
Albrecht v. Graefes Arch. klin. exp. 0phthM. 172, 112--124 (1967)
Adaptive Parameter der Blauverschiebung der Spektralsensitivit~it des Kaninehenauges*
1~. KSINSlK Abteilung flit Experimentelle Ophthalmologie (II. Physiol. Abt.)
des W. G. Kerckhoff-Instituts der Max-Planck-Gesellschaft, Bad Nauheim
Eingegangen am 3. November 1966
Beim Kaninchen wird eine Purkinje-Verschiebung vermiBt [6, 9], obwohl seine 2qetzhaut ein sowohl Stabchen wie Zapfen enthaltendes Sinnesepithel aufweist [17, 21]. Messungen der spektralen Empfindlich- keit einzelner Retinaganglienzellen des Kaninchens zeigen jedoch neben Werten ahnlich der Absorptionskurve des Sehpurpurs andere mit einem Nebenmaximum ira Blau und Grtin [5, 15]. Uber ahnliche Verteilungen der spektralen Reizwerte wird bei Verwendung optomotorischer Metho- den berichtet [20]. Nachdem Retinaganglienzellen mit besonderer Blau- empfindlichkeit besonders bei helladaptierten Kaninchen beobachtet wurden [5], stellt sich die folgende Untersuchung die Aufgabe, die spektrale Empfindlichkeit des gleichen Tieres bei stufenweise gestei- getter Helladaptation elektroretinographisch zu messen, um die adap- riven Bedingungen der Blauversehiebung nigher kennenzulernen.
Methodik Pr~iparation. ~3ber 2qacht dunkelad~ptierte, 2,0--3,8 kg schwere Kaninchen
wurden bei rotem Licht (RG 5) mi~ Urethan (25%ige LSsung, 1,5 g/kg KSrper- gewicht) narkotisiert. Iqach Kaniilierung der Trachea wurde das Tier mit einem Kopfhalter fixier~ und nach ErSffnung des Sch~dels durch Absaugen des GroB- hirns dezerebriert. Nickhaut und Augenlider sowie der Bulbus (an der Insertions- stelle der Augenmuskeln) wurden mit F~den fixiert. Zur Pupillenerweiterung und Ausschaltung der Pupillomotorik diente Atrop. sul/. (1%ig). Die in den meisten F~Lllen naeh Dezerebrierung zu beobachtende Erh5hung der KSrpertemperatur auf etwa 41~ machte die W~rmezufuhr dutch ein Heizkissen in der Regel iiber- fliissig.
Elelrtrische Ableitung und Verstdrlcung. Das Elektroretinogramm wurde mit zwei Silber-Silberchlorid-Ringer-Baumwoll-Elektroden zwisehen Limbus und ektro- pioniertem Oberlid abgeleite~. Bei Vcrwendung einer Haftschale bestand die diL ferente Elektrode aus Silber-Silberchlorid. Die abgeleiteten PotentiMe wurden naeh Gleichstromverstgrkung (Vorverstarker yon J.F. TSimies-Freiburg sowie Tektronix Typ 2 A 63) auf einem Ka~hodenstrahloscillogTaphen (Tektronix Typ 565) dargestellt und photographiert. In einigen Versuehen wurden die abgeleiteten Potentiale einem Computer o/ Average Transients (Technical Measurement Cor- poration, Typ 400 C, North Haven, Conn.) zugefiihrt. Elektronisch gemittelte
* Mit Untersttitzung der Deutschen Forschungsgemeinsehaft.

Adaptive Parameter der Blauversehiebung 113
Potentiale, ,,konventionelles" Elektrore~inogramm und Lichtreizmarkierung wurden auf drei Strahlen des Oscillographen simultan darges~ell~ und photo- graphisch registriert (vgl. Abb. 4). Elektronenreehner und Liehtreiz wurden so gesteuer~, dab die elektronische Mittelwertsbildung 70 msee vor Beginn des Lieh~- reizes einsetzte.
Lichtreizung. Verwandt wurde ein Zweistrahl-Adaptometer mit einer gleich- strombetriebenen Xenon-Kurzbogenlampe (Typ XBO 150 W/l , Osram). Zur Aus- filterung monochromatisehen Lichts zwischen 401 und 630 m~z dienten Interferenz- Bandfilter (Typ AL, Sehot~ & Gem, Mainz) bekannter spek~raler Durchl~ssigkei~. Die energetische Eiehung der monoehromatisehen Liehter effolgte mit einem geeiehten S~rahlungsthermoelemen~ (Typ B, Pyro-Werk, Hannover) ; ein l~ultiflex- Galvanometer (Typ MGF, Dr. Lange, Berlin) diente Ms Anzeige. Zur selektiven Adaptation dienten Absorptionsfilter (OG 5, in Einzelf/illen BG 25, Schott & Gen., M~inz). Die Beleueh~ung der Iqe~zhaut erfolgte in einigen Versuehen nach dem l~inzip tier 1~Iaxwellsehen Beobaeh~ung bei einer Tes~feldgrSl3e yon 7 ~ in anderen Versuehen mit Hilfe einer weii]mattierten HaftsehaIe (Ganzfeldbe]euchtung). In der Regel betrug die Reizdauer 0,1 see, zur getrennten Darstellung yon on- und off-Effekt in einigen Versuchen 0,2 see. Unterbreehung und Freigabe der Lichtreize erfolg~e fiber einen Lichthahn (Drehmagnet), der yon einem Reeh~eek-S~imulator (Grass Typ S-4) gesteuert wurde. Zur Kontrolle der Reizdauer diente eine Photo- zelle, die yon einer planparallelen Glasplatte im Teststrahl beliehtet wurde.
Photometrie. Die maximale Leuehtdicht~ des zur Sehwellenmessung und Adap- tation verwendeten weiBen Lieh~s betrug 104 Nit; sie wurde jeweils bei Versuchs- ende mit einem S.E.I. Exposure Photometer (Salford Electrical Instruments, Ilford) auf der dem Auge zugekehrten Seite der mattierten Haftschale gemessen. Durch Neutralfilter bekannter spektraler ])urehliissigkeit konnte die Leuchtdiehte in Stufen yon 10~ 45 in einem Bereich yon insgesamt 10 s abgeschw~cht werden. Bezfiglich der Bestrahlungsst~rken monochromatisehen Liehts vgl. Abb. 3.
Yersuchsgang. l~aeh Preparation des Tieres wurde die Dunkeladaptation er- neuert, bis Konstanz der Reizschwelle im Elektroretinogramm eintra~. Gemessen wurde die HShe der b-Welle yon der Nnllinie bzw. (falls vorhanden) von tier Senke der a-Welle. Der Abstand zwischen zwei aufeinanderfolgenden Testreizen betrug 5--10 sec (helladaptierte Augen) bzw. 30 see (dunkeladaptierte Augen). Zur elek- tronischen Mittelung yon Netzhautpotentialen unter ttelladaptation wurde mit einer Beliehtungsfrequenz yon 1,5 bzw. 7 Hz gearbeitet. Zur Bestimmung der Kennlinie der b-Welle (b-Wellenamplitude als Funktion der Testreiz-Leuchtdich~e) wurden die benutzten Reizwellenl~ngen (473 und 546 mtz ) alternierend dargeboten. ~nderungen im Erregungszustand wurden somit g]eiehm~l]ig verteilt. Die Dauer der Helladaptation vor der Messung betrug jeweils 2 rain und mehr. Adaptive Ein- fl/isse w~hrend der Messung wurden dutch regelm~Gige Beliehtungen mi~ einer Kontroll-Wellenl~nge effal]t.
Ergebnisse
Die elek~rore~inographische Messung der Spek t ra l sens i t iv i t s des d u n k e l a d a p t i e r t e n Kan inchenauges ergibt ein M~ximum im blaugrf inen Bereieh [24], wobei die l~eizwerte be im p igmen t i e r t en Kan inehen (beim Alb inokan inehen ers~ naeh Ausseha l tung der L ieh t s t r euung ve to reflek- t i e renden Augenhin te rgrund) sieh i~hnlieh der Absorp t ionskurve des Sehpurpurs ver te i len [10, 11]. Eine ~hnliche spek t ra le Ver te i lung zeigt das K a n i n c h e n bei f requenter i n t e rmi t t i e r ende r IAchtreizung, bei der

114 1~. KSlI~SlK:
die ebenfalls yon St~bchen dominierte Katzennetzhaut eine Purkinje- Verschiebung erkennen 1/~t]t [9]. Das Fehlen eJuer Purkinje-Verschiebung beim Kaninchen wurde auch in der Dunkeladaptat ionskurve [6] und w~hrend stufenweise gesteigerter adaptiver Beleuchtung festgestellt [4].
Man kann dieses Resultat in verschiedener Weise interpretieren. So kann die Spektralsensitivits des skotopischen Prozesses mit der des photopischen Prozesses iibereinstimmen, wenn beide Prozesse ein- und dasselbe Photopigment benutzen. In diesem FMle ist ffir den skotopischen und den photopischen Prozel~ zw~r verschiedene absolute Schwelle, doch gleiche Spektralempfindlichkeit zu erwarten. Die spektrale Zusammen- setzung des Adaptationslichts ist in diesem Fall fiir den eingetretenen Empfindlichkeitsverlust ohne Bedeutung. Eine andere Erkl~rung ver~ sucht die fehlende Purkinje-Verschiebung mit der Existenz yon zwei Prozessen mit verschiedener absoluter und spektraler Empfindlichkeit zu vereinen: Die Verschiebung der l~eizwerte kann gering sein oder ausbleiben, wenn die Liehtfarbe der zur Helladaptat ion benutzten adaptiven Beleuchtung dem spektralen Empfindlichkeitsmaximum des weniger sensitiven Prozesses entspricht. Bei einer Netzhaut mit normaler Purkinje-Verschiebung ist diese Situation bei Helladaptat ion mit lang- welligen Lichtern gegeben; dagegen unterscheiden sich blaues und weiBes JAcht nur geringfiigig in der Fgh~gkeit, eine Purkinje-Verschiebung herbei- zuffihren [8]. U m die genannten MSglichkeiten beim Kaninchen zu prfifen, wurde die Farbe des zur Helladaptat ion benutzten weiBen (Xenon-)Lichts in einigen Versuchen mit Hi]fe yon Absorptionsffltern vergndert. Zur Entfernung des Blauanteils aus dem Adaptationslicht diente ein Farbglas (OG 5, Schott & Gen., Mainz) mit stefler Absorp- t ionskante gegenfiber kfirzeren Wellenl/~ngen.
Der spek~rale DurchlaBgrad yon OG 5 in 2 mm Dieke betr~g~ bei 535 m~ 0,5; unterhalb 520 m~ ist die Durehl~ssigkeit kleiner als 0,005. Beim Kaninehen, fiir dessen Netzhaut Liehter yon 500 m~ den gr61]ten Reizwe~ besitzen, wird hierdureh der wirksame Strahlungsan~eil merkbar verminder~, wEhrend eine deut- liche Leuchtdiehteverminderung f~r das helladaptierte mensehliche Auge nicht eintritt. Da sieh die pho~ometrischen GrSBen auf das spek~rale Verhal~en des helladaptierten menschliehen Auges beziehen~ dessen Empfindlichkeitsmaximum (560 m~) stark yon dem des Kaninehens (500 m~) abweieht, wurde zur Bestimmung der Leuchtdiehte blaufreien Liehts seine wEhrend Dunkeladap~ation bestimmte Wirks~mkeit mit einem weiBen Lich~ bekannter Leuchtdiehte vergliehen, ttierzu wurden die Amplituden der b-Welle in AbhEngigkeit yon der Testliehtintensit~t vergleiehsweise ffir weil]es und blaufreies Lieht aufgezeichne~. Dabei ist das blau- freie Lieh~ beim Kaninchen etwa viermM weniger wirksam als das weiBe Lieht. Um diesen Betrug wurde die Abszisse der D~rstellungen, deren D~ten unter blau- freier ~dap~iver Beleuehtung gewonnen wurden, nach links versehoben (,,skotopiseh ~quiv~lente Liehter").
Zur Untersuchung der relativen spektralen Empfindliehkeit der K~ninchermetzhaut bei verschiedenem Adapt~tionszustand wurde das
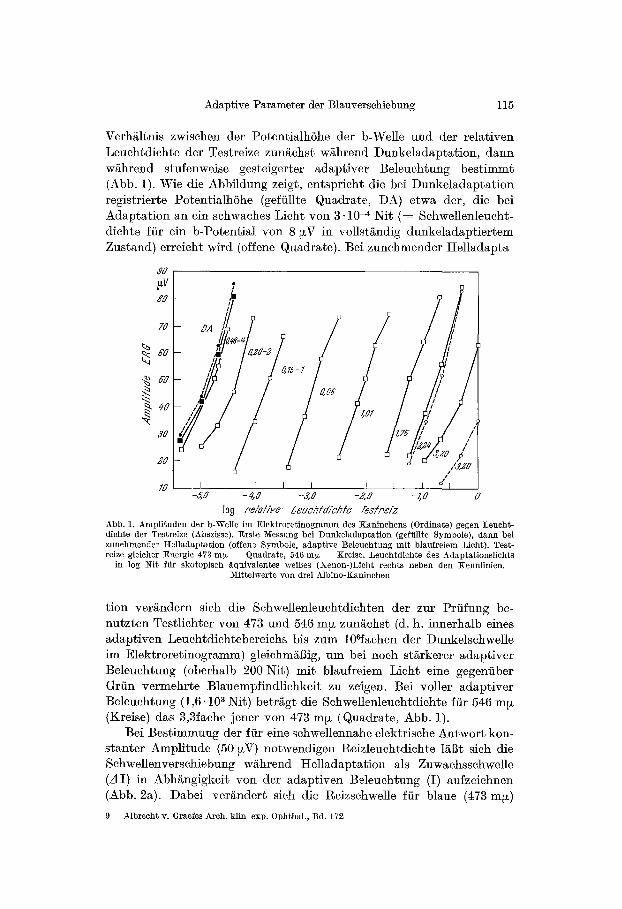
Adaptive Parameter der Blauverschiebung 115
Verh~ltnis zwisehen der Potentialh6he der b-Welle und der relativen Leuchtdichte der Testreize zun~ehst w~hrend Dunkeladaptation, dann wghrend stufenweise gesteigerter adaptiver Beleuchtung bestimmt (Abb. 1). Wie die Abbildung zeigt, entspricht die bei Dunkeladaptation registrierte PotentiMhShe (gefiillte Quadrate, DA) etwa der, die bei Adaptation an ein schwaches Licht von 3" 10 -4 Nit ( = Schwellenleucht- dichte ffir ein b-Potential yon 8 ~V in vollst~ndig dunkeladaptiertem Zustand) erreicht wird (offene Quadrate). Bei zunehmender Itelladapta
80 lxV
70
%
I . z/O
80
BO
10
J f 711~+8-~1 ? i~ l u, w-z l
I I 1 I -$o - 4,0
D ////I I 1" d /I
. x
I I I I I 4 1 I -LO -AO -1,,o 0
log pelaliye leucbldichle Tesfpeiz Abb. 1. Amplituden der b-Welle im Elektroretinogramm des ~:aninchens (Ordinate) gegell Leucht- dichte der Testreize (Abszisse). El'ste Messung bei nunkeladaptation (gefiillte Symbole), dann bei zunehmencler ttelladaptation (offene Symbole, adaptive Beleuchtang mit blaufreiem Licht). Test- reize gleieher Energie 473 m~ - - Quadrate, 546 nip - - Kreise. Leuchtdichte des Adaptationslichts
in log Nit fiir skotopisch ~quivalentes wciBes (Xenon-)Lieht rechts neben den Kennlinien. Mittelwerte yon drei Albino-Kaninchen
tion ver/~ndern sieh die Schwellenleuchtdiehten der zur Prfifung be- nutzten Testliehter yon 473 und 546 m~ zun/tchst (d. h. innerhalb eines adaptiven Leuchtdichtebereichs bis zum 10Gfachen der Dunkelsehwelle im Elektroretinogramm) gleiehm/iBig, um bei noeh stgrkerer adaptiver Beleuchtung (oberhalb 200 Nit) mit blaufreiem Lieht eine gegenfiber Grfin vermehrte B]auempfindlichkeit zu zeigen. Bei roller adaptiver Beleuchtung (1,6.10 a Nit) betrggt die Sehwe]lenleuchtdiehte Ifir 546 m~ (Kreise) das 3,3fache jener yon 473 m~ (Quadrate, Abb. 1).
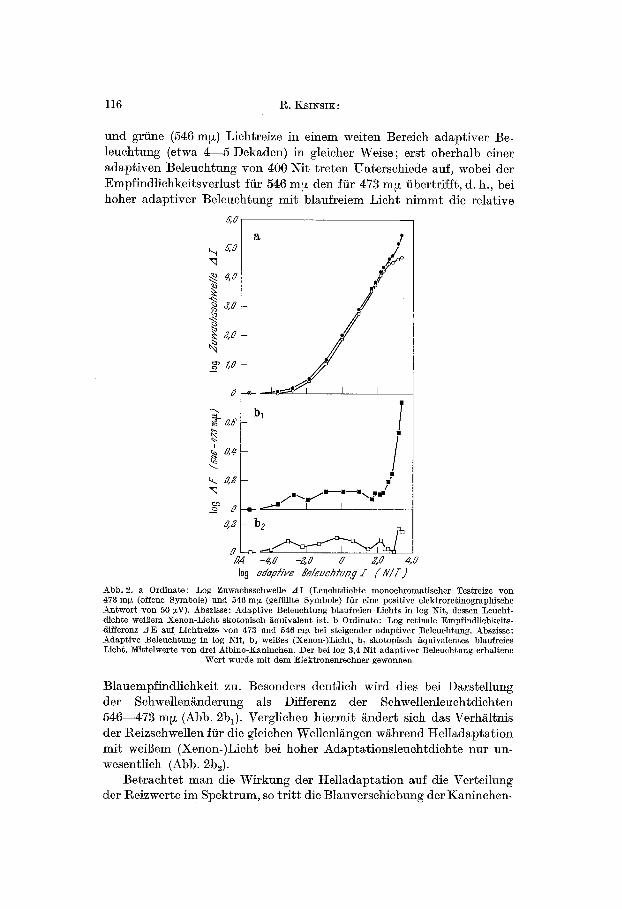
Bei Bestimmung der fiir eine sehwellennahe elektrische Antwort kon- stanter Amplitude (50 ~V) notwendigen t~eizleuehtdiehte li~Bt sich die Schwellenversehiebung w/~hrend Helladaptation als Zuwaehsschwelle (A I) in Abhgngigkeit yon der adaptiven Beleuchtung (I) aufzeichnen (Abb. 2a). Dabei ver~ndert sieh die l~eizsehwe]le fiir blaue (473 m~)
9 Albrecht v. Graefes Arch. klin, exp. Ophthal., Bd. 172
116 I~. KSINSIK :
und gr/ine (546 m~) Lichtreize in einem weiten Bereich adaptiver Be- leuchtung (etwa 4- -5 Dekaden) in gleicher Weise; erst oberhalb einer adaptiven Beleuchtung yon 400 Ni~ treten Unterschiede auf, wobei der Empfindliehkeitsverlust ffir 546 m~z den f/it 473 m~t fibertrifft, d. h., bei hoher adaptiver Be]euchtung mit blaufreiem Licht nimint die relative
6,o
5,o
/ - / , 0
s,o
f,o
0
0,0 ~
i
o o,z
bl f
_ ~ . / ' " ~ / ' - - ~ - ' 'Z'J"
b~.
DA -~,o -z,o o 2,0 4,0 log adaptive #eleuchfun# Z (N/T)
Abb. 2. a Ordinate: Log Zuwaehssehwelle zlI (Leuehtdiehte monoehromatiseher Testreize yon 473 mix (offene Symbole) und 546 m~ (geftillte Symbole) fOr eine positive elektroretino~aphische Antwort yon 50 gN). Abszisse: Adaptive :Beleuchtung blaufreien Lichts in log Nit, dessert Leucht- diehte weiBem Xenon-Licht skotopiseh f~quivalent is~. b Ordinate: Log retinale Empfindliehkeits- differenz A E attf Liehtreize yon 473 und 546 m~ bei steigender adaptiver :Beleuehtung. Abszisse: Adaptive Beleuchtung in log Nit, b~ weiges (Xenon-)Licht, b1 skotopisch /iquivalentes blaufreies Licht. Mittelwerte yon drei Albino-:Kaninchen. Der bei log 3,4 Nit adaptiver Beleuchtung erhaltene
Weft wurde mit dent Elektronenreehner gewonnen
Blauempfindlichkeit zu. Besonders deutlich wird dies bei Darstellung der Schwellengnderung als Differenz der Sehwellenleuehtdiehten 546 .473 m~ (Abb. 2bl). Vergliehen hiermit gndert sich das Verhgltnis der t~eizsehwellen ffir die gleiehen Wellenl/~ngen wghrend I tel ladaptat ion mit weiBem (Xenon-)Licht bei hoher Adaptationsleuehtdichte nur un- wesentlich (Abb. 2b~).
Betraehtet man die Wirkung der I tel ladaptat ion auf die Verteilung der Reizwerte im Spektrum, so t r i t t die Blauverschiebung der Kaninchen-
A d a p t i v e P ~ r a m e t e r d e r B l a u v e r s c h i e b u n g 117
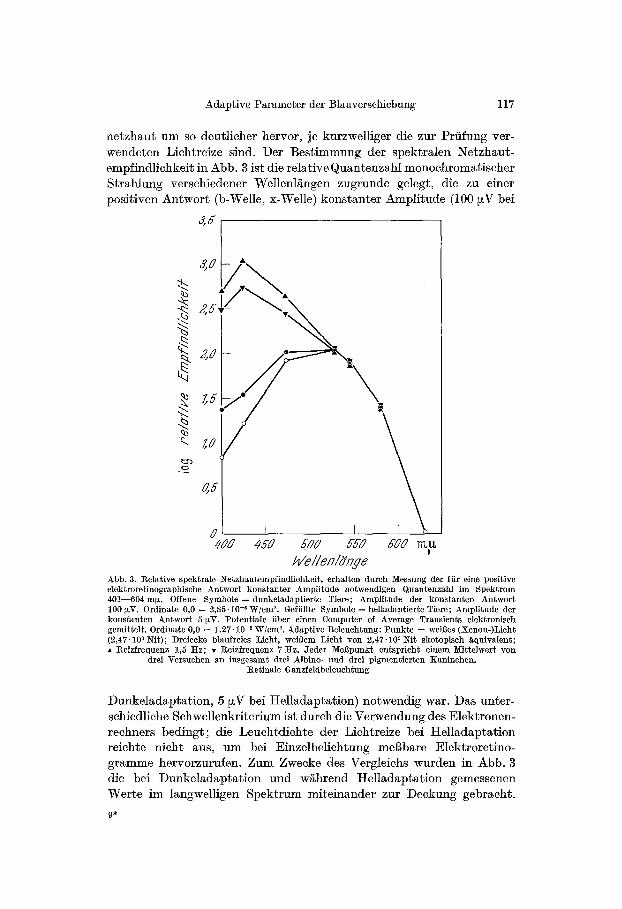
netzhaut um so deutlicher hervor, je kurzwelliger die zur Prtifung ver- wendeten Lichtreize sind. Der Bestimmung der spektralen Netzhaut- empfindlichkeit in Abb. 3 ist die relative Quantenzahl monochrom~tischer Strahlung verschiedener Wellenl~ngen zugrunde gelegt, die zu einer positiven Antwort (b-Welle, x-Welle) konstanter Amplitude (100 ~V bei
J,5
~ 2 f -
r..
0 l I I [ ~ dO0 ~gO gO0 5#0 800 ~
Wellenldn#e Abb. 3. l~elative spektrale Netzhautempfindliehkeit, erbalteu dutch Messung der ffir eine positive elektroretinographische Antwort konstanter Amplitude notwendigen Quantenzahl im Bpektntm 401--604 m~. Offene Symbole = dunkeladaptier~e Tiere; Amplitude der konstanten Antwort 100 ~V. Ordinate 0,0 = 2,85"10 -6 WIomK Geffillte Symbole = helladaptierte Tiere; Amplitude der konstanten Antwort 5 VV. Potentiale fiber einen Computer of Average Transients elektroniseh gemittelt. Ordinate 0,0 = 1,27.10 -2 W/cm ~. Adaptive Beleuehtung: Punkte = weil3es (Xenon-)Lieht (~,47. l0 s Nit); Dreieeke blaufreies Lieht, wei~em Lieht yon 2,47.10 s Nit skotopiseh ~qdivalent; �9 l~eizfrequenz 1,5 Itz; , l~eizfrequenz 7 ttz. Jeder MeBpunkt entsprieht einem Mittelwert yon
drei Versuehen an insgesamt drei Albino- und drei pigmentierten Xaninehen. ]~etiuale Ganzfeldbeleuchtung
Dunkelad~pt~tion, 5 ~V bei Helladapt~tion) notwendig war. Das unter- schiedliche Schwellenkriterium ist durch die Verwendung des Elektronen- rechners bedingt; die Leuchtdichte der Lichtreize bei t telladaptation reichte nicht aus, u m b e i Einzelbelichtung meBbare Elektroretino- gramme hervorzurufen. Zum Zwecke des Vergleichs wurden in Abb. 3 die bei Dunkelad~ptation und w~hrend Helladaptation gemessenen Werte im ]angwel]igen Spektrum miteinander zur Deckung gebracht.
9*

118 R. KSINSIK :
Die Darstellung zeigt bei Helladaptation mit. unbuntem Lieht (Abb. 3, Punkte) f/Jr kurzwellige Lichtreize die bereits von anderen Autoren be-
580
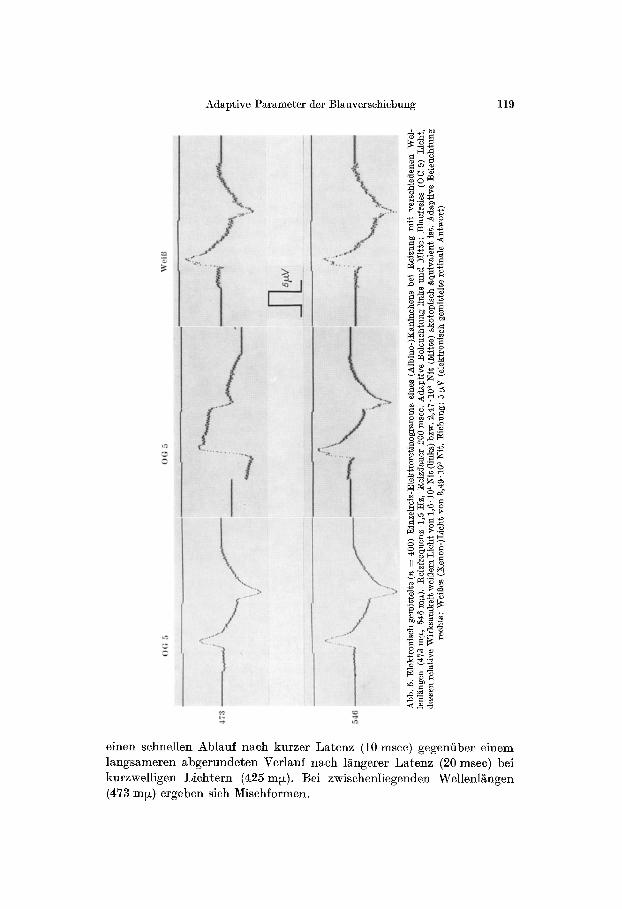
Abb. 4. Lichtreizmarkieruag (oben), elektro- nisch gemittelte :Einzelreiz-s me (Mitre) und ]~inzelreiz-]glektroretinogram- me (unten) des Kaninchens bei Reizung mit verschiedenen Wellenlgngen wiihrend Hell- adaptation mit blaufreiem Lieht, das weigem Lieh~ ~on 2,47.10 s Nit skotopisch ~quivalent ist. Reizdauer 200 msec, Reizfrequenz 1,5 Ha. g ichung: 300 g u (Einzel-Elektroretino- gramm), 5 pN (elektronisch gemitteltes ]~lek- troretinogramm, n = 400). Einzelversuch.
Ptgmentiertes Kaninchen
schriebene geringe Empfindlich- keitssteigerung [5,19, 23]. Vergliehen hiermit ist die bei Helladaptation mit blaufreiem Licht (Abb. 3, Drei- ecke) zu beobaehtende Empfind- lichkeitssteigerung im kurzwelligen Spektrum um ein Mehrfaches grS- Ber. Dureh die Empfindliehkeits- steigerung im kurzwelligen Spektrum verschiebt sich das Sensitivitits- maximum unter tte]ladaptation mit blaufreiem Lieht yon Blaugriin nach Violett.
Eine Besonderheit tier unter Helladaptation mit blaufreiem Licht hervortretenden Blauempfindlieh- keit ist ihre besondere ]Prequenz- abhgngigkeit: Erfolgt die elektro- nische Mittelwertsbildung der Retina- potentiale bei einer Lichtreizfre- quenz von 1,SHz (Abb. 3) auf- rechte Dreiecke), so ist die Empfind- lichkeit im kurzwelligen Spektrum-- bezogen auf die im lgngerwelligen Spektrum erhaltenen Werte-- relativ gr6Ber als bei einer Lichtreizfrequenz yon 7 Hz (abw/irtsgeriehtete Drei- eeke). Diese Abh/~ngigkeit der I~eiz- sehwelle yon der Belichtungsfrequenz wird verst/~ndlieh, wenn man die bei ttelladaptation je nach Reizwellen- lgnge untersehiedliehe Potentialform betrachtet, deren Registrierung die Verwendung des Elektronen- reehners ermSglieht. Die in Abb. 4 bei einer adaptiven Beleuchtung yon 2470 Nit (etwa dem 109f~ehen der Absolutschwelle des mensehliehen Auges entspreehend [18]) registrier- ten Retinapotentiale zeigen bei 1/~n- gerwelligen Lichtreizen (507,580 m~)
Adaptive Parameter der Blauverschiebung 119
�9
i ~iiii!i 2:/
:ii
~/!i:~ ~!i!'ii~ i ~ ~/ii ~ii~i~I~~'!'i ii~i~ ~ !~i,~i~, ~
. ~
~
~.~ ~
~ . .
o
einen schnellen Ablauf n~ch kurzer Latenz (10 msec) gegeniiber einem langsameren abgerundeten Verlanf nach langerer Latenz (20 msec) bei kurzwelligen Lichtern (425 ml~ ). Bei zwischenliegenden Wellenl~ngen (473 m~) ergeben sich Mischformen.

120 R. KsI~SlK:
Der Einflul~ farbiger Helladaption auf die Netzhautpotentiale des Kaninchens kann folgendermal~en beschrieben werden: Im adaptiven Leuchtdichtebereich unterhalb yon 100 Nit ist die Farbe der Adaptations- liehter ohne Bedeutung fiir die Potentialform (blaufreie Helladaptation, Abb. 5, links). Bei ttelladaptation oberhalb 400 Nit erh~lt man bei Verwendung blaufreien Adaptationslichtes (Abb. 5, Mitre) auf Lichtreize von 473 m~ skotopische, auf Reize yon 546 m~ photopische Potential- formen, tfelladaptation mit weil~em (Xenon-)Licht hoher Leuchtdichte fiihrt auch auf Liehtreize yon 473 m~ zu photopischen Potentialen (Abb. 5, reehts), doch sind auf kiirzere Wellenl/~ngen (401,425 m~) auch unter WeiBadaptation skotopische Potentiale registrierbar. J~hnliche Befunde wie bei Helladaptation mit weil~em Licht ergeben sich bei Ver- wendung rotfreien Lichts (BG 25).
Die Dauer der Helladaptation hat auf die Form der abgeleiteten Potentiale keinen deutlichen EirdluB. Nach 15 rain adaptiver Beleueh- tung mit 104Nit war bei Wechsel zu blaufreier Helladaptation mit 2,47-103 Nit die in Abb. 5 (Mitre) beschriebene Potential/~nderung un- mittelbar erkennbar.
Diskussion Zur Deutung der Blauversehiebung der spektralen l~eizwerte der
Kaninchennetzhaut bei hoher adaptiver Beleuehtung bestehen folgende MSglichkeiten:
1. Helladaptation bewirkt eine Veranderung der absorptiven Eigen- schaften des Sehpurpurs. Die entstehenden Produkte sind selbst licht- empfindlich und absorbieren bei kiirzeren Wellenlangen (Ein-Pigment- Hypothese der Blauverschiebung).
2. Die Verschiebung der I~eizwerte bei Helladaptation beruht auf einer Aktivierung von Netzhautelementen mit besonderer Blauempfind- lichkeit. Helladaptation mit blaufreiem Licht f6rdert die Bleichung des Sehpurpurs, w~hrend das Blau absorbierende Photopigment geschont wird (Zwei-Pigment-Itypothese der Blauverschiebung).
Zur Halbbleichung des Sehpurpurs werden in der menschlichen Netz- haut 6,8 log Troland.sec ben6tigt [22]; bei einer Einwirkungsdauer des Lichts yon 45 see entspricht dies einer retinalen Beleuchtung yon 140000 Troland. Beim Kaninchen hat die gleiche retinale Beleuchtung gr6Bere photochemische Wirksamkeit, da die Durchl/~ssigkeit dcr brechenden Medien bei 500 m~ um den Betrag 0,84/0,50, d. h. um das 1,68lathe gr61]er ist [16]. Weiterhin ist die retinale Beleuchtungssti~rke des Xaninchenauges aufgrund seiner ~efraktion yon 95 Dioptrien [1] gegeniiber dem menschlichen Auge um den Faktor (95/60) 3, d.h. um das 2,5fache h6her. Beim Kaninchen diirften bei gleicher Photosensitivi- tat des Sehpurpurs somit zur Halbbleiehung bereits 140 000/1,68.2,5 =

Adaptive Parameter der Blauverschiebung 121
33 000 Troland ausreichen. Da Troland = Nit. Pupillenfl/iche in Quadrat- millimeter ist, ergeben sich bei einer Pupillengr6$e yon 78,5 mm ~ f/it das atropinisierte Kaninehenauge bei der hier maximal verffigbaren Beleuchtnng yon 2470 Nit etwa 200000 , ,Kaninchen-Trolands" (gleieh 800000 ,Menschen-Trolands"). Bei einer Helladaptation yon 45 sec erreehnet sich hieraus zur Halbbleichung des Sehpurpurs im Kaninehen- auge eine adaptive Beleuchtung yon 420 Nit. Dieht oberhalb dieses Wertes (450 Nit) setzt die Blauversehiebung ein.
Obwohl die errechnete Bleiehung des Sehpurpurs mit dem Auftreten der Blauversehiebung gut fibereinstimmt, ist die Ein-Pigment-Hypo- these - - zumindest in der vorliegenden Form - - wenig wahrseheinlieh. So besteht eine besondere Blausensitivitgt einze]ner Ganglienzellen der Kaninchennetzhaut bereits w/ihrend Dunkeladaptation [5], wenn auch - - wie ein Vergleich zwisehen der spektralen Dunkelempfindliehkeits- kurve der b-We]le mit der Absorptionskurve des Sehpurpurs zeigt [13] - - die Zahl solcher Elemente relativ klein sein dfirfte. Bei Aufzeiehnung der Erregungsvorg/inge der zwei ersten Netzhautneurone im Elektro- retinogramm treten sie daher erst naeh Bleiehung des Sehpurpurs deutlich hervor. Da aueh der Blaumechanismus seinerseits der blei- ehenden Wirkung der adaptiven Beleuehtung unterliegt, ist zu seiner Darstellung blaufreies Adaptationslieht wirksamer als das bl/iulich- weiBe Xenonlieht, das zwisehen 450 und 492 m~ einige Emissionsbanden besitzt [ld]. M6glieherweise ist der untersehiedliehe Blauanteil ,,weiBen" Adaptationslichtes der versehiedenen Liehtquellen (Glfihlampe, Xenon- lampe) ffir die hinsichtlieh der Blausensitivit/it der Xaninchennetzhaut verschiedenartigen friiheren Beobaehtungen [4, 5] mitverantwortlieh.
Zwar 1/iBt sich das besondere Hervortreten des Blaumeehanismus unter blaufreier tIelladaptation aueh in die Ein-Pigmenthypothese ein- ordnen, da das photosensible Bleiehungsprodukt selbst aueh der blei- ehenden Wirkung weigen Adaptationslichtes unterliegen wird. Doeh spreehen versehiedene Befunde wie z.B. die ,,skotopisehe" Potential- form sowie die besondere Abh/ingigkeit der PotentialgrSBe yon der Be- liehtlmgsfrequenz bei blauvioletten I{eizen f/ir nervSse Faktoren. Form und Ablauf der elektroniseh summierten Potentiale auf Blaureiz ghnelt den Antworten reiner St/~bchennetzh/~ute [2, 7], w/~hrend die Antworten auf mittel- und langwellige Liehtreize sieh kaum yon den photopisehen Antworten gemisehter Netzh&ute mit Purkinje-Verschiebung unter- scheiden. An der Spektralsensitivit/s des heltadaptierten Kaninchens sind somit sowohl Sehpurpurelemente als aueh blauempfindliche Emp- f/~ngerstrukturen beteiligt. Zur Aussonderung der Blauelemente 1/s sich intermittierende Belichtung [9], zur Aussonderung der Sehpurpur- elemente blanfreies Adapta~ions]ieht verwenden.

122 1%. KSI~SIK:
Bisher ist nicht bekannt, inwieweit sich die blauempfindlichen Netzhautelemente des Kaninchens ~hnlich den grfinen St/ibehen der Frosehnetzhaut [3, 12] bei geeigneter Beleuchtung lichtmikroskopisch yon den Sehpurpurelementen unterscheiden.
Zusammenfassung Mit der Methode der konstanten elektrischen Antwort wurde die
spektrMe Verteilung der Augenempfindlichkeit dezerebrierter Kaninchen in Abh/~ngigkeit v o n d e r Leuchtdichte und der spektralen Zusammen- setzung der adaptiven Beleuchtung gepr/ift. In Ubereinstimmung mit bei Ableitung yore Sehnerven des Kaninchens gemaehten Beobachtungen zeigt sich elektroretinographisch bei I-Ielladaptation anstelle einer Pur- kinje-Verschiebung der Spektralempfindlichkeit zu l~ngeren Wellen- I/ingen eine Verschiebung zu kiirzeren Wellenlgngen (,,Blauverschie- bung"). Die Verschiebung erfordert eine adaptive Beleuchtung yon 1000 Nit (106.Dunkelschwelle im Elektroretiuogramm) und ist bei Ver- wendung blaufreier Adaptationslichter wesentlich starker als f/Jr weiges Xenonlicht gleicher skotopischer Wirksamkeit. Elektronische Mittelung der PotentiMe mit I-Iilfe eines Digitalrechners ergibt auf kurzwellige und 1/~ngerwellige Lichtreize verschiedene PotentiMformen. Die Ampli- tuden der als Antwort auf kurzwellige Liehtreize beobachteten Poten- time zeigen bei intermittierender geizung eine st/Lrkere Frequenzab- h/ingigkeit als bei Reizung mit mittel- und l~ngerwelligen Lichtern.
Summary The spectral performance of the electroretinal response of the rabbit
was investigated at widely different states of adaptation in deeerebrated animals.
Proceeding from dark to light adaptation the increment threshold A I (relative luminance of test light necessary to produce a small constant b-wave in the electroretinogram) was measured at different levels of adaptive illumination I between 0.0003 Nit (electroretinogram threshold for a b-wave of 8 ~V) and 2,500 Nit. For blue (473 m~) and green (546 m~) test lights the A I / I ratio was found to be the same up to 400 Nit irre- spective of the colour of the adapting light. With light adaptation of higher luminances the zJ I / I ratio was smaller for 473 m~ than for 546 m~. This was seen especially with adaptive illumination of blue-free visible light.
Instead of a Purkinje-shift to longer wavelengths during light adaptation there is, in the rabbit 's spectral sensitivity function, a shift towards shorter wavelengths. The shift is relatively small and about equal with illumination of blue light and with white light of the xenon

Adaptive Parameter der Blauverschiebung 123
arc. W i t h a d a p t a t i o n to blue-free visible l ight the blue shif t is bigger as c o m p a r e d to a d a p t a t i o n of whi te l ight of the xenon are of equal scotopie efficiency.
By using a Computer o/ Average Trans ien ts the photopic response el ici ted b y tes t l ights of med ium and long wavelengths was seen to consist of fas t r ising " p h o t o p i e " po ten t ia l s wi th sharp contours and of s lowly rising " s c o t o p i e " po ten t ia l s wi th tes t l ights of shor t wave- lengths.
W i t h i n t e r m i t t e n t l ight a lower fusion f requency is ob ta ined in t he r a b b i t ' s e lee t rore t inogram in response to blue-viole t l ight as compared to green l ight of equal luminance.
Literatur [1] ARDEN, G. B., and O. P. Gt~EAVES: The reversible alterations of the electro-
retinogram of the rabbit after occlusion of the retinal circulation. J. Physiol. (Lond.) 133, 266--274 (1956).
[2] BROWN, K.T. , and K. WATA~AB~: Rod receptor potential from the retina of the night monkey. Nature (Lond.) ]9G, 547--550 (1962).
[3] DE~TO~, E. J., and J. H. WYLLIE: Study of the photosensitive pigments in the pink and green rods of the frog. J. Physiol. (Lond.) 127, 81--89 (1955).
[4] DODT, E., K. ECttTE U. K. H. JSSSEN: Elektroretinographische Messung der Unterschiedsschwelle (increment threshold) hei Steigerung der adaptiven Beleuchtung. 63. Zusammenkunit Dtsch. Ophth. Ges. Berlin 1960, S. 319-- 322.
[5] - - , u. V. EI, E~us: Spektrale Sensitivitgt einzelner Elemente der Kaninchen- netzhaut. Pflfigers Arch. ges. Physiol. 262, 301--306 (1956).
[ 6 ] - - - Change of threshold during dark adaptation measured with orange and blue light in cats and rabbits. Experientia (Basel) 14, 313 (1960).
[ 7 ] - - , u . J. HEcK: Retinapotentiale der zapfenfreien Netzhaut des Gecko (Sphaerodactylus muralis). Pflfigers Arch. ges. Physiol. 2~9, 226--230 (1954).
[8] - - , and K. H. J~SSEN: Change of threshold during light and dark adaptation following exposure to spectral lights of equal scotopic and equal photopie efficiencies. J. opt. Soc. Amer. 51, 1269--1274 (1961).
[9 ] - - , and J. B. W~LTH~: Photopie sensitivity mediated by visual purple. Experientia (Basel) 14, 142 (1958).
[ 1 0 ] - - - Spektrale Sensitivit~t und Blutreflexion. Pflfigers Arch. ges. Physiol. 266, 187--192 (1958).
[11] - - - - Elektroretinographische Messung der Spektralsensitivit~t yon Albino- augen bei direkter und diaskleraler Beleuehtung. Pflfigers Arch. ges. Physiol. 268, 435 443 (1959).
[12] DO~ER, K. 0., and T. I~UTE~: The spectral sensitivity and photopigment of the green rods in the frog's retina. Vision Res. 2, 357~372 (1962).
[13] Ei~E~rxus, V. : I~ecovery in the dark of the rabbit 's electroretinogram. Aeta physiol, scan& 44, Suppl. 150 (1958).
[14] F ~ i ~ r s G , tt.-G., W. MtiNc~ u. M. I~IC~E~: Die Eignung der Xenonlampe als Standardlichtquelle fiir Strahlungs- und Farbmessungen. Farbe 5, 41~68 (1956).

124 R. KSIZqSIK: Adaptive Parameter der Blauverschiebung
[16] I-IrLL, R. M. : Unit responses of the rabbit lateral geniculate nucleus to mono- chromatic light on the retina. Science 135, 98--99 (1962).
[16] KI~sEY, V. E. : Spectral transmission of the eye to ultraviolet radiations. Arch. Ophthal. 39, 508--513 (1948).
[17] KR~VSn, W. : Die Anatomie des Kaninehens. Leipz:g: W. Engelmann 1884. [18] LIT~, G. It. M. vA~q: Simultane Bestimmung der elektroretinographisehen und
sensorisehen Reizschwelle. Vision Res. 6, 185--197 (1966). [19] iVA[oN~IER, M., A. ScIr~V~Z, and P. JORDAIq: Spectral sensitivity of retina and
visual cortex in the rabbit. Vision Res. 2, 189--200 (1962). [20] NvBOE~, J. F. W. : Heterochromous photometry in the rabbit. Acta physiol.
pharmacol, neerl. ]8, 209 (1965). [21] Owo, J . : Anatomic et physiologic de l'ceil clans la serie animale. Paris: Alcan
1927. [22] RVSHTO~, W. A. H. : Dark adaptation and the regeneration of rhodopsin.
J . Physiol. (Lond.) 156, 166--178 (1961). [23] VATT~.~, 0., Tm KO~]~I~ u. M. MO~NIn~: Die spektrale Sensitivit~t der Retina
und des optischen Cortex beim Kaninchen. Vision Res. 4, 329--343 (1964). [24] WIRT~, A. : Electroretinographie eva]uation of the scotopic visibility function
in cats and albino rabbits. Acta physiol, scand. 29, 22--30 (1953).
Professor Dr. E. DODT W. G. Kerckhoff-Institut 6350 Bad Nauheim
Recommended

















