Embed Size (px)
Citation preview

Aus der
Abteilung für Neuroanatomie und Molekulare Hirnforschung
der Ruhr-Universität Bochum
Prof. Dr. med. R. Dermietzel
Regulation von Pannexin1 durch Phosphorylierung
Inaugural-Dissertation
zur
Erlangung des Doktorgrades der Medizin
einer
Hohen Medizinischen Fakultät
der Ruhr-Universität Bochum
vorgelegt von
Laura Twardowski
aus Düsseldorf
2011

Dekan: Prof. Dr. med. K. Überla
Referent: Prof. Dr. med. R. Dermietzel
Korreferent: PD Dr. rer. nat. H. Rottensteiner
Tag der Mündlichen Prüfung: 15.05.2012

Abstract Laura Twardowski Regulation von Pannexin1 durch Phosphorylierung Problem: Pannexin1 spielt eine wichtige Rolle bei physiologischen und pathologischen
Vorgängen, wie der Regulation der Immunabwehr, oder dem ischämischen Zelltod.
Mechanismen zur Regulation von Pannexin1-Kanal-Aktivität sind bisher noch wenig
erforscht. Da bekannt ist, dass der Lebenszyklus der aus der Gap junction-
Proteinfamilie stammenden und mit den Pannexinen nah verwandten Connexine auf
verschiedenste Weise durch Phosphorylierung gesteuert wird, wurden in vorliegender
Arbeit potentielle Phosphorylierungsstellen von Pannexin1 untersucht.
Methode: Mittels gezielter Mutagenese wurden neun potentielle
Phosphorylierungsstellen von Pannexin1 durch Aminosäureaustausch mutiert. Die
resultierenden Mutanten wurden mit Hilfe eukaryoter Expressionsvektoren in N2A-
Zellen exprimiert. Anhand von Western Blot-Analysen, konfokaler
Fluoreszenzmikroskopie und elektrophysiologischen Messungen wurden transfizierte
N2A-Zellen anschließend untersucht.
Ergebnis: Es zeigte sich, dass alle mit den verschiedenen Panx1-Mutanten
transfizierten Zellen potentiell überlebensfähig waren und funktionsfähige Panx1-
Kanäle bildeten. Phosphorylierung scheint somit kein Mechanismus zur Regulation der
Panx1-Biosynthese, oder des Transports der Kanäle zur Membran zu sein. Mutationen
der Aminosäuren Y10 im N-Terminus, sowie S343 und S328 im C-Terminus führten
zur Ausbildung eines permanent offenen Kanals und Zelltod. Zudem zeigten Y324F-
mutierte Panx1-Kanäle eine gesteigerte Leitfähigkeit. Die übrigen Mutationen
verursachten im Vergleich zum Pannexin1 Wildtyp-Kanal keine wesentlichen
Änderungen der Zelleigenschaften.
Diskussion: Pannexin1 ist sowohl an physiologischen als auch an pathologischen
Prozessen in Zellen beteiligt. Die vorgelegte Arbeit zeigt erstmals, dass
posttranslationale Phosphorylierung von Panx1 ein wichtiger Mechanismus zur
Regulation von Zelleigenschaften ist. Ein besseres Verständnis dafür, wie Pannexine
reguliert werden, kann einen wichtigen Beitrag zur Untersuchung und Therapie
Pannexin1 bedingter krankhafter Zustände, wie dem entzündungsinduzierten
neuronalen Zelltod im ischämischen Gehirn leisten.

I
Inhaltsverzeichnis
1 EINLEITUNG 1
1.1 Gap Junctions 1 1.1.1 Connexine 3 1.1.2 Innexine 5 1.1.3 Pannexine 6
2 ZIELSETZUNG 13
3 MATERIAL 14
3.1 Organismen 14 3.1.1 N2A-Zelllinie 14
3.2 Vektoren 14
3.3 Medien und Lösungen 14 3.3.1 Zellkulturmedien und Lösungen 14 3.3.2 Lösungen für die Protein-Biochemie 15 3.3.3 Lösungen für die Elektrophysiologie 15
3.4 Chemikalien und Antikörper 16 3.4.1 Chemikalien 16 3.4.2 Antikörper 17 3.4.3 Verwendete Kits 17
3.5 Geräte 18 3.5.1 Apparaturen für die Zellkultur 18 3.5.2 Apparaturen für die Protein-Biochemie 18 3.5.3 Apparatur für die Konfokalmikroskopie 18 3.5.4 Apparaturen für die Elektrophysiologie 18
4 METHODEN 20
4.1 Zellkultur 20
4.2 Transfektion 20
4.3 Vektoren 20
4.4 Seitenspezifische Mutagenese 21
4.5 Gelelektrophorese von Gesamtproteinextrakt 22
4.6 Western Blot 22
4.7 Konfokalmikroskopie 23

II
4.8 Elektrophysiologie 23 4.8.1 Elektrophysiologie in transfizierten N2A-Zellen 23 4.8.2 Analyse elektrophysiologischer Daten 24
5 ERGEBNISSE 26
5.1 Expressionsanalyse 27
5.2 Zelluläres Verteilungsmuster und Überlebenszeit 29
5.3 Elektrophysiologie in N2A-Zellen 30 5.3.1 Vergleich der Untersuchungsmethoden IV-Kurve und Rampe 31 5.3.2 Einfluss der Vektoren pEGFP-N3 und pIRES2-EGFP auf die
Zelleigenschaften 33 5.3.3 Zelleigenschaften 35 5.3.4 Carbenoxoloneinfluss 39
6 DISKUSSION 46
6.1 Elektrophysiologische Analyse der Zelleigenschaften transfizierter N2A-Zellen 47
6.2 Die Verwendung der Vektoren pEGFP-N3-m-Panx1 und pIRES2-EGFP-mPanx1 hat keinen Einfluss auf die Eigenschaften von mPanx1 48
6.3 Phosphorylierung spielt eine wichtige Rolle in der Regulation von Panx1 49
6.3.1 Alle Mutanten bilden funktionsfähige Pannexin1-Kanäle aus 50 6.3.2 Die N-terminale Mutante Y10F führt zu strukturellen Veränderungen
des Panx1-Kanals 51 6.3.3 Mutationen putativer Phosphorylierungsstellen in der intrazellulären
Schleife von Panx1 haben keinen Einfluss auf die Eigenschaften des Kanals im Vergleich zu Panx1 wt 53
6.3.4 C-Terminale Panx1-Phosphorylierungsstellen sind wichtig für die Regulation von Panx1-Kanaleigenschpaften 55
7 ZUSAMMENFASSUNG 59
8 LITERATURVERZEICHNIS 61
9 ANHANG 69
10 LEBENSLAUF 78

III
Abkürzungsverzeichnis A Ampere
ACSF artifizielle cerebrospinale Flüssigkeit
APS Ammoniumpersulfat
Aqua dest. destilliertes Wasser
cAMP zyklisches Adenosin-Mono-Phosphat
Cbx Carbenoxolon
Cx Connexin
D-MEM Dulbecco´s modifiziertes Eagle Medium
EDTA Ethylendiamintetraessigsäure (engl.
„ethylene diaminetetraacetic acid“)
EGFP enhanced green fluorescence protein
EGTA Ethylenglycoltetraessigsäure (engl.
„ethylene glycol tetraacetic acid“)
ER Endoplasmatisches Retikulum
FCS fetales Kälberserum (engl. „fetal calf
serum“)
FFA Flufenaminsäure
HEPES 2-(4-(2-Hydroxyethyl)-1-piperazinyl)-
ethansulfonsäure
Inx Innexin
IRES interne ribosomale Eintrittsstelle (engl.
„internal ribosome entrysite“)
IP3 Inositol-Tris-Phosphat
LCC Kanäle mit grosser Leitfähigkeit (engl.
„large conductance channels“)
LY Lucifer Yellow
m Maus
NEA nicht-essentielle Aminosäuren (engl.
„non-essential amino acids“)
nt nicht transfizierte Zellen
Panx Pannexin

IV
PBS Phosphatpuffer (engl. „phosphate
buffered saline“)
PMSF Phenylmethylsulfonylfluorid
SDS-PAGE Natriumdodecylsulfat Polyacrylamid
Gelelektrophorese (engl. „sodium
dodecylsulfate polyacryamide gel
electrophoresis“)
TEMED N,N,N`,N`-Tetramethylethylendiamin
Tris Tris(hydroxymethyl)-aminomethan
wt Wildtyp
V Volt
vo „vector only“

V
Tabellenverzeichnis Tabelle 1: Auflistung von Vektoren, die mittels in vitro Rekombination gebildet
wurden. 14
Tabelle 2: Liste der Erstantikörper für die Western Blots. 17
Tabelle 3: Liste der Zweitantikörper für die Western Blots. 17
Tabelle 4: Oligonukleotide für die seitenspezifische Mutagenese. 21

VI
Abbildungsverzeichnis Abbildung 1: Struktureller Vergleich der drei Gap junction-Proteinfamilien. 2
Abbildung 2: Struktureller Aufbau von Gap junctions. 4
Abbildung 3: Lokalisation potentieller Phosphorylierungsstellen. 26
Abbildung 4: Expressionsanalyse von mPanx1-Mutanten mit modifizierten potentiellen Phosphorylierungsstellen in N2A-Zellen. 28
Abbildung 5: Zelluläres Verteilungsmuster von mPanx1 und Überlebenszeit von mPanx1-Mutanten mit modifizierten potentiellen Phosphorylierungsstellen in N2A-Zellen. 29
Abbildung 6: Vergleich der Stimulationsprotokolle „IV-Kurven“ und „Rampen“ anhand der maximal ausgelösten Membranströme, gemessen in transfizierten N2A-Zellen. 32
Abbildung 7: Vergleich der Messergebnisse pIRES2-EGFP-mPanx1-transfizierter N2A-Zellen mit denen pEGFP-N3-mPanx1-transfizierter N2A-Zellen am Beispiel der Maximalströme der IV-Kurven. 34
Abbildung 8: Elektrophysiologische Charakterisierung der Panx1 wt und der mutierten mPanx1-Kanäle in N2A-Zellen mittels IV-Kurven. 35
Abbildung 9: Quantitative Analyse der mittleren maximalen Stromamplituden der nur den Vektor exprimierenden Zellen (vo) und des Panx1 wt und seiner Mutanten in N2A-Zellen. 37
Abbildung 10: Quantitative Analyse der mittleren Eingangswiderstände der nur den Vektor exprimierenden Zellen (vo) und des Panx1 wt und seiner Mutanten in N2A-Zellen. 38
Abbildung 11: Quantitative Analyse des mittleren S-Index von nur den Vektor exprimierenden Zellen (vo) und dem Panx1 wt und seinen Mutanten in N2A-Zellen. 39
Abbildung 12: Einfluss von Cbx auf gemittelte IV-Kurven der nur den Vektor exprimierenden Zellen (vo) und des mPanx1 wt seiner Mutanten in N2A-Zellen. 41

VII
Abbildung 13: Vergleich des Einflusses von Cbx auf die IV-Kurven der nur den Vektor exprimierenden Zellen (vo) und des Panx1 wt und seine Mutanten in N2A-Zellen. 42
Abbildung 14: Quantitative Analyse des Cbx-Effekts auf die mittleren maximalen Stromamplituden der nur den Vektor exprimierenden Zellen (vo), und des Panx1 wt und seine Mutanten in N2A-Zellen. 44
Abbildung 15: Quantitative Analyse des Cbx-Effekts auf den mittleren S-Index der nur den Vektor exprimierenden Zellen (vo) und des Panx1 wt und seine Mutanten in N2A-Zellen. 45

1
1 Einleitung
1.1 Gap Junctions Die Entstehung vielzelliger Organismen erforderte die Entwicklung neuer
Kommunikationswege zwischen den Zellen eines Zellverbandes. Die Stelle, an
der die Zytoplasmata zweier benachbarter Zellen direkt mit einander in
Verbindung treten, wird als „Gap junction“ bezeichnet. Fast alle Zelltypen der
Vertebraten bilden Gap junctions aus, mit wenigen Ausnahmen: Erythrozyten,
Spermien und reife Skelettmuskelzellen kommunizieren nicht über Gap
junctions (Dermietzel und Spray, 1993). Der Begriff Gap junction bezeichnet
einen fleckförmigen Bereich einer Zellmembran, innerhalb dessen zahlreiche
Einzelkanäle nebeneinanderliegen. Die Entfernung zwischen den
aneinandergrenzenden Zellen im Gewebe, die normalerweise etwa 20 nm
beträgt, ist hier auf 3,5 nm verringert (Revel und Karnovsky, 1967). Die Gap
junctions überbrücken diese Entfernung, indem sich zwei
membrandurchspannende Hemikanäle der gegenüberliegenden Membranen
aneinander lagern. Sie bilden eine gemeinsame Kanalpore mit einem relativ
großen Durchmesser von bis zu 1,2 nm aus (Bennett und Zukin, 2004; s.
Review Maeda und Tsukihara, 2011). Aufgrund dieses relativ grossen
Durchmessers gelangen nicht nur Ionen direkt von einer Zelle in die nächste,
sondern auch Moleküle mit einer Größe von bis zu 1 kDa (s. Review Maeda
und Tsukihara, 2011). Dazu gehören Metabolite wie Glukose (Tabernero et al.,
1996), sekundäre Botenstoffe wie Kalzium, Inositoltriphosphat (IP3), oder
zyklisches Adenosin-Mono-Phosphat (cAMP) (Tsien und Weingart, 1974) und
Nukleotide (Pitts und Simms, 1977). Zudem ist über Gap junctions eine
bidirektionale Fortleitung von elektrischen Signalen möglich.
Entdecker der Gap junctions waren Furshpan und Potter, die 1959 bei der
Untersuchung von Flusskrebs Riesenmotorneuron-Synapsen nach elektrischer
Reizung eine kurze Latenz zwischen präsynaptischem Stimulus und
postsynaptischer Antwort beobachteten, die sich nicht durch chemische
Übertragung erklären ließ. Seither unterscheidet man in Neuronen chemische

2
und elektrische Synapsen, die Gap junctions. An elektrischen Synapsen sind
prä- und postsynaptische Zellen direkt miteinander verbunden, das heißt es
besteht eine zytoplasmatische Kontinuität. Vorteil eines solchen funktionellen
Synzytiums ist die unverzögerte Fortleitung elektrischer Signale, die z.B. für
Flucht- und Abwehrreaktionen von entscheidender Bedeutung sein kann. Bei
der Meeresschnecke Aplysia zum Beispiel kann ein Stimulus über elektrisch
gekoppelte Motorneurone eine synchronisierte Aktivierung der Tintendrüse und
Ausschüttung einer schützenden Tintenwolke bewirken (Carew und Kandel,
1976). Aber nicht nur die Funktion und Synchronisation neuronaler Netzwerke
(s. Review Zoidl und Dermietzel, 2002) bis hin zum epileptischen Anfall (Traub
et al., 2002; Volman et al., 2011), sondern auch die synchronisierte Aktivität von
Kardiomyozyten und glatten Muskelzellen, z.B. im Uterus, beruht auf der
schnellen elektrischen Signalfortleitung via Gap junctions. Eine veränderte
Anzahl, Zusammensetzung, Verteilung oder Aktivierung von Gap junctions im
Herzen kann infolgedessen zu Erkrankungen des Herzens, wie etwa einer
erhöhten Arrhythmogenität, oder Herzinfarkten führen (Kanno und Saffitz, 2001;
Severs et al., 2001; s. Review Salameh und Dhein, 2011).
Jeder Hemikanal besteht aus sechs identischen Protein-Untereinheiten, jede
dieser Untereinheiten wiederum aus vier Transmembrandomänen (Kumar und
Gilula, 1996).
Bisher konnten drei verschiedene Proteinfamilien identifiziert werden:
Connexine, Innexine und Pannexine.
Abbildung 1: Struktureller Vergleich der drei Gap junction-Proteinfamilien. In Vertebraten werden die Gap junctions von der Proteinfamilie der Connexine gebildet. Innexine und Pannexine gehören zu einer Superfamilie, sie kommen in Invertebraten und Vertebraten vor. Alle drei Proteintypen bestehen aus vier Transmembrandomänen (M1-M4), zwei extrazellulären Schleifen (EL1, EL2) und einer intrazellulären Schleife, sowie dem intrazellulär lokalisierten Amino- und Carboxyterminus. Connexine tragen in den extrazellulären Schleifen jeweils drei hochkonservierte Cysteine (markiert mit „C“), die für die interzelluläre Ausbildung der Gap junction essentiell sind. Innexine und Pannexine weisen hier dagegen lediglich jeweils zwei Cysteine auf. (Grafik freundlicherweise zur Verfügung gestellt von Helga Schulze).

3
1.1.1 Connexine In Vertebraten werden Gap junctions von der Proteinfamilie der
„Connexine“ (Cx) gebildet. Das menschliche Genom enthält 21 Gene für
Connexine, das der Maus 20. Davon sind 19 sequenz-ortholog (Söhl und
Willecke, 2003). Man unterscheidet verschiedene Connexin-Typen, die mit
„Cx“ und einem Zusatz bezeichnet werden, der auf das vorhergesagte
Molekulargewicht des Connexins hinweist (Beyer et al., 1987). So hat zum
Beispiel das so genannte „Cx43“ ein Molekulargewicht von 43.036 Da. Alle
Connexine haben dieselbe Sekundärstruktur: sie bestehen aus vier α-helikalen
Transmembrandomänen, je einem intrazellulären C- und N-Terminus, sowie
einer intrazellulären und zwei extrazellulären Schleifen (Hertzberg et al., 1988; s.
Review Yeager und Harris, 2007). Die extrazellulären Schleifen tragen drei
hochkonservierte Cysteine, die für die interzelluläre Ausbildung der Gap
junctions essentiell sind (Kumar und Gilula, 1996; Bao et al., 2004). Die
Transmembrandomänen und die extrazellulären Schleifen sind hochkonserviert,
während die intrazelluläre Schleife und der C- und N-Terminus sowohl in ihrer
Länge als auch in ihrer Sequenz variieren können (s. Review Yeager und Harris,
2007). Aufgrund dieser Variationen werden die Connexine in drei Unterklassen
eingeteilt: α, β und γ. Sechs Connexine mit jeweils vier Transmembrandomänen
lagern sich zu einem „Connexon“, dem Hemikanal zusammen (Makowski et al.,
1977). Connexone können heteromer oder homomer sein und haben dann
jeweils unterschiedliche Eigenschaften (Elfgang et al., 1995). Als
„homomer“ bezeichnet man Connexone, die aus einem Typ Connexin bestehen,
als „heteromer“ dementsprechend solche, die aus verschiedenen Unterklassen
der Connexine zusammengesetzt sind (Dermietzel, 1998). Zwei Hemikanäle
aneinandergrenzender Zellen können letztlich eine Gap junction bilden. Diese
Kanäle können wiederum heterotyp oder homotyp sein. Als „homotyp“ werden
solche Gap junctions bezeichnet, die aus zwei identischen Hemikanälen
gebildet werden, „heterotyp“ sind Kanäle aus einer Kombination
unterschiedlicher Hemikanäle. Ihre Zusammensetzung scheint die Leitfähigkeit,
die Permeabilität und die Spannungsteuerung der Gap junctions zu
beeinflussen (Manthey et al., 2001).

4
Abbildung 2: Struktureller Aufbau von Gap junctions. (A) Jedes Connexin besteht aus vier Transmembrandomänen (M1-M4) und zwei extrazellulären Schleifen (EL1, EL2). Der N- und C-Terminus liegen intrazellulär. (B) Jeweils sechs Connexine lagern sich zu einem Connexon, dem Hemikanal zusammen. Zwei Connexone gegenüberliegender Zellen bilden die membrandurchspannende interzelluläre Gap junction. (C) Meist liegen in der Zellmembran mehrere Einzelkanäle fleckförmig in so genannten Gap junction Plaques zusammen. Die Entfernung zwischen aneinandergrenzenden Zellen ist hier von normalerweise 20 nm auf 3,5 nm verringert. In 1 und 3 sind homomere Connexone, die aus einem Typ Connexin bestehen, dargestellt, Beispiel 2 zeigt heteromere Connexone, die aus verschiedenen Connexinen gebildet werden. Wird eine Gap junction aus zwei Hemikanälen derselben Zusammensetzung geformt (Beispiel 1 und 2), so heißt dies homotyp, eine Zusammensetzung verschiedener Connexone zu einer Gap junction wird hingegen als heterotyp bezeichnet (Beispiel 3) (Grafik freundlicherweise zur Verfügung gestellt von Helga Schulze).
Connexine existieren nicht nur als interzelluläre Gap junctions, also in
aneinandergrenzenden Zellmembranen, sondern auch als funktionelle
Hemikanäle nicht-kommunizierender Membranen. Die Hemikanäle sind hier
Kommunikationswege zwischen dem Zytoplasma und dem Extrazellulärraum
und haben z.B. an der Regulation des Zellvolumens, oder der parakrinen
Freisetzung von ATP, NAD+ oder Glutamat, Teil (Goodenough und Paul, 2003;
s. Review Goodenough und Paul, 2009). Neu synthetisierte Connexine werden
ko-translational in die Membran des endoplasmatischen Retikulums eingefügt
und oligomerisieren im Golgi-Apparat zu Hemikanälen. Von dort werden sie in
Transportvesikeln mittels Mikrofilamenten und Mikrotubuli zur Zellmembran

5
transportiert (Olk et al., 2009). Der Abbau von Connexonen erfolgt aus der
Mitte eines Gap junction Plaques in Vesikeln, die „annular junctions“ genannt
werden, sowohl lysosomal aus der Plasmamembran als auch proteosomal aus
dem endoplasmatischen Retikulum (Jordan et al., 2001; Laird, 2006). Da
Connexine eine sehr kurze Halbwertszeit zwischen 1,5 und 5 Stunden haben
(Laird, 2006), kann über die Geschwindigkeit von Synthese und Abbau der
Connexine auch direkt die Funktion der Zellen und Zellverbände reguliert
werden. Es sind sowohl verschiedenste transkriptionelle, als auch
posttranskriptionelle Regulationsmechanismen zur exakten Einstellung der
mRNA-Menge beschrieben (s. Review Sáez et al., 2003).
Die meisten Connexine sind Phosphoproteine (Cooper et al., 2000), so dass
weitere funktionelle Variabilität durch Phosphorylierung entsteht. Von der
Synthese, über den Transport zur Membran, den Zusammenbau, den Einbau in
die Membran, die Spannungssteuerung, bis zum Abbau der Kanäle werden
viele Phasen des Connexin-Lebenszyklus mittels Phosphorylierung gesteuert (s.
Review Sáez et al., 2003; s. Review Solan und Lampe, 2009).
1.1.2 Innexine Innexine (Inx) wurden ursprünglich in Drosophila melanogaster und
Caenorhabditis elegans entdeckt und man nahm an, dass sie die strukturellen
Komponenten der Invertebraten Gap junctions seien (Phelan et al., 1998b;
Phelan und Starich, 2001). Der Name Innexin leitet sich von „Invertebraten
Analogon der Connexine“ ab. Heute weiß man, dass Innexine auch in
Vertebraten vorkommen, und vermutet, dass Innexine und Pannexine zu einer
Superfamilie gehören (Yen und Saier, 2007; Fushiki et al. 2010). Obwohl
Innexine keine Sequenzhomologie zu den Connexinen aufweisen, sind die
Proteinstrukturen dennoch ähnlich: genau wie Connexine bestehen Innexine
aus vier Transmembrandomänen, sowie intrazellulären N- und C-Termini
(Phelan und Starich, 2001). Im Gegensatz zu den Connexinen tragen Innexine
in ihren extrazellulären Schleifen jedoch nur zwei hochkonservierte Cysteine
(Phelan und Starich, 2001). Ihre physiologischen Eigenschaften ähneln
trotzdem denen der Connexine (Phelan et al., 1998a).

6
1.1.3 Pannexine Der Begriff „Pannexin“ leitet sich von griechisch „pan“ (alles, gesamt) und
lateinisch „nexus“ (Verbindung, Verknüpfung) ab und wurde von Panchin und
Mitarbeitern geprägt (Panchin et al., 2000), um die Innexine der Invertebraten
und ihre Vertebraten-Homologe als eine ubiquitär vorkommende Proteinfamilie
zusammenzufassen. Sowohl im Maus- als auch im menschlichen Genom sind
drei Pannexin-kodierende Gene vorhanden die als PANX1, PANX2 und PANX3
bezeichnet werden (Panchin et al. 2000). Die Panx1 mRNA kommt ubiquitär im
Körper vor, im zentralen Nervensystem, wie dem Cortex, dem Striatum, dem
Bulbus olfactorius, dem Hippocampus, dem Thalamus, dem Kleinhirn und der
unteren Olive konnte zusätzlich Panx2 mRNA nachgewiesen werden (Bruzzone
et al. 2003; Ray et al. 2005; Weickert et al., 2005). Auf zellulärer Ebene
unterscheidet sich jedoch die Verteilung der Panx1 und Panx2 mRNA: im
Hippocampus wird zum Beispiel sowohl Panx1 als auch Panx2 in GABAergen
Interneuronen exprimiert. Im Kleinhirn jedoch kommt Panx1 mRNA besonders
reichlich in der weißen Substanz vor während Panx2 mRNA in den Purkinje-
Zellen festgestellt werden konnte (Bruzzone et al., 2003). Panx3 mRNA scheint
dagegen nicht im ZNS vorzukommen sondern in der Haut, in Osteoblasten und
in Fibroblasten (Panchin et al., 2000; Bruzzone et al., 2003; Baranova et al.,
2004). Pannexine weisen, wie Innexine und Connexine auch, vier
Transmembrandomänen, eine intra- und zwei extrazelluläre Schleifen, sowie
einen intrazellulären N- und C-Terminus auf. Und wie Innexine, nicht jedoch
Connexine, tragen Pannexine nur jeweils zwei Cysteine in den extrazellulären
Schleifen (Phelan und Starich, 2001).
Neueren Untersuchungen von Dahl und Mitarbeitern (Ambrosi et al., 2010)
zufolge bilden sowohl Panx1, als auch Panx2 homomere Kanäle aus. Dabei
scheinen die homomeren Panx2-Kanäle keine hexamere Anordnung
aufzuweisen, wie dies für Panx1-Kanäle angenommen wird, sondern sie
scheinen oktamere Kanäle zu formen. Heteromere Kanäle aus Panx1 und
Panx2 sind dagegen sehr instabil und scheinen in vivo nicht funktionell zu sein.
Ob Panx3 ebenfalls homomere Kanäle ausbilden kann, bleibt fraglich.
Bruzzone et al. (2003) konnten in elektrophysiologischen Untersuchungen
zumindest keine funktionellen homomeren Panx3-Kanäle nachweisen.

7
Es ist erwiesen, dass Pannexine funktionelle nicht-junctionale Kanäle ausbilden
und so eine Aufgabe ähnlich normaler Ionenkanäle ausüben können, indem sie
das Zytoplasma einer Zelle mit dem Extrazellulärraum verbinden. Eine Funktion
als Gap Junction konnte dagegen bisher nicht nachgewiesen werden (Penuela
et al., 2007; Boassa et al., 2007; Sosinsky et al., 2011). Für Panx1 konnte
zudem von Zoidl und Mitarbeitern (Zoidl et al., 2007) mittels Western Blot und
elektronenmikroskopischer Immunhistochemie eine überwiegend
postsynaptische Lokalisation in Pyramidenzellen des Hippocampus gezeigt
werden. Dies unterstreicht die Funktion von Pannexinen als nicht-junctionale
Kanäle. Andererseits liegen sowohl Studien vor, die den Übertritt von
Farbstoffen von einer Zelle in die nächste über Pannexine, und damit Gap
junctions, beschreiben (Vanden Abeele et al, 2006), als auch solche, die einen
Farbstoffübertritt zwischen benachbarten Zellen verneinen (Huang et al., 2007).
Allerdings ist Panx1 unter physiologischen Bedingungen an extrazellulären
Positionen glykosyliert, was die Formation zweier Pannexin-Kanäle zu
funktionellen Gap junction verhindern kann (Boassa et al., 2007). Es ist also
wahrscheinlich, dass Pannexine in vivo ausschließlich als nicht-junctionale
Kanäle vorkommen.
Pharmakologische und physiologische Eigenschaften von Pannexinen Da zwei nicht verwandte Gap junction-Proteinfamilien in Vertebraten existieren,
ist anzunehmen, dass sie unterschiedliche Funktionen in Zellverbänden erfüllen.
Zur Unterscheidung zwischen Connexin- und Pannexin-Eigenschaften dienen
ihre pharmakologischen und physiologischen Merkmale. Flufenaminsäure
(FFA) ist ein bekannter Chlorid-Kanal-Blocker, der auch Connexin-Kanäle
reversibel blockiert (Harks et al., 2001). Pannexine werden dagegen kaum von
FFA beeinflusst, sondern effektiv durch Carbenoxolon (Cbx) reversibel blockiert.
Cbx blockiert zwar auch Connexone, aber nur in sehr hoher Konzentration.
Zudem ist der Cbx-Effekt bei Connexonen geringer, als bei Pannexonen
(Bruzzone et al., 2005; Weihong et al., 2009). 2004 wandten Bao et al. die
Patch-clamp-Technik an, um weitere Eigenschaften der Pannexin1-Kanäle in
Xenopus-Oozyten zu analysieren. Sie stellten fest, dass die
Einzelkanalleitfähigkeit bei ca. 500 pS liegt, während Connexin-Hemikanäle
lediglich eine Leitfähigkeit zwischen 18 pS (Cx32, Gomez-Hernandez et al.,

8
2003) und 220 pS (Cx43, Contreras et al., 2003) zeigen. Zudem stellte sich
heraus, dass die Pannexin1-Kanäle in fünf Öffnungszuständen vorkommen
können: Neben einem komplett offenen Zustand existieren vier weitere
Öffnungszustände mit geringerer Leitfähigkeit von 5%, 25%, 30% und 90% der
maximalen Leitfähigkeit (Bao et al., 2004). Da Pannexin-Kanäle eine sehr
große Kanalpore bilden, durch die wichtige Metabolite verloren gehen und
extrazelluläres Kalzium ungehindert einströmen könnte, liegen die Kanäle meist
in einem geschlossenen Zustand vor und die Öffnung des Kanals ist streng
reguliert. Bei einem negativen Haltepotential ist der Pannexin1-Kanal
geschlossen (Bruzzone et al., 2003), öffnet jedoch bei positiven
Haltepotentialen (Bao et al., 2004). Außerdem kann eine Öffnung des Kanals
durch mechanischen Stress (Bao et al., 2004), extrazelluläres ATP, Anstieg der
intrazellulären Kalziumkonzentration (Bruzzone et al., 2005), Sauerstoffmangel
oder Entzündungszustände hervorgerufen werden, während ein
zytoplasmatischer pH-Abfall zum Kanal-Verschluss führt (Locovei et al., 2006).
All diese Eigenschaften kommen, wenn überhaupt, nur in wenigen Connexonen
vor. Zusätzlich sind die Pannexin-vermittelten Ströme bei negativen
Haltepotentialen in Patch-clamp Versuchen grösser, als bei positiven
Haltepotentialen, so dass die Kanäle eine Gleichrichterfunktion ausüben
können (Bao et al., 2004).
Funktion von Pannexinen In tierischen Zellen hat Kalzium eine wichtige Bedeutung für die Erregbarkeit,
die Aktivierung von Enzymen und Hormonen, sowie als intrazellulärer second
messenger. Interzelluläre Kalziumwellen spielen eine Rolle in der Regulation
des Zilienschlags des Epithels der Atemwege (Sanderson et al., 1990), oder
der Modulation synaptischer Übertragung zwischen Neuronen (Newman, 2001).
Für die Fortleitung solcher Kalziumwellen über Zellgrenzen hinaus sind
Pannexine essentiell. Die mechanische Stimulation von Zellen führt zum
Anstieg intrazellulären IP3. IP3 kann seinerseits Kalzium aus IP3-sensitiven
Speichern freisetzen und über Gap junctions in benachbarte Zellen diffundieren,
wo es weitere Kalzium-Speicher öffnet und zur Ausbildung einer interzellulären
Kalziumwelle beiträgt (Sanderson et al., 1990). Bevor die Existenz von
Pannexinen bekannt war, nahm man an, dass die hierfür benötigten Kanäle aus

9
Connexonen gebildet werden. Connexine reagieren jedoch auf einen Anstieg
der intrazellulären Kalzium-Konzentration sehr sensitiv mit einem Verschluss
ihrer Kanalpore und könnten deshalb nur an einer sehr frühen Phase der
Kalziumwellen teilhaben (Sanderson et al., 1994). Pannexine andererseits
beantworten eine Erhöhung der intrazellulären Kalziumkonzentration mit einer
Öffnung ihrer Kanalpore und scheinen deshalb besser geeignet, Kalziumwellen
weiterzuleiten. Locovei et al. konnten 2006 nachweisen, dass Pannexin1-
Kanäle sowohl Kalziumwellen auslösen als auch fortleiten können. Mittels
elektrophysiologischer Ableitungen an Xenopus-Oozyten, die Pannexin und
P2Y1 exprimieren, konnten sie folgendes zeigen: Ein durch mechanischen
Stress geöffnetes Pannexon führt zur Freisetzung von ATP durch den
Pannexin-Kanal in den Extrazellulärraum. Extrazelluläres ATP aktiviert dann
einen purinergen Rezeptor vom Typ P2Y oder P2X, dies führt zum Anstieg der
intrazellulären IP3-Produktion und zur Freisetzung intrazellulären Kalziums. Mit
dem Anstieg des intrazellulären Kalziums werden weitere Pannexin-Kanäle
geöffnet und mehr ATP in den Extrazellulärraum freigesetzt, dort kann es zu
benachbarten Zellen diffundieren und diese über purinerge Rezeptoren
aktivieren. Diese Art der Ausbreitung von Kalziumwellen wäre also unabhängig
von der Existenz von Gap junctions und funktioniert auch mit Pannexinen, die
lediglich nicht-junctionale Ionenkanäle bilden.
Dieser Mechanismus der ATP-Freisetzung konnte schnell mit der bei Ischämie
auftretenden Kalzium-Dysregulation in Verbindung gebracht werden
(Thompson et al., 2006). Gleichzeitig können über Panx1- und P2X-Rezeptoren
G-Protein gekoppelte Rezeptoren aktiviert werden, die während und nach
ischämischen Bedingungen kardioprotektive Signalwege in den Zellen auslösen
(Vessey et al., 2011).
Die ATP-Freisetzung via Pannexin1-Kanäle scheint auch eine Rolle in der
Verarbeitung von Geschmacksinformationen in den Papillae vallatae der Maus
zu spielen. Vermutlich führt die Stimulation von Rezeptorzellen in
Geschmacksknospen zu einer Freisetzung von ATP durch Pannexin1-Kanäle.
Das ATP kann dann direkt afferente Axone der Geschmacksnerven oder
angrenzende präsynaptische Zellen aktivieren (Dando und Roper, 2009). Wie
die Pannexin1-Kanäle in den Geschmacksknospen aktiviert werden, konnten
2010 Huang und Roper aufdecken: eine Erhöhung der intrazellulären

10
Kalziumkonzentration oder eine Membrandepolarisation führen zur Öffnung der
Kanäle.
Außerdem kann ATP als lösliches „find-me“-Signal apoptotischer Zellen
Phagozyten anlocken und so zur Aufrechterhaltung der Gewebshomöostase
beitragen. 2010 konnten Chekeni et al. Pannexin1-Kanäle als Freisetzungsweg
für ATP in apoptotischen Zellen identifizieren. Pharmakologische Hemmung
des Panx1-Kanals konnte die ATP-Freisetzung vermindern, während die
Überexpression des Kanals zu einer verstärkten Freisetzung führte. Sie
identifizierten außerdem eine Caspasen-Schnittstelle in Panx1, die ebenfalls
eine wichtige Funktion in der Apoptose hat: Caspasen-Aktivität führt zur
Ausbildung eines permanent offenen Panx1-Kanals mit darauf folgendem
Zelltod.
Auch bei der Regulation der Immunabwehr spielt ATP und seine Freisetzung
via Pannexin1-Kanäle eine Rolle (Woehrle et al., 2010a; Woehrle et al., 2010b).
Pannexin1-Kanäle können als Reaktion auf einen Reiz mit hypertoner
Salzlösung ATP freisetzen. Das Nukleotid ATP führt über eine Bindung an P2X-
Rezeptoren zu einer gesteigerten T-Zell-Funktion, indem es z.B. die Produktion
des T-Zell-Wachstumsfaktors Interleukin-2 steigert. Die posttraumatische T-
Zell-Anergie und damit Immunsuppression wird somit über eine ATP-
Freisetzung über Pannexin1-Kanäle vermindert.
Kürzlich konnte mittels Patch-clamp-Technik eine weitere Funktion von
Pannexinen aufgedeckt werden: so genannte LCC-Kanäle („large conductance
channels“) wurden in kardialen Myozyten aufgrund ihrer pharmakologischen
Eigenschaften in elektrophysiologischen Experimenten und durch Expression
mittels adenoviraler Vektoren als Panx1-Kanäle identifiziert. Die Öffnung dieser
Kanäle könnte zu einem Kalziumeinstrom und Membrandepolarisationen mit
Auslösung spontaner Aktionspotentiale und damit arrhythmogener Aktivität im
Herzen beitragen (Kienitz et al., 2011).
Zusammenfassend kann man sagen, dass für Pannexin1 mittlerweile eine
Vielzahl an Funktionen sowohl in physiologischen, als auch in pathologischen
Reaktionen von Zellen auf verschiedenste Reize bekannt ist. So spielen, wie
bereits erwähnt, Panx1-Kanäle eine wichtige Rolle in der Freisetzung von ATP
und damit der Auslösung und Weiterleitung von Kalziumwellen, der Apoptose,

11
der Verarbeitung von Geschmacksinformationen, oder der Regulation der
Immunabwehr. Darüber hinaus wird für Panx1 eine Rolle in der Vasodilatation,
bei Entzündungsmechanismen, dem ischämischem Zelltod, der Epilepsie, und
der Tumorsuppression angenommen. Die spontane Öffnung von Panx1-
Kanälen im Herzen könnte außerdem zur Auslösung arrhythmogener Aktivität
beitragen.
Über die Aufgabe von Panx2 und Panx3 weiß man dagegen immer noch
wesentlich weniger. Um ihre Funktion im Gewebe bestimmen zu können,
müssen weitere Untersuchungen zu ihren physiologischen und
pharmakologischen Eigenschaften folgen. Einige wenige Versuche haben
bereits stattgefunden: Pannexin3 scheint die Chondrozyten-Proliferation und
Differenzierung im Knorpel zu regulieren. Zumindest konnte Panx3-mRNA in
der Knorpelwachstumszone nachgewiesen werden (Iwamoto et al. 2010). Die
Herunterregulation von Panx3 in Chondrozyten verhinderte die Differenzierung
spezieller Knorpelzellen (ATDC5), während die Transfektion mit Panx3-mRNA
die chondrogene Differenzierung förderte. Zudem konnte durch die Transfektion
mit Panx3-mRNA die Freisetzung von ATP in den Extrazellulärraum begünstigt
werden, so dass auch für den Panx3-Hemikanal eventuell eine Funktion in der
Freisetzung von ATP angenommen werden kann.
Panx3-mRNA konnte auch in der Haut nachgewiesen werden und könnte hier
für die Aufrechterhaltung der epidermalen Architektur zuständig sein. So
scheint die Überexpression von Panx3 in Keratinozyten der Rattenepidermis
zur Aufrechterhaltung des embryonalen Aufbaus der Epidermis zu führen
während die Überexpression von Panx1 eine Dysregulation der
Keratinozytendifferentierung auslöst (Celetti et al., 2010).
Auch Pannexin2 könnte eine Rolle bei der Differenzierung von Zellen spielen.
Swayne et al. konnten mit Hilfe kurzer Haarnadel-RNA im Jahr 2010 die Panx2-
Expression in Neuro2A-Zellen inhibieren und damit die Rate neuronaler
Differenzierung beschleunigen. Die posttranslationale Veränderung von Panx2
scheint also den Zeitpunkt mitzubestimmen, an dem Neuronenvorläuferzellen
sich zu differenzieren beginnen. In Gliomzellen könnte Panx2 außerdem als
negativer Wachstumsfaktor wirken, da die Panx2-Expression in
Genexpressionsanalysen von Lai et al. 2009 in C6 Gliom-Zellen von Ratten

12
erheblich herunterreguliert war. In vitro konnten Onkogenitätsparameter wie
Morphologie und die Zahl der Zell-Zell-Kontakte der Gliomzellen durch eine
verstärkte Panx2-Expression reduziert werden.

13
2 Zielsetzung
Pannexin1 spielt eine wichtige Rolle bei physiologischen und pathologischen
Vorgängen im Menschen wie z.B. der Regulation der Immunabwehr (Woehrle
et al., 2010a; Woehrle et al., 2010b), oder der Auslösung arrhythmogener
Aktivität im Herzen (Kienitz et al., 2010). Einblicke in die Art und Weise, wie
Pannexin1-Aktivität gesteuert wird, hat also einen bedeutenden Anteil an der
Erforschung solcher Prozesse.
Tatsächlich sind die Regulationsmechanismen von Pannexin1 bisher noch
nicht ausreichend untersucht, anders als etwa die Mechanismen
posttranslationaler Modifikation von Connexinen. Obwohl die Gap junction-
Proteinfamilien der Connexine und Pannexine keine Sequenzhomologie
aufweisen, ist ihre Sekundärstruktur gleich (Hertzberg et al., 1988; Phelan et al.,
1998a; Phelan und Starich, 2001) und sowohl Connexine, als auch Pannexin1
werden aus sechs Untereinheiten gebildet (Makowski et al., 1977; Boassa et al.,
2007). Es ist deshalb anzunehmen, dass die Kanaleigenschaften dieser beiden
Familien ähnlich reguliert werden. Während von Connexinen bekannt ist, dass
sie als Phosphoproteine eine große funktionelle Variabilität durch
Phosphorylierung erreichen können, weiß man in dieser Hinsicht noch wenig
über Pannexine (Review Sáez et al., 2003). Tatsächlich weisen die mittlere
zytoplasmatische Schleife und der C-Terminus Konsensus-Stellen für
Proteinkinasen auf, so dass man davon ausgehen kann, dass auch Pannexine
mittels Phosphorylierung in ihren Eigenschaften verändert werden können
(Barbe et al., 2006).
In vorliegender Arbeit wurde daher der Frage nachgegangen, ob Pannexin1
ebenfalls posttranslational phosphoryliert wird und welchen Einfluss diese
Modifikation auf die Zelleigenschaften hat. Zu diesem Zweck sollten potentielle
Phosphorylierungstellen in Panx1 mittels gezielter Mutagenese mutiert und mit
Hilfe eukaryoter Expressionsvektoren in N2A-Zellen transfiziert werden.
Anschließend sollten transfizierte Zellen anhand von Western Blot-Analysen,
konfokaler Fluoreszenzmikroskopie und elektrophysiologischen Messungen
untersucht werden.

14
3 Material
3.1 Organismen
3.1.1 N2A-Zelllinie N2A-Zellen sind Neuroblastom-Zellen aus Mus musculus (Klebe und Ruddle,
1969). Sie wurden freundlicherweise von Dr. David Spray (Albert Einstein
College, NY, USA) zur Verfügung gestellt.
Der Umgang mit dieser Eukaryoten-Zelllinie erfolgte in einem lizensierten S1-
Labor der Abteilung für Neuroanatomie und Molekulare Hirnforschung der
Ruhr-Universität Bochum, Deutschland.
3.2 Vektoren Tabelle 1 enthält eine Übersicht über die in dieser Arbeit verwendeten Vektoren. Tabelle 1: Auflistung von Vektoren, die mittels in vitro Rekombination gebildet wurden.
Vektor Eingefügtes Gen
Verwendungszweck
pEGFP-N3 mPanx1 eukaryotischer Expressionsvektor für ein Fusionsprotein aus mPanx1 (N-Terminus) und EGFP (C-Terminus)
pIRES2-EGFP
mPanx1 eukaryotischer Expressionsvektor für mPanx1 und EGFP (Transkription von bicistronischer mRNA)
3.3 Medien und Lösungen
3.3.1 Zellkulturmedien und Lösungen Der Phosphatpuffer und die Trypsin-EDTA-Lösung wurden zum Passagieren
der Zellen benötigt.
Phosphatpuffer (PBS, phosphate buffered saline)
0, 138 M NaCl, 0,0028 M KCl, 0,01 M Na2HPO4, 0,0018 M KH2PO4, pH 7,4

15
3.3.2 Lösungen für die Protein-Biochemie Folgende Puffer wurden für die Gelelektrophorese und die Western Blots
benötigt:
Laemmli-Probenpuffer
2% SDS (Natriumdodecylsulfat), 10% Glycerol, 10% ß-Mercaptoethanol, 50 mM Tris-HCl, pH 6,8
Laemmli-Laufpuffer
25 mM Tris-HCl pH 8,3, 192 mM Glycin, 0,1% SDS, ad 1l Aqua dest.
Transferpuffer
48 mM Tris, 39 mM Glycin, 0,037% SDS, 20% Methanol, ad 1l Aqua dest.
Lysispuffer
0,25 mM Natriumdesoxycholsäure, 150 mM NaCl, 2,0 mM EDTA, 0,1 mM Na3VO4, 10 mM NaF, 1 mM PMSF (Phenylmethylsulfonylfluorid), 1% Triton X 100, 50 mM Tris-HCl, pH 7,4
3.3.3 Lösungen für die Elektrophysiologie ACSF (artifizielle cerebrospinale Flüssigkeit)
Die ACSF wurde zur Perfusion der N2A-Zellen während der
elektrophysiologischen Messungen verwendet.
Zunächst wurden 25 ml der Lösung A mit Aqua dest. auf 437,5 ml aufgefüllt und
10 min mit Carbogen (95% O2; 5% CO2) begast. Anschließend wurden 25 ml
der Lösung B hinzugegeben, es folgten weitere 10 min Carbogen-Begasung,
um den pH-Wert einzustellen. Die Carbogen-Begasung der Lösung wurde
Trypsin-EDTA-Lösung (PAA)
0,05% Trypsin, 0,02% EDTA (Ethylendiamintetraessigsäure)
N2A-Kulturmedium Das N2A-Medium diente zur Kultivierung der N2A-Zellen. D-MEM (4,5 g/l Glucose, +L-Glutamin, +Pyruvat;
Gibco, Invitrogen, Carlsbad, CA, USA) + 5% FCS (fetales Kälberserum) + 1% L-Glutamin + 1% Penicillin/Streptomycin + 1% NEA (nicht essentielle Aminosäuren) + 1% Na+-Pyruvat

16
während der Perfusion der Zellen fortgesetzt um den pH-Wert konstant zu
halten und die Zellen mit Sauerstoff zu versorgen.
Lösung A 2,438 M NaCl, 0,049 M KCl, 0,039 M CaCl2, 0,025 M MgCl2, 0,024 M NaH2PO4, 0,244 M D-Glukose, 0,0078 M L-Ascorbat
Lösung B
0,488 M NaHCO3
Pipettenlösung Die Patch-Pipetten wurden mit folgender Pipettenlösung (Pelegrin et al., 2006)
gefüllt:
Stammlösung
0,147 M NaCl, 0,01 M HEPES (2-(4-(2-Hydroxyethyl)-1-piperazinyl)-ethansulfonsäure), 0,003 M MgCl2
Für die Messungen wurde 1 ml dieser Stammlösung in 10 ml Aqua dest. gelöst
und mit 0,038 g EGTA (Ethylenglycoltetraessigsäure) versetzt. Der pH-Wert
wurde anschließend mit KOH auf 7,3 eingestellt.
Pharmaka Carbenoxolon (3ß-Hydroxy-11-oxoolean-12-en-30-oic acid 3-hemisuccinate
disodium) ist ein Derivat der Glycyrrhizinsäure und wurde in einer Konzentration
von 20 µM als Kanal-Blocker eingesetzt.
3.4 Chemikalien und Antikörper
3.4.1 Chemikalien Acrylamid (AppliChem, Darmstadt, Deutschland), Ammoniumpersulfat (APS;
Sigma, Taufkirchen, Deutschland), Blocking Reagenz (Roche, Mannheim,
Deutschland), Calziumchlorid (J.T. Baker), Carbenoxolon (Sigma; MP
Biomedicals, Solon, OH, USA), D-Glukose (J.T. Baker, Freising, Deutschland),
Dinatriumhydrogenphosphat (J.T. Baker), Dulbecco´s modifiziertes Eagle
Medium (D-MEM; Invitrogen, Karlsruhe, Deutschland), Effectene Transfection
Reagent Kit (Qiagen; Hilden, Deutschland), Ethylendiamintetraessigsäure
(EDTA, Titriplex III; Merck, Darmstadt, Deutschland),
Ethylenglycoltetraessigsäure (EGTA;), fetales Kälberserum (FCS; BiochromKG,
17
Berlin, Germany), Glukose (J.T. Baker), Glycin (AppliChem), Glycerin (J.T.
Baker), Hoechst 33342 (Invitrogen), Kaliumchlorid (Merck),
Kaliumdihydrogenphosphat (J.T. Baker), Kaliumhydroxid, L-Ascorbat, L-
Glutamin (PAA, Cölbe, Deutschland), Magnesiumchlorid (J.T. Baker), ß-
Mercaptoethanol (Sigma), Methanol (J.T. Baker), Natriumchlorid (J.T. Baker),
Natriumdodecylsulfat (SDS; AppliChem), Natriumdihydrogencarbonat (J.T.
Baker), Natrium-Pyruvat (PAA), Nicht essentielle Aminosäuren (NEA; PAA),
Odyssey Blocking Puffer (Licor Biosciences, Lincoln, NE, USA), Penicillin (PAA),
Phenylmethylsulfonylfluorid (PMSF; Sigma), Penicillin/Streptomycin (PAA),
Tetramethylethylendiamin (TEMED; AppliChem), Tris(hydroxymethyl)-
aminomethan (Tris; AppliChem), Trypsin-EDTA (PAA), Tween20 (AppliChem)
3.4.2 Antikörper Tabelle 2 enthält eine Übersicht über die in dieser Arbeit für die Western Blots
verwendeten Erstantikörper. Tabelle 2: Liste der Erstantikörper für die Western Blots.
Name Spezies Hersteller Spezifität β-Actin (clone AC-15)
Maus Sigma β-Actin
Panx1 4515 Huhn Aves Labs (Tigard, OR, USA); freundliche Spende von Gerhard Dahl
mPanx1
Living Colors A.v. Peptide Antibody
Kaninchen Clontech EGFP, ECFP, EYFP
Die Zweitantikörper für die Western Blots sind in Tabelle 3 aufgelistet. Tabelle 3: Liste der Zweitantikörper für die Western Blots.
Name Spezies Hersteller
anti-chicken Cy 5.5 Ziege abcam
anti-mouse IRDye 800 Ziege Licor Biosciences
anti-rabbit IRDye 680 Ziege Licor Biosciences
3.4.3 Verwendete Kits Transfektion Effectene Transfection Reagent Kit
(Qiagen; Hilden, Deutschland)

18
3.5 Geräte
3.5.1 Apparaturen für die Zellkultur Inkubator Hera-cell (Heraeus, Hanau,
Deutschland)
Sterilbank LaminAir 1.2 (Heto-Holten, Allerød, Dänemark)
3.5.2 Apparaturen für die Protein-Biochemie Blotting-Kammer Trans-Blot SD Semi-dry Transfer Cell
(BioRad, Hercules, CA, USA)
Natriumdodecylsulfat Polyacrylamid Gelelektrophorese (SDS-PAGE)
PerfectBlueTM Vertikales Doppelgelsystem Twin ExWs (peqlab Biotechnologie GmbH, Erlangen, Deutschland)
Nitrozellulosemembran Hybond-ECL Nitrozellulosemembran (Amersham Biosciences, GE Healthcare, Buckinghamshire, GB)
Infrarot Western Blot Detektor System Odyssey Infrared Imaging System (Licor Biosciences)
3.5.3 Apparatur für die Konfokalmikroskopie Konfokales Laser Scanning Mikroskop LSM510-Meta (Zeiss)
3.5.4 Apparaturen für die Elektrophysiologie Fluoreszenz-Mikroskop Axioskop (Zeiss, Göttingen,
Deutschland)
Messpipetten Glass Capillary Tubing (1,5-1,1 mm, Typ G75150T-4, Warner-Instrument Corporation, Hamden, CT, USA)
Vertikalpuller PIP5 Vertikalpuller (HEKA, Lambrecht, Deutschland)
Verstärker Axopatch 200B (Axon Instruments, Forster City, CA, USA)

19
Frequenzfilter Humbug 50/60 Hz Noise Eliminator
(Quest Scientific, North Vancouver, BC, Kanada)
AD/DA-Wandler CV 203 BU AD/DA-Wandler (Axon Instruments)
Software WinWCP Software (Strathclyde Institut für Pharmazie und Biomedizinische Wissenschaften, Universität Strathclyde, GB) SigmaStat (SysStat Software GmbH, San José, CA, USA)

20
4 Methoden
4.1 Zellkultur Die N2A-Zellen wurden bei 37°C und 5% CO2 in N2A-Kulturmedium in 10 cm
Ø-Kulturschalen in einem CO2-Inkubator kultiviert. Alle zwei bis drei Tage
wurden sie passagiert. Hierzu wurden die Zellen zunächst mit 5 ml PBS
gewaschen und anschließend in 1,5 ml Trypsin-EDTA für 5 min bei 37°C im
Wärmeschrank inkubiert. Mit Hilfe von N2A-Medium konnten die Zellen dann
suspendiert und vereinzelt werden. Die Zellen wurden für 5 min bei 200 g
zentrifugiert und anschließend in 5 ml Medium resuspendiert. 0,3 ml (dies
entspricht etwa 1,5x106 Zellen) dieser Suspension wurden dann zu 10 ml
Kulturmedium hinzugefügt und in 10 cm Ø-Kulturschalen ausgesät.
4.2 Transfektion Die transiente Transfektion der N2A-Zellen wurde mittels des Effectene
Transfection Reagent Kits (Qiagen) den Herstellerangaben folgend
durchgeführt. Für die elektrophysiologischen Versuche wurden 15 000 Zellen
auf poly-(L)-Lysin-beschichteten Deckgläschen ausgesät. Für die Herstellung
von Proteinextrakten wurden je 30 000 Zellen pro well in 24-well Platten in N2A-
Kulturmedium inkubiert. Am nächsten Tag wurde die transiente Transfektion mit
200-400 ng Plasmid-DNA durchgeführt. Weitere 24 Stunden später wurde das
Medium gewechselt, um zytotoxische Effekte der Transfektions-Reagenzien zu
minimieren.
4.3 Vektoren N2A-Zellen wurden mit zwei verschiedenen eukaryotischen
Expressionsvektoren transfiziert. Zum einen wurde der pIRES2-EGFP-mPanx1-
Vektor eingesetzt. Er enthält ein Expressionskassette zur bicistronischen
Expression der beiden Proteine mPanx1 (Pannexin 1 (Maus)) und EGFP
21
(enhanced green fluorescent protein). Zum andern wurde der pEGFP-N3-
mPanx1-Vektor genutzt der eine Expressionskassette beinhaltet die für ein
Fusionsprotein aus mPanx1 und C-terminal lokalisiertem EGFP kodiert.
4.4 Seitenspezifische Mutagenese Die seitenspezifische Mutagenese basiert auf einer von Carter im Jahr 1987
entwickelten Methode. Zur Untersuchung der Funktion möglicher
Phosphorylierungsstellen wurden mit Hilfe gezielter Mutagenese diejenigen
Aminosäuren im mPanx1 mutiert, die laut NetPhos 2.0 Server
(http://www.cbs.dtu.dk/services/NetPhos/) einen Vorhersagewert von größer 0,5
hatten. Tabelle 4: Oligonukleotide für die seitenspezifische Mutagenese. Die mutierten Nukleotide sind mit fett gedruckten Großbuchstaben markiert. Die Sequenz ist in 5´ 3´Richtung angegeben.
Name DNA-Sequenz [5´- 3´] Aminosäure-Sequenz
Vorher-sagewert
mPanx1-Y10F ccacggagtTtgtgttctcgg LATEYVFSD 0,718 mPanx1-Y192F
ccacttcaagtTcccaatcgtgg SHFKYPIVE 0,663
mPanx1-Y324F
gtctgaaggctTcaatgacttgagcc KSEGYNDLS 0,941
mPanx1-T387A
cgtccctacagGccaagggagagg TSLQTKGED 0,780
mPanx1-S159A
ggctgccaagGCtgctcgagatttgg KAAKSARDL 0,998
mPanx1-S206A
caaaaaagaactctGCtcatttaatcatg KKNSSHLIM 0,728
mPanx1-S328A
ggctacaatgacttgGCcctctacaacc YNDLSLYNL 0,580
mPanx1-S343A
gtgagctcaaaGcgtacaagtgtctg SELKSYKCL 0,996
mPanx1-S405A
gatttggacctgAGcagcgaggctgc DLDLSSEAA 0,981
Die Plasmide mit der mutierten Pannexin-DNA wurden freundlicherweise von
Dr. Stefanie Bunse, Abteilung für Neuroanatomie und Molekulare Hirnforschung
der Ruhr-Universität Bochum, Deutschland, produziert und zur Verfügung
gestellt.

22
4.5 Gelelektrophorese von Gesamtproteinextrakt Mit Hilfe der SDS-Polyacrylamid Gelelektrophorese (SDS-PAGE) konnten
Proteine aus Proteinextrakten nach der von Laemmli 1970 eingeführten
Methode nach ihrem Molekulargewicht aufgetrennt werden (Laemmli, 1970).
Hierzu wurde das PerfectBlue vertikale Doppelgelsystem verwendet. Zur Lyse
von 30 000 transfizierten Zellen wurden 100 µl Lysispuffer auf die Zellen
gegeben. Anschließend wurde das Lysat bei 4°C und 13000 rpm für 5 min
zentrifugiert. Dann wurden 10 µl des Lysats mit 2,5 µl Laemmli-Probenpuffer
vermischt, für 5 min bei 95°C denaturiert und anschließend auf das Gel
aufgetragen. Die Gelelektrophorese wurde bei 30-40 mA durchgeführt. Um eine
Überhitzung des Gels zu vermeiden wurde eine Wasserkühlung (15°C)
angeschlossen.
Folgende Komponenten wurden benötigt:
Trenngel
8-10% Acrylamid, 380 mM Tris-HCl (Tris(hydroxymethyl)-aminomethan), pH 8,8, 0,1% SDS, 0,08% TEMED (Tetramethylethylendiamin), 0,08% APS (Ammoniumpersulfat)
Kammgel
5% Acrylamid, 125 mM Tris-HCl pH 6,8, 0,1% SDS, 0,08% TEMED, 0,15% APS
4.6 Western Blot Im Anschluss an die elektrophoretische Auftrennung der Proteine nach ihrem
Molekulargewicht wurden die Proteine auf eine Nitrozellulosemembran
übertragen. Dazu wurden vier Transferpuffer-getränkte Whatmann Papiere in
die Blotting-Kammer gelegt und darauf die Nitrozellulosemembran und das
SDS-Gel platziert. Darauf folgten zwei weitere puffergetränkte Whatmann
Papiere. Der Proteintransfer erfolgte dann bei 250 mA für 1 h. Anschließend
wurde die Nitrozellulosemembran dreimal für jeweils 10 min mit PBS
gewaschen, bevor unspezifische Bindung der Antikörper an die
Nitrozellulosemembran mittels Odyssey Blocking Reagenz blockiert wurde (1 h
bei Raumtemperatur). Anschließend wurde der Erstantikörper zugegeben und
die Membran wurde über Nacht bei 4°C inkubiert. Am zweiten Tag wurden alle

23
ungebundenen Antikörper entfernt in dem die Membran dreimal für jeweils 10
min mit PBS-Tween 0,1% gewaschen wurde. Im Anschluss wurde die Membran
mit Zweitantikörper, verdünnt mit der Odyssey Blocking Reagenz, für 1 h bei
Raumtemperatur unter Lichtabschluss inkubiert. Die Membran wurde dann
dreimal für jeweils 10 min mit PBS gewaschen, um die ungebundenen
Zweitantikörper zu entfernen. Mit Hilfe des Odyssey Infrarot Imaging Systems
konnten die Signale dann detektiert und dokumentiert werden.
4.7 Konfokalmikroskopie Die Analyse konfokaler Bilder wurde mit dem LSM 510-Meta System
durchgeführt. Dieses war mit Argon- und Helium-Neon-Lasern, einer Laser-
Diode (Emissionswellenlänge: 405 nm), einem 40x (NA 1,4) und einem 63x (NA
1,4) Ölobjektiv und der LSM 510-Meta Software ausgestattet. Die Aufnahmen
wurden im „single track“-, „multi track“-, oder „online fingerprinting“-Modus
durchgeführt. Die Bilder konnten dann mit 1024x1024 Pixeln gespeichert und
als TIFF-Datei in AdobePhotoShop exportiert und bearbeitet werden.
4.8 Elektrophysiologie
4.8.1 Elektrophysiologie in transfizierten N2A-Zellen Alle Messungen wurden mittels der Patch-Clamp-Technik als
Ganzzellableitungen gemäß der von Neher und Sakmann (1976) entwickelten
Methode durchgeführt.
Die mit transfizierten N2A-Zellen bedeckten Deckgläschen wurden in die
Messkammer eingelegt, die bei einer Flussgeschwindigkeit von 2 ml/min
kontinuierlich mit ACSF durchspült wurde. Mittels dieses Zulaufs konnte
außerdem der Hemikanalblocker Carbenoxolon Bad-appliziert werden.
Messpipetten aus Borosilikatglas wurden mit einem Vertikalpuller so hergestellt,
dass sie nach Füllen mit der Pipettenlösung Impedanzen zwischen 6 und 9 MΩ
aufwiesen. Mit Hilfe des Fluoreszenz-Mikroskops wurden unter visueller
Kontrolle grün (EGFP) fluoreszierende und damit transfizierte Zellen
ausgewählt. Unter Durchlichtbeleuchtung konnten die ausgewählten Zellen

24
anschließend in die Ganzzellkonfiguration gebracht und über die Membran
fließende Ströme gemessen werden.
Als Hauptverstärker wurde der Axopatch 200B eingesetzt, mit Hilfe eines
zusätzlichen Frequenzfilters konnten zudem weitere Störsignale gefiltert werden.
Die verstärkten und gefilterten analogen Messdaten wurden über den AD/DA-
Wandler digitalisiert und zum Computer übertragen. Sie konnten hier mit der
Software „Win WCP“ sichtbar gemacht und für die spätere Analyse gespeichert
werden.
Zur Charakterisierung elektrophysiologischer Eigenschaften der abgeleiteten
Zellen wurden zunächst Strom-Spannungskurven (IV-Kurven) aufgenommen.
Dazu wurde das Membranpotential der Zelle auf -30 mV geklemmt und dann
Spannungssprünge zwischen -50 mV und +60 mV in 10 mV Schritten ausgelöst.
Die Spannungssprünge hatten dabei eine Dauer von 500 ms. Außerdem
wurden 200 ms dauernde, kontinuierlich ansteigende Stromrampen verwendet,
wobei die Zellen von -100 mV bis +80 mV depolarisiert wurden. Für jede Zelle
wurden zunächst abwechselnd je zwei IV-Kurven und Stromantworten auf die
Stromrampen unter Kontrollbedingungen in Anwesenheit normaler ACSF
gemessen, bevor eine kontinuierliche Bad-Applikation von Carbenoxolon für 8
min erfolgte. Um den Cbx-Effekt verfolgen zu können, wurde während der
Applikation alle 2 min je eine IV-Kurve und die Antwort auf eine
Spannungsrampe aufgenommen. Dabei wurden nicht nur die
elektrophysiologischen Parameter der abgeleiteten Zellen mit mutierten
Hemikanäle bestimmt, sondern zur Kontrolle auch die Eigenschaften von Zellen,
die entweder nur das EGF-Protein enthielten, oder EGFP-mPx1 transfiziert
waren.
Abgeleitete Zellen konnten außerdem mit Hilfe einer Mikroskopkamera
fotografiert und ihre Morphologie anschließend verglichen werden.
4.8.2 Analyse elektrophysiologischer Daten Zur späteren Charakterisierung der elektrophysiologischen Parameter
abgeleiteter Zellen wurde der maximal über die Zellen fließende Strom bei
Verwendung der Spannungsrampe bzw. aus den IV-Kurven vor und während
Cbx-Applikation bestimmt. Hieraus konnte der Cbx-Effekt in % errechnet
werden. Zellen, die eine prozentuale Abnahme der Maximal-Amplitude unter

25
Cbx von weniger als 20% gegenüber dem Kontrollwert in normaler ACSF
aufwiesen, wurden in die Auswertung nicht einbezogen. Dies geschah unter der
Annahme, dass solche Zellen keine ausreichende Expression der Pannexin1-
Kanäle aufwiesen. Dieses Auswahlkriterium wurde nicht auf die pEGFP-N3-
transfizierten Kontrollzellen, sowie auf die pIRES2-EGFP-transfizierten
Kontrollzellen angewendet, da diese Zellen keine Kanalproteine exprimieren
konnten. Folglich musste hier jeder vermeintliche Cbx-Effekt auf physiologische
oder pathologische Schwankungen der Membranleitfähigkeiten unabhängig von
Pannexin1-Kanälen zurückzuführen sein.
Außerdem wurde zur Beurteilung der Spannungsabhängigkeit der
Membranleitfähigkeit der so genannte „S-Index“ ermittelt. Hierzu wurde
zunächst die Steigung der IV-Kurve zwischen den ersten beiden
Spannungssprüngen von -60 bis -50 mV (sneg) und zwischen den letzten beiden
Spannungssprüngen von +50 bis +60 mV (spos) bestimmt. Der S-Index wurde
dann mittels folgender Gleichung berechnet:
𝑆 =𝑠!"# − 𝑠!"# 𝑠!"# + 𝑠!"#
Die ersten beiden Messwerte unter Kontrollbedingungen und die Messungen
nach vier, sechs und acht Minuten Carbenoxolon-Applikation wurden jeweils
vor Berechnung des Maximalstroms, bzw. des S-Index gemittelt.
Statistische Signifikanz zwischen den verschiedenen Versuchsgruppen wurde
unter Verwendung der Statistiksoftware SigmaStat mit Hilfe des Student´s T-
Test, des Mann-Whitney Rangsummentests, des „paired t-tests“ oder des
„signed rank tests“ bestimmt.

26
5 Ergebnisse
Aus Experimenten zur Regulation von Connexinen weiß man, dass diese im
Laufe ihres Lebenszyklusses vielfach posttranslational modifiziert werden. Viele
Connexine sind Phosphoproteine und die Phosphorylierung von Connexinen ist
ein wichtiges Mittel zur Regulation biologischer Prozesse in Zellen als
Anpassungsmechanismus an veränderte Umweltbedingungen (siehe Review
Solan und Lampe, 2009). Es ist anzunehmen, dass auch Pannexine
posttranslational modifiziert werden. Da sowohl der N- und C-Terminus, als
auch die intrazelluläre Schleife konservierte Konsensusstellen für Kinasen
aufweisen, ist es sehr wahrscheinlich, dass Pannexine ebenfalls
posttranslational phosphoryliert werden (Barbe et al., 2006). Mit Hilfe gezielter
Mutagenese wurden für diese Arbeit potentielle Phosphorylierungsstellen in
mPanx1 mutiert. Alle mutierten Aminosäuren hatten dabei laut NetPhos2.0
Server einen Vorhersagewert von grösser 0,5. Das Panx1 wurde an insgesamt
neun Stellen genetisch verändert: Drei Tyrosine wurden zu Phenylalanin (Y10F,
Y192F und Y324F), ein Threonin zu Alanin (T387A) und fünf Serine zu Alanin
(S159A, S206A, S328A, S343A und S405A) mutiert, dabei wurde pro Plasmid
nur eine Mutation eingebracht.
Abbildung 3: Lokalisation potentieller Phosphorylierungsstellen. In der Grafik ist die Struktur des Panx1 schematisch dargestellt. Alle potentiellen Phosphorylierungsstellen sind farbig markiert und liegen intrazellulär. In Rot sind Tyrosine, in Blau Serine und in Gelb ein Threonin hervorgehoben. (Grafik freundlicherweise zur Verfügung gestellt von Helga Schulze).

27
5.1 Expressionsanalyse Zum Nachweis der Expression von mPanx1 in transfizierten N2A-Zellen wurde
ein Western Blot durchgeführt. Mit Hilfe spezifischer Erst- und Zweitantikörper
konnten so die mPanx1-Proteine markiert und mittels Odyssey Infrarot Imaging
System dargestellt werden. Für alle Versuche wurden N2A-Zellen entweder mit
dem pIRES2-EGFP- oder dem pEGFP-N3-Vektor transfiziert, die jeweils für den
Panx1 Wildtyp (wt), oder eine der neun Mutanten kodierten. Außerdem wurde
zur Kontrolle jeweils nur der leere Vektor ohne Information für den Panx1-Kanal
transfiziert. Der pIRES2-EGFP-mPanx1-Vektor ermöglicht eine bicistronische
Expression der Proteine mPanx1 und EGFP, und der pEGFP-N3-mPanx1-
Vektor die Expression von Fusionsproteinen aus mPanx1 und C-terminal
lokalisiertem EGFP. Zusätzlich wurden auch nicht transfizierte Zellen eingesetzt
(nt). Zur Kontrolle der aufgetragenen Proteinmenge wurde das β-Aktin aus den
Proben ebenfalls Antikörper gefärbt.
In Abbildung 4 sind die Western Blots gezeigt, dabei ist in Grafik 4 A der
Western Blot der Proteinlysate von den N2A-Zellen, die mit dem pIRES2-
EGFP-Vektor transfiziert waren und in B der Western Blot der Lysate von den
Zellen, die pEGFP-N3 transfiziert waren abgebildet. Grundlegende
Unterschiede waren zwischen den beiden eingesetzten Vektoren aus den
Western Blots nicht zu erkennen. In beiden Western Blots waren für die nicht
transfizierten Kontrollen und die mit dem leeren Vektor transfizierten Zellen
keine spezifischen Banden zu erkennen, damit war in diesen Zellen kein Panx1
nachweisbar. Anhand der Aktinbande ließ sich für alle aufgetragenen Proben
nachweisen, dass Protein aufgebracht wurde. In beiden Blots zeigten die
meisten Mutanten und der Wildtyp außerdem zwischen 36 und 55 kDa bei den
mit dem pIRES2-EGFP-Vektor-transfizierten Zellen und zwischen 55 und 95
kDa bei den mit dem pEGFP-N3-Vektor-transfizierten Zellen, das von Boassa et
al. (2007) als „multiple band pattern“ beschriebene Muster aus Mehrfachbanden,
das für Panx1 typisch ist. Die oberen beiden Banden repräsentierten dabei die
glykosylierten Formen, die untere die nicht glykosylierte Form des Panx1-
Proteins. Die oberste Bande, die die voll glykosylierte Form des Panx1-Proteins
kennzeichnete, wies dabei auf eine Lokalisation von Panx1 in der
Plasmamembran bei den meisten Mutanten (Panx1-Y192F, Y324F, S206
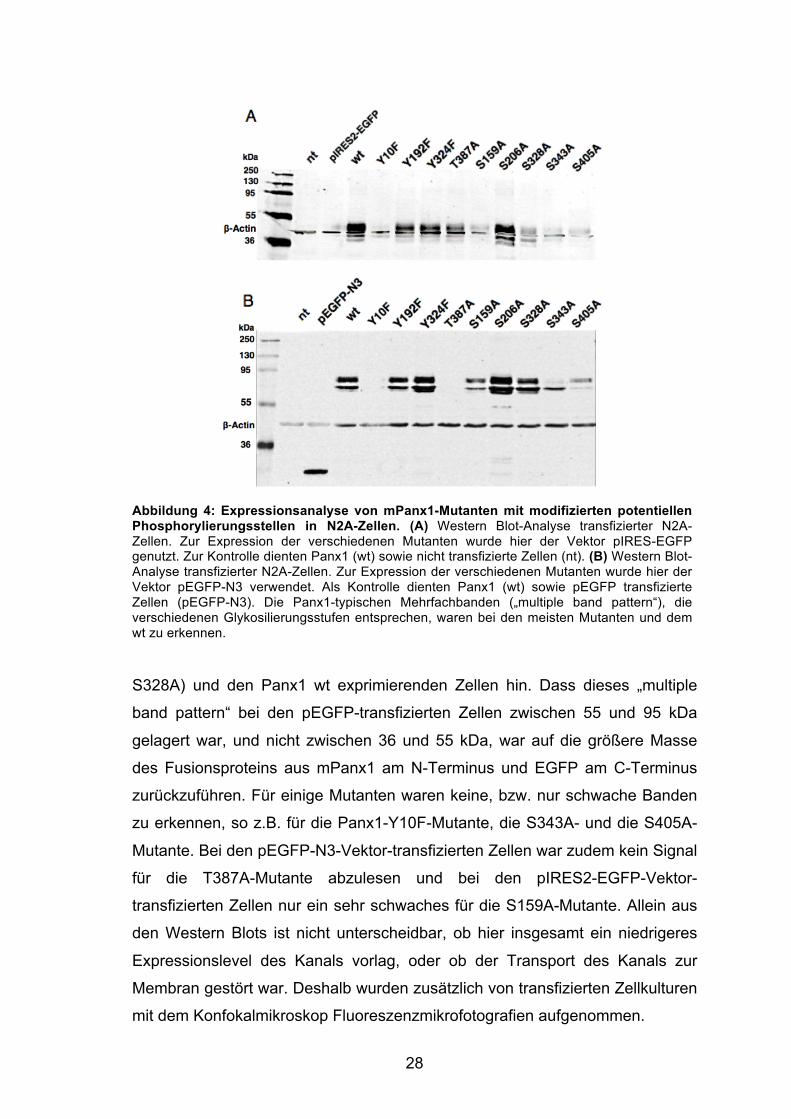
28
Abbildung 4: Expressionsanalyse von mPanx1-Mutanten mit modifizierten potentiellen Phosphorylierungsstellen in N2A-Zellen. (A) Western Blot-Analyse transfizierter N2A-Zellen. Zur Expression der verschiedenen Mutanten wurde hier der Vektor pIRES-EGFP genutzt. Zur Kontrolle dienten Panx1 (wt) sowie nicht transfizierte Zellen (nt). (B) Western Blot-Analyse transfizierter N2A-Zellen. Zur Expression der verschiedenen Mutanten wurde hier der Vektor pEGFP-N3 verwendet. Als Kontrolle dienten Panx1 (wt) sowie pEGFP transfizierte Zellen (pEGFP-N3). Die Panx1-typischen Mehrfachbanden („multiple band pattern“), die verschiedenen Glykosilierungsstufen entsprechen, waren bei den meisten Mutanten und dem wt zu erkennen.
S328A) und den Panx1 wt exprimierenden Zellen hin. Dass dieses „multiple
band pattern“ bei den pEGFP-transfizierten Zellen zwischen 55 und 95 kDa
gelagert war, und nicht zwischen 36 und 55 kDa, war auf die größere Masse
des Fusionsproteins aus mPanx1 am N-Terminus und EGFP am C-Terminus
zurückzuführen. Für einige Mutanten waren keine, bzw. nur schwache Banden
zu erkennen, so z.B. für die Panx1-Y10F-Mutante, die S343A- und die S405A-
Mutante. Bei den pEGFP-N3-Vektor-transfizierten Zellen war zudem kein Signal
für die T387A-Mutante abzulesen und bei den pIRES2-EGFP-Vektor-
transfizierten Zellen nur ein sehr schwaches für die S159A-Mutante. Allein aus
den Western Blots ist nicht unterscheidbar, ob hier insgesamt ein niedrigeres
Expressionslevel des Kanals vorlag, oder ob der Transport des Kanals zur
Membran gestört war. Deshalb wurden zusätzlich von transfizierten Zellkulturen
mit dem Konfokalmikroskop Fluoreszenzmikrofotografien aufgenommen.
29
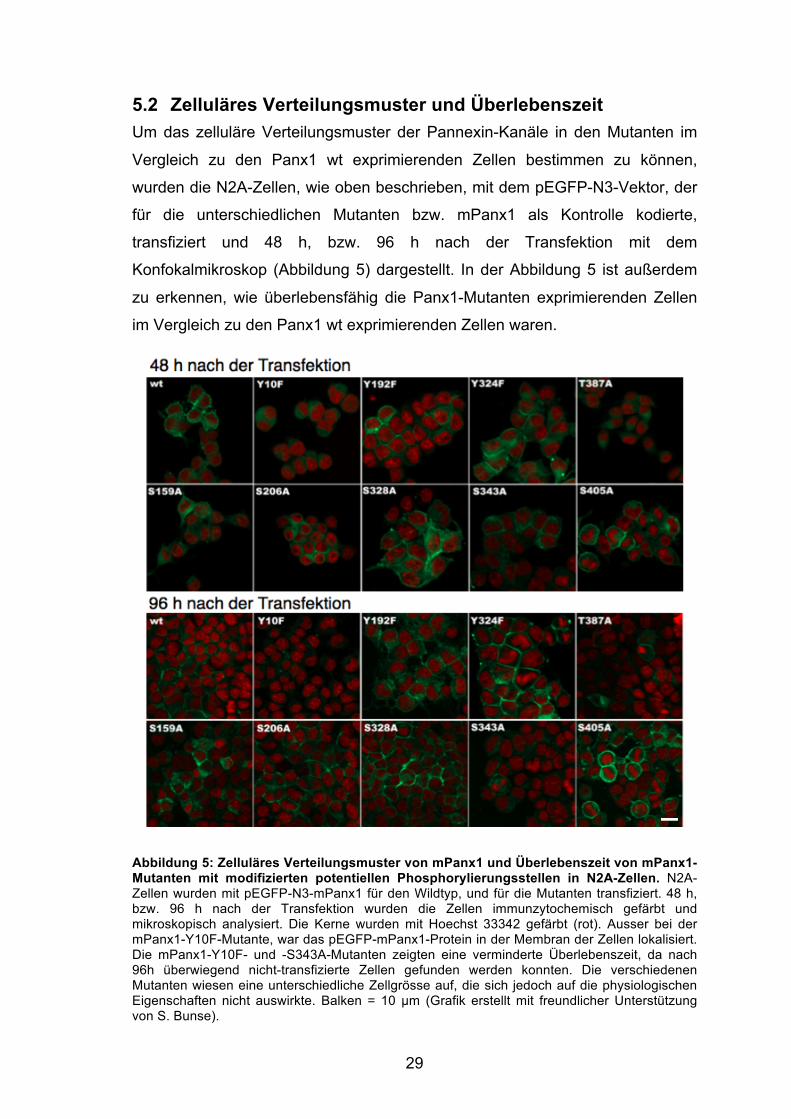
5.2 Zelluläres Verteilungsmuster und Überlebenszeit Um das zelluläre Verteilungsmuster der Pannexin-Kanäle in den Mutanten im
Vergleich zu den Panx1 wt exprimierenden Zellen bestimmen zu können,
wurden die N2A-Zellen, wie oben beschrieben, mit dem pEGFP-N3-Vektor, der
für die unterschiedlichen Mutanten bzw. mPanx1 als Kontrolle kodierte,
transfiziert und 48 h, bzw. 96 h nach der Transfektion mit dem
Konfokalmikroskop (Abbildung 5) dargestellt. In der Abbildung 5 ist außerdem
zu erkennen, wie überlebensfähig die Panx1-Mutanten exprimierenden Zellen
im Vergleich zu den Panx1 wt exprimierenden Zellen waren.
Abbildung 5: Zelluläres Verteilungsmuster von mPanx1 und Überlebenszeit von mPanx1-Mutanten mit modifizierten potentiellen Phosphorylierungsstellen in N2A-Zellen. N2A-Zellen wurden mit pEGFP-N3-mPanx1 für den Wildtyp, und für die Mutanten transfiziert. 48 h, bzw. 96 h nach der Transfektion wurden die Zellen immunzytochemisch gefärbt und mikroskopisch analysiert. Die Kerne wurden mit Hoechst 33342 gefärbt (rot). Ausser bei der mPanx1-Y10F-Mutante, war das pEGFP-mPanx1-Protein in der Membran der Zellen lokalisiert. Die mPanx1-Y10F- und -S343A-Mutanten zeigten eine verminderte Überlebenszeit, da nach 96h überwiegend nicht-transfizierte Zellen gefunden werden konnten. Die verschiedenen Mutanten wiesen eine unterschiedliche Zellgrösse auf, die sich jedoch auf die physiologischen Eigenschaften nicht auswirkte. Balken = 10 µm (Grafik erstellt mit freundlicher Unterstützung von S. Bunse).

30
Schon 48 h nach der Transfektion war die Lokalisation des pEGFP-Panx1-
Proteins an der Zelloberfläche (pEGFP-N3-mPanx1-wt, -Y192F, Y324F, S159A,
S206A, S328A, S343A, S405A) zu erkennen. Das Y10F- und T387A-Panx1-
Protein war hingegen nicht in der Zellmembran nachzuweisen. Eine Ursache
hierfür könnte eine Störung des Transports des Panx1-Proteins zur Membran
sein oder es könnte insgesamt eine niedrigere Expressionsrate vorliegen. Nach
96 h war im Vergleich zu den Panx1 wt exprimierenden Zellen und auch zu den
anderen Mutanten die Gesamtzahl der pEGFP-N3-Y10F und -S343A
transfizierten Zellen geringer. Da bei der Y10F-, und auch bei der S343A-
Mutante, folglich die Überlebensfähigkeit der Zellen herabgesetzt war, ist eine
andere Erklärung für die geringe Menge Panx1-Protein in der Zellmembran
wahrscheinlicher. Alle Zellen, die das Panx1-Y10F-Protein stark exprimierten
waren vermutlich zum Zeitpunkt der Aufnahme schon abgestorben so dass nur
schwach exprimierenden Zellen, die wenig Panx1-Protein gebildet hatten, in
den Bildern zu erkennen waren. Zusätzlich war die Fluoreszenzintensität bei
der S405A-Mutante stärker als beim Wildtyp.
Oben beschriebene Versuche bestätigen, dass von einer Membranständigkeit
des mPanx1-wt-Proteins und der mutierten Varianten in den transfizierten N2A-
Zellen ausgegangen werden konnte und damit dieses Expressionssystem für
elektrophysiologische Untersuchungen der Phosphorylierungsmutanten in N2A-
Zellen verwendet werden konnte.
5.3 Elektrophysiologie in N2A-Zellen Mittels Western Blot-Analysen und konfokaler Fluoreszenzmikroskopie konnten
eindeutige Hinweise darauf gewonnen werden, dass mit den
Expressionsvektoren für Panx1 und die mutierten Varianten transfizierte N2A-
Zellen potentiell überlebensfähig waren und Pannexin-Kanäle ausbildeten.
Bisher konnte kein Unterschied zwischen Zellen, die mit dem pEGFP-N3-Vektor
und solchen, die mit dem pIRES2-EGFP-Vektor transfiziert waren, festgestellt
werden.
Um die elektrophysiologischen Eigenschaften der Panx1-Mutanten im Vergleich
zum Panx1 wt untersuchen zu können, wurden die Expressionsvektoren in
N2A-Zellen transfiziert. Die GFP-Fluoreszenz konnte in den Ableitexperimenten

31
als Marker für erfolgreich transfizierte N2A-Zellen genutzt werden. Von solchen
grün fluoreszierenden Zellen wurden zunächst Strom-Spannungskurven, im
Folgenden mit „IV-Kurven“ bezeichnet, mittels Ganzzellableitungen
aufgenommen. Die Zellen wurden dafür auf ein Membranpotential von -30 mV
geklemmt. Dann wurde das Membranpotential in Spannungssprüngen mit einer
Dauer von 250 ms von zunächst -50 mV in 10 mV-Schritten stufenweise auf
+60 mV geändert. Zusätzlich wurden Stromverläufe bei kontinuierlich
ansteigendem Haltepotential über eine Dauer von 200 ms aufgenommen und
die Zellen dabei von -100 mV auf + 80 mV depolarisiert. Diese Stromverläufe
werden im Folgenden mit „Rampe“ bezeichnet. Jeweils im Anschluss wurde der
Ionenkanal-Blocker Carbenoxolon Bad-appliziert und sein Einfluss auf die
abgeleiteten Ganzzellströme während der Messung der IV-Kurven und der
linearen Spannungsverläufe bestimmt.
5.3.1 Vergleich der Untersuchungsmethoden IV-Kurve und Rampe Zur Bestimmung der elektrophysiologischen Eigenschaften von transfizierten
N2A-Zellen wurden bei den Ganzzellableitungen sowohl IV-Kurven als auch
Strom-“Rampen“ aufgezeichnet. Mit Hilfe der IV-Kurven kann der maximal über
die Zelle fließende Strom und die Spannungsabhängigkeit der Leitfähigkeit
gemessen werden. Unter Verwendung der Stromrampen kann dagegen besser
die Öffnungskinetik der Kanäle bestimmt werden.
Es stellte sich die Frage, welches Stimulationsprotokoll besser zur Analyse der
Zelleigenschaften transfizierter N2A-Zellen geeignet ist. In der statistischen
Auswertung aller Einzelzelldaten zeigte sich, dass für die Werte aus den
Stromrampen genauso häufig signifikante Unterschiede zwischen den Panx1-
Mutanten und den Panx1 wt exprimierenden Zellen festzustellen waren, wie für
die Werte aus den IV-Kurven (s. Tabelle A 2 im Anhang). Ein Vergleich der
Maximalströme, gemessen in den IV-Kurven und Rampen, ist in Abbildung 6
grafisch dargestellt.
Die Maximalströme zeigten für die Messungen in den IV-Kurven und Rampen
gleiche Tendenzen. So war der Maximalstrom der Zellen, die nur mit dem
Vektor transfiziert wurden („vector only“, vo), sowohl in den IV-Kurven, als auch
in den Rampen, signifikant kleiner als in den Panx1 wt exprimierenden Zellen
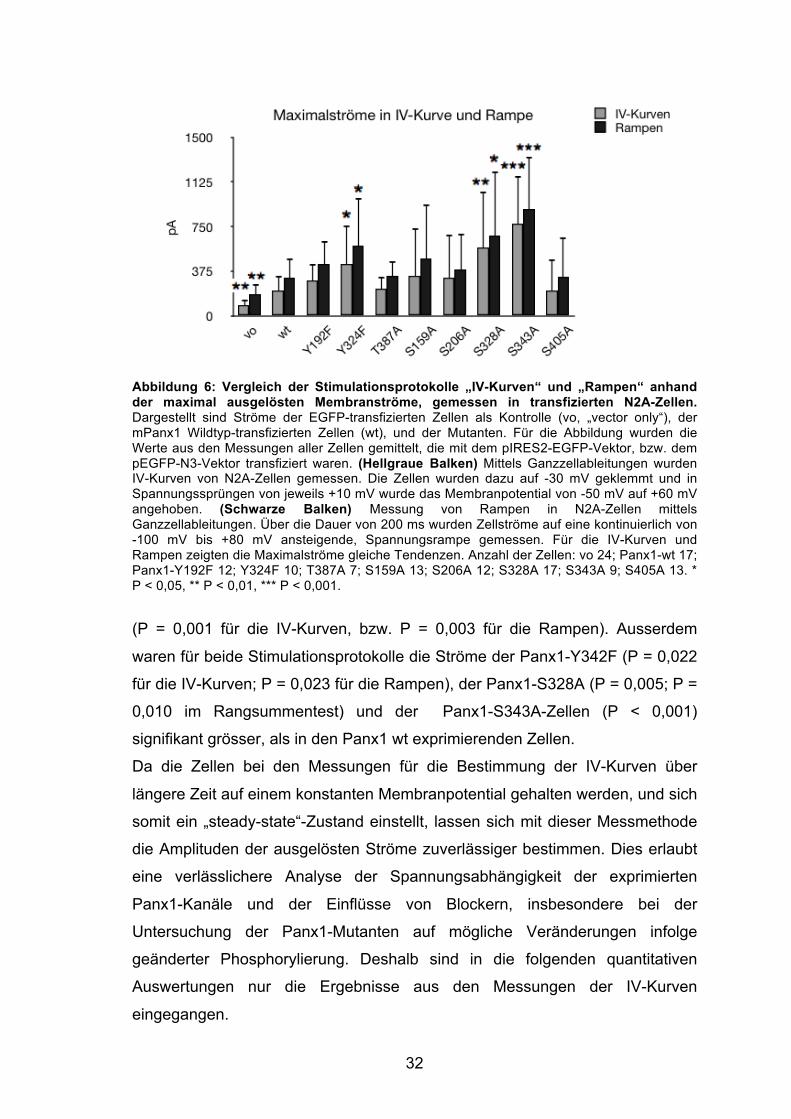
32
Abbildung 6: Vergleich der Stimulationsprotokolle „IV-Kurven“ und „Rampen“ anhand der maximal ausgelösten Membranströme, gemessen in transfizierten N2A-Zellen. Dargestellt sind Ströme der EGFP-transfizierten Zellen als Kontrolle (vo, „vector only“), der mPanx1 Wildtyp-transfizierten Zellen (wt), und der Mutanten. Für die Abbildung wurden die Werte aus den Messungen aller Zellen gemittelt, die mit dem pIRES2-EGFP-Vektor, bzw. dem pEGFP-N3-Vektor transfiziert waren. (Hellgraue Balken) Mittels Ganzzellableitungen wurden IV-Kurven von N2A-Zellen gemessen. Die Zellen wurden dazu auf -30 mV geklemmt und in Spannungssprüngen von jeweils +10 mV wurde das Membranpotential von -50 mV auf +60 mV angehoben. (Schwarze Balken) Messung von Rampen in N2A-Zellen mittels Ganzzellableitungen. Über die Dauer von 200 ms wurden Zellströme auf eine kontinuierlich von -100 mV bis +80 mV ansteigende, Spannungsrampe gemessen. Für die IV-Kurven und Rampen zeigten die Maximalströme gleiche Tendenzen. Anzahl der Zellen: vo 24; Panx1-wt 17; Panx1-Y192F 12; Y324F 10; T387A 7; S159A 13; S206A 12; S328A 17; S343A 9; S405A 13. * P < 0,05, ** P < 0,01, *** P < 0,001.
(P = 0,001 für die IV-Kurven, bzw. P = 0,003 für die Rampen). Ausserdem
waren für beide Stimulationsprotokolle die Ströme der Panx1-Y342F (P = 0,022
für die IV-Kurven; P = 0,023 für die Rampen), der Panx1-S328A (P = 0,005; P =
0,010 im Rangsummentest) und der Panx1-S343A-Zellen (P < 0,001)
signifikant grösser, als in den Panx1 wt exprimierenden Zellen.
Da die Zellen bei den Messungen für die Bestimmung der IV-Kurven über
längere Zeit auf einem konstanten Membranpotential gehalten werden, und sich
somit ein „steady-state“-Zustand einstellt, lassen sich mit dieser Messmethode
die Amplituden der ausgelösten Ströme zuverlässiger bestimmen. Dies erlaubt
eine verlässlichere Analyse der Spannungsabhängigkeit der exprimierten
Panx1-Kanäle und der Einflüsse von Blockern, insbesondere bei der
Untersuchung der Panx1-Mutanten auf mögliche Veränderungen infolge
geänderter Phosphorylierung. Deshalb sind in die folgenden quantitativen
Auswertungen nur die Ergebnisse aus den Messungen der IV-Kurven
eingegangen.

33
5.3.2 Einfluss der Vektoren pEGFP-N3 und pIRES2-EGFP auf die Zelleigenschaften
Die elektrophysiologischen Messungen wurden zunächst mit N2A-Zellen
durchgeführt, die mit dem pEGFP-N3-Vektor transfiziert waren. In diesen
Versuchen enthielt jede fluoreszierende Zelle Fusionsproteine aus Panx1 und
EGFP. Da nicht ausgeschlossen werden konnte, dass bei der Verwendung des
Fusionsproteins die Funktion des Pannexins durch das EGF-Protein beeinflusst
wurde, wurden die Versuche zusätzlich mit dem pIRES2-EGFP-Vektor
wiederholt. Dieser Vektor kodiert für ein bicistronisches mRNA-Transkript,
anhand dessen die Proteine Panx1 und EGFP separat von einander translatiert
werden. So konnte ein möglicher Einfluss des EGFP auf die Funktion des
Panx1 ausgeschlossen werden. Um einen möglichen Einfluss der verwendeten
Vektoren auf die Zelleigenschaften zu untersuchen, wurden die Ergebnisse aus
den Messungen der pIRES2-EGFP-mPanx1-wt-Zellen mit Hilfe des Student´s
T-Tests mit den Ergebnissen aus den Messungen der pEGFP-N3-mPanx1-wt-
Zellen verglichen. Die Ergebnisse aus den Messungen der nur mit den
verschiedenen Vektoren transfizierten Zellen wurden auf die gleiche Weise
statistisch gegenüber gestellt (s. Tabelle A 1 Anhang). Für die nur mit dem
Vektor transfizierten Zellen konnte für zwei Messparameter ein signifikanter
Unterschied zwischen den Vektoren festgestellt werden, zum einen für die
Maximalströme der Rampen (P = 0,009) und zum anderen für den Cbx-Effekt
auf den S-Index (P = 0,003), berechnet aus den Rampen. Dieser Unterschied
kann jedoch nicht auf einen Einfluss des Vektors auf den Panx1-Kanal
zurückzuführen sein, da diese Zellen kein Panx1-Protein exprimierten. Auch im
Vergleich der Ergebnisse aus den Messungen der Panx1-Wildtypzellen
ergaben sich signifikante Unterschiede zwischen den verwendeten Vektoren.
Abweichungen waren für die Maximalströme, gemessen in den IV-Kurven (P =
0,014) und die S-Indizes, berechnet aus den IV-Kurven (P = 0,018) und den
Rampen (P = 0,023) signifikant. Hier war ein Einfluss des verwendeten Vektors
auf die Eigenschaften des exprimierten Kanals folglich nicht auszuschließen.
Vergleicht man jedoch die Ergebnisse für die Messungen an den Panx1-
Mutanten transfizierten Zellen, so ist kein systematischer, und damit relevanter
Einfluss der Vektoren auf die Kanaleigenschaften der Panx1-Proteine
feststellbar.
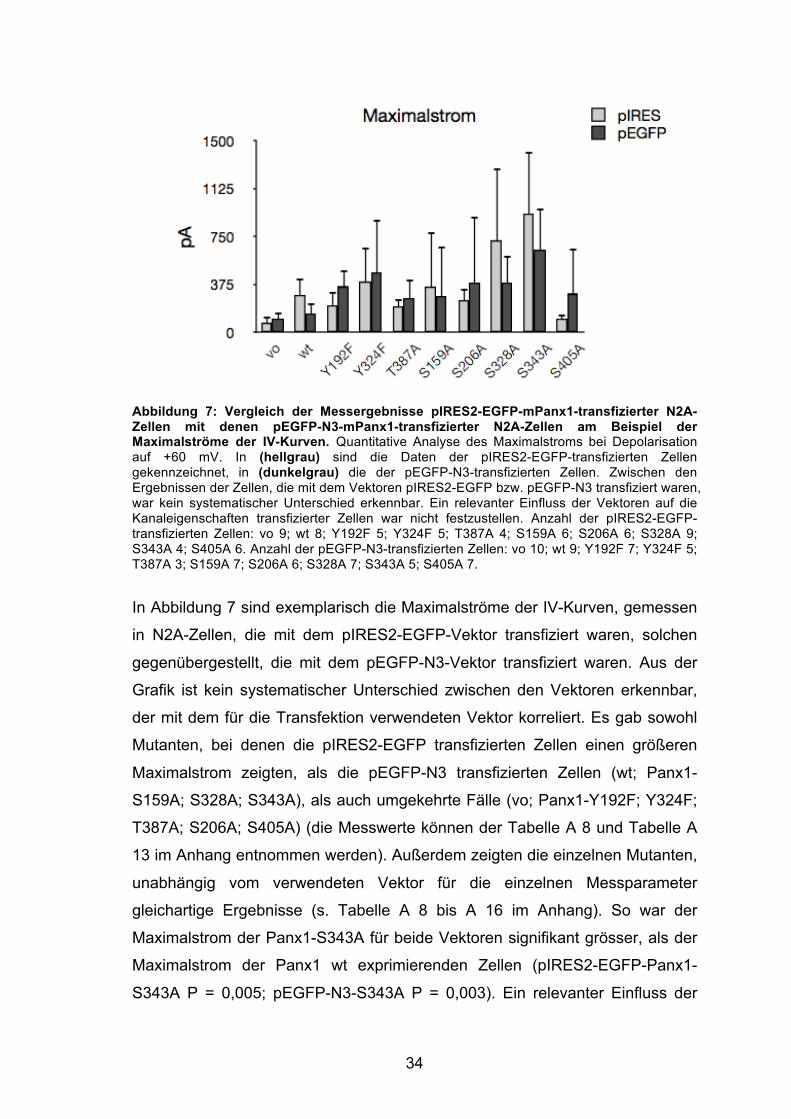
34
Abbildung 7: Vergleich der Messergebnisse pIRES2-EGFP-mPanx1-transfizierter N2A-Zellen mit denen pEGFP-N3-mPanx1-transfizierter N2A-Zellen am Beispiel der Maximalströme der IV-Kurven. Quantitative Analyse des Maximalstroms bei Depolarisation auf +60 mV. In (hellgrau) sind die Daten der pIRES2-EGFP-transfizierten Zellen gekennzeichnet, in (dunkelgrau) die der pEGFP-N3-transfizierten Zellen. Zwischen den Ergebnissen der Zellen, die mit dem Vektoren pIRES2-EGFP bzw. pEGFP-N3 transfiziert waren, war kein systematischer Unterschied erkennbar. Ein relevanter Einfluss der Vektoren auf die Kanaleigenschaften transfizierter Zellen war nicht festzustellen. Anzahl der pIRES2-EGFP-transfizierten Zellen: vo 9; wt 8; Y192F 5; Y324F 5; T387A 4; S159A 6; S206A 6; S328A 9; S343A 4; S405A 6. Anzahl der pEGFP-N3-transfizierten Zellen: vo 10; wt 9; Y192F 7; Y324F 5; T387A 3; S159A 7; S206A 6; S328A 7; S343A 5; S405A 7.
In Abbildung 7 sind exemplarisch die Maximalströme der IV-Kurven, gemessen
in N2A-Zellen, die mit dem pIRES2-EGFP-Vektor transfiziert waren, solchen
gegenübergestellt, die mit dem pEGFP-N3-Vektor transfiziert waren. Aus der
Grafik ist kein systematischer Unterschied zwischen den Vektoren erkennbar,
der mit dem für die Transfektion verwendeten Vektor korreliert. Es gab sowohl
Mutanten, bei denen die pIRES2-EGFP transfizierten Zellen einen größeren
Maximalstrom zeigten, als die pEGFP-N3 transfizierten Zellen (wt; Panx1-
S159A; S328A; S343A), als auch umgekehrte Fälle (vo; Panx1-Y192F; Y324F;
T387A; S206A; S405A) (die Messwerte können der Tabelle A 8 und Tabelle A
13 im Anhang entnommen werden). Außerdem zeigten die einzelnen Mutanten,
unabhängig vom verwendeten Vektor für die einzelnen Messparameter
gleichartige Ergebnisse (s. Tabelle A 8 bis A 16 im Anhang). So war der
Maximalstrom der Panx1-S343A für beide Vektoren signifikant grösser, als der
Maximalstrom der Panx1 wt exprimierenden Zellen (pIRES2-EGFP-Panx1-
S343A P = 0,005; pEGFP-N3-S343A P = 0,003). Ein relevanter Einfluss der
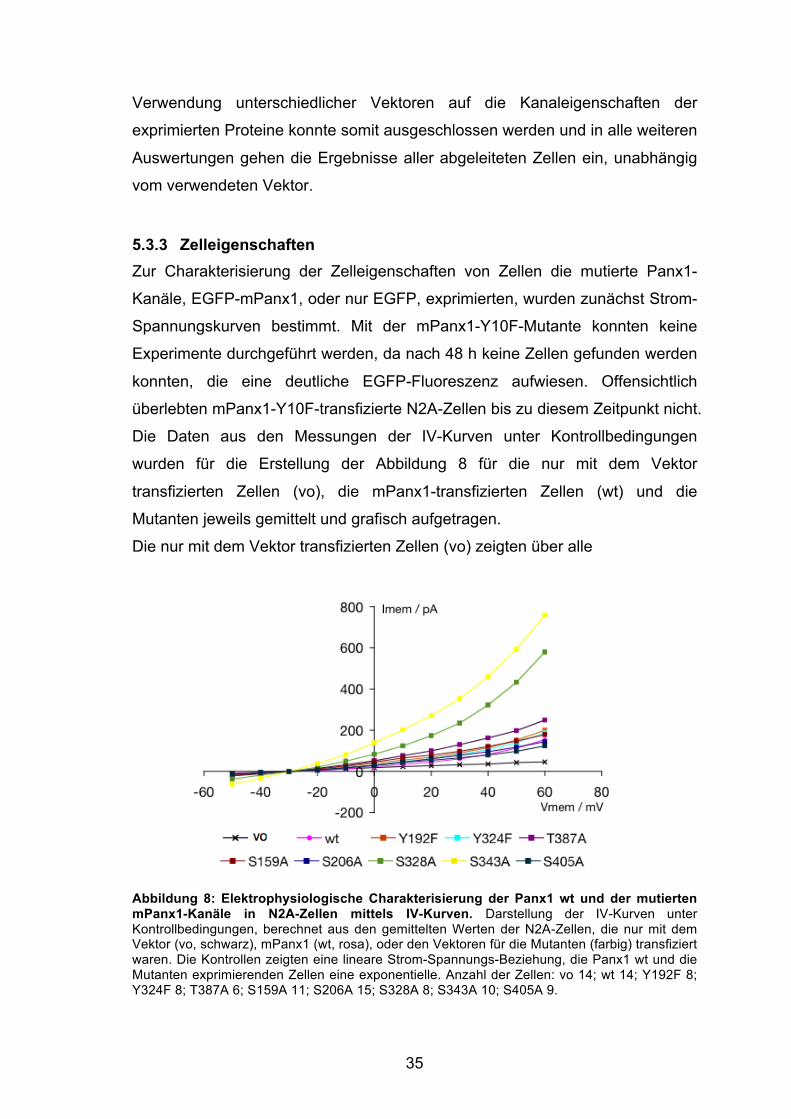
35
Verwendung unterschiedlicher Vektoren auf die Kanaleigenschaften der
exprimierten Proteine konnte somit ausgeschlossen werden und in alle weiteren
Auswertungen gehen die Ergebnisse aller abgeleiteten Zellen ein, unabhängig
vom verwendeten Vektor.
5.3.3 Zelleigenschaften Zur Charakterisierung der Zelleigenschaften von Zellen die mutierte Panx1-
Kanäle, EGFP-mPanx1, oder nur EGFP, exprimierten, wurden zunächst Strom-
Spannungskurven bestimmt. Mit der mPanx1-Y10F-Mutante konnten keine
Experimente durchgeführt werden, da nach 48 h keine Zellen gefunden werden
konnten, die eine deutliche EGFP-Fluoreszenz aufwiesen. Offensichtlich
überlebten mPanx1-Y10F-transfizierte N2A-Zellen bis zu diesem Zeitpunkt nicht.
Die Daten aus den Messungen der IV-Kurven unter Kontrollbedingungen
wurden für die Erstellung der Abbildung 8 für die nur mit dem Vektor
transfizierten Zellen (vo), die mPanx1-transfizierten Zellen (wt) und die
Mutanten jeweils gemittelt und grafisch aufgetragen.
Die nur mit dem Vektor transfizierten Zellen (vo) zeigten über alle
Abbildung 8: Elektrophysiologische Charakterisierung der Panx1 wt und der mutierten mPanx1-Kanäle in N2A-Zellen mittels IV-Kurven. Darstellung der IV-Kurven unter Kontrollbedingungen, berechnet aus den gemittelten Werten der N2A-Zellen, die nur mit dem Vektor (vo, schwarz), mPanx1 (wt, rosa), oder den Vektoren für die Mutanten (farbig) transfiziert waren. Die Kontrollen zeigten eine lineare Strom-Spannungs-Beziehung, die Panx1 wt und die Mutanten exprimierenden Zellen eine exponentielle. Anzahl der Zellen: vo 14; wt 14; Y192F 8; Y324F 8; T387A 6; S159A 11; S206A 15; S328A 8; S343A 10; S405A 9.

36
Spannungssprünge von -50 bis +60 mV einen linearen Stromverlauf (schwarze
Kurve). Sowohl die den Panx1 Wildtyp als auch die die Mutanten
exprimierenden Zellen (farbige Kurven) wiesen dagegen einen exponentiellen
Anstieg der IV-Kontrollkurve bei positiven Potentialen auf.
Die Befunde der nur mit dem Vektor transfizierten Zellen und der Panx1 wt
exprimierenden Zellen entsprechen damit früheren Studien von Bruzzone et al.
(2003), die elektrophysiologische Untersuchungen an Xenopus-Oozyten als
Expressionssytem durchführten. Bunse et al. (2010) konnten diese Strom-
Spannungsbeziehungen auch in N2A-Zellen als Expressionssystem
nachweisen. Die exponentielle Steigung der IV-Kurve der Panx1-transfizierten
Zellen ist auf die spannungsabhängige Öffnung der Panx1-Kanäle bei
Membranpotentialen positiver als 20 mV zurückzuführen. Die hier gezeigten IV-
Kurven können folglich als Beleg dafür dienen, dass alle aufgeführten Mutanten
einen funktionsfähigen Panx1-Kanal ausbildeten.
Auffällig waren zudem die sehr großen Maximalströme der mPanx1-S343A-
und der S328A-Mutanten, bei denen Ströme mit einer Amplitude von bis zu
1800 pA in einzelnen Zellen abgeleitet werden konnten, im Vergleich zu
maximalen Strömen von bis zu 490 pA bei den Panx1 wt exprimierenden Zellen.
Um die elektrophysiologischen Eigenschaften der mutierten und der wt Panx1-
Kanäle besser vergleichen zu können, wurden grundlegende Parameter aus
den IV-Kurven berechnet und quantitativ analysiert. Dazu gehörte die Analyse
des Maximalstroms, gemessen zwischen -30 und +60 mV, wie sie in Abbildung
9 dargestellt ist. Aus der Abbildung 9 ist zu erkennen, dass die maximalen
Stromamplituden der mPanx1-Y324F-Mutante mit 434,77 (± 334,97) pA
signifikant grösser waren (P = 0,022), als die der Panx1 wt exprimierenden
Zellen (213,67 ± 130,31 pA). Noch grösser waren die Maximalströme der
mPanx1-S328A- (573,86 ± 471,83 pA; P = 0,005) und der S343A-Mutanten
(773,01 ± 404,37 pA; P < 0,001 im Rangsummentest). Im Gegensatz dazu
waren die Ströme der nur mit dem Vektor transfizierten Zellen signifikant kleiner
(90,05 ± 53,73 pA; P = 0,001 im Rangsummentest), als die der Panx1 wt
exprimierenden Zellen. Diese Mutanten zeigten also, im Vergleich zum Panx1
Wildtyp eine gesteigerte Leitfähigkeit der Panx1-Kanäle.
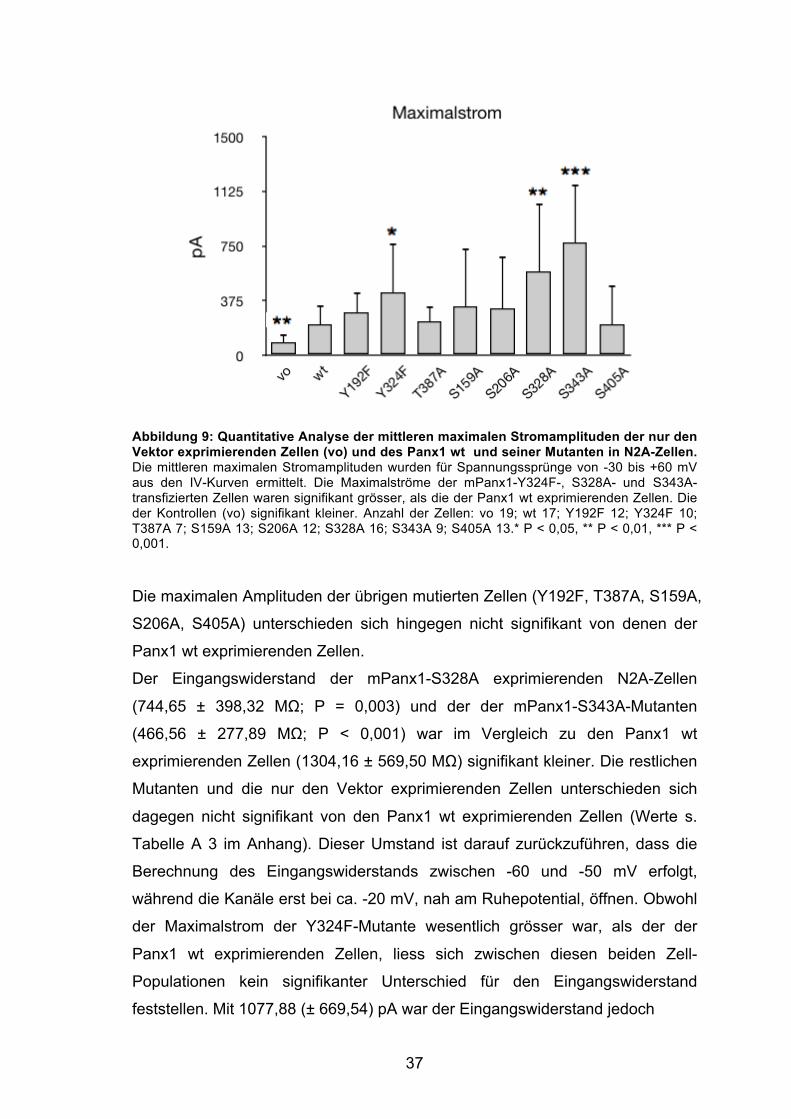
37
Abbildung 9: Quantitative Analyse der mittleren maximalen Stromamplituden der nur den Vektor exprimierenden Zellen (vo) und des Panx1 wt und seiner Mutanten in N2A-Zellen. Die mittleren maximalen Stromamplituden wurden für Spannungssprünge von -30 bis +60 mV aus den IV-Kurven ermittelt. Die Maximalströme der mPanx1-Y324F-, S328A- und S343A-transfizierten Zellen waren signifikant grösser, als die der Panx1 wt exprimierenden Zellen. Die der Kontrollen (vo) signifikant kleiner. Anzahl der Zellen: vo 19; wt 17; Y192F 12; Y324F 10; T387A 7; S159A 13; S206A 12; S328A 16; S343A 9; S405A 13.* P < 0,05, ** P < 0,01, *** P < 0,001.
Die maximalen Amplituden der übrigen mutierten Zellen (Y192F, T387A, S159A,
S206A, S405A) unterschieden sich hingegen nicht signifikant von denen der
Panx1 wt exprimierenden Zellen.
Der Eingangswiderstand der mPanx1-S328A exprimierenden N2A-Zellen
(744,65 ± 398,32 MΩ; P = 0,003) und der der mPanx1-S343A-Mutanten
(466,56 ± 277,89 MΩ; P < 0,001) war im Vergleich zu den Panx1 wt
exprimierenden Zellen (1304,16 ± 569,50 MΩ) signifikant kleiner. Die restlichen
Mutanten und die nur den Vektor exprimierenden Zellen unterschieden sich
dagegen nicht signifikant von den Panx1 wt exprimierenden Zellen (Werte s.
Tabelle A 3 im Anhang). Dieser Umstand ist darauf zurückzuführen, dass die
Berechnung des Eingangswiderstands zwischen -60 und -50 mV erfolgt,
während die Kanäle erst bei ca. -20 mV, nah am Ruhepotential, öffnen. Obwohl
der Maximalstrom der Y324F-Mutante wesentlich grösser war, als der der
Panx1 wt exprimierenden Zellen, liess sich zwischen diesen beiden Zell-
Populationen kein signifikanter Unterschied für den Eingangswiderstand
feststellen. Mit 1077,88 (± 669,54) pA war der Eingangswiderstand jedoch
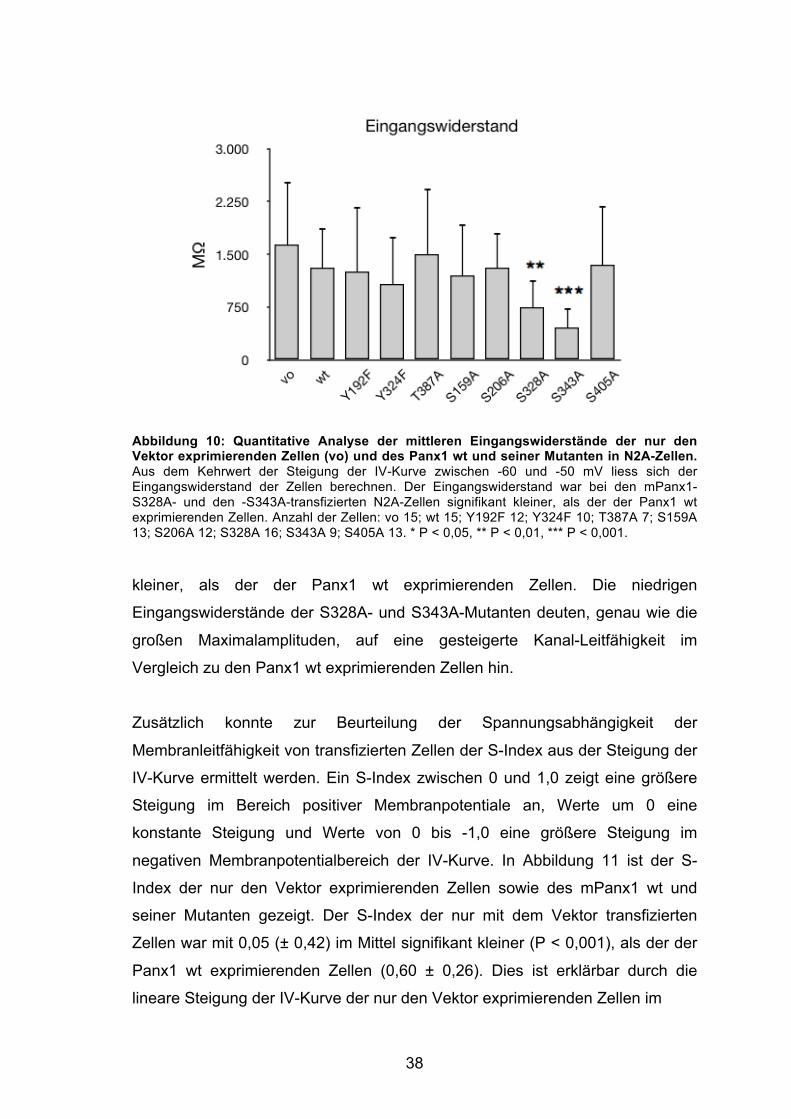
38
Abbildung 10: Quantitative Analyse der mittleren Eingangswiderstände der nur den Vektor exprimierenden Zellen (vo) und des Panx1 wt und seiner Mutanten in N2A-Zellen. Aus dem Kehrwert der Steigung der IV-Kurve zwischen -60 und -50 mV liess sich der Eingangswiderstand der Zellen berechnen. Der Eingangswiderstand war bei den mPanx1-S328A- und den -S343A-transfizierten N2A-Zellen signifikant kleiner, als der der Panx1 wt exprimierenden Zellen. Anzahl der Zellen: vo 15; wt 15; Y192F 12; Y324F 10; T387A 7; S159A 13; S206A 12; S328A 16; S343A 9; S405A 13. * P < 0,05, ** P < 0,01, *** P < 0,001.
kleiner, als der der Panx1 wt exprimierenden Zellen. Die niedrigen
Eingangswiderstände der S328A- und S343A-Mutanten deuten, genau wie die
großen Maximalamplituden, auf eine gesteigerte Kanal-Leitfähigkeit im
Vergleich zu den Panx1 wt exprimierenden Zellen hin.
Zusätzlich konnte zur Beurteilung der Spannungsabhängigkeit der
Membranleitfähigkeit von transfizierten Zellen der S-Index aus der Steigung der
IV-Kurve ermittelt werden. Ein S-Index zwischen 0 und 1,0 zeigt eine größere
Steigung im Bereich positiver Membranpotentiale an, Werte um 0 eine
konstante Steigung und Werte von 0 bis -1,0 eine größere Steigung im
negativen Membranpotentialbereich der IV-Kurve. In Abbildung 11 ist der S-
Index der nur den Vektor exprimierenden Zellen sowie des mPanx1 wt und
seiner Mutanten gezeigt. Der S-Index der nur mit dem Vektor transfizierten
Zellen war mit 0,05 (± 0,42) im Mittel signifikant kleiner (P < 0,001), als der der
Panx1 wt exprimierenden Zellen (0,60 ± 0,26). Dies ist erklärbar durch die
lineare Steigung der IV-Kurve der nur den Vektor exprimierenden Zellen im
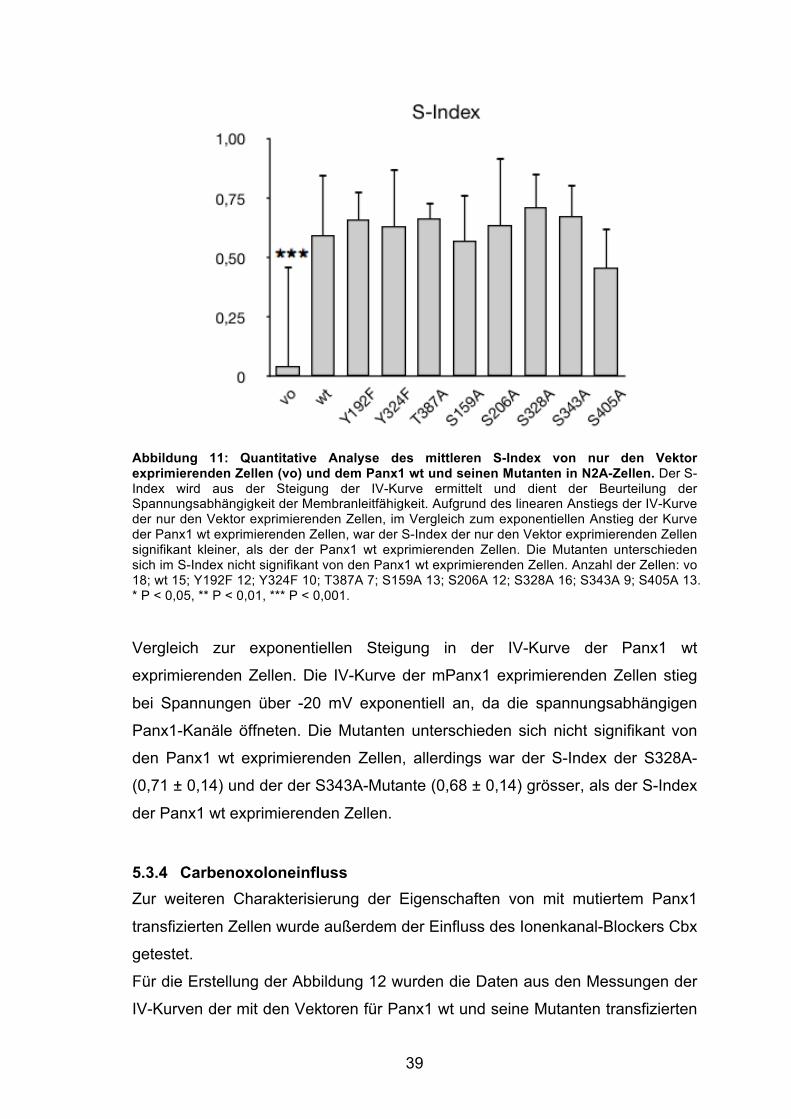
39
Abbildung 11: Quantitative Analyse des mittleren S-Index von nur den Vektor exprimierenden Zellen (vo) und dem Panx1 wt und seinen Mutanten in N2A-Zellen. Der S-Index wird aus der Steigung der IV-Kurve ermittelt und dient der Beurteilung der Spannungsabhängigkeit der Membranleitfähigkeit. Aufgrund des linearen Anstiegs der IV-Kurve der nur den Vektor exprimierenden Zellen, im Vergleich zum exponentiellen Anstieg der Kurve der Panx1 wt exprimierenden Zellen, war der S-Index der nur den Vektor exprimierenden Zellen signifikant kleiner, als der der Panx1 wt exprimierenden Zellen. Die Mutanten unterschieden sich im S-Index nicht signifikant von den Panx1 wt exprimierenden Zellen. Anzahl der Zellen: vo 18; wt 15; Y192F 12; Y324F 10; T387A 7; S159A 13; S206A 12; S328A 16; S343A 9; S405A 13. * P < 0,05, ** P < 0,01, *** P < 0,001.
Vergleich zur exponentiellen Steigung in der IV-Kurve der Panx1 wt
exprimierenden Zellen. Die IV-Kurve der mPanx1 exprimierenden Zellen stieg
bei Spannungen über -20 mV exponentiell an, da die spannungsabhängigen
Panx1-Kanäle öffneten. Die Mutanten unterschieden sich nicht signifikant von
den Panx1 wt exprimierenden Zellen, allerdings war der S-Index der S328A-
(0,71 ± 0,14) und der der S343A-Mutante (0,68 ± 0,14) grösser, als der S-Index
der Panx1 wt exprimierenden Zellen.
5.3.4 Carbenoxoloneinfluss Zur weiteren Charakterisierung der Eigenschaften von mit mutiertem Panx1
transfizierten Zellen wurde außerdem der Einfluss des Ionenkanal-Blockers Cbx
getestet.
Für die Erstellung der Abbildung 12 wurden die Daten aus den Messungen der
IV-Kurven der mit den Vektoren für Panx1 wt und seine Mutanten transfizierten

40
Zellen, sowie nur mit dem Vektor transfizierte N2A-Zellen jeweils gemittelt und
grafisch aufgetragen. Die IV-Kurven wurden zunächst unter
Kontrollbedingungen und dann nach Bad-Applikation von 20 µM Carbenoxolon
aufgenommen. Erwartungsgemäß zeigten die nur mit dem Vektor transfizierten
Zellen über alle Spannungssprünge von -50 bis +60 mV auch hier einen
linearen Stromverlauf. Sie wurden vom Einsatz des Kanalblockers nicht
beeinflusst, die linearen IV-Kurven überlagern sich in der Abbildung 12 A. Die
Panx1 wt exprimierenden Zellen und alle Mutanten (B bis J) wiesen dagegen
einen exponentiellen Anstieg der Kontrollkurve bei positiven Potentialen auf.
Nach Applikation von Cbx konnte bei allen Mutanten und den Panx1 wt
exprimierenden Zellen eine Abnahme der maximalen Amplitude und der
Steigung beobachtet werden.
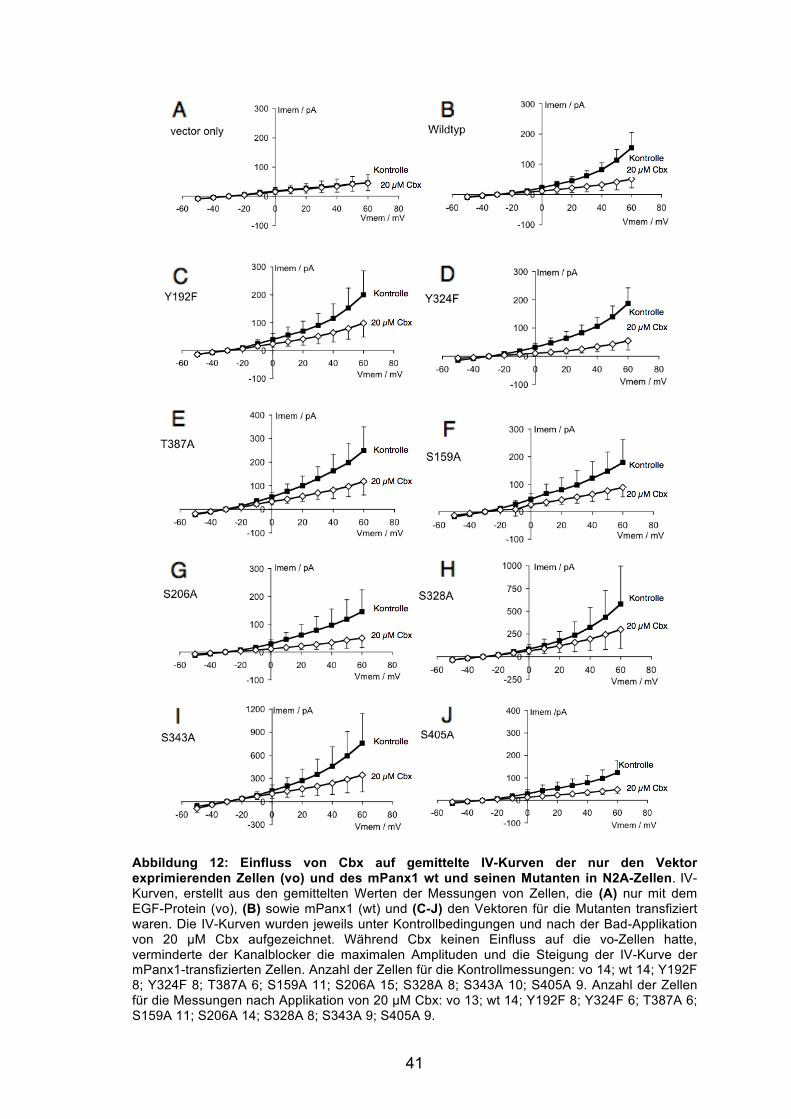
41
Abbildung 12: Einfluss von Cbx auf gemittelte IV-Kurven der nur den Vektor exprimierenden Zellen (vo) und des mPanx1 wt und seinen Mutanten in N2A-Zellen. IV-Kurven, erstellt aus den gemittelten Werten der Messungen von Zellen, die (A) nur mit dem EGF-Protein (vo), (B) sowie mPanx1 (wt) und (C-J) den Vektoren für die Mutanten transfiziert waren. Die IV-Kurven wurden jeweils unter Kontrollbedingungen und nach der Bad-Applikation von 20 µM Cbx aufgezeichnet. Während Cbx keinen Einfluss auf die vo-Zellen hatte, verminderte der Kanalblocker die maximalen Amplituden und die Steigung der IV-Kurve der mPanx1-transfizierten Zellen. Anzahl der Zellen für die Kontrollmessungen: vo 14; wt 14; Y192F 8; Y324F 8; T387A 6; S159A 11; S206A 15; S328A 8; S343A 10; S405A 9. Anzahl der Zellen für die Messungen nach Applikation von 20 µM Cbx: vo 13; wt 14; Y192F 8; Y324F 6; T387A 6; S159A 11; S206A 14; S328A 8; S343A 9; S405A 9.
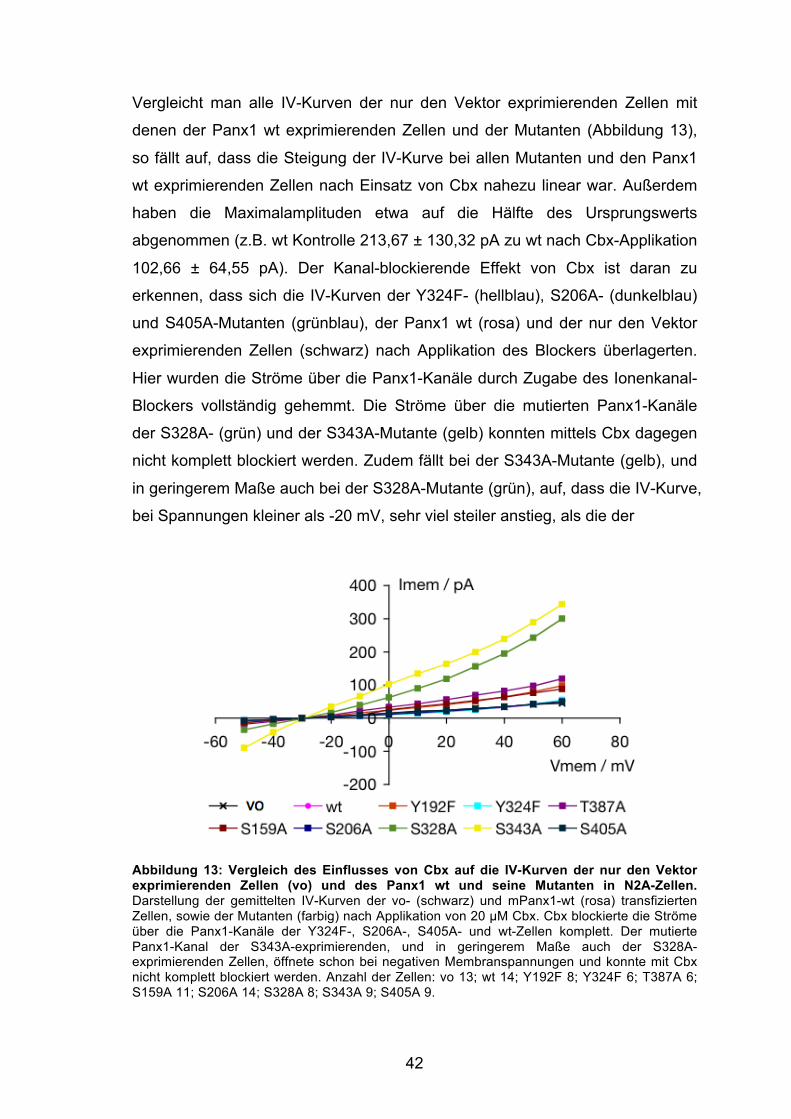
42
Vergleicht man alle IV-Kurven der nur den Vektor exprimierenden Zellen mit
denen der Panx1 wt exprimierenden Zellen und der Mutanten (Abbildung 13),
so fällt auf, dass die Steigung der IV-Kurve bei allen Mutanten und den Panx1
wt exprimierenden Zellen nach Einsatz von Cbx nahezu linear war. Außerdem
haben die Maximalamplituden etwa auf die Hälfte des Ursprungswerts
abgenommen (z.B. wt Kontrolle 213,67 ± 130,32 pA zu wt nach Cbx-Applikation
102,66 ± 64,55 pA). Der Kanal-blockierende Effekt von Cbx ist daran zu
erkennen, dass sich die IV-Kurven der Y324F- (hellblau), S206A- (dunkelblau)
und S405A-Mutanten (grünblau), der Panx1 wt (rosa) und der nur den Vektor
exprimierenden Zellen (schwarz) nach Applikation des Blockers überlagerten.
Hier wurden die Ströme über die Panx1-Kanäle durch Zugabe des Ionenkanal-
Blockers vollständig gehemmt. Die Ströme über die mutierten Panx1-Kanäle
der S328A- (grün) und der S343A-Mutante (gelb) konnten mittels Cbx dagegen
nicht komplett blockiert werden. Zudem fällt bei der S343A-Mutante (gelb), und
in geringerem Maße auch bei der S328A-Mutante (grün), auf, dass die IV-Kurve,
bei Spannungen kleiner als -20 mV, sehr viel steiler anstieg, als die der
Abbildung 13: Vergleich des Einflusses von Cbx auf die IV-Kurven der nur den Vektor exprimierenden Zellen (vo) und des Panx1 wt und seine Mutanten in N2A-Zellen. Darstellung der gemittelten IV-Kurven der vo- (schwarz) und mPanx1-wt (rosa) transfizierten Zellen, sowie der Mutanten (farbig) nach Applikation von 20 µM Cbx. Cbx blockierte die Ströme über die Panx1-Kanäle der Y324F-, S206A-, S405A- und wt-Zellen komplett. Der mutierte Panx1-Kanal der S343A-exprimierenden, und in geringerem Maße auch der S328A-exprimierenden Zellen, öffnete schon bei negativen Membranspannungen und konnte mit Cbx nicht komplett blockiert werden. Anzahl der Zellen: vo 13; wt 14; Y192F 8; Y324F 6; T387A 6; S159A 11; S206A 14; S328A 8; S343A 9; S405A 9.

43
übrigen Zellen. Der mutierte Panx1-Kanal der S343A- und eventuell auch der
der S328A-Zellen war folglich schon bei sehr niedrigen Membranspannungen
geöffnet und durch Cbx in der verwendeten Konzentration nicht blockierbar.
Eine weitergehende Darstellung des Cbx-Effekts auf die Maximalamplituden ist
in Abbildung 14 gezeigt. Die maximalen Stromamplituden, in % des
Ausgangswertes, konnten nach Applikation von 20 µM Cbx aus den IV-Kurven
für Spannungssprünge von -30 auf +60 mV berechnet werden. Der
blockierende Effekt von Carbenoxolon war bei allen Zellen, die mutierte Panx1-
Kanäle exprimierten und den Panx1 wt exprimierenden Zellen zu beobachten:
die mittleren maximalen Amplituden waren nach Cbx-Applikation auf ca. 50%
vermindert (s. Tabelle A 4 im Anhang). Im Gegensatz dazu waren die
Restamplituden der nur den Vektor exprimierenden Zellen nach Zugabe von
Cbx mit 103,45% (± 61,96%) grösser als die der übrigen Zellgruppen. Dieser
Wert ist mit P = 0,001 im Rangsummentest signifikant verschieden von den
Panx1 wt exprimierenden Zellen. Cbx konnte hier nicht wirken, da keine Panx1-
Kanäle vorhanden waren. Zwischen den Zellen, die mit mutierten Panx1
transfizierte waren und den Panx1 wt exprimierenden Zellen konnte hingegen
kein signifikanter Unterschied im Cbx-Effekt festgestellt werden. Die Y324F-
(42,22 ± 13,65%), S328A- (47,57 ± 15,04%) und die S343A-Mutante (45,93 ±
18,89%) wiesen jedoch eine kleinere Restamplitude auf, als die Panx1 wt
exprimierenden Zellen (52,39 ± 14,99%). Bei diesen Mutanten konnte Cbx
einen größeren Anteil des Stroms blockieren, als bei den Panx1 wt
exprimierenden Zellen. Die Einwirkung von Cbx auf die Panx1 wt
exprimierenden Zellen (P < 0,001 im paired t-test) und alle Mutanten (s. Tabelle
A 1) führte zu einer signifikanten Verminderung der Maximalamplituden im
Vergleich zu den Amplituden unter Kontrollbedingungen. Bei den nur mit dem
Vektor transfizierten Zellen veränderten sich die Amplituden dagegen nicht
signifikant nach Cbx-Applikation (P = 0,626 im paired t-test).
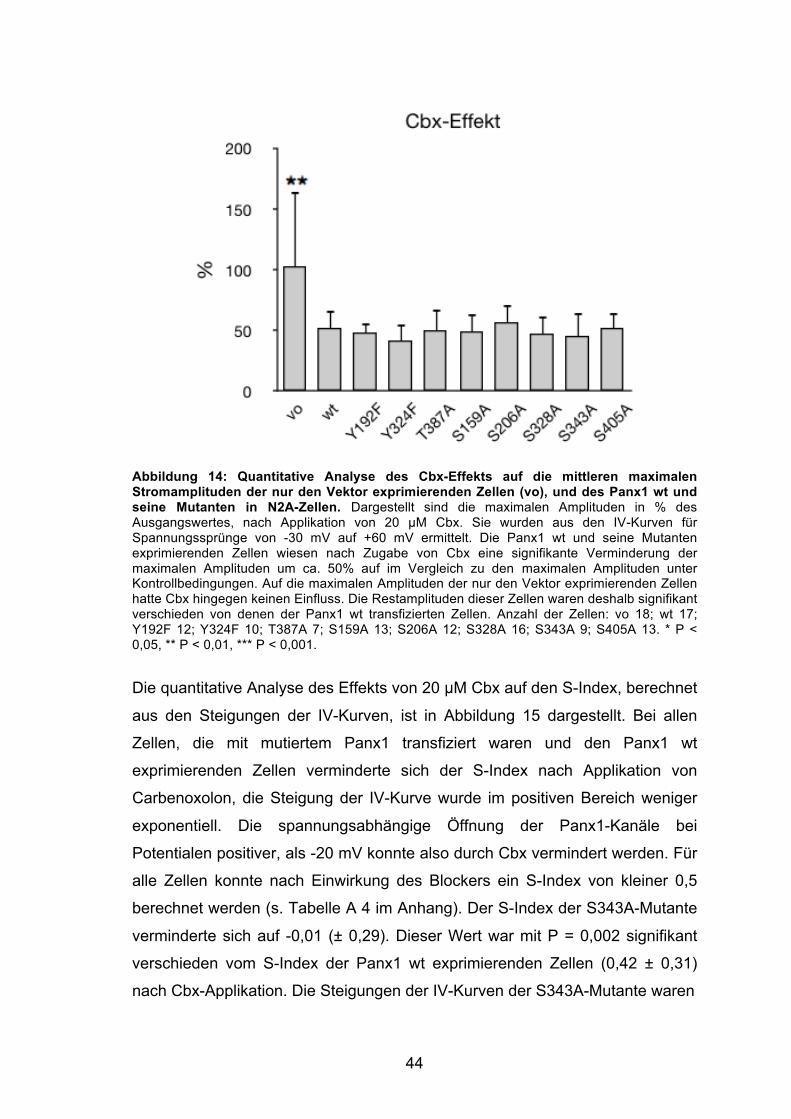
44
Abbildung 14: Quantitative Analyse des Cbx-Effekts auf die mittleren maximalen Stromamplituden der nur den Vektor exprimierenden Zellen (vo), und des Panx1 wt und seine Mutanten in N2A-Zellen. Dargestellt sind die maximalen Amplituden in % des Ausgangswertes, nach Applikation von 20 µM Cbx. Sie wurden aus den IV-Kurven für Spannungssprünge von -30 mV auf +60 mV ermittelt. Die Panx1 wt und seine Mutanten exprimierenden Zellen wiesen nach Zugabe von Cbx eine signifikante Verminderung der maximalen Amplituden um ca. 50% auf im Vergleich zu den maximalen Amplituden unter Kontrollbedingungen. Auf die maximalen Amplituden der nur den Vektor exprimierenden Zellen hatte Cbx hingegen keinen Einfluss. Die Restamplituden dieser Zellen waren deshalb signifikant verschieden von denen der Panx1 wt transfizierten Zellen. Anzahl der Zellen: vo 18; wt 17; Y192F 12; Y324F 10; T387A 7; S159A 13; S206A 12; S328A 16; S343A 9; S405A 13. * P < 0,05, ** P < 0,01, *** P < 0,001.
Die quantitative Analyse des Effekts von 20 µM Cbx auf den S-Index, berechnet
aus den Steigungen der IV-Kurven, ist in Abbildung 15 dargestellt. Bei allen
Zellen, die mit mutiertem Panx1 transfiziert waren und den Panx1 wt
exprimierenden Zellen verminderte sich der S-Index nach Applikation von
Carbenoxolon, die Steigung der IV-Kurve wurde im positiven Bereich weniger
exponentiell. Die spannungsabhängige Öffnung der Panx1-Kanäle bei
Potentialen positiver, als -20 mV konnte also durch Cbx vermindert werden. Für
alle Zellen konnte nach Einwirkung des Blockers ein S-Index von kleiner 0,5
berechnet werden (s. Tabelle A 4 im Anhang). Der S-Index der S343A-Mutante
verminderte sich auf -0,01 (± 0,29). Dieser Wert war mit P = 0,002 signifikant
verschieden vom S-Index der Panx1 wt exprimierenden Zellen (0,42 ± 0,31)
nach Cbx-Applikation. Die Steigungen der IV-Kurven der S343A-Mutante waren
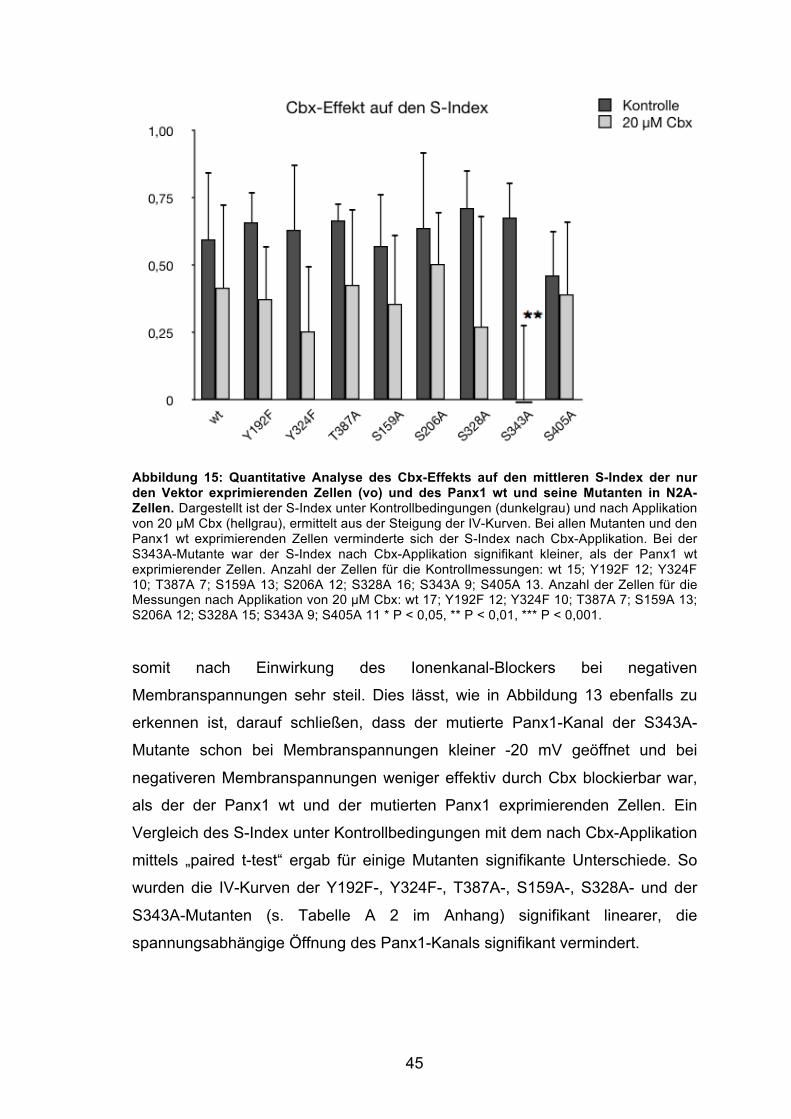
45
Abbildung 15: Quantitative Analyse des Cbx-Effekts auf den mittleren S-Index der nur den Vektor exprimierenden Zellen (vo) und des Panx1 wt und seine Mutanten in N2A-Zellen. Dargestellt ist der S-Index unter Kontrollbedingungen (dunkelgrau) und nach Applikation von 20 µM Cbx (hellgrau), ermittelt aus der Steigung der IV-Kurven. Bei allen Mutanten und den Panx1 wt exprimierenden Zellen verminderte sich der S-Index nach Cbx-Applikation. Bei der S343A-Mutante war der S-Index nach Cbx-Applikation signifikant kleiner, als der Panx1 wt exprimierender Zellen. Anzahl der Zellen für die Kontrollmessungen: wt 15; Y192F 12; Y324F 10; T387A 7; S159A 13; S206A 12; S328A 16; S343A 9; S405A 13. Anzahl der Zellen für die Messungen nach Applikation von 20 µM Cbx: wt 17; Y192F 12; Y324F 10; T387A 7; S159A 13; S206A 12; S328A 15; S343A 9; S405A 11 * P < 0,05, ** P < 0,01, *** P < 0,001.
somit nach Einwirkung des Ionenkanal-Blockers bei negativen
Membranspannungen sehr steil. Dies lässt, wie in Abbildung 13 ebenfalls zu
erkennen ist, darauf schließen, dass der mutierte Panx1-Kanal der S343A-
Mutante schon bei Membranspannungen kleiner -20 mV geöffnet und bei
negativeren Membranspannungen weniger effektiv durch Cbx blockierbar war,
als der der Panx1 wt und der mutierten Panx1 exprimierenden Zellen. Ein
Vergleich des S-Index unter Kontrollbedingungen mit dem nach Cbx-Applikation
mittels „paired t-test“ ergab für einige Mutanten signifikante Unterschiede. So
wurden die IV-Kurven der Y192F-, Y324F-, T387A-, S159A-, S328A- und der
S343A-Mutanten (s. Tabelle A 2 im Anhang) signifikant linearer, die
spannungsabhängige Öffnung des Panx1-Kanals signifikant vermindert.

46
6 Diskussion
Pannexin1-Kanäle konnten bisher mit verschiedenen pathologischen
Vorgängen beim Menschen in Verbindung gebracht werden. So kann
Neurodegeneration aufgrund von Entzündungen im post-ischämischen Gehirn,
bei Diabetes, oder M. Alzheimer auf eine Öffnung neuronaler Pannexin1-
Kanäle und den damit einhergehenden neuronalen Zelltod zurückgeführt
werden (Orellana et al., 2011). Auch für die Ausbildung bleomyzininduzierter
Lungenfibrose ist eine entzündungsbedingte Aktivierung von Pannexin1-
Kanälen als ursächlich anzunehmen (Riteau et al., 2010).
Ein besseres Verständnis der Regulation von Pannexinen kann deshalb einen
wichtigen Beitrag zur Erkennung und Therapie solcher Erkrankungen leisten.
Von Connexinen ist bereits bekannt, dass die posttranslationale Modifikation
durch Phosphorylierung, die ein wichtiges Mittel zur Regulation biologischer
Prozesse in Zellen darstellt verschiedene funktionelle Eigenschaften der
Connexin-Kanäle beeinflussen kann (Solan und Lampe, 2009). Da Pannexine
ebenfalls Phosphoproteine sind und Pannexine und Connexine die gleiche
Sekundärstruktur aufweisen, war davon auszugehen, dass auch Pannexine
phosphoryliert werden (Barbe et al., 2006). In vorliegender Arbeit wurden
deshalb mit Hilfe gezielter Mutagenese neun potentielle
Phosphorylierungsstellen von Pannexin1 mutiert und die daraus entstandenen
Panx1-Mutanten weitergehend untersucht.
Für N2A-Zellen, die mit den Expressionsvektoren für mPanx1 oder eine der
erzeugten Mutanten transfiziert waren, konnte mittels Western Blot und
konfokaler Fluoreszenzmikroskopie nachgewiesen werden, dass die meisten
Zellen potenziell überlebensfähig waren und Pannexin-Kanäle ausbildeten. Um
mögliche funktionelle Veränderungen zu charakterisieren, schloss sich eine
Untersuchung elektrophysiologischer Eigenschaften transfizierter Zellen an.

47
6.1 Elektrophysiologische Analyse der Zelleigenschaften transfizierter N2A-Zellen
Zur Untersuchung elektrophysiologischer Eigenschaften transfizierter N2A-
Zellen wurden Ganzzellableitungen mit zwei verschiedenen
Stimulationsprotokollen durchgeführt. Einerseits 500 ms andauernde
Spannungspulse, mit denen das Membranpotential der Zellen schrittweise von
-50 mV auf +60 mV eingestellt wurde, und andererseits Stromrampen, bei
denen das Haltepotential kontinuierlich von -100 mV auf +80 mV geändert
wurde. Es stellte sich die Frage, welches Stimulationsprotokoll besser zur
Analyse der Zelleigenschaften geeignet ist. Bei Verwendung der
Spannungspulse können besser der maximal über die Zelle fließende Strom
und die Spannungsabhängigkeit der Leitfähigkeit bestimmt werden.
Stromrampen erlauben hingegen auch die Ermittlung der Öffnungskinetik von
Ionenkanälen, abgesehen von prinzipiellen Limitierungen der Ganzzell-Methode.
Für die Ergebnisse aus den elektrophysiologischen Experimenten war kein
relevanter Unterschied zwischen den beiden Stimulationsprotokollen
feststellbar. Die Zellen, die mit den mutierten Panx1-Kanälen transfiziert waren,
zeigten für beide Untersuchungsmethoden gleiche Zelleigenschaften und
signifikante Unterschiede zwischen den Mutanten und den Panx1 wt
exprimierenden Zellen waren für beide Protokolle gleich häufig. Bei
Verwendung der Spannungspulse wurde das Membranpotential der
abgeleiteten Zelle jedoch über längere Zeit konstant gehalten, so dass sich ein
„steady-state“-Zustand einstellen und die Zellströme so zuverlässiger bestimmt
werden konnten. Dagegen kann die Spannungsabhängigkeit von Ionenkanälen
mittels Stromrampen nur unzureichend und auch nur qualitativ untersucht
werden, da verzögert reagierende Kanäle bei schnellen Änderungen des
Membranpotentials möglicherweise nicht erfasst werden. Auf die Darstellung
der Ergebnisse aus den Messungen der Stromrampen wurde in dieser Arbeit
deshalb zugunsten der Analyse der Daten aus den Spannungspulsen verzichtet.
Für die Beschreibung der elektrophysiologischen Eigenschaften der
transfizierten N2A-Zellen wurden daher aus den Stromantworten auf die
Spannungspulse Strom-Spannungsbeziehungen (IV-Kurven) bestimmt.
Eine zuverlässige und quantitative Analyse der Öffnungskinetik von einzelnen
Ionenkanälen erfolgt außerdem besser mit Hilfe von Einzelkanalmessungen,

48
anhand derer auch Öffnungszustände, Öffnungswahrscheinlichkeiten und
Inaktivierungszeiten von Kanälen bestimmt werden können.
Einzelkanalmessungen waren für die überblickartige Untersuchung mutierter
potentieller Phosphorylierungsstellen, wie sie in vorliegender Arbeit erfolgen
sollte, jedoch nicht geeignet. Zudem muss für eine Einzelkanalmessung ein
Stück der Membran aus der Zellwand herausgerissen werden und alle
Komponenten, die für die Phosphorylierung benötigt werden, würden so
verloren gehen. Es wäre also nicht feststellbar, ob die erzeugten Mutationen zu
veränderter Phosphorylierung führten.
6.2 Die Verwendung der Vektoren pEGFP-N3-m-Panx1 und pIRES2-EGFP-mPanx1 hat keinen Einfluss auf die Eigenschaften von mPanx1
Zur Transfektion von N2A-Zellen mit mutierten Pannexin1-Kanälen wurden zwei
verschiedene Expressionsvektoren eingesetzt. Zum einen der pEGFP-N3-
mPanx1-Vektor, der für Fusionsproteine aus mPanx1 und C-terminal
lokalisiertem EGFP kodierte, und zum andern der pIRES2-EGFP-mPanx1-
Vektor, der eine bicistronische Expression von mPanx1 und EGFP ermöglichte.
Aus früheren Arbeiten ist bereits bekannt, dass der C-Terminus eine wichtige
Rolle bei der Regulation von Kanal-Eigenschaften spielt. Untersuchungen an
Cx43 ergaben beispielsweise, dass eine Verkürzung des C-Terminus zu einer
intrazellulären Retention des Proteins und folglich verminderter Gap junction-
Bildung im Herzen führen und damit Vorhofflimmern auslösen kann (Thibodeau
et al., 2010). Auch von Pannexinen weiß man, dass eine Verkürzung des C-
Terminus veränderte Kanaleigenschaften verursachen kann. So bildet eine
Spaltung des Pannexin1-C-Terminus an einer spezifischen Caspase-
Schnittstelle über die Ausbildung eines permanent offenen Kanals einen
Zwischenschritt der Apoptose (Chekeni et al., 2010).
Um die Messergebnisse richtig interpretieren zu können, musste ein Einfluss
des EGFP am C-Terminus auf den Pannexin1-Kanal in Versuchen mit dem
pEGFP-N3-mPanx1-Vektor ausgeschlossen werden. Bisher wurde allerdings
von keiner Arbeitsgruppe ein Einfluss eines C-terminalen Markers, oder einer
Verlängerung des C-Terminus auf die Eigenschaften des Panx1-Kanals

49
beobachtet (Chekeni et al., 2010; unveröffentlichtes Material unserer
Arbeitsgruppe). Lediglich eine Verkürzung des Carboxy-Terminus zwischen den
Aminosäurepositionen 370-380 (unveröffentlichtes Material unserer
Arbeitsgruppe; Chekeni et al., 2010), bzw. Mutationen von Aminosäuren im C-
Terminus (Bunse et al., 2010; Wang und Dahl, 2010) führten zu veränderten
Zelleigenschaften. Deshalb sollte die Verwendung des pEGFP-N3-mPanx1-
Vektors keinen Einfluss auf die Zelleigenschaften haben.
Ein Vergleich der elektrophysiologischen Messungen von pEGFP-N3-mPanx1-
transfizierten Zellen mit denen von pIRES2-EGFP-mPanx1-transfizierten Zellen
ergab zudem keinen systematischen Unterschied, der mit dem für die
Transfektion verwendeten Vektor korreliert hätte. So waren z.B. die maximalen
Amplituden der IV-Kurven von Zellen die carboxyterminale Mutanten
exprimierten, nicht immer dann grösser, wenn der pEGFP-N3-mPanx1-Vektor
verwendet wurde. Bei den Y324F-, T387A- und S405A-Mutanten war zwar der
Maximalstrom der pEGFP-N3-mPanx1-transfizierten Zellen grösser, als der der
pIRES2-EGFP-mPanx1-transfizierten Zellen, bei den S328A- und S343A-
Mutanten war es jedoch umgekehrt. Zellen, die mit den Vektoren für mutierte
Panx1-Kanäle transfiziert waren, zeigten außerdem, unabhängig vom
verwendeten Vektor, gleichartige Tendenzen für die verschiedenen
Messparameter.
Darüber hinaus zeigte auch die Expressionsanalyse mittels Western Blot
keinen Unterschied zwischen den eingesetzten Vektoren. Bei allen Panx1
transfizierten Zellen, unabhängig vom eingesetzten Expressionsvektor, konnte
eine Membranständigkeit des Pannexin1-Kanals nachgewiesen werden.
Somit konnte umgekehrt auch ein Einfluss des separat vom Panx1-Kanal
translatierten EGFP-Proteins auf die Zelleigenschaften pIRES2-EGFP-mPanx1-
transfizierter Zellen ausgeschlossen werden.
6.3 Phosphorylierung spielt eine wichtige Rolle in der Regulation von Panx1
Die meisten Connexine sind Phosphoproteine und die Phosphorylierung von
Connexinen kann mit verschiedenen Mechanismen zur Regulation zellulärer
Kommunikation in Verbindung gebracht werden (Lampe und Lau, 2000). So

50
spielt Phosphorylierung eine wichtige Rolle in allen Abschnitten des Connexin-
Lebenszyklusses, wie der Biosynthese, dem Transport zur und den Einbau des
Proteins in die Membran, der Ausbildung von Gap junctions, dem Öffnen und
Schließen des Hemikanals und der Gap junctions, sowie dem Abbau des
Kanals (s. Review Sáez et al., 2003; Sáez et al., 2005; Solan und Lampe, 2009).
Pannexine sind gleichfalls Phosphoproteine und auch hier könnte der
Phosphorylierung eine wesentliche Bedeutung in der Regulation ihres
Lebenszyklusses zukommen (Barbe et al., 2006).
6.3.1 Alle Mutanten bilden funktionsfähige Pannexin1-Kanäle aus Die Überlebensfähigkeit der neun, an potentiellen Phosphorylierungsstellen des
Pannexin1-Kanals mittels gezielter Mutagenese erzeugten Mutanten und die
Lokalisation des Kanals in der Zellmembran transfizierter N2A-Zellen konnten
mittels Western Blot und konfokalen Aufnahmen nachgewiesen werden. Alle
Zellen, die mutierte Panx1-Kanäle exprimierten zeigten außerdem in den
Strom-Spannungs-Kurven ein Antwortverhalten ähnlich dem der Panx1 wt
exprimierenden Zellen, da sie alle bei positiven Membranpotentialen einen
exponentiellen Anstieg der IV-Kurve aufwiesen. Im Unterschied dazu zeigten
Zellen, die mit dem leeren Vektor transfiziert waren, hier einen linearen
Stromverlauf. Keine der neun Mutationen führte also zu einer Störung der
Ausbildung eines funktionsfähigen Pannexin1-Kanals. Die Biosynthese der hier
untersuchten Pannexin1-Kanäle und ihr Transport zur Zellmembran sind
folglich nicht eingeschränkt. Dieses Ergebnis ist überraschend, da von
Connexinen bekannt ist, dass Phosphorylierung ein wichtiger Mechanismus zur
Regulation des Transports zur Membran ist (Musil und Goodenough, 1991;
Martin et al., 2001). So konnte für Cx43 das Serin an Position 373 als wichtige
Lokalisation posttranslationaler Phosphorylierung zur Steuerung des
Proteintransports zur Membran identifiziert werden (Park et al., 2007). Auch die
Expression von Connexinen wird mittels Phosphorylierung kontrolliert (Salameh
et al., 2009). Es ist also erstaunlich, dass die in dieser Arbeit untersuchten
Phosphorylierungsstellen von Pannexin1 keine Aufgabe in Biosynthese und
Transport zu haben scheinen. Allerdings konnten im Rahmen dieser Arbeit
nicht alle potentiellen Phosphorylierungsstellen in Pannexin1 untersucht werden,
da die Mutagenese des Serin 394, das laut Netphos2.0 Server ebenfalls einen

51
Vorhersagewert von grösser 0,5 hatte, nicht erfolgreich war. Eine
posttranslationale Modifikation dieser Aminosäure könnte also zur Regulation
von Biosynthese und Transport von Bedeutung sein. Die Untersuchung der
Eigenschaften einer mPanx1-S394A-Mutante sollte deshalb in weiteren
Experimenten noch folgen. Außerdem ist es möglich, dass zur Regulation
dieser Prozesse die Phosphorylierung einer einzelnen Aminosäure nicht
ausreichend ist, sondern mehrere Aminosäuren gleichzeitig phosphoryliert
werden müssen. Hier könnten Studien mit Pannexin1-Kanälen, die an
mehreren potentiellen Phosphorylierungsstellen gleichzeitig mutiert sind,
weitere Erkenntnisse bringen.
6.3.2 Die N-terminale Mutante Y10F führt zu strukturellen Veränderungen des Panx1-Kanals
Innerhalb des N-Terminus des Pannexin1-Proteins gab es lediglich eine
Aminosäure, die als potentielle Phosphorylierungsstelle vorhergesagt wurde,
das Tyrosin an Position Y10. Es wurde mit Hilfe gezielter Mutagenese zu
Phenylalanin mutiert. Die Western Blot-Analyse dieser Mutante zeigte nur sehr
schwache Banden, so dass man annehmen könnte, dass hier der Kanal nicht in
die Zellmembran, oder das endoplasmatische Retikulum, eingebaut wurde. Wie
die Analyse mit Hilfe des Konfokalmikroskops zeigte, wiesen die Zellen
außerdem eine stark verkürzte Überlebenszeit von weniger als 96 h auf. Auch
in einer durchflusszytometrischen Untersuchung dieser Mutante war der Anteil
toter Zellen 24 h nach Transfektion grösser als der lebender Zellen
(unveröffentlichte Ergebnisse unserer Arbeitsgruppe). Aus diesem Grund waren
elektrophysiologische Messungen an mPanx1-Y10F transfizierten N2A-Zellen
nicht möglich. Bei einer Wiederholung der Experimente mit transfizierten
Xenopus-Oozyten (S. Bunse, pers. Mitteilung) zeigten sich die Oozyten jedoch
als überlebensfähiger als die N2A-Zellen und elektrophysiologische Messungen
waren möglich. Letale Mutanten führen in N2A-Zellen möglicherweise prinzipiell
früher zum Zelltod als in Oozyten da die N2A-Zelllinie weniger robust ist. Die
schwachen Banden in der Western Blot-Analyse sind also vermutlich darauf
zurückzuführen, dass stark exprimierende N2A-Zellen früh abstarben und für
die Blot-Analyse Protein aus Zellen ohne oder mit nur geringer Panx1-
Expression eingesetzt wurde. Um den Einbau des mutierten Kanals in die

52
Zellmembran von mPanx1-Y10F transfizierten Zellen endgültig nachweisen zu
können, müssten allerdings weitere immunzytochemische Untersuchungen von
transfizierten N2A-Zellen oder injizierten Oozyten folgen.
Auch in Versuchen mit injizierten Oozyten war die Überlebenszeit der Y10F-
Mutanten mit ca. 40 h gegenüber mehr als 100 h bei den Wildtyp-Zellen stark
herabgesetzt. Durch den Einsatz von Cbx konnte die Überlebenszeit jedoch auf
über 80 h verdoppelt werden (S. Bunse, pers. Mitteilung). Dieses Ergebnis lässt
auf eine gesteigerte Kanal-Aktivität der Y10F-Mutante schließen. Der
Zusammenbruch transmembranärer Ionengradienten, oder der Anstieg
intrazellulären Kalziums, sowie der Verlust wichtiger Metabolite durch eine
dauernde Öffnung des Kanals bei Ruhemembranpotentialen war vermutlich die
Ursache für den schnellen Zelltod transfizierter Zellen. Die gesteigerte
Leitfähigkeit des Panx1-Y10F-Kanals war auch in den elektrophysiologischen
Untersuchungen an injizierten Oozyten nachweisbar. Denn Maximalstrom, Cbx-
Effekt und Ruhemembranpotential waren bei der Y10F-Mutante signifikant
grösser, der Haltestrom jedoch signifikant niedriger, als bei den Panx1 wt
exprimierenden Oozyten (S. Bunse, pers. Mitteilung). Um die gesteigerte
Leitfähigkeit weitergehend zu untersuchen, könnten Farbstoffaufnahme-
Versuche mit Lucifer Yellow (LY) durchgeführt werden. Oozyten, die wt Panx1-
Kanäle exprimieren, nehmen spontan kein LY auf, da Panx1-Kanäle am
Ruhepotential geschlossen sind. Würden die Y10-F-Mutanten eine LY-
Aufnahme induzieren und wäre die Farbstoffaufnahme außerdem durch Cbx
blockierbar, so würde dies ebenfalls für das Vorhandensein dauernd geöffneter
Pannexin-Kanäle sprechen.
Es gibt verschiedene Erklärungsmöglichkeiten für eine gesteigerte Leitfähigkeit
der Y10F-Mutante. So könnte durch veränderte Phosphorylierung die
Offenwahrscheinlichkeit der Panx1-Kanäle, oder ihre mittlere Öffnungsdauer
vergrößert sein. Wahrscheinlicher ist jedoch ein Einfluss der N-terminalen
Mutation auf die Struktur des Kanals und eine daraus resultierende
Kanalöffnung am Ruhemembranpotential (z.B. durch Verlust der
Spannungsabhängigkeit für die der N-Terminus eine wichtige Rolle spielt).
Aufgrund der Ergebnisse einer SCAM-Analyse der Panx1-Struktur durch Wang
und Dahl (2010) kann davon ausgegangen werden, dass in Pannexin1 der N-
Terminus strukturelle Aufgaben erfüllt und nicht an der Bildung der Kanalpore

53
beteiligt ist. Dies steht im Gegensatz zu den Erkenntnissen aus
Untersuchungen an Connexinen. Für Cx26 konnte aufgrund von Analysen der
Kristallstruktur eine Beteiligung der N-terminalen Aminosäuren an der
Formation des cytoplasmatischen Anteils der Pore ermittelt werden (Maeda et
al., 2009).
Wenn in Pannexin1 also der N-Terminus wichtig für die korrekte Faltung des
Proteins und den Transfer, bzw. die Faltung im endoplasmatischen Retikulum,
den Transport zur, sowie die Funktion in der Plasmamembran hat (Wang und
Dahl, 2010), so hat auch die in dieser Arbeit untersuchte N-terminale Y10F-
Mutante einen Einfluss auf die Struktur des Pannexin1-Kanals. Und die
veränderte Struktur führt, unabhängig von Phosphorylierungseffekten, zur
Ausbildung eines Kanals mit größerer Leitfähigkeit oder größerer
Offenwahrscheinlichkeit. Dies könnte in Einzelkanalmessungen weitergehend
untersucht werden.
6.3.3 Mutationen putativer Phosphorylierungsstellen in der intrazellulären Schleife von Panx1 haben keinen Einfluss auf die Eigenschaften des Kanals im Vergleich zu Panx1 wt
Die Serine S159 und S206, sowie das Tyrosin Y192 sind potentielle
Phosphorylierungsstellen, die in der intrazellulären Schleife lokalisiert sind. Die
beiden Serine wurden zu Alanin und das Tyrosin zu Phenylalanin mutiert. In der
Western Blot-Analyse transfizierter N2A-Zellen konnten für diese drei Mutanten
die Pannexin1-typischen Mehrfachbanden („multiple band pattern“)
nachgewiesen werden, so dass man davon ausgehen kann, dass hier
membranständige Kanäle gebildet wurden. Zwar konnten für die pIRES2-
EGFP-mPanx1-S159A transfizierten N2A-Zellen nur sehr schwache Banden
festgestellt werden, jedoch ist hier die als Kontrolle dienende β-Aktin-Bande
ebenfalls nur sehr blass, so das anzunehmen ist, dass insgesamt nur wenig
Protein aufgebracht wurde. Konfokalmikroskopisch konnte die Lokalisation des
Panx1-Proteins an der Zelloberfläche dieser mutierten Zellen bestätigt werden
und ihre Überlebenszeit entsprach der der Panx1 wt exprimierenden Zellen.
Auch in den elektrophysiologischen Untersuchungen und quantitativen
Analysen unterschieden sich diese Mutanten nicht von den Panx1 wt
exprimierenden Zellen. Die Phosphorylierung von Aminosäuren der

54
intrazellulären Schleife kann folglich nicht zur Regulation von
Kanaleigenschaften, wie Leitfähigkeit, Spannungsabhängigkeit, oder die
Blockierbarkeit durch Cbx von Bedeutung zu sein. Ein Einfluss auf andere
Eigenschaften, wie die Kalzium-, oder pH-Abhängigkeit des Kanals ist im
Rahmen dieser Arbeit jedoch nicht untersucht worden und damit nicht
ausgeschlossen. Hier sollten weitere Experimente folgen, um die Funktion
intrazellulärer potentieller Phosphorylierungsstellen besser beurteilen zu
können.
Von Connexinen weiß man, dass Phosphorylierungstellen in der intrazellulären
Schleife für die Regulation von Bedeutung sind (Solan und Lampe 2009). So
konnte für die Connexine35 und 36 festgestellt werden, dass Phosphorylierung
am Serin 110 in der intrazellulären Schleife zu verminderter elektrischer
Kopplung über Gap junctions führt (Urschel et al., 2006; Kothman et al., 2007).
Außerdem wurde beschrieben, dass die Phosphorylierung zweier Serine einen
größeren Effekt hat, als die alleinige Modifikation einer Stelle. Es ist deshalb
anzunehmen, dass auch bei Pannexinen ein Phosphorylierungseffekt eventuell
erst durch Veränderung mehrerer Positionen beobachtet werden kann.
In bisher unveröffentlichten Versuchen unserer Arbeitsgruppe mit injizierten
Xenopus-Oozyten konnte außerdem festgestellt werden, dass die S206A-
Mutante eine signifikant größere Leitfähigkeit hatte, als die Panx1 wt
exprimierenden Zellen (S. Bunse, pers. Mitteilung). Es ist also möglich, dass die
Phosphorylierungsstellen der intrazellulären Schleife doch eine wichtige
Funktion in der Regulation der in dieser Arbeit untersuchten Zelleigenschaften
haben, und lediglich mit der verwendeten N2A-Zelllinie nicht detektiert werden
konnte. N2A-Zellen sind Neuroblastomzellen und die Expression neuronaler
Komponenten in diesen Zellen ist gegeben. Sie könnten im Vergleich zu den
Xenopus-Oozyten andere, z.B. spannungsabängige Ionenkanäle exprimieren,
die dazu führen, dass Pannexin1-vermittelte Ströme zumindest teilweise
maskiert werden.
Es gibt verschiedene Erklärungsmöglichkeiten für die gesteigerte Leitfähigkeit
mutierter N2A-Zellen. So könnten, durch einen erhöhten Transport von Kanälen
zur Membran oder einen verminderten Abbau, insgesamt mehr Kanäle in der
Membran vorhanden sein, so dass pro Zeiteinheit mehr Kanäle gleichzeitig
geöffnet wären. Auch eine gesteigerte Offenwahrscheinlichkeit, oder eine

55
längere mittlere Öffnungsdauer, sowie eine höhere Einzelkanalleitfähigkeit
könnten zu einer gesteigerten Leitfähigkeit beitragen. Für Connexine sind all
diese Mechanismen als Folge von Phosphorylierung bekannt (Moreno et al.,
1992; van Veen et al., 2000; Cottrell et al., 2003; Review Račkauskas et al.,
2010). Darüber hinaus konnte erhöhte Hemikanal-Leitfähigkeit als Folge von
Mutationen einzelner potentieller Phosphorylierungsstellen in Connexinen mit
seltenen Krankheiten in Verbindung gebracht werden. So wird die Okulo-dento-
digitale Dysplasie auf eine erhöhte Cx43-Hemikanal-Leitfähigkeit zurückgeführt.
Die mutierten Aminosäurepositionen G138R und G143S führen bei dieser
Krankheit zu einem veränderten Phosphorylierungszustand des Proteins,
dadurch kommt es zu einem verminderten Abbau von Cx43 und in Folge
dessen zu einer erhöhten Zahl von Kanälen in der Membran mit letztlich
gesteigerter Leitfähigkeit (Dobrowolski et al., 2007).
6.3.4 C-Terminale Panx1-Phosphorylierungsstellen sind wichtig für die Regulation von Panx1-Kanaleigenschaften
Fünf potentielle Phosphorylierungsstellen sind im C-Terminus von Pannexin1
lokalisiert und wurden ebenfalls mittels gezielter Mutagenese verändert. Das
Tyrosin Y324 wurde zu Phenylalanin, die Serine S328, S343 und S405, sowie
das Threonin T387 zu Alanin mutiert. Es ist bekannt, dass bei Connexinen
insbesondere der C-Terminus zur Regulation von Zelleigenschaften
phosphoryliert wird (Warn-Cramer et al., 1996; Review Solan und Lampe, 2009;
Review Račkauskas et al., 2010), deshalb wurde vermutet, dass auch bei
Pannexinen die Phosphorylierung des C-Terminus wichtig ist.
Die mPanx1-Y324F- und S405A-transfizierten N2A-Zellen verhielten sich
jedoch in den Experimenten ähnlich wie die Mutanten der intrazellulären
Schleife (s. 6.3.3), also sehr ähnlich den Panx1 wt exprimierenden Zellen. Bei
Expression der Y324F-Mutante waren die maximalen Ströme in den IV-Kurven,
und damit die Leitfähigkeit, signifikant grösser, als die der Panx1 wt
exprimierenden Zellen. Dieses Ergebnis konnte durch Messungen an Xenopus-
Oozyten (S. Bunse, pers. Mitteilung) bestätigt werden. Hier ergab sich darüber
hinaus für beide Mutanten eine größere Leitfähigkeit als für die Panx1 wt
exprimierenden Zellen. Folglich spielt auch bei diesen Mutanten, wie bei den
Mutanten der intrazellulären Schleife, Phosphorylierung eine Rolle in Transport

56
und Abbau der Kanäle, oder Kanaleigenschaften, wie Offenwahrscheinlichkeit,
mittlerer Öffnungsdauer und Einzelkanalleitfähigkeit. Bei der S405A-Mutante
konnte zudem eine starke Fluoreszenzintensität und ausgeprägte
Membranlokalisation in den fluoreszenzmikroskopischen Aufnahmen
festgestellt werden. Dies würde auf eine erhöhte Anzahl von Kanälen an der
Zelloberfläche, entweder durch einen schnelleren Transport zur Membran oder
einen verlangsamten Abbau, hindeuten. „Pulse-chase“- Experimente in
Verbindung mit Zellfraktionierung könnten weiteren Aufschluss über den hier
überwiegenden Mechanismus geben, denn mit dieser Methode kann der Weg
einzelner neu synthetisierter Verbindungen durch die Zelle verfolgt werden.
Panx1-T387A-transfizierte N2A-Zellen glichen in den elektrophysiologischen
Messungen ebenfalls den Panx1 wt exprimierenden Zellen, zeigten jedoch
Auffälligkeiten in den konfokalen Aufnahmen und den Western Blot-Analysen.
Konfokalmikroskopisch konnte nur wenig Panx1-Protein in der Zellmembran
nachgewiesen werden und bei den Blot-Analysen pEGFP-N3-mPanx1-T387A-
transfizierter Zellen konnten nur sehr schwache Banden abgelesen werden.
Deshalb wurden von unserer Arbeitsgruppe zusätzlich Versuche zu den
Überlebensraten von T387A-exprimierenden Oozyten durchgeführt (S. Bunse,
pers. Mitteilung). Hier zeigte sich eine erhöhte Sterberate von ca. 30%
innerhalb von 40 h nach Injektion (gegenüber wenigen Prozent bei den Panx1
wt exprimierenden Zellen) und eine Verlängerung der Überlebenszeit unter Cbx.
Eine mögliche Erklärung für die Ergebnisse aus den konfokalmikroskopischen
Aufnahmen und der Western Blot-Analyse wäre also, dass, ähnlich wie bei der
Y10F-Mutante, viele Zellen zum Untersuchungszeitpunkt bereits abgestorben
waren und nur Zellen eingesetzt wurden, die eine geringe Expressionsrate
aufwiesen. Dies unterstützen auch die Ergebnisse aus den
elektrophysiologischen Messungen injizierter Oozyten (S. Bunse, pers.
Mitteilung). Hier waren die maximalen Amplituden signifikant grösser und das
Ruhemembranpotential signifikant kleiner, als bei den Panx1 wt exprimierenden
Zellen. Eine erhöhte Kanal-Aktivität am Ruhemembranpotential mit Verlusten
von Metaboliten und dem Zusammenbruch transmembranärer Ionengradienten
könnte also auch hier für den schnellen Zelltod verantwortlich sein.
Die S343A- und die S328A-Mutante waren von den C-terminalen Mutanten am
auffälligsten. Bei der S343A-Mutante konnte in der Western Blot-Analyse nur

57
sehr schwache Banden gefunden werden und in den konfokalen Aufnahmen
konnte zwar Panx1-Protein an der Zelloberfläche nachgewiesen werden, die
Überlebenszeit war jedoch stark vermindert. Auch in Versuchen zur
Überlebenszeit injizierter Oozyten (S. Bunse, pers. Mitteilung), konnte eine
stark eingeschränkte Überlebensrate festgestellt werden. Schon 40 h nach
Injektion waren alle Oozyten, die die S343A-Mutante exprimierten, degeneriert.
Überraschenderweise konnte die Überlebenszeit auch in Anwesenheit von Cbx
nicht wesentlich verlängert werden. Entsprechend den Ergebnissen aus den
Versuchen zu den Überlebensraten injizierter Oozyten zeigten S343A-
transfizierte N2A-Zellen Auffälligkeiten in den elektrophysiologischen
Messungen. Mit bis zu 1800 pA in einzelnen Messungen waren die maximalen
Stromamplituden sehr groß und der Eingangswiderstand war signifikant kleiner,
als bei den Panx1 wt exprimierenden Zellen. Für diese Ergebnisse ist
vermutlich ebenfalls eine gesteigerte Aktivität des Panx1-S343A-Kanals
verantwortlich. Diese Aktivität ist atypisch, da der Kanal schon bei
Membranspannungen von kleiner -20 mV öffnete. Ein ähnliches Verhalten ist
von humanem Cx26 bekannt, das schon bei Membranpotentialen von -50 mV
öffnet (Gonzáles et al., 2006). Interessanterweise erhöhen einige Mutationen in
Cx26, die zu verschiedenen Hautkrankheiten und genetisch bedingter Taubheit
führen, ebenfalls die Hemikanal-Leitfähigkeit (Lee et al., 2009). Man könnte
vermuten, dass Mutationen von Panx1 genauso zu hereditären Krankheiten
führen können. Zur Bestätigung dieser Annahme müssen allerdings weitere
Untersuchungen über physiologische und pathologische Eigenschaften von
Pannexin1 folgen.
Auffallend war in den elektrophysiologischen Messungen, ähnlich wie in den
Versuchen zur Oozyten-Überlebenszeit, eine inkomplette Blockade durch Cbx.
Denn in den IV-Kurven wiesen die Ströme auch nach Cbx-Applikation noch
eine exponentielle Steigung bei positiven Potentialen auf. Trotzdem war die
Restamplitude nach Cbx-Applikation prozentual kleiner als die der Panx1 wt
exprimierenden Zellen, allerdings war dieser Unterschied nicht signifikant. Der
S-Index verminderte sich hingegen sogar signifikant gegenüber dem der Panx1
wt exprimierenden Zellen. Cbx scheint hier einen größeren Anteil des Stroms
blockiert zu haben, als bei den Panx1 wt exprimierenden Zellen. Man könnte
sich vorstellen, dass die Mutation S343A in einem veränderten

58
Kanaldurchmesser, geänderter Ladung der Kanalpore, oder anderen
Inaktivierungszeiten resultiert. Welcher Mechanismus tatsächlich für die extrem
große Leitfähigkeit und die inkomplette Blockade durch Cbx ursächlich ist, ist
mit heutigem Wissen nicht zu bestimmen, da die Wirkungsweise von Cbx noch
unbekannt ist. Hier müssen weitere Experimente zum besseren Verständnis
folgen.
Die S328A-Mutante verhielt sich im Prinzip wie die S343A-Mutante nur dass sie
in den konfokalmikroskopischen Aufnahmen und den Western-Blot-Analysen
unauffällig war. Untersuchungen von Oozyten-Überlebensraten (S. Bunse, pers.
Mitteilung) ergaben auch für diese Mutante eine eingeschränkte Überlebenszeit
und in den elektrophysiologischen Messungen zeigte sich eine sehr große
maximale Stromamplitude bei erniedrigtem Eingangswiderstand, was ebenfalls
auf eine erhöhte Leitfähigkeit schließen lässt. Auch hier war nur eine
inkomplette Blockade mit Cbx zu erreichen. Ob bei den S328A-transfizierten
Zellen hierfür der gleiche Mechanismus ursächlich ist, wie bei den S343A-
Mutanten, ließe sich z.B. mit Einzelkanalmessungen in weiteren Experimenten
bestimmen.
Zusammenfassend kann also gesagt werden, dass Phosphorylierung wichtig
für die Regulation von Panx1-Kanaleigenschaften ist. Aus den Ergebnissen
dieser Arbeit kann geschlossen werden, dass die Biosynthese und der
Transport der Kanäle zur Plasmamembran nicht qualitativ mittels
Phosphorylierung gesteuert werden. In Abhängigkeit von der Lokalisation
mutierter Aminosäuren im Protein konnten jedoch veränderte Zelleigenschaften
festgestellt werden. Die N-terminale Mutation an der Position Y10 führte zu
strukturellen Änderungen des Panx1-Kanals mit erhöhter
Einzelkanalleitfähigkeit und frühem Zelltod, während Mutationen in der
intrazellulären Schleife sich in N2A-Zellen wie Panx1 wt exprimierenden Zellen
verhielten. Die Mutationen Y324F, S328A und S343A im C-Terminus führten
ebenfalls zu einer erhöhten Leitfähigkeit transfizierter N2A-Zellen, die hier
jedoch auf Phosphorylierungseffekte zurückzuführen ist, welche eine erhöhte
Offenwahrscheinlichkeit, längere mittlere Öffnungsdauer, oder größere
Einzelkanalleitfähigkeit induzierte.

59
7 Zusammenfassung
Pannexine gehören, zusammen mit den Connexinen und Innexinen, zur Gap
junction-Proteinfamilie. Während Connexine gut untersucht sind, sind die
Regulationsmechanismen von Pannexinen noch nicht vollständig erforscht. Da
die Sekundärstruktur der Proteine (Hertzberg et al., 1988) und der Aufbau des
Connexin- und Pannexin-Kanals vergleichbar sind (Makowski et al., 1977;
Boassa et al., 2007), kann jedoch davon ausgegangen werden, dass die
Kanäle ähnlich reguliert werden. Angesichts der Tatsache, dass sowohl
Connexine, als auch Pannexine Phosphoproteine sind (Musil und Goodenough,
1992; Barbe et al., 2006) und der Connexin-Lebenszyklus mittels
Phosphorylierung reguliert wird (s. Review Sáez et al., 2003), sollte in dieser
Arbeit die Funktion potentieller Phosphorylierungsstellen in Panx1 untersucht
werden. Zu diesem Zweck wurden neun Aminosäuren in Panx1 mittels gezielter
Mutagenese verändert und die resultierenden Mutanten mit Hilfe eukaryoter
Expressionsvektoren in N2A-Zellen exprimiert. Unter Einsatz von Western Blot-
Analysen, konfokaler Fluoreszenzmikroskopie und elektrophysiologischen
Messungen wurden transfizierte Zellen untersucht.
Es zeigte sich, dass alle mit den unterschiedlichen Mutanten transfizierten
Zellen potentiell überlebensfähig waren und funktionsfähige Pannexin-Kanäle
gebildet wurden. Phosphorylierung scheint somit kein Mechanismus zur
Regulation der Panx1-Biosynthese oder des Transports der Kanäle zur
Membran zu sein. Während fünf der Mutationen keine Änderungen der
elektrophysiologischen Zelleigenschaften transfizierter N2A-Zellen bewirkten,
verursachte die Expression der N-terminalen Mutante Y10F und der C-
terminalen Mutanten S343A und S328A in N2A-Zellen, vermutlich über die
Ausbildung eines permanent offenen Kanals, einen frühen Zelltod. Die C-
terminale Y324F-Mutante führte zudem zu einer gesteigerten Leitfähigkeit
transfizierter N2A-Zellen. Vergleichbare Daten konnten auch aus bisher
unveröffentlichten Versuchen unserer Arbeitsgruppe mit Xenopus-Oozyten
gewonnen werden. Da bekannt ist, dass der N-Terminus in Panx1 strukturelle
Aufgaben erfüllt (Wang und Dahl, 2010), resultierten die veränderten

60
Kanaleigenschaften der Y10F-Mutante aus ihrem Einfluss auf die
Proteinstruktur. Die gesteigerte Kanal-Aktivität der C-terminalen Mutanten war
hingegen auf eine veränderte Regulation mittels Phosphorylierung
zurückzuführen.
In dieser Arbeit konnte somit erstmals gezeigt werden, dass posttranslationale
Phosphorylierung ein wichtiger Mechanismus zur Regulation der Panx1-
Kanaleigenschaften ist. Da Panx1 in physiologischen und pathologischen
Prozessen, wie der Steuerung der Immunabwehr (Woehrle et al., 2010), oder
dem post-ischämischen Zelltod (Orellana et al., 2011) eine wichtige Rolle spielt,
kann ein besseres Verständnis für Panx1-Regulationsmechanismen einen
Beitrag zur Untersuchung und Therapie solcher Vorgänge leisten.

61
8 Literaturverzeichnis
Ambrosi, C., Gassmann, O., Pranskevich, J.N., Boassa, D., Smock, A., Wang, J., Dahl, G., Steinem, C., Sosinsky, G.E. (2010). Pannexin1 and Pannexin2 Channels Show Quaternary Similarities to Connexons and Different Oligomerization Numbers from Each Other. J. Biol. Chem. 285(32), 24420-31 Bao, L., Locovei, S., Dahl, G. (2004). Pannexin membrane channels are mechanosensitive conduits for ATP. FEBS 572, 65-68 Bao, X., Chen, Y., Reuss, L., Altenberg, G.A. (2004). Functional expression in Xenopus oocytes of gap-junctional hemichannels formed by a cysteine-less connexin 43. J. Biol. Chem. 279, 9689-9692 Baranova, A., Ivanov, D., Petrash, N., Pestova, A., Skoblov, M., Kelmanson, I., Shagin, D., Nazarenko, S., Geraymovych, E., Litvin, O., Tiunova, A., Born, T.L., Usman, N., Staroverov, D., Lukyanov, S., Panchin, Y. (2004). The mammalian pannexin family is homologous to the invertebrate innexin gap junction proteins. Genomics 83, 706-716 Barbe, M.T., Monyer, H., Bruzzone, R. (2006). Cell-Cell Communication Beyond Connexins: The Pannexin Channels. Physiology 21, 103-114 Bennett, M.V.L. and Zukin, R.S. (2004). Electrical coupling and neuronal synchronizytion in the mammalian brain. Neuron 41, 495-511 Beyer, E.C., Paul, D.L. and Goodenough, D.A. (1987). Connexin 43: A protein from a rat heart homologous to a gap junction protein from liver. J. Cell. Biol. 105, 2621-2629 Boassa, D., Ambrosi, C., Qiu, F., Dahl, G., Gaietta, G., Sosinsky, G. (2007). Pannexin1 channels contain a glycosylation site that targets the hexamer to the plasma membrane. J. Biol. Chem. 282, 31733-31743 Bruzzone, R., Hormuzdi, S.G., Barbe, M.T., Herb, A., Monyer, H. (2003). Pannexins, a family of gap junction proteins expressed in brain. Proc. Natl. Acad. Sci. USA 100, 13644–13649 Bruzzone, R., Barbe, M.T., Jakob, N.J., Monyer, H. (2005). Pharmacological properties of homomeric and heteromeric pannexin hemichannels expressed in Xenopus oocytes. J. of Neurochem. 92, 1033-43 Bunse, S., Schmidt, M., Prochnow, N., Zoidl, G., Dermietzel, R. (2010). Intracellular cysteine C346 is essentially involved in regulating Panx1 channel activity. J. Biol. Chem. 285(49), 38444-52

62
Carew, T.J. and Kandel, E.R. (1976). Two functional effects of decreased conductance EPSP´s: synaptic augmentation and increased electrotonic coupling. Science 192, 150-153 Carter, P. (1987). Improved oligonucleotide-directed mutagenesis using M13 verctors. Methods Enzymol. 145, 382-402 Celetti, S.J., Cowan, K.N., Penuela, S., Shao, Q., Churko, J., Laird, D.W. (2010). Implications of pannexin 1 and pannexin 3 for keratinocyte differentiation. J. Cell Sci. 123, 1363-72 Chekeni, F.B., Elliott, M.R., Sandilos, J.K., Walk, S.F., Kinchen, J.M., Lazaroski, E.R., Armstrong, A.J., Penuela, S., Laird, D.W., Salvesen, G.S., Isakson, B.E., Bayliss, D.A., Ravichandran, K.S. (2010). Pannexin 1 channels mediate „find-me“ signal release and membrane permeability during apoptosis. Nature 467, 863-7 Contreras, J.E., Saez, J.C., Bukauskas, F.F., Bennett, M.V. (2003). Gating and regulation of connexin 43 (Cx43) hemichannels. Proc. Natl. Acad. Sci. USA 100, 11388-11393 Cooper, C.D., Solan, J.L., Dolejsi, M.K. and Lampe, P.D. (2000). Analysis of connexin phosphorylation sites. Methods 20, 196-204 Cottrell, G.T., Lin, R., Warn-Cramer, B.J., Lau, A.F., Burt, J.M. (2003). Mechanism of v-Src- and mitogen-activated protein kinase-induced reduction of gap junction communication. Am. J. Physiol. Cell Physiol. 384(2), C511-20 Dando, R. and Roper, S.D. (2009). Cell to cell communication in intact taste buds through ATP signalling from pannexin 1 gap junction hemichannels. J. Physiol. 587, 5899-906 Dermietzel, R. (1998). Gap junctions wiring: a „new“ principle in cell-to-cell communication in the nervous system? Brain. Res. Rev. 26, 176-183 Dermietzel, R. and Spray, D.C. (1993). Gap junctions in the brain: where, what type, how many und why? Trends Neuroscience 16, 186-192 Dobrowolski, R., Sommershof, A., Willecke, K. (2007). Some oculodentodigital dysplasia associated Cx43 mutations cause increased hemichannel activity in addition to deficient gap junction channels. J. Membr. Biol. 219, 9-17 Elfgang, C., Eckert, R., Lichtenberg-Frate, H., Butterweck, A., Traub, O., Klein, R.A., Hulser, D.F., Willecke, K. (1995). Specific permeanility and selective formation of gap junction channels in connexin-transfected HeLa cells. J. Cell. Biol. 129, 805-817 Furshpan, E.J. and Potter, D.D. (1959). Transmission at the giant motor synapses of the crayfish. J.Physiol. 145, 289-325

63
Fushiki, D., Hamda, Y., Yoshimura, R., Endo, Y. (2010). Phylogenetic and bioinformatic analysis of gap junction-related proteins, innexins, pannexins and connexins. Biomed. Res. 31, 133-142 Gomez-Hernandez, J.M., de Miguel, M., Larrosa, B., Gonzalez, D., Barrio, L.C. (2003). Molecular basis of calcium regulation in connexin-32 hemichannels. Proc. Natl. Acad. Sci. USA 100, 16030-16035 Gonzáles, D., Gómez-Hernández, J.M., Barrio, L.C. (2006). Species specificity of mammalian connexin-26 to form open voltage-gated hemichannels. FASEB J. 20, 2329-2338 Goodenough, D.A. and Paul, D.L. (2003). Beyond the gap: functions of unpaired connexon channels. Nat. Rev. Mol. Cell. Biol. 4, 285-294 Goodenough, D.A. and Paul, D.L. (2009). Gap junctions. Cold Spring Harb. Perspect. Biol. 1(1), a002576 Harks, E.G., deRoos, A.D., Peters, P.H., deHaan, L.H., Brouwer, A., Ypey, D.L., van Zoelen, E.J., Theuvenet, A.P. (2001). Fenamates: anovel class of reversible gap junction blockers. J. Pharmacol. Exp. Ther. 298(3), 1033-41 Hertzberg, E.L., Disher, R.M., Tiller, A.A., Zhou, Y., and Cook, R.G. (1988). Topology of the Mr 27,000 liver gap junction protein. Cytoplasmic localization of amino- and carboxyl termini and a hydrophilic domain which is protease-hypersensitive. J. Biol. Chem. 263, 19105-19111 Huang, Y., Grinspan, J.B., Abrams, C.K., Scherer, S.S. (2007). Pannexin1 is expressed by neurons and glia but does not form functional gap junctions. Glia 55, 46-56 Huang, Y.A., Roper, S.D. (2010). Intracellular Ca(2+) and TRPM5-mediated membrane depolarization produce ATP secretion from taste receptor cells. J. Physiol. 588, 2343-50 Iwamoto, T., Nakamura, T., Doyle, A., Ishikawa, M., de Vega, S., Fukumoto, S., Yamada, Y. (2010). Pannexin 3 regulates intracellular ATP/cAMP levels and promotes chondrocyte differentiation. J. Biol. Chem. 285, 18948-58 Jordan, K., Chodock, R., Hand, A.R., Laird, D.W. (2001). The origin of annular junctions: a mechanism of gap junction internalization. J. Cell Sci. 114, 763-73 Kanno, S. and Saffitz, J.E. (2001). The role of myocardial gap junctions in electrical conduction and arrhythmogenesis. Cardiovasc. Pathol. 10, 169-177 Kienitz, M.-C., Bender, K., Dermietzel, R., Pott, L., Zoidl, G. (2011). Pannexin 1 constitutes the large-conductance cation channel of cardiac myocytes. J. Biol. Chem. 286(1), 290-8

64
Klebe, R.J. and Ruddle, F.H. (1969). Neuroblastoma: Cell culture analysis of a differentiating stem cell system. J. Cell. Biol. 43, 69A Kothmann, W.W., Li, X., Burr, G.S., O’Brien, J. (2007). Connexin 35/36 is phosphorylated at regulatory sites in the retina. Vis. Neurosci. 24(3), 363-375 Kumar, N.M. and Gilula, N.B. (1996). The Gap junction communication channel. Cell 84, 381-388 Laemmli (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227(5259), 680-5 Lai, C.P., Bechberger, J.F., Naus, C.C. (2009). Pannexin2 as a novel growth regulator in C6 glioma cells. Oncogene 28, 4402-8 Laird, D.W. (2006). Life cycle of connexins in health and disease. Biochem. J. 394, 527-543 Lampe, P.D. and Lau, A.F. (2000). Regulation of gap junctions by phosphorylation of connexins. Arch. Biochem. Biophys. 384(2), 205-15 Lee, J.R., DeRosa, A.M., White, T.W. (2009). Connexin mutations causing skin disease and deafness increase hemichannel activity and cell death when expressed in Xenopus Oocytes. J. of Inv. Derm. 129, 870-878 Locovei, S., Wang, J., Dahl, G. (2006). Activation of pannexin 1 channels by ATP through P2Y receptors and by cytoplasmic calcium. FEBS Lett. 580, 239-244 Maeda, S. and Tsukihara, T. (2011). Structure of the gap junction channel and ist implications for its biological functions. Cell. Mol. Life Sci. 68, 1115-1129 Maeda, S., Nakagawa, S., Suga, M., Yamashita, E., Oshima, A., Fujiyoshi, Y., Tsukihara, T. (2009). Structure of the connexin 26 gap junction channel at 3.5 Å resolution. Nature 458, 597–602 Makowski, L., Caspar, D.L.D., Phillips, W.C. and Goodenough, D.A. (1977). Gap Junction Structures: II. Analysis of the X-ray diffraction Data. J.Cell. Biol. 74, 629-645 Manthey, D., Banach, K., Desplantez, T., Lee, C.G., Kozak, C.A., Traub, O., Weingart, R. and Willecke, K. (2001). Intracellular domains of mouse connexin26 and -30 affect diffusional and electrical properties of gap junction channels. J. Membr. Biol. 181, 137-148 Martin, P.E.M., Blundell, G., Ahmad, S., Errington, R.J., Evans, W.H. (2001). Multiple pathways in the trafficking and assembly of connexin 26, 32 and 43 into gap junction intercellular communication channels. J. of Cell Sc. 114, 3845-3855

65
Moreno, A.P., Fishman, G.I., Spray, D.C. (1992). Phosphorylation shifts unitary conductance and modifies voltage dependent kinetics of human connexin43 gap junction channels. Biophys. J. 62(1), 51-3 Musil, L.S. and Goodenough, D.A. (1991). Biochemical Analysis of Connexin43 Intracellular Transport, Phosphorylation, and Assembly into Gap Junctional Plaques. J. of Cell. Biol. 115(5), 1357-1374 Newman, E.A. (2001). Propagation of intercellular calcium waves in retinal astrocytes and Müller cells. J. Neurosc. 21, 2215-23 Olk, S., Zoidl, G., Dermietzel, R. (2009). Connexins, Cell Motility, and the Cytoskeleton. Cell Motil. 66, 1000-1016 Orellana, J.A., Froger, N., Ezan, P., Jiang, J.X., Bennett, M.V.L., Naus, C.C., Giaume, C., Sáez, J.C. (2011). ATP and glutamate released via astroglial connexin43 hemichannels mediate neuronal death through activation of pannexin 1 hemichannels. J. Neurochem. 118(5), 826-40 Panchin, Y., Kelmanson, I., Matz, M., Lukyanov, K., Usman, N., Lukyanov, S. (2000). A ubiquitous family of putative gap junction molecules. Curr. Biol. 10, 473-474 Park, J.D., Wallick, C.J., Martyn, K.D., Lau, A.F., Jin, C., Warn-Cramer, B.J. (2007). Akt Phosphorylates Connexin43 on Ser373, a „Mode-1“ Binding Site for 14-3-3. Cell Commun. Adhes. 14(5), 211-226 Penuela, S., Bhalla, R., Gong, X.Q., Cowan, K.N., Celetti, S.J., Cowan, B.J., Bai, D., Shao, Q., Laird, D.W. (2007). Pannexin 1 and pannexin 3 are glycoproteins that exhibit many distinct characteristics from the connexin family of gap junction proteins. J. Cell. Sci. 120(Pt 21), 3772-83 Phelan, P., Stebbings, L.A., Baines, R.A., Bacon, J.P., Davies, J.A., Ford, C. (1998a). Drosophila Shaking-B protein forms gap junctions in paired Xenopus oocytes. Nature 391, 181-184 Phelan, P., Bacon, J.P., Davies, J.A., Stebbings, L.A., Todman, M.G., Avery, L., Baines, R.A., Barnes, T.M., Ford, C., Hekimi, S., Lee, R., Shaw, J.E., Starich, T.A., Curtin, K.D., Sun, Y.A., Wyman, R.J. (1998b). Innexins: a family of invertebrate gap-junction proteins. Trends Genet. 14, 348-349 Phelan, P. and Starich, T.A. (2001). Innexins get into the gap. Bioessays 23, 388-396 Pitts, J.D. and Simms, J.W. (1977). Permeability of junctions between animal cells. Intercellular transfer of nucleotides but not of macromolecules. Exp. Cell Res. 104, 153-163 Račkauskas, M., Neverauskas, V., Skeberdis, V.A. (2010). Diversity and properties of connexin gap junction channels. Medicina (Kaunas) 46(1), 1-12

66
Ray, A., Zoidl, G., Weickert, S., Wahle, P., Dermietzel, R. (2005). Site-specific and developmental expression of pannexin 1 in the mouse nervous system. Eur. J. Neurosci. 21, 3277–3290 Revel, J.P., Karnovsky, M.J. (1967). Hexagonal array of subunits in intercellular junctions of the mouse heart and liver. J. Cell. Biol. 33, C7-C12 Riteau, N., Gasse, P., Fauconnier, L., Gombault, A., Couegnat, M., Fick, L., Kanellopoulos, J., Quesniaux, V.F., Marchand-Adam, S., Crestani, B., Ryffel, B., Couillin, I. (2010). Extracellular ATP is a danger signal activating P2X7 receptor in lung inflammation and fibrosis. Am. J. Respir. Crit. Care Med. 182(6), 774-83 Salameh, A. and Dhein, S. (2011). Adrenergic control of cardiac gap junction function and expression. Naunyn Schmiedbergs Arch. Pharmacol. 383(4), 331-46 Salameh, A., Krautblatter, S., Karl, S., Blanke, K., Rojas Gomez, D., Dhein, S., Pfeiffer, D., Janousek, J. (2009). The signal transduction cascade regulating the expression of the gap junction protein connexin43 by β-adrenoceptors. Br. J. Pharmacol. 158(1), 198-208 Sanderson, M.J., Charles, A.C., Dirksen, E.R. (1990). Mechanical stimulation and intercellular communication increases intracellular Ca2+ ein epithelial cells. Cell. Regul. 1, 585-596 Sanderson, M.J., Charles, A.C., Boitano, S., Dirksen, E.R. (1994). Mechanisms and function of intercellular calcium signaling. Mol.Cell. Endocrinol. 98, 173-87 Sáez, J.C., Berthoud, V.M., Brañes, M.C., Martínez, A.D. und Beyer, E.C. (2003). Plasma membrane channels formed by Connexins: Their regulation and functions. Physiol. Rev. 83, 1359-1400 Sáez, J.C., Retamala, M.A., Basiliob, D., Bukauskas, F.F., Bennett, M.V.L. (2005). Connexin-based gap junction hemichannels: Gating mechanisms. Biochem. Biophys. Acta 1711(2), 215-24 Severs, N.J., Rothery, S., Dupont, E., Coppen, S.R., Yeh, H.I., Ko, Y.S., Matsushita, T., Kaba, R., Halliday, D. (2001). Immunocytochemical analysis of connexin expression in the healthy and diseased cardiovascular system. Microsc. Res. Tech. 52, 301-322 Solan, J.L. and Lampe, P.D. (2009). Connexin43 phosphorylation: structural changes and biological effects. Biochem. J. 419, 261-272 Sosinsky, G.E., Boassa, D., Dermietzel, R., Duffy, H.S., Laird, D.W., Macvicar, B., Naus, C.C., Penuela, S., Scemes, E., Spray, D.C., Thompson, R.J., Zhao, H.B., Dahl, G. (2011). Pannexin channels are not gap junction hemichannels. Channels. 5(3),193-7

67
Söhl, G. and Willecke, K. (2003). An update on connexin genes and their nomenclature in mouse and man. Cell. Commun. Adhes. 10, 173-180 Swayne, L.A., Sorbara, C.D., Bennett, S.A. (2010). Pannexin 2 is expressed by postnatal hippocampal neural progenitors and modulates neuronal commitment. J. Biol. Chem. 285, 24977-86 Tabernero, A., Giaume, C., Medina, J.M. (1996). Endothelin-1 regulates glucose utilization in cultured astrocytes by controlling intercellular communication through gap junction. Glia 16, 187-195 Thibodeau, I.L., Xu, J., Li, Q., Liu, G., Lam, K., Veinot, J.P., Birnie, G.H., Jones, D.L., Krahn, A.D., Lemery, R., Nicholson, B.J., Gollop, M.H. (2010). Paradigm of genetic mosaicism and lone atrial fibrillation: physiological characterization of a connexin 43-deletion mutant identified from atrial tissue. Circulation 122(3), 236-44 Thompson, R.J., Zhou, N., MacVicar, B.A. (2006). Ischemia opens neuronal gap junction hemichannels. Science 312, 924-26 Traub, R.D., Draguhn, A., Whittington, M.A., Baldeweg, T., Bibbig, A., Buhl, E.H., Schmitz, D. (2002). Axonal gap junctions between principal neurons: a novel source of network oscillations, and perhaps epileptogenesis. Rev. Neurosci. 13(1), 1-30 Trexler, E.B., Bennett, M.V., Bargiello, T.A. and Verselis, V.K. (1996). Voltage gating and permeation in gap junction hemichannel. Proc. Natl. Acad. Sci. USA 93, 5836-41 Tsien, R.W. and Weingart, R. (1974). Proceedings: Cyclic AMP: cell-to-cell movement and inotropic effect in ventricular muscle, studied by a cut-end method. J. Physiol. 242, 95P-96P Urschel, S., Höher, T., Schubert, T., Alev, C., Söhl, G., Wörsdörfer, P., Asahara, T., Dermietzel, R., Weiler, R., Willecke, K. (2006). Protein kinase A-mediated phosphorylation of connexin36 in mouse retina results in decreased gap junctional communication between AII amacrine cells. J. Biol. Chem. 281(44), 33163-71 Vanden Abeele, F., Bidaux, G., Gordienko, D., Beck, B., Panchin, Y.V., Baranova, A.V., Ivanov, D.V., Skryma, R., Prevarskaya, N. (2006). Functional implications of calcium permeability of the channel formed by pannexin 1. J. Cell. Biol. 174, 535-546 van Veen, T.A., van Rijen, H.V., Jongsma, H.J. (2000). Electrical conductance of mouse connexin45 gap junction channels is modulated by phosphorylation. Cardiovasc. Res. 46(3), 496-510

68
Vessey, D.A., Li, L., Kelley, M. (2011). Ischemic preconditioning requires opening of pannexin-1/P2X7 channels not only during preconditioning but again after index ischemia at full reperfusion. Mol. Cell Biochem. 351(1-2), 77-84 Volman, V., Matjaž, P., Bazhenov, M. (2011). Gap junctions and epileptic seizures – two sides of the same coin? PLoS One 6(5), e20572 Wang, J. and Dahl, G. (2010). SCAM Analysis of Panx1 suggests a peculiar pore structure. J. Gen. Physiol. 136(5), 515-27 Warn-Cramer, B.J., Lampe, P.D., Kurata, W.E., Kanemitsu, M.Y., Loo, L.W., Eckhart, W., Lau, A.F. (1996). Characterization of the mitogen-activated protein kinase phosphorylation sites on the connexin-43 gap junction protein. J. Biol. Chem. 271(7), 3779-86 Weickert, S., Ray, A., Zoidl, G., Dermietzel, R. (2005). Expression of neural connexins and pannexin1 in the hippocampus and inferior olive: a quantitative approach. Brain Res. Mol. Brain Res. 133, 102–109 Weihong, M., Hui, H., Pelegrin, P., Surprenant, A. (2009). Pharmacological characterization of pannexin-1 currents expressed in mammalian cells. J. Pharmacol. Exp. Ther. 328, 409-18 Woehrle, T., Yip, L., Manohar, M., Sumi, Y., Yao, Y., Chen, Y., Junger, W.G. (2010a). Hypertonic stress regulates T cell function via pannexin-1 hemichannels and P2X receptors. J. Leukoc. Biol. 88, 1181-9 Woehrle, T., Yip, L., Elkhal, A., Sumi, Y., Chen, Y., Yao, Y., Insel, P.A., Junger, W.G. (2010b). Pannexin-1 hemichannel-mediated ATP release together with P2X1 and P2X4 receptors regulate T-cell activation at the immune synapse. Blood. 116, 3475-84 Yeager, M. and Harris, A.L. (2007). Gap junction channel structure in the early 21st century: facts and fantasies. Curr. Opin. Cell Biol. 19(5), 521-8 Yen, M.R. and Saier, M.H. Jr. (2007). Gap junctional proteins of animals: the Innexin/pannexin Superfamilie. Prog. Biophys. Mol. Biol. 94(1-2), 5-14 Zoidl, G. and Dermietzel, R. (2002). On the search fort he electrical synapse: a glimpse at the future. Cell Tissue Res. 310, 137-142 Zoidl, G., Petrasch-Scharwez, E., Ray, A., Meier, C., Bunse, S., Habbes, H.-W., Dahl, G., Dermietzel, R. (2007). Lokalization of the Pannexin1 Protein at postsynaptic sites in the cerebral cortex and hippocampus. Neuroscience 146, 9-16
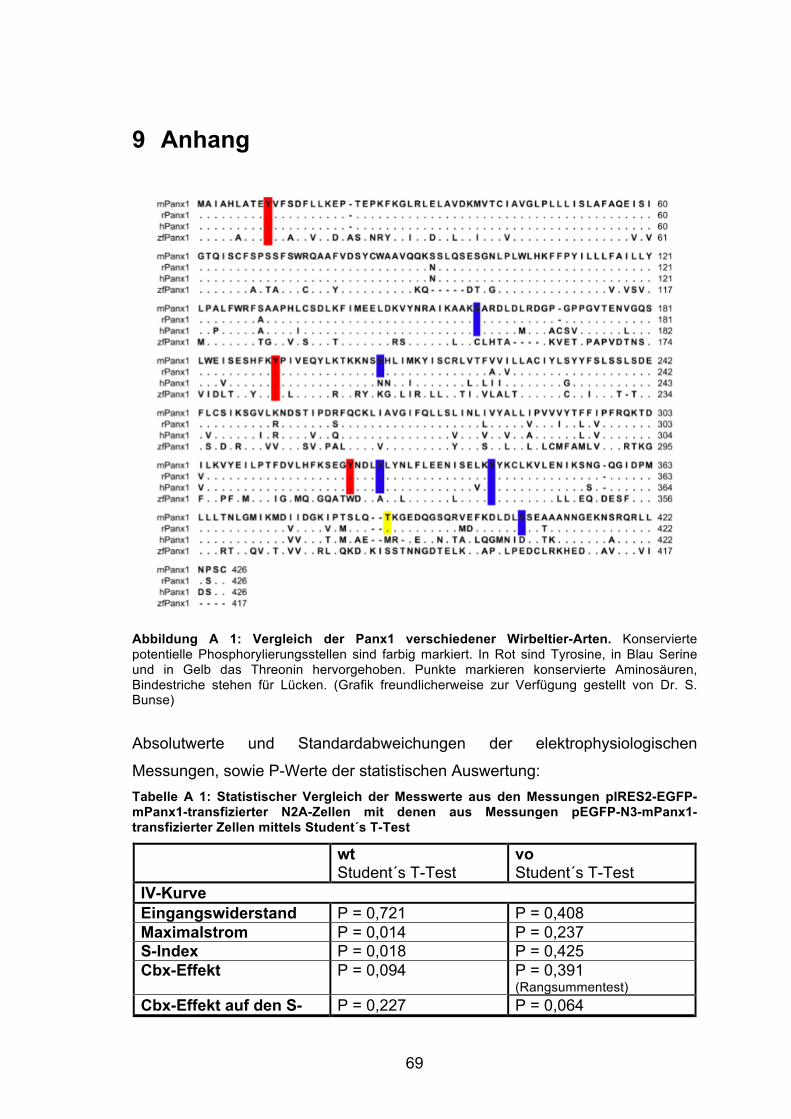
69
9 Anhang
Abbildung A 1: Vergleich der Panx1 verschiedener Wirbeltier-Arten. Konservierte potentielle Phosphorylierungsstellen sind farbig markiert. In Rot sind Tyrosine, in Blau Serine und in Gelb das Threonin hervorgehoben. Punkte markieren konservierte Aminosäuren, Bindestriche stehen für Lücken. (Grafik freundlicherweise zur Verfügung gestellt von Dr. S. Bunse)
Absolutwerte und Standardabweichungen der elektrophysiologischen
Messungen, sowie P-Werte der statistischen Auswertung: Tabelle A 1: Statistischer Vergleich der Messwerte aus den Messungen pIRES2-EGFP-mPanx1-transfizierter N2A-Zellen mit denen aus Messungen pEGFP-N3-mPanx1-transfizierter Zellen mittels Student´s T-Test
wt Student´s T-Test
vo Student´s T-Test
IV-Kurve Eingangswiderstand P = 0,721 P = 0,408 Maximalstrom P = 0,014 P = 0,237 S-Index P = 0,018 P = 0,425 Cbx-Effekt P = 0,094 P = 0,391
(Rangsummentest) Cbx-Effekt auf den S- P = 0,227 P = 0,064
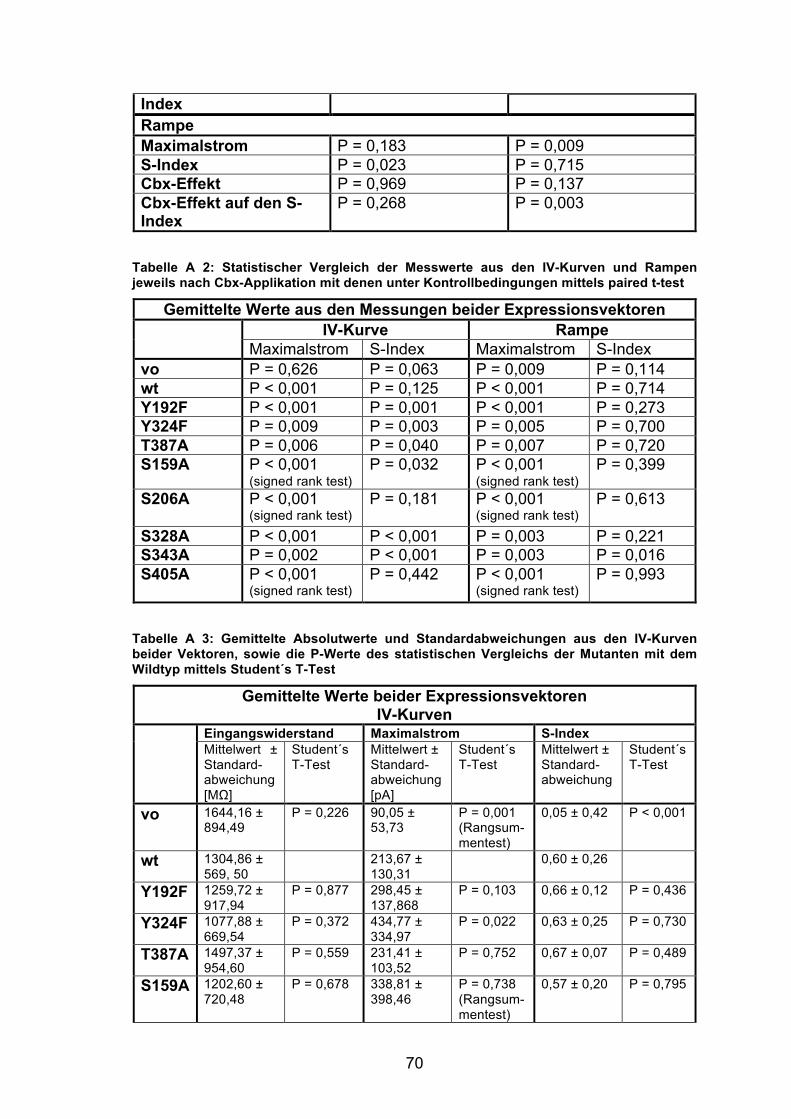
70
Index Rampe Maximalstrom P = 0,183 P = 0,009 S-Index P = 0,023 P = 0,715 Cbx-Effekt P = 0,969 P = 0,137 Cbx-Effekt auf den S-Index
P = 0,268 P = 0,003
Tabelle A 2: Statistischer Vergleich der Messwerte aus den IV-Kurven und Rampen jeweils nach Cbx-Applikation mit denen unter Kontrollbedingungen mittels paired t-test
Gemittelte Werte aus den Messungen beider Expressionsvektoren IV-Kurve Rampe
Maximalstrom S-Index Maximalstrom S-Index vo P = 0,626 P = 0,063 P = 0,009 P = 0,114 wt P < 0,001 P = 0,125 P < 0,001 P = 0,714 Y192F P < 0,001 P = 0,001 P < 0,001 P = 0,273 Y324F P = 0,009 P = 0,003 P = 0,005 P = 0,700 T387A P = 0,006 P = 0,040 P = 0,007 P = 0,720 S159A P < 0,001
(signed rank test) P = 0,032 P < 0,001
(signed rank test) P = 0,399
S206A P < 0,001 (signed rank test)
P = 0,181 P < 0,001 (signed rank test)
P = 0,613
S328A P < 0,001 P < 0,001 P = 0,003 P = 0,221 S343A P = 0,002 P < 0,001 P = 0,003 P = 0,016 S405A P < 0,001
(signed rank test) P = 0,442 P < 0,001
(signed rank test) P = 0,993
Tabelle A 3: Gemittelte Absolutwerte und Standardabweichungen aus den IV-Kurven beider Vektoren, sowie die P-Werte des statistischen Vergleichs der Mutanten mit dem Wildtyp mittels Student´s T-Test
Gemittelte Werte beider Expressionsvektoren IV-Kurven
Eingangswiderstand Maximalstrom S-Index Mittelwert ± Standard-abweichung [MΩ]
Student´s T-Test
Mittelwert ± Standard-abweichung [pA]
Student´s T-Test
Mittelwert ± Standard-abweichung
Student´s T-Test
vo 1644,16 ± 894,49
P = 0,226 90,05 ± 53,73
P = 0,001 (Rangsum-mentest)
0,05 ± 0,42 P < 0,001
wt 1304,86 ± 569, 50
213,67 ± 130,31
0,60 ± 0,26
Y192F 1259,72 ± 917,94
P = 0,877 298,45 ± 137,868
P = 0,103 0,66 ± 0,12 P = 0,436
Y324F 1077,88 ± 669,54
P = 0,372 434,77 ± 334,97
P = 0,022 0,63 ± 0,25 P = 0,730
T387A 1497,37 ± 954,60
P = 0,559 231,41 ± 103,52
P = 0,752 0,67 ± 0,07 P = 0,489
S159A 1202,60 ± 720,48
P = 0,678 338,81 ± 398,46
P = 0,738 (Rangsum-mentest)
0,57 ± 0,20 P = 0,795
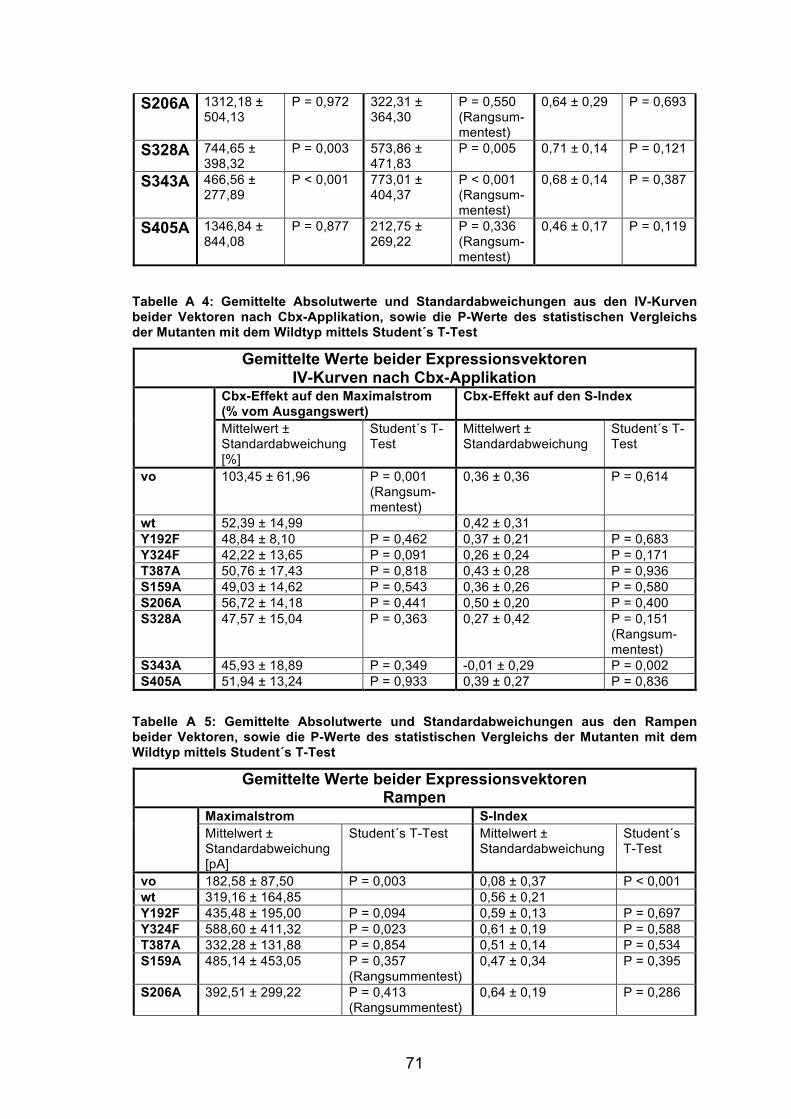
71
S206A 1312,18 ± 504,13
P = 0,972 322,31 ± 364,30
P = 0,550 (Rangsum-mentest)
0,64 ± 0,29 P = 0,693
S328A 744,65 ± 398,32
P = 0,003 573,86 ± 471,83
P = 0,005 0,71 ± 0,14 P = 0,121
S343A 466,56 ± 277,89
P < 0,001 773,01 ± 404,37
P < 0,001 (Rangsum-mentest)
0,68 ± 0,14 P = 0,387
S405A 1346,84 ± 844,08
P = 0,877 212,75 ± 269,22
P = 0,336 (Rangsum-mentest)
0,46 ± 0,17 P = 0,119
Tabelle A 4: Gemittelte Absolutwerte und Standardabweichungen aus den IV-Kurven beider Vektoren nach Cbx-Applikation, sowie die P-Werte des statistischen Vergleichs der Mutanten mit dem Wildtyp mittels Student´s T-Test
Gemittelte Werte beider Expressionsvektoren IV-Kurven nach Cbx-Applikation
Cbx-Effekt auf den Maximalstrom (% vom Ausgangswert)
Cbx-Effekt auf den S-Index
Mittelwert ± Standardabweichung [%]
Student´s T-Test
Mittelwert ± Standardabweichung
Student´s T-Test
vo 103,45 ± 61,96 P = 0,001 (Rangsum-mentest)
0,36 ± 0,36 P = 0,614
wt 52,39 ± 14,99 0,42 ± 0,31 Y192F 48,84 ± 8,10 P = 0,462 0,37 ± 0,21 P = 0,683 Y324F 42,22 ± 13,65 P = 0,091 0,26 ± 0,24 P = 0,171 T387A 50,76 ± 17,43 P = 0,818 0,43 ± 0,28 P = 0,936 S159A 49,03 ± 14,62 P = 0,543 0,36 ± 0,26 P = 0,580 S206A 56,72 ± 14,18 P = 0,441 0,50 ± 0,20 P = 0,400 S328A 47,57 ± 15,04 P = 0,363 0,27 ± 0,42 P = 0,151
(Rangsum-mentest)
S343A 45,93 ± 18,89 P = 0,349 -0,01 ± 0,29 P = 0,002 S405A 51,94 ± 13,24 P = 0,933 0,39 ± 0,27 P = 0,836
Tabelle A 5: Gemittelte Absolutwerte und Standardabweichungen aus den Rampen beider Vektoren, sowie die P-Werte des statistischen Vergleichs der Mutanten mit dem Wildtyp mittels Student´s T-Test
Gemittelte Werte beider Expressionsvektoren Rampen
Maximalstrom S-Index Mittelwert ± Standardabweichung [pA]
Student´s T-Test Mittelwert ± Standardabweichung
Student´s T-Test
vo 182,58 ± 87,50 P = 0,003 0,08 ± 0,37 P < 0,001 wt 319,16 ± 164,85 0,56 ± 0,21 Y192F 435,48 ± 195,00 P = 0,094 0,59 ± 0,13 P = 0,697 Y324F 588,60 ± 411,32 P = 0,023 0,61 ± 0,19 P = 0,588 T387A 332,28 ± 131,88 P = 0,854 0,51 ± 0,14 P = 0,534 S159A 485,14 ± 453,05 P = 0,357
(Rangsummentest) 0,47 ± 0,34 P = 0,395
S206A 392,51 ± 299,22 P = 0,413 (Rangsummentest)
0,64 ± 0,19 P = 0,286
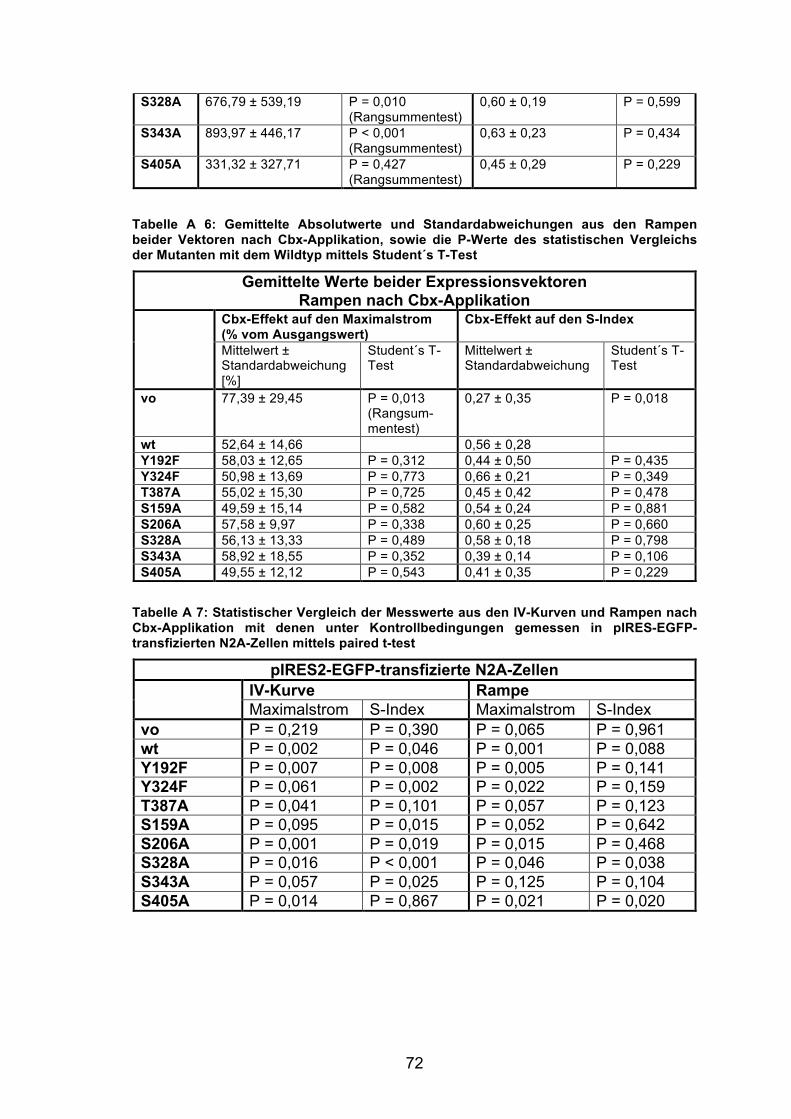
72
S328A 676,79 ± 539,19 P = 0,010 (Rangsummentest)
0,60 ± 0,19 P = 0,599
S343A 893,97 ± 446,17 P < 0,001 (Rangsummentest)
0,63 ± 0,23 P = 0,434
S405A 331,32 ± 327,71 P = 0,427 (Rangsummentest)
0,45 ± 0,29 P = 0,229
Tabelle A 6: Gemittelte Absolutwerte und Standardabweichungen aus den Rampen beider Vektoren nach Cbx-Applikation, sowie die P-Werte des statistischen Vergleichs der Mutanten mit dem Wildtyp mittels Student´s T-Test
Gemittelte Werte beider Expressionsvektoren Rampen nach Cbx-Applikation
Cbx-Effekt auf den Maximalstrom (% vom Ausgangswert)
Cbx-Effekt auf den S-Index
Mittelwert ± Standardabweichung [%]
Student´s T-Test
Mittelwert ± Standardabweichung
Student´s T-Test
vo 77,39 ± 29,45 P = 0,013 (Rangsum-mentest)
0,27 ± 0,35 P = 0,018
wt 52,64 ± 14,66 0,56 ± 0,28 Y192F 58,03 ± 12,65 P = 0,312 0,44 ± 0,50 P = 0,435 Y324F 50,98 ± 13,69 P = 0,773 0,66 ± 0,21 P = 0,349 T387A 55,02 ± 15,30 P = 0,725 0,45 ± 0,42 P = 0,478 S159A 49,59 ± 15,14 P = 0,582 0,54 ± 0,24 P = 0,881 S206A 57,58 ± 9,97 P = 0,338 0,60 ± 0,25 P = 0,660 S328A 56,13 ± 13,33 P = 0,489 0,58 ± 0,18 P = 0,798 S343A 58,92 ± 18,55 P = 0,352 0,39 ± 0,14 P = 0,106 S405A 49,55 ± 12,12 P = 0,543 0,41 ± 0,35 P = 0,229
Tabelle A 7: Statistischer Vergleich der Messwerte aus den IV-Kurven und Rampen nach Cbx-Applikation mit denen unter Kontrollbedingungen gemessen in pIRES-EGFP-transfizierten N2A-Zellen mittels paired t-test
pIRES2-EGFP-transfizierte N2A-Zellen IV-Kurve Rampe
Maximalstrom S-Index Maximalstrom S-Index vo P = 0,219 P = 0,390 P = 0,065 P = 0,961 wt P = 0,002 P = 0,046 P = 0,001 P = 0,088 Y192F P = 0,007 P = 0,008 P = 0,005 P = 0,141 Y324F P = 0,061 P = 0,002 P = 0,022 P = 0,159 T387A P = 0,041 P = 0,101 P = 0,057 P = 0,123 S159A P = 0,095 P = 0,015 P = 0,052 P = 0,642 S206A P = 0,001 P = 0,019 P = 0,015 P = 0,468 S328A P = 0,016 P < 0,001 P = 0,046 P = 0,038 S343A P = 0,057 P = 0,025 P = 0,125 P = 0,104 S405A P = 0,014 P = 0,867 P = 0,021 P = 0,020
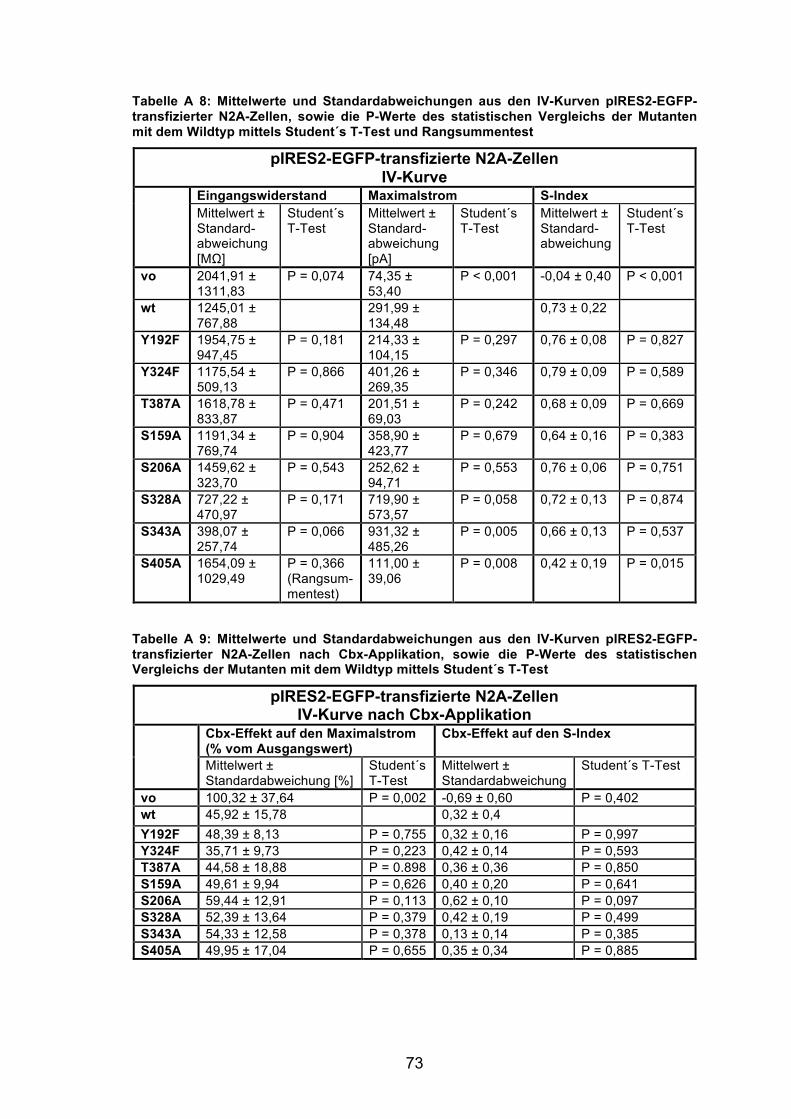
73
Tabelle A 8: Mittelwerte und Standardabweichungen aus den IV-Kurven pIRES2-EGFP-transfizierter N2A-Zellen, sowie die P-Werte des statistischen Vergleichs der Mutanten mit dem Wildtyp mittels Student´s T-Test und Rangsummentest
pIRES2-EGFP-transfizierte N2A-Zellen IV-Kurve
Eingangswiderstand Maximalstrom S-Index Mittelwert ± Standard-abweichung [MΩ]
Student´s T-Test
Mittelwert ± Standard-abweichung [pA]
Student´s T-Test
Mittelwert ± Standard-abweichung
Student´s T-Test
vo 2041,91 ± 1311,83
P = 0,074 74,35 ± 53,40
P < 0,001 -0,04 ± 0,40 P < 0,001
wt 1245,01 ± 767,88
291,99 ± 134,48
0,73 ± 0,22
Y192F 1954,75 ± 947,45
P = 0,181 214,33 ± 104,15
P = 0,297 0,76 ± 0,08 P = 0,827
Y324F 1175,54 ± 509,13
P = 0,866 401,26 ± 269,35
P = 0,346 0,79 ± 0,09 P = 0,589
T387A 1618,78 ± 833,87
P = 0,471 201,51 ± 69,03
P = 0,242 0,68 ± 0,09 P = 0,669
S159A 1191,34 ± 769,74
P = 0,904 358,90 ± 423,77
P = 0,679 0,64 ± 0,16 P = 0,383
S206A 1459,62 ± 323,70
P = 0,543 252,62 ± 94,71
P = 0,553 0,76 ± 0,06 P = 0,751
S328A 727,22 ± 470,97
P = 0,171 719,90 ± 573,57
P = 0,058 0,72 ± 0,13 P = 0,874
S343A 398,07 ± 257,74
P = 0,066 931,32 ± 485,26
P = 0,005 0,66 ± 0,13 P = 0,537
S405A 1654,09 ± 1029,49
P = 0,366 (Rangsum-mentest)
111,00 ± 39,06
P = 0,008 0,42 ± 0,19 P = 0,015
Tabelle A 9: Mittelwerte und Standardabweichungen aus den IV-Kurven pIRES2-EGFP-transfizierter N2A-Zellen nach Cbx-Applikation, sowie die P-Werte des statistischen Vergleichs der Mutanten mit dem Wildtyp mittels Student´s T-Test
pIRES2-EGFP-transfizierte N2A-Zellen IV-Kurve nach Cbx-Applikation
Cbx-Effekt auf den Maximalstrom (% vom Ausgangswert)
Cbx-Effekt auf den S-Index
Mittelwert ± Standardabweichung [%]
Student´s T-Test
Mittelwert ± Standardabweichung
Student´s T-Test
vo 100,32 ± 37,64 P = 0,002 -0,69 ± 0,60 P = 0,402 wt 45,92 ± 15,78 0,32 ± 0,4 Y192F 48,39 ± 8,13 P = 0,755 0,32 ± 0,16 P = 0,997 Y324F 35,71 ± 9,73 P = 0,223 0,42 ± 0,14 P = 0,593 T387A 44,58 ± 18,88 P = 0.898 0,36 ± 0,36 P = 0,850 S159A 49,61 ± 9,94 P = 0,626 0,40 ± 0,20 P = 0,641 S206A 59,44 ± 12,91 P = 0,113 0,62 ± 0,10 P = 0,097 S328A 52,39 ± 13,64 P = 0,379 0,42 ± 0,19 P = 0,499 S343A 54,33 ± 12,58 P = 0,378 0,13 ± 0,14 P = 0,385 S405A 49,95 ± 17,04 P = 0,655 0,35 ± 0,34 P = 0,885
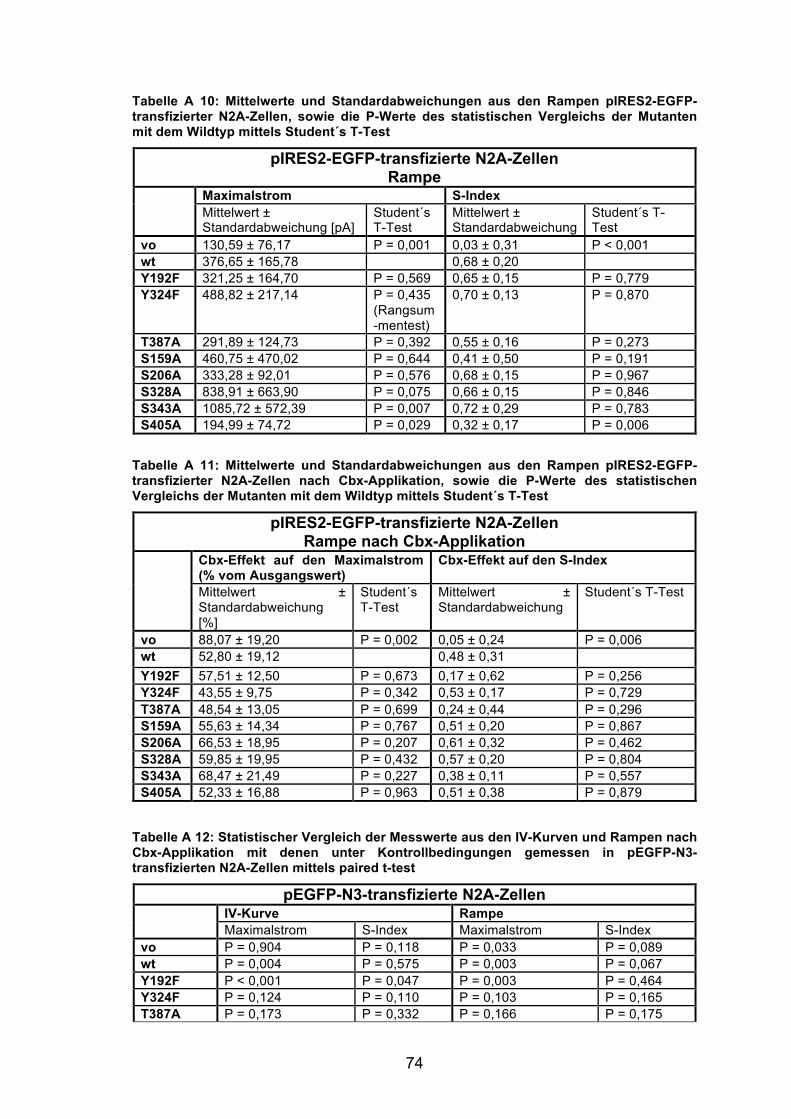
74
Tabelle A 10: Mittelwerte und Standardabweichungen aus den Rampen pIRES2-EGFP-transfizierter N2A-Zellen, sowie die P-Werte des statistischen Vergleichs der Mutanten mit dem Wildtyp mittels Student´s T-Test
pIRES2-EGFP-transfizierte N2A-Zellen Rampe
Maximalstrom S-Index Mittelwert ± Standardabweichung [pA]
Student´s T-Test
Mittelwert ± Standardabweichung
Student´s T-Test
vo 130,59 ± 76,17 P = 0,001 0,03 ± 0,31 P < 0,001 wt 376,65 ± 165,78 0,68 ± 0,20 Y192F 321,25 ± 164,70 P = 0,569 0,65 ± 0,15 P = 0,779 Y324F 488,82 ± 217,14 P = 0,435
(Rangsum-mentest)
0,70 ± 0,13 P = 0,870
T387A 291,89 ± 124,73 P = 0,392 0,55 ± 0,16 P = 0,273 S159A 460,75 ± 470,02 P = 0,644 0,41 ± 0,50 P = 0,191 S206A 333,28 ± 92,01 P = 0,576 0,68 ± 0,15 P = 0,967 S328A 838,91 ± 663,90 P = 0,075 0,66 ± 0,15 P = 0,846 S343A 1085,72 ± 572,39 P = 0,007 0,72 ± 0,29 P = 0,783 S405A 194,99 ± 74,72 P = 0,029 0,32 ± 0,17 P = 0,006
Tabelle A 11: Mittelwerte und Standardabweichungen aus den Rampen pIRES2-EGFP-transfizierter N2A-Zellen nach Cbx-Applikation, sowie die P-Werte des statistischen Vergleichs der Mutanten mit dem Wildtyp mittels Student´s T-Test
Tabelle A 12: Statistischer Vergleich der Messwerte aus den IV-Kurven und Rampen nach Cbx-Applikation mit denen unter Kontrollbedingungen gemessen in pEGFP-N3-transfizierten N2A-Zellen mittels paired t-test
pEGFP-N3-transfizierte N2A-Zellen IV-Kurve Rampe
Maximalstrom S-Index Maximalstrom S-Index vo P = 0,904 P = 0,118 P = 0,033 P = 0,089 wt P = 0,004 P = 0,575 P = 0,003 P = 0,067 Y192F P < 0,001 P = 0,047 P = 0,003 P = 0,464 Y324F P = 0,124 P = 0,110 P = 0,103 P = 0,165 T387A P = 0,173 P = 0,332 P = 0,166 P = 0,175
pIRES2-EGFP-transfizierte N2A-Zellen Rampe nach Cbx-Applikation
Cbx-Effekt auf den Maximalstrom (% vom Ausgangswert)
Cbx-Effekt auf den S-Index
Mittelwert ± Standardabweichung [%]
Student´s T-Test
Mittelwert ± Standardabweichung
Student´s T-Test
vo 88,07 ± 19,20 P = 0,002 0,05 ± 0,24 P = 0,006 wt 52,80 ± 19,12 0,48 ± 0,31 Y192F 57,51 ± 12,50 P = 0,673 0,17 ± 0,62 P = 0,256 Y324F 43,55 ± 9,75 P = 0,342 0,53 ± 0,17 P = 0,729 T387A 48,54 ± 13,05 P = 0,699 0,24 ± 0,44 P = 0,296 S159A 55,63 ± 14,34 P = 0,767 0,51 ± 0,20 P = 0,867 S206A 66,53 ± 18,95 P = 0,207 0,61 ± 0,32 P = 0,462 S328A 59,85 ± 19,95 P = 0,432 0,57 ± 0,20 P = 0,804 S343A 68,47 ± 21,49 P = 0,227 0,38 ± 0,11 P = 0,557 S405A 52,33 ± 16,88 P = 0,963 0,51 ± 0,38 P = 0,879
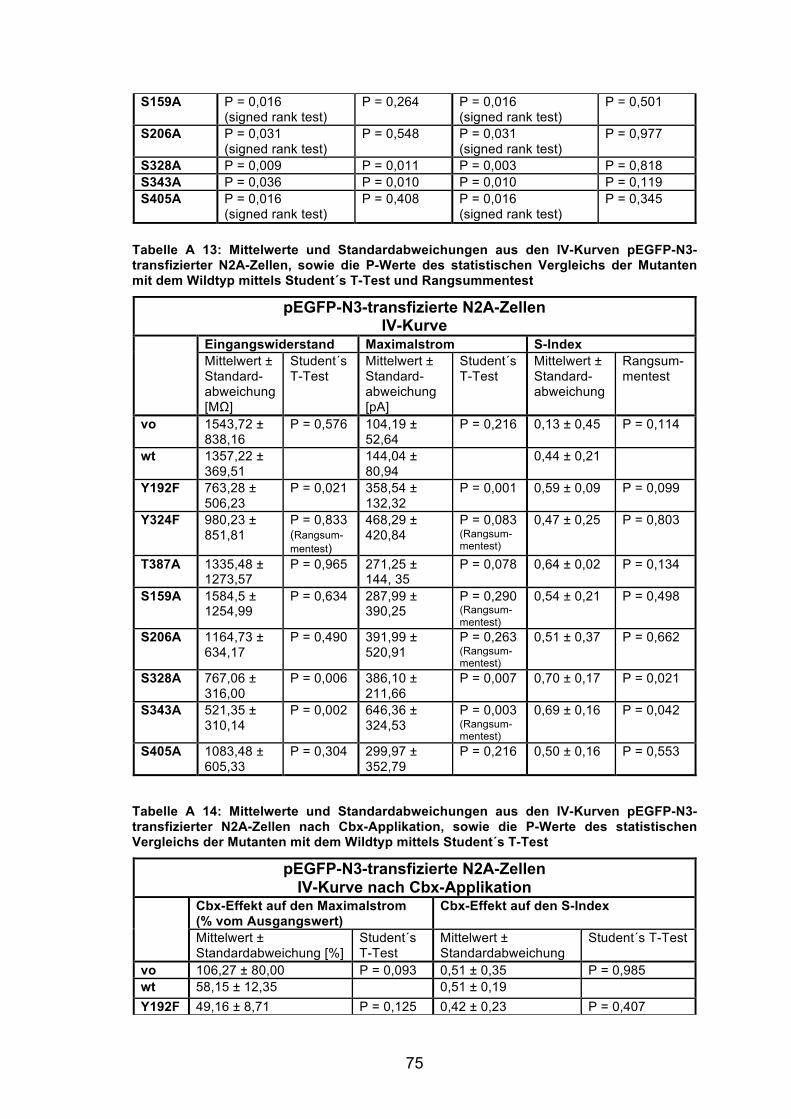
75
S159A P = 0,016 (signed rank test)
P = 0,264 P = 0,016 (signed rank test)
P = 0,501
S206A P = 0,031 (signed rank test)
P = 0,548 P = 0,031 (signed rank test)
P = 0,977
S328A P = 0,009 P = 0,011 P = 0,003 P = 0,818 S343A P = 0,036 P = 0,010 P = 0,010 P = 0,119 S405A P = 0,016
(signed rank test) P = 0,408 P = 0,016
(signed rank test) P = 0,345
Tabelle A 13: Mittelwerte und Standardabweichungen aus den IV-Kurven pEGFP-N3-transfizierter N2A-Zellen, sowie die P-Werte des statistischen Vergleichs der Mutanten mit dem Wildtyp mittels Student´s T-Test und Rangsummentest
pEGFP-N3-transfizierte N2A-Zellen IV-Kurve
Eingangswiderstand Maximalstrom S-Index Mittelwert ± Standard-abweichung [MΩ]
Student´s T-Test
Mittelwert ± Standard-abweichung [pA]
Student´s T-Test
Mittelwert ± Standard-abweichung
Rangsum-mentest
vo 1543,72 ± 838,16
P = 0,576 104,19 ± 52,64
P = 0,216 0,13 ± 0,45 P = 0,114
wt 1357,22 ± 369,51
144,04 ± 80,94
0,44 ± 0,21
Y192F 763,28 ± 506,23
P = 0,021 358,54 ± 132,32
P = 0,001 0,59 ± 0,09 P = 0,099
Y324F 980,23 ± 851,81
P = 0,833 (Rangsum-mentest)
468,29 ± 420,84
P = 0,083 (Rangsum-mentest)
0,47 ± 0,25 P = 0,803
T387A 1335,48 ± 1273,57
P = 0,965 271,25 ± 144, 35
P = 0,078 0,64 ± 0,02 P = 0,134
S159A 1584,5 ± 1254,99
P = 0,634 287,99 ± 390,25
P = 0,290 (Rangsum-mentest)
0,54 ± 0,21 P = 0,498
S206A 1164,73 ± 634,17
P = 0,490 391,99 ± 520,91
P = 0,263 (Rangsum-mentest)
0,51 ± 0,37 P = 0,662
S328A 767,06 ± 316,00
P = 0,006 386,10 ± 211,66
P = 0,007 0,70 ± 0,17 P = 0,021
S343A 521,35 ± 310,14
P = 0,002 646,36 ± 324,53
P = 0,003 (Rangsum-mentest)
0,69 ± 0,16 P = 0,042
S405A 1083,48 ± 605,33
P = 0,304 299,97 ± 352,79
P = 0,216 0,50 ± 0,16 P = 0,553
Tabelle A 14: Mittelwerte und Standardabweichungen aus den IV-Kurven pEGFP-N3-transfizierter N2A-Zellen nach Cbx-Applikation, sowie die P-Werte des statistischen Vergleichs der Mutanten mit dem Wildtyp mittels Student´s T-Test
pEGFP-N3-transfizierte N2A-Zellen IV-Kurve nach Cbx-Applikation
Cbx-Effekt auf den Maximalstrom (% vom Ausgangswert)
Cbx-Effekt auf den S-Index
Mittelwert ± Standardabweichung [%]
Student´s T-Test
Mittelwert ± Standardabweichung
Student´s T-Test
vo 106,27 ± 80,00 P = 0,093 0,51 ± 0,35 P = 0,985 wt 58,15 ± 12,35 0,51 ± 0,19 Y192F 49,16 ± 8,71 P = 0,125 0,42 ± 0,23 P = 0,407
76
Y324F 48,73 ± 14,79 P = 0,226 0,09 ± 0,21 P = 0,003 T387A 58,99 ± 14,09 P = 0,923 0,51 ± 0,18 P = 0,945 S159A 54,24 ± 23,60 P = 0,233 0,35 ± 0,30 P = 0,154 S206A 54,00 ± 16,07 P = 0,581 0,39 ± 0,21 P = 0,278 S328A 41,36 ± 15,41 P = 0,030 0,11 ± 0,55 P = 0,058 S343A 39,21 ± 21,62 P = 0,056 -0,12 ± 0,35 P < 0,001 S405A 53,65 ± 10,06 P = 0,488 0,43 ± 0,23 P = 0,499
Tabelle A 15: Mittelwerte und Standardabweichungen aus den Rampen pEGFP-N3-transfizierter N2A-Zellen, sowie die P-Werte des statistischen Vergleichs der Mutanten mit dem Wildtyp mittels Student´s T-Test
pEGFP-N3-transfizierte N2A-Zellen Rampe
Maximalstrom S-Index Mittelwert ± Standardabweichung [pA]
Student´s T-Test
Mittelwert ± Standardabweichung
Student´s T-Test
vo 229,38 ± 70,86 P = 0,486 0,11 ± 0,43 P = 0,036 wt 268,05 ± 155,11 0,45 ± 0,17 Y192F 517,07 ± 181,63 P = 0,010 0,54 ± 0,11 P= 0,247 Y324F 688,37 ± 555,54 P = 0,110
(Rangsum-mentest)
0,51 ± 0,21 P = 0,570
T387A 386,12 ± 145,70 P = 0,275 0,45 ± 0,12 P = 0,963 S159A 459,64 ± 458,64 P = 0,177 0,49 ± 0,21 P = 0,542 S206A 451,75 ± 425,56 P = 0,252 0,60 ± 0,22 P = 0,163 S328A 468,36 ± 221,53 P = 0,051 0,51 ± 0,21 P = 0,545 S343A 740,56 ± 293,63 P = 0,002 0,56 ± 0,15 P = 0,264 S405A 448,18 ± 419,07 P = 0,251 0,54 ± 0,34 P = 0,540
Tabelle A 16: Mittelwerte und Standardabweichungen aus den Rampen pEGFP-N3-transfizierter N2A-Zellen nach Cbx-Applikation, sowie die P-Werte des statistischen Vergleichs der Mutanten mit dem Wildtyp mittels Student´s T-Test
pEGFP-N3-transfizierte N2A-Zellen Rampe nach Cbx-Applikation
Cbx-Effekt auf den Maximalstrom (% vom Ausgangswert)
Cbx-Effekt auf den S-Index
Mittelwert ± Standardabweichung [%]
Student´s T-Test
Mittelwert ± Standardabweichung
Student´s T-Test
vo 67,77 ± 34,50 P = 0,220 0,52 ± 0,29 P = 0,372 wt 52,51 ± 10,48 0,57 ± 0,24 Y192F 58,41 ± 13,73 P = 0,345 0,66 ± 0,25 P = 0,955 Y324F 58,42 ± 13,73 P = 0,382 0,82 ± 0,14 P = 0,211 T387A 63,66 ± 15,85 P = 0,185 0,73 ± 0,19 P = 0,619 S159A 47,85 ± 16,80 P = 0,221 0,57 ± 0,28 P = 0,604 S206A 56,12 ± 12,07 P = 0,548 0,60 ± 0,18 P = 0,689 S328A 51.88 ± 9,63 P = 0,905 0,60 ± 0,15 P = 0,677 S343A 51,29± 13,32 P = 0,852 0,39 ± 0,18 P = 0,068 S405A 47,18 ± 6,51 P = 0,259 0,42 ± 0,33 P = 0,057

Danksagung An erster Stelle möchte ich Herrn Professor Dr. R. Dermietzel für die
Überlassung des Themas dieser Arbeit, die Möglichkeit in seinem Labor zu
arbeiten und die Übernahme des Referats danken.
Weiterhin danke ich Herrn PD Dr. M. Schmidt und Frau Dr. S. Bunse für die
Betreuung meiner Arbeit, ihre unaufhörliche Diskussionsbereitschaft, die
Beantwortung aller Fragen und dafür, dass sie meine Arbeit Korrektur gelesen
haben. Frau Dr. S. Bunse möchte ich insbesondere für die Herstellung der
Plasmide und die Hilfe bei den konfokalen Aufnahmen danken, Herrn Dr. M.
Schmidt für die Hilfe bei Problemen mit den elektrophysiologischen Messungen.
Auch bei Frau Dr. K. Engelhardt möchte ich mich dafür bedanken, dass Sie
meine Arbeit Korrektur gelesen hat.
Mein Dank gilt außerdem Helga Schulze für ihre Hilfe bei der Bildgestaltung,
Christiane Zoidl, Jeannette Willms, Sabine Peuckert und Sabine Schreiber-
Minjoli für ihre Hilfe in labortechnischen Angelegenheiten und PD Dr. G. Zoidl
für seine Ratschläge und sein Interesse an meiner Arbeit.
Außerdem danke ich meinen Eltern, meiner Schwester und Florian Blumeroth,
die immer für mich da sind.
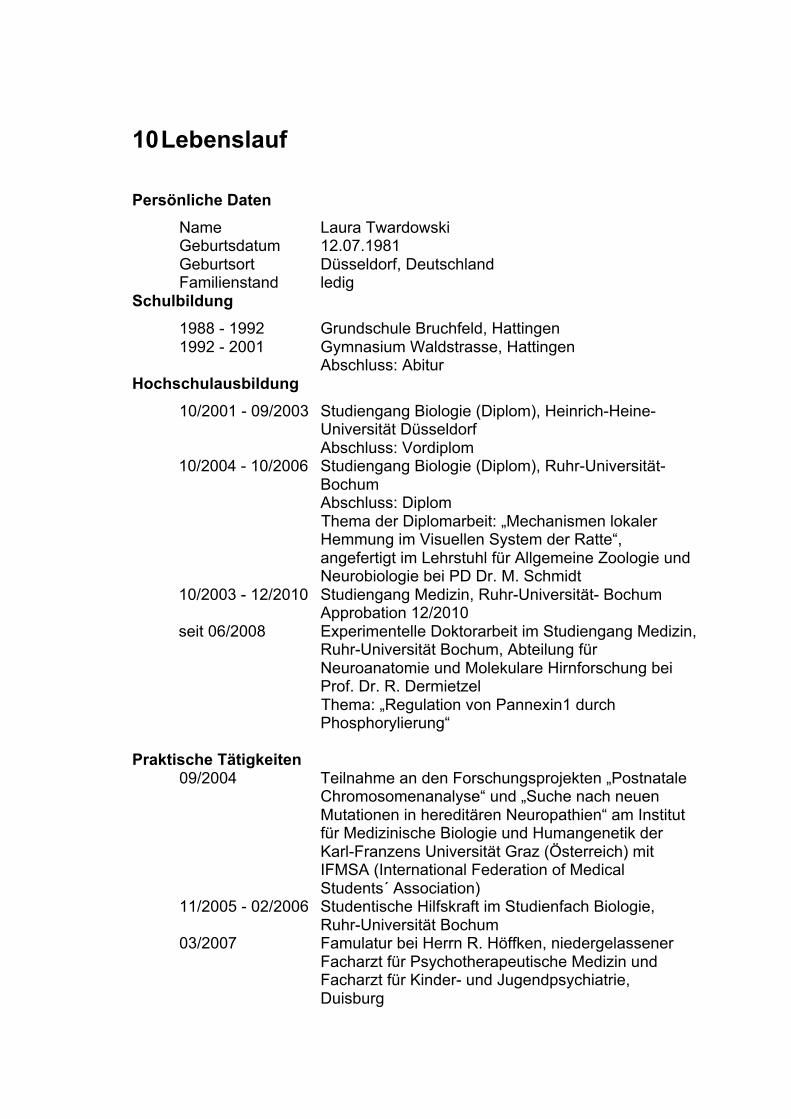
10 Lebenslauf
Persönliche Daten Name Laura Twardowski Geburtsdatum 12.07.1981 Geburtsort Düsseldorf, Deutschland Familienstand ledig
Schulbildung 1988 - 1992 Grundschule Bruchfeld, Hattingen 1992 - 2001 Gymnasium Waldstrasse, Hattingen Abschluss: Abitur
Hochschulausbildung 10/2001 - 09/2003 Studiengang Biologie (Diplom), Heinrich-Heine-
Universität Düsseldorf Abschluss: Vordiplom
10/2004 - 10/2006 Studiengang Biologie (Diplom), Ruhr-Universität-Bochum Abschluss: Diplom Thema der Diplomarbeit: „Mechanismen lokaler Hemmung im Visuellen System der Ratte“, angefertigt im Lehrstuhl für Allgemeine Zoologie und Neurobiologie bei PD Dr. M. Schmidt
10/2003 - 12/2010 Studiengang Medizin, Ruhr-Universität- Bochum Approbation 12/2010
seit 06/2008 Experimentelle Doktorarbeit im Studiengang Medizin, Ruhr-Universität Bochum, Abteilung für Neuroanatomie und Molekulare Hirnforschung bei Prof. Dr. R. Dermietzel Thema: „Regulation von Pannexin1 durch Phosphorylierung“
Praktische Tätigkeiten
09/2004 Teilnahme an den Forschungsprojekten „Postnatale Chromosomenanalyse“ und „Suche nach neuen Mutationen in hereditären Neuropathien“ am Institut für Medizinische Biologie und Humangenetik der Karl-Franzens Universität Graz (Österreich) mit IFMSA (International Federation of Medical Students´ Association)
11/2005 - 02/2006 Studentische Hilfskraft im Studienfach Biologie, Ruhr-Universität Bochum
03/2007 Famulatur bei Herrn R. Höffken, niedergelassener Facharzt für Psychotherapeutische Medizin und Facharzt für Kinder- und Jugendpsychiatrie, Duisburg

06/2007 - 10/2007 Wissenschaftliche Hilfskraft im Studienfach Biologie, Ruhr-Universität Bochum
07/2007 Studienaufenthalt im Rahmen der Doktorarbeit am Institut für Medizinische Grundlagenforschung der Universität von Oslo (Norwegen)
09/2007 Famulatur im Universitätskrankenhaus „Mater Misericordiae“ des Universitäts Colleges Dublin (Irland) im Fachbereich Pneumologie und Transplantationsmedizin
02/2008 Famulatur bei Dr. R. Müller, niedergelassener Facharzt für Radiologie, Hattingen
03/2008 Famulatur bei Dr. D. Metzler, niedergelassener Facharzt für Strahlentherapie, Hattingen
09/2008 Famulatur im Piteå Älvdals Krankenhaus (Schweden) in der Abteilung für Innere Medizin
08/2009 - 07/2010 Praktisches Jahr im Marien-Hospital Witten (Innere Medizin, Chirurgie, Gynäkologie)
seit 04/2011 Assistenzärztin in der Abteilung für Labormedizin des Robert-Bosch-Krankenhauses in Stuttgart