Embed Size (px)
Citation preview
BiomaterialienDOI: 10.1002/ange.201200591
Artifizielle Eierstiele, hergestellt aus rekombinant produziertemFlorfliegenseidenprotein**Felix Bauer und Thomas Scheibel*
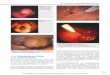
Seit Jahrzehnten hat Seide Wissenschaftler wegen ihrer me-chanischen Eigenschaften (Kombination aus Zugfestigkeit,Dehnbarkeit und Z�higkeit), ihrer Biokompatibilit�t undBioabbaubarkeit inspiriert. Mçgliche Anwendungen vonSeide reichen vom Fahrzeugbau (beispielsweise Airbags) biszu medizinischen Anwendungen (Nervenleitungen oderWirkstofftransport).[1–8] Florfliegen (Abbildung 1 A) verwen-den Seidenfasern als Eierstiele. Im Unterschied zu denmeisten anderen Seiden hat Eierstiel-Seide eine Cross-b-Struktur, die schon 1957 entdeckt wurde.[9] Seitdem wurdennur wenige zus�tzliche Cross-b-Seiden gefunden.[10] In derArt Chrysopa carnea enth�lt die Spinnlçsung haupts�chlichf�nf Proteine (Abbildung 1B), und in der Art Mallada signatawurden zwei Serin- und Glycin-reiche Proteine (MalXB1(86 kD) und MalXB2 (55.5 kD)) gefunden. Beide MalXB-Proteine haben hochrepetitive Kernsequenzen, flankiert vonkleinen terminalen Dom�nen.[11] Die vermeintliche Sekun-d�rstruktur der Kernsequenz beider Proteine ist b-Faltblatt-reich, mit einer Strangl�nge von vier Aminos�uren, gefolgtvon b-Schleifen (Abbildung 1 der Hintergrundinformatio-nen). Um einen Eierstiel zu produzieren, platziert die Flor-fliege einen Tropfen Spinnlçsung auf einer Oberfl�che unddr�ckt ein Ei in den Tropfen. Durch einfaches Ziehen am Eiwird ein Faden gezogen, der innerhalb weniger Sekundenaush�rtet. In ihrer nat�rlichen Umgebung h�ngen die Eier-stiele unter Bl�ttern, aber wenn man sie umdreht, verbiegensie sich nicht unter dem Gewicht des Eis (Abbildung 2B).Allerdings sind Eierstiele bei hoher Luftfeuchtigkeit bis zu600 % dehnbar.[11]
Um biomimetisch einen Eierstiel mit identischen Eigen-schaften zu produzieren, haben wir ein artifizielles Eierstiel-protein entworfen und rekombinant produziert. Durch denrepetitiven Charakter der bekannten Eierstielproteine war esmçglich, eine Konsensussequenz des Kernbereichs vonMalXB2 zu ermitteln, die wir [AS] nannten. Die Sequenzumfasst 48 Aminos�uren (Abbildung 1D; Abbildung 1 derHintergrundinformationen). Wie beim Herstellen von hochrepetitiven Spinnenseidenproteinen[12,13] wurde DNA, die f�r
dieses Modul kodiert, mithilfe einer nahtlosen Klonierungs-technik[12] multimerisiert, um die repetitive Sequenz nachzu-ahmen. Um ein f�r die Expression in E. coli optimiertes Genzu erhalten, wurden die Sequenzen, die f�r die terminalen
Abbildung 1. Nat�rliche und rekombinante Florfliegenseide. A) Florflie-ge Chrysopa carnea. B) Spinnlçsung aus der Dr�se einer weiblichenChrysopa carnea, durch SDS-PAGE getrennt und mit Silberf�rbung vi-sualisiert; M: Proteinmarker; G: Florfliegendr�sen Extrakt. C) Gereinig-tes rekombinantes N[AS]8C-Protein; P: gereinigtes N[AS]8C. D) Artifizi-elle Module.
Abbildung 2. Vergleich von nat�rlichen und artifiziellen Stielen.A) Schematisches Bild der Stielproduktion: 1) Stiel mit Pinzette ausdem Tropfen auf die Aluminiumfolie ziehen; 2) Entspannen des Stielsdurch Verschieben der Aluminiumfolie; 3) der Stiel kontrahiert w�h-rend der Nachbehandlung bei 60 8C und 70 % relativer Luftfeuchtigkeit.B) Bild eines artifiziellen Stiels mit einem St�ck Aluminiumfolie aufder Spitze (links) und ein nat�rlicher Stiel mit Ei an der Spitze(rechts).
[*] F. Bauer, Prof. Dr. T. ScheibelLehrstuhl Biomaterialien, Universit�t BayreuthUniversit�tsstraße 30, 95440 Bayreuth (Deutschland)E-Mail : [email protected]: http://www.fiberlab.de
[**] Wir danken Christopher Volker Synatschke f�r die Hilfe bei denMALDI-TOF-Messungen, Johannes Diehl und Claudia Bl�m f�r dieUnterst�tzung bei den SEM-Aufnahmen sowie Dr. Andrew Smith,Dr. Martin Humenik und Lukas Eisoldt f�r kritisches Lesen desManuskripts.
Hintergrundinformationen zu diesem Beitrag sind im WWW unterhttp://dx.doi.org/10.1002/ange.201200591 zu finden.
AngewandteChemie
6627Angew. Chem. 2012, 124, 6627 –6630 � 2012 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim
Dom�nen kodieren, mit dem Konstrukt ligiert. Das artifizi-elle Protein N[AS]8C enth�lt acht Wiederholungen desModuls [AS], beide nat�rlichen Termini und hat ein Mole-kulargewicht von 53 kD, was dem des nat�rlichen ProteinsMalXB2 entspricht.[11] Zwar zeigte eine SDS-PAGE-Analyseein hçheres Molekulargewicht als f�r N[AS]8C berechnet an,jedoch wurde die korrekte Masse mithilfe von Massenspek-trometrie best�tigt (Abbildung 2 der Hintergrundinforma-tionen).
Wegen der schlechten Lçslichkeit von N[AS]8C in w�ss-rigen Lçsungen wurde das Protein (10% (w/v)) in Hexaflu-oraceton (HFA) aufgenommen. Im Allgemeinen induziertHFA in gelçsten Proteinen a-helikale Struktur;[14] im Fall vonN[AS]8C wurde diese Sekund�rstruktur durch Circulardi-chroismus(CD)-Spektroskopie best�tigt (Abbildung 3 derHintergrundinformationen). Die Stielbildung aus Seidenachahmend, wurde eine Pinzette in einen Tropfen N[AS]8C-Lçsung gedippt und eine Faser aus dem Tropfen gezogen. DasEnde des Stiels wurde auf ein St�ck Aluminiumfolie geheftet,und anschließend wurde der Stiel getrocknet (Abbil-dung 2A). Zur Nachbehandlung wurden die Stiele in einemKlimaschrank �ber Nacht bei 60 8C und 70% relativer Luft-feuchtigkeit inkubiert. Weitere mçgliche Nachbehandlungs-methoden w�ren das �berschichten mit kosmotropen Salz-lçsungen, Ethanol, Methanol, Isopropanol oder das Anlegenhohen Drucks.[15, 16]
Die k�nstlichen Eierstiele haben eine glatte Oberfl�chewie die nat�rlichen Stiele (Abbildung 4 der Hintergrundin-formationen). Allerdings treten bei den nat�rlichen StielenEinschn�rungen auf, die durch Belastung unter hoher Luft-feuchtigkeit (> 30%) oder unter Wasser auftreten. DiesesPh�nomen f�hrt zu einem Verlust der Biegesteifigkeit in deneingeschn�rten Bereichen, begleitet von einer Umformungvon einer Cross-b- zu einer Parallel-b-Struktur.[9] SolcheEinschn�rungen wurden nicht bei artifiziellen Stielen beob-achtet.
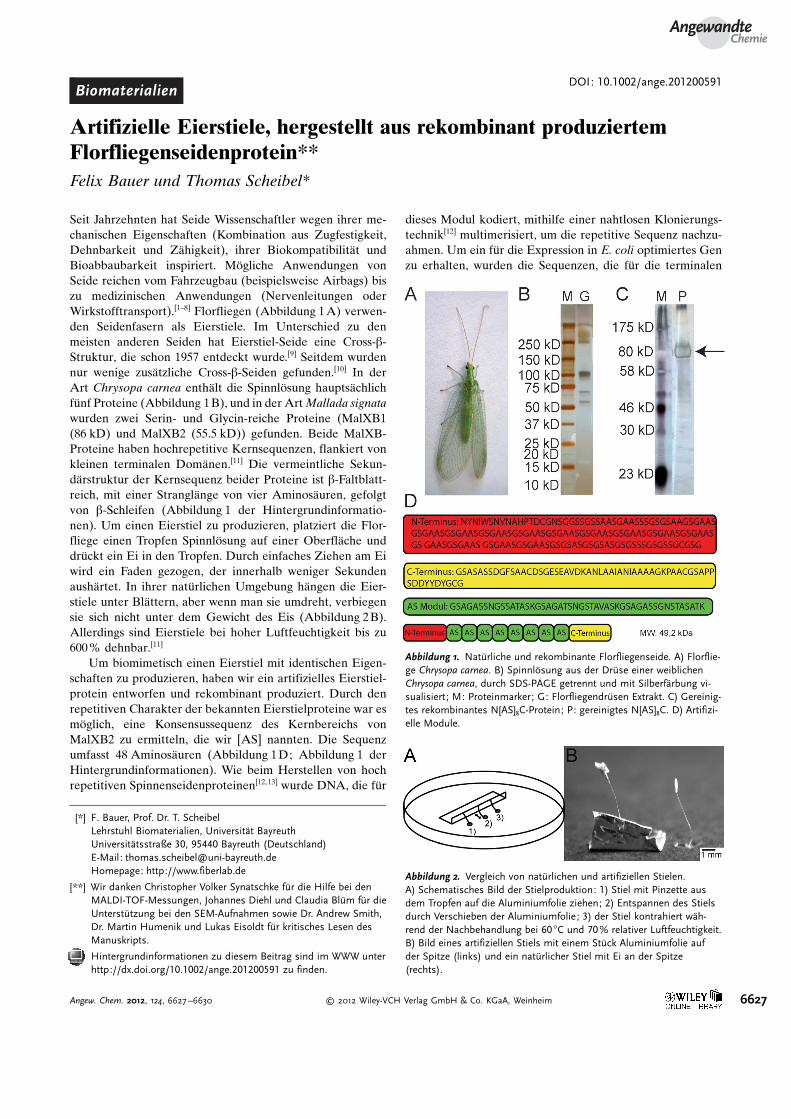
Die Sekund�rstruktur der artifiziellen und nat�rlichenStiele wurde mit FT-IR-Spektroskopie analysiert (Abbil-dung 3). Fourier-Selbstdekonvoluierung der Amid-I-Region[17] ergab einen niedrigeren b-Faltblatt-Anteil in denartifiziellen als in den nat�rlichen Stielen (Tabelle 1). FT-IR-
spektroskopische Messungen vor und nach Zugmessungenzeigten keine �nderung bei nat�rlichen Stielen und einenleichten Anstieg des b-Faltblatt-Gehalts bei reduziertem b-Schleifen-Gehalt bei artifiziellen Stielen. Der a-helikaleAnteil ist in beiden Stielen mit 6–9 % relativ gering. Auffal-
lenderweise war der helikale Anteil vor und nach Zugmes-sungen gleich. Die mithilfe polarisierter FT-IR-Spektroskopiegemessene Poly(l-alanin)-NCa-Streckschwingung[18] von na-t�rlichen Eierstielen bei einer Wellenzahl von 1167 cm�1 imWinkel zur Stielachse zeigt eine starke Orientierung der Po-ly(Ala)-b-Faltbl�tter im Winkel von 0 und 908 zur Stielachse(molekularer Ordnungsparameter[25] Smol : �0.44; Abbil-dung 5 der Hintergrundinformationen). Das k�nstlich her-gestellte Protein hat keine Poly(alanin)-Sequenz, weshalbdessen Orientierung nicht detektierbar war. Dies zeigt, dassbisher unbekannte, Poly(alanin)-reiche Proteine, mçglicher-weise eines der drei zus�tzlich in C. carnea vorkommenden(Abbildung 1B), stark am Proteinausrichtungsprozess in na-t�rlichen Stielen beteiligt sein kçnnten. Deshalb ist unserderzeitiger Ansatz nicht geeignet, die Eigenschaften von na-t�rlichen Eierstielen vollst�ndig nachzuahmen.
Sowohl der nat�rliche als auch der k�nstliche Stiel zeigenDoppelbrechung (Abbildung 4A,B), die auf strukturellerAnisotropie basiert. Die Doppelbrechung bei den nat�rlichenStielen ist stark, da hoch geordnete kristalline Bereiche mit b-Faltblatt-Struktur, angeordnet im rechten Winkel zur Faser-achse, vorhanden sind. K�nstliche Stiele zeigen schw�chereDoppelbrechung, was auf weniger stark geordnete Strukturenhinweist.
Abbildung 3. FT-IR-Absorptionsspektrum von nat�rlichen und artifiziel-len Stielen vor den Zugmessungen; Einsch�be: Fourier-Selbstdekonvo-luierung der Amid-I-Region von nat�rlichen und artifiziellen Stielen.
Tabelle 1: Durchschnittliche Sekund�rstruktur von nat�rlichen undk�nstlichen Eierstielen vor (vZM) und nach Zugmessungen (nZM), mitFT-IR-Spektroskopie gemessen und durch Fourier-Selbstdekonvoluie-rung der Amid-I-Bande quantifiziert.[17]
Sekund�rstruktur nat�rlicher Eierstiel k�nstlicher EierstielvZM [%] nZM [%] vZM [%] nZM [%]
b-Faltbl�tter 40�6 39�5 32�3 39�4a-Helices 6�4 7�1 9�2 7�1b-Schleifen 24�3 22�1 30�3 24�1Random Coils 30�4 32�2 29�1 30�2
Abbildung 4. Vergleich der Doppelbrechung von nat�rlichen undk�nstlichen Stielen. A) Mikroskopisches Bild eines nat�rlichen Florflie-gen-Eierstiels. B) Mikroskopisches Bild eines k�nstlichen Stiels. Oben:Hellfeld; unten: mit gekreuzten Polarisatoren; Maßstab: 50 mm.
.AngewandteZuschriften
6628 www.angewandte.de � 2012 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim Angew. Chem. 2012, 124, 6627 –6630
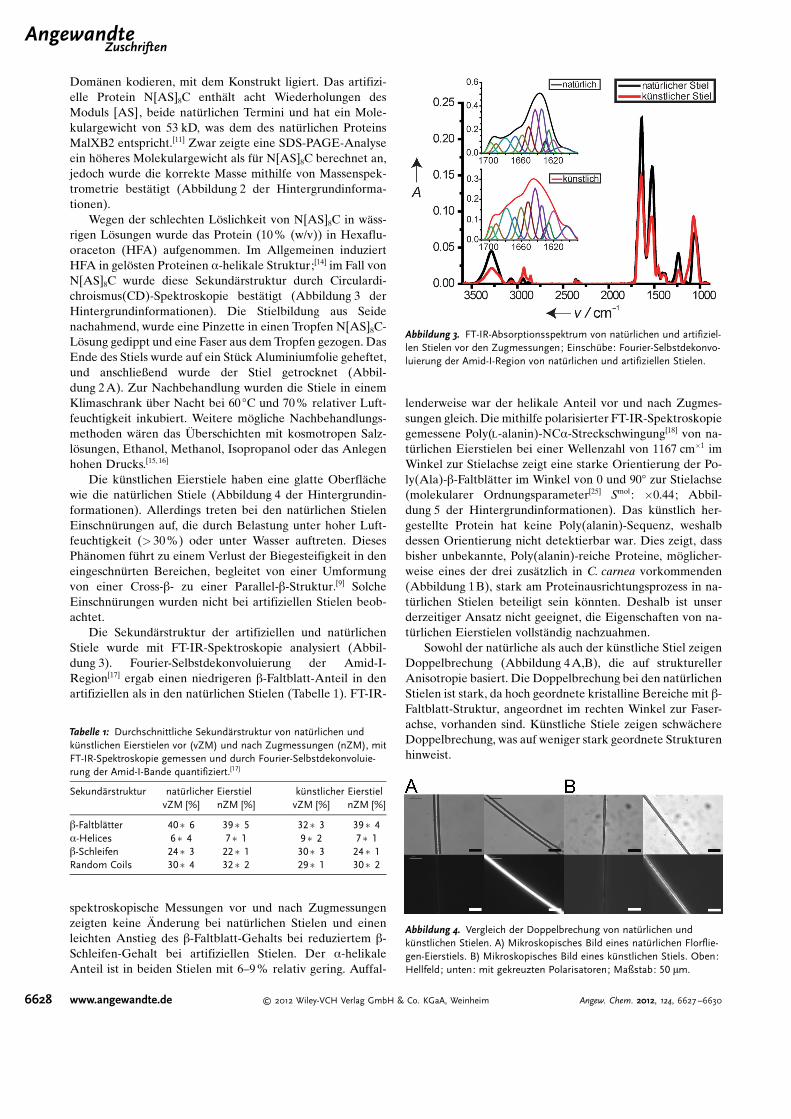
Die k�nstlichen Stiele habeneinen �hnlichen Durchmesser(10 mm) wie die nat�rlichen undzeigen auch Biegesteifigkeit (Ab-bildung 2 B). Die mechanischenEigenschaften wurden bei einer re-lativen Luftfeuchtigkeit (RH) von30 und 70% sowie bei einer Tem-peratur von 22 8C gemessen. DasKontrollieren der Luftfeuchtigkeitist sehr wichtig f�r Zugmessungenvon Seiden.[19–21] Die Zugfestigkeitvon nat�rlichen Eierstielen(C. carnea) ist unter den gew�hltenBedingungen mit 68 MPa (30%RH) und 155 MPa (70% RH) re-lativ gering gegen�ber den publizierten Werten von 186–375 MPa, gemessen bei 65% RH (Mallada signata undChrysopa spec. ;[11, 22] Tabelle 2). Bei 30 % RH reißen die Stielebei einer Dehnung von 2%, w�hrend sie bei 70 % RH beieiner Dehnung von 211 % reißen (Abbildung 5 und Tabel-le 2). Der Elastizit�tsmodul von nat�rlichen Eierstielen bei30% RH betr�gt 5.7 GPa, bei k�nstlichen Stielen hingegennur ca. 40 % dieses Wertes (2.3 GPa). Die Zugfestigkeit vonnat�rlichen Stielen betr�gt 68 MPa und die der k�nstlichenStiele 55 MPa (Tabelle 2). Wenn man die Porosit�t derk�nstlichen Stiele mit einbezieht, kann sogar eine Zugfes-tigkeit von ca. 63 MPa errechnet werden (30 % RH). Inter-essanterweise liegt die Z�higkeit von k�nstlichen Stielen bei30% RH bei 1.76 MJm�3 und damit leicht hçher als die dernat�rlichen Stiele mit 1.2 MJm�3.
Das artifizielle und rekombinant hergestellte Eierstiel-protein der Florfliege konnte zu k�nstlichen Stielen verar-beitet werden, die bei 30 % RH ann�hernd gleiche Eigen-schaften aufweisen wie die nat�rlichen Eierstiele. Die Stielesind wie die nat�rlichen Stiele biegesteif, was sie f�r An-wendungen, bei denen Steifigkeit in transversaler Richtungvon Bedeutung ist, interessant macht. Des Weiteren ermçg-licht es der rekombinante Ansatz, das molekulare Struktur-Funktions-Verh�ltnis in Eierstielen besser zu verstehen. Alsn�chstes ist es wichtig, die Poly(alanin)-reichen Proteine zuidentifizieren und w�ssrige Spinnlçsungen zu erzielen, wasnçtig ist, um eine Vorstrukturierung zu erhalten. Eine vor-strukturierte Spinnlçsung ist der Schl�ssel f�r F�den mitCross-b-Struktur.
Experimentelles und MethodenAlle Genmodule (N, [AS] und C) wurden mit einer 5’-BamHI- undeiner 3’-HindIII-Restriktionsschnittstelle konstruiert und bei Gene-art bestellt. Die Module wurden mithilfe der Enzyme BsaI, BsgI undBseRI geschnitten und anschließend ligiert.[12] Das vollst�ndigeN[AS]8C-Konstrukt wurde mithilfe von BamHI und HindIII in einenpET28-Vektor kloniert.
BL21(DE3)-Zellen wurden mit pET28-N[AS]8C transformiertund in einem Fermenter (Infors Minifors 2.5 L) nach einer zuvorbeschriebenen Vorschrift kultiviert.[23] Bei einem OD600-Wert von 72wurden die Bakterien drei Stunden mit 1 mm Isopropyl-b-d-thio-galactopyranosid (IPTG) induziert. Im Anschluss wurden die Zellengeerntet und dreimal gewaschen.
Die Reinigung erfolgte nach einer Vorschrift, die f�r rekombi-nante Seidenproteine beschrieben worden war.[24] Nach Zelllyse,Ans�uern und Zentrifugieren wurde der �berstand fraktionierendmit Ammoniumsulfat gef�llt (1.5 und 2.5m). Das gef�llte Proteinwurde mit Wasser gewaschen und lyophilisiert.
Zur Analyse des produzierten Proteins wurden 3 mgmL�1
N[AS]8C in 6m Guanidiniumthiocyanat gelçst und auf eine End-konzentration von 2m Guanidiniumthiocyanat verd�nnt. Vor demAuftragen der Proben wurden diese mit ZipTip-C4-Pipettenspitzenentsalzt und mit Matrixlçsung eluiert. MALDI-TOF-Massenspek-trometrie erfolgte mit einem Bruker Reflex III, ausger�stet mit
Tabelle 2: Zugmessungen von nat�rlichen (C. carnea) und k�nstlichen Eierstielen. Die Versuche wurdenbei 30% RH und 70% RH und bei einer Temperatur von 22 8C durchgef�hrt.
Dehnbarkeit [%] Zugfestigkeit smax [MPa] Elastizit�tsmodul [MPa] Z�higkeit [MJm�3]
Literaturwerte[a]
65% RH[11] 381 31065% RH[22] 249 ca. 375
nat�rliche Eierstiele30% RH 2�1 68�19 5777�1257 1.2�0.7270% RH 210�100 155�75 3175�1016 87�49
k�nstliche Eierstiele30% RH 5�2 55�14 2330�850 1.76�0.970% RH 6�3 25�11 1012�252 1.09�0.59
[a] F�r M. signata und C spec.
Abbildung 5. A) Spannungs-Dehnungs-Diagramm (Maximalwerte) vonnat�rlichen und k�nstlichen Eierstielen bei 30% RH. Die Unterschiedeim Rauschen sind durch die kleineren Durchmesser der k�nstlichenStiele zu erkl�ren. B) SEM-Bilder von Querschnittsfl�chen von nat�rli-chen (links) und k�nstlichen Stielen (rechts).
AngewandteChemie
6629Angew. Chem. 2012, 124, 6627 –6630 � 2012 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.angewandte.de

einem 337-nm-N2-Laser, im Linearmodus und mit 20 kV Beschleu-nigungsspannung. Als Matrix wurde Sinapins�ure (20 mgmL�1) in50% CH3CN und 0.1% Trifluoressigs�ure in H2O verwendet.
Die Stiele wurden aus seiner 10-proz. (w/v) Lçsung von N[AS]8Cin HFA gezogen und bei Raumtemperatur getrocknet. Anschließendwurde die Spannung aus den Stielen durch Ann�hern der beidenAuflagepunkte entfernt, und die Proben wurden bei 60 8C und 70%RH �ber Nacht in einem Klimaschrank nachbehandelt.
F�r die Rasterelektronenmikroskopie (SEM) wurden die Probendurch Sputtern mit Platin beschichtet (Dicke 2 nm). Die SEM-Bilderwurden mit einem Zeiss-1530-Ger�t unter der Verwendung desinlens-Detektors aufgenommen.
Fourier-transformierte Infrarotspektroskopie wurde an einemBruker tensor 27/pike MIRacle unter Verwendung der Hyperion-Einheit durchgef�hrt. Die Datenprozessierung und Fourier-Selbst-dekonvoluierung der Amid-I-Region erfolgten mit der Opus-6.5-Software.[17] Polarisierte FT-IR-spektroskopische Messungen undProzessierung der Daten wurden gem�ß Hagenau et al. durchge-f�hrt.[25]
Mikroskopische Aufnahmen erfolgten an einem Mikroskop(Leica DMI 3000B), das mit Polarisatoren ausgestattet war.
Proben f�r die Zugmessungen wurden mit Plastikklebstoff aufPlastikrahmen mit einem Spalt von 2 mm geklebt. Spannungs-Deh-nungs-Kurven wurden mit einer Zugmaschine (Bose Electroforce3220), ausgestattet mit einer 0.5-N-Kraftmessdose und einer Klima-kammer zur Einstellung der relativen Luftfeuchtigkeit, aufgenom-men. Die Stiele wurden mit einer Geschwindigkeit von 0.01 mms�1
bis zum Reißen gedehnt.CD-Spektren wurden von 250 bis 195 nm an einem Jasco-J-815-
CD-Spektrometer mit einem Intervall von 0.1 nm, einer Bandbreitevon 1 nm, einer Scangeschwindigkeit von 50 nmmin�1, einer DigitalIntegration Time (D.I.T.) von 1 s und drei Akkumulierungen aufge-nommen.
Eingegangen am 20. Januar 2012,ver�nderte Fassung am 10. April 2012Online verçffentlicht am 16. Mai 2012
.Stichwçrter: b-Faltbl�tter · Biomimetische Synthesen ·Faserproteine · Seide
[1] J. G. Hardy, T. R. Scheibel, Prog. Polym. Sci. 2010, 35, 1093 –1115.
[2] A. Leal-Egana, T. Scheibel, Biotechnol. Appl. Biochem. 2010, 55,155 – 167.
[3] A. Lammel, M. Schwab, M. Hofer, G. Winter, T. Scheibel, Bio-materials 2011, 32, 2233 – 2240.
[4] C. Allmeling, A. Jokuszies, K. Reimers, S. Kall, C. Y. Choi, G.Brandes, C. Kasper, T. Scheper, M. Guggenheim, P. M. Vogt,Cell Proliferation 2008, 41, 408 – 420.
[5] C. Allmeling, A. Jokuszies, K. Reimers, S. Kall, P. M. Vogt, J.Cell. Mol. Med. 2006, 10, 770 – 777.
[6] K. Spiess, A. Lammel, T. Scheibel, Macromol. Biosci. 2010, 10,998 – 1007.
[7] S. Weisman, V. S. Haritos, J. S. Church, M. G. Huson, S. T.Mudie, A. J. Rodgers, G. J. Dumsday, T. D. Sutherland, Bioma-terials 2010, 31, 2695 – 2700.
[8] J. G. Hardy, L. M. Romer, T. R. Scheibel, Polymer 2008, 49,4309 – 4327.
[9] K. D. Parker, K. M. Rudall, Nature 1957, 179, 905 – 906.[10] C. L. Craig, Annu. Rev. Entomol. 1997, 42, 231 – 267.[11] S. Weisman, S. Okada, S. T. Mudie, M. G. Huson, H. E. Trueman,
A. Sriskantha, V. S. Haritos, T. D. Sutherland, J. Struct. Biol.2009, 168, 467 – 475.
[12] D. Huemmerich, C. W. Helsen, S. Quedzuweit, J. Oschmann, R.Rudolph, T. Scheibel, Biochemistry 2004, 43, 13604 – 13612.
[13] C. Vendrely, T. Scheibel, Macromol. Biosci. 2007, 7, 401 – 409.[14] J. M. Yao, H. Masuda, C. H. Zhao, T. Asakura, Macromolecules
2002, 35, 6 – 9.[15] H. Peng, S. Zhou, J. Jiang, T. Guo, X. Zheng, X. Yu, J. Phys.
Chem. B 2009, 113, 4636 – 4641.[16] X. Zhang, M. R. Reagan, D. L. Kaplan, Adv. Drug Delivery Rev.
2009, 61, 988 – 1006.[17] X. Hu, D. Kaplan, P. Cebe, Macromolecules 2006, 39, 6161 –
6170.[18] W. H. Moore, S. Krimm, Biopolymers 1976, 15, 2465 – 2483.[19] A. Sch�fer, T. Vehoff, A. Glisovic, T. Salditt, Eur. Biophys. J.
Biophys. Lett. 2008, 37, 197 – 204.[20] T. Vehoff, A. Glisovic, H. Schollmeyer, A. Zippelius, T. Salditt,
Biophys. J. 2007, 93, 4425 – 4432.[21] G. R. Plaza, G. V. Guinea, J. Perez-Rigueiro, M. Elices, J. Polym.
Sci. Part B 2006, 44, 994 – 999.[22] H. R. Hepburn, H. D. Chandler, M. R. Davidoff, Insect Bio-
chem. 1979, 9, 69 – 77.[23] D. J. Korz, U. Rinas, K. Hellmuth, E. A. Sanders, W. D. Deck-
wer, J. Biotechnol. 1995, 39, 59 – 65.[24] X.-X. Xia, Z.-G. Qian, C. S. Ki, Y. H. Park, D. L. Kaplan, S. Y.
Lee, Proc. Natl. Acad. Sci. USA 2010, 107, 14059 – 14063.[25] A. Hagenau, P. Papadopoulos, F. Kremer, T. Scheibel, J. Struct.
Biol. 2011, 175, 339 – 347.
.AngewandteZuschriften
6630 www.angewandte.de � 2012 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim Angew. Chem. 2012, 124, 6627 –6630