Embed Size (px)
Citation preview

Auslosung von embryonalen Fehlbildungen beim Zebrabarbling 1. Einleitung
Angeborene Fehlbildungen (Mii3bildungen) sind ein Problem, das weiten Kreisen erst durch die Contergan-Tragodie vor etwa 20 Jahren bewui3t wurde. Die Haufigkeit menschlicher Fehlbildungen nahm aber schon vorher zu und scheint auch weiter im Steigen begriffen. Daher sollten die Prinzi- pien der Fehlentwicklung oder Teratogenese allgemein bekannt sein und im Unterricht entsprechend berucksichtigt werden. Einige wesentliche Prinzipien lassen sich aus Versu- chen mit Embryonen des Zebrabarblings ab- leiten. Dieser bekannte Aquarienfisch, der zunehmend auch zur Erfassung schadlicher Umwelteinflusse benutzt wird [7 ] , hat klei- ne, glasklare Eier, die sich hervorragend zur Beobachtung der Embryonalentwicklung mit einem normalen Schulermikroskop eignen [5, 161. Die hier beschriebenen Experimente belegen die fur viele Fehlentwicklungen cha- rakteristische Abhangigkeit von schadlichen Einfliissen wahrend einer bestimmten sensi- blen Entwicklungsphase. Sie lehren, dai3 eine einzige fruhe Entwicklungsanomalie eine ganze Kette von Folgevorgangen in falsche Bahnen lenken kann. Weiter zeigen diese Versuche, dai3 unterschiedliche schadigende Einflusse (Teratogene) zu vergleichbaren Anomalien fuhren konnen, wenn sie wah- rend ein und derselben Entwicklungsphase auf den Embryo einwirken. Schliei3lich lehrt das Spektrum der Versuchsergebnisse, dai3 die Reaktion auf einen bestimmten Fehlbil- dungsreiz individuell recht verschieden aus- fallen oder auch ganz unterbleiben kann.
Obschon unsere Experimente ein Wirbeltier betreffen, erscheinen sie bei Abwagung der Gegegebenheiten vertretbar. Die behandel- ten Fischembryonen erreichen namlich ent- weder uberhaupt nicht jenen Ausbildungs- grad des Zentralnervensystems, der irgendei- ne Form von Bewui3tsein erwarten lafit, oder man fixiert sie vor Erreichen dieser Stufe. Unter naturlichen Bedingungen wurden oh- nehin die meisten dieser Embryonen nicht einmal so weit kommen - ihre eigenen Vater oder andere Laichrauber hatten sie schon vorher verschlungen.
2. Technische Angaben
Halbwuchsige Zebrabarblinge (Danios) der Art Brachydanio rerio (Hamilton-Buchanan 1822) kauft man fur Groschenbetrage in der Zoohandlung, mindestens 2 Weibchen und 4 Mannchen (halbwuchsig nur vom Fachmann
zu unterscheiden). Man halt sie nach Ge- schlechtern getrennt bei ca. 26°C und futtert sie vorwiegend rnit Lebendfutter, bis die Weibchen durch Laichansatz stark ange- schwollen erscheinen. Dann setzt man je ein Weibchen und zwei Mannchen in einen Laichkasten mit einigen schwimmenden Wasserpflanzen und hangt diesen in ein Bek- ken (z. B. Kunststoffaquarium) ohne Boden- belag und Pflanzen, das auf dunkler Unterla- ge steht (Abbildung 1). Als Laichkasten eig- nen sich Kuhlschrank-Klarsichtbehalter mit Deckel (ca. 20 x 20 x 10 cm), deren herausge- sagter Boden durch Perlongaze (Vorhang- stoff) von etwa 2-3 mm Maschenweite er- setzt wird (Kunstharzkleber!). Das Ablai- chen erfolgt kurz nach dem Morgengrauen; man kann das Zeitprogramm der Fische um- stimmen, indem man eine Pappkiste uber die Halterungsaquarien stulpt und die Beleuch- tung jeden Tag etwa 20 min fruher oder spa- ter einschaltet, bis das kunstliche Morgen- grauen auf die gewunschte Ablaichstunde fallt. Dann setzt man die Trios wieder zum Ablaichen an, und zwar am Vorabend des Versuchstages im Dunklen. Der Laichvor- gang ist von aufgeregtem Verhalten der Fi- sche begleitet. Laichen pralle Weibchen uber mehrere Tage nicht ab, wechselt man die Mannchen aus - die Weibchen haben ihre Vorlieben und Antipathien! Nach dem Ab- laichen hebt man den Laichkasten samt Fi- schen und Pflanzen heraus und uberfuhrt ihn in ein anderes Aquarium oder trennt die Trios wieder. Temperaturspriinge sind fur Fische wie Eier nachteilig, daher mui3 stets gleich temperiertes Wasser bereitgehalten werden. Die Weibchen sind nach 2-3 Tagen wieder laichbereit. Pro Laichakt legt ein Weibchen durchschnittlich 100-200 Eier ab.
Die Eier, die beim Ablaichen durch die Gaze gefallen sind, heben sich im streifenden Licht gut vom dunklen Untergrund des Aquariums ab und konnen aufpipettiert oder dekantiert werden. Man ubertragt sie in halbhoch mit Wasser gefullte Plastik-Petrischalen, die man zur Warmeregelung auf einem Aquarium schwimmen lafit, falls kein Brutschrank mit ca. 26°C verfugbar ist. Bei tieferen Tempera- turen verlauft die Entwicklung zunehmend abnorm.
Zur Auslosung der Fehlbildungen benutzen wir organische Verbindungen, im Hauptver- such Ethylalkohol (Ethanol) und zur Ergan- zung ggf. Colcemid, eine Modifikation des Giftes der Herbstzeitlose (Colchicin), das die Zellteilung hemmt (im Chemikalienhandel
Btofogze tn unserer Zeit / 13. jahrg. 1983 / Nr. 3 0 Verlag Chemte GmbH, 0-6940 Wetnheim, 1983 0045-20JX/83/0306-0087 $02.50/0
87
88 Biologie in unserer Zeit / 13. Jahrg. 1983 / Nr. 3

erhaltlich). Als Losungsmittel dient Aqua- rienwasser, Stammlosungen sollte man je- doch mit Aqua dest. ansetzen. Die Eier mehrerer Entwicklungsstadien werden ver- schiedenen Konzentrationen dieser Stoffe ausgesetzt. Die hier empfohlenen Konzen- trationen (siehe Abbildung 9) konnen nur als Richtwerte dienen, weil Reaktionsschwelle und Optimalkonzentration bei den Eiern verschiedener Weibchen voneinander abwei- chen. Aus dem gleichen Grunde sollte man zum Vergleich verschiedener Stadien mog- lichst Eier desselben Weibchens oder sogar desselben Geleges heranziehen. Nach Ablauf der Behandlungszeit von 24 h bzw. 1-2 h werden die Eier durch mehrfachen Wasser- wechsel gewaschen und in Aquarienwasser bei 26°C aufgezogen. Unbehandelte Eier des gleichen Geleges bewahrt man unter sonst gleichen Bedingungen als Kontrollansatz auf, der etwaige Anomalien des Geleges erkennen laflt. Zur Eingrenzung der sensiblen Phase waren kiirzere Behandlungszeiten mit Etha- nol erwiinscht. Wegen der erforderlichen ho- heren Ethanolkonzentration geben sie jedoch weniger zuverlassige Ergebnisse als die vor- geschlagene 24stiindige Behandlung.
Zur Beobachtung benutzt man die schwache Mikroskopvergroflerung oder ein Stereomi- kroskop, notfalls langt auch eine starke Lu- pe. Versuchs- und Kontrolleier werden fi- xiert, ehe der Korper sich pigmentiert, am besten aber nach Pigmentierung der Augen- becher. Diese erfolgt nach etwa 1 1/2 Tagen, und zu diesem Zeitpunkt sollte man auch die weniger weit entwickelten Eier fixieren, die andernfalls unter Zerfallserscheinungen ab- sterben. Als Fixiermittel benutzt man Etha-
no1/40% Formaldehyd/Eisessig/H20 im Ver- haltnis 16 : 6 : 1 : 30. Kein Wasser mit den Eiern eintragen, sonst quellen sie. Fixierte Embryonen vor der Untersuchung wassern!
3. Der Ablauf der Embryonalentwicklung
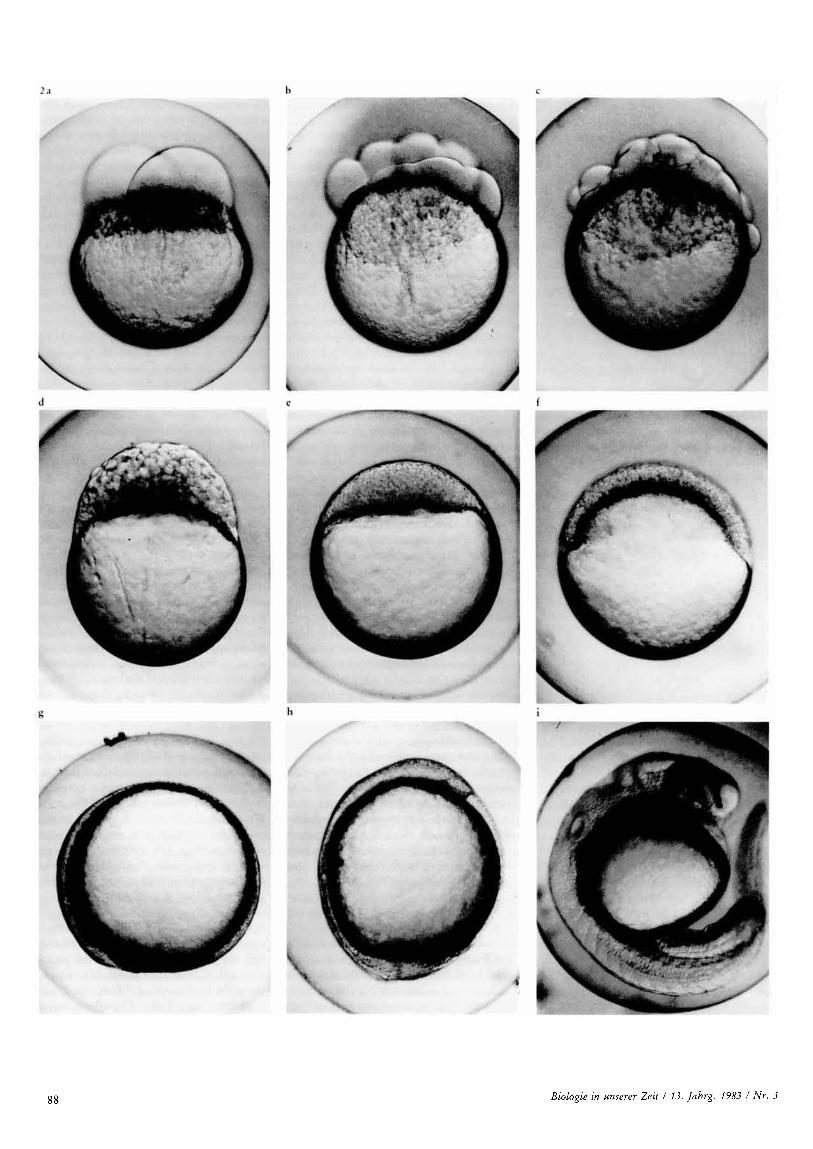
Die Embryonalentwicklung, die bei 26°C et- wa 4 Tage dauert, unterteilt man in eine An- zahl von Stadien (Abbildung 2 a-i) [5]. Sie beginnt mit einer typischen Discoidalfur- chung (Abbildung 2 a-d) [TI]. Die im Ver- gleich zu anderen Knochenfischen sehr gro- i3en Furchungszellen unterteilen sich zuneh- mend, bis tausende kleiner Zellen als kuppel- formige Keimscheibe auf dem verbleibenden, mit Dotter gefiillten Rest der Eizelle sitzen (Abbildung 2 e, f). Die Keimscheibe hat eine aui3ere Deckschicht aus flachen Zellen (Deckepithel), basal ist sie durch eine plas- mareiche, mit polyploiden Zellkernen durch- setzte Grenzschicht vom Dotter geschieden, die man als Periblast bezeichnet. Zwischen beiden Schichten sind in groi3er Zahl die rundlichen Innenzellen angehauft (auch "deep cells" genannt); aus diesen Zellen ent- stehen spater alle Organe des Embryos und vielleicht sogar Teile seiner Auflenflache. Die Keimscheibe wird bald dunner und dehnt sich ringsum mit gleicher Geschwindigkeit aus, bis sie die Dotterkugel vollig umschlieflt (Abbildung 2 e-g). Diese Gestaltungsbewe- gung beruht sowohl auf Zellteilungen als auch auf Zellverlagerung; man nennt sie Epi- bolie. Nachdem der Keimscheibenrand den Aquator der Dotterkugel iiberschritten hat, verdickt er sich zum Randring. Dieser weitet sich dann an einer Stelle zum Keimschild aus, der sich in Richtung auf das Zentrum der
Biologie in unserer Zeit / 13. Jahrg. 1983 1 Nr. 3 89
Keimscheibe streckt und damit die Langsach- se des zukunftigen Embryos markiert. Am Vorderende des Keimschilds entsteht aus den polnahen Innenzellen die Vorderkopfanlage. Die Rumpfanlage wachst gegenlaufig aus, und zwar in dem Mafle, wie der Randring bei der Epibolie zum unteren Pol hin fortschrei- tet (Abbildung 2 f-h). Der vom Randring umschlossene Bezirk entspricht dem Ur- mund, und die Epibolie ist eine stark abge- wandelte Form der Gastrulation. 1st sie voll- endet und der Urmund verschlossen, so er- scheint am unteren Pol eine Knospe (Abbil- dung 2 h), aus welcher der zukunftige Schwanzabschnitt des Fischkorpers hervor- geht; sie entsteht aus dem Hinterende des Keimschildes, also am Dovsalrand des soeben verschlossenen Urmunds. Am Kopf erschei- nen spater die Augenanlagen mit ihren kugel- formigen Linsen, und dahinter werden jeder- seits die schusselformigen Anlagen fur die Sinnesorgane des Innenohres sichtbar (Ab- bildung 2 i). In dem Mafle, wie sich der Kor- per des Embryos durch Zusammenstromen von Innenzellen herausmodelliert, wird der verbleibende Anteil der Keimscheibe ausge- dunnt. Er bildet die Anlage des Dottersacks. In seinen Wandschichten formt sich vor der Kopfanlage das Herz (Abbildung 2 i) und be- ginnt bereits zu schlagen, ehe es nach hinten unter den Kopf rutscht. Schon vor der Aus- bildung des Blutkreislaufs auf dem Dotter- sack und im Korper beginnt die Schwanzan- lage mit den sogenannten Pendelbewegun- gen. Spater sieht man die groflen Blutkorper- chen im Rhythmus des Herzschlags durch die Gefafle schieflen. Wenn das von Pigment- zellen gesprenkelte Fischlein sich schliefllich aus der Eihulle schnellt, ist es eigentlich noch eine Larve: sein Maul ist noch nicht zur Nah- rungsaufnahme geeignet, und es lebt noch ei- ne Weile von seinen Dottervorraten.
Zwei Eigenarten der Knochenfisch-Entwick- lung seien im Hinblick auf unsere Experi- mente besonders hervorgehoben, die ,,Kon- vergenzbewegung" der Innenzellen und das Prinzip der ,,Massiventwicklung", dem die Zellen dann beim Aufbau der Organe folgen. Bei der Epibolie breiten sich die Innenzellen zunachst radial, also gleichsam entlang den Meridianen der Polkappe aus. Spater wei- chen die meisten aber von ihrem jeweiligen Meridian ab, und zwar in Richtung auf den Keimschild und damit von beiden Seiten zur Mittellinie des Embryos hin (Abbildung 3 a). Dank dieser Konvergenz sammeln sie sich entlang der zukunftigen Korperachse an, ent- weder auf direktem Wege oder auf dem Um-
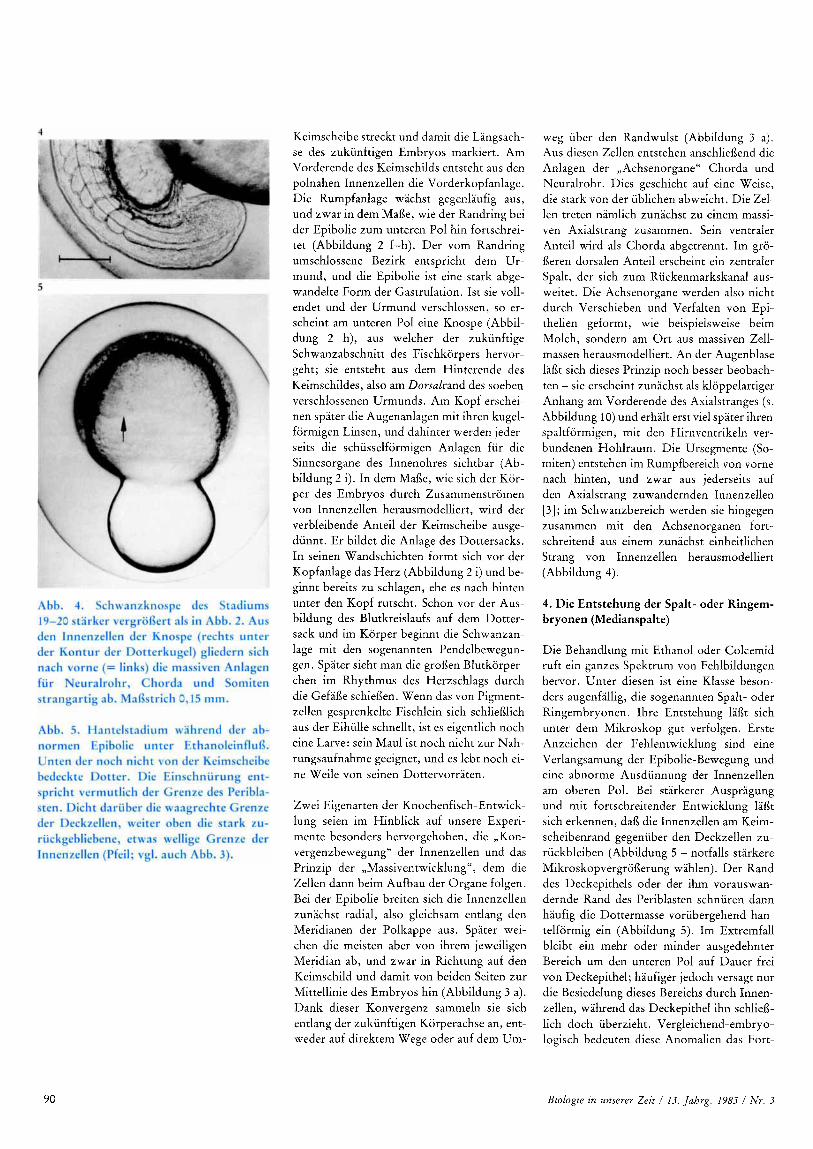
weg uber den Randwulst (Abbildung 3 a). Aus diesen Zellen entstehen anschlieflend die Anlagen der ,,Achsenorgane" Chorda und Neuralrohr. Dies geschieht auf eine Weise, die stark von der ublichen abweicht. Die Zel- len treten namlich zunachst zu einem massi- ven Axialstrang zusammen. Sein ventraler Anteil wird als Chorda abgetrennt. Im gro- fleren dorsalen Anteil erscheint ein zentraler Spalt, der sich zum Ruckenmarkskanal aus- weitet. Die Achsenorgane werden also nicht durch Verschieben und Verfalten von Epi- thelien geformt, wie beispielsweise beim Molch, sondern am O r t aus massiven Zell- massen herausmodelliert. An der Augenblase laflt sich dieses Prinzip noch besser beobach- ten - sie erscheint zunachst als kloppelartiger Anhang am Vorderende des Axialstranges (s. Abbildung 10) und erhalt erst vie1 spater ihren spaltformigen, rnit den Hirnventrikeln ver- bundenen Hohlraum. Die Ursegmente (So- miten) entstehen im Rumpfbereich von vorne nach hinten, und zwar aus jederseits auf den Axialstrang zuwandernden Innenzellen [ 3 ] ; im Schwanzbereich werden sie hingegen zusammen rnit den Achsenorganen fort- schreitend aus einem zunachst einheitlichen Strang von Innenzellen herausmodelliert (Abbildung 4).
4. Die Entstehung der Spalt- oder Ringem- bryonen (Medianspalte)
Die Behandlung rnit Ethanol oder Colcemid ruft ein ganzes Spektrum von Fehlbildungen hervor. Unter diesen ist eine Klasse beson- ders augenfallig, die sogenannten Spalt- oder Ringembryonen. Ihre Entstehung laflt sich unter dem Mikroskop gut verfolgen. Erste Anzeichen der Fehlentwicklung sind eine Verlangsamung der Epibolie-Bewegung und eine abnorme Ausdunnung der Innenzellen am oberen Pol. Bei starkerer Auspragung und mit fortschreitender Entwicklung lafit sich erkennen, dafi die Innenzellen am Keim- scheibenrand gegenuber den Deckzellen zu- ruckbleiben (Abbildung 5 - notfalls starkere Mikroskopvergroflerung wahlen). Der Rand des Deckepithels oder der ihm vorauswan- dernde Rand des Periblasten schnuren dann haufig die Dottermasse vorubergehend han- telformig ein (Abbildung 5). Im Extremfall bleibt ein mehr oder minder ausgedehnter Bereich um den unteren Pol auf Dauer frei von Deckepithel; haufiger jedoch versagt nur die Besiedelung dieses Bereichs durch Innen- zellen, wahrend das Deckepithel ihn schliefi- lich doch uberzieht. Vergleichend-embryo- logisch bedeuten diese Anomalien das Fort-
90 Biologie in unserer Zeit / 13. Jahrg. 1983 / Nr. 3
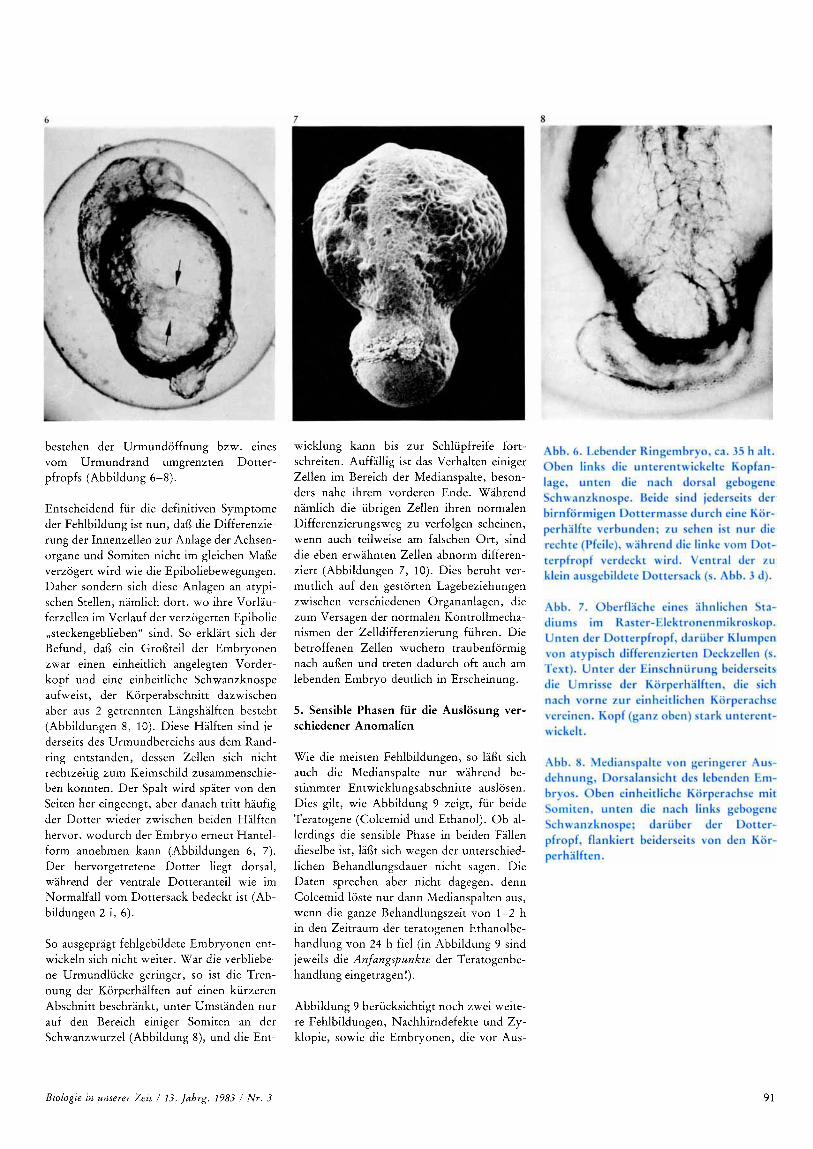
bestehen der Urmundoffnung bzw. eines vom Urmundrand umgrenzten Dotter- pfropfs (Abbildung 6-8).
Entscheidend fur die definitiven Symptome der Fehlbildung ist nun, dai3 die Differenzie- rung der Innenzellen zur Anlage der Achsen- organe und Somiten nicht im gleichen Mafie verzogert wird wie die Epiboliebewegungen. Daher sondern sich diese Anlagen an atypi- schen Stellen, namlich dort, wo ihre Vorlau- ferzellen im Verlauf der verzogerten Epibolie ,,steckengeblieben" sind. So erklart sich der Befund, dai3 ein Groi3teil der Embryonen zwar einen einheitlich angelegten Vorder- kopf und eine einheitliche Schwanzknospe aufweist, der Korperabschnitt dazwischen aber aus 2 getrennten Langshalften besteht (Abbildungen 8, 10). Diese Halften sind je- derseits des Urmundbereichs aus dem Rand- ring entstanden, dessen Zellen sich nicht rechtzeitig zum Keimschild zusammenschie- ben konnten. Der Spalt wird spater von den Seiten her eingeengt, aber danach tritt haufig der Dotter wieder zwischen beiden Halften hervor, wodurch der Embryo erneut Hantel- form annehmen kann (Abbildungen 6, 7). Der hervorgetretene Dotter liegt dorsal, wahrend der ventrale Dotteranteil wie im Normalfall vom Dottersack bedeckt ist (Ab- bildungen 2 i, 6).
So ausgepragt fehlgebildete Embryonen ent- wickeln sich nicht weiter. War die verbliebe- ne Urmundlucke geringer, so ist die Tren- nung der Korperhalften auf einen kurzeren Abschnitt beschrankt, unter Umstanden nur auf den Bereich einiger Somiten an der Schwanzwurzel (Abbildung 8), und die Ent-
wicklung kann bis zur Schlupfreife fort- schreiten. Auffallig ist das Verhalten einiger Zellen im Bereich der Medianspalte, beson- ders nahe ihrem vorderen Ende. Wahrend namlich die ubrigen Zellen ihren normalen Differenzierungsweg zu verfolgen scheinen, wenn auch teilweise am falschen Or t , sind die eben erwahnten Zellen abnorm differen- ziert (Abbildungen 7, 10). Dies beruht ver- mutlich auf den gestorten Lagebeziehungen zwischen verschiedenen Organanlagen, die zum Versagen der normalen Kontrollmecha- nismen der Zelldifferenzierung fuhren. Die betroffenen Zellen wuchern traubenformig nach aui3en und treten dadurch oft auch am lebenden Embryo deutlich in Erscheinung.
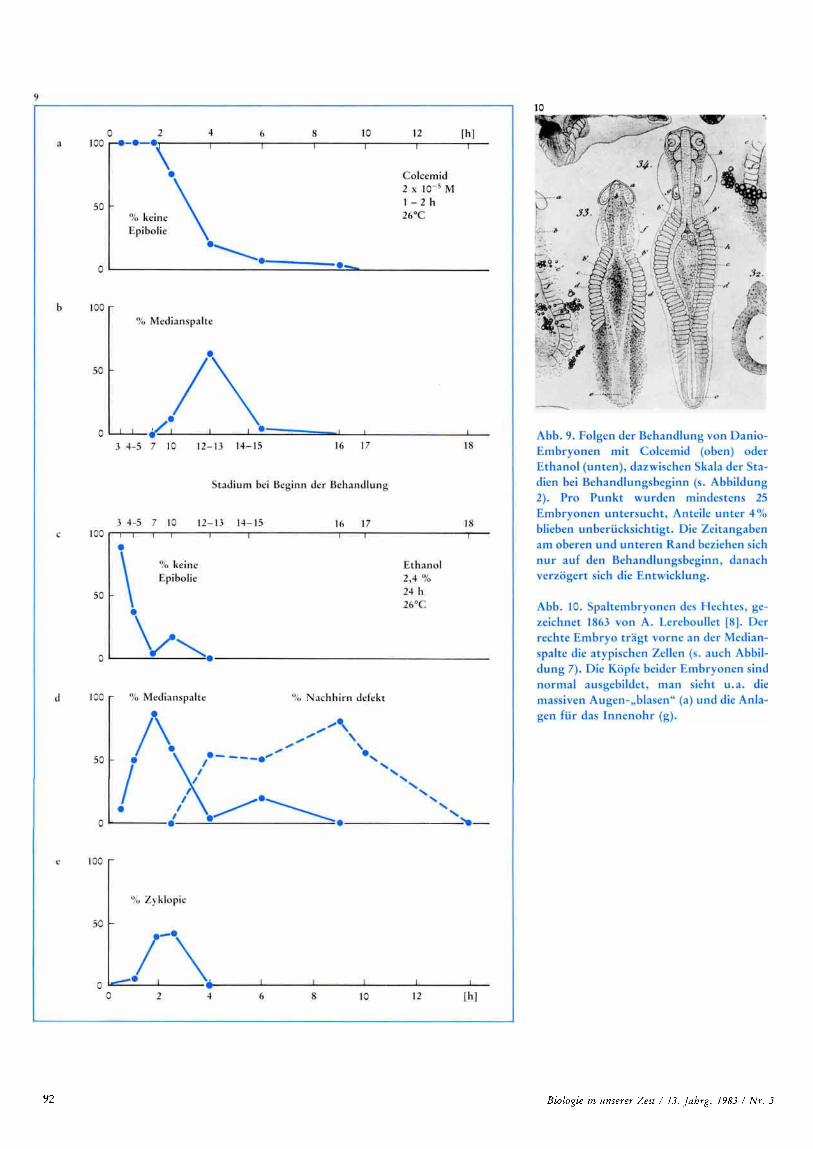
5. Sensible Phasen fur die Auslosung ver- schiedener Anomalien
Wie die meisten Fehlbildungen, so Iai3t sich auch die Medianspalte nur wahrend be- stimmter Entwicklungsabschnitte auslosen. Dies gilt, wie Abbildung 9 zeigt, fur beide Teratogene (Colcemid und Ethanol). O b al- lerdings die sensible Phase in beiden Fallen dieselbe ist, lai3t sich wegen der unterschied- lichen Behandlungsdauer nicht sagen. Die Daten sprechen aber nicht dagegen, denn Colcemid loste nur dann Medianspalten aus, wenn die ganze Behandlungszeit von 1-2 h in den Zeitraum der teratogenen Ethanolbe- handlung von 24 h fie1 (in Abbildung 9 sind jeweils die Anfungspunkte der Teratogenbe- handlung eingetragen!) .
Abbildung 9 berucksichtigt noch zwei weite- re Fehlbildungen, Nachhirndefekte und Zy- klopie, sowie die Embryonen, die vor Aus-
Biologie in unserer Zeit / 13. jahrg. 1983 / Nr. 3 91
92 Btologte in unserer Zett / 13. Jahrg. I983 / Nr. 3

bildung deutlicher Achsenorgane abstarben. Die Zyklopie hat ihren Namen aus der grie- chischen Mythologie. Wie der von Odysseus uberlistete Zyklop Polyphemos haben auch die zyklopischen Fischembryonen nur ein Auge, und das sitzt in der Mit te lhie des Kopfes. Ubergangsstufen der Fehlbildung belegen, dafl das Mittelauge aus der Ver- schmelzung der Augenanlagen beider Kor- perseiten entsteht; man sieht dies beim Auf- treten des Augenpigments sehr deutlich. Die Nachhirndefekte, die nur an fixierten Em- bryonen mit Sicherheit zu erkennen sind, be- stehen in einer Verbreiterung der Anlage des Neuralrohres in dieser Region. Dabei tritt ei- ne mediane Einsenkung der Dorsalwand auf, die eine unvollstandige Medianspaltung vor- tauscht.
Die sensible Phase fur die Auslosung von Nachhirn-Anomalien unterscheidet sich deutlich von derjenigen fur Medianspalten, zumindest in Bezug auf den Beginn der tera- togenen Ethanolbehandlung (Abbildung 9 d). Zwischen beiden besteht ein weiterer we- sentlicher Unterschied: die sensible Phase fur Medianspalten, gleich ob Colcemid- oder Ethanol-bedingt (Abbildung 9 b, d), wird zu jungeren Stadien hin durch die Mortalitat be- grenzt (Abbildung 9 a, c), die sensible Phase fur die Nachhirndefekte hingegen durch eine weitere Fehlbildung, die zeitlich fruher aus- losbare Medianspalte; beginnt die Teratogen- behandlung in der Obergangszeit, so entwik- keln einzelne Embryonen beide Anomalien. Medianspalte und Zyklopie belegen eine Re- gel, die fur die menschliche Fehlbildungsleh- re sehr wichtig ist: Teratogen-Dosen, die in der sensiblen Phase zu spezifischen Fehlent- wicklungen fiihren, wirken auf jungere Sta- dien in kurzer Frist todlich, wahrend sie bei Einwirkung nach Ablauf der sensiblen Phase ohne dauernde Folgen bleiben [13, 141.
6. Schluflbetrachtungen
Der Spalt zwischen rechter und linker Kor- perhalte ist die auffalligste der in unseren Versuchen ausgelosten Fehlbildungen. Die verlangsamte Epibolie und der zu lange fort- bestehende Urmund verhindern, dai3 sich die Vorlauferzellen der beiden Korperhalften rechtzeitig in der Mittellinie treffen. Dieses Versagen konnte darauf beruhen, dai3 die be- nutzten Zellgifte die Zellwanderung direkt hemmen. Wahrscheinlicher ist, daB die In- nenzellen sich weniger haufig teilen und da- her die Zellmenge nicht zur volligen Umhul- lung der Dotterkugel ausreicht. Die Zyklopie
laflt sich sicher als Folge der zu geringen Zell- zahlen im Bereich der Kopfanlage verstehen; die Ausdunnung der Zellschicht am oberen Pol ist ja das erste sichtbare Symptom nach Ethanolbehandlung. Bei Molchen kann man Zyklopie durch operative Verkleinerung der Kopfanlage auslosen [9], und als menschliche Fehlbildung wurde sie nach Behandlung von krebskranken Schwangeren mit Zellteilungs- hemmern beobachtet.
Die allgemeinen Entwicklungsanomalien, aus denen die gespaltene Anlage des Danio- Embryos in unseren Versuchen folgt, sind die verringerte Zellvermehrung und die Hemmung bestimmter Gestaltungsbewegun- gen. Die gleichen allgemeinen Anomalien werden fur zahlreiche Fehlbildungen beim Menschen verantwortlich gemacht. Direkte oder indirekte Hemmung der Zellvermeh- rung durfte sowohl bei der Unterentwick- lung von GliedmaBen (Contergan!) als auch, wie oben erwahnt, bei der Entstehung der Zyklopie eine Rolle spielen. Unzureichende Gestaltungsbewegungen und ihre Folgepro- zesse sind die haufigste Ursache fur mangeln- den Verschlui3 des Neuralrohres oder der Neuralbogen einiger Ruckenwirbel (Spina bifida) .
Es sei hier aber betont, dafl diese Gemein- samkeiten zwischen Fisch und Mensch nur die generellen Prinzipien betreffen und nicht die Einzelheiten der Fehlentwicklung oder ihrer Auslosung. Sie mussen beim Saugerem- bryo schon wegen seiner Abhangigkeit vom Stoffwechsel der Mutter in vielerlei Hinsicht verschieden sein. Vor allem aber weichen die Fruhstadien der Knochenfischentwicklung extrem von denen der Sauger ab [lo]. Daher umfaflt das ,,fetale Alkoholsyndrom" von Maus und Mensch vorwiegend andere Fehl- bildungen [ 11 ; eine Gemeinsamkeit konnte man am ehesten zwischen der Entwicklungs- verzogerung beim Danio und dem drastisch reduzierten Geburtsgewicht beim Menschen sehen.
Werfen wir abschlieflend einen Blick auf Fehlbildungen bei anderen Chordaten, die ebenfalls auf raumlich getrennter Differen- zierung der Korperhalften beruhen. Sie sind schon vom niedersten Chordatier bekannt, namlich vom Lanzettfischchen Branchiosto- ma (Amphioxus). Die fruhesten Beschreibun- gen stammen von Knochenfisch-Embryonen (Abbildung 10); im Gegensatz zu manchen spateren Arbeiten wird hier der Zusammen- hang zwischen gestorter Epibolie und Me-
dianspalte deutlich erkannt [8]. Die ersten Beschreiber von Ringembryonen bei Amphi- bien, unter ihnen der ,,Vater der Entwick- lungsphysiologie", Wilhelm Roux, sahen den Zusammenhang zwischen abnormer Gastru- lation und Fehlbildung sehr deutlich [4, 121 (Abbildung 11). Dafl Roux aus dem Entwick- lungsverlauf einen falschen Schlufl zog, ist nicht nur aus historischen Griinden interessant. Wir wurden namlich seinen Fehler wiederho- len, wenn wir annahmen, dai3 die Korper- halften des Danio-Embryos auch im norma- len Entwicklungsablauf beiderseits des Ur- munds angelegt wurden - sie entstehen aber dorsal vom Urmund im Keimschild, wie oben beschrieben (Abbildung 3).
O b axiale Spalten des gleichen Ursprungs auch beim Menschen vorkommen, mufl mangels Beobachtungsmoglichkeit wahrelid des Entwicklungsablaufs offen bleiben. Zur Erklarung der sehr seltenen volligen Wirbel- spaltung wurde schon recht fruh ein Versa- gen der Gastrulation herangezogen (Literatur siehe [15]), und diese Erklarung hat durch neue Befunde uber die Fruhentwicklung der Labormaus an Wahrscheinlichkeit gewon- nen. Bei der Maus stammen namlich die zu- kunftigen Darmzellen aus der Auflenschicht des Embryonalschildes [2] und nicht aus dem Entoderm des Dottersacks (siehe 2.B. [13]), konnen also im Gegensatz zur gangigen
Biologie in unserer Zeit / 13. Jahrg. 1983 / Nr. 3 93

BioFocus
Lehrbuchmeinung nur durch einen ausge- pragten Gastrulationsvorgang auf die Unter- seite (Dottersackseite) des Keimschildes gelangen, wo der Darm entsteht; versagt die Gastrulation, konnten die zukunftigen Darm- zellen als Hindernis in der Mitte der Korper- anlage liegen bleiben.
Als alternative Erklarung fur die regionale Spaltung der Achsenorgane bietet sich die Annahme an, dai3 eine einheitliche Korperan- lage sekundar entlang der Mi t te lh ie aufge- rissen sei. Erklarungen dieser Art wurden nicht nur fur die menschliche Fehlbildung, sondern auch mehrfach fur die Spaltem- bryonen der Knochenfische vorgeschlagen 161). Die Uberprufung dieser Hypothese fuhrte uns zu den hier geschilderten Experi- menten und Beobachtungen.
Literatur
[1] Chernuff, G . F . : The fetal alcohol syn- drome in mice: an animal model. Teratology 15,223-230 (1977).
[2] Gardner, R. L., Rossant, J.: Investig- ation of the fate of 4-5 day post-coitum mouse inner cell mass cells by blastocyst injection. J. Embryol. Exp. Morphol. 52, 141-152 (1979).
[3] Heinig, S.: Epiplatys dageti (Cyprino- dontidae): Embryonalentwicklung, Ent- wicklung der Keimblatter, Organdifferenzie- rung. Film E 2149, Institut fur den Wissen- schaftlichen Film, Gottingen 1975.
[4] Hertwig, 0.: Urmund und Spina bifida. Arch. Mikrosk. Anat. Entwicklungsmech. 39, 353-524 (1892).
[8] Lereboullrt, A.: Kecherches sur les mon- struosites du brochet observees dans I'oeuf et sur leur mode de production. Ann. nat. Ser. 4 ZOO^.) 20, 177-271 (1863).
[9] Mangold, 0.: Hans Spemann, ein Mei- ster der Entwicklungsphysiologie. Sein Le- ben und sein Werk. Wissenschaftl. Verlags- gesellschaft, Stuttgart 1953.
[lo] Ortmann, R. : Gastrula and gastrulation in vertebrates. In: Ontogenese und Phyloge- nese (Hrsg. R. Siewing), Beihefte Z. zool. Systematik und Evolutionsforschung 1, 57-72 (1979).
1111 Roosen-Runge, E. C.: Enibryonalent- wicklung von Brachydanio rerio, Film C 42, Institut fur den Wissenschaftlichen Film Gottingen, 1936.
1121 Roux, W.: Uber die Lagerung des Ma- terials des Medullarrohres im gefurchten Froschei. Anat. Anz. 3, 697-705 (1888).
[13] Schuhmacher, G . H . : Embryonale Ent- wicklung des Menschen (Taschenbuch). Gu- stav Fischer Verlag, Stuttgart 1975.
[14] Stengel, H.: Die Ursachen menschlicher Fehlbildungen. Naturw. Rundschau 27, 306-312 (1974).
[15] Theiler, K.: Beitrag zur Analyse von Wirbelkorperfehlbildungen: Experimente, Genetik und Entwicklung. 2. menschl. Ver- erbungs- und Konstitutionslehre 31, 271-322 (1953).
[16] Unterrichtsfilm FT 875 ,,Entwicklung eines Fischembryos"; Inst. f . Film u. Bild in Wissenschaft und Unterricht (FWU) Grun- waldlMunchen, oder Kreisbildstellen.
Klaus Sander. Martin Baumann [5] Hisaoka, H . K., Battle, H. I.: The nor- mal developmental stages of the zebrafish Bra- chydanio rerio (Hamilton-Buchanan). J. Morphol. 102, 311-326 (1958).
[6] Laale, H. W.: Ethanol induced noto- chord and spinal chord duplications in the embryo of the zebrafish (Brachydanio rerio). J. Exp. Zool. 177, 51-64 (1971).
Anschrift: [7] Laale, H . W.: The biology and use of zebrafisch, Brachydanio rerio, in fisheries research. A literature review. J. Fish Biol. 10, 121-173 (1977).
Prof. Dr. K . Sander, M. Baumann, Institut fur Biologie I (Zoologie) der Universitat, Al- bertstrai3e 21 a, D-7800 Freiburg i. Br.
Gezielte Erbanderungen bei Saugerzellen
In den letzten Jahren hat die Gentechnologie rapide Fortschritte gemacht. Heute stehen verlai3liche Methoden fur die Isolierung von bestimmten Genen und ihre beliebige Ver- mehrung in prokaryotischen Mikroorganis- men - das ,,Klonieren" - zur Verfugung. Die Bedeutung dieser neuen Methoden fur die biologische Grundlagenforschung ist schon jetzt gewaltig; das biologische Weltbild ist wieder einmal dabei, sich kraftig zu veran- dern. Andererseits hat sich aber die Massen- herstellung von medizinisch oder sonstwie bedeutsamen Genprodukten als unerwartet schwierig erwiesen. Das hangt letztlich zu- sammen mit den bei Prokaryoten und Euka- ryoten eben doch recht unterschiedlichen Sy- stemen fur die Regulation von Genaktivita- ten und die Erzeugung der endgultigen Gen- produkte; schliefllich ist auch die bloi3e Wiedereinfuhrung klonierter Gene in ent- sprechende Empfangerzellen keine einfache Sache. Daher ist beispielsweise die Heilung von Erbkrankheiten durch gezielte geneti- sche Manipulation von Stammzellen oder Keimzellen vorerst ein Wunschtraum geblie- ben, und entsprechendes gilt etwa auch fur die Moglichkeit, Kulturpflanzen durch den Einbau von Bakterien-Genen zur Stickstoff- Assimilation zu befahigen und damit auf- wendige Dungung mindestens teilweise uberflussig zu machen. So ist es verstandlich, dai3 die Kultivierungstechniken fur Sauger- und Human-Zellen in den letzten Jahren eine Renaissance erlebten und dai3 man versucht, auch diese Zellen genetisch in ahnlicher Wei- se zu manipulieren, wie das heute bei Bakte- rien durch die Plasmid- und Phagen-Techni- ken moglich ist. Fur die Grundlagenfor- schung sind solche Untersuchungen in jedem Fall von groatem Interesse. Wer etwa Aus- kunft uber die komplexen Regulationsvor- gange in Eukaryoten-Zellen (Eucyten) erhal- ten will, ist nun einmal auf die Untersuchung eben dieser Zellen angewiesen. Wie ist der Stand der Forschung und Technik in diesem Bereich?
Im BioFocus des letzten Heftes wurde davon berichtet, dai3 im Zuge der phylogenetischen Entwicklung zwischen den DNA-haltigen Kompartimenten von Eucyten Verlagerun- gen bestimmter DNA-Sequenzen vorgekom- men sind. Bei der genetischen Manipulation von Eucyten geht es darum, Gentransfer kunstlich und gezielt - gewohnlich zwischen verschiedenen Zellen - vorzunehmen. Die- sem Ziel dienten schon die altesten Zuch-
94 Btologae irr unserer Zeat / 13. Jahrg. 1983 / Nr. 3