Embed Size (px)
Citation preview

Aus der Universitätsklinik und Poliklinik für Innere Medizin III
des Universitätsklinikums Halle (Saale)
(Direktor: Prof. Dr. med. Stefan Frantz)
Der Ein�uss von Endotoxin auf die Herzfrequenz und
Herzfrequenzvariabilität bei Wildtyp-Mäusen � ein Modell der
kardialen autonomen Dysfunktion bei Intensivpatienten mit
Multiorgan-Dysfunktions-Syndrom (MODS)
Dissertation
zur Erlangung des akademischen Grades
Doktor der Medizin (Dr. med.)
Vorgelegt
der Medizinischen Fakultät
der Martin-Luther-Universität Halle-Wittenberg
von Sara Ruccius, geborene Hettrich,
geboren am 08.10.1984 in Berlin
Betreuer: APL. Prof. Dr. med. Henning Ebelt
Gutachter/Gutachterin:
APL. Prof. Dr. med. Henning Ebelt (Erfurt)
Prof. Dr. phil. habil. Kuno Hottenrott
PD Dr. med. Juliane Stieber (Erlangen)
03.11.2015
09.11.2016

Referat
Eine Einschränkung der Herzfrequenzvariabilität (HRV) ist bei Intensivpatienten mit ei-
nem Multiorgan-Dysfunktions-Syndrom (MODS) prognostisch ungünstig. Die Rolle des
Endotoxins bei der Entstehung dieser HRV-Einengung wurde mittels fortlaufender tele-
metrischer EKG- und Aktivitätsregistrierung durch intraperitoneal platzierte Biopotenti-
altransmitter bei adulten männlichen ICR-Wildtyp-Mäusen evaluiert, nachdem zunächst
die Einschätzung der Stabilisierung nach operativer Transmitterimplantation und der
autonomen Regulation der HRV bei den Tieren erfolgte.
Bei Wildtyp-Mäusen ist die Ruheherzfrequenz unmittelbar postoperativ erhöht, die
HRV und die Zirkadianität sind eingeschränkt. Während sich die Ruheherzfrequenz nach
5 Tagen normalisiert, benötigt die HRV-Stabilisierung mindestens 10 Tage. Atropin- und
Isoproterenolapplikation bewirken eine ausgeprägte Tachykardisierung. Propranololappli-
kation verursacht eine Bradykardie. Atropin reduziert alle und Propranolol keinen der
untersuchten HRV-Parameter. Eine autonome Blockade beein�usst weder die Herzfre-
quenz noch die HRV signi�kant. Isoproterenol senkt die Standardabweichung der nor-
malen RR-Intervalle (SDNN) und führt zu einer Reduktion der Low Freuqency/High
Frequency-Ratio (LF/HF-Ratio). Propranolol hebt die LF/HF-Ratio an. Endotoxin führt
zu einem signi�kanten Anstieg der Herzfrequenz und unter einer autonomen Blockade zu
einer Bradykardisierung. Es kommt zu einer hochgradig eingeschränkten HRV, vor allem
bei alleiniger Endotoxin-Applikation.
Die Experimente weisen auf einen postoperativen Stabilisierungszeitraum von minde-
stens 10 Tagen und auf einen ausgeglichenen autonomen Ruhetonus bei Wildtyp-Mäusen
hin. Der Parasympathikus ist entscheidend an der Entstehung der HRV beteiligt. Die
in Zellkulturexperimenten beschriebene direkte Hemmung des kardialen Schrittmacher-
stroms If und dessen Sympathikus-Sensibilisierung durch Endotoxin werden in den hier
vorgestellten in-vivo-Experimenten bei ICR-Wildtyp-Mäusen funktionell bestätigt. Die
Befunde unterstützen die Hypothese der Interaktion von Endotoxin mit den kardialen
Schrittmacherzellen und stellen deren zentralen Stellenwert bei der Entstehung der pro-
gnoserelevanten autonomen kardialen Dysfunktion bei Patienten mit MODS zur Diskus-
sion.
Ruccius, Sara: Der Ein�uss von Endotoxin auf die Herzfrequenz und Herzfrequenzvaria-bilität bei Wildtyp-Mäusen � ein Modell der kardialen autonomen Dysfunktion bei In-tensivpatienten mit Multiorgan-Dysfunktions-Syndrom (MODS). Halle (Saale), Martin-Luther-Univ., Med. Fak., Diss., 71 Seiten,12.08.2015.

I
Inhaltsverzeichnis
1 Einleitung 1
2 Zielstellung 7
3 Material und Methodik 8
3.1 Versuchstiere und Tierhaltung . . . . . . . . . . . . . . . . . . . . . . . . . 8
3.2 Implantation der Telemetrie-Transmitter . . . . . . . . . . . . . . . . . . . 8
3.3 Postoperative Beobachtung, pharmakologische Beein�ussung des
autonomen Nervensystems und Endotoxinapplikation . . . . . . . . . . . . 10
3.4 Datengewinnung und Datenaufzeichnung . . . . . . . . . . . . . . . . . . . 12
3.5 Datenverarbeitung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
4 Ergebnisse 21
4.1 Stabilisierung und Eintreten einer zirkadianen Rhythmik nach operativer
Transmitterimplantation . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
4.2 Autonome Regulation der Ruheherzfrequenz und der
Herzfrequenzvariabilität . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
4.3 Ein�uss von Endotoxin auf die Ruheherzfrequenz und die
Herzfrequenzvariabilität . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
5 Diskussion 34
5.1 Telemetrische Datengewinnung an Mäusen . . . . . . . . . . . . . . . . . . 34
5.2 Herzfrequenzvariabilität in Mensch und Maus � Rückschlüsse auf die Re-
gulation des autonomen Nervensystems . . . . . . . . . . . . . . . . . . . . 38
5.3 Die Einschränkung der Herzfrequenzvariabilität bei Sepsis und MODS . . 45
5.4 Die Rolle von Endotoxin in der gestörten Interorgankommunikation . . . . 48
5.5 Studienlimitierung und Forschungsexposé . . . . . . . . . . . . . . . . . . 54
6 Zusammenfassung 56
7 Literaturverzeichnis 58
8 Thesen 70

II
Abkürzungsverzeichnis
APACHE Acute Physiology And Chronic Health Evaluation
APR-1 Ambient Pressure Reference Monitor
ASDNN mittlere Standardabweichung normaler RR-Intervalle alle 5 Minuten
B6 siehe C57BL/6
Balb/c Mausstamm (Bagg Albino Jackson Laboratory)
BP Blood Pressure, Blutdruck
bpm beats per minute, Einheit der Herzfrequenz in Schlage/min
C57BL/6 Mausstamm (Stamm C57 Black/Rasse 6)
CHF Congestive Heart Failure
CK/CK-MB Creatinkinase, Isoenzym MB der Creatinkinase, spezi�sch für den Herzmuskel
CLP Cecal Ligation and Puncture
D.m. Diabetes mellitus
FD Frequency Domain, beinhaltet die Parameter HF, LF und LF/HF-Ratio
FFT Fast Fourier Transformation
FVB Mausstamm (Friend Leukemia Virus B)
GDA Glutardialdehyd
HCN Hyperpolarization-activated Cyclic Nucleotide-gated Cation Channel
hHCN2 Human Hyperpolarization-activated Cyclic Nucleotide-gated Cation Channel 2
HF High Frequency
HRV Heart Rate Variability, Herzfrequenzvariabilität
IBI Inter Beat Interval, siehe RR-Intervall
ICR Mausstamm (Institute of Cancer Research, Synonym CD-1 R©)
If Strom, der durch den kardialen Schrittmacherkanal HCN �ieÿt
IL-1β Interleukin-1β
i.p. intraperitoneal
LD50 Letale Dosis; Dosis einer Substanz, die in 50 % der applizierten Fälle tödlich ist
LF Low Frequency
LF/HF-Ratio Verhältnis der LF Power zur HF Power
LPS Lipopolysaccharid, Endotoxin
meanAct3 mittlere motorische Aktivität der untersuchten Wildtyp-Mäuse in 3 Minuten
meanAct33 mittlere motorische Aktivität der untersuchten Wildtyp-Mäuse in dem

III
die meanAct3 umgebenden 30 Minuten
meanNN arithmetischer Mittelwert aller normalen RR-Intervalle eines
EKGs in einem de�nierten Zeitraum (Kehrwert der Herzfrequenz)
MI Myokardinfarkt
MODS Multiorgan-Dysfunktions-Syndrom
MSOF Multiple-System-Organ-Failure
MW± SD arithmetisches Mittel ± Standardabweichung
NMRI Mausstamm (Naval Medical Research Institute)
NN-Abstand siehe RR-Intervall
nu normalized units
PAN Peripheres Autonomes Nervensystem
PBS Phosphate Bu�ered Saline
pNN6 relativer Anteil aufeinander folgender normaler RR-Abstände,
die über 6 ms schwanken
pNN50 relativer Anteil aufeinander folgender normaler RR-Abstände,
die über 50 ms schwanken
PNP Periphere Neuropathie
RAAS Renin-Angiotensin-Aldosteron-System
RLA Receiver-Modell
RMSSD Root Mean Square of Successive Di�erences,
Quadratwurzel der quadrierten Di�erenzen aufeinanderfolgender RR-Intervalle
RR-Intervall Zeitintervall zwischen den R-Zacken zweier normaler, aufeinander folgender
QRS-Komplexe im EKG, synonym gebraucht zu NN-Abstand und IBI
SIRS Systemic In�ammatory Response Syndrome
SDNN Standardabweichung aller normalen RR-Intervalle eines EKGs
in einem de�nierten Zeitraum
SK Sinusknoten
TD Time Domain, beinhaltet die Parameter meanNN, SDNN, RMSSD und
pNN6
TISS Therapeutic Intervention Scoring System
TNFα Tumor Necrosis Factor α
VLF Very Low Frequency

1
1 Einleitung
Das aktuelle Konzept der Pathogenese eines Multiorgan-Dysfunktions-Syndroms (MODS)
einschlieÿlich der kardialen Beteiligung im Sinne einer septischen Kardiomyopathie zeigt
unterschiedliche Mechanismen auf, die zur Erklärung dieses Krankheitsgeschehens heran-
gezogen werden können (Abb. 1). Diesem Modell zufolge, und in den aktuellen Sepsisleitli-
nien der Deutschen Sepsis-Gesellschaft und der Deutschen Interdisziplinären Vereinigung
für Intensiv- und Notfallmedizin erneut betont, verursache ein infektiöser Stimulus als
Auslöser einer Sepsis eine systemische, pro- und antiin�ammatorische Kaskaden enthal-
tende Wirtsreaktion (Müller-Werdan et al., 2006a; Reinhart et al., 2010). Eine in diesem
Rahmen überschieÿende, unkontrollierte und ungerichtete entzündungszell- und media-
torvermittelte Immunreaktion (Systemic In�ammatory Response Syndrome, SIRS) könne
im Sinne eines horror autotoxicus zu einer Schädigung des Wirts selbst führen (Baue,
1992; Redl et al., 1993). Dabei komme es durch das auslösende Agens, beispielsweise
Endotoxin (Loppnow et al., 1993; Schletter et al., 1995), über eine Aktivierung von Gra-
nulozyten, Makrophagen bzw. Monozyten (van der Meer et al., 1988) und einer Reihe
weiterer Wirtszellen zu einer Freisetzung zahlreicher Mediatoren, die das laborchemisch
beschriebene Phänomen des cytokine storm erzeugen (Clark, 2007; Wang and Ma, 2008).
Eine groÿe Anzahl dieser Mediatoren konnte seit den 1980er-Jahren identi�ziert werden
(Redl et al., 1993; Waage et al., 1993). Die Aktivität proin�ammatorischer Zytokine
wie beispielsweise des Tumor Necrosis Factor α (TNFα) (Damas et al., 1989) und des
Interleukin-1β (IL-1β, Okusawa et al., 1988) korrelieren eng mit der Entstehung eines
Multiorgan-Dysfunktions-Syndroms. Die direkt kardiodepressive Wirkung dieser Media-
toren konnte 1996 (Kumar et al., 1996) und in der Folge in verschiedenen Arbeiten
nachgewiesen werden (Müller-Werdan et al., 1998; Cain et al., 1999).
In klinischen Studien der 1970er-Jahre wurde eine verringerte kardiale Auswur�ei-
stung bei erhöhtem peripheren Widerstand im Sinne einer Myokarddepression bei Sep-
sis interpretiert und diese vorranging als eine reaktive, kardiovaskuläre Beteiligung des
Herzens im Rahmen des septischen Geschehens verstanden (Shoemaker, 1971; Nishiji-
ma et al., 1973; Hinshaw, 1974). Auch eine initiale hyperdyname Kreislaufsituation mit
normaler oder erhöhter Auswur�eistung und erniedrigtem peripheren Widerstand wurde
beobachtet, diese jedoch als noch initial adäquate Beteiligung des Herzens bei Sepsis ein-
geordnet (Hinshaw, 1974). Margaret M. Parker wies eine eingeschränkte linksventrikuläre

2
Abb. 1: Aktuelles Konzept der Pathogenese eines Multiorgan-Dysfunktions-Syndroms (MODS),
Ma.=Makrophagen, Gr.=Granulozyten (Müller-Werdan et al., 2006a).
Pumpfunktion bei Sepsis 1984 szintigraphisch nach (Parker et al., 1984). Bei genauer Be-
trachtung der Relation von kardialer Auswur�eistung und peripherem Widerstand zeigte
sich in der Folge, dass die Pumpfunktion, auch wenn sie absolut zunächst normal oder
erhöht erscheint, im Verhältnis zu der dramatischen Nachlastreduktion nicht adäquat ge-
steigert ist (Müller-Werdan et al., 2006a). Die septische Kardiomyopathie wurde zunächst
als vorrangig �sekundäre� verstanden (Hinshaw, 1974, verursacht durch z. B. peripheres
Pooling, Myokardminderperfusion aufgrund von Hypotension oder tachykardiebedingte
Minderversorgung). Im weiteren wurde auch von einer direkten Schädigung des Myokards
durch zirkulierende Mediatoren wie beispielsweise Zytokine oder hohe Catecholaminkon-
zentrationen ausgegangen (Müller-Werdan et al., 1996).
Das Konzept der septischen Kardiomyopathie, welches die Schädigung des Herzens
als eine eigene Entität im Rahmen der Systemerkrankung Sepsis begreift, gewinnt erst
in jüngerer Zeit zunehmend an Bedeutung (Müller-Werdan et al., 2006b). Sie ist vor-
rangig eine klinische Diagnose, die durch verschiedene apparativ-diagnostische Befunde
gestellt werden kann (Müller-Werdan et al., 2006b). Bisher konnten keine charakteristi-
schen pathomorphologisch-histologischen Befunde der septischen Kardiomyopathie ge-
funden werden, die Schädigung des Herzens ist unspezi�sch (Müller-Höcker and Haerty,

3
1993). Über 24 Stunden mit Endotoxin oder Tumor Necrosis Factor α (TNFα) inku-
bierte Rattenkardiomyozyten zeigen ebenso wenig morphologische Veränderungen unter
dem Phasen-Kontrast-Mikroskop (Müller-Werdan et al., 1998), wie eine 6-stündige Li-
popolysaccharid (LPS)-Inkubation humaner Vorhofkardiomyozyten (Zorn-Pauly et al.,
2007). Auf Zellebene jedoch konnten in den letzten Jahren verschiedene funktionelle Be-
einträchtigungen der Kardiomyozyten im Rahmen einer Sepsis aufgezeigt werden, die
unter anderem die Induktion in�ammatorischer Kaskaden, die Beein�ussung des Ca2+-
Haushaltes, eine Mitochondriendysfunktion sowie die Modulierung autonomer Ein�üs-
se umfassen und die Pumpfunktionseinschränkung mit erklären könnten (Flierl et al.,
2008; Kneyber et al., 2009). Eine hochgradig eingeschränkte Herzfrequenzvariabilität
(HRV) stellt ein typisches, klinisches Charakteristikum der septischen Kardiomyopathie
dar (Müller-Werdan et al., 2006b), dessen zelluläre Entstehungsmechanismen bisher un-
geklärt sind.
Zum ersten Mal berichtete Christopher S. Garrard von einer autonomen Dysfunk-
tion, die sich in einer eingeschränkten Herzfrequenzvariabilität im Rahmen einer Sepsis
bei Intensivpatienten ausdrückte (Garrard et al., 1993). Verschiedene klinische Arbeiten
validierten in der Folge diese Beobachtung und konnten gleichzeitig die negative progno-
stische Aussagekraft dieser HRV-Einengung aufzeigen (u. a. Piepoli et al., 1995; Ellenby
et al., 2001; Schmidt et al., 2005; Schmidt et al., 2008a). Der pathophysiologische Hinter-
grund der HRV-Einschränkung und ihrer ungünstigen prognostischen Bedeutung bleiben
weithin ungeklärt.
John J. Skillman beschrieb 1969 ein tödliches Syndrom mit einem gleichzeitigen Auf-
treten von respiratorischer Insu�zienz, Hypotension, Sepsis und Ikterus (Skillman et al.,
1969). Die Arbeitsgruppe um Nicholas L. Tilney berichtete 1973, dass die postoperative
Mortalität bei chirurgischen Patienten mit der Anzahl der dysfunktionalen Organsyste-
me korreliere � er beschrieb die Entwicklung eines MODS aus einem SIRS und die damit
einhergehende hohe Sterblichkeit (Tilney et al., 1973; Godin and Buchman, 1996).
Ausgangspunkt für die Hypothese Paul J. Godins und Timothy G. Buchmans zur Ent-
stehung eines MODS (Godin and Buchman, 1996) war das ab dem Ende der 1980er-Jahre
aufkommende Verständnis physiologischer, biologischer Systeme als nonlineare, komplexe
Gebilde untereinander kommunizierender, verschieden oszillierender Regulatoren (Gold-
berger and West, 1987b; Pool, 1989). Bereits 1973 beschrieb Saykrs, dass sich die in der
HRV widerspiegelnden Regulationsschleifen und Oszillationen der Blutdruckregulation,

4
der Temperatur und der Atmung nonlinear verhalten und daher auf überraschende Art
und Weise ineinander eingreifen und sich gegenseitig beein�ussen können (Saykrs, 1973).
Die HRV und deren Frequenzanalyse, insbesondere in der 1/f -Darstellung, ist nach
Ary L. Goldberger der Beweis für eine physiologische, chaotisch erscheinende Interkon-
nektivität im Körper (Kobayashi and Musha, 1982; Goldberger and West, 1987a). Seine
Hypothese würde um so mehr gestützt, als dass schwere Erkrankungen, wie die Cheyne-
Stokes-Atmung und Herzrhythmusstörungen mit einer eingeschränkten HRV einhergin-
gen (Goldberger et al., 1984).
Da in einem komplexen Netzwerk jede Komponente die anderen moduliert und jede
Komponente von allen anderen moduliert werden kann, zeigt das messbare biometri-
sche Ausgangssignal einer solchen Komponente eine sehr hohe Variabilität, wobei sich
in komplexen Netzwerken bestimmte bevorzugte Interaktionszustände ausbilden (Seely
and Christou, 2000), wie es beispielsweise an einem typischen, physiologischen HRV-
Spektrum erkennbar ist. Steven Pincus leitete 1994 aus seinem mathematischen Modell
der approximate entropy ab, dass eine zunehmende Regelmäÿigkeit des Ausgangssignals
einer Komponente aus einem komplexen, untereinander gekoppelten System als Isolation
derselben in diesem System interpretiert werden kann (Pincus, 1994). Er schlussfolger-
te, dass eine zunehmende Regelmäÿigkeit eines biometrischen Signals, beispielsweise der
Herzfrequenz, ein Zeichen seiner zunehmenden Isolation im Organismus sei. Eine star-
re Herzfrequenz könne somit als Zeichen einer Entkopplung verschiedener oszillierender
Organsysteme gewertet werden und damit auf eine gestörte Interorgankommunikation
hinweisen.
Godin und Buchman entwickelten daraufhin 1996 die Hypothese, dass die Entwick-
lung eines MODS aus einem initialen Stimulus, wie einer Infektion, durch solch eine
Entkopplung der Interorgankommunikation entstünde, die sich an der eingeschränkten
HRV bei diesem Krankheitsgeschehen demonstriere (Godin and Buchman, 1996). Neue-
re Arbeiten deuten darauf hin, dass eine intakte Interorgankommunikation im Rahmen
einer Infektion direkt durch das autonome Nervensystem, insbesondere durch den choli-
nergic antiin�ammatory pathway gewährleistet werde, welcher im Rahmen einer Sepsis
möglicherweise unterbrochen oder gestört sei (Tracey, 2007; Schmidt et al., 2008b).
Bisher ist sowohl unklar, wo die Interorgankommunikation unterbrochen wird, als
auch, welche Substanzen an dieser Beeinträchtigung im Rahmen eines MODS involviert
sind. Als eine Möglichkeit kommt die Störung des autonomen Ein�usses direkt an den

5
Zielzellen, am Herzen an den kardialen Schrittmacherzellen, in Betracht (Garrard et al.,
1993; Schmidt et al., 2008b).
Neben TNFα, dessen Serumspiegel 2009 mit einer späten, langanhaltenden HRV-
Einschränkung bei Sepsis in Korrelation gebracht werden konnte (Fairchild et al., 2009),
könnten die eine Sepsis auslösenden Pathogene direkt für die septische Kardiomyopathie
mit der für sie charakteristischen HRV-Einschränkung verantwortlich sein (Abb. 1).
1989 experimentell durchgeführte Endotoxinapplikationen an Menschen provozierten
bei den Probanden eine linksventrikuläre systolische Dysfunktion, die der Kardiomy-
opathie bei Patienten mit Sepsis gleicht (Su�redini et al., 1989). In weiteren Studien
konnte eine deutliche dosisabhängige Einschränkung der HRV nach Endotoxinapplikati-
on in Tieren (Goldstein et al., 1995) und am Menschen beobachtet werden (Godin et al.,
1996). Die direkte Interaktion von Endotoxin, dem Lipopolysaccharid (LPS) gramnega-
tiver Bakterien, mit den Zielstrukturen des Herzens, wie beispielsweise dem kardialen
Schrittmacherkanal, könnte somit ein entscheidender Mechanismus in der Genese der
HRV-Einschränkung bei Sepsis sein (Schmidt et al., 2007; Zorn-Pauly et al., 2007).
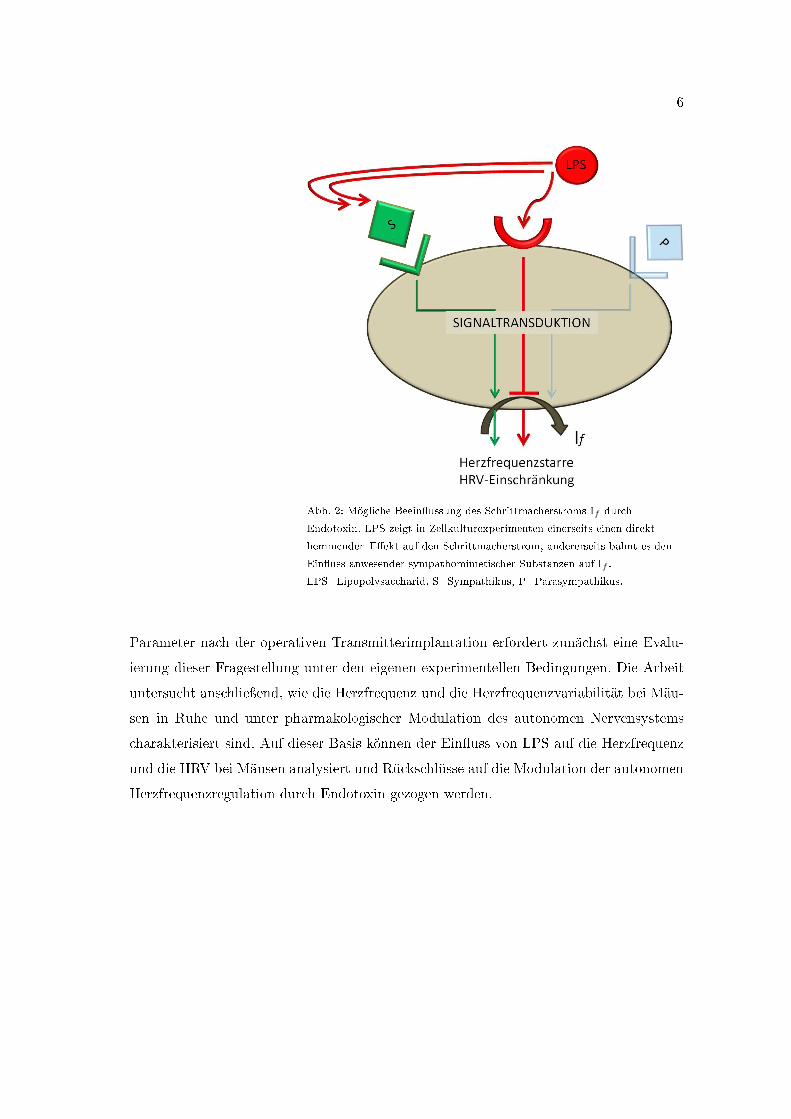
Die vorliegende Dissertation analysiert die Interaktion von Endotoxin mit den Schritt-
macherzellen des Herzens in vivo als eine mögliche Ursache der gestörten Organkommu-
nikation, welche sich in einer HRV-Einschränkung ausdrückt (Abb. 2). In Zellkulturex-
perimenten konnten im Voraus zwei mögliche Mechanismen der direkten Beein�ussung
des If -Stroms durch Endotoxin aufgezeigt werden (Zorn-Pauly et al., 2007). Zum einen
hemmt Endotoxin in Abwesenheit autonomer Ein�üsse den If -Strom direkt, indem es den
Strom�uss bei Membranpotentialen > -80 mV hemmt und die Aktivierung des Kanals
verlangsamt (Abb. 2). Dies sollte theoretisch in einer Bradykardisierung resultieren. Zum
anderen bahnt LPS in Anwesenheit sympathomimetischer Substanzen deren Ein�uss auf
den Schrittmacherstrom, wodurch sich eine Tachykardie ausbilden sollte. Beide Mecha-
nismen könnten daher an der Entstehung einer prognoserelevanten HRV-Einschränkung
bei Intensivpatienten mit MODS beteiligt sein und eine Ursache für die Unterbrechung
der Interorgankommunikation auf Endorganebene darstellen.
In dieser Arbeit werden die im Zellkulturexperiment nachgewiesenen Mechanismen in
vivo am Modell der Wildtyp-Maus untersucht. Dabei werden die EKGs der Mäuse mit-
tels intraperitoneal platzierter Telemetrietransmitter erfasst, und aus ihnen verschiedene
etablierte Parameter der Herzfrequenzvariabilität ermittelt (Malik et al., 1996; Thireau
et al., 2008). Die unterschiedliche Datenlage zur Stabilisierung der Tiere und der HRV-
6
Abb. 2: Mögliche Beein�ussung des Schrittmacherstroms If durch
Endotoxin. LPS zeigt in Zellkulturexperimenten einerseits einen direkt
hemmenden E�ekt auf den Schrittmacherstrom, andererseits bahnt es den
Ein�uss anwesender sympathomimetischer Substanzen auf If .
LPS=Lipopolysaccharid, S=Sympathikus, P=Parasympathikus.
Parameter nach der operativen Transmitterimplantation erfordert zunächst eine Evalu-
ierung dieser Fragestellung unter den eigenen experimentellen Bedingungen. Die Arbeit
untersucht anschlieÿend, wie die Herzfrequenz und die Herzfrequenzvariabilität bei Mäu-
sen in Ruhe und unter pharmakologischer Modulation des autonomen Nervensystems
charakterisiert sind. Auf dieser Basis können der Ein�uss von LPS auf die Herzfrequenz
und die HRV bei Mäusen analysiert und Rückschlüsse auf die Modulation der autonomen
Herzfrequenzregulation durch Endotoxin gezogen werden.

7
2 Zielstellung
Ziel der Dissertation ist es, den vorangegangenen in-vitro-Nachweis der direkten Inter-
aktion von Endotoxin mit dem Schrittmacherkanal des Herzens, in vivo am Modell der
Wildtyp-Maus zu evaluieren und damit einen Beitrag zur Aufklärung der Entstehung
einer hochgradig eingeschränkten Herzfrequenzvariabilität (HRV) bei Intensivpatienten
mit Multiorgan-Dysfunktions-Syndrom (MODS) zu liefern. Für die Interpretation der
durch Biopotentialtransmitter telemetrisch gewonnenen HRV-Daten ist die Beurteilung
der postoperativen Stabilisierung und der autonomen Regulation der Ruheherzfrequenz
der Wildtyp-Mäuse wesentlicher Ausgangspunkt. Daher werden folgende Fragestellungen
bearbeitet:
1. Welcher postoperative Erholungszeitraum ist nach Implantation der EKG-Transmitter
für die Stabilisierung der Tiere notwendig?
2. Wie hoch ist die telemetrisch bestimmte Ruheherzfrequenz und wo liegen die Ruhe-
HRV-Parameter bei ICR-Wildtyp-Mäusen?
3. Wie werden die Ruheherzfrequenz und die HRV bei ICR-Wildtyp-Mäusen durch
das autonome Nervensystem moduliert?
4. Welchen Ein�uss hat Endotoxin mit und ohne autonome Blockade auf die Herzfre-
quenz und die HRV?
Anhand dieser Fragestellungen soll die Entstehung der HRV-Einschränkung im Rahmen
eines septischen MODS diskutiert und die mögliche Rolle des Endotoxins als Störfaktor
der HRV-Regulation auf Endorganebene erörtet werden.

8
3 Material und Methodik
3.1 Versuchstiere und Tierhaltung
Die Experimente fanden im Zentrum für Medizinische Grundlagenforschung (ZMG) an
der Martin-Luther-Universität Halle-Wittenberg statt. Das Referat Verbraucherschutz,
Veterinärangelegenheiten des Landesverwaltungsamtes Sachsen-Anhalt genehmigte die
Durchführung der Versuche an den lebenden Tieren. Zur telemetrischen Datengewinnung
wurden zehn adulte männliche ICR-Wildtyp-Mäuse (CD-1 R©-Mäuse, Harlan Winkelmann
GmbH Borchen) operativ mit einem intraperitonealen Transmittersystem versorgt. Nach
15-tägiger postoperativer Beobachtung und durchgängiger Registrierung der motorischen
Aktivität sowie der Herzfrequenz begann die Durchführung der Injektions-Experimente.
Die konventionelle Haltung der Tiere erfolgte im ZMG. Sie richtete sich nach den
Empfehlungen für Versuchstierhaltung der Europäischen Kommission von 2007 (Dimas,
2007). Während der Aufzeichnungsphase waren die Tiere in Einzelhaltung in Typ II-
Normkä�gen (207 x 265 x 140 cm, insgesamt 363 cm2 Boden�äche) untergebracht. Die
Polycarbonat-Kä�ge besitzen einen Edelstahlgitter-Deckel mit einer Futterraufe. Der Bo-
den ist mit Weichholzfasern und Zellsto� ausgelegt; zudem be�ndet sich in jedem Kä�g
ein Maus-Iglu (Polycarbonat, 100 x 90 x 55 cm, rot-transparent).
Die Mäuse erhielten Allgemeinfuttermittel (Altromin 1324, Altromin Spezialfutter
GmbH&CoKG, Im Seelenkamp 20, D-32791 Lage) und Leitungswasser über eine 300 ml-
Trink�asche ad libitum. Die Raumtemperatur betrug 22± 1 ◦C bei einer Luftfeuchtigkeit
von 64± 1 %. Im Wechsel von zwölf Stunden wurde der Tag-Nacht-Rhythmus simuliert.
3.2 Implantation der Telemetrie-Transmitter
Die Telemetrie-Transmitter PhysioTelTM EA-F20 der Firma Data Science International
(s. Kapitel 3.4, S. 12) wurden unter Inhalationsnarkose in die Bauchhöhle implantiert.
Als Anästhetikum diente Iso�uran in einer Einleitungskonzentration von 4 % und einer
Erhaltungskonzentration von 1,5 � 2,5 % in Sauersto� (van Zutphen et al., 1995.). Die
Mäuse wurden gewogen, in Rückenlage auf eine Wärmeplatte bei 38 ◦C gelegt und �xiert.
Die Analgesierung erfolgte mittels Carprofen (Rimadyl R©) 5 mg/kg Körpergewicht (van
Zutphen et al., 1995.), das subkutan in die Beinfalte appliziert wurde. Das Fell der ge-
samten Bauch�äche und der Fixationspunkte der Telemetrieelektroden wurde groÿ�ächig
rasiert und mit Povidon-Iod-Lösung (Braunol R©, B. Braun Melsungen AG) desin�ziert.

9
Abb. 3: Subkutane Tunnelung mittels Metallstab und Verlegung der
Transmitterelektroden durch die Plastikhülse (Data Science International,
2008; S. 3-6, Abb. 5).
Abb. 4: Endgültige Lage des
Transmitters (Data Science
International, 2008; S. 3-5
Abb. 3).
Die Operation erfolgte nach Anleitung des Hardwareherstellers (Data Science In-
ternational, 2008). Zunächst wurde eine mediane Laparotomie durchgeführt: Entlang
der Medianlinie erfolgte ein zwei Zentimeter langer Hautschnitt; das Peritoneum wurde
stumpf abpräpariert und entlang der Linea alba erö�net. Der Transmitter konnte nun
steril in die Bauchhöhle eingeführt werden. Das Peritoneum wurde daraufhin lateral des
Schnittes an zwei Punkten mit einer scharfen, groÿlumigen Kanüle durchstochen und die
Ableitelektroden durch die Kanüle hindurchgeführt, so dass diese nun extraperitoneal
zum Liegen kamen. Die Peritonealwunde wurde mittels chirurgischer Naht mit resorbier-
barem Nahtmaterial verschlossen (Polyglactin 910, VICRYL R©, 5-0).
Für die Platzierung der Ableitelektroden wurde ein Metallstab subkutan zu den Fi-
xationspunkten am rechten Brustkorb in Höhe der rechten Vorderpfote für die negative
Elektrode und unter den linken Rippenbogen für die positive Elektrode vorgeschoben und
durch jeweils einen kleinen Hautschnitt wieder ausgeführt (Abb. 3). Über diesen wurde
eine Plastikhülse gefädelt und der Metallstab herausgezogen. Die Ableitelektroden lieÿen
sich nun durch die Hülle hindurchführen. Ihre Ausrichtung entspricht der Extremitä-
tenableitung II nach Einthoven (Abb. 4). Die Sonden wurden mit dem tip cover an dem
Muskel angenäht und dadurch �xiert. Nach Verschluss der Hautwunden mittels nicht-

10
Tab. 1: Dosierung, Bezug und Literaturnachweis der verwendeten Pharmaka
Pharmaka Dosierung Injektions-
volumen*
Lieferant/Hersteller Hinweise zur Dosierung
Propranolol-
hydrochlorid
20 mg/kg ca. 900 µl MIBE GmbH Arzneimittel Weiergräber et al., 2005
Atropinsulfat 1 mg/kg ca. 200 µl B. Braun Melsungen AG Weiergräber et al., 2005
LPS (1/100
der LD50)
0,4 mg/kg ca. 200 µl Sigma Aldrich Logistik Corral et al., 2005
Isoproterenol-
hydrochlorid
0.5 mg/kg ca. 200 µl Sigma Aldrich Logistik Stieber et al., 2006
*Empfehlung der GV-SOLAS zur Substanzapplikation bei Versuchstieren (GV-SOLAS, 2008).
resorbierbaren Nahtmaterial (Polypropylen, Prolene R©) wurde die Narkose zeitnah been-
det. Die Tiere erlangten innerhalb von fünf bis zehn Minuten das Bewusstsein wieder. Die
Operation von Beginn der Narkose bis zur Ausleitung dauerte 35± 5 Minuten. Postope-
rativ erhielten die Mäuse für drei Tage zur Analgesie Carprofen 50 mg/l im Trinkwasser.
3.3 Postoperative Beobachtung, pharmakologische Beein�ussung des
autonomen Nervensystems und Endotoxinapplikation
Während der 15 ersten Tage nach der operativen Transmitterimplantation wurden die
Daten unter der Fragestellung der Entwicklung der Ruheherzfrequenz, der Bewegungs-
aktivität, der Stabilisierung der HRV-Parameter sowie des Eintretens zirkadianen Ver-
haltens kontinuierlich aufgezeichnet. Bei der statistischen Datenanalyse wurden der 1.,
5. und 10. postoperative Tag verglichen (Thireau et al., 2008). Während dieser Pha-
se fanden keine Injektions-Experimente statt; mit diesen wurde frühestens ab dem 16.
postoperativen Tag begonnen (Abb. 5, Leon et al., 2004).
Das autonome Nervensystem wurde durch intraperitoneale Injektion verschiedener
Pharmaka beein�usst. Eine Übersicht der verwendeten Substanzen sowie deren Dosie-
rungen gibt Tabelle 1. Propranololhydrochlorid und Atropinsulfat wurden als fertige
Lösungen vom Hersteller bezogen und bei Raumtemperatur gelagert.
Isoproterenolhydrochlorid und Endotoxin (Lipopolysaccharid, LPS) wurden als Pul-
ver bezogen, in PBS gelöst und als hochkonzentrierte Stocklösungen bei -20 ◦C gelagert.
Die Substanzen wurden 30 Minuten vor den Injektionen auf Raumtemperatur aufge-
taut. Alle applizierten Pharmaka wurden unmittelbar vor den Versuchen mit Phosphate

11
Abb. 5: Versuchsablauf.
Bu�ered Saline (PBS) auf die zu injizierenden Volumina verdünnt (Tab. 1).
Die insgesamt 16 Injektionen pro Maus wurden unter Kurznarkose mit 2,5 % Iso�u-
ran durchgeführt, wobei die Pharmaka über die Beinfalte oder bei Behinderung durch
die Lage des Transmitters über den linken Unterbauch intraperitoneal appliziert wur-
den. Um den Ein�uss physiologischer zirkadianer Schwankungen der Herzfrequenz und
der Aktivität der Mäuse auf die Versuche so gering wie möglich zu halten, wurden alle
Injektionen zwischen 16 und 18 Uhr durchgeführt. Sie erfolgten in drei Blöcken (Abb. 5).
Im ersten Block wurden je dreimal im Wechsel der nicht-selektive β-Blocker Propra-
nolol, das Anticholinergikum Atropin und beide in Kombination zur Herstellung einer
kompletten autonomen Blockade appliziert. Der zeitliche Mindestabstand dieser aufein-
ander folgenden Injektionen betrug 24 Stunden. In einem zweiten Block wurde der E�ekt
von LPS sowie LPS unter autonomer Blockade mittels Atropin und Propranolol auf die
Herzfrequenzvariabilität getestet. Da eine länger andauernder E�ekt von LPS auf den
Organismus und die HRV zu erwarten war (Godin et al., 1996; Fairchild et al., 2009),
wurde der zeitliche Mindestabstand zwischen den insgesamt vier Injektionen auf jeweils
48 Stunden erweitert.

12
Abb. 6: Transmitter PhysioTelTM EA-F20, Data Science International,
USA; Archiv der Verf.
Der dritte Versuchsblock umfasste die Injektion von Isoproterenol. Die Substanz wur-
de jeweils dreimal im Abstand von 24 Stunden verabreicht.
3.4 Datengewinnung und Datenaufzeichnung
Während der telemetrischen Datenaufzeichnung befanden sich die Mäuse in ihrer ge-
wohnten Umgebung und konnten sich frei bewegen. Daten zur Beurteilung der Stabi-
lisierung der HRV-Parameter und der Bewegung der Tiere nach der operativen Trans-
mitterimplantation wurden von neun Wildtyp-Mäusen ausgewertet, eine Maus verstarb
am 1.Tag nach der Transmitterimplantation. Von acht Wildtyp-Mäusen wurden Daten
aus den Injektionsversuchen gewonnen; eine Maus verstarb nach dem postoperativen Be-
obachtungzeitraum von 15 Tagen noch vor Initiierung der Injektions-Experimente. Die
für die Aufzeichnungen verwendete Hard- und Software wurde von Data Science In-
ternational (DSI) bezogen (Data Science International, PhysioTel R© and MultiplusTM
Implant, St. Paul, Minnesota, USA). Die verwendeten Transmitter PhysioTelTM EA-
F20 (Abb. 6) eignen sich für mehrere aufeinander folgende Aufzeichnungen. Sie wurden
nach Versuchsablauf und Tötung der Versuchstiere durch cervikale Dislokation aus der
Bauchhöhle entfernt, manuell gesäubert und sterilisiert. Die Sterilisation erfolgte für 24
Stunden in 2%iger Glutardialdehyd (GDA)-Lösung. Nach Waschen der Transmitter mit
sterilem PBS standen diese für eine neue Implantation zur Verfügung.
Die Transmitter wiegen zwischen 3,55 g und 3,80 g. Sie bestehen aus einem Silikonkör-
per (2 x 1 x 1 cm), der die Batterie und die elektronischen Bauelemente enthält (Abb. 6).
Diese detektieren über die Ableitelektroden die Biopotential-Veränderungen, die elektri-
sche Herzaktion, und senden die Daten mittels Radiowellen zu einem Receiver (Abb. 7).
Der Transmitter kann magnetisch an- und ausgeschaltet werden. Die zwei Ableitelektro-
den (biopotential leads) sind mit Silikon ummantelt um Störsignale abzuschirmen. Am

13
Abb. 7: Die Datengewinnung über das Telemetriesystem, mod�ziert nach Data Science International, 2010.
ECG=Electrocardiogram, APR-1=Ambient Pressure Reference Monitor.
Ende der Elektroden be�ndet sich ein kurzes Stück frei gewundenes Kabel (helix of wi-
re), über welches die Biopotentiale detektiert werden. Die Elektroden werden von jeweils
einer Hülse (tip cover) vor Irritationen am Implantationsort geschützt. Es standen zwei
Receiver (RLA) zur Verfügung, die in Form von Platten unter jeweils einem Mauskä�g
standen, so dass maximal zwei Aufzeichnungen parallel laufen konnten. Die Umwandlung
der analogen in digitale Daten erfolgt mit einer Sampling-Frequenz von 2000 Hz ohne Si-
gnal�lter (Malik et al., 1996; Thireau et al., 2008). Diese �ndet im Transmitter selbst
statt (Data Science International, 2008, vgl. Thireau et al., 2008).
Die Receiver leiten die digitalisierten Daten an die Data Exchange Matrix weiter,
die diese Informationen bündelt und nach den einzelnen Transmittern aufgeschlüsselt,
in einem für die dazugehörige Software (DSIs Dataquest A.R.T. Silver 3.1 Acquisition-
System) lesbaren Datenformat weiterleitet. Die Bewegung der Mäuse (activity, act) wird
durch die relative Änderung der Signalstärke der Radiowellen aus den Transmittern zu
den Receivern ermittelt, die sich aus der Änderung der räumlichen Lage der Mäuse im
Kä�g ergibt. Je nach der Änderung der Signalstärke leitet die Data Exchange Matrix

14
einen oder mehrere digitale Impulse an den Computer weiter (Data Science International,
2009).
Mittels der Acquisition-Software (Dataquest A.R.T. 3.1 Silver) wurden die verschie-
denen Transmitter kalibriert und den jeweiligen Receivern zugeordnet. Das EKG und
die motorische Aktivität wurden aufgezeichnet und online graphisch dargestellt, so dass
diese vom Benutzer aufzeichnungssynchron beobachtet werden konnten. Die Roh-Daten
wurden automatisch in zuvor de�nierten Ordnern auf der Festplatte des Computersy-
stems gespeichert, die dann für die Auswertung mittels der Analysis-Software abrufbar
waren. Die Datenaufzeichnung erfolgte für die ersten 15 Tage postoperativ kontinuierlich.
Sie wurde alle drei bis vier Tage kurz unterbrochen und unter einem neuen Speicherplatz
neu gestartet, um die Gröÿe der Datenpakete für die darauf folgende Bearbeitung hand-
habbar zu machen. Für die Injektionsversuche wurde die Aufzeichnung spätestens eine
halbe Stunde vor der Injektion neu gestartet und für mindestens eine Stunde nach der
Injektion fortgesetzt.
3.5 Datenverarbeitung
Die Daten wurden mit dem Programm Analysis (DSI) sowie Microsoft Excel 2003 bear-
beitet. Die EKG- und Bewegungsdaten wurden mittels zweier verschiedener Algorithmen
importiert; zum einen für die Langzeitaufzeichnungen im postoperativen Beobachtungs-
zeitraum, zum anderen für die Beurteilung der Pharmakae�ekte. Für beide wurden sowohl
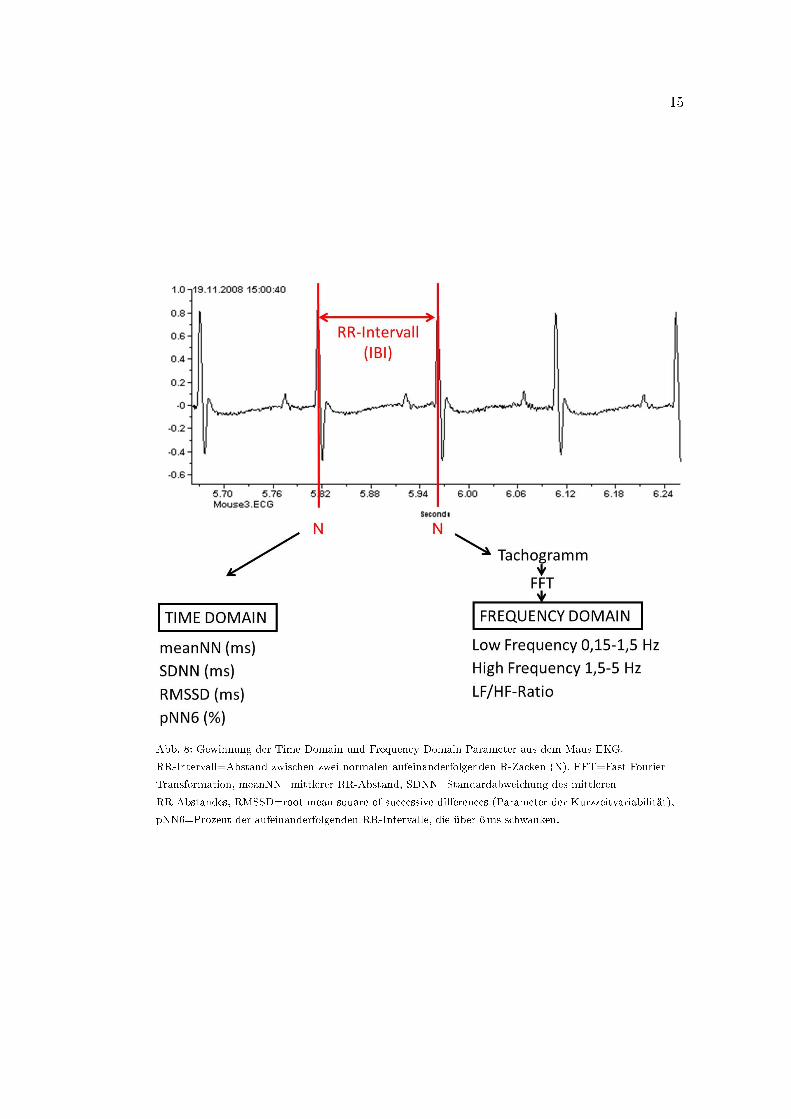
Time Domain als auch Frequency Domain Parameter gewonnen (Abb. 8).
Für die HRV-Analyse mussten zunächst die RR-Intervalle (NN-Abstände, Inter Beat
Intervalle) aus dem EKG extrahiert werden (Abb. 8, Malik et al., 1996). Dafür wurden
die QRS-Komplexe des importierten EKGs mittels automatischer R-Zacken-Erkennung
detektiert und markiert. Mittels der Markierung konnten die RR-Intervalle errechnet
werden und standen in einem neuen Datenpaket gebündelt für die Gewinnung der HRV-
Parameter zur Verfügung.
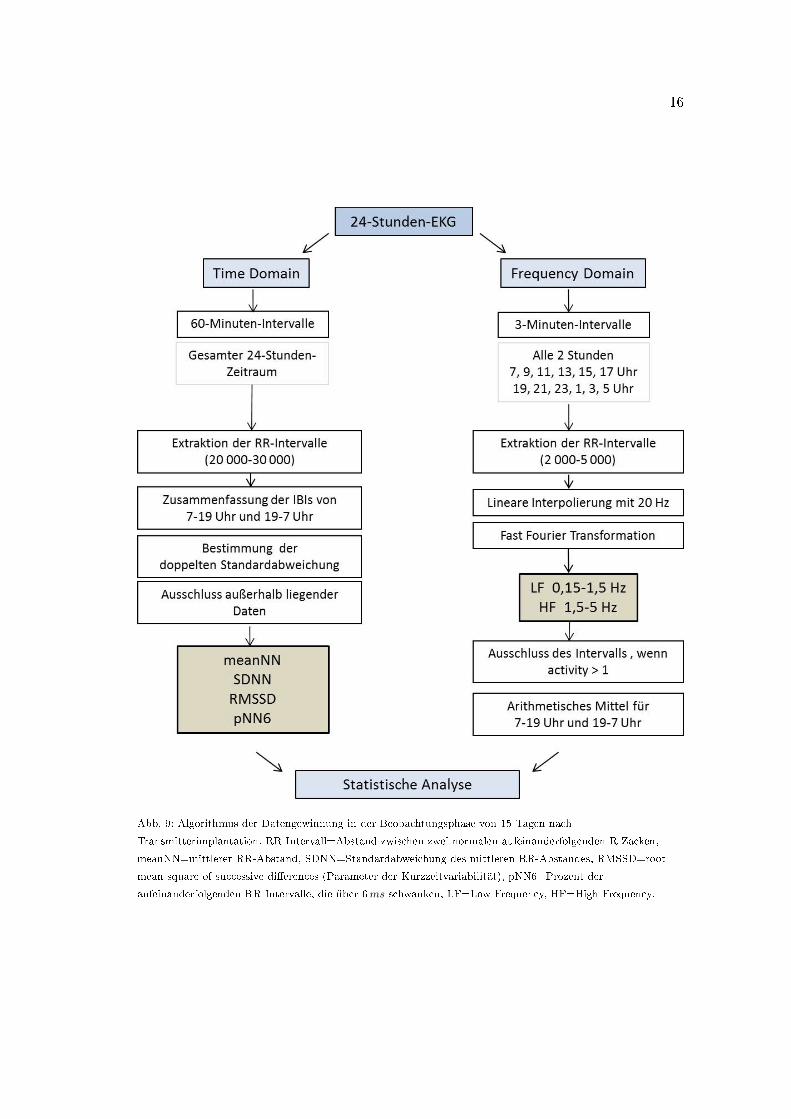
Stabilisierung nach operativer Transmitterimplantation Die Stabilisierung nach der ope-
rativen Transmitterimplantation wurde anhand der Entwicklung von Herzfrequenz und
HRV in der Ruhephase (Helligkeitsphase, 7�19 Uhr) des 1., 5. und 10. postoperativen Tags
beurteilt (Thireau et al., 2008). Zudem wurde das Eintreten einer zirkadianen Rhythmik
beobachtet. Für die Gewinnung der Time Domain Parameter wurden die EKG-Daten
15
Abb. 8: Gewinnung der Time Domain und Frequency Domain Parameter aus dem Maus-EKG.
RR-Intervall=Abstand zwischen zwei normalen aufeinanderfolgenden R-Zacken (N), FFT=Fast Fourier
Transformation, meanNN=mittlerer RR-Abstand, SDNN=Standardabweichung des mittleren
RR-Abstandes, RMSSD=root mean square of successive di�erences (Parameter der Kurzzeitvariabilität),
pNN6=Prozent der aufeinanderfolgenden RR-Intervalle, die über 6ms schwanken.
16
Abb. 9: Algorithmus der Datengewinnung in der Beobachtungsphase von 15 Tagen nach
Transmitterimplantation. RR-Intervall=Abstand zwischen zwei normalen aufeinanderfolgenden R-Zacken,
meanNN=mittlerer RR-Abstand, SDNN=Standardabweichung des mittleren RR-Abstandes, RMSSD=root
mean square of successive di�erences (Parameter der Kurzzeitvariabilität), pNN6=Prozent der
aufeinanderfolgenden RR-Intervalle, die über 6ms schwanken, LF=Low Frequency, HF=High Frequency.

17
in 60-Minuten-Paketen importiert und die Ausgabe der RR-Intervalle, ca. 20 000�30 000
pro Paket, als Text-Datei gespeichert (Abb. 9). Zwölf 60-Minuten-Pakete wurden entspre-
chend der Hell- und Dunkelphasen, von 7�19 und 19�7 Uhr, zusammenhängend in eine
Excel-Arbeitsmappe importiert. Aus den RR-Intervallen eines Zwölf-Stunden-Abschnitts
wurde der Mittelwert und die doppelte Standardabweichung bestimmt und Werte ent-
fernt, die auÿerhalb der doppelten Standardabweichung lagen (Thireau et al., 2008). Aus
den selektierten Daten lieÿen sich die Time Domain Parameter meanNN, SDNN, RMSSD
und pNN6 ermitteln (Tab. 2). Die Herzfrequenz (HR) wurde aus der meanNN bestimmt
(HR [min−1]=60∗1000/meanNN [ms]).
Für die Frequency Domain Analyse wurden im Abstand von zwei Stunden (7, 9, 11,
13, 15, 17, 19, 21, 23, 1, 3 und 5 Uhr) jeweils drei 3-Minuten-Segmente aus den EKGs
ausgewählt (Abb. 9). Die Bewegungsaktivität wurde für diese drei Minuten sowie für den
umgebenden 30-Minuten-Zeitraum (insgesamt 33 Minuten) bestimmt. Für die Berech-
nungen wurden die EKG-Segmente nur dann verwendet, wenn die zugehörige Aktivität
der Mäuse < 1 betragen hatte. Nach Extraktion der RR-Intervalle (ca. 2 000-2 500) aus
dem EKG wurde ein Tachogramm erstellt, welches mit 20 Hz linear interpoliert wurde
(Thireau et al., 2008). Für die anschlieÿende Fast Fourier Transformation (FFT) wurde
der Low Frequency (LF) Bereich von 0,15 bis 1,50 Hz festgelegt, der High Frequency
(HF) Bereich von 1,5 bis 5,0 Hz (Just et al., 2000; Thireau et al., 2008). Die Power
wurde in absoluten Einheiten in ms2 für beide Bereiche aufgelistet und gespeichert. Das
Ergebnis der FFT wurde von der Software automatisch mit einer Segmentlänge von 2048
Punkten graphisch in einem linearen Koordinatensystem dargestellt, wobei auf der Ordi-
nate die Power Spektrum Density in ms2/Hz gegen die Frequenz in Hz auf der Abszisse
abgetragen wurde. Aus dieser Darstellungsweise ist die Power in ms2 als Fläche unter
der Kurve als Integral erkennbar (Malliani et al., 1991). Nach dem zwölfstündigen Hell-
Dunkel-Rhythmus unterteilt, wurden die Daten separat in Excel importiert. Neben der
absoluten LF und HF erfolgte hier die Ermittlung der LF/HF-Ratio (Tab. 3).
Autonomes Nervensystem und LPS-E�ekt Die Erfassung der Kontrolldaten wurde aus
je drei 3-Minuten-Segmenten der Ruhe-EKGs gewonnen, die 25�10 Minuten vor den
Injektionen aufgezeichnet wurden. 20�40 Minuten nach der Injektion der verschiedenen
Substanzen wurden drei weitere 3-Minuten-Abschnitte des EKGs ausgewählt, um den
E�ekt der Applikationen zu beurteilen (Abb. 10, Thireau et al., 2008).
18
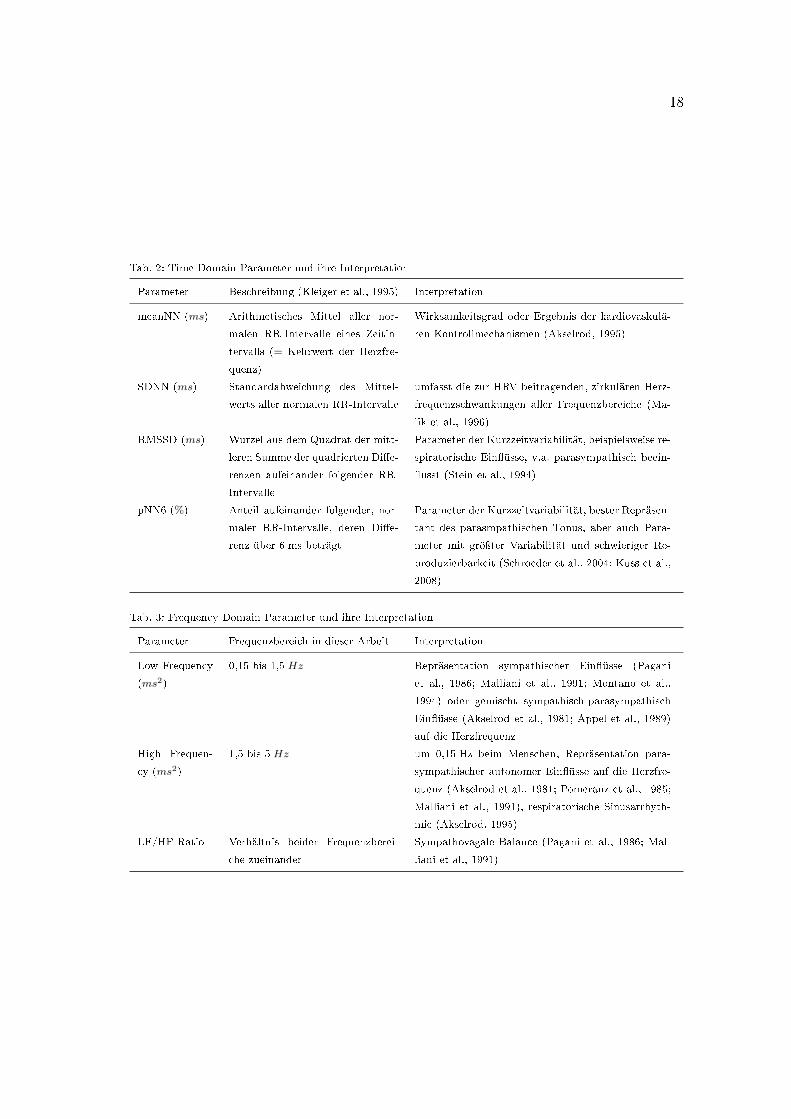
Tab. 2: Time Domain Parameter und ihre Interpretation
Parameter Beschreibung (Kleiger et al., 1995) Interpretation
meanNN (ms) Arithmetisches Mittel aller nor-
malen RR-Intervalle eines Zeitin-
tervalls (= Kehrwert der Herzfre-
quenz)
Wirksamkeitsgrad oder Ergebnis der kardiovaskulä-
ren Kontrollmechanismen (Akselrod, 1995)
SDNN (ms) Standardabweichung des Mittel-
werts aller normalen RR-Intervalle
umfasst die zur HRV beitragenden, zirkulären Herz-
frequenzschwankungen aller Frequenzbereiche (Ma-
lik et al., 1996)
RMSSD (ms) Wurzel aus dem Quadrat der mitt-
leren Summe der quadrierten Di�e-
renzen aufeinander folgender RR-
Intervalle
Parameter der Kurzzeitvariabilität, beispielsweise re-
spiratorische Ein�üsse, v.a. parasympathisch beein-
�usst (Stein et al., 1994)
pNN6 (%) Anteil aufeinander folgender, nor-
maler RR-Intervalle, deren Di�e-
renz über 6 ms beträgt
Parameter der Kurzzeitvariabilität, bester Repräsen-
tant des parasmpathischen Tonus, aber auch Para-
meter mit gröÿter Variabilität und schwieriger Re-
produzierbarkeit (Schroeder et al., 2004; Kuss et al.,
2008)
Tab. 3: Frequency Domain Parameter und ihre Interpretation
Parameter Frequenzbereich in dieser Arbeit Interpretation
Low Frequency
(ms2)
0,15 bis 1,5 Hz Repräsentation sympathischer Ein�üsse (Pagani
et al., 1986; Malliani et al., 1991; Montano et al.,
1994) oder gemischt sympathisch-parasympathisch
Ein�üsse (Akselrod et al., 1981; Appel et al., 1989)
auf die Herzfrequenz
High Frequen-
cy (ms2)
1,5 bis 5 Hz um 0,15 Hz beim Menschen, Repräsentation para-
sympathischer autonomer Ein�üsse auf die Herzfre-
quenz (Akselrod et al., 1981; Pomeranz et al., 1985;
Malliani et al., 1991), respiratorische Sinusarrhyth-
mie (Akselrod, 1995)
LF/HF-Ratio Verhältnis beider Frequenzberei-
che zueinander
Sympathovagale Balance (Pagani et al., 1986; Mal-
liani et al., 1991)

19
Die Auswahl der Segmente erfolgte manuell nach dem Kriterium der möglichst voll-
ständigen Bewegungsfreiheit. Segmente mit einer motorischen activity > 1 wurden von
der Auswertung ausgeschlossen. Aus den 3-Minuten-EKGs wurden die RR-Intervalle ex-
trahiert und für die Berechnung der Time Domain Parameter in Excel als Textdatei
gespeichert. Für die Frequency Domain Analyse verblieben die RR-Intervalle in der EKG-
Analyse-Software, wo sie mittels FFT weiterverarbeitet wurden.
Statistik Die statistischen Analysen wurden mit dem ProgrammWinStat 2001.1 fürMi-
crosoft Excel 2003 durchgeführt. Die Auswertung des postoperativen Beobachtungszeit-
raums beruht auf der Datenauswahl von neun Wildtyp-Mäusen. Nicht in die Auswertung
einbezogen werden konnte eine Wildtyp-Maus, die in Folge der Transmitterimplantation
am zweiten postoperativen Tag verstarb. Nach Testung auf Normalverteilung mittels des
Kolmogorov-Smirnov-Anpassungstests erfolgte die Bestimmung der Signi�kanz mit dem
t-Test für verbundene Stichproben.
Die Beurteilung der Zirkadianität erfolgte anhand der Schwankung der meanNN (und
Herzfrequenz) sowie der motorischen Aktivität von 7�19 Uhr und 19�7 Uhr des 1., 5. und
10. postoperativen Tages. Aus den jeweils sechs 3-Minuten- und umgebenden 30-Minuten-
Aktivitäts-Intervallen pro Ruhe- (7�19 Uhr) und Aktivitätsphase (19�7 Uhr) wurde das
arithmetische Mittel gebildet.
Die Stabilisierung der Ruhe-HRV wurde anhand der während der Ruhephase der
Tiere (7�19 Uhr) am 1., 5. und 10. postoperativen Tag gewonnenen Daten untersucht.
Ein Vergleich der Time Domain erfolgte aus den Langzeitaufzeichnungen. Für die statisti-
sche Analyse der Frequency Domain wurden aus den jeweils sechs 3-Minuten-Abschnitten
der Ruhephase (7�19 Uhr) die Intervalle mit einer 3-Minuten-Aktivität < 1 count/min
ausgewählt. Die Aktivität in den umgebenden 30 Minuten lag bei den ausgewählten In-
tervallen unter 5 counts/min. Aus den verbliebenen Intervallen wurde das arithmetische
Mittel gebildet.
Für die Auswertung der Injektionsexperimente wurden alle Intervalle mit einer acti-
vity < 1 eingeschlossen. Als Kontrollwert diente für jede Maus der Mittelwert der jeweili-
gen 3-Minuten-Intervalle, die vor den Injektionen gewonnen wurden und eine activity < 1
aufwiesen. Nach Testung auf Normalverteilung erfolgte die Bestimmung des Signi�kanz-
niveaus mit dem t-Test für verbundene Stichproben. Signi�kante Unterschiede der E�ekte
verschiedener Pharmaka untereinander wurden mit dem t-Test für unverbundene Stich-
proben bestimmt. Als signi�kant wurde ein p-Wert < 0,05 festgelegt. Alle in den Tabel-
len und Abbildungen dargestellten Daten werden als Mittelwert± Standardabweichung
(MW± SD) angegeben.
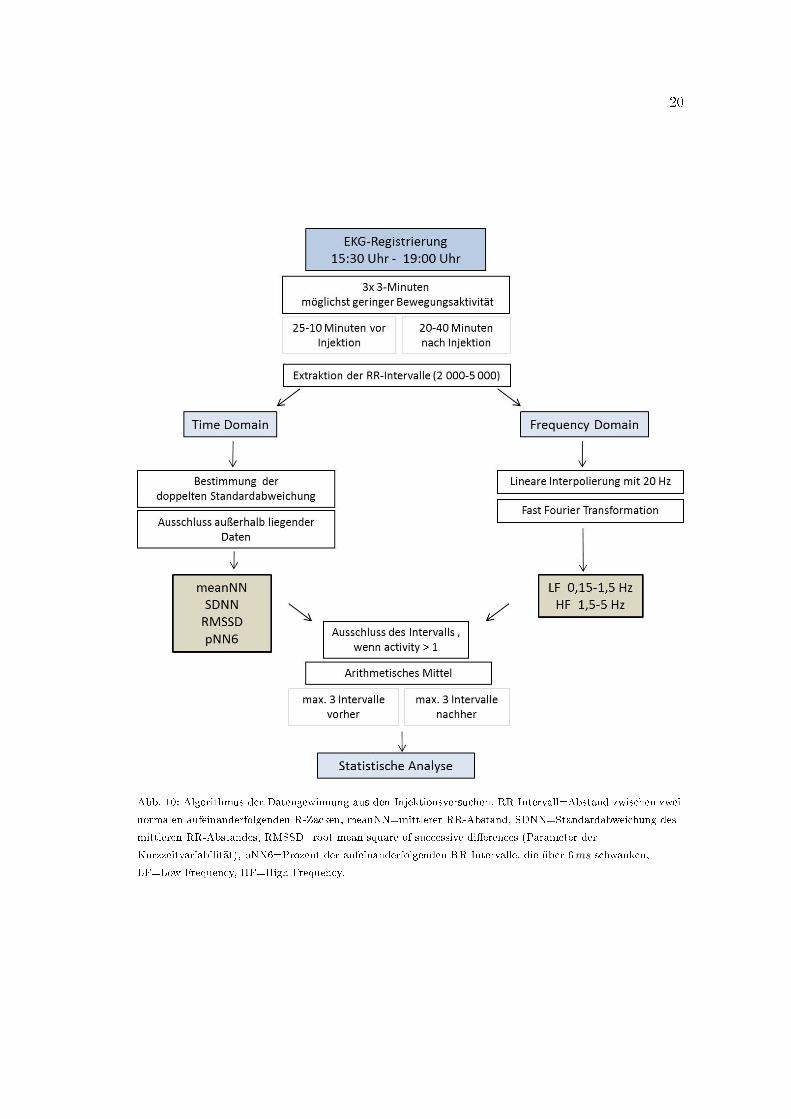
20
Abb. 10: Algorithmus der Datengewinnung aus den Injektionsversuchen. RR-Intervall=Abstand zwischen zwei
normalen aufeinanderfolgenden R-Zacken, meanNN=mittlerer RR-Abstand, SDNN=Standardabweichung des
mittleren RR-Abstandes, RMSSD=root mean square of successive di�erences (Parameter der
Kurzzeitvariabilität), pNN6=Prozent der aufeinanderfolgenden RR-Intervalle, die über 6ms schwanken,
LF=Low Frequency, HF=High Frequency.

21
4 Ergebnisse
4.1 Stabilisierung und Eintreten einer zirkadianen Rhythmik nach operativer Transmit-
terimplantation
Neun von zehn ICR-Wildtyp-Mäusen zeigen unmittelbar nach der Transmitterimplantati-
on einen guten Allgemeinzustand und im weiteren postoperativen Erholungszeitraum von
15 Tagen keine Au�älligkeiten (Hawkins, 2002). Anhand der 24-Stunden-Herzfrequenz-
Aufzeichnungen ist die wellenförmige, helligkeitssynchrone Schwankung der Herzfrequenz
und mit ihr die zirkadiane Herzfrequenzregulation erkennbar (Abb. 11). Zudem lässt sich
die Herzfrequenzvariabilität an der Breite des schwarzen Aufzeichnungsstreifens orientie-
rend erkennen.
Das Gewicht ist bei den meisten Tieren nach 15 Tagen stabilisiert. Die zwei Monate
alten Wildtyp-Mäuse nehmen deutlich an Gewicht zu (Tab. 4, > 10 %). Das Elektro-
kardiogramm und die Bewegungsaktivität der Mäuse können unmittelbar nach Ende der
Operation durch die Telemetrieanlage aufgezeichnet und beobachtet werden, wobei die p-
Wellen und QRS-Komplexe deutlich di�erenzierbar sind. Wie bereits bei der erstmaligen
Aufzeichnung eines Maus-EKGs beschrieben (Goldbarg et al., 1968), ist die T-Welle auch
in den hier durchgeführten EKG-Aufzeichnungen nicht gesondert abgrenzbar (Abb. 8).
Entladene Transmitterbatterien führten zu einer unvollständigen Datenaufzeichnung in-
nerhalb des postoperativen Beobachtungszeitraums bei zwei Mäusen, bei weiteren zwei
Tab. 4: Eigenschaften der Versuchstiere
Maus (N=10) Alter (Mo) Gewicht vor Transmitterimplantation (g) Gewicht nach 15 Tagen (g)
1 4 47,5 46,1
2 4 40,2 40,5
3 13 44,2 41,9
4 7 37,2 40
5 11 40 40,1
6 11 39 37,4
7 2 30 36
8 2 34 38
9 2 31,5 35
10 8 38,8 -?
Es wurden 10 Wildtyp-Tiere untersucht. Alle Tiere waren männlich. Das Alter ist in Monaten (Mo) angegeben.?Tod der Maus am 2. postoperativen Tag.
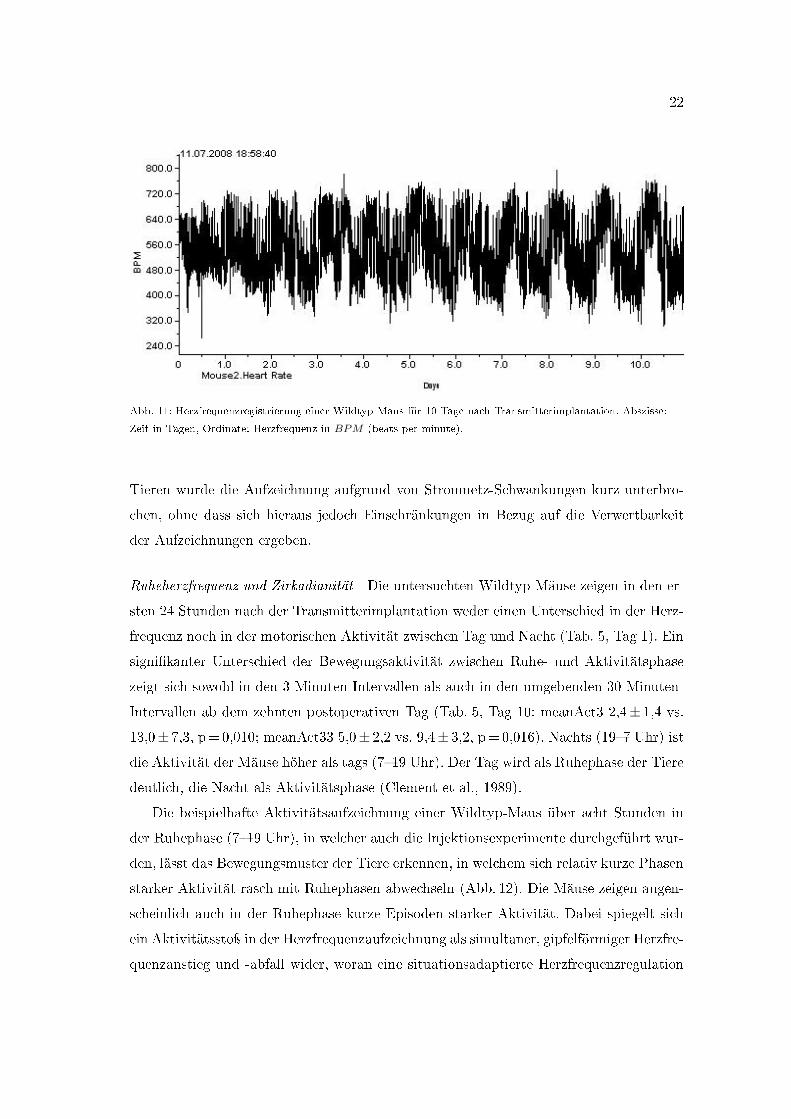
22
Abb. 11: Herzfrequenzregistrierung einer Wildtyp-Maus für 10 Tage nach Transmitterimplantation. Abszisse:
Zeit in Tagen, Ordinate: Herzfrequenz in BPM (beats per minute).
Tieren wurde die Aufzeichnung aufgrund von Stromnetz-Schwankungen kurz unterbro-
chen, ohne dass sich hieraus jedoch Einschränkungen in Bezug auf die Verwertbarkeit
der Aufzeichnungen ergeben.
Ruheherzfrequenz und Zirkadianität Die untersuchten Wildtyp-Mäuse zeigen in den er-
sten 24 Stunden nach der Transmitterimplantation weder einen Unterschied in der Herz-
frequenz noch in der motorischen Aktivität zwischen Tag und Nacht (Tab. 5, Tag 1). Ein
signi�kanter Unterschied der Bewegungsaktivität zwischen Ruhe- und Aktivitätsphase
zeigt sich sowohl in den 3-Minuten-Intervallen als auch in den umgebenden 30-Minuten-
Intervallen ab dem zehnten postoperativen Tag (Tab. 5, Tag 10: meanAct3 2,4± 1,4 vs.
13,0± 7,3, p=0,010; meanAct33 5,0± 2,2 vs. 9,4± 3,2, p=0,016). Nachts (19�7 Uhr) ist
die Aktivität der Mäuse höher als tags (7�19 Uhr). Der Tag wird als Ruhephase der Tiere
deutlich, die Nacht als Aktivitätsphase (Clement et al., 1989).
Die beispielhafte Aktivitätsaufzeichnung einer Wildtyp-Maus über acht Stunden in
der Ruhephase (7�19 Uhr), in welcher auch die Injektionsexperimente durchgeführt wur-
den, lässt das Bewegungsmuster der Tiere erkennen, in welchem sich relativ kurze Phasen
starker Aktivität rasch mit Ruhephasen abwechseln (Abb. 12). Die Mäuse zeigen augen-
scheinlich auch in der Ruhephase kurze Episoden starker Aktivität. Dabei spiegelt sich
ein Aktivitätsstoÿ in der Herzfrequenzaufzeichnung als simultaner, gipfelförmiger Herzfre-
quenzanstieg und -abfall wider, woran eine situationsadaptierte Herzfrequenzregulation
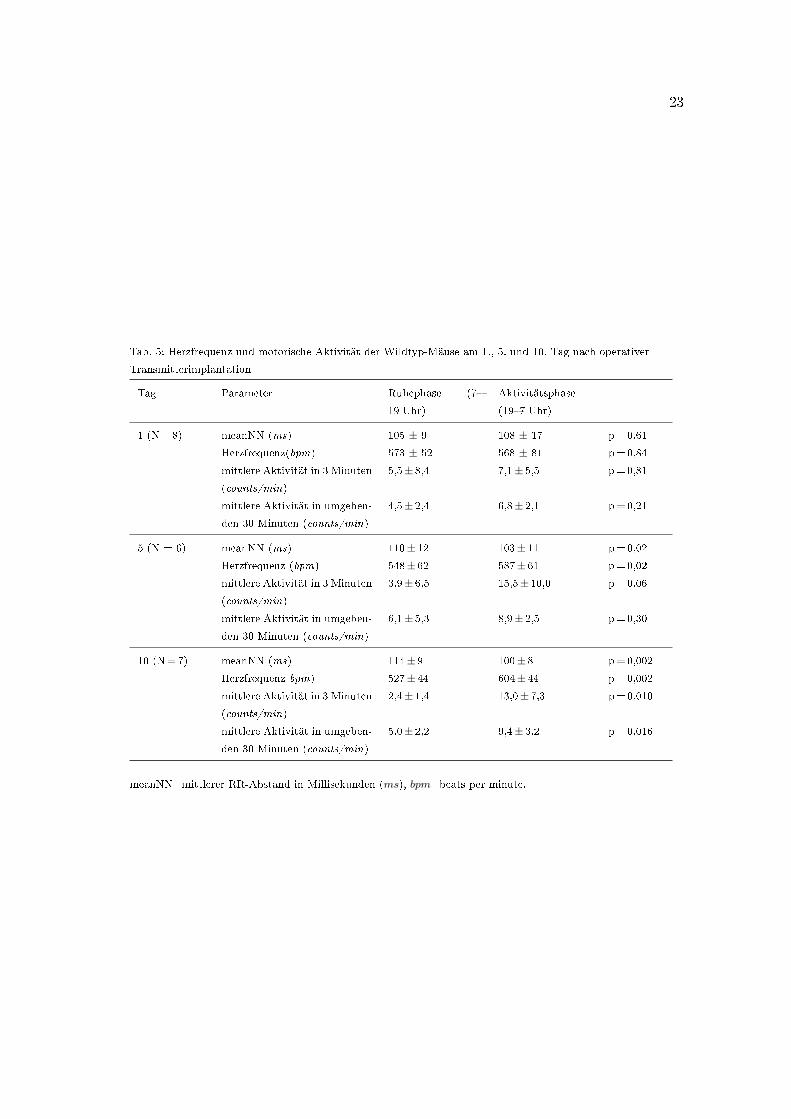
23
Tab. 5: Herzfrequenz und motorische Aktivität der Wildtyp-Mäuse am 1., 5. und 10. Tag nach operativer
Transmitterimplantation
Tag Parameter Ruhephase (7�
19 Uhr)
Aktivitätsphase
(19�7 Uhr)
1 (N=8) meanNN (ms) 105 ± 9 108 ± 17 p=0,61
Herzfrequenz(bpm) 573 ± 52 568 ± 81 p=0,84
mittlere Aktivität in 3 Minuten
(counts/min)
5,5± 8,4 7,1± 5,5 p=0,81
mittlere Aktivität in umgeben-
den 30 Minuten (counts/min)
4,5± 2,4 6,8± 2,1 p=0,21
5 (N = 6) meanNN (ms) 110± 12 103± 11 p=0,02
Herzfrequenz (bpm) 548± 62 587± 61 p=0,02
mittlere Aktivität in 3 Minuten
(counts/min)
3,9± 6,5 15,5± 10,0 p=0,06
mittlere Aktivität in umgeben-
den 30 Minuten (counts/min)
6,1± 5,3 8,9± 2,5 p=0,30
10 (N=7) meanNN (ms) 114± 9 100± 8 p=0,002
Herzfrequenz bpm) 527± 44 604± 44 p=0,002
mittlere Aktivität in 3 Minuten
(counts/min)
2,4± 1,4 13,0± 7,3 p=0,010
mittlere Aktivität in umgeben-
den 30 Minuten (counts/min)
5,0± 2,2 9,4± 3,2 p=0,016
meanNN=mittlerer RR-Abstand in Millisekunden (ms), bpm=beats per minute.
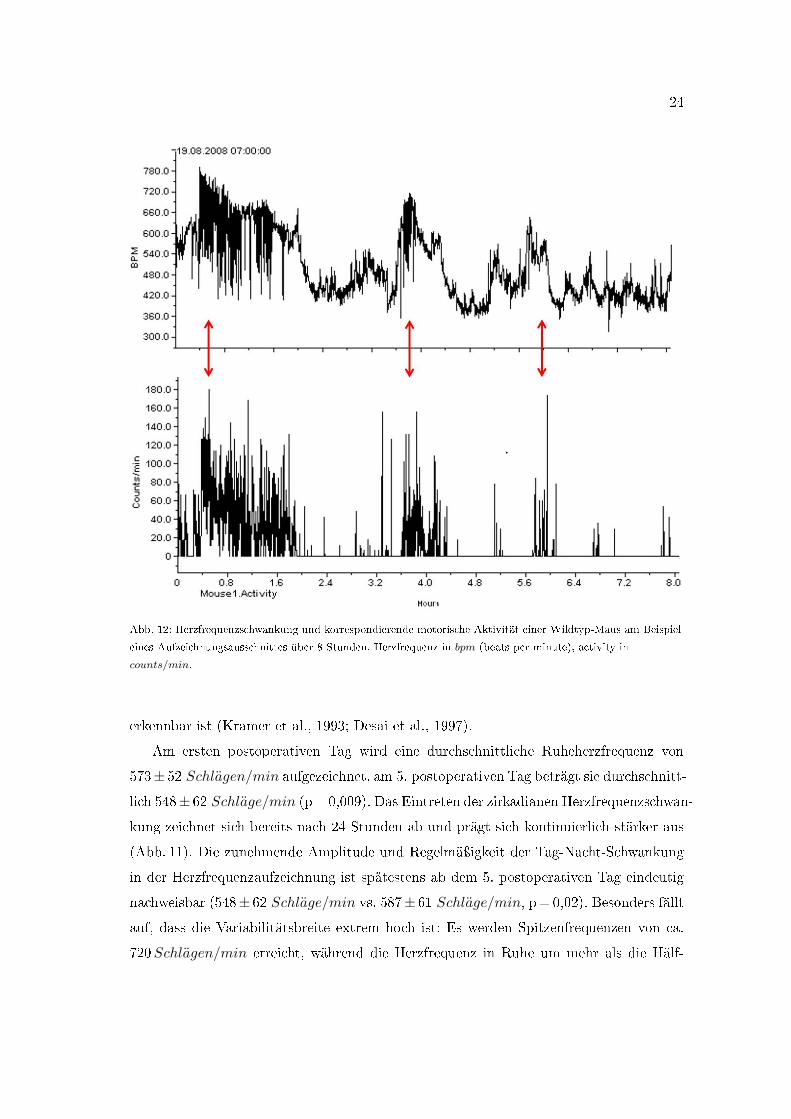
24
Abb. 12: Herzfrequenzschwankung und korrespondierende motorische Aktivität einer Wildtyp-Maus am Beispiel
eines Aufzeichnungsausschnittes über 8 Stunden. Herzfrequenz in bpm (beats per minute), activity in
counts/min.
erkennbar ist (Kramer et al., 1993; Desai et al., 1997).
Am ersten postoperativen Tag wird eine durchschnittliche Ruheherzfrequenz von
573± 52 Schlagen/min aufgezeichnet, am 5. postoperativen Tag beträgt sie durchschnitt-
lich 548± 62 Schlage/min (p=0,009). Das Eintreten der zirkadianen Herzfrequenzschwan-
kung zeichnet sich bereits nach 24 Stunden ab und prägt sich kontinuierlich stärker aus
(Abb. 11). Die zunehmende Amplitude und Regelmäÿigkeit der Tag-Nacht-Schwankung
in der Herzfrequenzaufzeichnung ist spätestens ab dem 5. postoperativen Tag eindeutig
nachweisbar (548± 62 Schlage/min vs. 587± 61 Schlage/min, p= 0,02). Besonders fällt
auf, dass die Variabilitätsbreite extrem hoch ist: Es werden Spitzenfrequenzen von ca.
720Schlagen/min erreicht, während die Herzfrequenz in Ruhe um mehr als die Hälf-
25
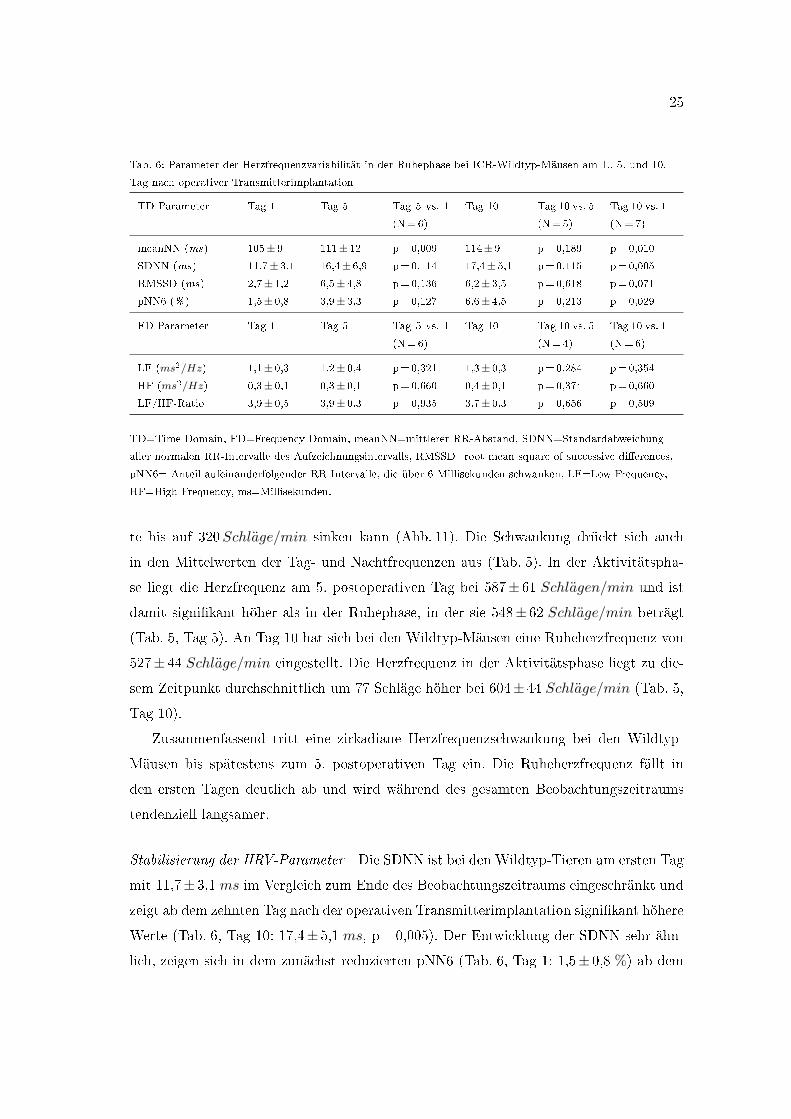
Tab. 6: Parameter der Herzfrequenzvariabilität in der Ruhephase bei ICR-Wildtyp-Mäusen am 1., 5. und 10.
Tag nach operativer Transmitterimplantation
TD Parameter Tag 1 Tag 5 Tag 5 vs. 1
(N=6)
Tag 10 Tag 10 vs. 5
(N=5)
Tag 10 vs. 1
(N=7)
meanNN (ms) 105± 9 111± 12 p=0,009 114± 9 p=0,189 p=0,010
SDNN (ms) 11,7± 3,1 16,4± 6,9 p=0,114 17,4± 5,1 p=0,115 p=0,005
RMSSD (ms) 2,7± 1,2 6,5± 4,8 p=0,136 6,2± 3,5 p=0,618 p=0,071
pNN6 (% ) 1,5± 0,8 3,9± 3,3 p=0,127 6,6± 4,5 p=0,213 p=0,029
FD Parameter Tag 1 Tag 5 Tag 5 vs. 1
(N=6)
Tag 10 Tag 10 vs. 5
(N=4)
Tag 10 vs. 1
(N=6)
LF (ms2/Hz) 1,1± 0,3 1,2± 0,4 p=0,321 1,3± 0,3 p=0,284 p=0,354
HF (ms2/Hz) 0,3± 0,1 0,3± 0,1 p=0,660 0,4± 0,1 p=0,374 p=0,660
LF/HF-Ratio 3,9± 0,5 3,9± 0,3 p=0,935 3,7± 0,3 p=0,656 p=0,509
TD=Time Domain, FD=Frequency Domain, meanNN=mittlerer RR-Abstand, SDNN=Standardabweichung
aller normalen RR-Intervalle des Aufzeichnungsintervalls, RMSSD=root mean square of successive di�erences,
pNN6= Anteil aufeinanderfolgender RR-Intervalle, die über 6 Millisekunden schwanken, LF=Low Frequency,
HF=High Frequency, ms=Millisekunden.
te bis auf 320Schlage/min sinken kann (Abb. 11). Die Schwankung drückt sich auch
in den Mittelwerten der Tag- und Nachtfrequenzen aus (Tab. 5). In der Aktivitätspha-
se liegt die Herzfrequenz am 5. postoperativen Tag bei 587± 61 Schlagen/min und ist
damit signi�kant höher als in der Ruhephase, in der sie 548± 62 Schlage/min beträgt
(Tab. 5, Tag 5). An Tag 10 hat sich bei den Wildtyp-Mäusen eine Ruheherzfrequenz von
527± 44 Schlage/min eingestellt. Die Herzfrequenz in der Aktivitätsphase liegt zu die-
sem Zeitpunkt durchschnittlich um 77 Schläge höher bei 604± 44 Schlage/min (Tab. 5,
Tag 10).
Zusammenfassend tritt eine zirkadiane Herzfrequenzschwankung bei den Wildtyp-
Mäusen bis spätestens zum 5. postoperativen Tag ein. Die Ruheherzfrequenz fällt in
den ersten Tagen deutlich ab und wird während des gesamten Beobachtungszeitraums
tendenziell langsamer.
Stabilisierung der HRV-Parameter Die SDNN ist bei den Wildtyp-Tieren am ersten Tag
mit 11,7± 3,1 ms im Vergleich zum Ende des Beobachtungszeitraums eingeschränkt und
zeigt ab dem zehnten Tag nach der operativen Transmitterimplantation signi�kant höhere
Werte (Tab. 6, Tag 10: 17,4± 5,1 ms, p= 0,005). Der Entwicklung der SDNN sehr ähn-
lich, zeigen sich in dem zunächst reduzierten pNN6 (Tab. 6, Tag 1: 1,5± 0,8 %) ab dem
26
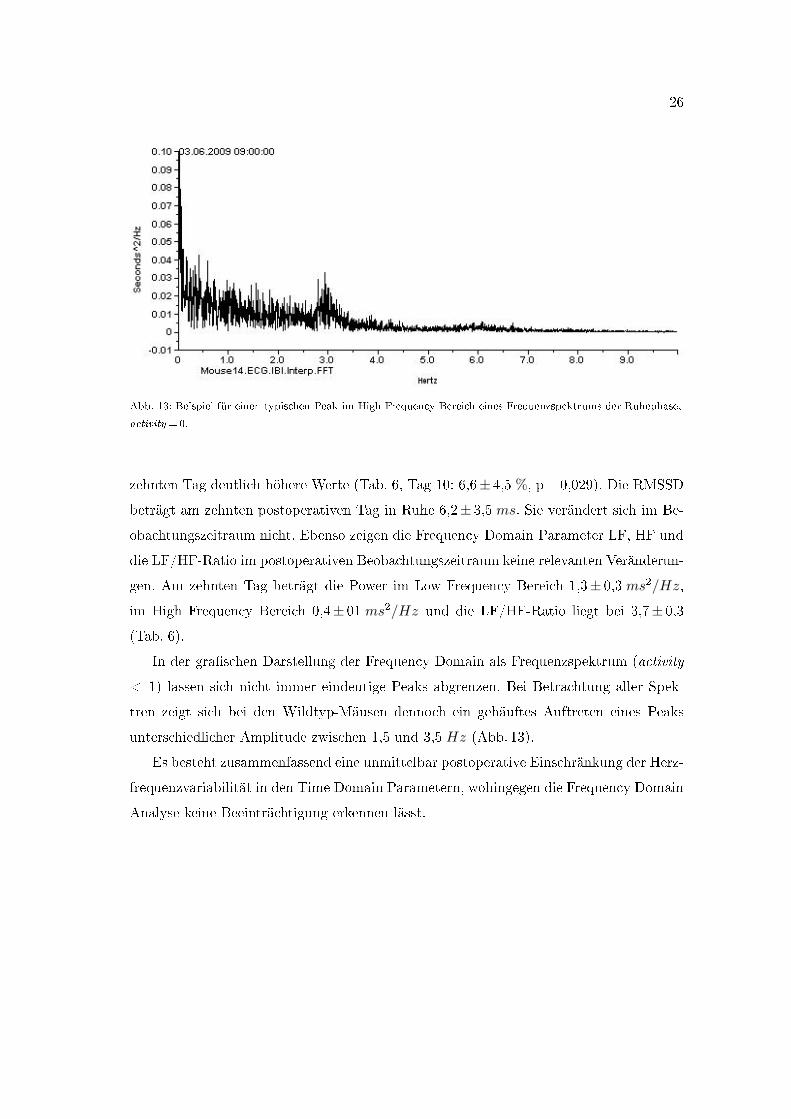
Abb. 13: Beispiel für einen typischen Peak im High Frequency Bereich eines Frequenzspektrums der Ruhephase,
activity =0.
zehnten Tag deutlich höhere Werte (Tab. 6, Tag 10: 6,6± 4,5 %, p= 0,029). Die RMSSD
beträgt am zehnten postoperativen Tag in Ruhe 6,2± 3,5 ms. Sie verändert sich im Be-
obachtungszeitraum nicht. Ebenso zeigen die Frequency Domain Parameter LF, HF und
die LF/HF-Ratio im postoperativen Beobachtungszeitraum keine relevanten Veränderun-
gen. Am zehnten Tag beträgt die Power im Low Frequency Bereich 1,3± 0,3 ms2/Hz,
im High Frequency Bereich 0,4± 01 ms2/Hz und die LF/HF-Ratio liegt bei 3,7± 0,3
(Tab. 6).
In der gra�schen Darstellung der Frequency Domain als Frequenzspektrum (activity
< 1) lassen sich nicht immer eindeutige Peaks abgrenzen. Bei Betrachtung aller Spek-
tren zeigt sich bei den Wildtyp-Mäusen dennoch ein gehäuftes Auftreten eines Peaks
unterschiedlicher Amplitude zwischen 1,5 und 3,5 Hz (Abb. 13).
Es besteht zusammenfassend eine unmittelbar postoperative Einschränkung der Herz-
frequenzvariabilität in den Time Domain Parametern, wohingegen die Frequency Domain
Analyse keine Beeinträchtigung erkennen lässt.
27
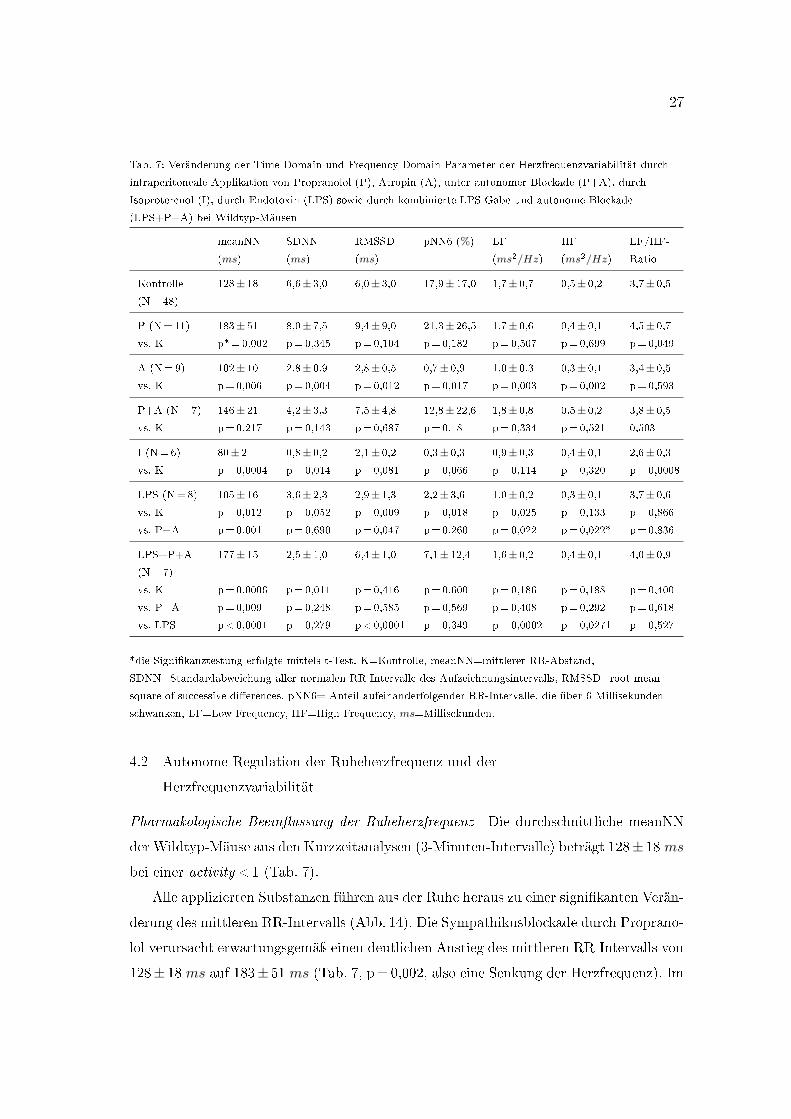
Tab. 7: Veränderung der Time Domain und Frequency Domain Parameter der Herzfrequenzvariabilität durch
intraperitoneale Applikation von Propranolol (P), Atropin (A), unter autonomer Blockade (P+A), durch
Isoproterenol (I), durch Endotoxin (LPS) sowie durch kombinierte LPS-Gabe und autonome Blockade
(LPS+P+A) bei Wildtyp-Mäusen
meanNN
(ms)
SDNN
(ms)
RMSSD
(ms)
pNN6 (%) LF
(ms2/Hz)
HF
(ms2/Hz)
LF/HF-
Ratio
Kontrolle 128± 18 6,6± 3,0 6,0± 3,0 17,9± 17,0 1,7± 0,7 0,5± 0,2 3,7± 0,5(N=48)
P (N=11) 183± 51 8,0± 7,5 9,4± 9,0 21,3± 26,5 1,7± 0,6 0,4± 0,1 4,5± 0,7vs. K p*=0,002 p=0,345 p=0,104 p=0,182 p=0,507 p=0,699 p=0,049
A (N=9) 102± 10 2,8± 0,9 2,8± 0,5 0,7± 0,9 1,0± 0,3 0,3± 0,1 3,4± 0,5vs. K p=0,006 p=0,004 p=0,012 p=0,017 p=0,003 p=0,002 p=0,593
P+A (N=7) 146± 21 4,2± 3,3 7,5± 4,8 12,8± 22,6 1,8± 0,8 0,5± 0,2 3,8± 0,5vs. K p=0,217 p=0,143 p=0,687 p=0,18 p=0,334 p=0,521 0,503
I (N=6) 80± 2 0,8± 0,2 2,1± 0,2 0,3± 0,3 0,9± 0,3 0,4± 0,1 2,6± 0,3vs. K p=0,0004 p=0,014 p=0,081 p=0,066 p=0,114 p=0,320 p=0,0008
LPS (N=8) 105± 16 3,6± 2,3 2,9± 1,3 2,2± 3,6 1,0± 0,2 0,3± 0,1 3,7± 0,6vs. K p=0,012 p=0,052 p=0,009 p=0,018 p=0,025 p=0,133 p=0,866
vs. P+A p=0,001 p=0,690 p=0,047 p=0,260 p=0,022 p=0,022* p=0,836
LPS+P+A
(N=7)
177± 15 2,5± 1,0 6,4± 1,0 7,1± 12,4 1,6± 0,2 0,4± 0,1 4,0± 0,9
vs. K p=0,0006 p=0,011 p=0,416 p=0,600 p=0,186 p=0,188 p=0,400
vs. P+A p=0,009 p=0,248 p=0,585 p=0,569 p=0,408 p=0,292 p=0,618
vs. LPS p<0,0001 p=0,279 p<0,0001 p=0,349 p=0,0002 p=0,0271 p=0,527
*die Signi�kanztestung erfolgte mittels t-Test. K=Kontrolle, meanNN=mittlerer RR-Abstand,
SDNN=Standardabweichung aller normalen RR-Intervalle des Aufzeichnungsintervalls, RMSSD=root mean
square of successive di�erences, pNN6= Anteil aufeinanderfolgender RR-Intervalle, die über 6 Millisekunden
schwanken, LF=Low Frequency, HF=High Frequency, ms=Millisekunden.
4.2 Autonome Regulation der Ruheherzfrequenz und der
Herzfrequenzvariabilität
Pharmakologische Beein�ussung der Ruheherzfrequenz Die durchschnittliche meanNN
der Wildtyp-Mäuse aus den Kurzzeitanalysen (3-Minuten-Intervalle) beträgt 128± 18 ms
bei einer activity <1 (Tab. 7).
Alle applizierten Substanzen führen aus der Ruhe heraus zu einer signi�kanten Verän-
derung des mittleren RR-Intervalls (Abb. 14). Die Sympathikusblockade durch Proprano-
lol verursacht erwartungsgemäÿ einen deutlichen Anstieg des mittleren RR-Intervalls von
128± 18 ms auf 183± 51 ms (Tab. 7, p=0,002, also eine Senkung der Herzfrequenz). Im
28
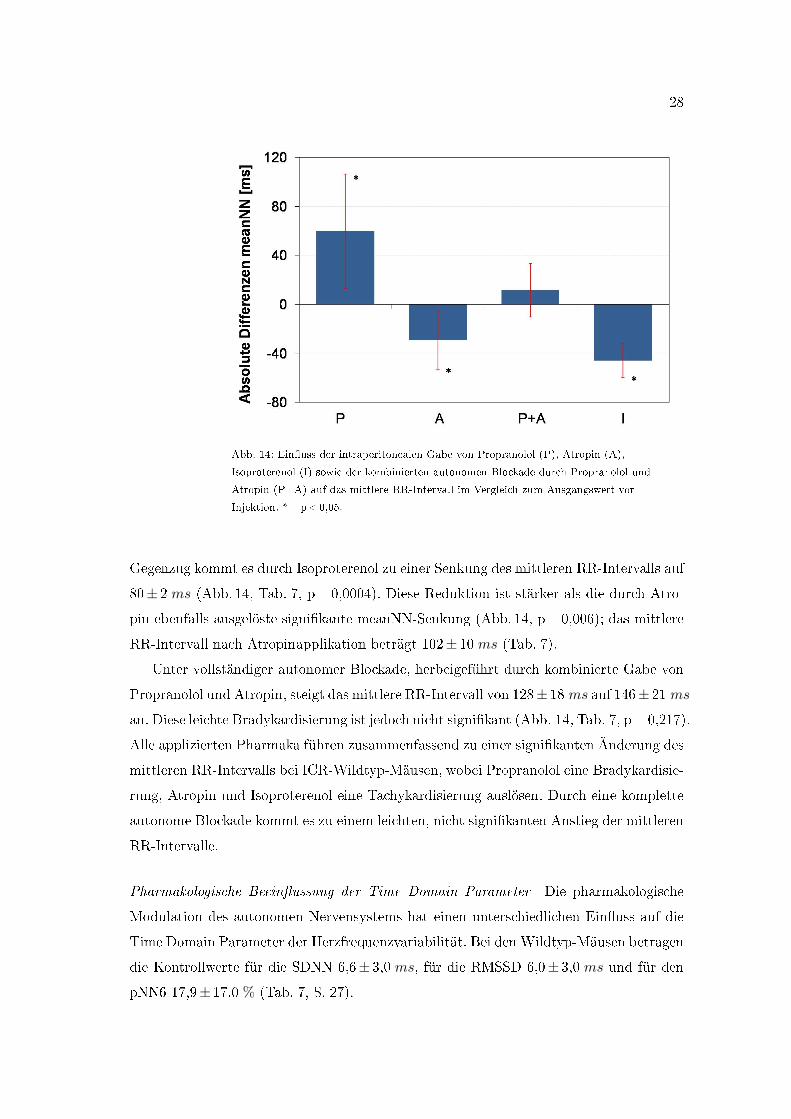
Abb. 14: Ein�uss der intraperitonealen Gabe von Propranolol (P), Atropin (A),
Isoproterenol (I) sowie der kombinierten autonomen Blockade durch Propranolol und
Atropin (P+A) auf das mittlere RR-Intervall im Vergleich zum Ausgangswert vor
Injektion. *=p<0,05.
Gegenzug kommt es durch Isoproterenol zu einer Senkung des mittleren RR-Intervalls auf
80± 2 ms (Abb. 14, Tab. 7, p=0,0004). Diese Reduktion ist stärker als die durch Atro-
pin ebenfalls ausgelöste signi�kante meanNN-Senkung (Abb. 14, p=0,006); das mittlere
RR-Intervall nach Atropinapplikation beträgt 102± 10 ms (Tab. 7).
Unter vollständiger autonomer Blockade, herbeigeführt durch kombinierte Gabe von
Propranolol und Atropin, steigt das mittlere RR-Intervall von 128± 18 ms auf 146± 21 ms
an. Diese leichte Bradykardisierung ist jedoch nicht signi�kant (Abb. 14, Tab. 7, p=0,217).
Alle applizierten Pharmaka führen zusammenfassend zu einer signi�kanten Änderung des
mittleren RR-Intervalls bei ICR-Wildtyp-Mäusen, wobei Propranolol eine Bradykardisie-
rung, Atropin und Isoproterenol eine Tachykardisierung auslösen. Durch eine komplette
autonome Blockade kommt es zu einem leichten, nicht signi�kanten Anstieg der mittleren
RR-Intervalle.
Pharmakologische Beein�ussung der Time Domain Parameter Die pharmakologische
Modulation des autonomen Nervensystems hat einen unterschiedlichen Ein�uss auf die
Time Domain Parameter der Herzfrequenzvariabilität. Bei den Wildtyp-Mäusen betragen
die Kontrollwerte für die SDNN 6,6± 3,0 ms, für die RMSSD 6,0± 3,0 ms und für den
pNN6 17,9± 17,0 % (Tab. 7, S. 27).
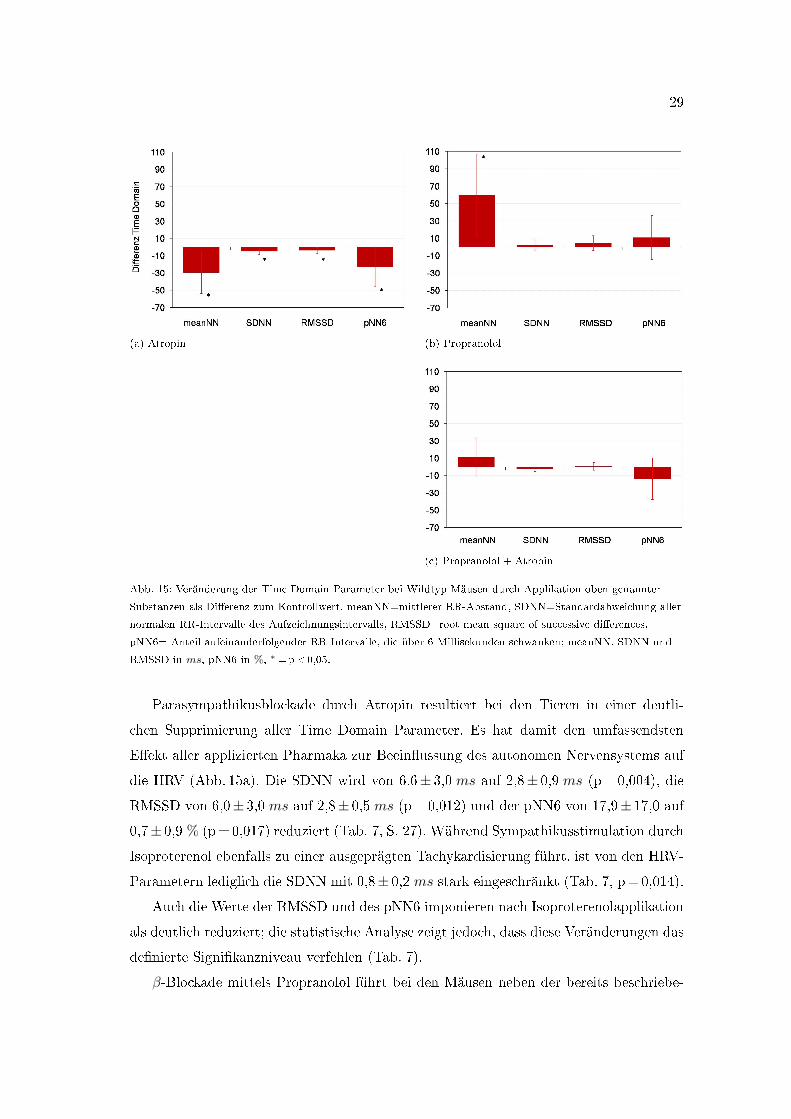
29
(a) Atropin (b) Propranolol
(c) Propranolol + Atropin
Abb. 15: Veränderung der Time Domain Parameter bei Wildtyp-Mäusen durch Applikation oben genannter
Substanzen als Di�erenz zum Kontrollwert. meanNN=mittlerer RR-Abstand, SDNN=Standardabweichung aller
normalen RR-Intervalle des Aufzeichnungsintervalls, RMSSD=root mean square of successive di�erences,
pNN6= Anteil aufeinanderfolgender RR-Intervalle, die über 6 Millisekunden schwanken; meanNN, SDNN und
RMSSD in ms, pNN6 in %, ∗=p<0,05.
Parasympathikusblockade durch Atropin resultiert bei den Tieren in einer deutli-
chen Supprimierung aller Time Domain Parameter. Es hat damit den umfassendsten
E�ekt aller applizierten Pharmaka zur Beein�ussung des autonomen Nervensystems auf
die HRV (Abb. 15a). Die SDNN wird von 6,6± 3,0 ms auf 2,8± 0,9 ms (p=0,004), die
RMSSD von 6,0± 3,0 ms auf 2,8± 0,5 ms (p=0,012) und der pNN6 von 17,9± 17,0 auf
0,7± 0,9 % (p=0,017) reduziert (Tab. 7, S. 27). Während Sympathikusstimulation durch
Isoproterenol ebenfalls zu einer ausgeprägten Tachykardisierung führt, ist von den HRV-
Parametern lediglich die SDNN mit 0,8± 0,2 ms stark eingeschränkt (Tab. 7, p=0,014).
Auch die Werte der RMSSD und des pNN6 imponieren nach Isoproterenolapplikation
als deutlich reduziert; die statistische Analyse zeigt jedoch, dass diese Veränderungen das
de�nierte Signi�kanzniveau verfehlen (Tab. 7).
β-Blockade mittels Propranolol führt bei den Mäusen neben der bereits beschriebe-

30
nen Bradykardisierung zu keiner signi�kanten Veränderung der Time Domain Parameter
(vgl. Tab. 7). Ebenso verursacht die autonome Blockade durch kombinierte Gabe von
Propranolol und Atropin keine signi�kanten Änderungen in der Time Domain (Abb. 15c,
Tab. 7).
Zusammenfassend ist die Atropingabe bei den Wildtyp-Mäusen mit einer starken
Supprimierung aller Time Domain Parameter der Herzfrequenzvariabilität verbunden,
während Propranolol und die kombinierte autonome Blockade keine Veränderungen be-
wirken. Isoproterenolapplikation führt zu einer leichten Einschränkung der HRV mit einer
signi�kanten Reduktion der SDNN.
Pharmakologische Beein�ussung der Frequency Domain Parameter In Ruhe wird die
Dominanz der Power (ms2) im Low Frequency (LF) Bereich gegenüber dem High Fre-
quency (HF) Bereich deutlich. Die LF Power beträgt bei denWildtyp-Tieren 1,7± 0,7 ms2,
die HF Power 0,5± 0,2 ms2, das Verhältnis der beiden wird durch die LF/HF-Ratio aus-
gedrückt und liegt in Ruhe bei 3,7± 0,5 (Tab. 7, S. 27).
Atropin führt zu einer signi�kanten Senkung der Power in beiden Frequenzabschnit-
ten (Tab. 7). Die LF wird von 1,7± 0,7 ms2 auf 1,0± 0,3 ms2 (p=0,003) und die HF von
0,5± 0,2 ms2 auf 0,3± 0,1 ms2 (p=0,002) reduziert. Die Blockierung des parasympathi-
schen Ein�usses durch Atropin führt zu einer Abschwächung der Power im gesamten Fre-
quenzspektrum, wobei die Wirkung auf die verschiedenen Bereiche gleichsinnig ist. Das
Verhältnis beider Frequenzbereiche zueinander, die LF/HF-Ratio, bleibt durch parasym-
pathische Modulation unbeein�usst (nach Atropinapplikation 3,4± 0,5 vs. Kontrollwert
3,7± 0,5, p=0,593). Der Parasympathikus ist somit entscheidend an der Entstehung der
Power in den untersuchten Frequenzbereichen bei Wildtyp-Mäusen beteiligt.
Die sympathische Modulation des Power Density Spektrums ist weniger durch eine
eindeutige Veränderung eines einzelnen Frequenz-Bereichs charakterisiert, als durch ei-
ne Verschiebung des Spektrums, die sich in der deutlichen Änderung der LF/HF-Ratio
zeigt. Durch β-Blockade wird die LF/HF-Ratio von 3,7± 0,5 auf 4,5± 0,7 angehoben
(Tab. 7, S. 27, p=0,049), durch sympathoadrenerge Erregung hingegen auf 2,6± 0,3 ge-
senkt (Tab. 7, S. 27, p=0,0008).
Atropin führt somit bei den untersuchten Wildtyp-Mäusen zu einer Einschränkung
der Frequency Domain in beiden untersuchten Frequenzbereichen. Der Ein�uss des Sym-
pathikus wird durch die LF/HF-Ratio charakterisiert. Kombinierte autonome Blockade
beein�usst die Frequency Domain nicht nachweisbar.
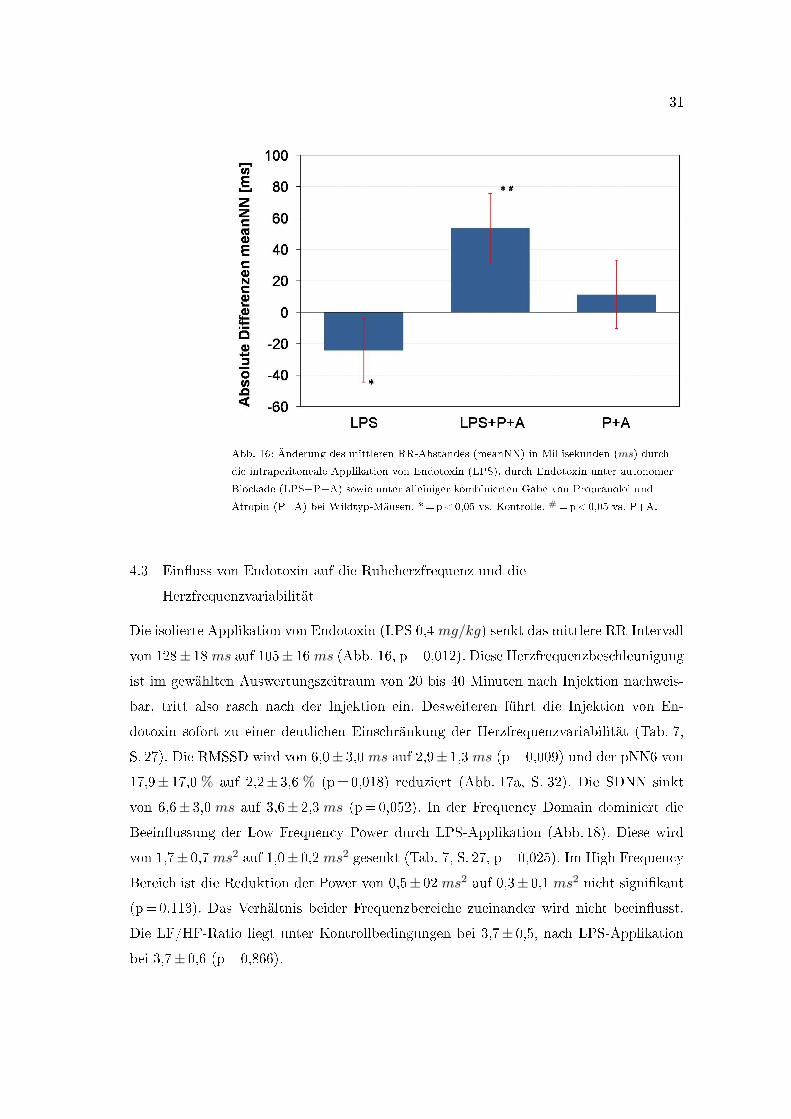
31
Abb. 16: Änderung des mittleren RR-Abstandes (meanNN) in Millisekunden (ms) durch
die intraperitoneale Applikation von Endotoxin (LPS), durch Endotoxin unter autonomer
Blockade (LPS+P+A) sowie unter alleiniger kombinierten Gabe von Propranolol und
Atropin (P+A) bei Wildtyp-Mäusen. *=p<0,05 vs. Kontrolle. #=p<0,05 vs. P+A.
4.3 Ein�uss von Endotoxin auf die Ruheherzfrequenz und die
Herzfrequenzvariabilität
Die isolierte Applikation von Endotoxin (LPS 0,4 mg/kg) senkt das mittlere RR-Intervall
von 128± 18 ms auf 105± 16 ms (Abb. 16, p=0,012). Diese Herzfrequenzbeschleunigung
ist im gewählten Auswertungszeitraum von 20 bis 40 Minuten nach Injektion nachweis-
bar, tritt also rasch nach der Injektion ein. Desweiteren führt die Injektion von En-
dotoxin sofort zu einer deutlichen Einschränkung der Herzfrequenzvariabilität (Tab. 7,
S. 27). Die RMSSD wird von 6,0± 3,0 ms auf 2,9± 1,3 ms (p=0,009) und der pNN6 von
17,9± 17,0 % auf 2,2± 3,6 % (p=0,018) reduziert (Abb. 17a, S. 32). Die SDNN sinkt
von 6,6± 3,0 ms auf 3,6± 2,3 ms (p=0,052). In der Frequency Domain dominiert die
Beein�ussung der Low Frequency Power durch LPS-Applikation (Abb. 18). Diese wird
von 1,7± 0,7 ms2 auf 1,0± 0,2 ms2 gesenkt (Tab. 7, S. 27, p=0,025). Im High Frequency
Bereich ist die Reduktion der Power von 0,5± 02 ms2 auf 0,3± 0,1 ms2 nicht signi�kant
(p=0,113). Das Verhältnis beider Frequenzbereiche zueinander wird nicht beein�usst.
Die LF/HF-Ratio liegt unter Kontrollbedingungen bei 3,7± 0,5, nach LPS-Applikation
bei 3,7± 0,6 (p=0,866).
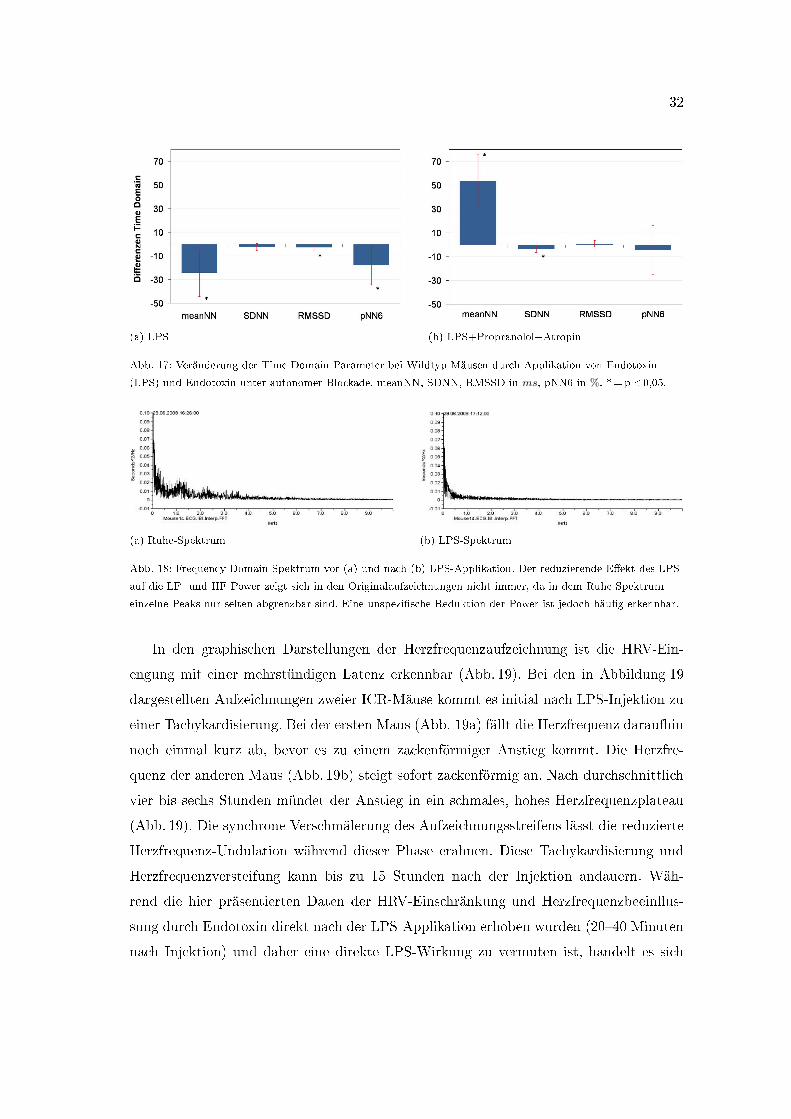
32
(a) LPS (b) LPS+Propranolol+Atropin
Abb. 17: Veränderung der Time Domain Parameter bei Wildtyp-Mäusen durch Applikation von Endotoxin
(LPS) und Endotoxin unter autonomer Blockade. meanNN, SDNN, RMSSD in ms, pNN6 in %. *=p<0,05.
(a) Ruhe-Spektrum (b) LPS-Spektrum
Abb. 18: Frequency Domain Spektrum vor (a) und nach (b) LPS-Applikation. Der reduzierende E�ekt des LPS
auf die LF- und HF Power zeigt sich in den Originalaufzeichnungen nicht immer, da in dem Ruhe-Spektrum
einzelne Peaks nur selten abgrenzbar sind. Eine unspezi�sche Reduktion der Power ist jedoch häu�g erkennbar.
In den graphischen Darstellungen der Herzfrequenzaufzeichnung ist die HRV-Ein-
engung mit einer mehrstündigen Latenz erkennbar (Abb. 19). Bei den in Abbildung 19
dargestellten Aufzeichnungen zweier ICR-Mäuse kommt es initial nach LPS-Injektion zu
einer Tachykardisierung. Bei der ersten Maus (Abb. 19a) fällt die Herzfrequenz daraufhin
noch einmal kurz ab, bevor es zu einem zackenförmiger Anstieg kommt. Die Herzfre-
quenz der anderen Maus (Abb. 19b) steigt sofort zackenförmig an. Nach durchschnittlich
vier bis sechs Stunden mündet der Anstieg in ein schmales, hohes Herzfrequenzplateau
(Abb. 19). Die synchrone Verschmälerung des Aufzeichnungsstreifens lässt die reduzierte
Herzfrequenz-Undulation während dieser Phase erahnen. Diese Tachykardisierung und
Herzfrequenzversteifung kann bis zu 15 Stunden nach der Injektion andauern. Wäh-
rend die hier präsentierten Daten der HRV-Einschränkung und Herzfrequenzbeein�us-
sung durch Endotoxin direkt nach der LPS-Applikation erhoben wurden (20�40 Minuten
nach Injektion) und daher eine direkte LPS-Wirkung zu vermuten ist, handelt es sich
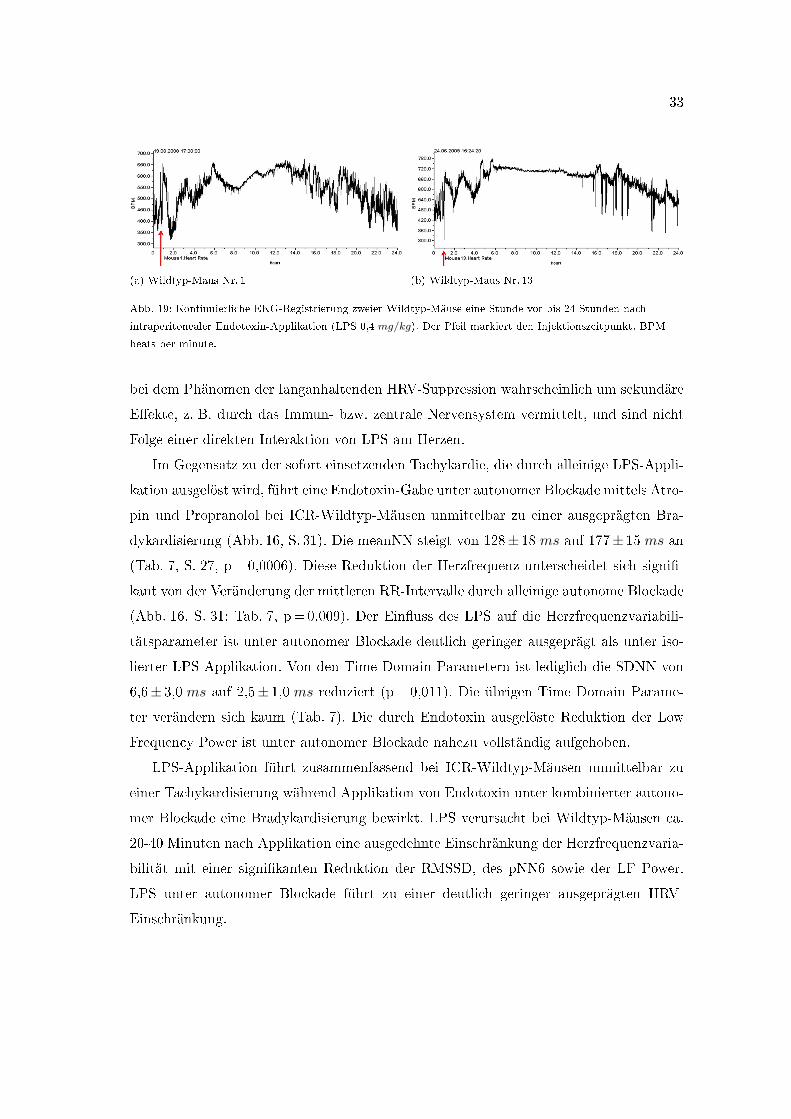
33
(a) Wildtyp-Maus Nr. 1 (b) Wildtyp-Maus Nr. 13
Abb. 19: Kontinuierliche EKG-Registrierung zweier Wildtyp-Mäuse eine Stunde vor bis 24 Stunden nach
intraperitonealer Endotoxin-Applikation (LPS 0,4 mg/kg). Der Pfeil markiert den Injektionszeitpunkt. BPM=
beats per minute.
bei dem Phänomen der langanhaltenden HRV-Suppression wahrscheinlich um sekundäre
E�ekte, z. B. durch das Immun- bzw. zentrale Nervensystem vermittelt, und sind nicht
Folge einer direkten Interaktion von LPS am Herzen.
Im Gegensatz zu der sofort einsetzenden Tachykardie, die durch alleinige LPS-Appli-
kation ausgelöst wird, führt eine Endotoxin-Gabe unter autonomer Blockade mittels Atro-
pin und Propranolol bei ICR-Wildtyp-Mäusen unmittelbar zu einer ausgeprägten Bra-
dykardisierung (Abb. 16, S. 31). Die meanNN steigt von 128± 18 ms auf 177± 15 ms an
(Tab. 7, S. 27, p=0,0006). Diese Reduktion der Herzfrequenz unterscheidet sich signi�-
kant von der Veränderung der mittleren RR-Intervalle durch alleinige autonome Blockade
(Abb. 16, S. 31; Tab. 7, p=0,009). Der Ein�uss des LPS auf die Herzfrequenzvariabili-
tätsparameter ist unter autonomer Blockade deutlich geringer ausgeprägt als unter iso-
lierter LPS-Applikation. Von den Time Domain Parametern ist lediglich die SDNN von
6,6± 3,0 ms auf 2,5± 1,0 ms reduziert (p=0,011). Die übrigen Time Domain Parame-
ter verändern sich kaum (Tab. 7). Die durch Endotoxin ausgelöste Reduktion der Low
Frequency Power ist unter autonomer Blockade nahezu vollständig aufgehoben.
LPS-Applikation führt zusammenfassend bei ICR-Wildtyp-Mäusen unmittelbar zu
einer Tachykardisierung während Applikation von Endotoxin unter kombinierter autono-
mer Blockade eine Bradykardisierung bewirkt. LPS verursacht bei Wildtyp-Mäusen ca.
20-40 Minuten nach Applikation eine ausgedehnte Einschränkung der Herzfrequenzvaria-
bilität mit einer signi�kanten Reduktion der RMSSD, des pNN6 sowie der LF Power.
LPS unter autonomer Blockade führt zu einer deutlich geringer ausgeprägten HRV-
Einschränkung.

34
5 Diskussion
5.1 Telemetrische Datengewinnung an Mäusen
Mit der Einführung kommerziell verfügbarer Telemetrie-Systeme und der damit verbun-
denen Möglichkeit, Operations- und Aufzeichnungstechniken sowie die telemetrische Da-
tengewinnung zu standardisieren und vergleichbar zu machen, ist es seit dem Ende der
1980er-Jahre gelungen, eine Vielzahl biometrischer Daten von wachen, sich frei bewe-
genden Labormäusen zu gewinnen (Kramer and Kinter, 2003). John G. Clement führte
1989 die ersten telemetrischen Aufzeichnungen der Bewegungsaktivität mittels eines solch
kommerziell verfügbaren Systems an männlichen ICR-Mäusen durch (Clement et al.,
1989). Die Bewegungsaktivität und Körpertemperatur seiner Tiere hatten sich vier Tage
nach dem operativen Eingri� stabilisiert.
Über die erste telemetrische Aufzeichnung eines Maus-EKGs und der Herzfrequenz
von Mäusen berichtete 1993 Klaas Kramer (Kramer et al., 1993). Er beobachtete eine
Normalisierung der Ruheherzfrequenz bereits einen Tag nach der operativen Systemim-
plantation, während die Normalisierung des Trink- und Fressverhaltens sowie des Be-
wegungsmusters nach vier Tagen eintrat. Auf die Notwendigkeit einer Stabilisierungs-
und Erholungsphase nach der initial durch die Operation und die Anästhesie ausgelö-
sten Stresssituation wiesen sowohl Clement als auch Kramer hin und empfahlen einen
Regenerationszeitraum von einer Woche.
Über die Vorteile der telemetrischen Datengewinnung besteht Einigkeit (Kramer and
Kinter, 2003; Hoyt et al., 2007). Mittels der Telemetrie ist es möglich, physiologische
Parameter sich frei bewegender Tiere unter Reduktion experimenteller Stressfaktoren zu
erfassen und damit möglichst realitätsnahe Messungen zu erhalten. Es werden jedoch
unterschiedliche Zeitpunkte für den Beginn der Experimente nach operativer Transmit-
terimplantation als günstig erachtet. Mit den experimentellen Durchführungen begannen
Studien 1 Tag (Desai et al., 1997), 3 bis 6 (Uechi et al., 1998), 3 bis 7 Tage (Gehrmann
et al., 2000; Ishii et al., 1996), 6 (Shusterman et al., 2002), 7 Tage (Clement et al., 1989),
10 (Thireau et al., 2008) oder 14 Tage (Baudrie et al., 2007) nach der Implantation der
Telemetrietransmitter.
Diese abweichenden Angaben ergeben sich aus der Nutzung unterschiedlicher Para-
meter, die zur Einschätzung der Beeinträchtigung der Tiere sowie der Stabilität der auf-
gezeichneten Daten herangezogen werden. Sie umfassen u. a. die Stabilisierung des Blut-

35
drucks, der Herzfrequenz, der Herzfrequenzvaiabilität (HRV) und des Körpergewichts,
das Eintreten eines zirkadianen Verhaltensmusters, das Putz-, Trink- und Fressverhal-
ten, die Bewegungsaktivität, das Schmerzverhalten sowie die Gewöhnung an den im-
plantierten Fremdkörper (Baumans et al., 2001; Leon et al., 2004). Weiterhin gelten
die Verwendung verschiedener Operationstechniken (Gehrmann et al., 2000) oder un-
terschiedlicher Materialien (Baumans et al., 2001), das unterschiedliche postoperative
Schmerz- und Tierhaltungs-Management (Leon et al., 2004), der Einsatz verschiedener
Mausstämme (beispielsweise ICR, C57BL/6J und FVB) oder di�erierende Auswertungs-
algorithmen als Grund für die unterschiedlichen Berichte. Einige Studien begründen die
Wahl der Länge der Stabilisierungsphase nicht (Joaquim et al., 2004; Witte et al., 2004).
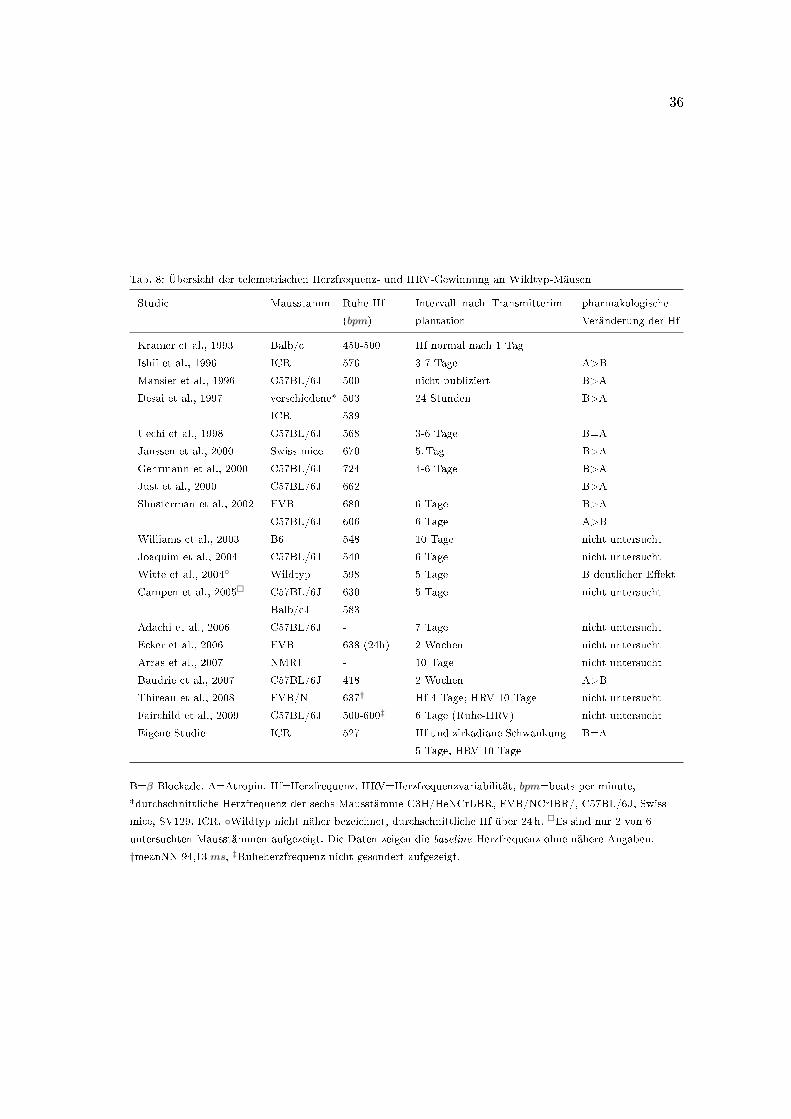
Eine Übersicht über Studien an verschiedenen Wildtyp-Stämmen, die die telemetrische
Herzfrequenz-Aufzeichnung mit einschlieÿen, bietet Tabelle 8.
Während Kramer über eine Stabilisierung der Herzfrequenzaufzeichnungen bereits am
1. Tag nach Transmitterimplantation berichtet (Kramer et al., 1993), beobachtet Ben J.
A. Janssen eine stabile 24-Stunden-Schwankung der Herzfrequenz ab dem 5. Tag (Janssen
et al., 2000). Thireau unterbreitete 2008 den Vorschlag eines standardisierten Verfahrens
zur Erfassung der Herzfrequenz und Herzfrequenzvariabilität bei Mäusen (Thireau et al.,
2008), dem in der vorliegenden Arbeit gefolgt wurde. In diesem Zusammenhang berichtet
er über eine Stabilisierung der Herzfrequenz am 4. Tag nach dem Eingri�. Durch die
operative Implantation des Telemetrietransmitters würde die Herzfrequenzvariabilität
hingegen zunächst überschätzt und zeige erst nach mindestens 10 Tagen eine ausreichende
Stabilisierung.
Die hier untersuchten ICR-Wildtyp-Mäuse zeigen eine signi�kante zirkadiane Herz-
frequenzschwankung ab dem 5. Tag nach der intraperitonealen Transmitterimplantation
mit hohen Herzfrequenzen in der Nacht und niedrigeren Herzfrequenzen am Tag. Die
zirkadiane Schwankung der motorischen Aktivität ist ab dem 10. postoperativen Tag
signi�kant.
Die Ruheherzfrequenz der hier untersuchten ICR-Wildtyp-Mäuse ist am 1. Tag nach
der Transmitter-Implantation, möglicherweise als Zeichen des operativen Stresses, er-
höht (Just et al., 2000). Sie sinkt signi�kant bis zum 5. Tag und liegt 10 Tage nach der
Transmitterimplantation in dem Spektrum der für Mäuse als typisch geltenden Ruhe-
herzfrequenzen von 450�550 Schlage/min (Desai et al., 1997; Hoyt et al., 2007).
Die Herzfrequenzvariabilität ist in den eigenen Untersuchungen direkt nach der Trans-
36
Tab. 8: Übersicht der telemetrischen Herzfrequenz- und HRV-Gewinnung an Wildtyp-Mäusen
Studie Mausstamm Ruhe-Hf
(bpm)
Intervall nach Transmitterim-
plantation
pharmakologische
Veränderung der Hf
Kramer et al., 1993 Balb/c 450-500 Hf normal nach 1 Tag
Ishii et al., 1996 ICR 576 3-7 Tage A>B
Mansier et al., 1996 C57BL/6J 500 nicht publiziert B>A
Desai et al., 1997 verschiedene* 503 24 Stunden B>A
ICR 539
Uechi et al., 1998 C57BL/6J 568 3-6 Tage B=A
Janssen et al., 2000 Swiss mice 670 5. Tag B>A
Gehrmann et al., 2000 C57BL/6J 724 4-6 Tage B>A
Just et al., 2000 C57BL/6J 662 B>A
Shusterman et al., 2002 FVB 680 6 Tage B>A
C57BL/6J 606 6 Tage A>B
Williams et al., 2003 B6 548 10 Tage nicht untersucht
Joaquim et al., 2004 C57BL/6J 540 6 Tage nicht untersucht
Witte et al., 2004◦ Wildtyp 598 5 Tage B deutlicher E�ekt
Campen et al., 2005� C57BL/6J 630 5 Tage nicht untersucht
Balb/cJ 583
Adachi et al., 2006 C57BL/6J - 7 Tage nicht untersucht
Ecker et al., 2006 FVB 638 (24h) 2 Wochen nicht untersucht
Arras et al., 2007 NMRI - 10 Tage nicht untersucht
Baudrie et al., 2007 C57BL/6J 418 2 Wochen A>B
Thireau et al., 2008 FVB/N 637† Hf 4 Tage; HRV 10 Tage nicht untersucht
Fairchild et al., 2009 C57BL/6J 500-600‡ 6 Tage (Ruhe-HRV) nicht untersucht
Eigene Studie ICR 527 Hf und zirkadiane Schwankung
5 Tage, HRV 10 Tage
B=A
B=β-Blockade, A=Atropin. Hf=Herzfrequenz, HRV=Herzfrequenzvariabilität, bpm=beats per minute,
*durchschnittliche Herzfrequenz der sechs Mausstämme C3H/HeNCrLBR, FVB/NCrlBR/, C57BL/6J, Swiss
mice, SV129, ICR. ◦Wildtyp nicht näher bezeichnet, durchschnittliche Hf über 24 h. �Es sind nur 2 von 6
untersuchten Mausstämmen aufgezeigt. Die Daten zeigen die baseline-Herzfrequenz ohne nähere Angaben.
†meanNN 94,13 ms, ‡Ruheherzfrequenz nicht gesondert aufgezeigt.

37
mitterimplantation entgegen den Beobachtungen der Arbeitsgruppe Thireaus, deutlich
eingeschränkt (Thireau et al., 2008). Dies deutet darauf hin, dass die HRV-Parameter
unmittelbar nach Transmitterimplantation nicht den physiologischen autonomen Tonus
der Tiere widerspiegeln, sondern deren postoperative Stress-Reaktion. Die SDNN und der
pNN6 zeigen erst ab dem 10.Tag nach Transmitterimplantation signi�kant höhere Werte.
Der Anstieg der HRV, vor allem des pNN6 deutet auf die zunehmende Bedeutung des
Parasympathikus in der Regulation der Ruheherzfrequenz mit der zunehmenden post-
operativen Erholung hin und lässt umgekehrt den Schluss zu, dass in der postoperativen
Phase zunächst ein stressbedingt erhöhter Sympathikotonus vorherrscht (Malliani et al.,
1994; Ecker et al., 2006; Arras et al., 2007).
Eine Komponente dieses Stresses kann u. a. Schmerz sein. Margarete Arras verweist
darauf, dass eine Tachykardisierung und HRV-Einschränkung bei leichten bis modera-
ten Schmerzen bei normal erscheinendem Verhalten und Aussehen der Tiere das einzige
Hinweiszeichen auf Schmerz sein kann (Arras et al., 2007). Obwohl die Herzfrequenz und
die HRV von vielen Faktoren beein�usst werden, besteht bei den untersuchten Tieren
trotz Analgesierung somit möglicherweise für mindestens 24 Stunden ein �Restschmerz�.
Ein intensiviertes Schmerzregime für die ersten drei postoperativen Tage sollte daher bei
zukünftigen Implantationen erwogen werden.
Die zirkadiane Schwankung der Herzfrequenz entsteht vor allem durch das konti-
nuierliche Absinken der Ruheherzfrequenz. Die Herzfrequenz in der Aktivitätsphase ist
im postoperativen Überwachungszeitraum durchgängig hoch: in Übereinstimmung mit
der Entwicklung der HRV zu Beginn möglicherweise als Korrelat des operativ beding-
ten Stresses, mit der steigenden Bewegungsaktivität zum 10.Tag hin mehr und mehr
als physiologisch situationsadaptiert. Diese Absenkung der Ruheherzfrequenz, die HRV-
Steigerung und die situationsadaptierte Herzfrequenzerhöhung können als Zeichen der
postoperativen Erholung der Tiere interpretiert werden. In Einklang mit Thireau und
Veronique Baudrie stabilisieren sich die Herzfrequenz und die zirkadianen Schwankung
bis zum 5. postoperativen Tag; die HRV-Normalisierung benötigt auch in den hier vorge-
stellten Experimenten die empfohlene Mindestdauer von 10 Tagen (Baudrie et al., 2007;
Thireau et al., 2008).
Lisa Leon verwies darauf, dass trotz der in ihrer Arbeit ebenfalls beobachteten Stabi-
lisierung der zirkadianen Bewegungsaktivität ab dem 5. postoperativen Tag die Norma-
lisierung einer Vielzahl von Körperfunktionen erst deutlich später einträte (Leon et al.,

38
2004). So normalisiere sich beispielsweise das Körpergewicht erst nach 15 Tagen. Frühere
Studien berichten über ein Wiedererlangen des Ausgangsgewichts nach 18�20 Tagen (Cle-
ment et al., 1989; Kramer et al., 1993). Baumans geht von einer vollständigen Erholung
der Tiere aus, wenn das Gewicht nach 14 Tagen wieder die Ausgangswerte erreicht hat
(Baumans et al., 2001). Die Normalisierung aller Körperfunktionen bei den untersuchten
Mäusen ist umso wichtiger, als dass eine Reduktion der Herzfrequenz und eine Erhöhung
der HRV u. a. durch verminderte Nahrungszufuhr ausgelöst werden können (Williams
et al., 2003). Der Nachweis des Eintritts zirkadianen Verhaltens reiche Leon zufolge da-
her nicht aus, um die postoperative Erholung zu beurteilen, auch die Herzfrequenz- und
HRV-Stabilisierung wären nur ein Hinweis (Arras et al., 2007). In den hier vorgestellten
Untersuchungen war die postoperative Entwicklung der Körperfunktionen nicht Gegen-
stand der Untersuchung. Mit dem Verweis auf die Arbeit Leons und Baumans wurden
nach der Stabilisierung der aufgezeichneten Parameter bis zum 10. Tag weitere 5 Ta-
ge zur Erholung der Körperfunktionen hinzugefügt (Baumans et al., 2001; Leon et al.,
2004). Da die Mäuse in diesem Zeitraum keine Verhaltensau�älligkeiten zeigten und der
Gewichtsverlust nach 15 Tagen deutlich unter 10 % lag, wurde davon ausgegangen, dass
sich die Tiere am Ende der 15 Tage von der Operation komplett erholt hatten.
5.2 Herzfrequenzvariabilität in Mensch und Maus � Rückschlüsse auf die Regulation
des autonomen Nervensystems
Die Herzfrequenzvariabilität (HRV) beschreibt die physiologische Schwankungsbreite des
Sinusrhythmus, die sich zum einen in einer adäquaten situationsadaptierten Herzfre-
quenzanpassung, zum anderen in der Variation der einzelnen, aufeinander folgenden Sinu-
serregungen als Reaktion auf interne und externe Ein�üsse in Ruhe ausdrückt (Abb. 20).
Sie ist das Ergebnis der direkten Modulation des Schrittmacherstroms If des Herzens
durch das autonome Nervensystem. In Interaktion mit dem zentralen Nervensystem,
Hormonen, Zytokinen und weiterer Mediatoren vermittelt es physiologische Regulati-
onsmechanismen im Körper, die sich in der HRV abbilden (Saykrs, 1973; Malik et al.,
1996). Die HRV als messbares Ausgangssignal eines komplexen Netzwerks entsteht somit
aus der intakten Kommunikation des Körpers mit seiner Umwelt und der Integrität des
Körpers selbst (Pincus, 1994). Eine Einschränkung der physiologischen Variationsbrei-
te der Herzfrequenz tritt beispielsweise mit zunehmendem Alter ein, zudem scheint es
eine geschlechtsspezi�sche Variation zu geben (Moodithaya and Avadhany, 2012). Eine

39
Abb. 20: Entstehungsmodell der Herzfrequenzvariabilität (HRV). Auswahl
an Ein�ussfaktoren und Studien. ZNS=Zentrales Nervensystem,
SK=Sinusknoten, PAN=peripheres autonomes Nervensystem,
RAAS=Renin-Angiotensin-Aldosteronsystem, �Akselrod et al., 1981.
*Furlan et al., 1990; Fallen and Kamath, 1995. •Saykrs, 1973; Pagani et al.,
1995. ‡Saykrs, 1973; La Rovere et al., 1995. #Williams et al., 2003.$Hottenrott et al., 2006. †Saykrs, 1973; Aysin and Aysin, 2006.§Moodithaya and Avadhany, 2012. **Pagani et al., 1986.
hochgradige Minderung der HRV hat pathologische Bedeutung und wird im Rahmen
verschiedener Krankheitsbilder beobachtet (z. B. Odemuyiwa, 1995; Casolo, 1995; Rech-
lin, 1995; Malik et al., 1996; Wang et al., 2000). Sie hat sich u. a. als starker negativer
Prädiktor für den klinischen Verlauf von Patienten mit Multiorgan-Dysfunktionssyndrom
herausgestellt (Schmidt et al., 2005).
Initiale Beobachtungen der Kopplung und gegenseitigen Beein�ussung physiologi-
scher Regulationskreise wurden bereits im 18. Jh. für Herzfrequenz und Atmung und
im 19. Jh. für Herzfrequenz und Blutdruck datiert (Berntson et al., 1997). Die Verbin-
dung von HRV und autonomen Nervensystem wurde Gary G. Berntson zufolge Anfang
des 20. Jh. erkannt, wobei die Bedeutung der vagalen Kontrolle hervorgehoben wurde.
Matthew N. Levy betonte 1971 die entscheidende Rolle zentraler autonomer Ein�üsse

40
für die Funktion des Herzens und deren komplexe Interaktion (Levy, 1971). Er stellte
dabei heraus, dass die parasympathische Regulation der Herzfunktion die sympathische
übergeordnet beein�ussen kann und dem N. vagus daher die dominierende Rolle in der
autonomen Kontrolle beim Menschen zukomme. Mit der Entwicklung der Spektralana-
lyse seit dem Ende des 19. Jh. und der Etablierung automatisierter, computergestützter
Verfahren der Fast Fourier Transformation vor allem ab den 1960er-Jahren (Kay and
Marple, 1981) wurde diese zur Untersuchung von Herzfrequenzschwankungen genutzt
und ein 1/f -Spektrum mit typischen Peaks beobachtet (Akselrod, 1995; Cerutti et al.,
1995). Vor dem Hintergrund der groÿen Bedeutung des autonomen Nervensystems in der
Herzfrequenzregulation stellten sich eine Vielzahl an Studien die Frage nach der Präsen-
tation der autonomen Partner in den HRV-Parametern (beispielsweise Chess et al., 1975;
Akselrod et al., 1981; Pomeranz et al., 1985; Pagani et al., 1986; Hayano et al., 1991).
Die HRV-Guidelines der Task Force of The European Society of Cardiology and The
Northern American Society of Pacing and Electrophysiology unterscheiden die zeitabhän-
gige Analyse (Time Domain) und die frequenzabhängige Analyse (Frequency Domain)
der HRV. In der Time Domain geben der mittlere RR-Abstand (meanNN) und die Stan-
dardabweichung der mittleren RR-Abstände (SDNN) einen allgemeinen Hinweis auf die
HRV und berücksichtigen die Langzeit- und die Kurzzeitvariabilität (Malik et al., 1996).
Die Höhe der Herzfrequenz kann dabei ein entscheidender Hinweis auf die Dominanz des
autonomen Partners geben � eine hohe Herzfrequenz ist unter physiologischen Bedin-
gungen Hinweis auf die Dominanz des Sympathikus, eine niedrige Frequenz auf die des
Parasympathikus (Malliani et al., 1994).
Über die Ruhe-meanNN kann die Reaktion der Herzfrequenz auf die experimentelle
Applikation von Pharmaka, die den autonomen Ein�uss am Herzen modi�zieren, abge-
schätzt werden (Shusterman et al., 2002). Die RMSSD (Wurzel aus dem Quadrat der
Summe der quadrierten Di�erenzen aufeinanderfolgender RR-Abstände) und der pNN50
(Anteil der aufeinanderfolgenden RR-Abstände, die über 50 ms di�erieren) präsentie-
ren beim Menschen vor allem den parasympathischen Ein�uss auf die Herzfrequenz und
mit diesem den Atemrhythmus (Malik et al., 1996). Die parasympathische Dominanz in
sämtlichen HRV-Parametern wurde ebenfalls beschrieben (Hayano et al., 1991).
Die Frequenz-Analyse der Herzfrequenzvariabilität zeigt mehrere typische Peaks, die
beim Menschen klassisch in die Low Frequency Domain von 0,04�0,15Hz und die High
Frequency Domain von 0,15�0,4Hz eingeteilt werden (Malik et al., 1996). Hinzu kommt

41
die Very Low Frequency im Bereich sehr niedriger Frequenzen (Cerutti et al., 1995). Die
High Frequency Power entsteht in Übereinstimmung vieler Studien beim Menschen durch
den Parasympathikus. Die vagale Präsentation in der High Frequency bei Menschen und
groÿen Säugetieren wurde mittels elektrischer Vagusstimulation (Akselrod et al., 1981),
durch Blockade muskarinerger Acetylcholinrezeptoren (Pomeranz et al., 1985; Hayano
et al., 1991) und durch Vagotomie (Malliani et al., 1991) gezeigt.
Über den Ein�uss der autonomen Partner auf die Entstehung des Low Frequency
Peaks bestehen unterschiedliche Meinungen. Viele Autoren verstehen die LF vorrangig
als sympathische Präsentation (Rimoldi et al., 1990; Malliani et al., 1991; Montano et al.,
1994; Pagani et al., 1986). Andere kommen zu dem Schluss, dass neben dem Sympathikus
auch der Parasympathikus an der Entstehung der LF Power beteiligt ist und sie daher
beide autonome Partner repräsentiert (Akselrod et al., 1981; Appel et al., 1989; Hayano
et al., 1991; Houle and Billman, 1999).
Die LF/HF-Ratio wird dem zufolge von einigen Autoren als Parameter interpretiert,
der die sympathovagale Balance widerspiegelt (Malliani et al., 1994), andere Autoren
halten die LF/HF-Ratio nicht uneingeschränkt für einen geeigneten Parameter zur Mes-
sung dieses Verhältnisses (Eckberg, 1997). Die VLF wird durch viele Regelkreise, wie
beispielsweise die Thermoregulation und die Plasma-Renin-Aktivität (Akselrod et al.,
1981) beein�usst. Aktuellere Studien beschreiben eine Reduktion der VLF als negativen
prognostischen Marker bei Sepsis (Schmidt et al., 2005).
Die Ruhe-HRV bei Menschen wird durch den parasympathischen Ein�uss dominiert
(Hayano et al., 1991). 1996 wurde die übergeordnete Bedeutung des Parasympathikus in
der Regulation der HRV in den HRV-Guidelines betont (Malik et al., 1996). Eine HRV-
Reduktion, die als negativer Marker für die Entwicklung vieler Erkrankungen und die
Entstehung von Komplikationen zunehmend an diagnostischer Bedeutung gewinnt, kann
als Zeichen einer herabgesetzten parasympathischen Aktivität beim Menschen interpre-
tiert werden (Malliani et al., 1994). Der Parasympathikus scheint somit für die Integrität
der Körperfunktionen eine entscheidende Bedeutung zu haben. Ein Verlust der Parasym-
pathikusaktivität bedeutet möglicherweise eine Unterbrechung der Interorgankommuni-
kation, die zu einer Entkopplung der Organfunktionen mit ungünstiger Prognose führen
kann (Schmidt et al., 2001).
Über die autonome Regulation der Ruheherzfrequenz und der Herzfrequenzvariabili-
tät bei Mäusen existieren viele Studien mit unterschiedlichen Ergebnissen und Schlussfol-

42
gerungen. Neben der Untersuchung von Mäusen mit Mutationen im sympathischen und
parasympathischen kardialen Signaltransduktionsweg (Uechi et al., 1998; Gehrmann et al.,
2002; Ecker et al., 2006), gewannen eine Vielzahl von Arbeiten ihre Erkenntnisse durch
pharmakologische Experimente an Wildtyp-Mäusen, mit deren Hilfe das autonome Ner-
vensystem moduliert wurde.
Véronique Baudrie schlussfolgerte beispielsweise aus einem relativ stärkeren Anstieg
der Herzfrequenz nach Atropin-Gabe im Gegensatz zu der weniger stark ausgeprägten
Senkung durch β-Blockade, dass der Parasympathikus bei Wildtyp-Mäusen in Ruhe do-
miniert (Baudrie et al., 2007). Diese Vermutung sei durch eine im Vergleich zu anderen
Arbeiten niedrigere Herzfrequenz der Mäuse gestützt (Tab. 8). Über einen gegenüber dem
β-Blocker stärkeren Atropine�ekt berichten ebenfalls Ishii und Shusterman (Ishii et al.,
1996; Shusterman et al., 2002). Auch eine ausgeglichene Reaktion auf pharmakologische
Sympathikus- und Parasympathikus-Blockade wurde beobachtet und auf einen ausgegli-
chenen autonomen Ruhetonus bei Wildtyp-Mäusen geschlossen (Uechi et al., 1998).
Aus der Mehrzahl der HRV-Untersuchungen an Wildtyp- und transgenen Mäusen
leitet sich ein sympathisch dominierter Ruhetonus ab (Ishii et al., 1996; Desai et al.,
1997; Gehrmann et al., 2000; Janssen et al., 2000; Just et al., 2000; Thireau et al.,
2008), der daher in der Literatur als für Mäuse charakteristisch beschrieben wird (Hoyt
et al., 2007). Dieser zeige sich in einer Herzfrequenzsenkung bei kombinierter autonomer
Blockade. Josef Gehrmann schloss auf das Vorherrschen des Sympathikus in der Ruhe-
herzfrequenz, da weder Atropin- noch Isoproterenol-Applikation zu einer Herzfrequenz-
erhöhung führt, wobei die Mäuse in seiner Arbeit eine vergleichsweise hohe Herzfrequenz
von 724Schlagen/min aufwiesen (Gehrmann et al., 2000). Bei Arnim Just bewirkte
Atropin-Applikation eine moderate Tachykardie und β-Blockade eine starke Bradykardie
bei einer Ruheherzfrequenz der Tiere von 662 Schlagen/min (Tab. 8, Just et al., 2000).
In der vorliegenden Untersuchung liegt die Ruheherzfrequenz der Wildtyp-Tiere in
dem beschriebenen, für Mäuse typischen Bereich (s. Tab. 8, Hoyt et al., 2007). In den
Injektionsexperimenten führt β-adrenerge Blockade mittels Propranolol zu einer deutli-
chen Herzfrequenzsenkung, während Atropin eine ebenfalls deutliche Tachykardisierung
bewirkt. Kombinierte autonome Blockade senkt die Herzfrequenz leicht, jedoch nicht
signi�kant. Dies deutet in Einklang mit Uechi auf einen ausgeglichenen autonomen Ein-
�uss auf die Ruheherzfrequenz bei den hier untersuchten ICR-Wildtyp-Mäusen hin (Uechi
et al., 1998).

43
Die Herzfrequenzvariabilität bei Mäusen wird entsprechend der HRV-Messung beim
Menschen und bei groÿen Säugetieren mit analog de�nierten Parametern bestimmt (Gehr-
mann et al., 2000; Thireau et al., 2008). Die Time Domain Parameter meanNN, SDNN
und RMSSD werden gleichartig errechnet. Dem pNN50 beim Menschen entspricht der
pNN6 bei Mäusen, der durch Atropin-Applikation bei Shusterman und Thireau sowie in
der vorliegenden Arbeit nahezu vollständig aufgehoben wird (Shusterman et al., 2002;
Thireau et al., 2008).
Das Frequency Domain Spektrum der HRV bei Mäusen wird ungefähr mit dem Zehn-
fachen des menschlichen Frequenzspektrums angesetzt (Just et al., 2000). Die Zuordnung
der verschiedenen Frequenzbereiche zu den konkreten Komponenten des autonomen Ner-
vensystems bei Mäusen ist uneinheitlich, ebenso die Abgrenzung der Bereiche (Baudrie
et al., 2007, Stauss, 2007). Die hier gewählte Grenzsetzung orientiert sich an den Emp-
fehlungen von Thireau zur standardisierten, telemetrischen Datengewinnung bei Mäusen
(Thireau et al., 2008). In Anlehnung an Vladimir Shusterman und Dwain L. Eckberg
wurden die LF und HF in absoluten Einheiten (ms2) ermittelt (Eckberg, 1997; Shu-
sterman et al., 2002). In Einklang mit Uechi und Shusterman überwiegt in den eigenen
Untersuchungen bei Wildtyp-Mäusen in Ruhe die LF Power gegenüber der HF Power;
die LF/HF-Ratio liegt bei 3,7.
Atropin-Applikation hat bei Wildtyp-Mäusen den deutlichsten E�ekt auf die HRV
und führt zu einer signi�kanten Reduktion aller Parameter. In der Frequency Domain
bewirkt die Substanz eine gleichgerichtete Einschränkung der Power im Low Frequency-
und High Frequency-Bereich ohne nennenswerte Beein�ussung der LF/HF-Ratio. Josef
Gehrmann beobachtete nach Atropin-Applikation bei Wildtyp-Mäusen einen komplett
reduzierenden E�ekt auf alle Time Domain Parameter, die LF und HF Power und die
LF/HF-Ratio (Gehrmann et al., 2000). Armin Just berichtet über ein Absinken der
HRV in allen Frequenzbereichen nach Atropin-Applikation mit abnehmender Intensität
< 1 Hz (Just et al., 2000), Véronique Baudrie über eine Einschränkung des gesamten
Frequenzspektrums mit dem stärksten E�ekt im LF Bereich (< 2,5 Hz, Baudrie et al.,
2007 ). Atropin-Applikation führt zu einer Einschränkung der LF und HF Power bei Keiji
Ishii und Thireau (Ishii et al., 1996; Thireau et al., 2008).
Aus diesen Beobachtungen kann auf den entscheidenden Ein�uss des Parasympathi-
kus in der Entstehung der HRV allgemein und insbesondere im gesamten Frequenzspek-
trum der Frequenzanalyse geschlossen werden. Über die komplette Reduktion der HRV

44
durch Atropin berichten in der Literatur viele Studien einstimmig (Ishii et al., 1996;
Gehrmann et al., 2000; Just et al., 2000; Baudrie et al., 2007; Thireau et al., 2008). Die
entscheidende Bedeutung des Parasympathikus in der physiologischen Regulation der
HRV bei Mäusen und damit die Ähnlichkeit zu der autonomen Modulation der HRV
beim Menschen kann somit als gesichert angesehen werden.
Eine Tachykardisierung per se ist in der Lage, die Total Power der Frequency Domain
zu reduzieren, was die gleichzeitige Einschränkung der LF und HF durch Atropin erklären
könnte (Pomeranz et al., 1985). Dass in den vorliegenden Experimenten die Tachykardi-
sierung allein die HRV-Einschränkung nicht erklären kann, verdeutlichen die Applikatio-
nen von Isoproterenol, welche bei Wildtyp-Mäusen eine ausgeprägten Tachykardisierung
ohne signiki�kante Beein�ussung der absoluten LF und HF Power hervorrufen.
Über den Ein�uss des Sympathikus auf die Herzfrequenzvariabilität bei Mäusen be-
stehen � analog der Diskussion beim Menschen � verschiedene Ansichten. In der vor-
liegenden Arbeit haben die Propranolol-Applikationen bei den Wildtyp-Mäusen weder
einen relevanten E�ekt auf die Time Domain Parameter noch auf die LF oder HF. Dies
ist vergleichbar mit Berichten am Mensch (Hayano et al., 1991) sowie anderen Studien
an der Maus (Ishii et al., 1996; Just et al., 2000; Baudrie et al., 2007). Isoproterenol
reduziert, wie auch in humanen Studien beobachtet, die SDNN signi�kant. Eine einge-
schränkte SDNN könnte demnach auf einen reduzierten parasympathischen Tonus oder
auf einen erhöhten Sympathikotonus hinweisen (Malliani et al., 1994).
Bei Mäusen wurde, wie oben beschrieben, eine gleichgerichtete Senkung der Power
im LF und im HF Bereich durch Parasympathikus-Blockade berichtet. Die LF/HF-Ratio
ist daher in Einklang mit Gehrmann, Ecker und Just nicht unbedingt als Marker der
sympathovagalen Balance bei Mäusen zu interpretieren. In der vorliegenden Arbeit cha-
rakterisiert die LF/HF-Ratio den Ein�uss des Sympathikus auf die Frequency Domain
der Herzfrequenzanalyse bei Wildtyp-Mäusen adäquat: Sympathikusstimulation redu-
ziert den Parameter, β-Blockade erhöht ihn. Auch diese Beobachtungen können mit bis-
herigen Studienergebnissen in Einklang gebracht werden. Bei Gehrmann kommt es durch
Propranolol-Applikation zu einer Anhebung der LF/HF-Ratio (Gehrmann et al., 2000).
Über eine deutlich reduzierte LF/HF-Ratio berichtet Uechi bei Mäusen mit einer Überex-
pression des kardialen Gsα-Proteins (Uechi et al., 1998), wohingegen Ecker bei insgesamt
erhöhter HRV eine reduzierte LF/HF-Ratio bei β1-knock-out-Mäusen beobachtet (Ecker
et al., 2006). Kombinierte autonome Blockade führt analog der Ergebnisse Janssens, aber

45
im Gegensatz zu den Ergebnissen Gehrmanns zu keiner Veränderung der HRV, was mög-
licherweise ein Resultat der tendenziell entgegengerichteten E�ekte von Propranolol und
Atropin ist.
In der Übersicht zeigt sich zusammengefasst eine starke Variation in den Ergebnissen
zum autonomen Ruhetonus und der Beein�ussung der HRV bei Wildtyp-Mäusen sowohl
zwischen den verschiedenen Studien als auch zwischen den verschiedenen Mausstämmen.
Aufgrund dieser Unterschiede erscheint es sinnvoll, dass in jeder Studie, die die HRV-
Analyse bei Mäusen zum Inhalt hat, zunächst der autonome Tonus des zu untersuchenden
Mausstamms unter den individuellen experimentellen Bedingungen bestimmt wird. Dies
ist umso wichtiger, solange sich ein standardisiertes Verfahren zur HRV-Gewinnung nicht
allgemein durchgesetzt hat.
5.3 Die Einschränkung der Herzfrequenzvariabilität bei Sepsis und MODS
In der ersten klinischen Arbeit, die von einer eingeschränkten Herzfrequenzvariabilität
bei Intensivpatienten mit einer Sepsis berichtete, zeigten 17 Patienten mit Sepsis eine
signi�kante Reduktion der Total Power und der LF in normalized units (nu), die invers
mit dem APACHE II und TISS Score korrelierte (Garrard et al., 1993, zu Sepsisscores
siehe Cullen et al., 1974 und Knaus et al., 1985).
Christopher S. Garrard beobachtete eine zunehmende HRV-Einengung mit zuneh-
mender Schwere der Erkrankung. Als mögliche Ebenen für die gestörte autonome Kon-
trolle der HRV wurden das ZNS, die Transmitterebene oder das veränderte Reaktions-
vermögen auf Endorganebene angesehen. Piepoli berichtete über ein annäherndes Ver-
schwinden der LFnu bei 12 Patienten im septischen Schock, von denen zwei verstarben.
Bei den Überlebenden kam es zu einemWiederanstieg dieses Wertes (Piepoli et al., 1995).
1999 berichteten Djillali Annane und Konstantin M. Heinroth über eine HRV-Ein-
schränkung inklusive einer Reduktion der LF/HF-Ratio, als häu�g genutzter Parameter
der sympathovagalen Balance bei Patienten mit septischen Schock (Annane et al., 1999)
und Patienten mit Multiorgan-Dysfunktions-Syndrom (Heinroth et al., 1999). Annane
zog Rückschlüsse auf die autonome Regulation mit einer ungenügenden sympathischen
Aktivierung zu Beginn des septischen Geschehens, während Heinroth die Betonung auf
die zunehmende HRV-Einengung mit der Erkrankungsschwere und deren prognostischen
Wert bei MODS legte, unabhängig von einer septischen oder nicht-septischen Genese.
Ste�en Schaefer stellte in seiner Dissertation ebenfalls eine signi�kant verminderte

46
HRV mit zunehmenden APACHE-Score heraus (Schaefer, 2005). In der Studie Marion
Korachs korrelierte eine eingeschränkte LF/HF-Ratio mit der Gesamtmortalität interni-
stischer Intensivpatienten, obwohl die mit diesem Parameter evaluierte sympathovagale
Balance individuell sehr unterschiedlich war (Korach et al., 2001). Sie schloss auf die
Nutzbarkeit dieses Wertes als diagnostischen Marker bei Sepsis, wenn die Ratio < 1,0
sei. Miles S. Ellenby stellte ergänzend bei pädriatrischen Patienten mit einer Sepsis die
ungünstige prognostische Bedeutung einer erniedrigten LF/HF-Ratio und einer ernied-
rigten Total Power fest (Ellenby et al., 2001).
Hendrik Schmidt konnte 2005 eine Einschränkung sämtlicher HRV-Parameter mit
Ausnahme der RMSSD bei MODS nachweisen (Schmidt et al., 2005). Er betonte eben-
falls, dass die beobachtete Abschwächung der gesamten autonomen Funktion, die sich
in der HRV-Einschränkung niederschlüge, prognostische Bedeutung habe: Als negativer
Marker für das Outcome von Patienten mit Herzinsu�zienz bei MODS sei besonders die
Reduktion der SDNN geeignet. Die lnVLF konnte von der Arbeitsgruppe als bester Pro-
gnosefaktor für die 28-Tage-Mortalität identi�ziert werden, wobei in der zitierten Studie
die Aussagekraft dieses Parameters der des APACHE II Scores entsprach.
1996 führte Godin experimentelle Endotoxin-Injektionen an gesunden Probanden
durch (Godin et al., 1996). Eine Einschränkung der HRV setzte nach ein bis zwei Stunden
ein, erreichte ihr Maximum nach vier bis fünf Stunden und hielt für maximal 8 Stunden
an. Nach experimenteller Cecal ligation and puncture (CLP) an Ratten stellte sich eine
Stunde später eine Tachykardisierung und eine Einschränkung der Total Power, der LF
und LF/HF-Ratio ein (Pancoto et al., 2008). Experimentelle Endotoxin-Applikationen
führten bei Mäusen zu einer vergleichbaren HRV-Einschränkung bis zu fünf Stunden
(Adachi et al., 2006) und ab einer bis neun Stunden (Fairchild et al., 2009). Die Be-
obachtungen lassen die Vermutung zu, dass die zugrundeliegenden Mechanismen der
HRV-Einschränkung während einer Sepsis bei Wildtyp-Mäusen, Ratten und Menschen
ähnlich sind.
Der verzögert eintretende und langanhaltende HRV-supprimierende E�ekt nach LPS-
Applikation wird dabei auf zentral und systemisch induzierte, mediatorvermittelte E�ek-
te zurückgeführt. Nach Endotoxinapplikation kann ein Anstieg des Serum-TNFα be-
reits nach ein bis zwei Stunden bei Mäusen und Ratten beobachtet werden (Redl et al.,
1993; Meng et al., 1998). Karin Fairchild konnte bei Mäusen die Serumkonzentration
von TNFα und anderen Zytokinen mit einer lang andauernden HRV-Suppression korre-

47
lieren (Fairchild et al., 2009). Diese langanhaltende Suppression und starke Zytokinaus-
schüttung zeigt Fairchild für eine LPS-Dosierung von 10 mg/kg i.p., der 25-fachen der
hier verwendeten Dosis. Die HRV-Einschränkung und die Dauer der Suppression sind in
Fairchilds Studie dosisabhängig bis zu einer Konzentration von 0,01 mg/kg (drei bis sechs
Stunden) nachweisbar, wobei die Zytokinausschüttung für diese niedrige LPS-Dosierung
nicht untersucht wurde. Eine niedrigere LPS-Dosierung löst jedoch auch eine geringere
in�ammatorische Reaktion aus (Corral et al., 2005).
Gegenüber den Langzeite�ekten verursacht Endotoxinapplikation in den hier durchge-
führten Experimenten an Wildtyp-Mäusen meist eine sofortige, ausgeprägte Tachykardi-
sierung. Diese geht einher mit einer hochgradig eingeschränkten Herzfrequenzvariabilität
(signi�kante Reduktion der RMSSD, der pNN6 und der LF Power). Diese Ergebnisse
ähneln denen aus Zellkulturexperimenten. Mit Endotoxin inkubierte neonatale Ratten-
Kardiomyozyten zeigen eine Reduktion des pNN50 und pNN30 sowie der RMSSD, bei
unbeein�usster SDNN (Schmidt et al., 2007). Es wird die Hypothese gestärkt, dass En-
dotoxin selbst eine entscheidende Rolle in der Entstehung der HRV-Einschränkung bei
Sepsis einnimmt (Zorn-Pauly et al., 2007). Eine direkte Endotoxin-Interaktion mit den
Zielzellen könnte im Gegensatz zu den mediatorvermittelten Langzeite�ekten des LPS
initial für die ausgeprägte HRV-Einschränkung und mit ihr für die Entkopplung der
Interorgan-Kommunikation verantwortlich sein.
Pincus Modell der approximate entropy, dessen zufolge die Regularität eines Signals in
einem System dessen Isolierung o�enbart, wurde in weiteren Studien über das Verhalten
gekoppelter Systeme ausgebaut. Paul J. Godin und Timothy G. Buchman stellten aus
dieser Entwicklung heraus 1996 die Theorie auf, dass die Entkopplung der Organe bei
Sepsis ursächlich für die Entstehung eines MODS sei (Godin and Buchman, 1996). Eine
eingeschränkte HRV im Sinne einer kardialen autonomen Dysfunktion kann diese bereits
eingetretene Entkopplung der Interorgankommunikation aufzeigen, die in der Konsequenz
möglicherweise zu einem MODS führt (Schmidt et al., 2008b; Werdan et al., 2008).
Die negative prognostische Stärke der eingetretenen HRV-Einschränkung in vielen
Studien wäre diesem Modell zufolge eine logische Schlussfolgerung: die HRV-Suppression
o�enbart sich nicht nur als Symptom oder diagnostisches Charakteristikum der septi-
schen Kardiomyopathie, sondern weist auch auf die gestörte Interogrankommunikation
als wahrscheinliche Teilursache dieser Krankheitsentität hin. Es handelt sich bei der septi-
schen Kardiomyopathie daher möglicherweise auch um eine Isolierungserkrankung, zu der

48
die spezi�sche Interaktion von LPS und den kardialen Schrittmacherzellen mit beitragen
könnte. Durch die gestörte Kommunikation wäre das Herz nicht in der Lage, sich aus ei-
ner sinnvollen sympathisch-parasympathischen Interaktion heraus, den Bedürfnissen des
Körpers adäquat anzupassen (Levy, 1971; Hainsworth, 1995). Die Reversibilität funktio-
neller Störungen bei der Erholung von der Erkrankung (Müller-Werdan et al., 2006a),
die mit einer Wiedererlangung der HRV als ein Zeichen einer einsetzenden Integration
des Herzens in den Organismus durch eine funktionierende Kommunikation einhergeht,
spricht für diese Annahme Godins.
Eine direkte Verminderung der myokardiale Kontraktilität durch Endotoxinapplikati-
on wird berichtet, die intrazellulären Mechanismen, die dies vermitteln, sind verschieden
(u. a. Flesch et al., 1999). Die Ursache für die charakteristische HRV-Einengung als Aus-
druck der autonomen Dysfunktion im Rahmen eines MODS ist somit möglicherweise
auch in der direkten Interaktion von Endotoxin mit den Schrittmacherzellen des Herzens
mitbegründet.
5.4 Die Rolle von Endotoxin in der gestörten Interorgankommunikation
1980 wurde über die gramnegative Sepsis als Hauptursache für das Multiple-System-
Organ-Failure (MSOF) mit konsekutiver hoher Sterblichkeit bei chirurgischen Patienten
auf der Intensivstation berichtet (Fry et al., 1980). Die Krankenhaussterblichkeit der
schweren Sepsis und des septischen Schocks betrug einer 2007 publizierten, multizentri-
schen Studie des Kompetenznetzwerks Sepsis (SepNet) zufolge im Jahr 2003 55%, die
Prävalenz in der Stichprobe der Intensivstationen lag bei 11% (Engel et al., 2007). Die
Inzidenz der Sepsis läge bei ca. 79 000 Einwohnern, die der schweren Sepsis und des sep-
tischen Schocks bei 75 000 Einwohnern pro Jahr (116 bzw. 110 pro 100 000, Reinhardt
et al., 2006; Engel et al., 2007). Septische Erkrankungen stellen demnach die dritthäu�g-
ste Todesursache in Deutschland dar, wobei jedoch nur in circa einem Drittel der Fälle
ein Erreger nachgewiesen werden kann (Reinhart et al., 2010).
Während Christoph Engel zufolge gramnegative Infektionen genauso häu�g auftreten
wie grampositive (Engel et al., 2007), machen sie in anderen Studien den Hauptanteil
der Infektionen aus und gehen dabei deutlich gehäuft mit schweren Verläufen der Sepsis,
wie dem septischen Schock einher (Alberti et al., 2002). Bereits früher wurde ein deutlich
ungünstigerer klinischer Verlauf eines septischen Schocks beobachtet, wenn der initiale
Fokus der Gastrointestinaltrakt darstellte (Nishijima et al., 1973). Aktuell nimmt der

49
Anteil gramnegativer Bakteriämien auf den Intensivstationen zu, wobei vor allem das
multiresistente Keimspektrum an Bedeutung gewinnt (Arndt et al., 2011). Ihr Stellenwert
bei der Entwicklung eines MODS und der damit assoziierten Mortalität ist als sehr hoch
einzuschätzen (Alberti et al., 2002).
1986 verwies Charles Natanson in einer Studie auf den direkt kardiodepressiven E�ekt,
den eine Endotoxinapplikation auslöst (Natanson et al., 1986). Er ergänzte 1989 durch
Experimente an Hunden, dass eine vergleichbare Myokarddepression ebenfalls durch
grampositive Keime ausgelöst werden kann (Natanson et al., 1989). Diese Beobachtung
bestätigte sich in klinischen Studien (Pilz et al., 1994). Natanson stellte die Hypothese
in den Vordergrund, dass der Myokarddepression bei einem durch Infektion ausgelösten
MODS ein allgemeingültiger, durch die Infektion ausgelöster Mechanismus zugrunde lie-
ge. Das Auftreten einer der septischen Kardiomyopathie ähnelnden Herzfunktionsein-
schränkung bei SIRS unterstützt diese Annahme (Smaïl et al., 1996; Müller-Werdan
et al., 2006b). Die Rolle reaktiv ausgeschütteter Zytokine, insbesondere des TNFα und
des IL-1, konnten in der Folge nachgewiesen und mit einer Myokarddeppression in Ver-
bindung gebracht werden (Hesse et al., 1988; Kumar et al., 1996; Meng et al., 1998; Cain
et al., 1999).
Neben diesen Faktoren ist es auch wahrscheinlich, dass Endotoxin selbst zur Entste-
hung eines MODS und zu der Isolierung des Herzens im Organismus beiträgt. Die experi-
mentelle Applikation von LPS an freiwilligen Probanden verursacht eine linksventrikuläre
systolische Dysfunktion, die der Kardiomyopathie bei Patienten mit Sepsis stark ähnelt
(Su�redini et al., 1989). Robert L. Danner zeigte 1991, dass ein MODS 10 mal häu�ger
und eine linksventrikuläre Dysfunktion 5 mal häu�ger bei Patienten mit gramnegativer
Endotoxinämie ausgelöst wurde, als bei Patienten mit septischen Schock ohne Endoto-
xinämie. Eine Endotoxinämie ging zudem mit einer erhöhten Mortalität einher (Danner
et al., 1991).
Verschiedene Studien konnten im Weiteren (neben den bereits genannten indirekten
kardiodepressiven Mechanismen) auch eine direkte Interaktion von Endotoxin und dem
Arbeitsmyokard als eine mögliche Ursache der septischen Kardiomyopathie aufzeigen.
LPS führte unter anderem zu einer Veränderung der Kardiomyozytenreagibilität auf au-
tonome Ein�üsse. Hierbei konnte sowohl eine erhöhte Sensibilisierung auf positiv inotrope
Ein�üsse durch β-adrenerge Stimulation (Rubin et al., 1994) als auch eine Abschwächung
beobachtet werden (Silverman et al., 1993). Ursula Müller-Werdan zeigte 1998 beispiels-

50
weise eine fehlende Steigerung der Kontraktionskraft auf Isoproterenol-Stimulation bei
Endotoxin-inkubierten Ratten-Kardiomyozyten, während die Reaktion auf α-adrenerge
Stimulation unbeein�usst ist (Müller-Werdan et al., 1998). Auch über eine Verstärkung
der inhibitorischen E�ekte von Acetylcholin mit einer Reduktion der Kontraktilität wurde
berichtet (Sulakhe et al., 1996). Endotoxin ist in der Lage, über verschiedene Mechanis-
men eine direkte Verkürzung des Aktionspotentials an Kardiomyocyten hervorzurufen,
die die Antwort auf autonome Ein�üsse modulieren könnten (diskutiert bei Zorn-Pauly
et al., 2007). Najah Abi-Gerges zeigte 1999 an isolierten Rattenkardiomyozyten einen
durch Endotoxin gesteigerten stimulierenden E�ekt von Isoproterenol auf ICa für 12
Stunden, der durch Stimulation der Adenylatzyklase nachgeahmt werden konnte. Der
antiadrenerge E�ekt von Acetylcholin auf ICa war erst nach 26 Stunden deutlich ge-
steigert (Abi-Gerges et al., 1999). Er äuÿerte die Hypothese, dass eine zeitabhängige
Veränderung des Adenylatzyklase-Pathways in den Kardiomyozyten über die autonome
Kontrolle von ICa zur myokardialen Dysfunktion bei Sepsis führen könnte.
Vor dem Hintergrund der hohen Katecholamin-Konzentrationen im Serum bei Sepsis
(Müller-Werdan et al., 1996; Annane et al., 1999) und dem therapeutischen Einsatz von
Katecholaminen mit dem gleichzeitig bekannten Phänomen der verminderten Responsi-
vität der Zielorgane könnte der Sensibilisierung der Kardiomyozyten für sympathische
Ein�üsse durch Endotoxin möglicherweise eine besondere Bedeutung zukommen. Diese
Potenzierung sympathischer Ein�üsse �ndet wahrscheinlich ebenfalls an den Schrittmach-
zellen des Herzens statt, was durch elektrophysiologische Untersuchungen an isolierten
Vorhofkardiomyozyten untermauert wird (Zorn-Pauly et al., 2007).
Der direkte Ein�uss von Endotoxin auf die Schrittmacherzellen des Herzens in vivo
wurde bisher noch nicht untersucht. Da experimentelle Endotoxin-Applikationen wieder-
holt zu einer ausgeprägten Tachy- oder Bradykardisierung mit hochgradig eingeschränk-
ter HRV führten, was ein wesentliches prognoserelevantes Charakteristikum der septi-
schen Kardiomyopathie darstellt, ist die Frage nach einer direkten Interaktion von be-
sonderem Interesse. Dem Schrittmacher-Kanal als taktgebende Zielstruktur des Herzens,
der sympathisch und parasympathisch moduliert wird (Baruscotti et al., 2005; Barbu-
ti and DiFrancesco, 2008), kommt dabei als möglicher Ort der Beeinträchtigung au-
tonomer Ein�üsse am Herzen während einer Sepsis und einem MODS eine besondere
Bedeutung zu. Auf die Möglichkeit der direkten Interaktion von LPS und dem Schritt-
macherstrom If verwies Schmidt 2007, in dessen Untersuchungen es nach experimenteller

51
LPS-Applikation zu einer Verminderung der HRV an isolierten, spontan kontrahierenden
Rattenkardiomyocyten kam (Schmidt et al., 2007). Die Interaktionsmechanismen von
LPS mit dem If -Strom, die Klaus Zorn-Pauly in Zellkulturexperimenten an humanen
Vorhofkardiomyozyten 2007 nachwies, stellen möglicherweise einen entscheidenden Me-
chanismus in der Entstehung der HRV-Einschränkung bei Sepsis dar (Zorn-Pauly et al.,
2007; Werdan et al., 2008; Ebelt and Werdan, 2010).
Mittels elektrophysiologischer Untersuchungen konnten zwei spezi�sche E�ekte des
LPS auf die hyperpolarization-activated cyclic nucleotide-gated cation channels (HCN-
Kanäle) als Träger des If aufgezeigt werden (Abb. 21). Zum einen führt LPS-Applikation
in Anwesenheit der sympathomimetischen Substanz Isoproterenol zu einer Potenzierung
des sympathischen Ein�usses an den Zellen (Abb. 21a). Dieser für den Sympathikus sen-
sibilisierende E�ekt könnte für die bereits oben beschriebene Tachykardisierung nach
LPS-Applikation bei den hier untersuchten Mäusen verantwortlich sein. Alleinige Isopro-
terenolapplikation reduziert die HRV-Parameter zwar deutlich, jedoch nicht signi�kant.
Die HRV-supprimierende Wirkung des Endotoxins könnte somit als eine Bahnung des
sympathischen Ein�usses interpretiert werden.
Der genaue Mechanismus dieser Beein�ussung des If -Stroms ist derzeit unklar. Eine
Rolle könnte die in Kardiomyocyten beschriebene Beein�ussung der second-messenger-
Kaskaden durch Endotoxin spielen. Eine direkte Interaktion von Endotoxin mit dem
human hyperpolarization-activated cyclic nucleotide-gates channel 2 (hHCN2) in-vitro
konnte bereits ebenfalls nachgewiesen werden (Klöckner et al., 2014).
Für den direkten Ein�uss des Endotoxins auf den Schrittmacherkanal spricht die
Geschwindigkeit, mit der der E�ekt nachweisbar ist. Die HRV-Einschränkung nach LPS-
Applikation tritt in der vorliegenden Arbeit bereits nach 20�40 Minuten ein. Auch wird
über die Schnelligkeit anderer durch LPS-Applikation hervorgerufener Kreislaufe�ekte
berichtet (Savci and Ulus, 1997). Gestützt wird die Vermutung einer ursächlich direkten
Interaktion von LPS und If weiterhin durch die Beobachtungen Javier Corrals, der bei
Verwendung einer identischen subletalen LPS-Dosierung wie in der vorliegenden Arbeit
kaum eine in�ammatorische Reaktion bei seinen untersuchten Mäusen feststellen konnte
(Corral et al., 2005). Xianzhong Meng beobachtete einen Anstieg des Serum-TNFα bei
Ratten nach LPS-Applikation in einer Dosierung von 0,5mg/kg frühestens nach 1 bis 2
Stunden (Meng et al., 1998), Heinz Redl bei Mäusen nach frühstens einer Stunde (Redl
et al., 1993).

52
(a) Alleinige LPS-Applikation erhöht die
Ansprechbarkeit des If -Stroms auf
sympathische Reize. Dies resultiert in einer
ausgeprägten Tachykardisierung. Die
Herzfrequenzvariabilität (HRV) ist hochgradig
eingeschränkt.
(b) Unter autonomer Blockade mit Propranolol und
Atropin (=Sterne) kann die Beein�ussung des
Sympathikus durch Endotoxin ausgeblendet werden. Es
stellt sich eine ausgeprägte Bradykardisierung ein, welche
das Ergebnis des in Zellkulturexperimenten
nachgewiesenen direkt hemmenden E�ekts auf If sein
könnte. Die HRV ist weniger stark eingeschränkt.
Abb. 21: Mögliche Beein�ussung des Schrittmacherstroms If durch Endotoxin nach Zorn-Pauly et al., 2007.
LPS=Lipopolysaccharid (Endotoxin), P=Parasympathikus, S=Sympathikus, If=Schrittmacherstrom,
HRV=Herzfrequenzvariabilität, Sterne=Propranolol und Atropin.
Die Tachykardisierung und HRV-Einschränkung sind in der vorliegenden Arbeit ca. 20
Minuten nach LPS-Applikation nachweisbar, was eine indirekte, z. B. zytokinvermittelte
Ursache weniger wahrscheinlich macht. Ergänzend zeigt sich bei der klinisch beobach-
teten Einschränkung der kardialen Pumpfunktion im Rahmen eines Traumas (SIRS) in
einigen Studien keine Beein�ussung der Herzfrequenz (Smaïl et al., 1996). Die die Tachy-
kardisierung begleitende HRV-Einschränkung könnte additiv zur Sympathikusbahnung
das Resultat der direkten LPS-If -Interaktion sein und ginge mit der von Zorn-Pauly be-
richteten verringerten Reagibilität des If -Kanals auf autonome, oder zumindest vagale
Reize unter LPS-Ein�uss einher.
Einen zweiten E�ekt an den untersuchten Zellen zeigte Zorn-Pauly in seinen Zellkul-
turexperimenten: eine direkte, spezi�sche Hemmung des Schrittmachstroms, die per se
zu einer Bradykardisierung führen sollte (Abb. 21b). Diese Situation wurde in der vorlie-
genden Arbeit in vivo durch die LPS-Applikation in Abwesenheit autonomer Ein�üsse
simuliert, hervorgerufen durch kombinierte autonome Blockade. Die Wildtyp-Mäuse zei-
gen in dieser Situation nach LPS-Gabe tatsächlich eine ausgeprägte Bradykardisierung,

53
die sich signi�kant von der Herzfrequenzänderung nach alleiniger autonomer Blockade
unterscheidet, und eine weniger stark ausgeprägte HRV-Einschränkung. Die vorliegen-
de in vivo-Arbeit bestätigt damit in funktioneller Hinsicht den im Zellkulturexperiment
gezeigten hemmenden E�ekt von LPS auf die kardialen Schrittmacherzellen.
Am Modell der Wildtyp-Maus können in der vorliegenden Arbeit beide von Zorn-
Pauly aufgezeigten LPS-E�ekte funktionell bestätigt werden. Die Arbeit trägt damit
zur Erklärung der Genese einer prognoserelevanten HRV-Einschränkung mit gleichzeitig
bestehender Tachykardie im Rahmen einer Sepsis bei.
Auf dem Konzept Godins beruhend, demzufolge das MODS als Resultat einer gestör-
ten Organkommunikation zu verstehen ist, rückte in den letzten Jahren die Bedeutung
einer Beeinträchtigung des cholinergic antiin�ammatory pathways als Ursache für die-
se gestörte Organkommunikation in den Blickpunkt (Schmidt et al., 2008b). Hinweise
auf die Aktivierung des Parasympathikus sowie auf die antiin�ammatorische Potenz des
intakten Parasympathikus während einer Infektion lieferten verschiedene Studien (Boro-
vikova et al., 2000; Hansen et al., 2001; Saeed et al., 2005; Bauhofer and Torossian, 2007;
Huston et al., 2007; Tracey, 2007; Bruchfeld et al., 2010). Schmidt schlussfolgerte 2005
aus einer reduzierten Barore�exaktivität und der HRV-Einschränkung bei intensivme-
dizinischen Patienten mit Multiorgan-Dysfunktions-Syndrom auf eine reduzierte vagale
Aktivität bei MODS (Schmidt et al., 2005) als ein Zeichen der Abschwächung dieses cho-
linergic antiin�ammatory pathways. Warum es im Rahmen eines infektiösen Geschehens
zu einer Störung dieser postulierten protektiven Rückkopplungsschleife kommen kann
und wo genau diese Störung lokalisiert ist, die in der Konsequenz zu der Entstehung
eines MODS und der septischen Kardiomyopathie beitragen könnte, ist bisher unklar
(Schmidt et al., 2008b).
Die beiden in dieser Arbeit funktionell bestätigten E�ekte des Endotoxins, Sympa-
thikusbahnung und direkte Hemmung an den kardialen Schrittmacherzellen in vivo sind
womöglich eine Ursache für die inadäquate Reaktion des Herzens auf vagale Ein�üsse:
als reduziert zeigten sich nach LPS-Applikation bei Wildtyp-Mäusen die typischerweise
den Parasympathikus präsentierenden Parameter RMSSD und pNN6 sowie die LF-Power
(die bei den Mäusen in der vorliegenen Arbeit ebenfalls vom Parasympathikus reguliert
wird). Die Reaktion ähnelt am ehesten der Atropinapplikation. Der sympathikusbahnen-
de E�ekt (Tachykardie und möglicherweise auch intensivere Suppression der HRV) mag
die autonome Reagibilität in der septischen Kardiomyopathie ebenfalls vermindern.

54
Vor dem Hintergrund der zugrunde liegenden Zellkuturexperimente, deren Kompo-
nenten der LPS-Interaktion hier einzeln in vivo nachvollzogen werden konnten, scheint
die Endotoxin-induzierte HRV-Einschränkung durch ein Zusammenwirken beider Inter-
aktionsmechanismen am HCN-Kanal zu entstehen. Die Arbeit stellt deren möglichen
zentralen Stellenwert bei der Entwicklung der autonomen Dysfunktion des Herzens im
Rahmen eines MODS zur Diskussion.
5.5 Studienlimitierung und Forschungsexposé
Mit den Leitlinien zur standardisierten HRV-Messung beim Menschen entwickelte die
Task Force of The European Society of Cardiology and The North American Society of
Pacing and Electrophysiology 1994 eine einheitliche Nomenklatur und reproduzierbare
Methoden der HRV-Erfassung, die eine Vergleichbarkeit und diagnostische Nutzbarkeit
der HRV-Daten aus klinischen und experimentellen Situationen ermöglichte und beför-
derte (Malik et al., 1996). Obwohl die HRV-Messung auch in tierexperimentellen Studien
für unterschiedlichste Fragestellungen genutzt wird, haben sich standardisierte Versuchs-
und Messprotokolle bisher nicht durchgesetzt. Eine Vergleichbarkeit der Ergebnisse aus
den verschiedenen Studien ist daher nur begrenzt möglich und zeigt sich beispielsweise
in den unterschiedlichen Daten zur autonomen Regulation der Herzfrequenz bei Mäusen.
Die vorliegende Dissertation folgte den vorgeschlagenen Richtlinien von Thireau et al.
zur HRV-Messung bei Mäusen (Thireau et al., 2008), um die Idee einer standardisierten
Datengewinnung bei Versuchstieren zu unterstützen und eine bessere Vergleichbarkeit der
Daten zu ermöglichen. Die Bestimmung der Time Domain und der Frequency Domain
Parameter mittels Fast Fourier Transformation (FFT) nach dem dort vorgeschlagenen
Algorithmus erbringt unter den gleichen technischen Voraussetzungen valide Ergebnisse,
die eine Einschätzung der postoperativen Stabilisierung, der pharmakologischen E�ekte
und insbesondere der E�ekte des Endotoxins zulassen. Die eigenen Ergebnisse zur auto-
nomen Regulation der Herzfrequenz reihen sich in frühere Arbeiten zur HRV bei Mäusen
ein, wobei nur ein Teil der Studien zu ähnlichen Ergebnissen und Schlussfolgerungen
gekommen war wie die vorliegende Arbeit. Vor dem Hintergrund, dass es aufgrund der
hohen Herzfrequenz bei Mäusen schwierig ist, ausreichend lange Intervalle mit einer aus-
reichenden Signalstabilität zu sammeln, die für die Durchführung der FFT notwendig
sind (Cerutti et al., 1995), stellen die von Thireau et al. vorgeschlagenen 3 Minuten in
absoluter Ruhe zur Erfassung der FD einen guten Kompromiss zwischen Datenausbeute

55
und Auswertungsaufwand dar (Thireau et al., 2008). Die ähnliche Reaktion der Mäuse
auf die autonome Modulation und auf die LPS-Applikation zu einer Vielzahl muriner
und humaner Studien bestätigte die Nutzbarkeit der Methodik zur Evaluierung pharma-
kologischer und toxikologischer Fragestellungen und erlaubt Rückschlüsse auf humane
Funktionsmechanismen.
In den hier durchgeführten Experimenten wurden keine zusätzlichen Kreislauf- oder
Laborparameter zur Beurteilung der Schwere des durch Endotoxin-Applikation hervor-
gerufenen Krankheitsbildes bei den Mäusen herangezogen. Es lässt sich daher nicht von
der verwendeten subletalen Endotoxindosis auf eine Ähnlichkeit der Kreislaufreaktion
zu der klinisch beobachteten Sepsis mit MODS schlieÿen. Vor dem Hintergrund, dass
Endotoxin-Injektionen an Tieren ähnliche E�ekte auslösen, wie das septische Geschehen
bei Patienten auf der Intensivstation (beispielsweise Savci and Ulus, 1997), fokussier-
te sich die vorliegende Arbeit auf den in-vivo-Nachweis der in Zellkulturexperimenten
nachgewiesenen Interaktion von LPS mit den kardialen Schrittmacherzellen. Als Marker
der septischen Kardiomyopathie weist die hochgradige HRV-Einschränkung der ICR-
Wildtyp-Mäuse durch die intraperitoneale Endotoxin-Applikation groÿe Ähnlichkeit zu
anderen experimentellen und klinischen Beobachtungen bei Patienten mit Sepsis und
MODS auf. Um den E�ekt der Interaktion von LPS und dem HCN-Kanal näher zu
charakterisieren, wurde zwischenzeitlich bereits mit der Fortsetzung der Experimente an
HCN4-Knock-out-Mäusen begonnen. Desweiteren könnte in diesem Rahmen untersucht
werden, ob pharmakologische Hemmer des If (z. B. Ivabradin) den Ein�uss von LPS auf
die HRV beein�ussen.

56
6 Zusammenfassung
Vor dem Hintergrund der prognoserelevanten autonomen kardialen Dysfunktion bei Pa-
tienten mit einem Multiorgan-Dysfunktions-Syndrom (MODS) wird in der vorliegenden
Dissertation der Fragestellung des direkten Ein�usses von Endotoxin auf die Schrittma-
cherzellen des Herzens nachgegangen und dessen Auswirkung auf die Herzfrequenzva-
riabilität (HRV) bei ICR-Wildtyp-Mäusen untersucht. Mittels telemetrischer Aufzeich-
nungsverfahren können die Bewegungsaktivität und die Herzfrequenz der Tiere in vivo
unter Normalbedingungen erfasst werden. Die HRV wird nach international etablierten
Methoden und Parametern ermittelt, hierbei wird ein Auswertungsalgorithmus zur stan-
dardisierten HRV-Gewinnung bei Mäusen von 2008 angewendet (Thireau et al., 2008).
Auf diese Weise ist es möglich, die Stabilisierung nach der Transmitterimplantation und
das autonome Nervensystem der untersuchten Tiere zu charakterisieren sowie den Ein-
�uss von Endotoxin auf die Herzfrequenz und die HRV zu beurteilen.
Eine zirkadiane Schwankung der Herzfrequenz setzt frühzeitig bis spätestens zum
5.Tag nach operativer Transmitterimplantation ein, während die HRV-Stabilisierung bis
zum 10. postoperativen Tag andauert. Die Ergebnisse zur Erholung und Stabilisierung
der HRV-Parameter nach der Transmitterimplantation bestätigen die Befunde einer Rei-
he von Arbeitsgruppen, wobei die Ergebnisse die Notwendigkeit eines längeren post-
operativen Erholungszeitraums verdeutlichen.
Die Experimente zeigen einen ausgeglichenen autonomen Ein�uss zwischen Sympathi-
kus und Parasympathikus bei der Regulation der Ruheherzfrequenz bei Wildtyp-Mäusen.
Die Reduktion sämticher HRV-Parameter inklusive der Low und High Frequency Power
durch Atropin bei nur geringem E�ekt der Propranolol-Applikationen deutet auf die vor-
nehmliche Bedeutung des Parasympathikus in der Entstehung der HRV hin. Der Ein�uss
des Sympathikus zeigt sich in der LF/HF-Ratio, wobei sympathomimetische Stimulation
die Ratio senkt, β-Blockierung die Ratio anhebt.
Endotoxin führt unmittelbar (d. h. innerhalb von 20 Minuten) nach Injektion bei
Wildtyp-Mäusen zu einem signi�kanten Anstieg der Herzfrequenz. Unter autonomer
Blockade bewirkt es dagegen eine ausgeprägte Bradykardisierung. Die HRV ist nach
Endotoxin-Applikation unmittelbar hochgradig eingeschränkt. Unter autonomer Blocka-
de ist diese LPS-induzierte HRV-Einschränkung weniger ausgeprägt.
Die Applikation von LPS verursacht Veränderungen der Herzfrequenz und der HRV,

57
wie sie in klinischen Studien sowie bei Mensch und Tier in experimenteller Situation
im Rahmen einer Sepsis, eines MODS oder einer künstlich hervorgerufenen Endoto-
xinämie bereits zuvor beobachtet wurden. Als ein möglicher Mechanismus dieser charak-
teristischen Herzfrequenzveränderung und HRV-Einschränkung bei Patienten mit MODS
kommt die direkte Interaktion von Endotoxin mit den Schrittmachzellen, insbesondere
dem Schrittmacherstrom If in Betracht. Die in Zellkulturexperimenten beschriebene di-
rekte Hemmung des If -Schrittmacherstroms und dessen Sympathikus-Sensibilisierung
durch Endotoxin können in den hier vorgestellten in vivo-Experimenten an Wildtyp-
Mäusen funktionell bestätigt werden.

58
7 Literaturverzeichnis
Literatur
Abi-Gerges N, Tavernier B, Mebazaa A, Faivre V, Paqueron X, Payen D, Fischmeister R, Méry PF (1999)
Sequential changes in autonomic regulation of cardiac myocytes after in vivo endotoxin injection in
rat. Am J Respir Crit Care Med 160:1196�1204.
Adachi Y, Nakajima Y, Satomoto M, Morita K, Doi M, Sato S (2006) The heart rate variability in mice:
telemetric evaluation of endotoxin shock. Masui 55:436�440.
Akselrod S: Components of Heart Rate Variability: Basic Studies. In: Malik M, Camm AJ (eds): Heart
Rate Variability. Futura Publishing Company, Inc., New York, 1995, pp 147�163.
Akselrod S, Gordon D, Ubel FA, Shannon DC, Barger AC, Cohen RJ (1981) Power spectrum analysis of
heart rate �uctuation: a quantitative probe of beat to beat cardiovascular control. Science 213:220�222.
Alberti C, Brun-Buisson C, Burchardi H, Martin C, Goodman S, Artigas A, Sicignano A, Palazzo M,
Moreno R, Boulmé R, Lepage E, Le Gall JR (2002) Epidemiology of sepsis and infection in ICU
patients from an international multicentre cohort study. Intensive Care Med 28:108�121.
Annane D, Trabold F, Sharshar T, Jarrin I, Blanc AS, Raphael JC, Gajdos P (1999) Inappropriate
sympathetic activation at onset of septic shock: a spectral analysis approach. Am J Respir Crit Care
Med 160:458�465.
Appel ML, Berger RD, Saul JP, Smith JM, Cohen RJ (1989) Beat to beat variability in cardiovascular
variabilities: noise or music? J Am Col Cardiol 14:1139�1148.
Arndt S, Lauf H, Weiss G, Lodes U, Mroczkowski P, Schulz HU, Lippert H, König W, Meyer F (2011)
Erreger- und Resistenzspektrum einer chirurgischen ITS im systematischen 10-Jahres-Vergleich 1996-
2005 im Rahmen des mikrobiologischen Routinemonitorings. Zentralbl Chir 136:152�158.
Arras M, Rettich A, Paolo C, Kasermann HP, Burki K (2007) Assessment of post-laparotomy pain in
laboratory mice by telemetric recording of heart rate and heart rate variability. BMC Vet Res 3:16.
Aysin B, Aysin E (2006) E�ect of respiration in heart rate variability (HRV) analysis. Conf Proc IEEE
Eng Med Biol Soc 1:776�779.
Barbuti A, DiFrancesco D (2008) Control of cardiac rate by �funny� channels in health and disease. Ann
N Y Acad Sci 1123:213�223.
Baruscotti M, Bucchi A, Difrancesco D (2005) Physiology and pharmacology of the cardiac pacemaker
(�funny�) current. Pharmacol Ther 107:59�79.
Baudrie V, Laude D, Elghozi JL (2007) Optimal frequency ranges for extracting information on cardio-
vascular autonomic control from the blood pressure and pulse interval spectrograms in mice. Am J
Physiol Regul Integr Comp Physiol 292:R904�R912.

59
Baue AE (1992) The horror autotoxicus and multiple-organ failure. Arch Surg 127:1451�1462.
Bauhofer A, Torossian A (2007) Mechanical vagus nerve stimulation�a new adjunct in sepsis prophylaxis
and treatment? Crit Care Med 35:2868�2869.
Baumans V, Bouwknecht JA, Boere H, Kramer K, Van Lith HA, Van de Weerd HA, Van Herck H (2001)
Intra-abdominal transmitter implantation in mice: e�ects on behaviour and body weight. Anim Wel
10:291�302.
Berntson G, Bigger JT, Eckberg DL, Grossman P, Kaufmann PG, Malik M, Nagaraja HN, Porges SW,
Saul JP, Stone PH, van der Molen MW (1997) Heart rate variability: origins, methods, and interpretive
caveats. Psychophysiology 34:623�648.
Borovikova LV, Ivanova S, Zhang M, Yang H, Botchkina GI, Watkins LR, Wang H, Abumrad N, Eaton
JW, Tracey KJ (2000) Vagus nerve stimulation attenuates the systemic in�ammatory response to
endotoxin. Nature 405:458�462.
Bruchfeld A, Goldstein RS, Chavan S, Patel NB, Rosas-Ballina M, Kohn N, Qureshi AR, Tracey KJ
(2010) Whole blood cytokine attenuation by cholinergic agonists ex vivo and relationship to vagus
nerve activity in rheumatoid arthritis. J Intern Med 268:94�101.
Cain BS, Meldrum DR, A DC, Meng X, Joo KS, Banerjee A, Harken AH (1999) Tumor necrosis
factor-alpha and interleukin-1 beta synergistically depress human myocardial function. Crit Care
Med 27:1309�1318.
Campen MJ, Tagaito Y, Jenkins TP, Balbir A, O'Donnell CP (2005) Heart rate variability responses to
hypoxic and hypercapnic exposures in di�erent mouse strains. J Appl Physiol 99:807�813.
Casolo G: Heart Rate Variability in Patients with Heart Failure. In: Malik M, Camm AJ (eds): Heart
Rate Variability. Futura Publishing Company, Inc., New York, 1995, pp 449�466.
Cerutti S, Bianchi AM, Mainardi LT: Spectral Analysis of the Heart Rate Variability Signal. In: Malik
M, Camm AJ (eds): Heart Rate Variability. Futura Publishing Company, Inc., New York, 1995, pp
63�74.
Chess GF, Tam RMK, Calaresu FR (1975) In�uence of cardiac neural inputs on rhythmic variations of
heart period in the cat. Am J Physiol 228:775�780.
Clark IA (2007) The advent of the cytokine storm. Immunol Cell Biol 85:271�273.
Clement JG, Mills P, Brockway B (1989) Use of telemetry to record body temperature and activity in
mice. J Pharmacol Methods 21:129�140.
Corral J, Yélamos J, Hernández-Espinosa D, Monreal Y, Mota R, Arcas I, Miñano A, Parrilla P, Vi-
cente V (2005) Role of lipopolysaccharide and cecal ligation and puncture on blood coagulation and
in�ammation in sensitive an resistant mice models. Am J Pathol 166:1089�1098.

60
Cullen DJ, Civetta JM, Briggs BA, Ferrara LC (1974) Therapeutic intervention scoring system: a method
for quantitative comparison of patient care. Crit Care Med 2:57�61.
Damas P, Reuter A, Gysen P, Demonty J, Lamy M, Franchimont P (1989) Tumor necrosis factor and
interleukin-1 serum levels during severe sepsis in humans. Crit Care Med 17:975�978.
Danner RL, Elin RJ, Hosseini JM, Wesley RA, Reilly JM, Parillo JE (1991) Endotoxemia in human
septic shock. Chest 99:169�175.
Data Science International (2008) ETA, CTA, EA or CA Device Surgical Manual. 4211 Lexington Avenue
North Suite 2244. St.Paul, MN 55126-6164 U.S.A.
Data Science International (2009) DSI Data Exchange Matrix. URL http://www.datasci.com/products/
implantable-telemetry/telemetry-hardware/data-exchange-matrix, eingesehen am 24.09.2009.
Data Science International (2010) DSI System Brochure. URL http://www.datasci.com/pdf/products/
DSISystemBrochure.pdf, eingesehen am 15.10.2010.
Desai KH, Sato R, Schauble E, Barsh GS, Kobilka BK, Bernstein D (1997) Cardiovascular indexes in
the mouse at rest and with exercise: new tools to study models of cardiac disease. Am J Physiol
272:H1053�1061.
Dimas S (2007) Empfehlung der Kommission vom 18. Juni 2007 mit Leitlinien für die Unterbringung
und P�ege von Tieren, die für Versuche und andere wissenschaftliche Zwecke verwendet werden
(2007/526/EG). Amtsblatt der Europäischen Union vom 30.07.2007.
Ebelt H, Werdan K (2010) Sepsis und Herz. Internist 51:844�849.
Eckberg DL (1997) Sympathovagal balance: a critical appraisal. Circulation 96:3224�3232.
Ecker PM, Lin CC, Powers J, Kobilka BK, Dubin AM, Bernstein D (2006) E�ect of targeted deletions
of β1- and β2-adrenergic-receptor subtypes on heart rate variability. Am J Physiol Heart Circ Physiol
290:H192�H199.
Ellenby MS, McNames J, Lai S, McDonald BA, Krieger D, Sclabassi RJ, Goldstein B (2001) Uncoupling
and recoupling of autonomic regulation of the heart beat in pediatric septic shock. Shock 16:274�277.
Engel C, Brunkhorst FM, Bone HG, Brunkhorst R, Gerlach H, Grond S, Gruendling M, Huhle G,
Jaschinski U, John S, Mayer K, Oppert M, Oltho� D, Quintel M, Ragaller M, Rossaint R, Stuber F,
Weiler N, Welte T, Bogatsch H, Hartog C, Loe�er M, Reinhart K (2007) Epidemiology of sepsis in
germany: results from a national prospective multicenter study. Intensive Care Med 33:606�618.
Fairchild KD, Saucerman JJ, Raynor LL, Sivak JA, Xiao Y, Lake DE, Moorman JR (2009) Endotoxin
depresses heart rate variability in mice: cytokine and steroid e�ects. Am J Physiol Regul Integr Comp
Physiol 297:R1019�R1027.

61
Fallen EL, Kamath MV: Circadian Rhythms of Heart Rate Variability. In: Malik M, Camm AJ (eds):
Heart Rate Variability. Futura Publishing Company, Inc., New York, 1995, pp 293�310.
Flesch M, Kilter H, Cremers B, Laufs U, Südkamp M, Ortmann M, Müller FU, Böhm M (1999) E�ects
of endotoxin on human myocardial contractility�involvement of nitritc oxid and peroxynitirite. J Am
Col Cardiol 33:1062�1070.
Flierl MA, Rittirsch D, Huber-Lang MS, Sarma JV, Ward PA (2008) Molecular events in the cardiomy-
opathy of sepsis. Mol Med 14:327�336.
Fry DE, Pearlstein L, Fulton RL, Polk HCJ (1980) Multiple system organ failure. The role of uncontrolled
infection. Arch Surg 115:136�140.
Furlan R, Guzzetti S, Crivellaro W, Dassi S, Tinelli M, Baselli G, Cerutti S, Lombardi F, Pagani M,
Malliani A (1990) Continuous 24-hour assessment of the neural regulation of systemic arterial pressure
and RR variabilities in ambulant subjects. Circulation 81:537�547.
Garrard CS, Kontoyannis DA, Piepoli M (1993) Spectral analysis of heart rate variability in the sepsis
syndrome. Clin Auton Res 3:5�13.
Gehrmann J, Hammer PE, Maguire CT, Wakimoto H, Triedman JK, Berul CI (2000) Phenotypic scree-
ning for heart rate variability in the mouse. Am J Physiol Heart Circ Physiol 279:733�740.
Gehrmann J, Meister M, Maguire CT, Martins DC, Hammer PE, Neer EJ, Berul CI, Mende U (2002)
Impaired parasympathetic heart rate control in mice with a reduction of functional G protein βγ-
subunits. Am J Physiol Heart Circ Physiol 282:H445�H456.
Godin PJ, Buchman TG (1996) Uncoupling of biological oscillators: a complementary hypothesis con-
cerning the pathogenesis of multiple organ dysfunction syndrome. Crit Care Med 24:1107�1116.
Godin PJ, Fleisher LA, Eidsath A, Vandivier RW, Preas HL, Banks SM, Buchman TG, Su�redini AF
(1996) Experimental human endotoxemia increases cardiac regularity: results from a prospective,
randomized, crossover trial. Crit Care Med 24:1117�1124.
Goldbarg AN, Hellerstein HK, Bruell JH, Daroczy AF (1968) Electrocardiogram of the normal mouse,
mus musculus: general considerations and genetic aspects. Cardiovasc Res 2:93�99.
Goldberger AL, West BJ (1987a) Applications of nonlinear dynamics to clinical cardiology. Ann N Y
Acad Sci 504:195�213.
Goldberger AL, West BJ: Chaos in Physiology: Health or Disease? In: Degn H, Holden AV, Olsen LF
(eds): Chaos in Biological Systems. Plenum Press, New York, 1987b, pp 1�4.
Goldberger AL, Findley LJ, Blackburn MR, Mandell AJ (1984) Nonlinear dynamics in heart failure:
implications of long-wavelength cardiopulmonary oscillations. Am Heart J 107:612�615.

62
Goldstein B, Kempski MH, Stair D, Tipton RB, DeKing D, DeLong DJ, DeAsla R, Cox C, Lund N,
Woolf PD (1995) Autonomic modulation of heart rate variability during endotoxin shock in rabbits.
Crit Care Med 23:1694�1702.
GV-SOLAS (2008) Empfehlung der GV-SOLAS zur Substanzapplikation bei Versuchstieren. URL http:
//www.bv-solas.de/auss/tie/index.html, eingesehen am 01.07.2008.
Hainsworth R: The Control and Physiological Importance of Heart Rate. In: Malik M, Camm AJ (eds):
Heart Rate Variability. Futura Publishing Company, Inc., New York, 1995, pp 3�19.
Hansen MK, O'Connor KA, Goehler LE, Watkins LR, Maier SF (2001) The contribution of the vagus
nerve in interleukin-1β-induced fever is dependent on dose. Am J Physiol Regul Integr Comp Physiol
280:R929�934.
Hawkins P (2002) Recognizing and assessing pain, su�ering and distress in laboratory animals: a survey
of current practice in the UK with recommendations. Lab Anim 36:378�395.
Hayano J, Sakakibara Y, Yamada A, Yamada M, Mukai S, Fujinami T, Yokoyama K, Watanabe Y,
Takata K (1991) Accuracy of assessment of cardiac vagal tone by heart rate variability in normal
subjects. Am J Cardiol 67:199�204.
Heinroth KM, Kuhn C, Stache N, Witthaut R, Müller-Werdan U, Werdan K, Prondzinsky R (1999) Ein-
geschränkte Herzfrequenzvariabilität bei Patienten mit septischem und nicht-septischem Multiorgan-
Dysfunktions-Syndrom. Intensivmed 36:436�445.
Hesse DG, Tracey KJ, Fong Y, Manogue KR, Palladino MAJ (1988) Cytokine appearance in human
endotoxemia and primate bacteremia. SGO 166:147�153.
Hinshaw LB (1974) Role of the heart in the pathogenesis of endotoxin shock. J Surg Research 17:134�145.
Hottenrott K, Hoos O, Esperer HD (2006) Herzfrequenzvariabilität und Sport. Herz 31:544�552.
Houle MS, Billman GE (1999) Low-frequency component of the heart rate variability spectrum: a poor
marker of sympathetic activity. Am J Physiol 276:H215�H223.
Hoyt RFJ, Hawkins JV, St Claire MB, Kennet MB: Mouse Physiology. In: Fox G (ed): The Mouse in
Biomedical Research. Elsevier Amsterdam, Boston, Heidelberg and others, 2007, pp 23�90.
Huston JM, Gallowitsch-Puerta M, Ochani M, Ochani K, Yuan R, Rosas-Ballina M, Ashok M, Goldstein
RS, Chavan S, Pavlov VA, Metz CN, Yang H, Czura CJ, Wang H, Tracey KJ (2007) Transcutaneous
vagus nerve stimulation reduces serum high mobility group box 1 levels and improves survival in
murine sepsis. Crit Care Med 35:2762�2768.
Ishii K, Kuwahara M, Tsubone H, Sugano S (1996) Autonomic nervous function in mice and voles
(microtus arvalis): investigation by power spectral analysis of heart rate variability. Lab Anim 30:359�
364.

63
Janssen BJA, Leenders PJA, Smits JFM (2000) Short-term and long-term blood pressure and heart rate
variability in the mouse. Am J Physiol Regul Integr Comp Physiol 278:R215�R225.
Joaquim LF, Farah VM, Bernatova I, Fazan RJ, Grubbs R, Morris M (2004) Enhanced heart rate
variability and barore�ex index after stress and cholinesterase inhibition in mice. Am J Physiol Heart
Circ Physiol 287:H251�H257.
Just A, Faulhaber J, Ehmke H (2000) Autonomic cardiovascular control in conscious mice. Am J Physiol
Regul Integr Comp Physiol 279:R2214�R2221.
Kay SM, Marple SL (1981) Spectrum analysis�a modern perspective. Proc IEEE 69:1380�1914.
Kleiger RE, Stein PK, Bosner MS, Rottman JN: Time-Domain Measurements of Heart Rate Variability.
In: Malik M, Camm AJ (eds): Heart Rate Variability. Futura Publishing Company, Inc., New York,
1995, pp 33�45.
Klöckner U, Rueckschloss U, Grossmann C, Matzat S, Schumann K, Ebelt H, Müller-Werdan U, Loppnow
H, Werdan K, Gekle M (2014) Inhibition of cardiac pacemaker channel hHCN2 depends on intercala-
tion of lipopolysaccharide into channel-containing membrane microdomains. J Physiol 592:1199�1211.
Knaus WA, Draper EA, Wagner DP, Zimmerman JE (1985) APACHE II: a severity of disease classi�-
cation system. Crit Care Med 13:818�829.
Kneyber MCJ, Gazendam RP, Niessen HWM, Kuiper JW, Dos Santos CC, Slutsky AS, Plötz FB (2009)
Mechanical ventilation during experimental sepsis increases deposition of advanced glycation end
products and myocardial in�ammation. Crit Care 13:R87.
Kobayashi M, Musha T (1982) 1/f �uctuation of heartbeat period. IEEE Trans Biomed Eng 29:456�457.
Korach M, Sharshar T, Jarrin I, Fouillot JP, Raphaël JC, Gajdos P, Annane D (2001) Cardiac variability
in critically ill adults: in�uence of sepsis. Crit Care Med 29:1380�1385.
Kramer K, Kinter LB (2003) Evaluation and applications of radiotelemetry in small laboratory animals.
Physiol Genomics 13:197�205.
Kramer K, van Acker SA, Voss HP, Grimbergen JA, van der Vijgh WJ, Bast A (1993) Use of telemetry
to record electrocardiogram and heart rate in freely moving mice. J Pharmacol Toxicol Methods
30:209�215.
Kumar A, Thota V, Dee L, Olson J, Uretz E, Parrillo JE (1996) Tumor necrosis factor α and interleukin
1β are responsible for in vitro myocardial cell depression induced by human septic shock serum. J
Exp Med 183:949�958.
Kuss O, Schumann B, Kluttig A, Greiser KH, Haerting J (2008) Time domain parameters can be
estimated with less statistical error than frequency domain parameters in the analysis of heart rate
variability. J Electrocardiol 41:287�291.

64
La Rovere MT, Mortara A, Pinna GD, Bernardi L: Barore�ex Sensitivity and Heart Rate Variability
in the Assessment of the Autonomic Status. In: Malik M, Camm AJ (eds): Heart Rate Variability.
Futura Publishing Company, Inc., New York, 1995, pp 189�206.
Leon LR, Walker LD, DuBose DA, Stephenson LA (2004) Biotelemetry transmitter implantation in
rodents: impact on growth and circadian rhythms. Am J Physiol Regul Integr Comp Physiol 286:R967�
R974.
Levy MN (1971) Sympathetic-parasympathetic interactions in the heart. Circ Res 29:437�445.
Loppnow H, Flad HD, Rietschel ET, Brade H: The Active Principle of Bacterial Lipopolysaccharides
(Endotoxins) for Cytokine Induction. In: Schlag G, Redl H (eds): Pathophysiology of Shock, Sepsis,
and Organ Failure. Springer Berlin, Heidelberg, New York, 1993, pp 405�416.
Malik M, Bigger JT, Camm AJ, Kleiger RE, Malliani A, Moss AJ, Schwartz PJ (1996) Task Force of The
European Society of Cardiology and The North American Society of Pacing and Electrophysiology:
Heart rate variability standards of measurement, physiological interpretation, and clinical use. Euro
Heart J 17:354�381.
Malliani A, Pagani M, Lombardi F, S C (1991) Cardiovascular neural regulation explored in the frequency
domain. Circulation 84:482�492.
Malliani A, Lombardi F, Pagani M (1994) Power spectrum analysis of heart rate variability: a tool to
explore neural regulatory mechanisms. Heart 71:1�2.
Mansier P, Médigue C, Charlotte N, Vermeiren C, Coraboeuf E, Deroubai E, Ratner E, Chevalier B,
Clairambault J, Carré F, Dahkli T, Bertin B, Briand P, Strosberg D, Swynghedauw B (1996) Decre-
ased heart rate variability in transgenic mice overexpressing atrial β1-adrenoceptors. Am J Physiol
271:H1465�H1472.
van der Meer JWM, Endres S, Lonnemann G, Cannon JG, Ikejima T, Okusawa S, Gelfand J, Dinarello
CA (1988) Concentrations of immunoreactive human tumor necrosis factor alpha produced by human
mononuclear cells in vitro. J Leuk Biol 43:216�223.
Meng X, Ao L, Meldrum DR, Cain BS, Shames BD, Selzman CH, Banerjee A, Harken AH (1998) TNF-α
and myocardial depression in endotoxemic rats: temporal discordance of an obligatory relationship.
Am J Physiol 275:R502�R508.
Montano N, Ruscone TG, Porta A, Lombardi F, M P, Malliani A (1994) Power spectrum analysis of
heart rate variability to assess the changes in sympathovagal balance during graded orthostatic tilt.
Circulation 90:1826�1831.
Moodithaya S, Avadhany ST (2012) Gender di�erences in age-related changes in cardiac autonomic
nervous function. J Aging Res .

65
Müller-Höcker J, Haerty A: Pathomorphological Aspects of the Heart in Septic Patients. In: Schlag G,
Redl H (eds): Pathophysiology of Shock, Sepsis, and Organ Failure. Springer Berlin, Heidelberg, New
York, 1993, pp 853�858.
Müller-Werdan U, Reithmann C, Werdan K: Cytokines and the Heart: Molecular Mechanisms of Septic
Cardiomyopathy. Springer New York, 1996, pp 21�52.
Müller-Werdan U, Schumann H, Loppnow H, Fuchs R, Darmer D, Stadler J, Holtz J, Werdan K (1998)
Endotoxin and tumor necrosis factor α exert a similar proin�ammatory e�ect in neonatal rat cardio-
myocytes, but have di�erent cardiodepressant pro�les. J Mol Cell Cardiol 30:1027�36.
Müller-Werdan U, Buerke M, Christoph A, Flieger RR, Heinroth K, Herklotz A, Ruÿ M, Schlitt A,
Schmidt H, Sö�ker G, Werdan K (2006a) Septische Kardiomyopathie. Intensivmed 43:486�497.
Müller-Werdan U, Buerke M, Ebelt H, Heinroth KM, A H, Loppnow H, Ruÿ M, Schlegel F, Schlitt A,
Schmidt HB, Sö�ker G, Werdan K (2006b) Septic cardiomyopathy�A not yet discovered cardiomy-
opathy? Exp Clin Cardiol 11:226�236.
Natanson C, Fink MP, Ballantyne HK, MacVittie TJ, Conklin JJ, Parrillo JE (1986) Gram-negative
bacteremia produces both severe systolic and diastolic cardiac dysfunction in a canine model that
simulates human septic shock. J Clin Invest 78:259�270.
Natanson C, Danner R, Elin R, Hosseini J, Peart K, Banks S, MacVittie T, Walker R, Parrillo J (1989)
Role of endotoxemia in cardiovascular dysfunction and mortality. J Clin Invest 83:243�251.
Nishijima H, Weil MH, Shubin H, Cavanilles J (1973) Hemodynamic and metabolic studies on shock
associated with gram negative bacteremia. Medicine 52:287�294.
Odemuyiwa O: Heart Rate Variability and Risk Strati�cation after Acute Myocardial Infarction: Change
in Heart Rate Variability after Acute Myocardial Infarction. In: Malik M, Camm AJ (eds): Heart Rate
Variability. Futura Publishing Company, Inc., New York, 1995, pp 363�368.
Okusawa S, Gelfand JA, Ikejima T, Connolly RJ, Dinarello CA (1988) Interleukin 1 induces a shock-like
state in rabbits. Synergism with tumor necrosis factor and the e�ect of cyclooxygenase inhibition. J
Clin Invest 81:1162�1172.
Pagani M, Lombardi F, Guzzetti S, Rimoldi O, Furlan R, Pizzinelli P, Sandrone G, Malfatto G, Dell'Orto
S, Piccaluga E (1986) Power spectral analysis of heart rate and arterial pressure variabilities as a
marker of sympatho-vagal interaction in man and conscious dog. Circ Res 59:178�193.
Pagani M, Lucini D, Rimoldi R, Furlan R, Piazza S, Biancardi L: E�ects of Physical and Mental Exercise
on Heart Rate Variability. In: Malik M, Camm A (eds): Heart Rate Variability. Futura Publishing
Company, Inc., New York, 1995, pp 245�266.
Pancoto JA, Corrêa PB, Oliveira-Pelegrin GR, Rocha MJ (2008) Autonomic dysfunction in experimental
sepsis induced by cecal ligation and puncture. Auton Neurosci 138:57�63.

66
Parker M, Shelhamer J, Bacharach S, Green M, Natanson C, Frederick T, Damske B, Parrillo J (1984)
Profound but reversible myocardial depression in patients with septic shock. Ann Intern Med 100:483�
490.
Piepoli M, Garrard CS, Kontoyannis DA, Bernardi L (1995) Autonomic control of the heart and peri-
pheral vessels in human septic shock. Intensive Care Med 21:112�119.
Pilz G, McGinn P, Boekstegers P, Kääb S, Weidenhöfer S, Werdan K (1994) Pseudomonas sepsis does not
cause more severe cardiovascular dysfunction in patients than non-Pseudomonas sepsis. Circ Shock
42:174�182.
Pincus SM (1994) Greater signal regularity may indicate increased system isolation. Math Biosci 122:161�
181.
Pomeranz B, Macaulay RJB, Caudill MA, Kutz I, Adam D, Gordon D, Kilborn KM, Barger AC, Shannon
DC, Cohen RJ, Benson H (1985) Assessment of autonomic function in humans by heart rate spectral
analysis. Am J Physiol 248:H151�H153.
Pool R (1989) Is it healthy to be chaotic? Science 243:604�607.
Rechlin T (1995) Die Bedeutung von Herzfrequenzanalysen bei psychiatrischen Fragestellungen. Fortschr
Neurol Psychiatr 63:106�120.
Redl H, Schlag G, Bahrami S, Davies J, Waage A, Ceska M, Buurman WA, Adolf G: The Cytokine
Network in Trauma and Sepsis I: TNF and IL-8. In: Schlag G, Redl H (eds): Pathophysiology of
Shock, Sepsis, and Organ Failure. Springer Berlin, Heidelberg, New York, 1993, pp 468�490.
Reinhardt K, Brunkhorst FM, Bone HG, Gerlach H, Gründling M, Kreymann G, Kujath P, Marggraf G,
Mayer K, Meier-Hellmann A, Peckelsen C, Putensen C, Stüber F, Quintel M, Ragaller M, Rossaint R,
Weiler N, Welte T, Werdan K (2006) Diagnose und Therapie der Sepsis, S-2 Leitlinien der Deutschen
Sepsis-Gesellschaft e.V. (DSG) und der Deutschen Interdisziplinären Vereinigung für Intensiv- und
Notfallmedizin (DIVI). Clin Res Cadiol 95:429�454.
Reinhart K, Brunkhorst FM, Bone HG, Bardutzky J, Demp�e CE, Forst H, Gastmeier P, Gerlach H,
Gründling M, John S, Kern W, Kreymann G, Krüger P W Kujath, Marggraf G, Martin J, Mayer
K, Meier-Hellmann A, Oppert M, Putensen C, Quintel M, Ragaller M, Rossaint R, Seifert H, Spies
C, Stüber F, Weiler N, Weimann A, Werdan K, Welte T (2010) Prävention, Diagnose, Therapie und
Nachsorge der Sepsis. S2-Leitlinien der Deutschen Sepsis-Gesellschaft und der Deutschen Interdiszi-
plinären Vereinigung für Intensiv- und Notfallmedizin. URL http://www.awmf.org/leitlinien/detail/
ll/079-001.html, eingesehen am 12.09.2012.
Rimoldi O, Pierini S, Ferrari A, Cerutti S, Pagani M, Malliani M (1990) Analysis of short-term oscillations
of R-R and arterial pressure in conscious dogs. Am J Physiol 258:H967�H976.
Rubin LJ, Keller RS, Parker JL, Adams HR (1994) Contractile dysfunction of ventricular myocytes
isolated from endotoxemic guinea pigs. Shock 2:113�120.

67
Saeed RW, Varma S, Peng-Nemero� T, Sherry B, Balakhaneh D, Huston J, Tracey KJ, Al-Abed Y,
Metz CN (2005) Cholinergic stimulation blocks endothelial cell activation and leukocyte recruitment
during in�ammation. J Exp Med 201:1113�1123.
Savci V, Ulus IH (1997) Cardiovascular e�ects of central choline during endotoxin shock in the rat. J
Cardiovasc Pharmacol 30:667�675.
Saykrs BM (1973) Analysis of heart rate variability. Ergonomics 16:17�32.
Schaefer S (2005) Arrythmien und eingeschränkte Herzfrequenzvariabilität bei Patienten mit Score-
quanti�zierter Sepsis und Score-quanti�ziertem MODS�eine prospektive Studie auf einer internisti-
schen Intensivstation. Dissertation, Martin-Luther-Universität Halle-Wittenberg.
Schletter J, Heine H, Ulmer AJ, Rietschel ET (1995) Molecular mechanisms of endotoxin activity. Arch
Microbiol 164:383�389.
Schmidt H, Müller-Werdan U, Ho�mann T, Francis DP, Piepoli MF, Rauchhaus M, Prondzinsky R,
Loppnow H, Buerke M, Hoyer D, Werdan K (2005) Autonomic dysfunction predicts mortality in
patients with multiple organ dysfunction syndrome of di�erent age groups. Crit Care Med 33:1994�
2002.
Schmidt H, Saworski J, Werdan K, Müller-Werdan U (2007) Decreased beating rate variability of sponta-
neously contracting cardiomyocytes after co-incubation with endotoxin. J Endotoxin Res 13:339�342.
Schmidt H, Hoyer D, Hennen R, Heinroth K, Rauchhaus M, Prondzinsky R, Hottenrott K, Buerke M,
Müller-Werdan U, Werdan K (2008a) Autonomic dysfunction predicts both 1- and 2-month mortality
in middle-aged patients with multiple organ dysfunction syndrome. Crit Care Med 36:967�970.
Schmidt H, Müller-Werdan U, Werdan K: The Consequences of Cardiac Autonomic Dysfunction in
Multiple Organ Dysfunction Syndrome. In: Vincent J (ed): Yearbook of Intensive Care and Emergency
Medicine. Springer Berlin, Heidelberg, New York, 2008b, pp 55�64.
Schmidt HB, Werdan K, Müller-Werdan U (2001) Autonomic dysfunction in the ICU patient. Curr Opin
Crit Care 7:314�322.
Schroeder EB, Whitsel EA, Evans GW, Prineas RJ, Chambless LE, Heiss G (2004) Repeatability of
heart rate variability measures. J Electrocardiol 37:163�172.
Seely AJE, Christou NV (2000) Multiple organ dysfunction syndrome: exploring the paradigm of complex
nonlinear systems. Crit Care Med 28:2193�2200.
Shoemaker WC (1971) Cardiorespiratory patterns in complicated and uncomplicated septic shock: phy-
siologic alterations and their therapeutic implications. Ann Surg 174:119�125.
Shusterman V, Usiene I, Harrigal C, Lee JS, Kubota T, Feldman AM, London B (2002) Strain-speci�c
patterns of autonomic nervous system activity and heart failure susceptibility in mice. Am J Physiol
Heart Circ Physiol 282:H2076�H2083.

68
Silverman HJ, Penaranda R, Orens JB, Lee NH (1993) Impaired β-adrenergic receptor stimulation of cy-
clic adenosine monophosphate in human septic shock: association with myocardial hyporesponsiveness
to catecholamines. Crit Care Med 21:31�39.
Skillman JJ, Bushnell LSB, Goldman H, Silen W (1969) Respiratory failure, hypotension, sepsis, and
jaundice. Am J Surg 117:523�530.
Smaïl N, Descorps Declère A, Duranteau J, Vigué B, Samii K (1996) Left ventricular function after
severe trauma. Intensive Care Med 22:439�442.
Stauss HM (2007) Power spectral analysis in mice�what are the appropriate frequency bands? Am J
Physiol Regul Integr Comp Physiol 292:R902�R903.
Stein PK, Bosner MS, Kleiger RE, Conger BM (1994) Heart rate variability: a measure of cardiac
autonomic tone. Am Heart J 127:1376�1381.
Stieber J, Wieland K, Stöckl G, Ludwig A, Hofmann F (2006) Bradycardic and proarrhythmic properties
of sinus node inhibitors. Mol Pharmacols 69:1328�1337.
Su�redini AF, Fromm RE, Parker MM, Brenner M, Kovacs JA, Wesley RA, Parrillo JE (1989) The car-
diovascular response of normal humans to the administration of endotoxin. New Eng J Med 321:280�
287.
Sulakhe PV, Sandirasegarane L, Davis JP, Vo XT, Costain WJ, Mainra RR (1996) Alterations in ino-
tropy, nitric oxide and cyclic GMP synthesis, protein phosphorylation and ADP-ribosylation in the
endotoxin-treated rat myocardium and cardiomyocytes. Mol Cell Biochem 163:305�318.
Thireau J, Zhang B, Poisson D, Babuty D (2008) Heart rate variability in mice: a theoretical and practical
guide. Exp Physiol 93:83�94.
Tilney NL, Bailey GL, Morgan AP (1973) Sequential system failure after rupture of abdominal aortic
aneurysms: an unsolved problem in postoperative care. Ann Surg 178:117�122.
Tracey KJ (2007) Physiology and immunology of the cholinergic antiin�ammatory pathway. J Clin Invest
117:289�296.
Uechi M, Asai K, Osaka M, Smith A, Sato N, Wagner TE, Ishikawa Y, Hayakawa H, Vatner DE, Shannon
RP, Homcy CJ, Vatner SF (1998) Depressed heart rate variability and arterial barore�ex in conscious
transgenic mice with overexpression of cardiac Gsα. Circ Res 82:416�423.
Waage A, Redl H, Schlag G, Schade U: The Cytokine Network in Sepsis II: IL-1 and IL-6. In: Schlag G,
Redl H (eds): Pathophysiology of Shock, Sepsis, and Organ Failure. Springer Berlin, Heidelberg, New
York, 1993, pp 491�501.
Wang H, Ma S (2008) The cytokine storm and factors determining the sequence and severity of organ
dysfunction in multiple organ dysfunction syndrome. Am J Emerg Med 26:711�715.

69
Wang YH, Huang TS, Lin JL, Hwang JJ, Chan HL, Lai JS, Tseng YZ (2000) Decreased autonomic
nervous system activity as assessed by heart rate variability in patients with chronic tetraplegia. Arch
Phys Med Rehabil 81:1181�1184.
Weiergräber M, Henry M, Südkamp M, de Vivie ER, Hescheler J, Schneider T (2005) Ablation of
Cav2.3/E-type voltage-gated calcium channel results in cardiac arrhythmia and altered autonomic
control within the murine cardiovascular system. Basic Res Cardiol 100:1�13.
Werdan K, Schmidt H, Ebelt H, Zorn-Pauly K, Koidl B, Hoke RS, Heinroth K, Müller-Werdan U (2008)
Impaired regulation of cardiac function in sepsis, SIRS, and MODS. Can J Physiol Pharmacol 87:266�
274.
Williams TD, Chambers JB, Gagnon SP, Roberts LM, Henderson RP, Overton JM (2003) Cardiovascular
and metabolic responses to fasting and thermoneutrality in Ay mice. Physiol Behav 78:615�623.
Witte K, Engelhardt S, Janssen BJ, Lohse M, Lemmer B (2004) Circadian and short-term regulation of
blood pressure and heart rate in transgenic mice with cardiac overexpression of the β1-adrenoceptor.
Chronobiol Int 21:205�216.
Zorn-Pauly K, Pelzmann B, Lang P, Mächler H, Schmidt H, Ebelt H, Werdan K, Koidl B, Müller-Werdan
U (2007) Endotoxin impairs the human pacemaker current If. Shock 28:655�661.
van Zutphen L, Baumans V, Beynen A: Grundlagen der Versuchstierkunde: ein Beitrag zum tierschutz-
gerechten Umgang mit Versuchstieren und zur Qualität tierexperimenteller Ergebnisse. Stuttgart,
1995.

70
8 Thesen
1. Die prognoserelevante Einschränkung der Herzfrequenzvariabilität (HRV) bei In-
tensivpatienten mit einem Multiorgan-Dysfunktions-Syndrom (MODS) kann das
Resultat einer gestörten Organkommunikation durch Fehlregulation des autono-
men Nervensystems sein.
2. Die im Zellkulturexperiment nachgewiesene direkte Interaktion von Lipopolysac-
charid (LPS) mit dem Schrittmacherstrom If stellt einen möglichen Mechanismus
dar, der an der gestörten Herzfrequenzregulation beteiligt sein könnte.
3. Für die Überprüfung dieser Hypothese am Mausmodell, wird ein standardisiertes
Verfahren für die telemetrische Registrierung von Biopotentialen an Mäusen als
tierexperimentelle Methode am Universitätsklinikum Halle (Saale) etabliert.
4. Die postoperative Stabilisierung der Mäuse nach Implantation der EKG-Transmitter
und der Aufzeichnungen sind mit den Ergebnissen anderen Arbeitsgruppen ver-
gleichbar.
5. Die für die Beurteilung des LPS-E�ekts notwendige Charakterisierung der autono-
men Regulation der Herzfrequenz unter Normalbedingungen ergibt einen ausgegli-
chenen autonomen Ruhetonus bei ICR-Wildtyp-Mäusen.
6. Der Parasympathikus hat einen entscheidenden Ein�uss auf die Entstehung der
HRV.
7. Endotoxinapplikation führte unmittelbar zu einer Tachykardisierung sowie zu einer
starken Reduktion sämtlicher HRV-Parameter bei ICR-Wildtyp-Mäusen.

71
8. Endotoxin unter pharmakologischer autonomer Blockade führt dagegen zu einer
ausgeprägten Bradykardisierung, die HRV-Einschränkung ist weniger stark ausge-
prägt.
9. Die in Zellkuturexperimenten nachgewiesene sympathikusbahnende Wirkung des
Endotoxins und der direkt blockierende E�ekt auf den If -Strom, erkennbar un-
ter autonomer Blockade, können somit in vivo in funktioneller Hinsicht bestätigt
werden.

Anhang
Curriculum Vitae
Name Sara Ruccius, geb. Hettrich
Geburtsdatum 08.10.1984
Geburtsort Berlin
Email [email protected]
Aktuelle Tätigkeit
Seit 01/2014 Assistenzärztin am Kardiocentrum der Klinik Rotes Kreuz, Frank-
furt am Main
10/2012-12/2013 Assistenzärztin in der Klinik für Kardiologie und Nephrologie im
Helios Klinikum Berlin-Buch
Ausbildung
04/2012�06/2012 2. Abschnitt der Ärztlichen Prüfung, Note 1,5
10/2010�09/2012 Studium der Humanmedizin an der Charité Berlin
08/2007�09/2007 1. Abschnitt der Ärztlichen Prüfung, Note 1,5
10/2005�09/2010 Studium der Humanmedizin an der Martin-Luther-Universität
Halle-Wittenberg
10/2004�09/2005 Studium der Publizistik und Musikwissenschaft an der HU und FU
Berlin
06/2004 Abitur an der Georg-Friedrich-Händel-Oberschule Berlin, Note 1,3
Stipendien
03/2008�06/2012 Stipendiatin der Studienstiftung des deutschen Volkes
Sprachen
Englisch Gute Kenntnisse in Wort und Schrift
Latein Caesar-Lektüre
Frankfurt, den 10.08.2015 Sara Ruccius

Selbständigkeitserklärung
Hiermit erkläre ich an Eides statt, dass ich die vorliegende Dissertation selbstständig und
nur unter Verwendung der angegeben Literatur und Hilfsmittel angefertigt habe.
Frankfurt, den 10.08.2015 Sara Ruccius

Erklärung über frühere Promotionsversuche
Hiermit erkläre ich, dass ich bisher keine Promotionsversuche unternommen habe und
dass an keiner anderen Fakultät oder Universität ein Promotionsverfahren anhängig ist.
Frankfurt, den 10.08.2015 Sara Ruccius

Danksagung
Ich bedanke mich besonders bei meinem Doktorvater Prof. Dr. med. Henning Ebelt für
die vielen zielführenden Ratschläge und richtungsweisenden Denkanstöÿe, nicht zuletzt
für die langanhaltende Geduld. Weiterhin danke ich allen Kollegen und Kommilitonen,
insbesondere Prof. Dr. med. Karl Werdan, Prof. Dr. rer. nat. Harald Loppnow, Dr. rer.
medic. Ying Zhang, Volker Otto und Dr. Ing. Holger Lehnich, für die gemeinsame For-
schungsarbeit, auÿerdem dem Doktorandenkolloquium der Universitätsklinik Halle (Saa-
le), in welchem ebenfalls anregend diskutiert werden konnte. Ich bedanke mich sehr bei
meinem Mann und meinem Sohn, meinen Eltern und meiner Schwester.
Frankfurt, den 10.08.2015 Sara Ruccius


















![Ernährungstherapie des Intensivpatienten · o Di-oder Tripeptide im oberen Dünndarm besser resorbiert als einzelne AS o keine Ballaststoffe [1] Ernährungstherapie des Intensivpatienten](https://img.pdfslide.org/doc/110x75/5d4695c888c993a5648c8031/ernaehrungstherapie-des-o-di-oder-tripeptide-im-oberen-duenndarm-besser-resorbiert.jpg)










