Embed Size (px)
Citation preview

A sketch of chemistry behind known carbon-based life on earth
http://www.nap.edu/catalog/11919.html
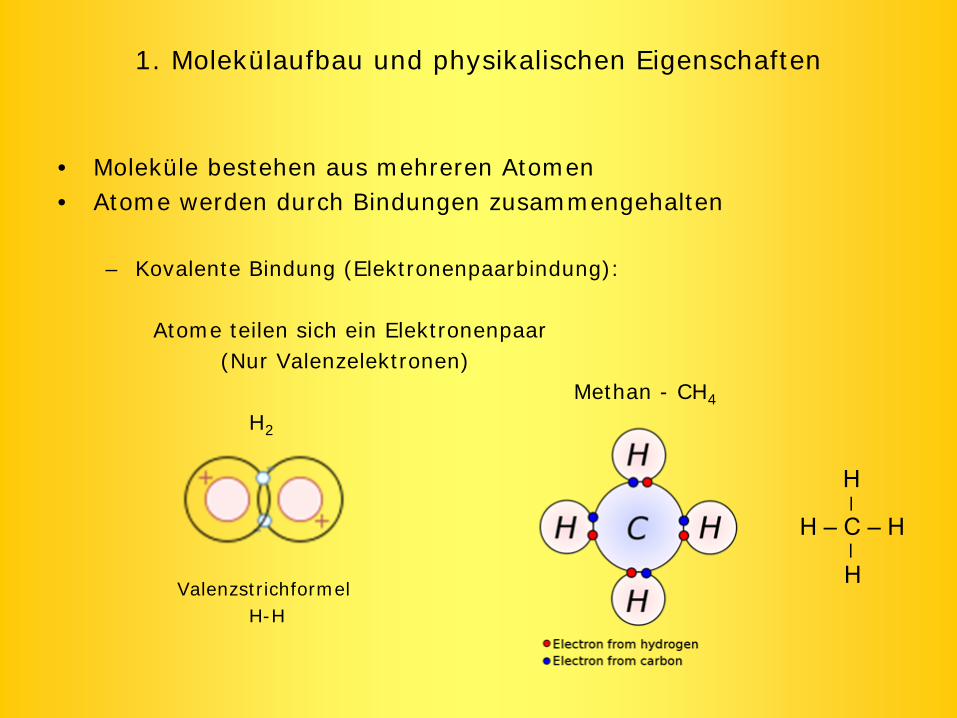
1. Molekülaufbau und physikalischen Eigenschaften
• Moleküle bestehen aus mehreren Atomen• Atome werden durch Bindungen zusammengehalten
– Kovalente Bindung (Elektronenpaarbindung):
Atome teilen sich ein Elektronenpaar(Nur Valenzelektronen)
Methan - CH4
H2
ValenzstrichformelH-H
H – C – H__H
H
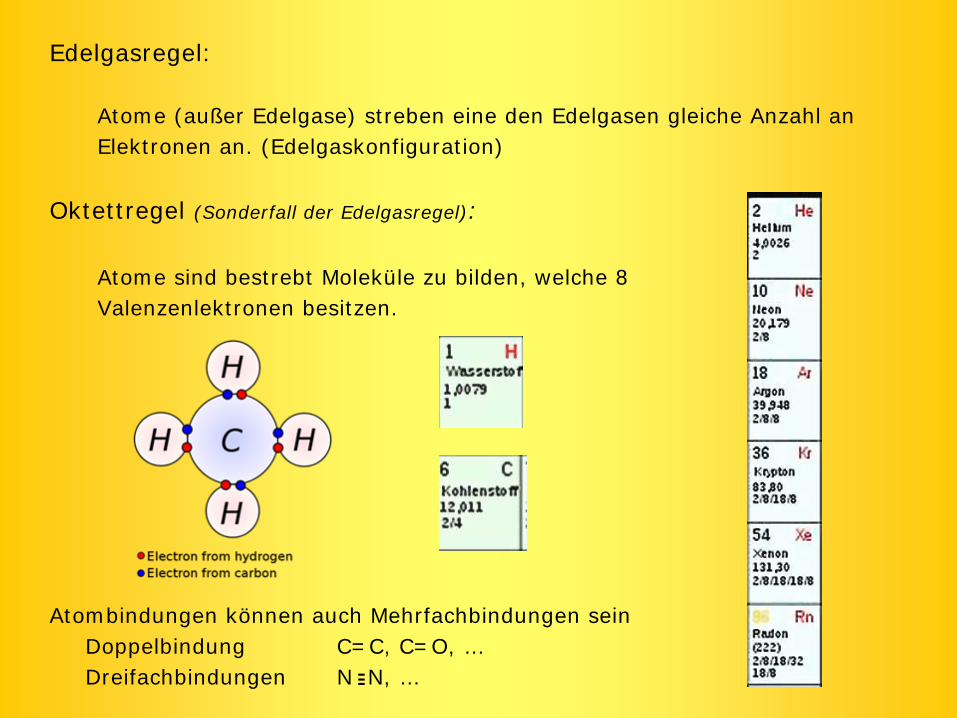
Edelgasregel:
Atome (außer Edelgase) streben eine den Edelgasen gleiche Anzahl anElektronen an. (Edelgaskonfiguration)
Oktettregel (Sonderfall der Edelgasregel):
Atome sind bestrebt Moleküle zu bilden, welche 8 Valenzenlektronen besitzen.
Atombindungen können auch Mehrfachbindungen seinDoppelbindung C=C, C=O, …Dreifachbindungen N N, …---

• Ladungsverteilung führt zu den physikalischen Eigenschaften
– Moleküle mit gleicher Anzahl an Protonen/Elektronen sind elektrisch neutral.
– Allerdings müssen die Elektronen nicht gleichmäßig im 3D-Raum verteilt sein
Dipolmomenten
Elektronegativität:
• Bestimmt durch die Kernladungszahl Z=n*e und dem Atomradius r• Anhaltspunkt für die Polarität• Atome mit ähnlicher EN teilen sich die Elektronen gleichmäßig
Relatives Maß für die Fähigkeit das Bindungselektronenpaar an sich ziehen zu können !
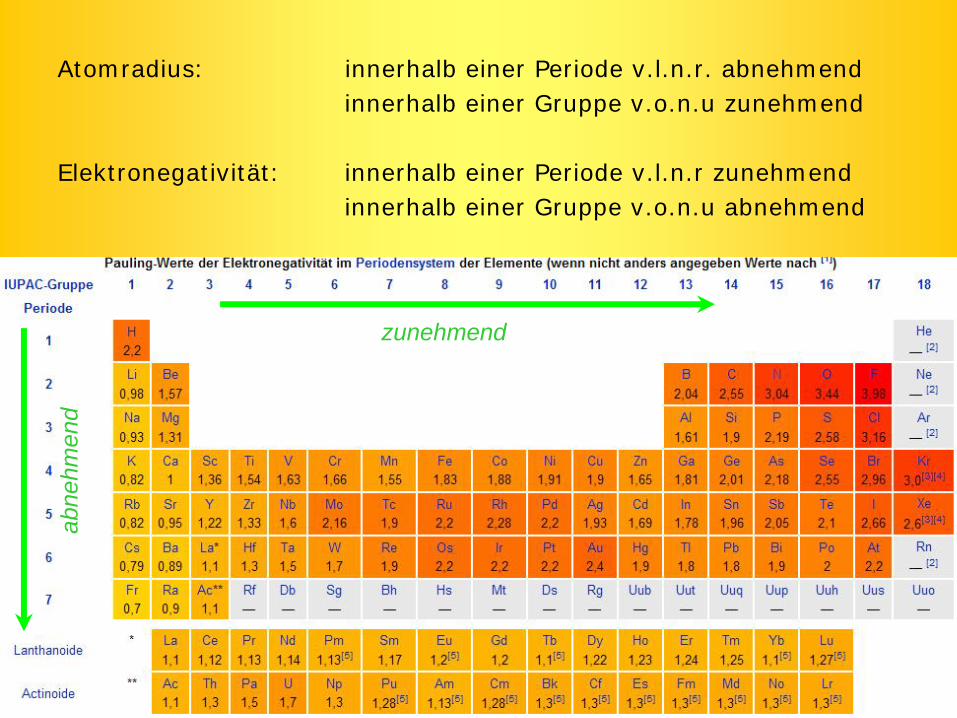
Atomradius: innerhalb einer Periode v.l.n.r. abnehmendinnerhalb einer Gruppe v.o.n.u zunehmend
Elektronegativität: innerhalb einer Periode v.l.n.r zunehmendinnerhalb einer Gruppe v.o.n.u abnehmend
zunehmend
abne
hmen
d
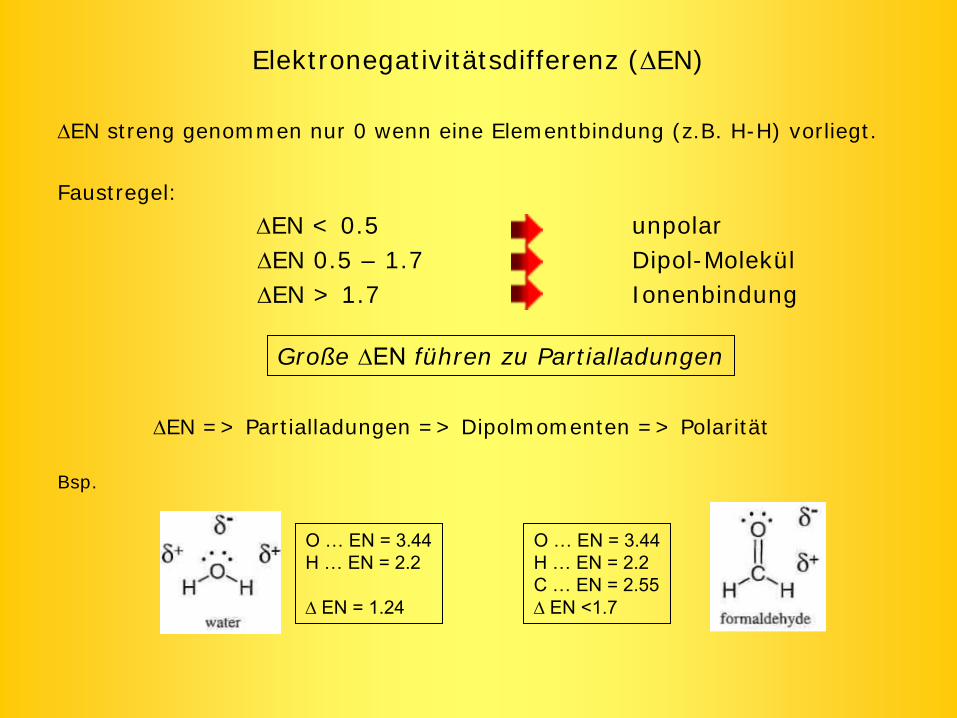
Elektronegativitätsdifferenz (EN)
EN streng genommen nur 0 wenn eine Elementbindung (z.B. H-H) vorliegt.
Faustregel:
EN < 0.5 unpolarEN 0.5 – 1.7 Dipol-MolekülEN > 1.7 Ionenbindung
EN => Partialladungen => Dipolmomenten => Polarität
Bsp.
Große EN
führen zu Partialladungen
O …
EN = 3.44H …
EN = 2.2
EN = 1.24
O …
EN = 3.44H …
EN = 2.2C …
EN = 2.55EN <1.7
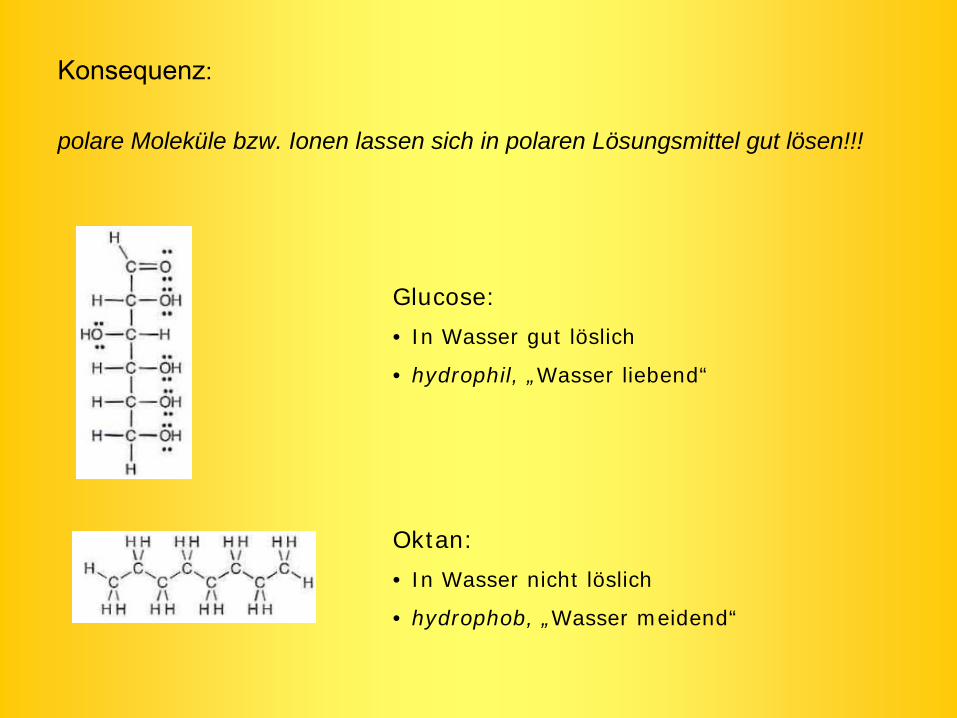
Konsequenz:
polare Moleküle bzw. Ionen lassen sich in polaren Lösungsmittel gut lösen!!!
Glucose:
• In Wasser gut löslich
• hydrophil, „Wasser liebend“
Oktan:
• In Wasser nicht löslich
• hydrophob, „Wasser meidend“

2. Molekülreaktionen
• Moleküle mit C-H bzw. C-C Bindungen reagieren bei Standard- temperatur rel. schlecht. (Standardtemperatur: Wasser flüssig bei 1 Atm)
Abhilfe:
d.h. C-Heteroatome aktivieren C-C und C-H Bindungen.
Die wichtigsten Heteroatomen in irdischen biochemischen Reaktionensind:
O, N, S
C-H bzw. C-C Bindungen mit Molekülen, welche C-Heteroatomen beinhalten, reagieren lassen
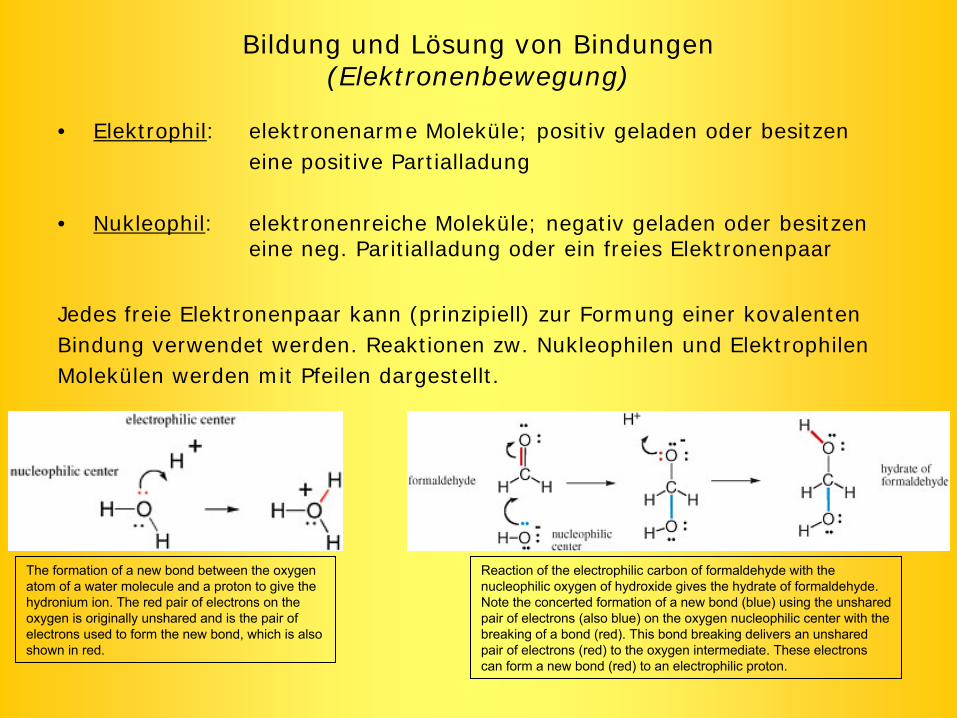
Bildung und Lösung von Bindungen (Elektronenbewegung)
• Elektrophil: elektronenarme Moleküle; positiv geladen oder besitzen eine positive Partialladung
• Nukleophil: elektronenreiche Moleküle; negativ geladen oder besitzen eine neg. Paritialladung oder ein freies Elektronenpaar
Jedes freie Elektronenpaar kann (prinzipiell) zur Formung einer kovalentenBindung verwendet werden. Reaktionen zw. Nukleophilen und ElektrophilenMolekülen werden mit Pfeilen dargestellt.
Reaction
of the
electrophilic
carbon
of formaldehyde
with
the
nucleophilic
oxygen
of hydroxide
gives
the
hydrate
of formaldehyde. Note the
concerted
formation
of a new
bond
(blue) using
the
unshared
pair of electrons
(also blue) on the
oxygen
nucleophilic
center
with
the
breaking
of a bond
(red). This
bond
breaking
delivers
an unshared
pair of electrons
(red) to the
oxygen
intermediate. These electrons
can
form a new
bond
(red) to an electrophilic
proton.
The
formation
of a new
bond
between
the
oxygen
atom
of a water
molecule
and a proton
to give
the
hydronium
ion. The
red pair of electrons
on the
oxygen
is
originally
unshared
and is
the
pair of electrons
used
to form the
new
bond, which
is
also shown
in red.
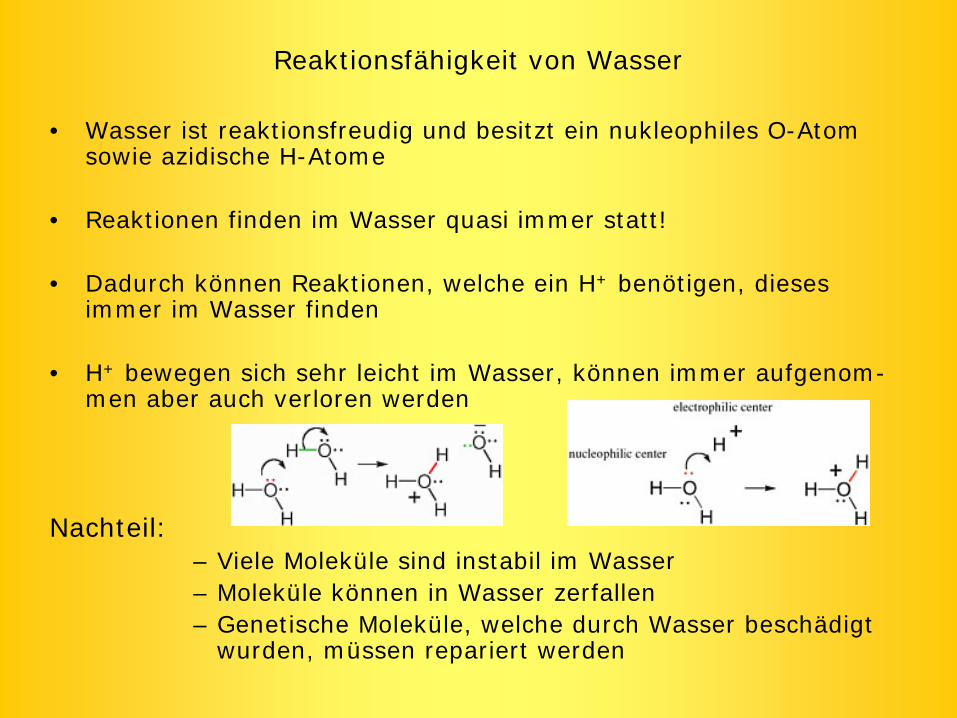
Reaktionsfähigkeit von Wasser
• Wasser ist reaktionsfreudig und besitzt ein nukleophiles O-Atom sowie azidische H-Atome
• Reaktionen finden im Wasser quasi immer statt!
• Dadurch können Reaktionen, welche ein H+ benötigen, dieses immer im Wasser finden
• H+ bewegen sich sehr leicht im Wasser, können immer aufgenom- men aber auch verloren werden
Nachteil:– Viele Moleküle sind instabil im Wasser– Moleküle können in Wasser zerfallen– Genetische Moleküle, welche durch Wasser beschädigt
wurden, müssen repariert werden
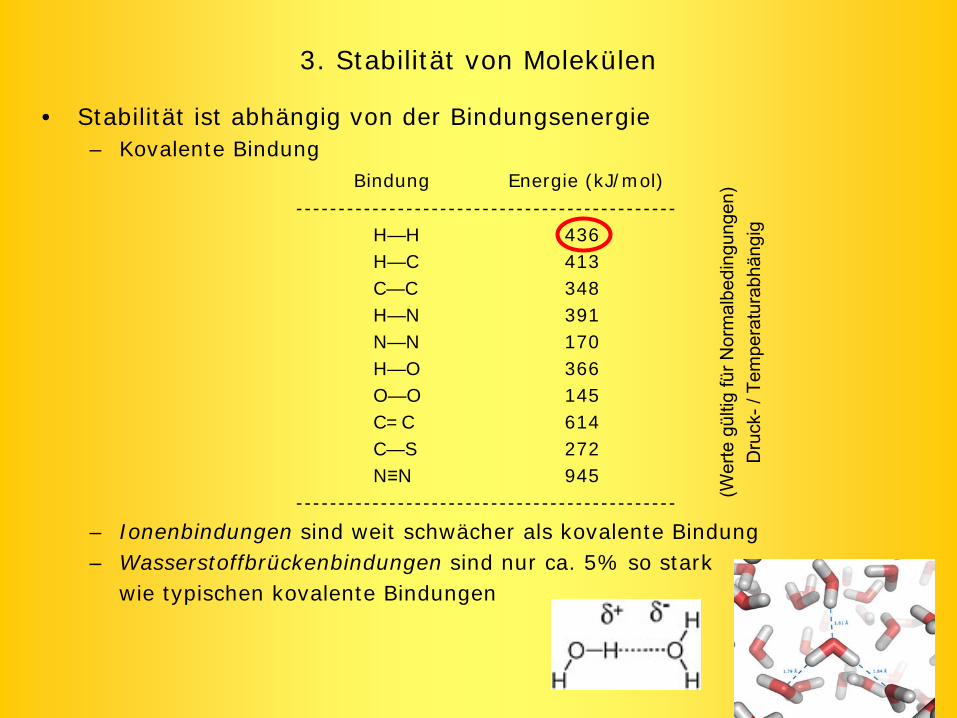
3. Stabilität von Molekülen
• Stabilität ist abhängig von der Bindungsenergie– Kovalente Bindung
Bindung Energie (kJ/mol)---------------------------------------------
H—H 436H—C 413C—C 348H—N 391N—N 170H—O 366O—O 145C=C 614C—S 272N≡N 945
---------------------------------------------
– Ionenbindungen sind weit schwächer als kovalente Bindung– Wasserstoffbrückenbindungen sind nur ca. 5% so stark
wie typischen kovalente Bindungen
(Wer
te g
ültig
für N
orm
albe
ding
unge
n)D
ruck
-/ T
empe
ratu
rabh
ängi
g
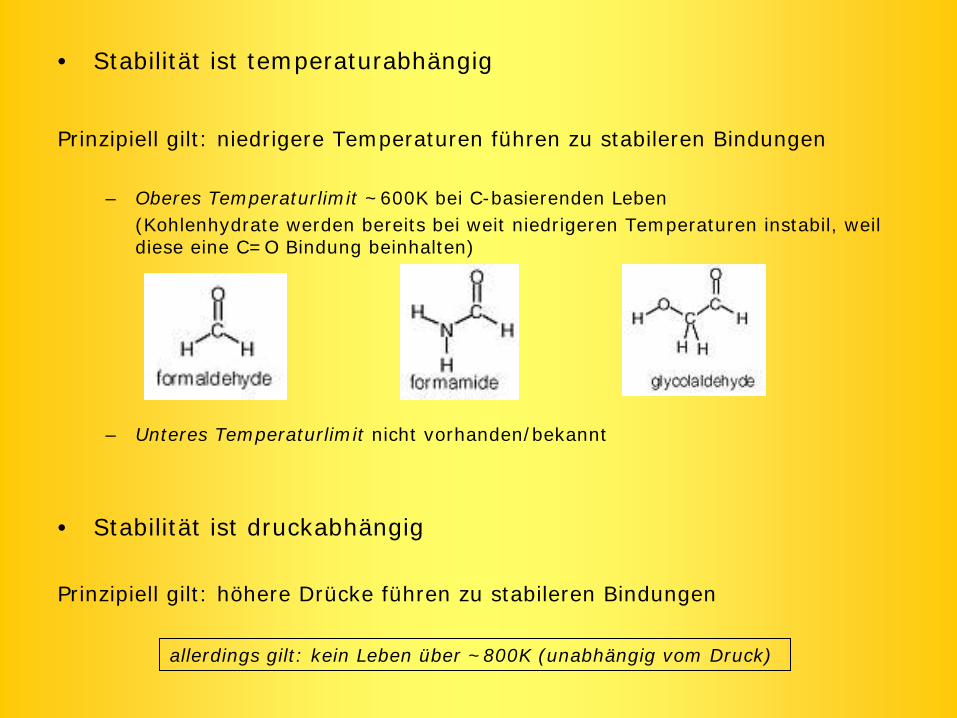
• Stabilität ist temperaturabhängig
Prinzipiell gilt: niedrigere Temperaturen führen zu stabileren Bindungen
– Oberes Temperaturlimit ~600K bei C-basierenden Leben(Kohlenhydrate werden bereits bei weit niedrigeren Temperaturen instabil, weil diese eine C=O Bindung beinhalten)
– Unteres Temperaturlimit nicht vorhanden/bekannt
• Stabilität ist druckabhängig
Prinzipiell gilt: höhere Drücke führen zu stabileren Bindungen
allerdings gilt: kein Leben über ~800K (unabhängig vom Druck)
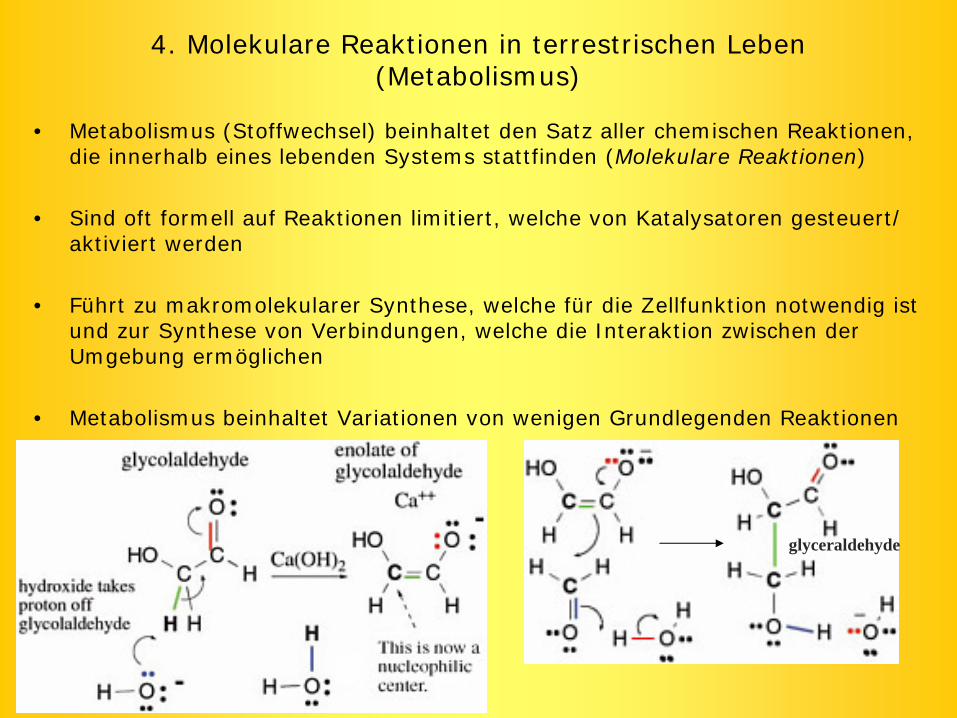
4. Molekulare Reaktionen in terrestrischen Leben (Metabolismus)
• Metabolismus (Stoffwechsel) beinhaltet den Satz aller chemischen Reaktionen, die innerhalb eines lebenden Systems stattfinden (Molekulare Reaktionen)
• Sind oft formell auf Reaktionen limitiert, welche von Katalysatoren gesteuert/ aktiviert werden
• Führt zu makromolekularer Synthese, welche für die Zellfunktion notwendig ist und zur Synthese von Verbindungen, welche die Interaktion zwischen der Umgebung ermöglichen
• Metabolismus beinhaltet Variationen von wenigen Grundlegenden Reaktionen
glyceraldehyde

Energiebedarf
• Metabolismus kann nicht im thermodynamischen GGW stattfinden• Molekülsynthese und die Erzeugung der Zellen benötigen Energie
Pflanzen – Energie durch Photosynthese aus Photonen
Tiere – Energie aus der Umwandlung von energiereiche Verbindungen, welchedurch die Nahrung aufgenommen wurden
• Energie wird üblicherweise nicht sofort benötigt, sondern gespeichert:
1. Durch Änderung der Konzentration von chemischen Spezies
(verschiedene immer miteinander in einem Reaktionsgleichgewicht auftretende Formen einer Substanz) (z.B. biologische Membrane)
2. In Molekülen (ATP-Adenosintriphosphat)
Die Energie erhalten die Organismen aus deren chemischen Umgebung, welche mitProzessen im Organismus gekoppelt sein müssen.

5. Katalysatoren
• Selbst durch die Aktivierung von C-C, C-H Verbindungen, laufen die Reaktionen sehr langsam ab
• Zur Beschleunigung der Reaktionen kommen Katalysatoren zur Anwendung• Diese können aus der Erbinformation abgeleitet werden und ermöglichen
dadurch die Genetik und den Darwinismus (Evolutionstheorie)
• Biologischen Polymerkatalysatoren nennt man Enzyme• Enzyme sind meist Proteine und diese exzellente Katalysatoren. Typische
Bakterien beinhalten ~2000 Proteinenzyme
• Enzyme steuern die biochemischen Reaktionen (Verdauung, kopieren und transkribieren der Erbinformation (DNA-Polymerase, RNA-Polymerase)
• Proteine verleihen den Zellen Struktur und transportieren Stoffe
Halbwertszeit für Reaktionen im Wasser Reaktion bei 25°C
---------------------------------------------------------------------------------------Hydration von Fumarsäure 7 × 105 JahreTriosephosphatisomerase 2 TageDekarboxylierung von Orotidine 8 × 107 JahreHydrolyse einer Peptidbindung 450 JahreHydrolyse einer Phosphodiesterbindung >13 × 106 Jahre
Beschleunigung zwischen 7 und 19 Größenordnungen

6. Makromolekulare Strukturen in terrestrischem Leben
Makromoleküle bestehen aus mehreren bis zu ca. 100.000 Bausteinen.Viele Funktionen irdischer Organismen werden durch Makromoleküle ermöglicht:
Proteine -> Katalyse, DNA -> Speicherung der vererbbaren Information, Doppellipidmembranen -> Zellkompartimentierung
• DNA ist gut geeignet weil:
– sie aus Doppelsträngen besteht und sich diese ergänzen (einfache Nachbildung)– die Information redundant vorkommt und im Falle einer Beschädigung eines Strangs
die Erbinformation nicht verloren ist– die Wechselwirkung zwischen den Strängen (durch Wasserstoffbrückenbindung)
stark genug ist, um die Struktur zu erhalten, aber nicht zu stark, um die Nachbildung der Stränge zu gewährleisten.
• Proteine sind gut geeignet wegen:
– der strukturellen Eigenschaft des Polymer der Aminosäuren
• die Polyamideverbindung der Proteine ist neutral• die Verbindung hat einen Dipol, um Wasserstoffbrückenbindungen zu bilden
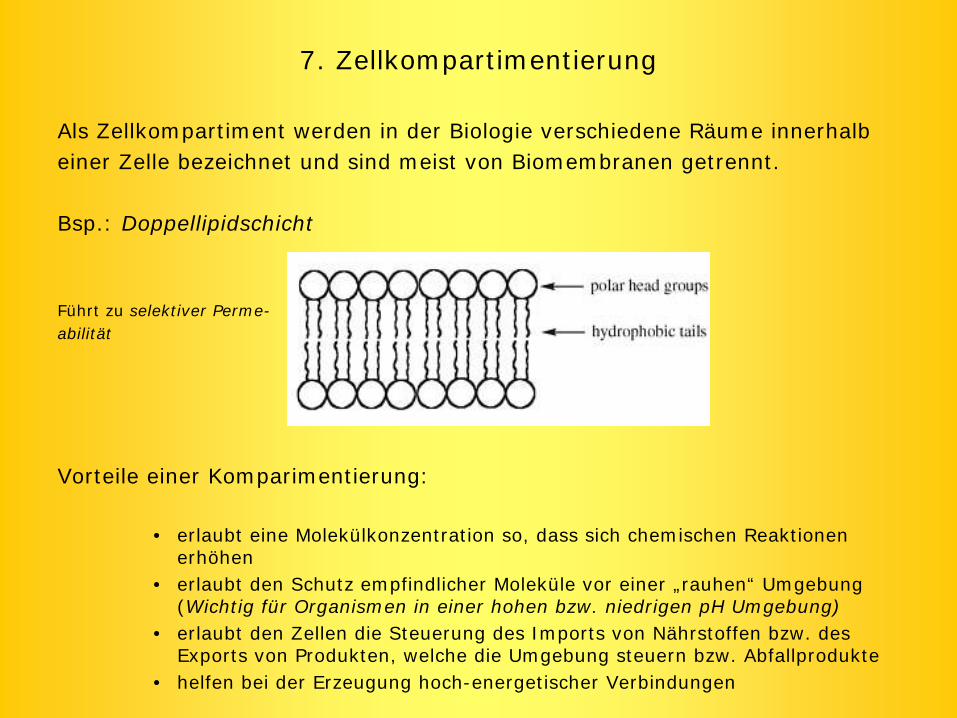
7. Zellkompartimentierung
Als Zellkompartiment werden in der Biologie verschiedene Räume innerhalb einer Zelle bezeichnet und sind meist von Biomembranen getrennt.
Bsp.: Doppellipidschicht
Führt zu selektiver Perme-abilität
Vorteile einer Komparimentierung:
• erlaubt eine Molekülkonzentration so, dass sich chemischen Reaktionen erhöhen
• erlaubt den Schutz empfindlicher Moleküle vor einer „rauhen“ Umgebung (Wichtig für Organismen in einer hohen bzw. niedrigen pH Umgebung)
• erlaubt den Zellen die Steuerung des Imports von Nährstoffen bzw. des Exports von Produkten, welche die Umgebung steuern bzw. Abfallprodukte
• helfen bei der Erzeugung hoch-energetischer Verbindungen

Zusammenfassung
1. Kovalente Bindungen ist die wesentliche Bindung in terrestrischem Leben
2. Kovalente Bindungen sind sehr stabil (hohe Energie) können durch Aktivierung gebrochen werden
3. Ladung ist der Schlüssel zu den Eigenschaften der Moleküle (Elektronegativität)
4. C-C, C-H Verbindungen sind unpolar -> schwer löslich -> reaktionsarm
5. Molekülreaktionen werden durch Heteroatome angeregt (aktiviert)
6. Wichtigsten Heteroatomen sind: O, N, S
7. Oberstes Temperaturlimit: ~800K
8. Katalysatoren zur Beschleunigung der Reaktionen notwendig
9. Proteine sind exzellente Katalysatoren
10. Zellkompartimentierung erfolgt durch Membrane (hydrophob und hydorphil)

Quellen:
Breslow
R., 1959, On the
mechanism
of the
formose
reaction, Tetrahedron Lett. 21:222-226; Butlerow, A., 1861, Bildung einer zuckerartigen Substanz durch Synthese. Annalen 120:295-298; and Ricardo, A., Carrigan, M.A., Olcott, A.N., and Benner, S.A., 2004, orate
minerals
tabilize
ibose, Science 03:1196.
Weber, A.L. 2001. The
sugar
model: Catalysis
by
amines
and amino
acid
products. Orig. Life Evol. Biosph. 31:71-886.
Wolfenden, R., and Snider, R. 2001.The depth
of chemical
time and the
power of enzymes
as catalysts. Accounts Chem. Res. 34:938-945.
Benner, S.A., and Hutter, D. 2002. Phosphates, DNA, and the
search
for
nonterran
life: A second generation
model
for
genetic
molecules. Bioorg. Chem. 30:62-880.Richert, C., Roughton, A.L., and Benner, S.A., 1996. Nonionic
analogs
of RNA with
dimethylsulfone
bridges. J. Am. Chem. Soc. 118:4518-4531.
Takano, Y., Marumo, K., Ebashi, T., Gupta, L.P., Kawahata, H., Kobayashi, K., Yamagishi, A., and Kuwubara, T. 2005. In situ
ore
formation
experiment: Amino
acids
and amino
sugars
trapped
in artificial
chimneys
on deep-ssea
hydrothermal systems
at Suiyo
Seamount, Izu-Bonin
Arc, Pacific Ocean. Bull. Chem. Soc. Jpn. 8:638-651.Martin, W., and Russell, M. 2003. On the
origin
of cells. A hypothesis
for
the
evolutionary
transitions
from
abiotic
geochemistry
to chemoautotrophic
prokaryotes, and from
prokaryotes
to nucleated
cells. Phil. Trans. Roy. Soc. Lond. 58:559-885.
Ban, N., Nissen, P., Hansen, J., Moore, P.B., and Steitz, T.A. 2000. The
complete
atomic
structure
of the
large ribosomal
subunit
at 2.4 Å
resolution. Science 89:905-920.Yusupov, M.M., Yusupova, G.Z., Baucom, A., Lieberman, K., Earnest, T.N., Cate, J.H.D., and Noller, H.F. 2001. Crystal
structure
of the
ribosome
at 5.5 Å
resolution. Science 292:883-896.Ban, N., Nissen, P., Hansen, J., Moore, P.B., and Steitz, T.A. 2000. The
complete
atomic
structure
of the
large ribosomal
subunit
at 2.4 Å
resolution. Science 89:905-920.Frick, L., Mac Neela
J.P., and Wolfenden, R. 1987. Transition
state
stabilization
by
deaminases. Rates of nonenzymic
hydrolysis
of adenosine
and cytidine. Bioorg. Chem. 5:100-108.Levy, M., and Miller, S.M. 1998. The
stability
of the
RNA bases: Implications
for
the
origin
of life. Proc. Natl. Acad. Sci. U.S.A. 95:7933-7938.