Embed Size (px)
Citation preview

4. Arbeitstreffen deutschsprachiger
Echinodermenforscher
4th Workshop of German & Austrian
Echinoderm Research
24.-26. October 2008
ABSTRACTS
Andreas Kroh & Brigitta Schmid
Naturhistorisches Museum Wien


4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Ei
n.
Fü
Re
A-1010 Wien, Österreich (Austria)
Te
@nhm-wien.ac.at
e-
entwurf: Andreas Kroh
Ti
W
Impressum
gentümer, Herausgeber und Verleger:
© 2008 Natuhistorisches Museum Wien
Alle Rechte vorbehalte
r den Inhalt der Beiträge sind die Autoren verantwortlich.
daktion:
Naturhistorisches Museum Wien
Burgring 7,
l.: +43 (1) 521 77 / 576 (Kroh); +43 (1) 521 77 / 564 (Schmid)
Fax: +43 (1) 521 77 / 459
e-mail: andreas.kroh
mail: [email protected]
Redaktionelle Bearbeitung (EDV, Layout, Grafik): Andreas Kroh
Umschlag
telbild: Coelopleurus (Keraiophorus) exquisitus COPPARD & SCHULTZ, 2006;
Neukaledonien (NHMW 2005z0277/0016; Photo von Alice SCHUMACHER, NHM
ien).
24.-26. October 2008 iii

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Inhalt
Encrinus sp. cf. E. robustus (Crinoidea, Encrinidae) aus dem Unteren
M
Devonian) (Poster)
anisms and life style
(P
lkalk-Crinoide Encrinus liliiformis -
V
rschiedenen Seeigeln und ihr bionisches
Po
)
B
H.C. Hotchkiss & Alexander Glass..............................................23
In
Tithonian of the Klippenbelt (Jurassic, Austria) (Poster)
uschelkalk von Niedersachsen (Poster)
Ulrich Bielert, Friedrich Bielert & Hans Hagdorn...........................................1
Classification of pre- and postmortem ossicular modifications of the
Cupressocrinitid skeletons (Crinoidea,
Jan Bohatý ............................................................................................3
Dental insights into ophiuroids: Feeding mech
oster)
Karin Boos...........................................................................................14
Ökophänotypbildung der Musche
ergleich zweier Populationen aus Nord-Württemberg (Vortrag)
Janina F. Dynowski & James H. Nebelsick.................................................15
Stereomdifferenzierung in ve
tenzial (Vortrag)
Nils Großmann & James H. Nebelsick.......................................................17
Die Stereome diverser regulärer Seeigelstachel (Poster
Nils Großmann & James H. Nebelsick.......................................................19
30 Years of Research on Crinoids / 30 Jahre Crinoidenforschung
(Keynote)
Hans Hess ...........................................................................................20
Pattern formation in starfish: arm stumps, regeneration models, and
evolution (Poster)
Frederick H.C. Hotchkiss........................................................................21
dellacoma in the Hunsrück Slate (Lower Devonian, Germany):
reidentification of Urasterella verruculosa (Asteroidea, Bdellacomidae)
(Poster)
Frederick
dex of Living and Fossil Echinoids: 1971 - present (Poster)
Andreas Kroh.......................................................................................25
Crinoids from the
Andreas Kroh & Alexander Lukeneder......................................................26
iv 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Temperature effect on feed consumption, absorption and assimilation
efficiencies and production in the sea urchin Strongylocentrotus
in
son L.
oster)
..............................................30
M
mplexen Strukturen (Vortrag)
Ec
............................................................................36
Ph
equence data (Vortrag)
Marleen Perseke, Guido Fritzsch, Detlef Bernhard, Peter F. Stadler & Martin
Schlegel ..............................................................................................38
New insights into early diversification of sea cucumbers as inferred
from calcareous ring material (Ordovician/Silurian, Baltoscandia)
(Vortrag)
Mike Reich...........................................................................................39
A nearly articulated holothurian from the Santonian (Cretaceous) of
England (Poster)
Mike Reich...........................................................................................40
termedius (Poster)
John M. Lawrence, Xuebin Cao, Yaqing Chang, Ping Wang, Y. Yu, Addi
Lawrence, Stephen A. Watts ..................................................................28
Amount of arm loss and rate of arm regeneration by Luidia clathrata
(Echinodermata: Asteroidea) (P
J.M. Lawrence & C.M. Pomory ................................................................29
Seastars (Asteroidea) from Madagascar (Vortrag)
Horst Moosleitner ...................................
orphologie und Faziesverteilung des indo-pazifischen Seeigels
Jacksonaster depressum (Vortrag)
James Nebelsick & Atef Abd El-Hamied A. S. Elattaar.................................32
Taphonomie von Jacksonaster depressum (Echinoidea: Clypeasteroida)
aus dem Roten Meer (Poster)
James Nebelsick & Atef Abd El-Hamied A. S. Elattaar.................................33
Skelett-Endosymbionten fossiler und rezenter Echinodermen: Von
Bioklaustrationen zu ko
Christian Neumann ...............................................................................34
hinoids from the Neogene of Portugal mainland: Systematics,
Palaeoecology, Palaeobiogeography (Vortrag)
Pedro Pereira ...........
ylogenetic relationships within Echinodermata based on mitochondrial
s
24.-26. October 2008 v

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Unusual holothurians (Echinodermata) from the Late Ordovician of
Sweden (Poster)
Mike Reich...........................................................................................42
Signs of predation on Middle Triassic crinoids form Poland (Poster)
Mariusz A. Salamon & Przemysław Gorzelak .............................................43
Are that sea urchins ???? - The strange morphology of the echinoid
family Pourtalesiidae (Vortrag)
Heinke Schultz .....................................................................................45
The early history of echinoderms: new insights from disarticulated early
and middle Cambrian ossicles (Keynote)
Andrew B Smith ...................................................................................48
Rhaetian ophiuroids from the Netherlands: a preliminary report (Poster)
Ben Thuy, Adiël A. Klompmaker & John W. M. Jagt....................................49
Irregular echinoids from the Turonian Seewen Limestone of the Allgäu
area (S. Germany) (Vortrag)
Frank Wiese & Nils Schlüter ...................................................................51
Antiquity and Longevity of a deep-sea oyster/crinoid association (Azores
Archipelago) (Vortrag)
Max Wisshak, Christian Neumann, Matthias López Correa, Serge Gofas,
Carmen Salas, Marco Taviani, Joachim Jakobsen, André Freiwald ................53
Bromierte Anthrachinon-Pigmente aus der rezenten Seelilie Proisocrinus
ruberrimus (Vortrag)
Klaus Wolkenstein, Wolfgang Schoefberger, Norbert Müller & Tatsuo Oji.......55
vi 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
24.-26. October 2008 vii

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
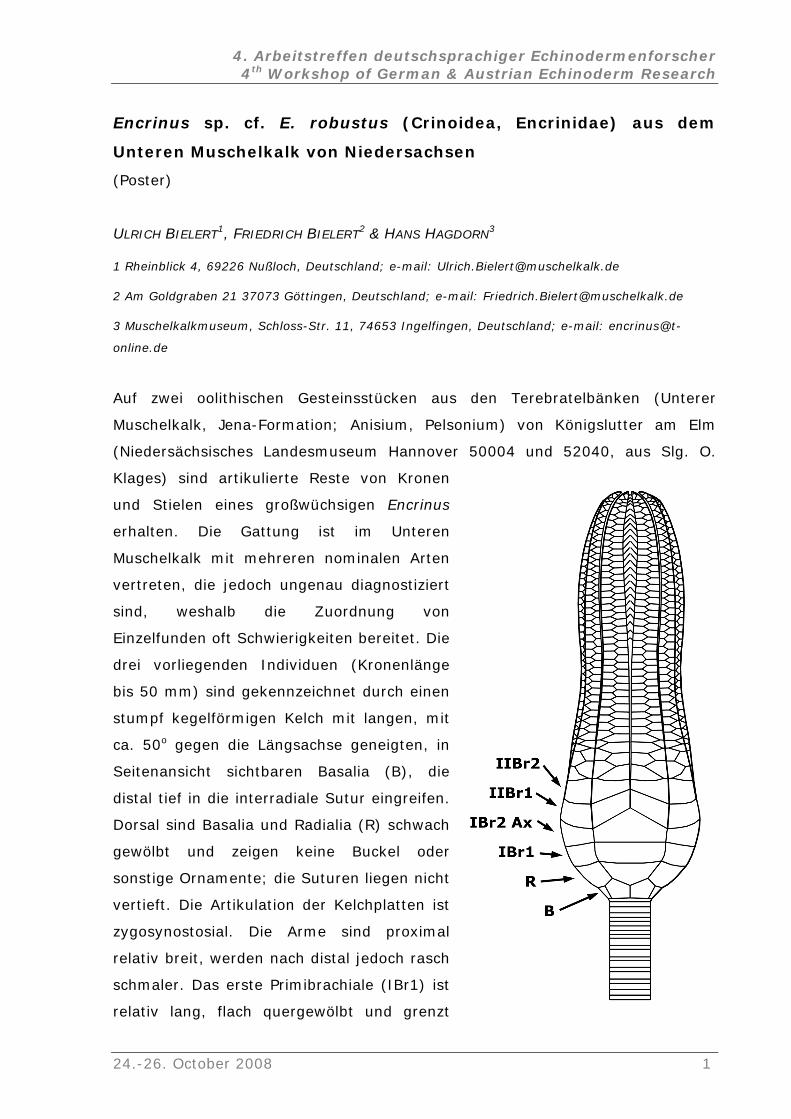
Encrinus sp. cf. E. robustus (Crinoidea, Encrinidae) aus dem
Unteren Muschelkalk von Niedersachsen
(Poster)
ULRICH BIELERT1, FRIEDRICH BIELERT2 & HANS HAGDORN3
1 Rheinblick 4, 69226 Nußloch, Deutschland; e-mail: [email protected]
2 Am Goldgraben 21 37073 Göttingen, Deutschland; e-mail: [email protected]
3 Muschelkalkmuseum, Schloss-Str. 11, 74653 Ingelfingen, Deutschland; e-mail: encrinus@t-
online.de
Auf zwei oolithischen Gesteinsstücken aus den Terebratelbänken (Unterer
Muschelkalk, Jena-Formation; Anisium, Pelsonium) von Königslutter am Elm
(Niedersächsisches Landesmuseum Hannover 50004 und 52040, aus Slg. O.
Klages) sind artikulierte Reste von Kronen
und Stielen eines großwüchsigen
erhalten. Die Gattung ist im Unteren
Muschelkalk mit mehreren nominalen Arten
vertreten, die jedoch ungenau diagno
sind, weshalb die Zuordnung von
Einzelfunden oft Schwierigkeiten bereite
drei vorliegenden Individuen (Kronenlänge
bis 50 mm) sind gekennzeichnet durch einen
stumpf kegelförmigen Kelch mit langen, mit
ca. 50o gegen die Längsachse geneigten
Seitenansicht sichtbaren Basalia (B), di
distal tief in die interradiale Sutur eingreifen.
Dorsal sind Basalia und Radialia (R) schw
gewölbt und zeigen keine Buckel
sonstige Ornamente; die Suturen liegen
vertieft. Die Artikulation der Kelchplatten
zygosynostosial. Die Arme sind proxima
relativ breit, werden nach distal jedoch ra
schmaler. Das erste Primibrachiale (IBr1)
relativ lang, flach quergewölbt und grenzt
Encrinus
stiziert
t. Die
, in
e
ach
oder
nicht
ist
l
sch
ist
24.-26. October 2008 1

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
mit langer Sutur gegen die Nachbarglieder. Auch das Primaxillare (IBr2Ax) ist
relativ lang und hat wie das IBr1 keine dorsalen Buckel. Zusammen mit den
sten Sekundibrachialia (IIBr1, IIBr2) wirken Kelch und proximale Arme
allonförmig aufgetrieben. Ab dem dritten Sekundibrachiale (IIBr3) setzt abrupt
eiligkeit der Armglieder ein. Die Armzweige sind zunächst dorsal noch
dorsal gerundet. Die Brachialia sind dorsal
nnähernd glatt und unornamentiert. Die Pinnularia sind schlecht erhalten. Am
ch schwankt, ist der
iagnostische Wert des Kelches mit den langen Basalia höher einzuschätzen als
) und E. robustus aus Oberschlesien
SSMANN 1926) und Thüringen (BIELERT & BIELERT 2000) haben einen
BEYRICH, E., 1857. Über die Crinoiden des Muschelkalks. – Abhandlungen der Akadademie
wei Arten der Gattung
Encrinus im Unteren Muschelkalk am Nordrand des Thüringer Beckens. – Beiträge
zur Geologie von Thüringen, Neue Folge 7: 137-145.
beiden er
b
echte Zweiz
schwach quergewölbt und haben scharf abgegrenzte Seitenflächen, werden ab
dem IIBr3 aber schmaler und
a
Stiel fällt auf, dass auch die proximalen, zirrenlosen Nodalia kaum länger und
dicker sind als die Internodalia und flache Seiten ohne Randwulst haben. Die
Nähte zwischen den durchweg runden Columnalia sind kaum vertieft. Wie bei
Populationen von E. liliiformis, wo die dorsale Ornamentierung von Kelch und
Armgliedern als ökophänotypisches Merkmal erhebli
d
die dorsale Ornamentierung von Kelch- und Armgliedern. Nur die altersgleichen
E. brahli aus Rüdersdorf (BEYRICH 1857
(A
flachkonischen Kelch und Armglieder ohne dorsale Skulptur. Den
halbkugelförmigen dorsalen Auftreibungen der Primaxillaria beim schlesischen
und thüringischen Material von E. robustus messen wir nur geringe
diagnostische, sondern ökophänotypische Bedeutung zu. Der Status von E.
brahli, dessen Typusmaterial (subadulte Individuen) verschollen ist, bedarf
weiterer Untersuchungen. Die hier vorgestellten Encriniden werden als Encrinus
sp. cf. E. robustus ASSMANN, 1925 bezeichnet.
Literatur
ASSMANN, P., 1926. Die Fauna der Wirbellosen und Diploporen der ober- schlesischen
Trias mit Ausnahme der Brachiopoden, Lamellibranchiaten, Gastropoden und
Korallen. – Jahrbuch der preußischen Geologischen Landes-Anstalt für 1925, 46:
504-527.
der Wissenschaften Berlin, 1: 1-49.
BIELERT, U. & BIELERT, F., 2000. Gemeinsames Auftreten von z
2 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Classification of pre- and postmortem ossicular modifications of
the Cupressocrinitid skeletons (Crinoidea, Devonian)
Poster)
ithout classifiable causes; 3, premortem ossicle anomalies as a
action of external interferences; 4, pre- and postmortem borings and bite
es without recognizable external influences –
generic" abnormalities (Figs. 1-2):
with regular grown
xial canals (~1500 skeletons analyzed). Further, individuals with additional or a
(
JAN BOHATÝ
Institut für Geologie und Mineralogie der Universität zu Köln, Zülpicher Str. 49a, 50674 Köln,
Germany; e-mail: [email protected]
Skeletal anomalies on Devonian cupressocrinitids (Crinoidea, Cladida) are herein
classified as: 1, growth anomalies without external influences; 2, growth
anomalies w
re
marks; and 5, pre- and postmortem epizoan encrusting.
1. Growth anomali
"
Growth anomalies without recognizable external influences are predominantly
distinguished by the reduction of thecal or brachial-ossicles respectively by
additional intermediary plates. These abnormalities could not be attributed to
injuries or involved regeneration and are obviously "genetically modified
anomalies" (BOHATÝ 2001). Most common are variances of the columnal axial
canal (Fig. 1), which occurs at the rate of ~1: 30 compared
a
reduced number of ossicles are recognized. Cupressocrinitids with a developed
quadrangular or hexagonal symmetry (Figs. 1-2) are relatively rare and occur at
several localities with an average rate of ~1: 70 compared with regularly
developed skeletons (~700 aboral cups and ~300 crowns analyzed). Due to the
abundance of anomalously grown axial canals or symmetry aberrations within
one fossil-horizon, the genetic basis of these interferences is assumed. In this
case, the appropiative rates of detectable growth anomalies compared with
normal individuals, could be higher than above-mentioned.
24.-26. October 2008 3

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
2. Growth anomalies without classifiable causes – without indications of
external influences (Fig. 3):
In some cases it is not possible to determine a cause for a growth anomaly. The
individuals with one additional or missing plate (Fig. 3), with an inexplicable
ossicular-swelling, or a modified exobrachial layer are not recognizable as
regeneration of the skeleton, "wound healings", or as documented "generic"
abnormalities. No direct evidence of predatory influences like borings or bite
marks can be recognized. Therefore, these modifications are summarized as
growth anomalies without classifiable causes – without indications of external
influences.
3. Premortem ossicular anomalies as a reaction of external interferences
– "wound healing" and skeletal regeneration of thecal or brachial
injuries (Figs. 4-5):
"Wound healing": Different sized anomalies in numerous small ossicles were
recognized on ~5 % of the studied cupressocrinitids (~700 aboral cups and
~300 crowns analyzed). These anomalies are obviously "wound healings" of
nonlethal injured individuals. Possible causes of these anomalies could be injuries
caused by predators or possibly by impact-injuries with suspended clastic
material. The affected regions may be small or large (Fig. 4). The maxim
observed injury affects up to 80 % of the surface of the cup.
Regeneration: Regenerations of echinoderm skeletons was recently reconsidered
by MOZZI et al. (2006), exemplified by the regenerative processes of the
editerranean Featherstar" Antedon mediterranea (LAMARCK, 1816). AMEMIYA &
JI (1992) described the crinoid regeneration processes. The regeneration in
1890. Direct interconnections between
"M
O
fossil crinoids was also discussed by GAHN & BAUMILLER (2005). For example, they
showed arm regeneration of Rhodocrinites kirbyi (WACHSMUTH & SPRINGER, 1889)
and Dichocrinus cinctus MILLER & GURLEY,
the increase of shell-breaking predators and the number of observed arm
regenerations of nonlethal injured crinoids were recognized (GAHN & BAUMILLER
4 24.-26. October 2008
4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
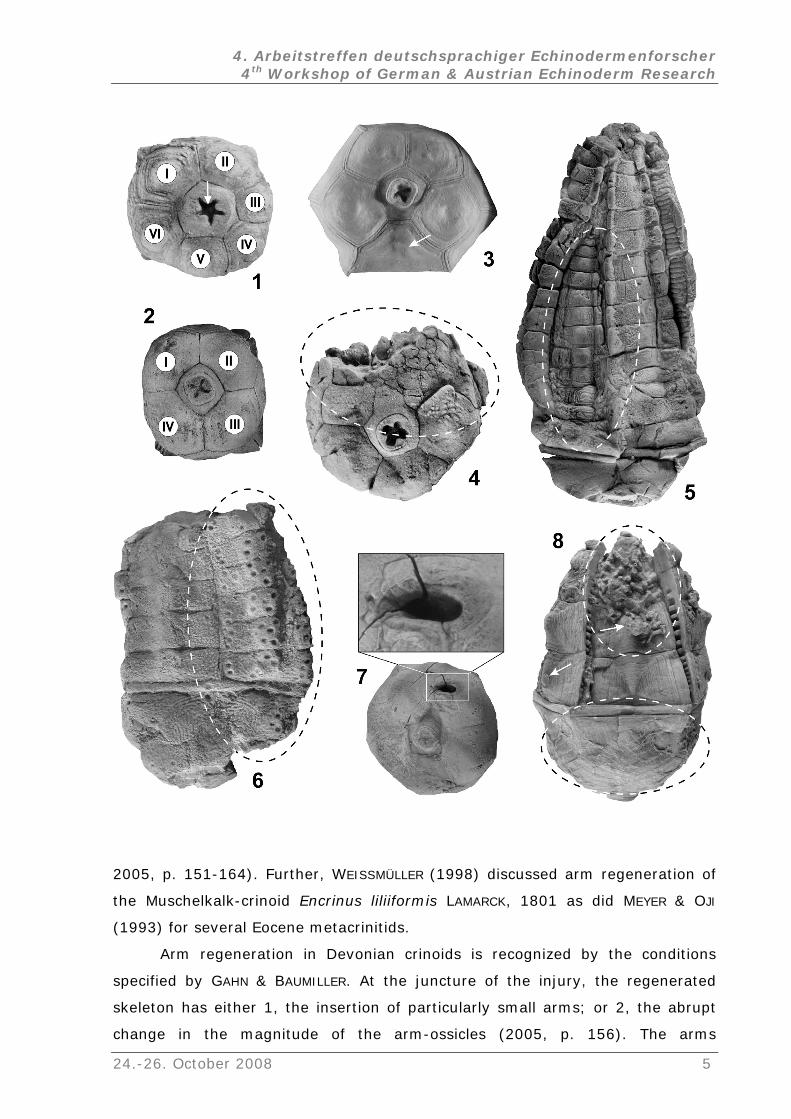
2005, p. 151-164). Further, WEISSMÜLLER (1998) discussed arm regeneration of
the Muschelkalk-crinoid Encrinus liliiformis LAMARCK, 1801 as did MEYER & OJI
(1993) for several Eocene metacrinitids.
Arm regeneration in Devonian crinoids is recognized by the conditions
specified by GAHN & BAUMILLER. At the juncture of the injury, the regenerated
skeleton has either 1, the insertion of particularly small arms; or 2, the abrupt
change in the magnitude of the arm-ossicles (2005, p. 156). The arms
24.-26. October 2008 5

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
recognized as regenerated were all smaller than regularly developed arms (Fig.
5). Nevertheless, the arms of the relevant individuals are enclosed in the typical
prescu socrinitid-like resting or avoidance posture, whereas the adjoining, normal
longer arms closed about the smaller one and are tangent distally above the
regenerated arm.
4. Pre- and postmortem borings and bite marks (Figs. 6-7):
Postmortem multi-borings: Almost 90 % of ~50 analyzed skeletons of C.
elongatus were covered by borings (SIEVERTS-DORECK 1963; BOHATÝ 2001, p. 8;
20 pl. 10, figs. 1-3) [Fig. 6]. More infrequently, specimens with multiple
borings were identified on the crowns of C. crassus (2006, pl. 10, fig. 8b). Both
species are covered by a thin and monolamellar exoplacoid layer, which
apparently offered less resistance against boring organisms, in contrast to the
multilamellar layers of Abbreviatocrinites. Generally, these borings were
restricted to the non-embedded
06,
side of the relevant skeletons and trend in
ordinated lines from the cup (or also from the preserved stem) and over one or
nnot be verified.
in
several arms. Presumably, the borings occurred soon after death. The skeletons
are articulated and covered by the unsheared exoplacoid layer on the one hand,
but on the other, the borings are restricted to the non-embedded side of the
crown. Platyceratid gastropods were discussed as a possible causer of the
borings (SIEVERTS-DORECK 1963). This theory ca
Pre- and postmortem incurred single borings: Single borings are present on the
ssicles of A. abbreviatus abbreviatus, A. geminatus, and Robustocrinites n. sp.
d by a trepostome bryozoan
Eostenopora sp.). Because the boring is positioned on the non-embedded side
o
In abbreviatocrinitids, they are normally restricted to the plates with sheared
exoplacoid layers and, therefore, most likely occurred postmortem. The single
boring of an unknown organism at the surface of the monolamellar exoplacoid
layer, observed in one affected robustocrinitid, is fille
(?
of the crown and runs across several plate boundaries, it is assumed to have
occurred postmortem.
BAUMILLER & MACURDA (1995) and BAUMILLER (1990, 1993) documented
borings on Paleozoic blastoids and crinoids. Also in this case, platyceratid
gastropods were discussed as the possible borers. A significant bit of evidence for
6 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
this theory is perhaps documented in the combined fossil evidence of a borehole,
positioned next to a gastropod valve (BAUMILLER 1990).
SEM-observations of thin cross-sections of the multilamellar exoplacoid
layer of A. geminatus exhibits potentially premortem microendolithic borings.
These meandering single borings have an averag proportion of 20 µm in width to
al boring(?) on the basal plate of A.
bbreviatus. The visible stereomatic reaction in the form of an annulus-like
arks
300 µm length. They were presumably lined with biogenous matter and
ultimately resulted in a secondary sulphide-ion surplus. Through this, the borings
are lined with marcasite-crystal agglomerates (FeS2). Microendolithic borings
could be observed in ~70 % of the studied multilamellar exoplacoid layers, but in
less than 20 % of the basal, radial, or brachial plates (30 thin sections analyzed).
Fig. 7 presumably has a deep, ov
a
swelling indicates that the single-boring occurred most likely premortem. But
isolated placoderm teeth from the same location also permit the assumption that
this trace may to the bite of a larger predator instead of a boring organism, like a
gastropod with specialised radula.
Premortem bite m : Bite marks at cupressocrinitids are rare and could be
bserved in less than 3 % of the studied individuals (~1500 skeletons analyzed).
n the crown-ossicles of cupressocrinitids.
o
They are possibly attributed to cephalopods, placoderms or arthropods.
Premortem bite marks are recognized as nonlethal injuries, because the bite
marks are accompanied by "wound healings".
5. Pre- and postmortem incurred epizonal encrusting (Fig. 8):
The epibiontic encrusting of Devonian crinoids, exemplified by Upper Eifelian
columnals, was recently discussed by Gluchowski (2005). Bryozoa,
Microconchida, Crinoidea, Tabulata, Rugosa, and Stromatoporida are also
identified o
Bryozoa: "Cyclostome bryozoans" (Hederella sp.) apparently preferentially
encrusted the crown-ossicles of Abbreviatocrinites nodosus (Fig. 8). Unlike other
cupressocrinitids, nearly 95 % of the observed A. nodosus-skeletons from the
lausbach and Nims members bear encrustings (~80 skeletons analyzed). The
growth of the hederellids most likely occurred instantaneously postmortem,
K
24.-26. October 2008 7

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
because some articulated crowns retain unsheared exoplacoid layers. In contrast,
the "cyclostome bryozoans" settled beyond the primary movable ossicle
boundaries of the endoskeleton. Presumably, the hederellids had a rapid rate of
growth. Gluchowski (2005, figs. 4F-H) also documented the hederellid-encrusting
of Upper Eifelian crinoid columnals.
Hederella is presumably not a true bryozoan (A. Ernst, personal commun.;
as colonial,
also see WILSON & TAYLOR 2001). TAYLOR & WILSON (2007) favored a close
relationship with phoronids, tentatively interpreting hederelloids
phoronid-like invertebrates with retractable lophophores. Along with
microconchids and cornulitids, hederelloids may have been part of a mid-
Paleozoic acme of lophophorate "worms".
Trepostome bryozoans: One brachial of a completely preserved A. nodosus
rown, one cup of an also entire Abbreviatocrinites schreueri crown, and one c
theca of P. gracilis were encrusted postmortem by trepostome bryozoans
(?Eostenopora sp.). The boring trace of an affected Robustocrinites arm is also
populated by (?)Eostenopora sp.; in this case, the colony settled in a non-
exposed position.
Fenestrate bryozoans: Especially within the uppermost Ahbach Formation
(lowermost Lower Givetian) of the "Wotan" quarry (Hillesheim syncline, Eifel,
enis
eath the attached stem, allows the presumption of a premortem
ettlement (compare BOHATÝ 2005, fig. 3B). In contrast, some shorter stem
ted that the bryozoans lived
Rh h Massif, Germany), several longer stems of A. geminatus and P. gracilis
were found encrusted by fenestrate bryozoans. The length of the overgrown
columnals as well as some observed embedding patterns of bryozoans located
undern
s
fragments or other disarticulated cupressocrinitid-ossicles were usually encrusted
postmortem. This assumption is based on the entire enclosure of some skeletal
elements.
Similarly holdfasts of most likely rhomboporid bryozoans attached to the
columnals of Schyschcatocrinus creber DUBATOLOVA, 1975, as reported by
GLUCHOWSKI (2005, figs. 3A-B). Gluchowski indica
attached to the fragmented dead stems that lay horizontally on the sea floor.
8 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Crinoidea: One observed cup of A. abbreviatus abbreviatus as well as ne
isolated radial and arm plate of A. geminatus were encrusted by the holdfasts of
other cladid crinoids (?P. gracilis). The roots settled postmortem on the shearing
of the multilamelar exoplacoid layer, at the interior of a radial plate, or at the
multilamellar exobrachial layer of an isolated brachial. This association was
observed on less than 3 % of the studied crinoids (~1500 skeletons analyzed).
GLUCHOWSKI (2005, p. 322) documented the postmortem encrusting of several
o
all
the columns of crinoid hosts, as well as dendritic
oldfasts distributed on all sides of the column were reported from Silurian strata
sm crinoid holdfasts attached to Upper Eifelian crinoid columnals.
Various attachments of crinoid juveniles to living or dead adults are known
from the Silurian to the Mississippian (see MEYER & AUSICH 1983). Coiling stems,
modified discoid holdfasts on
h
by FRANZÉN (1977) and PETERS & BORK (1998).
Lophophorata (?) - Microconchida (Fig. 8): Microconchids with unstructured or
ornamented valves encrusted the ossicles of nearly 40 % of the studied
cupressocrinitids (~1500 skeletons analyzed). It is remarkable, that larger
individuals are rare and isolated (compare Bohatý 2006, pl. 5, fig. 8), whereas
numerous smaller microconchids encrusted the crinoids. Supposeably, the
s encrusting of microconchids on the columnals of
antalocrinus scutellus LE MENN, 1985 and Schyschcatocrinus creber DUBATOLOVA,
colonization occurred immediately postmortem, before shearing of the exoplacoid
layers and ossicle disarticulation.
The single-specie
T
1975, represent additional settlement examples (GLUCHOWSKI 2005, p. 323, figs.
5I-L).
Anthozoa - Tabulata: The predominantly postmortem settlement of tabulate
orals was recognized on 5-10 % of the studied cupressocrinitids (~1500
zed). The most common epibiontic tabulates were auloporids like
s. serpens
of A. sus.
coral ( D RBIGNY, 1850) was found within the lower part
of the Loogh Formation (Lower Givetian) in the "Wotan" quarry (Hillesheim
c
skeletons analy
Aulopora cf. A. serpens minor (GOLDFUSS, 1829) and A. cf. A.
(GOLDFUSS, 1829), settling on isolated crown-ossicles, completely preserved cups
geminatus, and on the crowns of A. nodo
Furthermore, one cup of A. a. abbreviatus with an encrusting favositid
Favosites cf. F. goldfussi 'O
24.-26. October 2008 9

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
syncline). GLUCHOWSKI (2005) documented small colonies of Favosites sp.
hed to Pentagonostipes petaloides MOORE & JEFattac FORDS, 1968 and
the a
living crin orted from the Upper Silurian (HALLECK
1973; B
GALLE s (compare MEYER & AUSICH 1983).
assoc
(GLUC AUSICH 1983; POWERS & AUSICH 1990
Rugo
Tantalocrinus scutellus LE MENN, 1985 and discussed the possible growth along
xis of the upright stalk of a living host. Favositids that lived attached to
oid hosts have also been rep
RETT & ECKERT 1982; PETERS & BORK 1998), Lower Devonian (GALLE 1978;
& PROKOP 2000), and Lower Carboniferou
Other tabulate corals (e.g. Antholites, Cladochonus, and Emmonsia)
iated with living crinoids are known from Devonian–Mississippian strata
HOWSKI 2005, p. 319; also see MEYER &
and DONOVAN & LEWIS 1999).
sa: Within the Ahbach and Loogh formations (Eifelian/Givetian threshold) in
cupre
CHLÜTER, 95
additional s,
ings of
respo
Porife
the "Wotan" quarry (Hillesheim syncline), rugose corals settled on disarticulated
ssocrinitid stems and isolated ossicles, including Glossophyllum soetenicum
(S 18 ) and Thamnophyllum caespitosum (GOLDFUSS, 1826). The
recovery of a completely overgrown theca (stromatoporoid suffusion
see below) documents a further epibiontic settlement by an indeterminable
juvenile stadium of a rugose coral. All settlements occurred postmortem.
GLUCHOWSKI (2005, p. 317-319) detected the premortem encrust
the rugose coral (?)Adradosia sp. on Schyschcatocrinus creber by the stereomic
nse of the crinoid.
ra(?) - Stromatoporida: Some non-disarticulated cups of A. a. abbreviatus
pletely encrusted by indeterminable stromatopowere com roid suffusions. These
encrusting ose corals,
Refe
AMEM
BAUMI
BAUMI
thaia, 26: 41-47.
s could be settled again by chaetetids, tabulate and rug
microconchids, and bryozoans.
rences:
IYA, S. & OJI, T. 1992. Regeneration in sea lilies. Nature, 357: 546-547.
LLER, T.K. 1990. Non-predatory drilling of Mississippian crinoids by platyceratid
gastropods. Palaeontology, 33/3: 743-748.
LLER, T.K. 1993. Boreholes in Devonian blastoids and their implication for boring by
platyceratids. Le
10 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
BAUMILLER, T.K. & MACURDA, D.B. JR. 1995. Borings in Devonian and Mississippian Blastoids
(Echinodermata). Journal of Paleontology, 69/6: 1084-1089.
BOHATÝ, J. 2001. Wachstumsanomalien mitteldevonischer Crinoidenkelche aus den
Kalkmulden der Eifel (Rheinisches Schiefergebirge). Greifswalder
Geowissenschaftliche Beiträge, 9: 7-9.
Ý, J. 2005. Doppellagige Kronenplatten: Ein neues anatomisches Merkmal
paläozoischer Crinoiden und Revision der Familie Cupressocrinitidae (Devon).
Paläontologische Zeitschrift, 79/2: 201-225.
Ý, J. 2006. Neue Cupressocrinitidae (Crinoidea) aus den mitteldevo
BOHAT
BOHAT nischen
BRETT
DONO
FRANZ , 10: 219–234.
GALLE . Favositidae (Tabulata) from the Devonian of Bohemia. Sbornik
GALLE
ublic.
GLUCH
olonica, 50/2: 315-328.
nt iconibus et
rn GRAFEN
d
Co, Düsseldorf.
Kalkmulden der Eifel (linksrheinisches Schiefergebirge, Deutschland).
Senckenbergiana lethaea, 86/2: 151-189.
, C.E. & ECKERT, J.D. 1982. Paleoecology of a well-preserved crinoid colony from the
Silurian Rochester Shale in Ontario. Life Science Contribution, Royal Ontario
Museum, 131: 1-20.
VAN, S.K. & LEWIS, D.N. 1999. An epibiont and the functional morphology of the
column of a platycrinitid crinoid. Proceedings of the Yorkshire Geological Society,
52: 321-323.
DUBATOLOVA, YU. A. 1975. Devonian crinoids of the Minusin Basin [in Russian]. Trudy
Instituta Geologii i Geofiziki AN SSSR, Sibirskoe Otdelenie, 272: 1-58.
ÉN, C. 1977. Crinoid holdfasts from the Silurian of Gotland. Lethaia
GAHN, F.J. & BAUMILLER, T.K. 2005. Arm regeneration in Mississippian crinoids: evidence of
intense predation pressure in the Paleozoic?. Paleobiology, 31/1: 151-164.
, A. 1978
Geologických Věd, Paleontologie 20: 33-62.
, A. & PROKOP, R.J. 2000. Complex parasitism and symbiosis of crinoid, subepidermal
parasite, and tabulate coral. Lower Devonian (Pragian), Barrandian, Czech Rep
Věstnik Českého Geologického Ústavu, 75: 441-444.
OWSKI, E. 2005. Epibionts on upper Eifelian crinoid columnals from the Holy Cross
Mountains, Poland. Acta Palaeontologica P
GOLDFUSS, G.A. 1826-44. Petrefacta Germaniae tam ea, quae in museo universitatis
regiae Borussicae Fridericiae Wilhelmiae Rhenanae servantur, quam alia
quaecunque in Museis Hoeninghusiano, Muensteriano aliisque exta
descriptionibus illustrate (Abbildungen und Beschreibungen der Petrefacten
Deutschlands und der angrenzenden Länder, unter Mitwirkung des Her
GEORG ZU MÜNSTER, herausgegeben von Dr. AUG. GOLDFUSS) – 1 (1826-33): Divisio
secunda: Radiariorum Reliquiae – Strahlenthiere der Vorwelt, p. 115-221. Arnz an
24.-26. October 2008 11

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
HALLECK, M.S. 1973. Crinoids, hardgrounds, and community succession: The Silurian
Waldron−Laurel contact in southern Indiana. Lethaia, 6: 239-252.
CK, J.BLAMAR .P.A. de 1801. Systême des animaux sans vertèbres, ou tableau général des
n de leurs rapports naturels et
LAMAR maux sans vertèbres, présentant les
enfin, l'exposition
des principes fondamentaux de la zoologie, Vol. 2. Verdière, Paris, 568 p.
LE MENN, J. 1985. Les Crinoides du Dévonien inferieur et moyen du Massif armoricain.
Mémoires de la Societé géologique et minéralogique de Bretagne, 30: 1-268.
MEYER, D.L. & AUSICH, W.I. 1983. Biotic interactions among recent and among fossil
crinoids, p. 377–427. In M.J.S. TEVESZ & P.L. MCCALL (eds.), Biotic Interactions in
Recent and Fossil Benthic Communities. Plenum Press, New York.
MEYER, D.L. & OJI, T. 1993. Eocene crinoids from Seymour Island, Antarctic Peninsula:
paleobiogeographic and paleoecologic implications. Journal of Paleontology, 67/2:
250-257.
MILLER, S.A. & GURLEY, W.F.E. 1890. Description of some new genera and species of
Echinodermata from the Coal Measures and Subcarboniferous rocks of Indiana,
Missouri, and Iowa. Journal Cincinnati Society of Natural History, 13/1: 3-25.
MOORE, R.C. & JEFFORDS, R.M. (1968): Classification and nomenclature of fossil crinoids
based on studies of dissociated parts of their columns. The University of Kansas
Paleontological Contributions, 46: 1-86.
MOZZI, D., DOLMATOV, I.Y., BONASORO, F., & CARNEVALI, M.D.C. 2006. Visceral regeneration
in the crinoid Antedon mediterranea: basic mechanisms, tissues and cells involved
in gut regrowth. Central European Journal of Biology, 1/4: 609-635.
PETERS, S.E. & BORK, K.B. 1998. Secondary tiering on crinoids from the Waldron Shale
(Silurian: Wenlockian) of Indiana. Journal of Paleontology, 72: 887-893.
POWERS, B.G. & AUSICH, W.I. 1990. Epizoan associations in a Lower Mississippian
Paleocommunity (Borden Group, Indiana, U.S.A.). Historical Biology, 4: 245-265.
classes, des ordres et des genres de ces animaux; présentant leurs caractères
essentiels et leur distribution, d'après la considératio
de leur organisation, et suivant l'arrangement établi dans les galeries du Muséum
d'Hist. Naturelle, parmi leurs dépouilles conservées; précédé du discours
d'ouverture du Cours de Zoologie, donné dans le Muséum National d'Histoire
Naturelle l'an 8 de la République. Lamarck & Deterville, Paris, viii + 432 p.
CK, J.B.P.A. de 1816. Histoire naturelle des ani
caractères généraux et particuliers de ces animaux, leur distribution, leurs classes,
leurs familles, leurs genres, et la citation des principales espèces qui s'y rapportent;
précédée d'une introduction offrant la détermination des caractères essentiels de
l'animal, sa distinction du végétal et des autres corps naturels,
12 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
SCHLÜTER, C. 1885. Über einige neue Anthozoen aus dem Devon. Verhandlungen des
naturhistorischen Vereins der preussischen Rheinlande und Westphalens, 44: 114-
151.
RTS-DORECK, H. 1963. Über Missbildungen bei Cupressocrinus elongatus aus dem
Mitteldevon der Eifel. Decheniana, 115/2: 239-244.
s",
McKinney (eds.), 14th Meeting of the
International Bryozoology Association, 02.-07.07.2007, Abstracts with Program.
SIEVE
TAYLOR, P.D.T. & WILSON, M.A. 2007. Morphology and affinities of hederelloid "bryozoan
p. 88. In: S. J. Hageman and F. K.
Boone, North Carolina, 2007, <http: //www.iba.appstate.edu/IBAfiles/
AbstrastWprogram.pdf> (IBA Website, p. 88).
WACHSMUTH, C. & SPRINGER, F. 1889. Crinoids. In S. A. MILLER (ed.), North American
geology and paleontology. Western Methodist Book Concern, Cincinnati, 664 p.
WILSON, M. A. & TAYLOR, P.D.T. 2001. "Pseudobryozoans" and the problem of encruster
diversity in the Paleozoic. PaleoBios, 21/ 2 addendum: 134-135.
24.-26. October 2008 13

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Dental insights into ophiuroids: Feeding mechanisms and life style
(Poster)
KARIN BOOS
r and Marine Research, PO Box
g
ious studies have considered
phiuroids to be generally omnivorous macro- or microphageous feeders,
iuroids performing different lifestyles (epibenthic,
Biologische Anstalt Helgoland/ Alfred Wegener Institute for Pola
180, 27483 Helgoland; e-mail: [email protected]
Ophiuroid echinoderms are highly specific towards different habitats reflectin
lifestyles and feeding mechanisms. While prev
o
different feeding mechanisms have evolved according to an epibenthic or
infaunal lifestyle, e.g. predation, scavenging, deposit feeding or filter feeding.
Most ophiuroids have been reported to show more than one feeding mechanism
along with their main feeding habit. The presence and morphology of teeth, oral
and dental papillae from oph
infaunal or cryptic) was compared and discussed in relation to reported feeding
mechanisms and diets.
14 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Ökophänotypbildung der Muschelkalk-Crinoide Encrinus liliiformis
- Vergleich zweier Populationen aus Nord-Württemberg
(Vortrag)
JANINA F. DYNOWSKI1 & JAMES H. NEBELSICK2
1 Staatliches Museum für Naturkunde Stuttgart, Rosenstein 1, 70191 Stuttgart, Deutschland; e-
mail: [email protected]
2 Institut für Geowissenschaften, Eberhard Karls Universität Tübingen, Sigwartstraße 10, 72076
Tübingen, Deutschland; e-mail: [email protected]
Die vorliegende Arbeit untersucht die Ökophänotypbildung von Encrinus liliiformis
aus dem süddeutschen Muschelkalk (Mitteltrias) von Nord-Württemberg. Wie
schon von früheren Bearbeitern beobachtet wurde, scheinen sich Kronen aus
flacheren Meeresbereichen in ihrer Morphologie von solchen aus tieferen
Bereichen zu unterscheiden.
Um diese Unterschiede in der Morphologie der Kronen zu erfassen, wurden
zwei Populationen von Encrinus liliiformis miteinander verglichen, die aus
unterschiedlichen Gebieten des Germanischen Beckens stammen: Crailsheim
stellt einen Lebensraum auf einer flach einfallenden Karbonatrampe in geringer
Wassertiefe dar, während die Population aus Neckarwestheim aus einem
Lebensraum in tieferen Beckenbereichen stammt.
Die Vermessung der Kronen und die darauf folgende statistische
Auswertung der Daten führte zu einer Differenzierung von zwei verschiedenen
Ökophänotypen:
1) Kronen mit kürzeren Armen und weniger stark ornamentierter Oberfläche aus
geringerer Wassertiefe;
2) Kronen mit längeren Armen und stärker ornamentierter Oberfläche aus
größeren Wassertiefen.
Diese Unterschiede können als Anpassungen an Prädationsdruck und
unterschiedliche Strömungsverhältnisse interpretiert werden.
Der Prädationsdruck ist im Allgemeinen im flacheren Wasser höher als in
größeren Tiefen, weswegen hier eine Verstärkung des Körperbaus als Schutz vor
Fressfeinden erwartet wird. Während die Encrinus-Kronen aus Crailsheim
allerdings eine weniger stark ausgeprägte Ornamentierung zeigen als die Kronen
24.-26. October 2008 15

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
aus Neckarwestheim, sind die Arme der Crinoiden in Crailsheim kürzer als in
Neckarwestheim. Letzteres kann demnach durchaus in Zusammenhang mit
erschied im Prädationsdruck stehen.
Die Länge der Arme kann auch ein Hinweis auf die Strömungsstärke sein,
ie Crailsheim höher ist als in tieferen
,
acheren Lebensräumen. Bei geringerer Strömungsenergie wären für Crinoiden
Nähe der
einem Unt
die in flacheren Meeresbereichen w
Regionen. Damit sind Unterschiede im Nahrungsangebot verbunden, das in
tieferen Bereichen mit weniger Strömung niedriger ist als in küstennahen
fl
als passive Suspensionsfresser längere Arme, und somit ein größerer
Filtrationsfächer, von Vorteil, um mehr Nahrungspartikel aus dem Wasser filtern
zu können. Hier könnte auch die stärkere Ornamentierung einen Einfluss haben,
zum Beispiel durch eine Änderung im Strömungsregime in direkter
Krone, um das Wasser effektiver zu filtern.
16 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Stereomdifferenzierung in verschiedenen Seeigeln und ihr
bionisches Potenzial
(Vortrag)
NILS GROßMANN1 & JAMES H. NEBELSICK2
1, 2 Institut für Geowissenschaften, Eberhard Karls Universität Tübingen, Sigwartstraße 10, 72076
Tübingen, Deutschland; e-mail: [email protected], [email protected]
für Textil- und
rfah
er Arten und der Gruppen) und bezüglich ihres Habitats, also
hinsichtlich Temperatur, Bewegungsverhalten, Größe und Funktionen der
Stacheln, miteinander verglichen.
Die Stacheln, wie auch die Schalen von Echinodermen weisen eine poröse
Mikrostruktur (Stereom) aus Mg-Calcit mit geringen Einlagerungen (unter 1%)
von organischem Material auf. Diese sehr feste und trotzdem leichte Struktur ist
außerordentlich stabil und charakteristisch für die Gruppe der Echinodermen. In
Verbindung mit den sehnigen Fasern an der Basis der Stachel wird eine
zusätzliche Stabilität erreicht.
Zur Durchführung des Projekts werden mehrere Seeigelarten aus den
Familien Echinidae, Echinometridae und Cidaridae in eigens dafür geschaffenen
Aquarien unter realen Umweltbedingungen (Salzgehalt 36‰; 25°C etc.)
gehalten.
Anhand von CT- und REM-Aufnahmen wird die Stereomstruktur in den Stacheln
dokumentiert, auch die Kritische-Punkt-Trocknung wird verwendet. Diese
Datensätze können inzwischen bei Druck- und Biegeversuchen zur Interpretation
des strukturellen Versagens herangezogen werden. In weiteren Untersuchungen
Die Biomimetik (Bionik), die als relativ junges interdisziplinäres Forschungsgebiet
gilt, beschäftigt sich mit der Nutzbarkeit biologischer Phänomene bzw.
Eigenschaften für eine alltägliche technische Umsetzung. Als Beispiel dient der
Lotus-Effekt, welcher u. a. in der Automobilbranche Anwendung findet.
Das Projekt „Neue Materialien für leichte, stoffdurchlässige
Einschlagschutzsysteme - Seeigel als Modellsystem”, unter finanzieller Leitung
der Landesstiftung Baden-Württemberg, wird in Kooperation mit der AG
Angewandte Mineralogie (Universität Tübingen) und dem Institut
Ve renstechnik Denkendorf durchgeführt. Bisher wurden die morphologischen
Unterschiede in den Festigkeiten der einzelnen Stereomtypen untereinander
(innerhalb d
24.-26. October 2008 17

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
wurden die Magnesium- und Calciumgehalte an verschiedenen Positionen in den
gemessen und es zeigte sich, dass diese unterschiedlich sind.
Stacheln von Heterocentrotus mammillatus dienen dem Verkeilen im
Seeigeln, wie bspw. Eucidaris metularia
it
er Aufgabe der Stacheln steht, bleibt noch zu prüfen.
sten Bruchversuchen deutet
ich an
Stacheln
Riffgestein, während sie bei anderen
oder Plococidaris verticillata der Abwehr dienen. Ob die dabei festgestellte
Unterschiedlichkeit in der Mikrostruktur in einem direkten Zusammenhang m
d
Das Stereom ist im zentralen Bereich der Stacheln großporig und bei
Heterocentrotus mammillatus zusätzlich labyrinthisch, bei Plococidaris verticillata
gleichmäßig angeordnet. Die Kritische-Punkt-Trocknung zeigt innerhalb des
Stereoms eine geringe Anzahl von Weichteilen, die meist aus verschiedenen
Zelltypen und Fasern bestehen. Auch gibt es Hinweise, dass die Porenräume mit
einer dünnen organischen Schicht ausgekleidet sind. Anhand der
Computertomographie-Aufnahmen werden besonders bei Heterocentrotus
mammillatus die Wachstumsringe deutlich. Bei er
s , dass diese Verdickungen zur Stabilität beitragen können.
Im weiteren Projektverlauf werden auch etwaige Unterschiede in der
Morphologie der Stacheln bzw. Schalen zwischen Warm- und Kaltwasserarten
untersucht.
18 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Die Stereome diverser regulärer Seeigelstachel
(Poster)
NILS GROßMANN1 & JAMES H. NEBELSICK2
, 72076
e-mail: [email protected], [email protected]
der Riffrand- oder Riffdachbereiche bewohnen. Als
ethoden werden REM-Aufnahmen und zusätzlich das Verfahren Kritische-Punkt-
trukturen und keine
Wachstumsringe im Stereom besitzt.
Insgesamt weisen die Stacheln wie auch die Schalen von Echinodermen,
eine poröse Mikrostruktur (Stereom) aus Mg-Calcit mit geringen Einlagerungen
(unter 1%) von organischem Material auf.
Weiterhin erfolgen Untersuchungen in der Morphologie der Stacheln bzw.
Schalen zwischen Warm- und Kaltwasserarten.
1, 2 Institut für Geowissenschaften, Eberhard Karls Universität Tübingen, Sigwartstraße 10
Tübingen, Deutschland;
Das Forschungsziel ist eine detaillierte Darstellung der Stereomstrukturen ganzer
Seeigelstachel. Es werden Stacheln von diversen Arten der Familien Echinidae,
Echinometridae und Cidaridae in Bezug auf ihre Mikrostrukturen untersucht und
Habitatsfaktoren wie Größe der Stacheln, Funktionsweise und
Umgebungstemperatur verglichen. Hierbei eignen sich besonders Vergleiche
zwischen Arten, die entwe
M
Trocknung sowie Computer-tomografie verwendet.
Unterschiede zeigen sich bisher nicht nur zwischen den einzelnen Familien,
sondern auch zwischen den Arten. So weisen Heterocentrotus mammillatus und
Echinometra mathaei unterschiedliche Ausprägungen von Wachstumsringen auf,
während Phyllacanthus imperialis trotz ähnlichem Habitat wie Heterocentrotus
mammillatus eher gleichmäßig angeordnete S
24.-26. October 2008 19

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
30 Years of Research on Crinoids / 30 Jahre Crinoidenforschung
(Keynote)
HANS HESS
; Im Gerstenacker 8, 4102 Binningen, Schweiz; e-mail:
978 erschienen die drei Bände von Part T, Echinodermata 3 (Crinoidea) des
Naturhistorisches Museum Basel
1
Treatise on Invertebrate Paleontology. Die Revision unter Leitung von W. I.
AUSICH ist im Gang, und das Manuskript von Band 3 (Articulata, Mesozoikum -
heute) liegt nun vor. Die wesentlichen Fortschritte und Aenderungen in der
Systematik werden kurz vorgestellt und mit einigen Beispielen, insbesondere von
Trias-Crinoiden, illustriert.
The three volumes of Part T, Echinodermata 3 (Crinoidea) of the Treatise on
Invertebrate Paleontology were published in 1978. The revision under the
Editorship of W. I. AUSICH is ongoing, and the manuscript of the revised vol. 3
(Articulata, Mesozoic-Recent) is now available. Significant progress in
systematics has been made and the major changes are briefly presented,
illustrated by examples of Triassic crinoids and others.
20 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Pattern formation in starfish: arm stumps, regeneration models,
and evolution
(Poster)
FREDERICK H.C. HOTCHKISS
Marine and Paleobiological Research Institute, PO Box 1016, Vineyard Haven, MA 02568 USA; e-
mail: [email protected]
Objective: To interpret a Linckia laevigata and an Asterias rubens that have an
arm stump that healed and did not regenerate the missing arm (specimens from
nature). Method: Interpret using positional information theory and regeneration
models from amphibians, insects, and planarians. Observations: Wound closure
was by symmetrical midline joining through distal folding of both sides of the
arms. In contrast, in specimens that regenerate the arm, wound closure is by
downward sliding of the aboral surface. First finding: Failure of these arm stumps
to regenerate indicates symmetrical positional information in the starfish arm
nd indicates wound closure and healing along the line of symmetry. Thereby no a
positional value gaps/confrontations were formed, and intercalary gap-filling
regeneration was not stimulated. Second finding: The best fit model for starfish
arm regeneration is the distalization followed by intercalation model of AGATA et
al. (2003, 2007).
Proposals: It is proposed that arm regeneration is initiated by formation of a
istalized signaling center just behind the regenerated terminal plate. The signal
duces distal intercalary regeneration. It is further proposed that distal
tercalary gap-filling growth is the normal growth mechanism of the arm; thus
reestablishes the growth mechanism of intact arms. According to
and d the terminal plate; this
d
in
in
regeneration
this idea, there is always a positional gap between the last-formed section of arm
the distalized signaling center located just behin
gap is never bridged, and arm growth is indeterminate. Evo-Devo Extrapolation:
could cause the signaling center behind the terminal plate to induce Mutations
formation of additional or fewer series of plates in the arm. This may help to
explain the sudden origins and losses of novel plate series such as the sublateral
podial plates of Paleozoic Eopentaroida. Once the organism possesses this type
24.-26. October 2008 21

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
of signaling mechanism, then mutations could cause it to function in more places
than just behind the terminal plate. Thus plate series that intercalate in other
parts of the body wall could have this type of origin (such as asteroid actinal
intermediate plates, intermarginal plates, dorsolateral plates, etc.). Mosaic
evolution and recurrent appearance/loss of intercalary plates within and between
echinoderm lineages are expected under this proposal.
References
AGATA, K., T. TANAKA, C. KOBAYASHI, K. KATO & Y. SAITOH, 2003. Intercalary regeneration in
planarians. Developmental Dynamics 226: 308-316.
AGATA, K., Y. SAITO & E. NAKAJIMA, 2007. Unifying principles of regeneration I:
Epimorphosis versus morphallaxis. Development, Growth and Differentiation 49:
73-78.
22 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Bdellacoma in the Hunsrück Slate (Lower Devonian, Germany):
reidentification of Urasterella verruculosa (Asteroidea,
Bdellacomidae)
(Poster)
FREDERICK H.C. HOTCHKISS1 & ALEXANDER GLASS2
1 Marine and Paleobiological Research Institute, PO Box 1016, Vineyard Haven, MA 02568 USA; e-
Division of Earth and Ocean Sciences, Box 90227, Duke University, Durham, North Carolina
list on the fauna of the Hunsrück Slate has listed Bdellacoma. The
olution: In the Natural History Museum, London, are specimens BMNH E13627
rom the Hunsrück Slate labeled Bdellacoma. The specimens have
Bdell H specimens also match the Hunsrück
of the holotype of U. verruculosa confirmed the presence of bursulella-type
pedicella
is of with madreporiform
Bdell ea, Bdellacomidae). The fossil record
of Bdellacoma, including isolated pedicellariae reported as “Bursulella” ostracod
microfossils, ranges from Early Silurian to Early Carboniferous, with localities in
England, Gotland, Poland, Germany and USA. Improved understanding of
Bdellacoma (especially SUTTON et al. 2005) permits identifying specimens that do
not have pedicellariae, namely unpublished Bdellacomidae sp. collected by
THAYER (1972) from the Upper Devonian Genesee Group, Cortland County, New
York: Yale Peabody Museum 5-rayed YPM 211365 with bursulella-type
pedicellariae evident; YPM 204172-204186 with pedicellariae not evident but
otherwise resembling YPM 211365; and the remarkable YPM 211610 with 16 or
17 rays and preserving at least one bursulella-type pedicellaria. Occurrence of
Bdellacoma in the Lower Silurian (Llandovery) Gutterford Burn Starfish Bed,
mail: [email protected]
2
27708
The Problem: Occurrence of Bdellacoma SALTER, 1857, in the Hunsrück Slate was
mentioned by W. K. SPENCER (1940), but no specimens were cited. On the other
hand, no specia
S
and E13630 f
highly distinctive bursulella-type pedicellariae that confirm identification as
acoma sp. Next Problem: The BMN
Slate asteroid Urasterella verruculosa LEHMANN, 1957. The Solution: Examination
riae and establishes that it is a species of Bdellacoma. The madreporite
asteroid type: aboral, interradial, convex, large, and
markings. The Hunsrück Slate asteroid Urasterella verruculosa is reidentified as
acoma verruculosa n. comb. (Asteroid
24.-26. October 2008 23

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Pentland Hills, Scotland, was mentioned by SPENCER (1940: 529), but no
s were cited. Significance: Bdellacoma is confirmed present in the
unsrück Slate. Assignment of Bdellacoma to the Asteroidea by SUTTON et al.
rted, except that future research should investigate the sublateral
gements: The importance of papers by
OCZAROWSKI (2001) and by SUTTON et al. (2005) cannot be overstated. We thank
starfish with three-dimensionally preserved soft parts from the Silurian of England.
specimen
H
(2005) is suppo
plates described by SPENCER (1940) in Bd. vermiformis; sublaterals are not yet
recognized in Bd. verruculosa. Acknowled
B
D. N. LEWIS (BMNH), C. MACCLINTOCK (YPM), M. SANDER (Institute for
Paläontologie, Rheinische Friedrich-Wilhelms Universität, Bonn), and A. B. SMITH
(BMNH) for loans/visits; D. CLAIN, M. CLAIN, A. P. HOTCHKISS, M. SGAN and G.
SGAN for support; and D. NICHOLLS for composing the poster. The poster was
prepared for the 12th IEC, August 2006, Durham, New Hampshire; contribution
No. 23 of Project Nahecaris; contribution No. 2 of MPRI; this abstract is new for
this workshop.
References
BOCZAROWSKI, A. 2001. Isolated sclerites of Devonian non-pelmatozoan echinoderms.
Palaeontologia Polonica No. 59: 1-219.
SPENCER, W. K. 1940. A monograph of the British Palaeozoic Asterozoa, Part. X, pp.
495-540, pls. 33-37, Palaeontographical Society for 1940.
SUTTON, M. D, D. E. G. BRIGGS, DAVID J. SIVETER, DEREK J. SIVETER, & D. J. GLADWELL. 2005. A
Proceedings of the Royal Society, Series B 272: 1001-1006.
THAYER, C. W. 1972. Marine paleoecology of the Upper Devonian Genesee Group of New
York. Ph.D. dissertation, Yale University.
24 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Index of Living and Fossil Echinoids: 1971 - present
(Poster)
ANDREAS KROH
Naturhistorisches Museum Wien, Geologisch-Paläontologische Abteilung, Burgring 7, 1010 Wien,
Österreich; e-mail: [email protected]
In 1978 Porter M. KIER & Mary H. LAWSON published their "Index of Living and
Fossil Echinoids 1924-1970" which listed all species- and genus-level taxa
erected since the publication of the “Essai de Nomenclature Raisonnée des
Echinides” by Jules LAMBERT & Paul THIÉRY (1909-25). Both books have been an
outstanding resource for taxonomic and (palaeo-)biodiversity research and are
among the most consulted volumes by echinoid specialists. KIER & LAWSON (1978)
in the introduction of their Index announced a supplement for the years 1971-75
but this, apparently, has never been published.
rature for new taxa and by cross checking this list with the
e Paleobiology Database, the need for published indices
ay s
Data collection for such an supplement was started in 1999 by the present
author, while doing research on the biogeography of Cenozoic echinoids, but
soon was extended to include all epochs. The list was prepared by culling the
recent echinoderm lite
Zoological Record and various other offline and online sources. Each paper
included was consulted in the original. Citations and taxa before 1971 were
included if they were absent in LAMBERT & THIÉRY (1909-25) and KIER & LAWSON
(1978). In contrast to KIER & LAWSON (1978) the present index also includes
detailed locality data and repository of the types.
With the increasing availability of taxonomic databases such as the
Zoological Record or th
m eem obsolete. Many of these resources, however, are incomplete and/or
accessible for subscribers only. Despite the benefits of an online database, there
are many instances where an old-fashioned catalogue is quite handy, be it in the
storage area of a museum or during field work in remote places.
KIER, P.M. & LAWSON, M.H. 1978. Index of living and fossil echinoids 1924-1970. –
Smithsonian Contributions to Paleobiology, 34: 1-182.
LAMBERT, J. & THIÉRY, P. 1909-1925. Essai de Nomenclature Raisonnée des Echinides. –
iii+607 pp. Chaumont (L. Ferrière).
24.-26. October 2008 25

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Crinoids from the Tithonian of the Klippenbelt (Jurassic, Austria)
(Poster)
ANDREAS KROH1 & ALEXANDER LUKENEDER2
1, 2 rhistorisches Museum Wien, Geologisch-Paläontologische Abteilung, Burgring 7, 1010 Wien,
Österreich; e-mail: [email protected]
Natu
f strongly tilted, inverse, well-bedded marls and limestones of the
Blassenstein Formation. At the base marls and marly limestone bands dominate,
being replaced by increasingly pure limestones towards the top. Based on
preliminary data from ammonites, palaeomagnetics, nannofossils and
calpionellids (LUKENEDER, in press; PRUNER et al., in press; REHÁKOVÁ et al., in
press) the section comprises a continuous succession of Lower Tithonian to
Middle Berriasian strata.
Microfacies analysis of the more strongly lithified parts of the section
revealed high abundances of crinoidal remains, particularly in the lower part.
Based on cross section shape these fragments could be tentatively assigned to
saccocomid crinoids.
In order to obtain three-dimensional specimens of the crinoids, as well as
other microfossils commonly observed in the thin sections (namely foraminifera,
ostracods, rhyncholiths, small aptychi, ophiuroid remains, etc.) the marly parts
of the succession (i.e. the lower 8 m) were sampled intensely. Employing a
combination of traditional (hydrogene-superoxide) as well as specialized agents
(the tenside Rewoquat) it was possible to disaggregate the bulk samples and
clean the microfossils.
A first survey of the residues shows high abundances of the pelagic crinoid
Saccocoma tenella (GOLDFUSS, 1831) in the lower 6 metres of the section. The
crinoids are represented by isolated radial and brachial ossicles mainly. The
former being characterized by their arrow-head like shape with serrated edges
and coarse reticulate sculpture, the latter by its disc-like wings (in the proximal
brachials). Above, calyxes of the cyrtocriid Phyllocrinus belbekenis ARENDT, 1974
appear, while saccocomid remains become rare and vanish. Additionally, juvenile
columnals tentatively referred to the Margocrinus/Balanocrinus-group
sporadically occur in the lower part of the succession.
1, [email protected] 2
The Nutzhof section, 5 km north of Hainfeld, comprises an 18 metre long
succession o
26 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
In the upper part of the section bulk sampling was unsuccessful due to low clay
content and high lithification of the rocks. Thin sections, however, show that the
saccocomid-rich microfacies is replaced by pure calpionellid limestone, that can
eted as more distally situated pelagic facies.
Similar successions have been documented from other parts of Austria,
related to changes of current patterns, possibly
caused by geodynamically induced palaeogeographic changes and basinal
be interpr
Czech Republic and Poland as well. It is likely that the observed changes in
lithology and microfauna are
deepening.
24.-26. October 2008 27

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Temperature effect on feed consumption, absorption and
assimilation efficiencies and production in the sea urchin
locentrotus intermedius
oster)
ON L.
WATTS4
ail:
ood or absorption
efficiency. Temperature had little effect on assimilation efficiency except for a
significant decrease at 22 °C. The gonad index was consistently least at 22 °C.
Organic matter production in the gonads was greatest and earliest (April) at 12
and 17 °C. Because S. intermedius has a great ability to acclimate to
temperature, temperature should not be a great concern in land aquaculture.
Under the conditions of this experiment, S. intermedius in culture produced
gonads of marketable size (gonad index >15) within one month.
Strongy
(P
JOHN M. LAWRENCE1,*, XUEBIN CAO2, YAQING CHANG2, PING WANG2, Y. YU2, ADDIS
LAWRENCE3, STEPHEN A.
1 Department of Biology, University of South Florida, Tampa Florida 33620, U.S.A; e-m
la
2 Key Laboratory of Mariculture and Biotechnology, Dalian Fisheries University, 52 Heishijiao Street,
Dalian, Liaoning Province, 116023, People’s Republic of China
3 Texas Agrilife Research, Texas A&M System, 1300 Port Street, Port Aransas, Texas 78373, U.S.A.
4 Department of Biology, University of Alabama at Birmingham, Birmingham, Alabama 35294,
U.S.A.
Strongylocentrotus intermedius is one of the most economically important
species of sea urchins and has great potential for aquaculture. It is essential to
understand how temperature affects utilization of nutrients and production as
controlled temperature may be an important expense in land aquaculture and
affect optimization of production. We maintained large (5 cm test diameter) S.
intermedius in the laboratory and fed them a formulated feed from 28 February
when the gonads were small (gonad index, GI, ~10) and sea water temperature
was low (7 °C) until 27 June when sea water temperature was high (24 °C)
under five temperature treatments (environmental temperature, 7, 12, 17 and
22 °C. Temperature had little effect on rate of consumption of f
28 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Amount of arm loss and rate of arm regeneration by Luidia
(Echinodermata: Asteroidea)
Poster)
logy, University of South Florida, Tampa, Florida 33620, U.S.A.; e-mail:
.38 ± 1.97 mm.
l regenerated arms had mean lengths (±SE) of 10.96 ± 0.90,
t phase
f intact arms and of arms regenerating
clathrata
(
J.M. LAWRENCE1 & C.M. POMORY2
1 Department of Bio
la
2 Department of Biology, University of West Florida, Pensacola, Florida 32514, U.S.A.
Three contiguous arms of Luidia. clathrata were amputated. The initial mean
lengths (±SE) of the two intact arms were 57.36 ± 2.13 and 57
The initial mean lengths of the amputated arm stumps were 12.18 ± 0.59
(proximal), 25.91 ± 1.67 (medial) and 42.25 ± 1.37 (distal) mm. Buds appeared
on all amputated arms after approximately 8 days. After 54 days proximal,
medial and dista
7.69 ± 0.61 and 3.99 ± 0.38 mm, mean dry weights (±SE) of 41.75 ± 6.79,
20.95 ± 3.18, 8.07 ± 1.13 mg, and mean amounts of organic matter (pooled
samples) of 6, 3 and 1 mg, respectively. All three arm positions are statistically
different from one another (P<0.001). Appearance of arm buds is the firs
of arm regeneration and is independent of level of amputation. Growth of
regeneration is the second phase and is dependent on level of amputation.
Studies have reported rate of growth o
from the disc also declines as the asymptotic length of the arm is reached. This
suggests similar mechanisms of control of growth occur in all three situations, all
depending on the relative position of the regenerating arm tip.
24.-26. October 2008 29

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Seastars (Asteroidea) from Madagascar
(Vortrag)
HORST MOOSLEITNER
Organismische Biologie, Universität Salzburg, Hellbrunnerstr. 34, 5020 Salzburg, Austria; e-mail:
About a small collection of shallow water seastars from NW-Madagascar
The inspection of a small collection of seastars, Mr. Hinterkircher (Munich) had
sampled 2007 in NW-Madagascar, showed astonishing results:
The collection included not only seastars known from that region (LORIOL 1885),
like Protoreaster linckii (DE BLAINVILLE, 1834), Fromia milleporella (LAMARCK,
1816) and Monachaster sanderi (MEISSNER, 1892), but some species not known
from there, too.
One of them was Fromia indica (PERRIER, 1869). Its discovery is
this species was hitherto known only from places east of the
aldives and the Lakkadives (CLARK & ROWE 1971, MOOSLEITNER 1997, SASTRY
ese islands lie more than 3000 km away from Madagascar.
One spe
confirm to any species known. It was sent to Mr. O’L (Australia), a
specialist for tha
as the discovery of the seastar Ophidiaster
seast n from the Indian
arma of the two species was
applicable. The seastar from Madagascar fits not only to the key for that species,
but also in all important details to O. granifer. Exact details are given in the
presentation. O. granifer was hitherto known from the easternmost part of the
Indian Ozean and from the western–central Pacific (to Japan und to the island of
the South Pacific). This finding expands the range of occurrence across the whole
Indian Ocean to western Madagascar.
astonishing because
M
2007), and th
cimen of Aquilonastra sp. could not be classified as it did not
OUGHLYN
t genus, for inspection.
The most astonishing result w
granifer, LÜTKEN 1872. The determination first seemed to be impossible, as the
ar did not confirm to the description of any species know
Ocean. Frank ROWE gave me the crucial tip to include Ophidiaster granifer and O.
tus into my investigations. He was right, the first
30 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Ophidiaster granifer
References
C , A.M. & F.W.E. R , 1971. Monograph of Shallow-Water Indo-West Pacific LARK OWE
Echinoderms. Trusteet of the British Museum (Nat. Hist.), London. 238 pp.
DE LORIOL P., 1885. Catalogue Raisonne des Echinodermes recueillis par M.V.de ROBILLARD
a l’ile Maurice, II Stellerides. Mémoires de la Société de Physique et d'Histoire
Naturelle de Genève 29/4: 1-84.
MOOSLEITNER, H., 1997. Shallow water Asteroidea (Echinodermata) from the coral reefs of
the Maldive Islands. Revue française d'Aquariologie 24: 3-14.
SASTRY, D.R.K., 2007. Echinodermata of India: An annotated list. Records of the
Zoological Survey India, Occassional Paper 271: 1-387.
24.-26. October 2008 31

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Morphologie und Faziesverteilung des indo-pazifischen Seeigels
Jacksonaster depressum
(Vortrag)
d. Diese Seeigel sind kaum untersucht worden und
pikalsystem, Petalodium, Lage und Ausmaß des Peristoms
JAMES NEBELSICK1 & ATEF ABD EL-HAMIED A. S. ELATTAAR2
1 Institut für Geowissenschaften, Universität Tübingen, Sigwartstrasse 10, 72076
Tübingen, Deutschland; e-mail: [email protected] 2 Geology Department, Sohag Faculty of Science, Sohag University, Sohag, P. O.
Box 82524, Egypt; e-mail: [email protected]
Jacksonaster depressum ist ein irregulärer indopazifischer Seeigel der Ordnung
Clypeasteroida. Das bearbeitete Material stammt aus der Nördlichen Bucht von
Safaga, Rotes Meer, Ägypten, wo dieser Seeigel am häufigsten in schlammigen
Sanden in Tiefen von 20 – 50 Meter anzutreffen ist. Der Seeigel erreicht Größen
von bis zu 3 cm und zeigt die für diese Ordnung charakteristische abgeflachte
Schale mit internen Stützelementen, die in dieser Gattung auf den äußeren Rand
des Tieres beschränkt sin
leben wahrscheinlich infaunal als Detritivor.
Wir haben die Skelettmorphologie, Taphonomie und ökologische Verteilung von
zahlreichen Exemplaren innerhalb des Untersuchungsgebietes studiert. Hierbei
wurden sowohl lebende wie auch tote Schalen inkludiert, die durch verschiedene
Methoden aufgesammelt worden sind. Vermessungen von spezifischen
morphologischen Merkmalen wurden durchgeführt, z.B. die allgemeinen
Schalenparameter, A
und Periprokts usw.. Außerdem wurden REM - Aufnahmen von Stacheln,
Pedicellarien, Tuberkeln wie auch von den Poren gemacht. Zusätzlich wurden
Differenzierungen im Stereom notiert. Die Variation dieser morphologische
Parameter zwischen lebenden und toten Tiere wurde analysiert, und zwar
innerhalb der Gesamtpopulation, wie auch zwischen unterschiedlichen
Probenpunkten und Faziestypen.
32 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Taphonomie von Jacksonaster depressum (Echinoidea:
Clypeasteroida) aus dem Roten Meer
(Poster)
JAMES NEBELSICK1 & ATEF ABD EL-HAMIED A. S. ELATTAAR2
2076
gen.de
Geology Department, Sohag Faculty of Science, Sohag University, Sohag, P. O.
e Zerstörung der Schale könnte sowohl an „kompletten“
au
rozesse
1 Institut für Geowissenschaften, Universität Tübingen, Sigwartstrasse 10, 7
Tübingen, Deutschland; e-mail: nebelsick@uni-tuebin2
Box 82524, Egypt; e-mail: [email protected]
Jacksonaster depressum ist ein irregulärer indopazifischer Seeigel der Ordnung
Clypeasteroida. Das bearbeitete Material stammt aus der Nördlichen Bucht von
Safaga, Rotes Meer, Ägypten, wo dieser Seeigel am häufigsten in schlammigen
Sanden in Tiefen von 20 – 50 Meter anzutreffen ist. Dieser Seeigel zeigt die für
diese Ordnung charakteristische abgeflachte Schale mit internen Stützelementen,
die für die Erhaltung signifikant sind. Die Taphonomie dieser Seeigel ist bis jetzt
kaum untersucht worden.
Die taphonomisch
als ch an Fragmenten erfolgen. Die taphonomische Merkmale können anhand
von der Veränderungen von Oberflächenmerkmalen wie Tuberkulation oder
Stereomtypen charakterisiert werden. Eine Vielzahl von taphonomischen
Prozessen konnte beobachtet werden, u.a. Disartikulation, Fragmentation,
Inkrustation wie auch Bioerosion. Angriffe durch Räuber spielen auch eine
wichtige Rolle bei der Zerstörung der Schale und der Produktion von
Schalenfragmenten.
Eine Reihe von Fragen stellen sich bezüglich der Taphonomie von
Jacksonaster: 1) Wie beeinflusst die Morphologie von Jacksonaster die
beobachteten taphonomischen Signaturen? 2) Welche taphonomischen P
sind für die Zerstörung wie auch Erhaltung dieser Gattung von Bedeutung, und
3) wie ist das Erhaltungspotential dise Seeigel in Vergleich zu anderen
Clypeasteroiden des Roten Meeres, wie Clypeaster, Echinodiscus, Fibularia und
Echinocyamus? Charakteristisch für Jacksonaster ist z.B. ihre Erhaltung als
ringförmige Schalenreste, ein Erhaltungsform, die bei den anderen oben
aufgeführten Seeigeln kaum in Erscheinung tritt.
24.-26. October 2008 33

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Skelett-Endosymbionten fossiler und rezenter Echinodermen: Von
Bioklaustrationen zu komplexen Strukturen
(Vortrag)
CHRISTIAN NEUMANN
Museum für Naturkunde, Humboldt Universität Berlin, Invalidenstrasse 43, D-10115 Berlin,
Deutschland; e-mail: [email protected]
Als Symbiose wird das Zusammenleben von Organismen unterschiedlicher
Artzugehörigkeit bezeichnet. Evolutionsbiologisch sind Symbiosen sehr
interessant, denn sie erfordern in der Regel einen hohen Grad an Spezialisierung
mindestens bei einem der beiden Partner. Verschiedene Fragen tun sich auf: Wie
sind diese oft komplexen Wirt/Symbiont-Systeme entstanden? Wie haben sich
Symbiosen über geologische Zeiträume verändert? Inwiefern sind dabei Prozesse
der Koevolution beteiligt? Fragen, die bisher schwer zu beantworten waren, denn
Symbiosen sind im Fossilbericht kaum überliefert. Ausnahmen sind solche
Interaktionen, die Spuren an den erhaltungsfähigen Hartteilen (Schalen,
Skelette) hinterließen.
auf ihrer
Körperoberfläche festgesetzt haben, mit ihrem Skelettgewebe zu überwachsen
(Neoplasie, Callus-Bildung). Diese Abwehrreaktion machen sich manche
Symbionten zu Nutze: Das „Überwuchert-werden“ durch das Kalzitskelett des
Wirtes bietet Vorteile, wie etwa einen besseren Schutz vor Fressfeinden.
So sind Crinoiden bevorzugte Wirte von Symbionten. Als passive Filtrierer
besitzen sie einen Stiel, der eine günstige Position des Fressapparates innerhalb
der Wasserströmung gewährleistet. Für andere filtrierende Organismen bilden die
Stiele eine willkommene Struktur, um sich darauf festzusetzen und so
„kostenlos“ in die Meeresströmung zu gelangen(„secondary tiering“). Als
Abwehrreaktion versucht die Seelilie oftmals, die unerwünschten Untermieter mit
ihrem Skelett geschwulstartig zu überwachsen. Bereits im Paläozoikum passen
Echinodermen bieten sich modellhaft für die Untersuchung fossiler
Symbiosen an, denn bei ihrem Innenskelett handelt es sich um ein lebendes
Gewebe, welches direkt und plastisch auf Irritationen (wie etwa einen
Parasitenbefall) reagieren kann. Diese Gewebeveränderungen sind fossil
erhaltungsfähig und somit vom Paläontologen interpretierbar. So versuchen
Stachelhäuter oft, unerwünschte Fremdorganismen, die sich
34 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
sich Vertreter unterschiedlichster taxonomischer Zugehörigkeit an diese
Wirtsreaktion an und gehen zu einer endosymbiontischen Lebensweise über. Um
ändige Einkapselung durch das Wirtsgewebe zu verhindern, bedarf es
ewisser Anpassungen von Seiten des Symbionten. So entstanden schließlich aus
ett-Einbettungen (Bioklaustrationen) hochkomplexe gallenartige
zen.
und Funktionsweise solcher Gallen lässt sich beispielhaft an
allen parasitischer Schnecken (Fam. Eulimidae) in den Stacheln von
icht überlieferten
eine vollst
g
einfachen Skel
Strukturen, welche physiologischen Prozesse des Symbionten unterstüt
Die Bildungs-
G
Lanzenseeigeln(Cidaroida) untersuchen: Hier nimmt der Parasit wesentlichen
Einfluss auf die Art und Weise der Gewebereaktion seines Wirtes. Je nach
Schneckenart sind die Gallen in ihrer Struktur höchst unterschiedlich und oftmals
hochkomplex. Es ist anzunehmen, dass die Morphologie der Gallen vom Genom
der Schnecken kontrolliert wird, obwohl die Galle selbst eine Bildung des Wirtes
ist („extended phenotype“). Sollte diese Hypothese zutreffen, offenbart sich darin
die Chance, anhand fossiler Gallen die Phylogenie ihrer n
Bewohner zu rekonstruieren und schließlich durch den Vergleich mit der
Phylogenie ihrer Wirte koevolutionäre Prozesse (z.B. Kospeziation)
nachzuweisen.
Mit Hilfe Computer-tomographischer Methoden wurden dreidimensionale
Einblicke in die Internstruktur des Skeletts befallener fossiler und rezenter
Echinodermen gewonnen, ohne dass dabei die Objekte zerstört werden mussten.
Dabei wurde sowohl Neutronen- als auch hochauflösende Synchrotronstrahlung
eingesetzt und spektakuläre Einblicke in das verborgene Innenleben längst
vergangener Lebensgemeinschaften gewonnen.
24.-26. October 2008 35

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Echinoids from the Neogene of Portugal mainland: Systematics,
Palaeoecology, Palaeobiogeography
(Vortrag)
PEDRO PEREIRA
Universidade Aberta, Rua Fernão Lopez, 9, 1° Esa, 1000-132 Lisboa, Portugal; e-mail:
The Portuguese Neogene echinoid fauna is reviewed in order to establish a
taxonomically standardised echinoid database. 41 taxa, including one new genus
and new species, from the Burdigalian and Middle Miocene of the Lower Tagus
Basin and Middle Miocene of Algarve and two taxa from the Middle Pliocene of
Mondego Basin are described. 25 taxa are reported for the first time from
Portugal mainland, while 23 of the previously reported Miocene species were
excluded (for different reasons) from the accepted species list.
This study uses echinoid diversity and the present geographic distribution
of genera that already existed during Neogene to assess climatic changes in the
Neogene of Portugal.
The presently available data suggest tropical conditions for the Burdigalian
(Neogene Cycle I) and Lower Langhian (Early Neogene Cycle II) of the Lower
Tagus Basin, followed by a temperature drop, coincident with the end of the
Upper Burdigalian-Lower Langhian Climatic Optimum. This temperature drop is
reflected by the reduction of the echinoid diversity, the disappearance of several
species, particularly among clypeasteroids and cassiduloids, and by the
appearance and following dominance of euechinoids (mainly, Psammechinus)
and Echinocyamus.
In Algarve the echinoid data suggest, for the Middle Miocene (Neogene
Cycle II), tropical conditions for that region and, for the Upper Tortonian
(Neogene Cycle III), probably subtropical conditions.
During the Neogene Cycle II there was a clear temperature gradient between
Algarve (warmer) and Lower Tagus Basin (less warm) which is obviously related
to the more southerly position of Algarve.
For the Middle Pliocene (Neogene Cycle III) of the in Mondego Basin the
echinoid data are two few to allow a palaeoclimate interpretation, but gastropod
data suggest subtropical conditions.
36 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
The echinoid fauna of Portugal mainland is closely related to that of the
Mediterranean region. In fact, the biogeographic investigation of the Portuguese
una shows that a major part of the Portuguese species is composed by
migrants from the Mediterranean area (42.9% of the fauna in the Burdigalian
ecies not exceeding 25% of total Portuguese echinoid fauna.
echinoid fa
im
and 60.9% in the Middle Miocene). Endemism is low during Miocene, with
endemic sp
24.-26. October 2008 37

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Phylogenetic relationships within Echinodermata based on
mitochondrial sequence data
(Vortrag)
MARLEEN PERSEKE1, GUIDO FRITZSCH2, DETLEF BERNHARD1, PETER F. STADLER2 & MARTIN
Interdisciplinary Center for Bioinformatics, University of Leipzig, Haertelstrasse 16-18, 04107
the ophiuroids are strongly
idea becomes
extended dataset for phylogenetic
analyses, the Echinodermata and all five echinoderm groups are clearly
recovered as monophyletic groups. Within the Echinodermata only the
relationships between Echinoidea, Asteroidea, and Holothuroidea are unstable
depending on the analyses method used. Most analyses show Holothuroidea as
sistergoup of a clade consisting of Echinoidea and Asteroidea. However, the basal
branching of Ophiuroidea within echinoderms persists in all analyses with strong
support and keeps the main problem within the mitochondrial dataset.
SCHLEGEL1
1 Institute of Biology II, Molecular Evolution and Animal Systematics, University of Leipzig,
Talstrasse 33, 04103 Leipzig, Deutschland; e-mail: [email protected]
2
Leipzig, Deutschland
Reconstruction of phylogenetic relationships within Echinodermata seems to be
difficult using mitochondrial datasets. Especially
different compared to the other echinoderm groups. However, only two complete
mitochondrial gene orders of ophiuroids are known. In our analyses this dataset
was expanded to a total of six mitochondrial genomes of ophiuroids with 3 new
families: Ophiocomidae, Amphiuridae and Gorgonocephalidae. With these new
mitochondrial gene orders, a consensus gene order of Ophiuro
apparent with only moderate rearrangement events within this group.
Additionally, new complete mitochondrial genomes from other echinoderm
groups were determined: an irregular echinoid and a further holothuroid. Both
mitochondrial genomes show no gene order rearrangements compared to the
known gene order of echinoids.
Using the amino acid sequences of this
38 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
New insights into early diversification of sea cucumbers as
inferred from calcareous ring material (Ordovician/Silurian,
ndia)
Vortrag)
n & Geopark,
oldschmidtstr. 1-5, 37077 Göttingen, Deutschland; e-mail: [email protected]
g tissue.
as stem group representatives.
well as providing
Baltosca
(
MIKE REICH
Geowissenschaftliches Zentrum der Universität Göttingen, Museum, Sammlunge
G
Compared to other modern Echinodermata, the early evolutionary history of
holothurians is poorly understood. In part, this is due to their disjunct
endoskeleton with ossicles and calcareous ring elements, which are released
following decomposition of the surroundin
Newly sampled Silurian sections on Gotland, Sweden, yield well-preserved
echinozoan echinoderms, including more than 1000 isolated calcareous ring
elements. The studied material is Telychian to Ludfordian (Early to Late Silurian)
in age and reveals the presence of several new taxa of arthrochirotid (extinct)
and apodid sea cucumbers as well
The calcareous pharyngeal ring (CR) is a defining feature of sea cucumbers
and is possessed by nearly all extant species. The CR supports the pharynx, the
water vascular system and tentacle ampullae in part, as
insertion points for the muscles. Therefore calcareous rings are distinct for
systematics and phylogeny, and important evolutionary structures.
The new material shows, that the Holothuroidea diversified significantly
through Late Ordovician and Silurian times.
24.-26. October 2008 39

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
A nearly articulated holothurian from the Santonian (Cretaceous)
of England
(Poster)
M EICH
Geowissenschaftliches Zentrum der Universität Göttingen, Museum, Sammlungen & Geopark,
Goldschmidtstr. 1-5, 37077 Göttingen, Deutschland; e-mail: [email protected]
With the exception of the Maa
IKE R
strichtian, only scarce data exist on fossil
ontsec area, Spain; both referred to the
podida and Dendrochirotida.
port the discovery of a nearly articulated holothuroid specimen
exac
ossic
likely to the family Holothuriidae.
ristic (Mertensiothuria), H.
(Microthele), and H. (Semperothuria) and H. (Theelothuria).
According to the decomposition of the surrounding tissue after death of the
animal, only a part of the former body skeleton is preserved; the anterior part of
the skeleton (calcareous ring) is missing. Due to the well-preserved body-wall
ossicles and the formation of the ossicles of the new find, I can exclude a
digestion and excretion of the sea cucumber by any predator.
After comparison with modern members of the Holothuriidae, we can
probably speculate that the body of our new Santonian species was also
cylindrical, elongate and scattered with numerous podia over the entire body,
dorsally as papillae, ventrally as cylindrical tube feet.
A wider distribution of the new genus / species can also be assumed,
because comparable long button-shaped sclerites were also known from Early
and Late Maastrichtian chalk sediments of the islands of Møn (Denmark) and
Rügen (NE Germany) as well as the Baltic Sea.
holothurians from the Upper Cretaceous, especially the Santonian. Up to now,
only two records have been mentioned worldwide: (1) from the Münsterland
area, Germany, and (2) from the M
A
Here I re
from the “Upper Chalk” (Santonian; BMNH E 48764) of England (unfortunately
t locality unknown). The unique specimen shows two different types of
les: (A) tables and (B) buttons, demonstrating aspidochirote affinities, very
Concerning the morphology of the buttons the new find share characte-
s with tube feet and papillae ossicles of Holothuria
40 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Holothuriid sea cucumbers are globally distributed and primarily deposit-feeders.
Modern members of the Holothuriidae inhabit the tropical eulittoral, usually coral
adjacent sandy areas; only a few species were known from temperate
hallows and deeper waters (bathyal depths).
dering how important aspidochirote sea cucumbers are today (nearly
y
tly investigated
y KERR et al. (2005) and SAMYN et al. (2005), unfortunately without detailed
SAMYN, Y., APPELTANS, W. & KERR, A.M., 2005. Phylogeny of Labidodemas and the
irotida) as inferred from morphology.
Z
reefs and
s
Consi
¼ of all known modern species), their fossil record is meagre and poorl
understood. The phylogeny of the family Holothuriidae was recen
b
consideration to the fossil record.
The origin and early diversification of the Holothuriidae can be located in
the Late Palaeozoic. There is still a hidden Mesozoic diversity since detailed
investigations are missing or sparse.
References
KERR, A.M., JANIES, D.A., CLOUSE, R.M., SAMYN, Y., KUSZAK, J. & KIM, Junhyong, 2005.
Molecular Phylogeny of Coral-Reef Sea Cucumbers (Holothuriidae: Aspidochirotida)
Based on 16S Mitochondrial Ribosomal DNA Sequence. Marine Biotechnology 7/1:
53-60.
Holothuriidae (Holothuroidea: Aspidoch
oological Journal of the Linnean Society 144/1: 103-120.
24.-26. October 2008 41

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Unusual holothurians (Echinodermata) from the Late Ordovician of
oster)
Geopark,
gen, Deutschland; e-mail: [email protected]
his study shows that the Holothuroidea diversified significantly through Late
rdovician times.
Sweden
(P
MIKE REICH
Geowissenschaftliches Zentrum der Universität Göttingen, Museum, Sammlungen &
Goldschmidtstr. 1-5, 37077 Göttin
Compared to other modern echinoderms, the early evolutionary history of sea
cucumbers is poorly understood. In part, this is due to their disjunct
endoskeleton with ossicles and calcareous ring elements, which are released
following decomposition of the surrounding tissue.
Newly sampled öjlemyr flints from the northwestern part of Gotland,
Sweden, yield well-preserved echinozoan echinoderms, including holothurian
ossicles. The studied material is Ashgill (upper Pirgu stage) in age and reveals
the presence of several new or poorly known taxa of elasipodid, aspidochirote,
and apodid holothurians or stem group representatives. The minute ossicles,
around 70–400 µm in length / diameter, exhibit an impressive morphology with
arms, spires, perforations, teeth etc.
The new material differs from previous described Palaeozoic, Mesozoic and
modern material by unusual symmetry.
T
O
42 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Signs of predation on Middle Triassic crinoids form Poland
(Poster)
MARIUSZ A. SALAMON1 & PRZEMYSŁAW GORZELAK2
1, 2 University of Silesia, Faculty of Earth Sciences, Department of Palaeontology & Biostratigraphy,
Bedzinska Str. 60, 41-200 Sosnowiec, Poland; e-mail: [email protected] ,
Observations of both extant and fossil crinoids indicate that the range of their
natural predators includes fishes, arthropods, asteroids and echinoids (Baumiller
et al., 2008 and the references cited therein). Using large samples of
disarticulated fossil crinoids from numerous Middle Triassic localities, we looked
for evidence of bite marks produced by predators. A large proportion of
columnals possessed characteristic scratches and pits imprinted in stereom.
Some of these signs are very similar to those found on Late Jurassic and Late
Cretaceous crinoids and ascribed to predation (comp. JAGT 1999, pl. 35, figs. 5,
10; JAGT & SALAMON 2007, pl. 3, fig. 4; GORZELAK & SALAMON in press, fig. 2).
These scratches are relatively shallow (0.1-1.3 mm deep), but clean, cuts in the
tereom of the ossicle surface. They are 0.1-1.1 mm wide and up to 2.1 mm
are usually perpendicular to the surface of the ossicle and their
s
long. The pits
outline is circular or slightly oval. Their diameter varies form 0.15 to 0.7 mm.
A,B – bite marks (dotted line) and C – traces of regeneration (arrow) on Middle
Triassic crinoids from Poland [Gogolin Beds (Lower Anisian, Muschelkalk); Upper
Silesia area). Scale bars equal 1mm.
24.-26. October 2008 43

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
They are relatively shallow (0.2-1.4 mm) and taper towards the bottom. In most
ossicles these marks occur on the external part of the lateral faces of well
columnals/pluricolumnals which are distict from abrasion suggesting
at the crinoid material was fresh and probably live, when the marks were made
s evidence of predation. This is also consistent with the fact that
hological
nomalies and regeneration traces.
e crinoids that
ds and holocrinids),
34/1: 22-34.
s
he southeast Netherlands and northeast Belgium – Part 2: Crinoids. -
Scripta Geologica, 116: 59-255.
id crinoids from
southern Poland – preliminary observations. - Scripta Geologica, 134: 61-76.
preserved
th
and we treat it a
certain ossicles, which possess bite marks, comprise also some pat
a
It has been argued that predation increases over geological time should
result in increases in defensive adaptations in prey taxa. It seems that most of
the major functional "experiments" in post-Paleozoic articulat
occurred during the Triassic can be described in terms of predation including
planktonic microcrinoids (roveacrinids), pseudoplanktonic stalked crinoids
(traumatocrinids and pentacrinids), strongly anti-predatory specialised
cyrtocrinids, stalk and arm shedding benthic crinoids (isocrini
free moving crinoids (paracomatulids). The high bite mark frequency on Early
Mesozoic crinoids from Poland proves that the evolution of morphologies and
behaviors of the above mentioned groups was stimulated by their interactions
with predators.
References
BAUMILLER, T.K., MOOI, R. & MESSING, C.G. (2008): Urchins in the meadow: paleobiological
and evolutionary implications of cidaroid predation on crinoids. - Paleobiology,
GORZELAK, P. & SALAMON, M.A. (in press): Signs of benthic predation on Late Jurassic
talked crinoids; preliminary data. - Palaios.
JAGT, J.W.M. (1999): Late Cretaceous-Early Palaeogene echinoderms and the K/T
boundary in t
JAGT, J.W.M. & SALAMON, M.A. (2007): Late Cretaceous bourgueticrin
44 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Are that sea urchins ???? - The strange morphology of the echinoid
e deep-sea.
During the Challenger-Expedition 1873 – 1876 most of them were dredged
The typical features for all species are a deep peristomial groove
expressed in juveniles, but
ecome progressively more and more obvious during ontogeny. These changing
made explanations on plate homologies rather problematic.
ry (EAT) by MOOI et al. (1994), DAVID & MOOI
for new
interpretations.
family Pourtalesiidae
(Vortrag)
HEINKE SCHULTZ
Dorfstr. 32, 25485 Hemdingen, Deutschland; e-mail: [email protected]
The family consists of species with very strangely transformed architectures
looking like a bottle or a vase rather than a pentameral echinoid. At least seven
extant genera belong to the pourtalesiids, living in th
for the first time and were described by A. AGASSIZ (1881). LOVÉN (1883) and
MORTENSEN (1907, 1950) covered the pourtalesiids exhaustively – and all studies
based on a very small number or even on a single specimen.
Russian research expeditions have brought new specimens of abyssal
species, which were described by A. MIRONOV in the seventies and nineties. This
year (2008) he wrote a publication on echinoids from the Mid-Atlantic Ridge,
which were dredged at depths of about 620 to 3460 m.
anteriorly, a disjunct apical system and on the oral side an interrupted
interambulacrum IA 5. The strange shapes are not
b
plate patterns
The Extraxial-axial Theo
(1996, 1998), including the Ocular Plate Rule, was the base
Two areas on the surface of pourtalesiids are of special interest:
1. The apical system on the aboral side: The holasteroid apical system is typically
ated, but compact. Behielong nd the single ocular plate OIII follow towards the
OII), sterior oculars (OI
posterior two genital plates (G2 + G3), then the two anterior oculars (OIV and
the two posterior genital plates (G1 + G4) and the po
and OV).
24.-26. October 2008 45

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
scheme is modified in the members of the pourtalesiids:
e genus Echinosigra there are only gonopores
This
In th in the two anterior genital
perfo y stay in contact with the posterior oculars.
apica
two perforating the following ocular plates. The posterior genital plates are
separated and without function, and the posterior ocular plates are again
separated from the genitals and from each other.
2. The plastron on the oral side
plates, but the posterior genitals shift towards the posterior. They are not
rated by a pore and the
In the genus Pourtalesia the adult specimens display another “model” of
l pattern: There are four gonopores, two in the anterior genital plates and
: The holasteroid plastron (interambulacrum IA 5)
is usually composed by a labral plate, followed by a single sternal plate and then
two episternal plates. In very young pourtalesiids this pattern is still present, but
during ontogeny the adjacent ambulacral plates shift towards the midline
separating the sternum from the labrum. In adult Pourtalesia (and Cystocrepis)
the ambulacral plates of each column are positioned one in front of the other
pushing the labrum towards the anterior into the peristomial groove.
Detailed and illustrated descriptions of the pourtalesiid species will be displayed
in the book: Sea urchins of the deep-sea (in prep.).
References
(more references in the book)
AGASSIZ, A., 1881. Report on the Challenger-Expedition. Zool. 3(9): 1-321.
DAVID, B. 1987. Dynamics of plate growth in the deep-sea echinoid Pourtalesia miranda
Agassiz: a new architectural interpretation. Bulletin of Marine Science 40(1): 29-
47.
LOVÉN, S., 1883. On Pourtalesia. A genus of Echinoidea. Kongl. Svenska Vetenskaps-
Akademiens Handlingar 19/7: 1-95.
MIRONOV, A.N., 2008. Pourtalesiid sea urchins of the northern Mid-Atlantic Ridge. Marine
Biology Research 4: 3-24.
MOOI, R. & DAVID, B., 1996. Phylogenetic analysis of extreme morphologies: deep-sea
holasteroid echinoids. Journal of Natural History 30: 913-953.
MOOI, R. & DAVID, B. 1998. Evolution within a bizarre phylum: Homologies of the first
echinoderms. American Zoologist 38: 965-974
46 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
MOOI, R., DAVID, B. & MARCHAND, D., 1994. Echinoderm skeletal homologies.: Classical
morphology meets modern phylogenetics. Proc. 18th Intern. Echinoderm
, Dijon 1994. A.A. Balkema, Rotterdam.
MORTENSEN, T., 1907. The Danish Ingolf-Expedition. Vol. IV Echinoidea (Part II). - 200
pp., Copenhagen.
. A Monograph of the Echinoidea. V, 1. Spatangoida I. Protosternata,
eridæ, Hemiasteridæ. – 432 pp., Copenhagen (C. A. Reitzel).
Conference
MORTENSEN, T., 1950
Meridosternata, Amphisternata I. Palæopneustidæ, Palæostomatidæ, Aëropsidæ,
Toxasteridæ, Micrast
24.-26. October 2008 47

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
The early history of echinoderms: new insights from disarticulated
d middle Cambrian ossicles.
eynote)
eum, Cromwell Road, London SW7 5BD, UK;
ur current view of the early history of echinoderms is based largely on
early an
(K
ANDREW B SMITH
Department of Palaeontology, The Natural History Mus
e-mail: [email protected]
O
articulate specimens from Cambrian deposits of North America and Europe.
However, Cambrian deposits in Australia and North Africa that are rich in
echinoderm debris suggests this is a very incomplete picture and provide
important glimpses into an unseen diversity. Furthermore this material is often
exquisitely preserved, allowing the detailed study of skeletal histology and
reconstruction of the soft-tissue anatomy of primitive echinoderms. Middle
Cambrian columnals show unexpected diversity and provide information on the
ligamental structure of the stem, while Australian material includes various
elements of a pelmatozoan with uniserial arms. These finds are leading to a
reinterpretation of distinction between Crinozoa and Blastozoa and the earliest
history of the Pelmatozoa.
48 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Rhaetian ophiuroids from the Netherlands: a preliminary report
(Poster)
BEN THUY1, ADIËL A. KLOMPMAKER2 & JOHN W. M. JAGT3
eowissenschaften, Eberhard Karls Universität Tübingen, Sigwartstraße 10, 72076
3 Natuurhistorisch Museu
Nethe
For the first t
Winterswijk, eastern Netherlands. Strata excavated at the quarry comprise
mainl
their body fossils and traces. Recently, a subrosion pipe fill
early
in prep.). Also, similar, early to middle Rhaetian dark shales occur in situ in the
prep ollected from both shale
appearances of the quarry by splitting the soft slabs. Specimens generally are
rare and occur individually or in groups of three to four individuals.
The material consists of more than 15 complete and articulated specimens
and disc fragments, plus some 30 arm fragments. Specimen size (disc diameter)
ranges from about 1 to 6 mm. Most specimens are at least partially pyritised.
Compaction and recrystallisation, however, tend to blur morphological details of
many specimens. To make matters worse, small gypsum needles and
encrustations formed some weeks after recovery of the slabs, preferably on the
ophiuroid remains. However, in thoroughly pyritised specimens, brief exposure to
a 20% H2O2-solution turned out to be succesfull - disc and arm fragments could
be freed from the matrix and from thin clayey encrustations. This preparation
method leaves specimens prone to fragmentation but it does render them
perfectly exposed and suitable for SEM-examination.
The prepared specimens exhibit enough morphological detail to be
taxonomically assessed. First observations suggest that the faunule is
monospecific. There are affinities with species of the genus Aplocoma D’ORBIGNY,
1852, and ongoing investigations aim at a precise taxonomic assignment and
1, 2 Institut für G
Tübingen, Deutschland; e-mail: [email protected]
m Maastricht, de Bosquetplein 6-7, NL-6200 AW Maastricht, The
rlands; e-mail: [email protected]
ime, ophiuroids are recorded from the quarry complex at
y Lower Muschelkalk (Anisian, Middle Triassic) limestones, renowned for
abundant vertebrate
in the Muschelkalk (OOSTERINK et al. 2006) yielded dark shales being dated as
to middle Rhaetian on bivalve and palynological evidence (KLOMPMAKER et al.
northern part of the quarry complex (HERNGREEN et al. 2005; KLOMPMAKER et al. in
.). The ophiuroid faunule presented herein was c
24.-26. October 2008 49

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
comparison with other Rhaetian ophiuroids from Europe and especially from the
lithologically comparable Penarth Group in Britain (HESS 1965; SWIFT & MARTILL
, G.F.W., V K - C , J.H.A. & O , H.W., 2005. New
quarry, the eastern Netherlands. Netherlands Journal of Geosciences 84: 409-413.
Mitteilungen aus der Bayerischen Staatssammlung für Paläontologie und historische
1999).
References
HERNGREEN AN ONIJNENBURG VAN ITTERT OSTERINK
geological data (Middle Triassic, Rhaetian-Liassic and Oligocene) of the Winterswijk
HESS, H., 1965. Trias-Ophiuren aus Deutschland, England, Italien und Spanien.
Geologie 5: 151-177.
KLOMPMAKER, A.A., HERNGREEN, G.F.W. & OOSTERINK, H., (in prep.). Uppermost Triassic
(Rhaetic) sediments and subrosion pipe age and stratigraphy at Winterswijk, the
eastern Netherlands.
OOSTERINK, H.W., SIMON, T., HAGDORN, H. & WINKELHORST, H., 2006. A subrosion pipe fill in
the Lower Muschelkalk, Winterswijk Quarry, Eastern Netherlands. Netherlands
Journal of Geosciences 85: 293-297.
SWIFT, A. & MARTILL, D. M., 1999. Fossils of the Rhaetian Penarth Group. Palaeontological
Association, Field Guides to Fossils, 9, 312 pp. London.
50 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Irregular echinoids from the Turonian Seewen Limestone of the
Allgäu area (S. Germany)
(Vortrag)
FRANK WIESE1 & NILS SCHLÜTER2
str. 74-
00, Gebäude Haus D, 12249 Berlin, Deutschland; e-mail: [email protected]
area
dinoc
macro
7 sp
strat
inter Echinoid Level, EL, 1), where Infulaster excentricus FORBES and
(COT hinocorys gravesi (DESOR)
occur. EL 2 consists of two Micraster leseki (DESMOULINS), and M. ex gr.
precursor/normanniae have collected from the top of the exposed rocks (EL 3).
δ13C stratigraphy dates EL 1 as terminal Mid-Turonian to lowermost Upper
Turonian, EL 2 as middle Upper Turonian (level Hitchwood Event = reussianum
Fauna of England & Germany) and EL 3 as Lower Coniacian, also confirmed by
associated inoceramids. Specifically the assemblage of AL 1 consist of taxa which
are characristic of more Boreal areas such as the Anglo Paris Basin, NW Germany
or – in parts – intermediate regions like northern Spain. Although single
elements of the assemblage are known to occur in southern areas (e.g.
Infulaster: Calcareous Alps, Austria: KROH & WAGREICH 2007; Scaglia, Italy; Betic
Cordillera, Spain) the assemblage as a whole spreads a decent flair of Boreal
influence on the distal south Tethyan shelf. Although southward migration of
northern faunas in the course of ocean water cooling is recorded from the Upper
Turonian onwards (WIESE & VOIGT 2002), the Middle Turonian shows still well-
developed biogeographic units among ammonites and echinoids of the Central
European shelf seas and a clear N-S temperature gradient (HART 2007). Thus,
the somehow disjunct position of AL 1 from the areas of its main occurrence may
not be expression of a cooler water zone in the Helvetian shelf seas. Given the
1, 2 Institut für Geologische Wissenschaften, Fachrichtung Paläontologie, FU Berlin, Malteser
1
The pelagic Seewen Limestone of the Burgberg quarry near Sonthofen, Allgäu
(Turonian-?Middle Coniacian, Alpide Helvetic Zone = north Tethyan shelf; c-
yst dominated biosedmentary system) is generally regarded as poorly
fossiliferous. Nevertheless ca. 30 specimens of irregular echinoids
belonging to ecies were collected during a field campaign for isotope
igraphic studies. Most of the taxa come from a stratigraphically limited
val (
Sternotaxis plana (MANTELL) are dominant. In addition, (?)Sternotaxis icaunensis
TEAU), Conulus subrotundus (MANTELL) and Ec
24.-26. October 2008 51

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
variable facies types in which the representatives of AL 2 occur (e.g.Chalk,
Pläner Limestones of NW Germany, marl/limestone alternations, marls), it may
argued that nutrient suppy in context with a distinct diet may have
iggered the development of this assemblage.
mens.
,
YWOOD, A.M., GREGORY, F.J. & SCHMIDT, D.N. (eds), Deep-Time Perspectives on
Climate Change: Marrying the Signal from Computer Models and Biological Proxies,
The Micropalaeontological Society, Special Publications. The Geological Society,
of the Northern Calcareous Alps. - Austrian Journal of Earth Sciences 100: 154-
rather be
tr
AL 2 & 3 cannot be interpreted due to the very low number of speci
References
HART, M. B. 2007. Late Cretaceous climates and foraminiferid distributions. In: WILLIAMS
M., HA
London, 235-250.
KROH, M. & WAGREICH, M. 2007. Infulaster (Echinodermata, Echinoidea) from the Turonian
161.
WIESE, F. & VOIGT, S. 2002. Late Turonian (Cretaceous) climate cooling in Europe: faunal
response and possible causes. – Geobios 35: 65-77.
52 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Antiquity and Longevity of a deep-sea oyster/crinoid association
(Azores Archipelago)
(V ag)
MAX WISSHAK
ortr
Rebikoff-Niggeler Foundation, Rocha Vermelha, 9900-451 Horta, Faial, Azores, Portugal
edrock overhangs in
420-500 m water depth under stable environmental conditions.
Radiocarbon dating of multi-generation oyster stacks (an effective
measure to optimise shell stability with minimal biomineralisation effort) reveals
that individuals reach an impressive lifespan of one to more than four centuries,
placing them among the longest-lived molluscs known to date. This pronounced
longevity in concert with a distinct incremental growth pattern, exhibiting a
proposed reproductive cyclicity of few years (conchiolin-rich growth breaks), and
an annual cyclicity potentially triggered by seasonal varying nutrient flux (distinct
Mg and S fluctuations in the shell calcite), make N. zibrowii a promising
geochemical archive (stable isotopes and trace element signatures) for the upper
bathyal.
Submersible investigations showed that the direct association of C. foresti
and N. zibrowii is not obligatory but that the crinoid is preferentially settling on
oysters, which provide benefits for the crinoid in terms of substrate availability
and/or participation in the oysters’ active feeding current. The upside down
1, CHRISTIAN NEUMANN2, MATTHIAS LÓPEZ CORREA1, SERGE GOFAS3,
CARMEN SALAS3, MARCO TAVIANI4, JOACHIM JAKOBSEN5, ANDRÉ FREIWALD1
1 GeoZentrum Nordbayern, Erlangen University, Loewenichstr. 28, D-91054 Erlangen, Deutschland,
e-mail: [email protected]
2 Museum für Naturkunde, Humboldt University Berlin, Invalidenstrasse 43, D-10115 Berlin,
Deutschland
3 Departamento de Biología Animal, Universidad de Málaga, E-29071 Málaga, Spain
4 CNR-Istituto di Scienze Marine, Via Gobetti 101, I-40129 Bologna, Italy.
5
From the steep submarine slope of the southern Faial Channel (Azores
Archipelago), a conspicuous archibenthic community of two ‘living fossils’,
comprising the up to 30 cm sized deep-sea oyster Neopycnodonte zibrowii and
the sessile cyrtocrinid Cyathidium foresti, is reported. Both taxa thrive in
particularly high densities concealed underneath volcanic b
24.-26. October 2008 53

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
position of both species provides protection from background sedimentation and
possibly shelter from predators.
ely related ancestors of both species are also found as fossils more
an 60 million years old in the Danian (Paleocene) of Faxe in Denmark –
und
Azorean occurrence, on pycnodontine
ryptic habitat preference was already established in the early Palaeogene. This
tion’. Sessile crinoids almost completely
nish
Clos
th
associated with deep-water coral limestone. There Cyathidium holopus is fo
settling in impressive numbers settling on various substrata, such as
scleractinian corals or, analogous to the
oysters. This remarkable resemblance suggests that this association and semi-
c
stands in contrast to the Cretaceous (Cenomanian to Maastrichtian) records of
Cyathidium which were exclusively reported from shallow-water
palaeoenvironments. We thus assume a habitat-shift of these cyrtocrinids toward
deeper waters during the transition of the Mesozoic and Cenozoic erathems. This
reflects the escape from enhanced predation pressure (particularly teleost fishes)
during the ‘Mesozoic Marine Revolu
va ed from shallow waters and were replaced by mobile and often strictly
nocturnal comatulids. The Cyathidium/(Neo)pycnodonte-association did not only
survive the Cretaceous/Palaeogene mass extinction, but the whole Cenozoic and
can today be appreciated as a ‘living-fossil community’.
54 24.-26. October 2008

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
Bromierte Anthrachinon-Pigmente aus der rezenten Seelilie
socrinus ruberrimus
rag)
Proi
(Vort
KLAU 1
1 Insti tät Linz, Altenbergerstraße 69, 4040
Linz, Österreich; e-mail: [email protected]
4 Department of Earth and Planetary Science, University of Tokyo, Hongo, Tokyo 113-0033, Japan
Die rezente Seelilie Proisocrinus ruberrimus ist für ihre intensive orangerote
Färbung bekannt (CLARK 1910). Bis heute gab es über die chemische Natur ihrer
Pigmente jedoch keine Informationen. Die Entdeckung von P. ruberrimus in etwa
1800 m Meerestiefe im Okinawa-Trog (Japan) mit dem Tauchboot „Shinkai 6500“
(OJI & KITAZAWA 2008) ermöglichte nun die chemische Untersuchung eines
Rohextrakts dieser seltenen Seelilie.
Die färbenden Verbindungen eines wässrigen Extrakts von P. ruberrimus
konnten durch Festphasenextraktion aufkonzentriert und gereinigt und die
Hauptverbindungen durch semipräparative Hochleistungsflüssigkeits-
chromatographie (HPLC) isoliert werden. Mittels hochauflösender
Massenspektrometrie konnten die Summenformeln der Pigmente bestimmt
werden (C18H12Br3NaO9S bis C20H15Br4NaO9S). Die charakteristischen
Isotopenmuster der Verbindungen bestätigten dabei das Vorkommen von drei
bzw. vier Bromatomen. Die vollständige Aufklärung der Strukturen durch ein-
und zweidimensionale Kernresonanzspektroskopie ermöglichte schließlich die
Zuordnung der neuen Verbindungen zur Gruppe der Anthrachinon-Pigmente.
Anthrachinone wurden bisher in zahlreichen Comatuliden nachgewiesen, nicht
jedoch bei Seelilien. Soweit bekannt, handelt es sich bei den Pigmenten um die
ersten bromierten Anthrachinone natürlicher Herkunft. Zudem besteht eine
Verwandtschaft der Pigmente aus P. ruberrimus mit den ebenfalls bromierten
Gymnochromen (Phenanthroperylenchinon-Pigmente) aus der Cyrtocrinide
Gymnocrinus richeri (DE RICCARDIS et al. 1991).
Literatur
CLARK, A.H., 1910. Proisocrinus, a new genus of recent crinoids. Proceedings of the
United States National Museum 38: 387-390.
S WOLKENSTEIN , WOLFGANG SCHOEFBERGER1, NORBERT MÜLLER1 & TATSUO OJI2
tut für Analytische Chemie, Johannes Kepler Universi
24.-26. October 2008 55

4. Arbeitstreffen deutschsprachiger Echinodermenforscher 4th Workshop of German & Austrian Echinoderm Research
56 24.-26. October 2008
DE RICCARDIS, F., IORIZZI, M., MINALE, L., RICCIO, R., RICHER DE FORGES, B. & DEBITUS, C.,
1991. The gymnochromes: Novel marine brominated phenanthroperylenequinone
pigments from the stalked crinoid Gymnocrinus richeri. Journal of Organic
Chemistry 56: 6781-6787.
OJI, T. & KITAZAWA, K., 2008. Discovery of two rare species of stalked crinoids from
Okinawa Trough, southwestern Japan, and their systematic and biogeographic
implications. Zoological Science 25: 115-121.





























