Embed Size (px)
Citation preview
brief communications
VASP dynamics during lamellipodia protrusion Klemens Rottner*, Barbara Behrendt†, J.Victor Small*‡ and Jurgen Wehland†
*Institute of Molecular Biology, Austrian Academy of Sciences, Billrothstrasse 11, A-5020 Salzburg, Austria†Gesellschaft für Biotechnologische Forschung, Abteilung Zellbiologie, Mascheroder Weg 1, D-38124 Braunschweig, Germany
‡e-mail: [email protected]
he continuous remodelling of the actin cytoskeleton is a prerequi-site for many cells to move and alter their shape. These activitiesare dependent on the highly regulated and site-specific formation
of protein complexes that act as adaptors to link external signals withactin assembly. The members of the Ena/VASP protein family, VASP(for vasodilator-stimulated phosphoprotein), Mena and Evl, have beenimplicated in the temporal and spatial control of actin-filament dynam-ics. These proteins not only localize to sites of actin assembly, such asfocal-adhesion sites, membrane ruffles and neuronal growth cones1–3,but are also involved in platelet aggregation4, axon guidance5 and theactin-based motility of the intracellular bacterial pathogen Listeriamonocytogenes2,6–8. By generating a stable melanoma cell line expressingVASP fused to green fluorescent protein (GFP), we now show thatVASP not only co-localizes to adhesion sites with the adaptor proteinsvinculin and zyxin (ref. 3 and data not shown), but is also recruited tothe tips of lamellipodia in amounts that are directly proportional to therate of protrusion. These data indicate that VASP may be an adaptormolecule involved in actin-based cell motility. They also raise impor-tant questions about the spatial relationships of the different compo-nents earmarked to have roles in actin-filament dynamics.
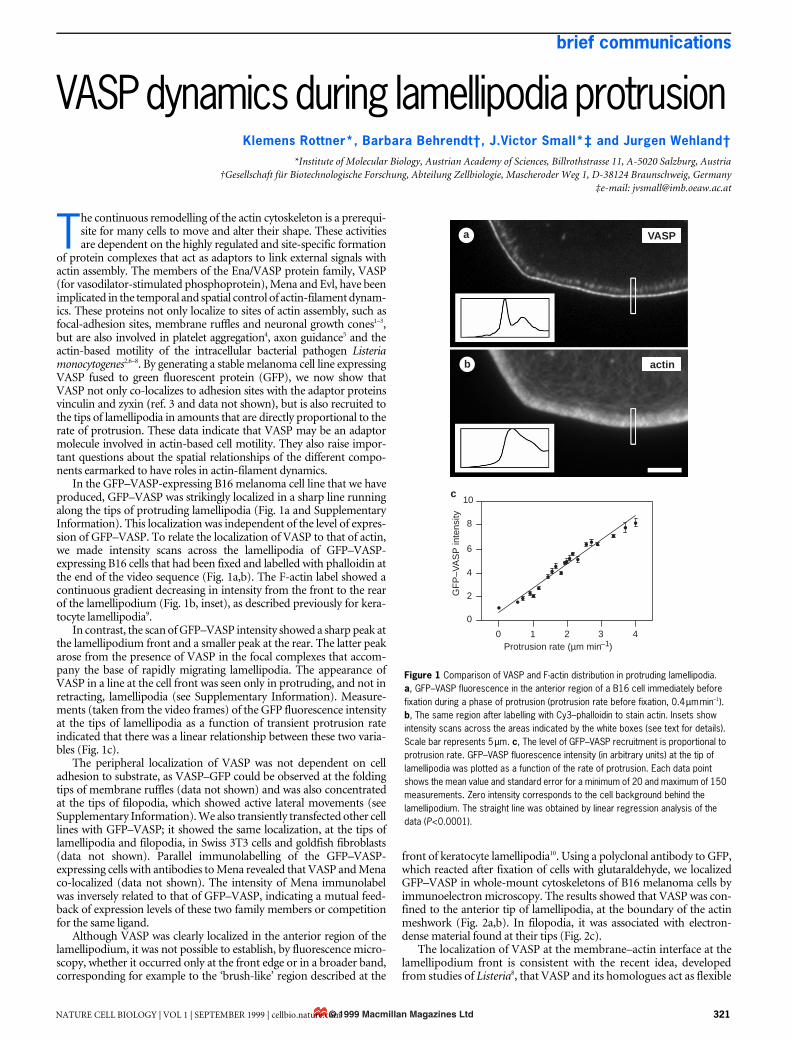
In the GFP–VASP-expressing B16 melanoma cell line that we haveproduced, GFP–VASP was strikingly localized in a sharp line runningalong the tips of protruding lamellipodia (Fig. 1a and SupplementaryInformation). This localization was independent of the level of expres-sion of GFP–VASP. To relate the localization of VASP to that of actin,we made intensity scans across the lamellipodia of GFP–VASP-expressing B16 cells that had been fixed and labelled with phalloidin atthe end of the video sequence (Fig. 1a,b). The F-actin label showed acontinuous gradient decreasing in intensity from the front to the rearof the lamellipodium (Fig. 1b, inset), as described previously for kera-tocyte lamellipodia9.
In contrast, the scan of GFP–VASP intensity showed a sharp peak atthe lamellipodium front and a smaller peak at the rear. The latter peakarose from the presence of VASP in the focal complexes that accom-pany the base of rapidly migrating lamellipodia. The appearance ofVASP in a line at the cell front was seen only in protruding, and not inretracting, lamellipodia (see Supplementary Information). Measure-ments (taken from the video frames) of the GFP fluorescence intensityat the tips of lamellipodia as a function of transient protrusion rateindicated that there was a linear relationship between these two varia-bles (Fig. 1c).
The peripheral localization of VASP was not dependent on celladhesion to substrate, as VASP–GFP could be observed at the foldingtips of membrane ruffles (data not shown) and was also concentratedat the tips of filopodia, which showed active lateral movements (seeSupplementary Information). We also transiently transfected other celllines with GFP–VASP; it showed the same localization, at the tips oflamellipodia and filopodia, in Swiss 3T3 cells and goldfish fibroblasts(data not shown). Parallel immunolabelling of the GFP–VASP-expressing cells with antibodies to Mena revealed that VASP and Menaco-localized (data not shown). The intensity of Mena immunolabelwas inversely related to that of GFP–VASP, indicating a mutual feed-back of expression levels of these two family members or competitionfor the same ligand.
Although VASP was clearly localized in the anterior region of thelamellipodium, it was not possible to establish, by fluorescence micro-scopy, whether it occurred only at the front edge or in a broader band,corresponding for example to the ‘brush-like’ region described at the
front of keratocyte lamellipodia10. Using a polyclonal antibody to GFP,which reacted after fixation of cells with glutaraldehyde, we localizedGFP–VASP in whole-mount cytoskeletons of B16 melanoma cells byimmunoelectron microscopy. The results showed that VASP was con-fined to the anterior tip of lamellipodia, at the boundary of the actinmeshwork (Fig. 2a,b). In filopodia, it was associated with electron-dense material found at their tips (Fig. 2c).
The localization of VASP at the membrane–actin interface at thelamellipodium front is consistent with the recent idea, developedfrom studies of Listeria8, that VASP and its homologues act as flexible
T
Figure 1 Comparison of VASP and F-actin distribution in protruding lamellipodia. a, GFP–VASP fluorescence in the anterior region of a B16 cell immediately before fixation during a phase of protrusion (protrusion rate before fixation, 0.4 µm min–1). b, The same region after labelling with Cy3–phalloidin to stain actin. Insets show intensity scans across the areas indicated by the white boxes (see text for details). Scale bar represents 5 µm. c, The level of GFP–VASP recruitment is proportional to protrusion rate. GFP–VASP fluorescence intensity (in arbitrary units) at the tip of lamellipodia was plotted as a function of the rate of protrusion. Each data point shows the mean value and standard error for a minimum of 20 and maximum of 150 measurements. Zero intensity corresponds to the cell background behind the lamellipodium. The straight line was obtained by linear regression analysis of the data (P<0.0001).
c
a
b
VASP
actin
10
8
6
4
2
0
0 1 2 3 4
GF
P–V
AS
P in
tens
ity
Protrusion rate (µm min )–1
© 1999 Macmillan Magazines LtdNATURE CELL BIOLOGY | VOL 1 | SEPTEMBER 1999 | cellbio.nature.com 321

brief communications
adaptors, linking membrane ligands with polymerizing actin fila-ments. It has also been shown that VASP is an important cofactorrequired for efficient Listeria movement in mixtures of purified pro-teins (T.P. Loisel, R. Boujimaa, D. Pantaloni and M.F. Carlier, per-sonal communication). In this context, VASP-family members maybe essential for docking the quickly growing ends of actin filaments ata defined peripheral location; this docking is necessary for the con-trolled formation of a laminar actin meshwork. Recent studies haverevealed that other components that may be involved in actin-basedmotility, such as the Arp2/3 complex, which promotes filamentextension, are localized to lamellipodia9,10; such studies have lent sup-port to the idea that a significant population of growing filament endsis removed from the membrane interface11. It remains to be seenwhether this model can be reconciled with the spatial localizations ofVASP and other proteins, such as N-WASP, SCAR (WAVE) and pro-filin, that have been implicated in actin-driven motility8,12–15. h
MethodsCells and transfection.Mouse melanoma cells (B16F1) were from American Type Culture Collection and maintained in DMEM
medium (Life Technologies) with 10% FCS (PAA Laboratories, Austria) at 37 °C in the presence of 5% CO2.
The vector for GFP–VASP was provided by U. Carl15 and comprised a full-length construct of human VASP
complementary DNA fused into the Clontech vector EGFP-C2. B16F1 cells were transfected using
lipofectamine (Life Technologies) essentially as described16. Stable cell lines were produced by selection with
1.5 mg ml–1 G418. Single colonies were further expanded in growth media containing 1 mg ml–1 G418. For video
microscopy, B16F1 cells were plated in Ham’s F12 medium containing 10% FCS onto acid-washed glass
coverslips coated first with 100 µg ml–1 polylysine and then with 25 µg ml–1 laminin (Sigma-Aldrich, Austria).
Video microscopy.Cells were observed in an open, heated chamber (Warner Instruments, Reading, UK) at 37 °C on an
inverted microscope (Axiovert 135TV, Zeiss) equipped for epifluorescence and phase-contrast
microscopy, using a ×100/NA 1.4 plan-apochromatic objective with or without 1.6 optovar intermediate
magnification. Tungsten lamps were used for both transmitted and epi-illumination.
Data were acquired using a back-illuminated, cooled charge-coupled-device camera (Princeton
Research Instruments, New Jersey) driven by IPLab software (Scanalytics, Massachusetts) and with
computer-controlled shutters (Optilas GmbH, Puchheim, Germany) in the transmitted and epi-
fluorescence light paths to minimize radiation damage to the cells.
Data were stored as 16-bit digital images and analysed on a Power Macintosh G3 using IPLab, Scion
Image 1.60c (Scion, Maryland) or Adobe Photoshop 5.0 software.
Intensity versus velocity analysis.To correlate lamellipodial protrusion rate with the GFP–VASP signal, we measured the fluorescence
intensity (recorded as counts per pixel) at the tip of the lamellipodium in consecutive frames, 22 s or 30
s apart. In practice, segments of lamellipodia that exhibited uniform rates of protrusion, as judged by the
displacement in the preceding frame, were used for analysis. Segments were typically 2–5 µm wide.
Intensity measurements were recorded for each pixel along the segment and the pixel values for each
protrusion rate were pooled. Measurements were taken from six cells and amounted to between 20 and
150 measurements per data point in Fig. 1c. For each cell, the average intensity in the region behind the
lamellipodium (the lamellum) was taken as background and subtracted from all measurements. To
normalize the measurements of cells with various GFP–VASP expression levels, we set the intensity of
GFP–VASP fluorescence at an intermediate protrusion rate of 2 µm min–1 to the same arbitrary unit for
each cell. Statistical analysis was performed using Microsoft Excel ’97 and Sigma Plot 4.0.
Immunolabelling.For immunoelectron microscopy, B16 cells were plated for 2–24 h onto 15-mm coverslips coated with
formvar films that had been further coated with laminin as described above. Immunolabelling was done
with cells on the filmed coverslips, using a polyclonal anti-GFP antibody from Clontech (1:100 dilution)
in conjunction with a secondary, 10-nm-gold/goat anti-rabbit conjugate from Biocell (Cardiff, UK; 1:30
dilution). In control experiments, we showed, using immunofluorescence microscopy, that the anti-GFP
polyclonal antibody reacted after fixation of cells with glutaraldehyde and revealed the same localization
as for GFP–VASP in living cells (data not shown). The same fixation protocol was used for electron
microscopy, as follows: 1 min fixation in a mixture of 0.5% glutaraldehyde and 0.25% Triton-X100 in
cytoskeleton buffer (150 mM NaCl, 5 mM MgCl2, 5 mM EGTA, 5 mM glucose, 100 µg ml–1 streptomycin,
10 mM MES, pH 6.1) at room temperature, followed by fixation in 1% glutaraldehyde in cytoskeleton
buffer for 15 min. Both fixation mixtures contained phalloidin at 5 µg ml–1. After a brief, 30-s incubation
in 0.5 mg ml–1 sodium borohydride and washing in cytoskeleton buffer, coverslips were incubated for 10
min in 5% horse serum, 1% BSA in PBS containing 5 µg ml–1 phalloidin, and transferred into the same
mixture containing the polyclonal anti-GFP antibody, but lacking horse serum, for 40 min. This was
followed by 2 × 5 min washes in PBS, 1 × 5 min wash in 1% BSA in PBS, and incubation in the second,
gold-conjugated antibody (goat anti-rabbit conjugate) in 1% BSA in PBS containing 5 µg ml–1 phalloidin
for 30 min. After 2 × 5 min washes in 1% BSA in PBS and in PBS alone, the coverslips were transferred to
a Petri dish in PBS for the final step. The films were removed from the coverslips with forceps, inverted
and floated onto the liquid surface as described in ref. 17; grids, made adhesive by coating with
polybutene, were then added to the film.
The filmed grids were then retrieved with parafilm and the cell side immediately rinsed with 100 µg
ml–1 bacitracin followed by 2% sodium silicotungstate; excess stain was removed by blotting with filter
paper and the grids were air-dried. Electron microscopy was performed in a Zeiss 10A electron
microscope operating at 80 kV.
For correlated phalloidin labelling, GFP–VASP-expressing cells were fixed on the microscope stage
with the same mixture as described for electron microscopy, and then incubated with 0.2 µg ml–1 Cy3–
phalloidin (a gift from H. Faulstich) for 60 min at room temperature. Pictures of cells fixed and stained
on the microscope were taken in cytoskeleton buffer containing 100 mM dithioerythritol to avoid
photobleaching.
RECEIVED 21 JUNE 1999; REVISED 19 JULY 1999; ACCEPTED 19 JULY 1999; PUBLISHED 17 AUGUST 1999.
1. Reinhard, M. et al. EMBO J. 11, 2063–2070 (1992).
2. Gertler, F. B., Niebuhr, K., Reinhard, M., Wehland, J. & Soriano, P. Cell 87, 227–239 (1996).
3. Hüttelmaier, S. et al. Curr. Biol. 8, 479–488 (1998).
4. Aszodi, A. et al. EMBO J. 18, 37–48 (1999)
5. Lanier, L. M. et al. Neuron 22, 313–325 (1999).
6. Chakraborty, T. et al. EMBO J. 14, 1314–1321 (1995).
7. Smith, G. A. et al. J. Cell Biol. 144, 1245–1258 (1999).
8. Small, J. V., Herzog, M. & Anderson, K. I. J. Cell Biol. 129, 1275–1286 (1995).
9. Svitkina, T. M. & Borisy, G. G. J. Cell Biol. 145, 1009–1026 (1999).
10. Bailly, M. et al. J. Cell Biol. 145, 331–345 (1999).
11. Mullins, R. D., Heuser, J.A. & Pollard, T. D. Proc. Natl Acad. Sci. USA 95, 6181–6186 (1998).
12. Machesky, L. M. & Insall, R. H. Curr. Biol. 8, 1347–1356 (1998).
13. Miki, H., Suetsugu, S. & Takenawa, T. EMBO J. 17, 6932–6941 (1998).
14. Rohatgi, R. et al. Cell 97, 221–231 (1999).
15. Carl, U. D. et al. Curr. Biol. 9, 715–718 (1999).
16. Ballestrem, C., Wehrle-Haller, B. & Imhof, B. A. J. Cell Sci. 111, 1649–1658 (1998).
17. Buckley, I. K. & Porter, K. R. J. Microsc. 104, 107–120 (1975).
ACKNOWLEDGEMENTS
We thank M. Schmittner for technical assistance and photography, E. Eppacher for manuscript
processing and T. Stradal for help with statistics. This work was funded in part by the Austrian Science
Research Foundation, the Austrian National Bank and the Seegen Stiftung of the Austrian Academy of
Sciences (to J.V.S) and in part by the Deutsche Forschungsgemeinschaft and the Fonds der Chemische
Industrie (to J.W.).
Correspondence and requests for materials should be addressed to J.V.S.
Supplementary Information is available on Nature Cell Biology’s World-Wide Web site (http://
cellbio.nature.com).
Figure 2 GFP–VASP is localized at the tips of lamellipodia and microspikes/filopodia. The figure shows electron micrographs of a whole-mounted cytoskeleton from a GFP–VASP-expressing cell that was labelled with an anti-GFP antibody followed by a secondary antibody conjugated to 10-nm gold. The boxed area in b is shown at higher magnification in a. c, Filopodium, showing label at the tip. ms, microspike; lam, lamellipodium. Scale bars in a, c represent 0.1 µm and in b, 0.5 µm.
a
b
c
ms
lam
© 1999 Macmillan Magazines Ltd322 NATURE CELL BIOLOGY | VOL 1 | SEPTEMBER 1999 | cellbio.nature.com


![Technical document HB510 [DE] - itoh-denki.comitoh-denki.com/images/TechnicalDocument/HB510/Technical document HB510... · Die Steuerplatine ist mit anderen Platinen des Typs HBxxx](https://img.pdfslide.org/doc/110x75/5e1def632e129127850187ea/technical-document-hb510-de-itoh-denkicomitoh-denkicomimagestechnicaldocumenthb510technical.jpg)












![Untitled Document [] · Title: Untitled Document Created Date: 1/23/2001 9:43:51 AM](https://img.pdfslide.org/doc/110x75/601a87c13516f3400634077d/untitled-document-title-untitled-document-created-date-1232001-94351-am.jpg)












