Embed Size (px)
Citation preview

Analyse von Strom-Spannungskurven
und Markov-Schalten des K+-Kanals in Chara in Hinblick
auf die Beteiligung schnellen Schaltens beim anomalen
Molfraktionseffekt in Anwesenheit von Tl+
Diplomarbeit der
Mathematisch-Naturwissenschaftlichen Fakultät der
Christian-Albrechts-Universität zu Kiel
vorgelegt von
Afshin Farokhi
Kiel, im Januar 1998

i
Inhaltsverzeichnis1.Einleitung ............................................................................................................................ 1
2. Biologische Grundlagen und Patch-Clamp-Technik ..................................................... 3
2.1. Biologische Grundlagen ......................................................................................... 32.1.1. Biologische Membran ................................................................................ 32.1.2. Membran-Transport-Klassifizierung ......................................................... 42.1.3. Kanäle und Carrier ..................................................................................... 42.1.4. Chara corallina .......................................................................................... 42.1.5. Aufbau eines Membranproteins ................................................................. 5
2.2. Die Patch-Clamp-Technik ...................................................................................... 62.2.2. Patchkonfigurationen ................................................................................. 6
3. Der anomale Molfraktionseffekt ...................................................................................... 8
3.1. Bisherige Meßergebnisse ........................................................................................ 83.1.1. Ergebnisse von Hagiwara ........................................................................... 83,1.2. Ergebnisse von Draber ............................................................................... 93.1.3. Ergebnisse von Albertsen ......................................................................... 103.1.3. Ergebnisse von Keunecke ........................................................................ 11
4. Modelle für die Beschreibung der Transport-vorgänge in Membrankanälen .......... 14
4.1. Markov-Modelle ................................................................................................... 144.2. Multi-Ion-Single-File-Modell ............................................................................... 154.3. Das enzymkinetische lazy-state-Modell ............................................................... 174.4. Wechselwirkungsmodell von Wu ......................................................................... 18
5. Veränderung des Schaltverhaltens beim Block durch Metallionen ........................... 20
5.1. Veränderung der Leitfähigkeit durch schnelles Schalten ..................................... 205.2. Reduzierte scheinbare Leitfähigkeit bei der Cs+-Blockade des K+-Kanals .......... 225.3. Der Na+-Effekt beim K+-Kanal und der Nachweis des schnellen Schaltens in Chara ................................................ 23
6. Experimenteller Aufbau und Methoden ....................................................................... 27
6.1. Der Aufbau ........................................................................................................... 276.2. Meßkammer .......................................................................................................... 286.3. Bearbeiten und Ziehen der Pipetten ..................................................................... 29

ii
6.4. Erd- und Pipettenelektrode ................................................................................... 296.5. Der Patch-Clamp Verstärker ................................................................................ 306.6. Das Patchen .......................................................................................................... 316.7. Die Kühlung ......................................................................................................... 326.8. Temperaturmessung und Probleme durch die Kühlung ....................................... 326.9. Auswerteprogramm .............................................................................................. 33
6.9.1. Bestimmung von Kanalzahl, Niveaus und Standardabweichung ............ 336.9.2. Die Rekonstruktion der Zeitreihe mit dem Hinkley-Detektor ................ 34
7. Meßergebnisse ................................................................................................................. 37
7.1. Strom-Spannungskurven ...................................................................................... 377.1.1. Bestimmung der Einzelkanalströme ........................................................ 377.1.2. Aufstellung von Strom-Spannungskurven ............................................... 39
7.1.3. Der anomale Molfraktionseffekt im scheinbaren Einzelkanalstrom ................. 397.2. Motivation zur Untersuchung des Zusammenhangs zwischen schnellem Schalten und scheinbarem Einzelkanalstrom ............................ 427.3. Verweildauerhistogramme ................................................................................... 43
7.3.1. Analyseprogramme .................................................................................. 437.3.2. Auswahl eines geeigneten Markov-Modells für den K+-Kanal in Chara 457.3.3. Der Vergleich von C<=>O<=>C- und O<=>O<=>C<=>C<=>C-Modelle ...................................................... 477.3.4. Betrachtung der Fitergebnisse bei den verschiedenen Modellen ............ 507.3.5. Analyse auf der Basis des Modells S<=>O<=>G<=>C<=>Z ................. 51
7.3.5.1. Änderung der Ratenkonstanten bei unterschiedlichen Temperaturen ............................................ 517.3.5.2. Änderung der Ratenkonstanten unter Einfluß von Tl+ ............... 57
7.3.5.2.1. Mittelung im Burstbereich ............................................. 577.3.6. Betrachtung der Besetzungswahrscheinlichkeiten der einzelnen Zustände im blockierten und unblockierten Kanal ..................... 64
7.3.6.1. Definition der Besetzungswahrscheinlichkeit ........................... 647.3.6.2. Besetzungswahrscheinlichkeiten im reduzierten Modell .......... 64
7.3.7. Auswirkungen des schnellen Schaltens auf die Reduzierung des Einzelkanalstroms .......................................... 677.3.8. Die Anwendbarkeit des Mittelungsmodells ........................................... 687.3.9. Untersuchung der Offenwahrscheinlichkeiten ....................................... 717.3.10. Kontrolle der Ratenkonstanten der Zeitreihe ausdem Target-Fit mit der Methode des direkten Fits der Zeitreihemit einem Hidden-Markov-Modell (HMM-Fit) .............................................. 75
8. Fazit und Ausblick ....................................................................................................... 77
9. Zusammmenfassung .................................................................................................... 79
Literaturverzeichnis ......................................................................................................... 81
Danksagung ....................................................................................................................... 86
Erklärung .......................................................................................................................... 87

1 EinleitungDer Transport von Ionen in pflanzlichen und tierischen Zellen ist ein interessanter und
wichtiger Bestandteil des Stoffwechsels. Unterschiedliche physikalische Mechanismen undModelle sind entwickelt worden, um den Transport durch Ionenkanäle zu erklären. ZurÜberprüfung der Modelle können unterschiedliche Effekte, Phänomene und Eigenschaften desKanalverhaltens benutzt werden. Die Leistungsfähigkeit eines Modells zeigt sich darin,inwieweit es die beobachteten Effekte erklären kann.
Der anomale Molfraktionseffekt (AMFE) ist einer dieser Effekte: In einer Mischung vonunterschiedlichen Ionen (z.B. K+ und Tl+) ist die Leitfähigkeit in der Mischlösung kleiner alsin den reinen Lösungen. Der anomale Molfraktionseffekt wurde für K+- und Tl+-Ionen zumersten Mal von Hagiwara et al. 1977 an den Eizellen eines Fisches entdeckt. Dieser Effektkonnte durch das Multi-Ion-Single-File-Pore-Modell (Hille und Schwarz, 1978) quantitativbeschrieben werden. Der Kanal hat in diesem Modell in seinem Inneren mehrereBindungsstellen. Die Ionen können durch Platzwechsel durch den Kanal springen, aber sichnicht aneinander vorbeibewegen. Das Modell sagt bei einem bestimmten Mischungsbereichzweier permeierender Ionensorten ein Minimum der Leitfähigkeit, d.h. den anomalenMolfraktionseffekt, voraus.
Schultze (1990) fand heraus, daß das Hille-Schwarz-Modell den anomalenMolfraktionseffekt nur in einem Spannungsbereich von ca. 30 mV erklären kann. Dies stehtim Gegensatz zu den experimentellen Daten von Draber (1990; Draber et al., 1991). Sie stelltefest, daß der Effekt für alle Spannungen beobachtet werden kann. Als Konkurrenz zum Hille-Schwarz-Modell schlugen Draber et al. (1991) ein enzymkinetisches Modell mit „lazy-state“vor. Damit wurde der AMFE durch ein Gating-Modell erklärt, bei dem schnelles Schalten(Gating, Wechseln von aktiven zu inaktiven Zuständen) den Stromfluß unterbricht und dieMittelung im Filter der Aufzeichnungsanlage zu einem reduzierten scheinbarenEinzelkanalstrom führt.
Draber (1990) konnte mit der damaligen Technologie dieses hypothetische schnelleSchalten bei einer Abtastrate von 5 kHz nicht nachweisen. Albertsen (1992) baute eine sehrviel schnellere Datenerfassungsanlage, die Aufnahmeraten von 200 kHz ermöglichte. Diehiermit aufgenommenen Daten zeigen ein sehr hohes Rauschen. Die Analyse dieser Datenerforderte neue Methoden. Durch die Entwicklung eines neuen Sprungdetektors (Hinkley-Detektor 8.Ordnung, Schultze und Draber, 1993) war man in der Lage, auch bei schlechtemSignal/Rauschverhältnis die rauschfreie Originalzeitreihe des Kanals zu rekonstruieren. Mitdiesen neuen Werkzeugen zeigte Draber (1994, Draber und Hansen 1994), daß der Blockdurch Cs+ auf das Gating und nicht auf den Einzelkanalstrom wirkt.
Stimuliert durch diesen Erfolg griffen Albertsen (1994) und später Keunecke (1995) dieSuche nach dem schnellen Schalten beim AMFE wieder auf. Sie fanden aber nur indirekteHinweise auf das hypothetische schnelle Schalten im AMFE.
Keunecke (1995) versuchte deshalb als Alternative noch einmal eine Erklärung mit einemPermeationsmodell, das direkt auf den Einzelkanalstrom wirkt. Sie wandte das Wu-Modell

Kapitel 1: Einleitung
2
(1991, 1992) auf ihre Daten von Chara an, um den anomalen Molfraktionseffekt zu erklären.Ähnlich wie das Hille-Schwarz-Modell (1978) war die Ion-Ion Ion-Wasser Wechselwirkungvon Wu (1991, 1992) nur in der Lage, für einen begrenzten Spannungsbereich den Effekt zuerzeugen.
Inzwischen entwickelte Blunck (1996) zwei neue Zugänge zur Untersuchung schnellerSchaltvorgänge. Er benutzte die Abkühlung auf 7°C als eine Möglichkeit, das Schalten desdurch Na+ blockierten K+-Kanals zu verlangsamen. Dabei entdeckte er, daß der Unterschiedzwischen den Einzelkanalströmen des blockierten und unblockierten Kanals verschwand undführte das darauf zurück, daß jetzt die zeitliche Auflösung reichen würde. Außerdem arbeiteteer das Analyse-Programm von Kijima und Kijima (1987b) so um, daß mit einem Target-Fitdie Ratenkonstanten des zugehörigen Markov-Modells auch aus Mehrkanalaufzeichnungengewonnen werden konnten. Kirst (1997) schuf dann eine sehr bequem zu handhabendeWindows-Version dieses Verfahrens und erweiterte die Anzahl der zulässigen Markov-Zustände des Einzelkanals auf 5.
Nachdem diese neuen Werkzeuge bereitgestellt worden waren, schien es sinnvoll, das alteProblem des AMFE wieder aufzugreifen. Das Ziel dieser Arbeit ist, durch hohe Abtastraten undTemperaturreduzierungen den Nachweis des schnellen Schaltens beim anomalenMolfraktionseffekt mit Tl+-Ionen zu erbringen. Es soll gezeigt werden, daß der Tl+-Block nureine scheinbare Reduzierung des Stroms des Kaliumkanals verursacht. Es werden dieExperimente, Methoden und die unterschiedlichen Analyseverfahren beschrieben, mit derenHilfe das schnelle Schalten aufgelöst werden soll.
Hinter dieser Arbeit steht auch der Wunsch, die Hypothese zu stützen, daß dieunterschiedlichen Metallionen alle über denselben Mechanismus wirken (an der gleichenBindungsstelle angreifen), aber aufgrund ihrer unterschiedlichen chemischen Natur verschiedeneEffekte verursachen. Für den Cs+- und Na+-Block wurde diskutiert, ob beide Ionen an derBindungsstelle für Ca2+ angreifen. Wenn jetzt für Tl+ auch der Nachweis gelänge, daß derAMFE auf schnellem Schalten beruht, wäre das dritte Ion in diese Hypothese eingereiht.

2 Biologische Grundlagen und Patch-Clamp-Technik
2.1 Biologische Grundlagen
2.1.1 Biologische Membran
Die Zelle ist die kleinste noch selbständig lebensfähige Einheit. Alle Organismenbestehen entweder aus einer einzigen, mehreren oder vielen Zellen. Die eukaryotische Zellebeinhaltet unterschiedliche Zellorganellen. Zu den Zellorganellen gehören der Zellkern, dieChloroplasten, Vakuolen und u.a. auch die Mitochondrien. Sie alle sind in das Cytoplasmaeingebettet. Wichtigstes Strukturelement sind Membranen aus Lipiden, die nicht nurProtoplasten nach außen abgrenzen, sondern auch die Zellorganellen im Protoplastenumschließen. Zellkern, Plastiden und Mitochondrien sind vom Cytoplasma durch eineDoppelmembran abgegrenzt, die aus zwei Elementarmembranen besteht. Damit wird dieRegel erfüllt, daß auf der einen Seite der Membran eine cytoplasmatische, auf der andereneine lumenale (nicht lebende) Phase ist. Bei pflanzlichen Zellen nennt man die äußereZellwand Plasmalemma. Die innere Membran, die die zentrale Vakuole vom Cytoplasmaabgrenzt, Tonoplast. Bei tierischen Zellen fehlt im Gegensatz zu den pflanzlichen Zellen eineZellulose-Zellwand und die Vakuole.Transportstudien finden fast ausschließlich am Plasmalemma und am Tonoplasten statt. Diesebestehen wie alle Membranen aus einer Doppellipidschicht. Zwischen die Lipidmolekülensind verschiedene Transportmoleküle eingelagert, mit deren Hilfe der Stoffaustausch über dieMembran stattfindet (Fig. 2.1).
Fig. 2.1: Schema einer biologischen Membran.

Kapitel 2: Biologische Grundlagen und Patch-Clamp-Technik
4
2.1.2 Membran-Transport-Klassifizierung
Ionentransportprozesse können in verschiedene Gruppen eingeteilt werden, die sich durch dieEnergiequellen für den Transport unterscheiden.
a) Passiver Transporter: Er ermöglicht den Transport von Ionen entlang eines Gradienten.Die Triebkraft ist die Potentialdifferenz plus der Nernstspannung des Ions.
b) Pumpen: Sie können unter Aufwendung chemisch gespeicherter Energie Ionen gegenKonzentrations- oder Potentialgradienten transportieren. Bei der Hydrolyse von ATP in ADPwird eine Energie pro Molekül von ca. 450 meV frei, die für den Transport durch dieMembran benutzt wird (Hansen et al., 1981; Bowman und Bowman, 1986; Serranno, 1988).
c) Cotransporter: Sie transportieren eine Ionenart passiv durch die Membran, und diegewonnene Energie wird für den Transport anderer Ionen verwendet. Der Transport beiderIonenarten in die gleiche Richtung wird als Symport bezeichnet, in entgegengesetzterRichtung als Antiport (Sanders et al., 1984). Der Gradient für das antreibende Ion wirdmeistens durch Pumpen erzeugt.
2.1.3 Kanäle und Carrier
Neben der Klassifizierung nach der Energieversorgung gibt es eine nach demMechanismus. Hier gibt es zwei Modelle für die Beschreibung der Transportmoleküle: Kanalund Carrier.
Kanäle können als wassergefüllte Poren betrachtet werden, die die Lipiddoppelschichtüberbrücken (Fig. 2.1). Ihre Transportrate beträgt etwa 107 (Moleküle oder Ionen) proSekunde (Läuger, 1980). Der Kanal kann sich öffnen und schließen, was mit Hilfe der Patch-Clamp-Technik beobachtet werden kann (Abschnitt 2.2).
Carrier sind lipidlösliche Moleküle, die sich innerhalb der Membran frei bewegen und dieIonen im Huckepackprinzip von einer Seite der Membran auf die andere schleusen. Sieerreichen Transportraten von 102 bis 104 Molekülen oder Ionen pro Sekunde (Läuger, 1980).
Carrier sind auf molekularer Ebene noch nicht als intrinsische Membranproteinegefunden worden. Deshalb werden viele Transporter, die früher als Carrier bezeichnetwurden, heutzutage als Kanäle betrachtet. Carrier sind bisher nur bei biologischenKampfstoffen wie Valinomycin, die Pilze gegen Bakterien verwenden, nachgewiesen worden(Mueller und Rudin, 1967).
2.1.4 Chara corallina
Chara corallina ist eine Süßwasseralge aus derFamilie der Characeen. Sie besteht aus Axialzellenund Internodialzellen (Fig. 2.2). DieInternodialzellen, die eine Länge bis zu 10 cm undeinen Durchmesser bis zu 1.2 mm haben, sind fürdie elektrophysiologische Untersuchung vonBedeutung. Die Internodialzelle bestehthauptsächtlich aus der Vakuole, die vomTonoplasten begrenzt ist.
Aus Chara lassen sich sehr leichtProtoplasmatropfen isolieren. Sie bilden sich am Fig.2.2: Chara corallina.

Kapitel 2: Biologische Grundlagen und Patch-Clamp-Technik
5
Boden einer Petrischale, wenn man eine Charazelle mit der Schere aufschneidet und dasCytoplasma ausströmen läßt. Die sich hierbei bildenden Tropfen sind von einerTonoplastenmembran umgeben (Lühring, 1986).
2.1.5 Aufbau eines Membranproteins
Membrankanäle bestehen aus vier bis sechs Makromolekülen. Sie bilden die Pore in derMembran und lagern sich so zusammen, daß in der Mitte der Durchgang entsteht (Fig. 2.3b).Bei spannungsgesteuerten Kaliumkanälen vom „Shaker-Typ“ sind es vier (Mac Kinnon,1991). Die Makromoleküle bestehen aus sechs Segmenten (α Helices S1-S6), die dieMembran durchsetzen. Die Segmente werden durch lineare Aminosäuresequenzenmiteinander verbunden (Fig. 2.4a). Am bekanntesten sind zwei Funktionseinheiten, derSelektivitätsfilter und der Spannungssensor.
Für die Selektivität des Kanals sind die entscheidenen Segmente die zwei β-sheet (p-loops, pore-loops), die die S5- und S6-Segmente brückenartig miteinander verbinden (Miller,1991; Catterall, 1992) und in die Pore hineinragen. Die vier P-Loops der Makromolekülebilden in der Pore einen Ring mit ihren Bindestellen (Fig.2.3b). Die Unterscheidung zwischenpermeierenden und nicht permeierenden Ionen wird durch diese Bindungsstellen getroffen.Sie können sehr deutlich unterscheiden zwischen ähnlichen Ionen wie Na+, K+, Tl+ und Cs+.Damit funktioniert der P-Loop als Selektivitätsfilter (Miller, 1991; Jan und Jan, 1992).
Die S4-Helix trägt positive Ladungen und ist für die Spannungsabhängigkeit desTransportes zuständig. Die S4-Helix kann sich in der Membran im elektrischen Feldverschieben. Dadurch wird der Kanal geöffnet oder geschlossen (Miller, 1991; Jan und Jan;1992).

Kapitel 2: Biologische Grundlagen und Patch-Clamp-Technik
6
Fig. 2.3a: Prinzipieller Aufbau einesKanalproteins (nach Pongs, 1992).b: Bildung der Pore aus S5- und S6-Segmenten. Im Zentrum der Pore liegendie P-Loops, die die selektive Öffnungbilden.
2.2 Die Patch-Clamp-Technik
Die Technik der Patch-Clamp-Messungen wurde von Sakman und Neher entwickelt(Neher und Sakman, 1976). Dabei wird der Strom durch eine sehr kleine, wenige µm2 großeFläche (patch) der Membran mit nur wenigen Kanälen gemessen, an die eine vorgegebeneSpannung (clamp) angelegt wird. Der gemessene Strom liegt in der Größenordnung einigerPicoampere (pA). Um einen sehr kleinen Abschnitt der Zellmembran (patch) elektrisch vonseiner Umgebung zu isolieren, muß man eine sehr dichte Verbindung zwischen derZellmembran und dem Rand einer Glaspipette schaffen. Die Pipette als Meßelektrode wirdauf die Zelloberfläche aufgesetzt und ein leichter Unterdruck erzeugt, so daß die Membranetwas in die Pipette hineingezogen wird. Der Abstand zwischen den Lipiden der Membranund dem Glas ist weniger als 1 nm (Corey und Stevens, 1983). Die Wasserstoffbrücken,Salzbrücken, Ca+-Brücken und Van der Waals-Kräfte sind verantwortlich für die Dichte desSeals (Corey und Stevens, 1983; Ophasi und Webb, 1994). Liegt der Kriechwiderstandzwischen Glas und Membran im Bereich einiger Gigaohm (um die 10 GΩ), wird dieseVersiegelung als Gigaseal bezeichnet. Nachdem sich ein solches Gigaseal gebildet hat, kannman für verschiedene Spannungen über der Membran den Strom mit einem Patch-Clamp-Verstärker messen (siehe Abschnitt 6.5).
2.2.1 Patchkonfigurationen
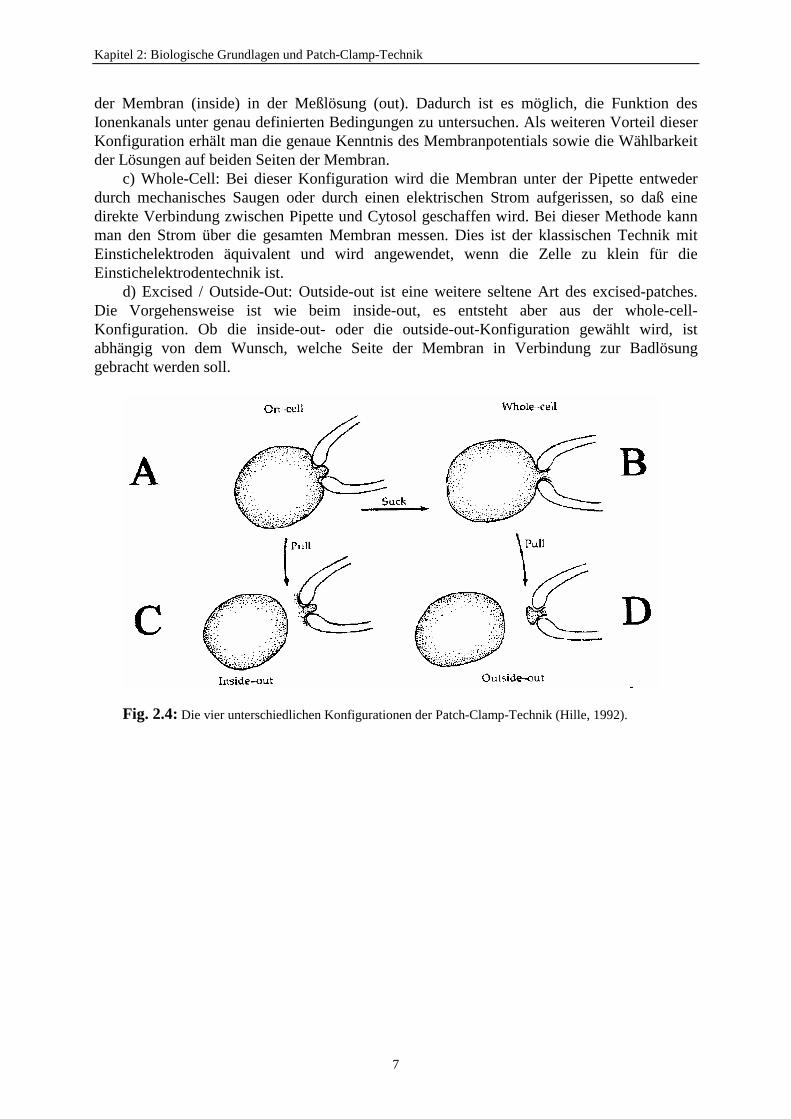
Es gibt vier wichtige Konfigurationen der Patch-Clamp-Technik (Fig. 2.4):a) Cell-Attached: Man setzt die Pipette auf die Membran, ohne daß die Verbindung zur
Gesamtzelle abreißt. Der große Vorteil ist, daß während der Messung die Zelle intakt bleibt,so daß die Funktion des Ionenkanals unter physiologischen Bedingungen untersucht werdenkann. Der Nachteil dieser Konfiguration ist, daß die Membranspannung am Patch unbekanntist, da die Spannung zwischen Zellinnerem und Badlösung in Serie im Meßpfad liegt. DieseTechnik gibt auch keine genaueren Informationen über die Ionenzusammensetzung im Innerender intakten Zelle.
b) Excised / Inside-Out: Durch das ruckartige Wegbewegen der Pipette kann, ohne daßhierdurch die hohe Abdichtung des Patches verloren geht, der Patch aus der Zelloberflächeherausgerissen werden. Bei dieser Konfiguration liegt der Patch mit der cytosolischen Seite
Kapitel 2: Biologische Grundlagen und Patch-Clamp-Technik
7
der Membran (inside) in der Meßlösung (out). Dadurch ist es möglich, die Funktion desIonenkanals unter genau definierten Bedingungen zu untersuchen. Als weiteren Vorteil dieserKonfiguration erhält man die genaue Kenntnis des Membranpotentials sowie die Wählbarkeitder Lösungen auf beiden Seiten der Membran.
c) Whole-Cell: Bei dieser Konfiguration wird die Membran unter der Pipette entwederdurch mechanisches Saugen oder durch einen elektrischen Strom aufgerissen, so daß einedirekte Verbindung zwischen Pipette und Cytosol geschaffen wird. Bei dieser Methode kannman den Strom über die gesamten Membran messen. Dies ist der klassischen Technik mitEinstichelektroden äquivalent und wird angewendet, wenn die Zelle zu klein für dieEinstichelektrodentechnik ist.
d) Excised / Outside-Out: Outside-out ist eine weitere seltene Art des excised-patches.Die Vorgehensweise ist wie beim inside-out, es entsteht aber aus der whole-cell-Konfiguration. Ob die inside-out- oder die outside-out-Konfiguration gewählt wird, istabhängig von dem Wunsch, welche Seite der Membran in Verbindung zur Badlösunggebracht werden soll.
Fig. 2.4: Die vier unterschiedlichen Konfigurationen der Patch-Clamp-Technik (Hille, 1992).

3 Der anomale MolfraktionseffektUnter dem anomalen Molfraktionseffekt (AMFE) versteht man folgendes Phänomen: Die
Einzelkanalleitfähigkeit durchläuft als Funktion des Verhältnisses der Konzentration von zweiIonenarten ein Extremum, wobei die Gesamtkonzentration konstant gehalten wird (Hille,1992).
3.1 Bisherige Meßergebnisse
Der anomale Molfraktionseffekt wurde sowohl bei tierischen als auch bei pflanzlichenMembranen beobachtet. Im folgenden werden die Ergebnisse des AMFEs in erster Linie beipflanzlichen Zellen und bei einer Mischung von K+ und Tl+ vorgestellt.
3.1.1 Ergebnisse von Hagiwara
Hagiwara et al. (1977) beobachteten den AMFE an Eizellen des Fisches Mediasteraaequalis bei externem Lösungswechsel. Die Lösung bestand aus einer Mischung von K+ undTl+ mit einer Gesamtkonzentration von 25 mM. Das Minimum der Abhängigkeit derLeitfähigkeit von der Konzentration, die sich bei verschiedenen Spannungen verschiebt, istbei der reinen K+-Konzentration dichter als bei der Tl+-Konzentration (Fig. 3.1).
Fig. 3.1: Membranleitfähigkeit an Eizellen des Fisches Mediastera aequalis in einer Mischlösung vonK+- und Tl+-Ionen. Der Tl+-Anteil im Verhältnis zur Gesamtkonzentration variiert zwischen 0 und 1. DieLeitfähigkeit geht durch das Minimum (Hagiwara et al., 1977).
Kapitel 3: Der anomale Molfraktionseffekt
9
Hagiwara erklärte den AMFE mit Hilfe eines Modells mit zwei Bindestellen (Hille undSchwarz, 1978). Somit wurde der AMFE als Beweis für das Multi-Ion-Porenmodell gesehen.Die verschiedenen Modelle für die Ionenkanäle werden in Kapitel 4 erklärt.
3.1.2 Ergebnisse von Draber
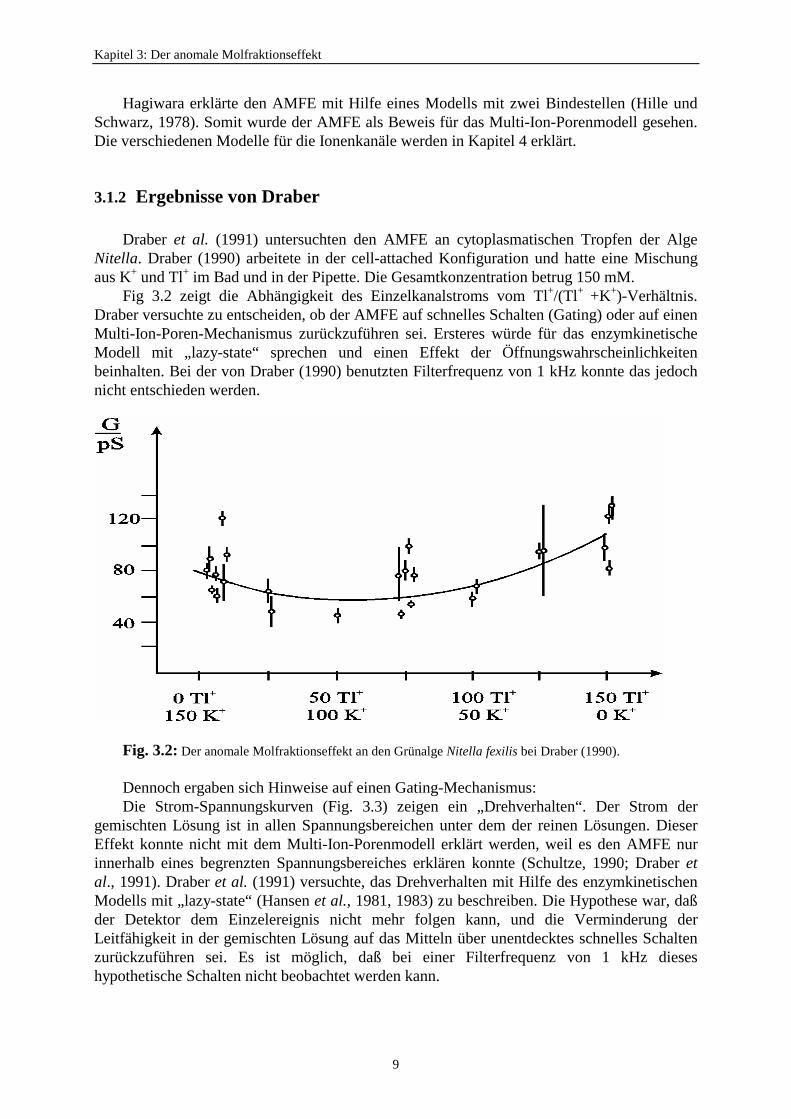
Draber et al. (1991) untersuchten den AMFE an cytoplasmatischen Tropfen der AlgeNitella. Draber (1990) arbeitete in der cell-attached Konfiguration und hatte eine Mischungaus K+ und Tl+ im Bad und in der Pipette. Die Gesamtkonzentration betrug 150 mM.
Fig 3.2 zeigt die Abhängigkeit des Einzelkanalstroms vom Tl+/(Tl+ +K+)-Verhältnis.Draber versuchte zu entscheiden, ob der AMFE auf schnelles Schalten (Gating) oder auf einenMulti-Ion-Poren-Mechanismus zurückzuführen sei. Ersteres würde für das enzymkinetischeModell mit „lazy-state“ sprechen und einen Effekt der Öffnungswahrscheinlichkeitenbeinhalten. Bei der von Draber (1990) benutzten Filterfrequenz von 1 kHz konnte das jedochnicht entschieden werden.
Fig. 3.2: Der anomale Molfraktionseffekt an den Grünalge Nitella fexilis bei Draber (1990).
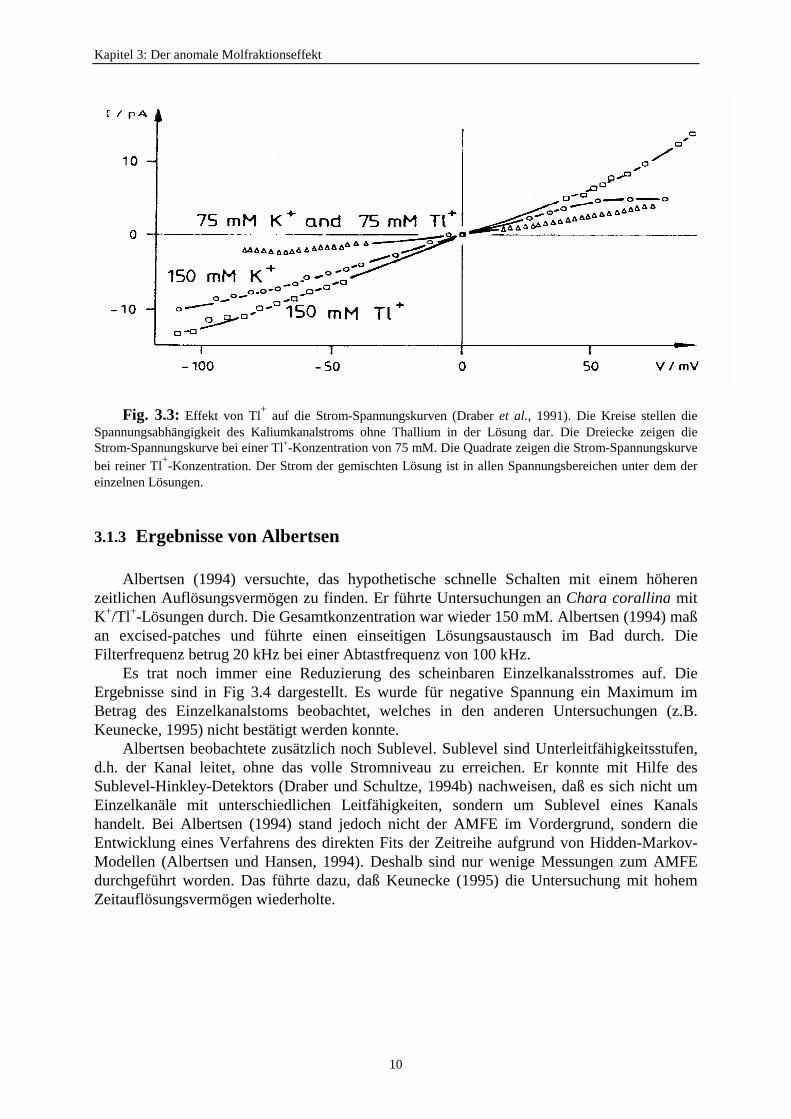
Dennoch ergaben sich Hinweise auf einen Gating-Mechanismus:Die Strom-Spannungskurven (Fig. 3.3) zeigen ein „Drehverhalten“. Der Strom der
gemischten Lösung ist in allen Spannungsbereichen unter dem der reinen Lösungen. DieserEffekt konnte nicht mit dem Multi-Ion-Porenmodell erklärt werden, weil es den AMFE nurinnerhalb eines begrenzten Spannungsbereiches erklären konnte (Schultze, 1990; Draber etal., 1991). Draber et al. (1991) versuchte, das Drehverhalten mit Hilfe des enzymkinetischenModells mit „lazy-state“ (Hansen et al., 1981, 1983) zu beschreiben. Die Hypothese war, daßder Detektor dem Einzelereignis nicht mehr folgen kann, und die Verminderung derLeitfähigkeit in der gemischten Lösung auf das Mitteln über unentdecktes schnelles Schaltenzurückzuführen sei. Es ist möglich, daß bei einer Filterfrequenz von 1 kHz dieseshypothetische Schalten nicht beobachtet werden kann.
Kapitel 3: Der anomale Molfraktionseffekt
10
Fig. 3.3: Effekt von Tl+ auf die Strom-Spannungskurven (Draber et al., 1991). Die Kreise stellen dieSpannungsabhängigkeit des Kaliumkanalstroms ohne Thallium in der Lösung dar. Die Dreiecke zeigen dieStrom-Spannungskurve bei einer Tl+-Konzentration von 75 mM. Die Quadrate zeigen die Strom-Spannungskurvebei reiner Tl+-Konzentration. Der Strom der gemischten Lösung ist in allen Spannungsbereichen unter dem dereinzelnen Lösungen.
3.1.3 Ergebnisse von Albertsen
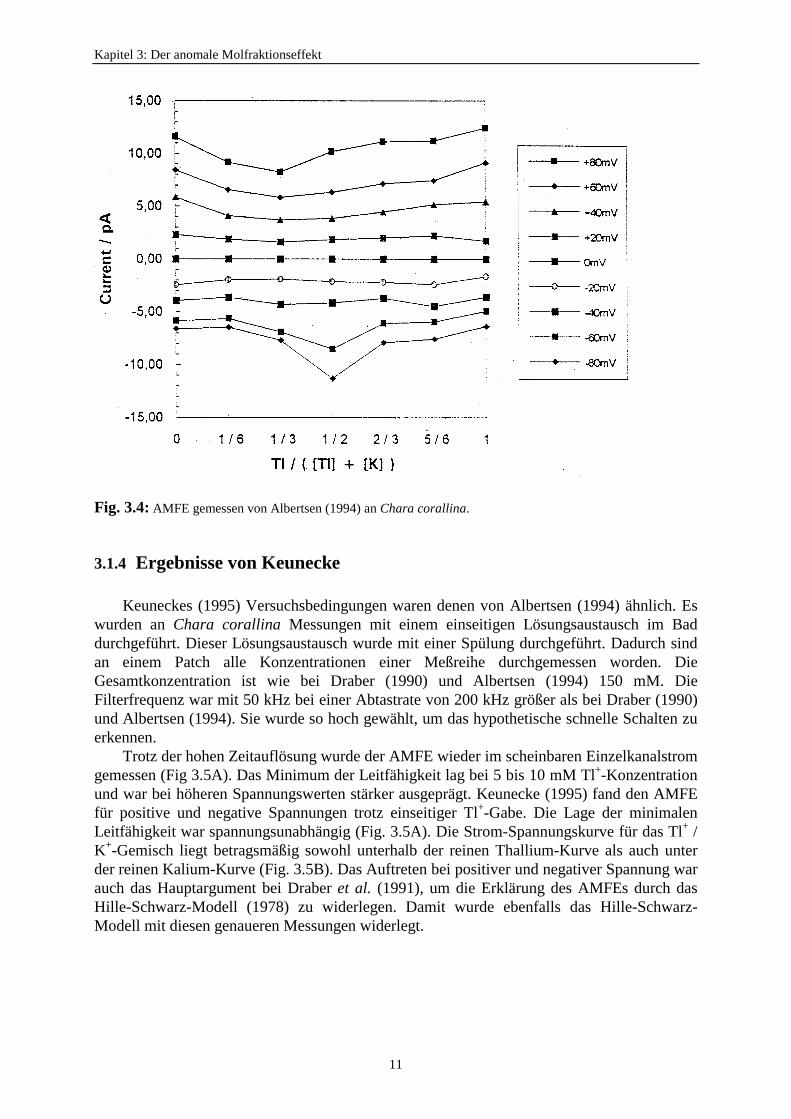
Albertsen (1994) versuchte, das hypothetische schnelle Schalten mit einem höherenzeitlichen Auflösungsvermögen zu finden. Er führte Untersuchungen an Chara corallina mitK+/Tl+-Lösungen durch. Die Gesamtkonzentration war wieder 150 mM. Albertsen (1994) maßan excised-patches und führte einen einseitigen Lösungsaustausch im Bad durch. DieFilterfrequenz betrug 20 kHz bei einer Abtastfrequenz von 100 kHz.
Es trat noch immer eine Reduzierung des scheinbaren Einzelkanalsstromes auf. DieErgebnisse sind in Fig 3.4 dargestellt. Es wurde für negative Spannung ein Maximum imBetrag des Einzelkanalstoms beobachtet, welches in den anderen Untersuchungen (z.B.Keunecke, 1995) nicht bestätigt werden konnte.
Albertsen beobachtete zusätzlich noch Sublevel. Sublevel sind Unterleitfähigkeitsstufen,d.h. der Kanal leitet, ohne das volle Stromniveau zu erreichen. Er konnte mit Hilfe desSublevel-Hinkley-Detektors (Draber und Schultze, 1994b) nachweisen, daß es sich nicht umEinzelkanäle mit unterschiedlichen Leitfähigkeiten, sondern um Sublevel eines Kanalshandelt. Bei Albertsen (1994) stand jedoch nicht der AMFE im Vordergrund, sondern dieEntwicklung eines Verfahrens des direkten Fits der Zeitreihe aufgrund von Hidden-Markov-Modellen (Albertsen und Hansen, 1994). Deshalb sind nur wenige Messungen zum AMFEdurchgeführt worden. Das führte dazu, daß Keunecke (1995) die Untersuchung mit hohemZeitauflösungsvermögen wiederholte.
Kapitel 3: Der anomale Molfraktionseffekt
11
Fig. 3.4: AMFE gemessen von Albertsen (1994) an Chara corallina.
3.1.4 Ergebnisse von Keunecke
Keuneckes (1995) Versuchsbedingungen waren denen von Albertsen (1994) ähnlich. Eswurden an Chara corallina Messungen mit einem einseitigen Lösungsaustausch im Baddurchgeführt. Dieser Lösungsaustausch wurde mit einer Spülung durchgeführt. Dadurch sindan einem Patch alle Konzentrationen einer Meßreihe durchgemessen worden. DieGesamtkonzentration ist wie bei Draber (1990) und Albertsen (1994) 150 mM. DieFilterfrequenz war mit 50 kHz bei einer Abtastrate von 200 kHz größer als bei Draber (1990)und Albertsen (1994). Sie wurde so hoch gewählt, um das hypothetische schnelle Schalten zuerkennen.
Trotz der hohen Zeitauflösung wurde der AMFE wieder im scheinbaren Einzelkanalstromgemessen (Fig 3.5A). Das Minimum der Leitfähigkeit lag bei 5 bis 10 mM Tl+-Konzentrationund war bei höheren Spannungswerten stärker ausgeprägt. Keunecke (1995) fand den AMFEfür positive und negative Spannungen trotz einseitiger Tl+-Gabe. Die Lage der minimalenLeitfähigkeit war spannungsunabhängig (Fig. 3.5A). Die Strom-Spannungskurve für das Tl+ /K+-Gemisch liegt betragsmäßig sowohl unterhalb der reinen Thallium-Kurve als auch unterder reinen Kalium-Kurve (Fig. 3.5B). Das Auftreten bei positiver und negativer Spannung warauch das Hauptargument bei Draber et al. (1991), um die Erklärung des AMFEs durch dasHille-Schwarz-Modell (1978) zu widerlegen. Damit wurde ebenfalls das Hille-Schwarz-Modell mit diesen genaueren Messungen widerlegt.
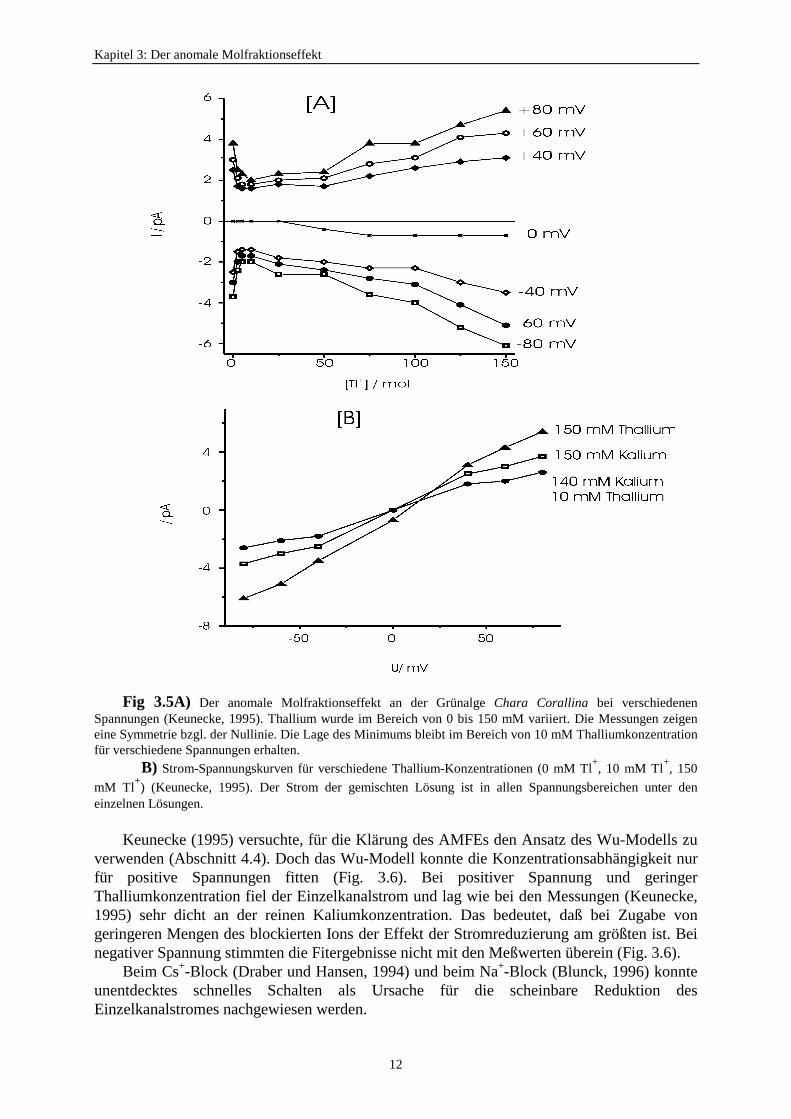
Kapitel 3: Der anomale Molfraktionseffekt
12
Fig 3.5A) Der anomale Molfraktionseffekt an der Grünalge Chara Corallina bei verschiedenenSpannungen (Keunecke, 1995). Thallium wurde im Bereich von 0 bis 150 mM variiert. Die Messungen zeigeneine Symmetrie bzgl. der Nullinie. Die Lage des Minimums bleibt im Bereich von 10 mM Thalliumkonzentrationfür verschiedene Spannungen erhalten.
B) Strom-Spannungskurven für verschiedene Thallium-Konzentrationen (0 mM Tl+, 10 mM Tl+, 150mM Tl+) (Keunecke, 1995). Der Strom der gemischten Lösung ist in allen Spannungsbereichen unter deneinzelnen Lösungen.
Keunecke (1995) versuchte, für die Klärung des AMFEs den Ansatz des Wu-Modells zuverwenden (Abschnitt 4.4). Doch das Wu-Modell konnte die Konzentrationsabhängigkeit nurfür positive Spannungen fitten (Fig. 3.6). Bei positiver Spannung und geringerThalliumkonzentration fiel der Einzelkanalstrom und lag wie bei den Messungen (Keunecke,1995) sehr dicht an der reinen Kaliumkonzentration. Das bedeutet, daß bei Zugabe vongeringeren Mengen des blockierten Ions der Effekt der Stromreduzierung am größten ist. Beinegativer Spannung stimmten die Fitergebnisse nicht mit den Meßwerten überein (Fig. 3.6).
Beim Cs+-Block (Draber und Hansen, 1994) und beim Na+-Block (Blunck, 1996) konnteunentdecktes schnelles Schalten als Ursache für die scheinbare Reduktion desEinzelkanalstromes nachgewiesen werden.
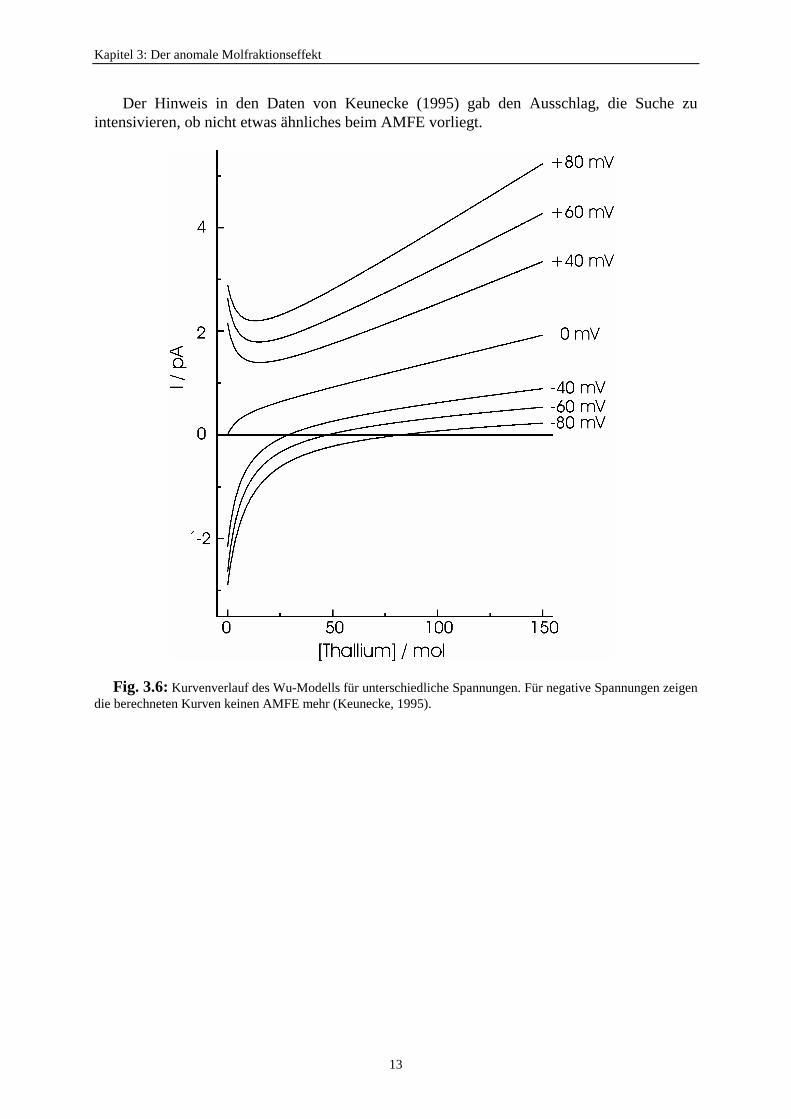
Kapitel 3: Der anomale Molfraktionseffekt
13
Der Hinweis in den Daten von Keunecke (1995) gab den Ausschlag, die Suche zuintensivieren, ob nicht etwas ähnliches beim AMFE vorliegt.
Fig. 3.6: Kurvenverlauf des Wu-Modells für unterschiedliche Spannungen. Für negative Spannungen zeigendie berechneten Kurven keinen AMFE mehr (Keunecke, 1995).

4 Modelle für die Beschreibung der Transport-vorgänge in Membrankanälen
Für die Beschreibung der Funktionsweise von Kanälen werden mathematische undphysikalische Modelle angewendet, die den Mechanismus des Transportes der Ionen durchden Kanal erklären. Eine grundsätzliche Unterscheidung ist die in Gating- und Permeations-modelle (Hansen et al., 1997). Permeationsmodelle beschreiben den Transport eines einzelnenIons durch den Kanal. Gating-Modelle beschäftigen sich mit der Unterbrechung des Stromspermeierender Ionen durch Übergänge zwischen aktiven und inaktiven Zuständen des Kanals.Hier gilt das besondere Interesse den Modellen, die die mechanische Erklärung des AMFEszum Ziel haben.
4.1 Markov-Modelle
Markov-Modelle sind formale Gating-Modelle, die das Schaltverhalten der Kanäle durchÜbergänge zwischen diskreten offenen und geschlossenen Zuständen beschreiben. Ziel derAnalyse ist die Ermittlung der Anzahl von „open“ und „closed“ Zuständen, ihre Anordnungzueinander und der Ratenkonstanten kij (Übergangswahrscheinlichkeit von Zustand i inZustand j).
Für eine stochastische Betrachtung der Schaltvorgänge der Kanäle auf der Grundlageeines Markov-Modells gibt es zwei einfache Voraussetzungen. Eine der Voraussetzungen ist,daß das System kein Gedächtnis hat. Das bedeutet, daß ein Übergang von einem Zustand ineinen anderen nicht von vorherigen Übergängen abhängt. Die andere Voraussetzung ist, daßdie kij nicht von der Besetzung der Zustände abhängig ist (Linearität des Systems, Colquhounund Hawkes, 1977).
Das Verhalten dieses Modells wird durch ein System von Differentialgleichungen ersterOrdnung beschrieben, das durch eine Exponentialfunktion gelöst wird.
KtPdt
tdP ⋅= )()((4.1)
KtetP =)( (4.2)
Die Elemente des Vektors P(t) sind die Besetzungswahrscheinlichkeiten jedes einzelnenZustandes, und die Matrix K bildet sich aus den Ratenkonstanten kij.
Für die Beschreibung der Aufenthaltszeiten in einem Zustand des Markov-Modells wirdmeistens eine Summe von Exponentialfunktionen benötigt (Colquhoun und Hawkes, 1977).Wenn mehrere Zeitkonstanten vorkommen, existieren mehrere Zustände, die voneinanderunabhängig sind.
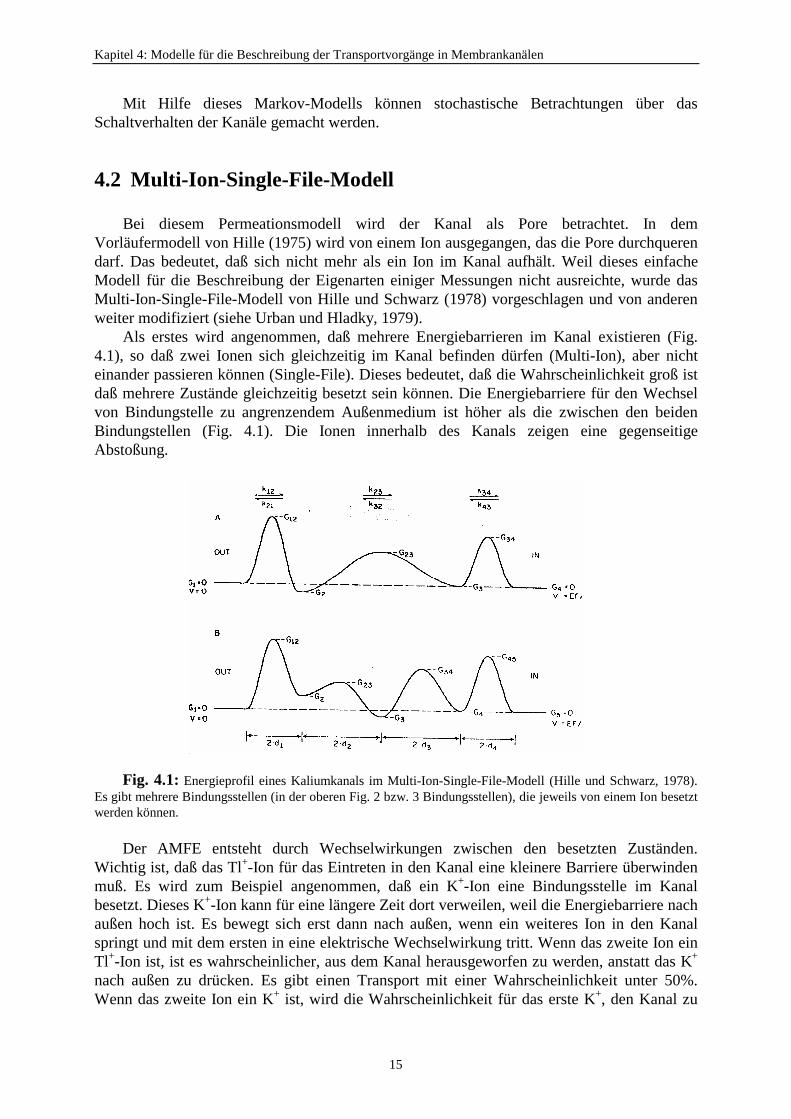
Kapitel 4: Modelle für die Beschreibung der Transportvorgänge in Membrankanälen
15
Mit Hilfe dieses Markov-Modells können stochastische Betrachtungen über dasSchaltverhalten der Kanäle gemacht werden.
4.2 Multi-Ion-Single-File-Modell
Bei diesem Permeationsmodell wird der Kanal als Pore betrachtet. In demVorläufermodell von Hille (1975) wird von einem Ion ausgegangen, das die Pore durchquerendarf. Das bedeutet, daß sich nicht mehr als ein Ion im Kanal aufhält. Weil dieses einfacheModell für die Beschreibung der Eigenarten einiger Messungen nicht ausreichte, wurde dasMulti-Ion-Single-File-Modell von Hille und Schwarz (1978) vorgeschlagen und von anderenweiter modifiziert (siehe Urban und Hladky, 1979).
Als erstes wird angenommen, daß mehrere Energiebarrieren im Kanal existieren (Fig.4.1), so daß zwei Ionen sich gleichzeitig im Kanal befinden dürfen (Multi-Ion), aber nichteinander passieren können (Single-File). Dieses bedeutet, daß die Wahrscheinlichkeit groß istdaß mehrere Zustände gleichzeitig besetzt sein können. Die Energiebarriere für den Wechselvon Bindungstelle zu angrenzendem Außenmedium ist höher als die zwischen den beidenBindungstellen (Fig. 4.1). Die Ionen innerhalb des Kanals zeigen eine gegenseitigeAbstoßung.
Fig. 4.1: Energieprofil eines Kaliumkanals im Multi-Ion-Single-File-Modell (Hille und Schwarz, 1978).Es gibt mehrere Bindungsstellen (in der oberen Fig. 2 bzw. 3 Bindungsstellen), die jeweils von einem Ion besetztwerden können.
Der AMFE entsteht durch Wechselwirkungen zwischen den besetzten Zuständen.Wichtig ist, daß das Tl+-Ion für das Eintreten in den Kanal eine kleinere Barriere überwindenmuß. Es wird zum Beispiel angenommen, daß ein K+-Ion eine Bindungsstelle im Kanalbesetzt. Dieses K+-Ion kann für eine längere Zeit dort verweilen, weil die Energiebarriere nachaußen hoch ist. Es bewegt sich erst dann nach außen, wenn ein weiteres Ion in den Kanalspringt und mit dem ersten in eine elektrische Wechselwirkung tritt. Wenn das zweite Ion einTl+-Ion ist, ist es wahrscheinlicher, aus dem Kanal herausgeworfen zu werden, anstatt das K+
nach außen zu drücken. Es gibt einen Transport mit einer Wahrscheinlichkeit unter 50%.Wenn das zweite Ion ein K+ ist, wird die Wahrscheinlichkeit für das erste K+, den Kanal zu
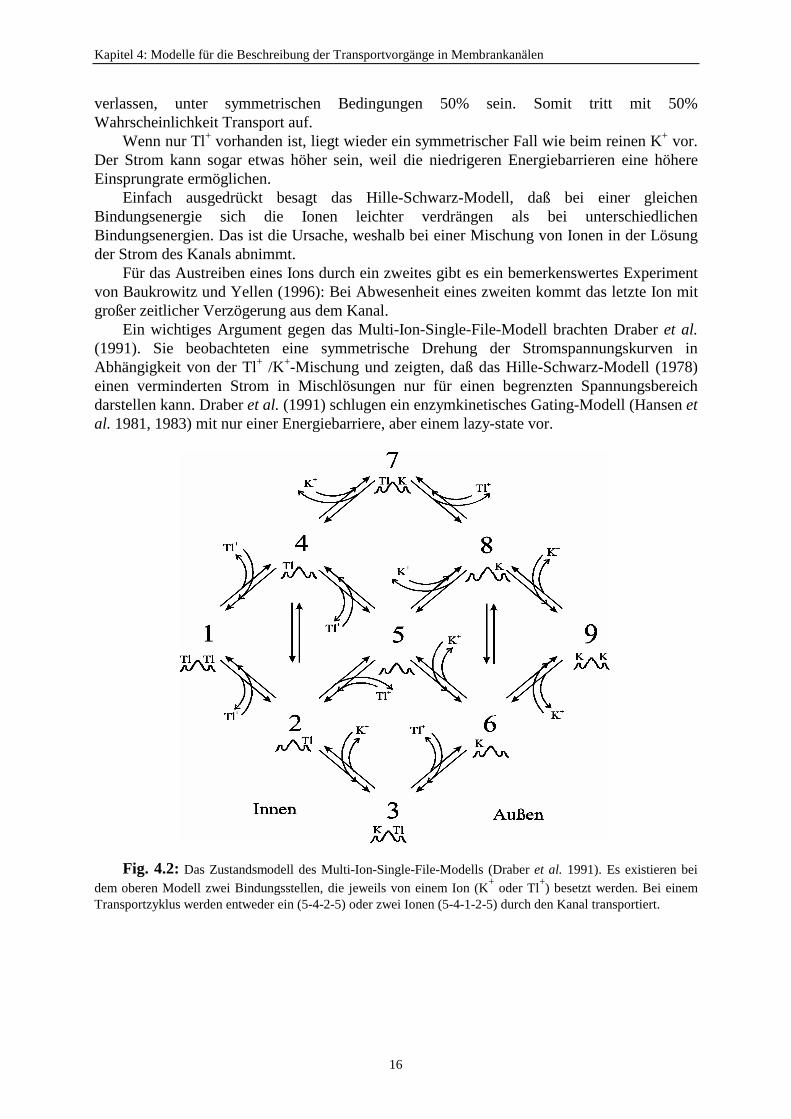
Kapitel 4: Modelle für die Beschreibung der Transportvorgänge in Membrankanälen
16
verlassen, unter symmetrischen Bedingungen 50% sein. Somit tritt mit 50%Wahrscheinlichkeit Transport auf.
Wenn nur Tl+ vorhanden ist, liegt wieder ein symmetrischer Fall wie beim reinen K+ vor.Der Strom kann sogar etwas höher sein, weil die niedrigeren Energiebarrieren eine höhereEinsprungrate ermöglichen.
Einfach ausgedrückt besagt das Hille-Schwarz-Modell, daß bei einer gleichenBindungsenergie sich die Ionen leichter verdrängen als bei unterschiedlichenBindungsenergien. Das ist die Ursache, weshalb bei einer Mischung von Ionen in der Lösungder Strom des Kanals abnimmt.
Für das Austreiben eines Ions durch ein zweites gibt es ein bemerkenswertes Experimentvon Baukrowitz und Yellen (1996): Bei Abwesenheit eines zweiten kommt das letzte Ion mitgroßer zeitlicher Verzögerung aus dem Kanal.
Ein wichtiges Argument gegen das Multi-Ion-Single-File-Modell brachten Draber et al.(1991). Sie beobachteten eine symmetrische Drehung der Stromspannungskurven inAbhängigkeit von der Tl+ /K+-Mischung und zeigten, daß das Hille-Schwarz-Modell (1978)einen verminderten Strom in Mischlösungen nur für einen begrenzten Spannungsbereichdarstellen kann. Draber et al. (1991) schlugen ein enzymkinetisches Gating-Modell (Hansen etal. 1981, 1983) mit nur einer Energiebarriere, aber einem lazy-state vor.
Fig. 4.2: Das Zustandsmodell des Multi-Ion-Single-File-Modells (Draber et al. 1991). Es existieren beidem oberen Modell zwei Bindungsstellen, die jeweils von einem Ion (K+ oder Tl+) besetzt werden. Bei einemTransportzyklus werden entweder ein (5-4-2-5) oder zwei Ionen (5-4-1-2-5) durch den Kanal transportiert.

Kapitel 4: Modelle für die Beschreibung der Transportvorgänge in Membrankanälen
17
4.3 Das enzymkinetische lazy-state-Modell
In diesem Gating-Modell wird der Kanal als ein großes Molekül, das wie ein Enzym mitden Ionen reagiert, betrachtet (Läuger, 1980, 1985, Hansen et al., 1981, Eisenberg 1990).Gating, also Ein- und Ausschalten des Transportes, wird durch einen sogenannten lazy-state(Hansen et al., 1983) eingeführt.
Der lazy-state, den Draber et al. (1991) für den K+-Kanal in Nitella flexilis eingeführthaben, wird durch eine weitere Bindungsstelle für das Kaliumion kontrolliert (rechte Delle amKanalsymbol, z.B. beim Zustand "4" in Fig. 4.3). Wenn das Kaliumion an diese Bindungstellebindet (Übergang 4 - 5 in Fig. 4.3), erhöht sich die Selektivität in K+. K+ wird weiterhintransportiert (Zyklus 3 - 8 - 9 - 3 in Fig. 4.3), doch Tl+ überführt den Kanal in den lazy-state 7.Wenn der Kontroll-Zustand nicht besetzt ist (Zustände 1 bis 6 in Fig. 4.3), können Tl+ und K+
gleichermaßen transportiert werden.Nach diesem Modell wird erwartet, daß die Wahrscheinlichkeit für den Durchgang der
Ionen durch den Kanal abnimmt, wenn die Offenwahrscheinlichkeit des Kanals in einerMischlösung aus K+ und Tl+ im Verhältnis zu einer reinen Lösung aus K+ untersucht wird.
Der Einzelkanalstrom sollte dann unabhängig von der K+ /Tl+-Mischung sein. DieseEigenschaft des Modells wurde jedoch durch die Messungen von Draber et al. (1991) nichtbestätigt. Zunächst scheint, daß deshalb dieses Modell nicht richtig sein kann. Doch es bestehtdie Möglichkeit, daß die Übergänge 3-7 in Fig. 4.3 so schnell sind, daß sie aufgrund einerbeschränkten zeitlichen Auflösung nicht erkannt werden, was auch Inhalt der vorliegendenArbeit ist. Dann wäre die gemessene Reduzierung des Einzelkanalstromes nur einescheinbare, da sie durch die Mittelung im Detektorfilter über einen konstanten, aber schnellschaltenden Einzelkanal entstanden sein könnte. Dann würde das Modell mit den gemessenenDaten übereinstimmen.
Fig 4.3: Das enzymkinetische lazy-state-Modell für den AMFE mit den von Draber et al. (1991) ermitteltenRatenkonstanten für den Transport von K+ oder Tl+. Es gibt eine allosterische Bindestelle für das K+ (Delle amKanalschema von Zustand 4). Ist diese Bindestelle nicht besetzt, kann sowohl K+ als auch Tl+ transportiertwerden (4-1-2 bzw. 4-5-6). Bei einer Besetzung der allosterischen Bindestelle mit K+ und derTransportbindungstelle mit Tl+ geht der Kanal in den Zustand 7 (lazy-state). Tl+ kann nicht transportiert werden.Der K+-Transport geht dagegen im Zyklus 3 - 9 - 8 weiter. Die Ratenkonstanten sind in Einheiten von 107 s-1
gegeben.

Kapitel 4: Modelle für die Beschreibung der Transportvorgänge in Membrankanälen
18
4.4 Wechselwirkungsmodell von Wu
Das Wechselwirkungsmodell von Wu (1991, 1992) ist ein Permeationsmodell und gehtvon einem Selektivitätsfilter aus, das mit verschiedenen Ionen unterschiedlich wechselwirkt.Dabei gelten für das Modell folgende Voraussetzungen:
Der Selektivitätsfilter liegt in der Pore mit nur einer Bindestelle, die aus symmetrischgeordneten Carbonylgruppen (C und O) besteht (Fig. 4.4). Ein Teil der Wassermoleküle wirdvon den Ionen während des Eintritts in den Kanal getrennt. Es kann ein weiteres Ion (B) inden Kanal eintreten, wenn ein Ion (A) an der Bindestelle des Selektivitätsfilters gebunden ist(Fig.4.4). Es treten dann zwischen den Ionen die Coulombwechselwirkungen auf. Das zweiteIon überträgt die kinetische Energie im Stoß auf das gebundene Ion. Nach den Stoßgesetzenerhält man den Energieübertrag zwischen den Ionen:
ABA
BAB E
mmmmcE 02)( +
= (4.3)
mA und mB: Massen von Ion (A) und Ion (B)c = KonstanteEB = Übertragungsenergie zwischen Ion (A) und Ion (B)E0A = Anfangsenergie von Ion (A)
Bei gleicher Masse ist die übertragene Energie am größten, womit auch dieWahrscheinlichkeit, daß EB größer als die Bindungsenergie ist, zunimmt. Das gebundene Ionkann somit aus dem Selektivfilter herausgeschlagen werden. Bei einer großen Abweichungdes Verhältnisses der Massen von eins nimmt die Wahrscheinlichkeit zu, daß dieEnergiebarriere nicht mehr überwunden werden kann und ein Block entsteht. Das bedeutet,daß der Stromfluß sehr stark abhängig von den Ionen ist, die miteinander in Wechselwirkungtreten. Bei unterschiedlichen Ionen nimmt somit die Wahrscheinlichkeit des Stromflusses abim Vergleich zu dem bei gleicher Ionenart.
Fig. 4.4: Wechselwirkungsmodell von Wu (nach Wu, 1991)C: Kohlenstoff der CarbonylgruppeO: Sauerstoff der CarbonylgruppeA: Ion, gebunden an den SelektivfilterB: Ein weiteres Ion gelangt in den Kanal, welches mit Ion A in Wechselwirkung trittW: Wassermoleküle, die mit Ion A in Wechselwirkung treten

Kapitel 4: Modelle für die Beschreibung der Transportvorgänge in Membrankanälen
19
Dieser Wirkungsmechanismus ist dem des Hille-Schwarz-Modells nicht unähnlich. Auchdort ist die Verdrängung durch ein zweites Ion für den Transport notwendig. Hier ist es dieÜbertragung kinetischer Energie, dort die elektrostatische Abstoßung im Kanal mit zweiBindungstellen. Der AMFE entsteht in beiden Fällen aus der größenVerdrängungswahrscheinlichkeit bei gleichen Ionen.Der von Wu (1992) betrachtete Mechanismus ist noch etwas komplizierter als hier dargestellt,weil die Wechselwirkungen zwischen dem Ion und den Wasserdipolen im Kanal sowie demIon mit dem extern angelegten elektrischen Feld berücksichtigt werden.Wie gut dieses Modell ist, wurde in der Arbeit von Keunecke (1995) für den anomalenMolfraktionseffekt untersucht (siehe Abschnitt 3.1.4).

5 Veränderung des Schaltverhaltens beim Blockdurch Metallionen
Bei einer Reduzierung des Gesamtstroms unter physiologischen Ruhebedingungen sprichtman vom Blockieren eines Kanals. Es wird zwischen Blockieren von Kanälen durch Kationen(H+, Alkali und Erdalkali) und durch Proteine (z.B. Kaliotoxin, Margatoxin) (Aiyar et al.,1995) unterschieden. Die Kationen halten sich zumeist in der cytosolischen und vacuolärenLösung auf und stehen mit dem transportierten Ion in Konkurrenz. Die blockierten Proteinebinden sich an ein bestimmtes Kanalprotein und blockieren den Kanal. In dieserArbeitsgruppe sind bislang die Blockierungen durch die Kationen Cs+, Na+ und Tl+ untersuchtworden. Hansen et al. (1997) vermuten, daß die unterschiedlichen Effekte durch dieMetallionen alle auf demselben Mechanismus beruhen. Dies ist die Bindung an die gleicheStelle im Kanal, wobei die beobachteten Phänomene von der Art des blockierenden Ionsabhängen. In allen Fällen wird angenommen, daß dieser Block zu dem im folgendenbeschriebenen schnellen Schalten führt.
5.1 Veränderung der Leitfähigkeit durch schnelles Schalten
Fig. 5.1: Darstellung des Blocks bei unterschiedlichen Blockgeschwindigkeiten (Blunck, 1996).
Permeations- und Gating-Modelle (Hansen et al., 1997) behandeln ganz unterschiedlichePhänomene. Die Entscheidung, welche Art der Modelle angewendet werden muß, kannschwierig werden, wenn in der Untersuchung „schnelles Schalten“ (Flickern) auftritt.Unzureichendes Auflösungsvermögen führt zu Informationsverlust.
In Fig. 5.1 werden Daten mit unterschiedlichen Blockierungsdauern gezeigt. Beimlangsamen Block gibt es keine Änderung des Stromniveaus, sondern nur eine Veränderungder Offenwahrscheinlichkeit im Vergleich zum unblockierten Kanalzustand. Der Verlauf desBlocks (Geschlossenzeiten) ist gut erkennbar.
Kapitel 5: Veränderung des Schaltverhaltens beim Block durch Metallionen
21
Beim mittleren Block kann das Auftreten von Bursts erkannt werden. Diese sind kurzeSchließungen (Gaps) innerhalb einer Offenperiode. Es kann zu einer Reduzierung desgemessenen (scheinbaren) Stromniveaus durch Mittelung im Tiefpaß des Meßsystems führen.
Solange das Auflösungsvermögen des Aufnahmesystems noch ausreicht, hat dasBurstverhalten keine Auswirkungen auf die gemessene Größe der Leitfähigkeit desEinzelkanals. Es verändert sich nur das Schaltverhalten. Bei Messungen am whole-cell-Patchführen die Effekte von Fig. 5.1a und b auf eine flachere Kennlinie und einen geringerenSättigungsstrom. Dies wird durch die Ensemblemittelung über die vielen verschiedenenKanäle verursacht. Je weniger Kanäle pro Zeiteinheit geöffnet sind, desto weniger Ionenkommen durch und der resultierende Gesamtstrom pro Zeiteinheit nimmt ab.
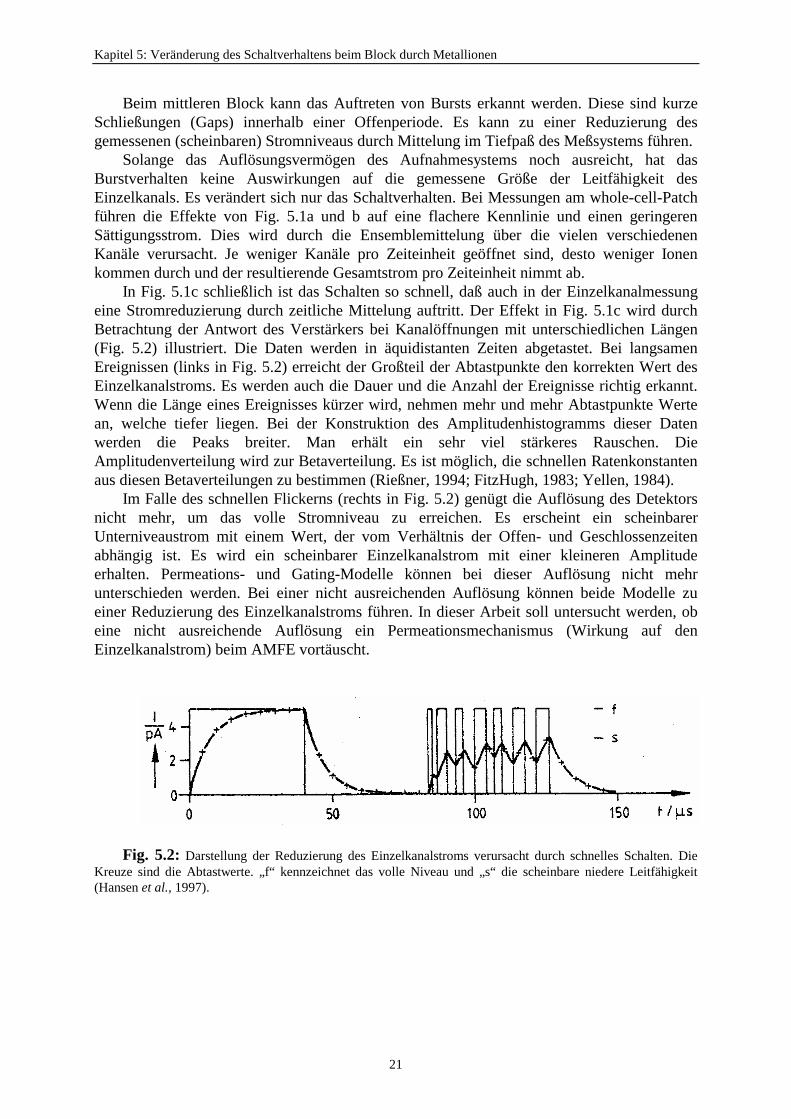
In Fig. 5.1c schließlich ist das Schalten so schnell, daß auch in der Einzelkanalmessungeine Stromreduzierung durch zeitliche Mittelung auftritt. Der Effekt in Fig. 5.1c wird durchBetrachtung der Antwort des Verstärkers bei Kanalöffnungen mit unterschiedlichen Längen(Fig. 5.2) illustriert. Die Daten werden in äquidistanten Zeiten abgetastet. Bei langsamenEreignissen (links in Fig. 5.2) erreicht der Großteil der Abtastpunkte den korrekten Wert desEinzelkanalstroms. Es werden auch die Dauer und die Anzahl der Ereignisse richtig erkannt.Wenn die Länge eines Ereignisses kürzer wird, nehmen mehr und mehr Abtastpunkte Wertean, welche tiefer liegen. Bei der Konstruktion des Amplitudenhistogramms dieser Datenwerden die Peaks breiter. Man erhält ein sehr viel stärkeres Rauschen. DieAmplitudenverteilung wird zur Betaverteilung. Es ist möglich, die schnellen Ratenkonstantenaus diesen Betaverteilungen zu bestimmen (Rießner, 1994; FitzHugh, 1983; Yellen, 1984).
Im Falle des schnellen Flickerns (rechts in Fig. 5.2) genügt die Auflösung des Detektorsnicht mehr, um das volle Stromniveau zu erreichen. Es erscheint ein scheinbarerUnterniveaustrom mit einem Wert, der vom Verhältnis der Offen- und Geschlossenzeitenabhängig ist. Es wird ein scheinbarer Einzelkanalstrom mit einer kleineren Amplitudeerhalten. Permeations- und Gating-Modelle können bei dieser Auflösung nicht mehrunterschieden werden. Bei einer nicht ausreichenden Auflösung können beide Modelle zueiner Reduzierung des Einzelkanalstroms führen. In dieser Arbeit soll untersucht werden, obeine nicht ausreichende Auflösung ein Permeationsmechanismus (Wirkung auf denEinzelkanalstrom) beim AMFE vortäuscht.
Fig. 5.2: Darstellung der Reduzierung des Einzelkanalstroms verursacht durch schnelles Schalten. DieKreuze sind die Abtastwerte. „f“ kennzeichnet das volle Niveau und „s“ die scheinbare niedere Leitfähigkeit(Hansen et al., 1997).
Kapitel 5: Veränderung des Schaltverhaltens beim Block durch Metallionen
22
5.2 Reduzierte scheinbare Leitfähigkeit bei der Cs+-Blockade desK+-Kanals
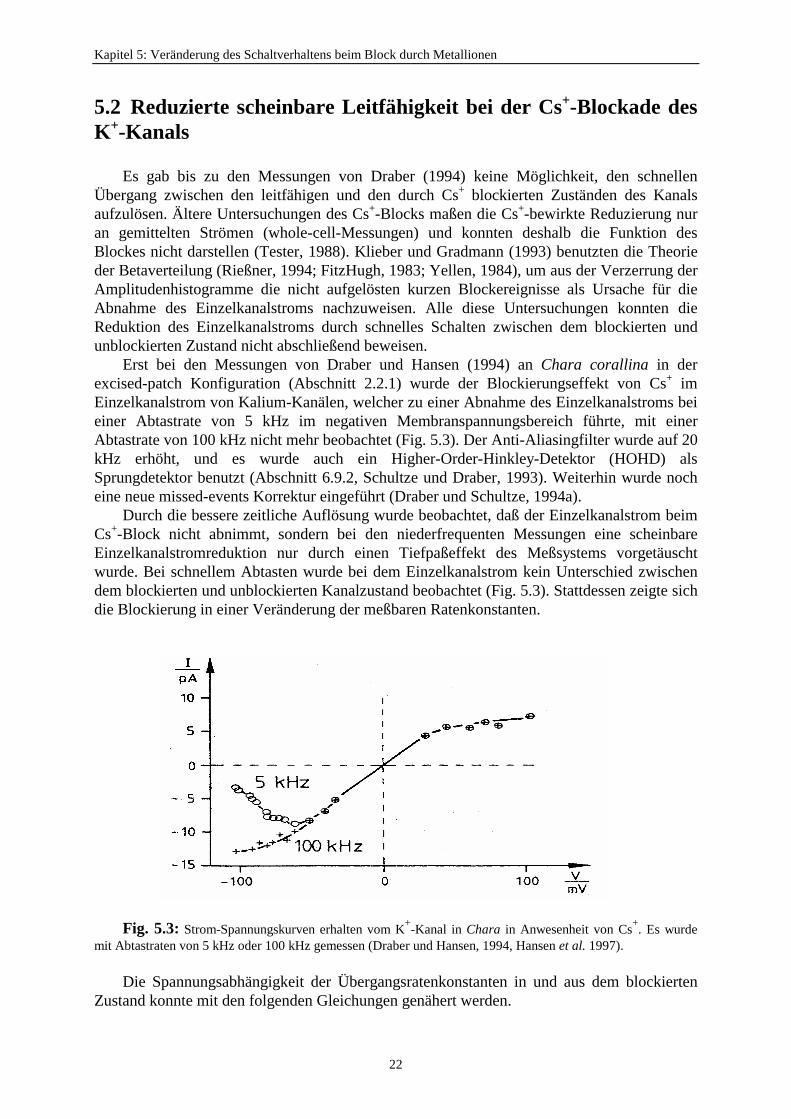
Es gab bis zu den Messungen von Draber (1994) keine Möglichkeit, den schnellenÜbergang zwischen den leitfähigen und den durch Cs+ blockierten Zuständen des Kanalsaufzulösen. Ältere Untersuchungen des Cs+-Blocks maßen die Cs+-bewirkte Reduzierung nuran gemittelten Strömen (whole-cell-Messungen) und konnten deshalb die Funktion desBlockes nicht darstellen (Tester, 1988). Klieber und Gradmann (1993) benutzten die Theorieder Betaverteilung (Rießner, 1994; FitzHugh, 1983; Yellen, 1984), um aus der Verzerrung derAmplitudenhistogramme die nicht aufgelösten kurzen Blockereignisse als Ursache für dieAbnahme des Einzelkanalstroms nachzuweisen. Alle diese Untersuchungen konnten dieReduktion des Einzelkanalstroms durch schnelles Schalten zwischen dem blockierten undunblockierten Zustand nicht abschließend beweisen.
Erst bei den Messungen von Draber und Hansen (1994) an Chara corallina in derexcised-patch Konfiguration (Abschnitt 2.2.1) wurde der Blockierungseffekt von Cs+ imEinzelkanalstrom von Kalium-Kanälen, welcher zu einer Abnahme des Einzelkanalstroms beieiner Abtastrate von 5 kHz im negativen Membranspannungsbereich führte, mit einerAbtastrate von 100 kHz nicht mehr beobachtet (Fig. 5.3). Der Anti-Aliasingfilter wurde auf 20kHz erhöht, und es wurde auch ein Higher-Order-Hinkley-Detektor (HOHD) alsSprungdetektor benutzt (Abschnitt 6.9.2, Schultze und Draber, 1993). Weiterhin wurde nocheine neue missed-events Korrektur eingeführt (Draber und Schultze, 1994a).
Durch die bessere zeitliche Auflösung wurde beobachtet, daß der Einzelkanalstrom beimCs+-Block nicht abnimmt, sondern bei den niederfrequenten Messungen eine scheinbareEinzelkanalstromreduktion nur durch einen Tiefpaßeffekt des Meßsystems vorgetäuschtwurde. Bei schnellem Abtasten wurde bei dem Einzelkanalstrom kein Unterschied zwischendem blockierten und unblockierten Kanalzustand beobachtet (Fig. 5.3). Stattdessen zeigte sichdie Blockierung in einer Veränderung der meßbaren Ratenkonstanten.
Fig. 5.3: Strom-Spannungskurven erhalten vom K+-Kanal in Chara in Anwesenheit von Cs+. Es wurdemit Abtastraten von 5 kHz oder 100 kHz gemessen (Draber und Hansen, 1994, Hansen et al. 1997).
Die Spannungsabhängigkeit der Übergangsratenkonstanten in und aus dem blockiertenZustand konnte mit den folgenden Gleichungen genähert werden.

Kapitel 5: Veränderung des Schaltverhaltens beim Block durch Metallionen
23
−= +−
− TkeVCssk
Bopengap 92.0exp][6.68 1 (5.1.a)
−= +−
− TkeVCssk
Bgapopen 17.0exp][43900 1 (5.1.b)
Hierbei haben e, V, T und kB ihre normale Bedeutung.Die Spannungsabhängigkeit eines Blocks wird häufig mit dem Woodhull-Modell (1973)
beschrieben. Die Ionen diffundieren nach diesem Modell in eine wassergefüllte Pore (Kanal),in der eine Bindungsstelle existiert. Das blockierende Ion springt über eine Energiebarriere indiese Bindungsstelle hinein. Die Ratenkonstanten für den Bindungsschritt des blockierendenIons haben eine Spannungsabhängigkeit entsprechend der Boltzmann Statistik. Für dasVerhältnis der Ratenkonstanten für die Hin- und Rückreaktion gilt die folgende Relation:
−∝
TkeVd
kk
B
exp2
1 (5.2)
Dabei ist d der Hin- und Rückweg im elektrischen Feld, den das blockierende Ion in derPore durchqueren muß. Das Woodhull-Modell (1973) war nicht in der Lage, die starkeSpannungsabhängigkeit für die Ratenkonstanten (d = 0.92 +0.17 > 1) (Formel 5.2) zuerklären.
Das Multi-Ion-Single-File-Pore Modell von Hille und Schwarz (1978) wäre theoretisch inder Lage, durch die zwei Bindestellen in der Membran eine höhere Spannungsabhängigkeit zuerklären, da die Exponenten der Spannungsabhängigkeit gekoppelter Ionen sich addieren.Draber und Hansen (1994) argumentieren, daß molekulare Daten (De Biasi et al., 1993) einemaximale Eindringtiefe des blockierenden Ions von maximal 20 % zu lassen. Damit fällt dieVoraussetzung für das Woodhull-Modell (1973) und auch die Ergänzung durch das Hille-Schwarz-Modell (1978) fort.
Weiterhin spricht gegen das Hille-Schwarz-Modell, daß es die von Draber et al. (1991)gemessene Wirkung des AMFEs nicht erklären kann (Abschnitt 3.1.2).
5.3 Der Na+-Effekt beim K+-Kanal und der Nachweis desschnellen Schaltens in Chara
Bertl (1989) untersuchte den Einfluß des Natriums auf den Block des Kaliumkanals imTonoplasten von Chara corallina. Dort betrug die Abtastrate 5 kHz gekoppelt mit einemAnti-Aliasingfilter von 1 kHz. Wie bei Cs+-Ionen tritt bei Na+-Ionen in der Lösung beiRaumtemperatur eine Reduktion der scheinbaren Einzelkanalleitfähigkeit auf. Die Idee ist wiebei Cs+, daß das unzureichende Auflösungsvermögen für die scheinbare Reduktion desEinzelkanalstroms verantwortlich ist. Der Nachweis im Falle des Na+ gestaltete sich aberschwieriger, da trotz der schnellen Abtastraten von 200 kHz und einem Anti-Aliasingfiltervon 50 kHz bei Blunck (1996) die Reduktion des Einzelkanalstroms beobachtet werdenkonnte. Weil das Öffnen und Schließen der Kanäle auf einer Konformationsänderung imProtein beruht, kann man durch Temperatursenkung die Geschwindigkeit der Übergängeverringern. Dadurch reicht die Auflösung aus, und man ist in der Lage, die unterschiedlichenZustände miteinander zu vergleichen.
Kapitel 5: Veränderung des Schaltverhaltens beim Block durch Metallionen
24
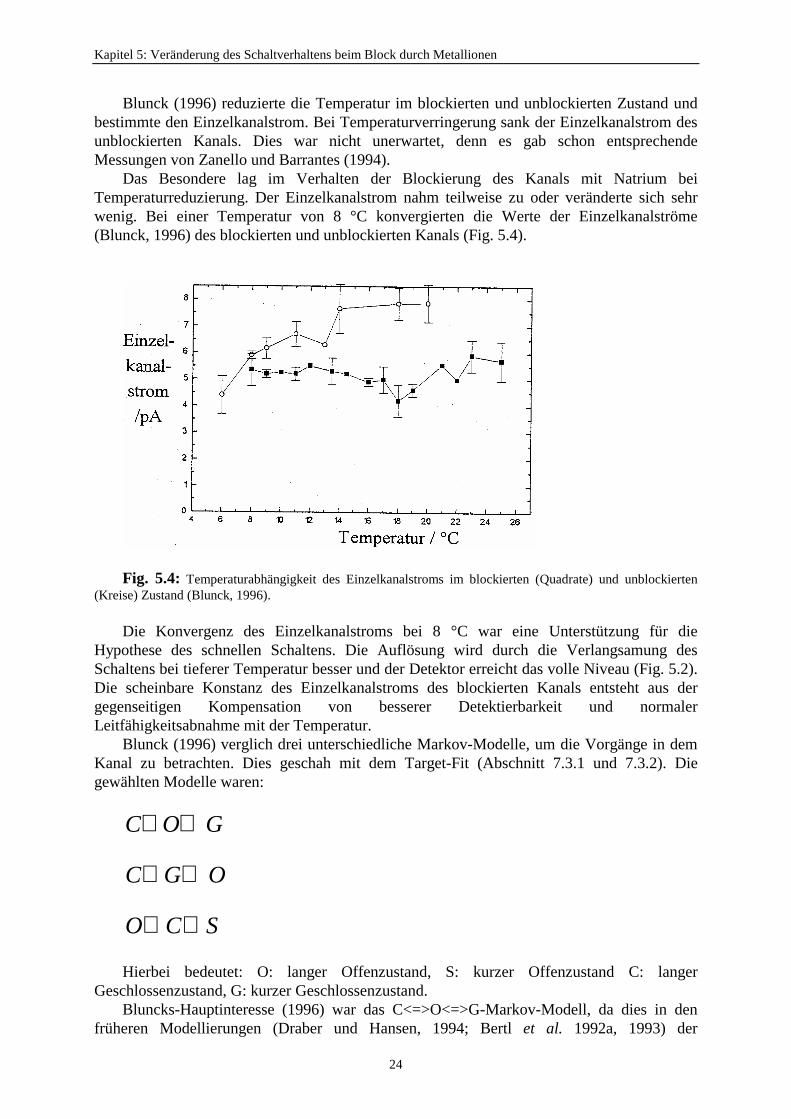
Blunck (1996) reduzierte die Temperatur im blockierten und unblockierten Zustand undbestimmte den Einzelkanalstrom. Bei Temperaturverringerung sank der Einzelkanalstrom desunblockierten Kanals. Dies war nicht unerwartet, denn es gab schon entsprechendeMessungen von Zanello und Barrantes (1994).
Das Besondere lag im Verhalten der Blockierung des Kanals mit Natrium beiTemperaturreduzierung. Der Einzelkanalstrom nahm teilweise zu oder veränderte sich sehrwenig. Bei einer Temperatur von 8 °C konvergierten die Werte der Einzelkanalströme(Blunck, 1996) des blockierten und unblockierten Kanals (Fig. 5.4).
Fig. 5.4: Temperaturabhängigkeit des Einzelkanalstroms im blockierten (Quadrate) und unblockierten(Kreise) Zustand (Blunck, 1996).
Die Konvergenz des Einzelkanalstroms bei 8 °C war eine Unterstützung für dieHypothese des schnellen Schaltens. Die Auflösung wird durch die Verlangsamung desSchaltens bei tieferer Temperatur besser und der Detektor erreicht das volle Niveau (Fig. 5.2).Die scheinbare Konstanz des Einzelkanalstroms des blockierten Kanals entsteht aus dergegenseitigen Kompensation von besserer Detektierbarkeit und normalerLeitfähigkeitsabnahme mit der Temperatur.
Blunck (1996) verglich drei unterschiedliche Markov-Modelle, um die Vorgänge in demKanal zu betrachten. Dies geschah mit dem Target-Fit (Abschnitt 7.3.1 und 7.3.2). Diegewählten Modelle waren:
GOC ⇔⇔
OGC ⇔⇔
SCO ⇔⇔
Hierbei bedeutet: O: langer Offenzustand, S: kurzer Offenzustand C: langerGeschlossenzustand, G: kurzer Geschlossenzustand.
Bluncks-Hauptinteresse (1996) war das C<=>O<=>G-Markov-Modell, da dies in denfrüheren Modellierungen (Draber und Hansen, 1994; Bertl et al. 1992a, 1993) der

Kapitel 5: Veränderung des Schaltverhaltens beim Block durch Metallionen
25
experimentellen Daten benutzt worden war. Blunck (1996) betrachtete die Ratenkonstantendes gewählten C<=>O<=>G-Markov-Modells. In der Tabelle 5.1 wurden dieseRatenkonstanten bei 20 °C und 10 °C miteinander verglichen. Die schnelle Ratenkonstante indiesem Zustandmodell war kGO.
T / °C kCO / Hz kOC / Hz kOG / Hz kGO / HzK+ + Na+ 10 50 25 100 1300K+ + Na+ 20 150 150 370 3300K+ 10 50 15 60 850K+ 20 150 75 100 1350
Tabelle 5.1: Vergleich der Ratenkonstanten des C<=>O<=>G-Modells für den blockierten undunblockierten Zustand bei 20 °C und 10 °C (Blunck, 1996).
Blunck (1996) konzentrierte sich nur auf die schnelle Ratenkonstante kGO und ist davonausgegangen, daß die anderen Übergangskonstanten (kCO, kOC und kOG) keinen Unterschied imblockierten und unblockierten Zustand zeigen. Die kGO Ratenkonstante bei 20 °Cunterscheidet sich sehr deutlich beim blockierten und beim unblockierten Zustand. DieseRatenkonstante wird geringer durch Temperatursenkung, und bei 10 °C gleichen die Wertesich für den blockierten und unblockierten Zustand an. Blunck (1996) ging davon aus, daß dieRatenkonstante kGO im blockierten Zustand verantwortlich für das schnelle Schalten undsomit auch verantwortlich für die nicht ausreichende Auflösung der schnellen Übergänge beiRaumtemperatur ist.
Blunck (1996) benutzte auch ein weiteres Markov-Modell und fand, daß dasO<=>C<=>S-Modell eine bessere Anpassung für die offen Dwell-Time-Histogramme zeigte.Das Modell C<=>O<=>G war außerdem nicht in der Lage, in langsamen Bereichen dieexperimentiellen Daten des Open-Dwell-Time-Histogramms zu beschreiben. Blunck (1996)ist davon ausgegangen, daß das C<=>O<=>G-Modell die beiden Offenzustände mit einerkleinen Abweichungen zusammenfassen kann. Es deutet sich an, daß im wahren Modell diebeiden Offenzustände zwei benachbarte Zustände sind. Die Existenz zweier Offenzuständestimmten mit den Beobachtungen der Ergebnisse von Tyerman (1992), Laver und Walker(1991), Laver (1992) und Albertsen (1994) überein. Tyerman (1992) hatte schon das ModellC<=>O<=>O<=>C vorgeschlagen. Albertsen (1994) meinte, daß einer der beidenOffenzustände in einen Sublevel übergeht. Beim schnellen Schalten kann durch Summierungüber die schnellen Schaltvorgänge bei nicht ausreichendem zeitlichen Auflösungsvermögen(Fig. 5.2) ein Offenzustand in einen Sublevel übergehen. So kann der Kanal zwischen einemschnellen (G<=>S) und einem langsameren Schaltvorgang (C<=>O) wechseln. Blunck (1996)hat das folgende Modell vorgeschlagen:
2
1
C
COSG!
⇔⇔⇔
Das O<=>G<=>C-Modell, das von Zanello und Barrantes (1994) vorgeschlagen wurde,hat bei Blunck (1996) zu keinen zufriedenstellenden Ergebnissen geführt. Die theoretischeKurve hat sich nicht den Meßdaten angepaßt. Es traten negative Ratenkonstanten auf. In den

Kapitel 5: Veränderung des Schaltverhaltens beim Block durch Metallionen
26
wenigen Fällen, bei denen gute Anpassung erfolgte, war der kurze Geschlossenzustand amEnde der Kette (G<=>C<=>O).
Der Grund für die obigen widersprüchlichen Ergebnisse wird in dem folgenden Kapitel 7deutlich: Es ist mindestens ein 5-Zustandsmodell notwendig, um den K+-Kanal in Chara zubeschreiben. Die oben genannten Autoren verfügten aber noch nicht über geeignete Softwareum 5-Zustandsmodelle an die Daten zu fitten und mußten deshalb die genanntenKompromisse eingehen.

6 Experimenteller Aufbau und Methoden
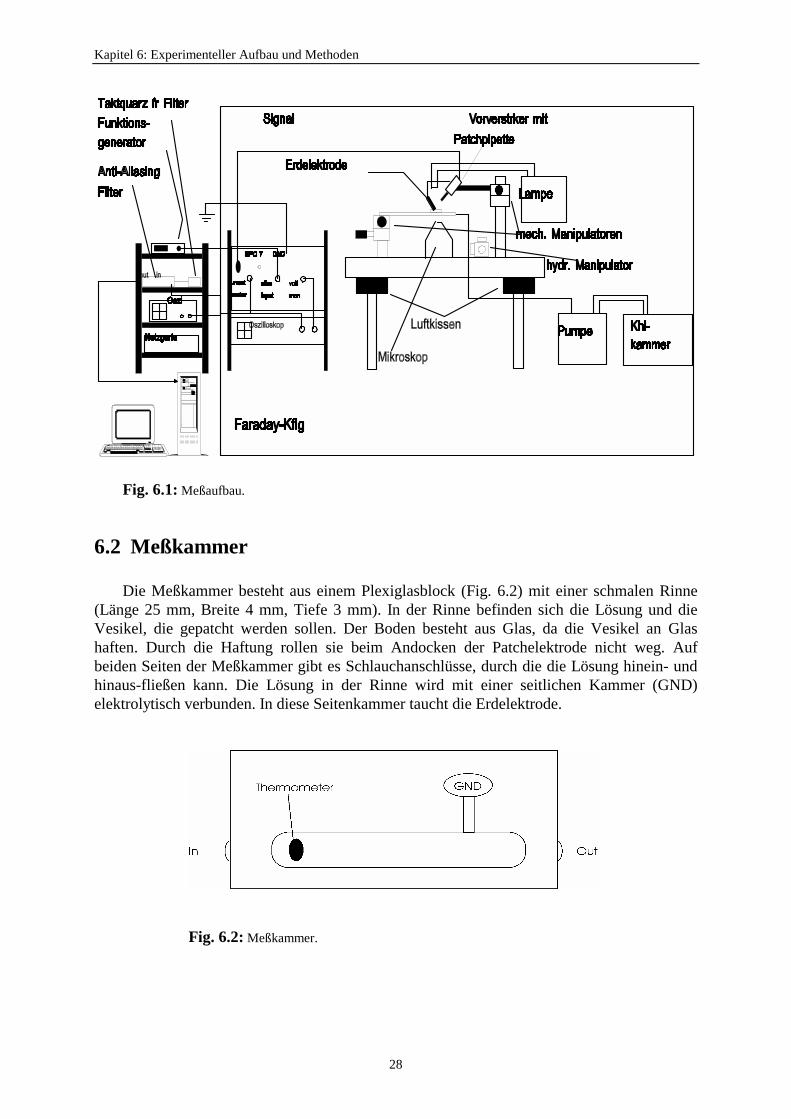
6.1 Der Aufbau
Der Versuchsaufbau wurde bereits von Albertsen (1992), Draber (1994), Blunck (1996)und Sutter (1996) benutzt. Er steht in einem Faraday-Käfig, da die Messungen sehrempfindlich gegenüber Störsignalen sind (Fig. 6.1). Ein Charakteristikum der Patch-Clamp-Technik ist die extreme Empfindlichkeit des Strom-Spannungswandlers, mit dessen Hilfekleine elektrische Ströme gemessen werden können.
Diese kleinen Signale werden leicht vom elektrischen Hintergrundrauschen überlagert. Eswird versucht, das Signal-Rausch-Verhältnis zu optimieren. Eine der wesentlichen Störquellenist die Einstreuung des elektromagnetischen Wechselfeldes (öffentliches Spannungsnetz).Diese Störung wird durch den Faraday-Käfig stark reduziert (Fig. 6.1). Die Geräte innerhalbdes Käfigs werden mit Gleichstrom (±15 V) betrieben. Die Netzgeräte dafür stehen außerhalbdes Käfigs (links in Fig. 6.1). Jedes einzelne Gerät sowie der gesamte Meßaufbau werden aneinem zentralen Punkt geerdet. Alle Leitungen, die eine Verbindung vom Innenraum zumAußenraum des Faraday-Käfigs haben, werden abgeschirmt, und die dazugehörigen Gerätegeerdet.
Das inverse Mikroskop und der Meßaufbau stehen auf einem luftgelagerten Tisch (Mittein Fig. 6.1), um mechanische Erschütterungen von außen (z.B. Schritte oder Schließen vonTüren) abzuschirmen. Sehr niederfrequente Schwingungen dämpft zusätzlich eine extremschwere Tischplatte.
Die Meßkammer liegt auf einem Objekttisch, in die gekühlte Flüssigkeit mit Hilfe einerPumpe (unten rechts in Fig. 6.1) hineinfließt. Der Objekttisch kann mit einemMikromanipulator (Fig. 6.1, Micro Control, Frankreich) in zwei Richtungen (vorwärts-rückwärts, rechts-links) bewegt werden. Der Pipettenhalter (Fig. 6.1) ist auf einem drei-dimensionalen Manipulator befestigt. Für die grobe Positionierung des Pipettenmanipulatorssteht noch ein zweidimensionaler Mikromanipulator zur Verfügung, weil für das Aufsetzender Pipette auf die Membran der dreidimensionale Manipulator aufgrund seiner hohenGenauigkeit einen zu engen Einstellbereich hat.
Das Licht zur Kontrolle des Patchvorgangs wurde von einer Halogenlampe HLX 64634(Osram) von oben eingestrahlt (oben in Fig. 6.1).
Kapitel 6: Experimenteller Aufbau und Methoden
28
Fig. 6.1: Meßaufbau.
6.2 Meßkammer
Die Meßkammer besteht aus einem Plexiglasblock (Fig. 6.2) mit einer schmalen Rinne(Länge 25 mm, Breite 4 mm, Tiefe 3 mm). In der Rinne befinden sich die Lösung und dieVesikel, die gepatcht werden sollen. Der Boden besteht aus Glas, da die Vesikel an Glashaften. Durch die Haftung rollen sie beim Andocken der Patchelektrode nicht weg. Aufbeiden Seiten der Meßkammer gibt es Schlauchanschlüsse, durch die die Lösung hinein- undhinaus-fließen kann. Die Lösung in der Rinne wird mit einer seitlichen Kammer (GND)elektrolytisch verbunden. In diese Seitenkammer taucht die Erdelektrode.
Fig. 6.2: Meßkammer.

Kapitel 6: Experimenteller Aufbau und Methoden
29
6.3 Bearbeiten und Ziehen der Pipetten
Die Patchpipette ist bereits in Fig. 2.4 vorgestellt worden. Für die Durchführung derVersuche werden als erstes die Pipetten bereitgestellt. Von den 1 m langen Glasröhrchen(Firma Hilgenberg, Malsfeld) werden ca. 7 cm abgesägt und mit Sigmacot (Sigma,Deisenhofen) von innen mit einer Spritze beschichtet. Diese Behandlung schirmt dieOberflächenladung ab. Hierdurch wird auch das Rauschen vermindert (Keunecke, 1995) unddie sogenannte Ω-Bildung (Einkriechen des Patches in die Pipette) verhindert, welche dieLebensdauer eines Patches verringert. Anschließend werden die Glasröhrchen zwei Tage langunter dem Abzug getrocknet, damit das nicht gebundene Sigmacot verdampft. Es wird wegender Giftigkeit des Sigmacots unter dem Abzug mit Handschuhen gearbeitet.
Die Pipetten werden am Tag der Versuchsdurchführung mit einem speziellenMikroelektrodenziehgerät L/M-3P-A (List, Darmstadt) ausgezogen. Das Glasröhrchen wird indie Ziehvorrichtung eingespannt und mit einem Gewicht belastet. Die Mitte des Röhrchensumgibt eine Heizspule mit einstellbarem Heizstrom. Durch die Hitze schmilzt dasGlasröhrchen und wird in die Länge gezogen. Der Pipettenziehvorgang besteht aus zweiStufen. In der zweiten Stufe wird das Glasröhrchen abgerissen und es entstehen zwei Pipettenmit einem Öffnungsdurchmesser von etwa 1 µm.
Die Pipetten verschmutzen sehr schnell an der Spitze. An einer verschmutzten Pipettebilden sich keine Seals. Deshalb wurden sie abgedeckt und in einem staubfreien Gehäuseaufbewahrt.
Die Pipetten werden anschließend poliert, um durch die dadurch entstandene glattereOberfläche eine bessere Haftung zwischen Pipettenspitze und Membran zu erreichen. Dazuwird die Pipettenspitze vor einem glühenden Draht unter einem Mikroskop angeschmolzen.Der Draht ist mit Glas beschichtet, damit keine Ionen emittiert werden, die die Pipettenspitzeverschmutzen könnten.
6.4 Erd- und Pipettenelektrode
Für die Herstellung der Erdelektrode (Fig. 6.2) wird zuerst eine Lösung aus 250 mMKNO3 , 5 mM Ca(NO3)2 und 3% Agar-Agar aufgekocht. Die Elektrodenlösung darf keinengroßen Einfluß auf die Lösung in der Meßkammer haben. Das Agar-Agar unterbindet deshalbden Ausfluß der Lösung aus der Erdelektrode. Die gekochte Lösung wird in einePasteurpipette gegeben und abgekühlt. Er wird darauf geachtet, daß sich keine Luftbläschenbilden, damit durchgehende Leitung gewährleistet ist. Sie wird über Nacht gekühlt, und amnächsten Tag wird ein chlorierter Silberdraht (Ag/AgCl; ∅ =1mm) bis zur Hälfte in diePasteurpipette gesetzt. Die Chlorierung führt dazu, daß der Draht nicht polarisierbar wird. DiePatchpipette muß eine elektrische Verbindung von der Membran zumStromspannungswandler bilden. Dazu wird sie in der vorderen Hälfte mit einer Salzlösung(250 mM KCl + 5 mM Ca(Cl)2) gefüllt. In die Salzlösung taucht ein chlorierter Silberdraht alsnicht polarisierbare Elektrode, der mit einem Ende an einen BNC-Stecker gelötet ist. Dieserkann direkt auf den Patchverstärker aufgesteckt werden.

Kapitel 6: Experimenteller Aufbau und Methoden
30
6.5 Der Patch-Clamp Verstärker
Der Patch-Clamp-Verstärker besteht aus einem Strom-Spannungswandler, der dasStromsignal der Pipette mißt und als Spannung ausgibt, und dem Nachverstärker, der dasSignal verstärkt und filtert. Der Strom-Spannungswandler steht stabil und erschütterungsfreiauf dem Pipettenmanipulator nahe der Meßkammer.
Die Ströme, die die Membran durchfließen, liegen im Bereich einiger Picoampere.Deshalb muß ein sehr empfindlicher Strom-Spannungskonverter mit hoher Bandbreite benutztwerden. Eines der Standardgeräte ist der L/M-EPC7 (List, Darmstadt) mit einer Bandbreitevon 100 kHz und bei einem Rückkopplungswiderstand Rf von 10 GΩ.
In Fig. 6.3 ist ein vereinfachtes Schaltbild des Patchverstärkers dargestellt. Der OP regeltden Unterschied zwischen Upip und Usoll auf Null. Das geschieht so, daß die Differenz derEingangsspannungen extrem verstärkt wird. Es entsteht zwischen Punkt 1 und Punkt 2 einSpannungsunterschied, so daß durch Rf ein Strom mit einer Spannung von Uf = Rf I fließt.Dieser Strom fließt in der Pipette und nicht in den OPA (OPA hat einen unendlich hohenEingangswiderstand). Der Strom ändert das Potential an Punkt 1 (Upip), bis dieEingangsspannungen gleich sind. Die Spannung am Punkt 1 ist gleich der Pipetten- bzw.Membranspannung, da der Spannungsabfall über der Pipette vernachlässigt wird. DieAbweichung zwischen der Pipettenspannung und der Sollspannung wird ausgeglichen. DieSpannung über Rf ist proportional zum Strom in der Pipette. Usoll wird vom zweitenOperationsverstärker abgezogen. Das Signal geht von dort zum Nachverstärker.
Der Stromspannungswandler verursacht Rauschen, das im Allgemeinen aber keine Rollespielt, da der entscheidene Beitrag des Rauschens durch die Pipette (Keunecke, 1995)entsteht.
Fig. 6.3: Vereinfachtes Schaltbild eines Patch-Clamp-Verstärkers.
Das Signal wird vom Verstärker zu einem 8-poliges Besselfilter außerhalb des Faraday-Käfigs weitergeleitet. Es dient als Anti-Aliasingfilter für die Datenerfassung. Der digitaleBesselfilter wird mit einer ±5 V Rechteckspannung getaktet (Albertsen, 1992). DieAbknickfrequenz ist 1/65 der Taktfrequenz. Sie betrug bei dieser Messung 25 kHz. Das

Kapitel 6: Experimenteller Aufbau und Methoden
31
Ausgangssignal des Besselfilters wird dann an einen digitalen Signal-Prozessor weitergeleitet(Dalanco Spry). Dieses Board kann das Signal bis zu 200 kHz abtasten. Es war bei den indieser Arbeit durchgeführten Messungen auf 100 kHz eingestellt. Durch das 25-kHz Filterkam es nicht zu einem Verstoß gegen das Abtasttheorem.
Die Daten werden per Knopfdruck durch das Aufnahmeprogramm „Sample“ auf dieFestplatte eines PC’s (486DX66) gespeichert. Es werden bei verschiedenen SpannungenAufnahmen gemacht. Die Dauer jeder Aufnahme betrug 20 Sekunden. Je länger die Meßzeitist, desto kleiner ist die Streuung der Dwell-Time Histogramme, aber desto kleiner wird dieWahrscheinlichkeit, daß das Gigaseal überlebt, besonders bei hoher Haltespannung. In demAufnahmeprogramm werden die Samplefrequenz, der Dateiname und die Länge des Meßfilesdokumentiert. Nach der Auswertung werden die Files auf eine CD gebrannt.
6.6 Das Patchen
Aus einem Anzuchtbecken werden die Algen Chara corallina entnommen, und dieRadialzellen werden von den Internodialzellen entfernt. Die Internodialzellen der Alge werdendann für die Vorbereitung des Versuchs, nachdem sie 10 Minuten getrocknet worden sind,aufgeschnitten, und der Zellinhalt wird vorsichtig mit zwei Fingern in der Lösung derMeßkammer ausgedrückt. Es bilden sich Vesikel (Tröpfchen), die am Boden der Rinne derMeßkammer haften. Je kleiner die Krümmung der Vesikeloberfläche, desto größer ist dieWahrscheinlichkeit einer Bildung des Gigaseals. Da die Vesikel einen Durchmesser um die 50µm haben, wird ein invertierendes Mikroskop (Zeiss ID02) mit einem Objektiv von 5×, 10×und 20× benutzt.
Danach wird die Pipette auf den Halter gesteckt und in die Lösung getaucht. Mit demMund wird über einen seitlich angebrachten Schlauch ein Überdruck erzeugt. Durch diesenÜberdruck wird der Schmutz an der Oberfläche weggeblasen, so daß die Pipettenspitze beimDurchfahren der Lösung nicht durch herumschwimmende Tropfentrümmer verunreinigt wird.Bei eingetauchter Pipette wird der Offsetstrom im EPC7-Verstärker (Abschnitt 6.5) auf Nullkompensiert. Dann folgt das Anlegen einer Rechteckspannung für die Kontrolle desSealvorganges. Der von ihr erzeugte Strom wird kleiner, wenn das Seal die Pipetteverschließt. Die Pipette wird mit Hilfe des Mikromanipulators in die Nähe des Vesikelsgebracht. Die Pipette wird schräg bis senkrecht von oben auf das Vesikel zu bewegt. Es wirdkurz vor Berührung der Membran ein leichter Unterdruck erzeugt, so daß das Vesikel an diePipette herangezogen wird. Am Oszillographen wird beobachtet, daß bei Sealbildung derrechteckige Pipettenstrom zusammenschrumpft. Bei einem hieraus ermittelten Widerstand imGiga-Ohm-Bereich ist die Verbindung zwischen dem Glasrand der Pipettenöffnung und derMembran fest. Man spricht von einem Gigaseal. Die Pipette wird dann ruckartigzurückbewegt und es bildet sich ein „excised Patch“. Das am Pipettenmund klebendeMembranstück (Patch) wird aus der Zelle herausgerissen (Abschnitt 2.2.1). Danach wird derPatch dicht unter die Oberfläche gebracht und somit das Rauschen reduziert (Keunecke,1995). Der kapazitive Rauschstrom fließt hauptsächlich durch die Kapazität der benutztenPipettenwand. Somit ist das Rauschen sehr stark von der Eintauchtiefe des Patches abhängig.Das Rauschen nimmt bei einer Eintauchtiefe von 20 µm um den Faktor 2.2 ± 0.5 ab(Keunecke, 1995). Dann wird die Aufnahme durch den Rechner außerhalb des Faraday-Käfigsüber eine Fernbedienung von innen gestartet. Vorher wird noch die Empfindlichkeit desVerstärkers auf 100 mV/pA erhöht.

Kapitel 6: Experimenteller Aufbau und Methoden
32
Die Abfolge der Messung war meist so, daß zuerst die Messung bei Raumtemperaturdurchgeführt, dann die Platte gekühlt und schließlich nach zwanzig Minuten die Messung bei7°C durchgeführt wurde.
6.7 Die Kühlung
Die Kühlung spielt eine tragende Rolle in dieser Arbeit, da versucht werden soll, beitiefen Temperaturen die hypothetischen Schaltprozesse so weit zu verlangsamen, daß sie einerdirekten Beobachtung zugänglich werden.
Der Tauchkühler und die Pumpe für den Kühlkreislauf (Fig. 6.1) wird während derMessung nicht in Betrieb genommen und führt dadurch nicht zu Störungen. Um die Messungauch bei tieferer Temperatur durchzuführen, wird vorausgesetzt, daß die Badlösung schnellabgekühlt wird, die Temperaturschwankung gering gehalten wird und keine zusätzlichenStörungen entstehen. Diese Voraussetzungen lassen sich am einfachsten durch die direkteAbkühlung des Objekttisches erfüllen. Die Kühldauer betrug 20 min. Dadurch wurde derWunsch nach einer schnellen Kühlung nicht ganz erfüllt, aber die meisten Seals hatten eineLebensdauer von mehr als 30 min. Der entscheidende Vorteil war jedoch, daß der Seal nichtunter mechanischer Belastung stand. Die Methode des Lösungsaustauschkreislaufs mit derMikropumpe zur Kühlung der Lösung (Blunck, 1996) hatte den großen Nachteil, daß der Sealsehr häufig durch die mechanische Belastung beim Lösungsaustausch zusammenbrach. ImGegensatz zu Bluncks Kühlplatte war die Meßkammer (Abschnitt 6.2) nicht nur auf der Plattemontiert, sondern wurde von der Platte auf allen Seiten eingeschlossen Der Objekttisch warwährend der gesamten Meßdauer auf die tiefe Temperatur gekühlt. So war die Meßtemperaturwährend der gesamten Aufnahme einer Zeitreihe konstant.
6.8 Temperaturmessung und Probleme durch die Kühlung
Die Temperatur wurde mit einem NTC-Thermometer gemessen. Der größte Vorteildieses Thermometers war, daß es eine kleine Spitze hatte und somit in die schmale Rinne derMeßkammer paßte. Es zeigte die Temperatur mit einer Abweichung von ±0.5 °C.
Da sich nur der Objekttisch abkühlte und die Umgebung des Meßaufbaus sich weiterhinin Raumtemperatur befand, traten folgende Probleme auf: An Teilen der Meßapparatur,besonders der Meßkammer und an der Erdelektrode kondensierte Wasser. Es entstandendadurch Kriechströme an der Meßkammer und Erdelektrode zu den Metallteilen des Aufbausoder des Drahtanschlusses. Die Offsetspannung änderte sich und konnte nicht mehrkompensiert werden. Durch das Kondenswasser am Objektiv konnte weder die Spitze derPipette beobachtet werden, noch konnte man die Bewegung der Pipette an die Oberfläche derLösung kontrollieren.
Die Meßkammer und ein Teil des Objekttisches wurden deshalb mit Siliconöleingerieben. Dies ergab eine sehr gute Isolation der Meßkammer. Außerdem wurdenExsikkationsperlen in Teebeuteln rund um die Meßkammer gelegt, damit nur getrocknete Luftdie Meßkammer erreicht. Diese Trockenperlen wurden ein- bis zweimal am Tag ausgetauschtund verloren ihre Feuchtigkeit in einem Trockenofen (100 °C, 6 h) wieder.

Kapitel 6: Experimenteller Aufbau und Methoden
33
6.9 Auswerteprogramm
In Fig. 6.4 ist eine Zeitreihe der Grünalge Chara corallina mit Tl+-haltiger Lösung beiRaumtemperatur dargestellt. Die Zeitreihe ist bei einer Haltespannung von -80 mVaufgenommen worden. Es werden drei unterschiedliche Niveaus erkannt. Das obere Niveauist das Nullniveau, bei dem alle Kanäle geschlossen sind. Diese Zeitreihe besteht aus zweigleichartigen Kanälen mit je einem Einzelkanalstrom von -3.3 pA. Das Rauschen beträgt ± 1.2pA. Auf der linken Seite von Fig. 6.4 ist das Schalten der beiden Kanäle deutlich erkennbar,und auf der rechten Seite von Fig. 6.4 wird festgestellt, daß die Kanäle für eine längere Zeitgeschlossen bleiben. Diese Phasen, in denen der Kanal hauptsächlich geschlossen ist, werdenin vielen Zeitreihen beobachtet. Diese langen geschlossenen Zustände werden dann durchkurze Offenereignisse unterbrochen (die ersten beiden Offenereignisse von der rechten Seiteder Fig. 6.4).
Die schnellen Ereignisse (Bursts) werden besonders deutlich, wenn beide Kanäle offensind. Es wird sichtbar, daß besonders bei zwei offenen Kanälen der Offenzustand durchschnelle Folgen von sehr kurzen Geschlossenzustände (Gaps) des einen Kanals unterbrochenwerden. Einige Burst sind teilweise nicht gut erkennbar und die Gaps erreichen die Nullinienicht mehr. Dies wird in gewissen Bereichen von Fig. 6.4 erkannt. Dieses Verhalten deutet aufein schnelles Schalten hin, in dem die Offenereignisse nur für eine kurze Zeit existieren.
Wie man solche Zeitreihen behandelt und weiter auswertet, wird im folgenden Abschnittund im Kapitel 7 erläutert.
Fig. 6.4: Ein typischer Zeitverlauf des Einzelkanalstromes in einer gemischten Lösung aus K+- und Tl+-Ionen bei Raumtemperatur und Vhold = -80 mV. Die Gesamtkonzentration ist 250 mM (230 mM KNO3 +20 mMTlNO3). Der Einzelkanalstrom ist mit 100 kHz abgetastet und mit 25 kHz gefiltert. Die hellen Linien sindCursoreinstellungen.
6.9.1 Bestimmung von Kanalzahl, Niveaus und Standardabweichung
Für die Auswertung der Zeitreihe (Fig. 6.4) wird das Programm „day“ (Kirst, 1996)benutzt. Es werden zuerst die Stromniveaus und das Rauschen bestimmt. Das Programmbietet zwei Möglichkeiten, die Einzelkanalstromniveaus zu ermitteln. Erstens besteht dieMöglichkeit, daß die gesamte oder nur ein Abschnitt der Zeitreihe betrachtet wird. Es wird einAbschnitt gewählt, in dem das Schalten gut erkennbar ist (Fig. 6.4). Die Niveaus werden nunmit dem Cursor von Hand eingestellt. Danach wird die Zeitreihe als Amplitudenhistogrammdargestellt und das Rauschen bestimmt (Fig 7.3). Im Amplitudenhistogramm können die

Kapitel 6: Experimenteller Aufbau und Methoden
34
Niveaus auch mit einem Simplex nachgefittet werden. Hierzu muß die Anzahl der Kanälebekannt sein, und es besteht die Möglichkeit, daß sich der Fit bei schlechten Startwertenverläuft. Daher wird die erste Methode (Fit-by-eye) vorgezogen, und dann imAmplitudenhistogramm mit diesen Startwerten und mit dem Simplex (entwickelt von Nedlerund Mead, 1965; als Programm dargestellt von Caceci und Cacheris, 1984) verifiziert. DieNachkorrektur durch das Fitten ist zumeist sehr gering. Nachdem die Niveaus, ihre Anzahlund die Standardabweichung bestimmt worden sind, wird die Zeitreihe durch den Hinkley-Detektor ausgewertet (Abschnitt 6.9.2). Das ergibt die idealisierte Zeitreihe, die für dieErstellung der Verweildauerhistogramme benötigt wird (Fig. 7.6, 7.7 und 7.8).
6.9.2 Die Rekonstruktion der Zeitreihe mit dem Hinkley-Detektor
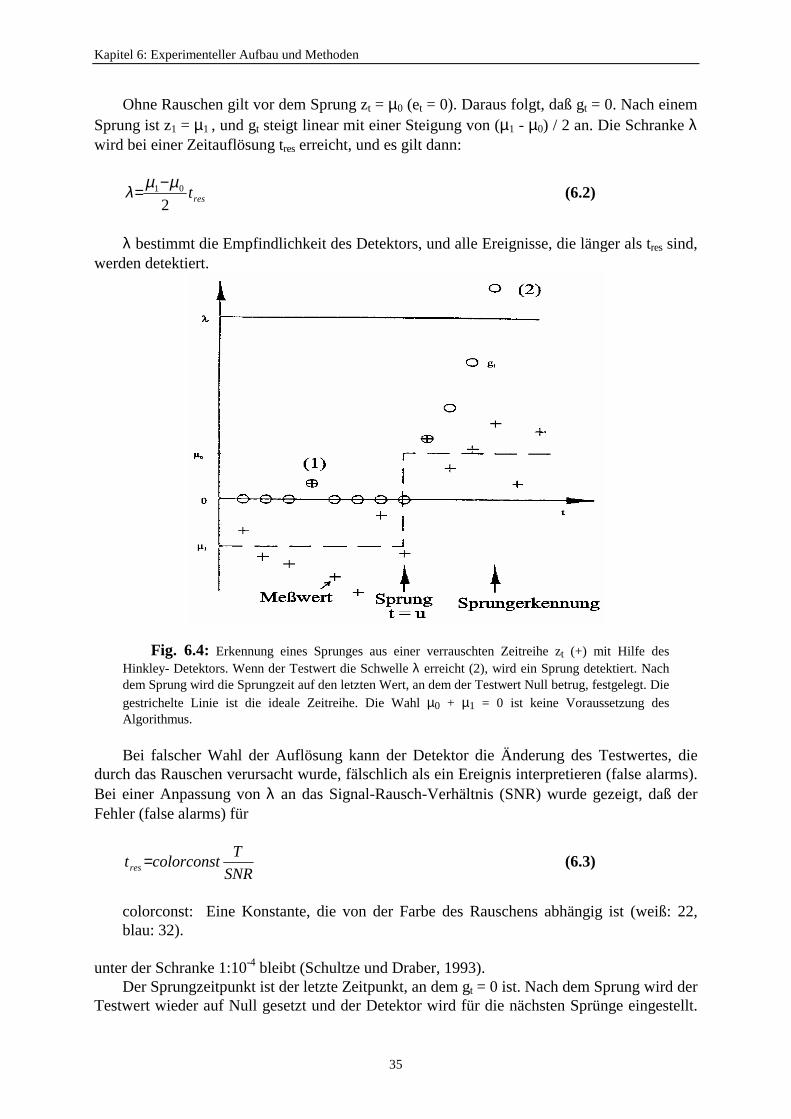
Um die biophysikalischen Mechanismen des Schaltverhaltens eines Kanals mit Markov-Modellen (Abschnitt 4.1) beschreiben zu können, muß mit Hilfe eines Algorithmus aus derverrauschten Zeitreihe des Stroms durch die Membran (Fig. 6.4) eine ideale Zeitreihe (ohneRauschen) möglichst fehlerfrei rekonstruiert werden.
Der am häufigsten verwendete Algorithmus ist ein Tiefpaßfilter mit nachfolgendemSchwellenwertdetektor, der für die Detektion eines Sprunges auf die halbe Sprunghöheeingestellt ist. Schultze und Draber (1993) führten den Hinkley-Detektor erster und höhererOrdnung mit einem besseren Auflösungsvermögen in die Patch-Clamp-Analyse ein. Dies istfür diese Arbeit eine entscheidende Voraussetzung, um den Nachweis der schnellenSchaltvorgänge beim anomalen Molfraktionseffekt (Kapitel 3) mit Thallium im Kaliumkanalzu erbringen. Somit wird der Hinkley-Detektor für diese Arbeit benutzt (Fig. 6.4).
Der Hinkley-Detektor benötigt wie alle anderen Detektoren für die Detektion einesSprunges zwischen zwei Niveaus die Anzahl und die Lage der Stromniveaus. Diese werdenmit Hilfe des Programms „day“ entweder aus dem Amplitudenhistogramm oder direkt durchdie visuelle Inspektion der Zeitreihe (Abschnitt 6.9.1) festgelegt. Der Algorithmus desHinkley-Detektors funktioniert so, daß aus den Meßpunkten der Zeitreihe zt ein Testwert gtberechnet wird, der aus der Summe der vorherigen Testwerte g(t-1) und der Differenz desMeßwertes vom Absprungsniveau et sowie dem Mittelwert p der Niveaus i und j entsteht undschließlich diesen Testwert mit einer Schranke λ vergleicht (Schwellenwertdetektor) (Fig.6.4). Bei negativen Testwerten wird er gleich Null gesetzt. Dadurch werden allevorhergehenden Werte nicht mehr berücksichtigt. Der Detektor bleibt immer „wach“ und estreten keine unnötigen Verzögerungen der Sprungdetektion auf.
−±+
=±−±
0)(
max 1 pegg tt
t (6.1)
mit
g0 = 0p = (µ1 - µ0) / 2µ0: Absprungsniveauµ1: Zielniveauet = zt - µ0± Sprung nach oben (+) bzw nach unten (-)
Kapitel 6: Experimenteller Aufbau und Methoden
35
Ohne Rauschen gilt vor dem Sprung zt = µ0 (et = 0). Daraus folgt, daß gt = 0. Nach einemSprung ist z1 = µ1 , und gt steigt linear mit einer Steigung von (µ1 - µ0) / 2 an. Die Schranke λwird bei einer Zeitauflösung tres erreicht, und es gilt dann:
rest2
01 µµλ −= (6.2)
λ bestimmt die Empfindlichkeit des Detektors, und alle Ereignisse, die länger als tres sind,werden detektiert.
Fig. 6.4: Erkennung eines Sprunges aus einer verrauschten Zeitreihe zt (+) mit Hilfe desHinkley- Detektors. Wenn der Testwert die Schwelle λ erreicht (2), wird ein Sprung detektiert. Nachdem Sprung wird die Sprungzeit auf den letzten Wert, an dem der Testwert Null betrug, festgelegt. Diegestrichelte Linie ist die ideale Zeitreihe. Die Wahl µ0 + µ1 = 0 ist keine Voraussetzung desAlgorithmus.
Bei falscher Wahl der Auflösung kann der Detektor die Änderung des Testwertes, diedurch das Rauschen verursacht wurde, fälschlich als ein Ereignis interpretieren (false alarms).Bei einer Anpassung von λ an das Signal-Rausch-Verhältnis (SNR) wurde gezeigt, daß derFehler (false alarms) für
SNRTcolorconsttres = (6.3)
colorconst: Eine Konstante, die von der Farbe des Rauschens abhängig ist (weiß: 22,blau: 32).
unter der Schranke 1:10-4 bleibt (Schultze und Draber, 1993).Der Sprungzeitpunkt ist der letzte Zeitpunkt, an dem gt = 0 ist. Nach dem Sprung wird der
Testwert wieder auf Null gesetzt und der Detektor wird für die nächsten Sprünge eingestellt.

Kapitel 6: Experimenteller Aufbau und Methoden
36
Bei einem Niveau, in dem Sprünge nach oben und nach unten möglich sind, werden zweiparallel laufende Detektoren, die gleichzeitig nach oben und nach unten schauen, benötigt.
Beim Higher-Order-Hinkley-Detektor wird anstelle der einfachen Ordnung desAlgorithmus eine höhere Ordnung benutzt. Diese unterdrückt die hochfrequentenRauschanteile stärker als bei der ersten Ordnung des Algorithmus. Der Testwert der höherenOrdnung läßt sich berechnen als:
≥+
=−
−
sonstgfürTgg
g ti
ti
tit 0
0)1()1()(1)( (6.4)
gti :: Der Testwert i-ter Ordnung.
Bei einer Überschreitung des Testwertes gi über die Schranke λ wird ein Sprung erkannt.Die Testwerte werden alle wie beim Hinkley-Detektor erster Ordnung gleich Null gesetzt,wenn gi < 0 ist. Das Prinzip des Higher-Order-Hinkley-Detektors entspricht dem desursprünglichen Hinkley-Detektors.
Diese Auswertung wird für alle Membranspannungsbereiche ausgeführt. Nur dieSpannungswerte von ±20 mV wurden wegen des schlechten Signal-Rausch-Verhältnissesnicht für die Verweildauerhistogramme benutzt. Für die Strom-Spannungskurven sind aberalle Daten verwendet worden. Die genauere Auswertung der Stromniveaus und die Dwell-Time-Histogramme werden in dem folgenden Kapitel beschrieben.

7 Meßergebnisse
7.1 Strom-Spannungskurven
7.1.1 Bestimmung der Einzelkanalströme
Die Untersuchungen wurden am Plasmatropfen der Süßwasseralge Chara corallina(Abschnitt 2.1.4) unter ähnlichen Bedingungen durchgeführt wie in den Untersuchungen vonKeunecke (1995) und Albertsen (1994). Dies bezieht sich insbesondere auf denLösungsaustausch auf nur einer Seite der Membran.
Die Messungen fanden bei Raumtemperatur oder bei 7°C statt, wie im folgendenangegeben. Für die Lösungen im Bad und in der Pipette wurde eine Gesamtkonzentration von250 mM statt 150 mM wie bei Albertsen (1994) und Keunecke (1995) benutzt. Die höherenSalzkonzentrationen führten auf höhere Einzelkanalströme und damit auf ein besseres Signal-Rauschverhältnis.Die Zusammensetzung der Tl+-freien Lösung war bei den folgenden Messungen
250 mM KNO3 + 5 mM Ca(NO3 )2 .
In der Tl+-haltigen Lösung war die Zusammensetzung so, daß das Verhältnis vonThallium zu Kalium dem Minimum der Leitfähigkeit in den Messungen von Keunecke (1995)entsprach. Das ergab eine Zusammensetzung von
230 mM KNO3 + 20 mM TlNO3 + 5 mM Ca(NO3 )2 .
Ein Unterschied gegenüber den Messungen von Keunecke (1995) bestand darin, daß fürden Lösungsaustausch nicht gespült wurde. Der Seal brach sehr häufig durch die mechanischeBelastung beim Lösungsaustausch. Das setzte voraus, daß Messungen mit verschiedenenBadlösungen an verschiedenen Seals durchgeführt werden mußten. Dies erhöht die statistischeSchwankung allerdings nicht allzu sehr, weil die Einzelkanalströme sehr reproduzierbar sind.
Die Aufnahme der Daten und die Durchführung der Versuche folgte dem in Abschnitt 6.5und 6.9 dargestellten Protokoll. Die Abknickfrequenz des Anti-Aliasingfilters lag bei 25 kHz.Sie war um den Faktor zwei kleiner als bei Keunecke (1995).
Fig.7.1 stellt gemessene Zeitreihen für je eine Aufzeichnung in reiner K+-Lösung(Fig.7.1.A) und in K+/Tl+-Mischung dar (Fig.7.1.B). Es handelt sich um einen Patch mit zweiKanälen. Gut zu erkennen ist die Höhe der Sprünge von einem Niveau zum anderen. DieSprunghöhe ergibt den Einzelkanalstrom.
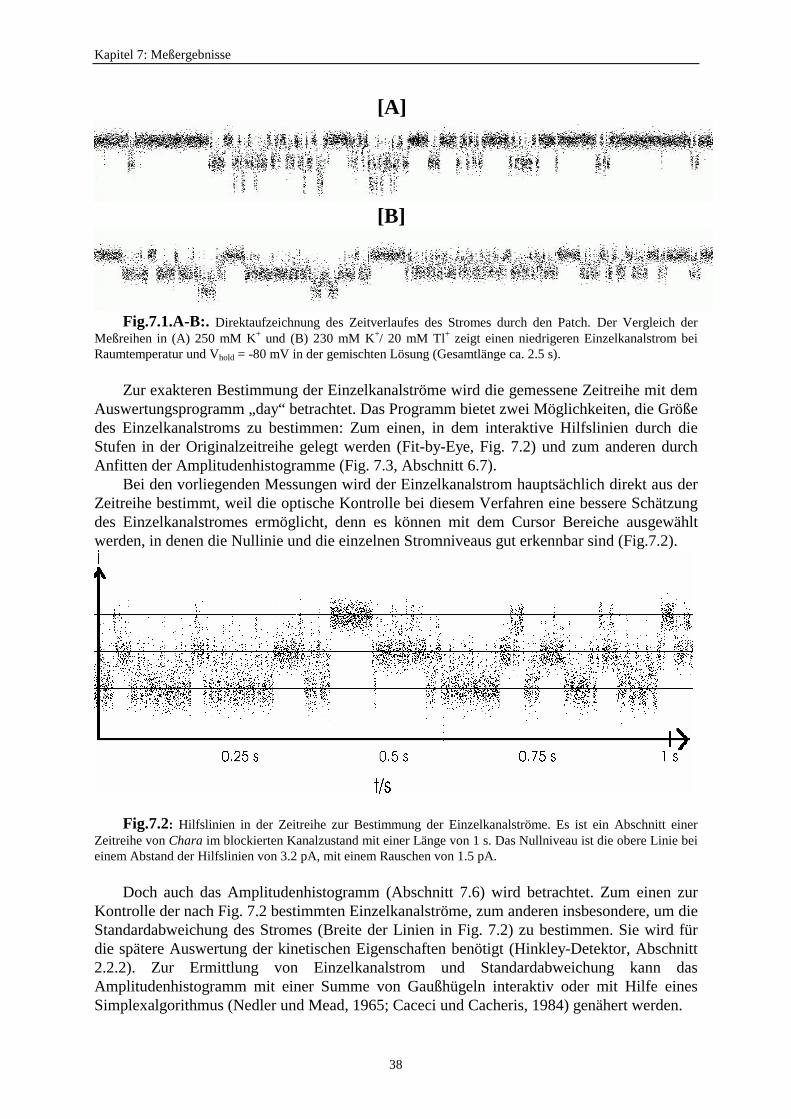
Kapitel 7: Meßergebnisse
38
[A]
[B]
Fig.7.1.A-B:. Direktaufzeichnung des Zeitverlaufes des Stromes durch den Patch. Der Vergleich derMeßreihen in (A) 250 mM K+ und (B) 230 mM K+/ 20 mM Tl+ zeigt einen niedrigeren Einzelkanalstrom beiRaumtemperatur und Vhold = -80 mV in der gemischten Lösung (Gesamtlänge ca. 2.5 s).
Zur exakteren Bestimmung der Einzelkanalströme wird die gemessene Zeitreihe mit demAuswertungsprogramm „day“ betrachtet. Das Programm bietet zwei Möglichkeiten, die Größedes Einzelkanalstroms zu bestimmen: Zum einen, in dem interaktive Hilfslinien durch dieStufen in der Originalzeitreihe gelegt werden (Fit-by-Eye, Fig. 7.2) und zum anderen durchAnfitten der Amplitudenhistogramme (Fig. 7.3, Abschnitt 6.7).
Bei den vorliegenden Messungen wird der Einzelkanalstrom hauptsächlich direkt aus derZeitreihe bestimmt, weil die optische Kontrolle bei diesem Verfahren eine bessere Schätzungdes Einzelkanalstromes ermöglicht, denn es können mit dem Cursor Bereiche ausgewähltwerden, in denen die Nullinie und die einzelnen Stromniveaus gut erkennbar sind (Fig.7.2).
Fig.7.2: Hilfslinien in der Zeitreihe zur Bestimmung der Einzelkanalströme. Es ist ein Abschnitt einerZeitreihe von Chara im blockierten Kanalzustand mit einer Länge von 1 s. Das Nullniveau ist die obere Linie beieinem Abstand der Hilfslinien von 3.2 pA, mit einem Rauschen von 1.5 pA.
Doch auch das Amplitudenhistogramm (Abschnitt 7.6) wird betrachtet. Zum einen zurKontrolle der nach Fig. 7.2 bestimmten Einzelkanalströme, zum anderen insbesondere, um dieStandardabweichung des Stromes (Breite der Linien in Fig. 7.2) zu bestimmen. Sie wird fürdie spätere Auswertung der kinetischen Eigenschaften benötigt (Hinkley-Detektor, Abschnitt2.2.2). Zur Ermittlung von Einzelkanalstrom und Standardabweichung kann dasAmplitudenhistogramm mit einer Summe von Gaußhügeln interaktiv oder mit Hilfe einesSimplexalgorithmus (Nedler und Mead, 1965; Caceci und Cacheris, 1984) genähert werden.
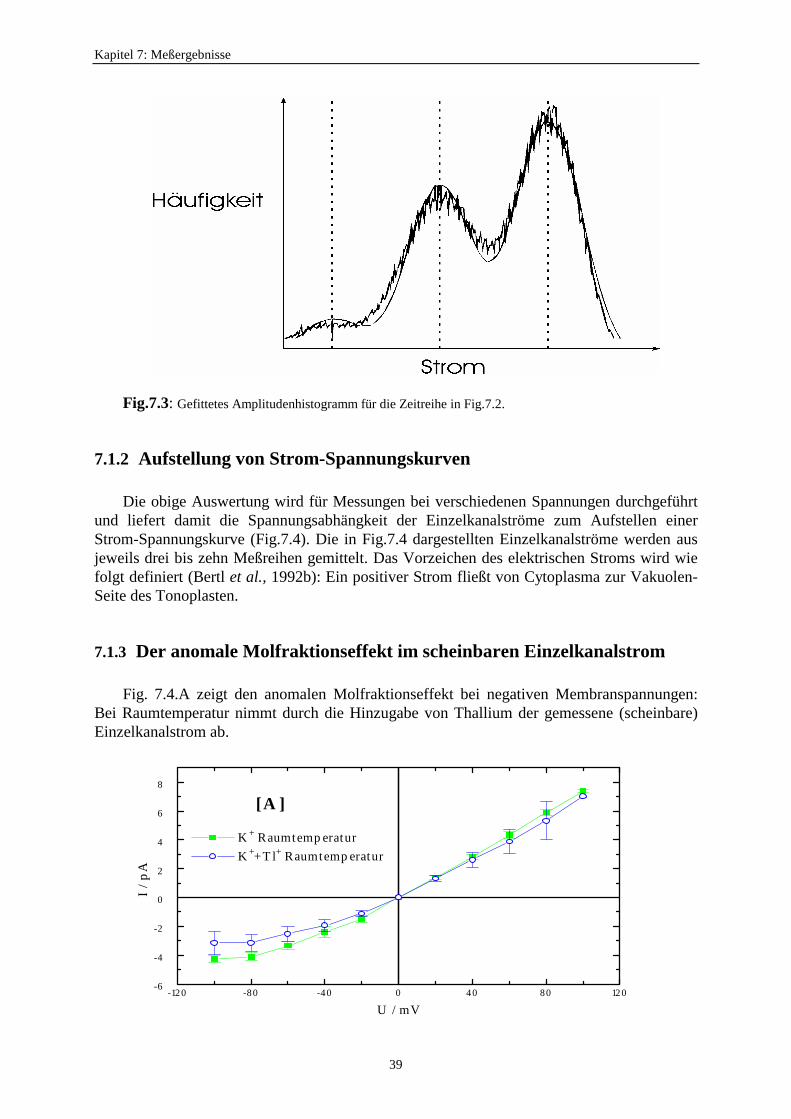
Kapitel 7: Meßergebnisse
39
Fig.7.3: Gefittetes Amplitudenhistogramm für die Zeitreihe in Fig.7.2.
7.1.2 Aufstellung von Strom-Spannungskurven
Die obige Auswertung wird für Messungen bei verschiedenen Spannungen durchgeführtund liefert damit die Spannungsabhängkeit der Einzelkanalströme zum Aufstellen einerStrom-Spannungskurve (Fig.7.4). Die in Fig.7.4 dargestellten Einzelkanalströme werden ausjeweils drei bis zehn Meßreihen gemittelt. Das Vorzeichen des elektrischen Stroms wird wiefolgt definiert (Bertl et al., 1992b): Ein positiver Strom fließt von Cytoplasma zur Vakuolen-Seite des Tonoplasten.
7.1.3 Der anomale Molfraktionseffekt im scheinbaren Einzelkanalstrom
Fig. 7.4.A zeigt den anomalen Molfraktionseffekt bei negativen Membranspannungen:Bei Raumtemperatur nimmt durch die Hinzugabe von Thallium der gemessene (scheinbare)Einzelkanalstrom ab.
-120 -80 -40 0 40 80 120-6
-4
-2
0
2
4
6
8
[A ]
K + Raumtemp eratur K ++T l+ Raumtemp eratur
I / p
A
U / mV
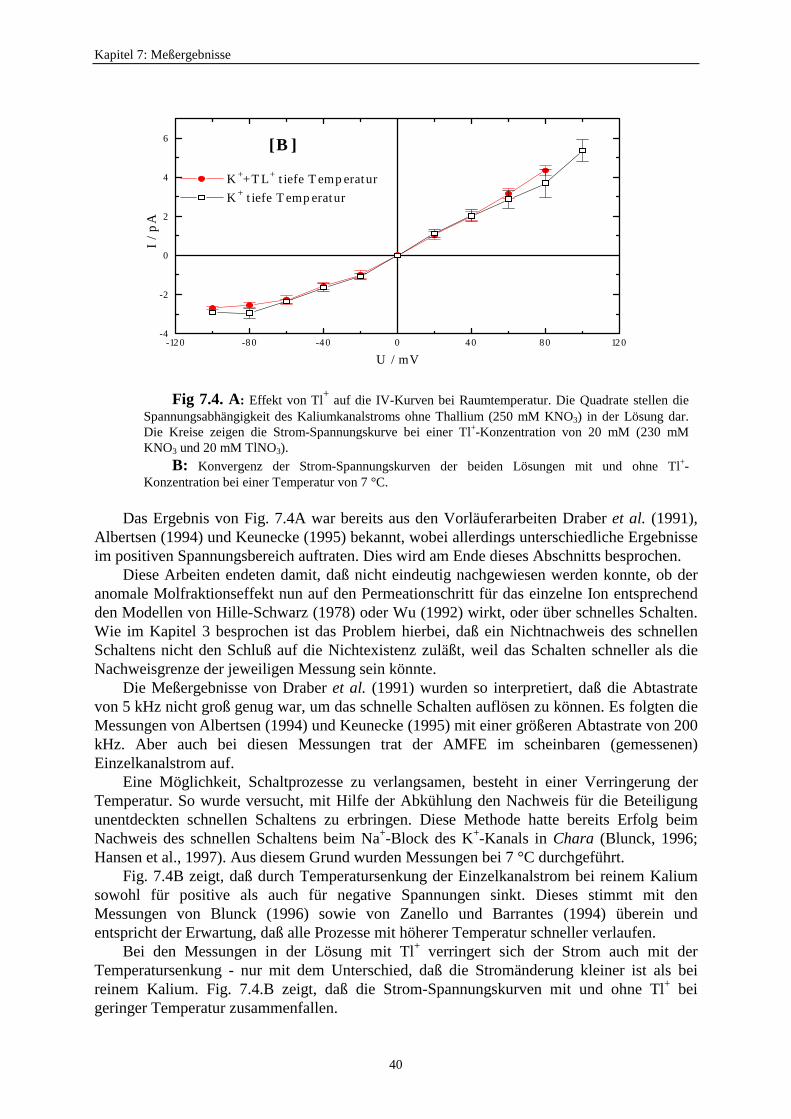
Kapitel 7: Meßergebnisse
40
Das Ergebnis von Fig. 7.4A war bereits aus den Vorläuferarbeiten Draber et al. (1991),Albertsen (1994) und Keunecke (1995) bekannt, wobei allerdings unterschiedliche Ergebnisseim positiven Spannungsbereich auftraten. Dies wird am Ende dieses Abschnitts besprochen.
Diese Arbeiten endeten damit, daß nicht eindeutig nachgewiesen werden konnte, ob deranomale Molfraktionseffekt nun auf den Permeationschritt für das einzelne Ion entsprechendden Modellen von Hille-Schwarz (1978) oder Wu (1992) wirkt, oder über schnelles Schalten.Wie im Kapitel 3 besprochen ist das Problem hierbei, daß ein Nichtnachweis des schnellenSchaltens nicht den Schluß auf die Nichtexistenz zuläßt, weil das Schalten schneller als dieNachweisgrenze der jeweiligen Messung sein könnte.
Die Meßergebnisse von Draber et al. (1991) wurden so interpretiert, daß die Abtastratevon 5 kHz nicht groß genug war, um das schnelle Schalten auflösen zu können. Es folgten dieMessungen von Albertsen (1994) und Keunecke (1995) mit einer größeren Abtastrate von 200kHz. Aber auch bei diesen Messungen trat der AMFE im scheinbaren (gemessenen)Einzelkanalstrom auf.
Eine Möglichkeit, Schaltprozesse zu verlangsamen, besteht in einer Verringerung derTemperatur. So wurde versucht, mit Hilfe der Abkühlung den Nachweis für die Beteiligungunentdeckten schnellen Schaltens zu erbringen. Diese Methode hatte bereits Erfolg beimNachweis des schnellen Schaltens beim Na+-Block des K+-Kanals in Chara (Blunck, 1996;Hansen et al., 1997). Aus diesem Grund wurden Messungen bei 7 °C durchgeführt.
Fig. 7.4B zeigt, daß durch Temperatursenkung der Einzelkanalstrom bei reinem Kaliumsowohl für positive als auch für negative Spannungen sinkt. Dieses stimmt mit denMessungen von Blunck (1996) sowie von Zanello und Barrantes (1994) überein undentspricht der Erwartung, daß alle Prozesse mit höherer Temperatur schneller verlaufen.
Bei den Messungen in der Lösung mit Tl+ verringert sich der Strom auch mit derTemperatursenkung - nur mit dem Unterschied, daß die Stromänderung kleiner ist als beireinem Kalium. Fig. 7.4.B zeigt, daß die Strom-Spannungskurven mit und ohne Tl+ beigeringer Temperatur zusammenfallen.
-120 -80 -40 0 40 80 120-4
-2
0
2
4
6 [B ]
K ++T L+ t iefe T emp eratur K + t iefe T emp eratur
I / p
A
U / mV
Fig 7.4. A: Effekt von Tl+ auf die IV-Kurven bei Raumtemperatur. Die Quadrate stellen dieSpannungsabhängigkeit des Kaliumkanalstroms ohne Thallium (250 mM KNO3) in der Lösung dar.Die Kreise zeigen die Strom-Spannungskurve bei einer Tl+-Konzentration von 20 mM (230 mMKNO3 und 20 mM TlNO3).
B: Konvergenz der Strom-Spannungskurven der beiden Lösungen mit und ohne Tl+-Konzentration bei einer Temperatur von 7 °C.
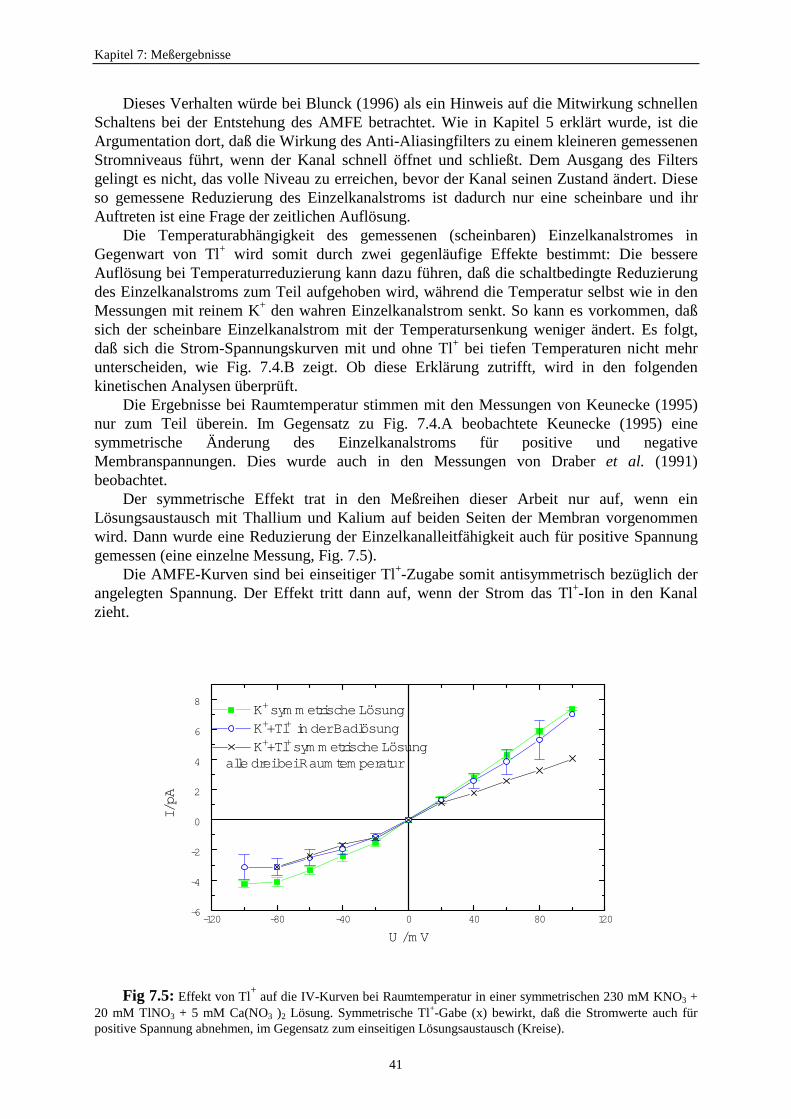
Kapitel 7: Meßergebnisse
41
Dieses Verhalten würde bei Blunck (1996) als ein Hinweis auf die Mitwirkung schnellenSchaltens bei der Entstehung des AMFE betrachtet. Wie in Kapitel 5 erklärt wurde, ist dieArgumentation dort, daß die Wirkung des Anti-Aliasingfilters zu einem kleineren gemessenenStromniveaus führt, wenn der Kanal schnell öffnet und schließt. Dem Ausgang des Filtersgelingt es nicht, das volle Niveau zu erreichen, bevor der Kanal seinen Zustand ändert. Dieseso gemessene Reduzierung des Einzelkanalstroms ist dadurch nur eine scheinbare und ihrAuftreten ist eine Frage der zeitlichen Auflösung.
Die Temperaturabhängigkeit des gemessenen (scheinbaren) Einzelkanalstromes inGegenwart von Tl+ wird somit durch zwei gegenläufige Effekte bestimmt: Die bessereAuflösung bei Temperaturreduzierung kann dazu führen, daß die schaltbedingte Reduzierungdes Einzelkanalstroms zum Teil aufgehoben wird, während die Temperatur selbst wie in denMessungen mit reinem K+ den wahren Einzelkanalstrom senkt. So kann es vorkommen, daßsich der scheinbare Einzelkanalstrom mit der Temperatursenkung weniger ändert. Es folgt,daß sich die Strom-Spannungskurven mit und ohne Tl+ bei tiefen Temperaturen nicht mehrunterscheiden, wie Fig. 7.4.B zeigt. Ob diese Erklärung zutrifft, wird in den folgendenkinetischen Analysen überprüft.
Die Ergebnisse bei Raumtemperatur stimmen mit den Messungen von Keunecke (1995)nur zum Teil überein. Im Gegensatz zu Fig. 7.4.A beobachtete Keunecke (1995) einesymmetrische Änderung des Einzelkanalstroms für positive und negativeMembranspannungen. Dies wurde auch in den Messungen von Draber et al. (1991)beobachtet.
Der symmetrische Effekt trat in den Meßreihen dieser Arbeit nur auf, wenn einLösungsaustausch mit Thallium und Kalium auf beiden Seiten der Membran vorgenommenwird. Dann wurde eine Reduzierung der Einzelkanalleitfähigkeit auch für positive Spannunggemessen (eine einzelne Messung, Fig. 7.5).
Die AMFE-Kurven sind bei einseitiger Tl+-Zugabe somit antisymmetrisch bezüglich derangelegten Spannung. Der Effekt tritt dann auf, wenn der Strom das Tl+-Ion in den Kanalzieht.
-120 -80 -40 0 40 80 120-6
-4
-2
0
2
4
6
8 K+ sym m etrische Lösung K++Tl+ in der Badlösung K++Tl+ sym m etrische Lösung
alle drei bei Raum tem peratur
I / pA
U / m V
Fig 7.5: Effekt von Tl+ auf die IV-Kurven bei Raumtemperatur in einer symmetrischen 230 mM KNO3 +20 mM TlNO3 + 5 mM Ca(NO3 )2 Lösung. Symmetrische Tl+-Gabe (x) bewirkt, daß die Stromwerte auch fürpositive Spannung abnehmen, im Gegensatz zum einseitigen Lösungsaustausch (Kreise).

Kapitel 7: Meßergebnisse
42
7.2 Motivation zur Untersuchung des Zusammenhangs zwischenschnellem Schalten und scheinbarem Einzelkanalstrom
Fig. 7.4B enthält einen Hinweis, daß normalerweise (bei Raumtemperatur) schnellesSchalten Ursache für die scheinbare Reduktion des Einzelkanalstromes ist. Im folgenden solluntersucht werden, ob diese Aussage durch eine Analyse der schnellen Übergangsraten deszum Kanal gehörenden Markovmodells bestätigt werden kann.
Im Falle des Cs+-Blockes haben Draber und Hansen (1994) gezeigt, daß die Reduktionder scheinbaren Einzelkanalströme, die bei 5 kHz-Abtastrrate gemessen wurden, einenMittelungseffekt im Anti-Aliasing Filter über schnelles Schalten zurückgeführt werden konnte(Fig. 5.2, rechte Seite).
Bevor die Frage aufgegriffen wird, ob hier im Falle des AMFEs ein ähnlicher Effektvorliegt, soll der Zusammenhang zwischen Übergangsraten und scheinbaremEinzelkanalstrom anhand eines einfachen Markovmodells O<=>C (Abschnitt 4.1) und eineseinzelnen Kanals dargestellt werden. Dies ermöglicht eine bessere Veranschaulichung derwesentlichen Mechanismen als die später folgende Analyse für das erheblich kompliziertereModell des vorliegenden Kanals.
Sei pj( ∞ ) die Wahrscheinlichkeit, daß sich der Kanal im Zustand j im Steady-Stateaufhält, und kij sei die Übergangsrate zwischen dem i-ten und j-ten Zustand. Als Steady-Statewird der Zustand bezeichnet, in dem der Kanal sich in einem stationären Zustand befindet.Das bedeutet, daß der Kanalstrom im zeitlichen Mittel konstant ist. Das Prinzip derMikroreversibilität besagt, daß jeder Reaktionsschritt im zeitlichen Mittel in beide Richtungengleich häufig durchlaufen wird (Hinfluß = Rückfluß). So gilt für das Modell O<=>C imSteady-State die folgende Gleichung:
COCOCO kpkp )()( ∞=∞ (7.1)
OCCO
CO
OC kkk
pp +=
∞+∞∞⇒
)()()(pO (7.2)
Der scheinbare Einzelkanalstrom ergibt sich durch den Mittelungsprozeß des Meßfiltersüber die schnell umschaltenden Offen- und Geschlossenzeiten (Zustand O und C) des Kanals.Damit ist der Strom proportional zur relativen Offenwahrscheinlichkeit:
)()()(pO
∞+∞∞∝⇒
OC ppi (7.3)
Die Gleichungen (7.2) und (7.3) liefern die Beziehung zwischen den Übergangsraten unddem mittleren Strom. Sie lautet:
OCCO
CO
kkki+
∝⇒ (7.4)
Formel (7.4) stellt einen direkten Zusammenhang zwischen den Übergangsraten und demStromniveau bei dem Zustandsmodell O<=>C und einem Einzelkanal im Falle unentdecktenschnellen Schaltens her.

Kapitel 7: Meßergebnisse
43
Reale Messungen haben aber mehr als einen Kanal und meistens mehr Zustände als dasO<=>C Modell. Dies gestaltet die kinetische Untersuchung eines realen Kanals sehrschwierig.
Um zu überprüfen, ob der Mechanismus von Draber und Hansen (1994) auch hier zutrifft,wird zuerst versucht, mit Hilfe der Dwell-Time-Histogramme (Abschnitt 7.3) das richtigeMarkov-Modell zu finden. Prinzipiell ist dieses Problem nicht eindeutig (Kienker, 1989).Allerdings genügt es für die Fragestellung hier, eine korrekte kinetische Beschreibung zugewinnen. Es ist für die Berechnung des mittleren Stromes nicht notwendig, das "wahre"Modell aus einer Klasse kinetisch äquivalenter Modelle zu bestimmen.
Die erste Information über die Anzahl der Zustände des Modells gewinnt man aus derAnzahl der Zeitkonstanten, die notwendig für den Fit der Dwell-Time-Histogramme sind. Dieentsprechenden Diagramme ergeben so die Anzahl der Offen- und Geschlossenzustände.
Danach werden die Ratenkonstanten ermittelt und die in verschiedenen Badlösungenerhaltenen Werte miteinander verglichen. Daraus gewinnt man die in obigen Gleichungenwichtigen Besetzungswahrscheinlichkeiten jedes einzelnen Zustands. Ausgehend von derBetrachtung der Übergangsraten und Besetzungswahrscheinlichkeiten wird ein Teilsystem desMarkov-Modells ausgewählt, das verantwortlich für das schnelle Schalten ist. Für diesesTeilmodell werden wieder die Besetzungswahrscheinlichkeiten bestimmt. Mit Hilfe einerähnlichen Beziehung kann dann wie in Formel 7.3 der scheinbare (gemittelte)Einzelkanalstrom abgeschätzt werden. Mit dieser Abschätzung wird versucht, die Hypothesedes schnellen Schaltens für die scheinbaren Stromreduzierungen zu untermauern.
7.3 Verweildauerhistogramme
7.3.1 Analyseprogramme
Im obigen Abschnitt wurde der Weg skizziert, um aus der kinetischen Analyse dieHypothese der Einzelkanalstromreduzierung durch schnelles Schalten zu verifizieren. Hierzumuß das Zeitverhalten des Kanals mathematisch beschrieben werden. Im Abschnitt 4.1 sinddie für diesen Zweck benutzten Markov-Prozesse dargestellt.
Das Schaltverhalten wird mit Hilfe der Ratenkonstanten der Übergänge zwischen denZuständen des Kanals direkt beschrieben. Im Rahmen dieser Arbeit interessiert die Änderungder Ratenkonstanten bei den unterschiedlichen Lösungen und Temperaturen.
Es gibt zwei Möglichkeiten, die Ratenkonstanten zu bestimmen. Die erste Methode istder direkte Fit der Zeitreihe mit einem Hidden-Markov-Modell (HMM-Fit). Das Attribut„Hidden“ bekommt ein Markov-Modell, wenn mehrere kinetisch verschiedene Zustände zurgleichen elektrischen Leitfähigkeit führen (mehrere O- oder C-Zustände) und/oder dasAusgangssignal durch Rauschen gestört ist. Beides trifft hier zu.
Beim direkten Zeitreihenfit wird ausgehend vom Zustand des Kanals zum Zeitpunkt nTdie Wahrscheinlichkeit des Zustandes zum Zeitpunkt (n+1)T mit einem 1-SchrittPrädiktionsalgorithmus geschätzt und mit dem gemessenen Strom verglichen (Horn undLange 1983; Fredkin und Rice, 1992; Albertsen und Hansen, 1994; Klein et al., 1997). DieRatenkonstanten des verwendeten Modells werden so lange mit einem Simplex Algorithmus(Nedler und Mead, 1965; Caceci und Cacheris, 1984) variiert, bis das Maximum derLikelihood für die gemessene Zeitreihe erreicht ist.
Der Nachteil dieser Analyse ist wie bei allen Verfahren, daß das Modell bekannt seinmuß, so daß verschiedene Modelle sequentiell getestet werden müssen, um das mit dermaximalen Maximum-Likelihood auszuwählen. Daraus ergibt sich das Hauptproblem: der

Kapitel 7: Meßergebnisse
44
Zeitbedarf, der zwischen 30 min bis 100h pro 10-s Meßreihe liegen kann (Albertsen, 1994;Albertsen und Hansen, 1994). Der Vorteil ist, daß zur Berechnung der Maximum-Likelihood-Funktion die Originalstromwerte aus der gemessenen Zeitreihe entnommen werden. Dadurchkönnen die schnellen Ratenkonstanten genauer bestimmt werden, weil die Fehler einesSprungdetektors (Hinkley-Detektor, Abschnitt 6.9.2) umgangen werden. Dies löst dasProblem der missed-events (Draber und Schulze, 1994) weit besser. Doch wegen des hohenZeitaufwands beim Fitten wurde dieses Analyseverfahren nur zu Kontrollzweckenangewendet.
Der Vorteil der Vermeidung eines Sprungdetektors geht bei der zweiten Methode, derAuswertung von Dwell-Time-Histogrammen mit dem Target-Fit (Kijima und Kijima, 1987b;Blunck, 1996; Kirst, 1997), verloren. Die Dwell-Time-Histogramme geben dieWahrscheinlichkeitsverteilungen der Aufenthaltszeiten in den verschiedenen Stromniveauswieder. Um diese Dwell-Time-Histogramme zu erhalten, muß die gemessene Zeitreihe einenSprungdetektor (Hinkley-Detektor achter Ordnung, Abschnitt 6.9.2, Schultze und Draber,1993) durchlaufen. Der Sprungdetektor stellt die rauschfreie, rekonstruierte Zeitreihe her. EinBuchhalterprogramm des Hinkley-Detektors stellt die Anzahl der gefundenen Verweildauernpro Stromniveau zu Histogrammen zusammen.
Das Dwell-Time-Histogramm gibt an, wie häufig die Verweilzeit zwischen t und t+ t∆auftrat. Die Dwell-Time-Histogramme, die der Sprungdetektor ausgibt, sind in 500 “bins“(Abszissenbereiche t∆ ) eingeteilt. Die Größe eines Bins ist 40 µs. Um die statistischenAbweichungen im Bereich der längeren Meßereignisse zu reduzieren, werden die Daten inexponentiell wachsenden Zeitintervallen gemittelt. Die Anzahl der Bins reduziert sich auf 50.
Die Bin-Weite, die der Hinkley-Detektor benutzt, um die Dewll-Time-Histogramme zuerstellen, war wie oben erwähnt 40 µs. Dies ist aber nicht der kleinste Abszissenwert imHistogramm. Nach Colquhoun et al. (1996) bewirken die missed-events, daß die Dwell-Time-Histogramme mit einer Funktion tn exp(-t / τ) gefittet werden müssen. Dadurch erhält maneinen ansteigenden Teil in den Dwell-Time-Histogrammen. Colqhoun et al. (1996) habengezeigt, daß man das τ gut bestimmen kann, wenn man die ersten 2-3 Werte am ansteigendenAst fortläßt. Deshalb beginnen die Dwell-Time-Histogramme bei 80 oder 120 µs. Dies gibteinen groben Anhaltspunkt für den Bereich, in dem die Zeitkonstanten noch einigermaßen gutbestimmt werden können.
Der Sprungdetektor ist das kritische Element in dieser Analyse. Allerdings steht mit demvon Schultze und Draber (1993) entwickelten Hinkley-Detektor höherer Ordnung imProgramm „day“ der zur Zeit beste Detektor zur Verfügung. Sein Vorteil ist die Anpassungder Detektorschwelle an das Rauschen. Hierzu bestimmt man durch den bereits erwähnten Fit(Abschnitt 7.1.1) der Amplitudenhistogramme die Streuung der Stromwerte pro Niveau. DieAnalyse der Histogramme wird ungenau, wenn nicht genügend Daten zur Verfügung stehen.
Einige der Meßreihen werden zum Schluß mit dem direkten Zeitreihen-Fit (HMM-Fit)analysiert. Die Übergangsratenkonstanten werden somit mit zwei unterschiedlichen Methodenbestimmt. Zur Analyse der großen Datenmengen kommt eine relativ schnelle, aber nicht sehrgenaue Methode zum Einsatz, und dann eine langsame, aber genauere Methode für bestimmteausgewählte Daten. Die langsame Methode liefert damit eine Abschätzung für dieGenauigkeit der mit der schnellen Methode bestimmten Ratenkonstanten.

Kapitel 7: Meßergebnisse
45
7.3.2 Auswahl eines geeigneten Markov-Modells für den K+-Kanal inChara
Beide oben beschriebenen Auswertungsverfahren richten sich direkt auf dieÜbergangsraten kij im Markov-Modell des Kanals. Die wichtigste Frage ist dabei die Auswahldes „richtigen“ Modells. Um zu sehen, welche vorgegebenen Markov-Modelle mit dengemessenen Daten übereinstimmen, wird der Menüpunkt „5-state-model“ imAuswerteprogramm „day“ (Kirst, 1996) aufgerufen. Dieses ermöglicht die Wahl vonModellen mit maximal 5 Zuständen und den anschließenden Fit der Dwell-Time-Histogramme auf der Basis dieses Modells. Folgende Modelle wurden auf ihre Eignunguntersucht (Figur. 7.6):
C1 ⇔ O ⇔ C2
O ⇔ C1 ⇔ C2 ⇔ C3
O1 ⇔ O2 ⇔ C1 ⇔ C2 ⇔ C3
O1 ⇔ O2 ⇔ C1 ⇔ C2
O1 ⇔ C1 ⇔ C2 ⇔ C3
!O2
C1 ⇔ O1 ⇔ C2 ⇔ O2 ⇔ C3
Fig. 7.6: Mögliche Modelle für den Kalium-Kanal in Chara corallina. O bezeichnet einen Zustand, beidem der Ionenkanal offen ist. C sind die Geschlossenzustände. Die Pfeile repräsentieren die Übergänge zwischenzwei Zuständen. Jedem Übergang wird eine Übergangsrate für die Hin- und Rückreaktion zugeordnet.
Das Modell, welches die experimentellen Daten am besten annäherte und somit genauerbetrachtet wurde, ist das Modell O<=>O<=>C<=>C<=>C. Dieses Ergebnis ergab sich aus denfolgenden Untersuchungen:
Die Modelltestung geschah sowohl an K+-Daten als auch an K+/Tl+-Daten. Hierfürwurden aus den Zeitreihen nach Fig. 7.1 die Dwell-Time-Histogramme erstellt. Diesgeschieht, wie in Abschnitt 7.3.2 beschrieben, dadurch, daß mit einem Sprungdetektor (sieheAbschnitt 6.9.2) die Verweilzeiten in den einzelnen Stromniveaus bestimmt und von einemBuchhalterprogramm in Dwell-Time-Histogrammen zusammengestellt werden.
Blunck (1996) modifizierte die Theorie der Auswertung von Dwell-Time-Histogrammenvon Kijima und Kijima (1987a,b) derart, daß eine Mehrkanalanalyse einfach wurde. Hierbeiwerden die Dwell-Time-Histogramme mit Summen von e-Funktionen gefittet, wie es aus derTheorie der Markov-Prozesse folgt (Abschnitt 4.1).
∑ −=i
ii taty )/exp()( τ (7.5)

Kapitel 7: Meßergebnisse
46
Während in der Literatur allgemein die Zeitkonstanten iτ und die Amplitudenfaktoren ai dasEnde der Analyse darstellen, geht der Target-Fit weiter. Das Ziel der Analysen sind dieRatenkonstanten kij. Deshalb läuft das Fitprogramm so ab, daß für ein festes Modell die kijunter der Lenkung eines Simplexalgorithmus (Nedler und Mead, 1965; Caceci und Cacheris,1984) geraten und ai und iτ daraus berechnet werden. Die ai und iτ werden im Fitprogrammmit den folgenden Gleichungen aus den Ratenkonstanten berechnet.
( ) ( ) )(),(),()( . mTimTimmi Kvv =τ (7.6.a)
∑ ∑ ∑∈ ∈ ∉
∞=m m mLr Lr Ls
mtrrs
imr
mrm
mi fvRSa )(2),()()( /)(.)(2 κ (7.6.b)
∑ ∑∈ ∉
∞=m mLr LS
srrsm
rm
tr Rf κκ.)(2 )()((7.6.c)
m Die Anzahl der offenen Kanäle.Rr Aufenthaltswahrscheinlichkeiten für den Makrokanal. Ein Makrokanal ist ein
Ensemble von Einzelkanälen, die dann wie ein einziger Kanal mit vielenZuständen behandelt wird.
Rrm( ) Zustände Rr von einem Zustand Ym .
Ym Alle kinetisch unterschiedlichen Zustände Rr mit derselben Leitfähigkeit(gleiche Anzahl von offenen Kanälen) werden in einem Zustand Ymzusammengefaßt.
Lm Die Menge der Indizes r, die zum Zustand Ym gehören.Rr( ∞ ) Besetzungswahrscheinlichkeit im stationären Zustand.
rsκ Ratenkonstanten des Makrokanals. Sie stehen mit den Ratenkonstanten desEinzelkanals wie folgt im Beziehung:
ijr
irs kb .)(=κ (7.7.a)
∑∑∑=
≠=
≠=
==−Q
i
Q
ijj
ijr
i
B
rss
rsrr kb1 1
)(
1.κκ (7.7.b)
K(m) Die Matrix der Ratenkonstanten des Makrokanals.v(i) v(i) sind die Eigenvektoren zur reduzierten Ratenmatrix des absorbierenden
Makrokanalmodells zum Stromniveau m (Gleichung 7.6.a). Der hochgestellteIndex (i) numeriert die verschiedenen Eigenvektoren durch.
Sm Normierungsfaktor.f tr
m( ) Die mittlere Übergangswahrscheinlichkeit für den Makrokanal. Sie wird ausder Gleichung 7.6.c erhalten.
b(r) = ( , ... , , , ... , )( ) ( ) ( ) ( )b b b brL
rL
rQ
r1 1+ Vektor der Besetzungszahlen für die Zustände im
Makrozustand mit b r1( ) , ... , bL
r( ) Offenzustände, bLr+1
( ), ... , bQ
r( )
Geschlossenzustände.

Kapitel 7: Meßergebnisse
47
Die Eigenwertgleichung 7.6.a wurde aus einer Differentialgleichung für einenMakrokanal (eine ähnliche Formel wie die Gleichung 4.1) erhalten.
Der Target-Fit hat mehrere Vorteile gegenüber dem allgemeinen Zeitkonstantenfit. DieBerechnung der Ratenkonstanten aus den Zeitkonstanten und Amplitudenfaktoren ist sehrkompliziert und mitunter ungenau, auch wenn neuerdings Jackson (1997) einen neuenAlgorithmus dafür vorgeschlagen hat. Noch wichtiger ist, daß die Dwell-Time-Verteilung füralle Niveaus in einer Mehrkanalaufzeichnung gleichzeitig gefittet werden müssen. Besonderswichtig für die Genauigkeit ist dies, wenn der vermutete Effekt des schnellen Schaltens durcheine einzelne Ratenkonstante verursacht wird.
Die zugehörigen Fitroutinen sind im Programm „Day“ eingebaut (Kirst, 1997).
7.3.3 Der Vergleich von C<=>O<=>C- und O<=>O<=>C<=>C<=>C-Modell
Das erste Modell hat historische Gründe. Eigentlich könnte das C<=>O<=>C-Modell vonvornherein ausgeschlossen werden. Doch das Modell C<=>O<=>C wurde von Blunck (1996)zum Nachweis des schnellen Schaltens beim Na+-Block des K+-Kanals in Chara gewählt.
Hierbei wurde von nur einem offenen Zustand ausgegangen. Die Verweilzeit in ihm wirddurch Übergänge in zwei geschlossene Zustände unterbrochen. Nach den Ergebnissen vonBlunck (1996) ist der Kanal in dem einen Zustand über einen längeren Zeitraum geschlossen,während die Geschlossen-Erreignisse des zweiten Zustandes durch sehr kurze Öffnungenunterbrochen werden.Der Ausschluß dieses Modells ergibt sich aus der Regel, daß die Anzahl der Offenzuständegleich der Anzahl der τ ’s in dem Offen-Dwell-Time-Histogramm und die Anzahl derGeschlossen-Zustände gleich der τ ’s in dem Geschlossen-Dwell-Time-Histogramm ist. Dabei dem Offen-Dwell-Time-Histogramm der Fit der Exponentialfunktion zwei Zeitkonstantenbenötigt, sollte ein Modell mit zwei Geschlossenzuständen ausgewählt werden.
Die Folge der zu geringen Zustandszahl ist, daß die theoretischen Kurven nichtausreichend an die Meßdaten angepaßt werden konnten (Fig 7.7A-B-C). Die Ergebnisseweichen besonders bei Fits mit einem Offenzustand und Fits mit zwei Offenzuständen sehrstark von den Meßdaten ab.
Bei dem O<=>O<=>C<=>C<=>C-Modell sind die Zustände ebenfalls in einer Reihe. DerUnterschied ist, daß es insgesamt fünf statt der vorherigen drei Zustände gibt (zwei offene unddrei geschlossene Zustände) und die Übergänge auch innerhalb der offenen und dergeschlossenen Zustände liegen können. Der Übergang zwischen geschlossenem und offenemZustand ist sehr schnell, was eine Grundvoraussetzung für das schnelle Schalten ist (dieRatenkonstanten werden im Abschnitt 7.3.5 noch genauer betrachtet).
Das entscheidende Argument ist aber, daß die theoretische Kurve am besten an dieMeßdaten angepaßt werden konnte (Fig 7.8A-B-C). Es wird beobachtet, daß die Meßdatenvon der Fitfunktion eine deutlich geringere Abweichung haben als bei dem C<=>O<=>C-Modell. Deshalb kommt das Modell C<=>O<=>C für die weitere Untersuchung derRatenkonstanten nicht mehr in Frage.
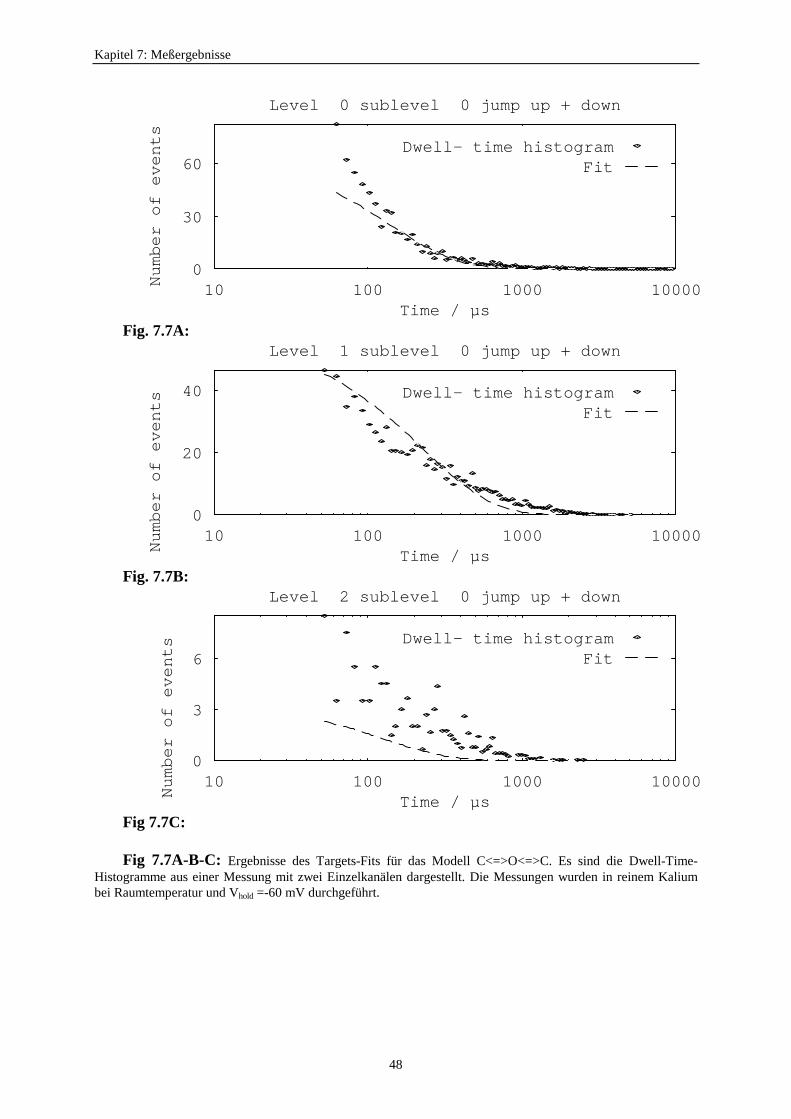
Kapitel 7: Meßergebnisse
48
0
30
60
10 100 1000 10000Number of events
Time / µs
Level 0 sublevel 0 jump up + down
Dwell- time histogramFit
Fig. 7.7A:
0
20
40
10 100 1000 10000
Number of events
Time / µs
Level 1 sublevel 0 jump up + down
Dwell- time histogramFit
Fig. 7.7B:
0
3
6
10 100 1000 10000
Number of events
Time / µs
Level 2 sublevel 0 jump up + down
Dwell- time histogramFit
Fig 7.7C:
Fig 7.7A-B-C: Ergebnisse des Targets-Fits für das Modell C<=>O<=>C. Es sind die Dwell-Time-Histogramme aus einer Messung mit zwei Einzelkanälen dargestellt. Die Messungen wurden in reinem Kaliumbei Raumtemperatur und Vhold =-60 mV durchgeführt.
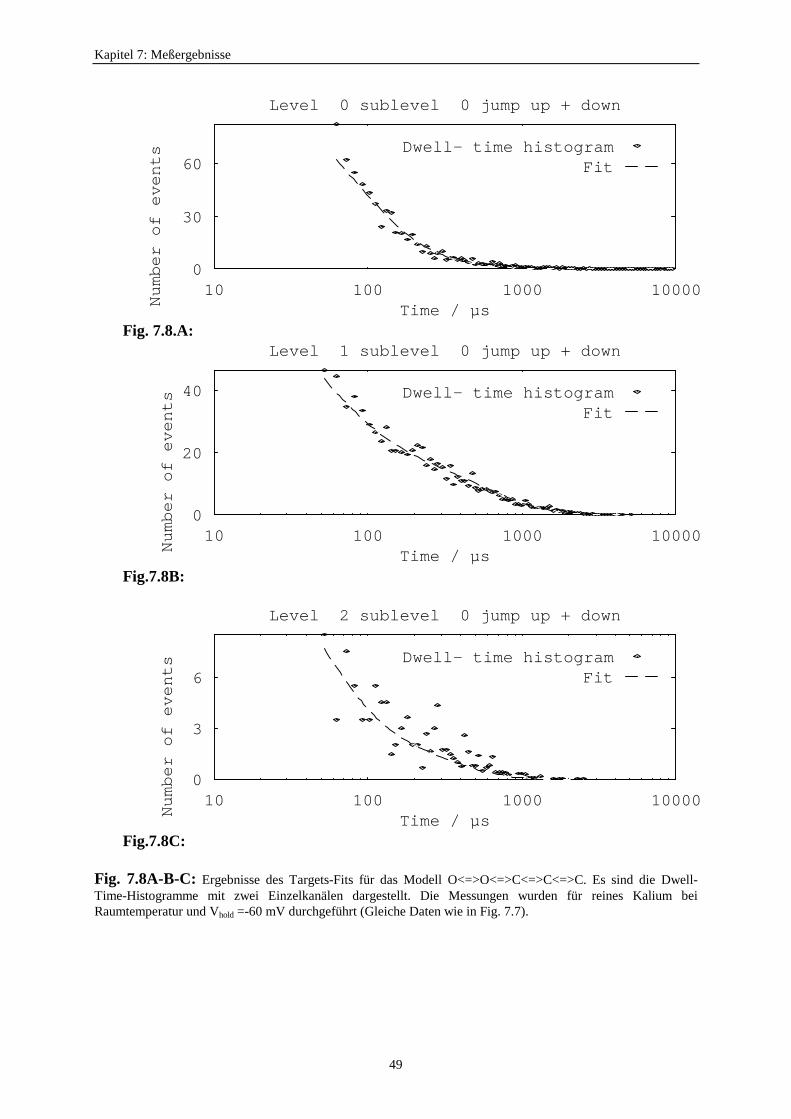
Kapitel 7: Meßergebnisse
49
0
30
60
10 100 1000 10000
Number of events
Time / µs
Level 0 sublevel 0 jump up + down
Dwell- time histogramFit
Fig. 7.8.A:
0
20
40
10 100 1000 10000
Number of events
Time / µs
Level 1 sublevel 0 jump up + down
Dwell- time histogramFit
Fig.7.8B:
0
3
6
10 100 1000 10000
Number of events
Time / µs
Level 2 sublevel 0 jump up + down
Dwell- time histogramFit
Fig.7.8C:
Fig. 7.8A-B-C: Ergebnisse des Targets-Fits für das Modell O<=>O<=>C<=>C<=>C. Es sind die Dwell-Time-Histogramme mit zwei Einzelkanälen dargestellt. Die Messungen wurden für reines Kalium beiRaumtemperatur und Vhold =-60 mV durchgeführt (Gleiche Daten wie in Fig. 7.7).
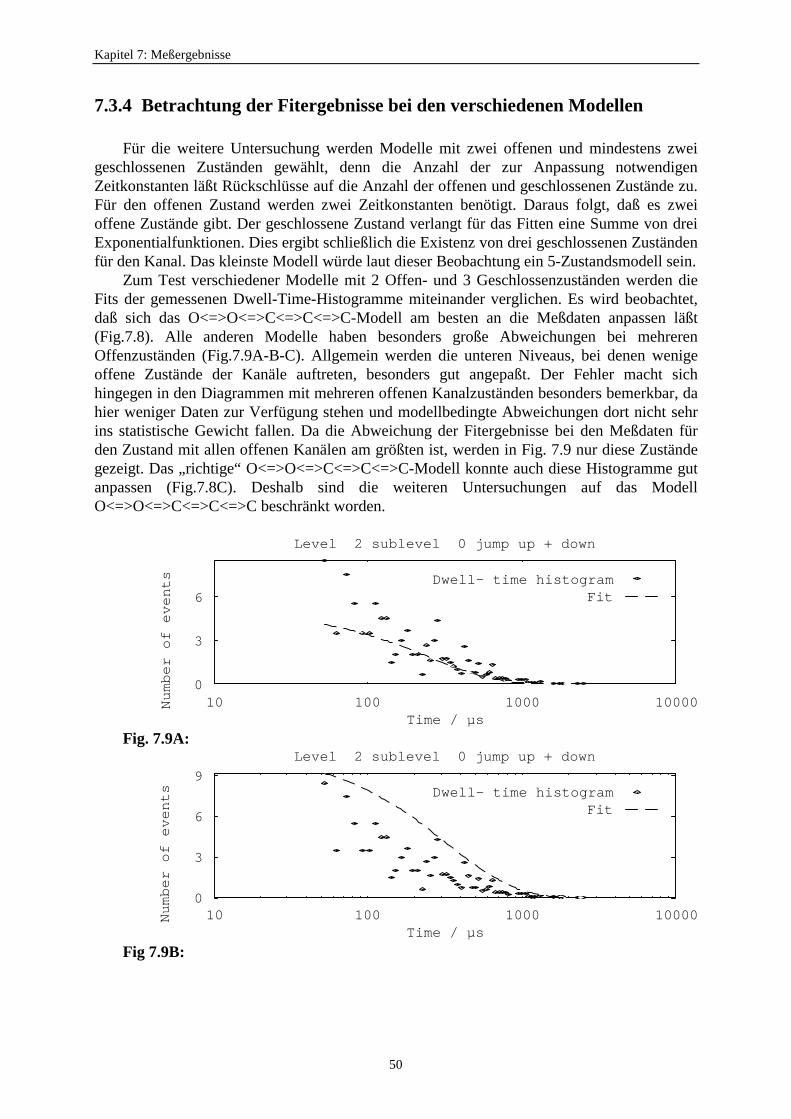
Kapitel 7: Meßergebnisse
50
7.3.4 Betrachtung der Fitergebnisse bei den verschiedenen Modellen
Für die weitere Untersuchung werden Modelle mit zwei offenen und mindestens zweigeschlossenen Zuständen gewählt, denn die Anzahl der zur Anpassung notwendigenZeitkonstanten läßt Rückschlüsse auf die Anzahl der offenen und geschlossenen Zustände zu.Für den offenen Zustand werden zwei Zeitkonstanten benötigt. Daraus folgt, daß es zweioffene Zustände gibt. Der geschlossene Zustand verlangt für das Fitten eine Summe von dreiExponentialfunktionen. Dies ergibt schließlich die Existenz von drei geschlossenen Zuständenfür den Kanal. Das kleinste Modell würde laut dieser Beobachtung ein 5-Zustandsmodell sein.
Zum Test verschiedener Modelle mit 2 Offen- und 3 Geschlossenzuständen werden dieFits der gemessenen Dwell-Time-Histogramme miteinander verglichen. Es wird beobachtet,daß sich das O<=>O<=>C<=>C<=>C-Modell am besten an die Meßdaten anpassen läßt(Fig.7.8). Alle anderen Modelle haben besonders große Abweichungen bei mehrerenOffenzuständen (Fig.7.9A-B-C). Allgemein werden die unteren Niveaus, bei denen wenigeoffene Zustände der Kanäle auftreten, besonders gut angepaßt. Der Fehler macht sichhingegen in den Diagrammen mit mehreren offenen Kanalzuständen besonders bemerkbar, dahier weniger Daten zur Verfügung stehen und modellbedingte Abweichungen dort nicht sehrins statistische Gewicht fallen. Da die Abweichung der Fitergebnisse bei den Meßdaten fürden Zustand mit allen offenen Kanälen am größten ist, werden in Fig. 7.9 nur diese Zuständegezeigt. Das „richtige“ O<=>O<=>C<=>C<=>C-Modell konnte auch diese Histogramme gutanpassen (Fig.7.8C). Deshalb sind die weiteren Untersuchungen auf das ModellO<=>O<=>C<=>C<=>C beschränkt worden.
0
3
6
10 100 1000 10000Number of events
Time / µs
Level 2 sublevel 0 jump up + down
Dwell- time histogramFit
Fig. 7.9A:
0
3
6
9
10 100 1000 10000Number of events
Time / µs
Level 2 sublevel 0 jump up + down
Dwell- time histogramFit
Fig 7.9B:
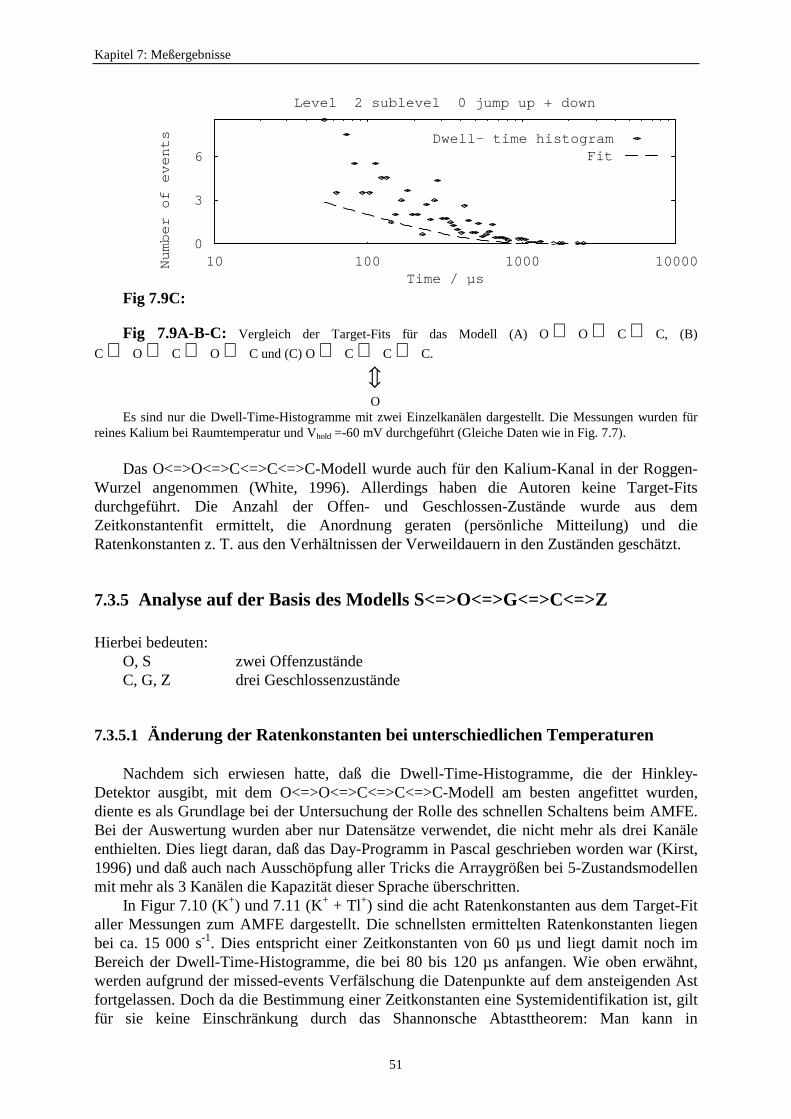
Kapitel 7: Meßergebnisse
51
0
3
6
10 100 1000 10000Number of events
Time / µs
Level 2 sublevel 0 jump up + down
Dwell- time histogramFit
Fig 7.9C:
Fig 7.9A-B-C: Vergleich der Target-Fits für das Modell (A) O ⇔ O ⇔ C ⇔ C, (B)C ⇔ O ⇔ C ⇔ O ⇔ C und (C) O ⇔ C ⇔ C ⇔ C.
! O
Es sind nur die Dwell-Time-Histogramme mit zwei Einzelkanälen dargestellt. Die Messungen wurden fürreines Kalium bei Raumtemperatur und Vhold =-60 mV durchgeführt (Gleiche Daten wie in Fig. 7.7).
Das O<=>O<=>C<=>C<=>C-Modell wurde auch für den Kalium-Kanal in der Roggen-Wurzel angenommen (White, 1996). Allerdings haben die Autoren keine Target-Fitsdurchgeführt. Die Anzahl der Offen- und Geschlossen-Zustände wurde aus demZeitkonstantenfit ermittelt, die Anordnung geraten (persönliche Mitteilung) und dieRatenkonstanten z. T. aus den Verhältnissen der Verweildauern in den Zuständen geschätzt.
7.3.5 Analyse auf der Basis des Modells S<=>O<=>G<=>C<=>Z
Hierbei bedeuten:O, S zwei OffenzuständeC, G, Z drei Geschlossenzustände
7.3.5.1 Änderung der Ratenkonstanten bei unterschiedlichen Temperaturen
Nachdem sich erwiesen hatte, daß die Dwell-Time-Histogramme, die der Hinkley-Detektor ausgibt, mit dem O<=>O<=>C<=>C<=>C-Modell am besten angefittet wurden,diente es als Grundlage bei der Untersuchung der Rolle des schnellen Schaltens beim AMFE.Bei der Auswertung wurden aber nur Datensätze verwendet, die nicht mehr als drei Kanäleenthielten. Dies liegt daran, daß das Day-Programm in Pascal geschrieben worden war (Kirst,1996) und daß auch nach Ausschöpfung aller Tricks die Arraygrößen bei 5-Zustandsmodellenmit mehr als 3 Kanälen die Kapazität dieser Sprache überschritten.
In Figur 7.10 (K+) und 7.11 (K+ + Tl+) sind die acht Ratenkonstanten aus dem Target-Fitaller Messungen zum AMFE dargestellt. Die schnellsten ermittelten Ratenkonstanten liegenbei ca. 15 000 s-1. Dies entspricht einer Zeitkonstanten von 60 µs und liegt damit noch imBereich der Dwell-Time-Histogramme, die bei 80 bis 120 µs anfangen. Wie oben erwähnt,werden aufgrund der missed-events Verfälschung die Datenpunkte auf dem ansteigenden Astfortgelassen. Doch da die Bestimmung einer Zeitkonstanten eine Systemidentifikation ist, giltfür sie keine Einschränkung durch das Shannonsche Abtasttheorem: Man kann in
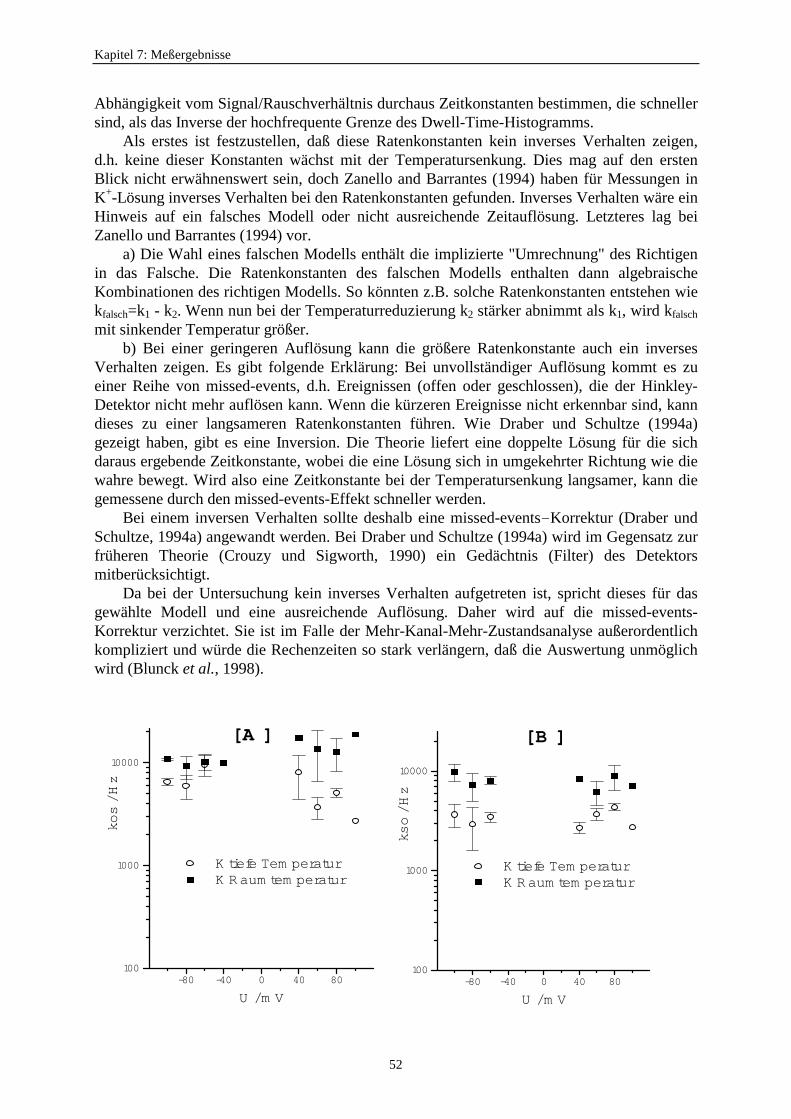
Kapitel 7: Meßergebnisse
52
Abhängigkeit vom Signal/Rauschverhältnis durchaus Zeitkonstanten bestimmen, die schnellersind, als das Inverse der hochfrequente Grenze des Dwell-Time-Histogramms.
Als erstes ist festzustellen, daß diese Ratenkonstanten kein inverses Verhalten zeigen,d.h. keine dieser Konstanten wächst mit der Temperatursenkung. Dies mag auf den erstenBlick nicht erwähnenswert sein, doch Zanello and Barrantes (1994) haben für Messungen inK+-Lösung inverses Verhalten bei den Ratenkonstanten gefunden. Inverses Verhalten wäre einHinweis auf ein falsches Modell oder nicht ausreichende Zeitauflösung. Letzteres lag beiZanello und Barrantes (1994) vor.
a) Die Wahl eines falschen Modells enthält die implizierte "Umrechnung" des Richtigenin das Falsche. Die Ratenkonstanten des falschen Modells enthalten dann algebraischeKombinationen des richtigen Modells. So könnten z.B. solche Ratenkonstanten entstehen wiekfalsch=k1 - k2. Wenn nun bei der Temperaturreduzierung k2 stärker abnimmt als k1, wird kfalschmit sinkender Temperatur größer.
b) Bei einer geringeren Auflösung kann die größere Ratenkonstante auch ein inversesVerhalten zeigen. Es gibt folgende Erklärung: Bei unvollständiger Auflösung kommt es zueiner Reihe von missed-events, d.h. Ereignissen (offen oder geschlossen), die der Hinkley-Detektor nicht mehr auflösen kann. Wenn die kürzeren Ereignisse nicht erkennbar sind, kanndieses zu einer langsameren Ratenkonstanten führen. Wie Draber und Schultze (1994a)gezeigt haben, gibt es eine Inversion. Die Theorie liefert eine doppelte Lösung für die sichdaraus ergebende Zeitkonstante, wobei die eine Lösung sich in umgekehrter Richtung wie diewahre bewegt. Wird also eine Zeitkonstante bei der Temperatursenkung langsamer, kann diegemessene durch den missed-events-Effekt schneller werden.
Bei einem inversen Verhalten sollte deshalb eine missed-events-Korrektur (Draber undSchultze, 1994a) angewandt werden. Bei Draber und Schultze (1994a) wird im Gegensatz zurfrüheren Theorie (Crouzy und Sigworth, 1990) ein Gedächtnis (Filter) des Detektorsmitberücksichtigt.
Da bei der Untersuchung kein inverses Verhalten aufgetreten ist, spricht dieses für dasgewählte Modell und eine ausreichende Auflösung. Daher wird auf die missed-events-Korrektur verzichtet. Sie ist im Falle der Mehr-Kanal-Mehr-Zustandsanalyse außerordentlichkompliziert und würde die Rechenzeiten so stark verlängern, daß die Auswertung unmöglichwird (Blunck et al., 1998).
-80 -40 0 40 80100
1000
10000
[ B ]
K tiefe Tem peratur K Raum tem peratur
kso / Hz
U / m V
-80 -40 0 40 80100
1000
10000
[ A ]
K tiefe Tem peratur K Raum tem peratur
kos / Hz
U / m V
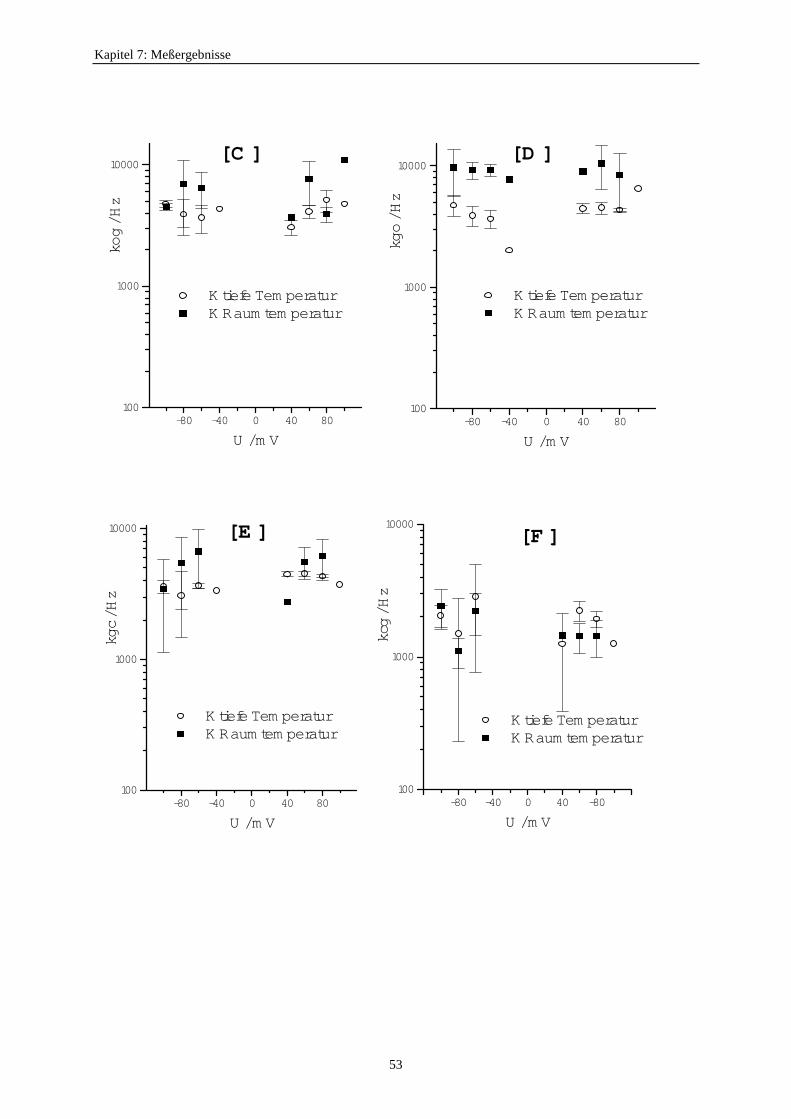
Kapitel 7: Meßergebnisse
53
-80 -40 0 40 80100
1000
10000[ C ]
K tiefe Tem peratur K Raum tem peratur
kog / Hz
U / m V-80 -40 0 40 80
100
1000
10000[ D ]
K tiefe Tem peratur K Raum tem peratur
kgo / Hz
U / m V
-80 -40 0 40 80100
1000
10000 [ E ]
K tiefe Tem peratur K Raum tem peratur
kgc / Hz
U / m V
-80 -40 0 40 -80100
1000
10000[ F ]
K tiefe Tem peratur K Raum tem peratur
kcg / Hz
U / m V
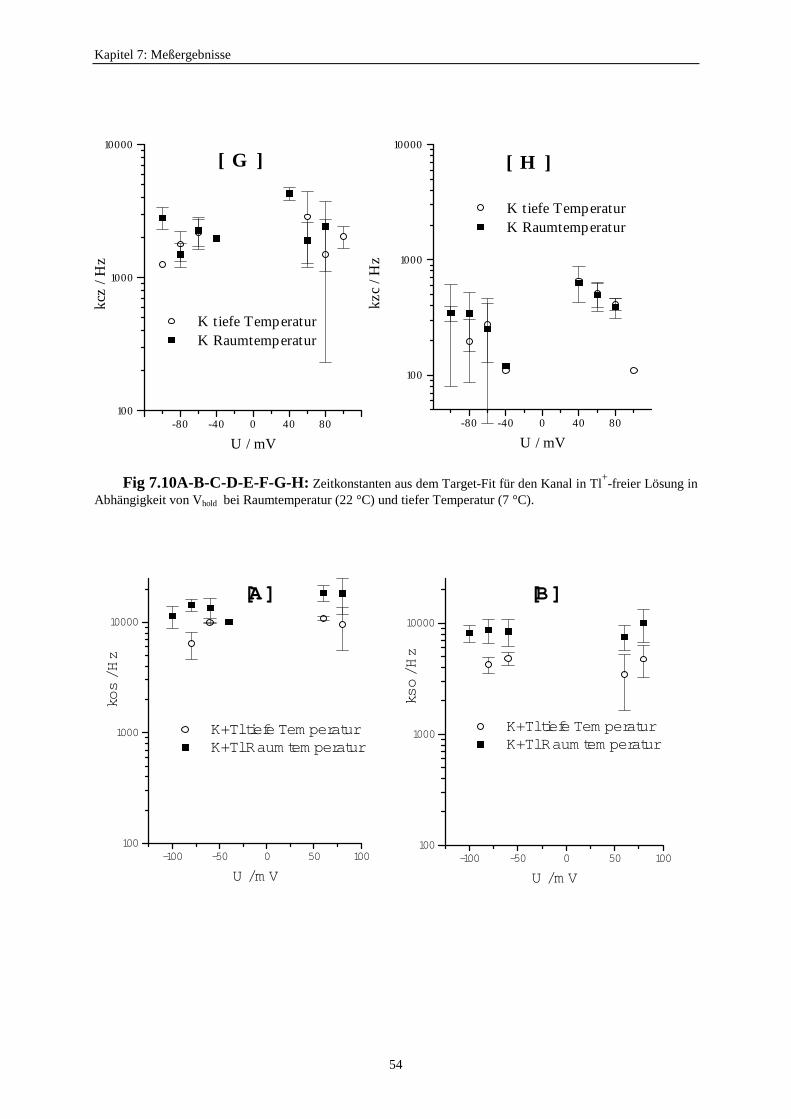
Kapitel 7: Meßergebnisse
54
-80 -40 0 40 80
100
1000
10000
[ H ]
K tiefe Temperatur K Raumtemperatur
kzc
/ Hz
U / mV-80 -40 0 40 80
100
1000
10000
[ G ]
K tiefe Temperatur K Raumtemperatur
kcz
/ Hz
U / mV
Fig 7.10A-B-C-D-E-F-G-H: Zeitkonstanten aus dem Target-Fit für den Kanal in Tl+-freier Lösung inAbhängigkeit von Vhold bei Raumtemperatur (22 °C) und tiefer Temperatur (7 °C).
-100 -50 0 50 100100
1000
10000
[A]
K+Tl tiefe Tem peratur K+Tl Raum tem peratur
kos / Hz
U / m V-100 -50 0 50 100
100
1000
10000
[B]
K+Tl tiefe Tem peratur K+Tl Raum tem peratur
kso / Hz
U / m V
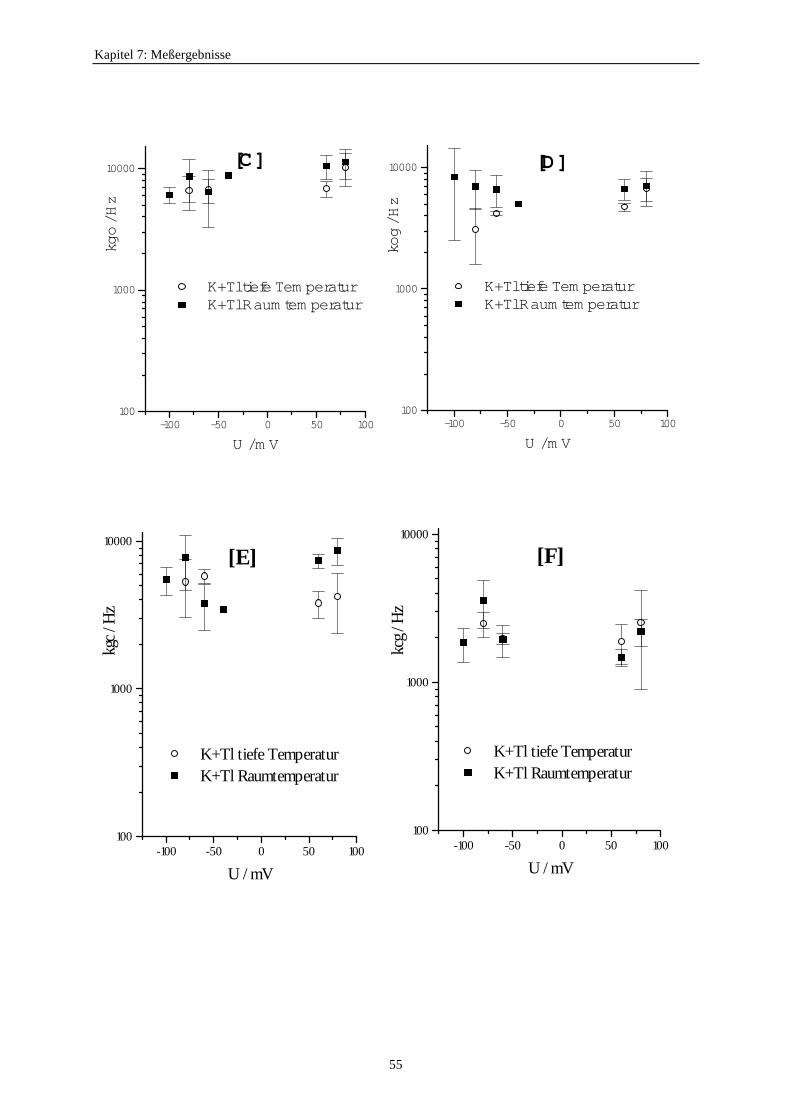
Kapitel 7: Meßergebnisse
55
-100 -50 0 50 100100
1000
10000 [D]
K+Tl tiefe Tem peratur K+Tl Raum tem peratur
kog / Hz
U / m V-100 -50 0 50 100
100
1000
10000 [C]
K+Tl tiefe Tem peratur K+Tl Raum tem peratur
kgo / Hz
U / m V
-100 -50 0 50 100100
1000
10000
[F]
K+Tl tiefe Temperatur K+Tl Raumtemperatur
kcg /
Hz
U / mV-100 -50 0 50 100
100
1000
10000[E]
K+Tl tiefe Temperatur K+Tl Raumtemperatur
kgc /
Hz
U / mV
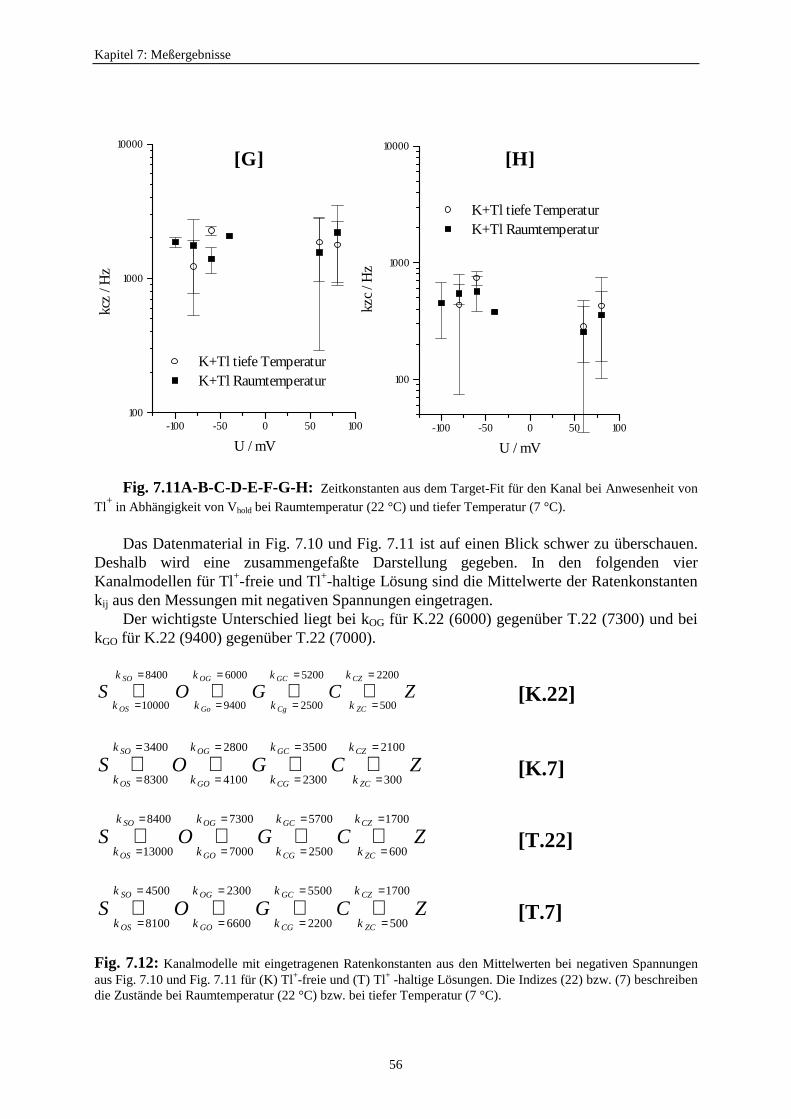
Kapitel 7: Meßergebnisse
56
-100 -50 0 50 100
100
1000
10000[H]
K+Tl tiefe Temperatur K+Tl Raumtemperatur
kzc /
Hz
U / mV-100 -50 0 50 100
100
1000
10000[G]
K+Tl tiefe Temperatur K+Tl Raumtemperatur
kcz
/ Hz
U / mV
Fig. 7.11A-B-C-D-E-F-G-H: Zeitkonstanten aus dem Target-Fit für den Kanal bei Anwesenheit vonTl+ in Abhängigkeit von Vhold bei Raumtemperatur (22 °C) und tiefer Temperatur (7 °C).
Das Datenmaterial in Fig. 7.10 und Fig. 7.11 ist auf einen Blick schwer zu überschauen.Deshalb wird eine zusammengefaßte Darstellung gegeben. In den folgenden vierKanalmodellen für Tl+-freie und Tl+-haltige Lösung sind die Mittelwerte der Ratenkonstantenkij aus den Messungen mit negativen Spannungen eingetragen.
Der wichtigste Unterschied liegt bei kOG für K.22 (6000) gegenüber T.22 (7300) und beikGO für K.22 (9400) gegenüber T.22 (7000).
ZCGOSCZ
ZC
GC
Cg
OG
Go
SO
OS
k
k
k
k
k
k
k
k
2200
500
5200
2500
6000
9400
8400
10000
=
=
=
=
=
=
=
=⇔⇔⇔⇔ [K.22]
ZCGOSCZ
ZC
GC
CG
OG
GO
SO
OS
k
k
k
k
k
k
k
k
2100
300
3500
2300
2800
4100
3400
8300
=
=
=
=
=
=
=
=⇔⇔⇔⇔ [K.7]
ZCGOSCZ
ZC
GC
CG
OG
GO
SO
OS
k
k
k
k
k
k
k
k
1700
600
5700
2500
7300
7000
8400
13000
=
=
=
=
=
=
=
=⇔⇔⇔⇔ [T.22]
ZCGOSCZ
ZC
GC
CG
OG
GO
SO
OS
k
k
k
k
k
k
k
k
1700
500
5500
2200
2300
6600
4500
8100
=
=
=
=
=
=
=
=⇔⇔⇔⇔ [T.7]
Fig. 7.12: Kanalmodelle mit eingetragenen Ratenkonstanten aus den Mittelwerten bei negativen Spannungenaus Fig. 7.10 und Fig. 7.11 für (K) Tl+-freie und (T) Tl+ -haltige Lösungen. Die Indizes (22) bzw. (7) beschreibendie Zustände bei Raumtemperatur (22 °C) bzw. bei tiefer Temperatur (7 °C).

Kapitel 7: Meßergebnisse
57
7.3.5.2 Änderung der Ratenkonstanten unter Einfluß von Tl+
7.3.5.2.1 Mittelung im Burstbereich
In den Vorläuferarbeiten (Draber et al., 1991; Draber und Hansen, 1994; Keunecke, 1995;Blunck, 1996) stand die Hypothese im Vordergrund, daß schnelles Schalten im Burstbereicheine scheinbare Reduzierung des Einzelkanalstromes erzeugt. Wie der Abschnitt 7.3.8 zeigt,ist die Situation hier anders, weil die Bestimmung der Einzelkanalströme und derRatenkonstanten vom gleichen Datenmaterial ausgeht, daß der gleiche Filter durchlaufen hat.
Doch um die Ergebnisse hier einmal im Lichte der Hypothese der Vorläufermodelle zubetrachten, soll so getan werden, als würde bei der Bestimmung des Einzelkanalstromes überdie Burstbereiche gemittelt. Diese Rechnungen haben neben dem genannten historischenGrund auch eine praktische Aussage. Viele Patcher arbeiten mit geringerer Zeitauflösung alsdie hier benutzte. Für diese Untersuchungen sind die folgenden Analysen von hoher Relevanz.
Es müssen also Bereiche herausgesucht werden, bei denen die Mittelung über vieleSchaltereignisse an einem zu niedrigeren geschätzten Einzelkanalstromwert führt. Fig. 7.1 und7.2 zeigen das Problem der Stromwertschätzung. Einzelne kurze Öffnungen können nicht fürdie Schätzung benutzt werden, wie z. B. die Werte bei mittleren Niveau bei 0.25 s in Fig. 7.2.Man braucht eine etwas längere „Wolke“, von ca. 1 bis 2 ms bei einemSignal/Rauschverhältnis von ca. 2 bis 3 um den Schwerpunkt für die Stromschätzung zutreffen. Diese Mittelzeit ist abhängig vom Signal/Rauschverhältnis der gemessenen Zeitreihe.Das Gleiche gilt für die Amplitudenhistogramme Fig. 7.3 Die Lage des Maximums ergibt sichaus der Betrachtung der Kurvenform der Verteilung. Auch dies ist ein Mittelungsprozeß.
Zur Veranschaulichung der damit verbundenen Problematik gehen wir erst einmal davonaus, daß ein Mittelungsprozeß wie in Fig. 5.2 vorläge. Die Richtigkeit dieser Annahme wirdim Licht der folgenden Ergebnisse dann noch einmal diskutiert.
Die Frage ist jetzt, welche Einheit für den Mittelungsprozeß betrachtet wird. So einAbschnitt schnellen Schaltens, wie er z. B. nach 0.25 s in Fig. 7.2 auf dem zweiten (untersten)Niveau auftritt, wird Burst genannt. Es ist deshalb zu untersuchen, ob ein Teilbereich desModells isoliert werden kann, der für die Bursts verantwortlich ist. Bursts sind Periodenschnellen Schaltens, die von etwas längeren Geschlossen-Zuständen unterbrochen werden. ImSinne des AMFEs ist dann zu untersuchen, ob eine Mittelung über die Offen- undGeschlossenzeiten im Burst die Reduzierung des scheinbaren Einzelkanalstroms beiAnwesenheit von Tl+ widerspiegelt.
Eine Inspektion der Kanalmodelle in Fig. 7.12 zeigt folgendes: Die Ratenkonstanten desÜberganges zwischen den benachbarten Offen- und Geschlossenzuständen (O und G) sindsehr hoch. Sie sind für das sogenannte Flickern im Burst verantwortlich. Die Frage ist, waseinen Burst beendet. Der Austausch O-S ist ebenfalls sehr schnell, so daß durch einen Sprungin S kein "Einfrieren" im Offenzustand das Flickern beenden würde.
Die Übergangsrate kGC ist deutlich größer als die Übergangsrate kCG sowohl in Tl+-freierwie in Tl+-haltiger Lösung bei Raumtemperatur (siehe Fig. 7.12.A und 7.12.B). DieRatenkonstante kCG ist für den unblockierten Kanal im Bereich 1000 bis 2500 Hz und kGC imBereich 3500 bis 6500 Hz. Für den blockierten Kanal liegt die Ratenkonstante kCG zwischen1800 und 3600 Hz und kGC zwischen 3800 und 7800 Hz. Der Rücksprung von C nach G hatdamit eine Ratenkonstante, die um den Faktor drei kleiner ist als die des Flickerns zwischenS, O und G und sie entspricht einer Zeitkonstante von ca. 3 ms, die zu den Lücken zwischenden Bursts paßt. Außerdem kann der Kanal mit gleicher Wahrscheinlichkeit in den Zustand Zspringen. Wenn der Kanal sich im Zustand Z befindet, ist er damit aus den schnellen

Kapitel 7: Meßergebnisse
58
Schaltvorgängen ganz heraus (auf dem obersten Niveau (Grundniveau) von 0.5 s in Fig. 7.2und die lange Stufe rechts in Fig. 6.4).
Die Ratenkonstante des Überganges von G nach C bestimmt die Länge des Bursts, dieVerweildauer in C oder Z die Pause zwischen den Bursts. Diese Verweildauer in C istbestimmt durch die Übergänge von Zustand C nach Zustand Z oder G. Beide Übergänge sinddeutlich langsamer als die schnellen Ratenkonstanten des Bursts. Deshalb wird für dieUntersuchung des Bursts nur das Teilmodell S<=>O<=>G vom GesamtmodellS<=>O<=>G<=>C<=>Z betrachtet.
Die Spannungsabhängigkeit der Ratenkonstanten steht nicht im Interesse dieser Arbeit.Dennoch ist bei verschiedenen Spannungen gemessen worden. Die Fig. 7.10 bis 7.11 zeigen,daß keine große Spannungsabhängigkeit im Bereich des AMFEs besteht. Durch dieMessungen bei unterschiedlichen Spannungen steht jedoch ein größeres Datenmaterial für denVergleich der Ratenkonstanten bei unterschiedlichen Temperaturen und für den blockiertenund unblockierten Fall zur Verfügung.
Für diesen Vergleich sind die Ratenkonstanten aus Fig. 7.10 und 7.11 noch einmal sozusammengestellt worden, daß K+- und K+ + Tl+-Werte direkt miteinander verglichen werdenkönnen. Auf den ersten Blick werden keine großen Unterschiede zwischen denRatenkonstanten in den unterschiedlichen Lösungen beobachtet. Der größte Unterschied wirdbei der schnellsten Ratenkonstante kos gefunden. In Raumtemperatur und für negativeSpannungen liegt die Ratenkonstante kos der K+/Tl+-Gemischlösung zwischen 11000 und15000 Hz. Bei reiner Kaliumlösung liegt sie zwischen 9000 und 11000 Hz. Bei einerTemperatur von 7 °C nimmt die Ratenkonstante kos der K+/Tl+-Gemischlösung zwischen 6500und 10000 Hz und der K+ Lösung zwischen 6000 und 9500 Hz ab (Fig. 7.13.C-D). Somitunterscheiden sich die Ratenkonstanten bei der Temperatur von 7 °C nicht mehr voneinander.Dieser Unterschied der Ratenkonstanten der S - O Übergänge kann eigentlich nicht beobachtetwerden, da es sich um einen Übergang zwischen zwei offenen Zuständen handelt. Er hat aberAuswirkung auf die Besetzungswahrscheinlichkeit der beiden offenen Zustände, worauf inAbschnitt 7.3.6.2 noch genauer eingegangen wird.
Die interessanten Übergangsraten sind die zwischen dem offenen und geschlossenenZustand (kOG, kGO). Bei der Betrachtung jeder dieser einzelnen Ratenkonstanten wird beiRaumtemperatur ein Unterschied zwischen dem blockierten und unblockierten Kanalbeobachtet, der im Mittelwert ca. 20 % ausmacht (Fig. 7.12). Dies wird noch deutlicher beider Betrachtung der Verhältnisse von kGO zu kOG (Fig. 7.14).
- 10 0 - 5 0 0 5 0 10 010 0
10 0 0
10 0 0 0
[ B ]
K + T l t ie f e T em p er a t u r K t ie fe T em p er a t u r
kso
/ H
z
U / m V- 10 0 - 5 0 0 5 0 10 0
10 0
10 0 0
10 0 0 0
[ A ]
K + T l R au m t em p er a t u r K R au m t em p er a t u r
kso
/ H
z
U / m V
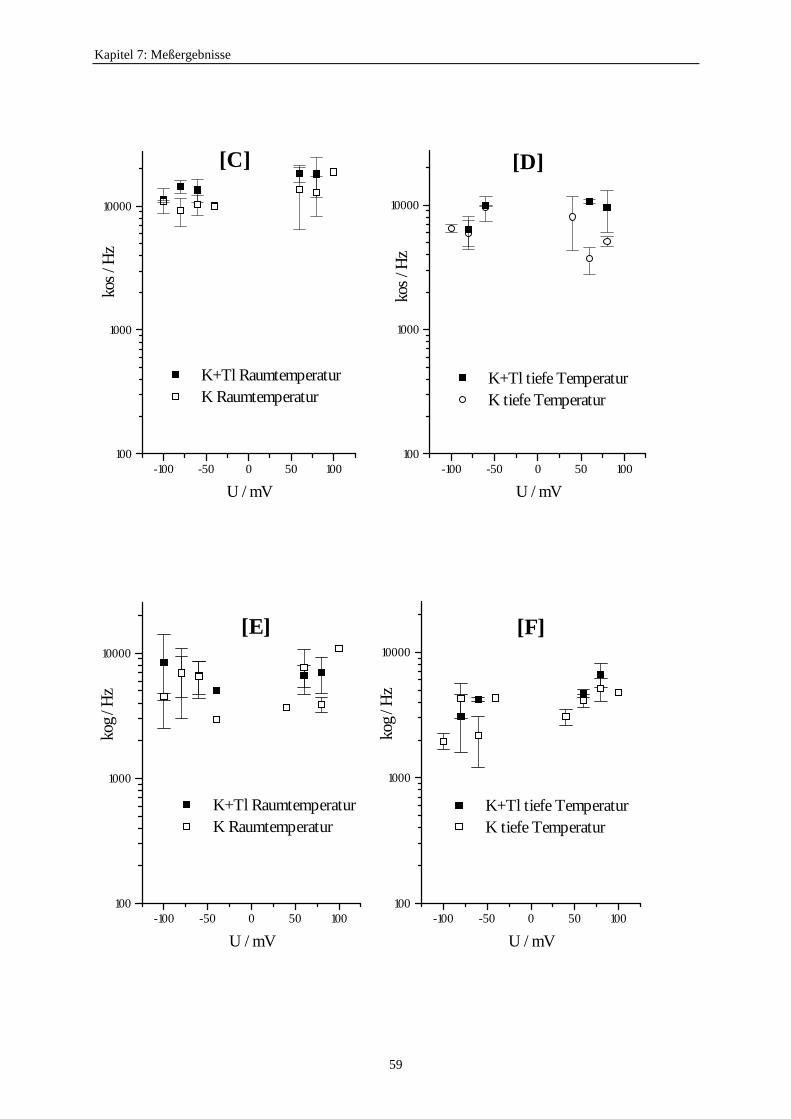
Kapitel 7: Meßergebnisse
59
-100 -50 0 50 100100
1000
10000
[C]
K+Tl Raumtemperatur K Raumtemperatur
kos /
Hz
U / mV-100 -50 0 50 100
100
1000
10000
[D]
K+Tl tiefe Temperatur K tiefe Temperatur
kos /
Hz
U / mV
-100 -50 0 50 100100
1000
10000
[E]
K+Tl Raumtemperatur K Raumtemperatur
kog /
Hz
U / mV-100 -50 0 50 100
100
1000
10000
[F]
K+Tl tiefe Temperatur K tiefe Temperatur
kog /
Hz
U / mV
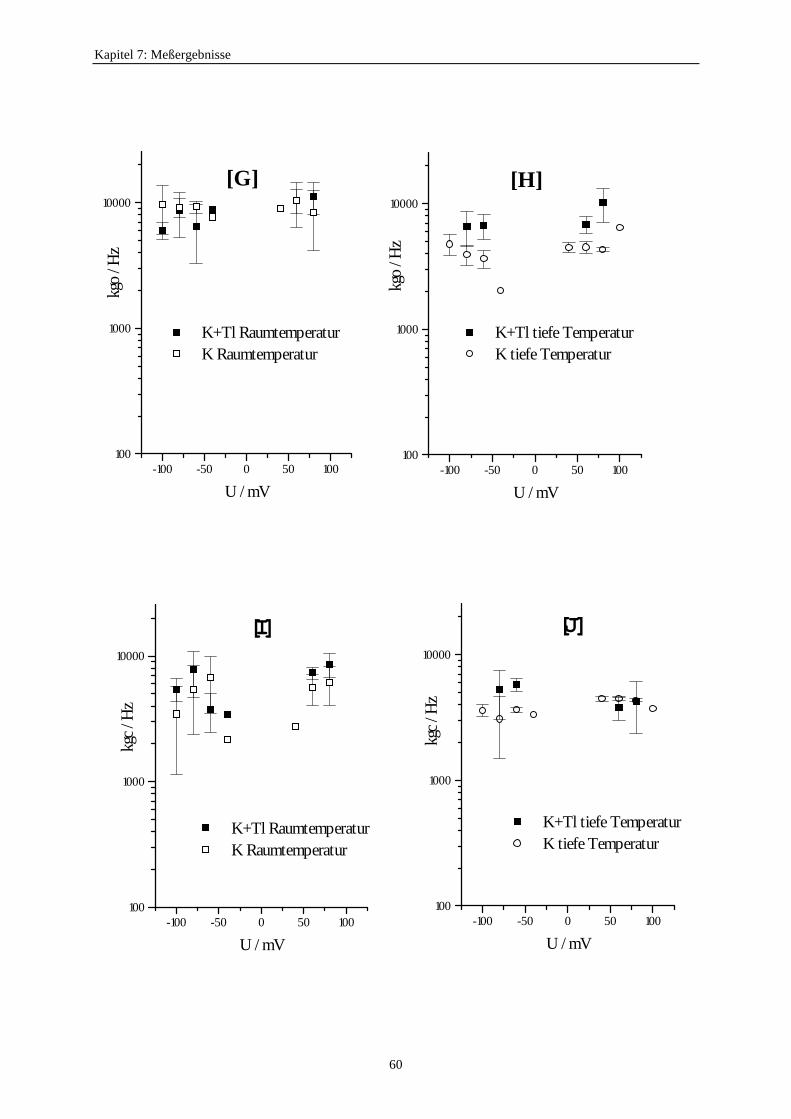
Kapitel 7: Meßergebnisse
60
-100 -50 0 50 100100
1000
10000
[H]
K+Tl tiefe Temperatur K tiefe Temperatur
kgo /
Hz
U / mV-100 -50 0 50 100
100
1000
10000
[G]
K+Tl Raumtemperatur K Raumtemperatur
kgo /
Hz
U / mV
-100 -50 0 50 100100
1000
10000
[J]
K+Tl tiefe Temperatur K tiefe Temperatur
kgc /
Hz
U / mV-100 -50 0 50 100
100
1000
10000
[I]
K+Tl Raumtemperatur K Raumtemperatur
kgc /
Hz
U / mV
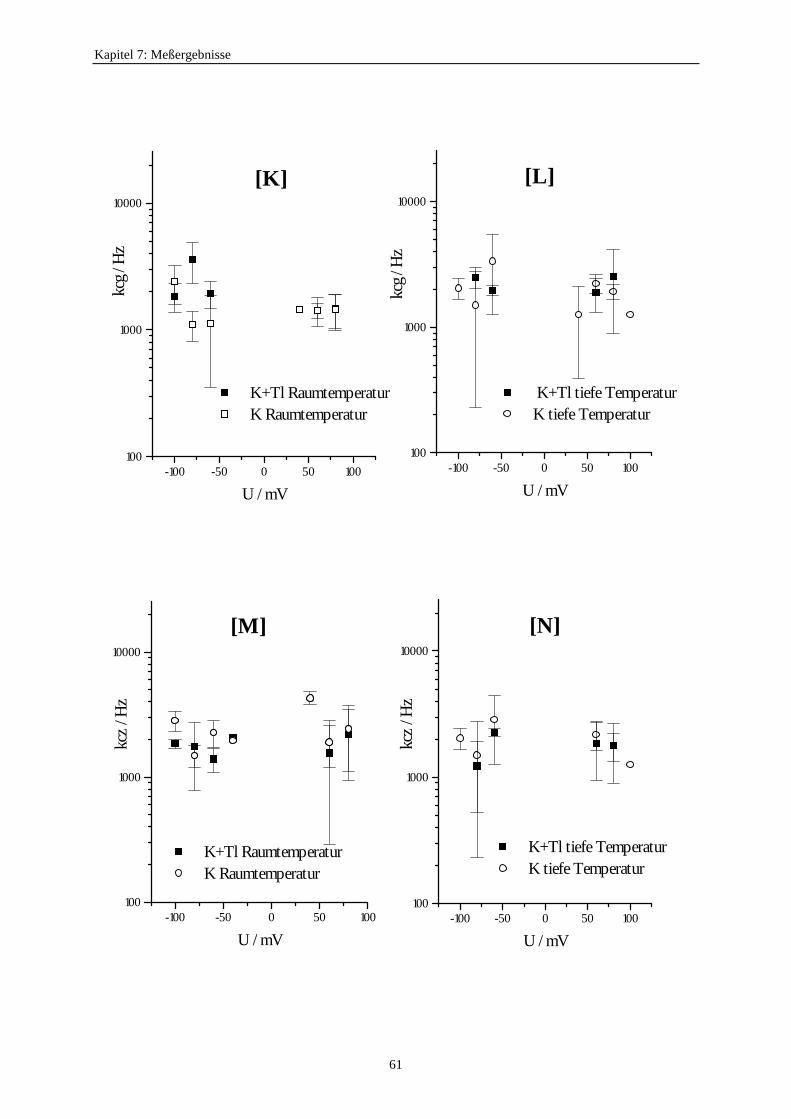
Kapitel 7: Meßergebnisse
61
-100 -50 0 50 100100
1000
10000
[L]
K+Tl tiefe Temperatur K tiefe Temperatur
kcg /
Hz
U / mV-100 -50 0 50 100
100
1000
10000
[K]
K+Tl Raumtemperatur K Raumtemperatur
kcg /
Hz
U / mV
-100 -50 0 50 100100
1000
10000
[N]
K+Tl tiefe Temperatur K tiefe Temperatur
kcz /
Hz
U / mV-100 -50 0 50 100
100
1000
10000
[M]
K+Tl Raumtemperatur K Raumtemperatur
kcz /
Hz
U / mV
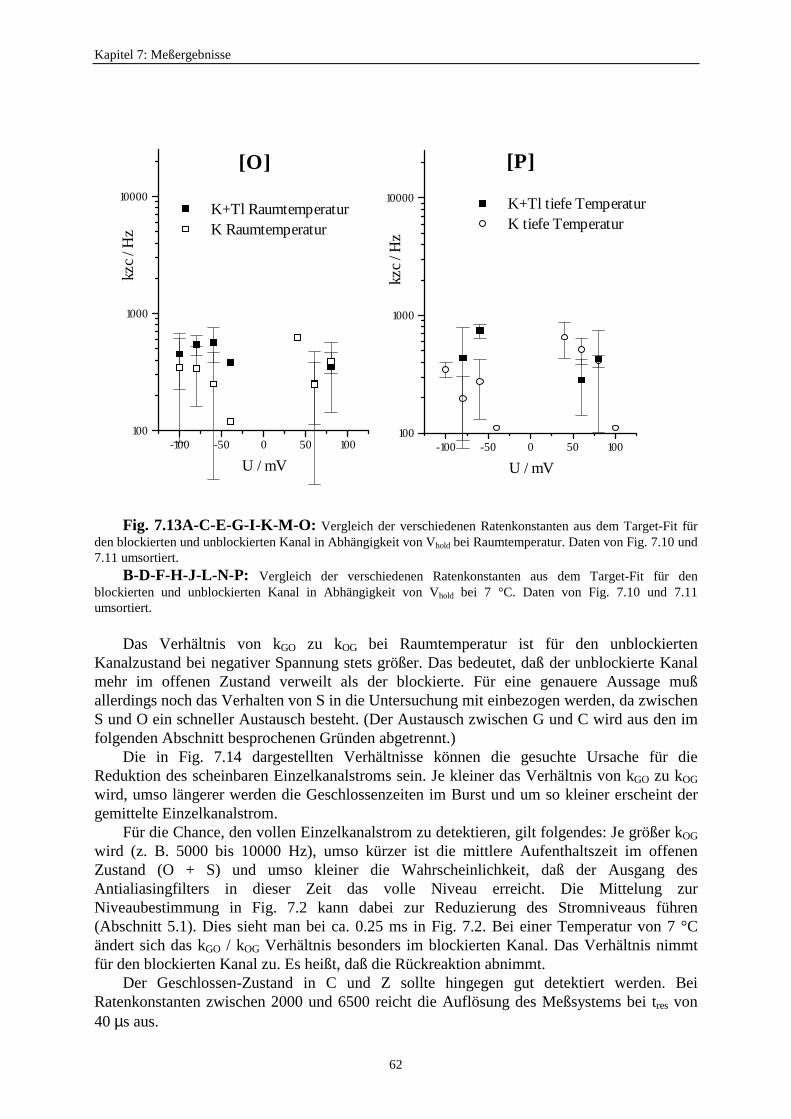
Kapitel 7: Meßergebnisse
62
-100 -50 0 50 100100
1000
10000
[P]
K+Tl tiefe Temperatur K tiefe Temperatur
kzc /
Hz
U / mV-100 -50 0 50 100
100
1000
10000
[O]
K+Tl Raumtemperatur K Raumtemperatur
kzc /
Hz
U / mV
Fig. 7.13A-C-E-G-I-K-M-O: Vergleich der verschiedenen Ratenkonstanten aus dem Target-Fit fürden blockierten und unblockierten Kanal in Abhängigkeit von Vhold bei Raumtemperatur. Daten von Fig. 7.10 und7.11 umsortiert.
B-D-F-H-J-L-N-P: Vergleich der verschiedenen Ratenkonstanten aus dem Target-Fit für denblockierten und unblockierten Kanal in Abhängigkeit von Vhold bei 7 °C. Daten von Fig. 7.10 und 7.11umsortiert.
Das Verhältnis von kGO zu kOG bei Raumtemperatur ist für den unblockiertenKanalzustand bei negativer Spannung stets größer. Das bedeutet, daß der unblockierte Kanalmehr im offenen Zustand verweilt als der blockierte. Für eine genauere Aussage mußallerdings noch das Verhalten von S in die Untersuchung mit einbezogen werden, da zwischenS und O ein schneller Austausch besteht. (Der Austausch zwischen G und C wird aus den imfolgenden Abschnitt besprochenen Gründen abgetrennt.)
Die in Fig. 7.14 dargestellten Verhältnisse können die gesuchte Ursache für dieReduktion des scheinbaren Einzelkanalstroms sein. Je kleiner das Verhältnis von kGO zu kOGwird, umso längerer werden die Geschlossenzeiten im Burst und um so kleiner erscheint dergemittelte Einzelkanalstrom.
Für die Chance, den vollen Einzelkanalstrom zu detektieren, gilt folgendes: Je größer kOGwird (z. B. 5000 bis 10000 Hz), umso kürzer ist die mittlere Aufenthaltszeit im offenenZustand (O + S) und umso kleiner die Wahrscheinlichkeit, daß der Ausgang desAntialiasingfilters in dieser Zeit das volle Niveau erreicht. Die Mittelung zurNiveaubestimmung in Fig. 7.2 kann dabei zur Reduzierung des Stromniveaus führen(Abschnitt 5.1). Dies sieht man bei ca. 0.25 ms in Fig. 7.2. Bei einer Temperatur von 7 °Cändert sich das kGO / kOG Verhältnis besonders im blockierten Kanal. Das Verhältnis nimmtfür den blockierten Kanal zu. Es heißt, daß die Rückreaktion abnimmt.
Der Geschlossen-Zustand in C und Z sollte hingegen gut detektiert werden. BeiRatenkonstanten zwischen 2000 und 6500 reicht die Auflösung des Meßsystems bei tres von40 µs aus.
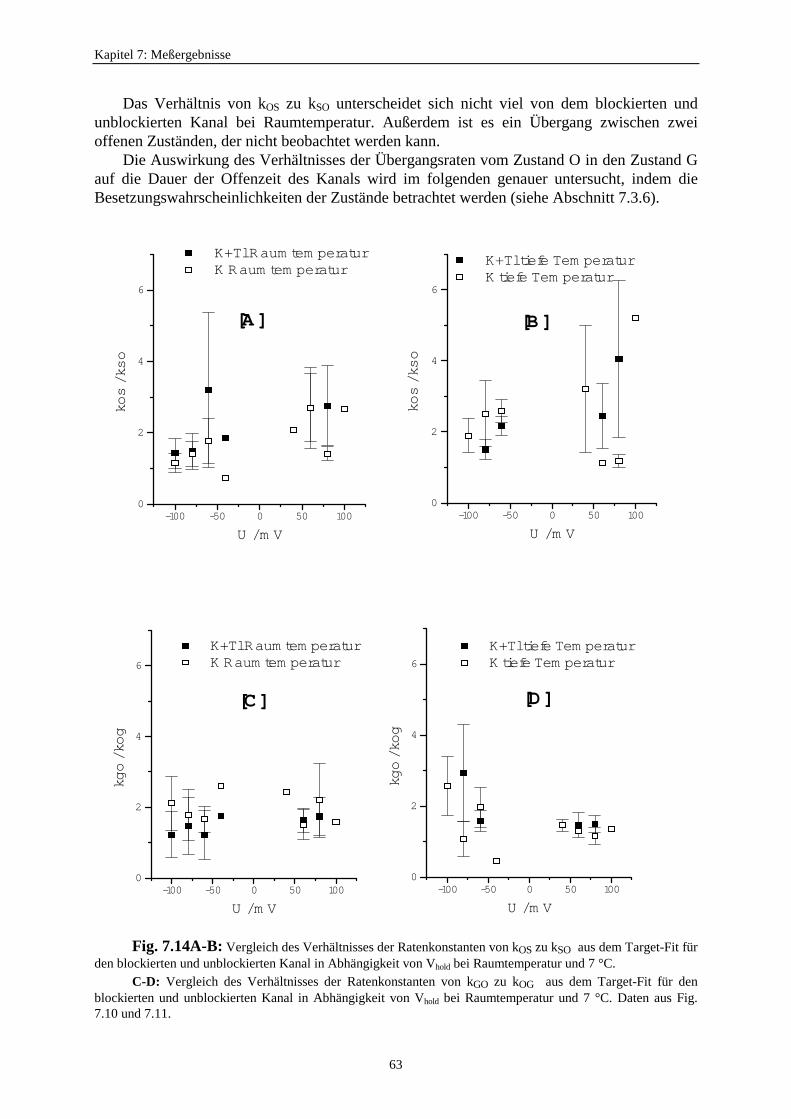
Kapitel 7: Meßergebnisse
63
Das Verhältnis von kOS zu kSO unterscheidet sich nicht viel von dem blockierten undunblockierten Kanal bei Raumtemperatur. Außerdem ist es ein Übergang zwischen zweioffenen Zuständen, der nicht beobachtet werden kann.
Die Auswirkung des Verhältnisses der Übergangsraten vom Zustand O in den Zustand Gauf die Dauer der Offenzeit des Kanals wird im folgenden genauer untersucht, indem dieBesetzungswahrscheinlichkeiten der Zustände betrachtet werden (siehe Abschnitt 7.3.6).
-100 -50 0 50 1000
2
4
6
[B]
K+Tl tiefe Tem peratur K tiefe Tem peratur
kos / kso
U / m V-100 -50 0 50 100
0
2
4
6
[A]
K+Tl Raum tem peratur K Raum tem peratur
kos / kso
U / m V
-100 -50 0 50 1000
2
4
6
[D]
K+Tl tiefe Tem peratur K tiefe Tem peratur
kgo / kog
U / m V-100 -50 0 50 100
0
2
4
6
[C]
K+Tl Raum tem peratur K Raum tem peratur
kgo / kog
U / m V
Fig. 7.14A-B: Vergleich des Verhältnisses der Ratenkonstanten von kOS zu kSO aus dem Target-Fit fürden blockierten und unblockierten Kanal in Abhängigkeit von Vhold bei Raumtemperatur und 7 °C.
C-D: Vergleich des Verhältnisses der Ratenkonstanten von kGO zu kOG aus dem Target-Fit für denblockierten und unblockierten Kanal in Abhängigkeit von Vhold bei Raumtemperatur und 7 °C. Daten aus Fig.7.10 und 7.11.

Kapitel 7: Meßergebnisse
64
7.3.6 Betrachtung der Besetzungswahrscheinlichkeiten der einzelnenZustände im blockierten und unblockierten Kanal
7.3.6.1 Definition der Besetzungswahrscheinlichkeit
Sei pi(t) der Besetzungszustand, der durch einen Vektor P(t) zusammengefaßt wird, unddie Übergangsratenkonstanten seien als Matrix K mit den Elementen kij dargestellt. So ergibtsich aus dem Massenwirkungsgesetz für die Flüsse in und aus den Zuständen die folgendeDifferentialgleichung:
KtPdt
tdP ⋅= )()((7.8)
Für die Berechnung der Steady-State-Verteilung pi(∞) im i-ten Zustand wirdvorausgesetzt, daß die Änderung der Besetzungswahrscheinlichkeiten Null ist. Aus derDifferentialgleichung (7.6) wird das folgende Bestimmungssystem mit dieser Voraussetzungerhalten:
( ) 0),...(),( 21 =⋅∞∞ Kpp (7.9)
Das Argument ∞ bedeutet stationäre Zustände. Außerdem gilt, daß die Summe über alleBesetzungswahrscheinlichkeiten gleich eins sein muß:
∑ =∞i
ip 1)( (7.10)
Mit Hilfe der Gleichungen (7.7 und 7.8) wird ein lineares Gleichungssystem erzeugt,welches für die Bestimmungen der Besetzungswahrscheinlichkeiten angewendet wird.
7.3.6.2 Besetzungswahrscheinlichkeiten im reduzierten Modell
Nach den Überlegungen des vorherigen Abschnitts 7.3.5.2 und, um die Auswirkungendes schnellen Schaltens auf die Reduzierung des Einzelkanalstroms zu untersuchen, wird einTeilbereich des Modells isoliert (siehe Abschnitt 7.3.5.2.1). Für die Betrachtung der Burstswird nur das reduzierte Markov-Modell S<=>O<=>G benutzt. Dies ist wie folgt begründet:Die Ratenkonstanten zwischen den Zuständen S<=>O<=>G sind sehr schnell, und ein Hin-und Herspringen zwischen diesen Zuständen erzeugt das im Burst (Fig. 6.4) vorliegendeschnelle Schalten. Springt das System jetzt von G nach C ist, wie in Abschnitt 7.3.5.2.1bereits erwähnt, der Burst beendet, denn der Rücksprung von C nach G ist langsamer und inder Zeitreihe des Stromes kann man die Lücke erkennen (siehe Fig. 7.2). Das TeilsystemS<=>O<=>G ist damit verlassen. Diese durch Lücken getrennten Bursts werden zurEinzelkanalstrombestimmung benutzt.
Für das reduzierte 3-Zustandsmodell S<=>O<=>G werden dann dieBesetzungswahrscheinlichkeiten der drei Zustände nach den Gleichungen 7.7 und 7.8 für denblockierten und unblockierten Kanal bestimmt und miteinander verglichen.
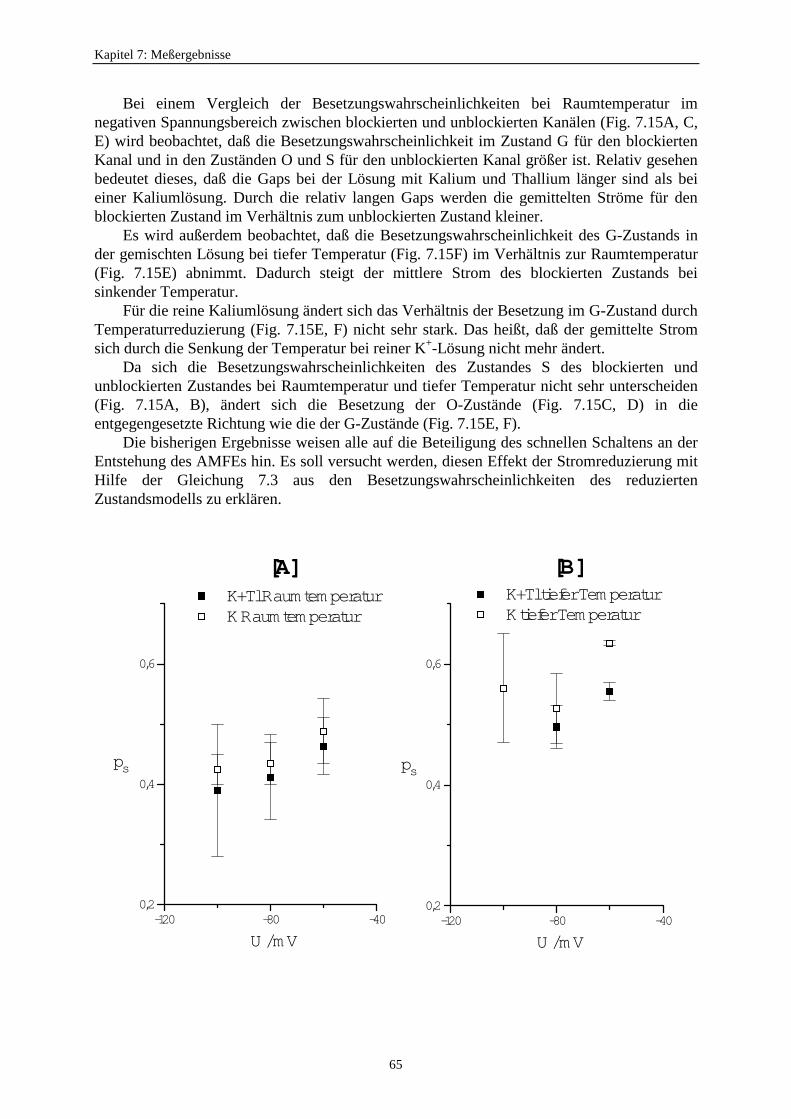
Kapitel 7: Meßergebnisse
65
Bei einem Vergleich der Besetzungswahrscheinlichkeiten bei Raumtemperatur imnegativen Spannungsbereich zwischen blockierten und unblockierten Kanälen (Fig. 7.15A, C,E) wird beobachtet, daß die Besetzungswahrscheinlichkeit im Zustand G für den blockiertenKanal und in den Zuständen O und S für den unblockierten Kanal größer ist. Relativ gesehenbedeutet dieses, daß die Gaps bei der Lösung mit Kalium und Thallium länger sind als beieiner Kaliumlösung. Durch die relativ langen Gaps werden die gemittelten Ströme für denblockierten Zustand im Verhältnis zum unblockierten Zustand kleiner.
Es wird außerdem beobachtet, daß die Besetzungswahrscheinlichkeit des G-Zustands inder gemischten Lösung bei tiefer Temperatur (Fig. 7.15F) im Verhältnis zur Raumtemperatur(Fig. 7.15E) abnimmt. Dadurch steigt der mittlere Strom des blockierten Zustands beisinkender Temperatur.
Für die reine Kaliumlösung ändert sich das Verhältnis der Besetzung im G-Zustand durchTemperaturreduzierung (Fig. 7.15E, F) nicht sehr stark. Das heißt, daß der gemittelte Stromsich durch die Senkung der Temperatur bei reiner K+-Lösung nicht mehr ändert.
Da sich die Besetzungswahrscheinlichkeiten des Zustandes S des blockierten undunblockierten Zustandes bei Raumtemperatur und tiefer Temperatur nicht sehr unterscheiden(Fig. 7.15A, B), ändert sich die Besetzung der O-Zustände (Fig. 7.15C, D) in dieentgegengesetzte Richtung wie die der G-Zustände (Fig. 7.15E, F).
Die bisherigen Ergebnisse weisen alle auf die Beteiligung des schnellen Schaltens an derEntstehung des AMFEs hin. Es soll versucht werden, diesen Effekt der Stromreduzierung mitHilfe der Gleichung 7.3 aus den Besetzungswahrscheinlichkeiten des reduziertenZustandsmodells zu erklären.
-120 -80 -400,2
0,4
0,6
[B][A] K+Tl Raumtemperatur K Raumtemperatur
ps
U / mV-120 -80 -40
0,2
0,4
0,6
K+Tl tiefer Temperatur K tiefer Temperatur
ps
U / mV
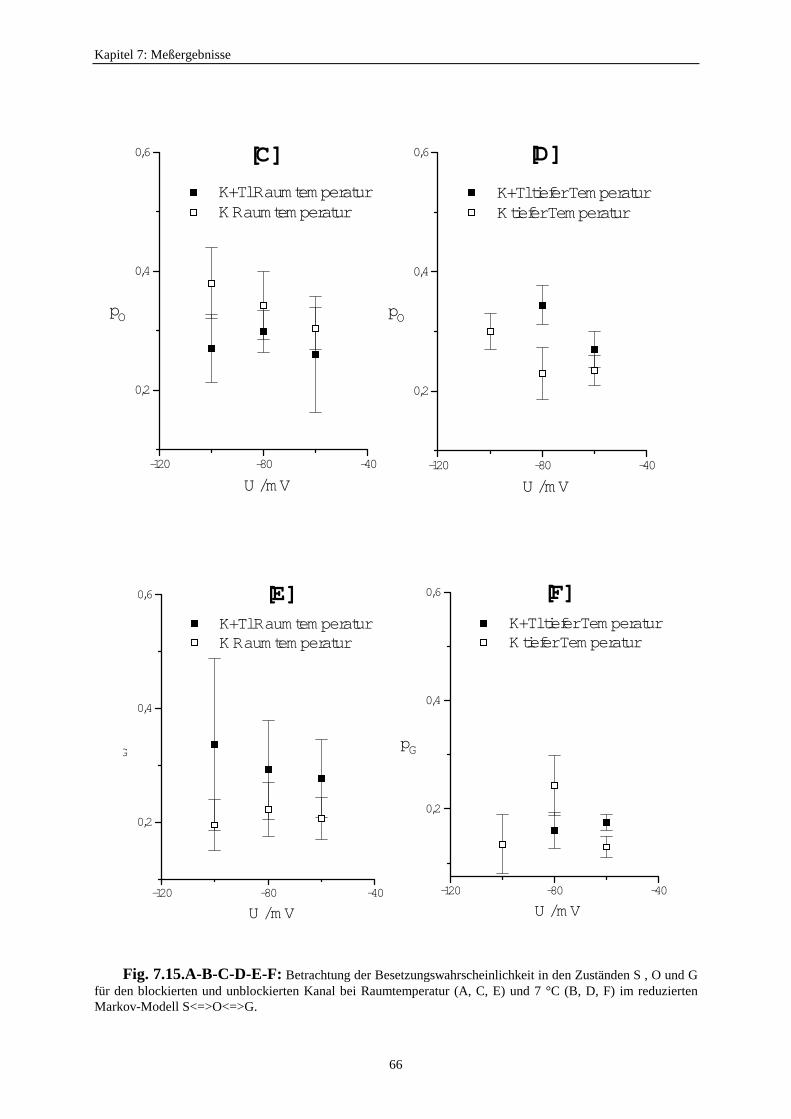
Kapitel 7: Meßergebnisse
66
-120 -80 -40
0,2
0,4
0,6 [D]
K+Tl tiefer Temperatur K tiefer Temperatur
pO
U / mV
-120 -80 -40
0,2
0,4
0,6 [C]
K+Tl Raumtemperatur K Raumtemperatur
pO
U / mV
-120 -80 -40
0,2
0,4
0,6
K+Tl tiefer Tem peratur K tiefer Tem peratur
pG
U / m V-120 -80 -40
0,2
0,4
0,6 [F][E] K+Tl Raum tem peratur K Raum tem peratur
G
U / m V
Fig. 7.15.A-B-C-D-E-F: Betrachtung der Besetzungswahrscheinlichkeit in den Zuständen S , O und Gfür den blockierten und unblockierten Kanal bei Raumtemperatur (A, C, E) und 7 °C (B, D, F) im reduziertenMarkov-Modell S<=>O<=>G.

Kapitel 7: Meßergebnisse
67
7.3.7 Auswirkungen des schnellen Schaltens auf die Reduzierung desEinzelkanalstroms
Wie oben ausgeführt sind die Besetzungswahrscheinlichkeiten der Offen- undGeschlossen-Zustände des reduzierten Teilmodells S<=>O<=>G von Wichtigkeit für dengemittelten Strom (Abschnitt 7.3.6.2).
Die Teilbehandlung der drei Zustände erfordert, daß die Summe der dreiAufenthaltswahrscheinlichkeiten auf 1 normiert wird (verweilt im Burst Teilsystem odernicht). Die Offenwahrscheinlichkeit dieses reduzierten Zustandsmodells ist damit proportionalzum Strom:
)()( ** ∞+∞∝ SO ppi (7.11)
So werden die Besetzungswahrscheinlichkeiten des reduzierten Markov-Modells, die imAbschnitt 7.3.6.2 bestimmt worden sind, entnommen, und daraus wird dieOffenwahrscheinlichkeit im Burstbereich bestimmt. Mit Hilfe der Beziehung (7.11) wird einedem Stromwert proportionale Größe erhalten. Es wird diese Größe für den blockierten undunblockierten Kanal bei Raumtemperatur und 7 °C verglichen. Weiterhin werden dieermittelten Werte aus der Beziehung 7.9 mit den gemessenen Stromwerten verglichen.
T / °C itheo σ SEW imess σ SEWK+ 20 0.79 0.07 0.02 0.79 0.1 0.03K+ + Tl+ 20 0.7 0.18 0.05 0.6 0.14 0.03K+ 7 0.81 0.07 0.03 0.81 0.1 0.03K+ + Tl+ 7 0.85 0.05 0.02 0.75 0.08 0.03
Tabelle 7.1: Vergleich der theoretischen (Gleichung 7.11) Stromwerte itheo mit den gemessenen imess
Sigma ist die Schwankung des Meßwertes, SEW ist die Schwankung des Erwartungswertes = σ / n . Bei derMittelung wurden die Daten von -60 mV, -80 mV und -100 mV zusammengefaßt, um die statistische Sicherheitzu erhöhen. In der Zeile imess sind die gemessenen Stromwerte für die direkte Abschätzung der kinetischenEffekte auf die jeweiligen itheor-Werte für K+ normiert worden, um die nicht interessierendeTemperaturabhängigkeit des wahren Einzelkanalstromes zu eliminieren.
Die Ergebnisse in Tab. 7.1 zeigen den gewünschten Effekt in Aufzeichnungen, die überdie Burstbereiche mitteln. Bei Raumtemperatur liegt der nach Gleichung 7.11 berechneteStrom für die K+ / Tl+-Mischung um 10% unter dem in K+. Bei 7 °C ist der Strom derMischung leicht größer als der im reinen K+. Während der Wert bei Raumtemperaturaußerhalb des Fehlerbereichs liegt, liegt er bei 7 °C innerhalb.
Bei den gemessenen Werten liegt der Strom für die K+ / Tl+-Mischlösung um mehr als20% unter dem in K+-Lösung. Auch bei tieferer Temperatur ist der Strom in der K+ / Tl+-Mischlösung auch unter dem der reinen K+-Lösung, allerdings ist dieser Unterschied um denFaktor 3 geschrumpft und kaum noch signifikant.
Die Frage, die sich in Anbetracht der großen Streuung der Ratenkonstanten (Fig. 7.13)stellt, ist die nach der Signifikanz der Aussage. Die Streuungen σ sind im Verhältnis zumUnterschied zwischen den itheo-Werten groß. Doch bei der Zuverlässigkeit eines Mittelwertesinteressiert die Schwankung des Erwartungswertes (SEW in Tab. 7.1). Betrachtet man denSEW in Tab. 7.1, so sieht man, daß sich die Fehlerbereiche bei Raumtemperatur nicht mehr
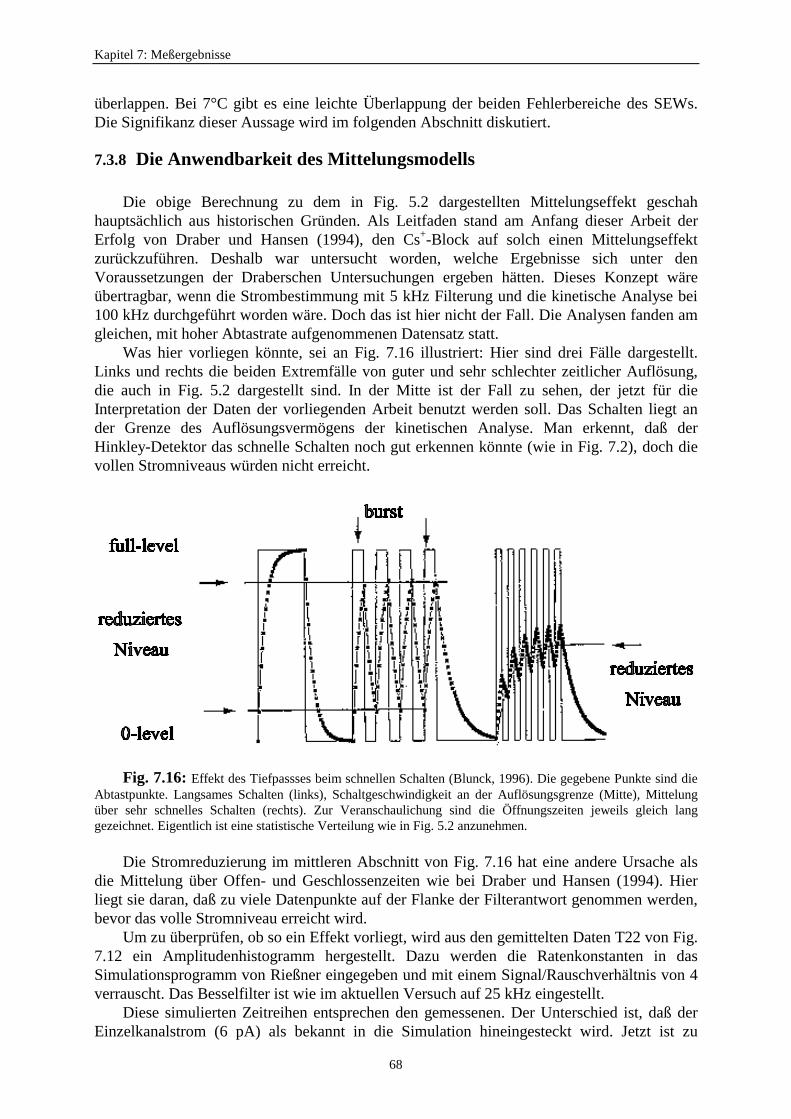
Kapitel 7: Meßergebnisse
68
überlappen. Bei 7°C gibt es eine leichte Überlappung der beiden Fehlerbereiche des SEWs.Die Signifikanz dieser Aussage wird im folgenden Abschnitt diskutiert.
7.3.8 Die Anwendbarkeit des Mittelungsmodells
Die obige Berechnung zu dem in Fig. 5.2 dargestellten Mittelungseffekt geschahhauptsächlich aus historischen Gründen. Als Leitfaden stand am Anfang dieser Arbeit derErfolg von Draber und Hansen (1994), den Cs+-Block auf solch einen Mittelungseffektzurückzuführen. Deshalb war untersucht worden, welche Ergebnisse sich unter denVoraussetzungen der Draberschen Untersuchungen ergeben hätten. Dieses Konzept wäreübertragbar, wenn die Strombestimmung mit 5 kHz Filterung und die kinetische Analyse bei100 kHz durchgeführt worden wäre. Doch das ist hier nicht der Fall. Die Analysen fanden amgleichen, mit hoher Abtastrate aufgenommenen Datensatz statt.
Was hier vorliegen könnte, sei an Fig. 7.16 illustriert: Hier sind drei Fälle dargestellt.Links und rechts die beiden Extremfälle von guter und sehr schlechter zeitlicher Auflösung,die auch in Fig. 5.2 dargestellt sind. In der Mitte ist der Fall zu sehen, der jetzt für dieInterpretation der Daten der vorliegenden Arbeit benutzt werden soll. Das Schalten liegt ander Grenze des Auflösungsvermögens der kinetischen Analyse. Man erkennt, daß derHinkley-Detektor das schnelle Schalten noch gut erkennen könnte (wie in Fig. 7.2), doch dievollen Stromniveaus würden nicht erreicht.
Fig. 7.16: Effekt des Tiefpassses beim schnellen Schalten (Blunck, 1996). Die gegebene Punkte sind dieAbtastpunkte. Langsames Schalten (links), Schaltgeschwindigkeit an der Auflösungsgrenze (Mitte), Mittelungüber sehr schnelles Schalten (rechts). Zur Veranschaulichung sind die Öffnungszeiten jeweils gleich langgezeichnet. Eigentlich ist eine statistische Verteilung wie in Fig. 5.2 anzunehmen.
Die Stromreduzierung im mittleren Abschnitt von Fig. 7.16 hat eine andere Ursache alsdie Mittelung über Offen- und Geschlossenzeiten wie bei Draber und Hansen (1994). Hierliegt sie daran, daß zu viele Datenpunkte auf der Flanke der Filterantwort genommen werden,bevor das volle Stromniveau erreicht wird.
Um zu überprüfen, ob so ein Effekt vorliegt, wird aus den gemittelten Daten T22 von Fig.7.12 ein Amplitudenhistogramm hergestellt. Dazu werden die Ratenkonstanten in dasSimulationsprogramm von Rießner eingegeben und mit einem Signal/Rauschverhältnis von 4verrauscht. Das Besselfilter ist wie im aktuellen Versuch auf 25 kHz eingestellt.
Diese simulierten Zeitreihen entsprechen den gemessenen. Der Unterschied ist, daß derEinzelkanalstrom (6 pA) als bekannt in die Simulation hineingesteckt wird. Jetzt ist zu
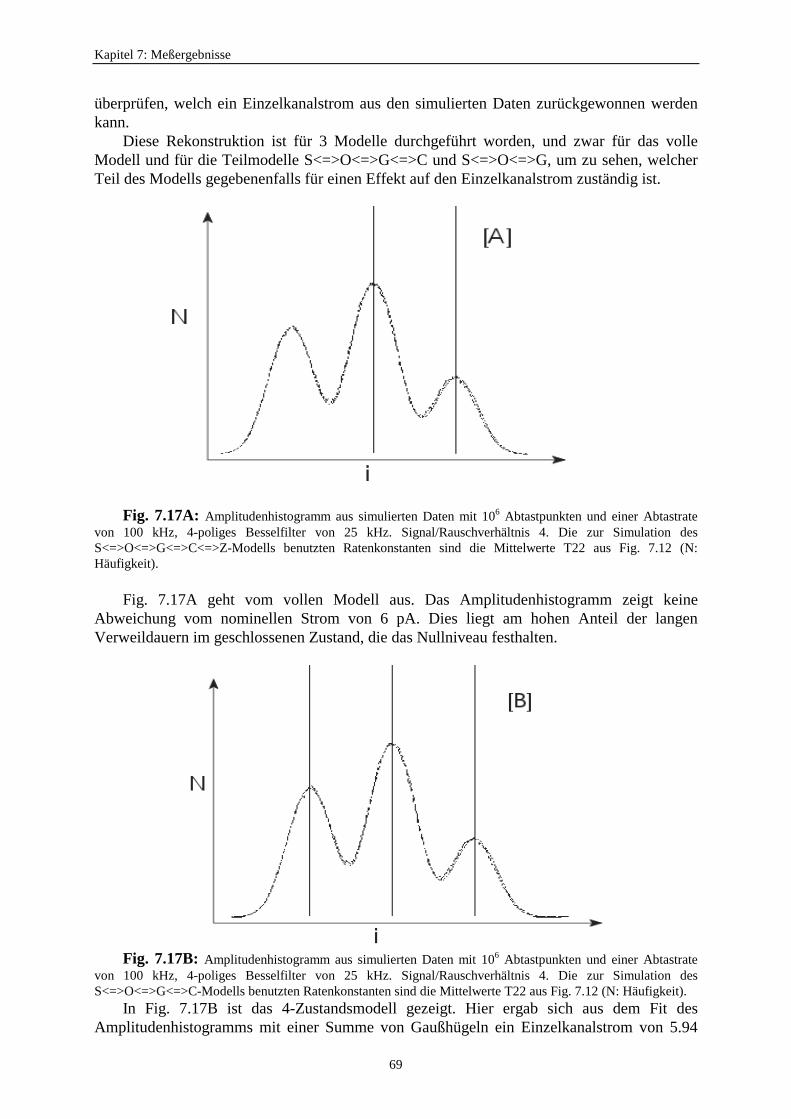
Kapitel 7: Meßergebnisse
69
überprüfen, welch ein Einzelkanalstrom aus den simulierten Daten zurückgewonnen werdenkann.
Diese Rekonstruktion ist für 3 Modelle durchgeführt worden, und zwar für das volleModell und für die Teilmodelle S<=>O<=>G<=>C und S<=>O<=>G, um zu sehen, welcherTeil des Modells gegebenenfalls für einen Effekt auf den Einzelkanalstrom zuständig ist.
Fig. 7.17A: Amplitudenhistogramm aus simulierten Daten mit 106 Abtastpunkten und einer Abtastratevon 100 kHz, 4-poliges Besselfilter von 25 kHz. Signal/Rauschverhältnis 4. Die zur Simulation desS<=>O<=>G<=>C<=>Z-Modells benutzten Ratenkonstanten sind die Mittelwerte T22 aus Fig. 7.12 (N:Häufigkeit).
Fig. 7.17A geht vom vollen Modell aus. Das Amplitudenhistogramm zeigt keineAbweichung vom nominellen Strom von 6 pA. Dies liegt am hohen Anteil der langenVerweildauern im geschlossenen Zustand, die das Nullniveau festhalten.
Fig. 7.17B: Amplitudenhistogramm aus simulierten Daten mit 106 Abtastpunkten und einer Abtastratevon 100 kHz, 4-poliges Besselfilter von 25 kHz. Signal/Rauschverhältnis 4. Die zur Simulation desS<=>O<=>G<=>C-Modells benutzten Ratenkonstanten sind die Mittelwerte T22 aus Fig. 7.12 (N: Häufigkeit).
In Fig. 7.17B ist das 4-Zustandsmodell gezeigt. Hier ergab sich aus dem Fit desAmplitudenhistogramms mit einer Summe von Gaußhügeln ein Einzelkanalstrom von 5.94
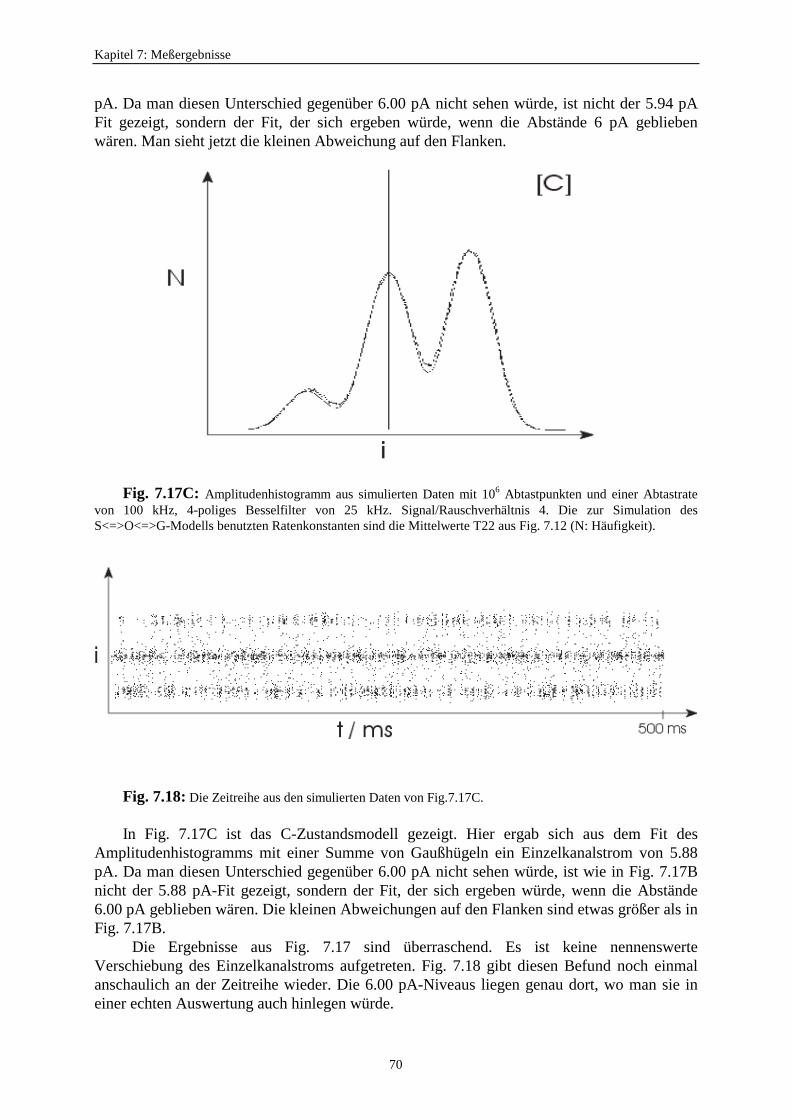
Kapitel 7: Meßergebnisse
70
pA. Da man diesen Unterschied gegenüber 6.00 pA nicht sehen würde, ist nicht der 5.94 pAFit gezeigt, sondern der Fit, der sich ergeben würde, wenn die Abstände 6 pA gebliebenwären. Man sieht jetzt die kleinen Abweichung auf den Flanken.
Fig. 7.17C: Amplitudenhistogramm aus simulierten Daten mit 106 Abtastpunkten und einer Abtastratevon 100 kHz, 4-poliges Besselfilter von 25 kHz. Signal/Rauschverhältnis 4. Die zur Simulation desS<=>O<=>G-Modells benutzten Ratenkonstanten sind die Mittelwerte T22 aus Fig. 7.12 (N: Häufigkeit).
Fig. 7.18: Die Zeitreihe aus den simulierten Daten von Fig.7.17C.
In Fig. 7.17C ist das C-Zustandsmodell gezeigt. Hier ergab sich aus dem Fit desAmplitudenhistogramms mit einer Summe von Gaußhügeln ein Einzelkanalstrom von 5.88pA. Da man diesen Unterschied gegenüber 6.00 pA nicht sehen würde, ist wie in Fig. 7.17Bnicht der 5.88 pA-Fit gezeigt, sondern der Fit, der sich ergeben würde, wenn die Abstände6.00 pA geblieben wären. Die kleinen Abweichungen auf den Flanken sind etwas größer als inFig. 7.17B.
Die Ergebnisse aus Fig. 7.17 sind überraschend. Es ist keine nennenswerteVerschiebung des Einzelkanalstroms aufgetreten. Fig. 7.18 gibt diesen Befund noch einmalanschaulich an der Zeitreihe wieder. Die 6.00 pA-Niveaus liegen genau dort, wo man sie ineiner echten Auswertung auch hinlegen würde.

Kapitel 7: Meßergebnisse
71
Obiges Ergebnis ist unerwartet. Auf der Grundlage der vorliegenden Daten ist alsoauszuschließen, daß schnelles Schalten den AMFE auf dem Einzelkanal vortäuscht. Die Frageist die nach der Zuverlässigkeit.1. Wenn schnellere unentdeckte Ratenkonstanten vorliegen würden, müßte man einen
Hinweis auf ihre Existenz in den Amplitudenhistogrammen sehen. Aber dieAmplitudenhistogramme der echten Zeitreihen (Fig. 7.3) ließen sich sehr gut durchGaußhügel fitten. Es trat keine Schiefe auf, die durch schnelles Schalten erzeugt würde.Die Gaußverteilungen werden dann zu Beta-Verteilungen (FitzHugh, 1983; Yellen, 1984).
2. Es könnte noch möglich sein, daß die Ratenkonstanten zu niedrig geschätzt wurden, weildie Dwell-Time-Analyse ohne missed-events-Korrektur durchgeführt wurde. Doch geradedie K+ + Tl+-Daten bei 20°C (Tab. 7.2) weichen im schnellen Bereich kaum von den mitdem HMM-Fit bestimmten ab, der weniger empfindlich gegenüber missed-events ist. ImFalle schnellen Schaltens müßten die Amplitudenverteilungen pro Niveau aus einerZerlegung von Beta-Verteilungen erhalten werden. Doch da hier keine Beta-Verteilungenvorliegen, erfüllt die im Programm implementierte Gaußzerlegung diese Aufgabezufriedenstellend.
3. Die Ratenkonstanten zeigen, wie schon oben erwähnt, kein inverses Verhalten (Zanellound Barrantes, 1994) wie es bei Messungen, in denen die Ratenkonstanten zu dicht an derAuflösungsgrenze liegen, vorkommt (Draber und Schultze, 1994a).
Nachdem die Amplitudenhistogramme in Fig. 7.17 die Hypothese der Reduzierung desscheinbaren Einzelkanalstroms durch schnelles Schalten widerlegt haben, ist zu fragen, wasdie Stromreduzierung in Tab. 7.1 bedeutet: Autoren, die mit langsamerenAufzeichnungsanlagen arbeiten, würden über die Burstbereiche mitteln und die in Tab. 7.1berechnete Reduzierung des Einzelkanalstromes zusätzlich zu den hier bei schnellenAbtastraten gefundenen messen.
7.3.9 Untersuchung der Offenwahrscheinlichkeiten
Durch den Hinkley-Detektor werden nicht nur die Dwell-Time-Histogramme erhalten,sondern auch die Mehrkanal-Offenwahrscheinlichkeiten. Sie entstehen aus der Summe derZeiten, in denen eine feste Anzahl von Kanälen gleichzeitig offen ist, und durch Divisiondurch die Gesamtzeit der analysierten Aufzeichnung.
Da die Wahrscheinlichkeitsverteilung der Anzahl der offenen Kanäle bei einemMehrkanal-Patch über die Binomialverteilung (Glasbey und Martin, 1988)
( ) kopen
kNopen pp
kN
kp −−
= 1)( (7.12)
mit der Offenwahrscheinlichkeit des Einzelkanals popen zusammenhängt, ergibt sich hier einezweite Möglichkeit (neben der kinetischen Analyse), die popen zu berechnen.
Hierbei ist N die Anzahl der Kanäle, k die Anzahl der offenen Kanäle, popen dieWahrscheinlichkeit, daß ein Kanal offen ist, und p(k) die Wahrscheinlichkeit, daß k-Kanäleoffen sind. Diese Gleichung wird dann mit Hilfe des Simplex-Algorithmus (Nedler und Mead,1965; Caceci und Cacheris, 1984) nach popen aufgelöst.
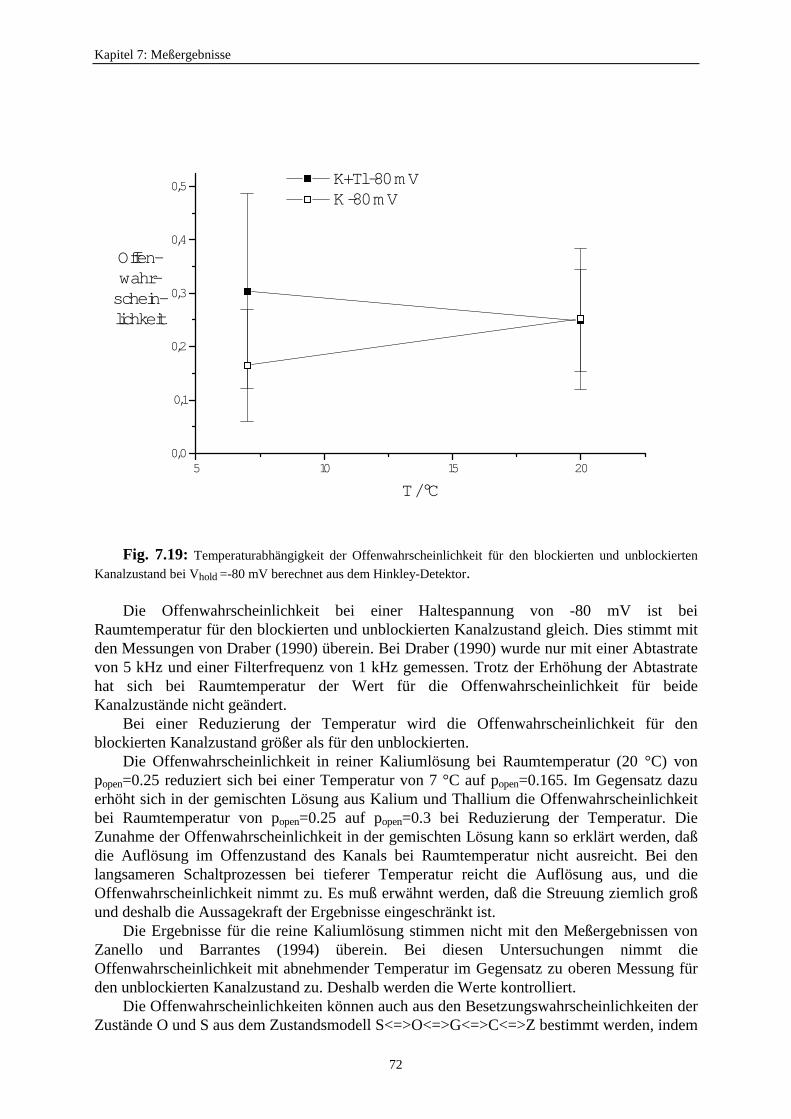
Kapitel 7: Meßergebnisse
72
5 10 15 200,0
0,1
0,2
0,3
0,4
0,5 K+Tl -80 mV K -80 mV
Offen-wahr-schein-lichkeit
T / °C
Fig. 7.19: Temperaturabhängigkeit der Offenwahrscheinlichkeit für den blockierten und unblockiertenKanalzustand bei Vhold =-80 mV berechnet aus dem Hinkley-Detektor.
Die Offenwahrscheinlichkeit bei einer Haltespannung von -80 mV ist beiRaumtemperatur für den blockierten und unblockierten Kanalzustand gleich. Dies stimmt mitden Messungen von Draber (1990) überein. Bei Draber (1990) wurde nur mit einer Abtastratevon 5 kHz und einer Filterfrequenz von 1 kHz gemessen. Trotz der Erhöhung der Abtastratehat sich bei Raumtemperatur der Wert für die Offenwahrscheinlichkeit für beideKanalzustände nicht geändert.
Bei einer Reduzierung der Temperatur wird die Offenwahrscheinlichkeit für denblockierten Kanalzustand größer als für den unblockierten.
Die Offenwahrscheinlichkeit in reiner Kaliumlösung bei Raumtemperatur (20 °C) vonpopen=0.25 reduziert sich bei einer Temperatur von 7 °C auf popen=0.165. Im Gegensatz dazuerhöht sich in der gemischten Lösung aus Kalium und Thallium die Offenwahrscheinlichkeitbei Raumtemperatur von popen=0.25 auf popen=0.3 bei Reduzierung der Temperatur. DieZunahme der Offenwahrscheinlichkeit in der gemischten Lösung kann so erklärt werden, daßdie Auflösung im Offenzustand des Kanals bei Raumtemperatur nicht ausreicht. Bei denlangsameren Schaltprozessen bei tieferer Temperatur reicht die Auflösung aus, und dieOffenwahrscheinlichkeit nimmt zu. Es muß erwähnt werden, daß die Streuung ziemlich großund deshalb die Aussagekraft der Ergebnisse eingeschränkt ist.
Die Ergebnisse für die reine Kaliumlösung stimmen nicht mit den Meßergebnissen vonZanello und Barrantes (1994) überein. Bei diesen Untersuchungen nimmt dieOffenwahrscheinlichkeit mit abnehmender Temperatur im Gegensatz zu oberen Messung fürden unblockierten Kanalzustand zu. Deshalb werden die Werte kontrolliert.
Die Offenwahrscheinlichkeiten können auch aus den Besetzungswahrscheinlichkeiten derZustände O und S aus dem Zustandsmodell S<=>O<=>G<=>C<=>Z bestimmt werden, indem
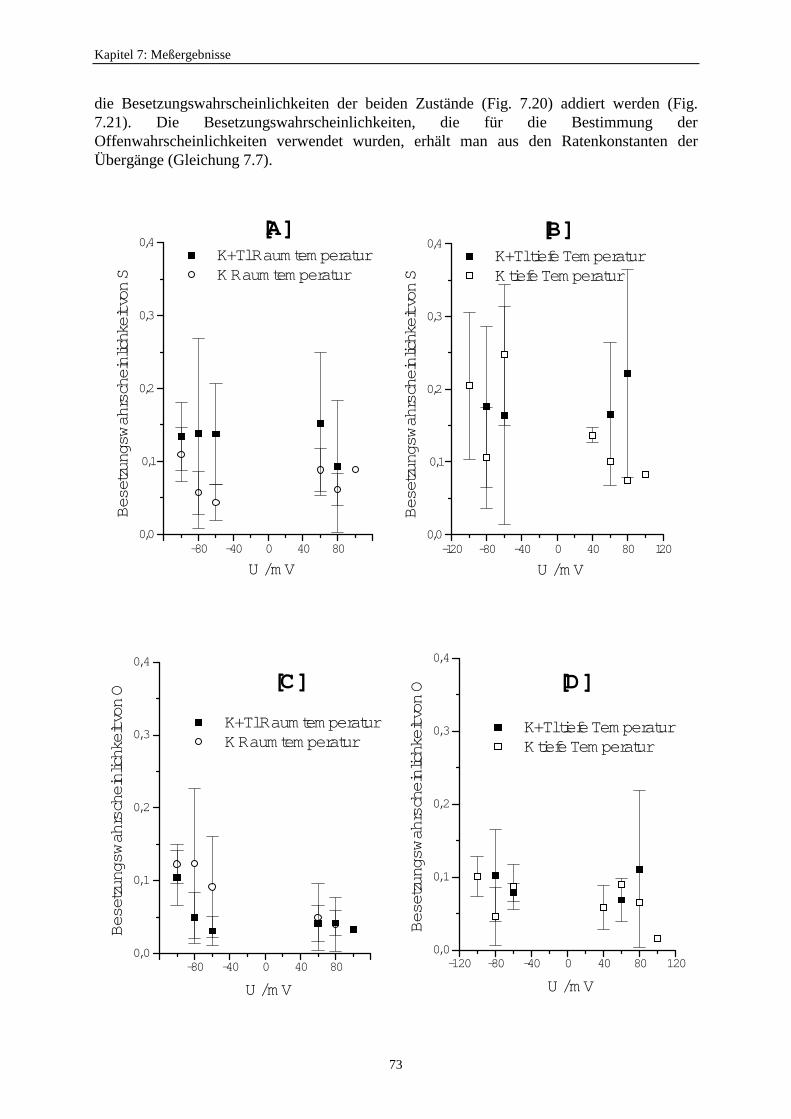
Kapitel 7: Meßergebnisse
73
die Besetzungswahrscheinlichkeiten der beiden Zustände (Fig. 7.20) addiert werden (Fig.7.21). Die Besetzungswahrscheinlichkeiten, die für die Bestimmung derOffenwahrscheinlichkeiten verwendet wurden, erhält man aus den Ratenkonstanten derÜbergänge (Gleichung 7.7).
-120 -80 -40 0 40 80 1200,0
0,1
0,2
0,3
0,4[B]
K+Tl tiefe Tem peratur K tiefe Tem peratur
Besetzungswahrscheinlichkeit von S
U / m V-80 -40 0 40 80
0,0
0,1
0,2
0,3
0,4[A]
K+Tl Raum tem peratur K Raum tem peratur
Besetzungswahrscheinlichkeit von S
U / m V
-80 -40 0 40 800,0
0,1
0,2
0,3
0,4
K+Tl Raum tem peratur K Raum tem peratur
Besetzungswahrscheinlichkeit von O
U / m V
-120 -80 -40 0 40 80 1200,0
0,1
0,2
0,3
0,4
[D][C]
K+Tl tiefe Tem peratur K tiefe Tem peratur
Besetzungswahrscheinlichkeit von O
U / m V
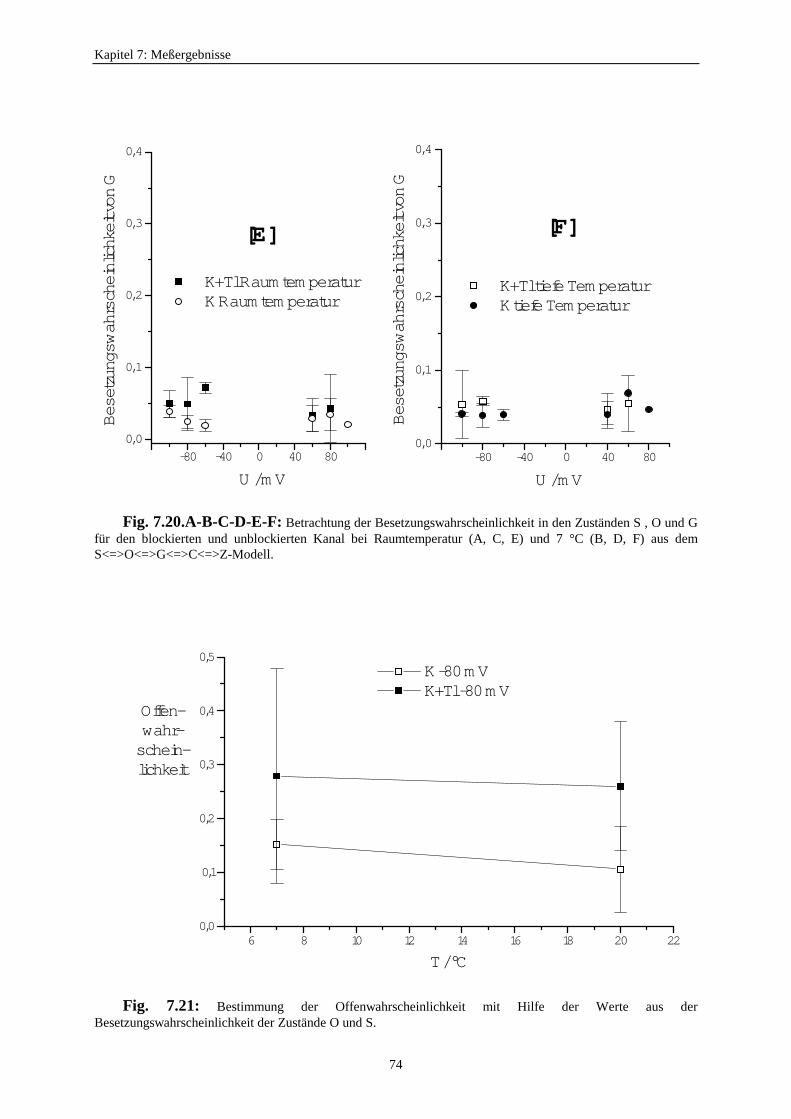
Kapitel 7: Meßergebnisse
74
-80 -40 0 40 80
0,0
0,1
0,2
0,3
0,4
[E]
K+Tl Raum tem peratur K Raum tem peratur
Besetzungswahrscheinlichkeit von G
U / m V
-80 -40 0 40 800,0
0,1
0,2
0,3
0,4
[F]
K+Tl tiefe Tem peratur K tiefe Tem peratur
Besetzungswahrscheinlichkeit von G
U / m V
Fig. 7.20.A-B-C-D-E-F: Betrachtung der Besetzungswahrscheinlichkeit in den Zuständen S , O und Gfür den blockierten und unblockierten Kanal bei Raumtemperatur (A, C, E) und 7 °C (B, D, F) aus demS<=>O<=>G<=>C<=>Z-Modell.
6 8 10 12 14 16 18 20 220,0
0,1
0,2
0,3
0,4
0,5 K -80 m V K+Tl -80 m V
Offen-wahr-schein-lichkeit
T / °C
Fig. 7.21: Bestimmung der Offenwahrscheinlichkeit mit Hilfe der Werte aus derBesetzungswahrscheinlichkeit der Zustände O und S.
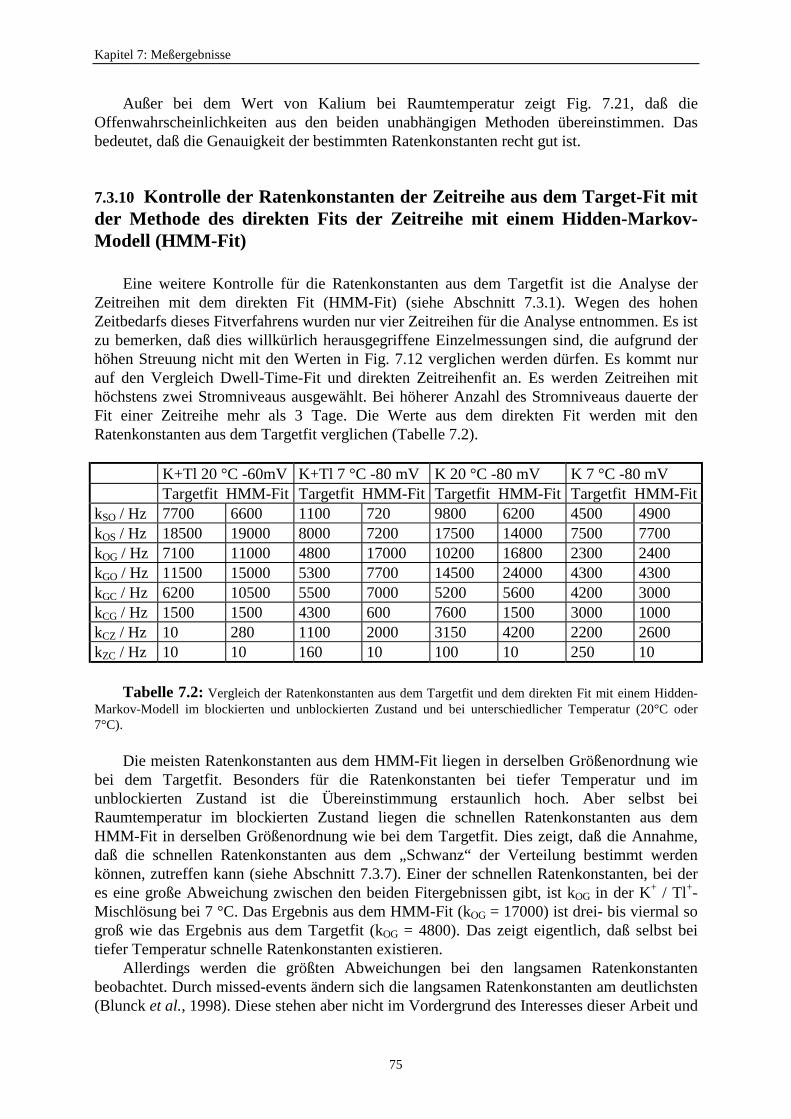
Kapitel 7: Meßergebnisse
75
Außer bei dem Wert von Kalium bei Raumtemperatur zeigt Fig. 7.21, daß dieOffenwahrscheinlichkeiten aus den beiden unabhängigen Methoden übereinstimmen. Dasbedeutet, daß die Genauigkeit der bestimmten Ratenkonstanten recht gut ist.
7.3.10 Kontrolle der Ratenkonstanten der Zeitreihe aus dem Target-Fit mitder Methode des direkten Fits der Zeitreihe mit einem Hidden-Markov-Modell (HMM-Fit)
Eine weitere Kontrolle für die Ratenkonstanten aus dem Targetfit ist die Analyse derZeitreihen mit dem direkten Fit (HMM-Fit) (siehe Abschnitt 7.3.1). Wegen des hohenZeitbedarfs dieses Fitverfahrens wurden nur vier Zeitreihen für die Analyse entnommen. Es istzu bemerken, daß dies willkürlich herausgegriffene Einzelmessungen sind, die aufgrund derhöhen Streuung nicht mit den Werten in Fig. 7.12 verglichen werden dürfen. Es kommt nurauf den Vergleich Dwell-Time-Fit und direkten Zeitreihenfit an. Es werden Zeitreihen mithöchstens zwei Stromniveaus ausgewählt. Bei höherer Anzahl des Stromniveaus dauerte derFit einer Zeitreihe mehr als 3 Tage. Die Werte aus dem direkten Fit werden mit denRatenkonstanten aus dem Targetfit verglichen (Tabelle 7.2).
K+Tl 20 °C -60mV K+Tl 7 °C -80 mV K 20 °C -80 mV K 7 °C -80 mVTargetfit HMM-Fit Targetfit HMM-Fit Targetfit HMM-Fit Targetfit HMM-Fit
kSO / Hz 7700 6600 1100 720 9800 6200 4500 4900kOS / Hz 18500 19000 8000 7200 17500 14000 7500 7700kOG / Hz 7100 11000 4800 17000 10200 16800 2300 2400kGO / Hz 11500 15000 5300 7700 14500 24000 4300 4300kGC / Hz 6200 10500 5500 7000 5200 5600 4200 3000kCG / Hz 1500 1500 4300 600 7600 1500 3000 1000kCZ / Hz 10 280 1100 2000 3150 4200 2200 2600kZC / Hz 10 10 160 10 100 10 250 10
Tabelle 7.2: Vergleich der Ratenkonstanten aus dem Targetfit und dem direkten Fit mit einem Hidden-Markov-Modell im blockierten und unblockierten Zustand und bei unterschiedlicher Temperatur (20°C oder7°C).
Die meisten Ratenkonstanten aus dem HMM-Fit liegen in derselben Größenordnung wiebei dem Targetfit. Besonders für die Ratenkonstanten bei tiefer Temperatur und imunblockierten Zustand ist die Übereinstimmung erstaunlich hoch. Aber selbst beiRaumtemperatur im blockierten Zustand liegen die schnellen Ratenkonstanten aus demHMM-Fit in derselben Größenordnung wie bei dem Targetfit. Dies zeigt, daß die Annahme,daß die schnellen Ratenkonstanten aus dem „Schwanz“ der Verteilung bestimmt werdenkönnen, zutreffen kann (siehe Abschnitt 7.3.7). Einer der schnellen Ratenkonstanten, bei deres eine große Abweichung zwischen den beiden Fitergebnissen gibt, ist kOG in der K+ / Tl+-Mischlösung bei 7 °C. Das Ergebnis aus dem HMM-Fit (kOG = 17000) ist drei- bis viermal sogroß wie das Ergebnis aus dem Targetfit (kOG = 4800). Das zeigt eigentlich, daß selbst beitiefer Temperatur schnelle Ratenkonstanten existieren.
Allerdings werden die größten Abweichungen bei den langsamen Ratenkonstantenbeobachtet. Durch missed-events ändern sich die langsamen Ratenkonstanten am deutlichsten(Blunck et al., 1998). Diese stehen aber nicht im Vordergrund des Interesses dieser Arbeit und

Kapitel 7: Meßergebnisse
76
das vorliegende Detektormaterial reicht auch nicht zur statistischen Sicherheit. Somit wird derEffekt nicht weiter untersucht.

8 Fazit und AusblickDies ist die vierte Arbeit, in der sich die Kieler Arbeitsgruppe mit dem Thema des
anomalen Molfraktionseffekts (aus K+- und Tl+-Mischung) beschäftigt. Das Ziel allerArbeiten war es, zu entscheiden, ob dem AMFE ein Permeations- oder Gatingmechanismuszugrunde liegt (Kapitel 4). Während der Nachweis der entscheidenden Rolle des Gatings beimBlock durch Cs+ (Draber und Hansen, 1994a) und Na+ (Blunck, 1996) gelang, konnte eineEntscheidung beim AMFE in den drei Vorgängerarbeiten noch nicht erreicht werden.
Nun ist eine Bilanz zu ziehen, wie der Stand am Ende dieser Arbeit ist. Ein Hinweis, daßder anomale Molfraktionseffekt durch schnelles Schalten verursacht wird, ergab sich aus denStrom-Spannungskurven bei tiefer Temperatur. Die Konvergenz der beiden Strom-Spannungskurven bei 7 °C unterstützt diese Hypothese.
Das Endziel ist jedoch der direkte Nachweis des schnellen Schaltens in kinetischenUntersuchungen. Im Gegensatz zu den Vorgängerarbeiten stand jetzt das neue Programm vonBlunck (1996) und Kirst (1997) zur Auswertung von Mehrkanal-Dwell-Time-Histogrammenzur Verfügung.
Die Untersuchungen auf der Grundlage des Markov-Modells S<=>O<=>G<=>C<=>Zerbrachte dann die Überraschung. Stellt man sich zuerst einmal dumm, indem man vergißt,daß die Niveaubestimmung von der gleichen zeitlichen Auflösung wie die Bestimmung derRatenkonstanten ausgeht, so ergeben die gemessenen Ratenkonstanten bei Raumtemperatur inTl+-haltiger Lösung tatsächlich durch Mittelung über die Burstbereiche eine scheinbareStromreduzierung. Sie beträgt zwar nur 10% statt der gemessenen 25%, aber in Anbetrachtder Fehler konnte man erst einmal zufrieden sein.
Doch diese Betrachtung hat nur Bedeutung für Wissenschaftler, die mit niedrigerenAbtastraten arbeiten. Ihnen könnte man aufgrund der Analyse hier sagen: 10% Eurergemessenen Stromreduzierung geht auf Kosten der Mittelung über schnelles Schalten.
Für die hier durchgeführte Analyse ist der in Abschnitt 7.3.8 beschrittene Weg der einzigrichtige. Man nimmt die Ratenkonstanten bei Tl+-Anwesenheit bei Raumtemperatur, steckt siein das Simulationsprogramm, überlagert die Zeitreihe mit dem angemessenen Rauschen undfiltert sie mit einem 4-poligen Besseltiefpaß wie im Versuch. Da wird untersucht, ob dieAnalyse den bei der Simulation benutzten Einzelkanalstrom von 6 pA wiedergibt. Selbst beiBeschränkung auf das kleine Burstmodell (S<=>O<=>G) verändert das schnelle Schalten denaus den Amplitudenhistogrammen gefundenen Strom nur auf 5.88 pA. Dieser Effekt von 2%liegt im Streubereich der Auswertung realer Daten. Er ist zu vernachlässigen gegenüber den25%, die als Stromreduzierung beim AMFE gemessen werden.
Dieses Ergebnis ist ein Meilenstein in der Geschichte des AMFEs im Kieler Labor. DieSuche nach schnellem Schalten begann mit der Diplomarbeit von Draber (1990), führte zugewaltigen Verbesserungen der Aufzeichnungsanlagen (Albertsen, 1992) und zurEntwicklung neuer effektiver Auswertealgorithmen (Schultze und Draber, 1993; Draber undSchultze, 1994a; Albertsen und Hansen, 1994; Blunck, 1996; Kirst, 1997) und nun ist aufeinen Schlag alles vorbei: Kein schnelles Schalten als Ursache.

Kapitel 8: Fazit und Ausblick
78
Die Frage ist, wie sicher obiges Ergebnis ist. Man könnte natürlich einwenden, es gäbenoch ein schnelles Schalten, das hier nicht entdeckt worden ist. Die Transferrate des Kanalsbei 6 pA ist 108 Ionen/s. Die schnellen Ratenkonstanten liegen bei 15 000 Hz. In diesemBereich mit einem Faktor 6000 kann sich noch einiges verbergen. Doch zur Zeit besteht keineHoffnung, etwas zu finden. Die Amplitudenverteilung der gemessenen Daten (Fig. 7.3) läßtsich gut mit Gaußhügeln nähern. Prozesse mit Zeitkonstanten, die um den Faktor 10 höherliegen, würden bereits sehr deutliche Abweichungen erzeugen und zu Beta-Verteilungenführen. Also muß ein hypothetisches Schalten schnellere Zeitkonstanten als ca. 5 µs,wahrscheinlich sogar 1 µs besitzen.
Dieser Bereich ist aber nur durch einen Technologiesprung zu erreichen, der sich imMoment nicht abzeichnet. Somit wird das Ergebnis dieser Arbeit für die nächste Zeit die letzteAussage zum schnellen Schalten beim AMFE sein.
In der Zukunft wird man sich also wieder stärker den Permeationsmodellen zuwenden.Hierfür ist wichtig, daß die symmetrischen Effekte auf die Strom-Spannungskurven, die vonDraber et al. (1991) und Keunecke (1995) gemessen wurden, hier nicht auftraten. DieSymmetrie der Wirkung des AMFEs auf die Strom-Spannungskurven im positiven undnegativen Spannungsbereich war das Hauptargument, das Permeationsmodell von Hille-Schwarz (1978) und Wu (1991,1992) auszuschließen und nach Gating-Modellen zu suchen.Albertsen (1994) und diese Arbeit fanden nur einen einseitigen Effekt.
Es ist nicht einsichtig, warum es zu diesen unterschiedlichen Ergebnissen kam.Insbesondere die Messungen von Keunecke (1995) und die Messungen dieser Arbeit fandenunter fast identischen Bedingungen statt. Die erkennbaren Unterschiede waren wie folgt:Höhere Osmolarität in den Messungen hier (250 mM KNO3) im Gegensatz zu 150 mM KNO3bei Keunecke (1995). Doch erste Messungen in dieser Arbeit mit 150 mM zeigten auchkeinen symmetrischen AMFE. Der einzige Unterschied ist, daß Keunecke einenLösungsaustausch durchführte und in der Kammer die Lösung wechselte, während hier fürjedes Mischungsverhältnis ein neuer Patch genommen wurde. Doch wie das den Unterschiederklären sollte, ist unverständlich.
Diese Symmetriefrage hat nicht nur Bedeutung für eine mögliche Erklärung mit den obengenannten Permeationsmodellen, sondern auch für die Art der Bindungsstelle. Die hiergefundene Erklärung, daß die symmetrische Wirkung nur auftritt, wenn die Lösungensymmetrisch sind, paßt gut zur Vorstellung, daß der Strom das Tl+-Ion in den Kanal zieht unddort die wirksame Bindungsstelle trifft. Außerdem hatte sich bei anderen Ionenblocks (Na+
Blunck, 1996; Cs+, Draber, 1994) gezeigt, daß die Wirkung einseitig war.Für die symmetrischen Wirkungen nahmen Draber et al. (1991) an, daß die
Bindungsstelle an der Außenmembran sitzt und das einmal gebundene Ion den Fluß in beideRichtungen beeinflußt. Keunecke (1995) benutzte inside-out-patches und Tl+-Lösung. Dannhätte die Bindungsstelle auf der cytosolischen Außenseite des Kanals liegen müssen, um densymmetrischen Effekt zu erklären.
Aufgrund der vorliegenden Ergebnisse fallen Argumente gegen das Modell von Wu(1991, 1992) und Hille-Schwarz (1978) fort. Die Modelle müßten überarbeitet werden, umeinige kleinere Widersprüche aufzulösen. Ein neuer Ansatz ist das Modell von Möller (1998),der in seiner Diplomarbeit eine Kombination aus Hille-Schwarz und dem enzymkinetischemModell (Hansen et al., 1981) vorgeschlagen hat. Es ist aber noch nicht auf den AMFEangewandt worden.
Es zeichnet sich also ab, daß die Reihe von AMFE-Arbeiten so bald nicht zu Ende ist.Hierbei ist auch zu klären, warum die Strom-Spannungskurven mit und ohne Tl+ bei 7°Ckonvergieren. Dies könnte auch den entsprechenden Befund von Blunck (1996) beim Na+-Block in anderem Licht erscheinen lassen.

9 ZusammenfassungIn dieser Arbeit wurde der anomale Molfraktionseffekt (AMFE) beim Kaliumkanal von
Tonoplasten der Grünalge Chara corallina mit Hilfe der Patch-Clamp-Technik untersucht.Unter dem AMFE versteht man, daß bei einer konstanten Gesamtkonzentration dieLeitfähigkeit der Mischlösung (K+- und Tl+-Ionen) kleiner ist als die der reinen Lösung. DieAufgabe bestand darin, zu untersuchen, ob der anomale Molfraktionseffekt durch die nichtausreichende Auflösung des Meßsystems entsteht, d.h., ob das schnelle Schalten im Anti-Aliasingfilter summiert wird und für die scheinbare Reduktion des Stromes verantwortlich ist.Deshalb wird mit Abtastraten von 100 kHz und mit einem Anti-Aliasingfilter von 25 kHzgearbeitet. Dazu wurden Strom-Spannungskurven in Abwesenheit und Gegenwart von 20 mMTlNO3 (bei 250 bzw. 230 mM KNO3) gemessen. Es wird bei dem blockierten Zustand trotzder hohen Abtastraten eine Reduktion der Leitfähigkeit um 25 % beobachtet.
Es wurden Messungen bei niedriger Temperatur (7°C) durchgeführt, um dieReaktionsgeschwindigkeit der Schaltvorgänge zu reduzieren. Die Abkühlung erfolgte durchdie direkte Kühlung des Objekttisches. Bei tiefer Temperatur (7 °C) konvergieren die beidenStröme des blockierten und unblockierten Zustands. Dieser Effekt wurde bei Untersuchungendes Na+-Blockes als ein wichtiger Hinweis auf die Beteiligung schnellen Schaltens an derEinzelkanalstromreduzierung gewertet.
Um dies zu überprüfen, wurden die Daten mit einem Hinkley-Detektor höherer Ordnunganalysiert, um mit Hilfe von Dwell-Time-Histogrammen das kinetische Verhalten des Kanalszu studieren. Der Detektor ist in der Lage, eine zeitliche Auflösung bis zu 40 µs für eineSprungdetektion zu erreichen.
Für eine stochastische Betrachtung der Schaltvorgänge der Kanäle wird ein Markov-Modell benutzt. Die Wahrscheinlichkeitsverteilung für die Verweildauern auf einem Niveauwird durch ein System von linearen Differentialgleichungen erster Ordnung beschrieben. DieGleichung läßt sich durch eine Summe von Exponentialfunktionen lösen.
Der neue Target-Fit aus Vorgängerarbeiten von Blunck (1996) und Kirst (1997)ermöglicht, durch Fitten der Dwell-Time-Histogramme die Ratenkonstanten deszugrundeliegenden Markov-Modells des Kanals zu ermitteln.
Die Wahl des geeigneten Markov-Modells ist der erste Schritt der Analyse. Die Analyseder Dwell-Time-Histogramme ergab, daß zwei Offenzustände und drei Geschlossenzuständefür den Target-Fit benötigt werden. Durch den Vergleich unterschiedlicher 5-Zustandsmodellewurde das beste Fitergebnis durch das S<=>O<=>G<=>C<=>Z-Modell (C, G und Zgeschlossen, O und S offen) erreicht. Die Pascal-bedingte Beschränkung des Adreßraumesließ den Fit mit dem vorliegenden Programm von Aufzeichnungen bis zu drei Kanälen zu.
Als nächstes wurden die Übergangsratenkonstanten kij in diesem Modell beiunterschiedlichen Temperaturen miteinander verglichen. Bei den Ratenkonstanten wurde keininverses Verhalten beobachtet. Inverses Verhalten bedeutet, daß keine der Ratenkonstantensich mit sinkender Temperatur steigert, was ein Hinweis auf ein falsches Modell oder einenicht ausreichende Zeitauflösung wäre. Bei einem inversen Verhalten wäre es notwendig

Kapitel 9: Zusammenfassung
80
gewesen, das gewählte Modell in Frage zu stellen oder eine missed-events-Korrektureinzuführen, da die Auflösung wahrscheinlich nicht ausreicht. Der größte Unterschied beiAufzeichnungen mit und ohne Thallium zeigte sich im Übergang O<=>G und bestan darin,daß der G-Zustand bei Anwesenheit von Tl+ länger wurde.
Die so ermittelten Ratenkonstanten wurden benutzt, um Zeitreihen zu simulieren. Zu denmit dem S<=>O<=>G<=>C<=>Z-Modell simulierten Zeitreihen wurde Rauschen mit SNR =4 addiert. Die Gewichtung mit einem 4-poligen Besselfilter (25 kHz) und die Abtastung mit100 kHz vervollkommnete die Korrespondenz zu den echten Zeitreihen. DerEinzelkanalstrom wurde bei der Simulation entsprechend den Messungen an Chara mit 6 pAfestgesetzt.
Amplitudenhistogramme der simulierten Zeitreihen dienten dem Studium der Wirkungdes Schaltens auf den gemessenen Einzelkanalstrom. Es ergab sich kein sichtbarer Effekt beider Simulation des vollständigen Modells. Die Beschränkung auf den Burstbereich durchSelektion des Teilmodells S<=>O<=>G mit den Ratenkonstanten der Tl+-Messungen beiRaumtemperatur erbrachte eine Reduzierung des gemessenen Einzelkanalstroms auf 5.88 pA.Dieser Effekt von 2% wäre in der Analyse realer Daten nicht detektierbar.
Das Ergebnis der Analyse ist damit eindeutig: Die Reduktion des Einzelkanalstromes imBereich des AMFE ist nicht auf schnelles Schalten zurückzuführen. Es wird diskutiert, ob esnoch unentdecktes schnelles Schalten geben könnte. Da die Amplitudenverteilungen derechten Daten keine signifikanten Abweichungen von Gaußverteilungen zeigen, ist zuschließen, daß solch ein hypothetisches schnelles Schalten weit über 200 kHz liegen müßteund mit der heutigen Technologie nicht zu verwirklichen ist.
Bevor dieser endgültige Schluß dargestellt wird, beschäftigte sich die Arbeit mit derFrage, ob bei Untersuchungen mit geringerer Zeitauflösung als der hier benutzten eineStromreduzierung durch schnelles Schalten auftreten könnte. Dazu wird wieder das für Burstsverantwortliche S<=>O<=>G Teilmodell betrachtet. Die Mittelung über die Offen- undGeschlossenzeiten in einem solchen Burst ergibt tatsächlich eine 10-prozentigeStromreduzierung bei Anwesenheit von Tl+ bei Raumtemperatur. Das bedeutet, daßMessungen mit langsameren Aufzeichnungsanlagen einen scheinbaren AMFE messenwürden, der um 10% höher liegt als der in diesen Messungen auftretende.
Als Kontrolle für die Dwell-Time-Analyse wurde mit Hilfe der Mehrkanal-Offenwahrscheinlichkeit des Hinkley-Detektors die Offenwahrscheinlichkeit eines Kanalsüber eine Binomialverteilung berechnet. Die Ergebnisse bei Raumtemperatur stimmen mit denErgebnissen von Draber et al. (1991) trotz der großen Streuung überein. Trotz der besserenAuflösung verglichen mit der von Draber et al. (1991) tritt keine Veränderung derOffenwahrscheinlichkeit bei Raumtemperatur für den blockierten und unblockierten Zustandauf. Die Offenwahrscheinlichkeit steigt mit Abnahme der Temperatur.
Die Offenwahrscheinlichkeit eines Kanals kann auch durch die Addition derBesetzungswahrscheinlichkeiten von Zustand S und O bestimmt werden, die sich aus derkinetischen Analyse (Dwell-Time-Fit) ergeben. Da die Besetzungswahrscheinlichkeiten ausden Ratenkonstanten bestimmt werden, wird sie auch als Kontrolle für die Ratenkonstantenverwendet. Weil die Werte aus den beiden unabhängigen Methoden größenordnungsmäßigübereinstimmen, ist dies ein Hinweis für die recht gute Bestimmung der Ratenkonstanten.
Außerdem wurde bei einigen Messungen ein direkter Zeitreihenfit mit dem Hidden-Markov-Modell (S<=>O<=>G<=>C<=>Z) durchgeführt. Die Ergebnisse der schnellenRatenkonstanten stimmen hier erstaunlich gut mit dem Target-Fit überein.

LiteraturverzeichnisAiyar, J., Withka, J. M., Rizzi, J. P., Singleton, D. H., Andrews, G. C., Lin, W., Boyd, J.,Hanson, D. C., Simon, M., Detlefs, B., Lee, C., Hall, J. E., Gutman, G. A., Chandy, K. G.(1995). Topology of the pore-region of a K+ channel revealed by the NMR-derived structures ofscorpion toxins. Neuron 15: 1169-1181.
Albertsen, A. (1992). Erfassung und Auswertung schneller Schaltvorgänge bei Patch-ClampUntersuchung. Diplomarbeit, Kiel.
Albertsen, A. (1994). Fast patch-clamp data acquistion and kinetic analysis of multi-channelrecords. Doktorarbeit, Kiel.
Albertsen, A., Hansen, U.-P. (1994). Estimation of kinetic rate constants from multi-channelrecordings by a direct fit of the time serie. Biophysical J. 67:1393-1403.
Baukrowitz, T., Yellen, G. (1996). Use-dependent blockers and exit rate of the last ion from amulti-ion pore of a K+ channel. Science 271: 653-656.
Bertl, A. (1989). Current-voltage relationships of a sodium-sensitive potassium channel in thetonoplast of Chara corallina. J. Membr. Biol. 109: 9-19.
Bertl, A., Gradmann, D., Slayman, C. L. (1992a). Calcium- and voltage-dependent ion channelin Saccharomyces cerevisiae. Philos. Trans. R. Soc. Lond. B 338: 63-72.
Bertl, A., Blumwald, E., Coronado, R. Eisenberg, R., Findlay, G., Gradmann, D., Hille, B.,Köhler, K., Kolb, H.-A., MacRobbie, E., Meissner, G., Miller, C., Neher, E., Palade, P.,Pantoja, O., Sanders, D., Schroeder, J., Slayman, C. L., Spanswick, R., Williams, A. (1992b).Electrical measurements on endomembrans. Science 258: 873-874.
Bertl, A., Slayman, C. L., Gradmann, D. (1993). Gating and conductance in an outward-rectifying K+ channel from the plasma membrane of Saccharomyces cerevisiae. J. MembraneBiol. 132: 183-199.
Blunck, R. (1996). Nachweis des schnellen Schaltens beim Na+-Block des K+-Kanals inChara. Diplomarbeit, Kiel.
Blunck, R., Kirst, U., Rießner, T., Hansen, U.P., (1998). How powerful is the dwell-timeanalysis of multi-channel records? submitted.

Literaturverzeichnis
82
Bowman, B. J., Bowman, E. J. (1986). H+ -ATPases from mitochondria, plasma membranes,and vacuoles of fungal cells. J. Membr. Biol. 94: 83-97.
Caceci, M. S., Cacheris, W. P. (1984). Fitting curves to data - the simplex algorithm is theanswer. BYTE 5/84: 340-362.
Catterall, W. A. (1992). Cellular and molecular biology of voltage-gated sodium channels.Physiol. Rev. 72: S15-S48.
Colquhoun, D., Hawkes, A. G. (1977). Relaxation and fluctuations of membrane currents thatflow through drug-operated channels. Proc. R. Soc. Lond. B 199: 231-262.
Colquhoun, D., Hawkes, A.G., Srodzinski, K. (1996). Joint distributions of apparent open timesand shut times of single ion channels and the maximum likelihood fitting of mechanisms. Phil.Transactions of the Royal Society of London A 354, 2555-1290.
Corey, D. P., Stevens, C. F. (1983). Science and technology of patch-recording electrodes. In:Single Channel Recording. Sakmann, B. and Neher, E., editor. Plenum Press, New York andLondon. 53-68.
Crouzy, S. C., Sigworth, F. J. (1990). Yet another approach to the dwell-time omissionproblem. Biophysical J. 58: 731-743.
De Biasi, M., Drewe, J. A., Kirsch, G. E., Brown, A. M. (1993). Histidin substitution identifies asurface position and confers Cs2+ selectivity on a K+ pore. Biophys. J. 65: 1235-1242.
Draber, S (1990). Untersuchung des anomalen Molfraktionseffektes mit Hilfe der Patch-Clamp Technik. Diplomarbeit, Kiel.
Draber, S. (1994). Methods for Analyzing Fast Single-Channel Data and Studies on the K+
Channel under Cs+ Blockade and under Control Conditions. PhD Thesis, Kiel
Draber, S., Schultze, R., Hansen, U.-P. (1991). Patch-Clamp studies on the anomalous molefraction effect of the K+ channel of cytoplasmic dropletsof Nitella: An attempt to distinguishbetween a multi-ion single-file pore and an enzyme kinetic model with lazy state. J.Membrane Biol. 123: 183-190.
Draber, S., Hansen, U.-P. (1994). Fast single-channel measurements resolve the effects of Cs+
on the K+ channel. Biophysical J. 67: 120-129.
Draber, S., Schultze, R. (1994a). Correction for missed events based on a realistic model of adetector. Biophys. J. 66: 191-201.
Draber, S., Schultze, R. (1994b). Detection of jumps in single-channel data containingsubductance levels. Biophysical J. 67: 1404-1413.
Eisenberg, R. S. (1990). Channels as enzymes. J. Membrane Biol. 115: 1-12.
FitzHugh, R. (1983). Statistical properties of the asymmetric random telegraph signal withapplication to single channel analysis. Mathematical Bioscience 64: 75-89.

Literaturverzeichnis
83
Fredkin, D. R., Rice, J. A. (1992). Maximum Likelihood estimation and identification directlyfrom single-channel recording. Proc R. Soc. Lond. B 249: 125-132.
Glasbey C. A., Martin, R. J. (1988). The distribution of numbers of open channels in multi-channels patches. J. Neurosci. Methods 24: 283-287.
Jackson, M. B. (1997). Inversion of Markov processes to determine rate constants from singlechannel data. Biophys. J. 73: 1382-1394.
Jan, L. Y., Jan, Y. N. (1992). Structural elements involved in specific K+ channel functions.Annu. Rev. Physiol. 54: 537-555.
Hagiwara, S., Miyazaki, S., Krasne, S., Ciani, S. (1977). Anomalous permeabilities of the eggcell membrane of a starfish in K+-Tl+ Mixtures. J. Gen. Physiol. 70: 269-281.
Hansen, U. P., Gradmann, D., Sanders, D., Slayman, C. L. (1981). Interpretation of current-voltage relationships for “activ” ion transport systems: I. Steady-state reaction-kinetic analysisof class-I mechanisms. J. Membrane Biol. 63: 165-190.
Hansen, U.-P.,Tittor, J., Gradmann, D. (1983). Interpretation of current-voltage relationshipsfor “activ” ion transport systems: II Nonsteady-state reaction-kinetic analysis of class-Imechanisms with one slow time-constant. J. Membrane Biol. 75: 141-169.
Hansen, U.-P., Keunecke, M., Blunck R., (1997). Gating and permeation models of plantchannels. J. of Experimentel Botany, 48: 365-382.
Hille, B. (1975). Ionic selectivity, saturation and block in sodium channels. A four barrier model. J. Gen. Physiol. 66: 535-560.
Hille, B. (1992). Ionic channels of excitable membranes. Sinauer Associates Inc., Sunderland,MA. 607 pp.
Hille, B., Schwarz, W. (1978). Potassium channels as multi-ion-single-file pores. J. Gen. Biol.72: 409-442.
Horn, R. A., Lange, K. (1983). Estimating kinetic constants from single channel data. Biophys.J. 43: 207-223.
Keunecke, M. (1995). Untersuchung des anomalen Molfraktionseffektes an der GrünalgeChara corallina. Diplomarbeit, Kiel.
Kienker, P. (1989). Equivalence of aggregated Markov models of ion-channel gating. Proc. Roy.Soc. Lond. Biol. 236: 269-309.
Kijima, S.,and Kijima, H. (1987a). statistical analysis of channel current from a membranepatch: I. Some stochastic properties of ion channels or molecular systems in equilibrium. J.theor. Biol. 128: 423-434.

Literaturverzeichnis
84
Kijima, S.,and Kijima, H. (1987b). statistical analysis of channel current from a membranepatch: II. Some stochastic theory of a multi-channel system in the steady-state. J. theor. Biol.128: 435-455.
Kirst, U. (1997). Modellierung und Softwareentwicklung für die statistische Analyse vonPatch-Clamp-Daten. Diplomarbeit, Kiel.
Klein, S., Timmer, J., Honerkamp, J. (1997). Analysis of multi channel patch clamp recordingsby Hidden Markov models. Biometrics 53: 870-884.
Klieber, H.-G., und Gradmann, D. (1993). Enzyme kinetics of the prime K+ channel in thetonoplast of Chara: selectivity and inhibition. J. Membr. Biol. 132: 253-265.
Läuger, P. (1980). Kinetic properties of ion carriers and channels. J. Membr. Biol. 57: 163-178.
Läuger, P. (1985). Ionic channels with conformational substates. Biophys. J. 47: 581-591.
Laver, D.R., Walker, N.A. (1991). Activation by Ca2+ and block by divalent ions of the K+
channel in the membrane of cytoplasmic drops from Chara australis. J. Membrane Biol. 120:131-139.
Laver, D.R. (1992). Divalent cation block and competition between divalent and monovalentcations in the large-conductance K+ channel from Chara australis. J. Gen. Physiol. 100: 269-300.
Lühring, H. E. (1986). Recording of single K+ channels in the membrane of cytoplasmic dropof Chara australis. Protoplasma 133: 19-28.
Mac Kinnon, R. (1991). Determination of the subunit stoichiometry of a voltage-activatedpotassium channel. Nature 350: 232-235.
Miller, C. (1991). Annus mirabilis of potassium channels. Science 252: 1092- 1096.
Möller, C. (1998). Theroetische und experimentelle Untersuchungen zur Modellierung vonStrom-Spannungs-Kurven aus Patch-Clamp-Messungen. Diplomarbeit, Kiel.
Mueller, P., Rudin, D. O. (1967). Development of K+-Na+ discrimination in experimentalbiomolecular lipid membranes by macrocyclic antibiotics. Biochem. Biophys. Res. Commun.26: 398.
Nedler, J.A., Mead, R. (1965). A simplex method for function minimization. Computer J. 7: 308
Neher, E., Sakmann, B. (1976). Single-channel currents recorded from membrane of denervatedfrog muscle fibres. Nature 260: 799-802.
Ophasi, L. R., Webb, W. W. (1994). Lipid-glass adhesion in giga-sealed patch-clampedmembranes. Biophys. J. 66: 75-79.

Literaturverzeichnis
85
Pongs, O. (1992). Molecular biology of voltage-dependent potassium channels. Physiol. Rev.72: S69-S88.
Rießner, T. (1994). Statistische Analyse von schnellen Schaltereignissen in Patch-Clamp Datenmit Hilfe von erweiterten Beta-Verteilungen. Diplom Arbeit Kiel.
Sanders, D., Hansen, U.-P., Gradmann, D., Slayman, C. L. (1984). Generalized kineticanalysis of ion-driven cotransport systems: a unified interpretation of selective ionic effects onMichaelis parameters. J. Membr. Biol. 77: 123-152.
Schultze, R. (1990). Untersuchung zum Auftreten des anomalen Molfraktionseffektes derStrom-Spannungskurven des biologischen Kaliumkanals mit Hilfe von Computersimulationenund analytischen Kriterien. Diplomarbeit, Kiel.
Schultze, R., Draber, S. (1993). A nonlinear filter algorithm for the detection of jumps inpatch-clamp data. J. Membrane Biol. 132: 41-52.
Serrano, R. (1988). Structure and function of proton translocating ATPase in plasma membranesof plants and fungi. Biochim. Biophys. Acta 947: 1-28.
Sutter, U. (1996). Biophysikalische Untersuchung an Xylemkontaktzellen: Präparation,Meßtechnik, Transporteigenschaften und Modellentwicklung. Diplomarbeit, Kiel.
Tester, M. (1988). Potassium channels in the plasmalemma of Chara corallina are multi-ionpores: voltage-dependent blockade by Cs+ and anomalous permeabilities. J. Membrane Biol.105: 87-94.
Tyerman, S.D. (1992). Anion channels in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 43:351-373.
Urban, B. W., Hladky, S. B. (1979). Ion transport in the simplest single ion pore. Biochim.Biophys. Acta 554: 410-429.
White, P. J. (1996). The permeation of ammonium through a voltage- indpendent K+ channelin the plasma membrane of rye. J. Membrane Biol. 152: 89-99.
Woodhull, A. M. (1973). Ionic blockade of sodium channels in nerve. J. Gen. Physiol. 61: 687-708.
Wu, J. (1991). Microscopic model for selective permeation in ion channels. Biopys. J. 60, 238-251.
Wu, J. (1992). Dynamic ion-ion and water-ion interactions in ion channels. Biophys. J. 61,1316-1331.
Yellen, G. (1984). Ionic permeation and blockade in Ca2+ activated K+ channels of bovinechromaffin cells. J. Gen. Physiol. 84: 157-186.
Zannello, L. P., Barrantes, F. J. (1994)Temperature sensivity of the K+ Cannel of Chara. Athermodynamic analysis. Plant Cell Pysiol. 35(2): 243-255.

86
DanksagungHerrn Prof. Dr. Ulf-Peter Hansen danke ich für das interessante Thema und die unermüdlicheund geduldige Betreuung. Ihm bin ich für den freundschaftlichen Umgang und die kritischenRatschlägen zum Dank verpflichtet.Maike Keuneke danke ich für die Einführung in die Patch-Clamp-Technik, für viele klärendeGespräche, erfrischende Hinweise, ihre Unterstützung und Hilfestellung.Thilo Rießner danke ich für die Lösungen aus den vielen Computerproblemen und diekompetente Diskussionsbereitschaft bei den vielen theoretischen Problemen. Er hat miraußerdem einige seiner Programme zur Verfügung gestellt.Mein Dank gilt auch den Mitgliedern der Arbeitsgruppe für Biophysik, die durch kritischeDiskussionen wesentlich zum Gelingen dieser Arbeit beigetragen haben.Einen ganz herzlichen Dank an alle, die bei der Korrektur der Arbeit mitgewirkt haben.Ganz besonders bedanke ich mich bei meinen Eltern, die mir das Studium ermöglichten undmich jederzeit tatkräftig unterstützten.

87
ErklärungHiermit versichere ich an Eides statt, daß ich die vorliegende Diplomarbeit selbständig undnur mit Hilfe meiner akademischen Lehrer sowie der angegebenen Literatur angefertigt habe.
Kiel, den ..........................
Afshin Farokhi





























