
1
WWU-MünsterProf. Jörg Kudla
Aufbau-Modul:
Zellbiologie, Physiologie und Genetik
Vorlesung:
Signaltransduktion durch sekundäre Botenstoffe II
Erzeugung spezifischer Ca2+-Signale in Tieren und Pflanzen 1. Einstrom
In Tieren:• Ausgelöst durch Ladungsänderung oder
Ligandenbindung (extra- oder intrazellulär z.B.IP3)
• Kanäle: CaV, TRP, CRAC, IP3R
• RTK: receptor tyrosine kinase
• GPCR: G protein-coupled receptor
• RyR: ryanodine receptors

2
Welche Kanäle ermöglichen Ca2+-Signaturen in Pflanzen?
Welche Kanäle ermöglichen Ca2+-Signaturen in Pflanzen?
3
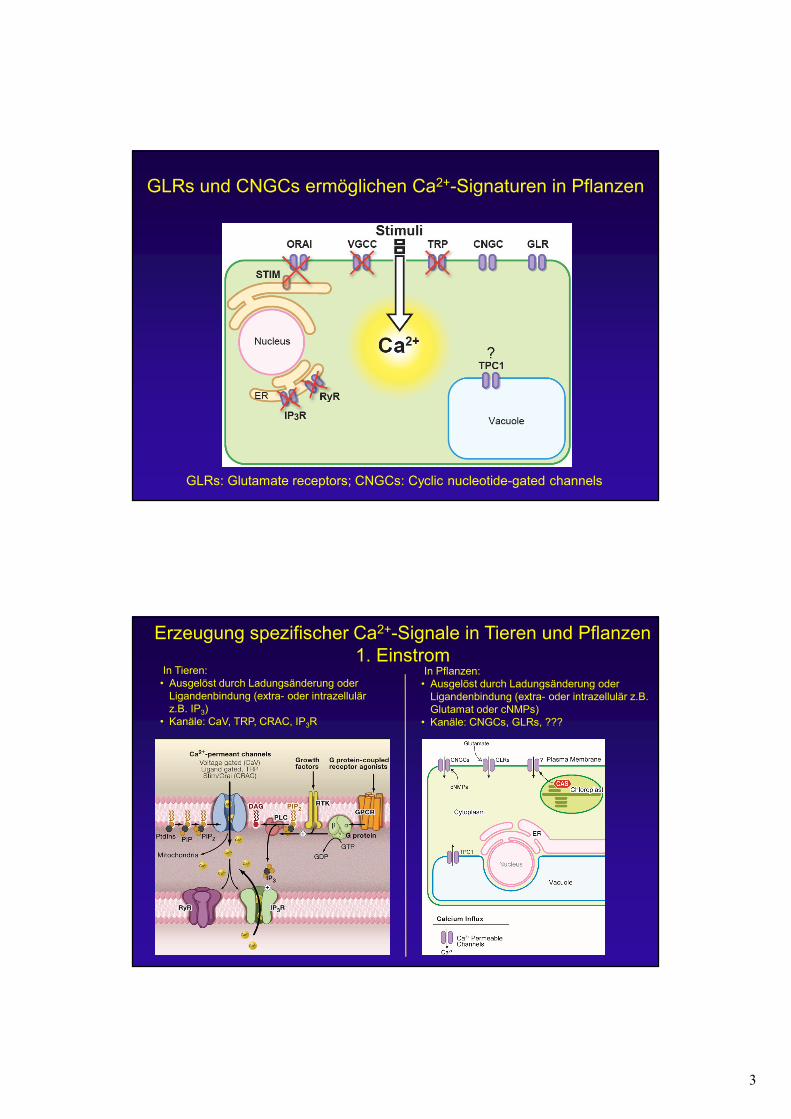
GLRs und CNGCs ermöglichen Ca2+-Signaturen in Pflanzen
GLRs: Glutamate receptors; CNGCs: Cyclic nucleotide-gated channels
In Pflanzen:• Ausgelöst durch Ladungsänderung oder
Ligandenbindung (extra- oder intrazellulär z.B. Glutamat oder cNMPs)
• Kanäle: CNGCs, GLRs, ???
Erzeugung spezifischer Ca2+-Signale in Tieren und Pflanzen 1. Einstrom
In Tieren:• Ausgelöst durch Ladungsänderung oder
Ligandenbindung (extra- oder intrazellulär z.B. IP3)
• Kanäle: CaV, TRP, CRAC, IP3R

4
In Tieren• Hauptspeicher im ER und extrazellulär• [Ca2+]cyt um 100nM • PMCA und SERCA hauptsächlich
verantwortlich für Ausstrom
• NCKX: Na+/Ca2+-K+ exchanger
• PMCA: plasma membrane Ca2+
ATPase
• NCX: Na+/Ca2+ exchanger
• SERCA: smooth endoplasmicreticular Ca2+ ATPase
• PLC: phospholipase C
Erzeugung spezifischer Ca2+-Signale in Tieren und Pflanzen 2. Ausstrom
In Pflanzen• Fast alle Kompartimente dienen als Ca2+-
Speicher • Aktiver Ausstrom durch CAX und ACA Proteine
Erzeugung spezifischer Ca2+-Signale in Tieren und Pflanzen 2. Ausstrom
In Tieren• Hauptspeicher im ER und extrazellulär• [Ca2+]cyt um 100nM • PMCA und SERCA hauptsächlich
verantwortlich für Ausstrom
5
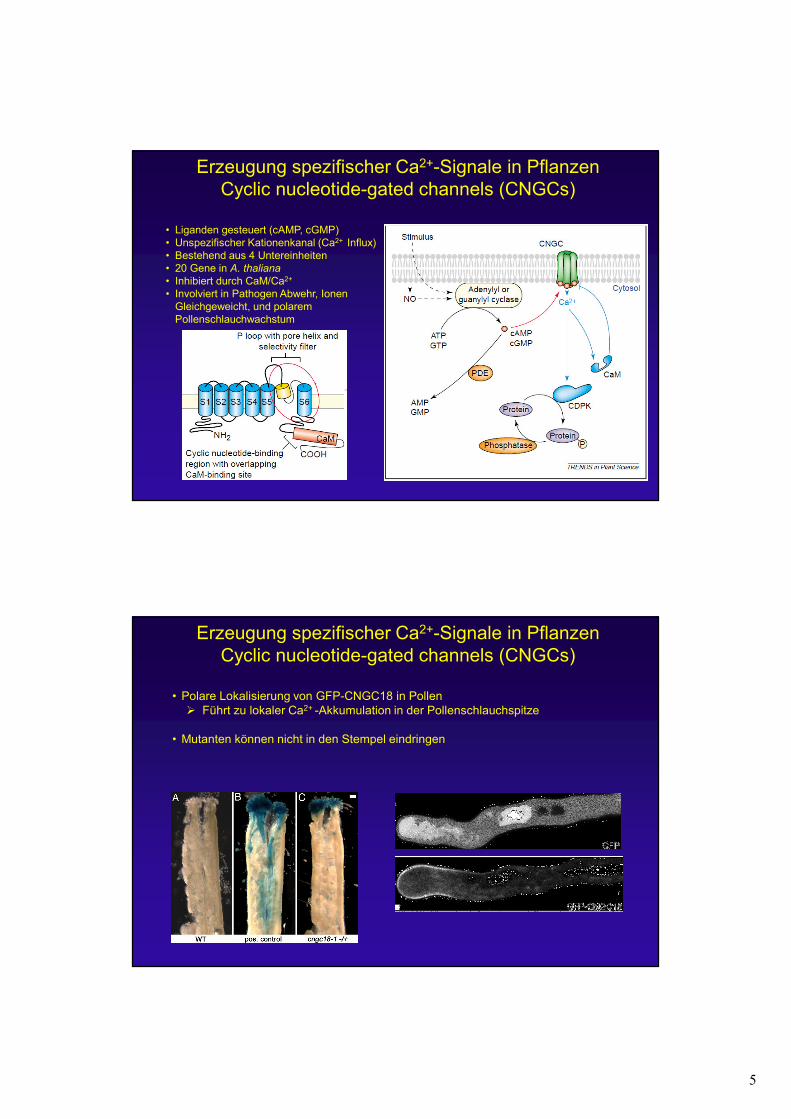
Erzeugung spezifischer Ca2+-Signale in PflanzenCyclic nucleotide-gated channels (CNGCs)
• Liganden gesteuert (cAMP, cGMP)• Unspezifischer Kationenkanal (Ca2+ Influx)• Bestehend aus 4 Untereinheiten• 20 Gene in A. thaliana
• Inhibiert durch CaM/Ca2+
• Involviert in Pathogen Abwehr, Ionen Gleichgeweicht, und polarem Pollenschlauchwachstum
• Polare Lokalisierung von GFP-CNGC18 in Pollen� Führt zu lokaler Ca2+ -Akkumulation in der Pollenschlauchspitze
• Mutanten können nicht in den Stempel eindringen
Erzeugung spezifischer Ca2+-Signale in PflanzenCyclic nucleotide-gated channels (CNGCs)

6
Erzeugung spezifischer Ca2+-Signale in PflanzenGlutamate receptors (GLRs)
• Liganden gesteuert (Glutamat, Glycin, andere AS)
• Unspezifischer Kationenkanal (Ca2+ Influx)
• 20 Gene in A. thaliana
• Involviert in Ca2+-Versorgung, Kältestress, ABA-Biosynthese, Wurzelentwicklung, Lichmorphogenese
LAOBP: lysine/arginine/ornithine-binding protein domainLIVBP: leucine/isoleucine/valine-binding protein domain
• GLRs sind u.a. am Pollenschlauchwachstum beteiligt
• Die Regulation erfolgt durch D-Serin aus dem Pistill
• Ca2+ akkumuliert in der Pollenschlauchspitze (stimuliert durch D-Ser)
WT glr1.2-1
Pollenschlauchdeformation in „knock-down“ Mutanten
Erzeugung spezifischer Ca2+-Signale in PflanzenGlutamate receptors (GLRs)
Ca2+-Einstrom wird durch D-Serin lokal induziert � Pollenschlauch wächst schneller

7
Erzeugung spezifischer Ca2+-Signale in PflanzenCation-proton exchanger (CAX)
• Protonengetriebener Ca2+ (Kationen) –Antiporter
• Geringe Affinität – hohe Kapazität
• 6 Gene in A. thaliana
• Verwandt mit „cation Ca2+ exchanger“
(CCX; 5 sequences)
• Vor allem an der Vakuole Lokalisiert (PM Aktivität allerdings auch berichtet)
• Involviert in Ca2+ Kompartimentierung, Zellwandmodifikation, Zellwachstum, Stomataregulierung
• CAX1 wird für die Ca2+-Kompartimentierung in die Vakuole benötigt
• Ca2+-Akkumulation im Apoplasten der cax1/cax3 knock-out Mutante
� Bewirkt limitiertes Wachstum und Stomataschluss
Erzeugung spezifischer Ca2+-Signale in PflanzenCation-proton exchanger (CAX)

8
Erzeugung spezifischer Ca2+-Signale in PflanzenAutoinhibitory Ca2+-ATPases (ACAs)
• ACAs sind PIIB-type ATPasen; Ca2+ wird gegen den Gradienten unter ATP verbrauch gepumpt
• Hohe Affinität – geringe Kapazität
• 10 Gene in A. thaliana
• Verwandt mit pflanzlichen ER Ca2+-ATPasen(ECAs; 4 Gene in A. thaliana) und tierischen PMCAs und SERCAs
• Lokalisieren an den meisten zellulären Membranen
• Reguliert durch Phosphorylierung und CaM-Bindung
• Involviert in Entwicklungsprozessen wie Pollenschlauchwachstum und Blütenentwicklung
ACA11
ACA4
ACA1
ACA2ACA7
ACA12ACA13
ACA8ACA9
ACA10
PM ER Tonoplast Chl Envelope UnKnown
cyt
out
D-PATP
Phenotypes: aca9: reduced Pollen tube growth and fertility; aca10 (cif1): Inflorescence and
rosette phenotypes
Erzeugung spezifischer Ca2+-Signale in PflanzenAutoinhibitory Ca2+-ATPases (ACAs)
9
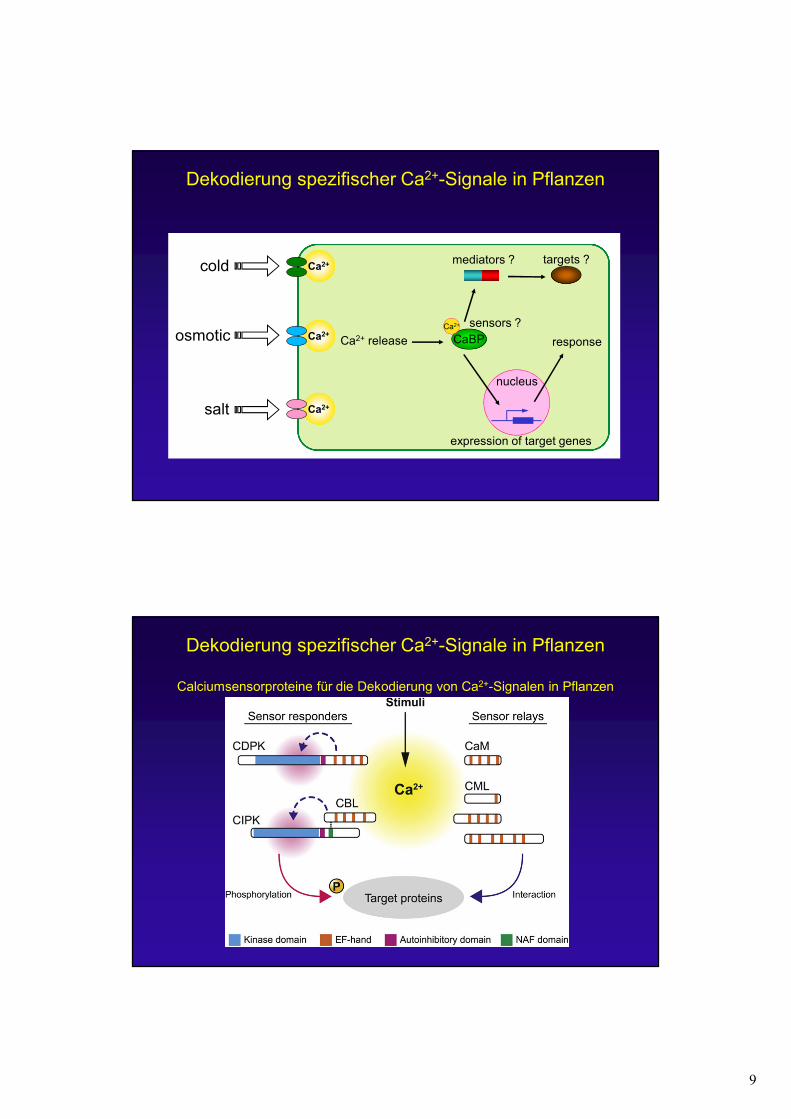
Dekodierung spezifischer Ca2+-Signale in Pflanzen
osmotic
nucleus
sensors ?
mediators ?
expression of target genes
response
targets ?
CaBP
salt
cold
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+ release
Calciumsensorproteine für die Dekodierung von Ca2+-Signalen in Pflanzen
Dekodierung spezifischer Ca2+-Signale in Pflanzen
10
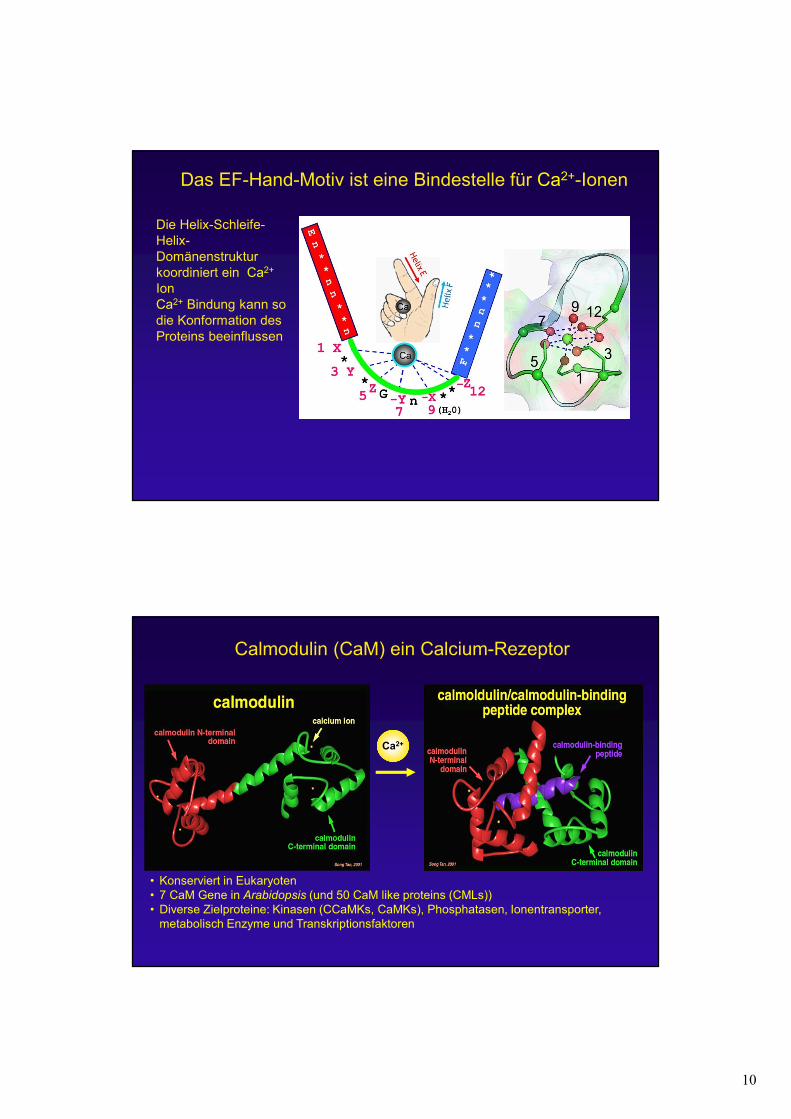
Das EF-Hand-Motiv ist eine Bindestelle für Ca2+-Ionen
Die Helix-Schleife-Helix-Domänenstruktur koordiniert ein Ca2+
IonCa2+ Bindung kann so die Konformation des Proteins beeinflussen
Calmodulin (CaM) ein Calcium-Rezeptor
Ca2+
• Konserviert in Eukaryoten• 7 CaM Gene in Arabidopsis (und 50 CaM like proteins (CMLs))• Diverse Zielproteine: Kinasen (CCaMKs, CaMKs), Phosphatasen, Ionentransporter,
metabolisch Enzyme und Transkriptionsfaktoren
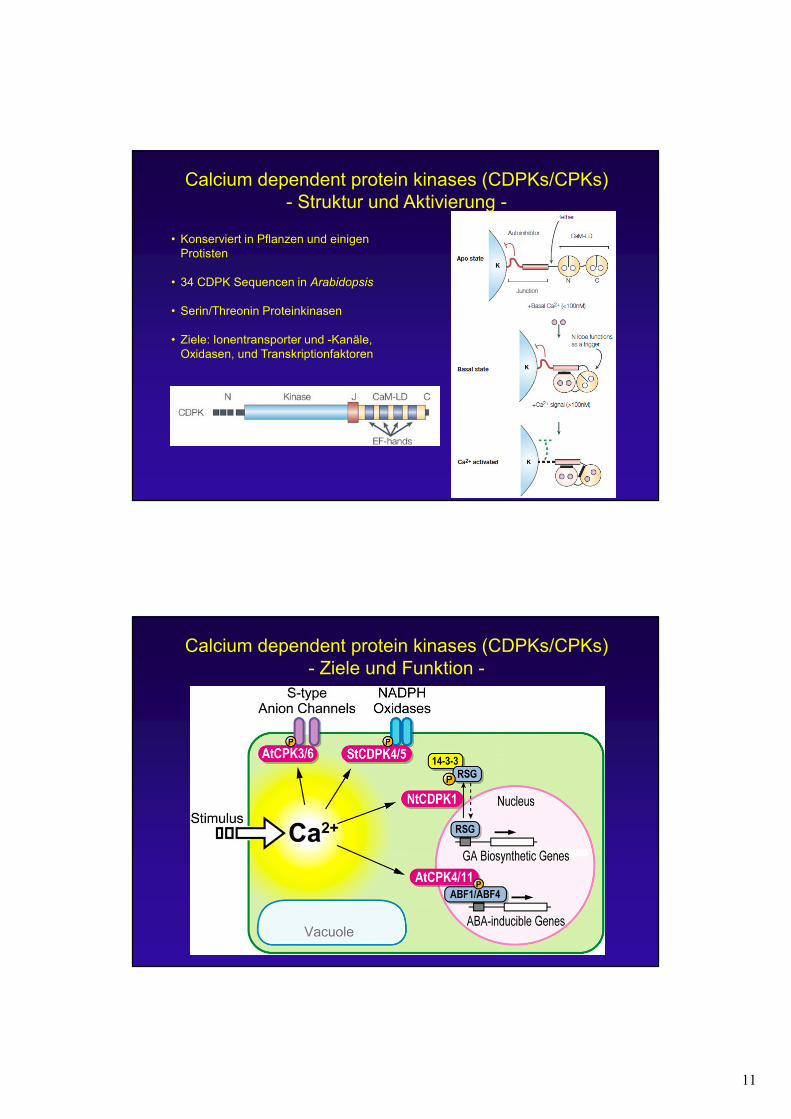
11
Calcium dependent protein kinases (CDPKs/CPKs)- Struktur und Aktivierung -
• Konserviert in Pflanzen und einigen Protisten
• 34 CDPK Sequencen in Arabidopsis
• Serin/Threonin Proteinkinasen
• Ziele: Ionentransporter und -Kanäle, Oxidasen, und Transkriptionfaktoren
Calcium dependent protein kinases (CDPKs/CPKs)- Ziele und Funktion -
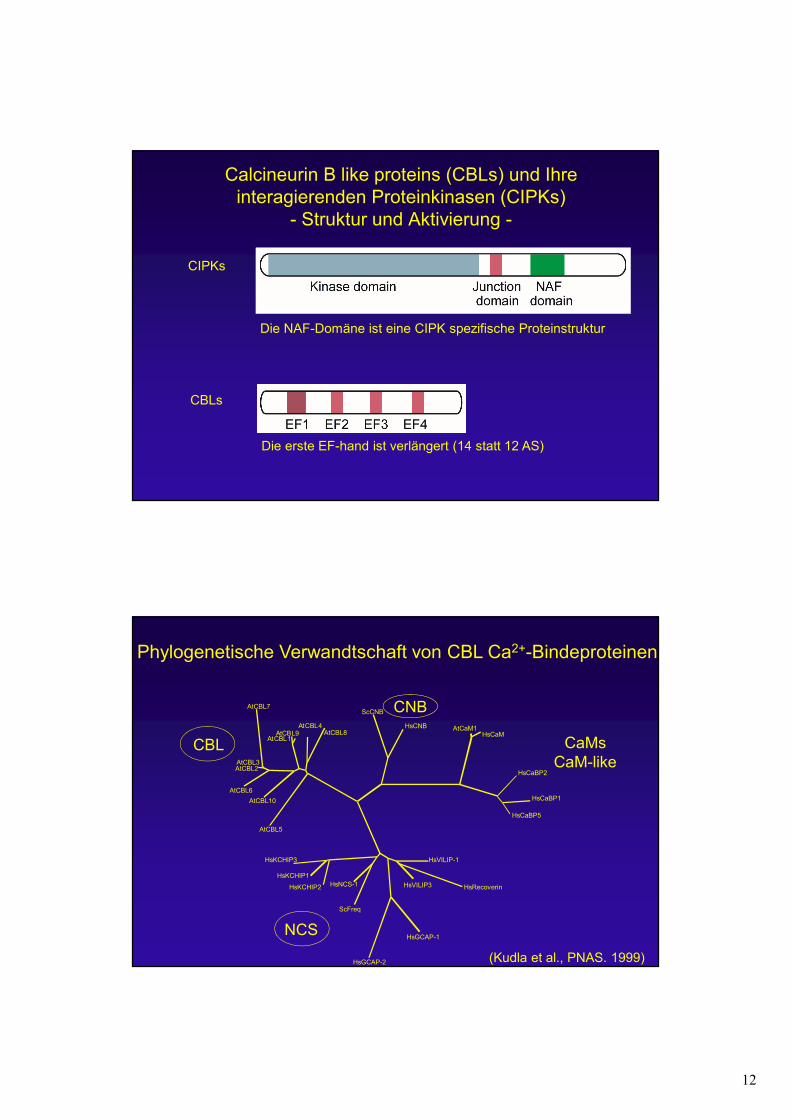
12
Calcineurin B like proteins (CBLs) und Ihre interagierenden Proteinkinasen (CIPKs)
- Struktur und Aktivierung -
CIPKs
CBLs
Die NAF-Domäne ist eine CIPK spezifische Proteinstruktur
Die erste EF-hand ist verlängert (14 statt 12 AS)
Phylogenetische Verwandtschaft von CBL Ca2+-Bindeproteinen
CBL
NCS
CNB
CaMsCaM-like
HsVILIP-1
HsRecoverinHsVILIP3
HsGCAP-1
HsGCAP-2
ScFreq
HsNCS-1HsKCHIP2
HsKCHIP1
HsKCHIP3
AtCBL5
AtCBL10
AtCBL6
AtCBL2AtCBL3
AtCBL7
AtCBL1AtCBL9
AtCBL4AtCBL8
ScCNB
HsCNB AtCaM1HsCaM
HsCaBP2
HsCaBP1
HsCaBP5
(Kudla et al., PNAS. 1999)
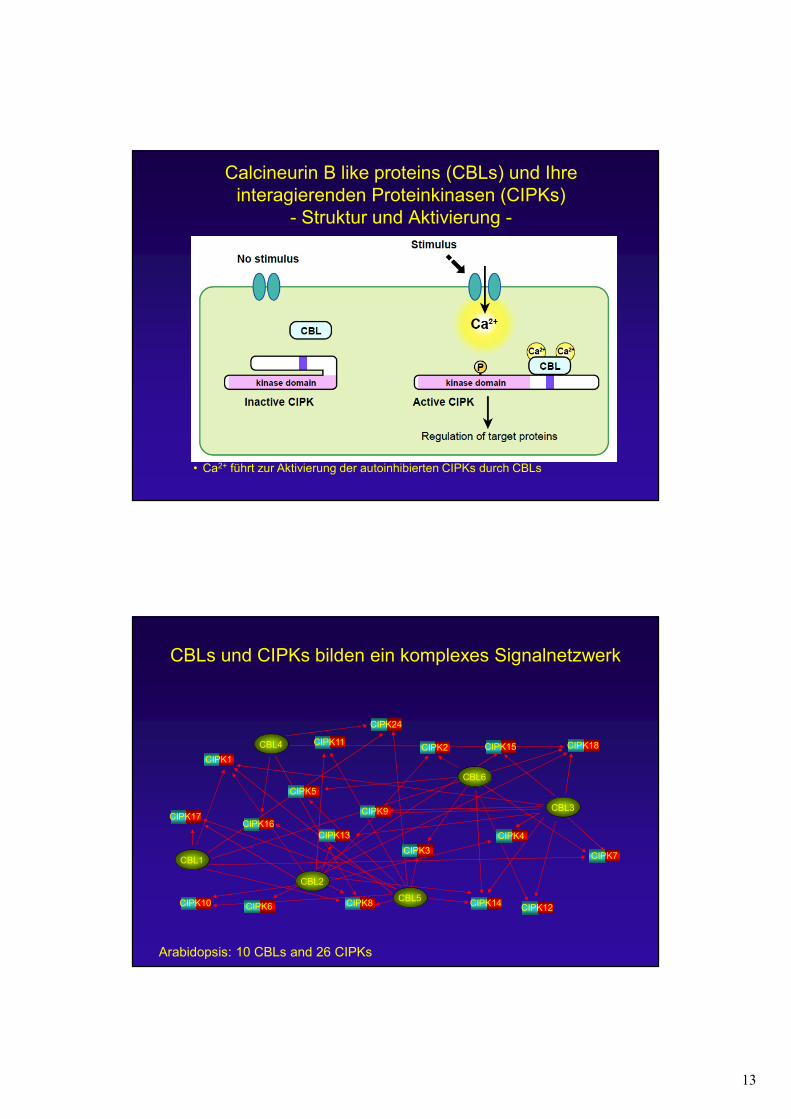
13
• Ca2+ führt zur Aktivierung der autoinhibierten CIPKs durch CBLs
Calcineurin B like proteins (CBLs) und Ihre interagierenden Proteinkinasen (CIPKs)
- Struktur und Aktivierung -
CBLs und CIPKs bilden ein komplexes Signalnetzwerk
CBL2
CBL1
CBL3
CBL4
CBL5
CBL6
CIPK1CIPK2
CIPK3
CIPK4
CIPK5
CIPK6
CIPK7
CIPK8
CIPK9
CIPK10
CIPK11
CIPK12
CIPK13
CIPK14
CIPK15 CIPK18
CIPK17
CIPK24
CIPK16
Arabidopsis: 10 CBLs and 26 CIPKs

14
Untersuchung eines komplexen Ca2+-Signalnetzwerks
Funktionale Prinzipien des CBL-CIPK-Netwerks(Mechanismen zur Generierung von Spezifität)• Unterschiedliche Ca2+-Affinität der CBLs• Spezifität und Gleichgewicht der CBL-CIPK Komplexformierung• Unterschiedliche Ca2+-Abhängigkeit der CBL-CIPK Komplexformierung• Lokalisierung der CBLs • Spezifische Lokalisierung der CIPKs durch CBLs
Funktion und Ziele spezifischer CBL-CIPK Komplexe
Untersuchung eines komplexen Ca2+-Signalnetzwerks
Funktionale Prinzipien des CBL-CIPK-Netwerks(Mechanismen zur Generierung von Spezifität)• Unterschiedliche Ca2+-Affinität der CBLs• Spezifität und Gleichgewicht der CBL-CIPK Komplexformierung• Unterschiedliche Ca2+-Abhängigkeit der CBL-CIPK Komplexformierung• Lokalisierung der CBLs • Spezifische Lokalisierung der CIPKs durch CBLs
Funktion und Ziele spezifischer CBL-CIPK Komplexe
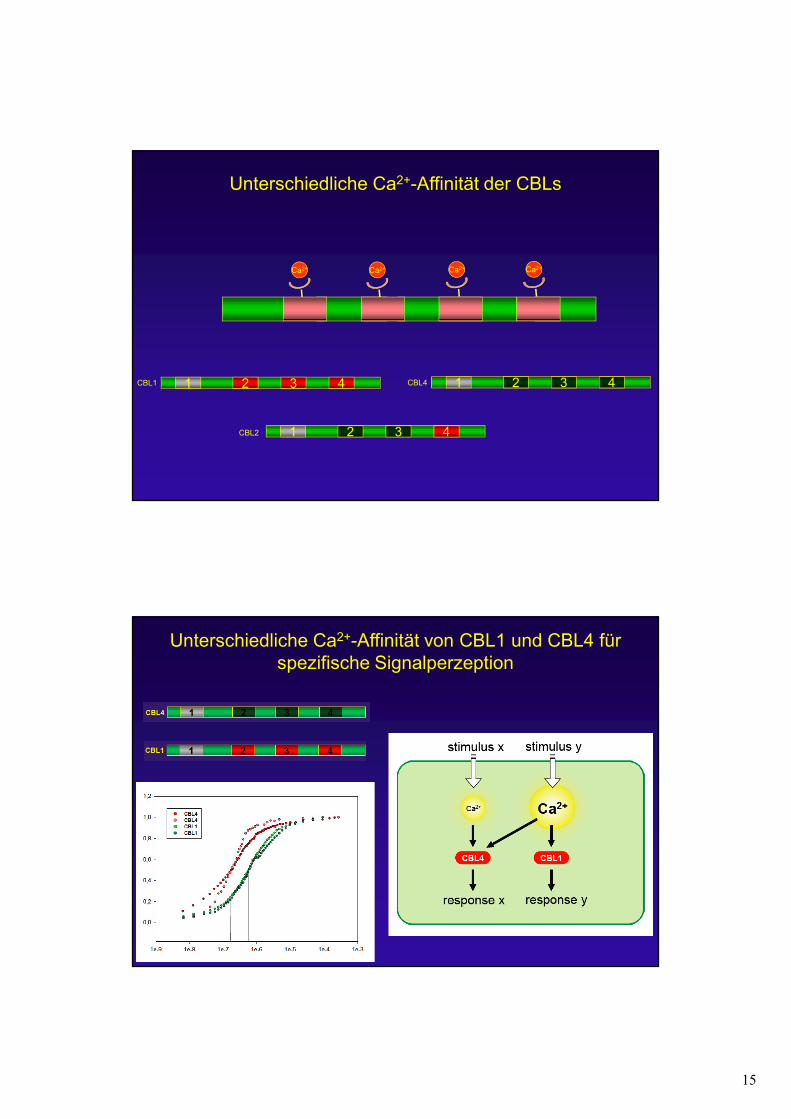
15
Ca2+ Ca2+ Ca2+
Unterschiedliche Ca2+-Affinität der CBLs
Ca2+
CBL1 1 2 3 4 CBL4 1 2 3 4
CBL2 1 2 3 4
Unterschiedliche Ca2+-Affinität von CBL1 und CBL4 für spezifische Signalperzeption

16
Untersuchung eines komplexen Ca2+-Signalnetzwerks
Funktionale Prinzipien des CBL-CIPK-Netwerks(Mechanismen zur Generierung von Spezifität)• Unterschiedliche Ca2+-Affinität der CBLs• Spezifität und Gleichgewicht der CBL-CIPK Komplexformierung• Unterschiedliche Ca2+-Abhängigkeit der CBL-CIPK Komplexformierung• Lokalisierung der CBLs • Spezifische Lokalisierung der CIPKs durch CBLs
Funktion und Ziele spezifischer CBL-CIPK Komplexe
Spezifität der CBL-CIPK Komplexformierung
LT AtCIPK1 AtCIPK2 AtCIPK4 AtCIPK6
AtCIPK14AtCIPK13AtCIPK12AtCIPK11AtCIPK9
AtCBL4
AtCBL1
AtCBL6AtCBL5
AtCBL3
AtCBL2
(Albrecht et al., EMBO J., 2001)
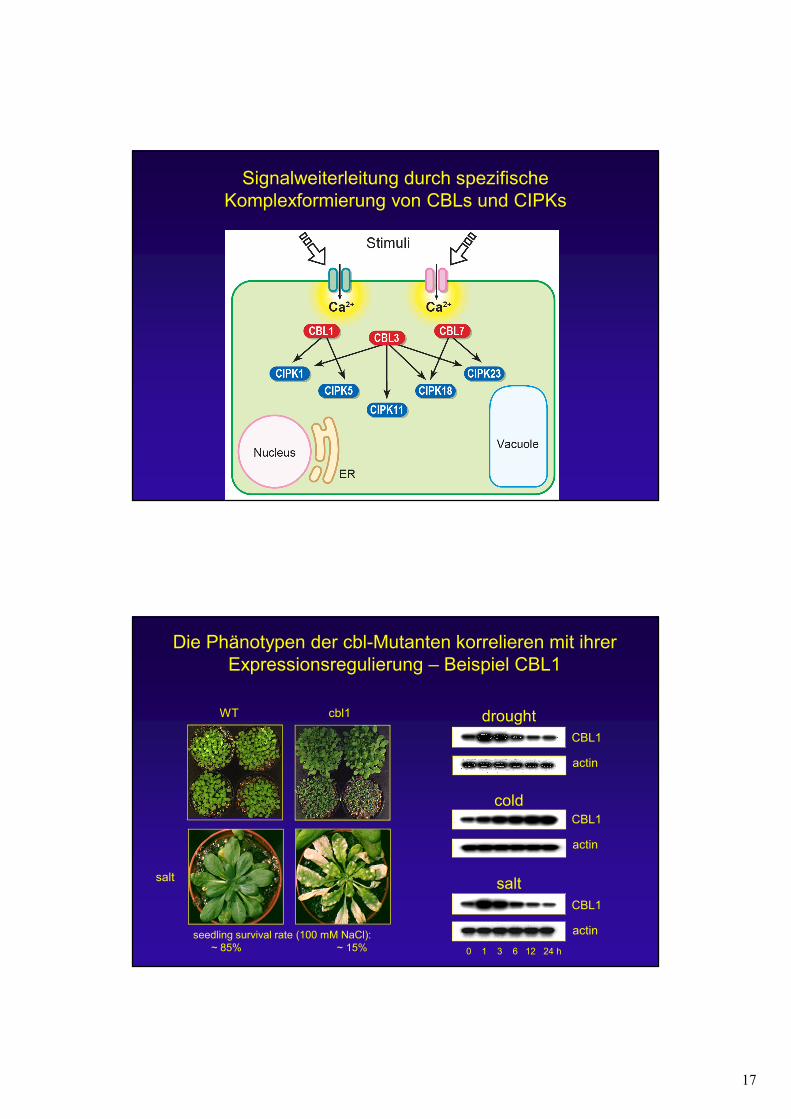
17
Signalweiterleitung durch spezifische Komplexformierung von CBLs und CIPKs
Die Phänotypen der cbl-Mutanten korrelieren mit ihrer Expressionsregulierung – Beispiel CBL1
WT
salt
cbl1
seedling survival rate (100 mM NaCl):~ 85% ~ 15%
drought
salt
cold
actin
CBL1
actin
CBL1
actin
CBL1
0 1 3 6 12 24 h

18
Hochregulierung von Signalkomponenten verschiebt die Balance der Komplexformierung und
Informationsweiterleitung
Untersuchung eines komplexen Ca2+-Signalnetzwerks
Funktionale Prinzipien des CBL-CIPK-Netwerks(Mechanismen zur Generierung von Spezifität)• Unterschiedliche Ca2+-Affinität der CBLs• Spezifität und Gleichgewicht der CBL-CIPK Komplexformierung• Unterschiedliche Ca2+-Abhängigkeit der CBL-CIPK Komplexformierung• Lokalisierung der CBLs • Spezifische Lokalisierung der CIPKs durch CBLs
Funktion und Ziele spezifischer CBL-CIPK Komplexe
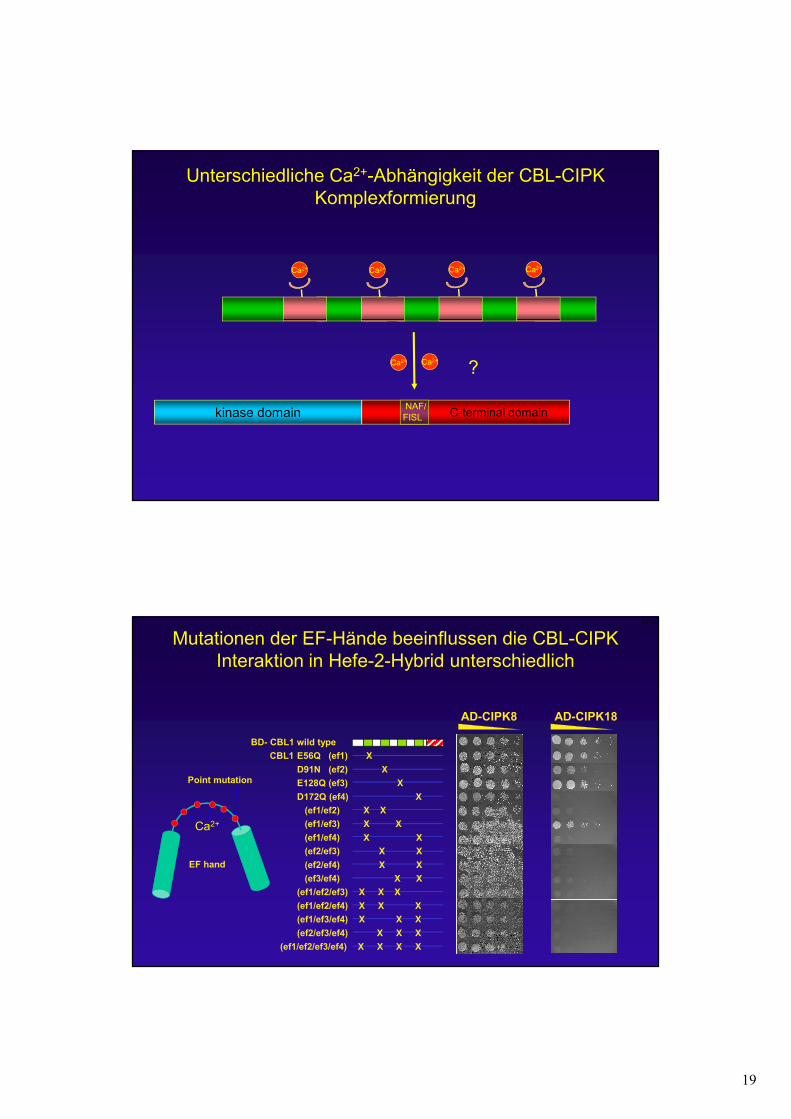
19
Ca2+ Ca2+ Ca2+
Unterschiedliche Ca2+-Abhängigkeit der CBL-CIPK Komplexformierung
kinase domain C-terminal domainNAF/FISL
Ca2+
Ca2+ Ca2+
?
Mutationen der EF-Hände beeinflussen die CBL-CIPK Interaktion in Hefe-2-Hybrid unterschiedlich
AD-CIPK8
Ca2+
Point mutation
EF hand
AD-CIPK18
CBL1 E56Q (ef1) X
D91N (ef2) X
BD- CBL1 wild type
E128Q (ef3) X
D172Q (ef4) X
(ef1/ef2) X X
(ef1/ef3) X X
(ef1/ef4) X X
(ef2/ef3) X X
(ef2/ef4) X X
(ef3/ef4) X X
(ef1/ef2/ef3) X X X
(ef1/ef2/ef4) X X X
(ef1/ef3/ef4) X X X
(ef2/ef3/ef4) X X X
(ef1/ef2/ef3/ef4) X X X X
EF mut

20
Unterschiedliche Ca2+-Abhängigkeit der CBL-CIPK Komplexformierung als Weiche in der Informationsverarbeitung
AD-CIPK1
AD-CIPK7
AD-CIPK8
AD-CIPK11
AD-CIPK17
AD-CIPK18
AD
AD-CIPK24
CBL1 WT CBL1 EF4
CIPK8
CIPK24
CIPK11
CIPK1
CIPK7/17
CIPK18
Unabhängig
Abhängig
Untersuchung eines komplexen Ca2+-Signalnetzwerks
Funktionale Prinzipien des CBL-CIPK-Netwerks(Mechanismen zur Generierung von Spezifität)• Unterschiedliche Ca2+-Affinität der CBLs• Spezifität und Gleichgewicht der CBL-CIPK Komplexformierung• Unterschiedliche Ca2+-Abhängigkeit der CBL-CIPK Komplexformierung• Lokalisierung der CBLs • Spezifische Lokalisierung der CIPKs durch CBLs
Funktion und Ziele spezifischer CBL-CIPK Komplexe
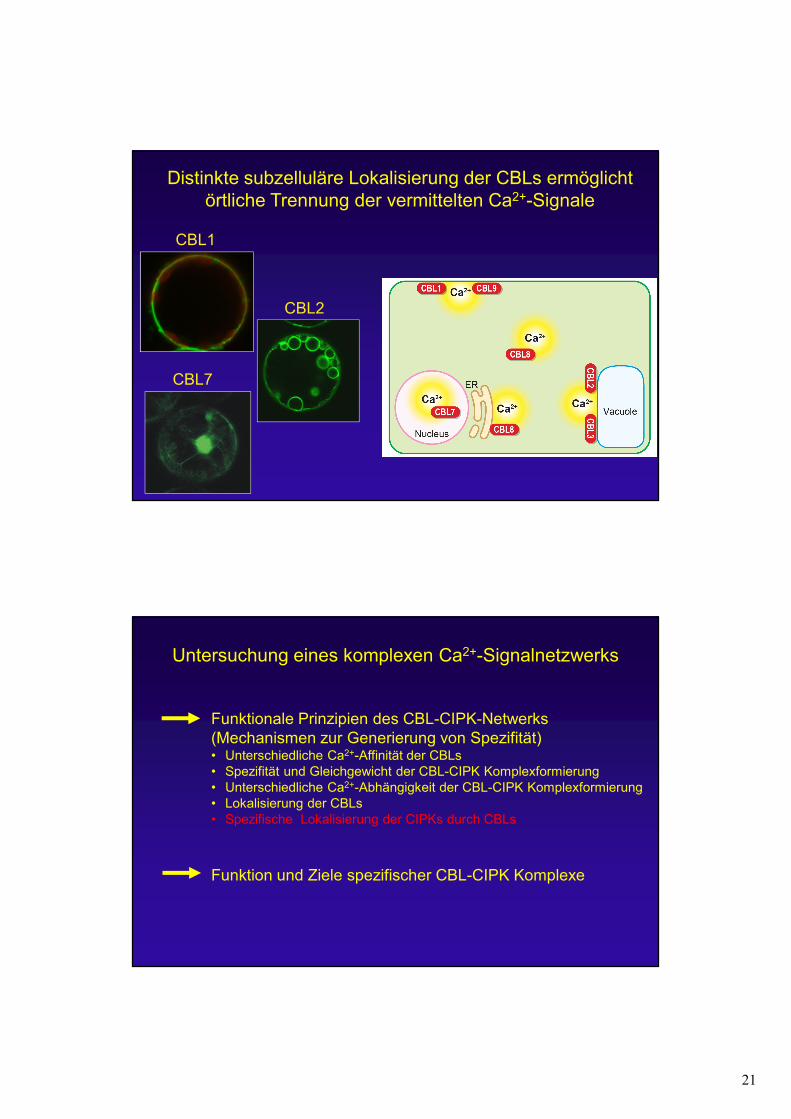
21
Distinkte subzelluläre Lokalisierung der CBLs ermöglicht örtliche Trennung der vermittelten Ca2+-Signale
CBL1
CBL2
CBL7
Untersuchung eines komplexen Ca2+-Signalnetzwerks
Funktionale Prinzipien des CBL-CIPK-Netwerks(Mechanismen zur Generierung von Spezifität)• Unterschiedliche Ca2+-Affinität der CBLs• Spezifität und Gleichgewicht der CBL-CIPK Komplexformierung• Unterschiedliche Ca2+-Abhängigkeit der CBL-CIPK Komplexformierung• Lokalisierung der CBLs • Spezifische Lokalisierung der CIPKs durch CBLs
Funktion und Ziele spezifischer CBL-CIPK Komplexe

22
Die Art der Lipidmodifikation der CBL-Proteine bestimmt die subzelluläre Lokalisierung der CBL/CIPK Komplexe
(Batistic et al., Plant Cell, 2008 &Batistic et al., Cell Research, 2012)
CIPK12mVenus +
mCherry
CIPK12mVenus +
CBL3mCherry
Alternative Komplexbildung von CIPK1 mit CBL1 oder CBL2 als Mechanismus der Dekodierung eines
Ca2+ Signals

23
Mechanisms that govern information processing in the CBL/CIPK network
(Kudla et al., Plant Cell, 2010)
Untersuchung eines komplexen Ca2+-Signalnetzwerks
Funktionale Prinzipien des CBL-CIPK-Netwerks(Mechanismen zur Generierung von Spezifität)• Unterschiedliche Ca2+-Affinität der CBLs• Spezifität und Gleichgewicht der CBL-CIPK Komplexformierung• Unterschiedliche Ca2+-Abhängigkeit der CBL-CIPK Komplexformierung• Lokalisierung der CBLs • Spezifische Lokalisierung der CIPKs durch CBLs
Funktion und Ziele spezifischer CBL-CIPK Komplexe
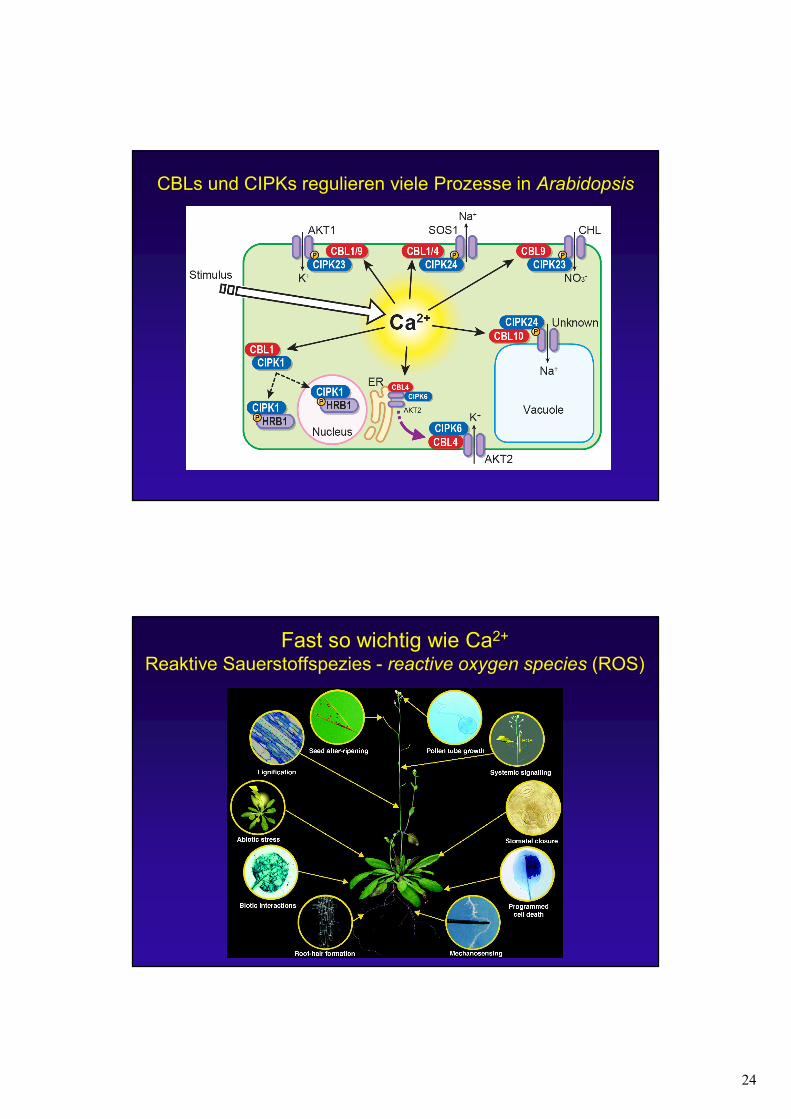
24
CBLs und CIPKs regulieren viele Prozesse in Arabidopsis
Fast so wichtig wie Ca2+
Reaktive Sauerstoffspezies - reactive oxygen species (ROS)

25
Eigenschaften von ROS
• ROS entstehen als Nebenprodukt der Photosynthese und Atmung
• Zusätzlich können ROS auch als Signalmoleküle synthetisiert werden
• ROS-Intermediate können andere Moleküle (Proteine, DNA, Lipide) oxidieren
und so schädigen
• Daher evolvierten effiziente ROS Entgiftungsmechanismen
�ROS-Level werden streng reguliert und sind im Ruhezustand sehr gering
�Zielgerichtete ROS-Produktion führt so zu lokalen Konzentrationsanstiegen
�ROS-Entgiftungsmechanismen beenden die Signale schnell
Warum sind ROS als Signalmoleküle geeignet?
Was sind reaktive Sauerstoffspezies?
3O2
1O2
•O2-
•OH
H2O2
H2O
• Triplettsauerstoff, Luftsauerstoff, reaktionsträge (Reaktivität begrenzt durch Spinpaarung und Reorientierung)
• Singulettsauerstoff; Anregungszustand durch Spinumkehr, sehr reaktiv
• Superoxidradikal; sehr reaktiv *
• Hydroxylradikal; hoch reaktiv und destruktiv *
• Wasserstoffperoxid; weniger reaktiv *
• Wasser, sehr reaktionsträge
* intermediäre Redoxzustände zwischen O2 und H2O
Reduktion(+ e-)

26
a) Energietransfer führt zu reaktivem Singulettsauerstoffb) Fortlaufender Elektronentransfer führt zu Formierung unterschiedlicher ROS
Superoxidradikal: Spontane oder katalysierte Transformation zu Peroxid
Wasserstoffperoxid: relativ stabil, kann Membranen durchdringen - passiv oder durch Aquaporine; kann aktiv abgebaut werden
Hydroxylradikal: sehr giftig – Lebenszeit im Bereich von Nanosekunden; daher nicht abbaubar
Fortlaufende Reduktion führt zu unterschiedlichen ROS
a)b) b) b) b)
Wichtige Spezies:Superoxid und Wasserstoffperoxid
Superoxid (•O2-) ist das primäre Produkt der Reduktion von
Singulett Sauerstoff, O2.
Es ist sehr unstabil und reagiert rasch zum stabileren Wasserstoffperoxid, H2O2 weiter (spontan oder enzymatisch durch die Superoxid-Dismutase katalysiert):
2 •O2- + 2 H+ → H2O2 + O2
Wasserstoffperoxid (H2O2 ) ist relativ stabil. Es kann Membranen durchdringen - entweder direkt durch die Membran oder durch Aquaporine. Wasserstoffperoxid kann aktiv abgebaut werden.

27
Entstehung von ROS in PflanzenzellenAls unerwünschte Nebenprodukte:
Chloroplast:
Direkter Elektronentransfer zu O2 am PSI (Mehler Reaktion)
Peroxisom:H2O2 Entstehung während der Glycolat Oxidation
Mitochondrium:
Direkter Elektronentransfer zu O2 in der Atmungskette
Gezielte Produktion:
Apoplast:
Zellwandperoxidasen erzeugen ROS für die Ligninsynthese
Plasmamembran:
NADPH Oxidasen (RBOH) produzieren ROS als Signalmoleküle
Oxidation von Zielmolekülendurch ROS
Beispiel: Oxidation von Lipiden durch Hydroxylradikale
Häufige Ziele: 1. C=C Dopperlbindungen in Biomolekülen:
� Cys, Met, His, Trp (Proteinschäden)� Guanosin (DNA Schäden)� Lipide (Membranschäden)
2. Oxidation von Thiolgruppen oder Metallgruppen in Proteinen
� Proteinschäden
ROS können wichtige Makromoleküle oxidieren
und somit deren Funktion zerstören!
Cytotoxizität von ROS

28
ROS Abbau in PflanzenzellenDer Glutathion-Ascorbat Zyklus
• H2O2 kann durch die Oxidation von Glutathione und Ascorbat abgebaut werden• Hierfür müssen die Moleküle in reduzierter Form vorliegen
GSH: Glutathion (reduziert) GSSG: Glutathion (oxidiert)ASC: Ascorbat MDA: MonodehydroascorbateDHA: Dehydroascorbat
GSSG + NADPH
GlutathionReduktase
GSH + NADP+
DHA + GSH
DHAreductase
Asc + GSSG
ROS Abbau in PflanzenzellenDer Glutathion-Ascorbat Zyklus
Um GSH und ASC zu regenerieren, müssen die Zellen mit NADPH “zahlen”
GSH: Glutathion (reduziert) GSSG: Glutathion (oxidiert)ASC: Ascorbat MDA: MonodehydroascorbatDHA: Dehydroascorbat NADPH: Nicotinamidadenindinukleotidphosphat

29
ROS Abbau in Pflanzenzellen Enzymatische Entgiftung durch Katalasen
H2O2
KatalaseH2O + ½ O2
• Katalase entgiftet H2O2 “kostenlos”, sie benötigt kein NADPH• Katalasen sind hauptsächlich in Peroxisomen lokalisiert
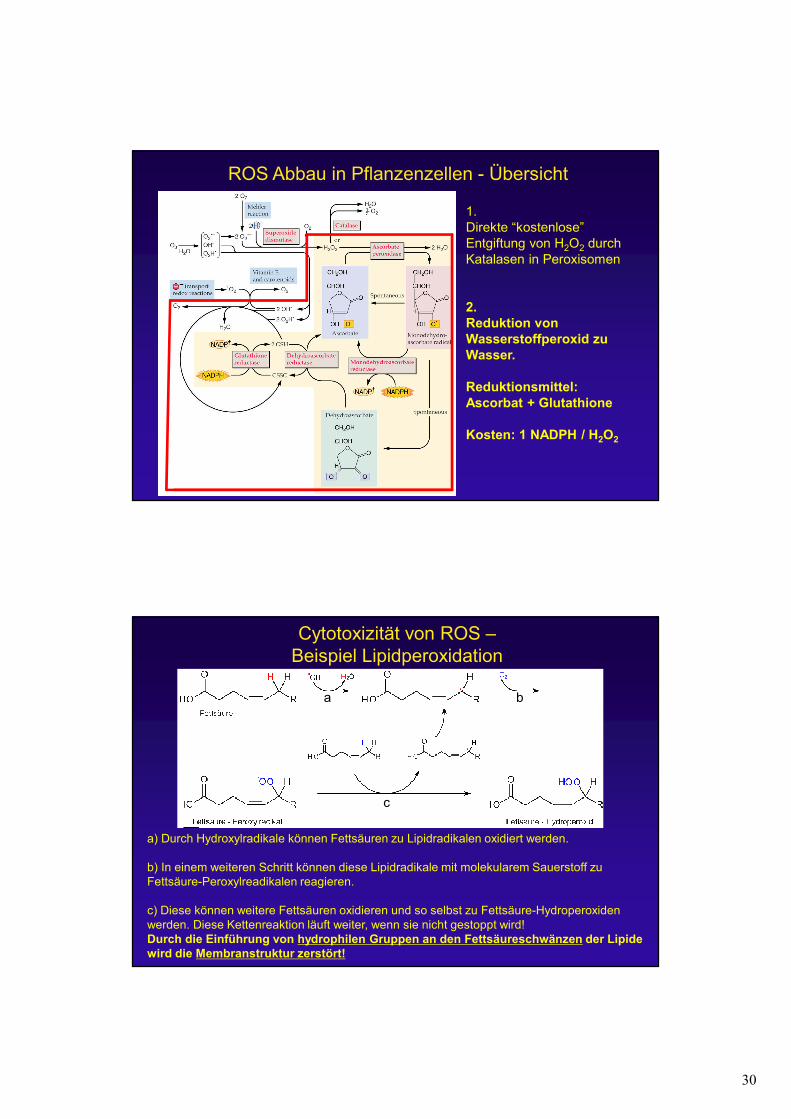
ROS Abbau in Pflanzenzellen - Übersicht
1.Direkte “kostenlose”
Entgiftung von H2O2 durch
Katalasen in Peroxisomen
30
1.Direkte “kostenlose” Entgiftung von H2O2 durch Katalasen in Peroxisomen
2.
Reduktion von
Wasserstoffperoxid zu
Wasser.
Reduktionsmittel:
Ascorbat + Glutathione
Kosten: 1 NADPH / H2O2
ROS Abbau in Pflanzenzellen - Übersicht
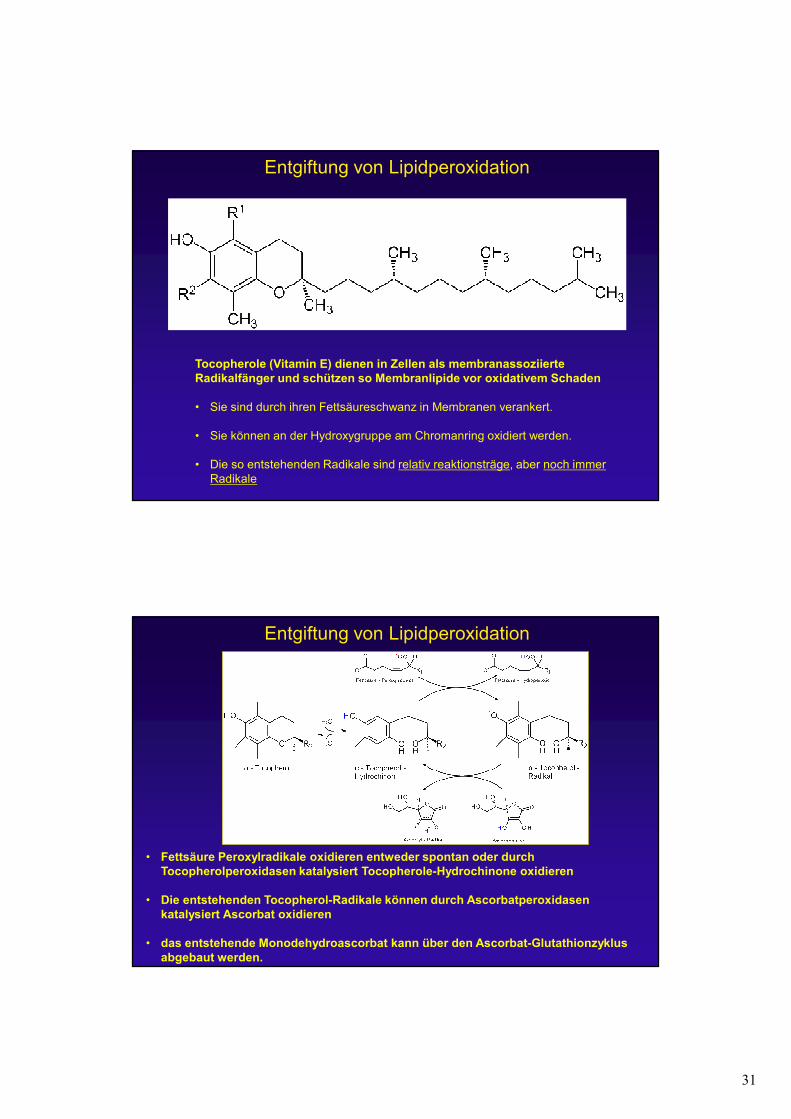
Cytotoxizität von ROS –Beispiel Lipidperoxidation
a) Durch Hydroxylradikale können Fettsäuren zu Lipidradikalen oxidiert werden.
b) In einem weiteren Schritt können diese Lipidradikale mit molekularem Sauerstoff zu Fettsäure-Peroxylreadikalen reagieren.
c) Diese können weitere Fettsäuren oxidieren und so selbst zu Fettsäure-Hydroperoxiden werden. Diese Kettenreaktion läuft weiter, wenn sie nicht gestoppt wird!Durch die Einführung von hydrophilen Gruppen an den Fettsäureschwänzen der Lipide
wird die Membranstruktur zerstört!
a b
c
31
Entgiftung von Lipidperoxidation
Tocopherole (Vitamin E) dienen in Zellen als membranassoziierte
Radikalfänger und schützen so Membranlipide vor oxidativem Schaden
• Sie sind durch ihren Fettsäureschwanz in Membranen verankert.
• Sie können an der Hydroxygruppe am Chromanring oxidiert werden.
• Die so entstehenden Radikale sind relativ reaktionsträge, aber noch immer Radikale
Entgiftung von Lipidperoxidation
• Fettsäure Peroxylradikale oxidieren entweder spontan oder durch
Tocopherolperoxidasen katalysiert Tocopherole-Hydrochinone oxidieren
• Die entstehenden Tocopherol-Radikale können durch Ascorbatperoxidasen
katalysiert Ascorbat oxidieren
• das entstehende Monodehydroascorbat kann über den Ascorbat-Glutathionzyklus
abgebaut werden.

32
RBOH Proteine:Die Quelle für ROS als Signalmoleküle
Regulation
• Ca2+ (durch EF Hände am N-Term)
• Phosphorylierung
• Rop (Rho verwandte GTPasen der Pflanzen)
• Phosphatidylsäure
• S-Nitrosylierung
• Homolog zur gp91phox Untereinheit von Rbos aus Säugern
• 10 AtRBOHs (A-J)
• Plasmamembran Lokalisation (6
Transmembrandomänen)
Plant “respiratory burst oxidases” (Rbohs) are NADPH oxidases related to the mammalian gp91phox subunit of NOX
Guard cells
Pathogen
response
Systemic signal
transduction
Wounding
Salt stress
Root hair
Germination
33
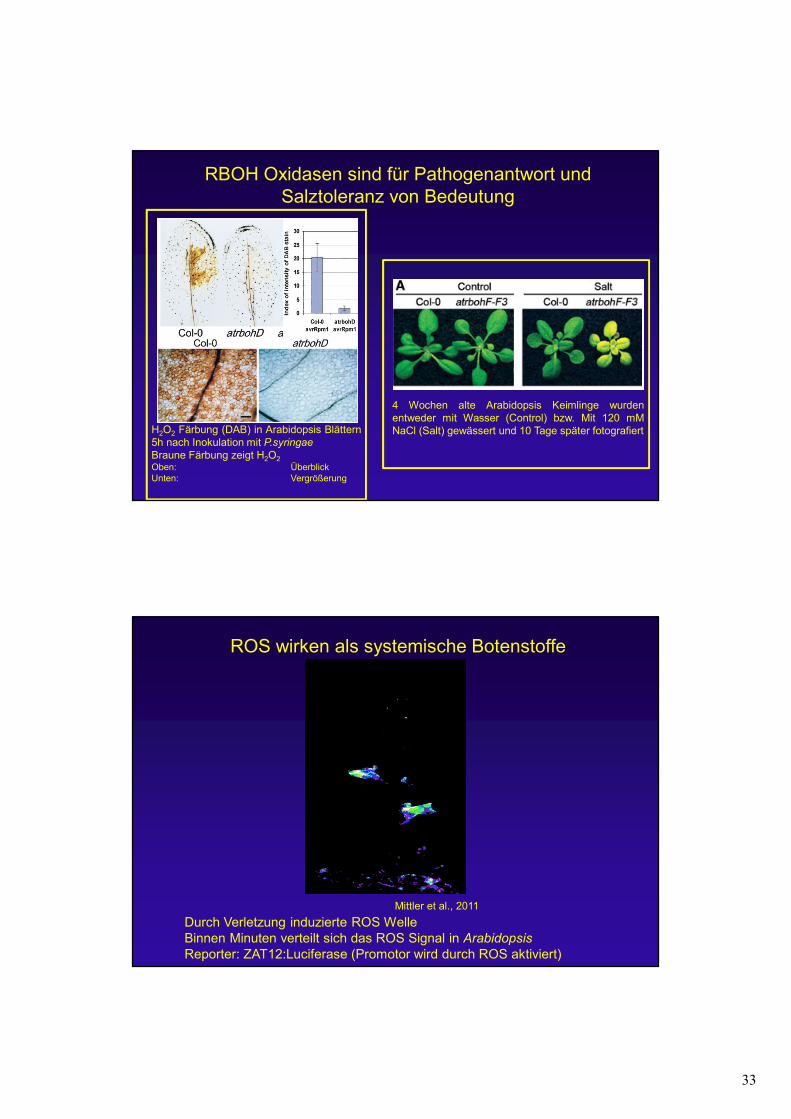
RBOH Oxidasen sind für Pathogenantwort und Salztoleranz von Bedeutung
H2O2 Färbung (DAB) in Arabidopsis Blättern5h nach Inokulation mit P.syringae
Braune Färbung zeigt H2O2Oben: ÜberblickUnten: Vergrößerung
4 Wochen alte Arabidopsis Keimlinge wurdenentweder mit Wasser (Control) bzw. Mit 120 mMNaCl (Salt) gewässert und 10 Tage später fotografiert
Mittler et al., 2011
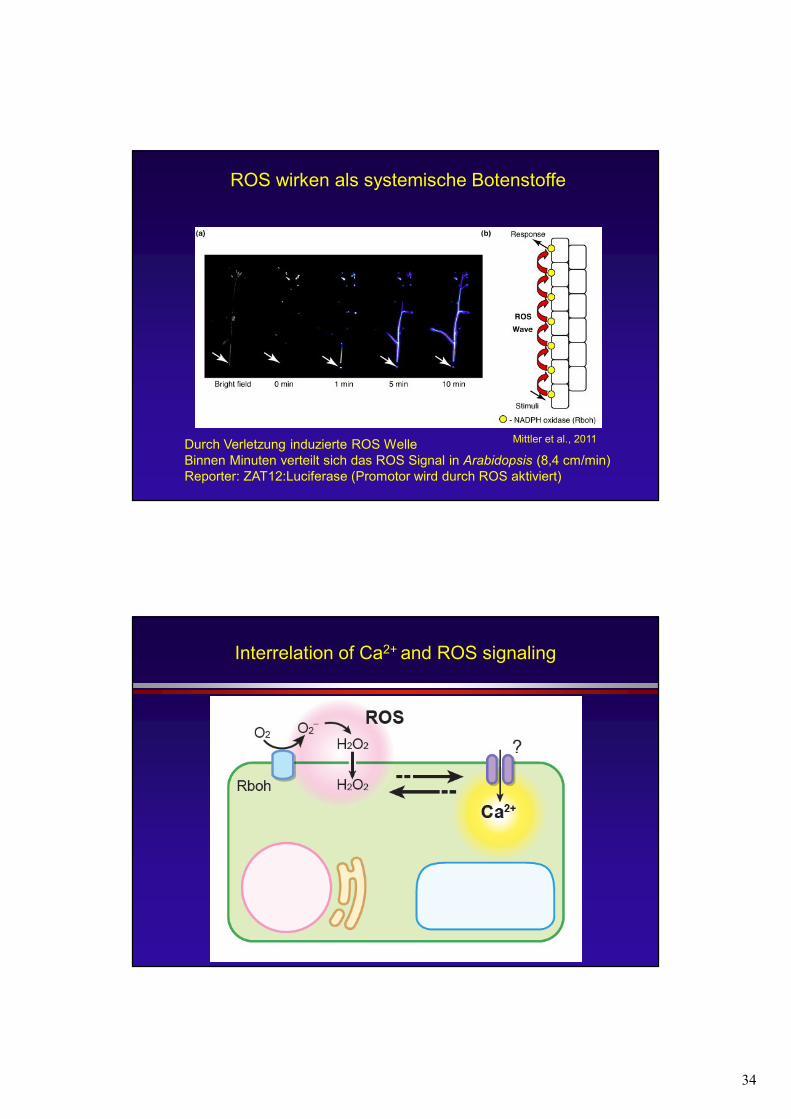
ROS wirken als systemische Botenstoffe
Durch Verletzung induzierte ROS WelleBinnen Minuten verteilt sich das ROS Signal in Arabidopsis
Reporter: ZAT12:Luciferase (Promotor wird durch ROS aktiviert)
34
ROS wirken als systemische Botenstoffe
Durch Verletzung induzierte ROS WelleBinnen Minuten verteilt sich das ROS Signal in Arabidopsis (8,4 cm/min) Reporter: ZAT12:Luciferase (Promotor wird durch ROS aktiviert)
Mittler et al., 2011
Interrelation of Ca2+ and ROS signaling
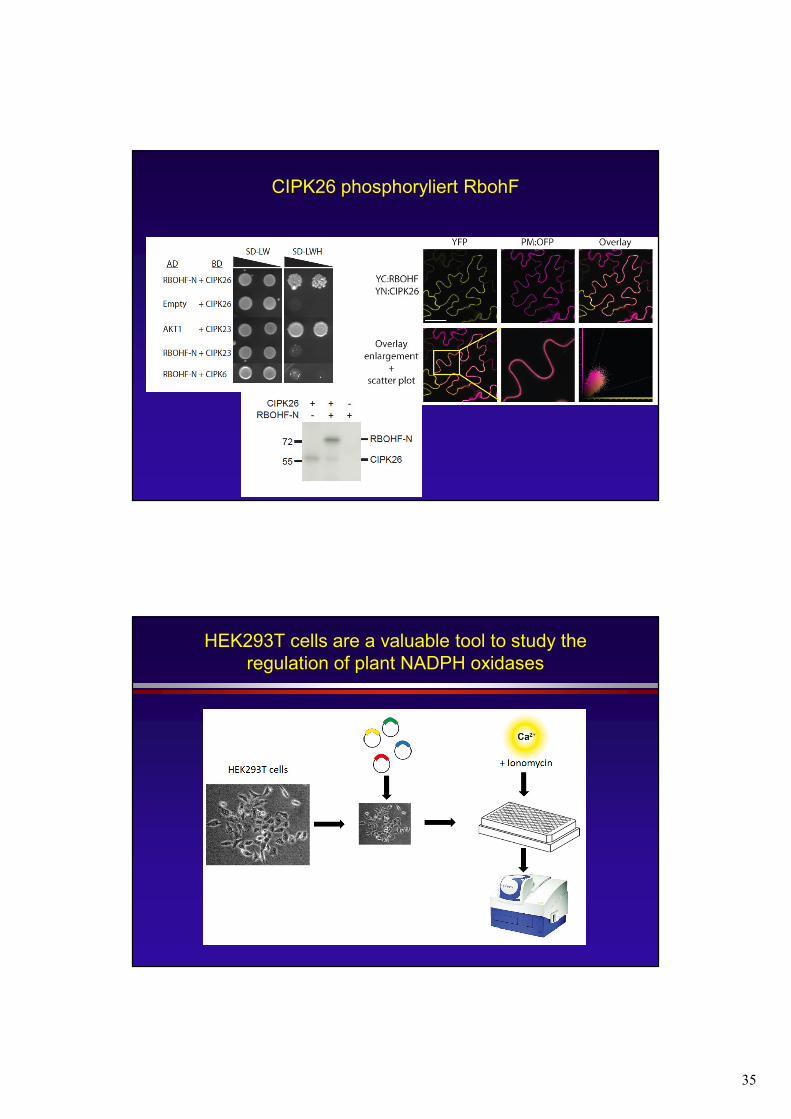
35
CIPK26 phosphoryliert RbohF
HEK293T cells are a valuable tool to study the regulation of plant NADPH oxidases
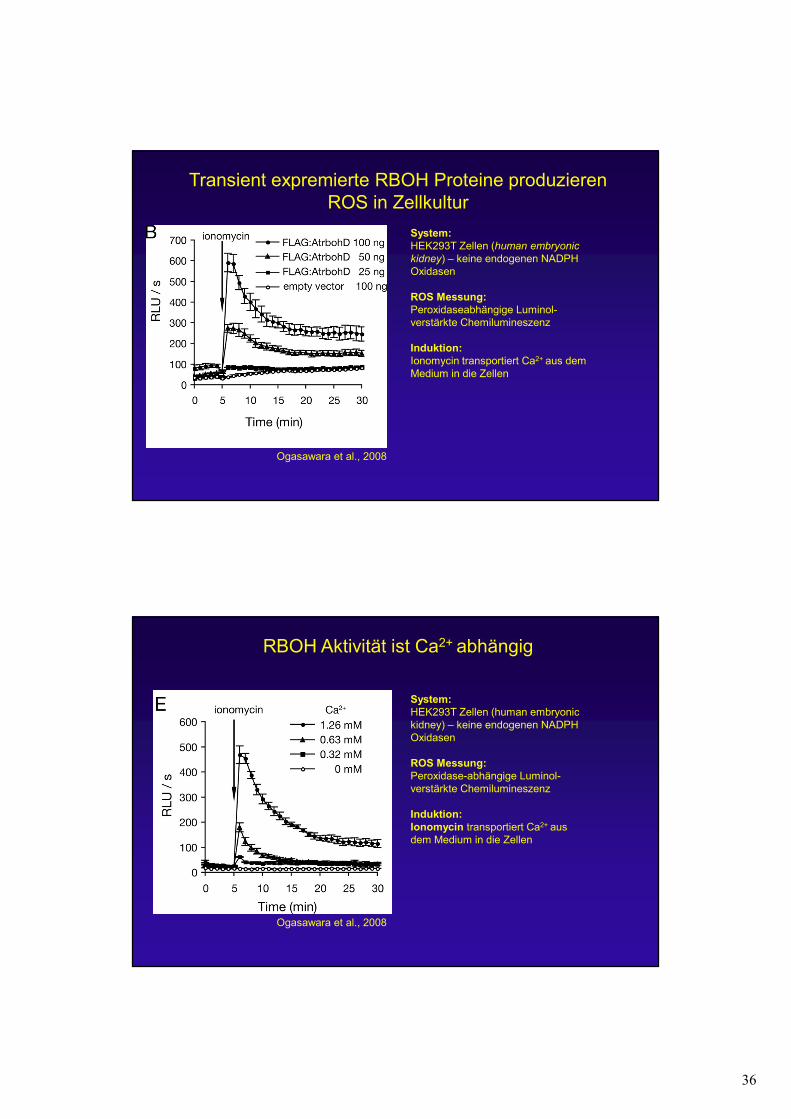
36
Transient expremierte RBOH Proteine produzieren ROS in Zellkultur
Ogasawara et al., 2008
System:
HEK293T Zellen (human embryonic
kidney) – keine endogenen NADPH Oxidasen
ROS Messung:
Peroxidaseabhängige Luminol-verstärkte Chemilumineszenz
Induktion:
Ionomycin transportiert Ca2+ aus dem Medium in die Zellen
RBOH Aktivität ist Ca2+ abhängig
Ogasawara et al., 2008
System:
HEK293T Zellen (human embryonickidney) – keine endogenen NADPH Oxidasen
ROS Messung:
Peroxidase-abhängige Luminol-verstärkte Chemilumineszenz
Induktion:
Ionomycin transportiert Ca2+ aus dem Medium in die Zellen
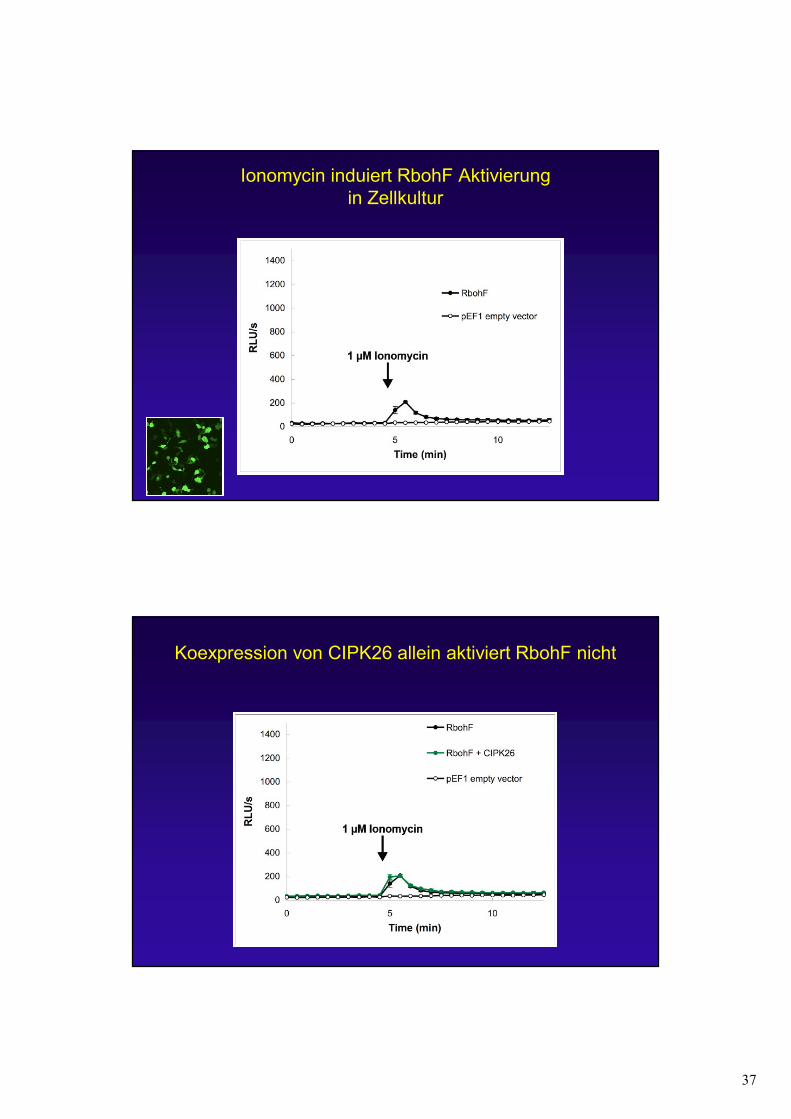
37
Ionomycin induiert RbohF Aktivierungin Zellkultur
Koexpression von CIPK26 allein aktiviert RbohF nicht
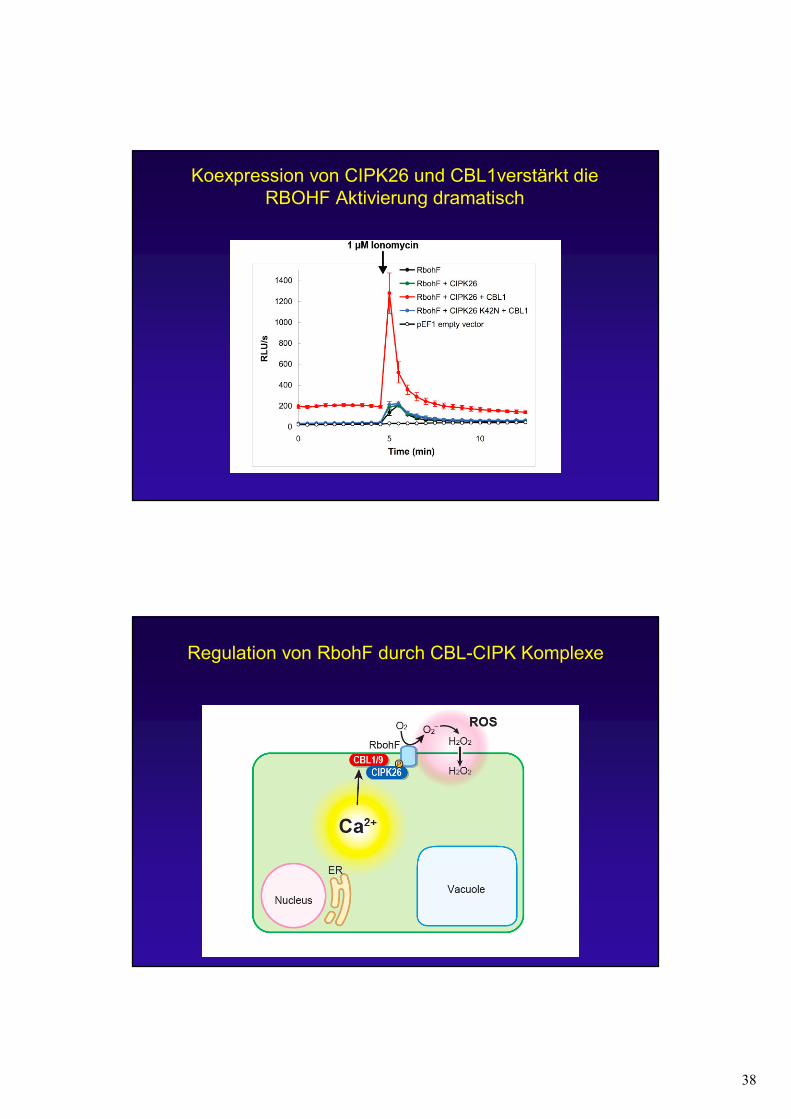
38
Koexpression von CIPK26 und CBL1verstärkt die RBOHF Aktivierung dramatisch
Regulation von RbohF durch CBL-CIPK Komplexe

39
Ca2+ abhängige Rboh Phosphorylierung alsSignalverstärker in Langstrecken-ROS Signalwegen?
Zusammenfassung:
Was charakterisiert die generellen Elemente der
pflanzlichen Signaltransduktion?
Jeder Stimulus wird von einem spezifischen Rezeptor erkannt.
Dieser Rezepter muss entweder direkt eine spezifische und integrierte Antwort auslösen (z.B. Kanal-Typ Rezeptoren) oder das Signal durch sekundäre Botenstoffe weiterleiten.
Die durch die sekundären Botenstoffe kodierten Informationen müssen durch weitere Signalweg-Komponenten entschlüsselt werden und zu Reaktionen führen, die dem ursprünglichen Stimulus entsprechen.

40
Zusammenfassung:
Wie funktioniert die Signalwahrnehmung und
Weiterleitung von verschiedensten Stimuli, mit einem
begrenzten Repertoire an Signaltransduktions-
Komponenten?
Ein sekundärer Botenstoff kodiert Informationen durch seine Konzentration, seine Lokalisation und die Frequenz seiner Präsenz.
Daher ist es notwendig, dass sekundäre Botenstoffe über strikt regulierte Ruhekonzentrationen verfügen.
Desweiteren ist notwendig, dass nach einem Reiz die Ruhekonzentrationen des Botenstoffes schnell wieder erreicht werden.
Recommended









![Vorlesung Zellbiologie Teil Biologie: Evolution – Zellbiologie – Entwicklung (Alberts, Kapitel 17) [ ppt ohne copyright-geschützte Abb.] Institut für Biologie](https://img.pdfslide.org/doc/110x75/55204d6949795902118bff28/vorlesung-zellbiologie-teil-biologie-evolution-zellbiologie-entwicklung-alberts-kapitel-17-ppt-ohne-copyright-geschuetzte-abb-institut-fuer-biologie.jpg)







