Embed Size (px)
Citation preview

Institut fur Allgemeine Botanik der Universitat Hamburg
Der EinfluB von Kinetin auf das Wachstum von Fritschiella tuberosa IYENG. (Chaetophorineae, Chlorophyceae) in axenischer Massenkultur
Effect of Kinetin on Growth of Fritschiella tuberosa lYENG.
(Chaetophorineae, Chlorophyceae) in axenic mass culture
MICHAEL MELKONIAN und ADOLF WEBER
Mit 1 Abbildung
Eingegangen am 5. Marz 1975
Summary
1. Using antibiotics axenic cultures of Fritschiella tuberosa were obtained. In contrast to
previous methods incubation with antibiotics was performed in darkness, the incubation medium and temperature being chosen as to stimulate bacterial metabolism. With this method high concentrations of antibiotics could be applied without damaging the alga to a greater extent.
2. Mass induction of zoospores of Fritschiella was obtained by supplying agar-grown cultures with liquid medium. Zoospores were used as a uniform inoculum for mass culture.
3. Under the conditions of semicontinuous culture a growth curve of Fritschiella was taken down and the growth rate at the chosen test conditions calculated to be about 0.7 (k).
4. Kinetin did not show any effect on growth rate during logarithmic growth over a wide range of concentrations (10-5-10-9 g/m!). High concentrations of Kinetin (2 X 10-5 g/ml) inhibited the growth rate by 40010, the contents of chloroplast pigments I dry weight were reduced by 20 0/0. These inhibitory effects were not observed with similar concentrations of Adenin (10-4 M).
5. Experiments have shown that Kinetin is probably decomposed by some bacteria and not stable in sterile aquous media for longer than two months in the light.
Key words: Axenic mass culture, Zoospore induction, Kinetin, Chlorophyceae, Fritschiella.
Zusammenfassung
1. Fritschiella tuberosa wurde durch Behandlung mit Antibiotika in axenische Kultur genommen.
2. Eine Methode zur Masseninduktion von Zoosporen bei Fritschiella, ausgehend von alteren Agar-Kulturen, wird beschrieben.
Z. Pflanzenphysiol. Bd. 76. S. 120-129. 1975.

Einflu6 von Kinetin auf Fritschiella 121
3. Unter den Bedingungen der Massenkultur wurde von Fritschiella tuberasa eine Wachsturnskurve aufgenornrnen und die Wachsturnsrate (k) bestirnrnt. Sie betragt w1lhrend der exponentiellen Phase unter den gew1lhlten Versuchsbedingungen 0,7.
4. Kinetin beeinflu6t die Wachsturnsrate in einern Konzentrationsbereich von 10-5 bis 10-9 g/rnl nicht. Hahere Kinetinkonzentrationen (2 X 10-5 g/rnl) hernrnen die Wachsturnsrate bereits urn 40 010. Hohe Adeninkonzentrationen (10-4 M) zeigen hingegen keinen Einflu6 auf die Wachsturnsrate.
5. Kinetin kann von Bakterien abgebaut werden und ist in w1l6riger, steriler Lasung irn Licht nicht l1lnger als zwei Monate stabil.
Einleitung
Cytokinin-Aktivitat ist in Algen-Extrakten mehrfach festgestellt worden (BENTLEYMOWAT und REID, 1968; HUSSAIN und BONEY, 1969; JENNINGS, 1969; VAN STADEN und BREEN, 1973), jedoch ist liber die Wirkung dieser Phytohormone auf Algen wenig bekannt (Zusammenfassung bei AUGIER, 1974).
Exogene Kinetingaben fOrdern das Wachs tum verschiedener hoch organisierter Braun-, Rot- und Grlinalgen (PROVASOLI, 1958; PEDERSEN, 1968; JENNINGS, 1969; FRIES, 1973), liber den Einflug von Kinetin auf einzellige Algen und einfache ZellFaden liegen jedoch widersprlichliche Ergebnisse vor: In einigen Fallen wird eine Forderung von Wachs tum und Zellteilung (BENTLEy-MoWAT und REID, 1967; OLSZEWSKA, 1958), in anderen kein Einflug von Kinetin liber einen weiten Konzentrationsbereich beobachtet (PROVASOLI, 1958; TAMIYA et al., 1962).
Die unterschiedlichen Untersuchungsergebnisse dlirften teilweise darauf zurlickzuflihren sein, dag die Versuchsbedingungen nur ungenligend standardisiert waren und das Untersuchungsmaterial sich nicht immer im gleichen physiologischen Zustand befand. So stellten LIEN et al., (1971) fest, dag bei Synchronkulturen von Chlorella pyrenoidosa im Gegensatz zu frliheren Ergebnissen kein fordernder Einflug von fJIndolyl-Essigsaure und Gibberellinsaure auf Wachstum und Zellteilung liber einen weiten Konzentrationsbereich auftrat. TAMIYA et al., (1962) waren die einzigen, die bei ihren Kinetin-Tests Synchronkulturen (von Chlarella ellipsoidea) verwendeten.
In der vorliegenden Arbeit wurde versucht, den Einflug von Kinetin auf das Wachs tum der hoch differenzierten Chaetophoracce Fritschiella tuberosa unter solchen Bedingungen zu prlifen, wie sie flir die Kultur einzelliger Algen schon seit langerem liblich sind. Hierflir muihen erhebliche methodische Schwierigkeiten liberwunden werden.
Material und Methoden
a) Kultur der Alge: Fritschiella tuberasa1) wurde entweder in beliifteten ErlenrneyerGef1l6en in einer Klirnakarnrner oder in entsprechenden Kulturrahren in Lichttherrnostaten der Fa. Kniese (Marburg) angezogen. Als Kulturrnediurn verwendeten wir eine anorganische
1) Herrn Dr. KOCH (Gattingen), der uns die Alge freundlicherweise zur Verfiigung stellte, danken wir sehr herzlich.
z. PJlanzenphysial. Bd. 76. S. 120-129. 1975.

122 M. MELKONIAN und A. WEBER
Nahrlosung (KESSLER und CZYGAN, 1970), deren pH-Wert vor dem Autoklavieren auf 5,5 eingestellt wurde. Die Be1euchtungsdauer betrug durchschnittlich 14 Stunden, die Lichtintensitat schwankte (abhangig von der Versuchsanordnung) zwischen 4 000 und 7500 Lux. Die Temperatur in der Klimakammer hie1ten wir zwischen 16 und 18°C, im Lichtthermostaten belief sie sich auf 20 ± 1 0c. Zur Herstellung fester Nahrboden wurde die anorganische Nahrlosung mit 1,5 Ofo Agar versetzt.
b) Herstellung axenischer Kulturen: Die iiblichen mechanischen Verfahren wie Ausplattieren bzw. Ausstreichen der Algen auf Agar oder die Isolierung mit Hilfe einer Mikrokapillare fiihrten nicht zum Erfolg. Eine Zerkleinerung der Fadenzellen durch Ultraschallbehandlung war ohne starke Schadigung der Zellen eben falls nicht moglich.
Wir haben deshalb die Algen mit Antibiotika-Losungen behandelt, jedoch im Gegensatz zu den iiblichen Verfahren unsere Versuche bei relativ hohen Temperaturen und im Dunkeln2)
durchgefiihrt. Es wurden folgende Antibiotika verwendet: Penicillin-G-Kaliumsalz (Fa. Serva), Streptomycinsulfat, Neomycinsulfat und Chloramphenicol (jeweils Fa. Boehringer).
Folgende Versuchsanordnung fiihrte bei alteren Kulturen (5 Wochen bis 9 Monate) zu axenischen Algen:
1. Behandlung mit Chloramphenicol (2 000 ,ug/ml) fiir 24 Stunden bei Zimmertemperatur. Dieser erste Schritt wurde in einer anorganischen Nahrlosung, der einige Tropfen einer 1 0/0-
igen Glucose-Pepton-Losung zugesetzt worden waren, durchgefiihrt. 2. Behandlung fiir 24 Stunden in dem angeg. Bakterien-Standard-Medium3) bei 30 °C unter
Zusatz folgender Antibiotika
Penicillin Streptomycin Neomycin
3 000 ,ug/ml 500 ,ug/ml 500 ,ug/ml
3. Erhohung der Streptomycin- und Neomycinkonzentration auf jeweils 1 500 ,ug/ml (Behandlung ebenfalls 24 Stunden).
Zwischen jedcr Behandlung wurden die Algen mit steriler, anorganischer Nahrlosung gewas chen und fiir 12 Stunden in antibiotikafreiem Bakterien-Medium bei 30 °C gehalten. Daran anschlieBend wurde das jeweilige Antibiotikagemisch aseptisch zupipettiert. Nach der 3. Behandlung wurden die Algen erneut gewaschen, in anorganisches Fliissigmedium iiberfiihrt, und im Licht (unter Beliiftung) weiterkultiviert.
c) Sterilitiitstests: Die Sterilitat der Stammkulturen wurde regelmaBig mit den folgenden Testmedien iiberpriift:
1. Bakterien-Standard-Nahrboden (in Anlehnung an DREWS, 1968): Die anorganische Fritschiella-Nahrlosung wurde mit folgenden Zusatzen versehen: 0,8 % Pep ton, 0,1 0/0
Fleischextrakt, 0,1 % Hefeextrakt, 0,1 % Glucose, 1,5 % Agar. Der pH-Wert wurde vor dem Sterilisieren auf 7 eingestellt.
2. Bakterien-Standard-Nahrboden (verdiinnt): Mit Ausnahme des Agars wurden alle Zusatze nur zu 1/100 der o. a. Menge der Nahrlosung zugesetzt.
3. BREWER-thioglycollate-medium (Difco). Die Medien 1. und 2. wurden auch als Fliissigmedien verwandt und dann ohne Agar
bereitet. Die Sterilitatstests wurden jeweils bei drei verschiedenen Temperaturen (16-18 °C im
Licht, 20 und 32°C im Dunkeln) iiber einen Zeitraum von 2 Wochen durchgefiihrt. d) Zoosporeninduktion: Die Zoosporenbildung konnte auf verschiedene Weise induziert
werden. Ausreichende Mengen wurden stets dann erhalten, wenn eine altere Algenkultur,
2) Urn den Photosynthese-Stoffwechsel der Alge weitgehend auszuschalten. 3) Urn moglichst alle Bakterien in Teilungsaktivitat zu versetzen.
Z. P/lanzenphysiol. Bd. 76. S. 120-129. 1975.

EinfluB von Kinetin auf Fritschiella 123
die auf einem festen Nahrboden gewachsen war, mit frischer Nahrlasung iiberschichtet und anschlieBend beliiftet wurde (naheres s. Kap. Ergebnisse).
e) Wachstumsmessungen - Versuche mit Kinetin: Als MaB fiir das Wachs tum diente die Trockengewichtszunahme/Zeit einer exponentiell wachsenden Kultur. Zur Trockengewichtsbestimmung wurden jeweils die Algen aus einer Kulturrahre auf einem Membranfilter (Cb 12 ,urn) gesammelt und bei 85°C bis zur Gewichtskonstanz getrocknet. Zur Ermittlung der Wachstumsrate wurden die Mittelwerte aus mindestens 5 Trockengewichtsbestimmungen im Abstand von 48 Stun den verwendet. Die Berechnung der Wachstumsraten erfolgte unter Verwendung von Logarithmen zur Basis 2 als «Zahl der Verdoppelungen pro Tag» (MONOD, 1949; HOOGENHOUT und AMESZ, 1965).
Urn eine fardernde oder hemmende Wirkung des Kinetins auf die Wachstumsrate zu priifen, wurde dieses in definierten Mengen dem Fritschiella-Anzuchtmedium als salzsaure Lasung vor der pH-Wert-Einstellung und vor dem Autoklavieren zugesetzt (MILLER, 1963). Mehr als 3 X 10-5 g/ml Kinetin lassen sich in der Nahrlasung nicht lasen.
Ergebnisse
a) Zoosporeninduktion: Fritschiella tuberosa ist eine terrestrische Griinalge, die unter natiirlichen Bedingungen ein sehr kompliziertes Fadensystem ausbildet (vgl. IYENGAR, 1932; SINGH, 1941, 1947). 1m Laborexperiment wird dieser Differenzierungsgrad kaum erreicht (PROSKAUER und BERMAN, 1970), vor allem dann nicht, wenn die Alge in Fliissigmedien kultiviert wird (McBRIDE, 1970). Es konnen allerdings, in Abhangigkeit vom Alter der Kultur, auch in Aquakulturen samtliche Zelltypen der «Wildform» beobachtet werden, jedoch nicht in der Form des fiir Fritschiella typischen Thallus (WEBER, 1975).
Fiir die im folgenden beschriebenen Kinetintests war die Morphologie unseres Objekts aus zwei Griinden interessant: Erstens wollten wir herausfinden, ob Kinetingaben den Differenzierungsgrad von Fritschiella tuberosa beeinflussen, zweitens muBten wir das Problem losen, von einer verzweigt-fadigen Alge ein homogenes Inokulum herzustellen.
Das einzige beschriebene Verfahren zur Gewinnung einer definierten Animpfmenge bei verzweigt-fadigen Griinalgen besteht darin, die Algen in einem Mixer zu homogenisieren (BOALCH, 1961; HINDAK, 1970). Diese Prozedur ist jedoch bei steriler Durchfiihrung auBerordentlich aufwendig und hat zudem den Nachteil, daB viele Zellen zerstort werden. Urn ein homogenes Inokulum herzustellen, benotigten wir ein Verfahren zur reproduzierbaren Gewinnung groBer Zoosporenmengen.
Die Bedingungen, die bei vielen Algen zur Bildung beweglicher Einzelzellen fiihren, sind mehrfach zusammenfassend beschrieben worden (z. B. KLEBS, 1896; ERBEN, 1962; DRING, 1974). Viele dieser Methoden fiihrten auch bei Fritschiella zur Induktion von Zoosporen; allerdings war die Ausbeute fiir unsere Zwecke meist zu gering. KLEBS (1896) erzielte bei den terrestrischen Algen Vaucheria rep ens, Protosiphon botryoides und Botrydium tuberosum Zoosporenbildung, wenn er Agar-Kulturen mit einem fliissigen Nahrmedium iiberschichtete. Mit einem ahnlichen Verfahren hatten wir auch bei Fritschiella Erfolg:
z. PJlanzenphysiol. Bd. 76. S. 120-129. 1975.
124 M. MELKONIAN und A. WEBER
Algen aus einer gut wachsenden, beliifteten Stammkultur wurden auf ein mineralisches Agar-Medium geimpft (in Petrischalen), und unter Bedingungen kultiviert, die den spateren Versuchsbedingungen entsprachen. Nach durchschnittlich 2 Wochen wurden bewachsene Agarstiicke4
) steril ausgeschnitten, in einen Erlenmeyer-Kolben mit frischer anorganischer Fritschiella-Nahrlosung gebracht und beliiftet. Die Induktion der Zoosporenbildung wurde am Lichtthermostaten in der Mitte der Lichtphase angesetzt. Das Alter der zum Beimpfen verwendeten Einzelzellen betrug jeweils 22-26 Stunden.
Diese Methode beansprucht ein Minimum an Arbeitsaufwand, die Induktion fiihrt zur Freisetzung von Zoosporen aus fast jeder Kurzzelle und eine Kontamination durch Fremdorganismen ist sehr sclten. Zellfaden wurden nur ausnahmsweise in der Suspension beobachtet.
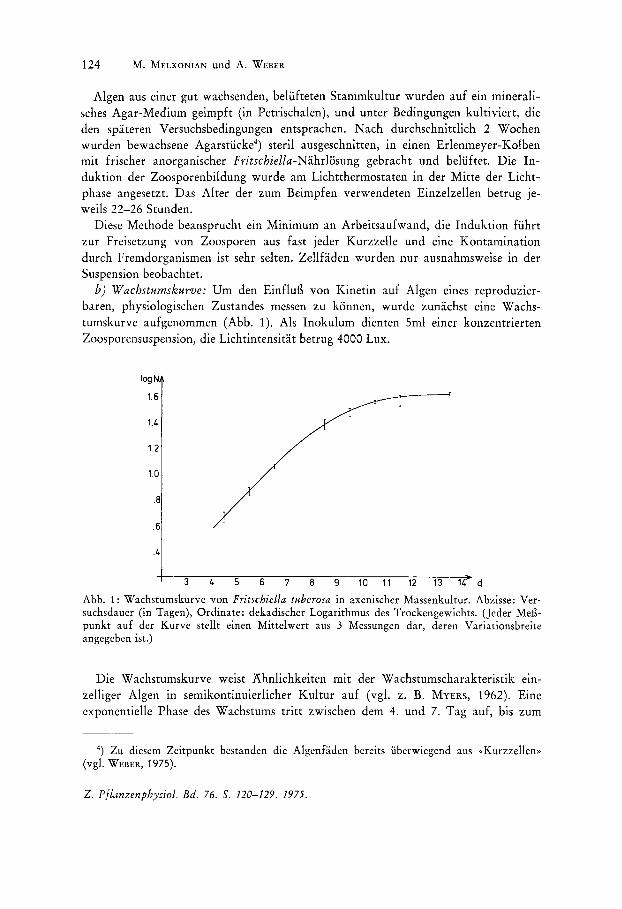
b) Wachstumskurve: Urn den EinfluB von Kinetin auf Algen eines reproduzierbaren, physiologischen Zustandes messen zu konnen, wurde zunachst eine Wachstumskurve aufgenommen (Abb. 1). Als Inokulum dienten Sml einer konzentrierten Zoosporensuspension, die Lichtintensitat betrug 4000 Lux.
(ogN
1.5
1.4
1.2
1.0
.8
.6
.4
3 4 5 6 7 8 9 10 11 12 13 14 d
Abb. 1: Wachstumskurve von Fritschiella tuberosa in axenischer Massenkultur. Abzisse: Versuchsdauer (in Tagen), Ordinate: dekadischer Logarithmus des Trockengewichts. (Jeder MeEpunkt auf der Kurve stellt einen Mittelwert aus 3 Messungen dar, deren Variationsbreite angegeben ist.)
Die Wachstumskurve weist Ahnlichkeiten mit der Wachstumscharakteristik e1l1-zelliger Algen in semikontinuierlicher Kultur auf (vgl. z. B. MYERS, 1962). Eine exponentielle Phase des Wachstums tritt zwischen dem 4. und 7. Tag auf, bis zum
4) Zu diesem Zeitpunkt bestanden die Algenfaden bereits iiberwiegend aus «Kurzzellen» (vgl. WEBER, 1975).
Z. Pjlanzenphysiol. Bd. 76. S. 120-129. 1975.

EinfluB von Kinetin auf Fritschiella 125
12. Tag schlie£h sich ein linearer Anstieg des Trockengewichts an und nach dem 12. Tag verandert sich das Trockengewicht nicht mehr (stationare Phase). Die Dbergange zwischen den einzelnen Ph as en sind gleitend. Aus dem Anstieg des Trockengewichts wahrend der exponentiellen Phase lie£~ sich eine Wachstumsrate von k = 0,7 berechnen.
Bei allen folgenden Versuchen wurden die Bestimmungen der Wachstumsraten wahrend der exponentiellen Wachstumsphase (am 4. und 5. bzw. 6. Tag) durchgefuhrt.
c) Vergleich des Wachstums steriler und unsteriler Kulturen: Zunachst wollten wir herausfinden, ob durch die Anwendung der Antibiotika-L6sungen Schaden in der physiologischen Leistung der Algen aufgetreten waren. Wir haben Cleshalb die Wachstumsraten axenischer und unbehandelter Kulturen verglichen (Tab. 1). Die Ergebnisse zeigen, dag eine dauerhafte Schadigung der Algen irn Zuge der Herstellung axenischer Kulturen nicht aufgetreten war.
Tab. 1: Vergleich des Wachstums zwischen sterilen und unsterilen Fritschiella-Kulturen w1ihrend der exponentiellen Wachstumsphase.
Erntetag sterile Kultur unsterile Kultur Trockengew. in mg Trockengew. in mg
4. Tag 5,5 (0,05)1) 6,2 (0,18) 6. Tag 15,9 (2,44) 16,5 (8,4) k 2
) 0,77 0,71
1) (): Varianz des Mittelwerts der Trockengewichte am jeweiligen Erntetag. 2) Wachstumsrate.
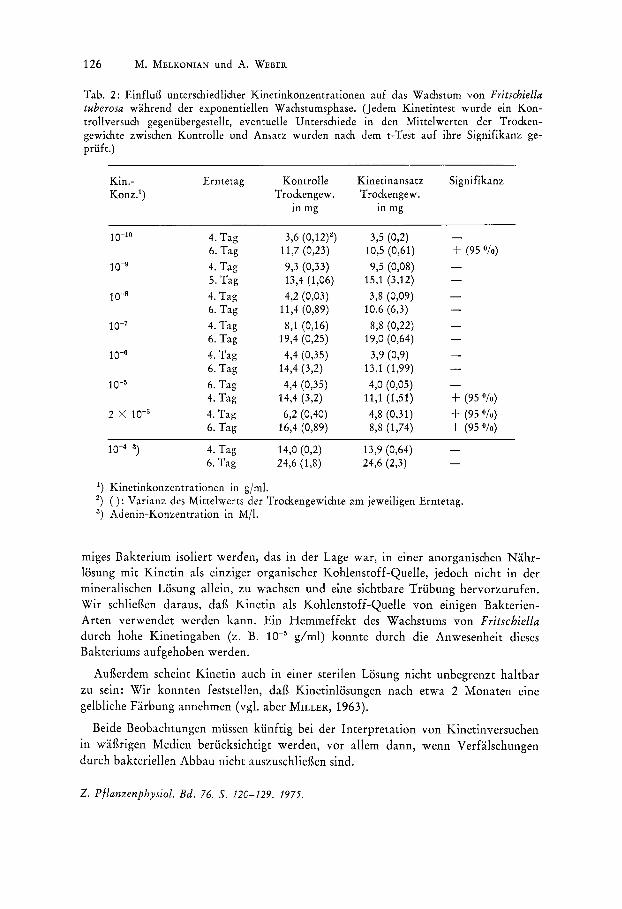
d) Einflufl von Kinetin auf das Wachstum: In Tab. 2 ist der Einflug von unterschiedlichen Kinetinkonzentrationen auf das Wachs tum von Fritschiella tuberosa wahrend der exponentiellen Wachsturnsphase angegeben.
Die Ergebnisse zeigen, dag Kinetin uber einen weiten Konzentrationsbereich (10-9
bis 10-6 g/rnl) keinen Einflug auf die Wachstumsrate von Fritschiella besitzt. Bei h6heren Kinetinkonzentrationen als 10-5 g/ml, dies scheint die Grenzkonzentration der Hemmung zu sein, kommt es zu einer Hemmung der Wachstumsrate urn durchschnittlich 40 Ofo (Mittelwert aus drei verschiedenen Versuchsansatzen). Noch h6here Kinetinkonzentrationen irn Nahrrnediurn (10-4 g/rnl) fuhren zu starken, rnorphologischen Veranderungen der Alge. Urn festzustellen, ob die Hemmung des Wachstums bei hohen Kinetinkonzentrationen ein unspezifischer Effekt ist, wurde ein Test mit einer Adenin-Konzentration gleicher Molaritat durchgefuhrt. 10-4 M Adenin irn Nahrmediurn fuhrt jedoch nicht zu einer Beeintrachtigung des Wachsturns von Fritschiella tuberosa.
e) Stabilitat des Kinetins in waflriger Losung: Aus einer durch Bakterien verunreinigten Fritschiella-Kultur konnte auf Bakterien-Standard-Agar ein stabchenf6r-
Z. P/lanzenphysiol. Bd. 76. S. 120-129. 1975.
126 M. MELKONIAN und A. WEBER
Tab. 2: Einflu13 unterschiedlicher Kinetinkonzentrationen auf das Wachstum von Fritschiella tuberosa wahrend der exponentiellen Wachstumsphase. (Jedem Kinetintest wurde ein Kontrollversuch gegeniibergestellt, eventuelle Unterschiedc in den Mittelwerten der Trockengewichte zwischen Kontrolle und Ansatz wurden nach dem t-Test auf ihre Signifikanz gepriift.)
Kin.Konz.1
)
10-5
Erntetag
4. Tag 6. Tag
4. Tag 5. Tag
4. Tag 6. Tag
4. Tag 6. Tag
4. Tag 6. Tag
6. Tag 4. Tag
4. Tag 6. Tag
4. Tag 6. Tag
1) Kinetinkonzentrationen in g/ml.
Kontrolle Trockengew.
mmg
3,6 (0,12?) 11,7 (0,23)
9,3 (0,33) 13,4 (1,06)
4,2 (0,03) 11,4 (0,89)
8,1 (0,16) 19,4 (0,25)
4,4 (0,35) 14,4 (3,2)
4,4 (0,35) 14,4 (3,2)
6,2 (0,40) 16,4 (0,89)
14,0 (0,2) 24,6 (1,8)
Kinetinansatz Trockengew.
mmg
3,5 (0,2) 10,5 (0,61)
9,5 (0,08) 15,1 (3,12)
3,8 (0,09) 10,6 (6,3)
8,8 (0,22) 19,0 (0,64)
3,9 (0,9) 13,1 (1,99)
4,0 (0,05) 11,1 (1,51)
4,8 (0,31) 8,8 (1,74)
13,9 (0,64) 24,6 (2,3)
Signifikanz
+ (95 Ofo)
+ (95 Ofo)
+ (95 Ofo) + (95 Ofo)
2) (): Varianz des Mittelwerts der Trockengewichte am jeweiligen Erntetag. 3) Adenin-Konzentration in MIl.
miges Bakterium isoliert werden, das in der Lage war, in einer anorganischen NahrlOsung mit Kinetin als einziger organischer Kohlenstoff-Quelle, jedoch nicht in der mineralischen Lasung allein, zu wachsen und eine sichtbare Triibung hervorzurufen. Wir schliegen daraus, dag Kinetin als Kohlenstoff-Quelle von einigen BakterienArten verwendet werden kann. Ein Hemmeffekt des Wachstums von Fritschiella
durch hohe Kinetingaben (z. B. 10-5 g/ml) konnte durch die Anwesenheit dieses Bakteriums aufgehoben werden.
Augerdem scheint Kinetin auch in einer sterilen Lasung nicht unbegrenzt haltbar zu sein: Wir konnten feststellen, dag KinetinlOsungen nach etwa 2 Monaten eine gelbliche Eirbung annehmen (vgl. aber MILLER, 1963).
Beide Beobachtungen mlissen kiinftig bei der Interpretation von Kinetinversuchen in wagrigen Medien beriicksichtigt werden, vor allem dann, wenn Verfalschungen durch bakteriellen Abbau nicht auszuschliegen sind.
z. Pjlanzenphysiol. Bd. 76. S. 120-129. 1975.

EinfluB von Kinetin auf Fritschiella 127
Diskussion
Etliche vielzellige, marine Algen reagieren auf eine Zufuhr von Phytohormonen mit einer Wachstumssteigerung. So fordert Kinetin das Wachs tum von Porphyra tenera (IWASAKI, 1965), Ectocarpus fasciculatus und Pylaiella litoralis (PEDERSEN, 1968), Ecklonia radiata und Hypnea musciformis (JENNINGS, 1969), Viva lactuca (PROVASOLI, 1958), Polysiphonia urceolata und Goniotrichum alsidii (FRIES, 1973) sowie Acetabularia mediterranea (ZETSCHE, 1963). Die wirksame Kinetinkonzentration liegt dabei im allgemeinen zwischen 10-6 und 10-8 g/m!. Interessant ist in dies em Zusammenhang, daB auch andere Purine wachstumsfordernd wirken konnen (IWASAKI, 1965; PROVASOLI, 1958; HAWKINS, 1966; FRIES, 1973). Kombinationen von Adenin und Kinetin zeigen haufig die beste Wirkung auf das Wachstum. Adenin wird dabei meistens in 20fach hoherer Konzentration als Kinetin angewandt.
Fur einzellige Algen liegen dagegen teilweise andere Ergebnisse vor. TAMIYA et a!. (1962) beobachteten weder einen fordernden noch einen hemmenden Einflug auf das Wachstum synchronisierter Chlorella ellipsoidea bei Kinetingaben in Hohe von 2 X 10-5 g/m!. BENTLEy-MOWAT und REID (1967) berichten dagegen, daB Kinetin bei einer Konzentration von 10-4 bis 10-5 g/ml fordernd auf das Wachstum zweier Phytoplankter wirkt (Gymnodinium splendens und Phaeodactylum tricornutum). Die genannten Autoren haben aber den EinfluB von Kinetin erst nach Ablauf der logarithmischen Wachstumsphase bestimmt. Auch ist nicht sicher, ob wahrend der Versuchsdurchfuhrung axenische Bedingungen herrschten. Bakterien konnten die Versuchsergebnisse aber entscheidend beeinflussen: Wir haben wahrend unserer Untersuchungen ein Bakterium isoliert, das vermutlich Kinetin abbaut. Urn Verfalschungen dieser Art zu vermeiden, haben wir ausschliemich axenische Massenkulturen fur unsere Tests herangezogen.
In der vorliegenden Arbeit wird erstmals ein Weg beschrieben, wie auch aus verzweigt-fadigen Grunalgen ein standardisiertes Inokulum fur axenische Massenkulturen hergestellt und anschlieBend der EinfluB eines Wuchsstoffes getestet werden kann.
Kinetin hat bei Gaben in Hohe von 10-9 bis 10-6 g/ml keinen EinfluB auf das Wachstum von Fritschiella tuberosa wah rend der logarithmischen Wachstumsphase. Hohere Kinetinkonzentrationen hemmen dagegen das Wachstum (vg!. Tab. 2). Wir konnten weiterhin feststellen, dag in dies em Fall auch der Gehalt an Chloroplastenpigmenten/Trockengewicht herabgesetzt ist (urn etwa 20 0/0). Es ist interessant, daB bei einer anderen hochorganisierten SuBwasseralge, Nitella hookeri (Charophyceae), STARLING et a!. (1974) eine Hemmung des Wachstums durch verschiedene Kinetinkonzentrationen feststellten.
Adenin in ahnlich hoher Konzentration (10-4 M) beeinfluBt dagegen die Wachsturns rate nicht. Der Hemmeffekt des Kinetins ist also nicht unspezifisch.
Erstaunlicherweise scheinen auch sehr niedrige Kinetinkonzentrationen (10-10 g/ml) einen hemmenden EinfluB auf das Wachs tum von Fritschiella zu haben (vg!. Tab. 2). Eine einleuchtende Interpretation dieses Ergebnisses ist uns z. Zt. noch nicht moglich.
Z. P/lanzenphysiol. Bd. 76. S. 120-129. 1975.

128 M. MELKONIAN und A. WEBER
Mit keiner einzigen Konzentration konnten wir ein schnelleres Wachstum von Fritschiella wiihrend der logarithmischen Wachstumsphase erzielen, d. h. Kinetin zeigt tiber weite Konzentrationsbereiche tiberhaupt keinen Effekt, manche Konzentrationen bewirken eine eindeutige Hemmung.
Fritschiella tuberosa verhiilt sich also nicht wie manche hoch differenzierten marinen Algen, sondern eher wie ChIarella ellipsaidea (TAMIYA et aI., 1962) oder Nitella haakeri (STARLING et aI., 1974).
Literatur
AUGIER, H.: Les phytohormones des algues. II. - Etude physiologique Ann. Sci. Naturelles, Bot., 12e S., T. 15, 120-180 (1974).
BENTLEy-MoWAT, J. A., and S. M. REID: Effect of gibberellins, kinetin and other factors on the growth of unicellular marine algae in culture. Bot. Mar. XII, 185-199 (1967).
- - Investigation of the radish leaf bioassay for kinetins and demonstration of kinetin-like substances in algae. Ann. Bot. (N. S.) 32, 23-32 (1968).
BOALCH, G. T.: Studies on Ectocarpus in culture. I. and II. J. Mar. Bio!. Ass. U. K., 41 (2), 279-304 (1961).
DREWS, G.: Mikrobiologisches Praktikum fiir Naturwissenschaftler. Berlin, Springer-Verlag, 1968.
DRING, M. J.: Reproduction. In: W. D. P. STEWART (Ed.) Algal Physiology and Biochemistry. p. 814-838. Blackwell, Oxford, 1974.
ERBEN, K.: Sporulation: In: R. A. LEWIN (Ed.) Physiology and Biochemistry of Algae. p. 701-707. Academic Press, New York and London, 1962.
FRIES, L.: Requirements for organic substances in seaweeds. Bot. Mar. XVI, 19-32 (1973). HAWKINS, E. K.: Induction of cell differentiation from dissociated cells and fragments of
Callithamnion roseum. Am. J. Bot. 53, 611 (1966). HINDAK, F.: Biomass production of filamentous green algae (Ulotrichales) in cultures. Algo!.
Stud. (Trebon) 1,77-109 (1970). HOOGENHOUT, H., and J. AMESZ: Growth rates of photosynthetic microorganisms in labora
tory cultures. Arch. Mikrobio!' 50, 10-24 (1965). HUSSAIN, A., and A. D. BONEY: Isolation of kinin-like substances from Laminaria digitata.
Nature (Lond.) 223, 504-505 (1969). IWASAKI, H.: Nutritional studies of the edible seaweed Porphyra tenera. I. The influence of
different B12 analogues, plant hormones, purines and pyrimidines on the growth of Conchocelis. Plant & Cell Physio!. 6, 325-335 (1965).
IYENGAR, M. O. P.: Fritschiella, a new terrestrial member of the Chaetophoraceae. New Phyto!' 31, 329-335 (1932).
JENNINGS, R. c.: Cytokinins as endogenous growth regulators in the algae Ecklonia (Phaeophyta) and Hypnea (Rhodophyta). Aust. J. Bio!. Sci. 22, 621-627 (1969).
KESSLER, E., und F.-C. CZYGAN: Physiologische und biochemische Beitrage zur Taxonomic der Gattung Chlorella. VI. Verwertung organischer Stickstoffverbindungen. Arch. Mikrobio!. 70,211-216 (1970).
KLEBS, G.: Bedingungen der Fortpflanzung bei einigen Algen und Pilzen. Fischer-Verlag, Jena, 1896.
LIEN, T., R. PETTERSEN, and G. KNUTSEN: Effects of Indole-3-acetic acid and Gibberellin on synchronous cultures of Chlorella fusca. Physio!. Plant. 24, 185-190 (1971).
McBRIDE, G. E.: Cytokinesis and ultrastructure in Fritschiella tuberosa. Arch. Protistenk. 112,365-375 (1970).
z. Pflanzenphysiol. Bd. 76. S. 120-129. 1975.

EinfluB von Kinetin auf Fritschiella 129
MILLER, C. 0.: Kinetin and kinetin-like compounds. In: K. PEACH und M. V. TRACEY (Eds.) Moderne Methoden der Pflanzenanalyse. Vol. 6, p. 194-202. Springer-Verlag, Wien, 1963.
MONOD, ].: The growth of bacterial cultures. Ann. Rev. Microbiol. 3, 371-394 (1949). MYERS, ].: Laboratory cultures. In: R. A. LEWIN (Ed.) Physiology and Biochemistry of
Algae. p. 603-613. Academic Press, New York and London, 1962. OLSZEWSKA, M. ].: L'influence de la kinetine sur l'allengement des cellules et sur l'intensite
de divisions cellulaire chez S pirogyra longata. Bull. Soc. Sci. et Lettr. Lodz, Sci. math. et nat. 9, (1),1-3 (1958).
PEDERSEN, M.: Ectocarpus Jasciculatus: marine brown alga requiring kinetin. Nature (Lond.) 218,776 (1968).
PROSKAUER, ]., and R. BERMAN: Agar culture medium modified to approximate soil conditions. Nature (Lond.) 227, 1161 (1970).
PROVASOLI, L.: Effect of plant hormones on Viva. BioI. Bull. 114, 375-384 (1958). SINGH, R. N.: On some phases in the life history of the terrestrial alga Fritschiella tuberosa
and its autecology. New Phytol. 40,170-182 (1941). - Fritschiella tuberosa IYENG. Ann. Bot. (N. S.) 11, 159-164 (1947). STARLING, M. B., V. ]. CHAPMAN, and]. M. A. BROWN: A contribution to the biology of
Nitella hookeri A. BR. in the Rotorua Lakes, New Zealand. II. Organic nutrients and physical factors. Hydrobiologia 45,157-168 (1974).
TAMIYA, H., Y. MORIMURA, and M. YOKOTA: Effects of various antimetabolites upon the life cycle of Chlorella. Arch. Mikrobiol. 42, 4-16 (1962).
VAN STADEN, ]., and C. M. BREEN: Cytokinins in freshwater algal cultures. Plant Sci. Letters 1, 325-330 (1973).
WEBER, A.: Chlorophylle und Carotinoide der Chaetophorineae (Chlorophyceae, Vlotrichales). 2. Der EinfluB unterschiedlicher Stickstoffkonzentrationen auf die Pigmentgarnitur und die Morphogenese der Grunalge Fritschiella tuberosa IYENGAR. Arch. MicrobioI. 102,45-42 (1975).
ZETSCHE, K.: Der EinfluB von Kinetin und Gibberellin auf die Morphogenese kernhaltiger und kernloser Acetabularien. Planta (Berl.) 59, 624-634 (1963).
M. MELKONIAN und A. WEBER, Institut fur Allgemeine Botanik der Universitat, D-2 Hamburg 36, ] ungi usstr. 6.
Z. PJlanzenphysiol. Bd. 76. S. 120-129. 1975.






















![[AWA 2017] Dr. Steffen de Sombre: Bildungsbürgertum und Massenkultur](https://img.pdfslide.org/doc/110x75/5a65d6ed7f8b9a723f8b477f/awa-2017-dr-steffen-de-sombre-bildungsbuergertum-und-massenkultur.jpg)






