicides. V. Effects on electron transport system of Ustilago maydis and Saccharomyces cerevisiae. J. Bacteriol., 110, 628.
United States Rubber Company, 1967. Neth. Appl. 6.605.525. (Cl C 07d) vom 27. 10.1966; Chem. Abstr., 66,95 055w.
WHITE, G. A., 1971. A potent effect of 1,4-oxathiin systemic fungicides on succinate oxidation by a particulate preparation from Ustilago maydis. Biochem. biophysic. Res. Commun., 44, 1212.
WHITE, G. A. and THORN, G. D., 1975. Structure-activity relationships of carboxamide fungicides and the succinic dehydrogenase complex of Cryptococcus laurentii and Ustilago maydis. Pestic. Biochem. Physiol., 6,380.
W m A M s o N , R. L. and METCALF, R. L., 1973. Salicylanilides: A new groupof active uncouplers of oxidative phosphorylation. Science, 168, 1694.
ZAUGQ, H. E. and RAPALA, R. T., 1947. o-Toluic acid. Org. Synth., 27,84.
Anschrift: Dr. T. SCHEWE Institut f i i r Physiologische und Biologische Chemie der Humboldt-Universitat DDR 104 Berlin, Hessische StraDe 3/4





















![Synthese neuer Benzo[c]phenanthridin-Derivate und deren ... · Synthese neuer Benzo[c]phenanthridin-Derivate . und deren Stickstoff-Analoga als potentielle Zytostatika . Dissertation](https://img.pdfslide.org/doc/110x75/5e20f36f5b31f87be65362b3/synthese-neuer-benzocphenanthridin-derivate-und-deren-synthese-neuer-benzocphenanthridin-derivate.jpg)







