Embed Size (px)
Citation preview

J. Anirn. Physiol. a. Anirn. Nutr. 69 (1993), 79-91 0 1993 Verlag Paul Parey, Hamburg und Berlin ISSN 0931-2439
Eingang des Ms. 6. 7. 1992
Aus dem Institut fur Ernahrungsphysiologie der Technischen Universitiit Mtinchen in Freising- Weihenstephan
Zum Einflufi von Zinkdepletion auf die Gehalte einzelner Phospholipide in der Erythrozytenmembran und deren Fettsaurezusammensetzung bei zwangsernahrten Ratten
Von K. EDER und M. KIRCHCESSNER
Einleitung
Mehrere Untersuchungen haben gezeigt, daB im Zinkmangel die Hamolyseresistenz von Rattenerythrozytenrnembranen erniedrigt ist (PATERSON und BETTCER 1985; O'DELL et al. 1987; ROTH und KIRCHCESSNER 1991 a, b). Im Zinkmangel war aber auch die Fluiditat der Erythrozytenmembran bei Ratten verandert (JAY et al. 1987). Da die Fluiditat biologischer Membranen in erster Linie durch ihre Lipidzusamrnensetzung bestimmt wird (STUBBS und SMITH 1984; SHINITZKY 1984), wurde auch eine rnogliche veranderte Lipidzusarnrnenset- zung der Erythrozytenrnembran als Ursache der verminderten Hamolyseresistenz unter- sucht UOHANNING und O'DELL 1989; DRISCOLL und BETTCER 1991; BEHRENS und PALLAUF 1992). Allerdings ist die Futteraufnahrne der Tiere bei diesen Versuchen sehr niedrig, was zu einem verminderten Wachstum, zu einem veranderten Lipidstoffwechsel und verander- tem Fettsauremuster in verschiedenen Geweben fuhrt (LEVEILLE 1970; KRAMER et al. 1984; SCHNEEMAN et al. 1986). Aufierdern werden die pair-fed Tiere zu Mahlzeitenfressern, die sich hauptsachlich in der postprandialen Phase befinden, wahrend die Mangeltiere als Nippler die vorgelegte Diat kontinuierlich aufnehmen, was ebenfalls den Lipidstoffwech- sel der Tiere beeinflufit (LEVEILLE 1970).
In der vorliegenden Arbeit sollte der EinfluB von Zinkmangel auf die Gehalte einzelner Phospholipide in der Erythrozytenmembran und deren Fettsaurezusarnmensetzung unter- sucht werden. Urn die oben genannten Probleme herkommlicher Zinkmangelversuche zu urngehen, wurden die Versuchstiere rnit Hilfe der Schlundsondentechnik wie bei S C H ~ L E I N et al. (1992) und KIRCHGESSNER et al. (1992) zwangsernahrt.
In den bisherigen Versuchen, in denen die Lipidzusammensetzung der Erythrozyten- rnernbran irn Zinkrnangel untersucht wurde, karn stets eine Diat zur Anwendung, deren Fettkomponente ausschlieBlich aus Maisol UOHANNING und O'DELL 1989, DRISCOLL und BETTCER 1992) oder Sojaol (BEHRENS und PALLAUF 1992) bestand und daher ein Mehrfaches des Bedarfes a n Linolsaure aufwies. Die Fettsaurezusarnrnensetzung der Diat beeinflufit die Fettsaurezusammensetzung der Erythrozytenmembran (PUTNAM et al. 1982; CARTWRICHT et al. 1985; BJERVE et al. 1987; GLATZ et al. 1989) und diirfte daher bedeutend fur die vorlie- gende Fragestellung sein. Deshalb wurde in der vorliegenden Untersuchung im Gegensatz zu den bisherigen Untersuchungen auch eine Diat eingesetzt, deren Linolsauregehalt bedarfsorientiert war.
Material und Methodik
24 mannliche Sprague-Dawley-Ratten mit einer durchschnittlichen Lebendmasse von 109 f 3 g wurden in 2 Behandlungsgruppen eingeteilt, von denen eine eine Diat mit einern
U.S. Copyright Clearance Center Code Statement: 0931-243919316902-0079 $ 02.5010

80 K. Eder und M. KirchgeJ3ner
Zinkgehalt von 118 k 2 ppm (Kontrollgruppe) und eine eine Diat mit einem Zinkgehalt von 2,4 f 0,6 ppm (Depletionsgruppe) erhielt. Alle Tiere wurden mit Hilfe der Schlund- sondentechnik zwangsernahrt (SCHULEIN eta]. 1992), wobei eine halbsynthetische Diat auf Caseinbasis eingesetzt wurde, deren Zusammensetzung in Tabelle 1 dargestellt ist. Die Fett- komponenten der Diat wurden im Hinblick auf eine bedarfsorientierte Linolsaurever- sorgung der Tiere ausgewahlt. Die Tiere wurden vierrnal am Tag (8.O0, 13.", 18.", 23."' Uhr) gefuttert und erhielten in den ersten vier Versuchstagen 14,5 g Futter. Da die Tiere bei einsetzendem Z i n h a n g e l keine so grogen Futtermengen mehr venver- ten konnen, wurde den Tieren ab dem 5. Tag 11,6 g Futter verabreicht.
Die Tiere wurden in einern vollklimati- siertem Raum bei einer Temperatur von 23°C und 60% relativer Luftfeuchte gehal- ten einzeln in Makrolonkafigen gehalten, deren Boden mit Plexiglaslaufrosten aus- gestattet war.
Nachdem bereits am Ende des 6. Tages 4 Tiere der Depletionsgruppe ausfielen, wurden die Tiere zu Beginn des 7. Tages im AnschluB an eine Ethernarkose dekapitiert. Das Blut wurde in heparinisierten Rohr- chen aufgefangen.
Die Bestirnmung der Gehalte einzelner Phospholipide und ihrer Fettsaurezusam- mensetzung erfolgte wie von EDER et al. (1992) beschrieben. Dazu wurde das Voll- blut zentrifugiert, die Erythrozyten rnit physiologischer NaCl gewaschen und ha- molysiert. Die Mernbranfragmente wurden mehrfach mit Tris-Puffer gewaschen und die Lipide der Erythrozytenmembranen mit Isopropanol extrahiert. Die Phospholipide des Extrakts wurden mittels HPLC (Fa. Merck-Hitachi, Darmstadt) getrennt, rnit einem Fraktionensammler (Fa. Gilson, Modell 201, Villiers-le-Be], Frankreich)
Tabelle I. Zusammensetzung der Versuchsdiat (Table 1. Composition of the experimental diet)
Casein Starke Saccharose Fett'
davon: Kokosfett Distelol
Cellulose Vitaminmischung' Mineralstoffmischung' DL-Methionin
200 328 300
80 68,6 11,4
30 20 40
2
Fettsaurezusammensetzung (in GewichtsVo, 8 Bestimmungen) Capronsaure (6 : 0) 0,47 f 0,lO; Caprylsaure (8:0)7,91 f0,93;Caprinsaure( l0:0)5,96* 0,79; Laurinsaure (12 :0) 41,22 f 2,83; Myri- stinsaure (14 :0) 15,24 f 0,96; Palmitinsaure (16:O) 8,30*0,88; Stearinsaure (18 0) 3,54 f 0.56; Olsaure (18: 1) 7,18 f 0.94; Linolsaure (18:2) 10,18f 1,14. ' Vitamine pro kg Diat: 5000 I.E. Retinol; 300 I.E. Cholecalciferol; 150 mg a-Tocophe- rolacetat; 5 mg Menadion-Natriumbisulfit; 5 mg Thiaminiumdichlorid; 10 mg kbofla- vin; 6 mg Pyridoxinhydrochlorid; 50 mg Ca- D-Pantothenat; 20 mg Nicotinsaure; 1000 mg Cholinchlorid; 0,2 mg Folsaure; 0,025 mg Vitamin B12; Zucker ad 20 g. ' Mineralstoffe pro kg Diat: NazHP04 . 2 H 2 0
6H'O 3,40 g; CaCO3 13,6 g; F e S 0 4 . 7 H 2 0 248,8 mg; C u S 0 4 . 7 H2O 47,2 mg; MnS04 . 5 H 2 0 123,l mg; KJ 9,O mg; NiS04 6 H 2 0 4,48 mg; N a M o 0 4 . 2 HzO 0.50 mg; SnCl2 2 HtO 0,57 mg; Na2Se03 . 5 H 2 0 0,67 mg; CrCI, 6 H 2 0 0,51 mg; NH4VO3 0,23 mg; NaSiO, 5 H 2 0 1,51 rng.
10,74 g; KH2P04 8,20 g; KCI 6.00 g; MgCI,
gesammelt und mit Natriurnmethoxid (Merck, Darmstadt) rnethyliert. Die Fettsaure- methylester wurden gaschromatographisch mit Heptadecansauremethylester als internern Standard quantitativ bestirnmt. Dazu wurde ein Gaschromatograph der Fa. Siemens (Modell Sichromat 2, Karlsruhe) mit temperaturprogrammierbarem Verdampfer (mV) und Flarnmenionisationsdetektor (FID) verwendet. Die Trennung der Fettsauremethylester erfolgte auf einer 50 m langen CP-Si1 88 Trennsaule der Fa. Chrompack (Middleburg, Nie- derlande). Die Aufbringung der Probe erfolgte iiber Split-Injektion, Split-Verhaltnis 1 :2. Die Mengen der einzelnen Phospholipide in den Erythrozytenmernbranen wurden aus der Summe ihrer Fettsauren berechnet.
Zur Bestimmung der Fettsaurezusammensetzung des Plasmas wurden 0,2 ml Plasma mit 1,5 rnl Chloroform-Methanol-Gemisch (2 : 1, v/v) versetzt. Dieser Ansatz wurde nach einer Stunde mittels eines HPLC-Einwegfilters filtriert, in ein verschlieflbares Methylierungsge-

Der ErnzuJ3 oon Zn-Depletion auf Erythrorytenmembranen 81
fag iiberfiihrt, das Losungsmittel unter Vakuum abgedampft und die Lipide rnit 1 ml Bor- tritluorid-Methanol-Reagens (purissimum, Fa. Fluka, Schweiz) versetzt. Nach 4stiindigem Erhitzen bei 90°C wurden die Fettsauremethylester durch Zugabe von 0,s ml Wasser und 1 ml Hexan extrahiert. Die Extraktion wurde nach Abheben der Hexanphase wiederholt. Die Fettsauremethylester der gesammelten Hexanphasen wurden gaschromatographisch bestimmt. Das Split-Verhaltnis bei der Injektion betrug 1 : 10.
Zur Bestimmung der Fettsaurezusammensetzung der Diit wurde 5 g fertiger Futterbrei mit 100 ml Chloroform-Methanol-Gemisch (2: 1, v/v) versetzt. Nach 5 h wurde ein Ali- quot dieses Ansatzes entnommen, die enthaltenen Lipide analog denen im Plasma methy- liert und die Fettsauremethylester gaschromatographisch bestimmt. Das Split-Verhaltnis bei der Injektion betrug 1 : 50.
Zur Bestimmung des Plasmazinkgehaltes wurden 0,2 ml Plasma mit bidest. Wasser im Verhaltnis 1 : 5 verdiinnt und der Zinkgehalt dieses Ansatzes direkt an einem Flammen- Atomabsorption-Spektralphotometer (Fa. Perkin-Elmer, Modell 303, Uberlingen) be- stimmt. Die Messung der Aktivitat der alkalischen Phosphatase erfolgte mit Hilfe einer Testkombination der Fa. Boehringer, Mannheim an einem Autoanalyzer (Fa. Hitachi, Modell 704). Die ermittelten Werte beziehen sich auf eine Inkubationstemperatur von 25°C.
Die statistische Auswertung der Versuchsergebnisse erfolgte mit Hilfe der einfaktoriel- len Varianzanalyse. Mittelwertsvergleiche wurden mit dem t-Test durchgefuhrt; unter- schiedliche Mittelwerte sind durch unterschiedliche Hochbuchstaben gekennzeichnet.
Gewicht (g) Tag 1 Tag 4 Tag 5 Tag 6 Tag 7
Zinkgehalt (pg/ml) , Alkalische Phosphatase (U/I)
Ergebnisse
Die Gewichtsentwicklung der Ratten sowie die Zinkgehalte und die Aktivitat der alkali- schen Phosphatase im Plasma sind in Tabelle 2 zusammengefagt. In den ersten vier Ver- suchstagen bestanden keine Unterschiede in der Gewichtsentwicklung zwischen Kontroll- und Depletionstieren. Am Tag 4 trat bei den Depletionstieren ein Riickgang der Gewichts- zunahme und ab Tag 5 ein Wachstumsstop auf. Der Zinkgehalt im Plasma war bei den Depletionstieren nach 7 Tagen um 71%, die Aktivitat der alkalischen Phosphatase um 66% gegeniiber den Kontrolltieren erniedrigt.
Die Gehalte einzelner Phospholipide in den Erythrozytenmembranen von Kontroll- und Depletionstieren sind in Tabelle 3 dargestellt. Die Erythrozytenmembranen der Depletionstiere wiesen hohere Gehalte an Phosphatidylcholin (PC), Phosphatidylethano-
Tab& 2. Gewichtsentwicklung sowie Zinkgehalt und Aktivitat der alkalischen Phosphatase im Plasma von Kontrolltieren und Depletionstieren am 7. Tag
(Table 2 Weight gain, zinc concentrations and activity of alkaline phosphatase in plasma of control and depletion animals of day 7)
Parameter
108,s f 3,2 109,6 f 2 3 123,9 + 2,O 123.8 f 2,9 129,4 f 3,5 126,5 f 4,4 136,4 + 2,9" 125,l f 5,9b 140,7 f 3,5" 124,l f 5Jb
358 +58' 123 +54b 1,38f 0,12" 0,40+ 0,06b

82 K. Eder und M. Kirchgejner
Tubelle 3. Gehalte und prozentuale Anteile einzelner Phospholipide in den Erythrozytenmembranen (pg PhosphoIipid/g gepackte Erythrozyten) von Kontrolltieren und Depletionstieren
(Tub/e 3. Contents and percentage distribution of individual phospholipids in erythrocyte membranes of control and depletion animals (pg phospholipids pe: g of packed erythrocvtes))
~ ~~
Phospholipid Zinkversorgung Kon trol le
(118 ppm)
PC 1160 f 98b (48,50hA) 1279 PE (Diacyl) 307 * 50' (12.7%) 357 PE (Plasmalogen) 351 * 69b (14,b0/o) 447 PS 273 f 85 (11,2%) 342 SM 311 f 56'(12,9%) 382 Summe 2402 f260' 2807
f 144' (45,6°/oB) f 52' (12,8O/o) f 79'(15,90/0) f 64 (12.3%) f 73'(13,6%) * 318"
PC/SM 3,80f 0,48 3,41 f 0.41
lamin (PE) in Diacyl-Form und in Plasmalogen-Form und Sphingomyelin (SM) auf. Die Summe dieser Phospholipide war bei den Depletionstieren ebenfalls erhoht. Die prozen- tualen Anteile einzelner Phospholipide waren zwischen Kontroll- und Depletionstieren ahnlich, jedoch warder Anteil von PC bei den Depletionstieren niedriger als bei den Kon- trolltieren. Das Mengenverhaltnis zwischen PC und SM, einem Parameter der die Fluiditat der Erythrozytenmernbran beeinflufit, war bei den Depletionstieren um 11 O/o erniedrigt, jedoch nicht signifikant.
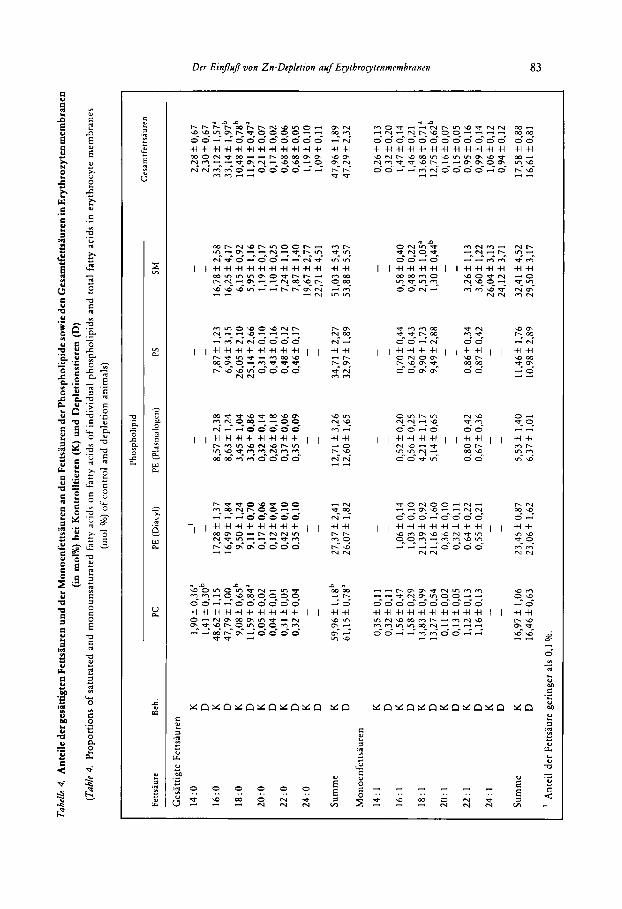
Die Anteile gesattigter und einfach ungesattigter Fettsauren an den gesamten Fettsauren der einzelnen Phospholipide sowie den Gesamtfettsauren der Erythrozytenmernbran sind in Tabelle 4 dargestellt. Beziiglich der Anteile der gesattigten Fettsauren traten Unter- schiede zwischen Kontroll- uiid Depletionstieren lediglich beim PC und bei den Gesamt- fettsauren der Erythrozytenmembran auf. Beim PC war bei der Depletionsgruppe der Anteil der Myristinsaure (14 : 0) erniedrigt, der Anteil der Stearinsaure (18 : 0) erhoht und die Summe der Anteile einzelner gesattigter Fettsauren ebenfalls erhoht. Bei den Gesamt- fettsauren der Erythrozytenmembran war bei der Depletionsgruppe der Anteil der Stearin- saure erhoht und der der Palmitinsaure vermindert. Die Summe der gesattigten Fettsauren war bei den Gesamtfettsauren jedoch nicht unterschiedlich. Bei den Phospholipiden PE (in Diacyl-Form und in Plasmalogen-Form), PS und SM traten bei den Anteilen der gesat- tigten Fettsauren keinerlei Unterschiede zwischen Kontroll- und Depletionstieren auf. Bezuglich der Monoenfettsauren traten Unterschiede lediglich beim SM und bei den Gesamtfettsauren der Erythrozytenmembran auf. In beiden Fraktionen war bei den Deple- tionstieren der Anteil der Olsaure (18 : 1) erniedrigt.
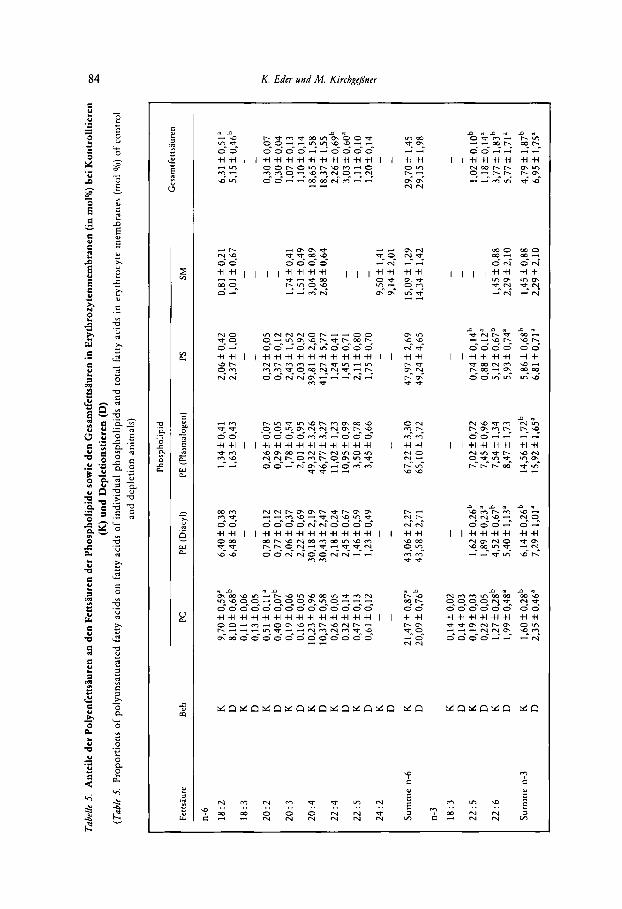
Die Anteile der mehrfach ungesattigten Fettsauren an den Gesamtfettsauren der Ery- throzytenmembran und den Fettsauren der einzelnen Phospholipide sind in Tabelle 5 dar- gestellt.
Bei den n-6 Fettsauren traten Unterschiede zwischen Kontroll- und Depletionsgruppe lediglich beim PC und bei den Gesamtfettsauren auf. Beim PC waren in der Depletions- gruppe die Anteile der Linolsaure (18 : 2), der Eicosadiensaure (20 : 2) und auch die Summe der Anteile einzelner n-6 Fettsauren vermindert. Bei den Gesarntfettsauren war der Anteil der Linolsaure bei der Depletionsgruppe vermindert, der Anteil der Docosatetraensaure (22 :.4) dagegen erhoht, wahrend die Summe der Anteile einzelner n-6 Fettsauren von der Zinkversorgung nicht beeinfluRt wurde.
Die starksten Unterschiede zwischen Kontroll- und Depletionstieren traten bei den Anteilen der n-3 Fettsauren auf. Der Anteil der Docosahexaensaure (22: 6) war bei den Phospholipiden PC, PE (Diacyl), PS und den Gesamtfettsauren erhoht, der Anteil der n-3 Form von Docosapentaensaure (22 : 5) war bei den Phospholipiden PE (Diacyl) und PS
Tubc
lle 4
. A
ntci
lc d
er g
csit
tigt
cn F
etts
aure
n un
d dc
r Mon
ocnf
etts
aure
n an
den
Fet
tsau
ren
der P
hosp
holip
idc s
owie
den
Ges
arnt
fctts
aurc
n in
Ery
thro
zytc
nmem
bran
en
(in
rnol
%) b
ci K
ontr
ollti
ercn
(K
) und
Dcp
lctio
nstic
rcn
(D)
(Tab
le 4.
Pr
opor
tions
of
satu
rate
d an
d m
onou
nsat
urat
ed fa
tty a
cids
on
fatty
aci
ds o
f in
divi
dual
pho
spho
lipi
ds a
nd to
tal
fatty
aci
ds i
n er
ythr
ocyt
e m
embr
anes
(m
ol O
/o) of
con
trol
and
dep
leti
on a
nim
als)
Phos
phol
ipid
G
erar
ntfe
ttra
uren
Fe
ttra
ure
Beh
. PC
PE (D
iacy
l)
PE (
Plas
mal
ogen
) PS
SM
Ges
attig
te F
etts
aure
n
14:O
K
1,90
f 0.
36'
D
1,41
f 0,
30b
16:O
K
48.6
2 f 1
.15
18:O
K
9,08
f 0,
65b
D
I1,5
9 f 0
,84'
20
:o
K 0.
05 f 0.
02
D
0.04
f 0.
01
22:o
K
0,3
I f 0,
05
D
0,32
f 0.
04
24:O
K D
-
Sum
me
K 59
,96f
1,1
8b
D
61,1
5 f 0,
78'
Mon
oenf
etts
aure
n
14:l
K
0,35
f 0
,ll
D
0,32
f 0
,ll
16:l
K
1,56
f 0,
47
D
1,58
f 0,
29
18:l
K
13,8
3 f 0,
99
D
13,2
7 f 0,
54
D
0.13
f 0,
05
22:l
K
1,12
f0,1
3 D
1,
16f0
,13
24:l
K D
Su
mm
e K
16,9
7f 1
.06
D
16,4
6 f 0,
63
' Ant
eil
der
Fetts
aure
ger
inge
r al
s 0.
1 %
.
D
47,7
9 f 1,
oo
-
2O:l
K
O,l
lf0,
02
-
-
I - -
17,2
8f 1
,37
16,4
9f 1
.84
9,50
* 1.
24
9.11
f 0.
70
0.17
f 0,
06
0.12
f 0.
04
0,42
f 0,
lO
0,35
f 0,
lO
-
-
27,3
7 f 2,
41
26.0
7 f 1
.82
-
-
1,06
f 0,
14
1,03
f 0.
10
2 1.3
9 f 0,
92
21,1
6f
1,60
0,
36f
0,lO
0.
32 f 0
,ll
0,64
f 0,
22
0,55
f 0,
21
-
-
23.4
5 f 0,
87
23.0
6 f 1
,62
-
-
8,57
f 2.
38
8.63
f 1.
24
3.45
f 1.
04
3.36
f 0
.86
0,32
f0,1
4 0,
26 k 0
, I8
0,37
f 0,
06
0,35
f 0
.09
-
-
12,7
1 f 3.
26
12,6
0f 1
,65
-
-
0,52
f 0,
20
0,56
f 0
,25
4,21
f 1
,17
5,14
f 0
,65
-
-
0,80
f 0
,42
0,67
f 0
,36
-
-
5,53
f 1
.40
6,37
f 1.
01
-
-
7,87
f 1
.23
6,94
f 3.
15
26.0
5 f 2.
10
25.1
4 f 2.
66
0,31
f 0.
10
0.43
f 0.
16
0,48
f 0.
12
0,46
f 0.
17
-
-
34,7
1 f 2
,27
32,9
7 f 1
,89
-
-
0,70
f 0,
44
0.62
f 0
.43
9,90
f 1
,73
9,49
f 2,
88
-
-
0,86
f 0,
34
0.87
f 0.
42
-
-
11,4
6f 1
.76
10,9
8 f 2.
89
- -
16
,~
* 2,
5a
16,2
5 f 4.
17
6, I5
f 0
,92
5,95
f 1
,16
1,19
* 0,
17
I,lO
f0,2
5 7,
24 f 1
,lO
7.87
f 1
,40
19,6
7 f 2
,77
22,7
1 f 4
3 1
5 LO
3 f 5
,43
53,8
8 f 5.
57
- -
0,58
f 0
,40
0.48
f 0.
22
2,53
f 1
,0Sa
1,
30f
0,44
b - -
3,26
f 1,
13
3,60
f 1,
22
26,0
4 f 3.
13
24,1
2 f 3
,71
32,4
1 f 4,
52
29.5
0f 3
.17
2,28
f 0
,67
2,30
f 0
,67
33,1
2f
1,57
' 33
,14f
1,
97b
10,4
8 f 0,
78b
11.9
1 f
0.47
' 0.
21 f 0.
07
0,17
f 0,
02
0.68
f 0,
06
0,68
f 0.
05
1.19
f 0,
lO
1.09
f 0.
1 1
47.9
6 f 1,
89
47.2
9 f 2.
32
0,26
f 0.
13
0,32
f 0.
20
1,47
f 0,
14
1,46
f 0,
21
13,6
8 f 0,
71'
12,7
5 f 0,
6Zb
0,16
f 0,
07
0.15
f 0,
05
0.95
f 0.
16
0.99
f 0,
14
1.06
f 0,
12
0,94
f 0.
12
17,5
8 f 0.
88
16,6
1 f 0.
81
Tube
lle 5
. A
ntei
le d
cr P
olye
nfet
tsau
ren
an d
en F
etts
aure
n de
r Ph
osph
olip
ide
sow
ie d
en G
esam
tfet
tsau
ren
in E
ryth
rozy
tenm
embr
anen
(in
mol
%) b
ci K
ontr
ollti
eren
(K
) und
Dep
letio
nstie
ren
(D)
(Tab
le 5
. Pr
opor
tions
of
poly
unsa
tura
ted
fatty
aci
ds o
n fa
tty a
cids
of
indi
vidu
al p
hosp
holi
pids
and
tota
l fa
tty a
cids
in
eryt
hroc
yte
mem
bran
es (
mol
%)
of c
ontr
ol
and
depl
etio
n an
imal
s)
Phos
phol
ipid
G
esam
tfet
tsau
ren
Fett
saur
e B
eh.
PC
PE (D
iacy
l)
PE (P
lasm
alog
en)
PS
SM
n-6
18:2
18:3
20:2
20:3
20:4
22:4
22 : 5
24:2
Sum
me
n-6
n-3
18:3
22:s
22:6
Sum
me
n-3
K D K D
K D
K
D
K
D
K D
K
D
K D K
D
K
D
K
D
K D
K
D
9.70
f 0,
59’
8.10
f 0,
68b
0.1
1 f 0
,06
0,13
f 0,
05
0.51
f 0
,ll”
0,
40 f 0,
07’
0.19
f 0,
06
0,16
f 0,
05
10.2
3 f 0,
96
10.3
7 f 0
,58
0,26
f 0,
05
0,32
f 0
,14
0,47
f 0,
13
0,61
f 0,
12
-
21,4
7 f 0,
87’
20,0
9 f 0,
76‘
0.14
f 0,
02
0.14
f 0,
03
0.19
f 0.
03
Oi2
2 f O
iO5
12
7 f 0,
28b
1.99
f 0,
48”
1.60
f 0
,28b
2,
35 f 0,
46’
6,40
f 0
,38
6.48
f 0
,43
- -
0,78
f 0
,12
0,77
f 0
,12
2.06
f 0
,37
2.22
f 0.
69
30,1
8*2,
19
30,4
3 f
2.47
2,
18 f 0
,24
2,45
f 0
,67
1,46
f 0,
59
1.23
f 0
,49
-
43.0
6 f
2,27
43
33 f 2,
71
-
1.61
f 0,
26b
1,89
f 0
,23”
4,
52 f 0.
67’
5,40
f 1,
13”
6,14
f 0,
26b
7,29
f l,O
la
1,34
f 0
.41
1,63
f 0
.43
-
-
0,26
f 0,
’07
0,29
f 0,
05
1,78
f 0,
54
2,O
l f 0
.95
49,3
2 f 3.
26
46,7
7 f 3
.27
11,0
2 f 1
,23
10,9
5 f 0,
99
3,50
f 0,
78
3,45
f 0,
66
-
67,2
2 f 3,
30
65,lO
f 3
.72
-
7,02
f 0
,72
7,45
f 0.
96
8,47
f 1
.73
14,5
6 f
1,7Z
b 15
.92 f 1
.65’
7,54
f 1
,34
2,06
f 0
,42
2.37
f 1
.00
-
-
0,32
f 0
,05
0,37
f 0
,12
2,43
f 1.
52
2,03
f 0,
92
39.8
1 f 2.
60
41,2
7f
5,77
1.
24 f 0,
41
1,45
f 0
,71
2,11
f 0,
80
1,75
f 0,
70
-
47,9
7 f 2,
69
49,2
4 f 4,
65
-
0,74
f 0.
14‘
0,88
f 0
,lL
” 5,
12 f 0
,67b
5.
93 f 0,
74’
5,86
f 0.
68‘
6.81
f0,
71”
0,8 1
f 0,
21
1,O
l It 0
,67
-
-
-
-
1,74
f 0
,4 1
1,51
f 0,
49
3.04
f 0
,89
2.68
f 0.
64
-
-
-
-
9,50
f 1
,41
9,14
f 2.
01
15,0
9 f 1
,29
14,3
4 f 1
,42
-
-
-
-
1,45
f 0
,88
2,29
f 2,
lO
1,45
f 0,
88
2.29
f 2,
lO
6,31
5.
15 f
f 0,
51”
0,46
b
-
-
0,30
f 0
,07
0.30
f 0,
04
1,07
f 0,
13
1,lO
f 0,1
4 18
,65 f
1,5
8 18
.37 f
1,55
2,
26 f 0,
69‘
3.03
f 0
,60d
1.
11 f 0
,lO
1,20
f 0.
14
29,7
0 f 1
,45
29.1
5 f 1
,98
- -
1.02
f 0,
lob
1,18
*0,1
4;
3,77
f 1
,83
5,77
f 1,
71a
4,79
f 1
,87‘
6,
95 f 1
,75‘
Drr Einflujr’ von Zn-Depletion auf Eythrocytenmrmbranm 85
sowie bei den Gesamtfettsiuren erhoht. Die n-3 Form der Linolensaure (18 : 3) war ledig- lich beim PC nachzuweisen und wurde durch Zinkdepletion nicht beeinflu&. Die Surnrne der n-3 Fettsauren war bei den Depletionstieren bei allen Phospholipiden (mit Ausnahrne des SM, wo n-3 Fettsauren quantitativ nur eine untergeordnete Rolle spielen) und den Gesamtfettsauren erhoht.
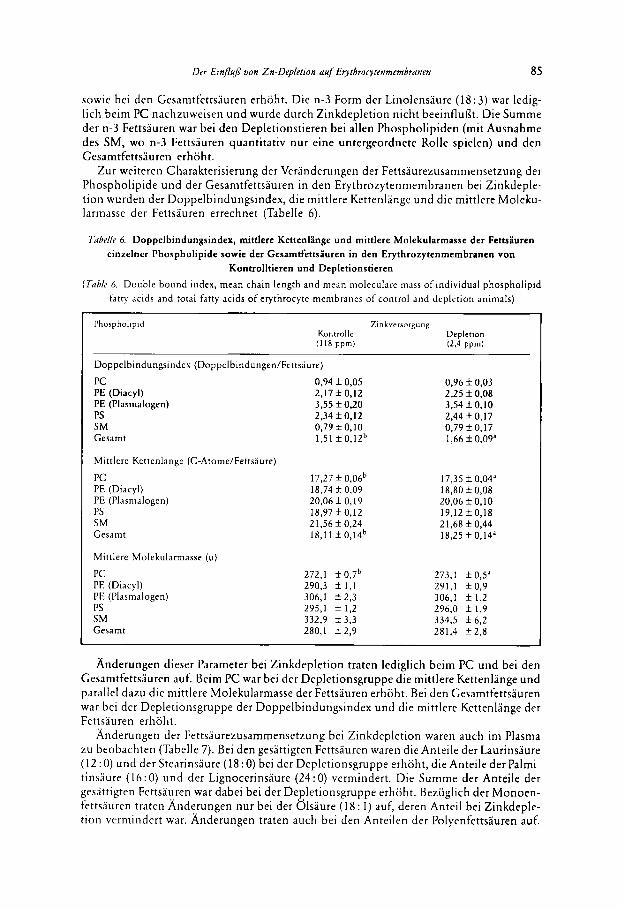
Zur weiteren Charakterisierung der Veranderungen der Fettsaurezusammensetzung der Phospholipide und der Gesamtfettsauren in den Erythrozytenrnembranen bei Zinkdeple- tion wurden der Doppelbindungsindex, die rnittlere Kettenlange und die mittlere Moleku- larmasse der Fettsauren errechnet (Tabelle 6).
Tubdr 6. Doppelbindungsindex, mittlere Kettenlange und mittlere Molekularmasse der Fettsauren einzelner Phospholipide sowie der Gesamtfettsauren in den Erythrozytenmembranen von
Kontrolltieren und Deplctionstieren (Tub/r 6. Double b o u n d index, mean chain length a n d mean moleculare mass of individual phospholipid
fatty acids and total fatty acids of erythrocyte membranes of control a n d depletion animals)
Phospholipid Zinkversorgung Kontrolle Depletion (118 ppm) (2.4 ppm)
Doppelbindungsindex (Doppelbindungen/Fettsaure)
P C PE (Diacvl)
SM Gesamt
Mittlere Kettenlinge (C-Atome/Fettsaure)
PC PE (Diacyl) PE (Plasmalogen) PS S M Gesamt
Mittlere Molekularmasse (u)
PC PE (Diacyl) PE (Plasmalogen) PS SM Gesamt
0.94 f 0.05 2.17 f 0.12 3.55 f 0.20 2.34 f 0.12 0.79 f 0.10 1.51 f 0 , 1 2 h
0.96 f 0.03 2.25 f 0.08 3,54fO,lO 2,44 + 0.17 0.79 f 0, I7 1,66 f 0.09’
17,27 f 0,06’ 17,35 f 0,04’ 18,74 f: 0,09 18.80 f 0,08 20,06 f 0,19 20.06 f 0.10 18,97 f 0,12 19.12 f0,18 21.56 f 0,24 21,68 f 0.44 18,lI f 0,14‘ 18,25 f 0,14d
272,l k 0 , 7 b 273.1 f0,5” 290,3 f 1 , l 291,l f 0 , 9 306,l f 2 , 3 306.1 f I,?. 295,l f 1,2 296.0 f 1,9 332,9 f 3,3 334,5 f 6 , 2 280,l f 2 , 9 281,4 k2.8
Anderungen dieser Parameter bei Zinkdepletion traten lediglich beim PC und bei den Gesamtfettsiuren auf. Beim PC war bei der Depletionsgruppe die mittlere Kettenlange und parallel dazu die mittlere Molekularmasse der Fettsauren erhoht. Bei den Gesamtfettsauren war bei der Depletionsgruppe der Doppelbindungsindex und die rnittlere Kettenlange der Fetttiuren erhoht.
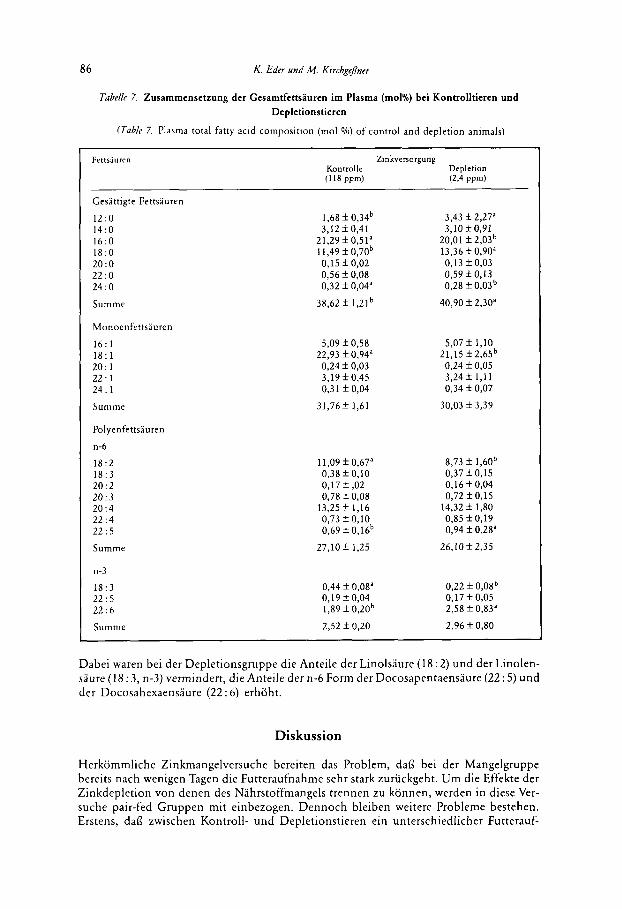
Anderungen der Fettsaurezusamrnensetzung bei Zinkdepletion waren auch irn Plasma zu beobachten (Tabelle 7). Bei den gesattigten Fettsauren waren die Anteile der Laurinsaure (12 : 0) und der Stearinsaure (18 : 0) bei der Depletionsgruppe erhoht, die Anteile der Palmi- tinsaure (16 : 0) und der Lignocerinsaure (24 : 0) vermindert. Die Sumrne der Anteile der gesattigten Fettsauren war dabei bei der Depletionsgruppe erhoht. Bezuglich der Monoen- fettsiuren traten Anderungen nur bei der Olsaure (18: 1) auf, deren Anteil bei Zinkdeple- tion verrnindert war. Anderungen traten auch bei den Anteilen der Polyenfettsauren auf.
86 K. Eder und M . Kirchgejner
Tube/le 7. Zusammensetzung der Gesamtfettsauren im Plasma (mol%) bei Kontrolltieren und Depletionstieren
(Table 7. Plasma total fatty acid composition (rnol O/o) of control and depletion animals)
Fettsduren Zinkversorgung Kontrolle Depletion
( 1 18 ppm) G 4 ppm)
Gesattigte Fettsauren
12 :o 14:O 16:O 18:O 2o:o 22 :o 24:O
Surnrne
Monoenfettsauren
1 6 : l 1 8 : l 2 0 : l 22 : 1 24 : 1
Surnrne
Pol yenfettsauren
n-6
18:2 18 :3 20:2 20:3 20:4 22:4 22:5
Surnme
n-3
18:3 22:5 2 2 : 6
Sumrne
1,68 f 0,34b 3 ,12 f0 ,41
21,29f0,51a 11.49 f 0,70b 0,15 f 0,02 0,56 f 0,08 0.32 f 0,04’
38,62f 1,21b
5,09 f 0,58 22,93 f 0,94“
0,24 f 0,03 3,19 f 0,45 0.31 f 0,04
31,76* 1,61
11,09 f 0,67” 0.38 f 0,lO 0,17 f ,02 0,78 f 0,08
13,25 f 1,16 0,73 f 0 , l O 0,69 f 0,16b
27,lO f 1,25
0,44 f 0,08’ 0,19 f 0,04 1,89 f 0,20b
2,52 f 0,20
3,43 f 2,27’ 3,lO f 0,91
20,Ol f 2,03b 13,36 f 0,90a 0,13 f 0,03 0,59 f 0,13 0,28 f 0,03b
40,90 f 2,30a
5,07 f 1,lO 21,15 f2 ,65b
0,24 f 0,05 3,24 f 1,11 0,34 f 0,07
30.03 f 3,39
8,73 f 1,60b 0.37 f 0,15 0,16*0,04 0,72 f 0 , 1 5
14,32 f 1 3 0 0,85 f 0,19 0,94 f 0,28”
26 ,10f2 ,35
0,22 f 0,08b 0,17 f 0,05 2,58 f 0,83”
2,96 f 0,80
Dabei waren bei der Depletionsgruppe die Anteile der Linolsaure (18 : 2) und der Linolen- saure (18 : 3 , n-3) vermindert, die Anteile der n-6 Form der Docosapentaensaure (22 : 5) und der Docosahexaensaure (22: 6) erhoht.
Diskussion
Herkommliche Zinkmangelversuche bereiten das Problem, daR bei der Mangelgruppe bereits nach wenigen Tagen die Futteraufnahme sehr stark zuruckgeht. U m die Effekte der Zinkdepletion von denen des Nahrstoffmangels trennen zu konnen, werden in diese Ver- suche pair-fed Gruppen mit einbezogen. Dennoch bleiben weitere Probleme bestehen. Erstens, daR zwischen Kontroll- und Depletionstieren ein unterschiedlicher Futterauf-

Der EinzuJ won Zn-Deplelion auf Erythrocylenmembranen 87
nahmerhythmus besteht und zweitens, daB sich die Tiere wegen der geringen Futteraui- nahme in einern unphysiologischen Zustand befinden (SCHULEIN et al. 1992). Um diese Probleme zu beseitigen, wurde in der vorliegenden Arbeit die Methode der Zwangsernah- rung angewendet, die bereits SCHULEIN et a]. (1992) sowie KIRCHGESSNER et a]. (1992) benutzten. Mit Hilfe dieser Methode konnte durch eine sehr hohe Futteraufnahme bereits innerhalb von 7 Versuchstagen ein starker Zinkmangel erzeugt werden, der durch die Gewichtsentwicklung sowie einem stark vermindertem Plasmazinkgehalt und einer stark verminderten Aktivitat der alkalischen Phosphatase im Plasma eindeutig belegt werden konnte.
Die Fragestellung der vorliegenden Untersuchung war, o b im Zinkmangel Anderungen der Gehalte einzelner Phospholipide in den Erythrozytenmembranen undloder ihrer Fett- saurezusamrnensetzung auftreten. Die Versuchsanstellung unterschied sich dabei irn wesentlichen in 4 Punkten von bisherigen Versuchen (JOHANNING und O'DELL 1989; DRIS- COLL und BETTGER 1991, BEHRENS und PALLAUF 1992), die die gleiche oder eine ahnliche Fragestellung zum Ziel hatten. Erstens, wie bereits oben envahnt, war die Futteraufnahme der Tiere wesentlich hoher als die der Tiere bei herkommlichen Zinkmangelversuchen. Zweitens war die Versuchsdauer wesentlich kurzer als bei herkomrnlichen Zinkmangelver- suchen. Drittens konnte rnit Hilfe der Zwangsernahrung eine Synchronisation der Futter- aufnahme erreicht werden. Viertens unterschied sich die Fettkomponente der Diat wesent- lich von den Fettkomponenten bisheriger Versuche. In den Versuchen VOnJOHANNlNG und O'DELL (1989), DRISCOLL und BETTGER (1991) sowie BEHRENS und PALLAUF (1992) wurden pflanzliche Ole mit einem sehr hohen Gehalt an Linolsaure als alleinige Fettkomponenten eingesetzt, so daB der Anteil der Linolsaure an den gesamten Fettsauren der Diat in diesen Untersuchungen iiber 50% betrug. Im vorliegendem Versuch wurde ein Gemisch aus Kokosfett und Distelol in einem Verhaltnis eingesetzt, das einen Linolsaureanteil von etwa 10% ergab. Die Linolsaureaufnahme der Tiere betrug damit in den ersten 4 Tagen taglich etwa 110 rng, in den weiteren Tagen taglich etwa 85 mg und entsprach damit dem Bedarf, der mit 20 bis 100 mg angegeben ist (NATIONAL RESEARCH COUNCIL 1978). Als Folge des geringeren Linolsauregehaltes in der Diat waren die Linolsaureanteile in den einzelnen Phospholipiden unabhangig von der Behandlung in der vorliegenden Arbeit niedriger als bei JOHANNING und O'DELL (1989) und DRISCOLL und BETTCER (1991). so betrug beispiels- weise der Linolsaureanteil im PC in der vorliegenden Arbeit im Mittel der Behandlungen 8,9%, bei DRISCOLL und BETTGER (1991) 19,00/0 und bei JOHANNING und O'DELL (1989) 18,2010. Die verminderten Linolsaureanteile der vorliegenden Untersuchung wurden hauptsachlich durch erhohte Gehalte an gesattigten Fettsauren ausgeglichen. Die Gehalte der ubrigen n-6 Fettsauren (20 : 4,22 : 4 ,22 : 5) in den einzelnen Phospholipiden, die in der vorliegenden Arbeit ermittelt wurden, stirnrnten weitestgehend mit denen der anderen genannten Arbeiten uberein.
Zinkdepletion fuhrte in der vorliegenden Untersuchung zu einer Erhohung der Gehalte der einzelnen Phospholipide in der Erythrozytenmembran. Irn Gegensatz dazu konnten DRISCOLL und BETTCER (1991) SOWieJOHANNlNG und O'DELL (1989) in ihren Untersuchun- gen keine veranderten Gehalte an Phospholipiden in der Erythrozytenmembran feststel- len. Die prozentualen Anteile der einzelnen Phospholipide in der Erythrozytenmembran stimmen groBteils mit den Ergebnissen von DRISCOLL und BETTCER (1991) uberein, jedoch war in der vorliegenden Untersuchung bei den Depletionstieren der PC-Anteil erniedrigt, bei DRISCOLL und BETTGER (1991) der PS-Anteil; bei BEHRENS und PALLAUF (1992) traten keine Unterschiede in den Anteilen einzelner Phospholipide auf. Das Mengenverhaltnis zwischen PC und SM, eines Parameters, der wesentlich die Fluiditat der Erythrozytenmem- bran bestimmt (STUBBS und SMITH 1984), lag in der vorliegenden Untersuchung bei den Kontrolltieren bei 3,8 und bei den Depletionstieren bei 3,4. In Ubereinstimmung mit die- sen Ergebnissen fanden DRISCOLL und BETTGER ein PC/SM-Verhaltnis von 4,O bzw. 3,O (bei pair-fed Kontrolltieren bzw. Depletionstieren). Im Gegensatz dazu zeigte sich bei BEHRENS

88 K. Eder und M . KirchgeJher
und PALLAUF (1992) bei den Depletionstieren ein groReres PUSM-Verhaltnis von 4,6 (gegenuber 4,l bei den pair-fed Kontrolltieren).
Neben den Gehalten war auch die Fettsaurezusammensetzung der einzelnen Phospholi- pide in der Erythrozytenmembran bei den Zinkdepletionstieren verandert. Dabei enthiel- ten die Phospholipide mit Ausnahme des PE in Plasmalogen-Form und des SM bei den Depletionstieren hohere Anteile an langkettigen n-3 Fettsauren. Erhohte Anteile an lang- kettigen n-3 Fettsauren im Zinkmangel fanden auch JOHANNING und O’DELL (1989) und DRISCOLL und BETTCER (1991). Unabhangig von der Behandlung lag das absolute Niveau der Anteile der n-3 Fettsauren in der vorliegenden Arbeit jedoch niedriger als bei den ande- ren Arbeiten. Der Grund dafur liegt im Energiemangel der Tiere bei den herkommlichen Zinkmangelversuchen. Bei Energiemangel steigen namlich die Anteile der langkettigen n-3 Fettsauren in verschiedenen Geweben an (KRAMER et al. 1984). Folgerichtig waren die Anteile dieser langkettigen n-3 Fettsiuren in den Phospholipiden in der Kontrollgruppe ahnlich wie bei den ad-libitum Kontrolltieren der der herkommlichen Zinkmangelver- suche. Beispielsweise betrug der Anteil der Docosahexaensaure im PE (Diacyl) in der vor- liegenden Arbeit bei der Kontrollgruppe 43 %, bei DRISCOLL und BETTGER bei der ad-libi- tum Kontrollgruppe 3,8%, bei der pair-fed Kontrollgruppe jedoch 5,9%; beim PE (Plasma- logen) betrugen die entsprechenden Anteile der Docosahexaensaure 7,5%, 7,8% bzw. 12,5%, beim PS 5,l Yo, 5,2% bzw. 8,5010 (Kontrollgruppe der vorliegenden Arbeit, ad-libi- tum- bzw. pair-fed Kontrollgruppe bei DRISCOLL und BETTCER 1991). Die Beobachtung, daB bei verminderter Futteraufnahme unphysiologisch hohe Anteile an langkettigen n-3 Fettsauren in der Erythrozytenmembran auftreten, kann ebenfalls als Beweis dafur gewertet werden, daR herkommliche Zinkmangelversuche zumindestens bei Untersuchungen im Bereich des Lipidstoffwechsel als sehr problematisch zu beurteilen sind.
Beim PC waren bei Zinkdepletion noch einige weitere Anderungen der Fettsaurezusam- mensetzung festzustellen, insbesondere der Anteile der Stearinsaure und der Linolsaure. Anderungen der Fettsaurezusammensetzung, die denen des PC der Erythrozytenmembran sehr ahnlich waren, traten auch im Plasma auf. Die prozentualen Anderungen bei der Depletionsgruppe betrugen fur die Palmitinsaure -6010 im Plasma bzw. -2% im PC der Erythrozytenmembran, fur die Stearinsaure +I6010 bzw. +28%, fur die Olsaure -8% bzw. -4010, fur die Linolsaure -21% bzw. -17% und fur die Docosahexaensaure +36% bzw. +57%. Diese Ergebnisse zeigen, dafl bei Zinkdepletion ein Zusammenhang zwischen den Anderungen der Fettsaurezusammensetzung des Plasmas und der des PC der Erythrozyten- membran besteht. Ein Hhnlicher Zusammenhang wurde bislang in der Literatur noch nicht aufgezeigt. Dieser Zusammenhang kann jedoch bei Betrachtung des Phospholipid-Turn- overs der Erythrozytenmembran erklart werden.
Generell sind Anderungen der Gehalte einzelner Phospholipide in den Erythrozyten- membranen und ihrer Fettsaurezusammensetzung nicht Folge einer de-Novo Synthese die- ser Lipide in den Erythrozyten, denn ausgereifte Erythrozyten sind nicht zur Lipidsyn- these befahigt (RENOIJ et al. 1976; RENOIJ und VAN GOLDE 1977; DISE et al. 1980). Der Turn- over der Phospholipide und ihrer Fettsauren in der Erythrozytenmembran wird durch ver- schiedene Prozesse bestimmt. Phospholipide der Erythrozytenmembran konnen mit denen des Plasmas ausgetauscht werden (SAKAGAMI et al. 1965; REED 1968). Dies trifft vor allem fur PC zu, das sich groBteils an der AuBenseite der Lipiddoppelschicht befindet (RENOIJ et al. 1976; RENOIJ und VAN GOLDE 1977). Dieser AustauschprozeR des PC zwi- schen Plasma und Erythrozytenmembran liefert auch eine mogliche Erklarung dafiir, daR sich die Anderungen der Fettsaurezusammensetzung bei Zinkdepletion in erster Linie in einer geanderten Fettsaurezusammensetzung des PC und weniger bei anderen Phospholi- piden der Erythrozytenmembran widerspiegelt. Wie Untersuchungen von SHOHET (1971) zeigten, wird mit Hilfe des Austauschprozesses taglich etwa ein Viertel des gesamten PC der Erythrozytenmembran ausgetauscht. Diese kurze Halbwertszeit des PC erklart auch, warum bereits innerhalb weniger Tage bei Zinkdepletion bereits relativ starke Veranderun-

Der Einzuji tion Zn-Depletion auf Erythrorytenmembranen 89
gen der Fettsaurezusammensetzung von PC auftraten. Andererseits findet aber auch nach langerer Zeit nur ein begrenzter Angleich der Fettsaurezusammensetzung zwischen dem PC der Erythrozytenmembran und dem PC des Plasmas statt (KUYPERS et al. 1988). Die Phospholipide, die sich an der Innenseite der Lipiddoppelschicht befinden (PE und PS) entstehen hauptsachlich aus Lysophospholipiden durch Veresterung mit freien Fettsauren, die ebenfalls aus dem Plasma stammen (MULDER und VAN DEENEN 1967; SHOHET et al. 1968; RENOIJ et al. 1976; RENOIJ und V A N GOLDE 1977). Bei diesen Phospholipiden waren bei den Depletionstieren hauptsachlich die Gehalte der langkettigen n-3 Fettsauren verandert. Dies spricht dafur, daR bei der Entstehung dieser Phospholipide aus ihren Lyso-Vorstufen bei Zinkdepletion vermehrt langkettige n-3 Fettsauren zur Veresterung venvendet werden.
Betrachtet man die Veranderungen der Phospholipide der Erythrozytenmembran bei den Zinkdepletionstieren, dann bieten sich zwei verschiedene Erklarungsansatze. Einer- seits ware es moglich, dai3 die beobachteten Veranderungen fur die verminderte Hamolyse- resistenz im Zinkmangel verantwortlich sind. Die Lipidzusammensetzung der Erythrozy- tenmembran bestimmt namlich wesentliche Eigenschaften der Erythrozyten, wie ihre Zell- form, Verformbarkeit, Permeabilitat und Hamolyseresistenz. Dabei ist vor allem das Men- genverhaltnis zwischen Cholesterin und Phospholipiden, das Mengenverhaltnis zwischen PC und SM sowie Kettenlange und Doppelbindungsindex der Fettsauren bedeutsam (STUBBS und SMITH 1984; SHINITZKY 1984; CULLIS et al. 1985). In der vorliegenden Unter- suchung war der Doppelbindungsindex bei den Gesamtfettsiuren im Zinkmangel um 10% erhoht, das Verhaltnis zwischen PC und SM um 11 Vo erniedrigt. Diese veranderten Para- meter konnten gemeinsam mit den erhohten Gehalten einzelner Phospholipide in der Erythrozytenmembran die verminderte Hamolyseresistenz im Zinkmangel erklaren.
Andererseits ware es moglich, daR die veranderte Lipidzusammensetzung der Erythro- zytenmembran nicht fur die verminderte Hamolyseresistenz ursachlich ist, sondern Folge einer Gegenregulation gegen andere zinkmangelspezifische, membrandestabilisierende Effekte ist. ROTH und KIRCHGESSNER (1991 a, 1992) untersuchten den EinfluR des Diatfettes bzw. einzelner ungesattigter Fettsauren der Diat auf die Himolyseresistenz von Rattenery- throzyten. Ihre Untersuchungen zeigten, daR die Zulage von Docosapentaensaure (20 : 5), einer langkettigen n-3 Fettsaure, zu einer linolsauredefizienten Diat bereits in geringen Mengen die Hamolyseresistenz von Rattenerythrozyten erheblich verbessert. Sie vermute- ten, daR dieser membranstabilisierende Effekt der Docosapentaensaure Folge des Einbaus dieser Fettsaure in die Erythrozytenmembran ist. Folgt man dieser Hypothese, dann durf- ten erhohte Anteile an langkettigen n-3 Fettsauren in der Erythrozytenmembran, wie bei Zinkdepletion beobachtet, die Hamolyseresistenz verbessern. Trotz der hoheren Anteile der n-3 Fettsauren ist im Zinkmangel die Hamolyseresistenz jedoch erniedrigt (PATERSON und BETTGER 1985; O’DELL et al. 1987; ROTH und KIRCHCESSNER 1991 a, b). Die erhohten Anteile membranstabilisierender langkettiger n-3 Fettsauren in der Erythrozytenmembran konnten demnach als Folge einer Gegenregulation mit dem Ziel, die im Zinkmangel ver- minderte Hamolyseresistenz zu verbessern, angesehen werden. Andere Untersuchungen zeigten namlich, daR es kompensatorische Veranderungen der Lipidzusammensetzung der Erythrozytenmembran mit dem Ziel der Aufrechterhaltung wichtiger Membraneigenschaf- ten gibt UAIN et al. 1982; SCHOUTEN et al. 1984; POPP-SNIJDERS et al. 1984; BEYNEN et a l . 1984). Beispielsweise kann ein erhohtes Verhaltnis zwischen Cholesterin und Phospholipi- den in der Erythrozytenmembran durch eine kompensatorische Erhohung der Anteile mehrfach ungesattigter Fettsauren ausgeglichen und damit die Fluiditat der Membran auf- recht erhalten werden (SCHOUTEN et al. 1984). Moglichenveise kann neben den erhohten Gehalten an n-3 Fettsauren auch der erhohte Gehalt einzelner Phospholipide in der Ery- throzytenmembran bei den Depletionstieren als eine Gegenregulation gegen zinkmangel- bedingte Membranveranderungen angesehen werden.

90 K. Eder und M. KircbgeJner
Zusammenfassung In der vorliegenden Arbeit wurde der EinfluR von Zinkdepletion auf die Gehalte einzelner Phospholipide in den Erythrozytenmembranen und deren Fettsaurenzusammensetzung unter- sucht. Mit Hilfe der Zwangsernahrung wurde innerhalb von 7 Versuchstagen ein starker Zink- mangel erzeugt, der durch einen um 71Yo verminderten Plasmazinkgehalt und eine um 66Yo verminderte Aktivitat der alkalischen Phosphatase irn Plasma belegt werden konnte. Die Deple- tionstiere zeigten erhohte Gehalte der Phospholipide Phosphatidylcholin, Phosphatidylethano- lamin (in Diacyl-Form und in Plasmalogen-Form), Sphingomyelin sowie der Summe der einzel- nen Phospholipide in den Erythrozytenmembranen. Die Fettsaurezusammensetzung der Phos- pholipide bei Zinkdepletion war gepragt durch erhohte Gehalte der langkettigen n-3 Fettsauren, insbesondere der Docosahexaensaure. Rechnerisch ergab sich fur die Gesamtfettsauren der Ery- throzytenmembran bei Zinkdepletion ein erhohter Doppelbindungsindex und eine erhohte mittlere Kettenlange. Die starksten Veranderungen der Fettsaurezusammensetzung bei Zink- depletion traten beim Phosphatidylcholin auf, wobei die Anteile der Stearinsaure, der Linol- saure, der Docosapentaensaure und der Docosahexaensaure verandert waren. Im Plasma traten Veranderungen der Fettsaurezusammensetzung auf, die denen des Phosphatidylcholin der Ery- throzytenmembran entsprachen. Die beobachteten Veranderungen der Gehalte der Erythrozy- tenmembranphospholipide und ihrer Fettsaurezusammensetzung konnten einen Erklarungs- ansatz fur die im Zinkmangel verminderte Hamolyseresistenz liefern, konnten aber auch Folge einer Gegenregulation gegen die im Zinkmangel verminderte Hamolyseresistenz sein.
Summary Influence of zinc defciency on the contents of individual phospholipids and their fatty acid compositions
in erythrocyte membranes of forcedfed rats
In the present study the influence of zinc deficiency o n the contents of individual phospholi- pids and their fatty acid compositions in erythrocyte membranes was investigated. Using the forced-feeding technique within seven days severe zinc deficiency was induced which was veri- fied by a reduced plasma zinc concentration (-71%) and by a reduced activity of plasma alka- line phosphatase (-66Vo). Erythrocyte membranes of zinc deficient animals had higher con- tents of the phospholipids phosphatidylcholine, diacyl phosphatidylethanolamine, plasmalo- gen phosphatidylethanolamine, sphingomyelin as well as total phospholipids. Erythrocyte membrane phospholipids of zinc deficient animals had higher contents o f long chain n-3 fatty acids, especially of docosahexaenoic acid. Erythrocyte membrane total fatty acids of zinc deficient animals also had a higher double bound index and a higher mean chain length than those of control animals. The largest changes occured in the fatty acid composition ofphospha- tidylcholin. In this phospholipid the contents of stearic acid, linolic acid, docosapentaenoic acid and docosahexaenoic acid where changed in zinc deficient animals. In zinc deficient ani- mals there were also changes o f the composition o f plasma fatty acids similar to those in ery- throcyte membrane phosphatidylcholine. The observed changes of the contents of phospholi- pids and their fatty acid compositions in erythrocyte membranes might give an explanations for the increased osmotic fragility of erythrocytes in zinc deficiency. O n the other hand, the changes of phospholipid contents and their fatty acid composition in zinc deficiency might be the result of a compensatory mechanism in order to decrease the osmotic fragility which is higher in zinc deficiency.
Literatur
BEHRENS, G.; PALLAUF, J., 1992: Kurzfassungen der Vortrage der 46. Tagung, Gesellschaft fur
BEYNEN, A. C.; SCHOUTEN, J. A.; POPP-SNIJDERS, C., 1984: Trends Biochem. Sci. 9, 474. BJERVE, K. S.; FISCHER, S.; ALME, K., 1987: Am. J. Clin. Nutr. 46, 570. CARTWRIGHT, I. J.; POCKLEY, A. G.; GALLOWAY, J. H.; GREAVES, M.; PRESTON, F. E., 1985:
Atherosclerosis 55, 267. CULLIS, P. R.; HOPE, M. J.; DE KRUIJEF, B.; VERKLEIJ, A. J.; TILCOCK, C. P. S., 1985: Structural
properties and functional roles of phospholipids in biological membranes. In: Phospho- lipids and cellular regulation. Ed. by J. F. Kuo. Vol. 1, pp. 1-60, CRC Press, Boca Raton.
Ernahrungsphysiologie, FrankfurtIMain, pp. 70-72.
DUE, C. A,; GOODMAN, D. B. P.; RASMUSSEN, H., 1980: J. Lipid Res. 21, 292.

Der EinfluJ3 von Zn-Depletion auf Erythrocytenmembranen 91
DRISCOLL, E. R.; BETTGER, W. J., 1991: Lipids 26, 459. EDER, K.; REICHLMAYR-LAIS, A. M.; KIRCHGESSNER, M., 1992: J. Chromatogr. 598, 33. GLATZ, J. F. C.; SOFFERS, A. E. M. F.; KATAN, M. B., 1989: Am. J. Clin. Nutr. 49, 269. JAIN, S . K.; MOHANDAS, N.; SENSABAUGH, G. F.; SHOJANIA, A. M.; SHOHET, S. B., 1982: J. Lab.
JAY, M.; STUART, S. M.; MCCLAIN, C . J.; PALMIERI, D. A.; BUTTERFIELD, D. A,, 1987: Biochim.
JOHANNING, G. L.; O’DELL, B. L., 1989: J. Nutr. 119, 1654. KIRCHGESSNER, M.; SCHULEIN, A.; ROTH, H. P.; SCHLIACK, G., 1992: J. Anim. Physiol. a. Anim.
KRAMER, T. R.; BRISKE-ANDERSON, M.; JOHNSON, S. B.; HOLMAN, R. T., 1984: J. Nutr. 114, 1224. KUYPERS, F. A,; ABRAHAM, S.; LUBIN, B.; CHIU, D., 1988: J. Lab. Clin. Med. 111, 529. LEVEILLE, G. A,, 1970: Fed. Proc. 29, 1294. MULDER, E.; VAN DEENEN, L. L. M., 1965: Biochim. Biophys. Acta 106, 106. NATIONAL RESEARCH COUNCIL, 1978: Nutrient requirements of laboratory animals, 3rd edition,
O’DELL, B. L.; BROWNING, J. D.; REEVES, P. G., 1987: J. Nutr. 117, 1883. PATERSON, P. G.; BETTGER, W. J., 1985: In: Trace Elements in man and animals. Ed. by C. F.
MILLS, I. BREMNER, J. K. CHESTERS. pp. 79-83, Commonwealth Agricultural Burreaux, Slough, UK.
POPP-SNIJDERS, C.; SCHOUTEN, J. A.; DE JONG, A. P.; VAN DER VEEN, E. A., 1984: Scand. J. Clin. Lab. Invest. 44, 39.
PUTNAM, J. C.; CARLSON, S. E.; DEVOE, P. W.; BARNESS, L. A,, 1982: Am. J. Clin. Nutr. 36, 106. REED, C. F., 1968: J. Clin. Invest. 47, 749. RENOIJ, W.; V A N GOLDE, L. M. G., 1977: Biochim. Biophys. Acta 470, 465. RENOIJ, W.; VAN GOLDE, L. M. G.; ZWAAL, R. F. A.; VAN DEENEN, L. L. M., 1976: Eur. J . Bio-
ROTH, H. P.; KIRCHGESSNER, M., 1992: Ann. Nutr. Metab. 36, 113.
-; -, 1991 b: J. Anim. Physiol. a. Anim. Nutr. 65, 273. SAKAGAMI, T.; MINARI, 0.; ORII, T., 1964: Biochim. Biophys. Acta 98, 111. SCHNEEMAN, B. 0.; LACY, D.; NEY, D.; LEFEVRE, M. L.; KEEN, C . L.; LONNERDAL, B.; HURLEY, S. L.,
SCHOUTEN, J. A,; BEYNEN, A. C.; POPP-SNIJDERS, C.; VAN DER VEEN, E. A,, 1984: Nutr. Rep. Int.
SCHULEIN, A,; KIRCHGESSNER, M.; ROTH, H. P., 1992: J. Anim. Physiol. a. Anim. Nutr. 67, 157. SHINITZKY, M., 1984: Membrane fluidity and cellular functions. In: Physiology of Membrane
SHOHET, S. B., 1971: J. Lipid Res. 12, 139. SHOHET, S . B.; NATHAN, D. G.; KARNOVSKY, M. L., 1968: J. Clin. Invest. 47, 1096. STUBBS, C. D.; SMITH, A. D., 1984: Biochim. Biophys. Acta 779, 89.
Anschrrft der Autoren: K. EDER und M. KIRCHGESSNER, Institut fur Ernahrungsphysiologie der
Clin. Med. 99, 816.
Biophys. Acta 897, 507.
Nutr. 67, 188.
pp. 7-38, Natl. Acad. Sci., Washington.
chem. 61, 53.
-. , - , 1991a: Z. Ernahrungsw. 30, 98.
1986: J. Nutr. 116, 1889.
29, 1223.
Fluidity. Ed. by M. SHINITZKY. Vol. 1, pp. 1-52, CRC Press, Boca Raton.
TU Miinchen, D-8050 Freising-Weihenstephan, FR Germany














![Kapitel 5) Lipide [Kompatibilitätsmodus] · Diese Lipide gehören zur Gruppe der Phospholipide. Dabei sind zwei Fettsäuren mit dem Glycerin verestert, am dritten C-Atom des Glycerins](https://img.pdfslide.org/doc/110x75/5e1985ec82bc82503761ec99/kapitel-5-lipide-kompatibilittsmodus-diese-lipide-gehren-zur-gruppe-der-phospholipide.jpg)













