Embed Size (px)
Citation preview

7 Das Nervensystem
Literatur zu diesem Kapitel:
• Der Körper des Menschen, A. Faller, M. Schün-ke; Thieme
• Biophysik, Hoppe, Lohmann, Markl, Ziegler,Kap. 15 Neurobiophysik, Kap. 16 Kybernetik
• Lehrbuch der Biophysik, E. Sackmann und R.Merkel, Kap. 14-17: Biophysik der Nervenlei-tung
• Physics in Biology and Medicine, P. Davido-vits, Kap. 13.1 The Nervous System
• Physics of the Body, J.R. Cameron, J.G. Sko-fronick, R.M. Grant, Kap. 9: Electrical Signalsfrom the Body
7.1 Information undKommunikation
7.1.1 Das menschliche Nervensystem
Es gibt im Körper eines Menschen im Wesentlichen2 Kommunikationssysteme: ein chemisches System,wo der Informationsfluss ausschließlich durch Bo-tenstoffe (Hormone) vermittelt wird, und ein Sy-stem mit festen Bahnen, das Nervensystem, das indiesem Kapitel behandelt wird.
Das Nervensystem hat zwei unterschiedliche Funk-tionsweisen: das animale (anima = Seele, Geist), so-matische oder willkürliche Nervensystem nimmtdie Umweltreize auf, verarbeitet sie, steuert bewus-ste Handlungen und erzeugt Vorstellungen. Das au-tonome oder vegetative Nervensystem ist für selb-ständige Körperfunktionen zuständig und kann nichtwillentlich beeinflusst werden. Die Zentren beiderSysteme befinden sich im Gehirn und Rückenmark.Im 19. Jahrhundert dachte man, die beiden Systemewären räumlich und funktionell voneinander weitge-hend unabhängig; aber inzwischen weiß man, dass
zwischen beiden zahlreiche anatomische und funk-tionelle Verbindungen bestehen und die Unterschei-dung nicht absolut ist.
Strukturell wird das Nervensystem in zwei Teile un-terteilt. Der erste Teil umfasst das Rückenmark unddas Gehirn und wird als zentrales oder cerebrospi-nales Nervensystem (ZNS) bezeichnet. Der zwei-te Teil des Nervensystems, das periphere Nervensy-stem (PNS) erstreckt sich in alle Bereiche des Kör-pers. Es ist weit verzweigt und besteht aus Ner-ven, Nervensträngen und kleineren Nervenansamm-lungen (Ganglien).
Abbildung 7.1: Prinzipieller Aufbau des sensori-schen und motorischen Systems.
Der Organismus tritt über das Nervensystem mit derUmwelt in Wechselwirkung. Die Umwelt wirkt aufden Organismus, und dies wird mit Hilfe des sen-sorischen Systems abgebildet. Dies beginnt an spe-ziell ausgebildeten Strukturen einer peripheren Ner-venzelle. Diese Umweltsensoren werden als Rezep-toren bezeichnet. Die Reaktionen der Rezeptoren
152

7 Das Nervensystem
werden als elektrische Signale über die angeschlos-senen Nervenfasern in das zentrale Nervensystemübertragen. Dort erfolgt die bewusste Empfindungund Wahrnehmung der Umwelt.
Das Nervensystem steuert auch den Informations-fluss in umgekehrter Richtung: vom motorischen Sy-stem werden über Nervenimpulse Signale an Mus-keln übertragen. Auf diese Weise erfolgt die Rück-wirkung des Organismus auf die Umwelt.
7.1.2 Informationsgehalt
Der Mensch nimmt Daten der Umgebung mit Hil-fe der Sinnesorgane auf und verarbeitet diese Infor-mation weiter. Um die Kapazität der Informations-aufnahme des Menschen zu quantifizieren, muss zu-nächst der Begriff der Information präzisiert werden.
Der Informationsgehalt einer Nachricht hängt vomWissensstand des Empfängers ab. Wenn man liest,die Sonne würde morgen aufgehen, dann lernt manvon dieser Aussage wesentlich weniger als wenn ge-sagt wird, dass sie nicht aufgehen wird. In der Infor-mationstheorie wird dieser “Überraschungsgehalt”quantifiziert. Wenn p(xi) die Wahrscheinlichkeit ei-ner Nachricht ist (zum Beispiel das Auftreten einesSymbols xi der Nachrichtenquelle), dann ist der zu-gehörige Informationsgehalt Ii gegeben durch
Ii := � log2 (p(xi)) .
Der Maßstab ist so festgelegt, dass eine Nachrichtmit p = 1/2 den Informationsgehalt 1 bit ergibt.1Gleich wahrscheinliche binäre Zeichen besitzenden Informationsgehalt von 1 bit pro Zeichen.
Dadurch ist es möglich, zum Beispiel den Infor-mationsgehalt einer Nachricht anzugeben oder auchdie maximale Nachrichtenmenge pro Zeiteinheit,die über einen Übertragungskanal gesendet werdenkann, wenn auch Störungen mit berücksichtigt wer-den. Der mittlere Informationsgehalt einer Nach-richtenquelle mit einem Vorrat aus N Symbolen er-gibt sich durch die Bildung des Erwartungswer-
1Wenn der natürliche Logarithmus zur Definition benutzt wird,dann spricht man nicht von Bits, sondern von Nats.
0 0.5 10
0.5
Abbildung 7.2: Entropie als Funktion der Wahr-scheinlichkeit p(x).
tes über die individuellen Informationsgehalte allerSymbole
H(x) := E{Ii} = �N
Âi=1
p(xi) · log2 (p(xi)) .
Diese Größe nennt man wegen ihrer formalen Ent-sprechung Entropie. Die Entropie ist bei gleichwahrscheinlichen Zeichen am größten: Hmax(x) =log2(N). Da dies auch die Anzahl der binären Ent-scheidungen ist, die notwendig sind, um eines der NZeichen auszuwählen, wird diese Größe auch Ent-scheidungsgehalt genannt.
Bei einem sprachlichen Text z.B. sind die einzelnenSymbole nicht unabhängig voneinander. Daher müs-sen lange Symbolfolgen betrachtet werden um dieEntropie zu berechnen,
H(x) = limn!•
1n
E{In}.
Für einen deutschen Text erhält man eine Entropievon 1.3 bit pro Buchstabe, bei einem englischen Text1.0 bit pro Buchstabe. Der Maximalwert für 30 Zei-chen wäre durch den Entscheidungsgehalt
log2(30) ⇡ 4.9
gegeben und ist 4.9 bit pro Buchstabe. Die Differenzzwischen Entscheidungsgehalt und Entropie nenntman Redundanz. Sie ist bei englischen Texten hö-her als bei deutschen (3.9 statt 3.6 bit pro Buchsta-be). Bei Kompressionsverfahren wird diese Redun-danz eliminiert und dadurch der Speicherbedarf re-duziert.
153

7 Das Nervensystem
7.1.3 Kapazität eines Übertragungskanals
Information wird zwischen unterschiedlichen Ortenübertragen, z.B. zwischen einer Sinneszelle und demGehirn. Man bezeichnet den Pfad zwischen Senderund Empfänger als Kommunikationskanal.
Sender EmpfängerH(x)
H(x|y) H(y|x)
H(y)T
Abbildung 7.3: Einseitig gerichteter Informations-oder Nachrichtenkanal.
Der sogenannte Shannon’sche Nachrichtenkanal isteinseitig gerichtet, das bedeutet die Nachrichten-quelle (Sender) ist unabhängig vom Empfänger. (DieKommunikation zwischen Lebewesen ist dagegennicht unabhängig, da die erzeugten Symbole von denempfangenen Symbolen statistisch abhängig sind.Dieser allgemeinere Fall, die bidirektionale Kom-munikation nach Marko ist für das Folgende abernicht weiter wichtig.) Wir bezeichnen die Informa-tionseinheiten an der Quelle mit xi, diejenigen, diebeim Empfänger eintreffen mit yi. Im Idealfall giltxi = yi 8i.
Informationsübertragung erfolgt in der Praxis nieperfekt, sondern unterliegt Störungen. Informations-technisch bedeutet eine Störung, dass ein Teil derempfangenen Zeichen y j sich von den gesendetenunterscheidet. Verschiedene xi können das gleicheZeichen y j ergeben (Rückschlussunsicherheit), diezugehörige Rückschlussentropie H(x|y) wird auchals Äquivokation bezeichnet.
Unter Irrelevanz oder Streuentropie H(y|x) verstehtman, dass aus einem Zeichen xi verschiedene y j wer-den. Sie ist gegeben als
H(a|b) := limn!•
1n
·E{p(a|b)} ,
d.h. durch die bedingte Wahrscheinlichkeit für dasAuftreten von a bei der Kenntnis von b. Von der
Entropie H(x) gelangt nur die Differenz T = H(x)�H(x|y) an das Kanalende. T ist der mittlere Trans-informationsgehalt, oder die Synentropie. Für denEmpfänger wirkt das Kanalende als Informations-quelle der Entropie H(y) = T +H(y|x). Für den un-gestörten Fall ist H(y) = T = H(x). Bei einem sehrstark gestörten Kanal werden die Zufallsvariablen xund y statistisch unabhängig, so dass mit H(x|y) =H(x) die Transinformation verschwindet. H(y) be-steht dann nur noch aus dem irrelevanten Anteil.
Der maximale Wert der Transinformation bestimmtdie Kanalkapazität C, also den Maximalwert desmöglichen Nachrichtenflusses (bei gegebenem Ka-nal). Wenn t die mittlere Übertragungszeit je Nach-richt (Symbol) ist, dann gilt
C =
✓Tt
◆
max.
Bei einem symmetrischen Binärkanal mit einer Feh-lerwahrscheinlichkeit q und einer Schrittdauer t ist
C =1t
[1+q log2 q+(1�q) log2 (1�q)] .
Für einen analogen Kanal mit der Bandbreite D f , derSendeleistung Ps und der Störleistung Pn ergibt sichfür Gauß’sche Verteilungen von Sende- und Störsi-gnal die Kanalkapazität
C = D f · log2
✓1+
Ps
Pn
◆.
7.1.4 Beispiele
Typische Werte für die Kanalkapazität technischerNachrichtensysteme sind:
• Telefonkanal: C = 5 ·104 bit/s
• Fernsehkanal: C = 7 ·107 bit/s
Diese Kanäle sind angepasst an die Informationska-pazität menschlicher Sinnesorgane, z.B.:
• Ohr: C = 4 ·104 bit/s
• Auge: C = 3 ·106 bit/s
154
7 Das Nervensystem
Die von Menschen bewusst verarbeiteten Kapazi-tätswerte sind erheblich kleiner. Beispiele:
• leise / laut lesen: C = 45 bit/s / C = 30 bit/s
• Schreibmaschine schreiben: C = 16 bit/s
• Klavierspielen: C = 23 bit/s
• Addieren / Multiplizieren: C = 12 bit/s
• Abzählen: C = 3 bit/s
Ein Mensch kann nur Informationsflüsse mit < 50bit/s bewusst aufnehmen und verarbeiten. Die not-wendige Informationsreduktion findet in der Deko-dierung der Sinneseindrücke durch das zentrale Ner-vensystem statt. Die größte Kompression findet beimAuge statt, wo ein Teil der Verarbeitung bereits inder Netzhaut geschieht (siehe 9.4).
7.2 Das Nervensystem
7.2.1 Übersicht
A!erenzen E!erenzen
Abbildung 7.4: Organisation des Nervensystems.
Das Nervensystem überträgt Signale von Sensoren(Sinneszellen) ans Zentralnervensystem und ‘Anord-nungen’ an Muskeln und andere Organe. Die Signal-wege zum Gehirn werden als “afferent” (=hinzufüh-rend) bezeichnet, die umgekehrte Richtung als “ef-ferent” (=wegführend).
Die somatischen Afferenzen umfassen den desGesichts-, Gehör- und Gleichgewichtsinns, dieBerührungsempfindung, Temperaturwahrnehmung,Schmerzwahrnehmung. Zu den viszeralen Afferen-zen gehören Signale von den inneren Organen, sowieGeruchs- und Geschmackswahrnehmungen.
Rezeptoren reagieren (nur) auf adäquate Reize undübersetzen diese in eine elektrische Erregung. Esmuss außerdem eine Reizschwelle überschrittenwerden, um die Reizempfindung auszulösen. Häu-fig reicht die Sensibilität von Rezeptoren bis an diephysikalischen Grenzen heran (siehe 8.3 und 9.3).So können bereits 5 Photonen als Lichtblitz wahrge-nommen werden.
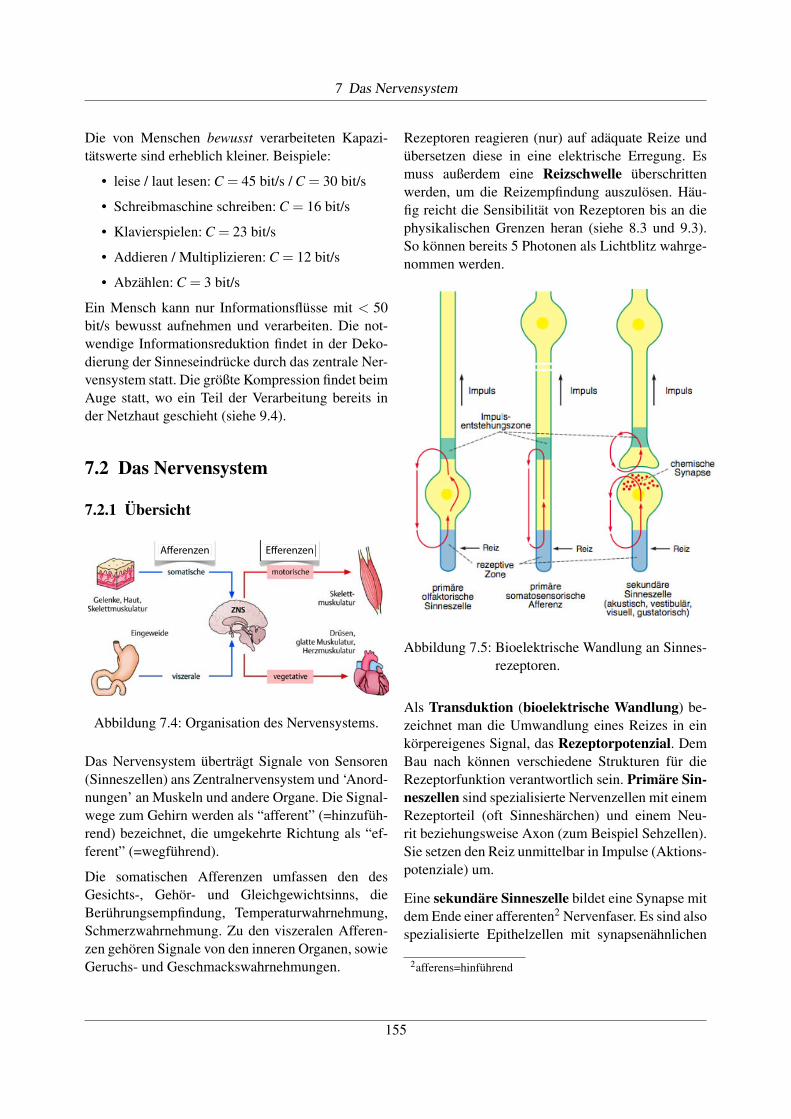
Abbildung 7.5: Bioelektrische Wandlung an Sinnes-rezeptoren.
Als Transduktion (bioelektrische Wandlung) be-zeichnet man die Umwandlung eines Reizes in einkörpereigenes Signal, das Rezeptorpotenzial. DemBau nach können verschiedene Strukturen für dieRezeptorfunktion verantwortlich sein. Primäre Sin-neszellen sind spezialisierte Nervenzellen mit einemRezeptorteil (oft Sinneshärchen) und einem Neu-rit beziehungsweise Axon (zum Beispiel Sehzellen).Sie setzen den Reiz unmittelbar in Impulse (Aktions-potenziale) um.
Eine sekundäre Sinneszelle bildet eine Synapse mitdem Ende einer afferenten2 Nervenfaser. Es sind alsospezialisierte Epithelzellen mit synapsenähnlichen
2afferens=hinführend
155

7 Das Nervensystem
Gebilden zur Reizweiterleitung an Nervenfaseren-den. Das Rezeptorpotenzial moduliert die Freiset-zung einer Transmittersubstanz, so dass der Impulsim nachgeschalteten afferenten Neuron entsteht.
7.2.2 Rezeptorpotenzial
Signale der Umwelt werden durch Rezeptoren auf-genommen und von Nerven weitergeleitet. Man un-terscheidet je nach Reiz zwischen:
• Mechano- (Haut, Ohr)
• Thermo- (Haut)
• Chemo- (Zunge, Nase)
• Photo- (Auge)
RezeptormembranMyelin
Entstehung von Aktionspotenzialen
Abbildung 7.6: Rezeptorpotenzial und Signalweiter-leitung.
In Rezeptoren wird durch den Reiz das Membranpo-tenzial geändert. Es entsteht ein Rezeptorpotenzial,dessen Amplitude die Stärke des Reizes widerspie-gelt.
Rezeptorpotenziale sind lokal, ihre Reichweite istnur wenige mm. Falls der Reiz eine bestimmteSchwelle überschreitet, löst er jedoch eine Abfolgevon Aktionspotenzialen aus, welche dem Nerv ent-lang wandern und so die Information überträgt.
Die Amplitude des Rezeptorpotenzials ist propor-tional zur Amplitude und zeitlichen Änderung desReizsignals. Nur wenn das Reizpotenzial einen Mi-nimalwert überschreitet, wird die Information weitergeleitet.
Die Frequenz des Aktionspotenzials hängt von derStärke des Rezeptorpotenzials ab. Dadurch wird ein
Haut Reiz
Stärke
Rezeptorpotenzial
Zeit
Aktions-potenzial
Abbildung 7.7: Reiz, Rezeptorpotenzial und Akti-onspotenzial.
analoges Signal (Reiz) in ein digitales Signal desAktionspotenzials umgesetzt.
7.2.3 Sinnesphysiologie
Die Sinnesorgane nehmen Reize auf und verarbeitendiese weiter zu (Nerven-)Impulsen, welche an daszentrale Nervensystem weitergeleitet werden. Daszentrale Nervensystem bekommt die Gesamtheit al-ler Impulse von verschiedenen Rezeptoren und aufdiese Weise kann ein Abbild der Umwelt entstehen.Die subjektive Verarbeitung eines einzelnen Sinnes-reizes führt zu einem Sinneseindruck. In einer Sin-nesempfindung sind mehrere Sinneseindrücke zueinem Gesamtbild zusammengefasst. Durch die Be-wertung dieser Empfindung und die Deutung mit derErfahrung gelangen wir zu der Wahrnehmung.
Das Gesamtbild einer Empfindung ist durch 4 Para-meter, die sogenannten ”Dimensionen”, beschreib-bar. Diese sind:
• Modalität. Zum Beispiel die klassischen 5 Sin-ne: Gesichts-, Gehör-, Tast-, Geschmacks- undGeruchssinn. Innerhalb einer Modalität kannman verschiedene Arten von Sinneseindrückenunterscheiden, die Submodalitäten.3
• Intensität. Eine Empfindung kann stark oderschwach sein.
3In der klassischen subjektiven Sinnesphysiologie auch als”Qualität” bezeichnet.
156

7 Das Nervensystem
Tabelle 7.1: Beispiele:Rezeptortyp Empfindungsmodalität Submodalität (Qualität)
Photorezeptoren Gesichtssinn Helligkeit, Dunkelheit, Farben, Form, BewegungThermorezeptoren Temperatursinn Kälte, WärmeChemorezeptoren Geruchssinn verschiedene Gerüche
Geschmackssinn Säure, Salze, Süße, BitterkeitMechanorezeptoren u.a.: Gehörsinn Tonhöhen
• Räumlichkeit. Eine Empfindung kann lokali-siert werden (z.B. Berührung der Haut).
• Zeitlichkeit. Eine Empfindung hat einen zeitli-chen Beginn und eine Abklingzeit.
7.3 Signale in Nerven
Unsere Vorstellung von der Signalübertragung inNerven wird zu einem wesentlichen Teil durch dieArbeiten von Hodking und Huxley [13, 11, 9, 12, 10]geprägt, welche als Hodgkin-Huxley Modell be-kannt sind.
7.3.1 Nervenzellen
Zellkörper
Nervenfaser
Schwann’sche Zelle
Dendriten
Zellkern
Axon
zum nächsten Neuron
zum nächsten NeuronSignalausbreitung
Abbildung 7.8: Nervenzelle.
Eine Nervenzelle (Neuron) ist eine auf Erregungs-weiterleitung spezialisierte Zelle. Sie gliedert sich in3 Abschnitte (entsprechend der Richtung des Erre-gungsablaufs):
• Dendriten: Rezeptive Strukturen,
• Neurit (Axon, Nervenfaser): Effektorische(nach außen leitende) Strukturen,
• Perikaryon (Soma): Zellleib; Stoffwechsel-zentrum.
Das Axon ist im periferen Nervensystem von Mye-lin (Markscheiden, Schwann’sche Zellen) umgeben.Dieses stellt eine isolierende Schicht dar (siehe7.4.4) und wird durch die Ranvier’schen Schnürringeunterbrochen, wo das Axon in direkter Verbindungmit dem extrazellulären Raum steht. Im zentralenNervensystem sind die Axone in der Regel zu Faser-bündeln zusammengefasst, im Bereich des periphe-ren Nervensystems zu Nervensträngen und Nerven.
7.3.2 Synapsen
Im Nervensystem werden Informationen in Sequen-zen von Aktionspotenzialen verschlüsselt und überdie Nervenfasern weitergeleitet. Zur weiteren Verar-beitung müssen diese Informationen auf andere Neu-ronen übertragen werden. Die Weitergabe der Infor-mation erfolgt an Strukturen, die unter dem Oberbe-griff Synapsen zusammengefasst sind.
In der Mitte der Abbildung 7.9 sind Bestandteile ei-ner synaptischen Verbindung und typische Lokalisa-tionen von Synapsen an einem Neuron gezeigt.
Es gibt zwei Typen von Synapsen (siehe Abbildung7.10): beim ersten Typ sind prä- und postsynaptischeAnteile über Proteine (Konnexone), die tunnelarti-ge Verbindungen (gap junctions) bilden, direkt mit-einander in Kontakt. Man spricht von elektrischenSynapsen, da die Erregungsübertragung durch elek-trotonische Ströme durch diese Tunnel erfolgt.
157
7 Das Nervensystem
axo-dendritische S.
axo-somatische S.
axo-axonale S.
axo-somatische S.
synaptischer Endkopf
postsynap-tische
Membran
synaptischer Spalt
Transmitter-bläschen
präsynaptische Membran
Abbildung 7.9: Überblick über verschiedene Typenvon Synapsen.
elektrische Synapse chemische Synapse
präsynaptische Faser
postsynaptische Membran
Konnexane
präsynaptische Faser
postsynaptische Membran
Membran-kanal
Rezeptor
Trans-mitter
Abbildung 7.10: Signalübertragung durch elektri-sche und chemische Synapse.
Beim zweiten Typ, der chemischen Synapse, sinddie Membranen der prä- und postsynaptischenStrukturen durch einen Spalt voneinander getrennt.Die Erregungsweiterleitung erfolgt über chemischeSubstanzen, die sogenannten Transmitter.
Es gibt exzitatorische (anregende) und inhibitorische(unterdrückende) Synapsen. Eine Synapse wird alsexzitatorisch bezeichnet, wenn ihre Aktivierung dieWahrscheinlichkeit erhöht, dass die postsynaptischeNervenzelle erregt wird, eine inhibitorische Synapsereduziert die Wahrscheinlichkeit.
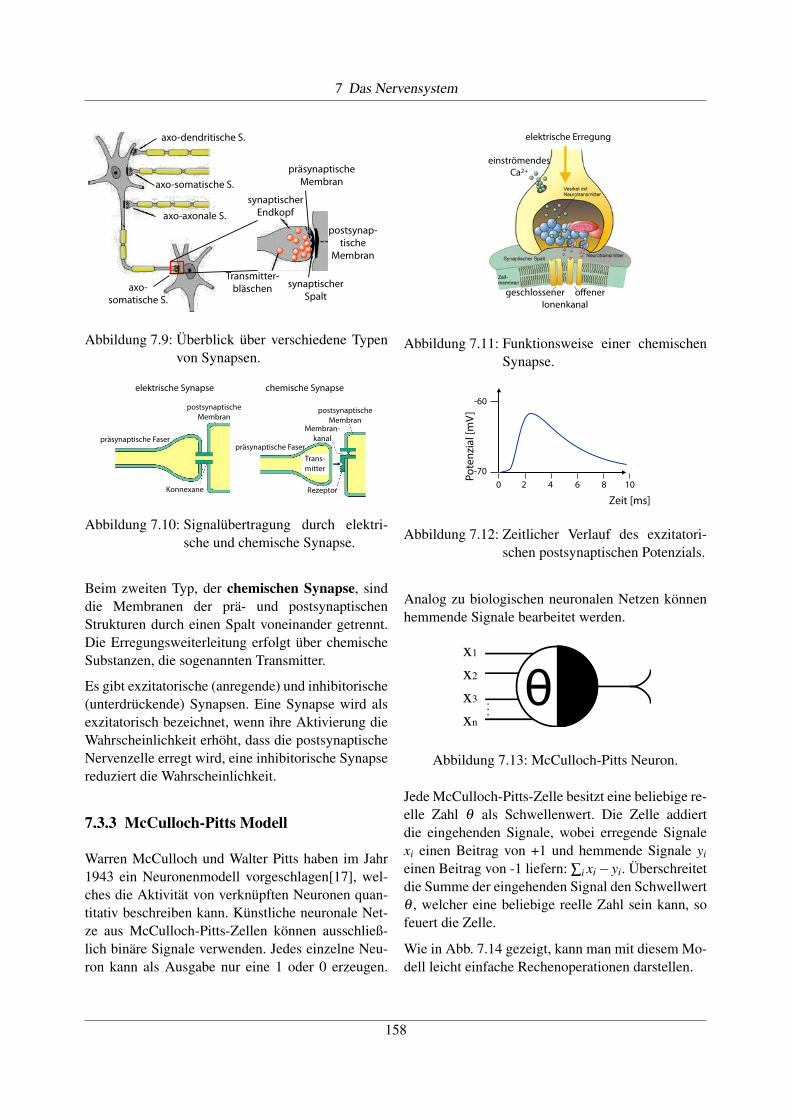
7.3.3 McCulloch-Pitts Modell
Warren McCulloch und Walter Pitts haben im Jahr1943 ein Neuronenmodell vorgeschlagen[17], wel-ches die Aktivität von verknüpften Neuronen quan-titativ beschreiben kann. Künstliche neuronale Net-ze aus McCulloch-Pitts-Zellen können ausschließ-lich binäre Signale verwenden. Jedes einzelne Neu-ron kann als Ausgabe nur eine 1 oder 0 erzeugen.
elektrische Erregung
einströmendes Ca2+
geschlossener o!enerIonenkanal
Abbildung 7.11: Funktionsweise einer chemischenSynapse.
Pote
nzia
l [m
V]
Zeit [ms]
-60
-700 2 4 6 8 10
Abbildung 7.12: Zeitlicher Verlauf des exzitatori-schen postsynaptischen Potenzials.
Analog zu biologischen neuronalen Netzen könnenhemmende Signale bearbeitet werden.
x1x2x3xn
Abbildung 7.13: McCulloch-Pitts Neuron.
Jede McCulloch-Pitts-Zelle besitzt eine beliebige re-elle Zahl q als Schwellenwert. Die Zelle addiertdie eingehenden Signale, wobei erregende Signalexi einen Beitrag von +1 und hemmende Signale yieinen Beitrag von -1 liefern: Âi xi �yi. Überschreitetdie Summe der eingehenden Signal den Schwellwertq , welcher eine beliebige reelle Zahl sein kann, sofeuert die Zelle.
Wie in Abb. 7.14 gezeigt, kann man mit diesem Mo-dell leicht einfache Rechenoperationen darstellen.
158

7 Das Nervensystem
AND OR NOT
Abbildung 7.14: Logische Gatter im McCulloch-Pitts Modell.
In einem erweiterten Modell kann jeder Synapse einGewicht wi zugeordnet werden. Das postsynaptischeNeuron feuert, wenn die gewichtete Summe allereingehenden Signale größer ist als ein Schwellen-wert Âi xiwi � q .
7.3.4 Ruhemembranpotenzial
Der Ursprung bioelektrischer Signale sind Potenzi-aldifferenzen zwischen dem Inneren und Äußerenbiologischer Zellen. Diese kommen durch Unter-schiede in der Konzentration der Ionen zustande undkönnen z.B. mit Hilfe von Mikroelektroden gemes-sen werden.
Zellen regulieren sehr sorgfältig ihre intrazellulä-re Ionen-Konzentration, um das Auftreten eines os-motischen Druckes zu vermeiden (Ausgleich vonKonzentrationsunterschieden). Als Konsequenz ha-ben die wichtigsten Ionen (Na+, K+, Cl� and Ca2+
verschiedene Konzentrationen im Extra- und Intra-zellularraum. Daraus ergibt sich eine Spannungsdif-ferenz über die Zellmembran.
Tintenfisch Riesen-Axon
Frosch Sartorius Muskel
Menschliche Rote Blutzellen
IntrazellulärIntrazellulärIntrazellulärIntrazellulär
Na+ 50 13 19
K+ 397 138 136
Cl- 40 3 78
ExtrazellulärExtrazellulärExtrazellulärExtrazellulär
Na+ 437 110 155
K+ 20 2.5 5
Cl- 556 90 112
Abbildung 7.15: Typische Ionenkonzentrationen inunterschiedlichen Zellen.
Im Wesentlichen gibt es zwei Arten von Zellen: er-regbare und nicht-erregbare. Alle Zellen haben eineMembran-Ruhe-Potenzial, aber nur erregbare Zellenkönnen es aktiv modulieren.
Zelle139 mMol Membran 1 µF/cm2
4 mMol
145 mMol
12 mMol
MP = -70 mV
Abbildung 7.16: Entstehung des Ruhemembranpo-tenzials. Eingezeichnet sind diewesentlichen Prozesse für denTransport von K+ und Na+ durchdie Zellmembran.
Die Doppellipidschicht einer Zelle ist elektrisch iso-lierend und besitzt eine hohe Kapazität von ungefähr1 µF/cm2 (Dicke der Zellmembran: ⇡ 5 nm). Diewichtigsten Ionenpumpen sind die Na+/K+ Pumpen(siehe 3.3.4). Andere Kanäle kontrollieren den passi-ven Transport für bestimmte Ionen. Außerdem ist dieZellwand für K+ Ionen teilweise durchlässig. Die-se erreichen deshalb eine Gleichgewichtsverteilungzwischen Innen und Außen.
Durch die Summe dieser Teilprozesse entsteht eineungleiche Ionenkonzentration zwischen dem Zellin-neren und Zelläußeren, und daher ein Potenzial, dasden Ruhezustand der Zelle charakterisiert. Dieseswird als Ruhemembranpotenzial bezeichnet.
Die Zelle insgesamt wie auch das Zelläußere sindelektrisch neutral, aber auf der Membran ergibt sichnetto ein negativer Ladungsüberschuss im Innern derZelle und ein positiver Überschuss außerhalb der
159
7 Das Nervensystem
Ladungsdichte
Ladungsdichte
Zelle
Extrazel-lulärraum
MP
= -7
0 m
V
Abbildung 7.17: Verlauf des elektrischen Feldes unddes elektrischen Potenzials über dieZellmembran.
Zelle. Abb. 7.17 zeigt, wie diese Ladungsdichtever-teilung ein elektrisches Feld über der Membran undeinen Gradienten des Potenzials erzeugt.
Das Ungleichgewicht der Ionenkonzentrationen imRuhezustand wird als Polarisation der Zellen be-zeichnet. Für eine Ionenart wird die daraus resultie-rende Potenzialdifferenz durch die Nernst’sche Glei-chung gegeben (siehe Kapitel 3.3.3). Der Konzentra-tionsgradient führt zu einem Strom von Na-Ionen insInnere der Zelle, für die K-Ionen ist der Konzentrati-onsgradient umgekehrt. Das elektrische Feld erzeugtfür beide positiven Ionen eine Kraft, welche ins In-nere der Zelle zeigt. Für Na addieren sich die beidenKräfte, beim K wirken sie entgegengesetzt und he-ben sich deshalb teilweise auf.
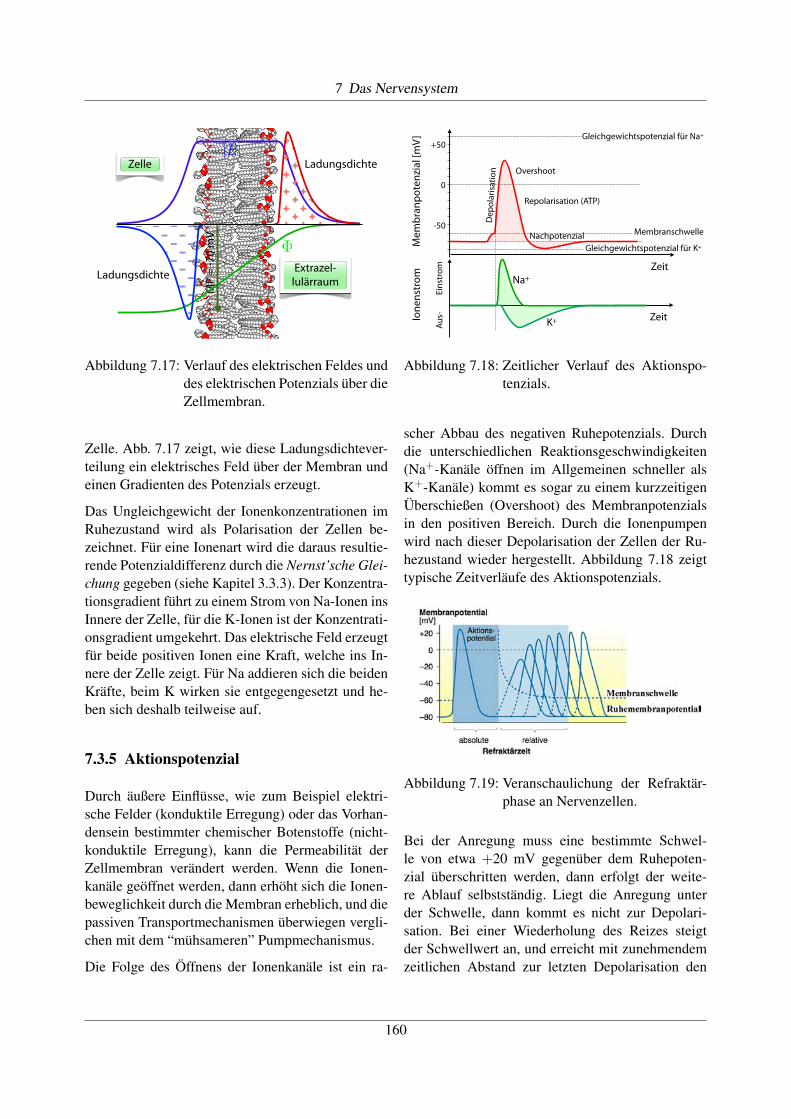
7.3.5 Aktionspotenzial
Durch äußere Einflüsse, wie zum Beispiel elektri-sche Felder (konduktile Erregung) oder das Vorhan-densein bestimmter chemischer Botenstoffe (nicht-konduktile Erregung), kann die Permeabilität derZellmembran verändert werden. Wenn die Ionen-kanäle geöffnet werden, dann erhöht sich die Ionen-beweglichkeit durch die Membran erheblich, und diepassiven Transportmechanismen überwiegen vergli-chen mit dem “mühsameren” Pumpmechanismus.
Die Folge des Öffnens der Ionenkanäle ist ein ra-
Gleichgewichtspotenzial für Na+
Membranschwelle
Gleichgewichtspotenzial für K+
Zeit
Mem
bran
pote
nzia
l [m
V] +50
0
-50 Dep
olar
isatio
n Overshoot
Nachpotenzial
Repolarisation (ATP)
Ione
nstr
om
Eins
trom
Aus- Zeit
Na+
K+
Abbildung 7.18: Zeitlicher Verlauf des Aktionspo-tenzials.
scher Abbau des negativen Ruhepotenzials. Durchdie unterschiedlichen Reaktionsgeschwindigkeiten(Na+-Kanäle öffnen im Allgemeinen schneller alsK+-Kanäle) kommt es sogar zu einem kurzzeitigenÜberschießen (Overshoot) des Membranpotenzialsin den positiven Bereich. Durch die Ionenpumpenwird nach dieser Depolarisation der Zellen der Ru-hezustand wieder hergestellt. Abbildung 7.18 zeigttypische Zeitverläufe des Aktionspotenzials.
Abbildung 7.19: Veranschaulichung der Refraktär-phase an Nervenzellen.
Bei der Anregung muss eine bestimmte Schwel-le von etwa +20 mV gegenüber dem Ruhepoten-zial überschritten werden, dann erfolgt der weite-re Ablauf selbstständig. Liegt die Anregung unterder Schwelle, dann kommt es nicht zur Depolari-sation. Bei einer Wiederholung des Reizes steigtder Schwellwert an, und erreicht mit zunehmendemzeitlichen Abstand zur letzten Depolarisation den
160

7 Das Nervensystem
Gleichgewichtswert. Es gibt eine Phase nach einemReiz, in dem keine neue Erregung möglich ist. Diesist die sogenannte absolute Refraktärphase. In dersich anschließenden relativen Refraktärphase isteine Erregung nur mit reduzierter Amplitude mög-lich und es gilt ein höherer Schwellenwert. Die Dau-er beider liegt bei etwa 2 ms, insgesamt also etwadoppelt so lang wie die Dauer des Aktionspotenzi-als.
7.3.6 Das Hodgkin-Huxley Modell
außen
innen
Cm Vm
Abbildung 7.20: Modell für die Leitfähigkeit derMembran nach Hodgking undHuxley.
Im Hodgkin-Huxley Modell ist die Spannung durchdie Membran durch die Nernst-Spannungen V00 derdrei wichtigsten Ionensorten gegeben, sowie durchdie geregelte Leitfähigkeiten g für die drei Ionen-sorten. Die Membrankapazität Cm liegt bei ⇡ 10�2
Fm�2.
Die Änderung des Membranpotenzials von etwa -75mV auf etwa +50 mV benötigt bei einer Membran-kapazität von Cm ⇡ 10�2 Fm�2 eine Ladung vonDV Cm ⇡ 1,25 ·10�3 Cm�2 oder
N =1,25 ·10�3 ·10�12
1,6 ·10�19 ⇡ 104
Ionen pro µm2. Dies entspricht einem relativ gerin-gen Anteil der vorhandenen Ionen.
Das Huxley-Hodgkin Modell beschreibt die Strom-dichte durch die Membran mit der Differentialglei-chung
j = CmdVm
dt+GKn4(Vm �Voo,K) (7.1)
+GNam3h(Vm �Voo,Na).
Hier stellen GK und GNa die maximale Leitfähig-keit für die beiden Ionensorten dar und n(t,Vm),m(t,Vm) und h(t,Vm) sind phänomenologische Funk-tionen, welche Leitfähigkeit als Funktion der Zeitund der Spannung beschreiben. Ihr Wert liegt zwi-schen 0 und 1. Beim Na stellt m die Wahrschein-lichkeit für die Aktivierung des Ionenkanals dar, hdie Wahrscheinlichkeit für die Deaktivierung. IhreWerte konnten für unterschiedliche Neuronen expe-rimentell bestimmt werden.
7.3.7 Messungen
Glaspipette
”Gigaseal”
Zellmembran
Ionenkanäle
Abbildung 7.21: Messung von einzelnen Ionenkanä-len mit Hilfe der Patch-ClampTechnik.
Der Strom durch einen einzelnen Ionenkanal kannmit Hilfe der sogenannten ”patch-clamp” Technikgemessen werden. Da der Widerstand eines einzel-nen Ionenkanals im Bereich von 1010 W liegt, mussdie Mikropipette einen sehr hohen Widerstand zwi-schen dem Inneren und dem Äußeren von � 1012 Waufweisen. Dafür muss die Zelle mit einem erhebli-chen Unterdruck angesogen werden.
Der Ionenstrom durch einen geöffneten Kanal liegtim Bereich von einigen pA.
Das Aktionspotenzial kann ebenfalls optisch gemes-sen werden, über die Änderung der Fluoreszenz vonMolekülen, die auf die Änderung des elektrischenFeldes reagieren. Der Kurvenverlauf in Abb. 7.23entspricht dem in Abb. 7.18 qualitativ gezeigten Ver-lauf und zeigt unter anderem den subkritischen Be-reich und das Nachpotenzial.
161
7 Das Nervensystem
Zeit [ms]
Kanal o!en
Kanal geschlossen
Abbildung 7.22: Strom durch einen einzelnen Io-nenkanal in geöffnetem oder ge-schlossenem Zustand.
Zeit [ms]
Abso
rptio
n [w
illk.
Ein
h.]
0 1 2 3 4
Abbildung 7.23: Messung des Aktionspotenzi-als über die Absorption einesFarbstoffs.
7.4 Signalübertragung
7.4.1 Erregungsleitung
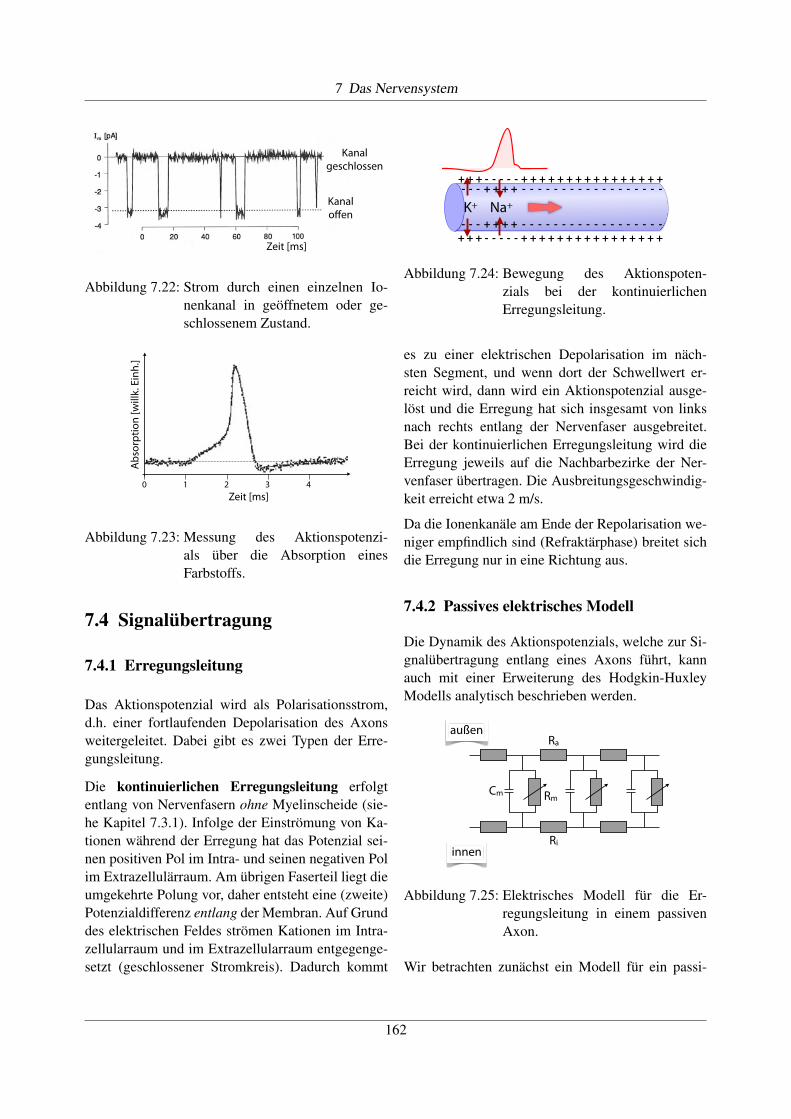
Das Aktionspotenzial wird als Polarisationsstrom,d.h. einer fortlaufenden Depolarisation des Axonsweitergeleitet. Dabei gibt es zwei Typen der Erre-gungsleitung.
Die kontinuierlichen Erregungsleitung erfolgtentlang von Nervenfasern ohne Myelinscheide (sie-he Kapitel 7.3.1). Infolge der Einströmung von Ka-tionen während der Erregung hat das Potenzial sei-nen positiven Pol im Intra- und seinen negativen Polim Extrazellulärraum. Am übrigen Faserteil liegt dieumgekehrte Polung vor, daher entsteht eine (zweite)Potenzialdifferenz entlang der Membran. Auf Grunddes elektrischen Feldes strömen Kationen im Intra-zellularraum und im Extrazellularraum entgegenge-setzt (geschlossener Stromkreis). Dadurch kommt
- - - - - - - - -- - - - - -- - - - - -+ + + ++ + + + + + + + ++ + + + + ++ + + +- - - - -
+ + + + + + + + ++ + + + + ++ + + +- - - - -- - - - - - - - -- - - - - -- - - - - -+ + + +K+ Na+
Abbildung 7.24: Bewegung des Aktionspoten-zials bei der kontinuierlichenErregungsleitung.
es zu einer elektrischen Depolarisation im näch-sten Segment, und wenn dort der Schwellwert er-reicht wird, dann wird ein Aktionspotenzial ausge-löst und die Erregung hat sich insgesamt von linksnach rechts entlang der Nervenfaser ausgebreitet.Bei der kontinuierlichen Erregungsleitung wird dieErregung jeweils auf die Nachbarbezirke der Ner-venfaser übertragen. Die Ausbreitungsgeschwindig-keit erreicht etwa 2 m/s.
Da die Ionenkanäle am Ende der Repolarisation we-niger empfindlich sind (Refraktärphase) breitet sichdie Erregung nur in eine Richtung aus.
7.4.2 Passives elektrisches Modell
Die Dynamik des Aktionspotenzials, welche zur Si-gnalübertragung entlang eines Axons führt, kannauch mit einer Erweiterung des Hodgkin-HuxleyModells analytisch beschrieben werden.
außen
innen
Cm Rm
Ra
Ri
Abbildung 7.25: Elektrisches Modell für die Er-regungsleitung in einem passivenAxon.
Wir betrachten zunächst ein Modell für ein passi-
162

7 Das Nervensystem
ves Axon, also für den Fall, dass die Spannung nichtausreicht für die Erregung eines Aktionspotenzials.Im Modell, das in Abb. 7.25 gezeigt ist, besteht derStrom durch die Membranwand aus einem kapaziti-ven Anteil und einem resistiven Anteil. Der Wider-stand Rm der Membran ist normiert auf die Längedes Abschnitts und hat damit die Dimension [Rm] =Wm. Die Widerstände Ra und Ri außerhalb, respekti-ve innerhalb des Axons, haben dagegen die Dimen-sion [Ra] = [Ri] = W/m.
Anwendung des Kirchhoff’schen Gesetzes zeigt,dass der Strom durch die Membran pro Längenein-heit
Im =Vm
Rm= �Cm
∂Vm
∂ t+
1Ri
∂
2Vm
∂x2 (7.2)
ist. Hier wurde angenommen, dass der Widerstandauf der Außenseite des Axons gering sei, Ra ! 0.Dies entspricht einer Diffusionsgleichung
1l
2∂
2Vm
∂x2 � t
∂Vm
∂ t= Vm.
Die beiden neu eingeführten Größen
l =
rRm
Ri, t = CmRm (7.3)
haben die Dimension einer Länge und einer Zeit.Das Verhältnis l
2/t = D entspricht einem Diffusi-onskoeffizienten.
Eine stationäre Lösung dieser Gleichung erhält man,wenn die Spannung an einem Ort x0 auf dem Wert V0festgehalten wird. In diesem Fall fällt die Spannungmit dem Abstand von diesem Punkt exponentiell ab,
Vm(x) = V0e�|x�x0|l .
Die mittlere Ausbreitungsgeschwindigkeit für eineunterschwellige Anregung ist damit
v̄ =
r2Dt
=p
2l
t
=
s2
RiRmC2m
. (7.4)
Da v̄ mit abnehmendem Innenwiderstand Ri zu-nimmt und dieser indirekt proportional zur Quer-schnittsfläche des Axons ist, ist die Signalübertra-gung in Axonen mit großem Durchmesser schneller.Beim Tintenfisch beträgt deshalb der Durchmessermehr als 200 µm.
7.4.3 Aktives Modell
Das Modell kann mit dem Hodgkin-Huxley Modellkombiniert werden, indem wir in Gleichung (7.2) dieStromdichte ersetzen durch Ausdruck (7.1):
ImRm = 2pr jRm = �t
∂Vm
∂ t+l
2 ∂
2Vm
∂x2
= 2prRm⇥GKn4(Vm �Voo,K)
+GNam3h(Vm �Voo,Na)⇤.
Hier stellt j die Stromdichte durch die Membrandar und r den Radius des Axons. Diese nichtlineareraum-zeitliche Differentialgleichung ist nicht analy-tisch, sondern nur numerisch lösbar.
Abbildung 7.26: Entwicklung der Pulse imNagumo-Modell und Gleich-gewichts-Amplituden.
Numerische Lösungen zeigen, dass auf diese Bewe-gungsgleichungen die Bildung von Pulsen beschrei-ben, welche oberhalb eines Schwellenwertes stabileFormen haben. Unterhalb des Schwellenwertes wer-den sie exponentiell gedämpft.
7.4.4 Saltatorische Erregungsleitung
Bei Menschen (allgemein bei höheren Organismen)sind die meisten Axone im peripheren Nervensy-stem von einer Myelinscheide mit einer Dicke voneinigen µm umhüllt. Diese werden gebildet durchdie Schwann’schen4 Zellen, eine spezielle Form derGliazellen.
Myelin ist eine Lipidschicht, welche als Isolatorwirkt. Sie reduziert damit die Leitfähigkeit der
4Theodor Schwann (1810–1882)
163

7 Das Nervensystem
Raniver-Schnürring
Myelin MyelinscheideSchwannzelle
Zellkern der Schwannzelle
Axon
Abbildung 7.27: Struktur eines Axons mitMyelinscheide.
Axon-Wand wie auch die Kapazität. Gemäß Glei-chung (7.4) ist die Ausbreitungsgeschwindigkeit
v̄ =1
Cm
r2
RiRm.
Für gegebene Materialeigenschaften steigt die Aus-breitungsgeschwindigkeit mit der Wurzel aus demQuerschnitt des Axons, v̄ µ
pr. Eine hohe Ge-
schwindigkeit könnte somit nur durch sehr großeDurchmesser erkauft werden. Die Myelinscheide er-niedrigt jedoch die Kapazität der Membran um 2-3 Größenordnungen und erhöht somit die Ausbrei-tungsgeschwindigkeit um den gleichen Betrag. Dagleichzeitig der Membranwiderstand erhöht wird,findet ein Ionenaustausch praktisch nur im Bereichder Schnürringe statt. Der erhöhte Membranwider-stand erhöht laut Gleichung (7.3) auch die Reichwei-te l der Depolarisation. Dadurch wird es möglich,die Distanz bis zum nächsten Schnürring zu über-springen.
+ ++ +
- -- -+ ++ +
Internodium (isolierend)
Abbildung 7.28: Saltatorische Erregungsleitung.
Die Myelinscheide wird in Abständen von eini-gen mm von den Ranvier’schen Schnürringen un-
terbrochen. Dadurch “springt” die Potenzialände-rung jeweils von einem Schnürring zum nächsten.Die Myelinscheide trägt deshalb zu einer wesent-lichen Beschleunigung der Nervenleitung bei: dieSprunglänge nimmt zu, die Kapazität der Mem-bran nimmt ab und damit die Zeitkonstante. Außer-dem benötigt die Signalübertragung dadurch weni-ger Energie. Es können Übertragungsgeschwindig-keiten von bis zu 180 m/s erreicht werden. Man be-zeichnet diese Art der Signalübertragung als salta-torische Erregungsleitung.
7.4.5 Effizienz
Wir können den Energiebedarf für die Ausbreitungeines Nervenimpulses über eine Distanz von 1 mmaus den elektrischen Eigenschaften abschätzen. Dieelektrische Kapazität wird dominiert von den Berei-chen der Ranvier’schen Schnürringe. Pro Abschnittliegt sie bei etwa C1 ⇡ 10�12 F. Die Energie, welchebenötigt wird, um diesen Kondensator um DV =0,11V aufzuladen, beträgt
E1 =12
DV 2C1 ⇡ 6 ·10�15 J. (7.5)
Insgesamt erlaubt der Übergang von der konti-nuierlichen zur saltatorischen Erregungsleitung ei-ne Erhöhung der Übertragungsgeschwindigkeit beigleichzeitiger Reduktion des Axon-Querschnitts:
Tintenfisch FroschSpez.Kapazität
10�2 Fm�2 2,5 ·10�5 Fm�2
Axon-Radius 240 µm 7 µmAusbreitungs-geschwindigk.
20 m/s 100 m/s
Diese Erhöhung der Übertragungseffizienz war einewesentliche Voraussetzung zur Entwicklung von hö-heren Organismen.
Schwann’sche Zellen wirken als Signalbeschleuni-ger, da die Polarisationswolke über die nichtleiten-den Schwann’schen Zellen springen müssen (salta-torische Leitung).
164

7 Das Nervensystem
Bei der Multiplen Sklerose - MS - zerfallen dieMyelinscheiden im Zentralnervensystem, bevorzugtum die Hirnventrikel. Dadurch wird die Erregungs-leitung verzögert und zum Teil sogar unterbrochen.Folge sind Störungen in allen Teilen des ZNS.
Es gibt Hinweise, dass Nervenimpulse nicht reinelektrisch übertragen werden, sondern dass auch me-chanische Prozesse involviert sind, so dass parallelzum elektrischen Impuls eine akustische Welle durchdem Axon entlang läuft.
7.4.6 Motoneuronen
Muskelzelle
Zellkern
Raniver’scher Schnürring
Axon
Schwann-Zelle
marklose Nervenfaser
Dendriten
Myelinscheide
Motorische Endplatte
Abbildung 7.29: Motoneuron.
Typische Beispiele von efferenten Nerven sind die-jenigen, welche den Bewegungsapparat steuern. DerZellkörper dieser Motoneuronen befindet sich meistim Rückenmark. Mehrere Axone verlassen dasRückenmark , teilen sich und laufen zu den einzel-nen Muskelzellen. Am Ende eines solchen Axonssteht eine spezielle Synapse, welche das Signal aufdie Muskelzelle überträgt. Sie wird als motorischeEndplatte bezeichnet und schüttet beim Eintreffeneines Aktionspotenzials den Neurotransmitter Ace-tylcholin aus, welches die Ionenkanäle in der Mus-kelfaser öffnet. Ein Motorneuron und die zugehöri-gen Muskelfasern (typischerweise 100-2000) stellenzusammen eine “motorische Einheit” dar.
parietalpräzentral
frontaltemporal
okzipital
Abbildung 7.30: Bezeichnung der wichtigsten Ge-hirnregionen.
7.5 Das Gehirn
7.5.1 Anatomie
Das Gehirn umfasst 2-3% des Körpergewichts, esgehört aber zu den aktivsten Organen und hatdementsprechend einen hohen Energieumsatz. Da-für benötigt es etwa 15% der Herzleistung, 20% desSauerstoffs und 25% der gesamten Glukose. Es be-steht aus 1012 Neuronen, welche über etwa 1014 Syn-apsen verbunden sind. Die Nervenbahnen im Cortexhaben keine Schwann‘schen Zellen.
Neben den Nervenzellen existiert eine vergleichba-re Zahl von Gliazellen. Diese sind kleiner als dieNervenzellen. Sie wurden in der Mitte des 19. Jahr-hunderts entdeckt und man vermutete zunächst, dasssie eine vor allem mechanische Unterstützung für dieNervenzellen darstellen. Später konnte gezeigt wer-den, dass Gliazellen maßgeblich am Stoff- und Flüs-sigkeitstransport sowie an der Aufrechterhaltung derHomöostase im Gehirn und an der Informationsver-arbeitung beteiligt sind.
Die Oberfläche des Gehirns entspricht einem Durch-messer von etwa 0,4 m, ist aber stark gefaltet, sodass sie im Schädel Platz findet. Der Faltungspro-zess kann mit Hilfe eines mechanischen Modells be-schrieben werden, welches davon ausgeht, dass dieNervenzellen unter Spannung stehen [6].
Im Gehirn sind bestimmte Bereiche des sensorischenwie auch des motorischen Cortex mit bestimmtenKörperregionen verbunden, wie in Abb. 7.31 dar-gestellt. Wie man aus der Größe der zugeordneten
165

7 Das Nervensystem
Sensorischer Cortex Motorischer Cortex
Abbildung 7.31: Zuordnung der Gehirnregionen imsensorischen und motorischen Cor-tex.
Bereiche sieht, sind beispielsweise Hand und Mundsehr viel stärker repräsentiert als andere Bereichewie die Beine. Die neuronale Repräsentation vonKörperregionen wird als Somatotopie bezeichnet.
7.5.2 Das Gehirn als physikalisches System
Das Gehirn erfüllt ähnliche Funktionen wie einRechner; es funktioniert jedoch zum Teil sehr vielanders. Die Anzahl der Neuronen (⇡ 1012) ist we-sentlich höher als die der Zahl der Elemente in ei-nem Rechnerchip (⇡ 109). Der größere Unterschiedliegt aber in der Zahl der Verbindungen (⇡ 1014 Syn-apsen) und in der Art der Verbindungen: im Ge-hirn kommunizieren einzelne Nervenzellen direkt,im Rechner kommunizieren die meisten Teile überein Bussystem, welches jeweils nur für eine Aufgabezur Verfügung steht. Ein typischer Mikroprozessorkann pro Sekunde etwa 1010 Operationen durchfüh-ren. Im Gehirn kann jede Synapse 10-100 mal proSekunde einen Impuls übertragen, insgesamt im Ge-hirn also etwa 1015 Operationen. Dies, obwohl dieeinzelnen Operationen sehr viel langsamer sind (⇡ 1ms) als bei einem Mikroprozessor, und es sind rela-tiv viele elementare Operationen notwendig, um z.B.zum Resultat einer einfachen Rechnung (2+2=4) zugelangen. Dafür laufen im Gehirn viele Prozesse par-allel, da die unterschiedlichen Gehirnregionen nichtsehr stark von einander abhängig sind.
Der Energieverbrauch des Gehirns (⇡ 20 W) istvergleichbar mit dem eines Mikroprozessors, sehrviel mehr als bei einem einfachen Taschenrechner,aber sehr viel weniger als bei einem Supercom-puter, der eine vergleichbare ‘Rechenleistung’ auf-weist für komplexe Probleme, wie z.B. das Schach-spiel. Betrachten wir die Energie E1 ⇡ 6 · 10�15 Jfür die Übertragung eines Signals zwischen zweiRanvier’schen Schnürringen als elementaren Prozess(siehe Gl. 7.5), so kann das Gehirn auf Grund desEnergieverbrauchs bis zu 1015 elementare Operatio-nen pro Sekunde durchführen - immer noch deutlichmehr als ein Mikroprozessor.
Die Schaltkreise des Gehirns sind dreidimensional.Dadurch sind viel mehr direkte Verbindungen mög-lich. Es besteht kein Bus-System. Dadurch könnensehr viel mehr Aufgaben parallel erledigt werden.
Das Gehirn ist plastisch: Verbindungen werden lau-fend neu geschaltet. Dadurch stellt sich das Gehirnauf neue Anforderungen ein und erlernt neue Fähig-keiten. Damit können auch die Aufgaben von be-schädigten Hirnarealen auf andere Bereich übertra-gen werden. Im menschlichen Nervensystem wirdsomit die Hardware auf die Anforderungen ange-passt. In einem Rechner hingegen ist die Hardwa-re für alle Teile (im Wesentlichen) identisch. Hierwird die Software angepasst, um die entsprechendenSteuerungen zu implementieren. Die DNA, also derBauplan für das Gehirn, definiert nicht die Lage dereinzelnen Neuronen und ihre Verbindungen.
Das Gehirn verhält sich wie ein kritisches System.Dies wird unter anderem durch die Zahl von Neu-ronen gezeigt, welche auf einen Stimulus reagieren,oder durch die Korrelationslänge von Gehirnaktivie-rungen. In einem solchen Zustand können Umwelt-reize einen großen Effekt haben und den Zustand desSystems rasch und stark verändern.
Im Rahmen des EU-Flagship Projekts “The HumanBrain Project” soll das Gehirn auf einem Supercom-puter simuliert werden. 5 Das Projekt soll minde-stens 10 Jahre laufen und pro Jahr mit bis zu 100MC finanziert werden.
5http://www.humanbrainproject.eu
166

7 Das Nervensystem
7.5.3 Neuron-Silizium Kopplung
Abbildung 7.32: Nervenzellen auf einem Silizium-Chip.
Abb. 7.32 zeigt einen Chip, welcher mit Nervenzel-len in Kontakt steht. Solche Kontakte wurden zu-nächst mit Blutegel-Neuronen auf Silizium herge-stellt, später mit Schnecken-Neuronen, welche sichdurch ihre Größe noch besser zur Untersuchung eig-nen, und schließlich mit Säugetier-Neuronen.
Hippocampus der Ratte
Die Elektroden sollen Kontakte
herstellen
Ausgangs-elektrode
Eingangselektrode
Abbildung 7.33: Experimentelle Hippocampus-Prothese. [New Scientist]
Solche Arbeiten sollen es erlauben, ausgefalleneHirnfunktionen zu ersetzen. Als Beispiel wurde aneiner Ratte ein Chip eingesetzt, welcher die Funktioneines geschädigten Bereichs des Hippocampus über-nehmen soll. Der Hippocampus codiert eingehendeErfahrungen und bereitet sie auf für die Speicherungim Langzeitgedächtnis. Fällt er aus, fällt das Lang-zeitgedächntis aus.
167