Embed Size (px)
Citation preview

Aus dem Institut für Bakteriologie und Mykologie der Veterinärmedizinischen Fakultät der Universität Leipzig
und dem Kreiskrankenhaus Altenburg
Untersuchungen zum Einfluss der Ernährung auf ausgewählte bakteriologische, enzymatische
und blutchemische Parameter beim Menschen
unter Gewichtsreduktion
Inaugural-Dissertation zur Erlangung des Grades eines
Doctor medicinae veterinariae (Dr. med. vet.) durch die Veterinärmedizinische Fakultät
der Universität Leipzig
eingereicht von Thomas Vorpagel aus Wernigerode
Leipzig, 2006

Mit Genehmigung der Veterinärmedizinischen Fakultät der Universität Leipzig Dekan: Prof. Dr. Karsten Fehlhaber Betreuer: Prof. Dr. Monika Krüger Prof. Dr. Gerhard Scholz Gutachter: Prof. Dr. Monika Krüger, Institut für Bakteriologie und Mykologie, Veterinärmedizinische Fakultät der Universität Leipzig Prof. Dr. Gerhard Scholz, Kreiskrankenhaus Altenburg Prof. Dr. Manfred Fürll, Medizinische Tierklinik, Veterinärmedizinische
Fakultät der Universität Leipzig Prof. Dr. Florian Schweigert, Institut für Ernährungswissenschaft der
Universität Potsdam
Tag der Verteidigung: 27. Februar 2006

Inhaltsverzeichnis
Inhaltsverzeichnis I
Abkürzungsverzeichnis V
Abbildungsverzeichnis VII
Tabellenverzeichnis VIII
1 Einleitung und Zielsetzung 1 2 Literaturübersicht 4 2.1 Aufbau der Magen-Darm-Flora 4
2.1.1 Begriffsklärung 4
2.1.2 Normalstatus Erwachsener 5
2.1.3 Die Entwicklung vom Neugeborenen zum Mischköstler 10
2.2 Die wandständige Flora und der Vergleich mit der Lumenflora 12
2.3 Stabilität, Kontrolle und Beeinflussbarkeit der Darmflora 14
2.3.1 Einblick in das Ökosystem des Gastrointestinaltraktes mit
Wechselwirkungen zwischen den Bakterien 14
2.3.2 Stabilität des Ökosystems 15
2.3.3 Einfluss der Ernährung des Wirtes auf die Darmflora 17
2.3.4 Einfluss von Prä- und Probiotika auf die Mikroflora 19
2.3.5 Einfluss des Alters des Wirtes 21
2.3.6 Zusammenhang von Erkrankungen des Darmtraktes und der
Mikroflora 23
2.3.7 Einfluss der Antibiose auf die Mikroflora 23
2.4 Interaktionen zwischen Darmflora und Makroorganismus 26
2.4.1 Metabolismus der Mikroorganismen und des Wirtes sowie gegen-
seitige Beeinflussung 26
2.4.2 Immunologie des Darmes 27
2.4.3 Kurzkettige Fettsäuren (SCFA) 31
2.4.3.1 Definition SCFA 31
2.5 Untersuchungsmöglichkeiten 33
2.5.1 Studienmodelle 33
2.5.1.1 In situ – Untersuchungen 33
2.5.1.2 Tierexperimentelle Untersuchungen 33
2.5.2 Nachweismethoden 35
2.6 Weitere Parameter 36
I

Inhaltsverzeichnis
2.6.1 Alkalische Phosphatase in Fäzes 36
2.6.1.1 Normbereiche der AP-Aktivität 37
2.6.1.2 Einflüsse auf die AP-Aktivität 38
2.6.2 MRS-Flora 38
2.6.3 Leptin 39
2.6.3.1 Wirkung auf das Körpergewicht 40
2.6.4 Body Mass Index (BMI) 40
2.6.5 Blutzucker und HbA1c 40
2.6.6 Gesamtcholesterol, LDL, HDL und Triglyceride 41
3 Material und Methoden 42
3.1 MIRA-Konzept: Mit Intelligenz richtig abnehmen 42
3.1.1 MIRA-PreCon und MIRA-DGE 42
3.1.2 Teilnehmer 42
3.2 Stuhlprobengewinnung und –verarbeitung 43
3.2.1 Kulturelle Keimzählung 43
3.2.1.1 Untersuchte Bakterienpopulationen und verwendete Nährböden 44
3.3 Keimzählung mit Hilfe der Fluoreszenz in situ-Hybridisierung (FISH) 44
3.3.1 Sonden für die Fluoreszenz in situ-Hybridisierung 45
3.3.2 Fixierung der Stuhlproben 45
3.3.3 FISH auf Objektträgern 46
3.3.4 Ermittlung der Zellzahlen mittels Fluoreszensmikroskopie 47
3.4 Alkalische Phosphatase 48
3.5 Fettsäurenbestimmung 49
3.6 Weiter Parameter 49
3.7 Biostatistische Auswertung 49
4 Ergebnisse 51 4.1 Dynamik der Keimzahlentwicklung von Fäkalkeimen,MIRA-
PreCon-Gruppe vs. MIRA-DGE-Gruppe im Untersuchungszeitraum 51
4.1.1 Ergebnisse der Stuhluntersuchungen, weibliche und männliche
Teilnehmer 51
4.1.1.1 Einfluss der Beprobungsmodalitäten 51
4.1.1.2 Im Plattenkulturverfahren ermittelte Keimzahlen 52
II

Inhaltsverzeichnis
4.1.2 Ergebnisse der Stuhluntersuchungen, nur weibliche Teilnehmer 56
4.1.2.1 Im Plattenkulturverfahren ermittelte Keimzahlen 56
4.1.3 Ergebnisse der Differenzierung der MRS-Gesamtkeimzahl 57
4.1.3.1 Keimzählung auf Spezialnährboden 57
4.1.3.2 Keimzahlermittlung mittels Fluoreszenz in situ-Hybridisierung (FISH) 58
4.2 Enzymatische Parameter 60
4.2.1 Ergebnisse der Untersuchungen auf Alkalische-Phosphatase-
Aktivität im Stuhl 60
4.2.1.1 MIRA-PreCon-Gruppe vs. MIRA-DGE-Gruppe, weibliche und
männliche Teilnehmer 60
4.2.1.2 MIRA-PreCon-Gruppe vs. MIRA-DGE-Gruppe, nur weibliche
Teilnehmer 60
4.3 Ergebnisse der Untersuchungen auf kurzkettige Fettsäuren (SCFA) 61
4.4 Body Mass Index und die Ergebnisse der blutchemischen
Untersuchungen 63
4.5 Untersuchungen zum Einfluss des BMI auf Stuhl- und Blutparameter 68
4.5.1 Im Plattenkulturverfahren ermittelte Keimzahlen, MIRA-I vs. MIRA-II 68
4.5.1.1 Kulturelle Differenzierung der MRS-Gesamtkeimzahl 71
4.5.2 Differenzierung der MRS-Gesamtkeimzahl mit der Fluoreszenz in
situ-Hybridisierung 72
4.5.3 Ergebnisse der Untersuchungen auf Alkalische-Phosphatase-
Aktivität, MIRA-I vs. MIRA-II 73
4.5.4 Ergebnisse der Untersuchungen auf kurzkettige Fettsäuren 73
4.5.5 Body Mass Index und Ergebnisse der blutchemischen
Untersuchungen 74
5 Diskussion 79
5.1 Methodenkritik 79
5.2 Keimzahlentwicklungen von Fäkalbakterien im Untersuchungs-
zeitraum 80
5.2.1 Aerobe Gesamtkeimzahl und aerobe Gram-negative Gesamt-
keimzahl im Plattenkulturverfahren 80
5.2.2 Candida spp. und Clostridium perfringens im Plattenkulturverfahren 84
5.2.3 MRS-Gesamtflora (Bifidobakterien, Laktobazillen, Enterokokken) 85
III

Inhaltsverzeichnis
5.2.3.1 Kulturelle Differenzierung der MRS-Gesamtflora 86
5.2.3.2 Differenzierung der MRS-Gesamtflora durch Fluoreszenz in situ-
Hybridisierung 87
5.3 Alkalische Phosphatase 89
5.4 Kurzkettige Fettsäuren 90
5.5. Body Mass Index 92
5.6 Blutchemische Parameter 94
5.6.1 Leptin 94
5.6.3 Blutzucker und HbA1c 94
5.6.4 Gesamtcholesterol, LDL, HDL und Triglyceride 95
5.7 Zusammenfassende Diskussion 96
6 Zusammenfassung 98
7 Summary 100
8 Literaturverzeichnis 102 Anhang auf CD-ROM, enthält die Ergebnisse der statistischen Tests,
Korrelationen und die Mittelwertsdifferenzen
Danksagung
IV

Abkürzungsverzeichnis
verwendete Abkürzungen
AP Alkalische Phosphatase
APC antigen presenting cell
BMI Body Mass Index (Körpergewicht in kg / Körpergröße in m2)
BZ Blutzucker
C2 Essigsäure, Acetat
C3 Propionsäure, Propionat
C4 Buttersäure, Butyrat
C5 Valeriansäure
C6 Capronsäure
C7 Heptansäure
DC Dentritische Zelle (dendritic cell)
DGE Deutsche Gesellschaft für Ernährung
DNA Desoxyribonukleinsäure
DSMZ Deutsche Sammlung von Mikroorganismen und Zellkulturen
FISH Fluoreszenz in situ-Hybridisierung
FOS Fructooligosaccharide
g centrifugal force
GALT gut associated lymphatic tissue
G-chol Gesamtcholesterol
GIT Gastrointestinaltrakt
HbA1c Hämoglobin A1c
HDL High Density Lipoprotein
HPA-Achse Hypothalamus-Hypophysen-Nebennieren-Achse
I-AP intestinale AP, auch alkalische Dünndarmphosphatase
IBD entzündliche Darmerkrankungen (inflammatory bowel disease)
iC4 iso-Buttersäure
iC5 iso-Valeriansäure
IEL intraepiteliale Lymphozyten
IL-1 Interleukin-1
kDa kilo-Dalton
L-AP Leber–AP
LDL Low Density Lipoprotein
V

Abkürzungsverzeichnis
LPL Lymphozyten der Lamina propria
M Molar (mol/Liter)
MALT mucosa associated lymphatic tissue
mM Millimol
ob-Gen obesity-Gen
OD Optische Dichte
OGTT oraler Glukosetoleranz-Test
RNA Ribonukleinsäure
s Standardabweichung
SCFA Short Chain Fatty Acid(s), flüchtige Fettsäure(n)
SIgA sekretorische Immunglobulin A-Antikörper
SIgM sekretorische Immunglobulin M-Antikörper
TG Triglyceride
TNF Tumornekrosefaktor
U/min Umdrehungen pro Minute
UC Colitis ulcerosa x Mittelwert
VI

Abbildungsverzeichnis
verwendete Abbildungen Abbildung Titel Seite
Abb. 1 Verlauf aerobe Gesamtkeimzahl, PreCon vs. DGE,
Kulturverfahren 53
Abb. 2 Verlauf aerobe Gram-negative Gesamtkeimzahl, PreCon
vs. DGE, Kulturverfahren 54
Abb. 3 Verlauf Keimzahl Clostridium perfringens, PreCon
vs. DGE, Kulturverfahren 54
Abb. 4 Verlauf Keimzahl Candida spp., PreCon vs. DGE,
Kulturverfahren 55
Abb. 5 Verlauf Gesamtkeimzahl Laktobazillen, Bifidobakterien
und Enterokokken, PreCon vs. DGE, Kulturverfahren 55
Abb. 6 Aktivitätsentwicklung Alkalische Phosphatase,
PreCon vs. DGE 60
Abb. 7 Keimzahlverlauf der mittels Fluoreszenz in situ-Hybridisierung
ermittelten Gesamtkeimzahl (Eub), Bifidobakterien (Bif),
Laktobazillen (Lab) und Enterokokken (Enterokok) der
MIRA-DGE (N=8) und MIRA-PreCon (N=15) 58
Abb. 8 Verlauf aerobe Gesamtkeimzahl, MIRA-I vs. MIRA-II,
Kulturverfahren 68
Abb. 9 Verlauf aerobe Gram-negative Gesamtkeimzahl, MIRA-I
vs. MIRA-II, Kulturverfahren 69
Abb. 10 Verlauf Keimzahl Clostridium perfringens, MIRA-I vs.
MIRA-II, Kulturverfahren 70
Abb. 11 Verlauf Keimzahl Candida spp., MIRA-I vs. MIRA-II,
Kulturverfahren 70
Abb. 12 Verlauf Gesamtkeimzahl Laktobazillen, Bifidobakterien
und Enterokokken, MIRA-I vs. MIRA-II, Kulturverfahren 71
Abb. 13 Aktivitätsentwicklung Alkalische Phosphatase, MIRA-I
vs. MIRA-II 73
Abb. 14 Verlauf Body Mass Index, MIRA-I vs. MIRA-II 74
VII

Tabellenverzeichnis
verwendete Tabellen
Tabelle Titel Seite
1 Besiedlung der Darmabschnitte, modifiziert nach
Berg, R.D. (1996) 6
2 Gesamtkeimzahlen der Anaerobier und fakultativen Anae-
robier; Zellzahlen Bifidobakterien und Bacteroides in Zäkum-
und Stuhlproben bei acht gesunden Personen; modifiziert
nach Marteau et al., 2001 7
3 Fäkale Mikroflora bei verschiedenen Ernährungsweisen,
modifiziert nach Finegold et al., 1983 9
4 Zusammenfassung der Keimzählungen bei 24 Biopsie-
proben von 12 Patienten, modifiziert nach Poxton et al, 1997 13
5 Keimzahlen von älteren Menschen mit Verstopfung 22
6 bakterielle Bildung und Metabolismus von SCFA, modifiziert
nach Gibson (1999) 32
7 Liste der für FISH verwendeten Oligonucleotidsonden 45
8 Untergruppen; Größe und durchschnittliche Zeit in Tagen
zwischen den Probenentnahmen 51
9 Keimzahlschwankungen am Beispiel der aeroben Gram-
negativen Gesamtkeimzahl in log10KbE/g zur Feststellung
des Einflusses des Abgabezeitpunktes 52
10 Keimzahlen auf Spezialnährboden ermittelt, in log10KbE/g 58
11 Keimzahlen getrennt nach MIRA-Gesamtgruppen auf
Spezialnährboden ermittelt, in log10KbE/g 58
12 Absolute Keimzahlen in log10KbE/g und prozentuale Anteile
der mittels FISH-Technik untersuchten Gattungen 59
13 Gehalt an volatilen Fettsäuren (SCFA) in µmol/g Stuhl 61
14 Dynamik der volatilen Fettsäuren (SCFA) in Prozent,
Woche 0 entspricht 100 Prozent 62
15 prozentuale Anteile der volatilen Fettsäuren (SCFA) 62
16 BMI und blutchemische Parameter 63
VIII

Tabellenverzeichnis
17 signifikante Unterschiede innerhalb und zwischen den Gruppen
beim BMI 64
18 signifikante Unterschiede innerhalb und zwischen den Gruppen
beim Leptin aus venösem Blut 64
19 signifikante Unterschiede innerhalb und zwischen den Gruppen
beim HbA1c 65
20 signifikante Unterschiede innerhalb und zwischen den Gruppen
beim Blutzucker 65
21 signifikante Unterschiede innerhalb und zwischen den Gruppen
beim Gesamt-Cholesterol 66
22 signifikante Unterschiede innerhalb und zwischen den Gruppen
beim HDL 66
23 signifikante Unterschiede innerhalb und zwischen den Gruppen
beim LDL 67
24 signifikante Unterschiede innerhalb und zwischen den Gruppen
bei den Triglyceriden 67
25 Keimzahlen auf Spezialnährboden ermittelt; in log10KbE/g,
darunter die Anteile in Prozent 72
26 mittels FISH erhobene Keimzahlen der MIRA-Untergruppen in
log10KbE/g, MIRA-I, N=12; MIRA-II, N=9 72
27 Kurzkettige Fettsäuren (SCFA) der MIRA-Untergruppen in mol% 74
28 Leptin in mmol/l 75
29 Blutparameter der MIRA-Untergruppen; HbA1c in %, Blutzucker
und –fette in mmol/l 75
IX


Einleitung und Zielsetzung
1. Einleitung und Zielsetzung
Das Interesse an der Magen-Darm-Flora, ihrer Zusammensetzung und ihren Stoff-
wechseleigenschaften ist in den vergangenen Jahrzehnten ständig gestiegen. Der
Zusammenhang von Darmbakterien, Umweltkeimen und Erkrankungen bei Tieren
und Menschen bestätigte die Wichtigkeit umfassender Untersuchungen, die zu
effektiven präventiven und kurativen Maßnahmen führen sollten.
Der menschliche Darm ist mit einer Oberfläche von 200-300m2 das größte Hohl-
organ im menschlichen Körper und seine größte Kontaktfläche zur äußeren Umwelt.
Durch Eintrag von außen gelangen in den Darm neben aufgenommenen Substraten
auch Bakterien. Auf diese reagiert der Wirt mit Abwehr oder Toleranz, so dass sich
im Säuglings- und Kleinkindalter eine individuelle und stabile Darmflora etabliert. Sie
schützt den Wirt über vielfältige Mechanismen vor der Besiedlung mit pathogenen
Keimen und den von ihnen ausgelösten Erkrankungen.
Die physiologische Darmflora verstoffwechselt zahlreiche mit der Nahrung aufgenom-
mene Substrate und stellt dem Wirt zusätzliche energiereiche Verbindungen zur
Verfügung. Sie verwertet durch mikrobielle Fermentation auch Nahrungsbestand-
teile, die durch die wirtseigenen Enzyme nicht spaltbar und daher unverdaulich sind.
Es werden u.a. flüchtige Fettsäuren gebildet, die dem Wirtsorganismus als Energie-
quellen dienen, sowie Vitamine wie Vitamin B12 und Vitamin K synthetisiert. Durch
die Abhängigkeit der Darmflora von den zugeführten Substraten bewirken Verände-
rungen der Ernährung Modulationen verschiedener Ausmaße. Regulierende Eingriffe
wurden mit Hilfe von prä- und probiotischen Zusätzen erreicht. Diese kommen
beispielsweise bei der Behandlung von Obstipationen und antibiotisch bedingten
Diarrhöen sowie vorbeugend gegen entzündliche und neoplastische Erkrankungen
vor allem des Dickdarmes zum Einsatz (GIBSON et al. 1995 und 1999, Tannock et
al. 2000).
Wie bereits angesprochen, schützt eine intakte physiologische Darmflora den Wirts-
organismus vor Erkrankungen. Zum einen stärkt sie die darmeigene Immunität durch
ständige Stimulation des GALT, in dem je nach Aktivität 30-80% der menschlichen
Immunzellen lokalisiert sind. Untersuchungen haben gezeigt, dass die lymphatischen
Organe des Darmes bei keimfreien Mäusen deutlich weniger aktiviert waren als bei
normal besiedelten Tieren (BERG 1996). Zum anderen verhindert die etablierte
Darmflora eine Fehlbesiedlung durch Kolonisation pathogener Keime und schützt
1

Einleitung und Zielsetzung
durch die dichte Besiedlung der Darmmukosa vor Translokationen der Erreger und
nachfolgenden Erkrankungen.
Die Diversität und Stabilität wird auch durch Wechselwirkungen zwischen Kompo-
nenten der Darmflora gewährleistet. Die fortwährende Konkurrenz um Nahrung und
Kolonisationsplätze führte zur Nutzung jeder Überlebensmöglichkeit innerhalb der
verschiedenen Habitate und Nischen. Dazu gehören z.B. die Nutzung von bakteri-
ellen Stoffwechselprodukten durch andere Spezies oder die Entwicklung transienter
Spezies. Durch den begrenzten Kolonisationsraum an der Darmmukosa wurden
Bakterien gezwungen, im Lumen zu überleben. Das realisierten diese durch An-
passung der eigenen Teilungsrate an die Passagezeit der Ingesta, was eine Aus-
scheidung verhinderte.
Störungen des Ökosystems Darm haben vielseitige Ursachen. Eingebrachte patho-
gene und potentiell pathogene Keime in ausreichender Keimzahl können z.B. durch
Toxinbildung und –freisetzung die physiologische Darmfunktion stören (z.B.Vibrio
cholerae oder enterotoxische E.coli). Ernährungsbedingte Erkrankungen treten vor
allem in der westlichen Welt gehäuft auf. Tumoröse Darmerkrankungen werden mit
nahrungsbedingten Floraverschiebungen in Zusammenhang gebracht. Auch Aller-
gien als Überreaktionen des Immunsystems sind oft ernährungsassoziiert (BRANDT-
ZAEG 2002).
Das Ernährungs- und Trainingszentrum ETZ-M.Scholz entwickelte in Kooperation mit
der Universität Leipzig das Konzept: „MIRA–Mit Intelligenz richtig abnehmen“, das
Gewichtsreduktionsprogramme auf ihre Praxistauglichkeit testen und Leitlinien für
künftige Programme erstellen soll. Basierend auf diesem Konzept wurde die MIRA-
Studie durchgeführt, bei der ein kommerzielles Gewichtsreduktionsprogramm (BCM-
Programm© der PreCon GmbH & Co. KG) und ein nicht kommerzielles Gewichts-
reduktionsprogramm („Ich nehme ab“ der Deutschen Gesellschaft für Ernährung
e.V.) auf Wirksamkeit untersucht wurden (Scholz G.H. et al., 2002). Die Programme
sahen als Basis für die gewün-schte Gewichtsreduktion eine jeweils unterschiedliche
Ernährungsgrundlage der teilnehmenden Personen vor.
Grundlage für die vorliegende Arbeit ist die mikrobiologische Auswertung von Stuhl-
proben der Teilnehmer der MIRA-Studie über den gesamten Studienzeitraum von 52
Wochen. Da die Darmflora sich mit der Ernährung verändert und die Ernährungs-
2

Einleitung und Zielsetzung
weisen in beiden Gewichtsreduktionsprogrammen verschieden waren, werden die
Unterschiede in der Entwicklung der Darmfloren untersucht.
Ziel ist es, die Zusammenhänge zwischen Ernährung und Ausbildung der Darmflora
zu erfassen. Des Weiteren werden Interaktionen zwischen Darmflora und Wirt durch
Betrachtung der Ergebnisse der blutchemischen Untersuchungen sowie der erfolgten
Gewichtsreduktion untersucht.
3

Literaturübersicht
2 Literaturübersicht 2.1 Aufbau der Magen-Darm-Flora 2.1.1 Begriffsklärung
Ein Ökosystem, auch das der Darmflora, setzt sich aus Lebensräumen, den Habita-
ten und Nischen zusammen (ALEXANDER 1971). Dabei werden Habitate als physi-
sche Räume im System, die von Gemeinschaften autochthoner Mikroorganismen
besetzt sind, definiert. Nischen sind die Überlebensstrategien, die im jeweiligen
Lebensraum von den einzelnen Organismen entwickelt wurden.
In einem Habitat sind autochthone, d.h. ständig anwesende Mikroorganismen sowie
allochthone, d.h. transiente Mikroben vertreten. Die allochthonen Mikroorganismen
beeinflussen das Zusammenwirken der ansässigen Flora im Allgemeinen nur wenig
und zeigen weder Kolonisation noch Wachstum im gesunden Wirtsorganismus
(SAVAGE 1977b). Nur bei Störungen des intestinalen Ökosystems kann es vorkom-
men, dass allochthone Mikroorganismen ein von autochthonen Mikroorganismen
verlassenes Habitat besiedeln.
Um die Verwendung definierender Begriffe zu vereinheitlichen und damit eine Grund-
lage der Verständigung zu schaffen, schlugen DUBOS et al. (1965) vor, die Mikro-
organismen des Gastrointestinaltraktes nach der Populationsdichte einzuteilen. Die
als „indigene Flora“ bezeichneten Mikroorganismen wurde in die „autochthone Flora“,
die „Normalflora“ und „echte Pathogene“ unterteilt. Die „autochthone Flora“ entwickel-
te sich gemeinsam mit dem Wirtsorganismus, passte sich dadurch im Laufe der
Evolution an und ist in allen Individuen einer Spezies vertreten. Die „Normalflora“
kommt bei allen Mitgliedern einer Gemeinschaft vor, ist aber nicht zwingend in allen
Gemeinschaften der gleichen Spezies vertreten und die „echten Pathogene“ sind im
Normalfall nicht vertreten, werden zufällig aufgenommen, treten plötzlich auf und
haben die Fähigkeit, in Geweben zu persistieren. Diese Einteilung war jedoch aus
mehreren Gründen nicht eindeutig. Die als indigen bezeichneten Mikroorganismen
schlossen sowohl die ansässigen nichtpathogenen Bakterien als auch ansässige und
transiente Pathogene mit ein. Auch wird die für die Einteilung als „autochthone
Mikroorganismen“ erforderliche Populationsdichte von einigen zweifellos autoch-
thonen Bakterien bei Erwachsenen normalerweise nicht erreicht, z.B. bei E. coli und
Enterococcus faecalis. Daher änderte SAVAGE (1977a,b) das Konzept dahin
4

Literaturübersicht
gehend, dass die Begriffe „indigen“ und „autochthon“ im Zusammenhang mit
Mikroorganismen als Synonym verwendet wurden. Er definierte „Normalflora“ als
Zusammensetzung aus autochthoner Flora und allochthoner Flora (auch transiente
Flora). Als „Autochthone Flora“ werden von ihm Mikroorganismen bezeichnet, die in
allen Gemeinschaften einer Spezies permanent vorkommen und folgende Eigen-
schaften aufweisen: anaerobes Wachstum im Gastrointestinaltrakt (GIT), ständige
Präsenz im GIT bei adulten Individuen dieser Spezies, Besiedelung spezieller gastro-
intestinaler Nischen, Stabilisierung der Darmflora und eine häufig enge Bindung an
das Mukosaepithel. Unter dem Begriff „Allochthone Flora“ fasst er Mikroorganismen
zusammen, die vorübergehend auftreten. Sie kommen daher nicht immer in allen
Gemeinschaften bzw. nicht allen Individuen einer Gemeinschaft einer Spezies vor
und können keine Habitate besiedeln außer unter unphysiologischen Bedingungen.
Außerdem ist die Einteilung der Pathogene als separate Gruppe nach DUBOS et al.
(1965) nicht unbeschränkt gültig. Es gibt durchaus potentiell pathogene Keime, die
als autochthone Flora den Wirt besiedeln und nur bei Störungen des Ökosystems
ihre pathogene Wirkung entfalten.
2.1.2 Normalstatus Erwachsener Die intestinale Mikroflora Erwachsener ist umfassend untersucht. Sie erfasst außer-
ordentlich komplexe Populationen von aeroben, fakultativ und strikt anaeroben
Spezies, wobei die nicht sporenbildenden Anaerobier dominieren. Hefen, Pilze und
Protozoen wurden bei den meisten Untersuchungen übersehen und daher nur selten
quantifiziert (BORIELLO 1986). Viele Bakteriologen sind der Meinung, dass Proto-
zoen nicht zur Normalflora gehören und nur im Krankheitsfall zu isolieren sind. Aller-
dings verlaufen die meisten Fälle dieser Parasitosen asymptomatisch. Siedeln sich
Protozoen im Menschen an und vermehren sich, ohne eine Krankheit auszulösen,
werden sie gemäß der ökologischen Definition als indigen bezeichnet. Weiterhin
wurde über eine Reihe kommensal lebender Protozoen berichtet (GROVE 1984). Die
Mundhöhle des erwachsenen und gesunden Menschen ist von ca. 200 Spezies
besiedelt (BERG 1996) und der menschliche Speichel enthält ca. 109 Bakterien/ml,
die v.a. von den oralen Schleimhäuten der Zunge und der Wangen abgeschilfert
werden. Im Zahnbelag können bis zu 1011 Bakterien/g nachgewiesen werden, wobei
hier auch eine vielseitige Anaerobierflora zu finden ist.
5

Literaturübersicht
Tab.1: Besiedlung der Darmabschnitte, modifiziert nach Berg, R.D. (1996)
Bakterien der Haut: 1012 Körperzellen: 1013 Gastrointestinaltrakt: 1014 Mundhöhle: ca. 200 aerobe und anaerobe Spezies Magen: geringe Keimzahlen Hefen, Laktobazillen, Helicobacter pylori Ileum: 106-108 Bakterien pro Milliliter Darminhalt Kolon: 1010-1011 log10KbE/g pro Gramm Darminhalt
400-500 überwiegend anaerobe Spezies, z.B. Bacteroides, Pepto- streptococcus, Eubacterium, Bifidobacterium, Ruminococcus, Bacillus, Fuso- bacterium, Clostridium, Lactobacillus, Enterococcus
KbE=koloniebildende Einheiten, Kolonspezies absteigend nach Häufigkeit geordnet
Bei gesunden Menschen wird der Mageninhalt als nahezu steril angesehen. Werden
Mikroorganismen nachgewiesen, dann nur wenige Vertreter in geringsten Keimzah-
len. GIANELLA et al. (1972) fanden im Mageninhalt </= 103 KbE/ml alphahämo-
lytische Streptokokken, anaerobe Kokken, Laktobazillen, Staphylococcus epider-
midis, und Candida albicans. Diese Autoren weisen auch auf die Rolle des Magen-
pH beim Wachstum bzw. der Degradierung der Mikroorganismen, die mit der
Nahrung oder anderweitig aufgenommen werden, hin. So wurde bei Patienten mit
Hypo- oder Achlorhydria, also verminderter oder nicht vorhandener Salzsäureprodu-
ktion, eine deutlich höhere Gesamtkeimzahl nachgewiesen, speziell Anaerobier wie
Bacteroides sowie Koliforme, die im gesunden Magen nicht vorkommen.
Die Besiedlung des humanen Dünndarms unterscheidet sich gravierend von Region
zu Region. Das proximale Duodenum ist steril oder sehr gering besiedelt, die Flora
des distalen Dünndarms dagegen ähnelt der des Kolons. Das Ileumende gilt auch als
Übergangszone zwischen Dünn- und Dickdarm. GORBACH et al. (1967) entnahmen
Dünndarminhalt mittels einer Nasensonde. Im oberen Duodenum erwachsener und
gesunder Probanden wurden 0 bis 104,5 KbE/ml Streptokokken, Staphylokokken,
Laktobazillen sowie Hefen gefunden. Dagegen konnten Bacteroides und Koliforme
nicht kultiviert werden. Die Flora des distalen Ileums besteht aus 103,5-106,3 KbE/ml
mit einer der Kolonflora ähnlichen Zusammensetzung. Vor allem waren hohe
Keimzahlen koliformer Keime und Bacteroides spp. enthalten. Bei diesen
Untersuchungen konnten die Autoren einen Einfluss der Sonde auf die
Darmphysiologie und damit auf die Darmflora nicht ausschließen. Die Ergebnisse
stimmen mit denen der Studie von THADEPALLI et al. (1979) überein. Die für diese
Studie entnommenen Proben von gesunden erwachsenen Studienteilnehmern nach
6

Literaturübersicht
Abdominaltraumata aus dem oberen Duodenum waren bei 82% der untersuchten
Personen steril, bei 18% wiesen die Proben eine geringe Keimzahl überwiegend
fakultativ anaerober Keime auf. Die Proben aus dem mittleren Ileum waren zu 55%
steril. Bei den restlichen 45% der Proben wurden 105-109 KbE/ml koliforme Keime
und bei 2 Patienten Clostridien bis 107 KbE/ml gefunden.
Die zäkale Mikroflora des Menschen ist nur wenig untersucht, da allein die Proben-
gewinnung äußerst schwierig und damit limitiert ist. MARTEAU et al. (2001) ver-
glichen die Zäkalflora mit der Fäkalflora von acht gesunden Probanden. Dabei wurde
entdeckt, dass die anaerobe Gesamtkeimzahl, die Bifidobakterien- und Bacteroides-
Keimzahlen in den Zäkumproben deutlich geringer waren als in den Stuhlproben. Die
Keimzahl der fakultativ anaeroben Keime unterschied sich nicht signifikant, aber der
prozentuale Anteil an der Gesamtflora ist im Zäkum mit 25% deutlich höher als in den
Stuhlproben, wo sie lediglich 1% ausmachten.
Tab.2: Gesamtkeimzahlen der Anaerobier und fakultativen Anaerobier, Zellzahlen
Bifidobakterien und Bacteroides in Zäkum- und Stuhlproben bei acht gesunden Personen, modifiziert nach Marteau et al. (2001)
Gesamtkeimzahlen Zäkum Fäzes Signifikanz
Anaerobier 8,0±0,22 10,4±0,12 p=0,02 fakultative Anaerobier
7,4±0,23 7,8±0,29 p=0,38
Bifidobakterien 6,7±0,45 8,9±0,22 p=0,02 Bacteroides 7,4±0,33 8,9±0,42 p=0,05
Keimzahlen als Mittel ± s, log10 KbE/ml
Die Gesamtlänge des Dickdarms bei Erwachsenen beträgt ca. 150 cm und besteht
aus dem Zäkum, dem aufsteigenden, transversen und absteigenden Kolon sowie
dem Rectum (MACFARLANE et al. 1991). Der Dickdarm hat ein Volumen von ca.
500 ml und fasst ca. 220 g Inhalt (CUMMINGS et al. 1990).
Im Dickdarm des Menschen gibt es extrem vielschichtige mikrobielle Populationen
und eine große Auswahl spezifischer ökologischer Nischen. Damit ist der Dickdarm
die am dichtesten besiedelte Region im humanen Verdauungstrakt. Nach BERG
(1996) ist das Kolon der Hauptort der mikrobiellen Kolonisation, durch die langsame
Peristaltik von bis zu 60 Stunden Durchflusszeit begünstigt. Es werden 400-500
verschiedene Bakterienspezies vermutet, dazu kommen noch Hefen, Pilze und
Protozoen. Untersuchungen berichten über Gesamtkeimzahlen von über 1011 KbE/g
7

Literaturübersicht
(MOORE et al. 1974). Das ist mehr als das Zehnfache der Anzahl menschlicher
Körperzellen (SAVAGE 1977a). Die Bakterien sind zu 99,9% Anaerobier, d.h. sie
stellen eine 100-1000fach höhere Population als die fakultativ anaeroben Mikroorga-
nismen. Die vorherrschenden Genera sind Bacteroides, Bifidobacterium, Clostridium,
Eubacterium, Fusobacterium, Peptostreptococcus, Peptococcus und Ruminococcus.
Die Zusammensetzung der Intestinalflora im Kolon ist nicht in allen Teilen gleich,
daher ist die Fäkalflora nur in Grenzen für die Kolonflora repräsentativ. BENTLEY et
al. (1972) verglichen die Floren von Colon transversus und terminalem Ileum mit der
Fäkalflora. Die höchste Keimzahl konnten sie in den Stuhlproben feststellen mit
durchschnittlich 109,6 KbE/g Anaerobier und durchschnittlich 107,5 KbE/g koliforme
Bakterien. Die Keimzahlen des Colon transversus liegen ca. 2-3 Potenzen niedriger
als in den Fäkal-Proben und das terminale Ileum wies noch niedrigere Keimzahlen
auf. Es wurden nur wenig Anaerobier festgestellt mit durchschnittlich 104,4 KbE/g.
MOORE u. HOLDEMAN (1974) fanden in kultivierten Stuhlproben vorwiegend Anae-
robier wie Bakteroides, Fusobakterien, Bifidobakterien, Eubakterien und Peptostrep-
tokokken mit einer Keimzahl bis 1010 KbE/g. Clostridien und fakultative Anaerobier
lagen bei einem Keimgehalt von 108 bis 109 KbE/g.
Die beim erwachsenen Menschen als normal definierte Fäkalflora soll anhand der
Studie von FINEGOLD et al. (1983) erläutert werden. In dieser Studie wurden Stuhl-
proben von 141 Probanden untersucht. Darunter waren 13 strikte Vegetarier, 15 Ja-
paner mit dem in Japan üblichen Nahrungsspektrum (vor allem Fisch und kein Rind-
fleisch) und 62 Probanden mit einer in der westlichen Welt üblichen Ernährungswei-
se. Erstaunlicherweise sind die Keimzahlen der Vertreter der Hauptgattungen bei
allen untersuchten Personen ungeachtet der diätetischen und der Altersunterschiede
sehr ähnlich, so dass die Autoren aufgetretene Veränderungen eher bei nichtdomi-
nanten Bakterienspezies vermuten.
In sehr hohen Keimzahlen wurde in allen Proben Bacteroides festgestellt, als Haupt-
spezies wurde B. thetaiotaomicron identifiziert. Weiterhin wurden auch B. vulgatus,
B. distasonis, B. fragilis und B. ovatus in hohen Keimzahlen gefunden, was mit den
Ergebnissen anderer Studien übereinstimmt (DUERDEN 1980). Die am häufigsten
identifizierten anaeroben Kokken waren Peptostreptokokken, Ruminokokken, Veillo-
nella und anaerobe Streptokokken. Häufigster Vertreter war hierbei Peptostrepto-
coccus productus, nachgewiesen bei 29% der untersuchten Personen mit durch-
schnittlich 5x109 KbE/g. Vertreter der Gattung Eubacterium kamen bei 94% der
8
Literaturübersicht
untersuchten Personen mit durchschnittlich 5x1010 KbE/g als häufigste Anaerobier
vor. Es wurden v.a. E. aerofaciens, E. contortum, E. cylindroides, E. lentum und
E. rectale identifiziert. Mit einer durchschnittlichen Keimzahl von 4x109 KbE/g bei
78% der untersuchten Personen wurden Laktobazillen isoliert mit L. acidophilus als
häufigsten Vertreter. Bei den Bifidobakterien wurden vor allem B. adolescentis, B.
infantis und B. longum isoliert. Andere Autoren (KIMURA et al. 1997, McCartney
1996) berichten von relativ einheitlichen Bifidobacterium-Keimzahlen von durch-
schnittlich 109-1010 KbE/g Feuchtmasse, während die Zahl der Laktobazillen stark
von 104 bis 108 KbE/g Feuchtmasse variierte. Außerdem wurde für jeden Teilnehmer
eine eigene Komposition der predominanten Spezies ermittelt. Bei allen von FINE-
GOLD et al. (1983) untersuchten Personen konnten Clostridium-Spezies mit einer
durchschnittlichen Keimzahl von 6,30x109 KbE/g festgestellt werden. Die häufigste
Spezies war hier C. ramosum mit durchschnittlich 1,30x109 KbE/g bei 54% der
Untersuchungsteilnehmer. Bei C. perfringens wurden Keimzahlschwankungen fest-
gestellt: Bei den japanischen Probanden war sie mit 4x107 KbE/g bei 73% der Teil-
nehmer hoch und nicht feststellbar dagegen bei den strikten Vegetariern. Diese
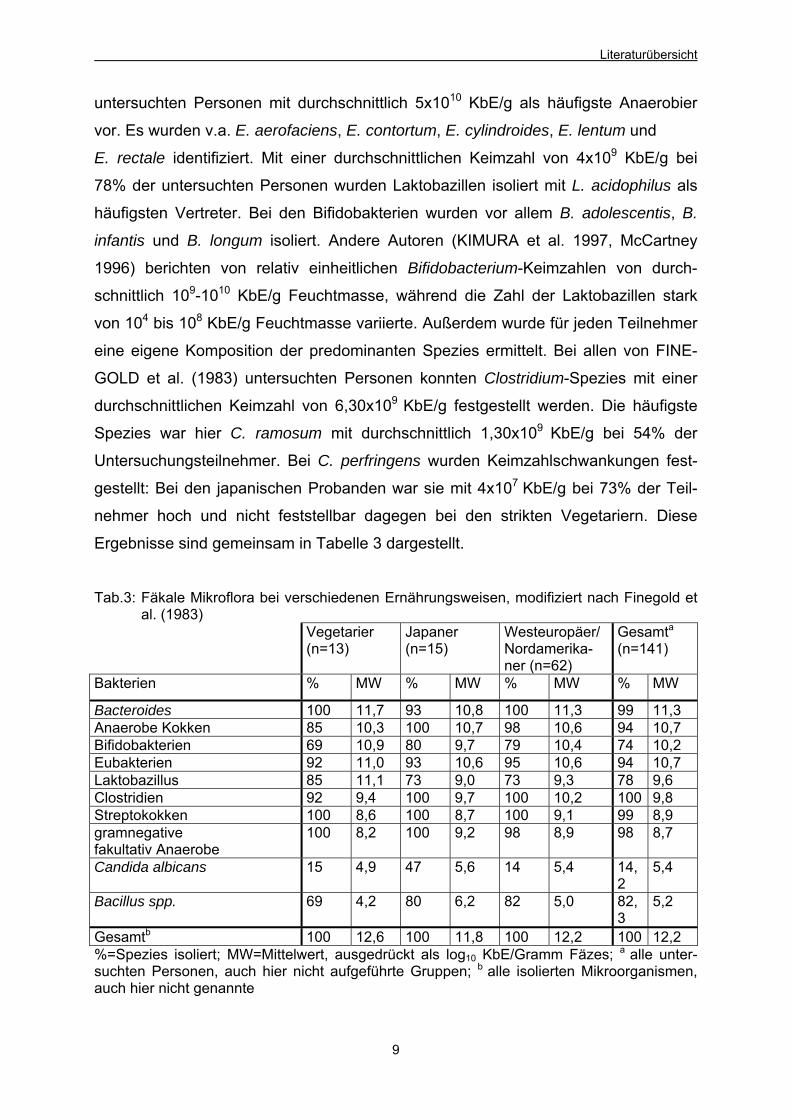
Ergebnisse sind gemeinsam in Tabelle 3 dargestellt.
Tab.3: Fäkale Mikroflora bei verschiedenen Ernährungsweisen, modifiziert nach Finegold et
al. (1983) Vegetarier
(n=13) Japaner (n=15)
Westeuropäer/Nordamerika-ner (n=62)
Gesamta(n=141)
Bakterien % MW % MW % MW % MW
Bacteroides 100 11,7 93 10,8 100 11,3 99 11,3 Anaerobe Kokken 85 10,3 100 10,7 98 10,6 94 10,7 Bifidobakterien 69 10,9 80 9,7 79 10,4 74 10,2 Eubakterien 92 11,0 93 10,6 95 10,6 94 10,7 Laktobazillus 85 11,1 73 9,0 73 9,3 78 9,6 Clostridien 92 9,4 100 9,7 100 10,2 100 9,8 Streptokokken 100 8,6 100 8,7 100 9,1 99 8,9 gramnegative fakultativ Anaerobe
100 8,2 100 9,2 98 8,9 98 8,7
Candida albicans 15 4,9 47 5,6 14 5,4 14,2
5,4
Bacillus spp. 69 4,2 80 6,2 82 5,0 82,3
5,2
Gesamtb 100 12,6 100 11,8 100 12,2 100 12,2 %=Spezies isoliert; MW=Mittelwert, ausgedrückt als log10 KbE/Gramm Fäzes; a alle unter-suchten Personen, auch hier nicht aufgeführte Gruppen; b alle isolierten Mikroorganismen, auch hier nicht genannte
9

Literaturübersicht
Andere Studien bezeichnen C. perfringens als überall vorkommende Spezies
(DRASAR et al. 1976a). Vor allem Enterokokken wurden von den grampositiven
fakultativ anaeroben Bakterien bei fast allen Probanden (99,3%) in relativ hoher
Keimzahl von durch-schnittlich 7,90x108 KbE/g isoliert. Am häufigsten war E. faecalis
vertreten. Koliforme Keime waren die häufigsten Vertreter der gramnegativen fakulta-
tiven Anaerobier, mit Schwerpunkt auf E. coli, mit einer Keimzahl von durchschnittlich
4x107 KbE/g bei 92,9% der Untersuchungsteilnehmer. Überall vertreten war Candida
albicans, innerhalb der japanischen Gruppe mit 4x105 KbE/g bei 47% der Untersu-
chungsteilnehmer und bei nur 14% der Untersuchungsteilnehmer mit einer westli-
chen Ernährungsweise. Pseudomonas aeruginosa war bei auffallend wenigen Teil-
nehmern zu isolieren. Die Ergebnisse von BUCK u. COOKE (1969) sagen ebenfalls
aus, dass gesunde Erwachsene wahrscheinlich resistent gegen eine Besiedlung mit
Pseudomonas aeruginosa sind. Bei nur 10,6% der hier untersuchten Probanden
wurden mit einer Keimzahl bis max. 105 KbE/g wenig Pseudomonas aeruginosa
gefunden.
2.1.3 Die Entwicklung vom Neugeborenen zum Mischköstler
Es wird allgemein angenommen, dass der Gastrointestinaltrakt (GIT) bei der Geburt
steril ist, allerdings berichtet eine Studie (MIMS et al., 1972) von per Kaiserschnitt
entwickelten Babies, in deren Mägen lebensfähige Bakterien gefunden wurden.
Beträchtliche Unterschiede existieren bei den Zeitspannen, die die Bakterien für die
Besiedlung der Mundhöhle und nachfolgend des GIT benötigen. Diese hängen von
der Art der Geburt und der Exposition während des Geburtsvorganges ab (SAVAGE
1977b). Das Neugeborene ist durch die Geburt mit Bakterien aus dem Anal- und
Genitaltrakt der Mutter und bei gestillten Babies zusätzlich durch den intensiven
Hautkontakt mit der stillenden Mutter besiedelt. Vor allem werden von Frühgebore-
nen und Kaiserschnittkindern Bakterien auch aus der Umwelt aufgenommen. So
werden Keime von anderen Säuglingen beispielsweise über Belüftungsanlagen ver-
breitet und durch das Betreuungspersonal und mit der Ausrüstung übertragen.
Bereits innerhalb der ersten 24 Lebensstunden konnten aus dem GIT der Neugebo-
renen Bakterien isoliert werden, darunter fakultative Anaerobier wie Enterobakterien,
Streptokokken, Laktobazillen und Clostridien, weiterhin Aerobier wie Mikrokokken
und strikte Anaerobier wie Bakteroides (MATA et al. 1971).
10

Literaturübersicht
Viele Studien über die Besiedlung Neugeborener mit Bakterien wurden in Abhängig-
keit von Ernährung und Alter durchgeführt. Interessanterweise lässt sich bei allen
Untersuchungen der verschiedenen Altersklassen ein Muster erkennen, was dem in
der Studie von STARK u. LEE (1982) ähnelt. Diese verglichen je eine Gruppe gesun-
der gestillter und eine Gruppe mit Muttermilchersatz gefütterter Säuglinge in den
ersten 12 Monaten. Es zeigte sich, dass fakultativ anaerobe Mikroorganismen die
Erstbesiedler sind, die die im Darm vorhandene Sauerstoffspannung reduzieren und
damit den strikten Anaerobiern die Kolonisation ermöglichen. Eine bestimmte Anzahl
Anaerobier folgt ihnen innerhalb der ersten 14 Tage. Nach 2 Tagen sind in beiden
Säuglingsgruppen Streptokokken und Enterobakterien mit Keimzahlspitzen am Ende
der ersten Woche präsent. Den größten Anteil der Anaerobier stellen in den ersten
Lebenstagen die Bifidobakterien, während Clostridien, anaerobe Streptokokken und
Bacteroides erst allmählich in den ersten 2 Lebenswochen auftreten. Die E. coli- und
Streptokokken-Keimzahlen erreichen ca. 109 KbE/g. Bei den gestillten Kindern gehen
die Keimzahlen von E. coli und Enterococcus nachfolgend stark zurück, während
Clostridien und Bacteroides vollständig verschwinden. Diese Entwicklung trat nicht
bei den mit Ersatzmilch gefütterten Kindern auf, so dass diese eine komplexere
Darmflora entwickelten. Im Alter von 4 Wochen besteht die Darmflora gestillter
Kinder vorwiegend aus Bifidobakterien mit ca. 4x1010 KbE/g, bis eine Zufütterung
einsetzt. Über eine dominante Bifidobakterien-Spezies gibt es keine Überein-
stimmung in diversen Studien, während bei Bacteroides die Spezies B. distasonis, B.
vulgatus und B. fragilis als vorherrschend beschrieben werden. Mit dem Beginn der
Aufnahme fester Nahrung verschwindet dieser Unterschied zwischen gestillten und
mit Muttermilchersatz gefütterten Babies. Mit 12 Monaten beginnt die anaerobe Flora
der Kleinkinder in Zahl und Zusammensetzung der der Erwachsenen zu ähneln.
Dieser Prozess ist im Alter von ca. 2 Jahren abgeschlossen und von einer Keimzahl-
verminderung, v.a. der Streptokokken und fakultativen gramnegativen Anaerobier,
begleitet, während die Anaerobier, v.a. Bacteroides und Laktobazillen, vorherrschend
werden.
11

Literaturübersicht
2.2 Die wandständige Flora und der Vergleich mit der Lumenflora
Quantitative Untersuchungen der mukosaassoziierten Darmflora sind selten. Erst in
jüngerer Vergangenheit wuchs das Interesse an Informationen über die Zusammen-
setzung der mit der Darmwand assoziierten Flora und deren Zusammenwirken mit
der Luminalflora. Grund dafür war beispielsweise die allein durch die luminale Flora
nur unzureichend erklärte Barrierefunktion des Darmes gegen Pathogene und Un-
kenntnis über die Rolle bei der Entstehung von Darmerkrankungen.
POXTON et al. (1997) untersuchten Biopsieproben von zwölf Personen. Davon
waren sechs an Ulzerativer Kolitis erkrankt und sechs wiesen keine entzündlichen
Darmveränderungen auf. Die Proben wurden im Rektum und im proximalen Kolon
entnommen. Zwischen den Proben einer Person wurden keine signifikanten Unter-
schiede festgestellt. Die Gesamtzahl der fakultativen Anaerobier lag bei 2,4x103 bis
1,3x106 KbE/Probe und die anaerobe Gesamtkeimzahl wurde mit bis zu 3,0x107
KbE/Probe ermittelt. In allen Proben waren Bacteroides spp. vorherrschend mit bis
zu 1,4x107 KbE/Probe, was einem Anteil an der anaeroben Gesamtflora von 66%
entspricht. Laktobazillen und Bifidobakterien wurden nicht bei allen Patienten und in
relativ geringen Keimzahlen festgestellt. Der Vergleich der Vorkommen von
Bacteroides spp. zeigt deutliche Unterschiede zwischen der lumen- und der
wandständigen Flora.
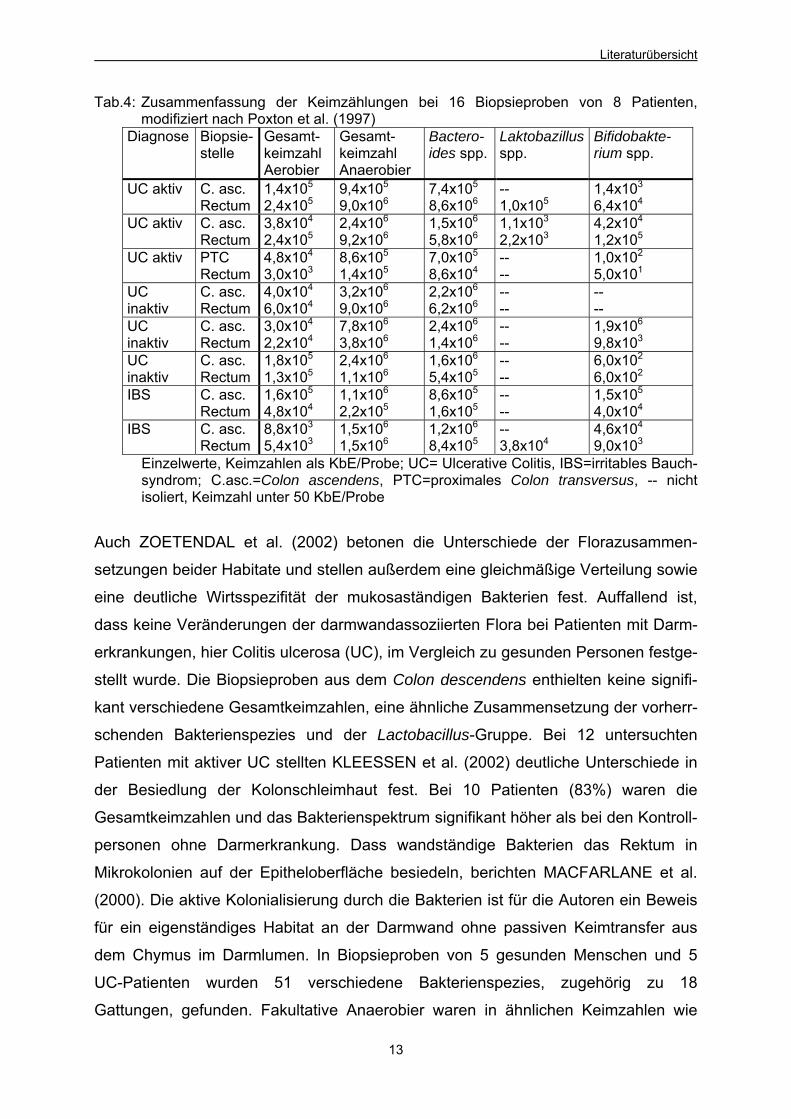
Tabelle 4 enthält als einen Ausschnitt der Gesamtergebnisse die Keimzahlen von 16
Proben, die bei 8 Patienten entnommen wurden.
12
Literaturübersicht
Tab.4: Zusammenfassung der Keimzählungen bei 16 Biopsieproben von 8 Patienten, modifiziert nach Poxton et al. (1997)
Diagnose Biopsie-stelle
Gesamt-keimzahl Aerobier
Gesamt-keimzahl Anaerobier
Bactero-ides spp.
Laktobazillus spp.
Bifidobakte-rium spp.
UC aktiv C. asc. Rectum
1,4x105
2,4x1059,4x105
9,0x1067,4x105
8,6x106-- 1,0x105
1,4x103
6,4x104
UC aktiv C. asc. Rectum
3,8x104
2,4x1052,4x106
9,2x1061,5x106
5,8x1061,1x103
2,2x1034,2x104
1,2x105
UC aktiv PTC Rectum
4,8x104
3,0x1038,6x105
1,4x1057,0x105
8,6x104-- --
1,0x102
5,0x101
UC inaktiv
C. asc. Rectum
4,0x104
6,0x1043,2x106
9,0x1062,2x106
6,2x106-- --
-- --
UC inaktiv
C. asc. Rectum
3,0x104
2,2x1047,8x106
3,8x1062,4x106
1,4x106-- --
1,9x106
9,8x103
UC inaktiv
C. asc. Rectum
1,8x105
1,3x1052,4x106
1,1x1061,6x106
5,4x105-- --
6,0x102
6,0x102
IBS C. asc. Rectum
1,6x105
4,8x1041,1x106
2,2x1058,6x105
1,6x105-- --
1,5x105
4,0x104
IBS C. asc. Rectum
8,8x103
5,4x1031,5x106
1,5x1061,2x106
8,4x105-- 3,8x104
4,6x104
9,0x103
Einzelwerte, Keimzahlen als KbE/Probe; UC= Ulcerative Colitis, IBS=irritables Bauch-syndrom; C.asc.=Colon ascendens, PTC=proximales Colon transversus, -- nicht isoliert, Keimzahl unter 50 KbE/Probe
Auch ZOETENDAL et al. (2002) betonen die Unterschiede der Florazusammen-
setzungen beider Habitate und stellen außerdem eine gleichmäßige Verteilung sowie
eine deutliche Wirtsspezifität der mukosaständigen Bakterien fest. Auffallend ist,
dass keine Veränderungen der darmwandassoziierten Flora bei Patienten mit Darm-
erkrankungen, hier Colitis ulcerosa (UC), im Vergleich zu gesunden Personen festge-
stellt wurde. Die Biopsieproben aus dem Colon descendens enthielten keine signifi-
kant verschiedene Gesamtkeimzahlen, eine ähnliche Zusammensetzung der vorherr-
schenden Bakterienspezies und der Lactobacillus-Gruppe. Bei 12 untersuchten
Patienten mit aktiver UC stellten KLEESSEN et al. (2002) deutliche Unterschiede in
der Besiedlung der Kolonschleimhaut fest. Bei 10 Patienten (83%) waren die
Gesamtkeimzahlen und das Bakterienspektrum signifikant höher als bei den Kontroll-
personen ohne Darmerkrankung. Dass wandständige Bakterien das Rektum in
Mikrokolonien auf der Epitheloberfläche besiedeln, berichten MACFARLANE et al.
(2000). Die aktive Kolonialisierung durch die Bakterien ist für die Autoren ein Beweis
für ein eigenständiges Habitat an der Darmwand ohne passiven Keimtransfer aus
dem Chymus im Darmlumen. In Biopsieproben von 5 gesunden Menschen und 5
UC-Patienten wurden 51 verschiedene Bakterienspezies, zugehörig zu 18
Gattungen, gefunden. Fakultative Anaerobier waren in ähnlichen Keimzahlen wie
13

Literaturübersicht
obligate Anaerobier präsent mit 106-107 Bakterien/cm2. Bacteroides und Bifidobac-
terium waren die häufigsten Gattungen mit den höchsten Keimzahlen. Eine gleich-
zeitige Besiedlung mit Bacteroides vulgatus und B. thetaiotaomicron kam nicht vor.
Daraus schlossen die Autoren, dass beide Spezies die gleichen oder sehr ähnliche
Nischen im Darm besiedeln. Bei allen gesunden Probanden wurde Bifidobacterium
adolescentis mit hohen Keimzahlen (bis 105 Bakterien/cm2) identifiziert, während
diese Spezies bei den UC-Patienten nicht nachgewiesen werden konnte. Eine
größere Patientengruppe mit entzündlichen Darmerkrankungen (IBD) wurde von
SWIDSINSKY et al. (2002) untersucht. 305 Patienten mit IBD mit verschiedenen
Ausprägungen und 40 Kontrollpersonen waren einbezogen. Auch hier wurden bei
IBD-Patienten höhere Keimzahlen in der Mukosa festgestellt als bei gesunden Pro-
banden, wobei die Konzentration der Bakterien mit dem Schweregrad der Erkran-
kung anstieg. Bei Patienten mit mehr als 1x104 KbE/µl wurde eine mit der Mukosa
verbundene Schicht aus Bakterien gefunden, die keine Zeichen von Translokationen
aufwiesen. Bei Erkrankten mit mehr als 5x104 KbE/µl wurden in der Lamina propria
Einschlüsse polymorpher Bakterien entdeckt. Die isolierten Bakterien waren fäkalen
Ursprungs. Die Autoren schlossen aus ihren Ergebnissen, dass die mikrobiologi-
schen Veränderungen keine Folge der Entzündung, sondern der spezifischen
Antwort des Wirtes ist. Sie vermuteten, dass die Mukosa im gesunden Zustand in der
Lage ist, die fäkalen Bakterien zurückzuhalten, während bei Patienten mit IBD diese
Funktion grundlegend gestört ist.
2.3 Stabilität, Kontrolle und Beeinflussbarkeit der Darmflora 2.3.1 Einblick in das Ökosystem des Gastrointestinaltraktes mit
Wechselwirkungen zwischen den Bakterien
Das Ökosystem des menschlichen Dickdarmes besteht aus vielfältigsten Komponen-
ten und unterliegt demzufolge zahlreichen Einflüssen. Eine Hauptkomponente ist die
Intestinalflora, deren zahlreiche und zum großen Teil noch unerforschte innere
Wechselwirkungen die Gesundheit des Wirtes und die Zusammensetzung und
Struktur der Intestinalflora selbst bestimmen. Andere Komponenten sind die folgen-
den vier Mikrohabitate, die im Kolon des Erwachsenen benannt werden können: Die
Oberfläche der Epithelzellen, die Mukusschicht auf den Villi und innerhalb der Kryp-
ten sowie der Darminhalt im Lumen. Die Rolle eines jeden Habitats ist sehr unter-
14

Literaturübersicht
schiedlich (FRETER 1983), jedoch stehen alle miteinander in Verbindung und beein-
flussen einander. Beispielsweise stellen die Krypten zwischen den Darmzotten bei
Darmabschnitten mit hoher Durchflussrate, wie z.B. dem Duodenum, wichtige Mikro-
habitate dar. Sie werden im Gegensatz zum Lumen deutlich weniger von der Peri-
staltik beeinflusst und ermöglichen so auch nichtwandständigen Bakterien die
Kolonisation (ROEDIGER 1986).
Ein Beispiel für die unüberschaubaren Interaktionen zwischen den Bakterien ist das
Bereitstellen zusätzlicher Energiequellen durch das Zerlegen der vom Wirt aufge-
nommenen Nahrung. Diese „versorgenden“ Interaktionen können eingeteilt werden
in die Verstoffwechselung von Primärsubstraten, die von extrazellulären Enzymen
verdaut wurden und der Metabolismus von Stoffwechselendprodukten anderer Mikro-
organismen. Diese Komplexität der verfügbaren Nahrungsquellen ermöglicht es
vielen Spezies, innerhalb des Darmes zu überleben und damit die große Diversität zu
erhalten (MACFARLANE und CUMMINGS 1991). Die Konkurrenz um Nahrung, also
um Kohlenstoff- und Energiequellen, ist bei FRETER (1983) der Hauptkontrollmecha-
nismus bei Bakterienpopulationen in einem kontinuierlich arbeitenden Kultursystem
mit Bakterien der Mauszäkalflora. Daraus entwickelte er die Theorie, dass jede
Spezies einige wenige Substrate besser verstoffwechseln kann als die anderen und
dass daher das Wachstum und die Aktivität einer Population durch die vorhandenen
Mengen dieser „begrenzenden Substrate“ kontrolliert werden.
Ein anderer Aspekt bakterieller Wechselwirkungen ist der Schutz des Wirtes vor
Krankheitserregern. Dazu gibt es zwei Diskussionsebenen: Zum einen den Effekt der
indigenen Mikroflora auf potentielle Pathogene und zum anderen die Anfälligkeit des
Ökosystems gegenüber Pathogenen (HENTGES 1992). Die Darmbesiedlung kann
vom Kampf um Nahrung und von den Metaboliten der Bakterien, die die Invasoren
antagonisieren, beeinflusst sein.
2.3.2 Stabilität des Ökosystems Das Kolon gesunder Erwachsener ist ein stabiles Ökosystem, in dem eindringenden
oder allochthonen Mikroorganismen das Überleben schwer fällt. In diesem Öko-
system sind alle Habitate von Vertretern der indigenen, auch autochthonen, Mikroflo-
ra besetzt (BERG 1996). Daher können transiente Spezies, die über Nahrung,
Wasser, Kontakt mit anderen Individuen etc. zugeführt werden, sich normalerweise
15

Literaturübersicht
nicht ansiedeln. Das Gleiche gilt für indigene Mikroorganismen. Jede Spezies ist oft
nur in einem oder wenigen Habitaten indigen. Nach Ablösung beispielsweise durch
Abschilferung werden sie transient und mit den Fäzes ausgeschieden. So sind die
Habitate im Darm von stabilen Mikroorganismen-Gemeinschaften besiedelt, was die
Stabilität der Darmflora in Populationsmenge und Spezieszusammensetzung erklärt.
Diese wird nur durch drastische Ernährungsumstellungen (DRASAR et al. 1974) oder
das Eindringen pathogener Mikroorganismen gestört. Die Stabilität der Florazusam-
mensetzung hängt auch von der Teilungsgeschwindigkeit der Bakterien vor allem im
Darmlumen ab. Mikroorganismen, die sich langsamer vermehren als die Exkretions-
rate des Wirtes, werden zwangsläufig ausgeschieden und können sich nicht ansie-
deln. Also können im Lumen nur Bakterien mit einer Teilungsrate überleben, die
mindestens so schnell ist wie die Exkretionsrate des Wirtes. Eine deutlich höhere
Vermehrungsrate der Bakterien oder eine verminderte Exkretionsrate führen dage-
gen zu einer Überwucherung der indigenen Flora, damit zu einer Störung des Gleich-
gewichtes und zu Erkrankungen des Darmtraktes wie z.B. dem „owergrowth-
syndrom“.
Das intestinale Ökosystem ist vielen Faktoren ausgesetzt, die dessen Stabilität
beeinflussen können. Einflusskriterien auf das Ökosystem des Kolon können grob
unterteilt werden in Ernährung, Wirtsphysiologie und die Mikroflora (SAVAGE
1977b). Bei einem gesunden Erwachsenen besteht zwischen diesen Komponenten
ein labiles dynamisches Gleichgewicht. Das Verständnis dieser Interaktionen könnte
die Reaktion des Wirtes auf verschiedene Umwelteinflüsse vorhersagen helfen. Auch
TANNOCK (1983) misst der Physiologie des Wirtes Bedeutung zu: Er fand heraus,
dass jeder externe Einfluss, der sich nachteilig auf ein Tier auswirkt, in der Lage ist,
durch Veränderungen in der Wirtsphysiologie das gastrointestinale Ökosystem als
Antwort auf diesen Einfluss zu verändern. So kann jede Veränderung der Umgebung
oder der Ernährung, die auf die optimalen Funktionsabläufe des Wirtes nachteilig ein-
wirkt, das gastrointestinale Ökosystem direkt oder indirekt beeinflussen.
Die vorhandene Diversität der indigenen Bakterien schützt die Stabilität des Öko-
systems (ALEXANDER 1971), erschwert allerdings auch das Verständnis der Kon-
trollmechanismen innerhalb des Systems enorm. Zu jeder Zeit ist jede Spezies
vielfältigen Einflüssen ausgesetzt, die bei Untersuchungen einzelner Bakterien nicht
auftreten. Diese Studien liefern somit keine Ergebnisse, die direkt auf das Gesamt-
system übertragbar sind. GORBACH et al (1967) beobachteten die Schwankungen
16

Literaturübersicht
einiger Spezies der untersuchten Fäkalflora im Wochentakt. Dabei waren die Verän-
derungen so gering, dass die Gesamtflora des Kolon auch über längere Zeiträume
als stabil betrachtet werden kann.
2.3.3 Einfluss der Ernährung des Wirtes auf die Darmflora
Die Ernährung des Wirtes ist auch Energiequelle für die intestinale Mikroflora. Der
Hauptanteil der Nahrung, die den oberen Verdauungstrakt unverdaut passiert,
besteht aus Kohlenhydraten. Bis zu 60 g Kohlenhydraten gelangen pro Tag ins
Kolon, davon sind bis zu 40 g pankreasamylaseresistente Stärke, die aber nicht
resistent gegen bakterielle Enzyme sein muss, sowie bis zu 18 g/d sind Nichtstärke-
Polysaccharide (CUMMINGS und MACFARLANE 1991). Der endogene Kohlen-
hydrat-Eintrag beispielsweise aus Sekreten verschiedener Darmdrüsen beträgt bis zu
3 g/d.
Kohlenhydrate werden im Allgemeinen in Form von Polysacchariden aufgenommen.
Die durch Enzyme daraus gespaltenen Einfachzucker werden im oberen Verdau-
ungstrakt absorbiert. Die unterschiedliche Verfügbarkeit der Substrate beruht auf den
unterschiedlich hohen Fermentationsraten der Kohlenhydrate in den verschiedenen
Darmabschnitten.
Proteine können ebenfalls verwendet werden (MACFARLANE et al. 1991). Den Dick-
darm erreichen bis zu 9 g/d aus der Nahrung und bis zu 6 g/d aus endogenen Enzy-
men. Die Proteinfermentation nimmt nach distal zu (MACFARLANE et al. 1992).
Diese uneinheitliche Verteilung der Nährstoffe spiegelt sich in der Zusammensetzung
der Intestinalflora in den verschiedenen Darmabschnitten wider.
Der Effekt der Nahrung auf die Fäkalflora ist intensiv untersucht worden, wobei aber
nur wenige direkte Einflüsse beobachtet wurden. Frühe Studien konnten keine Ver-
änderungen der Keimzahlen berichten (DRASAR et al. 1974, FINEGOLD et al.
1978), ausgenommen nach dem Verabreichen einer synthetischen Nahrung, deren
Bestandteile chemisch genau definiert waren (WINITZ et al. 1970). FINEGOLD et al.
(1983) beschäftigten sich mit dem Problem der Beeinflussbarkeit der Darmflora.
Alter, Ernährung, Herkunft bzw. Aufenthaltsort, also auch geographische Einflüsse,
wurden untersucht. Die festgestellten Verschiebungen innerhalb der untersuchten
Darmbakterien waren sehr gering. Es konnte jedoch nicht ausgeschlossen werden,
dass grundlegende Veränderungen der Lebensumstände des Wirtes wie Hunger
17

Literaturübersicht
oder ausschließlich parenterale Ernährung deutlichere Änderungen als die bei
FINEGOLD et al. (1983) festgestellten bewirken könnten. Bei der Auswertung dieser
Ergebnisse müssen die damaligen technischen Möglichkeiten in Betracht gezogen
werden, die die Identifizierung vieler Darmbakterien gar nicht ermöglichten und damit
ein unvollständiges Bild der Veränderungen lieferten. Die Ziele der Studien liefen
nicht auf eine Gesamterfassung der durch Ernährung hervorgerufenen Veränderun-
gen hinaus. Sie waren entweder beschränkt auf die Feststellung der Wirkung einzel-
ner Nahrungsbestandteile wie Protein- oder Fettmengen oder die Biotransformatio-
nen von potentiellen Karzinogenen (SIMON et al. 1981).
Eine detailliertere Untersuchung der Veränderung der Darmflora als Reaktion auf
verschiedene Ernährungsweisen wurde von FINEGOLD et al. (1983) durchgeführt.
Diese Studie wurde im Abschnitt „Aufbau der Darmflora“ vorgestellt, so dass hier
nicht weiter darauf eingegangen werden soll.
Grundlegende Änderungen der Ernährung allein scheinen wenig Einfluss auf die
Darmflora zu haben (TANNOCK 1983). Wenn man das Auftreten der Reisediarrhoe
betrachtet, könnte allerdings das Zusammenwirken mehrere Faktoren den Einfluss
der Ernährung verstärken. Der Reisedurchfall an sich ist in den meisten Fällen die
Folge einer Campylobacter-Infektion, die durch die Aufnahme weniger Keime ausge-
löst wird. Die Empfindlichkeit auf Darmpathogene ist oft bei Reisenden erhöht, die
gleichzeitig eine Nahrungsumstellung erfahren sowie in Menge und Art unbekannten
Pathogenen ausgesetzt sind und vielleicht durch lange Reisezeiten und eine unge-
wohnte Umgebung wie die Höhe der Temperatur oder der Luftfeuchtigkeit zusätzlich
gestresst sind. Die Vermutung liegt nahe, dass die Empfänglichkeit für Krankheits-
erreger bei Nahrungsumstellung und gleichzeitig verstärktem Erregerdruck und
Stress auch durch Überlastung der Immunabwehr steigt.
Die bei anderen Autoren (BORIELLO 1986) geäußerten Vermutungen, große
Mengen Rohfaser und Protein in der Nahrung rufen keine Veränderung der Mikro-
flora hervor, bestätigten sich nicht. Vielmehr bewirkten erhöhte Mengen Kohlenhydra-
te einen Anstieg der Bifidobakterien und eine fettreiche Diät erhöhte die Bacteroides-
Keimzahl, während eine Elementardiät ein Absinken von Enterokokken und Laktoba-
zillen bewirkte. Außerdem sind Enterobakterien und Laktobazillen Indikatoren für
Stress einschließlich Ernährungsstress, wobei die Laktobazillenzahlen absinken und
die Keimzahlen der anderen Bakterien ansteigen (TANNOCK 1983).
18

Literaturübersicht
Aus Tierversuchen ist bekannt, dass Nahrungsentzug Veränderungen der Mikroflora
hervorrufen kann (TANNOCK 1983). Diese Änderungen wurden nicht nur bei fäkalen
und luminalen, sondern auch bei mukosa-assoziierten Darmbakterien beobachtet.
Studien zu Veränderungen der mukosa-assoziierten Flora gibt es nur wenige. Bei
Nagern wurde beobachtet, dass Nahrungskomponenten keinen Effekt auf die lumin-
ale Mikroflora hatten, während deutliche Veränderungen bei den mukosa-assoziie-
rten Mikroorganismen beobachtet wurden.
2.3.4 Einfluss von Prä- und Probiotika auf die Mikroflora
Schon Metchnikoff vermutete 1908, dass reichhaltige, schwere Mahlzeiten und
Alkohol vermieden werden sollten, um nicht eine Schädigung des Darmes hervorzu-
rufen und dass die Aufnahme fermentierter Milch lebensverlängernd wirken könnte.
Seitdem wurden bei Menschen und Tieren zahlreiche Einflussmöglichkeiten ent-
deckt. Die Forschungen wurden ausgedehnt, da es für wichtig erachtet wurde, über
diese Mechanismen verfügen zu können, um z.B. die Folgen einer Antibiotika-Thera-
pie zu mildern. BENGMARK (1995) analysierte die Veränderungen der Ernährungs-
gewohnheiten der letzten 100 Jahren. Diese waren vor allem in Ländern der westli-
chen Welt häufig dramatisch. Genannt werden der stark gestiegene Konsum raffinier-
ten Zuckers und gesättigter Fette, die 10fache Aufnahme von Natrium und die
erheblich gesunkene Zufuhr pflanzlicher Fasern und von Mineralstoffen, Vitaminen,
ungesättigten Fetten und Antioxidantien. Er schlußfolgert, dass die in vielen Jahrtau-
senden an die urzeitliche Ernährungs- und Lebensweise der menschlichen Vorfahren
angepassten Gene sich nicht an die erfolgten rasanten Wechsel adaptieren konnten.
Darin sieht der Autor eine Ursache für das gehäufte Auftreten entzündlicher, degene-
rativer und neoplastischer Erkrankungen vor allem des Verdauungstraktes. Eine
mindestens ebenso große Bedeutung misst der Autor der dramatisch verminderten
Aufnahme von Bakterien bei. Der Rückgang beim Konsum fermentierter Produkte
und die vermehrt angebotenen mehrfach bearbeiteten Nahrungsmittel verringerten
die Aufnahme der heute probiotisch genannten Keime. Die daraus erfolgte Verände-
rung der Darmflora mit den entsprechenden Auswirkungen auf den MDT können zu
den genannten und weiteren Erkrankungen führen.
In Anbetracht der in den letzten Jahrzehnten immer stärker zunehmenden Resisten-
zen von Bakterien gegen antimikrobielle Wirkstoffe und der vor allem bei Langzeit-
19

Literaturübersicht
therapien auftretenden Nebenwirkungen finden andere Möglichkeiten der Abwehr-
steigerung und Krankheitsprävention und –behandlung wachsende Aufmerksamkeit.
Hier zu besprechen sind die Prä- und Probiotika. Präbiotika sind unverdauliche
Nahrungsbestandteile, die eine Energiequelle für eine Vielzahl von Kolonbakterien
darstellen und die Vermehrung und/oder die Aktivität dieser Mikroorganismen mit
positiven Auswirkungen auf den Wirt stimulieren (ROBERFROID 1998). Die bekann-
testen Präbiotika sind Inulin als unverdauliches Fruktooligosaccharid und Lactulose
als nichtspaltbares Disaccharid. Inulin und sein enzymatisches Spaltprodukt Oligo-
fruktose kommen in vielen Gemüsepflanzen (z.B. Zwiebeln, Spargel), in Getreide und
in Obst (z.B. Bananen) vor. In mehreren in vivo –Humanstudien ist der stimulierende
Einfluss von Inulin und Oligofruktose auf das Wachstum der Bifidobakterien belegt
(GIBSON et al. 1995; KLEESSEN et al. 1997; GIBSON 1999). Andere Autoren
beschreiben neben dem bifidogenen Effekt den Einfluss auf blutchemische Para-
meter wie Lipide (DAVIDSON und MAKI 1999) und Immunstimulation, gesteigerte
Vitaminsynthese und Senkung des Kolonkrebs-Risikos (JENKINS et al. 1999). Laktu-
lose ist ein synthetisch hergestelltes Disaccharid, bestehend aus Fruktose und
Galaktose. Seit über 40 Jahren wird es in der Medizin haupsächlich zur Behandlung
der portosystemischen Enzephalopathie und bei Verstopfung verwendet. Durch
seine lokale Wirkung ist es ein ausgesprochen mildes Therapeutikum (SCHUMANN
2002). Weiterhin wird Laktulose nur im Dickdarm von Bakterien gespalten, wobei vor
allem SCFA und Gase freigesetzt werden. Durch die einsetzende pH-Wert-Senkung
wird gemeinsam mit dem osmotisch gebundenen Wasser die Peristaltik stimuliert
und die Flora des Dickdarms beeinflusst. Es werden säuretolerante Spezies geför-
dert. Ein besonders positiver Einfluss ist auf Laktobazillen und speziell Lactobacillus
acidophilus beobachtet worden (SALMINEN und SALMINEN 1997).
Als Probiotika werden lebende Mikroorganismen bezeichnet, die nach Aufnahme die
Prävention bzw. Behandlung bestimmter Erkrankungen unterstützen. Der Trend geht
hier zur oralen Applikation lebender Mikroorganismen, v.a. Laktobazillen und Bifido-
bakterien zur Stabilisierung der Darmflora. Weiterhin eignen sich auch Stämme von
E.coli, Subspezies von Streptococcus, Enterococcus, Bacteroides, Bacillus, Propioni-
bacterium sowie verschiedene Pilze als Probiotika (ROLFE 2000). Die zu Verfügung
stehenden probiotischen Produkte unterscheiden sich z.T. erheblich in Konzentration
und Wirksamkeit. Um diese Unterschiede zu verdeutlichen, nehmen RUSCH und
RUSCH (2001) eine Aufteilung in Gruppen vor. Medizinische Probiotika sind Arznei-
20

Literaturübersicht
mittel, die als Wirkstoffe lebende oder/und abgetötete Mikroorganismen enthalten.
Sie werden gezielt bei entsprechender Indikation eingesetzt. Als Pharmazeutische
Probiotika werden in Apotheken erhältliche Produkte bezeichnet. Sie enthalten die
Mikroben als Lyophylisat und kommen als Nahrungsergänzung zum Einsatz. Zu den
Alimentären Probiotika werden die handelsüblichen Erzeugnisse wie Milchdrinks
oder Joghurt, die Bakterienkulturen enthalten und zur Gesundheitsunterstützung
empfohlen werden, gezählt. Das Wirkungsspektrum der Probiotika ist breit gefächert
Es wird von Prävention vor Diarrhöen sowie der Durchfallbehandlung sowohl bei
bakterieller (GOLDIN et al. 1992) als auch viraler Ursache berichtet und der Fähigkeit
der Probiotika, karzinogene Spaltprodukte der Darmbakterien zu binden (RUSCH
und RUSCH 2001). Die probiotischen Bakterien wirken antagonistisch gegen poten-
tiell pathogene Erreger. So sind Wechselwirkungen zwischen Laktobazillen und
Gram-negativen Keimen festgestellt worden, die zur Dezimierung der Gram-negati-
ven Flora führten und damit die Belastung des Wirtes durch z.B. bakterielles Endo-
toxin verringerten (BENGMARK 1998).
Synbiotika sind Produkte, die Prä- und Probiotika kombiniert enthalten. Sie ver-
bessern das Überleben und die Kolonisationsfähigkeiten der zugeführten Keime im
MDT und steigern die Wachstumsrate und/oder die Stoffwechselrate einiger limitier-
ter gesundheitsfördernder Mikroorganismen (DELZENNE und ROBERFROID 1994).
2.3.5 Einfluss des Alters des Wirtes
Die Veränderungen der Darmflora im fortgeschrittenen Alter sind nur wenig unter-
sucht worden. Somit ist es ausgesprochen schwierig, den Einfluss abzuschätzen,
den das Alter des Wirtes auf die eigene Darmbakterien ausübt. GORBACH et al.
(1967) untersuchten einige Probanden mit dem Ergebnis, dass weniger Bifidobakte-
rien feststellbar waren und Pilze und Koliforme vermehrt auftraten. Allerdings war der
Probenumfang gering und die Streuung der Ergebnisse sehr groß, so dass diese
Befunde vorsichtig bewertet werden müssen. Die Änderungen der Mikroflora des
Kolon beim Wechsel vom Stillkind zum Mischköstler haben ihre Ursache wohl eher in
dem massiven Diätwechsel als im Altersunterschied (MATA et al. 1971).
Aktuelle Daten über die Kolonflora älterer Menschen sind bei KLEESSEN et al.
(1997) zu finden. Hier wurde die Wirkung von Inulin und Laktulose auf bei dieser
Altersgruppe verbreitete Darmträgheit untersucht und der Status der Fäkalflora vor
21
Literaturübersicht
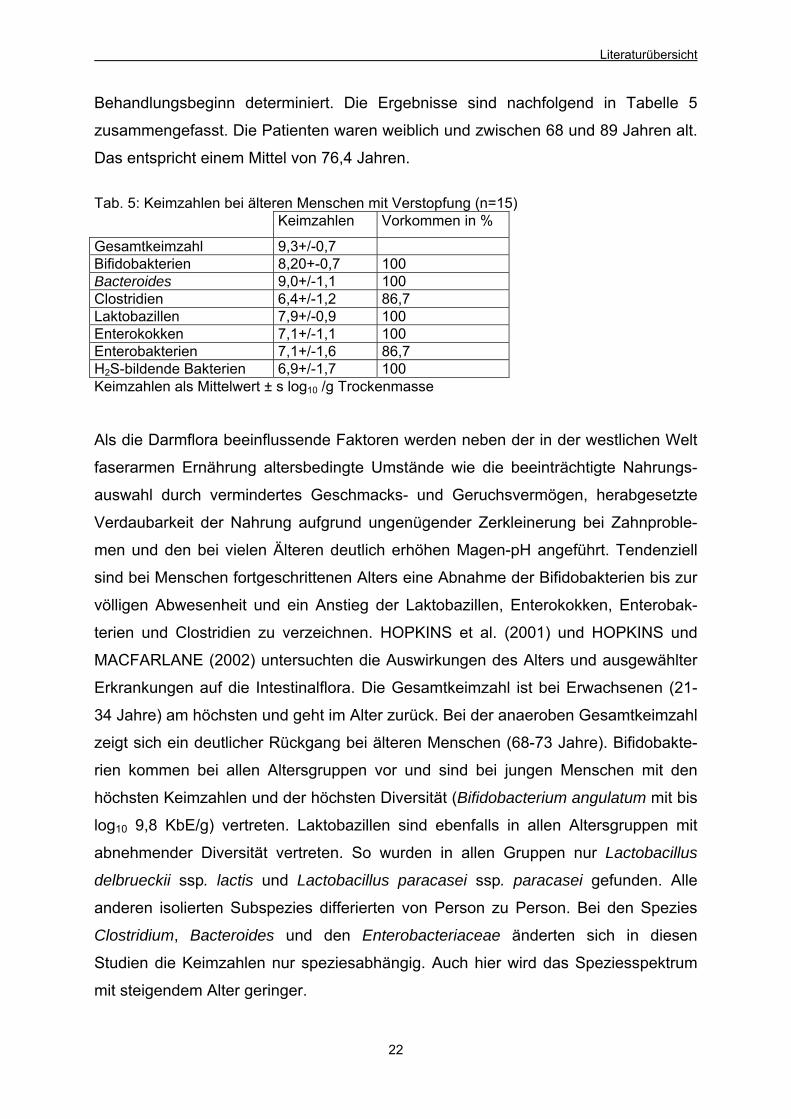
Behandlungsbeginn determiniert. Die Ergebnisse sind nachfolgend in Tabelle 5
zusammengefasst. Die Patienten waren weiblich und zwischen 68 und 89 Jahren alt.
Das entspricht einem Mittel von 76,4 Jahren.
Tab. 5: Keimzahlen bei älteren Menschen mit Verstopfung (n=15) Keimzahlen Vorkommen in %
Gesamtkeimzahl 9,3+/-0,7 Bifidobakterien 8,20+-0,7 100 Bacteroides 9,0+/-1,1 100 Clostridien 6,4+/-1,2 86,7 Laktobazillen 7,9+/-0,9 100 Enterokokken 7,1+/-1,1 100 Enterobakterien 7,1+/-1,6 86,7 H2S-bildende Bakterien 6,9+/-1,7 100 Keimzahlen als Mittelwert ± s log10 /g Trockenmasse
Als die Darmflora beeinflussende Faktoren werden neben der in der westlichen Welt
faserarmen Ernährung altersbedingte Umstände wie die beeinträchtigte Nahrungs-
auswahl durch vermindertes Geschmacks- und Geruchsvermögen, herabgesetzte
Verdaubarkeit der Nahrung aufgrund ungenügender Zerkleinerung bei Zahnproble-
men und den bei vielen Älteren deutlich erhöhen Magen-pH angeführt. Tendenziell
sind bei Menschen fortgeschrittenen Alters eine Abnahme der Bifidobakterien bis zur
völligen Abwesenheit und ein Anstieg der Laktobazillen, Enterokokken, Enterobak-
terien und Clostridien zu verzeichnen. HOPKINS et al. (2001) und HOPKINS und
MACFARLANE (2002) untersuchten die Auswirkungen des Alters und ausgewählter
Erkrankungen auf die Intestinalflora. Die Gesamtkeimzahl ist bei Erwachsenen (21-
34 Jahre) am höchsten und geht im Alter zurück. Bei der anaeroben Gesamtkeimzahl
zeigt sich ein deutlicher Rückgang bei älteren Menschen (68-73 Jahre). Bifidobakte-
rien kommen bei allen Altersgruppen vor und sind bei jungen Menschen mit den
höchsten Keimzahlen und der höchsten Diversität (Bifidobacterium angulatum mit bis
log10 9,8 KbE/g) vertreten. Laktobazillen sind ebenfalls in allen Altersgruppen mit
abnehmender Diversität vertreten. So wurden in allen Gruppen nur Lactobacillus
delbrueckii ssp. lactis und Lactobacillus paracasei ssp. paracasei gefunden. Alle
anderen isolierten Subspezies differierten von Person zu Person. Bei den Spezies
Clostridium, Bacteroides und den Enterobacteriaceae änderten sich in diesen
Studien die Keimzahlen nur speziesabhängig. Auch hier wird das Speziesspektrum
mit steigendem Alter geringer.
22

Literaturübersicht
2.3.6 Zusammenhang von Erkrankungen des Darmtraktes und der Mikroflora
Die Flora des Darmtraktes ist vielen Einflüssen ausgesetzt. Sie zeigt sich stabil
genug, um im gesunden Darm eindringende Bakterien abzuwehren. Bei Erkrankun-
gen des GIT, vor allem bei entzündlichen, ändern sich die Umgebungsbedingungen
wie z.B. die Durchblutung der Mukosa, Sekretion und Absorption. Damit wird die
Homöostase des Ökosystems Darm gestört. Dieses ermöglicht wiederum pathoge-
nen Bakterien die Kolonisation und führt zu entsprechenden Krankheitsbildern.
KLESSEN et al. (2002) untersuchten Mukosaproben von Patienten mit entzündlichen
Darmerkrankungen (IBD) mittels Fluoreszenz in situ-Hybridisierung im Vergleich zu
gesunden Personen. Eine Zunahme der Bakterien wurde festgestellt sowie eine Be-
siedlung der Submukosa bei Erkrankten. Das Keimspektrum änderte sich ebenfalls
deutlich in Abhängigkeit der einzelnen Erkrankungen. Auch bei Obstipation verschie-
denster Ätiologie verändert sich die Darmflora. SCHULZE (1992) berichtet über eine
Verringerung der Bifidobakterien-Keimzahl sowie der apathogener E. coli und der
Gesamtkeimzahl und dagegen über eine Zunahme der Bacteroides-Keimzahl.
Weiterhin kommt es durch die lange Verweildauer des Chymus durch Verringerung
der Peristaltik zur Akkumulation von mikrobiellen Enzymen und deren Abbaupro-
dukten (Phenole, Kresole, Ammoniak), denen z.T. kanzerogene Wirkungen zuge-
schrieben werden. Über eine deutliche Verminderung der Floradiversität bei an durch
Clostridium difficile verursachter Diarrhoe (CDAD) erkrankten Patienten berichteten
HOPKINS und MACFARLANE (2002). Außerdem unterschieden sich auch die Keim-
spektren. Es wurden v. a. Keime aus den Genera Lactococcus, Fusobacterium und
Actinomyces isoliert, die bei gesunden Erwachsenen nicht vorkamen.
2.3.7 Einfluss der Antibiose auf die Mikroflora Bis Anfang der fünfziger Jahre war es nicht bekannt, dass eine Therapie mit anti-
mikrobiellen Substanzen durch Störungen der Kolon-Mikroflora Diarrhöen und
Enteritiden auslösen kann. Eine Chemotherapie kann ebenfalls das Ökosystem im
Darm dramatisch stören. Die durch Antibiotika-Applikation mögliche Verminderung
der Kolonflora schwächt die Stabilität des Gleichgewichtes und prädisponiert den
Patienten für Infektionen mit pathogenen Keimen. Bei BRIEDIGKEIT (1992) wird von
23

Literaturübersicht
einer Überwucherung der Darmflora durch Clostridium difficile nach Antibiotika-
Therapie und anschließender Manifestierung einer schweren Enterokolitis gespro-
chen. Diese einer antimikrobiellen Therapie nachfolgenden Clostridium difficile-
Infektionen werden im Zusammenhang mit den verschiedensten Krankheitsbildern
erwähnt und bei KYNE et al. (2001) mit dem häufigen und unkritischen Gebrauch von
Antibiotika mit breitem Wirkspektrum in Verbindung gebracht. Ein überlegter Einsatz
der Antibiotika sowie ein Hygienesystem, das die Kontamination der Umwelt der
Erkrankten und Kreuzkontaminationen verhindert, wird von den Autoren zur Bekäm-
pfung nosokomialer Clostridium difficile-Erkrankungen empfohlen. Antimikrobielle
Wirkstoffe, die häufig vor dem Auftreten der Symptome einer Clostridium difficile-
Infektion angewendet wurden, sind Clindamycin, Ampicillin, Amoxycillin und Cephalo-
sporine, während bei der Bekämpfung dieser Erkrankung vor allem Metronidazol und
Vancomycin zum Einsatz gelangten (MYLONAKIS et al. 2001). Grundsätzlich sind
jedoch alle Antibiotika in der Lage, bei behandelten Patienten eine Clostridium diffi-
cile-Infektion hervorzurufen.
Die Applikation von Antibiotika hat nicht nur Einfluss auf die Zusammensetzung der
Darmflora, sondern auch auf die Ausbildung von Resistenzen indigener Bakterien
und Veränderungen charakteristischer physiologischer und biochemischer Eigen-
schaften des Ökosystems. In einer in-vitro Studie untersuchten BENDER et al.
(2001) mit Hilfe der Kolon-Simulationstechnik (Cositec) die Auswirkungen von
Vankomycin als Antibiotikum und Metronidazol als Chemotherapeutikum auf den
Stoffwechsel der Kolonflora bei Schweinen. In weiteren Modellen wurden eine
Störung des Kolonmilieus durch eine pH-Wert-Absenkung auf 5,1 ohne Einfluss anti-
mikrobieller Substanzen und der gleichzeitige Einfluss von Metronidazol und dem
sauren pH-Wert simuliert. Als Indikator für die Auswirkungen auf den Stoffwechsel
wurde die Bildung von Fettsäuren untersucht. Beide Wirkstoffe verursachten eine
signifikante Abnahme der kurzkettigen Fettsäuren (SCFA), was auf deutliche Stoff-
wechsel-Veränderungen schließen lässt. Niedrige Vankomycin-Konzentrationen
verminderten die Produktion von Acetat und Butyrat. Die Bildung von Propionat
wurde erst in höheren Konzentrationen von Vancomycin negativ beeinflusst. Metro-
nidazol unterband die Butyratbildung fast vollständig. Eine ähnliche Wirkung hatte die
pH-Wert -Absenkung ohne antimikrobielle Wirkstoffe, hier war besonders die Acetat-
bildung betroffen. Andere Autoren (MELLON et al. 2000) betonen vor allem die
Reduktion der Propionatbildung im Kolon durch oral appliziertes Metronidazol. Eine
24

Literaturübersicht
Verminderung der Propionatbildung um 43% innerhalb der ersten 24 Stunden und
die völlige Eliminierung dieser SCFA aus den Fäzes nach drei Wochen konnten fest-
gestellt werden und sind deutliche Hinweise auf Störungen des physiologischen
Kolonstoffwechsels.
SANDLER et al. (2000) beschäftigten sich weiterführend mit Auswirkungen einer
antimikrobiellen Therapie als mögliche Ursache des frühkindlichen Autismus. Durch
Berichte der Eltern betroffener Kinder kam zutage, dass den Symptomen des
Autismus häufig eine Behandlung mit Breitspektrum-Antibiotika, z.B. aufgrund einer
Mittelohrentzündung mit nachfolgender Diarrhoe, voranging. Daraus wurde die Hypo-
these entwickelt, dass diese Behandlung den Schutz der indigenen Flora vermindert
und dadurch Fremdkeime, hier vor allem Neurotoxinbildner, den geschädigten Darm
besiedeln.
Die Auswirkungen der einzelnen Gruppen antimikrobieller Wirkstoffe auf die Kolon-
flora sind verschieden. TAKESUE et al. (1999) untersuchten die Auswirkungen der
Antibiotika-Therapie bei Patienten mit postoperativer Infektion des Bauchraumes auf
die Kolonflora. Zur postoperativen Infektionsprophylaxe wurde bis drei Tage nach
dem Eingriff Cefazolin appliziert. Behandelt wurden auftretende Infektionen entweder
mit der Kombination Panipenem/Betamipron (Pp/Bm, n=5) oder Sulbactam/ Cefope-
razon (Sb/Cp, n=5). Die Stuhlproben der Patienten wurden vor der Behandlung
sowie am 4. und 7. Behandlungstag untersucht. Die anaerobe Gesamtkeimzahl ver-
minderte sich signifikant bei vier von fünf Patienten mit Sb/Cp-Therapie und die Zahl
der Bifidobakterien sank unter die Nachweisgrenze bei ebenfalls vier von fünf Patien-
ten dieser Gruppe. Die Behandlung mit Pp/Bm zeigte solche Auswirkungen nicht.
Sowohl die anaerobe Gesamtkeimzahl als auch die Bifidobakterien-Keimzahl blieben
relativ konstant. Die aerobe Gesamtkeimzahl änderte sich in beiden Gruppen nur
wenig. Allerdings wurden die Enterobacteriaceae weitestgehend durch Enterokokken
oder Candida spp. bei allen Patienten der Sb/Cp-Gruppe und bei einem Patienten
der Pp/Bm-Gruppe ersetzt. Die Autoren beenden ihre Ausführungen mit der Auffor-
derung an die Kliniker, diese Veränderungen und deren mögliche Auswirkungen vor
dem Therapiebeginn zu bedenken.
25

Literaturübersicht
2.4 Interaktionen zwischen Darmflora und Makroorganismus 2.4.1 Metabolismus der Mikroorganismen und des Wirtes sowie
gegenseitige Beeinflussung Der Dickdarm des Menschen wurde in der Literatur auch als kontinuierlich arbeiten-
der, anaerober Fermenter mit einem Volumen von ca. 500 ml und ca. 220 g Inhalt
beschrieben (CUMMINGS et al. 1990). Diese Bezeichnung ist sicher stark verein-
fachend. Sowohl die extreme Vielfalt der Bakterienspezies und die zum Teil weder
erkannten noch erforschten Wechselwirkungen innerhalb des Systems als auch der
Einfluss der Epithelialzellen durch Sekretion und Absorption auf das Ökosystem des
Kolons erschweren die Untersuchungen in vivo. Außerdem sind viele Auswirkungen
der Ernährung des Wirtes und des Immunsystems auf die Ökologie des Darms
unbekannt. Allgemein bekannt ist die Abhängigkeit der Struktur und Funktion der
Mikroflora von Umweltbedingungen wie Nahrungsangebot, pH, Redoxpotential und
mikrobiellen Interaktionen sowie Interaktionen mit dem Wirt.
In der jüngeren Vergangenheit wurde mehr und mehr dazu übergegangen, die bloße
Zählung und Bestimmung der kultivierten Bakterien durch die Untersuchung des
Zusammenwirkens zu ergänzen. Ziel war und ist es, Reaktionen im Ökosystem Dick-
darm auf bestimmte Einflüsse feststellen und die entsprechenden Entwicklungen
vorhersagen zu können. Die Untersuchung dieser Funktionszustände gelingt am
besten durch die Analyse bereits abgelaufener Prozesse.
Dabei ist es von Vorteil, die Mechanismen grob einzuteilen (MIDTVEDT 1989). Es
wird unterteilt in wirtsgebundene und den Mikroorganismen zuzuordnende Einflüsse.
SAVAGE (1977b) benennt die Ernährung des Wirtes als eigenständigen und hier
dritten Faktor. Die wirtsgebundenen Mechanismen wurden intensiv untersucht in
Studien mit keimfreien Tieren und normal besiedelten Kontrolltieren. Im Vergleich
von keimfreien Mäusen mit konventionellen Tieren erkennt BERG (1996) zahlreiche
Unterschiede in morphologischen, physiologischen, biochemischen und immuno-
logischen Merkmalen. Im Darm der keimfreien Mäusen konnte er beispielsweise eine
dünnere Lamina propria, verminderte Epithelzellerneuerung, erhöhte Transitzeit,
kleinere Peyersche Plaques und die verminderte Größe verschiedener Organe
feststellen.
Eine Energiequelle für die Mikroflora ist die Nahrung des Wirtes. Kolonbakterien
verwerten eine Vielzahl der aufgenommenen Bestandteile und erweitern durch z.B.
26

Literaturübersicht
Zerlegen von Komponenten und Abgabe von eigenen Stoffwechselprodukten das für
andere Bakterienspezies verfügbare Nahrungsspektrum erheblich (PRINS 1977).
Weiterhin werden Vitamine wie Vitamin B12 und Vitamin K synthetisiert. Zusätzlich
können Sekrete des Wirtes z.T. sehr schnell metabolisiert und als Energiequelle
genutzt werden (MACFARLANE et al. 1989a,b). Der Wirt sezerniert fermentierbare
Kohlenhydrate, z.B. Glykoproteine wie Mucin und Polysaccharide wie Chondroitin-
sulfate, die vor allem von Bifidobakterien, einigen Bacteroides spp. und Ruminokok-
ken verwendet werden können. Diese Kolonbakterien besitzen eine Glykosidase, die
die Glykoproteine abbaut und damit Zucker als Kohlenstoffquelle für andere Bakte-
rien bereitstellt (HOSKINS und BOULDING 1981). Verwertbare Membranglykopro-
teine werden von abgeschilferten Epithelzellen gewonnen. Von einigen Proteolyten
können Pankreasenzyme als Stickstoffquelle benutzt werden. In der Diskussion als
Nahrungsquelle steht auch die Mukusschicht, die das Dickdarmepithel bedeckt und
von spezialisierten Becherzellen sezerniert wird. Diese Schicht besteht überwiegend
aus Muzin, einem Glykoprotein von ca. 2000 kDa, sowie aus vielen weiteren kleinen
Proteinen, Glykoproteinen, Fetten und Glykolipiden. Diese Schicht dient zum einen
dem Schutz der Mukosa als Teil der antimikrobiellen Barriere (KLEESSEN et al.
2002). Bei Patienten mit Darmerkrankungen wie der IBD ist die Mukusschicht im
Kolon stark verringert, was die Exposition der Mukosa zu den Darmbakterien
verstärkt und was damit die Wahrscheinlichkeit der Besiedlung mit diesen Keimen
erhöht (KLEESSEN et al. 2002, MACFARLANE et al. 2000). Somit fungiert sie auch
als Trennschicht zwischen Darminhalt und den Bakterien, die die Darmmukosa
physiologischerweise in Mikrokolonien besiedeln. Dabei handelt es sich nach
MACFARLANE et al. 2000 um zahlreiche gram-positive und gram-negative Stäbchen
und Kokken, die 18 verschiedenen Gattungen zugeordnet werden konnten. 2.4.2 Immunologie des Darmes
Die Schleimhäute werden sofort nach der Geburt mit einer Vielzahl von Mikroorga-
nismen und einer breiten Spanne von Protein-Antigen konfrontiert. Das Immun-
system der Schleimhaut hat während der evolutionären Entwicklung zwei Hauptstra-
tegien zur Abwehr entwickelt (BRANDTZAEG 2002): Das sind zum einen der
Antigen-Ausschluß durch sIgA- und sIgM-Antikörper, die auch die Kolonisation von
eingebrachten Mikroorganismen regulieren und damit die Zusammensetzung der
27

Literaturübersicht
indigenen Mikroflora mitbestimmen. Des weiteren wird das Eindringen potentiell
gefährlicher löslicher Antigene aus dem Darmlumen verringert. Die zweite Abwehr-
strategie ist ein suppressiver Mechanismus, der Überempfindlichkeitsreaktionen auf
unschädliche Substanzen bei deren Kontakt mit der Schleimhaut vermeiden soll.
Diese Variante wird auch als „orale Toleranz“ bezeichnet, wenn mit der Nahrung
aufgenommenes, im Darm befindliches Antigen beteiligt ist (BISCHOFF et al. 2000).
Ähnliche Mechanismen wirken gegen Antigen aus der kommensalen Mikroflora
(HELGELAND et al. 2000).
Das Mukosa-assoziierte lymphatische Gewebe (MALT) besteht aus dem darm-
assoziierten lymphatischen Gewebe (GALT), den flächig ausgebreiteten lympha-
tischen Infiltrationen in der Lamina propria, dem Oberflächenepithel der Darm-
schleimhaut und zwischen den Epithelzellen gelegenen intraepithelialen Lympho-
zyten (DUCHMANN et al. 1999). Im GALT werden der Waldeyersche Rachenring mit
den Rachen- und Gaumenmandeln, der Blinddarm und spezielle Strukturen wie die
Peyer´schen Plaques zusammengefasst (BRANDTZAEG et al. 1999).
Zu Beginn einer normalen Immunantwort werden luminale Antigene über dünne
spezialisierte Mukosa-(M) Zellen mit Hilfe eines endozytotischen Prozesses trans-
epithelial aufgenommen. Durch die Lage des spezialisierten Epithels direkt über den
Peyer`schen Plaques erfolgt anschließend der erste Kontakt mit spezialisierten regu-
latorischen T-Zellen und antigenpräsentierenden Zellen (APC) wie B-Zellen und
follikuläre dentritische Zellen sowie die Einleitung einer mukosalen, auf die Produk-
tion von IgA und Immunneutralisation spezialisierten humoralen Immunantwort
(KELSALL et al. 1995). Ein Teil der aktivierten Lymphozyten tritt dann über die
mesenterialen Lymphknoten in die Zirkulation ein und kann von dort selektiv durch
Bindung an mukosale Adressine (BRISKIN et al. 1993) in die Lamina propria des
Darmes und andere mukosale Gewebe rezirkulieren („homing“). Studien an Lympho-
zyten der Lamina propria (LPL) und an IEL zeigen, dass sich beide Zellpopulationen
sowohl durch ihre unterschiedliche Lokalisation als auch durch zahlreiche phänoty-
pische und funktionelle Charakteristika unterscheiden (JAMES und ZEITZ 1994). LPL
ähneln in der phänotypischen Verteilung den Lymphozyten des peripheren Blutes. T-
Lymphozyten der Lamina propria (LP-T) sind in der überwiegenden Mehrzahl präakti-
vierte Zellen, die wichtige regulatorische und effektorische Funktionen wahrnehmen
(ZEITZ et al. 1991). Ihr besonderer Differenzierungszustand ist gekennzeichnet
durch die Expression eines festen Musters an Oberflächenmolekülen, die für die T-
28

Literaturübersicht
Zellaktivierung und deren Funktion von Bedeutung sind und einen höheren Grad der
Aktivierung im Vergleich zu zirkulierenden LP-T. Spezifisch sind weiterhin eine unter
definierten Bedingungen geringere Stimulierbarkeit über den T-Zellrezeptor und eine
vermehrte Fähigkeit, Zytokine zu synthetisieren und zu sezernieren. Intraepitheliale
Lymphozyten unterscheiden sich dagegen bezüglich ihrer phänotypischen
Zusammensetzung deutlich von den Lymphozyten des peripheren Blutes. So sind
IEL in der Mehrzahl CD8+ T-Zellen und exprimieren nahezu alle das αEß7-Integrin
(HML-1). Auch die Variabilität der auf der Zelloberfläche von IEL exprimierten
antigenspezifischen T-Zellrezeptoren ist im Gegensatz zu Lymphozyten des
peripheren Blutes oder Lymphozyten der intestinalen Lamina propria stark limitiert
(GROSS et al. 1994). Dies deutet darauf hin, dass IEL nur ein eingeschränktes
Spektrum von Antigenen erkennen. Die Herkunft und Funktion intraepithelialer T-
Lymphozyten beim Menschen ist noch nicht ausreichend geklärt. Es wird angenom-
men, dass die intraepithelialen T-Lymphozyten bei der Erstverteidigung gegen
luminales Antigen beteiligt sind und eng mit Epithelzellen interagieren.
Ein weiterer Abwehrmechanismus des Wirtes ist die Bereitstellung von Immunglobu-
lin A-Antikörpern (IgA) und IgM-Antikörpern (IgM). IgA und IgM werden von speziali-
sierten B-Zellen an Schleimhäuten gebildet, wo sie hauptsächlich wirksam sind.
Wichtigste Syntheseorgane sind vor allem der Darm, das respiratorische Epithel, die
laktierende Milchdrüse und andere exokrine Drüsen (GOLDMAN 1993). Auf die
Schleimhautoberflächen wird es als Dimer sezerniert, verbunden durch eine von den
Antikörpern synthetisierte J-Kette, die als „sekretorische Komponente“ bezeichnet
wird. Der Transport an die Schleimhautoberfläche wird von polymeren Ig-Rezeptoren
(pIgR) vermittelt. Nach dem Transport bleibt ein Fragment des pIgR mit dem Anti-
körper-Dimer in Verbindung und schützt es beispielsweise vor der Einwirkung proteo-
lytischer Enzyme im Darmlumen. Die so entstandenen sekretorischen Antikörper
sIgA und sIgM binden auf der Schleimhautoberfläche an Antigen und treten mit APC
in Kontakt. Die IgA-Antikörper bewirken einen Barriereeffekt, indem sie Viren, Toxine
und Bakterien am Anheften an die Epithelzelle durch Blockade der Adhäsionsrezep-
toren hindern. Ebenso schützt die IgA-Blockade vor der Absorption potentiell patho-
gener Substanzen.
Die Synthese des IgA wird durch Cytokine gesteuert. Von Antigen stimulierte T-
Zellen differenzieren vorwiegend zu CD4+ Helferzellen. Diese induzieren die Diffe-
renzierung von antigenspezifischen B-Zellen zu mehrheitlich IgA-bezogenen Plasma-
29

Literaturübersicht
zellen, wobei die CD4+-Helferzellen von DC und die Sekretion von Zytokinen wie den
Wachstumsfaktor TGF-β und das Interleukin 10 (IL-10) gesteigert wird (BRANDT-
ZAEG et al. 1999).
Die indigene Darmflora stimuliert die genannten Mechanismen der Immunabwehr
sowie die Mukussekretion und die Peristaltik (SAVAGE 1977b, FRETER 1983,
TANNOCK 1981) und schützt den Wirt durch bakteriellen Antagonismus. So kann
eine intakte indigene Darmflora die Kolonialisierung pathogener Keime verhindern,
z.B. durch Besetzen aller verfügbaren Habitate und durch Produktion antibakterieller
Substanzen. Beispielsweise siedelte sich Vibrio cholerae in größeren Mengen in
immunisierten keimfreien Tieren an als in den mit antagonistischen Bakterien besie-
delten Kontrollen (SHEDLOVSKY und FRETER 1974).
SHEDLOVSKY und FRETER (1974) stellten dar, dass die bakteriellen Antagonismen
mit der lokalen Immunantwort synergistisch gegen die Kolonisierung von Invasoren
wirken. Bei LEE (1985) wurden nach kritischer Bewertung der vorhandenen Literatur
keine Beweise für den Einfluss des Immunsystems auf Fluktuationen der Darmflora
gefunden. FRETER (1992) schlussfolgert ebenfalls, dass der Einfluss gering sein
muss, da es nur schwer vorstellbar sei, wie diese Mikroflora überhaupt existieren
könne, wenn sie vom Immunsystem beeinflusst würde. Möglicherweise spiele die
Immunantwort bei Erwerb und Entwicklung der Mikroflora eine Rolle (was mit den
Erkenntnissen von BRANTDZAEG (2002) übereinstimmt), aber sie habe wenig Effekt
auf die stabil entwickelte indigene Intestinalflora bei gesunden Erwachsenen.
Kurz nach der Geburt sind sowohl die Barrierefunktion der Schleimhaut als auch das
immunoregulatorische Netzwerk nur schwach ausgebildet. Deshalb ist diese Zeit
kritisch im Bezug auf die Schleimhautabwehr sowohl gegen Infektionen als auch bei
der Ausbildung von allergischen Erkrankungen. (HOLT et al. 2000). Der Zeitpunkt
des ersten Kontaktes mit dem jeweiligen Antigen (z.B. Kuhmilch) wie auch dessen
Dosis und vor allem die rasche Ausbildung einer physiologischen und stabilen indige-
nen Mikroflora beeinflussen die Ausbildung einer immunologischen Homöostase
maßgeblich (BRANDTZAEG et al. 1999).
30

Literaturübersicht
2.4.3 Kurzkettige Fettsäuren (SCFA) 2.4.3.1 Definition SCFA
Flüchtige oder volatile Fettsäuren sind kurzkettige Monokarbonsäuren. Die im Dick-
darm vorhandenen SCFA sind überwiegend bakteriellen Ursprungs. Sie werden von
mehreren Spezies unter Aufspaltung von fermentierbaren Kohlenhydraten und Pro-
teinen gebildet (CUMMINGS und MACFARLANE 1991) und geben einen Einblick in
die Stoffwechselleistungen der Mikroflora insgesamt. Der größte Teil der gebildeten
SCFA (90-95%) werden sehr schnell vor allem im Zäkum und aszendierenden Kolon
von den Mukosazellen absorbiert. Butyrat ist eine Hauptenergiequelle für die
Kolonozyten und wird von diesen zu Ketonkörpern und CO2 verstoffwechselt (BERG-
MANN 1990). Die anderen SCFA gelangen über die Pfortader in die Leber, wo sie
der Energiegewinnung oder Fettsynthese dienen. Propionat wird zu Glukose meta-
bolisiert und liefert damit dem Wirtsorganismus direkt nutzbare Energie. Weiterhin
können sie anderen Organen als Energielieferant zur Verfügung stehen (CUMMINGS
et al. 1991), wie z.B. Acetat in der Muskulatur (BERGMANN 1990, DELZENNE und
ROBERFROID 1994). So können volatile Fettsäuren bis zu 10% des menschlichen
Energiebedarfes abdecken (BERGMANN 1990). Dabei ist die SCFA-Produktion
direkt abhängig von der Art und der Menge aufgenommener Rohfaser. SCFA senken
im Kolon und damit auch in den Fäzes den pH-Wert und modifizieren dadurch die
Kolonflora. Unterstützt werden vor allem Bifidobakterien und andere säuretolerante
Spezies, die ihrerseits regulierend auf Bakterienspezies wie E.coli und Clostridium
einwirken (DELZENNE und ROBER-FROID 1994, SALMINEN und SALMINEN
1997). Dieser Effekt wurde vor allem nach Aufnahme von unverdaulichen Oligo- und
Polysacchariden beobachtet. Weiterhin haben SCFA einen protektiven Einfluss auf
die Darmmukosa (ROEDIGER 1980) und können darüber durch Regulierung der
Freisetzung von Entzündungsmediatoren bei entzündlichen Darmerkrankungen
heilend einwirken (SALMINEN und SALMINEN 1997, BENGMARK und JEPPSSON
1995). Damit haben die im Kolon gebildeten SCFA neben der Energieversorgung
des Wirtsorganismus eine große Bedeutung in der Abwehr pathogener und potentiell
pathogener Keime im Darm und bei der Aufrechterhaltung der Darmbarriere zur
Verhinderung der bakteriellen Translokation und systemischer Infektionen.
31

Literaturübersicht
Flüchtige Fettsäuren werden von vielen Bakterienspezies gebildet (Tabelle 6).
Tabelle 6: bakterielle Bildung und Metabolismus von SCFA, modifiziert nach Gibson (1999) SCFA an Bildung beteiligte Spezies Metabolismus Acetat : Bifidobakterien, Bacteroides, Clostridien, verstoffwechselt in Niere,
Eubakterien, Laktobazillen, anaerobe Muskulatur, Gehirn und Streptokokken, Veillonella, Fusobakterien, Herz Butyrovibrio u.a.
Propionat: Veillonella, Bacteroides, Propionibakterien Abbau in Leber, hemmt Cholesterolsynthese
Butyrat: Fusobakterien, Eubakterien, Clostridien, Energiequelle für Kolon- Butyrovibrio, Peptostreptokokken epithel , reguliert Zell-
wachstum und Differenzierung
Lokal wirken die drei Haupt-SCFAs Acetat, Propionat und Butyrat antidiarrhoisch
durch direkte oder indirekte Beteiligung an der Wasser- und Elektrolytresorbtion
(MIDTVEDT 1989). Acetat verbessert die Blutversorgung des Kolon und erhöht die
Motilität des Ileums (BERGMANN 1990). Butyrat ist eine bevorzugte Energiequelle
für die Kolonozyten und kann eine präventive Wirkung gegen verschiedene Arten
von Kolitis und Kanzerogenese entwickeln. Die in der westlichen Welt häufig auftre-
tenden Kolonerkrankungen sind oft eine Folge der hier üblichen stärke- und faser-
armen Ernährung. Diese führt zu einer geringeren SCFA-Produktion durch die
Kolonbakterien und damit einer Abnahme deren protektiven Einflusses (SCHEP-
PACH 1994).
FRETER et al. (1983) beschreiben Effekte von Fettsäuren auf die Verlängerung der
lag-Phase (= Latenzphase zwischen dem Einbringen der Keime in das Nährmedium
und Einsetzen der ersten Zellteilungen) von E. coli in anaerober Kultur. Diese Effekte
könnten zur Eliminierung eindringender nichtindigener Bakterien beitragen, indem
deren Vermehrungsrate verschoben wird. Die Teilung der Bakterien setzt später ein
und verläuft langsamer als ursprünglich, so dass sie ausgeschieden werden, bevor
sie sich vermehren konnten.
32

Literaturübersicht
2.5 Untersuchungsmöglichkeiten 2.5.1 Studienmodelle
Der Untersuchungsschwerpunkt wurde in älteren Studien vor allem auf die Analyse
der Florazusammensetzung und Zählung einzelner Mikroorganismen-Spezies gelegt,
während in heutigen Studien das Zusammenwirken und die Funktion der Bakterien
im Mittelpunkt stehen (MIDTVEDT 1989). Auch FRETER (1992) hebt das hervor. Die
im Stuhl gefundenen Bakterien müssen nicht dort angesiedelt sein, sondern können
aus höher gelegenen Darmabschnitten abgeschilfert und also transient sein. Von
diesen Bakterien geht keine nennenswerte Metabolitenproduktion aus, so dass das
Kolon – Gleichgewicht dadurch nicht beeinträchtigt wird. Weiterhin müssen im Lumen
gefundene Bakterien nicht dort vermehrungsfähig sein, sondern können inaktive
abgeschilferte Tochterzellen von Mukosa-assoziierten Populationen sein. Demzu-
folge sollten bei Untersuchungen nicht nur Keimzahl und Spezies, sondern auch die
Funktionen und Wechselwirkungen bestimmt werden.
Die Mikroflora des Kolon kann entweder außerhalb des Wirtes, in vitro, oder im
Makroorganismus, in vivo, untersucht werden. Im Tierversuch können Tiere mit
eigener oder assoziierter Flora besiedelt sein. In situ-Studien versuchen die aktuelle
Situation im Wirt zu fixieren und zu analysieren.
2.5.1.1 in situ-Untersuchungen
In Initialstudien wurden oft Fäkalproben statt Koloninhalt verwendet. Diese waren
leichter zu entnehmen, repräsentieren aber nur bedingt die Kolonflora.
Bei nichtinvasiven in situ-Studien kann die Darmflora recht einfach untersucht
werden, beispielsweise als Reaktion auf verschiedene Einflüsse wie bei Ernährungs-
umstellungen. Die hier gewonnenen Proben können ohne Verzögerung untersucht
werden. Ein weiterer Vorteil ist, dass die Proben von gesunden Probanden stammen,
die nicht vorbehandelt sind wie z.B. Chirurgie-Patienten, was die Magen-Darm-Flora
durchaus beeinflussen kann.
Sehr viel schwieriger gestaltet sich die Probengewinnung aus höher gelegenen
Darmabschnitten. Optimal wäre die direkte Entnahme, da sich bei Verwendung einer
Sonde die Bakterien dort anheften und somit verschleppt werden können. Die Ent-
nahme bei Unfallopfern oder bei eines natürlichen Todes Gestorbenen ist ethisch
33

Literaturübersicht
außerordentlich umstritten (CONWAY 1995). Die Materialgewinnung während einer
Operation ist dagegen relativ einfach möglich. Hier muss aber der Einfluss der
Narkose und einer eventuellen prophylaktischen bzw. therapeutischen antibiotischen
Vorbehandlung bedacht werden. Außerdem sind OP-Patienten in der Regel genüch-
tert (GORBACH 1967).
Für Untersuchungen am Tier sind nicht invasive und invasive in situ-Studien gleicher-
maßen geeignet und sehr viel einfacher durchführbar. Die Kotgewinnung ist vor allem
beim Großtier unproblematisch und für invasive Studien ist es speziell bei Schlacht-
vieh und entsprechenden Versuchstieren einfach, einzelne Darmabschnitte mit Inhalt
zu entnehmen oder den gesamten Darm zur Untersuchung zu erhalten.
2.5.1.2 Tierexperimentelle Untersuchungen
Keimfreie Tiere, bei BERG (1996) als „lebendes Teströhrchen“ bezeichnet, sind z.B.
für Untersuchungen geeignet, bei denen die Kolonisationsfähigkeit einzelner Bakter-
ien im Darm, Wechselwirkungen zwischen Bakterienspezies im Wirt und zwischen
Wirt und Flora und externe Einflüsse wie die Ernährung näher beleuchtet werden
soll. Diese Tiere werden steril durch Kaiserschnitt entwickelt, unter sterilen Bedin-
gungen gehalten und besitzen somit keine eigene Magen-Darm-Flora. Sie können für
Untersuchungen monoassoziiert, also mit einem bekannten Keim oder polyassoziiert,
d.h. mit mehreren bekannten Keimen besiedelt werden. Bei den erzielten Ergeb-
nissen muss aber stets beachtet werden, dass bei solcherart besiedelten Tieren die
Flora keinen natürlichen Bedingungen unterliegt und die festgestellten Reaktionen im
normal besiedelten Wirt nicht oder andersgeartet auftreten können.
HENTGES und FRETER (1962) berichten über Unterschiede bei Interaktionen von
Shigella flexneri mit verschiedenen Bakterien abhängig vom verwendeten in vitro-
System. Nur die CF-Kultur als einziges in vitro-System zeigte ähnliche bakterielle
Interaktionen wie die zuvor im Mausdarm beobachteten. Keimfreie Versuchstiere
können auch mit vollständigen Floren wie Humanflora assoziiert werden (KLEESSEN
et al. 2001). Die Handhabung der Tiere, die Überwachung z.B. der Ernährung und
die Probengewinnung sind hier deutlich einfacher als beim „Originalwirt“ Mensch.
34

Literaturübersicht
2.5.2 Nachweismethoden
Die Kultivierung ist die älteste Form des Bakteriennachweises und erfolgt durch
Anzüchtung lebensfähiger Zellen auf geeigneten Nährmedien und nachfolgender
Zählung und Identifizierung. Das Probenmaterial muss bei Darminhaltsproben in
jedem Fall schnellstmöglich in ein sauerstofffreies Transportmedium verbracht
werden. Transportzeit und -temperatur sind ebenfalls bestimmend für das Untersu-
chungsergebnis. Für die weitere Verarbeitung ist die Nutzung einer Anaerobier-
kammer nahezu unumgänglich. Das Arbeiten unter sauerstofffreien Bedingungen
minimiert die Verluste bei den sauerstoffempfindlichen Darmbakterien, auch Spezies
mit geringen Keimzahlen können kultiviert werden.
Mit der Verbesserung der anaeroben Nachweismethoden konnte die Magen-Darm-
Flora immer grundlegender analysiert werden. Damit ist die Untersuchung der
Zusammensetzung der Darmflora vor allem durch das Angebot geeigneter Medien
limitiert. Traditionell werden die Keime mittels Kulturmorphologie, biochemischen und
serologischen Eigenschaften identifiziert. Weiterführende Bestimmungen sind
molekularbiologisch möglich. FINEGOLD et al. (1983) konnten mittels Selektivmedi-
en und strikt anaerober Kultivierung ca. 10% der humanen Zäkumflora in vitro an-
züchten. Andere berichteten von der Kultivierung von 60% der mikroskopisch
gezählten Bakterien (HOLDEMAN et al. 1976).
Da sich nur relativ wenige Bakterienspezies unter z.T. enormem Aufwand kultivieren
lassen, sind Methoden, die ohne vorherige Kultivierung auskommen, von Vorteil.
Eine Möglichkeit, Bakterien in situ zu untersuchen, ist die Hybridisierung mit Oligo-
nucleotidsonden, die an ribosomale RNA der Bakterien binden. Damit können nicht
nur die Keimzahlen entsprechender Bakteriengruppen ermittelt werden, sondern die
Menge vorhandener rRNA liefert auch Informationen über die Wachstumsraten
einzelner Zellen. Ebenso geeignet ist die PCR als ein schnelles und genaues Verfah-
ren, das mit relativ wenig labortechnischem Aufwand betrieben werden kann (WANG
et al. 1996).
Bei KLEESSEN et al. (1999) wurden Proben aus unterschiedlichen Darmabschnitten
von Ratten untersucht. Dabei kamen vier verschiedenen Methoden zur Anwendung:
Die Kultivierung auf Nährböden, Fluoreszenz in situ-Hybridisierung (FISH) mit 16S
rRNA-Sonden, Färbung mit 4´,6-Diamidino-2-Phenylindol (DAPI) und die ebenfalls
auf Fluoreszenz basierende LIVE/DEAD® BacLight Viability Kit (L/D)-Färbung zur
35

Literaturübersicht
Zählung der lebenden und abgestorbenen Zellen. Mittels FISH und L/D wurden
ähnliche Zellzahlen ausgezählt. Nur 50% der mit DAPI gezählten Zellen wurden
durch FISH oder der Anfärbung lebender Zellen bestätigt. Die Färbung mit DAPI
kann für die Zählung aller vorhandenen Bakterien genutzt werden. Der Anteil der kul-
turell festgestellten Keimzahlen sinkt bei den gnotobiotischen Ratten auf 70 % und
bei den konventionellen Tieren auf 25 %. Diese Ergebnisse führen zu der Erkenntnis,
dass die Kultivierung auf Nährmedien ein unvollständiges Spektrum der wirklich
vorhandenen Bakterien liefert, während zur Zählung lebender Keime die FISH-
Technik und die L/D-Methode gute Alternativen sind.
2.6 Weitere Parameter
In diesem Abschnitt sollen die Parameter, die in die vorliegende Arbeit eingearbeitet
wurden, kurz vorgestellt werden.
2.6.1 Alkalische Phosphatase in Fäzes
Phosphatasen sind Enzyme, die die Hydrolyse von Phosphorsäure-Estern katalysie-
ren. „Alkalische Phosphatase“ wird so genannt, da ihr Aktivitätsoptimum im alkali-
schen pH-Bereich liegt. An Proteinen kann sie beispielsweise Phosphatreste von
phosphoryliertem Serin, Threonin oder Tyrosin abspalten.
Die fäkale AP ist kein einzelnes Enzym. Ihre Aktivität stellt die Summe individueller
Vorgänge der AP im Dünndarm, ggf. in der Plazenta und in Leber, Knochen und
Nieren dar, wobei postgenetische Formen die AP der Gallenwege und Tumor-Phos-
phatasen einschließen. Diese membrangebundenen Isoformen von Glykoproteinen
haben eine gemeinsame Proteinstruktur, unterscheiden sich aber im Kohlehydrat-
gehalt. Die AP-Aktivitätswerte in den Fäzes und im Serum werden vor allem zur
Diagnose von Leber- und Knochenerkrankungen genutzt (LEHMANN und ZENGEL
1980). Alle Isoformen der AP sind am Transport von Stoffwechselzwischenprodukten
durch die Zellmembranen beteiligt.
Die AP der Epithelzellen der Darmschleimhaut (intestinale AP) nimmt als Resorp-
tionsenzym eine wichtige Stellung ein. Eine Beteiligung an der Fettresorption ist
wahrscheinlich, da in Serum und Lymphe erhöhte AP-Spiegel nach fetthaltigen
Mahlzeiten beobachtet wurden (BENIC und FISHER-HERMANN 1970, ZHANG et al.
36

Literaturübersicht
1996). Eine weitere wichtige Funktion ist die Dephosphorylierung von bakteriellem
Endotoxin. POELSTRA et al. (1997) testeten in vivo Endotoxine von E. coli und
Salmonella Minnesota R595 mit dem Resultat, dass in beiden Fällen das jeweilige
Endotoxin durch die I-AP bei dem physiologischen pH von 7,5 dephosphoryliert
wurde. Weiterführend wurde Levamisol bei Ratten eingesetzt. Die einsetzende
Hemmung der I-AP-Aktivität führte zu einer signifikant reduzierten Überlebensrate
durch Septikämie von mit E.coli infizierten Ratten. Bei mit Staphylococcus aureus
infizierten Tieren gab es keinen Einfluss auf die Überlebensrate, was die I-AP-
Wirkung bei Infektionen mit endotoxin-produzierenden gramnegativen Bakterien
bestätigt und ihre Bedeutung beim Schutz des Wirtes verdeutlicht.
POSEN (1967) hielt es nach mehreren Untersuchungen für wahrscheinlich, dass die
AP in den Fäzes vorwiegend aus abgestoßenen Darmepithelzellen stammt, was
LEHMANN et al. (1981) am Rattenmodell bestätigten. Außer der alkalischen
Schleimhautphosphatase gelangen Phosphatasen aus der Galle und, durch Aus-
scheidung von Serumproteinen in den Darm, auch aus den Knochen, ggf. der
Plazenta u. a. Organen in den Darmkanal. Die Darmflora kann in vitro AP produ-
zieren (HORRIGAN und DANOVITCH 1984), wobei beweisende in-vivo-Studien
fehlen, da bisher keine Aktivitätsreduktion durch Antibiotika erreicht werden konnte.
Insgesamt muss daher die in den Fäzes gemessene AP-Aktivität als Gesamtaktivität
betrachtet werden. Eine Aufschlüsselung der Einzel-AP ist, wie bei BENIC und
FISHER-HERMANN (1970) beschrieben, z.B. durch Stärkegelelektrophorese
möglich. Dieselben Autoren berichten auch über den wahrscheinlichen Abbau
zumindest einzelner AP-Isoenzyme. Sie errechneten, dass die bei einem angenom-
menen minimalen Eintrag über die Galle allein in den Darm gelangte AP höher ist als
die in den Fäzes gemessene Gesamt-AP. Untersuchungen von LEHMANN und
HILLERT (1980) zeigten, dass die I-AP in den Fäzes stabil bleibt. Die Autoren gingen
daher von keiner nennenswerten Degradierung der Aktivität der I-AP durch Proteo-
lyse etc. aus, so dass vom Abbau andere Isoenzyme betroffen sein müssen.
2.6.1.1 Normbereiche der AP-Aktivität
Normalwerte werden bei LEHMANN und ZENGEL (1980) für die I-AP mit durch-
schnittlich 5,32 Unit/g Stuhl mit Minimalwerten bei 0,75 Unit/g und Maximalwerten bei
38,21 Unit/g angegeben. LEHMANN et al. (1981) zeigten am Rattenmodell, dass die
37

Literaturübersicht
AP der Darmschleimhaut unterschiedliche Anteile an der fäkalen Gesamt-AP einneh-
men kann. Die Aktivität der I-AP der Ratten lag zwischen 9,07 bis 22,56 Unit/24h und
deren Anteil bei 58,1% bis 85,7% der fäkalen Gesamt-AP. Die Untersuchungen von
LEHMANN und HILLERT (1980) geben einen Normwert der I-AP bei Menschen mit
durchschnittlich 47,5 Unit/g und der fäkalen Gesamt-AP mit durchschnittlich 60,8
Unit/g Stuhl an.
2.6.1.2 Einflüsse auf die AP-Aktivität
Bei verschiedenen Erkrankungen der Herkunftsorgane erfahren die spezifischen AP
deutliche Aktivitätsänderungen. So berichteten LEHMANN und HILLERT (1980) von
einer Abnahme der Aktivität der I-AP im Stuhl von Patienten mit Sprue-Syndrom von
bis zu 76%, bei Patienten mit totaler Zottenatrophie ein Absinken auf durchschnittlich
5,3 Unit/g und mit partieller Zottenatrophie auf durchschnittlich 14,3 Unit/g. Weiter
führten die Autoren aus, dass symptomfreie Patienten eine stabile AP-Aktivität
zeigten, während Erkrankungsschübe die Aktivität schwanken ließen.
Die Autoren stellten weiterhin fest, dass die Ernährung keinen schnellen und direkten
Effekt auf die Aktivität der I-AP hat. Ausgenommen sind hier Nahrungsbestandteile
(z.B. Gluten bei sensitiven Personen), die toxisch auf die Darmschleimhaut wirken
und damit sekundär ein Absinken der Aktivität bewirken. Behandlungen mit Cytosta-
tika dagegen bewirken einen kurzfristigen Anstieg der intestinalen AP, was auf den
zellzerstörenden Effekt der Medikamente zurückzuführen ist (LEHMANN et al. 1980).
2.6.2 MRS-Flora
Die Vertreter der MRS-Flora gehören zu den laktatbildenden Keimen oder Milchsäu-
rebakterien. Diese werden als gram-positive Stäbchen und Kokken beschrieben, die
Milchsäure als wichtigstes oder einziges Gärungsprodukt herstellen. Als Hauptver-
treter werden u.a. Laktobazillen (v.a. im Dünndarm), Bifidobakterien, Streptokokken
und Enterokokken angeführt. Diese Keime stellen hohe Ansprüche an Kultivierungs-
medien und Wachstumsbedingungen. Laktobazillen und Enterokokken sind zwar
aerotolerant, Bifidobakterien dagegen wachsen nur strikt anaerob. Der Vitaminbedarf
der Laktatbildner ist sehr hoch, höher als der des Menschen (MADIGAN et al. 2001).
Als Energiequelle werden überwiegend Kohlenhydrate genutzt, so dass vor allem die
38

Literaturübersicht
im Kolon ansässigen Bifidobakterien, aber auch alle anderen Kohlenhydratspalter,
auf Zufuhr von im Dünndarm nicht gespaltenen und resorbierten Kohlenhydraten
angewiesen sind. Vor allem Laktobazillen sind ausgesprochen säuretolerant. Sie
wachsen noch bei einem pH von 4 und wurden häufig im Magen nachgewiesen. Alle
milchsäurebildenden Bakterien sind ausgesprochen selten pathogen, wenn sie über-
haupt als Krankheitserreger isoliert werden können.
Über die gesundheitsfördernden Eigenschaften der Laktobazillen und Bifidobakterien
liegen zahlreiche Publikationen vor (ORRHAGE und NORD 2000, RUSCH und
RUSCH 2001, ISHIBASHI und YAMAZAKI 2001). Ihr Einsatz als Probiotikum erfolgt
beispielsweise zur Stabilisierung bei starken Störungen der Darmflora nach Antibio-
tikatherapie, postoperativ oder nach langer ausschließlich parenteraler Ernährung
(Bennet et al. 1996). In vitro-Studien zeigen, dass pathogene Keime (z.B. Clostridium
botulinum und C. difficile) durch etablierte Laktobazillen-, Bifidobakterien- und
Enterokokkenstämme an der Kolonisierung gehindert werden können (OHKUSA et
al. 1995). LIEVIN et al. (2000) beschreiben ähnliche Vorgänge in vitro und in axeni-
schen Mäusen. Hier wird die Assoziation und Invasion lethaler Salmonella typhimu-
rium-Stämme durch vorangesiedelte Bifidobakterium-Stämme verhindert. Die ausge-
prägte Fähigkeit der Laktobazillen und Bifidobakterien zur Adhärenz an der Mucosa
führen zur Bildung einer Barriere gegen pathogene und potentiell pathogene Keime.
Die Synthese antimikrobieller Substanzen, die Nahrungs- und Lebensraum-
konkurrenz der im Darm etablierten Stämme und die Senkung des Milieu-pH-Wertes
durch die gebildete Milchsäure sind Grundlagen der darmprotektiven Wirkung dieser
Spezies.
2.6.3 Leptin
Leptin ist ein Hormon, das in den Körperfettzellen vermittelt durch das ob-Gen
synthetisiert wird. Verschiedene Tiermodelle wiesen zunächst auf einen eindeutigen
Zusammenhang zwischen Adipositas und Leptin hin. Die daraus resultierende
Hoffnung vieler Wissenschaftler, Adipositas und die zahlreichen Folgeerkrankungen
schnell heilen zu können, hat sich nicht erfüllt. Leptin ist neben seinen Wirkungen auf
Energieverbrauch und Körpergewicht in zahlreiche Stoffwechselvorgänge eingebun-
den. Es zeigt vielfältige Beziehungen zu neuroendokrinen Regelkreisen wie der HPA-
Achse, Pubertät, Fruchtbarkeit und Gravidität sowie der Schilddrüsenfunktion.
39

Literaturübersicht
Weiterhin ist Adipositas ein überaus komplexes Krankheitsbild, was die Klärung der
verschiedenen Wirkungen und Einflüsse des Leptin zusätzlich erschwert (UHLMANN
2002).
2.6.3.1 Wirkung auf das Körpergewicht
Bei ausreichender Sekretion beeinflusst Leptin das Körpergewicht über Gewichts-
reduktion durch Hypophagie, eine effizientere Glukoseverwertung und Erhöhung des
Energieverbrauches sowie Sympathikusstimulation (Glukosestoffwechsel). Es regu-
liert zentral als Sättigungssignal die Nahrungs- und damit auch die Energieaufnah-
me, inhibiert die Lipogenese und stimuliert in den Adipozyten die Lipolyse (FRÜH-
BECK et al. 1997). Es besteht eine starke positive Korrelation mit dem Körpermasse-
index BMI (SCHOLZ et al. 1996), dem Körpergewicht, der vorhandenen Fettgewebs-
masse und der Adipozytengröße, was nicht verwunderlich ist, da Leptin überwiegend
von Fettzellen synthetisiert wird (HAMILTON et al. 1995). Daher geht eine Gewichts-
reduktion in der Regel mit einer Absenkung des Plasmaleptinspiegels einher (CON-
SIDINE et al. 1996). Die Leptinfreisetzung kann durch Insulin, Glukose, Dexametha-
son/Cortisol, TNF und IL-1 stimuliert werden.
2.6.4 Body Mass Index (BMI)
Definiert wird der BMI als ein Parameter für die Gewichtsbeurteilung als Quotient aus
dem Körpergewicht in Kilogramm und der Körpergröße in Metern zum Quadrat. Als
normalgewichtig gelten Menschen mit einem BMI von 20-25 kg/m2. Darunterliegende
BMI sprechen für Untergewicht und Werte darüber in Abstufungen für leichtes bis
deutliches und ab einem BMI von 40 kg/m2 für sehr starkes Übergewicht.
2.6.5 Blutzucker und HbA1c
Bei der Blutzuckermessung wird der Glukose-Gehalt des Blutserums, der so genann-
te Blutzuckerspiegel, ermittelt und wird vor allem zur Erkennung und Kontrolle des
Diabetes mellitus durchgeführt. Die in dieser Arbeit angeführten Werte wurden mit
einem Glukosebelastungstest (OGTT) bestimmt und sind Nüchternwerte. Normale
40

Literaturübersicht
Blutzuckerwerte liegen bei Erwachsenen bei 3,3-5,0 mmol/l oder 1,0-1,2 g/l. Dauer-
haft höhere Werte zeigen Diabetes an.
Eine Überprüfung der gemessenen Blutzuckerwerte kann durch die Bestimmung des
HbA1c erfolgen. Dieser wird auch als „Zuckerhämoglobin“ und „Blutzuckergedächt-
nis“. Mit diesem Test wird der Anteil des glykosierten, an Zucker gebundenen
Blutfarbstoffs Hämoglobin ermittelt. Es werden mit diesem Test die durchschnittlichen
Blutzuckerwerte der letzten sechs bis zwölf Wochen ermittelt. Der Normalwert liegt
bei 4-6 %. Das ist der Anteil des glykosierten Hämoglobins zum Gesamthämoglobin.
2.6.6 Gesamtcholesterol, LDL, HDL und Triglyceride
Wie Eiweiße und Zucker sind auch Fette eine wichtige Nahrungsquelle für den Orga-
nismus, wobei sie mit 39 Kilojoule je Gramm (kJ/g) den höchsten Energiegehalt
besitzen. Durch ihren hohen Energiegehalt sind Fette auch der mengenmäßig
wichtigste Speicherstoff des Organismus. Der Körper kann auch andere Stoffe wie
Eiweiße und Zucker in Fette umwandeln und speichern.
Der Hauptanteil der Nahrungsfette, ungefähr 90 Prozent, besteht aus Triglyzeriden
(Neutralfetten). Andere Nahrungsfette sind Cholesterin, fettlösliche Vitamine und
Phospholipide.
Da Fette sehr schlecht wasserlöslich sind, werden sie im Blut mithilfe von Lipoprote-
inen transportiert. Das sind Eiweiße, die Fette binden und damit als Emulgator in
wässriger Umgebung fungieren. In dieser Arbeit werden die während der MIRA-
Studie gemessenen LDL- und HDL-Werte interpretiert. LDL transportiert Cholesterin
aus der Leber in andere Gewebe für den Aufbau anderer Strukturen (Vitamin D,
Geschlechtshormone u.v.a.). Es kann sich bei Überangebot an den Gefäßwänden
anlagern und wird für das Entstehen von Atherosklerose und nachfolgenden Erkran-
kungen wie dem Herzinfarkt mitverantwortlich gemacht. Das HDL transportiert nicht
benötigtes Cholesterin zur Leber zurück und scheint damit die Gefäßwände vor
Ablagerungen zu schützen.
Normalwerte sind für Cholesterin 2,8-7,8 mmol/l, für Triglyceride 0,5-11 mmol/l, für
LDL <3,4 mmol/l und für HDL bis 0,9 mmol/l.
41

Material und Methoden
3 Material und Methoden 3.1 MIRA-Konzept: Mit Intelligenz richtig abnehmen
Das MIRA-Konzept bietet die Möglichkeit, bestehende Gewichtsreduktions- Pro-
gramme auf Leitlinienkonformität und Praxiseignung zu prüfen. Weiterhin soll dieses
Konzept Qualitätskriterien für die Analyse und Bewertung von Adipositas-
therapieprogrammen bereitstellen.
Die in dieser Arbeit vorgestellte Studie wurde als ambulantes Adipositastherapiepro-
gramm durchgeführt. Der Erfolg dieser Programme wird an folgenden Kriterien ge-
messen: Mindestens 50% der Teilnehmer sollten nach einem Jahr mindestens 5%
Gewicht reduziert und mindestens 20% der Teilnehmer wenigstens 10% Gewicht
reduziert haben (intention-to-treat-Analyse). Weiterhin wird eine Verbesserung von
eng mit Adipositas verbundenen Risikofaktoren (Hypertonie, Diabetes mellitus) ge-
fordert und das Absetzen von Medikamenten bzw. die Dosisreduktion und damit die
Kostensenkung für die medikamentöse Therapie bei hämodynamischen und metabo-
lischen Zielkriterien erwartet. (Scholz et al. 2002)
3.1.1 MIRA-PreCon und MIRA-DGE
Diese Studien waren „prospektive, kontrollierte, monozentrische, nicht medikamentö-
se Interventionsstudien mit finanzieller Eigenbeteiligung der Teilnehmer“ (Scholz et
al. 2002), mit deren Hilfe ein kommerzielles (BCM-Programm©, PreCon GmbH & Co
KG) und ein nicht kommerzielles, vorrangig als „Hilfe zur Selbsthilfe“ entworfenes
Programm („Ich nehme ab“ der Deutschen Gesellschaft für Ernährung e.V.) auf die
Erfüllung der o. g. Qualitätskriterien geprüft werden sollte.
An dieser Studie beteiligt waren das Ernährungsberatungs- und Trainingszentrum
(ETZ) M.Scholz, Leipzig in Zusammenarbeit mit der Universität Leipzig.
3.1.2 Teilnehmer
Insgesamt wurden 116 Teilnehmer (97 Frauen, 51±10 Jahre, BMI 33,0±5,2 kg/m2; 19
Männer, 54±8 Jahre, BMI 33,2±6,2kg/ m2) mit mindestens einem kardiovaskulären
Risikofaktor (Hypertonie, Koronare Herzkrankheit, Apoplexie, Varikosis, Thrombose,
Embolie) in diese Studie eingeschlossen. Stuhlproben wurden von insgesamt 60
42

Material und Methoden
Teilnehmern untersucht. Davon entfielen 29 Frauen und 8 Männer auf die MIRA-
PreCon-Gruppe und 21 Frauen und 2 Männer auf die MIRA-DGE-Gruppe. Der
durchschnittliche BMI lag bei der MIRA-PreCon-Gruppe bei 32,4 kg/m2 und in der
MIRA-DGE-Gruppe bei 32,5 kg/m2. In der weiteren Auswertung wurden Männer und
Frauen getrennt betrachtet, um eventuelle geschlechtsspezifische Unterschiede her-
ausstellen zu können. Auf Vergleiche Männer vs. Frauen wurde aus statistischen
Gründen verzichtet, da die Probandenanzahl zu ungleich ausfiel.
3.2 Stuhlprobengewinnung und -verarbeitung
Stuhlproben wurden zu Beginn der Studie (Woche 0) sowie 12, 24, 52 Wochen nach
Studienbeginn untersucht. Sie wurden von den Teilnehmern selbst entnommen und
bei +8°C gekühlt im ETZ M.Scholz abgegeben. Zwischen Entnahme und Abgabe
lagen maximal 12 Stunden. In der Regel gaben die Teilnehmer ihre Probe im Laufe
des Vormittags des Entnahmetages ab.
Die Verarbeitung der Stuhlproben erfolgte, wenn möglich, frisch. In Einzelfällen wa-
ren Proben bis maximal 5 Tage eingefroren. Vergleichende Untersuchungen im
Rahmen dieser Studie haben gezeigt, dass das Einfrieren auf die zu untersuchenden
bakteriologischen Parameter keinen signifikanten Einfluss hatte.
3.2.1 Kulturelle Keimzählung
Die Stuhlproben wurden frisch oder aufgetaut im Verhältnis 1:10 mit phoshatgepuf-
ferter Kochsalzlösung (PBS = Phosphat Buffered Saline) versetzt und nach Zugabe
von 6 bis 10 Glasperlen mit Hilfe eines Vortexers (MS1 Minishaker, IKA®) bis zur
homogenen Verteilung des Probenmaterials gemischt. Diese 1:10 Verdünnung wur-
de als Stammlösung für weitere Verdünnungsschritte genutzt. Dafür wurden in einer
Mikrotiterplatte (96, F-Form, Fa. J. Peske oHG, Aindling-Arnhofen) Verdünnungsstu-
fen von 1:102 bis maximal 1:107 durch Vorlegen von 270µl PBS in jede benötigte Ka-
vität und Zugabe von 30µl der nächstniedrigeren Verdünnung hergestellt. Aus diesen
Verdünnungen wurden jeweils 10µl entnommen, auf Universal- und Selektivnährbo-
den (3.2.1.1.) getropft und mithilfe eines sterilen Glasstabes ausgestrichen. Die
Nährböden wurden bei 37ºC aerob bzw. anaerob inkubiert. Die Keimzählung fand, je
nach untersuchter Keimart, nach 18-24 h bzw. nach 42-48h statt. Es wurden alle
43

Material und Methoden
Keimzählungen als Doppelbestimmung durch-geführt. Die ermittelten Keimzahlen
beziehen sich auf ein Gramm Feuchtmasse. Die untere Nachweisgrenze dieser Me-
thode liegt bei 2,70 log10(KBE/g).
3.2.1.1 Untersuchte Bakterienpopulationen und verwendete Nährböden
Als Summenparameter wurden die aerobe Gesamtkeimzahl (aGKZ) und die aerobe
Gram-negative Keimzahl (aK-) untersucht. Die Bestimmung der aGKZ erfolgte auf
Nähragar I (Sifin, Berlin) und die aeroben gramnegativen Bakterien wurden auf Mac-
Conkey-Agar (Sifin, Berlin) bestimmt. Beide Nährböden wurden nach Aufbringen der
Proben bei 37ºC unter aeroben Bedingungen für 18-24 h bebrütet.
Die Keimzahl an Bifidobakterien, Laktobazillen und Enterokokken wurde auf MRS-
Agar [(de Man, Rogosa, Sharpe), Sifin, Berlin] ermittelt und wird im Folgenden als
MRS-Keimzahl bezeichnet. Dieser Nährboden wurde bei 37ºC unter anaeroben Be-
dingungen für 42-48 h bebrütet. Das anaerobe Milieu wurde mit Hilfe eines sauer-
stoffzehrenden Mediums (Anaerocult®, Merck, Darmstadt) hergestellt.
Für die Bestimmung der Clostridium perfringens – Keimzahlen wurde Schafblutagar
mit Zusatz von Polymyxin und Neomycin verwendet. Dazu wurde 1l Nähragar 1 (Si-
fin, Berlin) nach Vorschrift hergestellt. Nach dem Abkühlen auf 45-50ºC wurden 50ml
Schafblut, 20g Glukose, 200mg Neomycin sowie 100mg Polymyxin B zugegeben.
Diagnostisch abgesichert wurden die anaerob gewachsenen Kolonien durch Gram-
färbung und Kultivierung auf Eigelb-Lactose-Agar.
Die Bestimmung der Candida-spp.-Keimzahl erfolgte auf Sabouraud-4%-Glukose-
Agar (Sifin, Berlin) nach fünftägiger aeroben Bebrütung bei 30°C. Differenziert wur-
den die gewachsenen Kolonien nach Gramfärbung und durch Anzucht auf CHRO-
Magar Candida (MAST DIAGNOSTICA, Reinfeld).
3.3 Keimzählung mit Hilfe der Fluoreszenz in situ-Hybridisierung (FISH)
Hybridisierung ist die komplementäre Basenpaarung von zwei einzelsträngigen Nuc-
leinsäuren. Die Identifizierung von Bakterien mittels in situ-Hybridisierung kann ohne
vorherige Kultivierung der Bakterien erfolgen. Hierzu werden die Bakterienzellen mit
Ethanol oder Paraformaldehyd fixiert, so dass ihre Zellstruktur erhalten bleibt.
44

Material und Methoden
Gleichzeitig jedoch wird dadurch ihre Zellmembran durchlässig, so dass die Sonden
penetrieren können. Bei der in situ-Hybridisierung in dieser Arbeit wurde die 16S
ribosomale RNA (rRNA) als Zielmolekül genutzt.
3.3.1 Sonden für die Fluoreszenz in situ-Hybridisierung
Für den Nachweis von Bakterien in Fäzes mittels in situ-Hybridisierung wurden Oli-
gonucleotidsonden (Interaktiva, Ulm) eingesetzt, die in Tabelle 6 aufgeführt sind. Die
Sonden waren am 5´-Ende mit dem Fluoreszensfarbstoff Cy3 (Indo-carbocyanin)
markiert. Das Emissionsmaximum von Cy3 liegt bei 565 nm.
Tabelle 7: Liste der für FISH verwendeten Oligonucleotidsonden Sondenname Sequenz (5´-3´) Referenz EUB 338 5´-GCT GCC TCC CGT AGG AGT-3´ Amann et al. 1990 EUB 785 5´-CTA CCA GGG TAT CTA ATC C-3´ Giovannoni et al. 1988 EUB 927 5´-ACC GCT TGT GCG GGC CC-3´ Lee et al. 1993 EUB 1055 5´-CAC GAG CTG ACG ACA GCC AT-3´ Lee et al. 1993 EUB 1088 5´-GCT CGT TGC GGG ACT TAA CC-3´ Lee et al. 1993 BIF164 5´-CAT CCG GCA TTA CCA CCC-3´ Langendijk et al. 1995 LAB 158 5´-GGT ATT AGC AYC TGT TTC CA-3´ Harmsen et al. 1999
Die in obiger Tabelle aufgeführten 5 Eubacteria–Oligonucleotidsonden wurden nach
Kleessen et al. (1999) in äquimolaren Mengen gemischt und als EUB – Mix bezeich-
net.
3.3.2 Fixierung der Stuhlproben
Ausgang für die hier notwendige Fixierung ist die unter 2.1. erwähnte mit PBS herge-
stellte Probenverdünnung. Diese wurde bei 300 x g zentrifugiert und der Überstand
für die Fixierung genutzt. Fixiert wurde zum einen mit Paraformaldehyd 4% (PFA).
Dabei wurde ein Teil Überstand mit 3 Teilen PFA 4% versetzt, bei +4ºC für 3-4 h in-
kubiert, anschließend aliquotiert und bei -20ºC gelagert. Zum Anderen kam eiskaltes
(-20ºC) vergälltes Ethanol zum Einsatz. Hier wurde ein Teil der Probe mit einem Teil
Ethanol gemischt und ebenfalls bei -20ºC gelagert.
Herstellung der Paraformaldehydlösung (PFA): Zwei Gramm PFA (Fluka, Buchs,
Schweiz) wurden in auf 60ºC vorgewärmtes Aqua bidest gelöst. Dazu wurde trop-
fenweise 2M NaOH gegeben, bis die Lösung klar war. Anschließend erfolgte die Zu-
gabe von 16,60ml 3x PBS- Puffer. Nach dem Einstellen des pH-Wertes auf 7,2 mit
45

Material und Methoden
HCl wurde mit Aqua bidest auf 50ml aufgefüllt. Die so hergestellte Lösung wurde ste-
rilfiltriert und bei 4°C maximal 24h gelagert.
Die Probenverdünnung und der jeweilige Fixationsprozess bestimmen die untere
Nachweisgrenze der Methode. Sie liegt bei PFA–fixierten Proben bei der Ver-
dünnung 1:20 (das entspricht einer Keimzahl von 5,97 log10(KBE/g)) und bei ethanol-
fixierten Proben bei der Verdünnung 1:5 (das entspricht einer Keimzahl von 5,37
log10 KBE/g)).
3.3.3 FISH auf Objektträgern
Die Fluoreszenz in situ-Hybridisierung wurde auf teflonbeschichteten Objektträgern
mit 8 Vertiefungen, sog. Wells [(Durchmesser 6mm) (Carl Roth GmbH, Karlsruhe)]
durchgeführt. Diese wurden nach MADDOX et al. (1987) vorher silanisiert, um die
Adhäsion der Bakterien am Objektträger zu erhöhen. Bei der Silanisierung wurden
die Objektträger über Nacht in 70%igem Ethanol bei Raumtemperatur entfettet, an-
schließend 1h in 60ºC warmem Aqua bidest gewaschen, kurz mit Aqua bidest
(Raumtemperatur) gespült und über Nacht bei 60ºC getrocknet. Die getrockneten
Objektträger wurden 7sec in 2%iger APES-Lösung (3-Aminotriethoxysilane, Sigma
Chemicals, Taufkirchen) in 99%igem Methanol inkubiert, 5min in 99%igem Methanol
gewaschen, 5min in Aqua bidest (Raumtemperatur) gespült und bei 37ºC getrocknet.
Bis zur Verwendung können die Objektträger bei Raumtemperatur lichtgeschützt ge-
lagert werden.
Je ein Aliquot von 10µl der fixierten Bakterienzellen wurde in die Aussparungen der
Objektträger getropft und gleichmäßig verteilt. In der vorliegenden Arbeit wurde die
Gesamtkeimzahl in der 1:50 oder 1:100-fachen Verdünnung, die Laktobazillen-Zahl
in der unverdünnten Fixierung oder 1:5-fachen Verdünnung und die Zahl der Bifido-
bakterien in der 1:10-fachen Verdünnung gezählt. Ausgang für die genannten Ver-
dünnungsstufen ist die jeweilige fixierte Probenlösung (s. 3.3.2.). Die aufgetragenen
Proben wurden 20min bei 46ºC getrocknet. Anschließend wurden 10µl Lysozymlö-
sung [(Puffer: 100mM Tris-HCl, Serva, Heidelberg (pH 8.0) + 50mM EDTA, Roth
GmbH & Co, Karlsruhe; Lysozym: Sigma Chemicals, Taufkirchen)] auf die Proben
gegeben und nach 10-15min durch vorsichtiges Spülen mit Aqua bidest (Raumtem-
peratur) entfernt. Die Zugabe von Lysozym dient dazu, die Permeabilität der Bakteri-
enzellwände besonders grampositiver Bakterien zu erhöhen. Danach wurden die Ob-
46

Material und Methoden
jektträger jeweils 5min in 50%, 70% und 96%iges Ethanol dehydriert. Nach dem Luft-
trocknen der Objektträger wurde die Eubacteria-spezifische Sondenmischung EUB-
Mix, die Bifidobacterium-spezifische Sonde BIF 164 oder die Laktobazillen- und En-
terokokken-spezifische Sonde LAB 158 (1µl der fluoreszenzmarkierten Oligonucleo-
tidsonde in 10µl Hybridisierungspuffer (0,9M NaCl, 10mM Tris-HCl, 0,01% SDS, Ro-
che, Mannheim)) in die Aussparungen pipettiert. Die Hybridisierung fand über Nacht
bei der für die jeweiligen Sonde günstigsten Temperatur in einer „Feuchten Kammer“
statt. Hierfür wurden die Objektträger in Zentrifugenröhrchen (50ml, TPP, Trasadin-
gen, Schweiz) verbracht, die mit Hybridi-sierungspuffer getränktem Zellstoff versehen
wurden. In dieser „Feuchten Kammer“ wurden die Objektträger anschließend inku-
biert. Nach der Hybridisierung wurden die Objektträger 20min in Hybridisierungspuf-
fer gewaschen. Die Temperatur des Puffers lag 2ºC über der Hybridisierungstempe-
ratur. Anschließend wurden die Objektträger kurz in Aqua bidest (Raumtemperatur)
gespült und luftgetrocknet. Nach dem Einbetten mit Antifading-Medium wurden die
Objektträger mit einem Deckglas abgedeckt und versiegelt. In der vorliegenden Ar-
beit wurde für die Versiegelung farbloser Nagellack verwendet. Für das Antifading-
Medium wurden 1,15g DABCO (1,4-diazabicyclo(2,2,2)octane, Sigma Chemicals,
Taufkirchen) in 5ml 200mM Tris-HCl gelöst und mit 45ml Glycerin gemischt.
Die Objektträger sind sofort oder bei lichtgeschützter Lagerung (bei Raum-
temperatur) innerhalb der folgenden zwei Tage auszuwerten.
3.3.4 Ermittlung der Zellzahlen mittels Fluoreszenzmikroskopie
Für die Quantifizierung der Zellzahlen mittels Fluoreszenzmikroskopie wurde ein Axi-
oskop (C. Zeiss, Jena) eingesetzt. Die Auszählung erfolgte bei 100-facher Objektiv-
Vergrößerung mit Hilfe eines Zählokulars (10x10 Teilfelder = 1 Zählquadrat mit einer
Fläche von 0,0001mm2). Es wurden von jeder Probe jeweils 2 Verdünnungen vorbe-
reitet. Von jeder dieser Verdünnungen wurden jeweils 12 Zählquadrate ausgezählt.
Für die Berechnung der Zellzahlen wurde folgende Formel verwendet.:
GZ = ZZ x FV x Fvol x FM
GZ = Gesamtzellzahl (ml-1)
ZZ = durchschnittliche Zellzahl pro Zählfeld
FV = Verdünnungsfaktor der eingesetzten Probe
47

Material und Methoden
Fvol = Volumenfaktor (das pro ml auf dem Objektträger aufgetragene
Probevolumen)
FM = Mikroskopierfaktor (hier 2827,40)
FM = Fläche einer Aussparung auf dem Objektträger/Zählfeldfläche
= 28,274mm2/0,01mm2
= 2827,40.
3.4 Alkalische Phosphatase
Die Aktivität der Alkalischen Phosphatase (AP) im Stuhl wurde mittels Mikrotiterplat-
tentest (96, F-Form) bestimmt. Hierzu wurden die Proben zu 10% in 1M Diethanola-
min-Puffer (1M Diethanolamin, 0,2mg/ml NaN3; 0,5mM MgCl2 x 6H2O, pH9,8, Fluka,
Buchs, Schweiz) aufgenommen und mehrmals homogenisiert. Anschließend wurde
bei 7375xg für 10 min bei Raumtemperatur zentrifugiert und der klare Überstand für
die AP-Bestimmung verwendet. Ausgehend von dieser Verdünnung wurden weitere
Verdünnungsschritte durchgeführt. Die Proben der vorliegenden Arbeit wurden
überwiegend 1:10.000 verdünnt.
Als AP-Substrat wurde p-Nitrophenylphosphat (pNPP, Fluka, Buchs, Schweiz) be-
nutzt und zu 30mM in Diethanolamin-Puffer aufgenommen. Eine Einheit (Unit) ent-
sprach dem Umsatz von 1µmol pNPP pro Minute bei pH 9,8 und 37°C.
Die Messung wurde wie folgt durchgeführt: Je Kavität wurden 10µl der verdünnten
Proben einpipettiert und danach erfolgte die Zugabe von 100µl pNPP-Substrat
(30mM) je Kavität. Die Endkonzentration des pNPP betrug 15mM. Daraufhin wurde
sofort der Blank-Wert (E0) bei 405nm photometrisch mittels Mitrotiterplatten-
Photometer (Anthos ht III, anthos labtech instruments, Salzburg) gemessen und da-
nach der Messansatz bei 37°C für 30min inkubiert. Nach der Messung des Substrat-
umsatzes (E1) bei 405nm (Substratendprodukt: p-Nitrophenol) erfolgte die Berech-
nung der Extinktionsdifferenz der Messungen E1 und E0. Unter Beachtung der Ver-
dünnungsfaktoren, der Inkubationszeit (min) und des Substratumsatzes (µmol) wurde
die AP-Aktivität in U/g berechnet.
48

Material und Methoden
3.5 Fettsäurenbestimmung
Die Bestimmung der SCFA wurde freundlicherweise im Institut für Physiologische
Chemie der Veterinärmedizinischen Fakultät der Universität Leipzig von Herrn Gerald
Valentin durchgeführt. Zur Vorbereitung der Fettsäurenbestimmung wurde das Pro-
benmaterial im Verhältnis 1:10 mit Aqua bidest homogen vermischt und für 15 min
bei 12.500xg zentrifugiert. Vom klaren Überstand wurden 200 µl entnommen und mit
25 µl Isocarpronsäure (0,02 M) versetzt, hier als Standard verwendet. Weiterhin wur-
den 280 µl Perchlor-säure (0,36 M) zur Keimabtötung und Proteinfällung sowie 270
µl Natriumhydroxid-Lösung (1 M) zur Neutralisierung der Säuren zugesetzt. Nach
sorgfältigem Mischen wurden die Proben bei –80°C eingefroren. Die Lyophyllisierung
erfolgte über Nacht (Lyophylle Fa. Christ, Alpha 1-2). Der Rückstand wurde in 100 µl
Ameisensäure (5 M) und 400 µl Aceton gelöst, anschließend bei 12.500xg zentrifu-
giert und der klare Überstand für die Gaschromatographie verwendet. Zur Verwen-
dung kam das Gerät GC3900 der Fa. Varian Deutschland GmbH mit der Methode
3900_SCFA.mth. Die verwendete Säule war eine Kapillarsäule (Fused Silica FFAP-
CB) mit den Werten lD=0,32 mm, df=0,5 µm und l=25 m.
Kalibriert wurde mit dem Kalibrationsstandard Superco No.46975-U „Volatile Acid
Standard Mix“ C1-C7 je 10 mM.
3.6 weitere Parameter
Die Bestimmung der folgenden Parameter erfolgte freundlicherweise vom Ernäh-
rungsberatungs-und Trainingszentrum M.Scholz., Leipzig. Blutzucker, HbA1c, Ge-
samtcholesterol, HDL, LDL und Triglyceride wurden aus venös entnommenem Blut
mit dem Automaten „Cob MIRA plus“ der Firma ABX (Frankreich) bestimmt. Die Lep-
tinergebnisse wurden mittels Radio-Immuno-Assay (RIA) ermittelt.
3.7 Biostatistische Auswertung
Die vorliegenden Ergebnisse wurden wie folgt mit dem Statistikprogrammpaket
SPSS 11.5 (SPSS Software GmbH München) statistisch bearbeitet.
Die Prüfung auf Normalverteilung der Werte wurde mit dem SHAPIRO-WILKS-TEST
durchgeführt. Die Prüfung ergab, dass bei den Untersuchungsparametern Gram-
49

Material und Methoden
negative Gesamtkeimzahl (AK), Clostridien (Clost.), Gesamtkeimzahl Bifidobakterien,
Laktobazillen und Enterokokken (MRS), Alkalische Phosphatase-Aktivität (AP),
Keimzahl Hefen, Fluoreszenz-Keimzahlen Bifidobakterien (Bif) und Lactobacillen
(Lac) teilweise signifikante Abweichungen von der Normalverteilung auftraten. Für
die aerobe Gesamtkeimzahl (aGKZ) und der Fluoreszenz-Gesamtkeimzahl (Eub) lag
Normalverteilung vor. Es wurden für alle Parameter der arithmetischen Mittelwert (x)
und die Standardabweichung (±s) berechnet.
Für AK, Clost., MRS, AP, Hefen, Bif und Lab wurden verteilungsunabhängige Prüf-
verfahren angewendet. Die Signifikanzprüfungen dieser untersuchten Parameter zwi-
schen den 4 Entnahmezeitpunkten erfolgte mit dem FRIEDMANN TEST. Wurden
hier Signifikanzen ermittelt, so erfolgte ein Paarvergleich durch den WILCOXON-
TEST. Die Signifikanzprüfungen dieser Untersuchungsparameter zwischen den ein-
zelnen Gruppen (Kontrollgruppe=MIRA-DGE und Versuchsgruppe=MIRA-PreCon)
wurden mit dem parameterfreiem U-TEST nach MANN-WHITNEY durchgeführt.
Für die normalverteilten Untersuchungsparameter (Eub, aGKZ) wurde zwischen den
beiden Behandlungsgruppen mit dem LEVENE-TEST und dem T-TEST die Signifi-
kanzprüfungen durchgeführt. Der Vergleich zwischen dem Entnahmezeiten wurde
beim Vorliegen einer Normalverteilung für die Parameter Eub und aGKZ mit allge-
meinen linearen Varianzanalyse mit Messwiederholung und dem Zwischensubjekt-
faktor „Gruppe“ berechnet. Hier werden nach Wechselwirkungen zwischen Zeit und
Gruppe gesucht. Bei signifikanter PILLAI-SPUR dieser Varianzanalyse erfolgten die
Einzelvergleiche zwischen den 4 Entnahmezeiten mit dem gepaarten T-TEST und
einer Korrektur nach dem BONFERRONI-Verfahren
Zusammenhänge wurden mit dem Korrelationskoeffizienten nach SPEARMAN (rs )
geprüft.
50
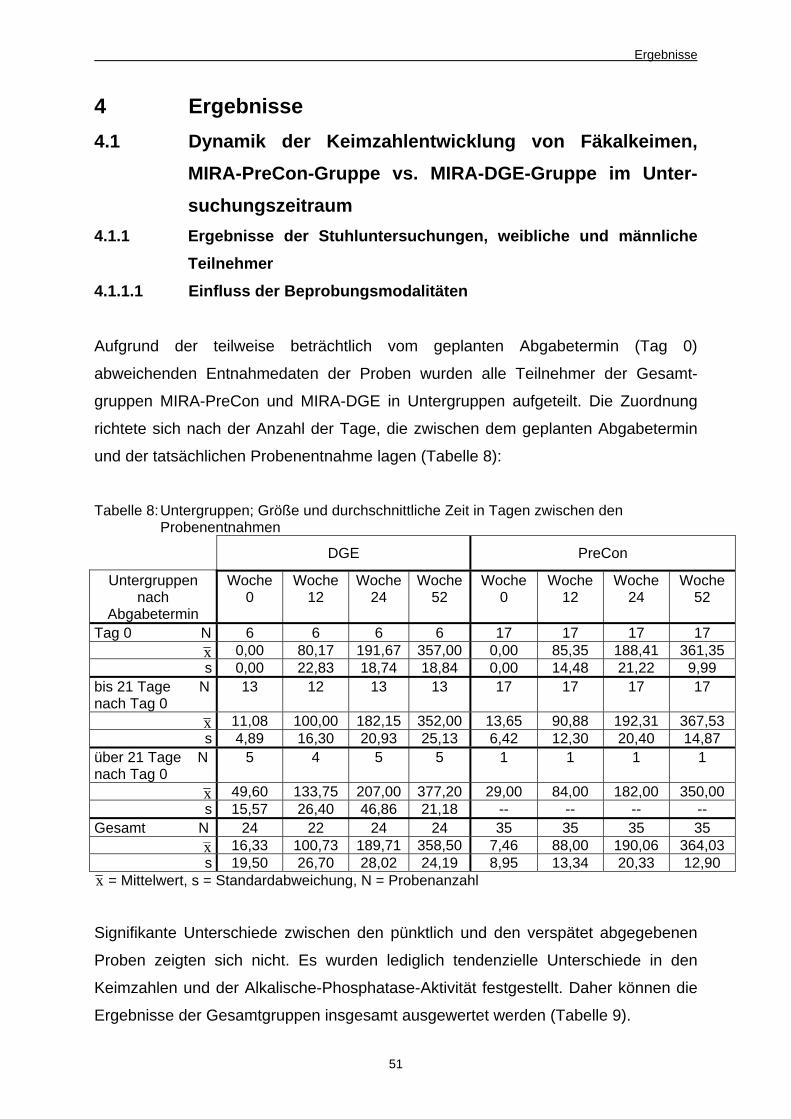
Ergebnisse
4 Ergebnisse 4.1 Dynamik der Keimzahlentwicklung von Fäkalkeimen,
MIRA-PreCon-Gruppe vs. MIRA-DGE-Gruppe im Unter-suchungszeitraum
4.1.1 Ergebnisse der Stuhluntersuchungen, weibliche und männliche Teilnehmer
4.1.1.1 Einfluss der Beprobungsmodalitäten
Aufgrund der teilweise beträchtlich vom geplanten Abgabetermin (Tag 0)
abweichenden Entnahmedaten der Proben wurden alle Teilnehmer der Gesamt-
gruppen MIRA-PreCon und MIRA-DGE in Untergruppen aufgeteilt. Die Zuordnung
richtete sich nach der Anzahl der Tage, die zwischen dem geplanten Abgabetermin
und der tatsächlichen Probenentnahme lagen (Tabelle 8):
Tabelle 8: Untergruppen; Größe und durchschnittliche Zeit in Tagen zwischen den Probenentnahmen
DGE PreCon
Untergruppen nach
Abgabetermin
Woche 0
Woche 12
Woche24
Woche 52
Woche 0
Woche 12
Woche 24
Woche 52
Tag 0 N 6 6 6 6 17 17 17 17 x 0,00 80,17 191,67 357,00 0,00 85,35 188,41 361,35 s 0,00 22,83 18,74 18,84 0,00 14,48 21,22 9,99
bis 21 Tage N nach Tag 0
13 12 13 13 17 17 17 17
x 11,08 100,00 182,15 352,00 13,65 90,88 192,31 367,53 s 4,89 16,30 20,93 25,13 6,42 12,30 20,40 14,87
über 21 Tage N nach Tag 0
5 4 5 5 1 1 1 1
x 49,60 133,75 207,00 377,20 29,00 84,00 182,00 350,00 s 15,57 26,40 46,86 21,18 -- -- -- --
Gesamt N 24 22 24 24 35 35 35 35 x 16,33 100,73 189,71 358,50 7,46 88,00 190,06 364,03 s 19,50 26,70 28,02 24,19 8,95 13,34 20,33 12,90
x = Mittelwert, s = Standardabweichung, N = Probenanzahl
Signifikante Unterschiede zwischen den pünktlich und den verspätet abgegebenen
Proben zeigten sich nicht. Es wurden lediglich tendenzielle Unterschiede in den
Keimzahlen und der Alkalische-Phosphatase-Aktivität festgestellt. Daher können die
Ergebnisse der Gesamtgruppen insgesamt ausgewertet werden (Tabelle 9).
51
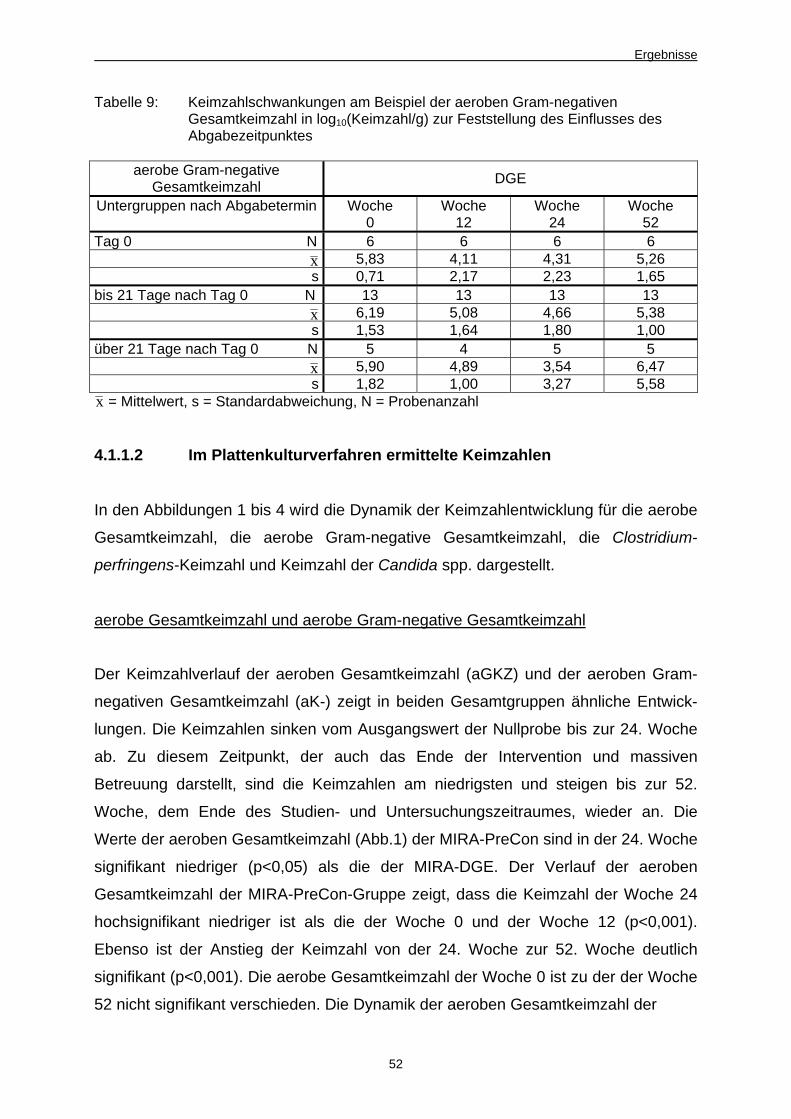
Ergebnisse
Tabelle 9: Keimzahlschwankungen am Beispiel der aeroben Gram-negativen Gesamtkeimzahl in log10(Keimzahl/g) zur Feststellung des Einflusses des Abgabezeitpunktes
aerobe Gram-negative
Gesamtkeimzahl DGE
Untergruppen nach Abgabetermin Woche 0
Woche 12
Woche 24
Woche 52
Tag 0 N 6 6 6 6 x 5,83 4,11 4,31 5,26 s 0,71 2,17 2,23 1,65
bis 21 Tage nach Tag 0 N 13 13 13 13 x 6,19 5,08 4,66 5,38 s 1,53 1,64 1,80 1,00
über 21 Tage nach Tag 0 N 5 4 5 5 x 5,90 4,89 3,54 6,47 s 1,82 1,00 3,27 5,58
x = Mittelwert, s = Standardabweichung, N = Probenanzahl
4.1.1.2 Im Plattenkulturverfahren ermittelte Keimzahlen
In den Abbildungen 1 bis 4 wird die Dynamik der Keimzahlentwicklung für die aerobe
Gesamtkeimzahl, die aerobe Gram-negative Gesamtkeimzahl, die Clostridium-
perfringens-Keimzahl und Keimzahl der Candida spp. dargestellt.
aerobe Gesamtkeimzahl und aerobe Gram-negative Gesamtkeimzahl
Der Keimzahlverlauf der aeroben Gesamtkeimzahl (aGKZ) und der aeroben Gram-
negativen Gesamtkeimzahl (aK-) zeigt in beiden Gesamtgruppen ähnliche Entwick-
lungen. Die Keimzahlen sinken vom Ausgangswert der Nullprobe bis zur 24. Woche
ab. Zu diesem Zeitpunkt, der auch das Ende der Intervention und massiven
Betreuung darstellt, sind die Keimzahlen am niedrigsten und steigen bis zur 52.
Woche, dem Ende des Studien- und Untersuchungszeitraumes, wieder an. Die
Werte der aeroben Gesamtkeimzahl (Abb.1) der MIRA-PreCon sind in der 24. Woche
signifikant niedriger (p<0,05) als die der MIRA-DGE. Der Verlauf der aeroben
Gesamtkeimzahl der MIRA-PreCon-Gruppe zeigt, dass die Keimzahl der Woche 24
hochsignifikant niedriger ist als die der Woche 0 und der Woche 12 (p<0,001).
Ebenso ist der Anstieg der Keimzahl von der 24. Woche zur 52. Woche deutlich
signifikant (p<0,001). Die aerobe Gesamtkeimzahl der Woche 0 ist zu der der Woche
52 nicht signifikant verschieden. Die Dynamik der aeroben Gesamtkeimzahl der
52
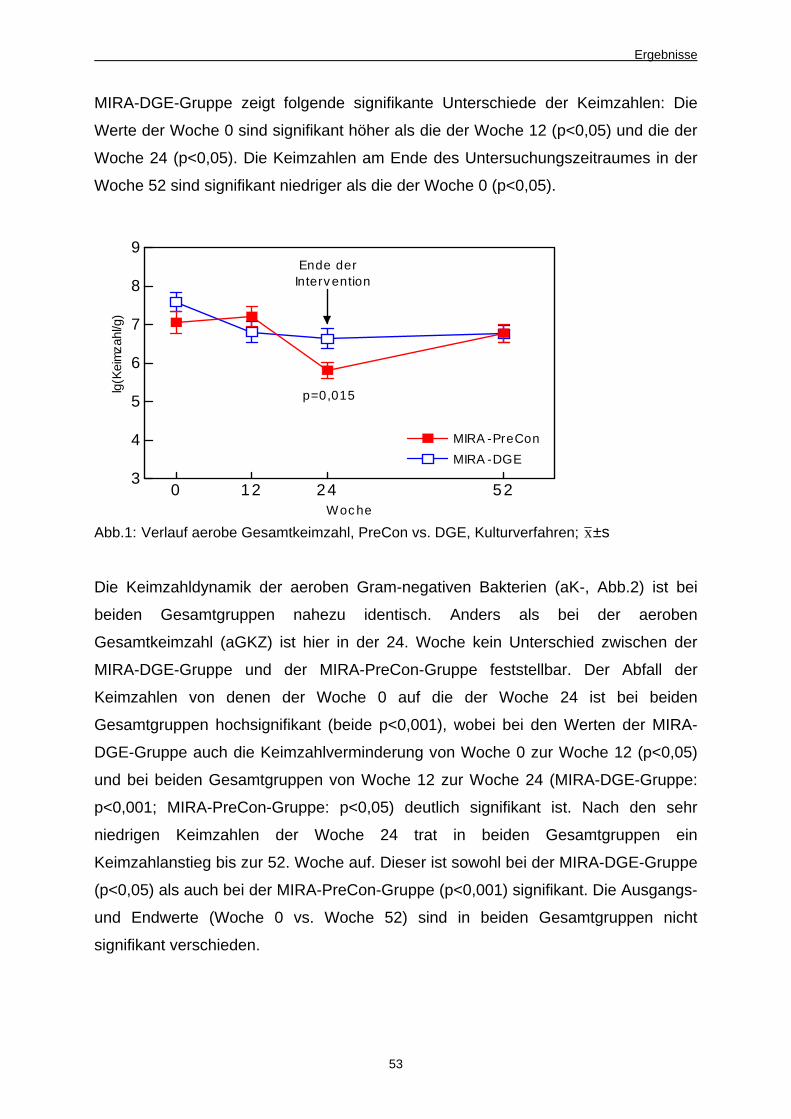
Ergebnisse
MIRA-DGE-Gruppe zeigt folgende signifikante Unterschiede der Keimzahlen: Die
Werte der Woche 0 sind signifikant höher als die der Woche 12 (p<0,05) und die der
Woche 24 (p<0,05). Die Keimzahlen am Ende des Untersuchungszeitraumes in der
Woche 52 sind signifikant niedriger als die der Woche 0 (p<0,05).
0 12 24 523
4
5
6
7
8
9
MIRA -PreConMIRA -DGE
Ende derInterv ention
p=0,015
Woc he
lg(K
eim
zahl
/g)
Abb.1: Verlauf aerobe Gesamtkeimzahl, PreCon vs. DGE, Kulturverfahren; x±s
Die Keimzahldynamik der aeroben Gram-negativen Bakterien (aK-, Abb.2) ist bei
beiden Gesamtgruppen nahezu identisch. Anders als bei der aeroben
Gesamtkeimzahl (aGKZ) ist hier in der 24. Woche kein Unterschied zwischen der
MIRA-DGE-Gruppe und der MIRA-PreCon-Gruppe feststellbar. Der Abfall der
Keimzahlen von denen der Woche 0 auf die der Woche 24 ist bei beiden
Gesamtgruppen hochsignifikant (beide p<0,001), wobei bei den Werten der MIRA-
DGE-Gruppe auch die Keimzahlverminderung von Woche 0 zur Woche 12 (p<0,05)
und bei beiden Gesamtgruppen von Woche 12 zur Woche 24 (MIRA-DGE-Gruppe:
p<0,001; MIRA-PreCon-Gruppe: p<0,05) deutlich signifikant ist. Nach den sehr
niedrigen Keimzahlen der Woche 24 trat in beiden Gesamtgruppen ein
Keimzahlanstieg bis zur 52. Woche auf. Dieser ist sowohl bei der MIRA-DGE-Gruppe
(p<0,05) als auch bei der MIRA-PreCon-Gruppe (p<0,001) signifikant. Die Ausgangs-
und Endwerte (Woche 0 vs. Woche 52) sind in beiden Gesamtgruppen nicht
signifikant verschieden.
53
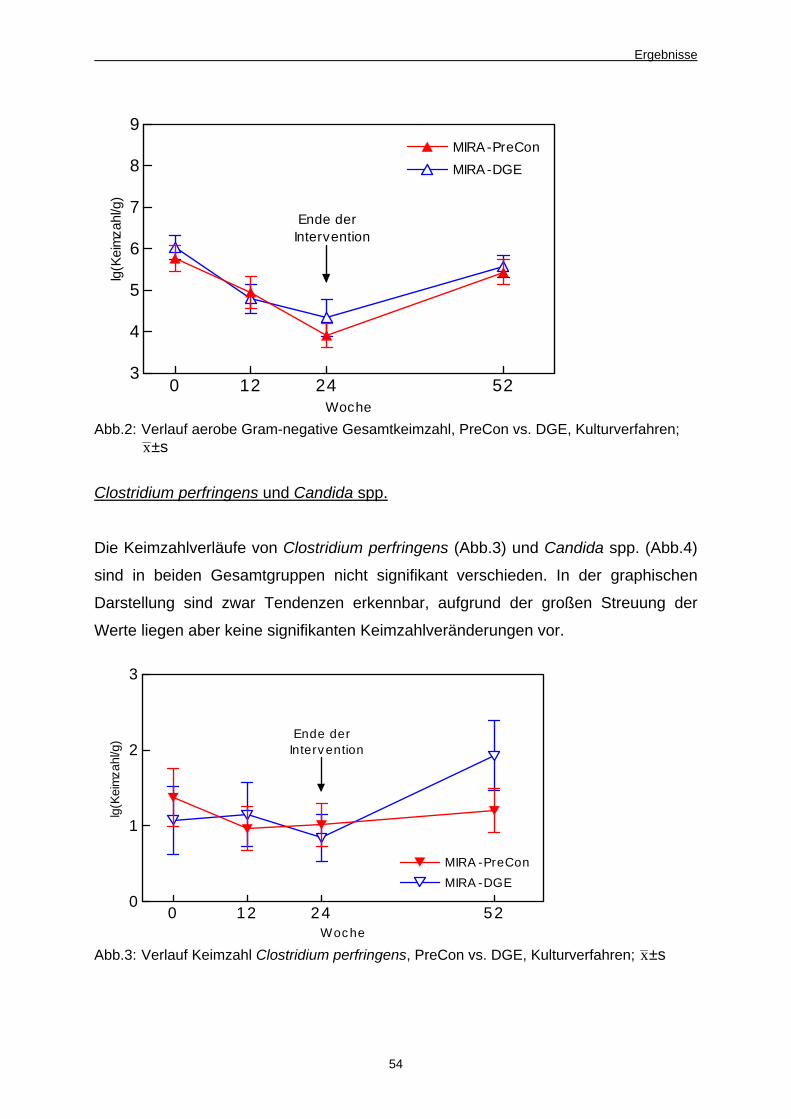
Ergebnisse
0 12 24 523
4
5
6
7
8
9MIRA-PreConMIRA-DGE
Ende derIntervention
Woche
lg(K
eim
zahl
/g)
Abb.2: Verlauf aerobe Gram-negative Gesamtkeimzahl, PreCon vs. DGE, Kulturverfahren; x±s
Clostridium perfringens und Candida spp.
Die Keimzahlverläufe von Clostridium perfringens (Abb.3) und Candida spp. (Abb.4)
sind in beiden Gesamtgruppen nicht signifikant verschieden. In der graphischen
Darstellung sind zwar Tendenzen erkennbar, aufgrund der großen Streuung der
Werte liegen aber keine signifikanten Keimzahlveränderungen vor.
0 12 24 520
1
2
3
MIRA -PreConMIRA -DGE
Ende derInterv ention
Woche
lg(K
eim
zahl
/g)
Abb.3: Verlauf Keimzahl Clostridium perfringens, PreCon vs. DGE, Kulturverfahren; x±s
54
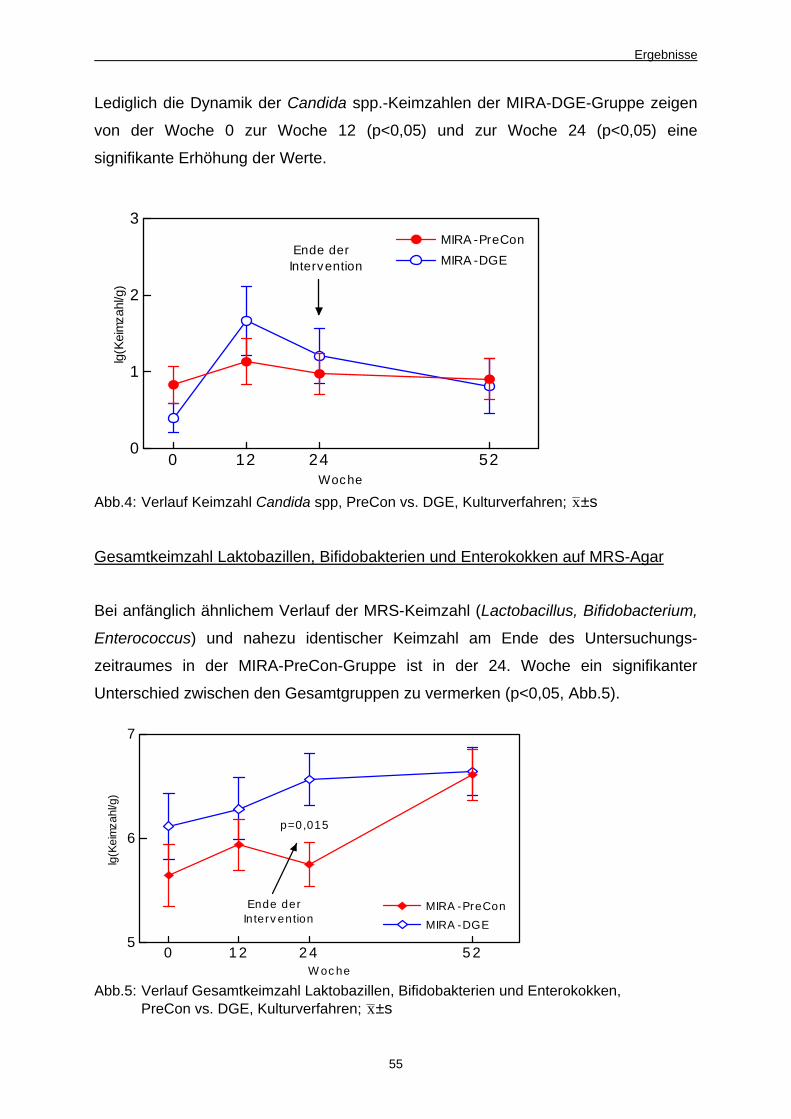
Ergebnisse
Lediglich die Dynamik der Candida spp.-Keimzahlen der MIRA-DGE-Gruppe zeigen
von der Woche 0 zur Woche 12 (p<0,05) und zur Woche 24 (p<0,05) eine
signifikante Erhöhung der Werte.
0 12 24 520
1
2
3MIRA -PreConMIRA -DGE
Ende derIntervention
Woche
lg(K
eim
zahl
/g)
Abb.4: Verlauf Keimzahl Candida spp, PreCon vs. DGE, Kulturverfahren; x±s
Gesamtkeimzahl Laktobazillen, Bifidobakterien und Enterokokken auf MRS-Agar
Bei anfänglich ähnlichem Verlauf der MRS-Keimzahl (Lactobacillus, Bifidobacterium,
Enterococcus) und nahezu identischer Keimzahl am Ende des Untersuchungs-
zeitraumes in der MIRA-PreCon-Gruppe ist in der 24. Woche ein signifikanter
Unterschied zwischen den Gesamtgruppen zu vermerken (p<0,05, Abb.5).
0 1 2 2 4 5 25
6
7
MIRA -PreConMIRA -DGE
Ende derInterv ention
p=0,015
W oc he
lg(K
eim
zahl
/g)
Abb.5: Verlauf Gesamtkeimzahl Laktobazillen, Bifidobakterien und Enterokokken, PreCon vs. DGE, Kulturverfahren; x±s
55

Ergebnisse
Der MRS-Keimzahlverlauf in der MIRA-DGE-Gruppe ist geprägt durch einen
signifikanten Anstieg der Werte. So ist der Wert der 52. Woche signifikant höher als
die Werte der Wochen 0 (p<0,05), 12 (p<0,05) und 24 (p<0,05).
Folgende Korrelationskoeffizienten wurden berechnet:
1) MIRA-PreCon: aGKZ/aK-: r=0,583**
aGKZ/MRS: r=0,422**
2) MIRA-DGE: aGKZ/aK-: r=0,597**
aGKZ/MRS: r=0,554**
(** = p<0,01, * = p<0,05)
4.1.2 Ergebnisse der Stuhluntersuchungen, nur weibliche Teilnehmer 4.1.2.1 Im Plattenkulturverfahren ermittelte Keimzahlen aerobe Gesamtkeimzahl und aerobe Gram-negative Gesamtkeimzahl
Folgende signifikante Unterschiede wurden berechnet: Die aerobe Gesamtkeimzahl
war bei den Teilnehmerinnen der MIRA-DGE-Gruppe in der Woche 24 signifikant
höher als bei den Teilnehmerinnen der MIRA-PreCon-Gruppe (p<0,05). Bei den
Teilnehmerinnen der MIRA-DGE-Gruppe ist der Keimzahlabfall von Woche 0 zur
Woche 12 (p<0,05) und zur Woche 24 (p<0,05) wie auch der Vergleich der Anfangs-
gegen die Endwerte (p<0,05) signifikant. Die Endwerte der Woche 52 liegen
signifikant niedriger als die der Woche 0. Die Werte der aeroben Gram-negativen
Gesamtkeimzahl sind zwischen den Unter-gruppen nicht signifikant verschieden. Der
Keimzahlabfall ist bei den Teilnehmerinnen der MIRA-DGE-Gruppe von den
Ausgangswerten der Woche 0 zu denen der Woche 12 (p<0,05) und der Woche 24
(p<0,001) signifikant. Die Keimzahlen der Woche 52 sind signifikant höher als die der
Woche 12 (p<0,05) und der Woche 24 (p<0,05). Die Keimzahlunterschiede der
Woche 0 gegen Woche 52 sind nicht signifikant.
56

Ergebnisse
Clostridium perfringens und Candida spp.
Die Keimzahlen Clostridium perfringens und Candida spp. unterscheiden sich nicht
zwischen den weiblichen DGE- und PreCon-Teilnehmern. Im Keimzahlverlauf
innerhalb der Teilnehmergruppen treten keine signifikanten Unterschiede auf.
Laktobazillen, Bifidobakterien und Enterokokken
Bei den Keimzahlen der Laktobazillen, Bifidobakterien und Enterokokken ist der Wert
der Woche 24 bei den Teilnehmerinnen der MIRA-DGE-Gruppe signifikant höher als
der Wert der MIRA-PreCon-Teilnehmerinnen (p<0,05). Während die Werte der
Teilnehmerinnen der MIRA-DGE-Gruppe im zeitlichen Verlauf keine signifikanten
Unterschiede aufweisen, ist bei denen der MIRA-PreCon-Gruppe der 52-Wochen-
Wert signifikant höher als der Wert der Woche 0 (p<0,05), der Woche 12 (p<0,05)
und auch der Woche 24 (p<0,05).
4.1.3 Ergebnisse der Differenzierung der MRS-Gesamtkeimzahl 4.1.3.1 Keimzählung auf Spezialnährboden
Auf MRS-Agar wurde eine Gesamtkeimzahl der hier untersuchten
laktatproduzierenden Bakterien ermittelt. Als eine Möglichkeit der Differenzierung der
Organismen kam die Anzüchtung auf einem Spezialnährboden in Betracht, auf dem
die einzelnen Gattungen durch spezifische Färbung unterschieden werden konnten
(„bifidobacteria agar“ nach Noack J. et al, 1998). Abgesichert wurde die Diagnose
durch eine Gramfärbung der jeweiligen Kolonien und durch Vergleich mit der
Koloniemorphologie entsprechender Referenzstämme (Lactobacillus acidophilus,
DSMZ 20079T; Bifidobacterium longum, DSMZ 20219T; Bifidobacterium adolescentis,
DSMZ 20083T; Enterococcus faecalis, DSMZ 20478T).
Die prozentualen Anteile liegen bei den Laktobazillen bei 27,7%, den Bifidobakterien
bei 55.5% und den Enterokokken bei 16.8%. Die Absolutwerte sind in Tabelle 10
dargestellt.
57
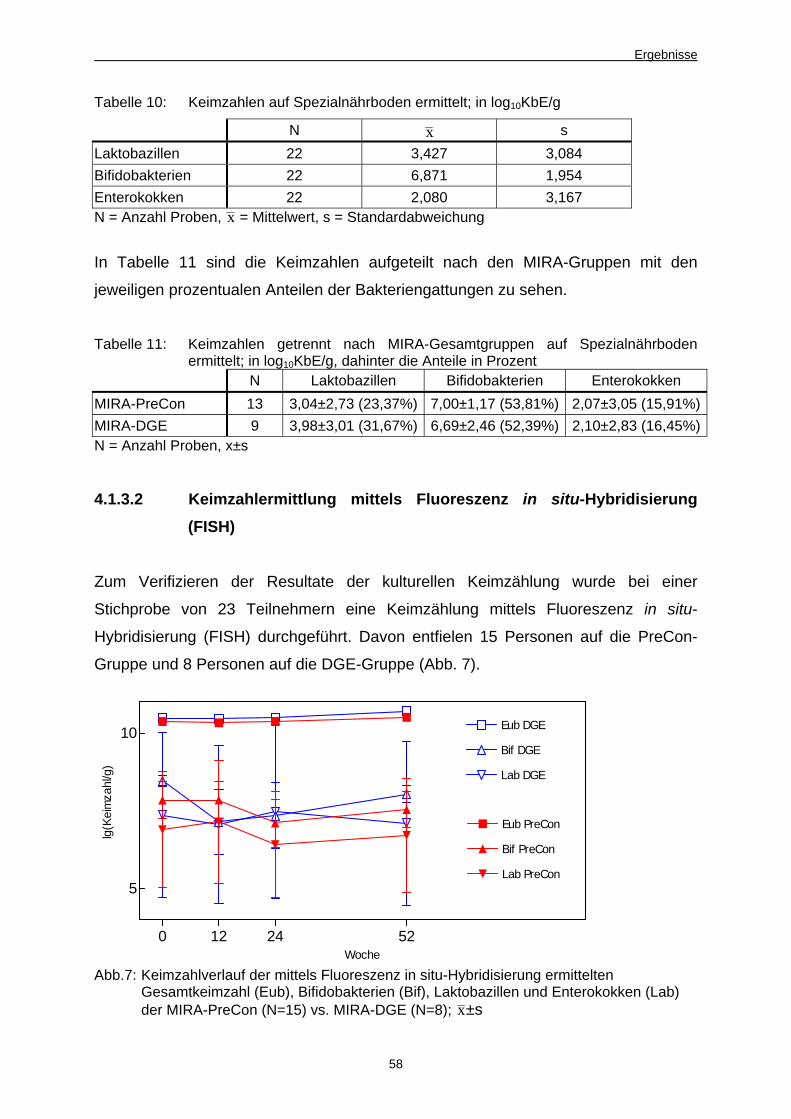
Ergebnisse
Tabelle 10: Keimzahlen auf Spezialnährboden ermittelt; in log10KbE/g
N x s Laktobazillen 22 3,427 3,084 Bifidobakterien 22 6,871 1,954 Enterokokken 22 2,080 3,167 N = Anzahl Proben, x = Mittelwert, s = Standardabweichung
In Tabelle 11 sind die Keimzahlen aufgeteilt nach den MIRA-Gruppen mit den
jeweiligen prozentualen Anteilen der Bakteriengattungen zu sehen.
Tabelle 11: Keimzahlen getrennt nach MIRA-Gesamtgruppen auf Spezialnährboden
ermittelt; in log10KbE/g, dahinter die Anteile in Prozent N Laktobazillen Bifidobakterien Enterokokken MIRA-PreCon 13 3,04±2,73 (23,37%) 7,00±1,17 (53,81%) 2,07±3,05 (15,91%)MIRA-DGE 9 3,98±3,01 (31,67%) 6,69±2,46 (52,39%) 2,10±2,83 (16,45%)N = Anzahl Proben, x±s
4.1.3.2 Keimzahlermittlung mittels Fluoreszenz in situ-Hybridisierung
(FISH)
Zum Verifizieren der Resultate der kulturellen Keimzählung wurde bei einer
Stichprobe von 23 Teilnehmern eine Keimzählung mittels Fluoreszenz in situ-
Hybridisierung (FISH) durchgeführt. Davon entfielen 15 Personen auf die PreCon-
Gruppe und 8 Personen auf die DGE-Gruppe (Abb. 7).
0 12 24 52
5
10
Eub PreCon
Bif PreCon
Lab PreCon
Eub DGE
Bif DGE
Lab DGE
Woche
lg(K
eim
zahl
/g)
Abb.7: Keimzahlverlauf der mittels Fluoreszenz in situ-Hybridisierung ermittelten Gesamtkeimzahl (Eub), Bifidobakterien (Bif), Laktobazillen und Enterokokken (Lab) der MIRA-PreCon (N=15) vs. MIRA-DGE (N=8); x±s
58
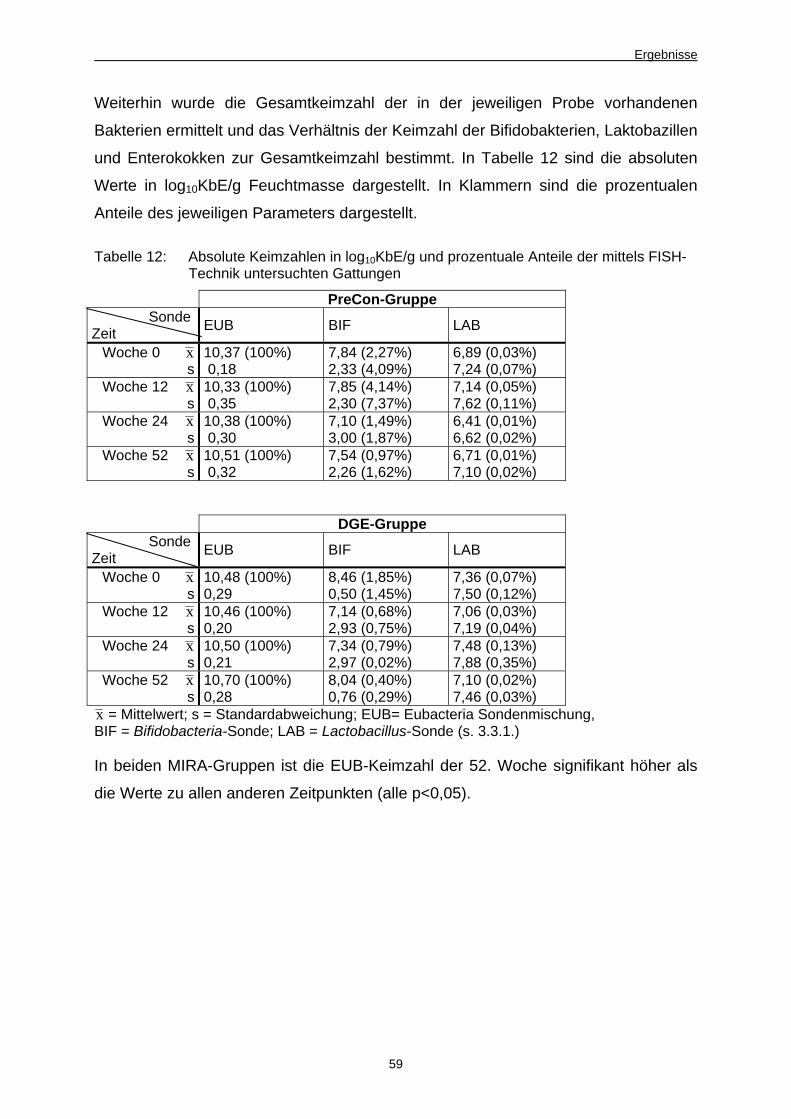
Ergebnisse
Weiterhin wurde die Gesamtkeimzahl der in der jeweiligen Probe vorhandenen
Bakterien ermittelt und das Verhältnis der Keimzahl der Bifidobakterien, Laktobazillen
und Enterokokken zur Gesamtkeimzahl bestimmt. In Tabelle 12 sind die absoluten
Werte in log10KbE/g Feuchtmasse dargestellt. In Klammern sind die prozentualen
Anteile des jeweiligen Parameters dargestellt. Tabelle 12: Absolute Keimzahlen in log10KbE/g und prozentuale Anteile der mittels FISH-
Technik untersuchten Gattungen
PreCon-Gruppe Sonde Zeit EUB BIF LAB
Woche 0 x s
10,37 (100%) 0,18
7,84 (2,27%) 2,33 (4,09%)
6,89 (0,03%) 7,24 (0,07%)
Woche 12 x s
10,33 (100%) 0,35
7,85 (4,14%) 2,30 (7,37%)
7,14 (0,05%) 7,62 (0,11%)
Woche 24 x s
10,38 (100%) 0,30
7,10 (1,49%) 3,00 (1,87%)
6,41 (0,01%) 6,62 (0,02%)
Woche 52 x s
10,51 (100%) 0,32
7,54 (0,97%) 2,26 (1,62%)
6,71 (0,01%) 7,10 (0,02%)
DGE-Gruppe Sonde Zeit EUB BIF LAB
Woche 0 x s
10,48 (100%) 0,29
8,46 (1,85%) 0,50 (1,45%)
7,36 (0,07%) 7,50 (0,12%)
Woche 12 x s
10,46 (100%) 0,20
7,14 (0,68%) 2,93 (0,75%)
7,06 (0,03%) 7,19 (0,04%)
Woche 24 x s
10,50 (100%) 0,21
7,34 (0,79%) 2,97 (0,02%)
7,48 (0,13%) 7,88 (0,35%)
Woche 52 x s
10,70 (100%) 0,28
8,04 (0,40%) 0,76 (0,29%)
7,10 (0,02%) 7,46 (0,03%)
x = Mittelwert; s = Standardabweichung; EUB= Eubacteria Sondenmischung, BIF = Bifidobacteria-Sonde; LAB = Lactobacillus-Sonde (s. 3.3.1.) In beiden MIRA-Gruppen ist die EUB-Keimzahl der 52. Woche signifikant höher als
die Werte zu allen anderen Zeitpunkten (alle p<0,05).
59
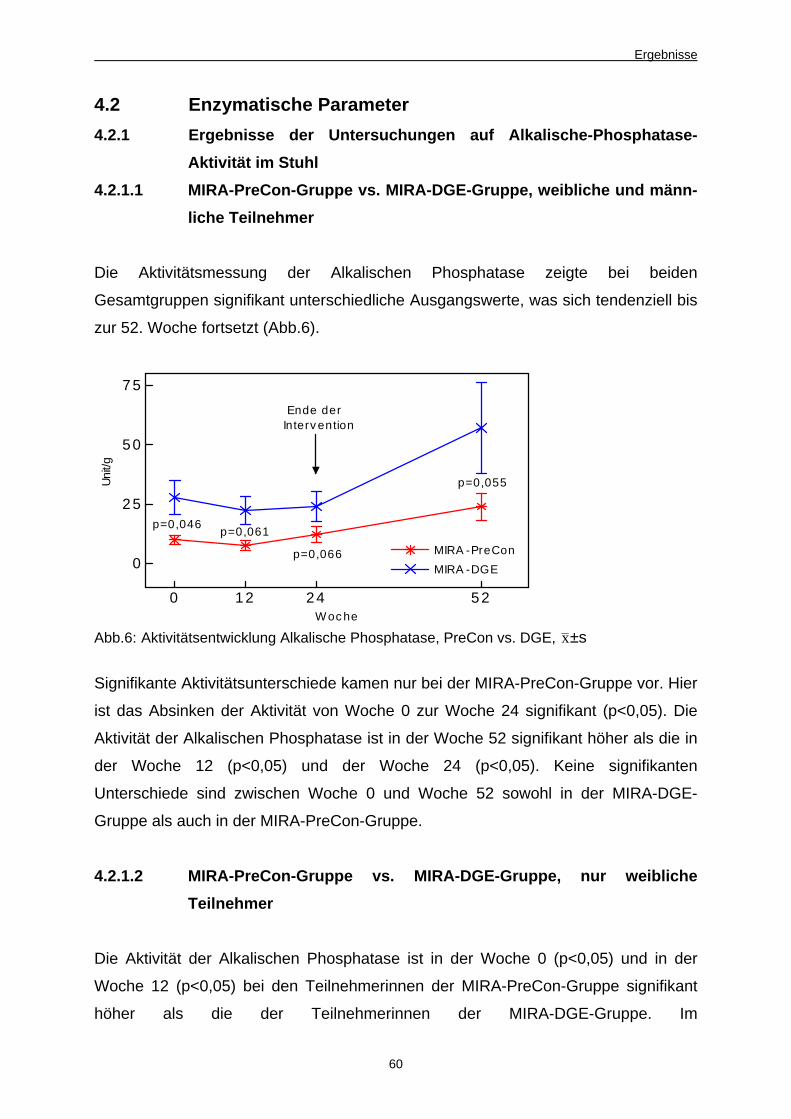
Ergebnisse
4.2 Enzymatische Parameter 4.2.1 Ergebnisse der Untersuchungen auf Alkalische-Phosphatase-
Aktivität im Stuhl 4.2.1.1 MIRA-PreCon-Gruppe vs. MIRA-DGE-Gruppe, weibliche und männ-
liche Teilnehmer Die Aktivitätsmessung der Alkalischen Phosphatase zeigte bei beiden
Gesamtgruppen signifikant unterschiedliche Ausgangswerte, was sich tendenziell bis
zur 52. Woche fortsetzt (Abb.6).
0 12 24 52
0
25
50
75
MIRA -PreConMIRA -DGE
Ende derInterv ention
p=0,046 p=0,061
p=0,066
p=0,055
W oc he
Unit/
g
Abb.6: Aktivitätsentwicklung Alkalische Phosphatase, PreCon vs. DGE, x±s
Signifikante Aktivitätsunterschiede kamen nur bei der MIRA-PreCon-Gruppe vor. Hier
ist das Absinken der Aktivität von Woche 0 zur Woche 24 signifikant (p<0,05). Die
Aktivität der Alkalischen Phosphatase ist in der Woche 52 signifikant höher als die in
der Woche 12 (p<0,05) und der Woche 24 (p<0,05). Keine signifikanten
Unterschiede sind zwischen Woche 0 und Woche 52 sowohl in der MIRA-DGE-
Gruppe als auch in der MIRA-PreCon-Gruppe.
4.2.1.2 MIRA-PreCon-Gruppe vs. MIRA-DGE-Gruppe, nur weibliche Teilnehmer
Die Aktivität der Alkalischen Phosphatase ist in der Woche 0 (p<0,05) und in der
Woche 12 (p<0,05) bei den Teilnehmerinnen der MIRA-PreCon-Gruppe signifikant
höher als die der Teilnehmerinnen der MIRA-DGE-Gruppe. Im
60
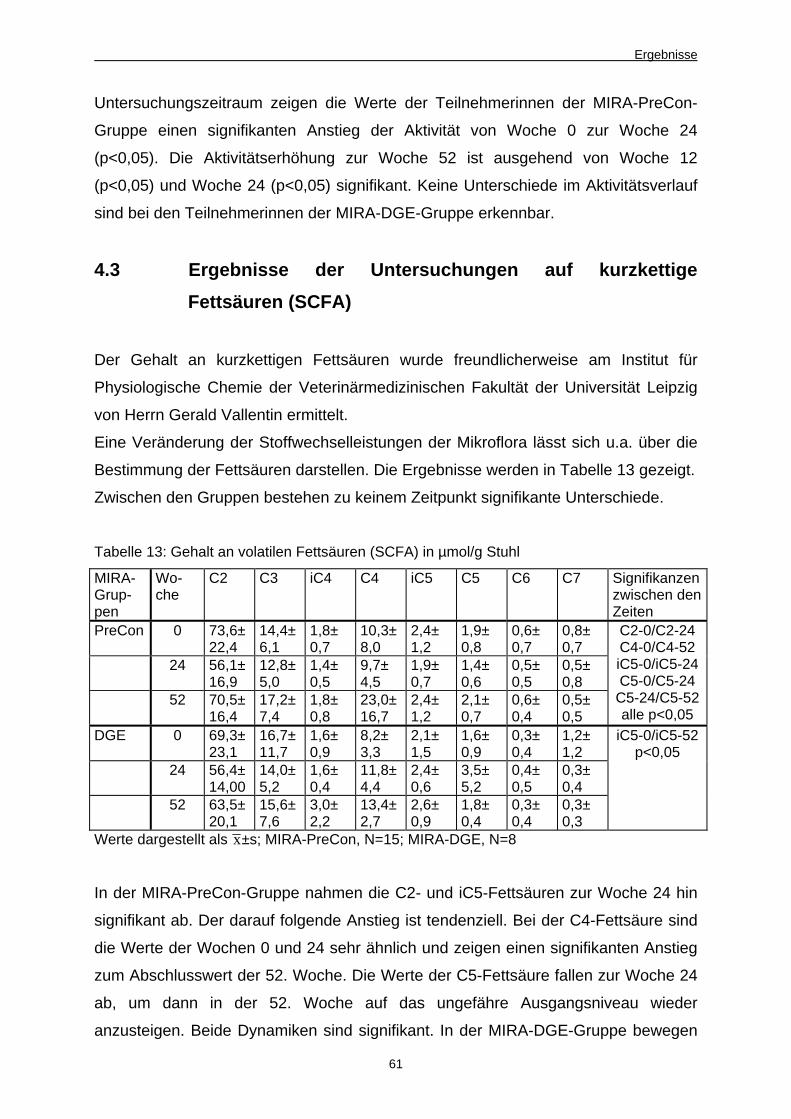
Ergebnisse
Untersuchungszeitraum zeigen die Werte der Teilnehmerinnen der MIRA-PreCon-
Gruppe einen signifikanten Anstieg der Aktivität von Woche 0 zur Woche 24
(p<0,05). Die Aktivitätserhöhung zur Woche 52 ist ausgehend von Woche 12
(p<0,05) und Woche 24 (p<0,05) signifikant. Keine Unterschiede im Aktivitätsverlauf
sind bei den Teilnehmerinnen der MIRA-DGE-Gruppe erkennbar.
4.3 Ergebnisse der Untersuchungen auf kurzkettige Fettsäuren (SCFA)
Der Gehalt an kurzkettigen Fettsäuren wurde freundlicherweise am Institut für
Physiologische Chemie der Veterinärmedizinischen Fakultät der Universität Leipzig
von Herrn Gerald Vallentin ermittelt.
Eine Veränderung der Stoffwechselleistungen der Mikroflora lässt sich u.a. über die
Bestimmung der Fettsäuren darstellen. Die Ergebnisse werden in Tabelle 13 gezeigt.
Zwischen den Gruppen bestehen zu keinem Zeitpunkt signifikante Unterschiede.
Tabelle 13: Gehalt an volatilen Fettsäuren (SCFA) in µmol/g Stuhl
MIRA-Grup-pen
Wo-che
C2 C3 iC4 C4 iC5 C5 C6 C7 Signifikanzenzwischen den Zeiten
PreCon
0 73,6± 22,4
14,4± 6,1
1,8± 0,7
10,3± 8,0
2,4± 1,2
1,9± 0,8
0,6± 0,7
0,8± 0,7
24 56,1± 16,9
12,8± 5,0
1,4± 0,5
9,7± 4,5
1,9± 0,7
1,4± 0,6
0,5± 0,5
0,5± 0,8
52 70,5± 16,4
17,2± 7,4
1,8± 0,8
23,0± 16,7
2,4± 1,2
2,1± 0,7
0,6± 0,4
0,5± 0,5
C2-0/C2-24 C4-0/C4-52 iC5-0/iC5-24 C5-0/C5-24
C5-24/C5-52alle p<0,05
DGE
0 69,3± 23,1
16,7± 11,7
1,6± 0,9
8,2± 3,3
2,1± 1,5
1,6± 0,9
0,3± 0,4
1,2± 1,2
24 56,4± 14,00
14,0± 5,2
1,6± 0,4
11,8± 4,4
2,4± 0,6
3,5± 5,2
0,4± 0,5
0,3± 0,4
52 63,5± 20,1
15,6± 7,6
3,0± 2,2
13,4± 2,7
2,6± 0,9
1,8± 0,4
0,3± 0,4
0,3± 0,3
iC5-0/iC5-52 p<0,05
Werte dargestellt als x±s; MIRA-PreCon, N=15; MIRA-DGE, N=8
In der MIRA-PreCon-Gruppe nahmen die C2- und iC5-Fettsäuren zur Woche 24 hin
signifikant ab. Der darauf folgende Anstieg ist tendenziell. Bei der C4-Fettsäure sind
die Werte der Wochen 0 und 24 sehr ähnlich und zeigen einen signifikanten Anstieg
zum Abschlusswert der 52. Woche. Die Werte der C5-Fettsäure fallen zur Woche 24
ab, um dann in der 52. Woche auf das ungefähre Ausgangsniveau wieder
anzusteigen. Beide Dynamiken sind signifikant. In der MIRA-DGE-Gruppe bewegen
61
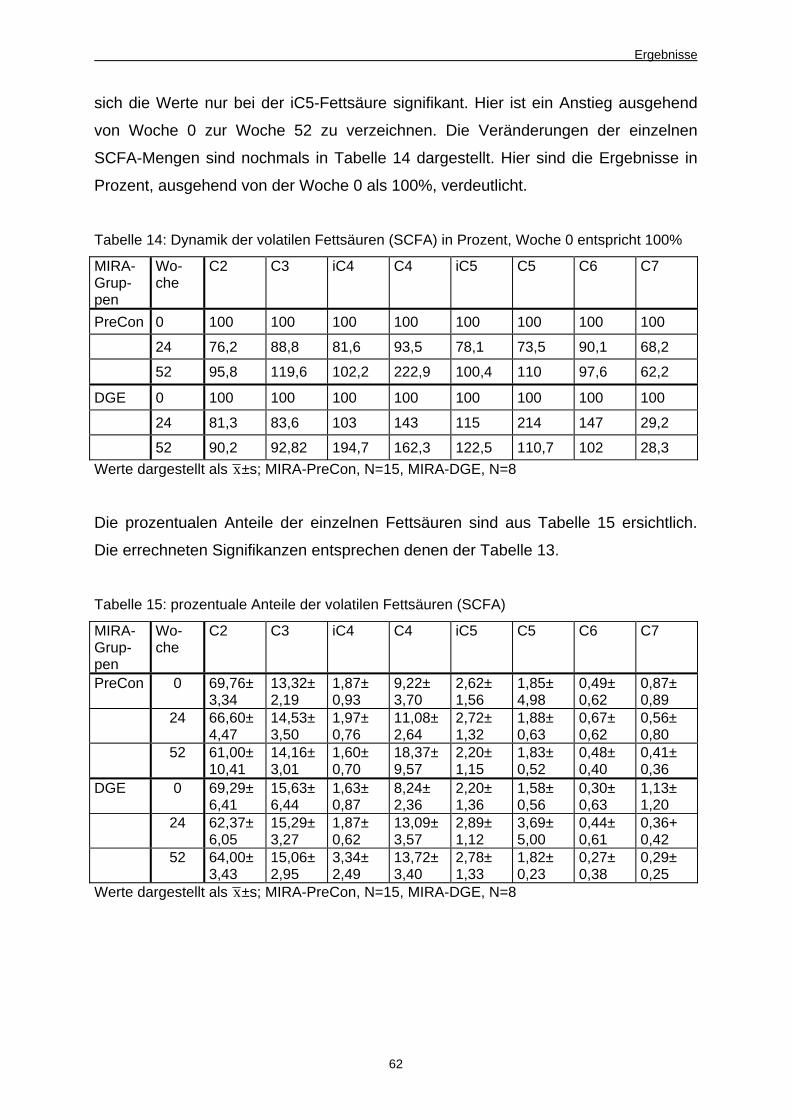
Ergebnisse
sich die Werte nur bei der iC5-Fettsäure signifikant. Hier ist ein Anstieg ausgehend
von Woche 0 zur Woche 52 zu verzeichnen. Die Veränderungen der einzelnen
SCFA-Mengen sind nochmals in Tabelle 14 dargestellt. Hier sind die Ergebnisse in
Prozent, ausgehend von der Woche 0 als 100%, verdeutlicht.
Tabelle 14: Dynamik der volatilen Fettsäuren (SCFA) in Prozent, Woche 0 entspricht 100%
MIRA-Grup-pen
Wo-che
C2 C3 iC4 C4 iC5 C5 C6 C7
PreCon 0 100 100 100 100 100 100 100 100
24 76,2 88,8 81,6 93,5 78,1 73,5 90,1 68,2
52 95,8 119,6 102,2 222,9 100,4 110 97,6 62,2
DGE 0 100 100 100 100 100 100 100 100
24 81,3 83,6 103 143 115 214 147 29,2
52 90,2 92,82 194,7 162,3 122,5 110,7 102 28,3 Werte dargestellt als x±s; MIRA-PreCon, N=15, MIRA-DGE, N=8
Die prozentualen Anteile der einzelnen Fettsäuren sind aus Tabelle 15 ersichtlich.
Die errechneten Signifikanzen entsprechen denen der Tabelle 13.
Tabelle 15: prozentuale Anteile der volatilen Fettsäuren (SCFA)
MIRA-Grup-pen
Wo-che
C2 C3 iC4 C4 iC5 C5 C6 C7
PreCon
0 69,76± 3,34
13,32± 2,19
1,87± 0,93
9,22± 3,70
2,62± 1,56
1,85± 4,98
0,49± 0,62
0,87± 0,89
24 66,60± 4,47
14,53± 3,50
1,97± 0,76
11,08± 2,64
2,72± 1,32
1,88± 0,63
0,67± 0,62
0,56± 0,80
52 61,00± 10,41
14,16± 3,01
1,60± 0,70
18,37± 9,57
2,20± 1,15
1,83± 0,52
0,48± 0,40
0,41± 0,36
DGE
0 69,29± 6,41
15,63± 6,44
1,63± 0,87
8,24± 2,36
2,20± 1,36
1,58± 0,56
0,30± 0,63
1,13± 1,20
24 62,37± 6,05
15,29± 3,27
1,87± 0,62
13,09± 3,57
2,89± 1,12
3,69± 5,00
0,44± 0,61
0,36+ 0,42
52 64,00± 3,43
15,06± 2,95
3,34± 2,49
13,72± 3,40
2,78± 1,33
1,82± 0,23
0,27± 0,38
0,29± 0,25
Werte dargestellt als x±s; MIRA-PreCon, N=15, MIRA-DGE, N=8
62
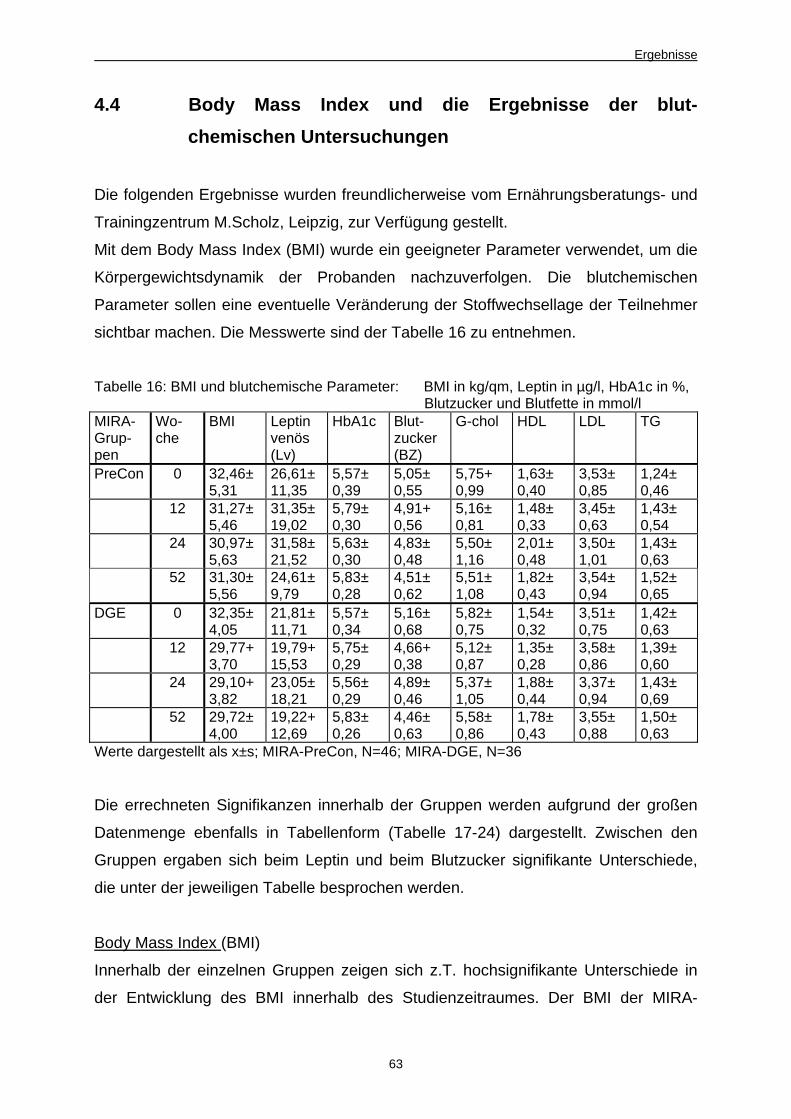
Ergebnisse
4.4 Body Mass Index und die Ergebnisse der blut-chemischen Untersuchungen
Die folgenden Ergebnisse wurden freundlicherweise vom Ernährungsberatungs- und
Trainingzentrum M.Scholz, Leipzig, zur Verfügung gestellt.
Mit dem Body Mass Index (BMI) wurde ein geeigneter Parameter verwendet, um die
Körpergewichtsdynamik der Probanden nachzuverfolgen. Die blutchemischen
Parameter sollen eine eventuelle Veränderung der Stoffwechsellage der Teilnehmer
sichtbar machen. Die Messwerte sind der Tabelle 16 zu entnehmen.
Tabelle 16: BMI und blutchemische Parameter: BMI in kg/qm, Leptin in µg/l, HbA1c in %, Blutzucker und Blutfette in mmol/l MIRA-Grup-pen
Wo-che
BMI Leptin venös (Lv)
HbA1c Blut- zucker (BZ)
G-chol HDL LDL TG
PreCon
0 32,46± 5,31
26,61± 11,35
5,57± 0,39
5,05± 0,55
5,75+ 0,99
1,63± 0,40
3,53± 0,85
1,24± 0,46
12 31,27± 5,46
31,35± 19,02
5,79± 0,30
4,91+ 0,56
5,16± 0,81
1,48± 0,33
3,45± 0,63
1,43± 0,54
24 30,97± 5,63
31,58± 21,52
5,63± 0,30
4,83± 0,48
5,50± 1,16
2,01± 0,48
3,50± 1,01
1,43± 0,63
52 31,30± 5,56
24,61± 9,79
5,83± 0,28
4,51± 0,62
5,51± 1,08
1,82± 0,43
3,54± 0,94
1,52± 0,65
DGE
0 32,35± 4,05
21,81± 11,71
5,57± 0,34
5,16± 0,68
5,82± 0,75
1,54± 0,32
3,51± 0,75
1,42± 0,63
12
29,77+ 3,70
19,79+ 15,53
5,75± 0,29
4,66+ 0,38
5,12± 0,87
1,35± 0,28
3,58± 0,86
1,39± 0,60
24 29,10+ 3,82
23,05± 18,21
5,56± 0,29
4,89± 0,46
5,37± 1,05
1,88± 0,44
3,37± 0,94
1,43± 0,69
52 29,72± 4,00
19,22+ 12,69
5,83± 0,26
4,46± 0,63
5,58± 0,86
1,78± 0,43
3,55± 0,88
1,50± 0,63
Werte dargestellt als x±s; MIRA-PreCon, N=46; MIRA-DGE, N=36
Die errechneten Signifikanzen innerhalb der Gruppen werden aufgrund der großen
Datenmenge ebenfalls in Tabellenform (Tabelle 17-24) dargestellt. Zwischen den
Gruppen ergaben sich beim Leptin und beim Blutzucker signifikante Unterschiede,
die unter der jeweiligen Tabelle besprochen werden.
Body Mass Index (BMI)
Innerhalb der einzelnen Gruppen zeigen sich z.T. hochsignifikante Unterschiede in
der Entwicklung des BMI innerhalb des Studienzeitraumes. Der BMI der MIRA-
63
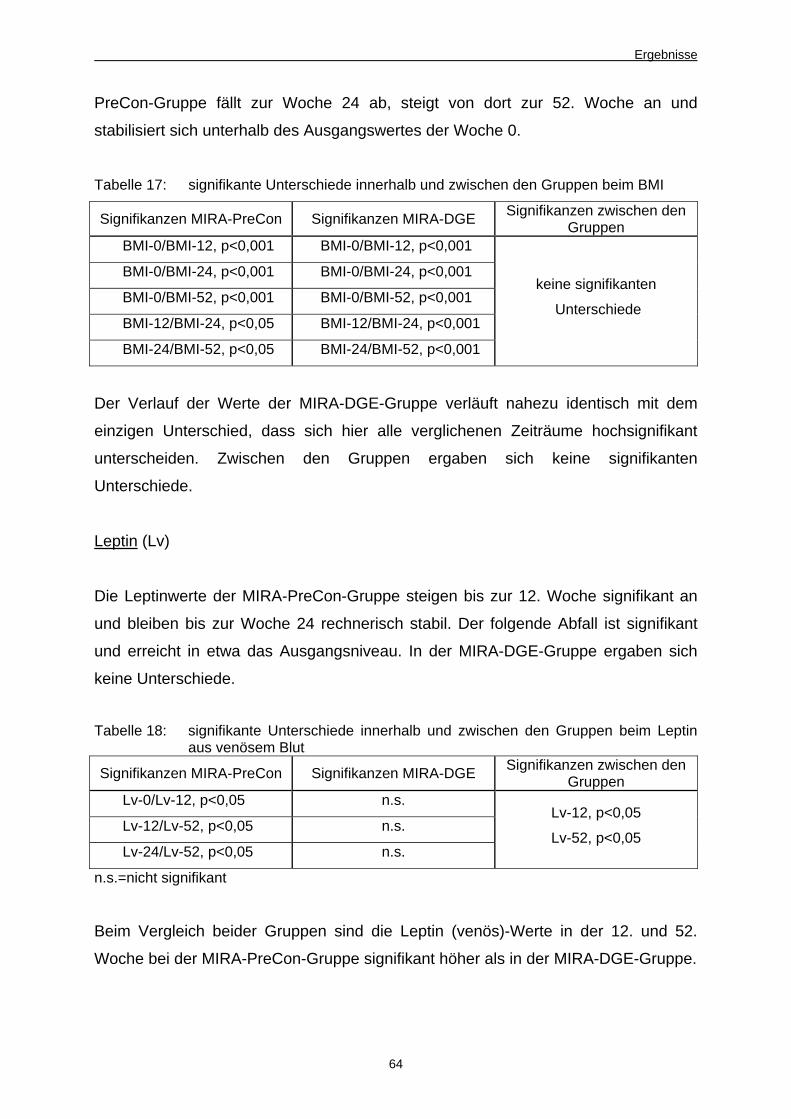
Ergebnisse
PreCon-Gruppe fällt zur Woche 24 ab, steigt von dort zur 52. Woche an und
stabilisiert sich unterhalb des Ausgangswertes der Woche 0.
Tabelle 17: signifikante Unterschiede innerhalb und zwischen den Gruppen beim BMI
Signifikanzen MIRA-PreCon Signifikanzen MIRA-DGE Signifikanzen zwischen den Gruppen
BMI-0/BMI-12, p<0,001 BMI-0/BMI-12, p<0,001
BMI-0/BMI-24, p<0,001 BMI-0/BMI-24, p<0,001
BMI-0/BMI-52, p<0,001 BMI-0/BMI-52, p<0,001
BMI-12/BMI-24, p<0,05 BMI-12/BMI-24, p<0,001
BMI-24/BMI-52, p<0,05 BMI-24/BMI-52, p<0,001
keine signifikanten
Unterschiede
Der Verlauf der Werte der MIRA-DGE-Gruppe verläuft nahezu identisch mit dem
einzigen Unterschied, dass sich hier alle verglichenen Zeiträume hochsignifikant
unterscheiden. Zwischen den Gruppen ergaben sich keine signifikanten
Unterschiede.
Leptin (Lv)
Die Leptinwerte der MIRA-PreCon-Gruppe steigen bis zur 12. Woche signifikant an
und bleiben bis zur Woche 24 rechnerisch stabil. Der folgende Abfall ist signifikant
und erreicht in etwa das Ausgangsniveau. In der MIRA-DGE-Gruppe ergaben sich
keine Unterschiede.
Tabelle 18: signifikante Unterschiede innerhalb und zwischen den Gruppen beim Leptin aus venösem Blut
Signifikanzen MIRA-PreCon Signifikanzen MIRA-DGE Signifikanzen zwischen den Gruppen
Lv-0/Lv-12, p<0,05 n.s.
Lv-12/Lv-52, p<0,05 n.s.
Lv-24/Lv-52, p<0,05 n.s.
Lv-12, p<0,05
Lv-52, p<0,05
n.s.=nicht signifikant
Beim Vergleich beider Gruppen sind die Leptin (venös)-Werte in der 12. und 52.
Woche bei der MIRA-PreCon-Gruppe signifikant höher als in der MIRA-DGE-Gruppe.
64
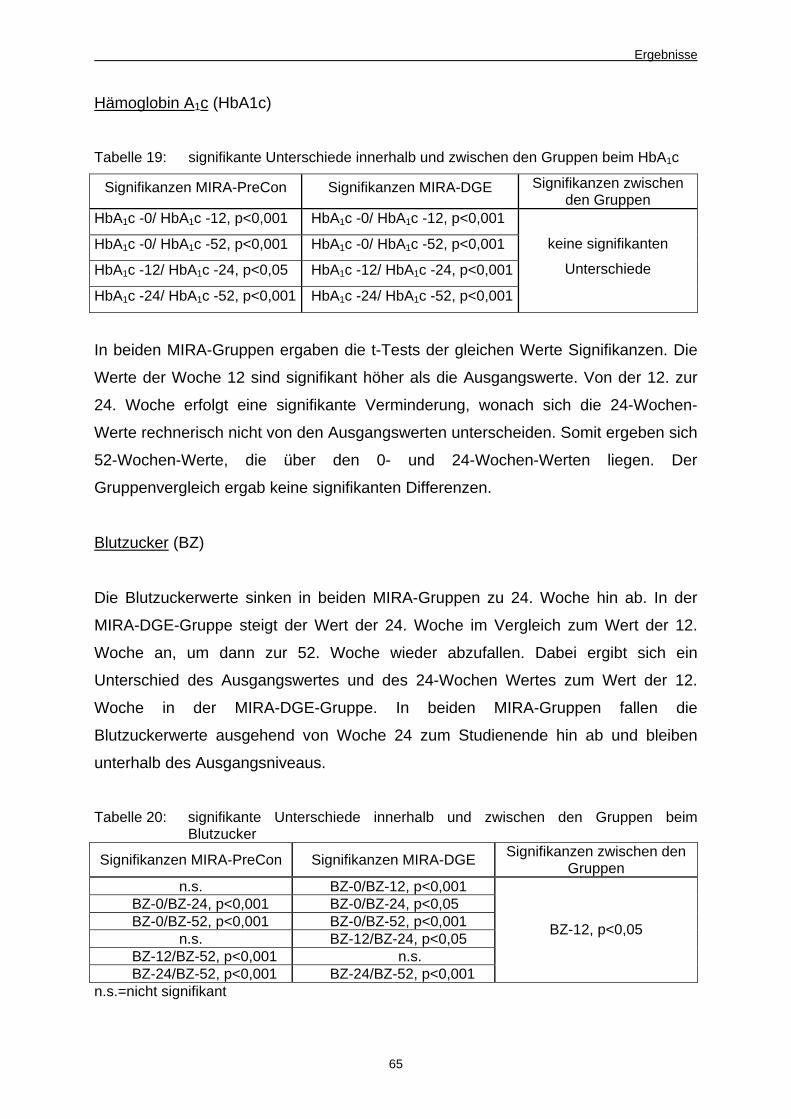
Ergebnisse
Hämoglobin A1c (HbA1c)
Tabelle 19: signifikante Unterschiede innerhalb und zwischen den Gruppen beim HbA1c
Signifikanzen MIRA-PreCon Signifikanzen MIRA-DGE Signifikanzen zwischen den Gruppen
HbA1c -0/ HbA1c -12, p<0,001 HbA1c -0/ HbA1c -12, p<0,001
HbA1c -0/ HbA1c -52, p<0,001 HbA1c -0/ HbA1c -52, p<0,001
HbA1c -12/ HbA1c -24, p<0,05 HbA1c -12/ HbA1c -24, p<0,001
HbA1c -24/ HbA1c -52, p<0,001 HbA1c -24/ HbA1c -52, p<0,001
keine signifikanten
Unterschiede
In beiden MIRA-Gruppen ergaben die t-Tests der gleichen Werte Signifikanzen. Die
Werte der Woche 12 sind signifikant höher als die Ausgangswerte. Von der 12. zur
24. Woche erfolgt eine signifikante Verminderung, wonach sich die 24-Wochen-
Werte rechnerisch nicht von den Ausgangswerten unterscheiden. Somit ergeben sich
52-Wochen-Werte, die über den 0- und 24-Wochen-Werten liegen. Der
Gruppenvergleich ergab keine signifikanten Differenzen.
Blutzucker (BZ)
Die Blutzuckerwerte sinken in beiden MIRA-Gruppen zu 24. Woche hin ab. In der
MIRA-DGE-Gruppe steigt der Wert der 24. Woche im Vergleich zum Wert der 12.
Woche an, um dann zur 52. Woche wieder abzufallen. Dabei ergibt sich ein
Unterschied des Ausgangswertes und des 24-Wochen Wertes zum Wert der 12.
Woche in der MIRA-DGE-Gruppe. In beiden MIRA-Gruppen fallen die
Blutzuckerwerte ausgehend von Woche 24 zum Studienende hin ab und bleiben
unterhalb des Ausgangsniveaus.
Tabelle 20: signifikante Unterschiede innerhalb und zwischen den Gruppen beim
Blutzucker
Signifikanzen MIRA-PreCon Signifikanzen MIRA-DGE Signifikanzen zwischen den Gruppen
n.s. BZ-0/BZ-12, p<0,001 BZ-0/BZ-24, p<0,001 BZ-0/BZ-24, p<0,05 BZ-0/BZ-52, p<0,001 BZ-0/BZ-52, p<0,001
n.s. BZ-12/BZ-24, p<0,05 BZ-12/BZ-52, p<0,001 n.s. BZ-24/BZ-52, p<0,001 BZ-24/BZ-52, p<0,001
BZ-12, p<0,05
n.s.=nicht signifikant
65
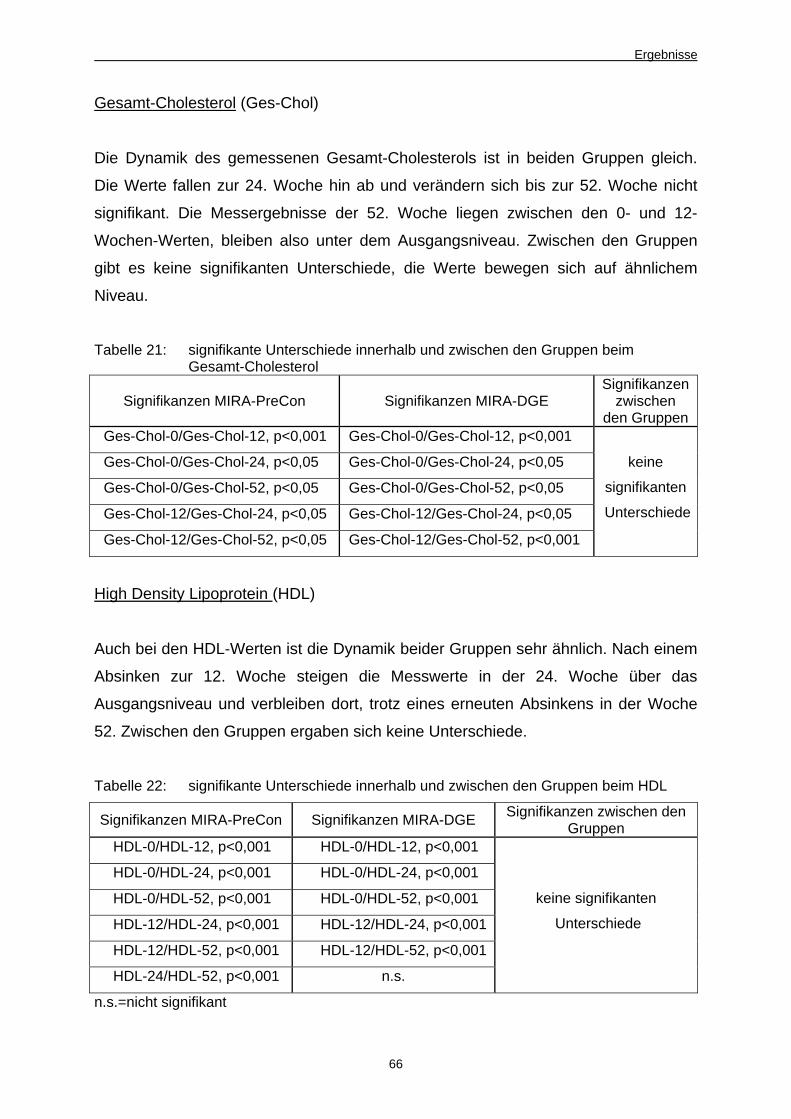
Ergebnisse
Gesamt-Cholesterol (Ges-Chol)
Die Dynamik des gemessenen Gesamt-Cholesterols ist in beiden Gruppen gleich.
Die Werte fallen zur 24. Woche hin ab und verändern sich bis zur 52. Woche nicht
signifikant. Die Messergebnisse der 52. Woche liegen zwischen den 0- und 12-
Wochen-Werten, bleiben also unter dem Ausgangsniveau. Zwischen den Gruppen
gibt es keine signifikanten Unterschiede, die Werte bewegen sich auf ähnlichem
Niveau.
Tabelle 21: signifikante Unterschiede innerhalb und zwischen den Gruppen beim Gesamt-Cholesterol
Signifikanzen MIRA-PreCon Signifikanzen MIRA-DGE Signifikanzen
zwischen den Gruppen
Ges-Chol-0/Ges-Chol-12, p<0,001 Ges-Chol-0/Ges-Chol-12, p<0,001
Ges-Chol-0/Ges-Chol-24, p<0,05 Ges-Chol-0/Ges-Chol-24, p<0,05
Ges-Chol-0/Ges-Chol-52, p<0,05 Ges-Chol-0/Ges-Chol-52, p<0,05
Ges-Chol-12/Ges-Chol-24, p<0,05 Ges-Chol-12/Ges-Chol-24, p<0,05
Ges-Chol-12/Ges-Chol-52, p<0,05 Ges-Chol-12/Ges-Chol-52, p<0,001
keine
signifikanten
Unterschiede
High Density Lipoprotein (HDL)
Auch bei den HDL-Werten ist die Dynamik beider Gruppen sehr ähnlich. Nach einem
Absinken zur 12. Woche steigen die Messwerte in der 24. Woche über das
Ausgangsniveau und verbleiben dort, trotz eines erneuten Absinkens in der Woche
52. Zwischen den Gruppen ergaben sich keine Unterschiede.
Tabelle 22: signifikante Unterschiede innerhalb und zwischen den Gruppen beim HDL
Signifikanzen MIRA-PreCon Signifikanzen MIRA-DGE Signifikanzen zwischen den Gruppen
HDL-0/HDL-12, p<0,001 HDL-0/HDL-12, p<0,001
HDL-0/HDL-24, p<0,001 HDL-0/HDL-24, p<0,001
HDL-0/HDL-52, p<0,001 HDL-0/HDL-52, p<0,001
HDL-12/HDL-24, p<0,001 HDL-12/HDL-24, p<0,001
HDL-12/HDL-52, p<0,001 HDL-12/HDL-52, p<0,001
HDL-24/HDL-52, p<0,001 n.s.
keine signifikanten
Unterschiede
n.s.=nicht signifikant
66
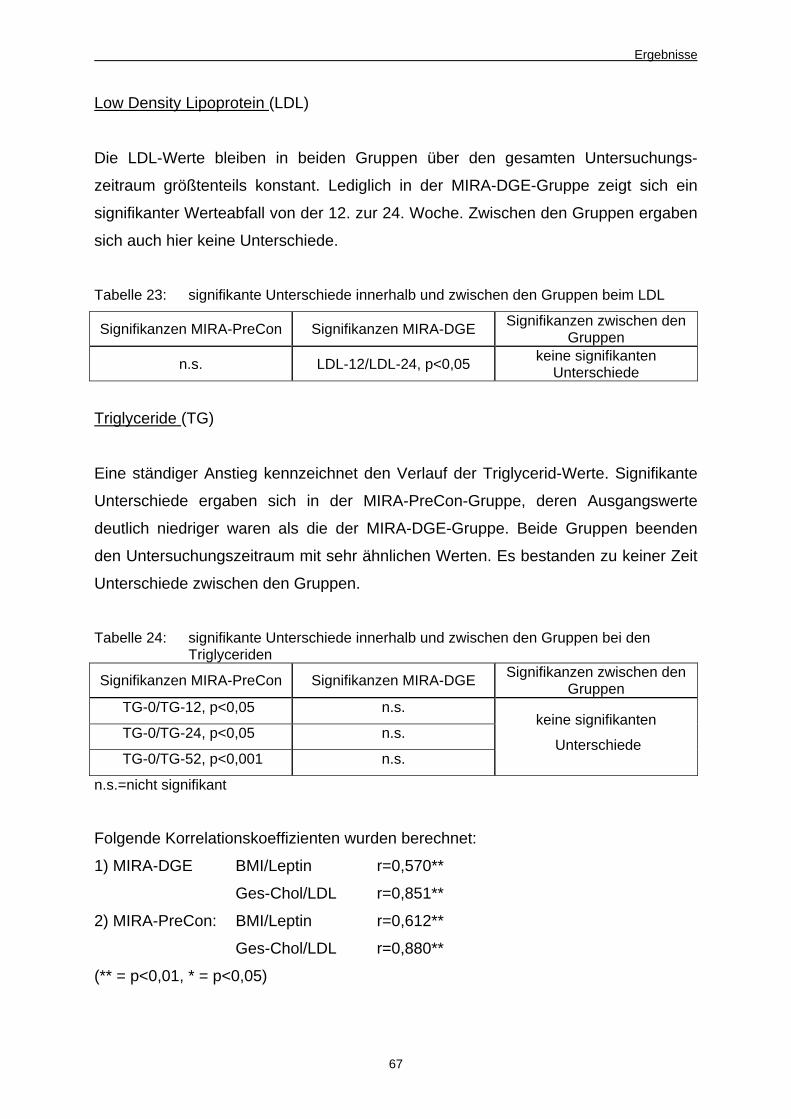
Ergebnisse
Low Density Lipoprotein (LDL)
Die LDL-Werte bleiben in beiden Gruppen über den gesamten Untersuchungs-
zeitraum größtenteils konstant. Lediglich in der MIRA-DGE-Gruppe zeigt sich ein
signifikanter Werteabfall von der 12. zur 24. Woche. Zwischen den Gruppen ergaben
sich auch hier keine Unterschiede.
Tabelle 23: signifikante Unterschiede innerhalb und zwischen den Gruppen beim LDL
Signifikanzen MIRA-PreCon Signifikanzen MIRA-DGE Signifikanzen zwischen den Gruppen
n.s. LDL-12/LDL-24, p<0,05 keine signifikanten Unterschiede
Triglyceride (TG)
Eine ständiger Anstieg kennzeichnet den Verlauf der Triglycerid-Werte. Signifikante
Unterschiede ergaben sich in der MIRA-PreCon-Gruppe, deren Ausgangswerte
deutlich niedriger waren als die der MIRA-DGE-Gruppe. Beide Gruppen beenden
den Untersuchungszeitraum mit sehr ähnlichen Werten. Es bestanden zu keiner Zeit
Unterschiede zwischen den Gruppen.
Tabelle 24: signifikante Unterschiede innerhalb und zwischen den Gruppen bei den Triglyceriden
Signifikanzen MIRA-PreCon Signifikanzen MIRA-DGE Signifikanzen zwischen den Gruppen
TG-0/TG-12, p<0,05 n.s.
TG-0/TG-24, p<0,05 n.s.
TG-0/TG-52, p<0,001 n.s.
keine signifikanten
Unterschiede
n.s.=nicht signifikant
Folgende Korrelationskoeffizienten wurden berechnet:
1) MIRA-DGE BMI/Leptin r=0,570**
Ges-Chol/LDL r=0,851**
2) MIRA-PreCon: BMI/Leptin r=0,612**
Ges-Chol/LDL r=0,880**
(** = p<0,01, * = p<0,05)
67
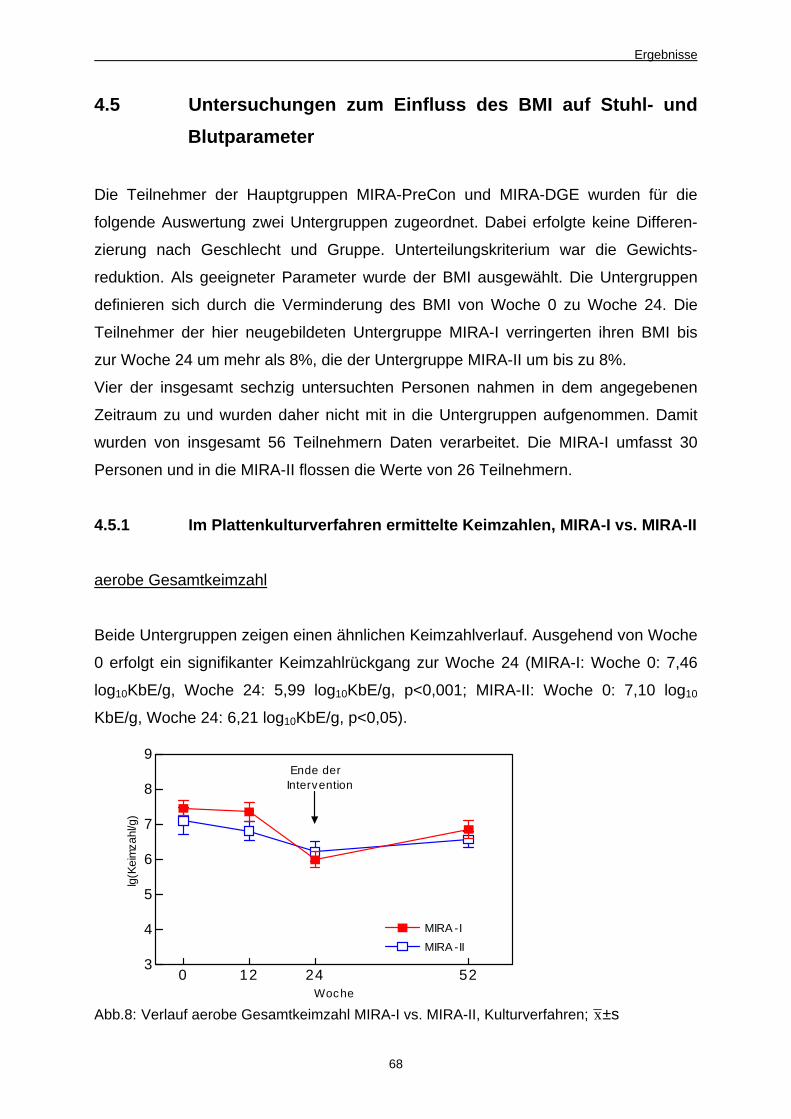
Ergebnisse
4.5 Untersuchungen zum Einfluss des BMI auf Stuhl- und Blutparameter
Die Teilnehmer der Hauptgruppen MIRA-PreCon und MIRA-DGE wurden für die
folgende Auswertung zwei Untergruppen zugeordnet. Dabei erfolgte keine Differen-
zierung nach Geschlecht und Gruppe. Unterteilungskriterium war die Gewichts-
reduktion. Als geeigneter Parameter wurde der BMI ausgewählt. Die Untergruppen
definieren sich durch die Verminderung des BMI von Woche 0 zu Woche 24. Die
Teilnehmer der hier neugebildeten Untergruppe MIRA-I verringerten ihren BMI bis
zur Woche 24 um mehr als 8%, die der Untergruppe MIRA-II um bis zu 8%.
Vier der insgesamt sechzig untersuchten Personen nahmen in dem angegebenen
Zeitraum zu und wurden daher nicht mit in die Untergruppen aufgenommen. Damit
wurden von insgesamt 56 Teilnehmern Daten verarbeitet. Die MIRA-I umfasst 30
Personen und in die MIRA-II flossen die Werte von 26 Teilnehmern.
4.5.1 Im Plattenkulturverfahren ermittelte Keimzahlen, MIRA-I vs. MIRA-II aerobe Gesamtkeimzahl
Beide Untergruppen zeigen einen ähnlichen Keimzahlverlauf. Ausgehend von Woche
0 erfolgt ein signifikanter Keimzahlrückgang zur Woche 24 (MIRA-I: Woche 0: 7,46
log10KbE/g, Woche 24: 5,99 log10KbE/g, p<0,001; MIRA-II: Woche 0: 7,10 log10
KbE/g, Woche 24: 6,21 log10KbE/g, p<0,05).
0 12 24 523
4
5
6
7
8
9
MIRA -IMIRA -II
Ende derIntervention
Woche
lg(K
eim
zahl
/g)
Abb.8: Verlauf aerobe Gesamtkeimzahl MIRA-I vs. MIRA-II, Kulturverfahren; x±s
68
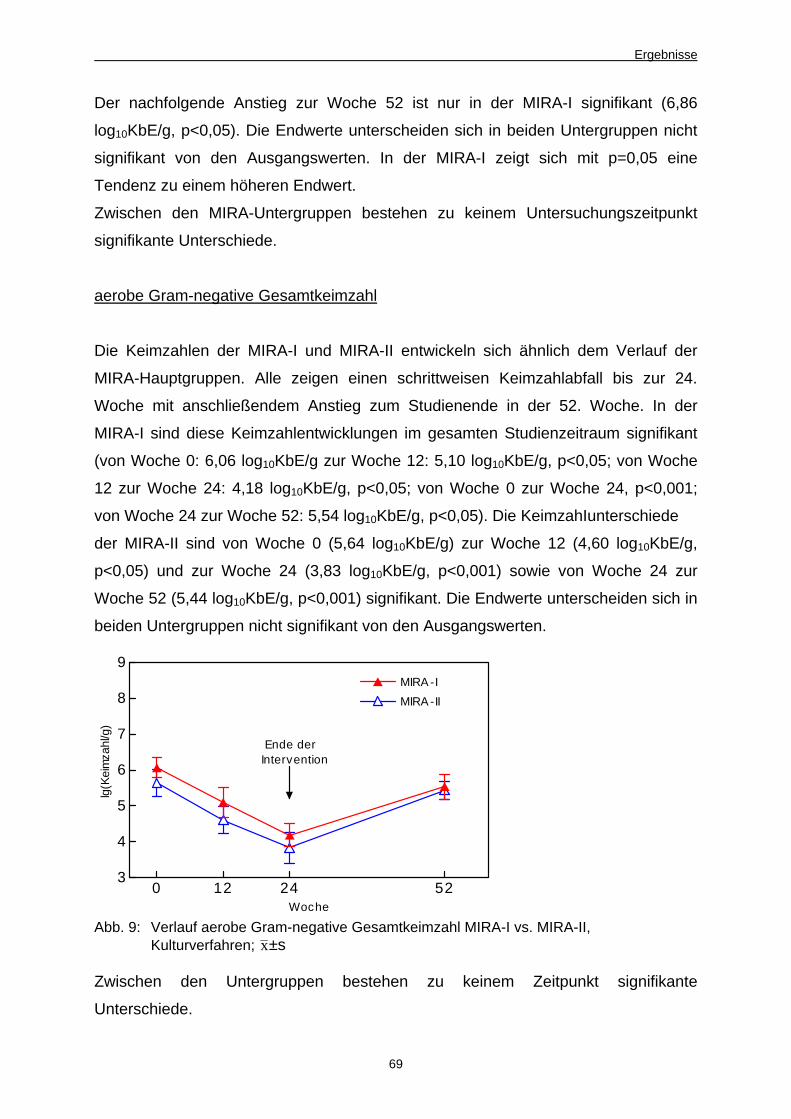
Ergebnisse
Der nachfolgende Anstieg zur Woche 52 ist nur in der MIRA-I signifikant (6,86
log10KbE/g, p<0,05). Die Endwerte unterscheiden sich in beiden Untergruppen nicht
signifikant von den Ausgangswerten. In der MIRA-I zeigt sich mit p=0,05 eine
Tendenz zu einem höheren Endwert.
Zwischen den MIRA-Untergruppen bestehen zu keinem Untersuchungszeitpunkt
signifikante Unterschiede.
aerobe Gram-negative Gesamtkeimzahl
Die Keimzahlen der MIRA-I und MIRA-II entwickeln sich ähnlich dem Verlauf der
MIRA-Hauptgruppen. Alle zeigen einen schrittweisen Keimzahlabfall bis zur 24.
Woche mit anschließendem Anstieg zum Studienende in der 52. Woche. In der
MIRA-I sind diese Keimzahlentwicklungen im gesamten Studienzeitraum signifikant
(von Woche 0: 6,06 log10KbE/g zur Woche 12: 5,10 log10KbE/g, p<0,05; von Woche
12 zur Woche 24: 4,18 log10KbE/g, p<0,05; von Woche 0 zur Woche 24, p<0,001;
von Woche 24 zur Woche 52: 5,54 log10KbE/g, p<0,05). Die KeimzahIunterschiede
der MIRA-II sind von Woche 0 (5,64 log10KbE/g) zur Woche 12 (4,60 log10KbE/g,
p<0,05) und zur Woche 24 (3,83 log10KbE/g, p<0,001) sowie von Woche 24 zur
Woche 52 (5,44 log10KbE/g, p<0,001) signifikant. Die Endwerte unterscheiden sich in
beiden Untergruppen nicht signifikant von den Ausgangswerten.
0 12 24 523
4
5
6
7
8
9MIRA -IMIRA -II
Ende derIntervention
Woche
lg(K
eim
zahl
/g)
Abb. 9: Verlauf aerobe Gram-negative Gesamtkeimzahl MIRA-I vs. MIRA-II, Kulturverfahren; x±s Zwischen den Untergruppen bestehen zu keinem Zeitpunkt signifikante
Unterschiede.
69
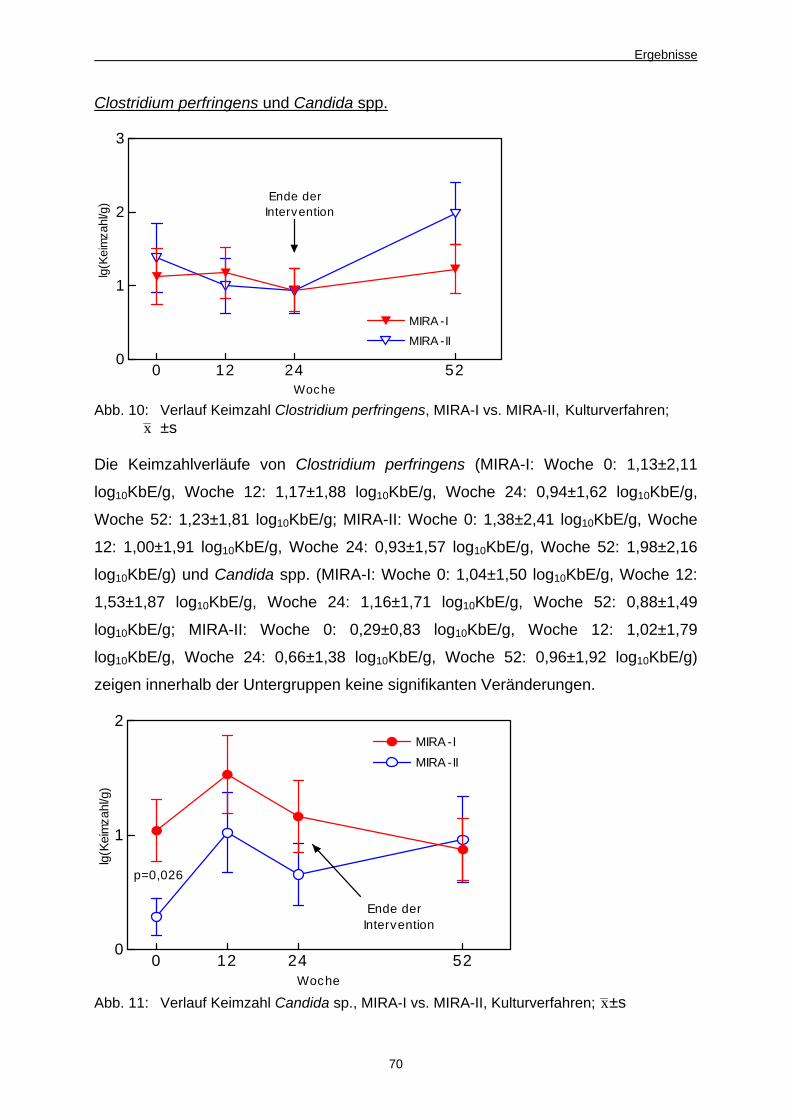
Ergebnisse
Clostridium perfringens und Candida spp.
0 12 24 520
1
2
3
MIRA -IMIRA -II
Ende derIntervention
Woche
lg(K
eim
zahl
/g)
Abb. 10: Verlauf Keimzahl Clostridium perfringens, MIRA-I vs. MIRA-II, Kulturverfahren; x ±s
Die Keimzahlverläufe von Clostridium perfringens (MIRA-I: Woche 0: 1,13±2,11
log10KbE/g, Woche 12: 1,17±1,88 log10KbE/g, Woche 24: 0,94±1,62 log10KbE/g,
Woche 52: 1,23±1,81 log10KbE/g; MIRA-II: Woche 0: 1,38±2,41 log10KbE/g, Woche
12: 1,00±1,91 log10KbE/g, Woche 24: 0,93±1,57 log10KbE/g, Woche 52: 1,98±2,16
log10KbE/g) und Candida spp. (MIRA-I: Woche 0: 1,04±1,50 log10KbE/g, Woche 12:
1,53±1,87 log10KbE/g, Woche 24: 1,16±1,71 log10KbE/g, Woche 52: 0,88±1,49
log10KbE/g; MIRA-II: Woche 0: 0,29±0,83 log10KbE/g, Woche 12: 1,02±1,79
log10KbE/g, Woche 24: 0,66±1,38 log10KbE/g, Woche 52: 0,96±1,92 log10KbE/g)
zeigen innerhalb der Untergruppen keine signifikanten Veränderungen.
0 12 24 520
1
2MIRA -IMIRA -II
Ende derIntervention
Woche
lg(K
eim
zahl
/g)
p=0,026
Abb. 11: Verlauf Keimzahl Candida sp., MIRA-I vs. MIRA-II, Kulturverfahren; x±s
70
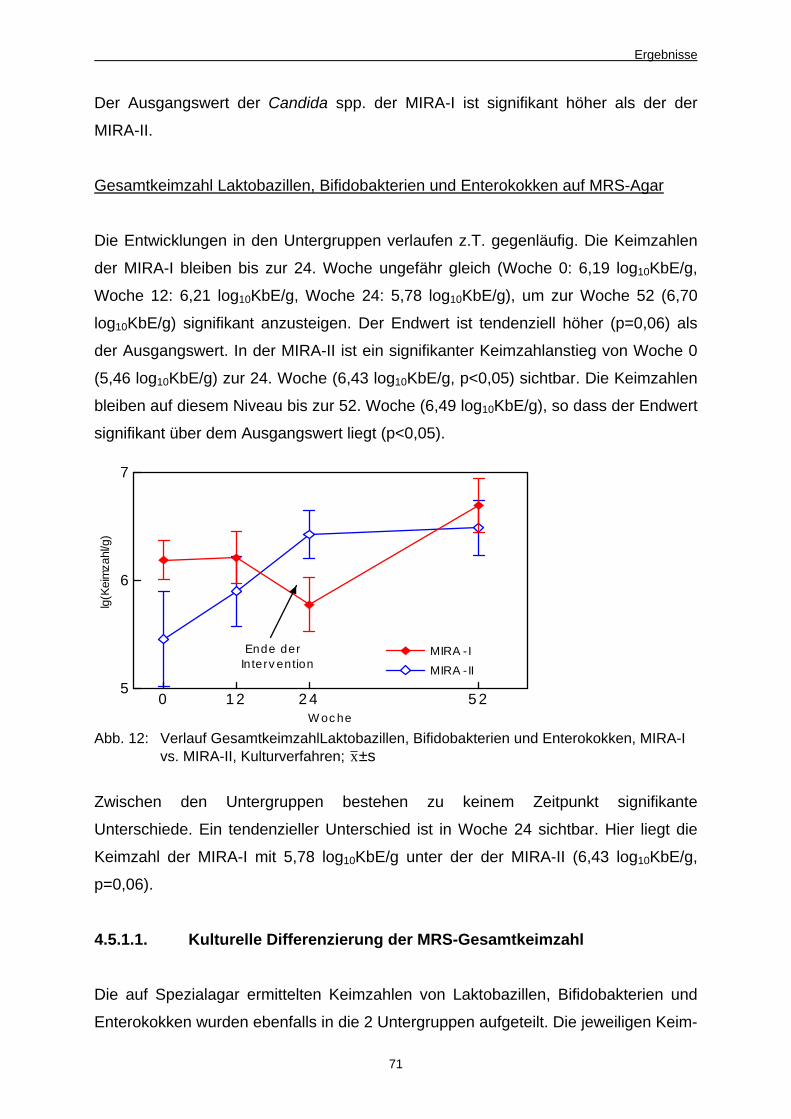
Ergebnisse
Der Ausgangswert der Candida spp. der MIRA-I ist signifikant höher als der der
MIRA-II.
Gesamtkeimzahl Laktobazillen, Bifidobakterien und Enterokokken auf MRS-Agar
Die Entwicklungen in den Untergruppen verlaufen z.T. gegenläufig. Die Keimzahlen
der MIRA-I bleiben bis zur 24. Woche ungefähr gleich (Woche 0: 6,19 log10KbE/g,
Woche 12: 6,21 log10KbE/g, Woche 24: 5,78 log10KbE/g), um zur Woche 52 (6,70
log10KbE/g) signifikant anzusteigen. Der Endwert ist tendenziell höher (p=0,06) als
der Ausgangswert. In der MIRA-II ist ein signifikanter Keimzahlanstieg von Woche 0
(5,46 log10KbE/g) zur 24. Woche (6,43 log10KbE/g, p<0,05) sichtbar. Die Keimzahlen
bleiben auf diesem Niveau bis zur 52. Woche (6,49 log10KbE/g), so dass der Endwert
signifikant über dem Ausgangswert liegt (p<0,05).
0 1 2 2 4 5 25
6
7
MIRA -IMIRA -II
Ende derInterv ention
W oc he
lg(K
eim
zahl
/g)
Abb. 12: Verlauf GesamtkeimzahlLaktobazillen, Bifidobakterien und Enterokokken, MIRA-I vs. MIRA-II, Kulturverfahren; x±s
Zwischen den Untergruppen bestehen zu keinem Zeitpunkt signifikante
Unterschiede. Ein tendenzieller Unterschied ist in Woche 24 sichtbar. Hier liegt die
Keimzahl der MIRA-I mit 5,78 log10KbE/g unter der der MIRA-II (6,43 log10KbE/g,
p=0,06).
4.5.1.1. Kulturelle Differenzierung der MRS-Gesamtkeimzahl
Die auf Spezialagar ermittelten Keimzahlen von Laktobazillen, Bifidobakterien und
Enterokokken wurden ebenfalls in die 2 Untergruppen aufgeteilt. Die jeweiligen Keim-
71
Ergebnisse
zahlen sind Tabelle 25 zu entnehmen. Zwischen beiden Untergruppen werden keine
signifikanten Veränderungen festgestellt.
Tabelle 25: Keimzahlen auf Spezialnährboden ermittelt; in log10KbE/g, darunter die Anteile
in Prozent N Laktobazillen Bifidobakterien Enterokokken
MIRA-I 10 2,46±2,70 (21,71%)
6,87±1,20 (60,71%)
1,99±3,30 (17,58%)
MIRA-II 10 4,49±3,26 (32,41%)
7,39±1,27 (53,31%)
1,98±3,20 (14,28%)
N = Anzahl Proben, Werte dargestellt als Mittelwert ±Standardabweichung
Folgende Korrelationskoeffizienten wurden berechnet:
1) MIRA-I: aGKZ/aK-: r=0,475**
aGKZ/MRS: r=0,574**
2) MIRA-II: aGKZ/aK-: r=0,715**
aGKZ/MRS: r=0,403**
(** = p<0,01, * = p<0,05)
4.5.2 Differenzierung der MRS-Gesamtkeimzahl mit der Fluoreszenz in
situ-Hybridisierung
Die Keimzahlverläufe der einzelnen Untergruppen sind in der Tabelle 26 dargestellt.
Tabelle 26: mittels FISH erhobene Keimzahlen der MIRA-Untergruppen in log10KbE/g; MIRA-I, N=12; MIRA-II, N=9
MIRA-I MIRA-II
Sonde Zeit EUB BIF LAB EUB BIF LAB
Woche 0 x s
10,42± 0,26
7,71± 2,58
6,95± 7,29
10,51± 0,24
8,40± 0,64
7,33± 7,48
Woche 12 x s
10,46± 0,16
7,83± 2,45
7,19± 7,67
10,24± 0,41
7,09± 2,58
7,09± 7,14
Woche 24 x s
10,35± 0,32
6,84± 3,27
6,39± 6,66
10,49± 0,23
7,35± 2,84
7,44± 7,86
Woche 52 x s
10,50± 0,24
7,47± 2,51
6,73± 7,15
10,66± 0,35
7,77± 0,25
7,10 7,43
x= Mittelwert; s = Standardabweichung; EUB = Eubacteria Sondenmischung (s.3.3.1.), BIF = Bifidobacteria-Sonde; LAB = Lactobacillus-Sonde
72
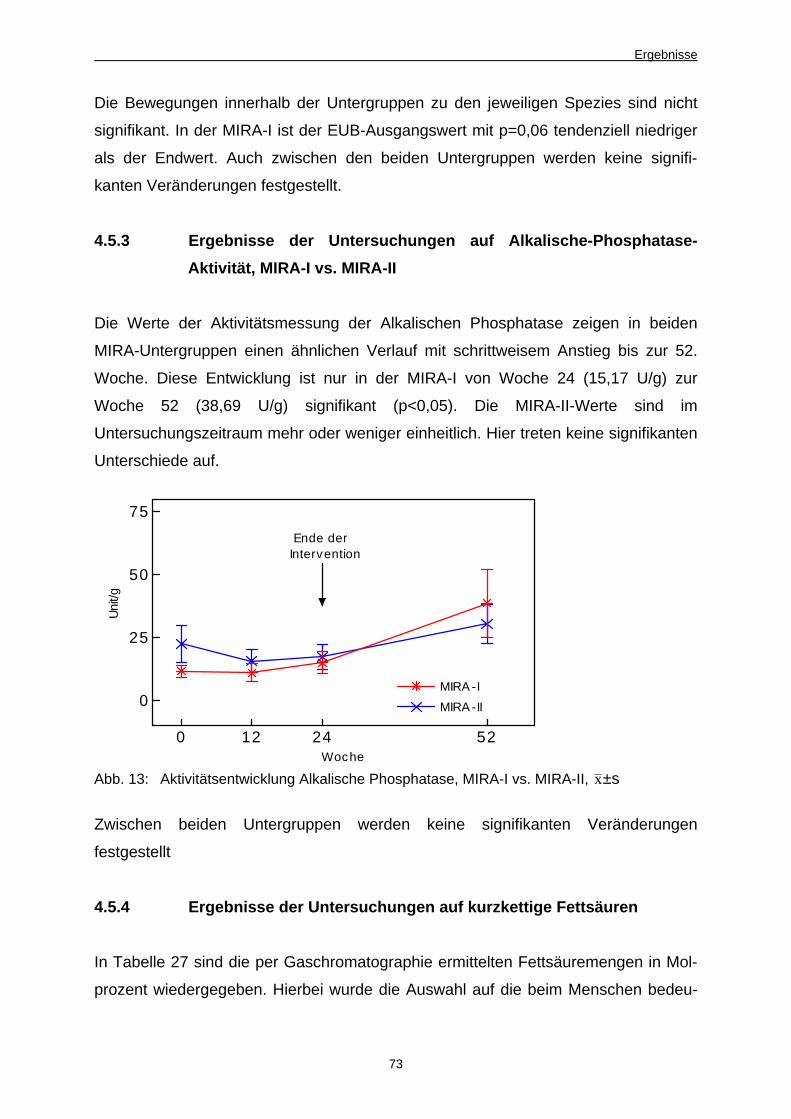
Ergebnisse
Die Bewegungen innerhalb der Untergruppen zu den jeweiligen Spezies sind nicht
signifikant. In der MIRA-I ist der EUB-Ausgangswert mit p=0,06 tendenziell niedriger
als der Endwert. Auch zwischen den beiden Untergruppen werden keine signifi-
kanten Veränderungen festgestellt.
4.5.3 Ergebnisse der Untersuchungen auf Alkalische-Phosphatase-Aktivität, MIRA-I vs. MIRA-II
Die Werte der Aktivitätsmessung der Alkalischen Phosphatase zeigen in beiden
MIRA-Untergruppen einen ähnlichen Verlauf mit schrittweisem Anstieg bis zur 52.
Woche. Diese Entwicklung ist nur in der MIRA-I von Woche 24 (15,17 U/g) zur
Woche 52 (38,69 U/g) signifikant (p<0,05). Die MIRA-II-Werte sind im
Untersuchungszeitraum mehr oder weniger einheitlich. Hier treten keine signifikanten
Unterschiede auf.
0 12 24 52
0
25
50
75
MIRA -IMIRA -II
Ende derIntervention
Woche
Unit/
g
Abb. 13: Aktivitätsentwicklung Alkalische Phosphatase, MIRA-I vs. MIRA-II, x±s
Zwischen beiden Untergruppen werden keine signifikanten Veränderungen
festgestellt
4.5.4 Ergebnisse der Untersuchungen auf kurzkettige Fettsäuren
In Tabelle 27 sind die per Gaschromatographie ermittelten Fettsäuremengen in Mol-
prozent wiedergegeben. Hierbei wurde die Auswahl auf die beim Menschen bedeu-
73
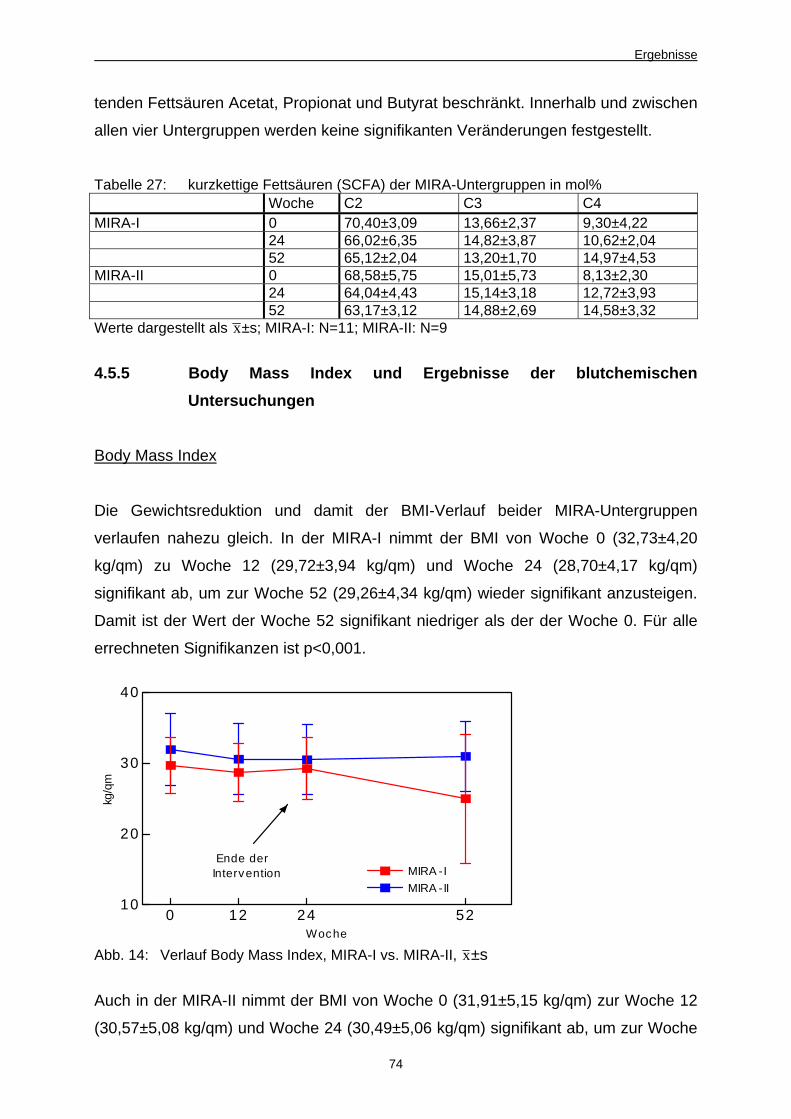
Ergebnisse
tenden Fettsäuren Acetat, Propionat und Butyrat beschränkt. Innerhalb und zwischen
allen vier Untergruppen werden keine signifikanten Veränderungen festgestellt.
Tabelle 27: kurzkettige Fettsäuren (SCFA) der MIRA-Untergruppen in mol% Woche C2 C3 C4 MIRA-I 0 70,40±3,09 13,66±2,37 9,30±4,22 24 66,02±6,35 14,82±3,87 10,62±2,04 52 65,12±2,04 13,20±1,70 14,97±4,53 MIRA-II 0 68,58±5,75 15,01±5,73 8,13±2,30 24 64,04±4,43 15,14±3,18 12,72±3,93 52 63,17±3,12 14,88±2,69 14,58±3,32 Werte dargestellt als x±s; MIRA-I: N=11; MIRA-II: N=9
4.5.5 Body Mass Index und Ergebnisse der blutchemischen Untersuchungen
Body Mass Index
Die Gewichtsreduktion und damit der BMI-Verlauf beider MIRA-Untergruppen
verlaufen nahezu gleich. In der MIRA-I nimmt der BMI von Woche 0 (32,73±4,20
kg/qm) zu Woche 12 (29,72±3,94 kg/qm) und Woche 24 (28,70±4,17 kg/qm)
signifikant ab, um zur Woche 52 (29,26±4,34 kg/qm) wieder signifikant anzusteigen.
Damit ist der Wert der Woche 52 signifikant niedriger als der der Woche 0. Für alle
errechneten Signifikanzen ist p<0,001.
0 12 24 5210
20
30
40
Ende derIntervention MIRA -I
MIRA -II
Woche
kg/q
m
Abb. 14: Verlauf Body Mass Index, MIRA-I vs. MIRA-II, x±s
Auch in der MIRA-II nimmt der BMI von Woche 0 (31,91±5,15 kg/qm) zur Woche 12
(30,57±5,08 kg/qm) und Woche 24 (30,49±5,06 kg/qm) signifikant ab, um zur Woche
74
Ergebnisse
52 (30,94±5,08 kg/qm) wieder signifikant anzusteigen. Damit ist der Wert der Woche
52 signifikant niedriger als der der Woche 0. Für alle errechneten Signifikanzen ist
p<0,001. Zwischen den Untergruppen werden keine signifikanten Veränderungen
festgestellt.
Leptin
Die Leptinwerte der MIRA-I nehmen von Woche 0 zu Woche 12 signifikant ab
(p<0,05). Danach erfolgen keine weiteren signifikanten Veränderungen. Die Werte
der MIRA-II zeigen einen mit p<0,05 signifikanten Anstieg von Woche 0 zu den
Wochen 12 und 24. Der Wert der 52. Woche ist signifikant niedriger als der Wert in
Woche 24. Die Leptinwerte der beiden MIRA-Untergruppen sind in Tabelle 28
dargestellt:
Tabelle 28: Leptin in mmol/l Woche Leptin Woche Leptin MIRA-I 0 25,00±9,11 MIRA-II 0 21,15±12,02 12 20,62±13,76 12 25,35±19,89 24 22,93±17,26 24 27,35±22,80 52 21,17±12,05 52 19,80±11,58 Werte dargestellt als x±s; MIRA-I: N=30; MIRA-II: N=26
Zwischen den Untergruppen werden keine signifikanten Veränderungen festgestellt.
Nachfolgend werden die in Tabelle 29 gemeinsam dargestellten Parameter im
Einzelnen besprochen. Dabei soll nur auf signifikante Entwicklungen eingegangen
werden.
Tabelle 29: Blutparameter der MIRA-Untergruppen; HbA1c in %, Blutzucker und –fette
in mmol/l Woche HbA1c Blut-
zucker Cholesterol gesamt
HDL LDL Triglyce-ride
MIRA-I 0 5,54±0,35 5,09±0,66 5,90±0,79 1,63±0,40 3,56±0,68 1,39±0,59 12 5,74±0,31 4,66±0,40 5,07±0,84 1,38±0,34 3,50±0,81 1,33±0,58 24 5,51±0,30 4,89±0,46 5,22±0,92 1,95±0,49 3,20±0,75 1,32±0,56 52 5,78±0,25 4,43±0,64 5,48±0,86 1,86±0,45 3,36±0,82 1,50±0,62 MIRA-II 0 5,62±0,37 5,13±0,61 5,66±0,91 1,52±0,31 3,44±0,88 1,31±0,60 12 5,79±0,29 4,89±0,54 5,10±0,82 1,40±0,25 3,52±0,77 1,43±0,57 24 5,65±0,29 4,80±0,51 5,41±1,02 1,88±0,41 3,53±1,00 1,46±0,67 52 5,87±0,29 4,57±0,54 5,51±0,98 1,84±0,72 3,63±0,89 1,47±0,66 Werte dargestellt als x±s; MIRA-I, N=30; MIRA-II, N=26
75

Ergebnisse
Hämoglobin A1c
Der Verlauf der HbA1c -Werte der MIRA-I zeigt einen signifikanten Anstieg von
Woche 0 zu Woche 12. Sie fallen in der Woche 24 signifikant auf das Ausgangs-
niveau zurück und steigen zur Woche 52 signifikant zur Woche 24 an. Der
Ausgangswert ist signifikant niedriger als der Endwert (alle p<0,001). Die Werte der
MIRA-II zeigen einen ähnlichen Verlauf. Die HbA1c -Werte signifikant an, um sich
dann in der 24. Woche wieder dem Ausgangswert anzunähern (beide p<0,05). Der
Anstieg von Woche 24 zu Woche 52 ist mit p<0,001 signifikant. Auch hier ist der
Endwert signifikant höher als der Ausgangswert (p<0,001). Zwischen den MIRA-
Untergruppen bestehen zu keinem Untersuchungszeitpunkt signifikante Unter-
schiede.
Blutzucker
Die Blutzuckerwerte der MIRA-I sind in der 12. und 24. Woche (beide p<0,001)
signifikant niedriger als der Ausgangswert, wobei von Woche 12 zur Woche 24 ein
mit p<0,05 signifikanter Anstieg zu verzeichnen ist. Ebenfalls signifikant niedriger ist
der 52-Wochen-Wert gegen die Werte der Wochen 0 und 24 (beide p<0,001). Die
MIRA-II zeigt eine kontinuierliche Abnahme der Blutzucker-Konzentration. Der
Ausgangswert ist zum Wert der Woche 12 mit p<0,05 und zum Wert der Woche 24
mit p<0,001 signifikant. Im weiteren Verlauf vermindert sich der Blutzuckerwert zur
Woche 52 signifikant (p<0,05). Der Ausgangswert liegt signifikant über dem Endwert
(p<0,001). Hier sind die Wertverminderungen von Woche 0 zu den Wochen 12 und
52 signifikant (beide p<0,05). Zwischen den MIRA-Untergruppen bestehen zu keinem
Untersuchungszeitpunkt signifikante Unterschiede.
Gesamt-Cholesterol
Die Gesamt-Cholesterolwerte erreichen in beiden MIRA-Untergruppen in der 12.
Woche ihren niedrigsten Stand. In der MIRA-I ist der Anfangswert zu den Werten der
Wochen 12 und 24 signifikant höher (p<0,001). Der Anstieg von Woche 24 zum
Endwert ist ebenfalls signifikant (p<0,05). Der Anfangswert liegt signifikant (p<0,05)
über dem Endwert. In der MIRA-II sind die Entwicklungen von Woche 0 zur Woche
76

Ergebnisse
12 (p<0,001) und von Woche 12 zur Woche 24 (p<0,05) signifikant . Zwischen den
MIRA-Untergruppen bestehen zu keinem Untersuchungszeitpunkt signifikante
Unterschiede.
High Density Lipoprotein
In der MIRA-I ist der Anfangswert signifikant niedriger als die Werte der Wochen 24
und 52 (beide p<0,001). Lediglich zur Woche 12 besteht ein signifikanter Abfall der
HDL-Werte, die zur Woche 24 wieder signifikant ansteigen (beide p<0,001). In der
MIRA-II erfolgt ein signifikanter Abfall der HDL-Werte (p<0,05), gefolgt von einem
signifikanten Anstieg zur 24. Woche (p<0,05). Der Ausgangswert ist signifikant
niedriger als die Werte der Wochen 24 (p<0,001) und 52 (p<0,05). Zwischen den
MIRA-Untergruppen bestehen zu keinem Untersuchungszeitpunkt signifikante
Unterschiede.
Low Density Lipoprotein
Die LDL-Werte der MIRA-I zeigen einen signifikante Werteabfall von Woche 12 zur
Woche 24. Damit liegt auch der Ausgangswert über dem der 24. Woche (beide
p<0,05). Hier tritt eine signifikante Wertverringerung von Woche 0 zu Woche 24 ein
mit anschließendem signifikanten Anstieg zur Woche 52 (beide p<0,05). In der
MIRA-II ist der Ausgangswert signifikant niedriger als der Endwert (p<0,05).
Zwischen den MIRA-Untergruppen bestehen zu keinem Untersuchungszeitpunkt
signifikante Unterschiede.
Triglyceride
Die Triglycerid-Werte der MIRA-I sind in der 24. Woche signifikant niedriger als in der
52. Woche (p<0,05). In der MIRA-II und zwischen den Untergruppen traten keine
signifikanten Veränderungen auf.
Korrelationen
Es wurden folgenden Korrelationen berechnet:
77

Ergebnisse
1) MIRA-I: BMI/Leptin: r=0,602**
BMI/BZ: r=0,529**
BMI/LDL: r=0,347**
Ges-Chol/LDL: r=0,818**
2) MIRA-II: BMI/Leptin: r=0,579**
BMI/LDL: r=-0,413**
Leptin/LDL: r=-0,417**
Ges-Chol/LDL: r=0,868**
(** = p<0,01, * = p<0,05)
78

Diskussion
5 Diskussion 5.1 Methodenkritik
In der vorliegenden Arbeit wurden folgende Aspekte berücksichtigt und der Schwer-
punkt bei den kulturellen Untersuchungen auf sauerstofftolerante Fäkalkeime, also
Aerobier und fakultative Anaerobier, gesetzt. Zur Untersuchung der Fäkalflora stehen
zahlreiche Methoden zur Verfügung. Die älteste und sehr häufig genutzte Technik ist
das Plattenkulturverfahren, das auch bei den Proben, die für die vorliegende Arbeit
untersucht wurden, Anwendung fand. Es ist eine sehr kostengünstige Methode, die
bei der Untersuchung aerober und fakultativ anaerober Keime ohne großen labor-
technischen Aufwand auskommt und innerhalb 24 Stunden schnelle Ergebnisse
liefert. Zur Untersuchung strikt anaerober Bakterien müssen allerdings eine Anaero-
bierkammer, spezielle Puffer, Anreicherungen und Nährböden zur Verfügung stehen,
um den Verlust vermehrungsfähiger Bakterien zu minimieren. Der Transport der
Proben muss anaerob und ohne Zeitverlust erfolgen und die Probenbearbeitung ist
sehr zeitintensiv, was den technischen Aufwand enorm erhöht. Im Zusammenhang
mit der trotz guter Untersuchungsbedingungen beträchtlichen Zahl nicht darstell-
barer Bakterien (Kleessen et al. 1999) zeigt sich, dass diese Methodik nur begrenzt
zur Flora-Charakterisierung mit unbekannten Keimen und Keimzahlen geeignet ist.
Durch Direktausstrich ohne Probenverdünnung können aber auch Bakterien, die in
geringen Keimzahlen vorkommen, nachgewiesen werden, sofern diese auf den
angebotenen Nährmedien kultivierbar sind. Diese niedrige Nachweisgrenze ist ein
großer Vorteil gegenüber anderen Methoden. Bei der Keimzählung von sporenbil-
denden Bakterien muss die ermittelte Keimzahl auf lebensfähige vegetative Mikro-
organismen bezogen werden. Die eventuell gebildeten Sporen werden nicht
nachgewiesen.
Mit der Fluoreszenz in situ-Hybridisierung können innerhalb von 48 Stunden Ergeb-
nisse ermittelt werden. Die Fluoreszenz-Mikroskopie erfordert kostenaufwändige
technische Voraussetzungen und z.T. teure Arbeitsmaterialien und ist damit nicht
überall anwendbar. Ein weiterer Nachteil ist die relativ hohe Nachweisgrenze, die aus
den Probenverdünnungen und dem Fixationsprozess resultiert (3.3.2) und nur in
Grenzen beeinflussbar ist. Mit der FISH-Technik können Nukleinsäuresequenzen,
also RNA- und DNA-Sequenzen, sichtbar gemacht werden (SWIGER und TUCKER
1996). Demzufolge ist diese Methode nicht auf vermehrungsfähige Bakterien ange-
79

Diskussion
wiesen und liefert im Vergleich mit anderen Untersuchungsmethoden realistische
Ergebnisse. Allerdings können abgestorbene Bakterien, deren RNA bereits durch
entsprechende Enzyme zerstört wurde, nicht nachgewiesen werden. Die Proben der
vorliegenden Arbeit wurden mit dieser Technik untersucht, um kulturell ermittelte
Parameter zu untermauern und in das Verhältnis zur Gesamtkeimzahl (aerob und
anaerob) zu setzen.
5.2 Keimzahlentwicklungen von Fäkalbakterien im Untersuchungszeitraum
5.2.1 Aerobe Gesamtkeimzahl und aerobe Gram-negative Gesamtkeim-zahl im Plattenkulturverfahren
Die Bakterien der aeroben Gram-negativen Gesamtkeimzahl sind in der aeroben
Gesamtkeimzahl enthalten. Das enge Verhältnis beider Parameter zueinander wird
durch die errechneten Korrelationskoeffizienten bestätigt (MIRA-PreCon-Gruppe:
r=0,583**, MIRA-DGE-Gruppe: r=0,597**). Bei den männlichen Teilnehmern der
MIRA-PreCon wird dieses Verhältnis mit r=0,784** noch deutlicher. Die Keimzahl-
verläufe beider Parameter sind erwartungsgemäß sehr ähnlich, daher sollen diese
beiden Parameter hier gemeinsam betrachtet werden.
Die Keimzahl der aeroben Gram-negativen Bakterien lag in den Proben dieser Studie
oft nur geringfügig unter der aeroben Gesamtkeimzahl. Deshalb kann davon ausge-
gangen werden, dass Spezies der Enterobacteriaceae, die als Hauptvertreter der
aeroben Gram-negativen Keimzahl identifiziert wurden, einen Großteil der aeroben
Gesamtkeimzahl ausmachen.
Mit den aeroben Gesamtkeimzahlen wurde ein äußerst geringer, aber bedeutender
Teil der Fäkalflora erfasst. Dieser Teil wird auch als „Begleitflora“ mit einem Anteil
von nur maximal 1% (ROLLE und MAYR 2002) bezeichnet. Die Bedeutung geht aus
mehreren Studien hervor. RUSCH und RUSCH (2001) stellen im Zusammenhang mit
einer Eignungsprüfung verschiedener Bakterien für die Verwendung zur Herstellung
von Autovaccinen fest, dass E.coli trotz der geringen Keimzahlen einen beachtlichen
bakteriziden Effekt erzielte. Das Serum von mit Kolibakterien besiedelten Tieren
hatte eine deutlich stärker abtötende Wirkung auf Salmonella Typhimurium als das
Serum von Tieren, die mit Laktobazillen oder Bifidobakterien besiedelt waren. Weiter
80

Diskussion
betonen die Autoren die umfassende Stimulierung der Immunzellen durch E.coli.
Durch Laktobazillen beispielsweise werden die T- und B-Zellen nicht aktiviert.
Die aeroben Gesamtkeimzahlen weisen die deutlichsten Veränderungen im Untersu-
chungszeitraum auf. Mit Beginn der Ernährungsumstellung in beiden Gruppen fallen
die hier untersuchten Keimzahlen bis zum Ende der massiven Intervention in der 24.
Woche signifikant ab (MIRA-PreCon-Gruppe: p<0,001; MIRA-DGE-Gruppe: p<0,05).
Das könnte auf ein reduziertes Nahrungsangebot und eine Veränderung des Kolon-
milieus, z.B. vermehrtes Wachstum einer anaeroben Spezies mit gleichzeitiger Hem-
mung der fakultativen Anaerobier, hindeuten. In Betracht kommen Laktobazillen und
Bifidobakterien. Nach Untersuchungen von BUTEL et al. (1998) und LIEVIN et al.
(2000) dominieren diese vor allem die Kolonflora durch ihre herausragende Fähig-
keit, am Mukus zu adhärieren und bakterizide Substanzen zu bilden. Des Weiteren
bestimmen sie den intestinalen pH-Wert mit durch die Bildung von Laktat.
Die Teilnehmer der MIRA-DGE-Gruppe nahmen in diesem Zeitraum deutlich mehr
Ballaststoffe zu sich als vor der Studie. Der Fettanteil der Nahrung sollte auf ein Mini-
mum begrenzt und die Aufnahme von leicht verdaulichen Kohlenhydraten beschränkt
werden. Die Teilnehmer der MIRA-PreCon-Gruppe nahmen bis zur 12. Woche
nahezu ausschließlich eine hypokalorische Nahrungsergänzungskost auf Milchei-
weißbasis zu sich (SCHOLZ et al. 2002) und stellten die Ernährung dann bis zur 24.
Woche schrittweise auf Mischkost nach den DGE-Richtlinien um. Der durchschnitt-
liche Ausgangswert der aeroben Gesamtkeimzahl stimmt mit ca. 107 KbE/g mit den
Werten von TANNOCK et al. (2000) überein und kann als Normalwert angesehen
werden.
Der signifikant stärkere Abfall der aeroben Gesamtkeimzahl der MIRA-PreCon-
Gruppe bis zur 24. Woche (von 7,06±1,69 auf 5,82±1,20 log10KbE/g; p<0,001) zeigt
einen größeren Einfluss der Ernährung. Allerdings sollte die unter Umständen
bessere Compliance dieser Gruppe gegenüber der MIRA-DGE-Gruppe und ihre
Auswirkung auf die ermittelten Ergebnisse nicht außer Acht gelassen werden. Von
der 24. bis zur 52. Woche sollten sich beide Gruppen gleichermaßen nach den DGE-
Richtlinien ernähren. Die in der MIRA-DGE-Gruppe nahezu gleichbleibenden Keim-
zahlen der fakultativen Anaerobier in diesem Zeitraum (Woche 24: aGKZ 6,63±1,29,
aK- 4,34±2,20; Woche 52: 6,76±1,10, aK- 5,58±1,32 log10KbE/g) lassen darauf
schließen, dass die in den ersten 24 Wochen umgestellte Ernährung beibehalten
wurde. Wenn die MIRA-DGE-Gruppe als Kontrollgruppe und die MIRA-PreCon-
81

Diskussion
Gruppe als Versuchsgruppe betrachtet werden, sollten sich bei gleicher Ernährung
die hier untersuchten Keimzahlen einander annähern. Tatsächlich unterscheiden sich
in der 52. Woche die aeroben Gesamtkeimzahlen und die aeroben Gram-negativen
Keimzahlen beider Gruppen nur unwesentlich voneinander (MIRA-PreCon-Gruppe:
Woche 24: aGKZ 5,82±1,20, aK- 3,91±1,74, Woche 52: aGKZ 6,78±1,37, aK-
5,44±1,79; MIRA-DGE-Gruppe: Woche 24: aGKZ 6,63±1,29, aK- 4,34±2,20; Woche
52: 6,76±1,10, aK- 5,58±1,32 log10KbE/g).
Die aerobe Gram-negative Keimzahl, in dieser Studie überwiegend aus den Entero-
bacteriaceae zugehörigen Spezies bestehend, zeigen zwischen den Gruppen zu
keinem Zeitpunkt signifikante Unterschiede. Demzufolge sind für die signifikant
stärkere Verminderung der aeroben Gesamtkeimzahl der MIRA-PreCon-Gruppe in
der 24. Woche andere Mikroorganismen verantwortlich, die nicht näher identifiziert
wurden. Da diese Keime auf MacConkey-Agar nicht gewachsen sind, kann man auf
Gram-positive Bakterien schließen. Nach SONNENBORN und GREINWALD (1991)
kommen aerobe Mikrokokken, gram-positive Aerobier wie Staphylokokken, Bacillus
spp. und andere in Frage. Im zeitlichen Verlauf sinken die aeroben Gram-negativen
Keimzahlen beider Gruppen zur 24. Woche hin signifikant ab, um sich bis zur 52.
Woche auf ein tendenziell niedrigeres Ausgangsniveau zurückzubewegen (MIRA-
PreCon-Gruppe: Woche 0: aGKZ 7,06±1,69, aK- 5,77±1,87, Woche 24: aGKZ
5,82±1,20, aK- 3,91±1,74, Woche 52: aGKZ 6,78±1,37, aK- 5,44±1,79; MIRA-DGE-
Gruppe: Woche 0: aGKZ 7,58±1,20, aK- 6,04±1,39, Woche 24: aGKZ 6,63±1,29, aK-
4,34±2,20; Woche 52: 6,76±1,10, aK- 5,58±1,32 log10KbE/g). Somit beeinflussen
beide Ernährungsumstellungen diese Parameter in beiden Gruppen, wenn auch in
ungleichem Ausmaß. Es wird aber auch deutlich, dass dieser Einfluss nicht dauerhaft
ist, sondern mit zunehmender Eigenverantwortung der Probanden nachlässt. Ein
möglicher Einfluss wäre das verstärkte Wachstum von Bakterienspezies, die Bakte-
riozine produzieren und gram-negative und gram-positive Keime hemmen können.
Von den in dieser Arbeit untersuchten Keimen kommen Bifidobakterien, Entero-
kokken und Laktobazillen in Frage. Durchgeführte Berechnungen ergeben signifikant
positive Korrelationen zwischen der Gesamtzahl der MRS-Bakterien und der aeroben
Gesamtkeimzahl sowie der aeroben Gram-negativen Gesamtkeimzahl. Die Keimzah-
len der Bifidobakterien nehmen zur 24. Woche hin leicht ab und steigen zum Stu-
dienende wieder an. Daher sind Einflüsse auf die aerobe Fäkalflora von dieser Seite
her möglich.
82

Diskussion
Der Einfluss beider Ernährungsweisen auf die aerobe Gesamtkeimzahl und die
aerobe Gram-negative Gesamtkeimzahl ist nachweislich vorhanden und besonders
bei der aeroben Gesamtkeimzahl je nach Ernährungsweise unterschiedlich stark
ausgeprägt. Die Auswertung der nach Geschlecht getrennten Gruppen innerhalb und
untereinander brachte keine neuen Ergebnisse. Besonders die weibliche Fraktion
spiegelte die Ergebnisse der Gesamtgruppen wieder, da die Gesamtgruppen weib-
lich dominiert waren. Daher wird auf diese Ergebnisse auch im Folgenden nicht
weiter eingegangen.
Die Unterteilung der Hauptgruppen nach BMI-Entwicklung zeigt bei den hier bespro-
chenen Parametern Keimzahlverläufe, die denen der jeweiligen Hauptgruppen
ähneln. Die bei den Teilnehmern mit geringer Gewichtsreduktion in der MIRA-II ver-
muteten „Ausreißer“, also vom Verlauf der Hauptgruppen stark abweichende Werte,
wurden nicht gefunden. Zwischen den Untergruppen gibt es im Gegensatz zu den
Hauptgruppen im gesamten Verlauf keine signifikanten Unterschiede. Sie verlaufen
erstaunlich ähnlich trotz der unterschiedlichen Gewichtsreduktion der Teilnehmer.
Die Keimzahlentwicklungen von Woche 24 zur Woche 52 sind in der MIRA-PreCon
gleich der MIRA-I wie auch in der MIRA-DGE gleich der MIRA-II. Das unterstreicht
die stärkere Gewichtsreduktion der Teilnehmer der MIRA-PreCon. Signifikante Unter-
schiede zwischen Ausgangs- und Endwerten kommen in MIRA-I und II nicht vor.
Lediglich eine Tendenz mit p=0,05 ist in der MIRA-I zu erkennen. Diese Angleichung
der Entwicklungen ist der Vermischung der Teilnehmer geschuldet und weist auf
einen stärkeren Einfluss der Ernährung auf die aerobe Gesamtkeimzahl im Gegen-
satz zum Einfluss des BMI hin.
Auch bei der aeroben Gram-negativen Gesamtkeimzahl ist die Dynamik der Unter-
gruppen der jeweiligen Hauptgruppe ähnlich. Alle Keimzahl-Bewegungen entspre-
chen denen der Hauptgruppen und zeigen keinen Unterschied bei BMI- und Ernäh-
rungseinfluss. Die enge Beziehung zwischen aerober Gesamtkeimzahl und aerober
Gram-negativer Gesamtkeimzahl ist in den Untergruppen ebenfalls deutlich. Der
Korrelationskoeffizient in der MIRA-I ist r=0,475** und in der MIRA-II r=0,715**.
Die Ergebnisse der Hauptgruppen MIRA-PreCon und MIRA-DGE konnten durch die
Auswertung der Untergruppen grundsätzlich bestätigt werden. Es wird deutlich, dass
die Teilnehmer mit guter und sehr guter Gewichtsreduktion (MIRA-I) wie auch die
Teilnehmer der MIRA-PreCon die eindrücklichsten Keimzahldynamiken aufweisen.
Das zeigt den Zusammenhang zwischen Gewichtsreduktion und Ernährungsweise
83

Diskussion
und spricht für einen stärker floramodulierenden Effekt der Zusatzkost der MIRA-
PreCon und für die konsequente Umsetzung und Einhaltung der Ernährungsvor-
schriften.
5.2.2 Candida spp. und Clostridium perfringens im Plattenkultur- verfahren
Ein Einfluss der jeweiligen Kost auf die Sprosspilze, hier speziell untersucht Candida
spp., lässt sich nicht eindeutig erkennen. Die ermittelten Keimzahlen sind sehr unein-
heitlich, weshalb die Gruppenwerte starke Streuungen aufweisen. Diese Varianz ver-
hindert trotz ungleicher Keimzahldynamik in den Gruppen signifikante Gruppenunter-
schiede. Die Candida spp.-Keimzahlen der MIRA-PreCon-Gruppe zeigen keine
größeren Schwankungen (Woche 0: 0,84±1,42, Woche 24: 0,98±1,60, Woche 52:
0,91±1,60 log10KbE/g) und lassen daher keinen Einfluss erkennen. Die MIRA-DGE-
Gruppe startete in Woche 0 mit niedrigeren Keimzahlen (0,40±0,92 log10KbE/g) und
zeigt einen signifikanten Anstieg zur 24. Woche (1,21±1,78 log10KbE/g, p<0,05).
Diese durchschnittliche Keimzahl ist mit unter 100 KbE/g immer noch sehr niedrig
und sollte daher nicht überbewertet werden. Es ist erkennbar, dass Candida spp. und
andere Sprosspilze zur physiologischen Magen-Darm-Flora gehören. Nach AUTEN-
RIETH (2000) kommen Candida spp. in Keimzahlen bis 103 KbE/g im physiolo-
gischen Dickdarm vor. Diese Angabe geht konform mit ROLLE und MAYR (2002),
die Hefen und auch Clostridium spp. als „Restflora“ mit einem Anteil unter 0,01%
bezeichnen. Nach einer Antibiotika-Therapie, bei Immunschwäche und Stoffwechsel-
störungen können Candida spp. im jeweiligen Habitat ansteigen und ein eigenes
Krankheitsbild (Candidose) verursachen. Häufige, wenn auch beim Menschen selten
auf Hefen zurückgeführte Erkrankungen sind Durchfall, Pneumomykosen und Soor.
Die Candida spp.-Keimzahlen beider MIRA-Gruppen bewegen sich über den gesam-
ten Untersuchungszeitraum im Normalbereich.
Clostridium perfringens wird bei DRASAR et al. (1976) als ubiquitäre Spezies bezei-
chnet. Aus den Proben beider MIRA-Gruppen wurden regelmäßig vegetative Formen
von Clostridium perfringens isoliert. Unabhängig von Zeitraum und Ernährungsre-
gime war in einigen Proben kein Nachweis möglich. Hier kann die Nachweisgrenze
von 103 log10KbE/g unterschritten worden sein oder die vorhandenen Keime sind von
der vegetativen Form in die sporulierte übergetreten und waren daher mit der kultu-
84

Diskussion
rellen Methode nicht nachweisbar. Die durchschnittlichen Clostridium spp.-Keim-
zahlen liegen bei FINEDOLD et al. (1983) zwischen 107 log10KbE/g bei fisch- und
fleischreicher Ernährung und nicht nachweisbar bei strikt vegetarischer Diät. Höhere
Clostridium spp.-Keimzahlen hängen daher offensichtlich mit der vermehrten Aufnah-
me tierischer Proteine zusammen. Clostridium perfringens allerdings fermentieren
überwiegend Kohlenhydrate (Zucker, Stärke, Pektin) unter Bildung volatiler Fett-
säuren, CO2, H2 und Ethanol (MADIGAN et al. 2001). Eine Veränderung der Keim-
zahl durch proteinreiche Ernährung war daher nur in geringem Umfang zu erwarten.
Die ermittelten durchschnittlichen Clostridium perfringens- Keimzahlen der MIRA-
PreCon- und MIRA-DGE-Gruppe liegen im gesamten Verlauf unter 100 KbE/g, was
der Zuordnung zur „Restflora“ entspricht. Veränderungen der beiden untersuchten
Parameter sind nicht zu erkennen, wofür starke Schwankungen der ermittelten Keim-
zahlen eine Ursache sind.
Die Auswertung der MIRA-Untergruppen bringt keine neuen Erkenntnisse. Die durch-
schnittlichen Candida spp.-Keimzahlen der Teilnehmer beider Untergruppen bewe-
gen sich zwischen 0 und 102 KbE/g ohne signifikante Differenzen. Die Ausgangs-
keimzahl der MIRA-I (1,04±1,50 log10KbE/g) ist signifikant höher als die der MIRA-II
(0,29±0,83 log10KbE/g). In der 12. Woche aber sind die Unterschiede nicht mehr
signifikant, so dass von einer Keimzahlangleichung beider Gruppen ausgegangen
werden kann, die nicht sichtbar durch die Ernährung beeinflusst wird. Die Clostridium
perfringens-Keimzahlen sind in beiden Untergruppen zu keinem Zeitpunkt signifikant.
Auf diese beiden Parameter hatte die aufgenommene Kost offenbar keine feststell-
baren Auswirkungen. Die Keimzahlen bewegen sich über den gesamten Untersu-
chungszeitraum im Normalbereich. Dies ist aber auch ein Hinweis auf die offenbar
intakte Darmflora der Probanden, da die potenziellen Krankheitserreger auf die
physiologische Menge begrenzt bleiben.
5.2.3 MRS-Gesamtflora (Bifidobakterien, Laktobazillen, Enterokokken)
Signifikant im Keimzahlverlauf der MIRA-Hauptgruppen PreCon und DGE ist nur der
Unterschied zwischen den Gruppen in der 24. Woche. Zu diesem Zeitpunkt liegen
die Keimzahlen der MIRA-DGE signifikant über denen der MIRA-PreCon. Innerhalb
der Gruppen gibt es im Verlauf der Keimzahlen keine signifikanten Unterschiede, so
85

Diskussion
dass die Werte im gesamten Studienverlauf als einheitlich betrachtet werden
müssen.
In beiden MIRA-Untergruppen entwickeln sich die MRS-Gesamtkeimzahlen ähnlich.
Es gibt zwischen ihnen keine signifikanten Differenzen. Lediglich ein mit p=0,06 ten-
denzieller Unterschied in der 24. Woche ist sichtbar. Hier ist die Keimzahl in der
MIRA-II mit 6,43 log10KbE/g tendenziell höher als die der MIRA-I mit 5,78 log10KbE/g.
Dieses Ergebnis ist dem der Hauptgruppen sehr ähnlich. Der Keimzahlunterschied in
der 24. Woche ist zwischen der MIRA-PreCon- und der MIRA-DGE-Gruppe signifi-
kant. Der in der MIRA-I von Woche 24 zur Woche 52 und in der MIRA-II von Woche
0 zur Woche 24 signifikante Anstieg ist in den MIRA-Hauptgruppen als Tendenz
sichtbar, aber nicht signifikant. Sowohl in den Haupt- als auch in den Untergruppen
korreliert die MRS-Gesamtkeimzahl mit der aeroben Gesamtkeimzahl. Offenbar
waren Vertreter beider Parameter auf den Nährböden vertreten und sind in die
Gesamtkeimzahlen mit aufgenommen worden.
5.2.3.1 Kulturelle Differenzierung der MRS-Gesamtflora
Die in den Untersuchungen zu dieser Arbeit ermittelten Anteile der Laktobazillen,
Bifidobakterien und Enterokokken an den MRS-Gesamtwerten liegen bei durch-
schnittlich 55,5% für Bifidobakterien, 27,7% für Laktobazillen und 16,8% für Entero-
kokken. Alle Ergebnisse stammen von Stichproben (N=22), die Proben aus allen
Zeitabschnitten und beiden MIRA-Gruppen enthalten. Die Keimzahlen der MIRA-
PreCon (N=13) und der MIRA-DGE (N=9) unterscheiden sich nicht signifikant. Diese
Werte sind etwas niedriger als die von KLEESSEN et al. (1997) mit der gleichen
Methode ermittelten Ergebnisse, die Verhältnisse der Spezies zueinander sind aber
ähnlich. Die Anteile an Bifidobakterien werden bei KLEESSEN et al. (1997) mit
63,27 %, an Laktobazillen mit 31,71 % und an Enterokokken mit 5,03 % ausgewie-
sen. Die Probanden waren ältere Menschen mit einem Altersdurchschnitt von 76,4
Jahren, daher sind die Prozentwerte mit den in der vorliegenden Arbeit errechneten
gut zu vergleichen. Das Durchschnittsalter der MIRA-Teilnehmer liegt bei den Frauen
bei 51±10 Jahren und bei den Männern bei 54±8 Jahren, womit die Probanden,
deren Proben für die vorliegende Arbeit untersucht wurden, etwas jünger als die von
KLEESSEN et al. (1997) untersuchten sind. Dieser geringe Altersunterschied kann
die Ergebnisse jedoch kaum beeinflusst haben. Gründe für die in dieser Arbeit
86

Diskussion
ermittelten niedrigeren Werte können zum einen die Mischung der Stichprobe aus
allen Zeitabschnitten der Studie und zum anderen die Kultivierung aus eingefrore-
nem Probenmaterial sein. Bei KLEESSEN et al. (1997) wurde mit frisch entnommen-
em Material gearbeitet, was Keimverluste entsprechend minimiert.
Die Unterteilung der MIRA-Hauptgruppen nach dem BMI in die MIRA-I und MIRA-II
ergab keine neuen Erkenntnisse gegenüber den bisher vorliegenden Ergebnissen.
Es werden keine signifikanten Unterschiede zwischen den Untergruppen zu den
einzelnen Bakterienspezies beobachtet. Die Keimzahlen sind in beiden Unter-
gruppen einheitlich und die prozentualen Werte unterscheiden sich nicht maßgeblich
von den bei KLEESSEN et al. (1997) benannten. Auch hier muss wie bei der Aus-
wertung der Hauptgruppen kritisch angemerkt werden, dass die Gruppenbildung
unter Berücksichtigung der Zeitpunkte nicht möglich war und eine exakte Auswertung
dieser Parameter erschwert. Es kann nur festgestellt werden, dass die Laktobazillen-,
Bifidobakterien- und Enterokokken-Keimzahlen wie in den MIRA-Hauptgruppen im
physiologischen Bereich liegen.
5.2.3.2 Differenzierung der MRS-Gesamtflora durch Fluoreszenz in situ- Hybridisierung
Die unter Anwendung der FISH-Technik ermittelten Keimzahlen sind ebenfalls aus
Stichproben ermittelt, konnten aber im Gegensatz zu den Werten der kulturellen
Differenzierung in den Gruppen im zeitlichen Versuchsverlauf betrachtet werden. Bei
den Keimzahldynamiken beider MIRA-Hauptgruppen und der Untergruppen MIRA-I
und MIRA-II wurden keine signifikanten Unterschiede innerhalb der Gruppen im
Verlauf und zwischen den Gruppen ermittelt. Die Tendenzen zeigen in beiden MIRA-
Hauptgruppen bei den Bifidobakterien eine Verminderung der Keimzahlen bis zur 24.
Woche und einen Anstieg bis zur 52. Woche (MIRA-PreCon-Gruppe: Woche 0:
7,84±2,33, Woche 24: 7,10±3,00, Woche 52: 7,54±2,26; MIRA-DGE-Gruppe: Woche
0: 8,46±0,50, Woche 24: 7,34±2,97, Woche 52: 8,04±0,76 log10 KbE/g). Ähnliche
tendenzielle Entwicklungen gibt es auch bei der Laktobazillen- und Enterokokken-
Keimzahl (LAB). Da vor allem die Bifidobakterien im Dickdarm auf fermentierbare
Kohlenhydrate angewiesen sind, enthalten vermutlich die Diäten beider MIRA-
Gruppen zu wenig Ballaststoffe, die das Kolon erreichen und den Bakterien als
Energiequelle dienen könnten, um eine signifikante Keimzahlsteigerung zu bewirken.
87

Diskussion
Das ist zumindest bei den Teilnehmern der MIRA-DGE-Gruppe unerwartet, da die
Ernährungsempfehlungen der DGE eine ballaststoffreiche Basis nahe legen. Hier
nimmt möglicherweise wieder die mangelnde Compliance einiger Teilnehmer der
MIRA-DGE-Gruppe Einfluss, indem die DGE-Richtlinien nur begrenzt umgesetzt
wurden. Andererseits wird das Wachstum von Bifidobakterien vor allem durch FOS
beeinflusst (DELZENNE und ROBERFROID 1994, GIBSON et al. 1995, GIBSON
1999, KRUSE et al. 1999). Diese FOS waren in den Diäten beider MIRA-Gruppen
offensichtlich nur gering enthalten. Die Zusatzkost der MIRA-PreCon-Gruppe enthielt
überwiegend leichtverdauliche Zucker, so dass ein positiver Einfluss vor allem auf die
hier untersuchten Bifidobakterien nicht zu erwarten war. Das wird durch die vorlie-
genden Ergebnisse bestätigt.
Laktobazillen kommen im menschlichen Darmtrakt vor allem im Dünndarm und dort
zum großen Teil wandständig vor (ZOETENDAL et al. 2002). Demzufolge sind die
Keimzahlen der aus den Fäzes isolierten Laktobazillen nur bedingt geeignete Para-
meter für die Gesamtkeimzahl-Bestimmung und für eine Dynamikbeurteilung unter
Ernährungseinfluss, da sie eine Mischung aus Keimen lumenständiger Spezies und
abgeschilferten Bakterien wandständiger Spezies in Dünndarm und Kolon darstellt.
Weiterhin kommt es häufig zu einem Wechsel der ansässigen Spezies und Verschie-
bung der jeweiligen Anteile innerhalb der Gesamtzahl (KIMURA et al. 1997; HOP-
KINS 2001). Diese wurden in der vorliegenden Arbeit nicht untersucht. Die Laktoba-
zillen-Gesamtkeimzahlen bei KIMURA et al. variierten stark von 104 bis 108 KbE/g.
Auch konnten Laktobazillen nicht in allen Stuhlproben isoliert werden. Damit stimmen
die entsprechenden Ergebnisse dieser Arbeit überein.
Vor allem die mittels FISH-Technik erstellten Bifidobakterien-Keimzahlen der MIRA-
Studie liegen um den Faktor 10-100 niedriger als in vergleichbaren Studien (TAN-
NOCK et al. 2000; MARTEAU et al. 2001). Bei beiden Autoren wurde Probenmaterial
von überwiegend jungen Teilnehmern untersucht. Bei MARTEAU et al. (2000) waren
die Teilnehmer 22-27 Jahre alt und bei TANNOCK et al. (2000) umfasste die Alters-
spanne 25-55 Jahre. Aus der Literatur ist bekannt, dass die Keimzahlen besonders
der Bifidobakterien, aber auch der Laktobazillen und die Gesamtzahl der anaeroben
Keime mit steigendem Lebensalter abnehmen und eine verminderte Spezies-
diversität autritt (Kleessen et al. 1997, HOPKINS et al. 2001, HOPKINS und
MACFARLANE 2002) Die von den genannten Autoren angewendeten
Untersuchungsmethoden unterscheiden sich zum Teil von denen der vorliegenden
88

Diskussion
Arbeit, so dass die dort ermittelten Keimzahlen nur bedingt vergleichbar sind. Die
Altersunterschiede zu den in dieser Arbeit untersuchten Gruppen werden als ein
Grund für die festgestellten Keimzahlunterschiede zu den Ergebnissen der oben
angeführten Autoren angenommen. Dazu kommen Lagerungsverluste, da die
Proben vor ihrer Untersuchung mit der FISH-Technik bereits tiefkühlgelagert waren.
Die Auswertung der FISH-Keimzahlen der MIRA-Untergruppen ergab keine von den
Hauptgruppen abweichenden Ergebnisse. Die Keimzahlen der einzelnen Parameter
unterliegen lediglich tendenziellen Schwankungen. Auch zwischen den Untergruppen
gibt es keine signifikanten Unterschiede.
Die hier untersuchten milchsäurebildenden Bakterien (Laktobazillen und Entero-
kokken, Bifidobakterien) weisen weder im Studienverlauf der einzelnen MIRA-
Gruppen noch zwischen den Gruppen signifikante Keimzahländerungen auf zu den
einzelnen Beprobungszeitpunkten. Die Kost der Probanden beider MIRA-Gruppen
beinhaltete nicht zwingend Substrate, die bekannterweise diese Bakterienspezies in
Wachstum und Stoffwechselaktivität stimulieren. Somit ist eine direkte Auswirkung
der aufgenommenen Nahrung auf dieses Keimspektrum nicht eindeutig nachweisbar.
Belegt wird die ausgesprochene Stabilität der untersuchten anaeroben Flora. Dass
die Schwankungen der aeroben Gesamtkeimzahl und der aeroben Gram-negativen
Gesamtkeimzahl hier nicht sichtbar werden, kann daran liegen, dass diese
Keimgruppen nur einen sehr geringen Anteil von ca. 0,01% an der Gesamtflora
haben.
5.3 Alkalische Phosphatase
In der vorliegenden Arbeit wurde die fäkale Gesamt-AP über eine photometrisch
gemessene Substratumsetzung bestimmt. Die MIRA-DGE-Gruppe begann die Studie
mit signifikant höheren AP-Werten als die MIRA-PreCon-Gruppe. Die nachfolgenden
Werte der MIRA-DGE-Gruppe zu den Untersuchungszeitpunkten waren signifikant
höher als die der MIRA-PreCon-Gruppe und zeigten im Verlauf eine tendenzielle
Steigerung bis zur 52. Woche. Diese Steigerung ist aufgrund der außerordentlich
starken Streuung der Werte nicht signifikant. Die Durchschnittswerte lagen bei der
MIRA-DGE-Gruppe mit 27,8±26,3 Unit/g in Woche 0 und mit 57,07±90,94 Unit/g in
Woche 52 unterhalb des von LEHMANN und HILLERT (1980) bei 71 gesunden Pro-
banden mit 60,80 Unit/g angegebenen Mittelwertes der fäkalen Gesamt-AP. Wie an
89

Diskussion
der Standardabweichung sichtbar wird, liegen die Werte vieler MIRA-Teilnehmer
höher und nähern sich damit dem genannten Normwert. Die Durchschnittswerte der
MIRA-PreCon-Gruppe liegen dagegen mit 10,0±8,6 Unit/g in Woche 0 und mit 24,0±
34,1 Unit/g in Woche 52 deutlich unter dem genannten Normalwert. Die Gesamt-AP-
Werte der einzelnen bei LEHMANN und HILLERT (1980) untersuchten gesunden
Probanden waren sehr unterschiedlich, so dass die dortige aus 71 Personen beste-
hende Kontrollgruppe eine große Streuung aufweist. Gleiches wurden auch bei
weiteren Untersuchungen festgestellt (HORRIGAN und DANNOVITCH 1974; LEH-
MANN und ZENGEL 1980). Insofern ist die niedrige AP-Gesamtaktivität der MIRA-
PreCon-Gruppe nicht zwangsläufig als das Ergebnis einer kollektiven Darmerkran-
kung zu werten, zumal über den gesamten Studienzeitraum keine ernsthaften
Beschwerden gemeldet wurden.
Die AP-Werte beider MIRA-Untergruppen bleiben ungefähr auf einem Niveau.
Zwischen und auch innerhalb der MIRA-Untergruppen bestehen im Gegensatz zu
den Hauptgruppen keine signifikanten Unterschiede. Die Ergebnisse der MIRA-
Untergruppen bestätigen zwar die Entwicklungen in den MIRA-Hauptgruppen, eine
Verdeutlichung der jeweiligen Aktivitätynamik entstand aber nicht. Methodische
Probleme sind Gründe für die ermittelten niedrigen Werte und die uneindeutige
Dynamik. Aufgrund einer Methodenumstellung wurden die Proben neu bearbeitet
und mussten dafür aufgetaut werden, was zu dem Aktivitätsverlust geführt haben
kann. In diesem Fall gibt die gemessene Aktivität der fäkalen Gesamt-AP daher
wenig Aufschluss über den Gesundheitszustand der Untersuchten oder über die
Herkunft weiterer Einflussfaktoren. Die somit bestehende Heterogenität der Mess-
werte beider MIRA-Gruppen erschweren die Interpretation der Ergebnisse. Um die
jeweiligen Quellen der fäkalen Gesamt-AP gesondert betrachten zu können, muss
sie in ihre einzelnen Isoenzyme aufgeschlüsselt werden.
5.4 Kurzkettige Fettsäuren
Die hauptsächlich im menschlichen Kolon gebildeten Fettsäuren sind Acetat, Propio-
nat und Butyrat. Es werden pro Tag ca. 200-400 mmol dieser SCFA gebildet. Das
Mengenverhältnis im gesunden Kolon wird mit Acetat zu Propionat zu Butyrat wie
3:1:1 oder konkret mit 65/18/11 Prozent (RASSMUSSEN et al. 1988) angegeben.
TANNOCK et al. (2000) untersuchten den Einfluss eines probiotischen Produktes
90

Diskussion
u.a. auf die SCFA-Bildung. Die dort ermittelten durchschnittlichen Ausgangswerte
von 10 Probanden (in mmol/kg) sind für Acetat 58,04±12,28, für Propionat 17,71±
5,97 und für Butyrat 19,46±6,89. Die in der vorliegenden Arbeit gemessenen SCFA-
Mengen geben wie die Werte von TANNOCK et al (2000) das von RASSMUSSEN et
al. (1988) genannte Verhältnis der Fettsäuren zueinander wieder.
Die Ausgangswerte der MIRA-PreCon-Gruppe (in µmol/g) mit Acetat 73,57±22,36,
Propionat 14,38±6,11 und Butyrat 10,33±8,02 und der MIRA-DGE-Gruppe (in µmol
/g) mit Acetat 69,34±23,14, Propionat 16,74±11,73 und Butyrat 8,24±3,33 sind den
bei RASSMUSSEN et al (1988) und TANNOCK und Mitarbeitern (2000) genannten
Konzentrationen ähnlich. Zwischen den Messergebnissen der MIRA-Gruppen beste-
hen zu keinem Zeitpunkt signifikante Unterschiede. Deshalb kann davon ausgegan-
gen werden, dass die hier angewendeten Ernährungsmuster die Fettsäurebildung
nahezu gleichermaßen beeinflussten. Lediglich die im zeitlichen Verlauf häufiger
auftretenden Signifikanzen der MIRA-PreCon-Gruppe könnten auf einen größeren
Einfluss hindeuten. Der signifikante Abfall der Acetat-Konzentration in der MIRA-
PreCon-Gruppe wird durch den im Vergleich zur MIRA-DGE-Gruppe höheren
Ausgangswert verursacht. Die Ernährungsumstellung war in der MIRA-PreCon-
Gruppe plötzlich und gravierend, während sich die Zusammensetzung der Nahrung
in der MIRA-DGE-Gruppe langsam änderte. Auch waren diese Änderungen nicht so
stark wie in der MIRA-PreCon-Gruppe, da sich die Anteile der Nahrungs-
hauptbestandteile nur verschoben und der Speiseplan in seinen Bestandteilen nur
wenig verändert wurde. Die Veränderungen der Acetat-Konzentrationen sind ein
Hinweis auf die in der MIRA-PreCon-Gruppe stärker auftretende Veränderungen des
Dickdarmmilieus, was die SCFA-Bildung beeinflusst haben kann. In der 24. Woche
ist die Acetat-Konzentration in beiden Gruppen fast identisch und auch der Anstieg in
der 52. Woche ist ähnlich. Nach einer Verringerung der Konzentration bis zur 24.
Woche zeigen die Butyrat-Werte einen enormen Anstieg von der 24. bis zur 52.
Woche auf mehr als das Doppelte des Ausgangswertes. Die Propionat-Werte
schwankten nur gering und waren daher nicht signifikant verschieden. Die Acetat-
und Propionat-Werte der MIRA-DGE-Gruppe folgen dem Abwärtstrend der Werte der
MIRA-PreCon-Gruppe bis zur 24. Woche mit nachfolgendem Anstieg. Die Butyrat-
Werte dagegen zeigen einen schrittweisen Anstieg mit einem 52-Wochen-Wert, der
deutlich unter dem der MIRA-PreCon-Gruppe liegt. Allerdings sind die Butyrat-Werte
91

Diskussion
der MIRA-DGE-Gruppe sehr viel einheitlicher (s: ±2,72) als die der MIRA-PreCon-
Gruppe (s: ±16,74).
Um die Beziehungen der flüchtigen Fettsäuren zu den hier untersuchten Bakterien-
spezies zu untersuchen, wurden Korrelationsberechnungen nach Pearson durch-
geführt. Weder korrelierte die Gesamtkonzentration der SCFA mit den Gesamtkeim-
zahlen noch einzelne Fettsäuren mit bakteriellen Einzelparametern. Es ist hier nicht
gelungen, den bakteriellen Ursprung der gebildeten Fettsäuren zu identifizieren.
Die Aufteilung nach BMI in die MIRA-Untergruppen brachte keine Verdeutlichung der
Entwicklungen. Die zeitlichen Veränderungen der jeweiligen Fettsäuremengen in den
einzelnen Untergruppen sind ebensowenig signifikant wie der Vergleich der Unter-
gruppen gegeneinander. Wie auch in den Hauptgruppen gibt es keine korrelierenden
Bewegungen innerhalb der ermittelten SCFA und auch keine Korrelationen mit den
bakteriologischen Parametern.
Im zeitlichen Verlauf der MIRA-Studie ist die Dynamik der SCFA-Konzentrationen in
allen Gruppen nahezu gleich. Es wird die Tendenz sichtbar, dass sich unter den
durchgeführten Ernährungsumstellungen die Menge der ausgeschiedenen SCFA
verändert hat, was in der Tendenz dem Verlauf der aeroben und anaeroben Gesamt-
keimzahl, der Keimzahlen der Bifidobakterien, Laktobazillen und Eubakterien ähnelt.
Die berechneten Korrelationskoeffizienten bestätigen diese Beziehungen aber nicht,
da keine signifikanten Korrelationen existieren. Um diesem Umstand zu begegnen,
sollten die Gruppen in weiterführenden Studien gleich groß und möglichst homogen
sein und es ist vorteilhaft, ein breites Keimspektrum mit geeigneten Methoden zu
untersuchen. Interessant wäre auch die Untersuchung der resorbierten SCFA-
Mengen und deren Metabolismus wie bei POUTEAU et al. (2003) beschrieben.
5.5 Body Mass Index
Die BMI-Werte und ihre Dynamik sind in beiden MIRA-Hauptgruppen so ähnlich,
dass keine Unterschiede zwischen ihnen errechnet werden konnten. Die Werte an
sich zeigen zwar einen zum Studienende hin abgesunkenen durchschnittlichen BMI,
der aber noch deutlich über dem Normalwert von 25 kg/qm liegt. Wie beim Leptin
unter 5.6.1.1 genannt, gibt es auch hier viele Einzelpersonen in den Gruppen, die
einen deutlichen Gewichtsverlust durch Diät und Bewegung erzielten. In den Ge-
92

Diskussion
samtwerten zeigt sich diese Dynamik aber nur als Tendenz, da die Werte von Teil-
nehmern mit geringeren Erfolgen überwiegen.
Die gemeinsame Betrachtung der Leptin- und BMI-Werte macht den Zusammenhang
beider Parameter deutlich. Der BMI korreliert deutlich mit dem Körperfettanteil des
Teilnehmers und darüber mit dem Leptinwert, da das Leptin in den Fettzellen des
Körpers gebildet wird. Diese BMI/Leptin-Korrelationen ergaben sich auch aus den
Messergebnissen der MIRA-Studie (MIRA-PreCon-Gruppe: r=0,612** , MIRA-DGE-
Gruppe: r=0,570**). Das Ziel, den BMI deutlich zu senken, damit das Risiko von
Herz-Kreislauf-Erkrankungen zu vermindern und die Lebenserwartung und –qualität
zu steigern, ist nur einzelnen Teilnehmern gelungen. Die Gruppenwerte bewegen
sich auch in der 52. Woche an der Grenze zum deutlichen Übergewicht. Hier wird
sichtbar, wie schwierig Ernährungsstudien zu interpretieren sind. Es kann nicht
davon ausgegangen werden, dass die Compliance aller Teilnehmer gesichert ist. Die
Teilnahme an Sportangeboten und die Intensität der sportlichen Betätigung in der
Freizeit waren sehr unterschiedlich, was die erzielten Ergebnisse einzelner Teil-
nehmer (nicht dargestellt) zeigen. Unterschiedliche Entwicklungen des BMI aufgrund
der verschiedenen Ernährungen werden lediglich tendenziell deutlich. Die Werte der
MIRA-PreCon-Gruppe fallen ausgehend von der 12. Woche mit höherer Signifikanz
weiter ab (p<0,001), während die BMI der MIRA-DGE-Gruppe in der 12. und 24.
Woche weniger signifikant (p<0,05) verschieden sind. Das ist ein Hinweis auf die
nachhaltiger gewichtsreduzierende Wirkung der Kost der MIRA-PreCon-Gruppe.
Die BMI-Entwicklung zeigt in beiden Untergruppen eine hochsignifikante Gewichtsre-
duktion bis zur 24. Woche (p<0,001). Ebenfalls in beiden Untergruppen ist der
Studienendwert in der 52. Woche ist zwar signifikant höher als in der 24. Woche,
aber auch signifikant niedriger als der Ausgangswert in Woche 0. Damit werden die
Ergebnisse der Hauptgruppen bestätigt. Der einzige Unterschied in der Werte-
entwicklung der Untergruppen besteht in der Dynamik von Woche 12 zur Woche 24.
Diese Entwicklung ist nur in der MIRA-I signifikant und verdeutlicht hier eine durch-
gehend hochsignifikante Gewichtsreduktion bis zur 24. Woche. Die erhoffte bessere
Körpergewichtsreduktion der MIRA-I-Teilnehmer lässt sich nicht nachweisen, da
keine signifikanten Unterschiede zwischen den Gruppen bestehen.
93

Diskussion
5.6 Blutchemische Parameter 5.6.1 Leptin
Die BMI- und Leptinergebnisse korrelieren miteinander. Die Koeffizienten sind für die
MIRA-I r=0,602** und für die MIRA-II r=0,579**. Der geringere Korrelationskoeffizient
der MIRA-II kann Auruck der z.T. gegenläufigen Entwicklung beider Parameter in
dieser Untergruppe sein.
Bei der Interpretation der Ergebnisse muss beachtet werden, dass die Gewichtsre-
duktion u.a. eine Folge und nicht die Ursache der Leptinsekretion ist. Die Leptin-
sekretion wiederum wird von vielen Faktoren gesteuert, die ihrerseits von zahlreichen
Einflüssen wie Ernährung und Stoffwechsellage abhängig sind. Die daraus resultie-
rende enge Beziehung zwischen BMI und Leptin ist sowohl in den MIRA-Hauptgrup-
pen als auch in den Untergruppen klar zu erkennen. Die Vielzahl der Steuerungs-
möglichkeiten der Leptinsekretion zeigt auch, dass die Entwicklung nicht punktuell
mit der BMI-Dynamik übereinstimmen muss, wie es in der MIRA-II beobachtet wer-
den kann.
5.6.3 Blutzucker und HbA1c
Die Gruppenwerte beider Parameter liegen sowohl vor der Studie als auch zum
Studienende im Normalbereich. Interessant ist die gegenläufige Entwicklung inner-
halb der Normwerte. Während der aktuelle Blutzuckergehalt in beiden Gruppen konti-
nuierlich absinkt, gibt es beim HbA1c einen signifikanten Anstieg von der 24. bis zur
52. Woche. Der Langzeitwert lässt eine gesteigerte Glukoseaufnahme und –bindung
an Hämoglobin nach der 24. Woche erkennen und die Rückkehr der Teilnehmer zu
ihren alten Essgewohnheiten, wenn auch in unterschiedlichem Ausmaß, vermuten.
Die bei den Gesamtgruppen beschriebene gegenläufige Entwicklung von HbA1c und
BZ ist auch in der MIRA-I und MIRA-II zu finden.
Die Dynamik der Werte beider Parameter ist in beiden Untergruppen sehr ähnlich.
Somit kann trotz einer Korrelation des BZ mit dem BMI in der MIRA-I (r=0,529**) kein
direkter Zusammenhang mit der Körpergewichtsreduktion hergestellt werden. Eher
ist die Ernährung hier ein Einflussfaktor. Alle Teilnehmer der MIRA-Studie waren
angehalten, auf leichtverdauliche Kohlenhydrate zu verzichten. Scheinbar wurde
diese Anweisung befolgt, denn die BZ-Werte sinken im Studienverlauf in allen
94

Diskussion
Gruppen. Der HbA1c-Anstieg bis zur 12. Woche ist das „Echo“ der bisherigen Ernäh-
rungsweise, bis sich dann in den Werten der Woche 24 die Umstellung der
Nahrungszusammensetzung auch in diesem Parameter niederschlägt.
5.6.4 Gesamtcholesterol, LDL, HDL und Triglyceride
Die Messergebnisse der Blutfette, ausser die des HDL, liegen in beiden Haupt-
gruppen zu allen Untersuchungszeitpunkten in den Normbereichen. Die Gesamt-
Cholesterolwerte in der 12. Woche sind in beiden MIRA-Untergruppen signifikant
niedriger als die jeweiligen Ausgangswerte. Nur in der MIRA-I ist das Gesamt-
Cholesterol in der 52. Woche signifikant niedriger als der Ausgangswert. In der
MIRA-II bewegen sich beide Werte auf dem gleichen Niveau.
Die Cholesterolkonzentration im Blut hängt nicht allein von der Aufnahme mit der
Nahrung ab. Die Resorption im Darm ist beim Menschen begrenzt und die Chole-
sterol-Synthese der Leber hängt wiederum von der aufgenommenen Cholesterol-
menge ab. Außerdem beeinflusst die Fettzusammensetzung die Serumcholesterol-
Konzentration. So erhöhen gesättigte Fettsäuren den Cholesterinspiegel, während er
durch ungesättigte Fettsäuren gesenkt wird (ULMER 1993). Die Ernährungsum-
stellung beider MIRA-Hauptgruppen beinhaltete unter anderem eine fettarme
Ernährung mit der Zufuhr lediglich essentieller und wertvoller ungesättigter Fettsäu-
ren. Das war in der MIRA-PreCon-Gruppe durch die Einnahme der Zusatzkost ein-
facher als für die Teilnehmer der MIRA-DGE-Gruppe, die erst lernen mussten,
welche Nahrungsmittel geeignet waren. Auch das Beibehalten der erlernten
Ernährungsweise beeinflusst die Gesamt-Cholesterolwerte und ist in der MIRA-I
besser gelungen als in der MIRA-II. Das kommt durch den signifikant niedrigeren
Endwert im Vergleich zum Studienbeginn zum Auruck.
Die Serum-LDL-Konzentrationen verändern sich in beiden Untergruppen signifikant.
Die Werte in der MIRA-I sinken zur 24. Woche hin ab und steigen anschließend
wieder an, während in der MIRA-II sich Ausgangs- und Endwert signifikant unter-
scheiden. LDL und auch HDL stellen Transportmöglichkeiten des Körpers für die
Cholesterolester, die Transport- und Speicherform des Cholesterols, dar. Ihr Auftre-
ten ist gebunden an die zu bewegende Cholesterolmenge, wobei die in den HDL
transportierten Cholesterolester großteils auf die LDL übertragen werden (JOHNSON
1987). In allen MIRA-Untergruppen korrelieren die Cholesterolwerte deutlich mit den
95

Diskussion
LDL-Konzentrationen (MIRA-I: 0,818**; MIRA-II: 0,868**). Hier ist dieser Zusammen-
hang eindeutig sichtbar.
Übereinstimmend in beiden MIRA-Untergruppen steigen die HDL-Konzentrationen
zur 24. Woche hin an und bleiben auf diesem Niveau. Das kann Auruck gestei-gerter
Cholesterolfreisetzung und –transport aus dem Speichergewebe sein. Über die
Nahrung wird weniger Fett in den Organismus gebracht, so dass dieser benötig-te
Energie- und Synthesegrundlagen aus den vorhandenen Speichern mobilisieren
muss. Die Gruppenwerte des HDL liegen über dem Normalwert von 0,9 mmol/l. Die-
ses Ergebnis ist äußerst positiv zu werten. Das auch als „gutes Cholesterol“ bezei-
chnete HDL wirkt protektiv auf die Gefäßwände, bindet Endotoxine und ist auch in
leicht erhöhten Konzentrationen lediglich Auruck eines verstärkten Transportes von
Cholesterolestern.
5.7 Zusammenfassende Diskussion
In der vorliegenden Arbeit wurden ernährungsbedingte Veränderungen der Keim-
zahlen der fakultativen Anaerobier festgestellt. Andere untersuchte bakterielle Para-
meter zeigten tendenzielle Veränderungen im Untersuchungszeitraum. Kein
Zusammenhang konnte zwischen den Dynamiken der Keimzahlen und denen der
gebildeten SCFA, der Blutfette, des Blutzucker- und Leptinspiegels sowie des BMI
hergestellt werden. Enge Beziehungen wurden zwischen Gesamtcholesterol und
LDL sowie zwischen BMI und Leptin sichtbar. Mittlere Korrelationen ergaben sich
zwischen der aeroben Gesamtkeimzahl und der aeroben Gram-negativen Gesamt-
keimzahl.
Die Frage, inwieweit die Zusammensetzung der aufgenommenen Nahrung oder
deren einzelne Bestandteile die Fäkalflora verändern können, wird unterschiedlich
beantwortet. Die Einschätzung der Stabilität der Kolonflora gegenüber exogenen
Faktoren spielt dabei eine wesentliche Rolle. Autoren, die die Mikroflora für in sich
außerordentlich stabil halten, vertreten oft die Ansicht, dass die Darmbakterien ihren
Energiebedarf unabhängig von der Nahrungszufuhr durch z.B. den Abbau des ente-
ralen Zelldetrius und der Muzine decken können (SAVAGE 1986). Die Auswirkungen
ballaststoffreicher Ernährung auf die Darmflora und die Darmphysiologie sind aber
durch zahlreiche Studien vor allem zum Einsatz von Probiotika belegt (ROBER-
96

Diskussion
FROID 1993, NINESS 1999). Die vorliegende Arbeit zeigt ebenfalls Veränderungen
der Darmflora, die mit einer Umstellung der Ernährung einhergehen.
Schlussfolgernd kann festgestellt werden, dass einige der hier untersuchten bakteri-
ellen Parameter von externer Energiezufuhr beeinflussbar sind und sich bei Änderun-
gen der Ernährungsweise dem Nährstoffangebot anpassen.
97

Zusammenfassung
6 Zusammenfassung
Thomas Vorpagel
Untersuchungen zum Einfluss der Ernährung auf ausgewählte bakteriologische,
enzymatische und blutchemische Parameter beim Menschen unter Gewichts-
reduktion
Institut für Bakteriologie und Mykologie der Veterinärmedizinischen Fakultät der Uni-
versität Leipzig und Kreiskrankenhaus Altenburg
Eingereicht im Februar 2005
116 Seiten, 14 Abbildungen, 29 Tabellen und 142 Literaturangaben
Schlüsselworte: Mensch, Ernährung, Bakteriologie, Gewichtsreduktion
Es war Ziel der vorliegenden Arbeit, Einflüsse der Ernährung auf die Fäkalflora des
Menschen anhand geeigneter Fäkalparameter zu untersuchen. Durch die Auswer-
tung ausgewählter blutchemischer Parameter wurde versucht, veränderte Stoff-
wechselsituationen der Probanden zu reflektieren.
In der MIRA-Studie des ETZ M.Scholz, Leipzig, die die Grundlage diese Arbeit bildet,
wurden zwei Gruppen mit unterschiedlichen Ernährungsregimen eingeschlossen. Die
Teilnehmer der MIRA-DGE-Gruppe gestalteten ihre Mahlzeiten nach den Richtlinien
der Deutschen Gesellschaft für Ernährung (DGE) und die Teilnehmer der MIRA-Pre-
Con-Gruppe erhielten eine hypokalorische Nahrungsergänzung der PreCon GmbH &
Co. KG. Beide Gruppen erhielten die gleiche Betreuung, wie Ernährungsberatung
und Auskunft über geeignete Bewegungsmöglichkeiten. Es wurden Proben von ins-
gesamt 60 Teilnehmern untersucht. Davon entfielen 29 Frauen und 8 Männer auf die
MIRA-PreCon-Gruppe und 21 Frauen und 2 Männer auf die MIRA-DGE-Gruppe.
Untersuchungszeitpunkte waren die Nullprobe vor Interventionsbeginn, die Wochen
12 und 24 sowie die Endprobe nach 52 Wochen. Der durchschnittliche BMI lag bei
der MIRA-PreCon-Gruppe bei 32,4 kg/m2 und in der MIRA-DGE-Gruppe bei 32,5
98

Zusammenfassung
kg/m2. Die Teilnehmer verfolgten das Ziel, Körpergewicht durch Ernährungsum-
stellung und körperliche Aktivität zu reduzieren.
Bei den ausgewählten bakteriologischen Parametern wurde in beiden Gruppen eine
signifikante Verminderung der fakultativen Anaerobier-Keimzahlen im Studienverlauf
beobachtet. Die anderen Bakterienspezies wiesen aufgrund der sehr starken Streu-
ung der Keimzahlen nur eine tendenzielle Entwicklung auf und konnten daher nicht
mit der jeweiligen Ernährungsumstellung in Verbindung gebracht werden.
Die Untersuchung der in den Fäzes ausgeschiedenen volatilen Fettsäuren ergab z.T.
signifikante Schwankungen innerhalb des Untersuchungszeitraumes. Eine Zuord-
nung der gebildeten Fettsäuren zu untersuchten Bakterien war nicht möglich.
Die blutchemischen Untersuchungen ergaben bei allen Untersuchungen Normal-
werte für die Blutfette und den Blutzucker sowie eine mittlere Korrelation zwischen
den ermittelten Leptinwerten und dem BMI.
Die Ergebnisse zeigen, dass die Ernährung die Fäkalflora beeinflusst. In weiterfüh-
renden Studien sollte ein breiteres Keimspektrum mit Schwerpunkt auf der strikt
anaeroben Flora untersucht werden, um Nahrungseinflüsse detaillierter darstellen zu
können.
99

Summary
7 Summary
Thomas Vorpagel
The influence of nutrition on particular bacteriological, enzymological, and other
metabolic parameters in man reducing its weight
Institute of Bacteriology and Mycology, Faculty of Veterinary Medicine, University of
Leipzig and Hospital of Altenburg
Submitted in February 2005
116 pages, 14 figures, 29 tables, 142 references
Keywords: man, nutrition, bacteriology, weight loss
The investigation how nutrition influences the faecal flora in man was aim of this
thesis. The examined samples of 60 mainly elderly people were divided into the
MIRA-PreCon group consisting of 29 women and 8 men and the MIRA-DGE group
consisting of 21 women and 2 men. The first blood and stool samples were collected
a short time before the MIRA-study began. The following samples were collected
after 12, 24, and the last after 52 weeks, respectively.
The average Body Mass Index (BMI) of the MIRA-PreCon group was 32,4 kg/m2 and
32,5 kg/m2 of the MIRA-DGE group. During the study, all participants tried to reduce
their body weight by changing their nutrition habits and by increasing their physical
activity. The participants of the MIRA-DGE group followed the recommendations of
the German Institute of Nutrition (DGE) and the participants of the MIRA-PreCon
group received a hypocaloric diet sold by the PreCon GmbH & Co KG.
In both MIRA groups a significant reduction of the facultative anaerobic counts
occurred. All other bacterial parameters investigated showed no distinctive develop-
ment. The metabolic parameters, e.g. the blood lipids, varied at all times of the
investigation within physiological limits. The concentration of BMI and Leptin medi-
cally correlated in both groups.
100

Summary
The results of this thesis prove the significant influence of nutrition on the faecal flora
in man. Comparing both MIRA groups, however, no significant differences occurred
throughout the study. Therefore, further studies in this field need to adequately
investigate mainly anaerobic species in order to detect minor changes in bacterial
counts.
101

Literaturverzeichnis
8 Literaturverzeichnis
1 Alexander M. Microbial ecology. New York: John Wiley and Sons; 1971.
2 Amann RI, Krumholz L, Stahl DA. Fluorescent-oligonucleotide probing of
whole cells for determinative, phylogenetic, and environmental studies in
microbiology. J Bacteriol. 1990;172(2):762-70.
3 Autenrieth IB. Die Ökologie der humanen Darmflora: Physiologische und
pathophysiologische Aspekte. J Ernährungsmed. 2000; 2(4):6-9.
4 Bender A Breves G, Stein J, Leonhard-Marek S, Schroder B, Winckler C.
Colonic fermentation as affected by antibiotics and acidic pH: Application of an
in vitro model. Z Gastroenterol. 2001;39(11):911-8.
5 Bengmark S, Jeppsson B. Gastrointestinal surface protection and mucosa
reconditioning. J Parenter Enteral Nutr. 1995;19(5):410-5.
6 Benic V, Fisher-Hermann M. Über alkalische Phosphatasen in menschlichen
Faeces. Z klin Chem u. klin Biochem. 1970;8.Jg.,Heft 5:458-64.
7 Bennet R, Gorbach SL, Goldin B. Treatment of relapsing Clostridium difficile
diarrhea with lactobacillus GG. Nutrition Today. 1996;31:35.
8 Bentley DW , Nicols RL, Condon RE, Gorbach SL. The Microflora of the
human ileum and intraabdominal colon: results of direct needle aspiration at
surgery and evaluation of the technique. J Lab Clin Med. 1972;79:421-9.
9 Berg RD. The indigenous gastrointestinal microflora. Trends in Microbiology.
1996;4(11):430-5.
10 Bergman EN. Energy contributions of volatile fatty acids from the gastroin-
testinal tract in various species. Physiol Rev. 1990;70(2):567-90.
102

Literaturverzeichnis
11 Bischoff SC, Mayer JH, Manns MP. Allergy and the gut. Int Arch Allergy
Immunol. 2000;121:270-83.
12 Boriello SP. Microbial Flora of the gastrointestinal tract, In: Hill MJ, editor.
Microbial metabolism in the digestive tract. Boca Raton, FL, CRC Press; 1986.
S. 2
13 Brandtzaeg P, Farstad IN, Johansen FE et. al. The B-cell system of human
mucosae and exocrine glands. Immunol Rev. 1999;171:45-87.
14 Brandtzaeg P. Current understanding of gastrointestinal immunoregulation and
its relation to food allergy. Ann NY Acad Sci. 2002;964:13-45.
15 Briedigkeit H. Obstipation und Darmflora. Z ärztl Fortbild. 1992;86:117-9.
16 Briskin MJ, McEvoxxy LM, Butcher EC. MAdCAM-1 has homology to
immunoglbulin and mucin-like adhesion receptors and IgA1. Nature.
1993;363:461-4.
17 Buck AC, Cooke EM. The fate of ingested Pseudomonas aeruginosa in normal
persons. J Med Microbiol. 1969;2:521-5.
18 Butel MJ, Roland N, Hilbert A, Popot F, Favre A, Tessedre AC et al. Clostridial
pathogenicity in experimantal necrotising enterocolitis in gnotobiotc quails and
protective role of bifidobacteria. J Med Microbiol. 1998;47(5):391-9.
19 Considine RV, Sinha MK, Heimann ML, Kriauciunas A, Stephens TW, Nyce
MR, et al. Serum immunoreactive-leptin concentrations in normal weight and
obese humans. N Engl J Med. 1996;334:292-5.
20 Conway LC. Microbial ecology of the human large intestine. In: Gibson GR
and Macfarlane GT, editors. Human colonic bacteria: role in nutrition,
physiology and pathology. Boca Raton: CRC Press; 1995.S. 4.
103

Literaturverzeichnis
21 Crowther JS, Drasar BS, Goddard P, Hill MJ, Johnson K. The effect of
chemically defined diet on the faecal flora and faecal steroid concentrations.
Gut. 1973;14:790.
22 Cummings JH, Banwell JG, Englyst HN, Coleman N, Segal I, Bersohn D. The
amount and composition of large bowel contents. Gastroenterology.
1990;98:A408.
23 Cummings JH, Macfarlane GT. A review: the control and consequence of
bacterial fermentation in the human colon. J Appl Bacteriol. 1991;70:443.
24 Davidson MH, Maki KC. Effects of inulin on serum lipids. J Nutr.
1999;129:1474-7.
25 Delzenne NM, Roberfroid MR. Physiological effects of non-digestible
oligosaccharides. Lebensm Wiss u Technol. 1994;27:1-6.
26 Drasar BS, Hill MJ. Human intestinal flora. New York: Academic Press, 1974.
27 Drasar BS, Jenkins DJA. Bacteria, diet and large bowel cancer. Am J Clin
Nutr. 1976;29:1410.
28 Drasar BS, Goddard P, Heaton S, Peach S ,West B. Clostridia isolated from
faeces. J Med Microbiol. 1976;9:63.
29 Dubos R, Schaedler RW, Costello R, Hoet P. Indigenous, normal and
autochthonous flora of the gastrointestinal tract. J Exp Med. 1965;122:67.
30 Duchmann R, Hoffmann J, Marth T, Schneider T, Stallmach A, Zeitz M.
Mukosales Immunsystem im Darm–Immunologische Grundlagen chronisch
entzündlicher Darmerkrankungen und der HIV–induzierten Enteropathie.
Magazin Forschung. 1999;1:47-64.
104

Literaturverzeichnis
31 Duerden BI. The isolation and identification of Bacteroides spp from normal
human faecal flora. J Med Microbiol. 1980;13:69.
32 Finegold SM, Sutter VL. Fecal flora in different populations, with special
reference to diet. Am J Clin Nutr. 1978;31:116.
33 Finegold SM, Sutter VL, Mathisen GE. Normal indigenous intestinal flora. In
Hentges DJ, editor. Human intestinal microflora in health and disease. London:
Academic Press, 1983:3.
34 Foo M, Lee A. Immunological response of mice to members of the
autochthoneous intestinal flora. Infect Immun. 1972;6:525.
35 Foo MC, Lee A, Cooper GN. Natural antibodies and the intestinal flora of
rodents. Aust J Exp Biol.Med. 1976;52:321.
36 Freter R. Mechanisms that control the microflora in the large intestine. In:
Hentges DJ, editor. Human intestinal microflora in health and disease. London:
Academic Press; 1983:S.33.
37 Freter R. Factors affecting the microecology of the gut. In: Fuller R, editor.
Probiotics: The scientific basis. London: Chapman and Hall; 1992:S111.
38 Frühbeck G, Aguado M, Martinez A. In vitro lipolytic effect of leptin on mouse
adipocytes: evidence for a possible autocrine/paracrine role of leptin. Biochem
Biophys Res Commun. 1997;240:590-4.
39 Gerhard NB, Iglewski WJ. Alterations in the fecal flora of an individual
frequently lacking coliforms. J Infect Dis. 1976;134:490-549.
40 Gianella RA, Broitman SA, Zamcheck N. Gastric acid barrier to ingested
microorganisms in man: studies in vivo and in vitro. Gut. 1972;13:251-6.
105

Literaturverzeichnis
41 Gibson GR, Beatty ER, Wang X, Cummings JH. Selective stimulation of
bifidobakteria in the human colon by oligofructose and inulin.
Gastroenterology. 1995;108:975-82.
42 Gibson GR. Dietary modulation of the human gut microflora using the
prebiotics oligofructose and inulin. J Nutr. 1999;129:1438-41.
43 Giovannoni SJ, DeLong EF, Olsen GJ, Pace NR. Phylogenetic group-specific
oligodeoxynucleotide probes for identification of single microbial cells. J
Bacteriol. 1988;170(2):720-6.
44 Goldin BR und SL Gorbach. Probiotics for humans. In: Fuller R, editor.
Probiotics: the scientific basis. London: Chapman and Hall; 1992.S. 355.
45 Goldman AS. The immune system of human milk: antimicrobial,
antiinflammatory and immunomodulating properties. Pediatr Infekt Dis J.
1993;12:664-71.
46 Gorbach SL, Nahas L, Lerner PI, Weinstein L. Studies of intestinal microflora.
I. Effects of diet, age, and periodic sampling on numbers of fecal
microorganisms in man. Gastroenterology. 1967;53:845-55.
47 Gross GC, Schwartz VL, Stevens C, Ebert EC, Blumberg RS, Balk SP.
Distribution of dominant t cell receptor ß chains in human intestinal mucosa. J
Exp Med. 1994;180:1337-44.
48 Grove D. Amoebal dysentery, intestinal protozoa and helminths. In: Goodwin
CS, editor. Microbes and infections of the gut. Oxford: Blackwell Scientific
Publications;1984.S.209.
49 Hamilton BS, Paglia D, Kwan AY, Deitel M. Increased obese mrna expression
in omental fat cells from massively obese humans. Nat Med. 1995;1:953-6.
106

Literaturverzeichnis
50 Harmsen D, Rothganger J, Singer C, Albert J, Frosch M. Intuitive hypertext-
based molecular identification of microorganisms. Lancet.
1999;353(9149):291.
51 Helgeland L, Brandtzaeg P. Development an function of intestinal b and t cells.
Microbiol Ecol Health Dis. 2000;Suppl.2:110-27.
52 Hentges DJ, Freter R. In vivo and in vitro Antagonism of intestinal bacteria
against Shigella flexneri. I. Correlation between various tests. J Infect Dis.
1962;110:30-7.
53 Hentges DJ. Gut flora and disease resistance. In: Fuller R, editor. Probiotics:
the scientific basis. London: Chapman and Hall, 1992.S.87.
54 Holdeman LV, Good IJ, Moore WEC. Human fecal flora: variation in bacterial
composition within individuals and a possible effect of emotional stress. Appl
Environ Microbiol. 1976;31:359.
55 Holt PG, Jones CA. The development of the immune system during pregnancy
and early life. Allergy. 2000;55:688-97.
56 Hopkins MJ, Sharp R, Macfarlane GT. Age and disease related changes in
intestinal bacterial populations assessed by cell culture, 16s rrna abundance,
and community cellular fatty acid profiles. Gut. 2001;48:198-205.
57 Hopkins MJ, Macfarlane GT. changes in predominant bacterial populations in
human faeces with age and with Clostridium difficile infection. J Med Microbiol.
2002;51:448-54.
58 Horrigan FD, Danovitch SH. The origin of human fecal alkaline phosphatase.
Dig Dis. 1984;19(7): 603-8.
107

Literaturverzeichnis
59 Hoskins LC, Boulding ET. Mucin degradation in human colon ecosystems.
Evidence for the existence and role of bacterial subpopulations producing
glycosides as extracelular enzymes. J Clin Invest. 1981;67:163.
60 Ishibashi N, Yamazaki S. Probiotics and safety. Am J Clin Nutr.
2001;73(Suppl):465-70.
61 James SP, Zeitz M. Human gastrointestinal mucosal t cells. In: Ogra,
Mestecky, Lamm, Strober, McGhee, Bienenstock, editors. Mucosal
immunology. Second edition. San Diego: Academic Press. 1994.S.275-85.
62 Jenkins DJA, Kendall CWC,. Vuksan V. Inulin, oligofructose and intestinal
function. J Nutr. 1999;129:1431-3.
63 Johnson LR. Physiology of the gastrointestinal tract, Vol.I+II, Second edition.
New York: Raven. 1987.
64 Kelsall BL, Strober W. Host defenses at mucosal surfaces. In: Rich, Fleischer,
Schwartz, Shearer, Strober, editors. Clinical immunology, principles and
practice. St. Louis: Mosby, 1995.S.299-322.
65 Kimura K, McCartney AL, McConnell MA, Tannock GW. Analysis of fecal
populations of bifidobacteria and lactobacilli and investigations of the
immunological responses of their human hosts to the predominant strains.
Appl Environ Microbiol. 1997;63(9):3394-8.
66 Kamin RA, Henry C, Fudenberg H. Suppressor cells in the rabbit appendix. J
Immunol. 1974;113:1151.
67 Kleessen B, Sykura B, Zunft H-J, Blaut M. Effects of inulin and lactose on fecal
microflora, microbial activity and bowel habit in elderly constipated persons.
Am J Clin Nutr.1997;65:1397-1402.
108

Literaturverzeichnis
68 Kleessen B, Noack J, Blaut M. Distribution of viable and nonviable bacteria in
the gastrointestinal tract of gnotobiotic and conventional rats. Microbiol Ecol
Health Dis. 1999;11:218-25.
69 Kleessen B, Hartmann L, Blaut M. Oligofructose and long-chain inulin:
Influence on the gut microbial ecology of rats associated with a human faecal
flora. B J Nutr. 2001;86:1-11.
70 Kleessen B, Kroesen AJ, Buhr HJ, Blaut M. Mucosal and invading bacteria in
patients with inflammatory bowel disease compared with controls. Scand J
Gastroenterol. 2002;37(9):1034-41.
71 Kruse H-P, Kleessen B, Blaut M. Effects of inulin on faecal bifidobacteria in
human subjects. Brit J Nutr. 1999;82:175-382.
72 Kyne L, Farrell RJ, Kelly CP. Clostridium difficile. Gastroenterol Clin North Am.
2001;30(3):753-777, ix-x.
73 Langendijk PS, Schut F, Jansen GJ, Raangs GC, Kamphuis GR, Wilkinson
MH, et al. Quantitative fluorescence in situ hybridization of Bifidobacterium
spp. with genus-specific 16s rna-targeted probes and its application in fecal
samples. Appl Environ Microbiol. 1995;61(8):3069-75.
74 Lee A. Neglected niches: The microbial ecology of the gastrointestinal tract. In:
Marshall KC, editor. Advances in microbial ecology. Vol. 8, New York: Plenum
Press.1985.S.115.
75 Lee FD, Kraszewski A, Gordon J, Howie JGR, McSeveney D, Harland WA.
Intestinal spirochaetosis. Gut. 1971;12:126-33.
76 Lehmann F-G, Zengel W-R. Eine Methode zur Bestimmung der alkalischen
Leberphosphatase im Stuhl und ihre Aktivität bei intra- und extra-hepatischem
Verschlußikterus. Z Gastroenterologie. 1980a;18:274-85.
109

Literaturverzeichnis
77 Lehmann F-G, Hillert U. Fecal intestinal alkaline phosphatase in coeliac
disease. Z Gastroenterologie. 1980b;18:381-8.
78 Lehmann F-G, Cramer P, Hillert U. Activity of small intestine alkaline
phosphatase in the feces in chronic intestinal diseases. Z Gastroenterol.
1980c;18(4):208-15.
79 Lehmann F-G, Hufnagel H, Lorenz-Meyer H. Fecal intestinal alkaline
phosphatase: a parameter for toxic damage of the small intestinal mucosa.
Digestion. 1981;21:156-62.
80 Lievin V, Pfeiffer I, Hudault S, Rochat F, Brassart D, Neeser J et al.
Bifidobacterium strains from resident infant human gastrointestinal microflora
exert antimicrobial activity. Gut.2000;47(5):646-52.
81 McCartney A, Wenzhi W, Tannock GW. Molecular analysis of the composition
of the bifidobacterial and lactobacillus microflora of humans. Appl Environ
Microbiol. 1996;62 (12):4608-13.
82 Macfarlane GT, Hay S, Gibson GR. Influence of mucin on glycosidase,
protease and arylamidase activities of human gut bacteria grown in a 3-stage
continuous culture system. J Appl Bacteriol. 1989a;66:407.
83 Macfarlane GT, Cummings JH, Macfarlane S, Gibson GR. Influence of
retention time on degradation of pancreatic enzymes by human colonic
bacteria grown in a 3-stage continuous culture system. J Appl Bacteriol.
1989b;67:521.
84 Macfarlane GT, Cummings JH. The colonic flora, fermentation and large bowel
digestive function In: Phillips SF, Pemberton JH, Shorter RG, editors. The
large intestine: physiology, pathophysiology and disease., New York: Raven
Press 1991.S.51.
110

Literaturverzeichnis
85 Macfarlane GT, Gibson GR, Cummings JH. Comparison of fermentation
reactions in different regions of the colon. J Appl Bacteriol. 1992;72:57.
86 Macfarlane S, Cummings JH, Macfarlane GT. Bacterial Populations on the
rectal mucosa in healthy and colitic subjects. Gastroenterology.
2000;118(Suppl.2), A101, 678.
87 Maddox PH, Jenkins D. 3-Aminopropyltriethoxysilane (APES): A new advance
in section adhesion. J Clin Pathol. 1987;40(10):1256-7.
88 Madigan MT, Martinko JM, Parker J. Mikrobiologie. Goebel W, editor., Berlin:
Spektrum Akademischer Verlag Heidelberg, 2001.
89 Marteau P, Pochart P, Dore J, Bera-Maillet C, Bernalier A, Corthier G.
Comparative study of bacterial groups within the human cecal and fecal
microbiota. Appl Environ Microbiol. 2001;67(10):4939-42.
90 Mata LJ, Urrutia JJ. Intestinal colonization of breast-fed children in a rural area
of low socioeconomic level. Am NY Acad Sci. 1971;176:93.
91 Midtvedt T. Monitoring of the functional state of the microflora. In: Hattori T,
Ishida Y, Maruyama Y, Morita R, Uchida A, editors. Recent advances in
microbial ecology. Japan Scientific Societies Press. Tokyo: 1989.S.515.
92 Mims LC, Medawar MS, Perkins JR, Grubb WR. Predicting neonatal infections
by evaluation of the gastric aspirate: a study in two hundred and seven
patients. Am J Obstet Gynecol. 1972;114:232.
93 Moore WEC, Holdeman LV. Human fecal flora: the normal flora of 20
japanese-hawaiians. Appl Microbiol 1974;27:961-79.
94 Mylonakis E, Ryan ET, Calderwood SB. Clostridium difficile-Associated
Diarrhea: A review. Arch Intern Med. 2001;161(4):525-33.
111

Literaturverzeichnis
95 Nelson DP, Mata LJ. Bacterial flora associated with the human gastrointestinal
mucosa. Gastroenterology. 1970;58:56-61.
96 Niness KR. Inulin and oligofructose: what are they? J Nutr. 1999;129(Suppl
7):1402-6.
97 Ohkusa T, Ozaki Y, Sato C, Mikuni K, Ikeda H. Long-term ingestion of
lactosucrose increases bifidobacterium sp. in human fecal flora. Digestion
1995;56:415-20.
98 Orrhage K und Nord CE. Bifidobacteria and lactobacilli in human health. Drugs
Exptl Clin Res. 2000;XXVI(3):95-111.
99 Poelstra K, Bakker WW, Klok PA, Kamps JA, Hardonk MJ, Meijer DK.
Dephosphorylation of endotoxin by alkaline phosphatase in vivo. Am J Pathol.
1997;151(4):1163-9.
100 Posen S. Alkaline phosphatase. Ann Entern Med. 1967;67(1):183-203.
101 Pouteau E, Nguyen P, Ballevre O, Krempf M. production rates and metabolism
of short-chain fatty acids in the colon and whole body using stable isotopes.
Proc Nutr Soc. 2003;62(1):87-93.
102 Poxton IR, Brown R, Sawyerr A, Ferguson A. The mucosal anaerobic gram-
negative bacteria of the human colon. Clin Infect Dis. 1997;(Suppl 2):111-3.
102 Prins RA. Biochemical activities of gut microorganisms. In: Microbial ecology
of the gut. Clarke RTJ, Bauchop T, editors., Academic Press. London:
1977.S.74.
103 Rassmussen HS, Holtung K, Ynggard C, Mortensen PB. Faecal
concentrations and production rates of short chain fatty acids in normal
neonates. Acta Paediatr Scand. 1988;77(3):365-8.
112

Literaturverzeichnis
104 Roberfroid MB. Dietary fiber, inulin, and oligofructose: a review comparing
their physiological effects. Crit Rev Food Sci Nutr. 1993;33(2):103-48.
105 Roberfroid MB. Prebiotics and synbiotics: concepts and nutritional properties.
Brit J Nutr. 1998;80(Suppl 2):197-202.
106 Robinett HG. Relationship of host antibody to fluktuations of Escherichia coli
serotypes in the human intestine. J Bacteriol. 1962;84:896-901.
107 Roediger WEW. Role of anaerobic bacteria in the metabolic welfare of the
colonic mucosa in man. Gut. 1980;21:793.
108 Roediger WEW. Interrelationship between bacteria and mucosa of the
gastrointestinal tract. In: Hill MJ, editor. Microbial metabolism in the digestive
tract. Boca Raton, FL: CRC Press; 1986.S.201.
109 Rolfe RD. The role of probiotic cultures in the control of gastrointestinal health.
J Nutr. 2000;130:396-402.
110 Rolle M, Mayr A. Medizinische Mikrobiologie, Infektions- und Seuchenlehre.
Mayr A, editor., Stuttgart: Enke-Verlag; 2002.
111 Rusch K, Rusch V. Mikrobiologische Therapie: Grundlagen und Praxis. Rusch
K und V,editors. Heidelberg: Haug; 2001.
112 Salminen S, Salminen E. Lactulose, lactic acid bacteria, intestinal micro-
ecology and mucosal protection. Scand J Gastroenterol. 1997;222:45-8.
113 Sandler RH, Finegold SM, Bolte ER, Buchanan CP, Maxwell AP, Väisänen M-
L, et al. short-term benefit from oral vancomycin treatment of regressive-onset
autism. J Child Neurol. 2000;15:429-35.
114 Savage, DC. Associations of indigenous microorganisms with gastrointestinal
mucosal epithelia. Am J Clin Nutr. 1970;23:1495-1501.
113

Literaturverzeichnis
115 Savage DC. Microbial ecology of the gastrointestinal tract. Ann Rev. Microbiol.
1977a;31:107.
116 Savage DC. Interactions between the host and its microbes. In: Clarke RTJ,
Bauchop T, editors. Microbial ecology of the gut. London: Academic Press,
1977b.S.277.
117 Savage DC. Gastrointestinal microflora in mammalian nutrition. Annu Rev
Nutr. 1986;6:155-178.
118 Scheppach W. Effects of short chain fatty acids on gut morphology and
function. Gut. 1994;35(Suppl1):35-8.
119 Scholz GH, Englaro P, Thiele I, Scholz M, Klusmann T, Kellner K, Rascher W,
Blum WF. Dissociation of serum leptin concentration and body fat content
during long term dietary intervention in obese individuals. Horm Metab Res.
1996;28:718-23.
120 Scholz GH, Flehmig G, Kahl Y, Gutknecht D, Schmidt U, Tolkmitt S, et al.
Proper Weight Loss with Intelligence (MIRA). 2 programs for weight reduction
in general practice trial. MMW Fortschr Med. 2002;144(40):28-32.
121 Schulze J. Obstipation und Darmflora. Z ärztl Fortbild. 1992;86:121-6.
122 Schumann C. Medical, nutritional and technological properties of lactulose.
Eur J Nutr. 2002;41(Suppl 1):17-25.
123 Shedlovsky S, Freter R. Synergism between ecologic and immunologic control
of mechanisms of intestinal flora. J Infect Dis. 1974;129:296.
124 Simon GL, Gorbach SL. Intestinal flora in health and disease. In: Johnson LR,
editor. Physiology of the gastrointestinal tract. New York: Raven Press,
1981.S.1361.
114

Literaturverzeichnis
125 Sonnenborn, U Greinwald R. Beziehungen zwischen Wirtsorganismus und
Darmflora. U. Sonnenborn, editor. Stuttgart; New York: Schattauer 1991.
126 Stark PL, Lee A. The microbial ecology of the large bowel of breast-fed and
formulae-fed infants during the first year of life. J Med Microbiol. 1982;15:189.
127 Swidsinsky A, Ladhoff A, Pernthaler A, Swidsinsky S, Loening-Baucke V,
Ortner M, et al. Mucosal flora in inflammatory bowel disease. Gastroenerology.
2002;122:44-54.
128 Swiger RR, Tucker JD. Fluorescence in situ hybridisation: a brief review.
Environ Mol Mutagen. 1996;27(4):245-54.
129 Takesue Y, Yokoyama T, Akagi S, Ohge H, Murakami Y, Sakashita Yet al.
Changes in intestinal flora after administration of anipenem/betamipron or
sulbactam/cefoperazone for treatment of postoperative infections in gastro-
ectomy patients. J Infect Chemother. 1999;5(1):52-7.
130 Tannock GW. Microbial interference in the gastrointestinal tract. Asian J Clin
Sci. 1981;2:2.
131 Tannock GW. Effect of dietary and environmental stress on the gastrointestinal
microbiota. In: Hentges, DJ editor. Human intestinal microflora in health and
disease. London: Academic Press; 1983.S.517.
132 Tannock GW, Munro K, Harmsen HJM, Welling GW, Smart J, Gopal PK.
Analysis of the fecal microflora of human subjects consuming a probiotic
product containing Lactobacillus rhamnosus DR20. Appl Environ Microbiol.
2000;66(6):2578-88.
133 Thadepalli H, Lou MA, Bach VT, Matsui TK,. Mandal AK Microflora of the
human small intestine. Am J Surg. 1979;138:845-50.
115

Literaturverzeichnis
134 Uhlmann K. Leptin und Nebenniere: Morphologie, physiologische und
molekulare Untersuchungen [Dissertation]. Universität Leipzig, 2002.
135 Ulmer HV. Ernährung. In: Physiologie des Menschen. Schmidt RF, Thews G,
editors. Berlin: Springer; 1993.S.719.
136 Van der Waaij D. The immunoregulation of the intestinal flora: experimental
investigations on the development and the composition of the microflora in
normal and thymus less mice. Microecol Ther. 1984;14:63.
137 Van Saene HKF, Van der Waaij D. A novel technique for detecting IgA coated
potentially pathogenic microorganisms in the human intestine. J Immunol
Meth. 1979;30:37.
138 Wang R-F, Cao W-W,. Cerniglia CE PCR detection and quantifikation of
predominant anaerobic bacteria in human an animal fecal samples. Appl
Environ Microbiol. 1996;62(4):1242-7.
139 Winitz M, Adams RF, Seedman DA, Davis PN, Jakyo LG, Hamilton JA.
Studies in metabolic nutrition employing chemically defined diets. II. Effects on
gut microflora populations. Am J Clin Nutr. 1970;23:546.
140 Zangh Y, Shao JS, Xie QM, Alpers DH. Immunolocalization of alkaline
phosphatase and surfactant-like particle proteins in rat duodenum during fat
absorption. Gastroenterology. 1996;110(2):478-88.
141 Zeitz M, Schieferdecker HL, Ullrich R, Jahn HU, James SP, Riecken EO.
Phenotype and function of lamina propria t lymphocytes. Immunol Res.
1991;10(3-4):199-206.
142 Zoetendal EG, Wright A v., Vilpponen-Salmela T, Ben-Amor K, Akkermans
ADL, de Vos WM. Mucosa-associated bacteria in the human gastrointestinal
tract are uniformly distributed along the colon and differ from the commuity
recovered from feces. Appl Environ Microb. 2002;68 (7):3401-7.
116

Danksagung
Danksagung
Ich danke Frau Prof. Dr. Monika Krüger für die Überlassung des Themas der
vorliegenden Dissertation sowie ihre fachliche Beratung und Unterstützung. Für die
Beteiligung an der MIRA-Studie und seine vielfältige Unterstützung danke ich Herrn
Prof. Dr. Gerhard Scholz.
Den beteiligten Mitarbeiterinnen und Mitarbeitern des Ernährungsberatungs- und
Trainingszentrums ETZ M.Scholz, Leipzig und dessen Leiterin, Frau Monika Scholz,
danke ich ebenfalls für ihre Unterstützung, v.a. bei der Entgegennahme und Aufbe-
wahrung der Proben.
Am Institut für Bakteriologie und Mykologie der Veterinärmedizinischen Fakultät
bekam ich umfassende Hilfe bei der Probenbearbeitung von Frau Dr. Brigitta
Kleeßen und Herrn Dr. Wieland Schrödl. Den Mitarbeiterinnen des bakteriologischen
Labors des Institutes danke ich für die Einführung in die bakteriologische Diagnostik.
Gedankt sei auch Frau Dippmann, die durch ihr unermüdliches Schaffen in der Nähr-
bodenküche die materiellen Grundlagen dieser Dissertation sicherte. Der Sekretärin
des Institutes, Frau Dagmar Schrödl, danke ich für ihren beratenden und oft wegwei-
senden Einsatz.
Besonders herzlich danke ich Herrn Andreas Richter aus der Ambulatorischen und
Geburtshilflichen Tierklinik für seine Hilfe bei der statistischen Auswertung meiner
Ergebnisse.
Ich danke meiner Familie für ihre Geduld und liebevolle Unterstützung.





























