Embed Size (px)
Citation preview

- 1 -
Der Kern des Design-Arguments in der Biologie
Warum sich in der Natur kein „intelligentes Design“ offenbart
MARTIN NEUKAMM
Inhalt
A. Einleitung
B. Der Kern des Design-Ansatzes
C. Kritik an der positiven Komponente
des Design-Arguments
C.1 Aus Naturteleologie folgt keine
(Handlungs-) Intentionalität
C.2 Lebewesen und technische Konstrukte haben radikal verschiedene Eigen-
schaften: Biosysteme sind Systeme der Selbstorganisation
C.3 Nicht reduzierbare Komplexität
C.4 Der Design-Ansatz ist ohne Spezifikation nicht prüfbar
C.5 Ein methodologischer Vergleich zwischen ID und der Archäologie
C.6 Zwei Eisenmassen in der Sahara
C.7 Design-Modelle, die sich zirkelfrei positiv testen lassen, fehlen
C.8 Die Kritik des Philosophen Elliott SOBER
D. Zielscheibenfehler: Plastizität und „programmierte Variabilität“ als Design-
Merkmale
E. Das Argument der Konstruktionsfehler
F. Kritik an der negativen Komponente des Design-Arguments
F.1 Nicht reduzierbare Komplexität als Einwand gegen Evolution
F.2 Reduzierbare Spezifität contra nicht reduzierbare Komplexität
F.3 Schätzungen zur Unwahrscheinlichkeit von Evolution
G. Der Design-Ansatz in der biologischen Forschung
G.1 Gibt es „gute“ Erklärungslücken?
G.2 Ist der Design-Ansatz ein „Science-Stopper“, oder stärkt ihn die Forschung?
H. Zusammenfassung
© AG EvoBio – Evolution in Biologie, Kultur und Gesellschaft 10/2018

- 2 -
A. Einleitung
Auf der Website des evangelikalen Vereins WORT UND WISSEN findet sich ein Grund-
satzartikel über das Design-Argument in der Biologie. Letzteres nennt sich auch Intelli-
gent Design, kurz: ID. Der Beitrag stammt von Markus WIDENMEYER und Reinhard
JUNKER und dient Menschen als Handreichung, die „methodisch sauber“ für Schöpfung
argumentieren wollen (WIDENMEYER & JUNKER 2016). Vollmundig heißt es, Kritiker wür-
den am Kern des Design-Arguments „scheitern“. Entsprechend wird der Text bewor-
ben – als ein „must read“ für alle, die argumentativ „up to date“ sein möchten.
Der Leser erwartet also eine argumentativ gewichtige Abhandlung zum intelligenten
Design. Doch die Erwartungen erfüllen sich nur teilweise. Tatsächlich unterscheidet
sich die Argumentation der Autoren nicht wesentlich von vorangegangenen Publikatio-
nen. Gleichwohl lohnt sich die Detail-Analyse.
Zum einen stellt der Beitrag Design-Argumente kompakt statt bücherfüllend dar, was die
Kritik erheblich erleichtert. Zum anderen arbeiten sich die Autoren erfreulicherweise we-
nig an biologischen Detailfragen ab. Dies kommt der Diskussion zugute, denn der Streit
darüber, inwiefern die Evolutionstheorie diesen oder jenen Entwicklungsschritt zufrie-
denstellend erklärt, ist fruchtlos. Die Frage, ob die Fakten den Design-Ansatz erhärten,
lässt sich nur auf dem Boden der Methodologie und Logik führen.
Es geht zuallererst um wissenschaftstheoretische Fragen: Ist Intelligent Design eine
vernünftige Alternative zur (naturalistischen) Evolutionstheorie? Ist es rational begrün-
det und durch Forschung zu untermauern? Sind die Einwände seiner Kritiker unzu-
reichend, wie die Anhänger des intelligenten Designs behaupten? Um diese Fragen zu
klären, widmen wir uns zunächst dem Kern der Argumentation nach WIDENMEYER &
JUNKER (2016) und kritisieren ihn dann systematisch.
B. Der Kern des Design-Ansatzes
Als „Design-Ansatz“ bezeichnen die Autoren die These, bestimmte Naturgegenstände
seien ursprünglich auf geistige Ursachen, das heißt auf Schöpfung, zurückführen (S.
2). Dieser Ansatz ist in logischer Hinsicht mit dem Design-Argument verknüpft – dem
Rückgriff auf wissenschaftliche Befunde und logische Aspekte. Design-Argumente
dienen dazu, den Design-Ansatz zu begründen (S. 1).

- 3 -
Nach WIDENMEYER & JUNKER besteht die Begründung für „eine geistige Verursachung
und damit für das Design-Argument“ aus zwei Komponenten. Erstens:
„1. Der Naturgegenstand zeigt definierte Kennzeichen von Planung bzw. Zielorientierung
(Teleologie), die wir in anderen Fällen ganz entsprechend unseren sonstigen Design-
Erfahrungen (Technik, Kunst) ausschließlich auf einen geistigen Urheber zurückführen
(wir sprechen dann von ‚Design-Indizien‘ oder ‚Design-Merkmalen‘ wie z. B. funktionale
Komplexität; s. u.)“ (S. 1-2).
Insbesondere seien „geistig hervorgebrachte“1 Gegenstände wie Computer „so gestal-
tet, dass ihre Teile in z. T. äußerst komplexen und vielschichtigen Zweck-Mittel-
Beziehungen stehen.“ Die Teile, ihre Form und Anordnung, seien „nur in Bezug auf
ihre Funktionen und letztlich das Ganze, den Zweck des Gegenstands, zu verstehen“.
„Damit ein Gegenstand eine Funktion ausüben kann, benötigt er in der Regel mehrere Bau-
elemente, die in spezifischer Weise jeweils konstruiert und zusammen aufeinander abge-
stimmt sein müssen. Dies betrifft oft ganz verschiedene Aspekte wie Materialeigenschaften,
Form, Steuerung u. a. Diese Bestandteile und ihre gegenseitigen Abstimmungen erfordern
sehr häufig eine hohe Komplexität, die nicht mehr verkleinert werden kann, ohne die in Rede
stehende Funktion vollständig zu verlieren (nichtreduzierbare Komplexität…). Diese Kon-
struktionen sind an sich klare Design-Kennzeichen…“ (S. 5).
Nicht reduzierbare Komplexität ist ein zentraler Begriff des Intelligent Designs. Ein zu-
sammengesetzter Gegenstand ist nicht reduzierbar komplex, wenn sich keines seiner
Bestandteile entfernen lässt, ohne dessen Funktion komplett zu zerstören (BEHE 1996, S.
39). Wir können auch sagen: Interagieren mehrere Elemente spezifisch miteinander, bil-
den sie eine funktional nicht reduzierbare Ganzheit, ein System.2 Stören wir das Zusam-
menspiel der Elemente, gehen seine emergenten Eigenschaften und Strukturen verloren.
Systeme besitzen zwar oft redundante Teile, deren Verlust ihre Aktivität nicht wesentlich
beeinträchtigt. Gleichwohl existiert ein „Kern“ an Elementen, der für das betreffende Sys-
1 Streng genommen stellt der Ausdruck „geistig hervorgebracht“ ein Oxymoron dar: Nie hat jemand ein
Stück „Geist“ etwas Gegenständliches hervorbringen sehen. Stets sind es (materielle) Systeme, die
Dinge hervorbringen, seien es Sterne oder vernunftbegabte Wesen mit komplexen Gehirnen.
2 In der ID-Literatur ist von nicht reduzierbar komplexen Systemen die Rede. Dies ist faktisch ein Pleo-
nasmus, eine überflüssige Doppelung von Wörtern mit gleicher Bedeutung. Wir können hier das Adjektiv
„nicht reduzierbar komplex“ weglassen. Denn es gibt kein System, das keine emergenten Strukturen be-
säße und in diesem Sinn nicht irreduzibel komplex wäre. Alle Systeme haben einen nicht reduzierbar
komplexen „Kern“. Andernfalls sind die betreffenden Elemente in unspezifischer Weise vereint, sodass sie
Aggregate bilden (vgl. MAHNER & BUNGE 2000, S. 26). Daher sprechen wir entweder von Systemen oder –
gleichbedeutend – von nicht reduzierbar komplexen Gegenständen, Merkmalen oder Strukturen.

- 4 -
tem konstitutiv ist. Lebewesen verfügen über zahlreiche komplexe Strukturen, die diese
Bedingung erfüllen. Die Autoren glauben, „die bekannten natürlichen Mechanismen“
seien „nach aller unserer Erkenntnis bei weitem überfordert“, sie hervorzubringen. Erklä-
rungen, die auf „überlegte Mittelwahl“ verzichten, ließen „a priori keine Merkmale … er-
warten, wie sie bei intentional organisierten Gegenständen vorliegen“ (S. 3).3 Damit sind
wir bei der zweiten, der negativen Komponente des Design-Arguments:
„2. Ein natürlicher Entstehungsvorgang des betrachteten Naturgegenstandes ist unbe-
kannt, und Erklärungsversuche scheitern trotz Wissenszuwachs (im Idealfall können so-
gar Gründe angegeben werden, warum sie scheitern). Ein konkretes Design-Argument
würde demnach geschwächt, wenn naturwissenschaftlich die Möglichkeit eines natürli-
chen Entstehungsvorgangs im Detail nachgewiesen würde, der zum betreffenden De-
sign-Merkmal führt (vgl. Abschnitt 4, Einwand 1). Damit würde das Design-Indiz seine
Kraft verlieren und bekäme Konkurrenz“ (S. 2).
Der Grund, weshalb eine Evolution nicht reduzierbar komplexer Strukturen unplausibel
sei, besteht WIDENMEYER & JUNKER zufolge darin,
„dass kein kontinuierlicher, kleinschrittiger und hinreichend wahrscheinlicher Weg von ei-
nem Zustand ohne diese spezielle Funktion (und ohne die speziell dafür nötige Konstrukti-
on) hin zu einem Zustand mit dieser Funktion (und der dafür nötigen Konstruktion) gedacht
[sic!] werden kann. Jeder einzelne Mutationsschritt müsste eine hinreichende statistische
Wahrscheinlichkeit haben. Er dürfte nicht selektionsnegativ sein, vielmehr müsste die ent-
sprechende Mutante in der Population konserviert und signifikant verbreitet werden. Dabei
müsste er sich dem in Rede stehenden Zustand annähern … Selbstverständlich kann hier
jedoch nicht (schrittweise) auf ein Ziel hin geplant und ‚gearbeitet‘ werden, da natürliche
Mechanismen zukunftsblind sind...“ (S. 7).
„Geistige Urheber“ unterliegen dieser Beschränkung nicht, „da sie zielgerichtet vorgehen
und vorab im Geiste die verschiedensten Aspekte gleichzeitig beachten und aufeinander
abstimmen können.“ Sie wären in der Lage, Organismen am Reißbrett neu zu konzipie-
ren, ohne auf stammesgeschichtliche „Altlasten“ Rücksicht nehmen zu müssen:
„Geistbegabte Wesen (Personen) haben Ich-Bewusstsein, Wertekategorien, Denkver-
mögen, setzen Ziele und verfolgen sie überlegt usw.“ Sie zeichneten sich aus durch:
„Zielsetzung (Zukunftsorientierung), Planung, Wahl (der Mittel), Überlegungen zu Zwi-
schenschritten, Einkalkulieren möglicher Hindernisse, Sich-Vorstellen von Weg und Ziel
(Intentionalität). Natürliche Vorgänge können das nicht“ (S. 2).
3 Woher die Autoren diese „A-priori-Erkenntnis“ haben, wissen wir nicht. Sie tun so, als sei durch evolutio-
näre Algorithmen de novo noch nie Funktional-Zweckmäßiges entstanden. Das ist aus der Luft gegriffen.
Ausfühlich beschäftigen sich MATT et al. (2014) mit diesem Thema. Beispiele darüber, was die Evolution
schon in kurzer Zeit zu leisten vermag, erläutern KRAKER & GERSHENZON (2011) und BEYER (2018).

- 5 -
Danach wäre der Schluss auf Intelligent Design ein empirischer Analogieschluss, ein
Schluss, der sich aus Erfahrungswissen speist:
„Nach aller unserer Erfahrung gilt: Organisierte Gegenstände entstehen durch den Ein-
satz von Intelligenz und Planung“ (S. 5).
Gleichwohl schließen WIDENMEYER & JUNKER eine natürliche Entstehung biologischer
Systeme nicht kategorisch aus. Sie halten sie lediglich für unplausibel derzeit.
C. Kritik an der positiven Komponente des Design-Arguments
Ist das Design-Konzept eine vernünftige Alternative zur naturalistischen Evolutionsthe-
orie? Ist der Schluss auf ein intelligentes Design in der Natur der Schluss auf die beste
Erklärung, wie seine Proponenten behaupten? Dagegen spricht, wie wir im Folgenden
sehen werden, eine Reihe von Gründen.
C.1 Aus Naturteleologie4 folgt keine (Handlungs-) Intentionalität
Es ist unstrittig: Die in Organismen ablaufenden Prozesse und deren Verhaltenswei-
sen sind komplex. Auch lassen sie sich im Lichte bestimmter „Zweck-Mittel-
Beziehungen“ betrachten. Im Rahmen seiner Naturteleologie verwendete KANT den
Begriff der inneren Zweckmäßigkeit. Dementsprechend sind naturteleologische Termi-
ni, die Zwecke und Ziele anzeigen, in der Biologie gang und gäbe: Das Wehrsekret
des Bombardierkäfers erfüllt „den Zweck“, Angreifer in die Flucht zu schlagen. Die raf-
finiert gestalteten Blütenstrukturen einiger Orchideen „dienen“ dazu, Bienen ihren Pol-
len aufzudrücken. Heliotrope Pflanzen wenden ihre Blätter und Knospen der Sonne zu,
„um“ die Energieproduktion zu steigern. Die Liste ließe sich endlos fortführen.
Um diese Form teleologischer Sprache zu vermeiden, bedarf es einer hohen Sprachdis-
ziplin. Möglich wäre es, wie MAHNER & BUNGE (2000, S. 357) nachweisen, doch die Vorzü-
ge einer guten, bildhaft-anschaulichen Sprache hindern uns meist daran. Dies ist im Prin-
zip harmlos. Problematisch ist das Vermengen von Naturteleologie und der in Artefakten
erkennbaren Handlungsteleologie eines Schöpfers dann, wenn sie Ausdruck einer onto-
logischen Position ist. Das ist der Fall, wenn der Betrachter auf gleiche Ursachen der Ent-
stehung schließt. So lesen wir bei WIDENMEYER & JUNKER (S. 1):
4 Zur Erläuterung: Der Begriff „Naturteleologie“ ist hier beschreibend gemeint. Er bezieht sich auf bioti-
sche Strukturen, die eine positive Auswirkung auf den Fortpflanzungserfolg ihres Trägers haben.

- 6 -
„1. Der Naturgegenstand zeigt definierte Kennzeichen von Planung bzw. Zielorientierung
(Teleologie), die wir in anderen Fällen ganz entsprechend unseren sonstigen Design-
Erfahrungen (Technik, Kunst) ausschließlich auf einen geistigen Urheber zurückführen
(wir sprechen dann von ‚Design-Indizien‘ oder ‚Design-Merkmalen‘ wie z. B. funktionale
Komplexität; s. u.)“ (S. 1-2).
Ein solches Parallelisieren von Natur- und Handlungsteleologie (Intentionalität) ist unstatt-
haft, da sich beide Formen kategorial voneinander unterscheiden: Eine Handlungsteleolo-
gie spiegelt sich in Objekten, an denen wir die gedankliche Vorwegnahme eines äußeren,
nicht dem Gegenstand dienlichen Ziels ablesen. Diese besitzen in aller Regel keine intern
zweckmäßigen Strukturen. Mit der Naturteleologie verhält es sich umgekehrt. Sie betrifft
Systeme, die „um ihrer selbst willen“ existieren: Ihre Strukturen dienen der Selbstorganisa-
tion und Reproduktion. Im Gegensatz zu Maschinen, die nur „bewegende Kraft“ haben,
haben Naturgegenstände „in sich bildende Kraft“ (TOEPFER 2004, S. 333). Wer sie trans-
zendiert, ihnen einen Schöpferwillen oder die Tendenz zur Selbstdarstellung der Lebewe-
sen voranstellt, überschreitet das empirisch Begründbare (vgl. TOEPFER 2004, S. 198).
Verdeutlichen wir dieses Argument anhand einiger Beispiele: Autos, Computer, Nuss-
knacker, Trinkgefäße und Uhren sind zweckmäßig und funktional. Sie sind es im Hin-
blick auf ein äußeres Ziel. Es sind fremddienliche Werkzeuge – von Menschen und
für Menschen gemacht, keine Werkzeuge der Selbstorganisation. In Kenntnis des De-
sign-Zwecks schließen wir ohne zu zögern auf einen Zwecksetzer. Bei Lebewesen ist
ein solcher Schluss nicht ohne weiteres möglich, weil sie selbstdienliche Strukturen
besitzen, jedoch keine fremddienlichen Werkzeuge, die den Schluss auf Planer recht-
fertigen. Diese Naturteleologie spiegelt sich exklusiv in den sich wechselseitig hervor-
bringen Teilen eines Organismus. Es stellt daher
„… eher eine Irreführung als eine gerechtfertigte methodische Parallelisierung dar, wenn
beide Bereiche mit der einheitlichen Begrifflichkeit der Teleologie, insbesondere mit dem
Wort Zweckmäßigkeit, erschlossen werden sollen. Nach dem Ergebnis meiner Arbeit
verliert die Zweckmäßigkeit in den Bereichen der Handlungs- und Naturteleologie die
Einheitlichkeit ihrer Bedeutung“ (TOEPFER 2004, S. 425).
Merke: Aus Naturteleologie folgt keine (Handlungs-) Intentionalität. Der Schluss
von der inneren Zweckmäßigkeit biotischer Prozesse auf einen handelnden
Zwecksetzer ist unstatthaft (TOEPFER 2004, S. 425f). Aus diesem Grund prägte
Colin S. PITTENDRIGH den Begriff „Teleonomie“, um diesen Effekt konzeptionell und
begrifflich sauber von intentionalem Telos zu unterscheiden.

- 7 -
C.2 Lebewesen und technische Konstrukte haben radikal verschiedene
Eigenschaften: Biosysteme sind Systeme der Selbstorganisation
Es kommt ein wichtiger Punkt dazu: Menschen kommen nicht umhin, Computer und
Uhren zu planen, zu konstruieren und zusammenzusetzen. Der Grund: Sie entwickeln
sich nicht aus Eizellen. Sie wachsen nicht, sie pflanzen sich nicht fort und unterliegen
keiner Mutation und Selektion. Daher kommen sie von vorn herein nicht als Produkt
evolutionärer Entwicklung in Betracht. Aber Lebewesen wachsen und vermehren sich.
Sie haben eine Generationenfolge. Und die Nachkommen unterscheiden sich von den
Eltern in Bezug auf Aussehen und Fitness (differenzielle Tauglichkeit der Varianten).
Mit einem Wort: Lebewesen sind evolutionsfähige Mehrgenerationen-Systeme.
Dies schließt zwar nicht aus, dass sie anfangs „designt“ wurden, aber ein solcher
Schluss ist nicht naheliegend. Er widerspricht all unseren Daten und Erkenntnissen.
Das wäre selbst dann der Fall, wenn es gelänge, Roboter zu konstruieren, die sich
„vermehren“. Denn es ist klar, dass Roboter kein natürliches, organisches Wachstum
mit zellulärer Differenzierung durchlaufen würden. Und sie wären nicht zeugungsfähig
im eigentlichen Wortsinn. Ihre „Nachkommen“ müssten sie händisch fertigen. Die An-
führungszeichen zeigen den Unterschied auf: Roboter unterscheiden sich kategorial
von allen Naturgegenständen: Es gibt hier keine Zellteilung, kein Wachstum, keine
Mutation, keine Rekombination, keine Selektion und keine natürliche Evolution.
Ein Beispiel, das BEYER (2018) anführt, veranschaulicht den fundamentalen Unter-
schied: Im Rahmen der Züchtung neuer, pollensteriler Maissorten entstand ein Protein
mit der prosaischen Namen T-urf13, das um 1970 entdeckt wurde. Im Verbund mit
einigen weiteren T-urf13-Molekülen bildet es Strukturen in der inneren Mitochondrien-
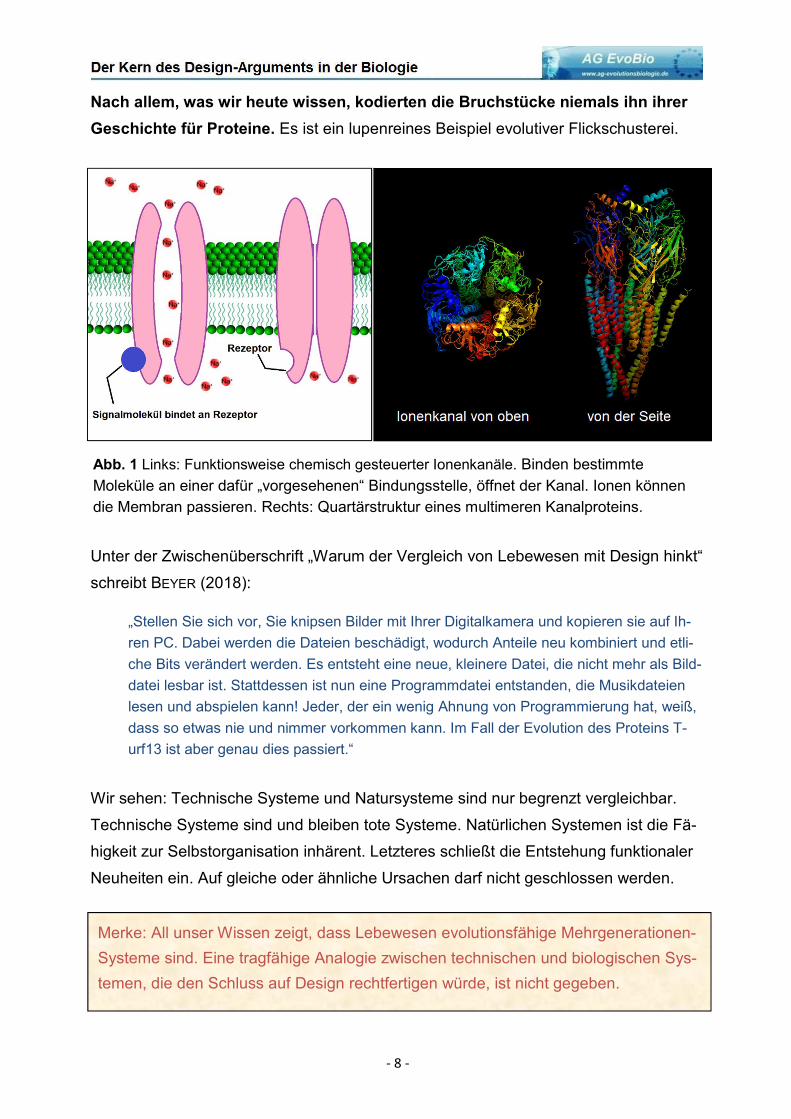
Membran, die einen chemisch gesteuerten Kanal formen: Er besitzt Bindestellen für
Moleküle, die dafür sorgen, dass der Kanal öffnet und Ionen die Membran passieren
lässt (Abb. 1). Dieser Mechanismus ist hochspezifisch: Er bewirkt zu einer definierten
Zeit der Blütenreifung der Mais-Pflanze „gezielt“ das Absterben der betreffenden Zel-
len in den Staubblättern. Die Pflanze bleibt vital, bildet aber keine Pollen mehr.
WIDENMEYER & JUNKER würden sagen, das System wirke hochgradig „zielgerichtet“:
Seine Teile stehen in einer spezifischen „Zweck-Mittel-Beziehung“, die die Züchter se-
lektierten. Doch der Clou ist, dass sie diese Struktur weder gezielt erschufen, noch
durch Mutation eines anderen Ionenkanals gewannen. Das Gen, welches für das
Protein kodiert, entstand zufällig durch Rekombinationen mitochondrialer DNA.
- 8 -
Nach allem, was wir heute wissen, kodierten die Bruchstücke niemals ihn ihrer
Geschichte für Proteine. Es ist ein lupenreines Beispiel evolutiver Flickschusterei.
Abb. 1 Links: Funktionsweise chemisch gesteuerter Ionenkanäle. Binden bestimmte
Moleküle an einer dafür „vorgesehenen“ Bindungsstelle, öffnet der Kanal. Ionen können
die Membran passieren. Rechts: Quartärstruktur eines multimeren Kanalproteins.
Unter der Zwischenüberschrift „Warum der Vergleich von Lebewesen mit Design hinkt“
schreibt BEYER (2018):
„Stellen Sie sich vor, Sie knipsen Bilder mit Ihrer Digitalkamera und kopieren sie auf Ih-
ren PC. Dabei werden die Dateien beschädigt, wodurch Anteile neu kombiniert und etli-
che Bits verändert werden. Es entsteht eine neue, kleinere Datei, die nicht mehr als Bild-
datei lesbar ist. Stattdessen ist nun eine Programmdatei entstanden, die Musikdateien
lesen und abspielen kann! Jeder, der ein wenig Ahnung von Programmierung hat, weiß,
dass so etwas nie und nimmer vorkommen kann. Im Fall der Evolution des Proteins T-
urf13 ist aber genau dies passiert.“
Wir sehen: Technische Systeme und Natursysteme sind nur begrenzt vergleichbar.
Technische Systeme sind und bleiben tote Systeme. Natürlichen Systemen ist die Fä-
higkeit zur Selbstorganisation inhärent. Letzteres schließt die Entstehung funktionaler
Neuheiten ein. Auf gleiche oder ähnliche Ursachen darf nicht geschlossen werden.
Merke: All unser Wissen zeigt, dass Lebewesen evolutionsfähige Mehrgenerationen-
Systeme sind. Eine tragfähige Analogie zwischen technischen und biologischen Sys-
temen, die den Schluss auf Design rechtfertigen würde, ist nicht gegeben.

- 9 -
C.3 Nicht reduzierbare Komplexität
Spätestens an dieser Stelle wenden ID-Vertreter ein, Biosysteme unterschieden sich
zwar hinsichtlich ihrer Fähigkeit zur Selbstorganisation von technischen Gegenstän-
den. Sie seien aber nicht so verschieden, dass sich der Schluss auf gleiche Ursachen
verbiete. Bereits LÖNNIG (1993) merkte hierzu an:
„Man hört hierzu manchmal den Einwand, dass sich die von Menschen erschaffenen ky-
bernetischen Systeme nicht fortpflanzen können. Dabei wird völlig übersehen, dass Mi-
tose und Meiose selbst ungeheuer komplexe kybernetische Systeme darstellen, deren
erfolgreiche Funktion unter anderem das genauestens koordinierte Zusammenspiel von
Hunderten von Genen erforder[t].“
Dieser Autor hebt implizit auf die Eigenschaft hochgradiger nicht reduzierbarer Kom-
plexität ab und deutet sie als Designindiz (so auch WIDENMEYER & JUNKER, S. 3). In
formalisierter Form könnte das Argument beispielsweise wie folgt lauten:
Formallogisch ist gegen dieses Argument nichts einzuwenden. Aber die Prämisse, ähn-
liche Eigenschaften (nicht reduzierbare Komplexität) hätten ähnliche (intelligente) Ursa-
chen (causa aequat effectum), ist nicht allgemein gültig (Martin MAHNER, pers. Mittei-
lung). Es ist nur begrenzt möglich, Rückschlüsse von einer bekannten Wirkung auf eine
unbekannte Ursache anzustellen. Selbstredend haben nicht reduzierbar komplexe tech-
nische Gegenstände intelligente Ursachen. Dennoch ist der Schluss über die unbelebte
Welt hinaus aus mehreren Gründen nicht stringent.
Zunächst trifft Randbedingung 2 in dieser Form nicht zu: Wir haben gesehen, dass mit
dem T-urf13-Protein ein hochspezifisches, nicht reduzierbar komplexes Merkmal evol-
vierte. Aber nehmen wir pro forma an, die Randbedingung wäre korrekt, der Ionenka-
Prämisse: Nach unserer Erfahrung gehen (hochgradig) nicht reduzierbar komplexe Merk-
male auf Intelligenz und Planung zurück.
Randbedingung 1: Lebewesen beinhalten (hochgradig) nicht reduzierbar komplexe organi-
sierte Merkmale.
Randbedingung 2: Die Suche nach natürlichen Erklärungen zur Entstehung von (hoch-
gradig) nicht reduzierbar komplexen Strukturen blieb bislang ergebnislos.
Folgerung: Der Schluss auf die beste Erklärung ist, dass Lebewesen erschaffen wurden.

- 10 -
nal aus irgendwelchen Gründen kein überzeugendes Beispiel. Dann wäre der Schluss
auf Design noch immer nicht gerechtfertigt. Denn es ist fraglich, ob nicht reduzierbare
Komplexität für den Analogie-Schluss überhaupt bedeutsam ist.
Zum einen ist Komplexität kein notwendiges Design-Kriterium. Wir benötigen sie nicht,
um Gegenstände wie Tonscherben, Mauerreste und Kleidung als Artefakte zu identifizie-
ren (MAHNER 2007, S. 341). Ein weiteres Beispiel stammt von den Autoren: Verwitterungs-
Strukturen in den Alpen im Vergleich zu den Menschenköpfen von der Osterinsel (Abb. 2):
„Das rechte Bild zeigt die berühmten Steinköpfe auf den Osterinseln. Hier wird niemand
auf die Idee kommen, sie alleine durch Erosion und Materialeigenschaften zu erklären.
Die Formen (Menschenfiguren) korrelieren hochspezifisch mit typischen Zielsetzungen
geistig begabter Wesen. Dies ist (hier) die bildhaft-abstrakte Darstellung von menschli-
chen Portraits. Dies rechtfertigt, a priori eine künstliche Entstehung anzunehmen. Wenn
bei hinreichender Kenntnis natürlicher Prozesse mechanistische Erklärungen für eine na-
türliche Entstehung derartiger Formen nicht vorliegen, haben wir gute Gründe, eine na-
türliche Entstehung auszuschließen“ (S. 4).
Abb. 2 Links: Felskopf in den Stubaier Alpen unterhalb des Kalbenjochs bei der Gemeinde
Trins. Rechts: Figuren auf den Osterinseln. Aus: WIDENMEYER & JUNKER (2016, S. 3).
Faktisch haben die Autoren hier Recht. Doch nicht Komplexität oder Funktionalität,
sondern das Wissen darüber, dass menschenähnliche Portraits durch Menschenhand
entstehen, stellt die Verbindung zu Design her. Wären menschliche Köpfe geformt wie
besagter Felskopf, vermuteten wir in dieser Struktur ein Konstrukt. Dann deutete ledig-
lich die Vielzahl der Blöcke auf den Osterinseln darauf hin, dass sie designt wurden.
Sämtlichen Analogie-Beispielen aus der Intelligent-Design-Literatur sehen wir ihre Tri-
vialität an: Autos, Computer, Motoren, Portraits, Raumschiffe, Uhren und Zahnräder
zeichnen sich durch ihren „Wiedererkennungswert“ aus. Dies gälte auch für so exoti-

- 11 -
sche Objekte wie extraterrestrische Radiowellen, denen die Kreiszahl Pi bis auf 1000
Nachkommastellen aufmoduliert wäre. SETI-Forscher schlössen daher auf einen künst-
lichen Ursprung, auf eine planmäßige Urheberschaft (NEUKAMM 2009, S. 49).
Andererseits ist selbst hochgradig nicht reduzierbare, funktionale Komplexität kein zu-
verlässiges Design-Indiz. Auch wenn wir nicht wüssten, wie sie zustande kam, wäre
der Design-Schluss unstatthaft. Ein Beispiel aus der nichtbelebten Welt: Der globale
Wasserkreislauf und dessen Verzahnung mit atmosphärischen und kosmischen Be-
dingungen ermöglicht Leben auf der Erde. Diese „Feinabstimmung“ lässt sich als eine
Form nicht reduzierbarer Komplexität auffassen (DRENDEL 2016). Trotzdem offenbart
sich in ihr nicht die Handschrift eines Designers. Sie entstand über Jahrmilliarden auf-
grund von Prozessen, die nicht abzusehen waren. Der Weg dahin verlief nicht zielori-
entiert geradlinig, und die Bedingungen auf der Erde sind ein kosmischer Glückstreffer.
Auf einen lebensfreundlichen Planeten kommen Tausende unbewohnbarer. Dieser
„sinnlose Aufwand“ (SCHMIDT-SALOMON 2005, S. 5) ist für Planung atypisch. Wie so oft,
fokussieren die ID-Anhänger auf den vermeintlichen „Sinn“ und übersehen den „Un-
sinn“. Treffend bemerkt CARROLL (2015):
„Die Sonne wird eines Tages kein Leben mehr auf der Erde zulassen. Schon heute lässt sie
auf den übrigen Planeten keinerlei Leben zu. Was folgt daraus für Design? Nichts. … Wir kön-
nen nicht leugnen, dass wir nicht existieren würden, würde unsere Existenz nicht durch Millio-
nen von Faktoren gestützt. Ja und? Viele dieser Faktoren existierten in der Vergangenheit
nicht und werden in Zukunft auf unserem Planeten nicht mehr existieren“ (ins Deutsche M.N.).
C.4 Der Design-Ansatz ist ohne Spezifikation nicht prüfbar
Wenden wir uns der Frage der Prüfbarkeit des Design-Ansatzes zu:
„Können wir empirisch bestätigte Fälle angeben, in denen mittels nicht-geistiger Prozes-
se organisierte Gegenstände aus nicht-organisierten Gegenständen entstanden sind (z.
B. aus einer komplexen Mixtur chemischer Verbindungen, die nicht wiederum biologi-
schen Ursprungs sind)? Da wie gesagt die Design-These die direkte Gegenthese zur
Merke: Weder im Alltag noch in der Wissenschaft ist nicht reduzierbare, funktiona-
le Komplexität ein zuverlässiger Indikator für intelligente Entstehungsursachen.
Auch wenn diesbezüglich eine stringente natürliche Erklärung aussteht, darf nicht
ohne weiteres auf einen artifiziellen Ursprung geschlossen werden. Dies ist nur
statthaft, wo natürliche Mechanismen von vorn herein (aufgrund bekannter physi-
kalisch-chemischer Gesetze) als Erklärung entfallen.

- 12 -
These einer nicht-geistigen Entstehung ist, ist eine Prüfung der These gleichzeitig eine
Prüfung der Gegenthese und umgekehrt“ (S. 5).
„Die Testbarkeit einer These entspricht der Testbarkeit der (genauen) Gegenthese. Wäre
der Design-Ansatz (als genaue Gegenthese einer naturalistischen Evolutionslehre) nicht
testbar, gölte dies ganz genauso für den Ansatz einer natürlich verlaufenden Evolution“
(S. 10, Fußn. 13).
Doch die These, der Design-Ansatz sei die „(genaue) Gegenthese einer naturalistischen
Evolutionslehre“, erweist sich als unbegründet. Warum? Nehmen wir an, es wäre möglich,
experimentell zu beweisen, dass der Mensch das Ergebnis natürlicher Evolution ist. Wäre
der Design-Ansatz geschwächt? Nur wenn die ID-Vertreter annehmen, der Befund stehe
im Widerspruch zu Design oder lasse den Design-Ansatz überflüssig erscheinen.
Einige Design-Anhänger setzen jedoch voraus, der Schöpfer verursache Mutationen
und arrangiere ihr unwahrscheinliches Zusammentreffen (RHONHEIMER 2007, S. 53).
Andere glauben, der Designer habe die Genome unserer Primaten-Vorfahren mit dem
Potenzial ausgestattet, sich zu Menschen zu entwickeln (↑ Abschnitt D, „programmier-
te Variabilität“). Der ID-Vertreter Michael BEHE (2008) geht noch weiter:
„Intelligentes Design ist durchaus kompatibel mit der Sichtweise, dass das Universum
ohne Überschreitung der Naturgesetze funktioniert, wobei das Design des Lebens viel-
leicht in seine ursprüngliche Struktur gepackt wurde“ (S. 166, ins Deutsche M.N.).5
Im Lichte dieser Spezifikationen würden experimentelle Beweise für eine lückenlose na-
türliche Evolution den Design-Ansatz nicht nur nicht schwächen, er ginge gestärkt dar-
aus hervor! Das heißt: Ohne das Einbeziehen von Hypothesen, die das Schöpfer-
handeln betreffen, lässt sich der Design-Ansatz nicht an konkreten Fällen prüfen.
Und je nach Spezifikation fällt das Prüfergebnis unterschiedlich aus. Doch wo liegt das
Problem? Selbstredend muss die Wissenschaft alle Theorien spezifizieren, damit sie
diese prüfen kann. Aus allgemeinen Theorien gewinnt sie theorieabhängige Modelle.
Das Problem ist, dass Schöpfungs-Modelle entweder keiner Prüfung standhalten, wie
die Vorstellungen des Kreationismus zeigen. Oder sie enthalten Hypothesen, die nicht
unabhängig vom Design-Ansatz prüfbar sind: Nichts Empirisches spricht dafür, ein
Schöpfer lanciere Mutationen oder habe das gesamte mögliche Variations-Potenzial in
die Genome von Organismen geschrieben. Wer dergleichen annimmt, um Design mit
5 Im Original: „Intelligent design is quite compatible with the view that the universe operates by unbroken
natural law, with the design of life perhaps packed into its initial set-up“.

- 13 -
Mikro- oder Makro-Evolution zu harmonisieren, setzt voraus, was er nicht unabhängig
vom Design-Ansatz belegen kann. Das Ergebnis ist ein fataler Zirkel wechselseitiger
Selbstbestätigung (circulus vitiosus): Das positive „Prüfergebnis“ des Design-Ansatzes
steckt bereits in der Prämisse. Erst der Nachweis entsprechender Designer und De-
sign-Methoden würde die Prämisse legitimieren. Gleiches gilt für die Prämisse der Au-
toren, eine lückenlose natürliche Evolution schwäche den Design-Ansatz.
Nun verstehen wir, warum die Forderung zur Spezifikation des Schöpferhandelns
sowie nach unabhängigen Belegen für den modus operandi des Designers unver-
zichtbar ist. Beides sind Voraussetzungen für eine kohärente (zirkelfreie) Prüfung des
Design-Ansatzes (ähnlich HEILIG 2011, S. 92f). Dieser Zusammenhang scheint WI-
DENMEYER & JUNKER entgangen zu sein. Sie meinen, „eine mögliche weitere Spezifizie-
rung“ würde „vom Design-Ansatz unbegründet verlangt“ (S. 12). Die Einsicht, dass wir
ohne diese weitergehende (kausale!) Spezifizierung Theorien weder auf reale Fälle
anwenden noch prüfen können, zählt zu den Grundlagen der Wissenschaftstheorie.
(Siehe das Schema der Operationalisierung nach MAHNER & BUNGE 2000, S. 91– 94).6
6 Auch die Evolutionstheorie ist auf die Spezifikation ihrer Mechanismen angewiesen: Wären die
„Großmechanismen“ der Evolution (Variation, Vererbung und Selektion) nicht unabhängig von der Evo-
lutionstheorie begründet, wäre eine zirkelfreie Prüfung der Evolutionstheorie unmöglich. Je nach „Tiefe“
der erklärenden Mechanismen ergeben sich teils unterschiedliche Vorhersagen.
Beziehen wir in die Evolutionstheorie die erwähnten Großmechanismen ein, entspricht die abgestufte
Ähnlichkeit der Arten ihren Erwartungen. Dagegen schwächt das gehäufte Auftreten von sogenannten
Konvergenzen und Parallelentwicklungen, die diese Ordnung stören, die so spezifizierte Theorie. Doch
integrieren wir verschiedene Erkenntnisse der Entwicklungsbiologie in die Evolutionstheorie („Evo-
Devo“), verfeinern sich die Vorhersagen. Dann sind auch Konvergenzen und Parallelentwicklungen
erwartbare Phänomene natürlicher Evolution (HALL 2012; MÜLLER & HASSEL 2018, S. 596–597, 626).
Merke: Der Design-Ansatz ist kein prüfbarer Entwurf zum Ursprung des Lebens. Es
gilt, ihn zu spezifizieren, das heißt mit zusätzlichen Hypothesen über das mutmaßli-
che Schöpferhandeln auszustatten. Erst dann läge ein (konkretes) Schöpfungs-
Modell vor, das sich an realen Fällen prüfen ließe.
In Ansätzen spezifizierte Design-Modelle gibt es. Denken wir an die „Sechs-Tage-
Schöpfung“ des Kreationismus oder an die Vorstellung, der Designer habe Lebe-
wesen mit „programmierter Variabilität“ ausgestattet. Einige Modelle nehmen an,
der Designer sei „menschenähnlich“, woraus folgen würde, dass seine Artefakte
ähnliche Merkmale aufwiesen, wie die des Menschen.
Doch die betreffenden Design-Modelle halten entweder keiner Prüfung stand, wie
der Kreationismus zeigt. Oder sie enthalten Zusatzhypothesen, die nicht unab-

- 14 -
In den folgenden Abschnitten wollen wir das, was wir in diesem Abschnitt theoretisch-
abstrakt behandelt haben, an Beispielen verdeutlichen.
C.5 Ein methodologischer Vergleich zwischen ID und der Archäologie
Das intelligente Design steht in der Biologie vor einem grundsätzlichen Problem: Es
erklärt nichts, sondern verlagert „die eigentliche Erklärung lediglich einen Schritt nach
hinten“ (MAHNER 2007, S. 350). Zwar würde ein intelligenter Designer gegebenenfalls
erklären, wie die ersten Lebewesen auf die Erde kamen. Doch die grundsätzlichere
Frage, wie Leben (und sei es in Gestalt des Designers) entsteht, lässt er unbeantwor-
tet. Außerdem kann ID keine potenziellen Designer vorweisen.
Gelegentlich kontern Design-Protagonisten mit Beispielen aus der Archäologie. So
lesen wir beim DISCOVERY INSTITUTE (2012), unser Wissen über intelligente Ursachen
erlaube Rückschlüsse auf Design, ohne Identität oder Herkunft der Designer zu ken-
nen. Nach Meinung der Autoren verkörpere die Archäologie „Intelligent Design in Ac-
tion“. Zum Beispiel sind moderne Technologien in der Lage, antike Verfahren zur Her-
stellung von Artefakten wie Tontafeln und Steinwerkzeugen zu rekonstruieren.
Dies wirft zwei fundamentale Fragen auf: Ist es unsinnig, Mechanismen und natür-
liche Erklärungen einzufordern, wo „geistige Entstehungsursachen“ gefragt
sind? Und: Zeigt die Archäologie, dass spezifisches Wissen über Designer und
ihre Methoden entbehrlich ist, um den Schluss auf Design zu ziehen?
Betrachten wir die methodologische Situation, stellen wir fest, dass dem nicht so ist:
hängig vom Design-Ansatz prüfbar sind. Die These von der „programmierten Va-
riabilität“ fällt ebenso in diese Kategorie wie die Annahme, der Designer handele
durch „Zufallsmutationen“ oder bringe Menschenähnliches hervor.
ID-Vertreter kombinieren den Design-Ansatz mit unterschiedlichsten, sich teils
gegenseitig ausschließenden Schöpfungs- und Schöpfervorstellungen. Sie sind
sich weder einig, was der Designer schuf, noch, inwieweit dieser in den Gang der
Welt eingriff. Da sie den Beitrag von Design auf völlig verschiedenen System-
ebenen vermuten (und diesen auch nicht spezifizieren können), ist es absurd
anzunehmen, es gäbe objektive Design-Indizien in der Biologie.

- 15 -
Abb. 3 Realwissenschaften wie die
Archäologie, die in bestimmten
Merkmalen von Steinen das Ergebnis
von Design erkennt, verweisen auf
empirisch nachgewiesene Urheber.
Sie spezifizieren deren Wirkmecha-
nismen und überprüfen sie experi-
mentell (hier: experimentelles Schla-
gen paläolithischer Faustkeile). So
liefern die Archäologen echte, kausa-
le Erklärungen. Das intelligente De-
sign der Biologie leistet das nicht.
Somit räumen wir explizit ein, dass designbasierte Ansätze im Rahmen der Naturwis-
senschaften Erklärungskraft entfalten (vgl. KOJONEN 2016, S. 86f). Doch aufgrund der
Allgemeinheit des Design-Ansatzes in der Biologie ist diesem kein Erfolg beschieden.
Intelligent Design unterscheidet sich in methodologischer Hinsicht dramatisch von florie-
renden wissenschaftlichen Disziplinen wie der Archäologie. Es operiert faktisch durch-
weg mit Unbekanntem: Designer, die infrage kämen, kennen wir nicht. Über ihre mut-
maßlichen Techniken, Fähigkeiten und Grenzen wissen wir nichts. Erforschen lässt sich
dergleichen nicht. Brauchbare Modelle, welche die Fertigungs-Mechanismen spezifizie-
ren, liegen nicht vor. Der Design-Ansatz bleibt spekulativ und inhaltsleer.
1. Die Archäologie befasst sich mit menschlichem Design. Das heißt, sie kennt
die potenziellen Urheber archäologischer Zeugnisse und kann deren Exis-
tenz und Aktivität unabhängig von der Objektebene nachweisen.
2. Die Archäologie bringt die menschlichen Zeugnisse mit bekannten (oder er-
forschbaren) Techniken in Verbindung. Sie kann Design-Modelle entwickeln,
die konkrete Fertigungs-Mechanismen (bestimmte Schlagtechniken) bein-
halten. In diesem Sinn liefert sie echte, mechanismische Erklärungen.
3. Die experimentelle Archäologie erforscht den Ursprung von Tontafeln und
Steinwerkzeugen. Ihr gelingt dies, indem sie die Fertigungs-Möglichkeiten
und Grenzen ihrer Urheber nachvollzieht.
4. Nur mithilfe dieses Wissens lassen sich Design-Vorstellungen rechtfertigen.
Ohne dieses können wir nicht prüfen, ob bestimmte Schlagmerkmale an
Steinen mit den Fertigkeiten des Urmenschen erklärbar sind (Abb. 3).

- 16 -
Analog zur Archäologie stehen dessen Anhänger vor der Aufgabe, potenzielle Techno-
logien in einem Modell zu konkretisieren. Und sie haben den Nachweis zu führen, dass
mögliche Urheber in den betreffenden Zeiträumen existierten. Erst dann ließen sich
unabhängige Belege für das betreffende Design beibringen. Dagegen führt Intelligent
Design das (vorgeblich) Unbekannte auf noch Unbekannteres zurück. Das ist, als woll-
ten wir ein Naturphänomen auf ein Alien zurückführen, dessen „Arbeitsweisen“ noch
unverstandener sind als das, was sie erklären sollen (Abb. 4).
Abb. 4 Das Design-Argument auf die Biologie anzuwenden ist, als führten Astrophysiker das
Explodieren von Sternen auf einen personalen Urheber, beispielsweise ein Alien, zurück. Ana-
log zu ID ließe sich argumentieren: 1.) Erfahrungsgemäß werden Explosionen meist von Men-
schenhand vorbereitet. 2.) Argument der „Feinabstimmung“: Supernovae erscheinen geplant,
denn es entstehen alle chemischen Elemente, die es für das Leben braucht. 3.) Längst nicht
alle kernphysikalischen Details zur Entstehung von Supernovae sind geklärt. – Ließe sich also
von einer Supernova auf einen intelligenten „Urheber“ schließen? Nur, wenn wir die Existenz
dessen voraussetzten, was es zu belegen gälte.
C.6 Zwei Eisenmassen in der Sahara
Angenommen, wir fänden in der Sahara zwei Eisenmassen. Die erste Masse offenbart
nach dem Anschliff, Polieren und Ätzen ein feines Martensit-Gefüge unter dem Mikro-
skop (Abb. 5, links). Die zweite zeigt nach der gleichen Behandlung ein regelmäßiges
Lamellen-Muster der Nickeleisen-Legierungen Kamacit und Taenit (Abb. 5, rechts).
Haben wir die Möglichkeit herauszufinden, wie diese Stücke entstanden?
Merke: Ohne Spezifikation des Design-Ansatzes in einem Design-Modell ist we-
der eine Anwendung auf reale Fälle noch eine Prüfung leistbar. Aus „unbekanntem
Design“ folgt nichts Konkretes. Der Design-Ansatz ist fruchtbar, wenn die Techno-
logien der mutmaßlichen Urheber bekannt oder prinzipiell rekonstruierbar sind.

- 17 -
Abb. 5 Kristallgefüge verschiedener Eisen-Nickel-Legierungen nach dem Ätzen mit Nital.
Links: Martensit-Gefüge. Rechts: Feines Lamellenmuster aus Kamacit und Taenit.
Das Martensit-Gefüge ist einfach strukturiert, könnte also natürlich entstanden sein.
Dagegen könnte es sich bei den komplexeren Figuren rechts um Leiterbahnen eines
elektronischen Bauteils für irgendeine Maschine handeln. Experimente zeigen, dass
sie sich nicht natürlich bilden. Braucht es darum eines geistigen Knowhows? Offen-
sichtlich bringt uns das Betrachten und Analogisieren dieser Objekte nicht weiter.
Erst das Wissen, dass das Martensit-Gefüge dem von gehärtetem Stahl entspricht, er-
laubt den Schluss auf ein Kunstprodukt. Erst die Kenntnis der Technologie gibt Anlass
zur Vermutung, dass die Eisenmasse designt ist. Erst die Erfahrung, welchen Zwecken
sie Menschen dient, rechtfertigt den Schluss auf eine Zwecksetzung. Analoges gilt für
das andere Eisen: Erst die Einsicht, dass sich bei extrem langsamem Abkühlen von Ni-
ckeleisen-Schmelzen Kamacit tafelförmig abscheidet, erlaubt den Schluss auf seine
kosmische Herkunft. Erst die Erkenntnis, dass das Muster WIDMANSTÄTTEN‘sche Figuren
repräsentiert, erlaubt den Schluss auf einen Meteoriten. Erst Wissen über die Anfänge
des Sonnensystems erhärtet den Schluss, dass Meteoriten natürliche Objekte sind.
Merke: Rein durch Betrachten von Objekten ist der Schluss auf Entstehungsursachen
nicht möglich. Es braucht zusätzliches Designer-Wissen oder Wissen über konkrete
Mechanismen, um auf Design oder natürliche Prozesse zu schließen. Im Fall von De-
sign benötigen wir Wissen über mögliche Intentionen oder Fertigungs-Techniken.

- 18 -
C.7 Design-Modelle, die sich zirkelfrei positiv testen lassen, fehlen
Oft hört man, Naturwissenschaft und Technik würden zeigen, dass sich durch Intelli-
genz Organismen manipulieren, züchten und für bestimmte Zwecke erschaffen ließen.
Allein dieser Umstand wird gerne und häufig als empirisches Argument für die Berech-
tigung des Design-Ansatzes in der Biologie gewertet:
„Nach aller unserer Erfahrung gilt: Organisierte Gegenstände entstehen durch den Ein-
satz von Intelligenz und Planung“ (S. 5).
Nun sind Gentechnologie und Züchtung zwar Beispiele, die belegen, dass moderne
Technologien Beachtliches leisten. Mehr noch: Prinzipiell ermöglicht intelligentes Planen
alles, was nicht gegen Naturgesetze verstößt und unsere Ressourcen übersteigt. Daher
ist es korrekt zu sagen: „Designer könnten im Prinzip Leben erschaffen“. Das Problem
ist nur: Diese Erkenntnis ist so allgemein, dass im Speziellen nichts für Design folgt.
Wie HEILIG (2015) dargelegt hat, muss in jedem konkreten Fall Rechenschaft über die
Plausibilität der (historischen) Randbedingungen abgelegt werden, bevor der De-
sign-Ansatz favorisiert werden kann. Das gilt unabhängig davon, ob ein „intelligenter
Prozess“ an sich bestimmte Produkte „erwarten“ lässt oder nicht.
So ist es beispielsweise hochproblematisch, einen (menschenähnlichen) Schöpfer an-
zunehmen, der zur Entstehungszeit der ersten Bakterien gewirkt haben soll. Auch wenn
wir wissen, dass hochintelligente Wesen prinzipiell Bakterien intelligent erzeugen könn-
ten, lässt doch gerade das Fehlen jedweden historischen Hinweises auf die Existenz
„präkambrischer Technologien“ diese Denkmöglichkeit höchst unplausibel erscheinen.7
Analoges gilt für natürliche Prozesse: Wenn ich beweisen kann, dass Proteine „von
selbst“ entstehen können, die irdischen Randbedingungen diese Möglichkeit aber nicht
hergäben, ließe sich nicht vernünftig auf eine chemische Evolution schließen.
Eine saubere Begründung für Design muss also folgende Punkte berücksichtigen:
7 Wie erwähnt gibt es zwei Ausnahmen: Finden wir Objekte, von denen wir von vorn herein wissen, dass
sie 1. erschaffen wurden oder 2., dass ihre Selbstorganisation unmöglich ist, ist Design evident. Bei
technischen Systemen sind in aller Regel beide Bedingungen erfüllt, bei Biosystemen keine einzige.
A. Nach unserem Wissen sind bestimmte Verfahrensweisen erforderlich, um
belebte Systeme gezielt zu verändern oder hervorzubringen.
B. Sprechen zwingende Gründe gegen die natürliche Evolution des Lebens?
C. Spricht Empirisches Wissen dafür, dass vor langer Zeit auf der Erde Tech-
nologien zum Einsatz kamen, die geeignet wären, Leben zu erzeugen?

- 19 -
C.8 Die Kritik des Philosophen Elliott SOBER
SOBER (2008) argumentiert, ein unbekannter Zufallsfaktor „erkläre“ die Herkunft des
Auges nicht weniger schlecht als ein Schöpfer, über dessen Absichten und Hand-
lungsoptionen wir nichts wissen. Es brauche zusätzliche Annahmen, um die spezifi-
schen Augenstrukturen mit einem theoretischen Entstehungsmodell logisch zu ver-
knüpfen (S. 144). Sinngemäß: Wer unspezifizierte „Design-Akte“ als Erklärung zulässt,
argumentiert nicht besser als jemand, der den blanken Zufall als „Erklärung“ für die
Herkunft biotischer Strukturen bemüht.
WIDENMEYER & JUNKER kontern mit einer Reductio ad absurdum, indem sie das Bei-
spiel des Auges exemplarisch durch das eines Computers ersetzen:
„Hypothese 1: Ein Computer ist durch intelligente Planung entstanden.
Hypothese 2: Ein Computer ist durch Zufall entstanden.
Entsprechend müsste SOBER folgern: ‚Da nach beiden Hypothesen ein Computer ent-
stehen kann, kann auf der bloßen Basis seiner Merkmale nicht entschieden werden,
welche Hypothese plausibler ist.‘ SOBER müsste jetzt konsequenterweise zusätzliche,
spezielle Merkmale fordern, die mit einer Design-These verbunden sind. Wir bräuchten
dann zusätzliches und unabhängiges Wissen über potentielle Computerhersteller und ih-
re (ggf. noch spezielleren) Absichten und Fähigkeiten (‚goals and abilities‘, s. o.) – über
die hochkomplexe, funktionale Struktur ihrer Produkte hinaus. Nur mit diesem Wissen
(wenn überhaupt!) könnte beurteilt werden, ob die Computer durch Zufall oder durch in-
telligente Planung entstanden sind.
Dieser Ansatz hat absurde Implikationen: SOBERs Prinzip bringt unter anderem mit sich,
dass man einfach behaupten könnte, dass Computer (oder beliebige andere technische
Gegenstände) durch Zufall entstehen könnten. Zudem: Woher wüssten wir von solchen
speziellen Absichten und Fähigkeiten des Herstellers? Nur über irgendwelche andere in
Raum und Zeit manifestierten Indizien: Wendet man SOBERs Ansatz konsequent an,
müsste man auch hier annehmen, dass sie durch Zufall zustande gekommen sein kön-
Merke: Was die Herkunft des Lebens anbelangt, so ist jedwede prähistorische Ver-
ortung des mutmaßlichen Designs gescheitert. Die historischen Wissenschaften
liefern keinerlei Hinweise auf intelligente Urheber oder Technologien, die auf der
frühen Erde hätten Leben hervorbringen können. Im Gegenteil: Die historischen
Randbedingungen lassen die Idee eines „präkambrischen Designers“ gänzlich un-
plausibel erscheinen. Intelligent Design ist nicht plausibler als die Phantasien eines
Erich von Däniken zur „Prä-Astronautik“, wonach Außerirdische vor Jahrtausenden
die Erde besucht und unsere zivilisatorische Entwicklung beeinflusst haben sollen.

- 20 -
nen, zumindest wenn nicht wieder weitere Indizien über die potenziellen Hersteller vor-
gebracht werden könnten. Und lägen diese vor, so könnten wieder weitere Indizien ge-
fordert werden, und dies ad infinitum. Es gibt hier zwei Hauptpunkte:
1. Egal, welche Indizien für einen letztlich beliebigen Sachverhalt vorliegen: Es
müssten nach SOBERs Ansatz immer noch weitere Indizien gefordert werden.
2. Wer behauptet, dass eine völlig unkonkrete Berufung auf „Zufall“ dieselbe Erklä-
rungskraft hat wie die Angabe einer konkreten systematischen Erklärung, der
leugnet effektiv die Testbarkeit jeder systematisch erklärenden These.
Im Gegensatz zu SOBERs Ansatz sind jedoch die komplexen konstruktiv-funktionalen
Merkmale als solche, die ein Computers oder jeder andere hochorganisierte Gegenstand
aufweist, für die Bevorzugung der Design-Hypothese völlig hinreichend: Es gibt sehr gute
Gründe, dass ein hochkomplex organisierter Gegenstand ein starkes Design-Indiz darstellt
(vgl. Abschnitt 2); und entsprechend gibt es keine guten Gründe, warum man für den
Schluss auf geistige Urheberschaft an sich zusätzliches Wissen über Motive und Fähigkei-
ten z. B. eines Computerherstellers bräuchte“ (S. 12).
Diese Erwiderung geht aus zwei Gründen fehl. Erstens: SOBERs Argument legt nicht
nahe, es sei realiter vernünftig anzunehmen, Computer entstünden durch Zufall. Viel-
mehr besagt das Argument, dass sich keine qualifizierte Entscheidung über die Frage
treffen lässt, welche Erklärung zutrifft. Das gilt, solange die erklärenden Faktoren (hin-
sichtlich des Wirkpotenzials und der Grenzen von Design) nicht durch wohlbegründete
Kenntnisse konkretisiert wurden. Grundsätzlich ließe sich dem unstrukturierten Zufall
genauso gut schöpferische Allmacht zuschreiben, wie einem unkonkreten Designer.
Denken wir an die nach dem Physiker Ludwig BOLTZMANN benannte Idee, dem Quan-
ten-Vakuum könne zufällig etwas so Komplexes wie ein Gehirn entspringen („BOLTZ-
MANN-Gehirn“). Wäre das eine zulässige Erklärung? Natürlich nicht.
SOBER zufolge ist der Verweis auf den Zufall nicht besser und nicht schlechter
als der Verweis auf einen dubiosen „Schöpfer“, über dessen Wirkgrenzen wir
nichts wissen. Ein solcher Joker ließe sich bei der Suche nach Erklärungen im-
mer bemühen. Weder die eine noch die andere „Erklärung“ liefert Mechanismen, die
eine logische Brücke zu den erklärungsbedürftigen Sachverhalten schlagen. Indem
also WIDENMEYER & JUNKER die Absurdität einer – sagen wir – „Zufallserklärung“ auf-
zeigen, führen sie ihren Lesern auch die Absurdität des Design-Arguments vor Augen.
Zweitens: Das von WIDENMEYER & JUNKER gewählte Beispiel enthält die von SOBER gefor-
derten Hilfsannahmen zur Konkretisierung des Schöpfungsvorgangs bereits in Form ver-
steckter Prämissen! Es handelt sich um das „unabhängige Wissen über potentielle Com-
puterhersteller und ihre (ggf. noch spezielleren) Absichten und Fähigkeiten…“.

- 21 -
Konkret: Wir wissen, dass Computer Menschenwerk sind. Wir wissen, welchen Zwecken
sie dienen. Wir wissen, wie sie sich herstellen lassen. Wir wissen, dass Computer nicht
evolvieren konnten. Und wir wissen, dass die Quantenmechanik den Zufall als realistische
Erklärung aussondert. Erst im Lichte dieses Wissens erscheint das Anliegen, den
Zufall als gleichberechtigte Alternative zum Design ins Auge zu fassen, absurd. Da-
her ist der von den Autoren konstruierte Vergleich irreführend. Ein Außerirdischer, dem
dieses Wissen fehlte, wäre nicht imstande, allein dem Ergebnis seiner Struktur-Funktions-
Analyse zu entnehmen, was Computer sind, geschweige denn, wie sie entstanden.
D. Zielscheibenfehler: Plastizität und „programmierte Variabilität“ als
Design-Merkmale
Die Individual-Entwicklung (Ontogenese) von Lebewesen ist plastisch. Darunter ver-
stehen wir, dass in Organismen ein enormes Variations-Potenzial steckt. Dieses tritt
durch bestimmte Umweltreize, Eingriffe in die Embryonal-Entwicklung oder durch Mu-
tationen zu Tage. Zum Beispiel ist das zentrale Nervensystem erstaunlich anpas-
sungsfähig: Die Funktionen ausgefallener Hirnbereiche übernehmen zu einem gewis-
sen Grad andere Hirnareale, und das Fehlen eines Sinns wird teils durch andere Sinne
kompensiert. Blinde orientieren sich oft gut mittels Reflexion von Geräuschen (Echoor-
tung). Transgene Mäuse erlernen dreifarbiges Sehen (vgl. JACOBS et al. 1999), usw.
Ein anderes Beispiel: Eine Behandlung von Schlammspringer-Embryos mit dem Hor-
mon Thyroxin hat vielschichtige Auswirkungen auf den Phänotyp. Die Brustflossen des
Fisches entwickeln sich zu beinchenartigen Extremitäten, die Haut wird dicker, die
Merke: Indem WIDENMEYER & JUNKER auf die Irrationalität beliebiger Zufallserklä-
rungen hinweisen, führen sie ihren Lesern unbeabsichtigt die Absurdität von Intel-
ligent Design vor Augen. Denn beide „Erklärungen“ sind methodologisch gleich-
wertig. Wer unbekanntes, gar wundersames Design zulässt, hat kein Argument,
um eine dubiose „Zufallsevolution“ zurückzuweisen. (Letztere hat, wohlgemerkt,
mit der Evolutionstheorie nichts zu tun, denn ihre Mechanismen sind spezifiziert
und empirisch belegt.)

- 22 -
Kiemen kleiner, die Luftatmung nimmt zu usw. Im Ergebnis können sich die Fische
länger außerhalb des Wassers aufhalten als normal (LORENZEN 1988).8
Auf zellulärer, organischer und entwicklungsgenetischer Ebene gibt es zahlreiche der-
artige „Stellschrauben“, die bestimmte Anpassungen erschweren oder ermöglichen
und somit evolutionäre Weichen stellen. Ein Grund ist, dass komplexe Entwicklungs-
prozesse nicht direkt genetisch codiert sind. Vielmehr organisieren sie sich selbst
durch wechselseitige Beeinflussung von Embryonalzustand und Genaktivierung.
Oft wirken kleine „Inputs“ wie „Signale“, die den Phänotyp vielschichtig ändern und die
Ontogenese in bestimmte Richtungen drängen. Beispielsweise verlängerten sich durch
eine Mutation bei den Vorfahren der Fledermäuse die Knochen ihrer Vordergliedma-
ßen. Da die ontogenetische Entwicklung „konzertiert“ abläuft, bedurfte es keiner weite-
ren Mutationen, um Blutgefäße, Haut, Muskulatur, Sehnen und Nerven passend zu
verlängern. Dies geschieht automatisch. Vergleichbares beobachten wir bei Hunden:
Ein erheblich modifizierter Körperbau erfordert wenige Mutationen, oft nur eine einzige.
KIRSCHNER & GERHART (2007) sprechen in diesem Zusammenhang von phänotypi-
scher Plastizität. Paradoxerweise deuten WIDENMEYER & JUNKER diese Plastizität in ih-
rem Sinne um: Aus einem natürlichen Phänomen, das evolutionäre Entwicklungen er-
leichtert, wird kurzerhand ein Schöpfungsindiz:
„Unter Plastizität (Formbarkeit) wird die Fähigkeit von Organismen verstanden, auf der
Basis desselben Genotyps (Erbguts) mehrere Phänotypen (gestaltliche Ausprägungen)
als Reaktion auf Umweltreize ausbilden zu können. Beispielweise kann beim Menschen
die Dicke der Hornhaut an den Händen oder Füßen abhängig von mechanischer Bean-
spruchung moduliert werden. Man weiß heute, dass ein Großteil der Merkmale der Le-
bewesen plastisch ist. Plastizität ist ein ausgesprochen teleologisches Konzept. Denn sie
beinhaltet die Fähigkeit des Organismus, auf genetische oder Umwelt-Änderungen zu
reagieren, um einen bestimmten Zustand aufrechtzuerhalten oder (wieder) zu erreichen.
Es wird also aktiv ein Ziel angesteuert oder beibehalten, indem Änderungen durch Kom-
pensationen ausgeglichen werden“ (S. 7).
Einer der Autoren (JUNKER 2014) vertritt gar die These, die Biologie führe durch Be-
rücksichtigung der Plastizität eine Art Zielorientierung in die Evolutionstheorie ein. Die
Frage nach einem Schöpfer stelle sich weiterhin, da zu erwarten sei, dass der wissen-
8 Entscheidend für die Evolutionstheorie ist, dass solche nichterblichen Modifikationen unter bestimmten
Voraussetzungen erblich werden können. So führen, um beim Schlammspringer zu bleiben, bestimmte
Mutationen zu Erhöhungen des Thyroxin-Spiegels.

- 23 -
schaftliche Fortschritt Hinweise auf geistige Ursachen liefere. In verschiedenen Publi-
kationen spricht der Autor diesbezüglich von „programmierter Variabilität“.
Gegen JUNKERs These greift Einwand C.1. Selbst wenn Organismen ein Ziel ansteuern
würden, folgt nicht, dass es einen Zwecksetzer brauchte. Dass auf natürliche Weise An-
passungen zustande kommen, die „antizipatorisch“ anmuten, ist empirisch gezeigt (vgl.
Fußnote 3). Die These, sie seien geistig programmiert, ist nichts als Spekulation.
Bereits aus systemtheoretischen Gründen weisen biotische Strukturen eine ho-
he Variabilität auf. Da sie durch ein komplexes Netz von sich gegenseitig beeinflus-
senden Mechanismen zustande kommen, sind auch die Auswirkungen genetischer
und äußerer Zustands-Änderungen komplex und vielfältig. Ein Teil der durch sie aus-
gelösten Variationen fällt in den Bereich der Teratologie, der Lehre von den körperli-
chen und organischen Fehlbildungen. Aber ein kleiner Teil der möglichen Variationen
hat unter bestimmten Voraussetzungen adaptiven Charakter.
Potenziell vorteilhafte Merkmale schlummern im Verborgenen: Manchmal zeigt sich,
dass Mutationen, die im Nachhinein als notwendige Bedingungen für bestimmte An-
passungen erkannt werden, allein noch keinen Effekt zeigen. Manchmal verschwinden
sie, ohne dass eine Anpassung zustande kam. Und zeigen sie, zusammen mit weite-
ren Mutationen, einen Effekt, bringen sie ihren Besitzern nicht unbedingt einen Vorteil.
Erst wenn die neuen Merkmale besser zur Umwelt passen oder ihr Besitzer aktiv ein
passenderes Habitat besetzt, liegt eine Anpassung vor. Doch oft verschwinden die
vorteilhaften Genvarianten durch genetische Drift – und die Neuerung ist dahin.
Das heißt: Ob bestimmte Variationen Bestand haben oder nicht, ist das Ergebnis
glücklicher Zufälle und verpasster Chancen. Anpassung zeigt sich retrospektiv, inso-
fern erweisen sich Entwicklungspotenzen nicht als vorausschauend! 99% der
Arten sind wieder ausgestorben, weil sie mit den ökologischen Veränderungen nicht
Schritt halten konnten. Oder die genetischen Änderungen wirkten sich langfristig nach-
teilig auf die Organismen aus. Mit Planung hat all dies herzlich wenig zu tun.
Intelligent-Design-Vertreter argumentieren wie Leute, die in SHAKESPEAREs Sonetten
geheime Botschaften entziffert zu haben glauben. Sie richten ihr Augenmerk nur auf
Passagen, für die bestimmte Dechiffrier-Algorithmen zufällig leserliche Resultate erga-
ben.9 Das ist als würde jemand blind Gewehrkugeln auf ein Scheunentor abfeuern, um
9 Siehe auch: Zielscheibenfehler (de.wikipedia.org/wiki/Zielscheibenfehler).

- 24 -
einige Einschüsse eine Zielscheibe herum pinseln und behaupten, er habe „ins
Schwarze“ getroffen. Die übrigen Schüsse werden nicht berücksichtigt. Dieser Denk-
fehler ist in der informellen Logik unter der Bezeichnung „Fehlschluss des texani-
schen Scharfschützen“ geläufig (Abb. 6).
Abb. 6 Intelligent-Design-Vertreter bege-
hen den Fehlschluss des „texanischen
Scharfschützen“: Zufällig Passendes wird
als Ergebnis einer Programmierung gedeu-
tet, Unpassendes, wie etwa Fehlbildungen,
der degenerativen Evolution angelastet.10
Fazit: Der Terminus „programmierte Variabilität“ ist eine Erfindung, die nur in Schöp-
fungsparadigmen Sinn ergibt. Auf Erfahrungswerten beruht er nicht. Das Konzept ist
auch nicht unabhängig von Design prüfbar: Wie ließe sich bestimmen, welche latenten
Phänotypen und welches Maß an Variabilität für welche Ziele „programmiert“ wurden?
Wer latent vorteilhafte Entwicklungspotenziale als Ergebnis von Planung deutet, sieht
sich mit absurden Konsequenzen konfrontiert: Der Evolutionshistoriker Thomas JUN-
KER (2004) verweist darauf, dass der Botaniker Asa GRAY, ein Freund DARWINs, in den
1860er Jahren eine ähnliche Ansicht vertrat. Er glaubte,
„… die Selektionstheorie mit religiösen Ideen vereinbaren zu können, indem er annahm,
dass die Variationen, die das Material für die natürliche Auslese darstellen, nicht zufällig,
sondern von Gott geplant seien. Solange ‚die physische Ursache der Variation völlig unbe-
kannt und geheimnisvoll ist,‘ könne man davon ausgehen, dass die ‚Variation an bestimm-
ten vorteilhaften Routen entlang geleitet wurde‘. Darwin entgegnete, dass man Grays Mo-
dell zufolge auch annehmen müsse, dass Gott die unzähligen Variationen der Haustiere
und Kulturpflanzen speziell für den Nutzen der Züchter vorherbestimmt habe; dass bei-
spielsweise Kropf und Schwanzfedern der Tauben variieren, damit die Taubenliebhaber ih-
re grotesken Formen züchten können, und dass Hunde in ihren geistigen Fähigkeiten vari-
ieren, damit man Kampfhunde züchten könne“ (Thomas JUNKER 2004, S. 11).
DARWINs Entgegnung veranschaulicht die Willkür dieses Konzepts. Die Annahme, ge-
genwärtige oder in der Zukunft liegende Anpassungen seien „voraus programmiert“, ist
10 Evidenzbasiert können wir auch umgekehrt argumentieren. Beispielsweise zeigt die Evolutionäre
Biotechnologie, dass das Wechselspiel aus Variation und Selektion bei der Erzeugung funktionaler
Biomoleküle ein „designfähiger Prozess“ ist (SCHUSTER 2014, S. 161). Wie SCHUSTER erläutert, sind
evolutionäre Algorithmen in bestimmter Hinsicht dem „rationalen Design“ überlegen (S. 162).

- 25 -
eine raffinierte Schutzhypothese. Sie dient dem Zweck, den Design-Ansatz an die
„unbequeme“ Tatsache der (beobachtbaren) Evolution anzupassen.
E. Das Argument der Konstruktionsfehler
Ein weiteres Argument gegen Intelligent Design betrifft sinnlose Umwege der Individu-
al-Entwicklung und funktionelle Mängel, die mit einer Herabsetzung der Fitness ihrer
Besitzer einhergehen. In einigen Fällen sind sie lebensgefährlich. So erfolgt die Geburt
just durch den nicht zu erweiternden Beckenring, der eine Steißgeburt ohne medizini-
schen Beitrag zur tödlichen Komplikation werden lässt. Die Kreuzung von Luft- und
Speiseröhre birgt die Gefahr der Aspiration mit potenziell tödlichem Ausgang. Die, ge-
messen am aufrechten Gang, verkehrt positionierten Abflusswege der Nasenneben-
höhlen verursachen Entzündungen und erhebliche gesundheitliche Risiken. Und ein
perforierter Wurmfortsatz gehörte bis ins Mittelalter zu den häufigsten Todesursachen.
Ein anderes Beispiel: Der dioptrische Apparat ist alles andere als der Paradefall eines
clever entworfenen Organs. Mindestens 40% der Erwachsenen sind auf eine Brille
angewiesen, da die Bildebene jenseits der Netzhaut liegt. Zudem sind bei den Wirbel-
tieren die Sehzellen dem Licht abgewandt (invers), während die Nervenzellen, die zum
Gehirn führen, nach außen gerichtet sind (Abb. 7). Die Folgen sind ein blinder Fleck
auf der Netzhaut sowie Blutgefäße und Nerven-Verdrahtungen vor (!) den Sehzellen.
All dies verlangt erhebliche Kompensationsleistungen von Auge und Gehirn.
Merke: „Programmierte Variabilität“ ist kein empirisches Faktum, sondern eine ge-
gen Überprüfung resistente Deutung, die ausschließlich im Schöpfungsparadigma
Sinn ergibt. Daher kann es sich um kein unabhängiges Design-Indiz handeln.
Aus der Tatsache, dass Lebewesen Variabilität und Plastizität zeigen, folgt nichts
für den Design-Ansatz. Vielmehr lässt sich daraus ein Argument gegen bestimmte
Formen der Evolutionskritik entwickeln: Plastizität erleichtert eine synorganisierte
Evolution (vgl. LORENZEN 1988). ID-Vertreter deuten diese jedoch in ihrem Sinne
um und stützen damit ihre Behauptung, dass Anpassungen, die sie „antizipato-
risch“ nennen, einen vorbereitenden Designer brauchen. Damit setzen sie voraus,
was es unabhängig vom Design-Ansatz zu belegen gilt.
- 26 -
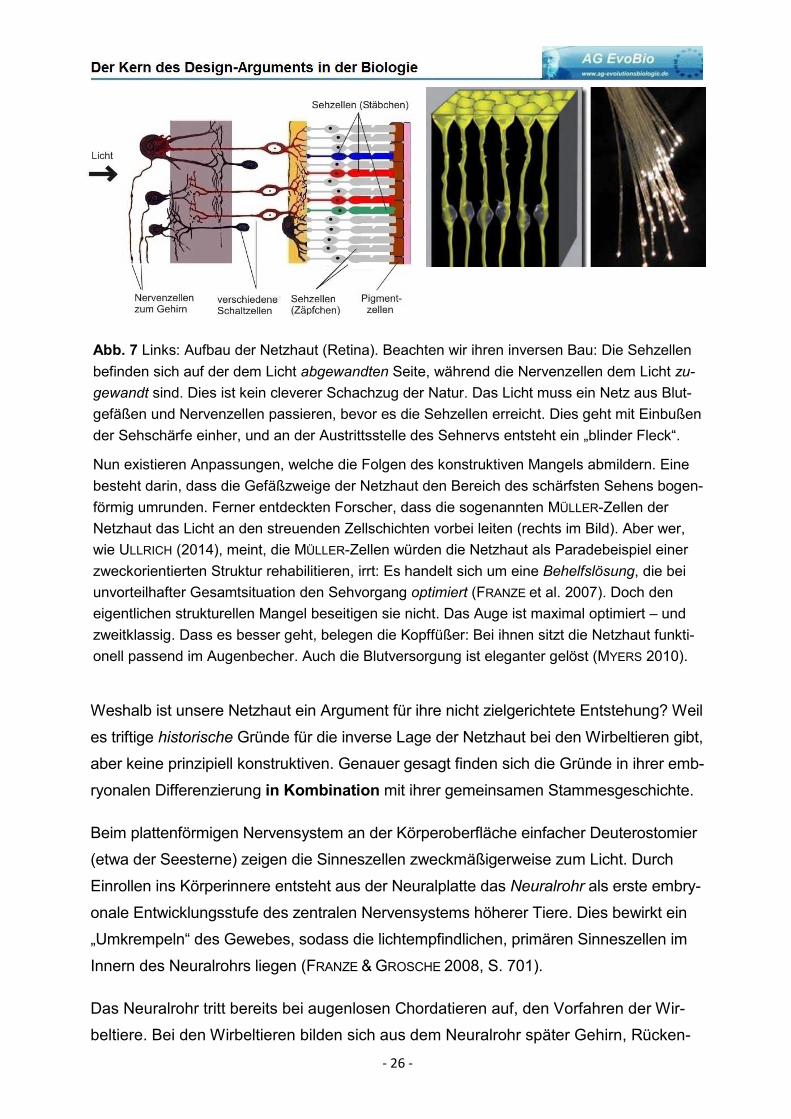
Abb. 7 Links: Aufbau der Netzhaut (Retina). Beachten wir ihren inversen Bau: Die Sehzellen
befinden sich auf der dem Licht abgewandten Seite, während die Nervenzellen dem Licht zu-
gewandt sind. Dies ist kein cleverer Schachzug der Natur. Das Licht muss ein Netz aus Blut-
gefäßen und Nervenzellen passieren, bevor es die Sehzellen erreicht. Dies geht mit Einbußen
der Sehschärfe einher, und an der Austrittsstelle des Sehnervs entsteht ein „blinder Fleck“.
Nun existieren Anpassungen, welche die Folgen des konstruktiven Mangels abmildern. Eine
besteht darin, dass die Gefäßzweige der Netzhaut den Bereich des schärfsten Sehens bogen-
förmig umrunden. Ferner entdeckten Forscher, dass die sogenannten MÜLLER-Zellen der
Netzhaut das Licht an den streuenden Zellschichten vorbei leiten (rechts im Bild). Aber wer,
wie ULLRICH (2014), meint, die MÜLLER-Zellen würden die Netzhaut als Paradebeispiel einer
zweckorientierten Struktur rehabilitieren, irrt: Es handelt sich um eine Behelfslösung, die bei
unvorteilhafter Gesamtsituation den Sehvorgang optimiert (FRANZE et al. 2007). Doch den
eigentlichen strukturellen Mangel beseitigen sie nicht. Das Auge ist maximal optimiert – und
zweitklassig. Dass es besser geht, belegen die Kopffüßer: Bei ihnen sitzt die Netzhaut funkti-
onell passend im Augenbecher. Auch die Blutversorgung ist eleganter gelöst (MYERS 2010).
Weshalb ist unsere Netzhaut ein Argument für ihre nicht zielgerichtete Entstehung? Weil
es triftige historische Gründe für die inverse Lage der Netzhaut bei den Wirbeltieren gibt,
aber keine prinzipiell konstruktiven. Genauer gesagt finden sich die Gründe in ihrer emb-
ryonalen Differenzierung in Kombination mit ihrer gemeinsamen Stammesgeschichte.
Beim plattenförmigen Nervensystem an der Körperoberfläche einfacher Deuterostomier
(etwa der Seesterne) zeigen die Sinneszellen zweckmäßigerweise zum Licht. Durch
Einrollen ins Körperinnere entsteht aus der Neuralplatte das Neuralrohr als erste embry-
onale Entwicklungsstufe des zentralen Nervensystems höherer Tiere. Dies bewirkt ein
„Umkrempeln“ des Gewebes, sodass die lichtempfindlichen, primären Sinneszellen im
Innern des Neuralrohrs liegen (FRANZE & GROSCHE 2008, S. 701).
Das Neuralrohr tritt bereits bei augenlosen Chordatieren auf, den Vorfahren der Wir-
beltiere. Bei den Wirbeltieren bilden sich aus dem Neuralrohr später Gehirn, Rücken-

- 27 -
mark und Augen. Unter der Herrschaft des PAX-6-Mastergens quellen aus dem hinte-
ren Bereich des Vorderhirns Wülste hervor, die sich zum Augenbecher umformen
(MÜLLER & HASSEL 2018, S. 475). Aufgrund dieses Entwicklungszwangs (engl.: deve-
lopmental constraints) wird die Netzhaut der Inversion unterzogen (Abb. 8).
Abb. 8 Entwicklung der inversen Netzhaut, ausgehend vom platten Nervensystem einfacher
Deuterostomier (z. B. Seesterne) zum ins Körperinnere eingesenkten Neuralrohr als Anlage
des Gehirns und Rückenmarks. Umgezeichnet nach FRANZE & GROSCHE (2008, S. 701).
Jene embryogenetische Weichenstellung, die die Inversion der Netzhaut herbeiführt,
haben also schon die Vorfahren der Chordatiere „getroffen“. Sie geht auf eine Zeit zu-
rück, bevor sich die Entstehung von Wirbeltieraugen abzeichnete. Diese „Vorentschei-
dungen“ ließen später keine Alternativen mehr zu. Denn ihr genetisches Erbe zu modi-
fizieren, bis die Sehzellen „richtig“ im Augenbecher liegen, machte es erforderlich, die
embryonale Anlage des Nervensystems von Grund auf umzugestalten.
Für Prozesse, die keinen Plan kennen, ist das nicht leistbar, und die ersten Wirbeltie-
rembryos, die Augen entwickelten, konnten nicht vorübergehend „wegen Umbaus
schließen“. Also blieb die Differenzierung von Stammzellen des Neuralrohrs in ihrer
besonderen topographischen Lage zu den Sinneszellen wie sie war.
Dagegen lässt sich nicht ansatzweise erklären, warum sich ein Designer Entwick-
lungszwängen unterwarf, die ihren Ursprung in historisch älteren Bauplänen haben. Er
soll doch in der Lage sein, überlegt zu handeln, die Merkmale planvoll, zielgerichtet
und „frei“ zu kombinieren. Warum soll er die Vorläufer-Sinneszellen erst ins Neuralrohr
wandern lassen, um sie dann invers aus dem hinteren Bereich des Vorderhirns her-
vorquellen zu lassen? Das ergibt nach gegenwärtiger Sachlage keinen Sinn.

- 28 -
Die Entwicklungsbiologen MÜLLER & HASSEL (2018, S. 627) resümieren:
„Die (nahezu) perfekte Konstruktion bestimmter Augen verleitet manche Menschen zu
der Annahme, dass ein intelligentes Wesen diese Augen geschaffen haben müsse, na-
türliche Evolution könne sie nicht hervorgebracht haben. Zunehmendes Wissen um Ho-
mologie auf molekularem Niveau, um Zwischenstufen in der Konstruktion wie auch um
suboptimale Lösungen (z. B. inverses Auge der Wirbeltiere, schlechte optische Qualität
des dioptrischen Apparates) machen Annahmen über ‚intelligentes Design‘ aber keines-
wegs zwingend. Im Gegenteil unterstützen die Daten … die Sicht, dass bekannte Me-
chanismen der Evolution letztlich eine Vielfalt brauchbarer Lösungen hervorbrachten.“
E.1 Einwände der Autoren
WIDENMEYER & JUNKER kontern mit folgenden Einwänden:
„Erstens zeigt eine Reihe von Untersuchungen, dass bei genauerer Kenntnis betreffen-
der Organe nennenswerte Mängel nicht nachweisbar sind. Dem Argument der Konstruk-
tionsfehler liegt oft nur ein Mangel an Kenntnissen über den betreffenden Gegenstand
zugrunde – und, wie es scheint, weltanschaulich motivierte Vorurteile.
Zweitens enthalten solche Argumente bestimmte (theologische) Annahmen über einen
hypothetischen Schöpfer, z. B. dass ein möglicher Schöpfer gute, wenn nicht zwingende
Gründe gehabt haben müsste, perfekte Strukturen (in unserem Sinne) hervorzubringen.
Diese meist stillschweigend gemachten Annahmen werden meistens nicht einmal ver-
sucht zu begründen (vgl. DILLEY 2013)“ (S. 9).
Bei JUNKER (2005) lesen wir, Unvollkommenheiten seien „kaum empirisch nachweisbar,
sondern … evolutionstheoretisch begründete Vermutungen, deren Plausibilität mit der
evolutionstheoretischen Voraussetzung steht oder fällt.“ Wie er zu dieser Auffassung kam,
ist unklar, jedenfalls ist sie falsch: Wenn wir die immanenten Funktionen der betreffenden
Systeme verstehen, sind wir in der Lage, qualifizierte Urteile über ihre Mängelstrukturen
zu fällen (KOJONEN 2016, S. 159). Dafür braucht es keine evolutiven Annahmen.
Konstruktive Mängel lassen sich an der verminderten Fitness von Organismen festma-
chen. Zum Beispiel kommt es aufgrund des Überkreuzens von Luft- und Speiseröhre, ins-
besondere bei Kleinkindern, regelmäßig zur Aspiration von Fremdkörpern und zum Ersti-
Merke: Anand biologischer Konstruktionsfehler lassen sich zwei Argumente entfalten:
(1) Intelligent Design kann die unvorteilhafte Gesamtsituation nicht zwanglos erklä-
ren, sondern ist auf willkürliche Zusatzannahmen angewiesen.
(2.) Die Mängel deuten auf einen nichtintendierten Entwicklungsprozess hin.

- 29 -
cken. Und der entzündete Wurmfortsatz des Blinddarms erhöhte früher die Sterblichkeit
deutlich, wobei die vermeintlichen Vorteile des Organs den Nachteil nicht wettmachen.
Allein aus medizinisch-technischer Sicht erweisen sich die Merkmale als nach-
teilig, reparaturbedürftig und alles andere als ein Ausweis an Planmäßigkeit.
Auch die Behauptung, dem Argument der Konstruktionsfehler läge ein Mangel an
Kenntnissen über den betreffenden Gegenstand zugrunde, trifft meist nicht zu. Die
inverse Retina gestaltet das Sehen suboptimal, wenngleich sie das unter den Umstän-
den Beste aus dem Sehvorgang macht.11 Beim Blinddarm lässt sich sogar beweisen,
dass sein Besitz mehr schadet als nützt (BURDA & BEGALL 2013, S. 350).
WIDENMEYER & JUNKERs zweiter Einwand läuft ebenfalls ins Leere. Auch wenn sie sich auf
den Philosophen Stephen DILLEY (2013) berufen: Einwände gegen Design, wie das Ar-
gument der Konstruktionsfehler, setzen nicht zwangsläufig theologische Annahmen vo-
raus. Zumindest benötigen wir keine, die Design-Vertreter nicht selber ins Spiel bringen.
Hier gilt es, zwei Spielarten des Arguments der Konstruktionsfehler zu unterschei-
den: Die sparsamere Variante setzt nicht voraus, ein Schöpfer habe zwingend perfekte
Strukturen hervorgebracht. Sie besagt nur, dass sich bestimmte Merkmals-Klassen mit
Design nicht erklären lassen, solange der Design-Ansatz nicht spezifiziert wird. Erste-
res ist ein theologisches Argument, letzteres ein methodologisches.
Die zweite, stärkere Variante des Arguments nimmt zwar auf das behauptete Schöp-
ferwirken Bezug. Doch sie gebraucht keine theologischen Annahmen; vielmehr orien-
tiert sie sich an den ureigenen Thesen des intelligenten Designs. Erinnern wir uns: De-
sign-Vertreter führen die „Zweck-Mittel-Beziehungen“ und „funktionale Komplexität“
biotischer Strukturen als Design-Merkmale an. Sie vergleichen das vermeintliche
„Knowhow“ mit Meilensteinen aus Medizin und Technik. Daher liegt es in der Natur
der Sache, dass Elemente, welche die Zweck-Mittel-Beziehungen stören, ihr Ar-
11 Eine weitreichende Kompensation der Sehschärfe-Einbußen ermöglichen auch die erwähnten MÜL-
LER-Zellen nicht. Andernfalls wäre die Gefäßarmut im Bereich des schärfsten Sehens unbegründet.
Zahlreiche Vögel verzichten auf eine vaskularisierte Netzhaut. Über ihren Fotorezeptoren winden sich
keine Blutgefäße, dadurch sehen sie schärfer. Zudem haben die MÜLLER-Zellen einen gravierenden
Nachteil: Sie neigen zur Hypertrophie, zur Bildung von Verdickungen und vermehrten Fortsätzen
(BRINGMANN & REICHENBACH 2001). Die sogenannte „Gliose“ führt unbehandelt zur Degeneration der
Netzhaut bis hin zum völligen Sehverlust.

- 30 -
gument schwächen: Teilstrukturen, die aus medizinisch-technischer Sicht nicht von
Zweckmäßigkeit zeugen, erwecken nicht den Anschein intelligenter Planung.12
Zwar ist es richtig, dass mit dem unspezifischen Design-Ansatz jegliche Art von Pfusch
verträglich wäre (↑ Abschnitt C.4). Es gibt unintelligente Designer, und schlechtes De-
sign wäre trotz allem Design (LUSKIN 2006). Doch ID-Vertreter stellen meist weit höhe-
re Ansprüche an die Qualität biotischer Strukturen als ein allgemeiner Design-Ansatz,
der auch stümperhaftes und sogar evolutionäres Design einschließen könnte!
KOJONEN (2016, S. 152f) bemerkt, dass schlechtes Design logisch mit einem Designer
kompatibel wäre. Im Weiteren betont er jedoch, dass dieser Einwand nicht das
Problem löst, dass schlechtes „Design“ besser zum unserem evolutionären Ver-
ständnis von der Welt passen könnte als zu planvollen, überlegten Prozessen.
Auch RAMMERSTORFER (2006) sieht das Problem. Suboptimale Strukturen oder „banale
Fehler“ würden den Design-Ansatz schwächen, denn:
„Stümperhaftes Design passt insgesamt (wie oben besprochen) hervorragend zu Pro-
zessen, die keinen Plan kennen. Damit würde auf dieser Ebene (‚Design-Fehler‘) ein gu-
tes Argument für Evolution und gegen Planung existieren“ (S. 90).
Das ist der Punkt. Machen wir uns klar: Würde ein Ingenieur die Strukturen des Auges
auf ein optisches Instrument übertragen, müsste er den Lichtsensor verkehrt in das
Gerät einbauen. Die ableitenden Kabel lägen im Strahlengang. Dann müsste er ein
Loch durch den Sensor bohren, um das Kabelgewirr hinter die optische Anordnung zu
führen. Das ist, um mit RAMMERSTORFER zu sprechen, ein „banaler Fehler“, der „her-
vorragend zu Prozessen“ passt, „die keinen Plan kennen“.
Wie befreit sich RAMMERSTORFER aus diesem Dilemma? Zunächst, indem er leugnet,
dass sich die inverse Retina nachteilig auf die Funktion des dioptrischen Apparats
auswirkt. Er zitiert Leute, die ernsthaft glauben, die Gesamtsituation sei vorteilhafter.
So würden die Netzhaut effektiver mit Blut versorgt, Stoffwechselabfälle besser ent-
12 Krass, dass die Autoren ihren Widerspruch nicht bemerken: Auf der einen Seite unterstellen sie De-
sign-Kritikern, sie würden „theologisch“ argumentieren, wenn sie feststellen, Suboptimalität spräche
gegen Design. Andererseits schreiben sie auf S. 2f, das „Merkmalsmuster“ geistig hervorgebrachter
Gegenstände sei „hochgradig speziell“ ausgeführt, „damit es (möglichst optimal) [sic!] entsprechende
Funktionen ausüben kann, also einer Zweck-Mittel-Beziehung entspricht“.
Hier hat KOJONEN (2016, S. 157) Recht, wenn er sagt: “Wenn ID-Vertreter tatsächlich manchmal auf die
Güte, Schönheit und auf den Grad der Perfektion der natürlichen Ordnung abheben, wäre es inkonsis-
tent, sich nicht auch dem Problem des schlechten Designs zu stellen“ (ins Deutsche M. N.).

- 31 -
sorgt (S. 71f). Als ob das ein Argument wäre: Die Netzhaut scharf sehender Vögel ist
aus gutem Grund nicht vaskularisiert. Trotzdem ist die Nährstoff-Versorgung effektiv.
In einer anderen Schrift (RAMMERSTORFER 2004, S. 3) behauptet er, aufgrund mangeln-
den Wissens über Entwicklungsbiologie sei derzeit „keine Urteilsbasis gegeben“, um auf
Suboptimalität zu schließen. Merkmale, die aus technischer Sicht wie Pfusch ausse-
hen, könnten aus entwicklungsbiologischer Sicht den bestmöglichen Kompromiss
darstellen. Denn die Ontogenese ist komplizierter als das „Verschrauben“ von Einzeltei-
len. Bei technischen Dingen entsteht die Funktion erst am Ende des Fertigungsprozes-
ses. Dagegen muss der Embryo in jedem Stadium seiner Entwicklung „funktionieren“.
Und er muss die Voraussetzungen für die jeweils nächsten Entwicklungsschritte schaf-
fen. Alles hängt mit allem zusammen. Daher ist es vorstellbar, dass ein Designer kurios
anmutende Strukturen in Kauf nahm, weil alternative Entwicklungswege gravierende
Probleme an anderer Stelle aufgeworfen hätten. Aber dieser Einwand muss scheitern.
Mit der Frage, ob derzeit eine entwicklungsbiologische „Urteilsbasis“ existiert, steht und
fällt auch das Design-Argument: Gibt es sie nicht, wissen wir auch nicht, ob die Teile
eines Systems „hochgradig speziell ausgeführt sind“ und inwieweit sie „einer Zweck-
Mittel-Beziehung“ entsprechen. Ähnlich argumentiert KOJONEN (2016, S. 159). Außer-
dem ist die Vermutung, dysfunktionale Strukturen seien die bestmöglichen Kompromis-
se, eine Schutzhypothese mala fide. Um beim Auge zu bleiben: Wir wissen, dass ele-
gantere Alternativen existieren. Der Augenbecher des Oktopusses quillt nicht aus sei-
nem Gehirn, sondern entsteht durch Einstülpung der embryonalen Außenhaut. Dann
versorgt ihn das Gehirn mit Nervenzellen. Daraus folgt, dass das Problem nicht kon-
struktionsbedingt ist. Das Problem ist, dass Embryos „unnötig erscheinende Remi-
niszenzen an die evolutive Vergangenheit“ zeigen (MÜLLER & HASSEL 2018, S. 506).
WIDENMEYER & JUNKER erheben den Einwand, auch die Evolutionstheorie würde keine
Erklärungen für Dysfunktionalität liefern. Stattdessen würden
„lediglich spekulative und pauschale evolutionstheoretische Szenarien postuliert, durch
die es zum betreffenden fehlkonstruierten Organ gekommen sein könnte – denen aber
erklärende … Inhalte und Belege weitgehend fehlen“ (S. 10).
Doch die Güte kausaler Erklärungen hat mit unserem Argument nichts zu tun! Wie er-
wähnt geht es darum, dass „stümperhaftes Design“ gut zu planlosen Prozessen passt.
Es bestätigt die Erwartungen natürlicher Evolution, passt aber nicht ohne weiteres zu
planmäßigem Handeln. Daher benötigen die ID-Vertreter passende Hilfsannahmen: Sie

- 32 -
mutmaßen, es könnte embryogenetische „Sachzwänge“ geben, die alternative Entwick-
lungsrouten ausschließen. Manche postulierten, der Designer arbeite wie ein Züchter im
Rahmen historisch limitierter Evolutionsprozesse (KOJONEN 2016, S. 157). Solche Ein-
wände haben Ad-hoc-Charakter. Zudem steht der Letztgenannte, wie KOJONEN bemerkt,
nur jener Minderheit an ID-Vertretern offen, die gemeinsame Abstammung akzeptieren.
Wir sehen: Sobald Analogien nicht ins Design-Konzept passen, leugnen sie die ID-
Vertreter. Dann behaupten sie, Imperfektes sei Ausdruck einer Kompromisslösung. Sie
legen nahe, das Imperfekte läge im Ermessensspielraum des Designers. Oder sie be-
haupten, Suboptimalität lasse sich nicht nachweisen. Wir haben gezeigt, dass es sich
um schwache Versuche von Kritikimmunisierung handelt.
Merke:
(1) Urteile über konstruktive Mängel bedürfen keiner evolutionären Interpretati-
on. Mängelstrukturen verursachen Fitness-Einbußen und erhöhte Mortalität.
Sie zeigen Merkmale, die aus technischer Sicht unsinnig erscheinen und der
Idee der Planmäßigkeit im Weg stehen.
(2) Das Argument der Konstruktionsfehler setzt keine theologischen Annahmen
voraus. Seine sparsamere Variante besagt lediglich, dass sich Mängel-
Strukturen nur mit natürlichen Prozessen zwanglos erklären lassen.
(3) Die starke Variante des Arguments nimmt zwar auf das behauptete Schöp-
ferwirken Bezug. Dafür wendet sie jedoch die Thesen des Intelligent Designs
an: Da ID-Vertreter mit Technik-Analogien arbeiten, ist es legitim, sich die-
sen Analogien zu stellen und zu prüfen, ob sie das Planmäßigkeits-
Argument schwächen. Denn: „Stümperhaftes Design passt … hervorragend
zu Prozessen, die keinen Plan kennen“ (RAMMERSTORFER 2006, S. 90.)
(4) „Die Planmäßigkeitsanaloge streicht den Sinn in der Natur heraus und
übersieht den Unsinn. Auf gleiche oder ähnliche [= intelligente] Ursachen
darf nicht geschlossen werden“ (MAHNER 1986, S. 79).

- 33 -
F. Kritik an der negativen Komponente des Design-Arguments
Wie eingangs erwähnt, umfasst die zweite Komponente des Design-Arguments die
Kritik an der Evolutionstheorie. Ihre skeptische Haltung gegenüber der natürlichen
Entstehung nicht reduzierbar komplexer Strukturen begründen die Autoren im Wesent-
lichen mit vorgeblicher Unwahrscheinlichkeit:
„Damit ein Gegenstand eine Funktion ausüben kann, benötigt er in der Regel mehrere Bau-
elemente, die in spezifischer Weise jeweils konstruiert und zusammen aufeinander abge-
stimmt sein müssen. Dies betrifft oft ganz verschiedene Aspekte wie Materialeigenschaften,
Form, Steuerung u. a. Diese Bestandteile und ihre gegenseitigen Abstimmungen erfordern
sehr häufig eine hohe Komplexität, die nicht mehr verkleinert werden kann, ohne die in Rede
stehende Funktion vollständig zu verlieren … Diese Konstruktionen sind an sich klare De-
sign-Kennzeichen … Gleichzeitig sind die bekannten natürlichen Mechanismen nach aller
unserer Kenntnis bei weitem überfordert, solche Konstrukte hervorzubringen.
Das evolutionstheoretische Grundproblem besteht darin, dass kein kontinuierlicher, klein-
schrittiger und hinreichend wahrscheinlicher Weg von einem Zustand ohne diese spezielle
Funktion (und ohne die speziell dafür nötige Konstruktion) hin zu einem Zustand mit dieser
Funktion (und der dafür nötigen Konstruktion) gedacht [sic!] werden kann. Jeder einzelne
Mutationsschritt müsste eine hinreichende statistische Wahrscheinlichkeit haben. Er dürfte
nicht selektionsnegativ sein, vielmehr müsste die entsprechende Mutante in der Population
konserviert und signifikant verbreitet werden. Dabei müsste er sich dem in Rede stehenden
Zustand annähern … Selbstverständlich kann hier jedoch nicht (schrittweise) auf ein Ziel hin
geplant und ‚gearbeitet‘ werden, da natürliche Mechanismen zukunftsblind sind: Da Selekti-
on immer nur die aktuellen, nicht aber potentielle zukünftige Funktionen ‚bewerten‘ kann, be-
sitzen hypothetische Vorstufen, die die Funktion des betreffenden Gebildes nicht besitzen,
im Hinblick auf diese spezifische Funktion keinen Selektionsvorteil“ (S. 5, S. 7).
Ist diese Argumentation überzeugend?
F.1 Nicht reduzierbare Komplexität als Einwand gegen Evolution
Zunächst haben WIDENMEYER & JUNKER (S. 7) Recht damit, dass die Selektion nicht
„potentielle zukünftige Funktionen ‚bewerten‘“ kann. Wie sollte sie auch? Was nicht
existiert, kann nicht „belohnt“ werden, und eine nicht vorhandene Funktion lässt sich
nicht verstärken. Die Evolution ist „zukunftsblind“ in dem Sinne, dass wir sagen: Der
Funktionszustand (A) eines Systems kann nicht vom Funktionszustand (B) eines evo-
lutiven Vorläufers aus direkt „anvisiert“ werden.

- 34 -
Die Grundzüge dieses Arguments stammen von dem Biochemiker Michael BEHE
(1996). Dieser beschreibt detailliert verschiedene biochemische Signalkaskaden und
„molekulare Maschinen“, die gemeinsam eine physiologische Funktion oder Reaktion
herstellen. Dann stellt er fest, dass die Wegnahme einer beliebigen Komponente zum
völligen Zusammenbruch der Funktionen führe: Die betreffenden Strukturen seien
nicht reduzierbar komplex. Daraus folgert BEHE, es sei unmöglich, sie in hinreichend
kleinen, von der Selektion belohnten Schritten aufzubauen. Denn wie sollen fortlaufend
komplexere Biosysteme evolviert sein, wenn sich das Komplexe an keiner Stelle ver-
einfachen lässt, ohne dass die Funktion wegbricht?
In diesem Zusammenhang sprechen ID-Vertreter vom „direkten DARWIN‘schen Weg“:
„Ein direkter darwinistischer Weg ist einer, wonach sich ein System durch natürliche Selek-
tion entwickelt, die eine gegebene Funktion schrittweise verstärkt. Während sich das Sys-
tem weiterentwickelt, ändert sich die Funktion nicht“ (DEMBSKI 2003; ins Deutsche M.N.).
So könnten sich die Strukturen des Auges im Prinzip stufenlos verändern. Die Selekti-
on „beurteilt“ nach jedem Variationsschritt, ob sich die Funktion verbessert oder ver-
schlechtert. Dieser Weg einer schrittweisen Optimierung bleibt der Evolution bei der
Konstruktion nicht reduzierbar komplexer Strukturen versperrt.
Michael BEHE ist sich im Klaren darüber, dass der Evolution indirekte Wege offenste-
hen, um nicht reduzierbar Komplexes hervorzubringen. Eine Möglichkeit ist die soge-
nannte Kooption (oder Kooptation). Darunter verstehen wir das Umgestalten von be-
reits existierenden (präadaptierten) Entwicklungswegen, die Inanspruchnahme von
vorhandenen Genen oder Bauelementen für eine neue Aufgabe. Dieses evolutive
Schema ist unter dem Stichwort Funktionswechsel geläufig. ID-Vertreter räumen ein,
dass die Evolution solche „indirekten“ Wege beschreiten kann, was leider viele Kritiker
(etwa BRIGANDT 2013, S. 233) übersehen. Sie bestreiten allerdings, dass dies bei sehr
komplexen Systemen erfolgversprechend sei (KOJONEN 2016, S. 65f).
MENUGE (2004, S. 104–105) begründet dies damit, dass die Transformation eines Vor-
läufersystems in eine neue, nicht reduzierbar komplexe Struktur an eine Reihe von Vo-
raussetzungen geknüpft sei. Zunächst müssten die Teile, aus denen die nicht reduzier-
bar komplexe „Maschine“ wird, in der Zelle vorhanden und für evolutionäre Variations-
Experimente zur Verfügung stehen. Sie dürfen bestehende Vitalfunktionen nicht beein-
trächtigen. Des Weiteren müsse sie die Evolution an den richtigen Ort transferieren und
passend (in der richtigen Reihenfolge) miteinander „verschalten“. Schließlich müsste sie

- 35 -
alle Teile in ihrer Struktur derart verändern und aufeinander „abstimmen“, dass diese die
neue Funktion herstellen. Kurz: Schon die präzise Lokalisation, Synchronisation und
Koordination einiger weniger Bauelemente soll vielfache Abstimmungen gleichzeitig er-
fordern. Deshalb werde die Entstehung komplizierterer molekularer „Maschinen“ und
biochemischer Regelkreise rasch unwahrscheinlich:
“Die darwinistische Evolution kann für einige nicht reduzierbar komplexen Strukturen ver-
antwortlich sein, aber je höher das Komplexitätsniveau des nicht reduzierbar komplexen
Systems ist, desto weniger plausibel ist es, dass sich dieses System durch darwinistische
Mechanismen entwickelt hat“ (Ins Deutsche M.N.).13
Was lässt sich auf sachlicher Ebene auf diese Argumentation entgegnen? Wir könnten
es uns einfach machen und auf die in Abschnitt C.2 beschriebene Evolution des
T-urf13-Proteins verweisen. Sie belegt, dass nicht reduzierbar komplexe Strukturen
mitunter in größeren Sprüngen entstehen, und das in überraschend kurzer Zeit.
Einen theoretischen Ansatz verfolgt der amerikanische Philosoph Paul DRAPER (2002),
dessen Arbeit zu den derzeit besten auf diesem Gebiet zählt. Seine Analyse und Wider-
legung umfassen drei Ebenen:
Er kritisiert die Behauptung, bestimmte Systeme seien sehr komplex.
Er entkräftet die Aussage, es sei unwahrscheinlich, dass sehr komplexe Sys-
teme auf indirektem Weg evolvieren könnten.
Er verwirft die These, irreduzibel komplexe Strukturen könnten nicht auf direk-
tem DARWIN‘schen Weg evolvieren.
Das Hauptproblem der ID-Argumentation: Die Komplexität heutiger Proteinkomplexe und
Regel-Kaskaden sagt nichts darüber, wie spezifisch das Arrangement der Teile in einem
wenig spezialisierten „Minimal-System“ zu sein brauchte. Auch sagt sie nichts darüber, in
wie vielen Schritten die Evolution sie hervorbringen konnte und wie „groß“ die Schritte sein
mussten. Letzteres hängt davon ab, welche Komponenten des Systems zuvor „richtig“
verortet und in passender Ausprägung vorhanden waren. Und sie hängt davon ab, wie
„weit“ die Funktionszustände des Systems und seines Vorläufers auseinander liegen.
Nehmen wir an, die Evolution optimiere die Funktion (A) eines Systems. Und sie füge
Teile zum bestehenden System hinzu. Dann kann es geschehen, dass der Komplex
unvermittelt eine zusätzliche Funktion (B) erfüllt. Wird später die Funktion (A) nicht
13 thegenomestale.wordpress.com/2011/09/14/revisiting-irreducible-complexity/

- 36 -
mehr gebraucht, kann das „Gerüst“, das die Entstehung der Funktion (B) ermöglichte,
verloren gehen. Die Eliminierung von Redundanz bewirkt, dass nur der nicht redu-
zierbar komplexe Zustand (B) übrigbleibt (THORNHILL & USSERY 2000; DRAPER 2002).
BURDA & BEGALL (2013, S. 288) erläutern das Prinzip an einem einfachen Beispiel: Die
in Nordamerika beheimatete Bolaspinne (Abb. 9) erzeugt einen Faden, an dem ein
Tropfen Klebstoff hängt. Gleichzeitig scheidet sie einen Stoff aus, der bestimmte Falter
anlockt. Die männlichen Falter fliegen zur Spinne und bleiben am Klebfaden hängen.
Das Arrangement ist nicht reduzierbar komplex, denn ohne Lockstoff ist der Faden
nutzlos und ohne Faden der Lockstoff. Dennoch können wir uns leicht vorstellen, wie
sich das System bildete, ohne beide Komponenten in einem Schritt hervorzubringen.
Abb. 9 Bolaspinnen produzieren ein Pheromon, das männliche Eulenfalter anlockt. Zudem er-
zeugen sie einen Klebfaden, an dem die Falter hängenbleiben. © Bernard DUPONT, CC BY-SA
2.0, www.flickr.com/photos/65695019@N07/9286584444 (Bild beschnitten und gedreht.)
Ein Vorfahr der Spinne produzierte Spinnennetze, die auch ohne Lockstoff funktionier-
ten. Später lockte die Sekretion eines Stoffs, der zufällig Zugang zum Kommunikati-
onssystem männlicher Eulenfalter fand, Beute an. In dem Augenblick, als der Lockstoff
ausreichte, um Beute anzulocken, war das „Gerüst“ des Spinnennetzes entbehrlich.
Die Evolution reduzierte es, bis nur noch ein Klebstoff-Faden übrigblieb. Im selben
Maß, wie das Netz schwand, gewann der Lockstoff an Bedeutung. Was ursprünglich
nur eine zufällige „verspielte“ Luxus-Ausstattung darstellte, erweist sich heute als un-
entbehrlich. Eine nicht reduzierbar komplexe Struktur ist entstanden.

- 37 -
Natürlich stößt dieses Beispiel an Grenzen. Wie erörtert, stellen ID-Vertreter die Mög-
lichkeit der „indirekten“ Evolution einfacher Systeme selten infrage. Zudem wendet
BEHE sein Argument auf molekulare Systeme an, wo es sich schärfer fassen lässt.
Doch das Beispiel eignet sich gut, um das Prinzip der Redundanz-Eliminierung zu il-
lustrieren: Sobald die „Gerüststrukturen“ verschwinden, sehen wir dem System nicht
mehr an, in welchen Schritten es entstand. Ebenso wenig sehen wir einer Kathedrale
das Holzgerüst an, das nötig war, um einen Bogen zu stützen. Und doch wissen wir,
dass es ein Fehlschluss wäre zu behaupten, die Konstrukteure hätten alle Steine in
einem Schritt verkeilen müssen, damit sie in einem Bogen zusammenhalten.
Die ID-Vertreter mögen einwenden, Steinbögen und Holzgerüste seien Beispiele für
„Design“, nicht für Evolution. Doch das gleiche Prinzip lässt sich auf die natürliche Evo-
lution übertragen – und sogar auf die Entstehung hochkomplexer molekularer Signal-
kaskaden (ROBINSON 1996; MILLER 2007, S. 154ff; NEUKAMM 2009, S. 219–224). An-
hand des Blutgerinnungssystems lässt sich zeigen, wie sich durch aufeinander folgen-
de Gen-Duplikationen mit Redundanz-Eliminierung und Funktionswandel einstufige
Prozesse in mehrstufige Kaskaden umwandeln lassen (Abb. 10).
Abb. 10 Bei der Blutgerinnung ist der wesentliche Schritt die Umwandlung des Glykoproteins
Fibrinogens in den „Wundklebstoff“ Fibrin. Dieser Prozess wird durch eine Protease (A) einge-
leitet, die an den Gewebsfaktor (TF) bindet und durch diesen aktiviert wird. Die aktivierte Pro-
tease (A*) überführt das Fibrinogen zum Fibrin und aktiviert ihrerseits viele weitere A-
Moleküle. Durch Gen-Duplikation entsteht eine Kopie der Protease A (links als B bezeichnet),
die ebenfalls an den Gewebefaktor TF bindet und die Effizienz der Fibrinerzeugung steigert.
Infolge einer Mutation kann sich die Spezifität der B-Protease ändern, sodass sie stärker an
den Gewebefaktor bindet und A rascher aktiviert als A*. Die Effizienz der Blutgerinnung nimmt
zu. Eine einfache Zweischrittkaskade ist entstanden (rechts). Durch weitere solche Schritte
lässt sich die Komplexität der Kaskade beliebig steigern. Aus NEUKAMM (2009, S. 222).

- 38 -
Generell können sich komplexe Stoffwechselwege schrittweise „von ihrem Ende
her“ (retrograd) herausbilden (LIGHT & KRAULIS 2004; FANI 2012). Daher sagt ihr
Maß an nicht reduzierbarer Komplexität nichts über die Plausibilität ihrer Evolu-
tion aus. Letztere hängt nur von der Plausibilität des unwahrscheinlichsten Einzel-
schritts ab. Wie die hier zitierten Autoren darlegen, gibt es auch andere Evolutionswe-
ge; bei den meisten spielen Gen-Duplikationen eine tragende Rolle. Und: Jedes der
Modelle wird durch Daten untermauert, die ihren Vorhersagen entsprechen.
Noch ein Beispiel aus der Mikrobiologie: Viele Bakterien und Archaeen verfügen über
Fortbewegungs-Organellen (sogenannte Flagellen, „Rotations-Motoren“). Damit sie
funktionieren, müssen Dutzende Proteine miteinander interagieren. Müssten wir vo-
raussetzen, dass sie die Evolution gleichzeitig aufeinander abstimmte, wären vermut-
lich nie Flagellen evolviert. Nun wissen wir, dass Bakterien und Archaeen sogenannte
Haft-Organellen (Pili) besitzen. Damit heften sie sich an Oberflächen oder binden be-
stimmte Moleküle. Für ihren Zusammenbau sorgt ein Membranprotein (Mem) sowie
ein membranassoziiertes Enzym, die sogenannte ATPase (Abb. 11).
Die ATPase liefert durch Spaltung des Energieträgers ATP die Energie für den Trans-
port der Protein-Untereinheiten aus der Zelle. Dabei beginnt sie strukturbedingt zu rotie-
ren. Das bei der Rotation erzeugte Drehmoment ist eine zufällige Eigenschaft der
ATPase, die für die Pili keine Rolle spielt. Doch die ATPase kann kein ATP spalten, oh-
ne zu rotieren: Die Spaltung von ATP sowie die Rotation sind gekoppelte Eigenschaf-
ten der ATPase, die eine Evolution in Richtung eines Rotations-Motors begünstigen
kann. Benötigt der Organismus die Haft-Funktion der Pili nicht mehr, kann er sie zur
Kraftübertragung des Drehmoments der ATPase auf das umgebende Medium nutzen.
Merke: Der Grad an nicht reduzierbarer Komplexität und Spezifität lässt keine
Rückschlüsse auf seinen Ursprung und seine Entstehungswahrscheinlichkeit zu.
So ist dieser nicht zu entnehmen, wie viele Schritte für ihre Bildung notwendig wa-
ren und welche Voraussetzungen sie begünstigten. Auch einer Kathedrale sehen
wir das einstige Holzgerüst nicht an, das einen Bogen stützte.
Ist das Gerüst verschwunden, könnten wir vermuten, die Konstrukteure hätten alle
Steine gleichzeitig verkeilen müssen, damit sie in einem Bogen zusammenhalten.
Das ist ein Irrtum. „Gerüststrukturen“, die eine allmähliche Entstehung des nicht
reduzierbar Komplexen erleichtern könnten, finden wir auch bei Lebewesen.

- 39 -
Wir vermuten, dass diese Modifikation Abstimmungen erfordert. Doch ein Großteil der
Flagellen-Komponenten sitzt bei verschiedenen Pili bereits „an der richtigen Stelle“. Da
sie auch für das Funktionieren von Flagellen erforderlichen Voraussetzungen erfüllen,
vereinfacht sich das evolutionäre Problem wesentlich. Die „Schrittweite“ ist weit kleiner
als verschiedene Design-Vertreter behaupten (↑ Abschnitt F.3).
Abb. 11 Modell der Zellmembran von Sulfolobus solfataricus. Die Vorläufer der Flagellen-,
Pili- und zuckerbindenden Proteine werden in die Membran integriert, durch Peptidase pro-
zessiert und in die verschiedenen Zelloberflächen-Strukturen eingegliedert. Die Assemblie-
rungs-Systeme bestehen in allen drei Fällen aus zwei Komponenten: einer ATPase und ei-
nem integralen Membranprotein. © Max-Planck-Institut für Terrestrische Mikrobiolo-
gie/ALBERS. Abdruck mit freundlicher Genehmigung von S.-V. ALBERS.
F.2 Reduzierbare Spezifität contra nicht reduzierbare Komplexität
Erörtern wir nun ein Beispiel, das in seiner Komplexität (Abb.13) atemberaubend ist.
Die Forschungs-Resultate der letzten Jahre führten dazu, dass sich die Einwände der
Evolutionsgegner nach und nach verflüchtigten. Wir sehen darin ein Paradebeispiel für
das Scheitern des Arguments der nicht reduzierbarer Komplexität. Worum geht es?
Einige Organellen moderner tierischer und pflanzlicher Zellen (Eucyten) entstanden
durch eine Reihe von Verschmelzungen verschiedener ursprünglicher Zellen. Die Biolo-
gie spricht von serieller Endosymbiose. Diesen Sachverhalt beschreibt die Endosym-
bionten-Theorie. Sie lehrt, dass eine bakterienähnliche Vorläuferzelle ein Sauerstoff

- 40 -
nutzendes Bakterium (Proteobakterium) aufnahm. Ein daraus hervorgehender Eukaryot
verleibte sich später ein Photosynthese betreibendes Cyanobakterium ein. Im Laufe von
Jahrmillionen büßten die aufgenommenen Bakterien (Symbionten) ihre Autonomie ein,
indem sie einen Großteil ihrer Gene in den Zellkern der Wirtszelle auslagerten. So wur-
de das Proteobakterium zu den Mitochondrien („Kraftwerken“) der modernen Zellen und
das Cyanobakterium zu den Chloroplasten der Pflanzen (Abb. 12).
Abb. 12 Evolution der Eucyte durch serielle Endosymbiose. Nach jedem Schritt lagerte sich ein
großer Teil der Symbionten-Gene in den Kern der Wirtszelle aus (Gentransfer). Auf dieser Wei-
se wurden sie zu Organellen, beispielsweise zu den Chloroplasten von Pflanzen, in denen die
Photosynthese stattfindet. (N: Nukleus = Zellkern). Mit freundlicher Genehmigung von R. BOCK.
Die Endosymbionten-Theorie ist etablierter Bestandteil der Evolutionstheorie und gut
belegt. Doch WORT UND WISSEN (2008) hält die „Co-Evolution“ von Endosymbiont und
Wirt für zu komplex und unwahrscheinlich. Ein funktionaler Gentransfer, so das Argu-
ment, erfordere gleichzeitig mehrere aufeinander abgestimmte Koordinationsschritte:
Die Symbionten-Gene müssen sich passend ins Kerngenom der Wirtszel-
le integrieren, damit sie „lesbar“ sind.
Die Produkte (Proteine) der in den Kern transferierten Gene haben über
die Membranen in die jeweiligen Organellen zu gelangen. Diesen „Re-
Import“ gewährleisten spezielle Transit-Peptide. Die Evolution hängte sie
den Proteinen an, indem sie den für sie kodierenden Genen passende
Zielsequenzen zuwies.
Ferner braucht es spezielle „Porenkomplexe“ mit einer Reihe passender
Protein-Untereinheiten, die es den Proteinen erlauben, sich in die jeweili-
gen Kompartimente „einzufädeln“. Diese sogenannten Translokasen müs-
sen am „richtigen Ort“ sein, damit die Verteilung der Proteine auf die Zell-
Kompartimente korrekt geschieht.
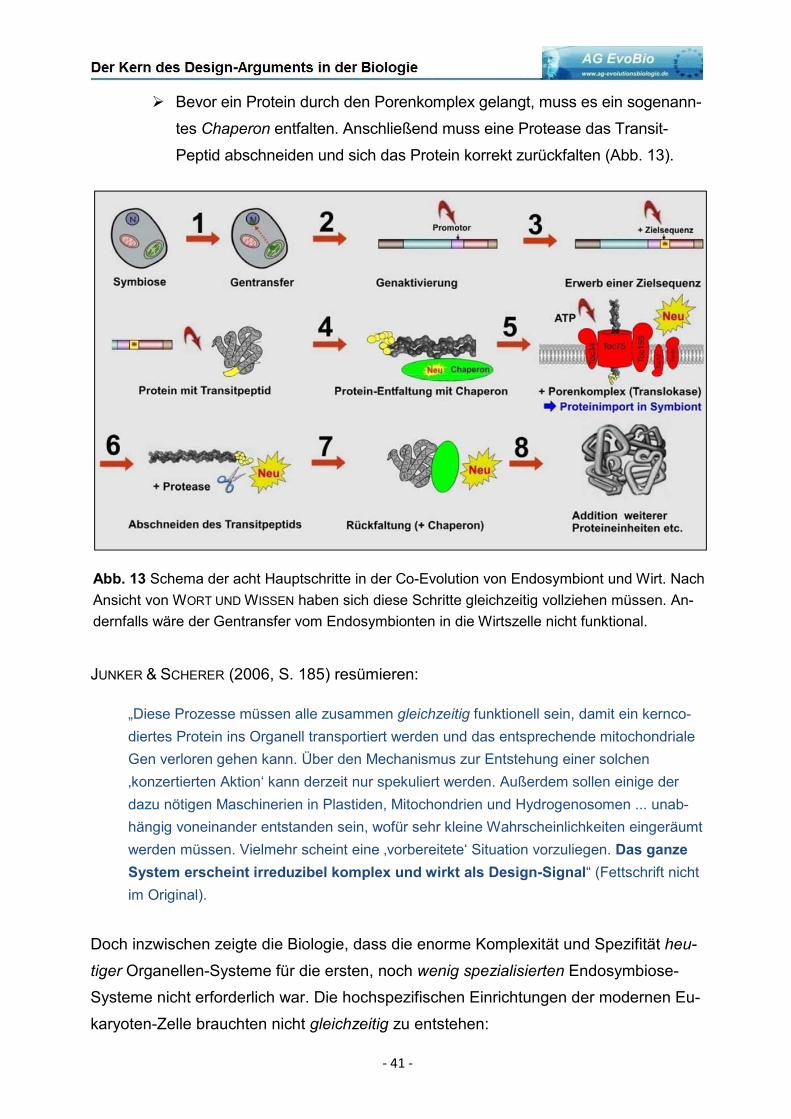
- 41 -
Bevor ein Protein durch den Porenkomplex gelangt, muss es ein sogenann-
tes Chaperon entfalten. Anschließend muss eine Protease das Transit-
Peptid abschneiden und sich das Protein korrekt zurückfalten (Abb. 13).
Abb. 13 Schema der acht Hauptschritte in der Co-Evolution von Endosymbiont und Wirt. Nach
Ansicht von WORT UND WISSEN haben sich diese Schritte gleichzeitig vollziehen müssen. An-
dernfalls wäre der Gentransfer vom Endosymbionten in die Wirtszelle nicht funktional.
JUNKER & SCHERER (2006, S. 185) resümieren:
„Diese Prozesse müssen alle zusammen gleichzeitig funktionell sein, damit ein kernco-
diertes Protein ins Organell transportiert werden und das entsprechende mitochondriale
Gen verloren gehen kann. Über den Mechanismus zur Entstehung einer solchen
‚konzertierten Aktion‘ kann derzeit nur spekuliert werden. Außerdem sollen einige der
dazu nötigen Maschinerien in Plastiden, Mitochondrien und Hydrogenosomen ... unab-
hängig voneinander entstanden sein, wofür sehr kleine Wahrscheinlichkeiten eingeräumt
werden müssen. Vielmehr scheint eine ‚vorbereitete‘ Situation vorzuliegen. Das ganze
System erscheint irreduzibel komplex und wirkt als Design-Signal“ (Fettschrift nicht
im Original).
Doch inzwischen zeigte die Biologie, dass die enorme Komplexität und Spezifität heu-
tiger Organellen-Systeme für die ersten, noch wenig spezialisierten Endosymbiose-
Systeme nicht erforderlich war. Die hochspezifischen Einrichtungen der modernen Eu-
karyoten-Zelle brauchten nicht gleichzeitig zu entstehen:

- 42 -
1. Der Gentransfer in den Kern der Wirtszelle kann unspezifisch erfolgen, solange
er Duplikate oder Gene betrifft, die aus dem unregulierten Zerfall gealterter
Symbionten stammen (NEUKAMM & BEYER 2011, S.17). Dies belegen beispiels-
weise Hülsenfrüchtler, bei denen verschiedene Zwischenstufen eines funktiona-
len Gentransfers existieren: Wir kennen für das Protein COX2 Gene, die nur in
den Mitochondrien auftreten. Es gibt Gene in Mitochondrien und Zellkern, die
nur in den Mitochondrien aktiv sind. Ferner existieren Gene, die sowohl in den
Mitochondrien als auch im Zellkern abgelesen werden, und Mitochondrien-
Gene, die nur im Kern aktiv sind (COVELLO & GRAY 1992; DALEY et al. 2002).
2. Experimente mit Pflanzen, in deren Chloroplasten Forscher bestimmte Resis-
tenz-Gene übertrugen, zeigen, dass die Gene überraschend häufig in den Zell-
kern „springen“. Ein Teil der Pflanzen schafft es, die nichtfunktional in den Zell-
kern transferierten Gene durch zufälliges Hinzufügen oder Entfernen eines
kleinen Stücks DNA zu aktivieren (STEGEMANN & BOCK 2006).
3. Zielsequenzen waren für die in den Kern der Wirtszelle ausgelagerten Symbionten-
Gene zunächst entbehrlich. Beispiele belegen, dass ein Teil der Gen-Produkte
nicht in die „vorgesehenen“ Zell-Kompartimente gelangt. Er verbleibt im Plasma
oder gelangt in andere Kompartimente (MARTIN et al. 2002; NOWITZKI 2002, S. 3).
4. Die später erworbenen Zielsequenzen, die das betreffende Gen markieren,
brauchten nicht besonders spezifisch zu sein: Sage und schreibe 20% aller co-
dierenden Gen-Bereiche und ein hoher Prozentsatz von Zufallssequenzen,
eignen sich grundsätzlich als Zielsequenz (TONKIN et al. 2008):
„Die geringe Komplexität [Sequenz-Spezifität; M.N.] des Transit-Peptids hat daher
wohl den Erwerb solcher Sequenzen über Exon-Shuffling oder einfach über den
Gebrauch von Zufallssequenzen erheblich erleichtert und damit den intrazellulä-
ren Gentransfer [von Organellen in den Kern] beschleunigt“ (TONKIN et al. 2008,
S. 4785; ins Deutsche M. N.)
5. Die Evolution profitiert davon, dass es „das richtige“ Kompartiment für den Pro-
tein-Import nicht gibt. Proteine können in jedes Organell importiert werden. Re-
sultiert daraus ein Vorteil, wird diese evolutive Veränderung selektiert und fixiert.
6. Ein einfaches Sortiersystem ist nicht auf die „enorme Komplexität und Spezifität“
(WORT UND WISSEN 2008) heutiger Porenkomplexe angewiesen. Zwar sind die
Protein-Untereinheiten heute nur in ihrer speziellen physiologischen Vernetzung
funktional. Wir wissen jedoch, dass ein Großteil davon ursprünglich unnötig war.

- 43 -
Ein Beispiel ist der Tic/Toc-Komplex in der Außenmembran von Chloroplasten. Des-
sen Kernstück ist das Porenprotein Toc75. Es steuert gemeinsam mit den Rezeptor-
Proteinen Toc34, Toc159 und Toc64 sowie mit einigen Regulator-Proteinen den Import
bestimmter, von den Chloroplasten benötigter Proteine (Abb. 14). Beim Cyanobakteri-
um Synechocystis findet sich das sehr ähnlich gebaute Protein SynToc75, allerdings
ohne die Untereinheiten des Tic-/Toc-Komplexes (BÖLTER et al. 1998).
Abb. 14 Toc- und Tic-Proteine. Nach Soll & Schleiff (2004, S. 204).
Dieses Protein sitzt beim Cyanobakterium (dem einstigen Proto-Endosymbionten) be-
reits an der „richtigen“ Stelle. Es ist in der Lage, bestimmte Proteine passieren zu las-
sen, wenngleich noch nicht mit der Spezifität heutiger Chloroplasten. Dies macht Syn-
Toc75 zum idealen Ausgangspunkt in der Evolution des Protein-Transporters. Die
Evolution konnte mit diesem noch weitgehend unspezialisierten Transportkanal begin-
nen und dessen Effizienz und Spezifität durch sukzessives Addieren von Rezeptoren
und Regulationsproteinen steigern (REUMANN et al. 2005; INABA & SCHNELL 2008; BO-
DYŁ et al. 2009, S. 1221f). Selbst bei hochentwickelten Pflanzen bewirkt der Knock-out
einiger Protein-Untereinheiten zwar eine Effizienz-Minderung des Proteintransports,
nicht aber dessen Zusammenbruch (KOVACHEVA et al. 2005).
Wieder ist der entscheidende Punkt, dass aus „nicht reduzierbarer Komplexität“
nicht folgt, dass alle Teile des Systems von Anfang an hochspezifisch miteinan-
der wechselwirken müssen. Um es mit DRAPER (2002) zu formulieren:

- 44 -
„Was Behe braucht, ist nicht nur Spezifität, sondern das, was ich als ‚nicht reduzierbare
Spezifität‘ bezeichnen möchte. Die Behauptung, dass die Spezifizität eines Systems nicht
reduzierbar sei, entspricht etwa der Behauptung, dass das System aufhört wirksam zu ar-
beiten, wenn seine Teile modifiziert werden. Diese Definition ist jedoch nur eine erste Nä-
herung, weil sie nicht reduzierbare Spezifität als qualitatives Konzept behandelt, während
sie in Wirklichkeit quantitativ ist – sie entsteht abgestuft, gradweise. Kein System funktio-
niert so, dass eine Veränderung seiner Teile, egal wie geringfügig sie auch sein mag, zu
einem totalen Funktionsverlust führen würde.“14
Nehmen wir an, die Wahrscheinlichkeit der Evolution nicht reduzierbar komplexer
Strukturen durch Funktionswechsel und Kooption wäre umso geringer, je komplizierter
sie sind. Dann dürfte sich ihr hoher Grad an Spezifität nicht verringern, ohne zum
Funktionsausfall zu führen. Das Endosymbiose-System zeigt exemplarisch, dass das
nicht der Fall ist.
F.3 Schätzungen zur Unwahrscheinlichkeit von Evolution
Wie gesehen bezeichnen JUNKER & SCHERER (2006, S. 185) die Evolution der eukaryo-
tischen Zelle als sehr unwahrscheinliche, „konzertierte Aktion“. Das System erscheine
„irreduzibel komplex“ und wirke „als Design-Signal“. Doch in der aktuellen Auflage
räumen sie ein, dass die Datenlage „eine schrittweise Entwicklung zu Endosymbionten
zunehmend plausibel“ mache (JUNKER & SCHERER 2013, S. 199). Von „Design-
Signalen“ und „konzertierten Aktionen“ ist nicht mehr die Rede!
14 Im Original: “So what Behe needs to rule out the sort of indirect route being considered is a system
that cannot function at all unless its parts are well matched. He needs, in other words, not just specifici-
ty, but what I will call ‘irreducible specificity.’ To claim that the specificity of a system is irreducible is,
roughly, to claim that the system would effectively cease to function if its parts were modified. This defi-
nition is, however, only a first approximation, because it treats irreducible specificity as a qualitative con-
cept, whereas in reality it is quantitative–it comes in degrees. No system is such that any modification of
its parts, no matter how slight, would result in a total loss of function.”
Merke: Nicht reduzierbare Komplexität wäre ein Argument gegen eine natürliche
Evolution, wenn gezeigt wäre, dass die betreffenden Systeme zugleich nicht redu-
zierbar spezifisch sind. Das Gegenteil ist der Fall. Die Annahme, die Evolution
einer nicht reduzierbar komplexen Struktur sei umso unwahrscheinlicher, je kom-
plizierter sie ist, ist unbegründet.

- 45 -
Kurioserweise hält dies die Autoren nicht davon ab, ihr gescheitertes Argument immer
wieder neu aufzulegen. Ihre Beispiele wechseln, die Struktur ihres Arguments bleibt
gleich. Sie behaupten, ein hinreichend wahrscheinlicher Weg zu einem nicht reduzier-
bar komplexen Gegenstand könne nicht einmal gedacht werden. Das mag in einzelnen
Fällen zutreffen – aber nur so lange, bis die Forschung das Problem durchdacht hat.
Wie anders lässt sich das Rückzugsgefecht bei JUNKER & SCHERER (2013, S. 199) er-
klären? Jedenfalls scheinen andere keine Schwierigkeiten damit zu haben, entspre-
chende Wege zu denken (ORR 1996; DRAPER 2002; NEUKAMM & BEYER 2011).
Auch das Kapitel über die Evolution bakterieller Fortbewegungs-Organellen (soge-
nannte Flagellen) haben die Autoren komplett revidiert. Seit BEHE (1996) zählt es zu
den Paradebeispielen nicht reduzierbar komplexer Strukturen in der ID-Literatur. Wäh-
rend JUNKER & SCHERER (2006) die Evolution der Flagellen transastronomisch un-
wahrscheinlich rechnen, findet sich in der aktuellen Auflage hierzu keine Kalkulation.
Statt der ehemals postulierten 160 sollen jeweils noch etwa zehn Mutationen zwischen
zwei positiv selektierbaren „Basis-Funktionszuständen“ liegen (SCHERER 2010, S. 20).
Dies würde eine drastische Erhöhung der Entstehungs-Wahrscheinlichkeit bedeuten.
Welche Werte bei solchen „Berechnungen“ herauskommen, hängt also enorm von den
Prämissen ab, die die Evolutionsgegner ihnen zugrunde legen. So gut wie keine Prä-
misse, die ID-Vertreter ihren Kalkulationen voranstellen, wird in der Fachwelt aner-
kannt. Damit ist klar, dass die Wahrscheinlichkeitswerte der ID-Vertreter höchst sub-
jektiv sind. Und je weniger wir über bestimmte Evolutionswege wissen, desto weniger
Sinn hat es, über Wahrscheinlichkeiten zu spekulieren. Jede neue Erkenntnis kann die
Einschätzungen zu Fall bringen – und so ist es tatsächlich in den meisten Fällen: Kei-
ne einzige Wahrscheinlichkeit, die JUNKER & SCHERER (1998–2006) in ihrem Schulbuch
präsentierten, taucht in der jeweils nächsten Auflage auf.
Ein anderes Beispiel: DENTON (2016, S. 226) behauptet, bereits das Rekrutieren eines
einzelnen Gens für eine neue Gen-Kaskade sei ein komplizierter Schritt. Das bloße An-
schalten des Gens erfordere enorm komplizierte Regulationsmechanismen, die gewähr-
leisten, dass das Gen am rechten Platz, zur rechten Zeit und in der richtigen Menge ex-
primiert werde. Andernfalls führe dies ins Chaos. Dergleichen ist aber nicht zwingend. In
vielen Fällen reicht es, wenn ein Gen überhaupt aktiv ist, und sei es in allen möglichen
Zellen. Die Optimierung der Genregulation kann oft später erfolgen.

- 46 -
Insbesondere den Einsichten der Systembiologie ist es zu verdanken, dass derartige
Spekulationen Zug um Zug an Plausibilität einbüßten. Zum Beispiel brauchen sich die
Komponenten eines Merkmalskomplexes („Moduls“) nicht unabhängig voneinander zu
verändern. Vielmehr gilt oft das Umgekehrte: Werden genetische „Schalter“ modifiziert,
verändert sich das Zusammenspiel aller untergeordneten Gene gleichzeitig. Infolge-
dessen evolviert mehr oder minder das ganze Modul. Und durch Übernahme eines
„Schalters“ in andere Gennetzwerke werden ganze Genkaskaden auf einen Schlag in
unterschiedlichen „Modulen“ aktiv (MONTEIRO & PODLAHA 2009).
Freilich ist es legitim zu fragen, ob die betreffende Evolution im konkreten Einzelfall
plausibel ist. Von wenigen Beispielen abgesehen, ist es unmöglich, dies experi-
mentell zu klären: Einerseits wissen wir über die Voraussetzungen der Realhistorie
zu wenig. Andererseits haben wir nicht genügend Zeit, eine Millionen Jahre lange Ent-
wicklungsreihe im Labor zu analysieren. Doch es gibt neue theoretische Einsichten.
Monte-Carlo-Simulationen zeigen, dass selbst dann gute Chancen bestehen, nicht
reduzierbar komplexe Strukturen hervorzubringen, wenn dies drei, vier oder noch mehr
„passende“ Mutationen gleichzeitig erfordern würde (TROTTER et al. 2014). Eine sol-
che Evolution ist insbesondere bei Genen erfolgversprechend, die unter keinem hohen
Selektionsdruck stehen. Hier können sich über lange Zeit selektionsneutrale Mutatio-
nen ansammeln (Abb. 15). Da der Anteil kryptischer Gene im Genom recht groß ist, ist
die Zahl komplexer Anpassungen unter bestimmten Bedingungen erheblich (Abb. 16).
Das mitochondriale Gen T-urf13 (↑ Abschnitt C.2) in bestimmten Mais-Sorten mag als
Beispiel genügen. Es besteht aus einer intergenischen Region, die nicht transkribiert
wird, und aus einem Fragment, das nicht in ein Protein übersetzt wird. Die Evolution
konnte mit diesen DNA-Stücken weitgehend selektionsneutral experimentieren.
Merke: Wahrscheinlichkeits-Abschätzungen der Evolutionsgegner, oft mit einer
mathematischen Scheinpräzision ausgestattet, täuschen Wissen über die Mög-
lichkeiten und Grenzen der Evolution vor, das wir nicht haben. Ein solches Wissen
existiert nicht ansatzweise. Andernfalls wüssten wir, welche Systeme sich unter
welchen Voraussetzungen in welche Richtungen entwickeln können und wie viele
Schritte notwendig sind. Wüssten wir dies alles, müssten die ID-Vertreter zeigen,
dass all die Voraussetzungen zu keiner Zeit und bei keiner Art realisierbar waren.
Sie bürden sich eine Beweislast auf, die sie nicht stemmen.
- 47 -
Abb. 15 Nicht reduzierbare Komplexität lässt sich als Tal zwischen zwei Erhebungen in der
Fitnesslandschaft interpretieren (links unten). Jede Erhebung repräsentiert einen Funktionszu-
stand, jeder Anpassungsgipfel ein lokales Fitness-Optimum. Da die Selektion Schritte mit ab-
nehmender Fitness unterbindet, endet die Optimierung auf einem der Gipfel. Doch Zufallsdrift
ermöglicht das Überqueren der Täler und so das Erreichen eines neuen Funktionszustandes.
Je „breiter“ das Tal zwischen zwei Funktionszuständen ist, desto schwerer lässt es sich über-
winden. Monte-Carlo-Simulationen zeigen, dass dies umso eher geschieht, je geringer der Ein-
fluss der Selektion ist. Besonders eignen sich kryptische Gene, die nur unter bestimmten Be-
dingungen abgelesen werden. So besteht bei dominanten Anpassungsgipfeln eine gute Chan-
ce, „Täler“ zu überwinden, die drei oder vier Mutationen „breit“ sind (rote Punkte). Bei rezessi-
ven Gipfeln ist dies schwieriger (blaue Quadrate). Grafik verändert nach TROTTER et al. (2014).
- 48 -
Abb. 16 Die erwartbare Zahl von Anpassungen in einer bestimmten Population als Funktion
der Talbreite. Wie in Abb. 15 symbolisieren die Linienfarben verschiedene Grade der „Kryptizi-
tät“. Die Autoren definieren sie als Stärke der Selektion, die auf die kryptischen Gen-Varianten
wirkt, im Verhältnis zur vollen Selektionsstärke, die auf die nicht-kryptischen Varianten wirkt.
Für jede Talbreite (j) führten die Autoren 100.000 Simulationen durch. Als Ergebnis erhielten
sie Monte-Carlo-Schätzungen für die Wahrscheinlichkeit, mit der sich j-dimensionale Anpas-
sungen in einer bestimmten Population durchsetzen würden. Diese Wahrscheinlichkeiten mul-
tiplizierten die Autoren mit der möglichen Zahl j-dimensionaler Anpassungen. Das Ergebnis:
Die Zahl der komplexen Anpassungen durch Rekombination polymorpher Gen-Varianten, die
in der Population fixiert wird, ist im Fall hoher Kryptizität erheblich. Aus: TROTTER et al. (2014).

- 49 -
G. Der Design-Ansatz in der biologischen Forschung
G.1 Gibt es „gute“ Erklärungslücken?
Auch wenn wir über die Evolution nicht reduzierbar komplexer Gegenstände nichts
wüssten, wäre die Annahme ihrer natürlichen Evolution nicht unplausibel. Warum?
Weil fehlendes Wissen kein Argument gegen eine Theorie sein kann: Es zeigt ledig-
lich, wo noch Forschungsbedarf besteht. Doch nehmen wir
„… einmal contra factum proprium an, der Beweis sei gelungen, dass die Evolutionstheo-
rie die Entstehung zweckmäßiger Strukturen und Funktionen nicht erklären kann. Intelli-
gent Design würde daraus nicht folgen. Mehrere andere Möglichkeiten lägen näher, zum
Beispiel der gute, alte Vitalismus des 19. und 20. Jahrhunderts, oder ein Stufenbau der
Natur, wie ihn Esoteriker annehmen, in dem höhere, feinstoffliche Seinsebenen die Bau-
muster für niedere, materielle Ebenen enthalten. Es gibt keinen Weg, von der Lücke auf
den Lückenbüßer zu schließen, solange man ein rein negatives Argument für die Lücke
entwickelt“ (HEMMINGER 2019).
Kurzum: Kritiker des Design-Ansatzes sprechen vom „Appell an das Nichtwissen“, vom
Fehlschluss des argumentum ad ignorantiam oder vom Lückenbüßer-Argument: De-
sign-Anhänger interpretieren die vermeintlichen oder tatsächlichen Erklärungs-Defizite
als Scheitern der Evolutionstheorie. Dann schieben sie vorschnell ihren Designer in die
Lücke, wodurch sie einer natürlichen Erklärung vorgreifen. Ist der Vorwurf berechtigt?
Merke: Es existiert keine logische Brücke von nicht reduzierbarer Komplexität zu
einem „intelligenten Design“. Entgegen Beteuerungen der ID-Vertreter lässt sich ein
solcher Weg nicht aufzeigen. Nicht reduzierbare Komplexität wäre ein Argument
gegen Evolution, wenn gezeigt wäre, dass sie für die Evolution eine nicht über-
windbare Hürde darstellte. Dieser Nachweis existiert nicht. Das Gegenteil ist der
Fall: Theoretische Überlegungen zeigen, dass die Entstehung komplexer Anpas-
sungen unter bestimmten Voraussetzungen sehr wahrscheinlich ist.
Doch nehmen wir an, Intelligent Design erbrächte diesen Nachweis. Dann wäre
nicht reduzierbare Komplexität nicht automatisch ein Argument für Design. Das wä-
re der Fall, wenn gezeigt wäre, dass der Design-Ansatz der Schluss auf die beste
Erklärung ist. Mangels überzeugender „positiver Komponente“ des Design-
Arguments ist auch dieser Nachweis nicht erbracht.

- 50 -
Es gibt Tatsachen, die sich prinzipiell nicht durch das Wechselspiel von Zufall und
Notwendigkeit erklären lassen. Dazu gehört die Konstellation der Steinblöcke von
Stonehenge. Wie natürliche Mechanismen die Steine derart spezifisch anordneten, ist
rätselhaft. Hier ist das Fehlen einer natürlichen Erklärung keine wirkliche Erklärungslü-
cke, sondern, wie manche behaupten, eine „gute Lücke“, die auf anerkanntem Wissen
beruht. Die beste Erklärung lautet, dass Menschen die Steine anordneten (Abb. 17).
Abb. 17 Die jungsteinzeitliche Megalith-Struktur Stonehenge. Bauwerke dieser Art sind in der
menschlichen Zivilisation nichts Ungewöhnliches. Doch bemerkenswert ist ihr Alter; die ge-
samte Anlage ist rund 5.000 Jahre alt. Nichts deutet auf eine natürliche Entstehung der spe-
ziell arrangierten Steinblöcke hin. Die Annahme beispielsweise, es handele sich um eiszeitli-
che „Findlinge“, die zufällig so zu liegen kamen, nachdem Gletscher schmolzen, wäre zu weit
hergeholt. Hier ist die, wenn wir es so ausdrücken wollen, Erklärungslücke positiv begründet.
Auch Design-Protagonisten behaupten, auf der Basis bekannten Wissens gegen Evolu-
tion und für Schöpfung zu argumentieren. Wer ihnen, so entgegnen sie, den Gebrauch
von Lückenbüßer-Argumenten unterstelle, ignoriere die positive Komponente des
Schlusses auf Design. Wir haben jedoch in den Abschnitten C. bis E. gesehen, dass
es keine überzeugende positive Komponente des Design-Arguments gibt. In der
Biologie existieren keine „guten Erklärungslücken“, um damit ID zu rechtfertigen.

- 51 -
Schlussendlich besinnt sich die Wissenschaft auf das, was sie über die Evolution weiß,
und nicht auf das, was sie nicht weiß: Wir kennen ein Arsenal natürlicher Evolutions-
mechanismen, die das Potenzial haben, komplexe Neuerungen entstehen zu lassen.
Theoretische und empirische Ergebnisse stützen diese Auffassung.15 Und solange
kein vernünftiger Grund vorliegt, warum Evolution an der nicht reduzierbaren Komple-
xität scheitern soll, bleibt die naturalistische Evolutionstheorie unangefochten.
G.2 Ist der Design-Ansatz ein „Science-Stopper“, oder stärkt ihn die Forschung?
Formulieren wir die Frage zunächst positiv: Bringt Intelligent Design die Forschung
voran? Bestenfalls insofern, als Design-Ansätze helfen, die Komplexität konkreter Bio-
systeme zu bestimmen. Doch da dieses Unternehmen in den Aufgabenbereich ver-
schiedener biologischer Disziplinen fällt, ist ein Mehrwert des Design-Ansatzes nicht
erkennbar. Günstigstenfalls zwingt er Biowissenschaftler, ihre Erklärungsmodelle de-
taillierter auszuarbeiten als bisher.16
HAFER (2015) weist nach, dass in ID-Publikationen empirische Aspekte, Modellbildung
und Vorhersagen eine untergeordnete Rolle spielen. Eigene Forschungsprogramme
haben sie nicht vorzuweisen, der kritische Aspekt überwiegt. Wie sollen sich unter die-
sen Umständen Biologie und der Design-Ansatz gegenseitig befruchten? Hinsichtlich
15 Siehe Fußnote 3.
16 Im ungünstigeren Fall müssen sie aufwändige Repliken schreiben, um Behauptungen über die Evolu-
tionstheorie gerade zu rücken, die ID-Vertreter in die Welt setzen. Einer der Negativ-Höhepunkte war
das unwürdige Gezänk um die Frage, ob Birkenspanner ein Beispiel für Selektion sind (vgl. PROFFITT
2007). Um Fragen mit Tragweite ging es nicht. Ideologisch aufgeladene Debatten der ID-Bewegung
behindern die wissenschaftliche Forschung zu sehr, als dass sie ein Stimulus wären.
Merke: Zwar ist der Vorwurf, die Evolutionsgegner bedienten sich des Lückenbüßer-
Arguments, vereinfacht. Doch mangels einer überzeugenden „positiven Komponen-
te“ des Design-Arguments spricht nichts für die Annahme, es gäbe ontologisch be-
gründete Erklärungslücken („gute Lücken“) für Intelligent Design.
Fehlende Erklärungen der Evolutionstheorie erhöhen nicht die Plausibilität des De-
sign-Ansatzes. Ebenso wenig schwächen sie die Evolutionstheorie, denn fehlendes
Wissen ist kein gutes Argument gegen Theorien. Da Design-Anhänger nach dieser
Logik argumentieren, ist es nicht verfehlt, ihnen den Gebrauch des Lückenbüßer-
Arguments zu unterstellen.

- 52 -
der Ursprungsfrage profitiert die Biologie erst recht nicht von Intelligent Design. Die
kausale Frage, die für die Wissenschaften konstituierend ist, klammert es aus. Belege
für Existenz potenzieller Akteure und für den Einsatz geeigneter Verfahrensweisen im
Präkambrium bleibt der Design-Ansatz schuldig. Ungeachtet dessen und trotz der
Rückzugsgefechte, zu denen sie die Datenlage zwang (↑ Abschnitt F.2 und F.3), be-
haupten die Autoren im Brustton der Überzeugung:
„Ein wichtiger Aspekt ist, dass der Fortschritt der Forschung das Design-Argument eher
gestärkt als geschwächt hat“ (S. 1).
Intelligent Design kann allenfalls damit punkten, dass sich herausstellte, dass die mo-
lekularbiologischen Prozesse komplizierter sind, als die Wissenschaft erwartet hatte.
Wir konzedieren, dass sich in bestimmten Fällen die Frage der Evolution in verschärf-
tem Maß stellt. Doch wie gezeigt, stärken weder ein hohes Maß an nicht reduzier-
barer Komplexität noch offene Fragen zur Evolution den Design-Ansatz.
Zudem vervollständigt sich der Erklärungsrahmen der Evolutionsbiologie stetig. Den
gegenteiligen Eindruck erweckt, wer entsprechende Beispiele ignoriert oder ihre Be-
deutung herunterspielt. Diese Strategie ist nicht überzeugend, da die oben diskutierten
Rückzugsgefechte auf Seiten der deutschen ID-Vertreter nicht irgendwelche Beispiele
betreffen. Sie betreffen Beispiele nicht reduzierbar komplexer Strukturen, die ID-
Vertreter gegen Evolution in Stellung brachten.
Kommen wir auf die Frage zurück, ob ID die Forschung behindert, das heißt einen
„Science-Stopper“ darstellt. Wie eingangs zitiert, schließen die Autoren eine natürliche
Entstehung biologischer Systeme nicht kategorisch aus. Sie halten diese lediglich für
unwahrscheinlich. Deshalb habe wissenschaftliches Forschen zu klären, ob das De-
sign-Argument davon profitiere oder an Plausibilität einbüße. Der Design-Ansatz sei
kein „Science-Stopper“, sondern auf Forschung angewiesen. Mit ihr lote Intelligent De-
sign die Grenzen der natürlichen Evolution aus.
Dieses Ansinnen erscheint legitim. Das Problem ist nur: Die Protagonisten des intelli-
genten Designs behaupten unverdrossen, natürliche Erklärungsversuche scheiterten
an der Ursprungsfrage. Für differenzierte Zwischentöne ist kein Platz. Viele verlangen,
Prozesse, die so langsam ablaufen, dass sie Jahrtausende und Jahrmillionen in An-
spruch nehmen, im Labor nachzustellen – eine absurde Forderung. Und vor allem: Sie
bringen Design ins Spiel, lange bevor der Naturalismus konsequent ausge-
schöpft ist. Wir sind noch nicht recht dabei, die Felder abzustecken, die es zu erfor-

- 53 -
schen gilt. Wie lange wird beispielsweise an der chemischen Evolution geforscht? Seit
65 Jahren. Wie alt ist das Forschungsfeld der „Evolutionären Entwicklungsbiologie“?
Kaum mehr als 30 Jahre! Was ist das im Vergleich zu 400 Jahren Physik?17
Darum hat HAUGHT (2004, S. 238) recht, der bemerkt:
„Intelligent Design ist ein ‚Wissenschafts-Stopper‘, da es bereits zu einem Zeitpunkt eine
Lückenbüßer-Erklärung anbietet, wo in der Forschung noch genügend Raum für weitere
wissenschaftliche Aufklärungen besteht“ (ins Deutsche M.N.)18
Dadurch, dass sich die wenigsten derer, die ID offensiv vertreten, an der Aufklärung
der betreffenden Mechanismen beteiligen, gewinnt dieser Einwand an Schärfe. Die
Protagonisten des intelligenten Designs verabschieden sich hier aus der Forschung.
Oft zeigen sie nicht das geringste Interesse an der Ursprungsfrage. In erfrischender
dogmatischer Klarheit bestätigt dies der „Baraminologe“ Todd WOOD (2010):
„Das ist der Grund, weshalb ich mich für die Entstehung des Lebens nicht interessiere
(und warum ich wahrscheinlich nie Meyer’s Buch zu Ende lesen werde). Ich weiß ja be-
reits, woher das Leben stammt. Ich schlage das Buch Genesis auf, und die Bibel teilt mir
genau mit, woher das Leben stammt. Darüber zu spekulieren, wie es in einem naturalis-
tischen Szenario entstanden sein könne, ist für mich reine Zeitverschwendung“ (ins
Deutsche M.N.)19
Wie der Evolutionshistoriker Thomas JUNKER (2009, S. 337) bemerkt, geht das Desin-
teresse so weit, dass die Kreationisten bezüglich der mutmaßlichen Schöpfung „von
einem Forschungsverbot fabulieren“. Zu beachten sei,
„dass der Schöpfungsakt an sich nicht naturwissenschaftlich beschrieben werden soll
und kann (JUNKER & SCHERER 2006, S. 296)“.
17 Falls die Evolutionsforschung in den nächsten 400 Jahren auf der Stelle treten sollte, können WIDEN-
MEYER & JUNKER gern wiederkommen und ihre Kritik neu auflegen. Vorher ist die Behauptung, der Natu-
ralismus scheitere an der Ursprungsfrage, und es gäbe diesbezüglich „gute Erklärungslücken“, abstrus.
18 „Intelligent Design is a ‚Science stopper‘, since it appeals to a God-of-the-gaps explanation at a point
in inquiry when there is plenty of room for further scientific elucidation.”
19 „That's why I don't care about the origin of life (and why I'll probably never finish reading Meyer's
book). I already know where life came from. I open the book of Genesis, and the Bible tells me exactly
where life came from. Speculating on how it might have happened in a naturalistic scenario seems like a
waste of time to me.“
Erwartungsgemäß retourniert WOOD diesen Einwand an die Adresse der „Atheisten“, die sich ihrerseits
für die Erforschung der „Arche Noah“ nicht interessierten. Aber das ist Blödsinn, denn wir können
schwerlich jemandem den Vorwurf machen, dass er nicht beliebige Phantasien erforscht.

- 54 -
Warum? Wäre der zur Rede stehende Schöpfungsakt ein wohlbegründeter Sachver-
halt, wäre nicht einzusehen, warum er nicht naturwissenschaftlich beschreibbar sein
sollte. Disziplinen wie die experimentelle Archäologie haben kein Problem damit, die
Schöpfungsakte der betreffenden Gegenstände zu erforschen. Vor diesem Problem
steht allein – Intelligent Design. Und weshalb? Weil der Design-Ansatz in der Biologie
offensichtlich einen fiktiven Sachverhalt behandelt.
H. Zusammenfassung
Die „positive“ Komponente des Design-Arguments kann nicht überzeugen, das heißt:
Der Schluss von der funktionalen Zweckmäßigkeit und „nicht reduzierbaren“ Komplexität
biotischer Merkmale und Prozesse auf einen Zwecksetzer ist keineswegs stichhaltig. All
unser Wissen zeigt nämlich, dass Lebewesen evolutionsfähige Mehrgenerationen-
Systeme sind. Tragfähige Analogien zwischen technischen und biologischen Systemen
gibt es nicht. Entgegen den Autoren ist nicht reduzierbare Komplexität kein Design-Indiz.
Des Weiteren ist der Design-Ansatz nur prüfbar, wenn ihn seine Protagonis-
ten spezifizieren: Er ist mit zusätzlichen Hypothesen über das mutmaßliche Schöpfer-
handeln zu versehen. Die Autoren von WORT UND WISSEN tun dies zwar in mancherlei
Hinsicht, beispielsweise, indem sie das Schöpferhandeln als menschenähnlich einstufen
und dann die strukturellen Ähnlichkeiten zwischen technischen Gegenständen und Bio-
systemen als Schöpfungs-Indiz deuten. Und selbstverständlich bietet ein intelligentes
Design Möglichkeiten, die im rein naturalistischen Rahmen so nicht existieren. Aber es
sind nur (Denk-) Möglichkeiten, keine empirisch begründeten Plausibilitäten. Warum?
Zum einen besteht das Problem darin, dass speziellere Design-Modelle entweder keiner
Prüfung standhalten, wie das Paradigma des Kreationismus zeigt. Oder sie enthalten
Hypothesen betreffs des Schöpferhandelns, die nicht unabhängig vom Design-Ansatz
prüfbar sind. Die Annahme, der Schöpfer stecke hinter Zufallsmutationen oder schreibe
das Repertoire künftiger Anpassungen in die Genome von Organismen („programmierte
Variabilität“) sind Beispiele. Die Annahme, eine durchgehend natürliche Evolution würde
den Design-Ansatz schwächen, ist ebenfalls eine Annahme, die nicht unabhängig vom
Design-Ansatz prüfbar ist.
Selbiges gilt für die Prämisse der Autoren, der Arten-Schöpfer sei in seinem Wir-
ken menschenähnlich oder gestalte die Dinge optimal: Wenn ich den postulierten
Schöpfer so spezifiziere, dass er Menschenähnliches schuf und Optimales hervorbrach-

- 55 -
te, werde ich angesichts teils menschenähnlicher und optimaler Strukturen in der Biolo-
gie immer an einen Schöpfer denken. Das ist trivial, da das Ergebnis schon in den Vo-
raussetzungen steckt. Ohne unabhängige Belege können die Voraussetzun-
gen beliebig gewählt werden.
In der Tat kombinieren Design-Anhänger den Design-Ansatz mit unterschiedlichsten,
sich teils gegenseitig ausschließenden Schöpfungs- und Schöpfervorstellungen. Sie
sind sich weder einig, was der Designer schuf, noch, wie er schuf oder inwieweit dieser
in den Gang der Welt eingriff (oder immer noch eingreift). Einige ID-Vertreter, wie die
Physikerin Barbara DROSSEL, betonen sogar, dass der Schöpferbeitrag mit einem
durchgehend natürlichen Verlauf der Evolution konform gehe, da sie „Design“ nur als
Zweitursache hinter der Natürlichkeit der Prozesse wähnen. Da sie also den Beitrag
von Design teils auf völlig verschiedenen Systemebenen vermuten, ist offensicht-
lich, dass es (derzeit) keine objektiven Design-Indizien in der Biologie gibt.
Wie unter anderem HEILIG (2015) gezeigt hat, besteht ein weiteres Problem des Design-
Ansatzes darin, dass Rechenschaft über die Plausibilität der (historischen) Randbedin-
gungen abgelegt werden muss, bevor der Design-Ansatz favorisiert werden kann.
Das gilt unabhängig davon, ob der „intelligente Prozess“ an sich das Produkt
„erwarten“ lässt oder nicht. So ist es beispielsweise sehr problematisch, ei-
nen menschenähnlichen Schöpfer anzunehmen, der zur Entstehungszeit der ersten
Bakterien vor mehreren Milliarden Jahren gewirkt haben soll. Das Fehlen jedweden his-
torischen Hinweises auf die Existenz „präkambrischer Technologien“ lässt den Schluss
auf Design gegenwärtig nicht plausibel erscheinen. Analoges gilt auch für natürliche
Prozesse: Auch wenn ich beweisen kann, dass Proteine prinzipiell „von selbst“ entste-
hen können, die irdischen Randbedingungen diese Möglichkeit aber äußerst unplausibel
erscheinen lassen, lässt sich nicht vernünftig auf eine chemische Evolution schließen.
Schlussendlich zeigt die Wissenschaft, dass der Design-Ansatz nur fruchtbar ist, wenn
die Verfahrensweisen der mutmaßlichen Urheber rekonstruierbar sind. Das heißt: Es
braucht zusätzliches Designer-Wissen, um vernünftig auf Design zu schließen. Diesbe-
züglich unterscheidet sich Intelligent Design dramatisch von florierenden Design-
Disziplinen wie der Archäologie. Der Design-Ansatz in der Biologie operiert durchweg
mit Unbekanntem: Designer, die infrage kämen, kennen wir nicht. Über ihre mutmaßli-
chen Techniken, Fähigkeiten und Grenzen wissen wir nichts. Erforschen lässt sich der-
gleichen nicht. Brauchbare Modelle, welche die Mechanismen spezifizieren, liegen nicht
vor. Der Design-Ansatz bleibt inhaltsleer und heuristisch steril (HEILIG 2011, S. 91).

- 56 -
Eine weitere Klasse von Einwänden gegen ID bezieht sich auf den Umstand, dass seine
Vertreter unentwegt den „Sinn“ in der Natur herausstreichen und den „Unsinn“ überse-
hen. Sie begehen den Fehlschluss des Texanischen Scharfschützen: Zufällig Passen-
des wird als Ergebnis von „Planung“ gedeutet, Unpassendes der degenerativen Evoluti-
on angelastet. Das ist als würde jemand blind Gewehrkugeln auf ein Scheunentor ab-
feuern, um einige Einschüsse eine Zielscheibe herum pinseln und behaupten, er habe
„ins Schwarze“ getroffen. Die übrigen Schüsse werden nicht berücksichtigt.
In der Biologie schwächt vor allem exzessives Auftreten von Mängelstrukturen das De-
sign-Argument. Entgegen der Behauptung der Autoren setzt dieses Argument keine
theologischen Annahmen voraus. Es geht darum, dass dysfunktionale Merkmale besser
zu Prozessen passen, die keinen Plan kennen. Um sie zu identifizieren, benötigen wir
keine evolutiven Annahmen. Mängelstrukturen verursachen Fitness-Einbußen und er-
höhte Mortalität. Sie sind an Merkmale geknüpft, die aus technischer Sicht unsinnig er-
scheinen und der Idee der Planmäßigkeit im Weg stehen. Die Einwände der Autoren
entpuppen sich als schwache Versuche von Kritikimmunisierung.
Auch die „negative“ Komponente, die den Schluss auf Design rechtfertigen soll, ist ge-
scheitert. Nicht reduzierbare Komplexität wäre ein Argument, wenn gezeigt wäre, dass
sie für die Evolution eine nicht überwindbare Hürde darstellte. Ein solcher Nachweis
existiert nicht, und wir können ihn aus offenen Fragen zur Evolution der betreffenden
Strukturen nicht ableiten. Im Gegenteil: Theoretische und empirische Argumente zeigen,
dass eine Evolution nicht reduzierbar komplexer Strukturen plausibel ist. Ob sie in jedem
Fall kleinschrittig und über selektionspositive Zwischenstufen verläuft, bleibt die Frage.
Doch daraus folgt nicht, die Evolution sei unwahrscheinlich.
Keines der Argumente, welche die Unwahrscheinlichkeit der betreffenden Evolution
begründen sollen, lässt sich schlüssig entfalten. Doch nehmen wir pro forma an, Intel-
ligent Design hätte ein solches Argument vorzuweisen. Dann wäre zwar nicht redu-
zierbare Komplexität ein Argument gegen natürliche Evolution. Aber mangels über-
zeugender positiver Komponente des Design-Arguments wäre der Schluss auf Design
nach wie vor nicht der Schluss auf die beste Erklärung. Wir könnten nur konstatieren,
dass es zum gegenwärtigen Zeitpunkt keine Erklärung gibt.
Zur Frage, inwieweit der Design-Ansatz die Forschung stimuliert, lässt sich festhalten:
Er kann ein Stimulus sein, etwa, wo es um Strukturaufklärung oder um die Frage der
Funktionalität komplexer Strukturen geht. Meist bewirkt er, dass sich Menschen aus

- 57 -
der Erforschung von Entstehungs-Mechanismen zurückziehen oder, wie der Fall von
Todd WOOD zeigt, jedes Interesse an der Entstehung und Entwicklung des Lebens
verlieren. Seine Anhänger propagieren das intelligente Design als Schluss auf die bes-
te Erklärung, lange bevor die Naturwissenschaften den Naturalismus konsequent aus-
schöpften. Daher ist die Feststellung, der Design-Ansatz sei eine Wissenschafts-
Bremse, zutreffend, und es ist legitim, die Frage nach seiner Motivation zu stellen.
Organisationen, die den Design-Ansatz als Instrument der Schöpfungs-Forschung
propagieren, verhehlen selten, worin sein Zweck besteht. „Zu keinem Zeitpunkt“, so
MATT et al. (2014, S. 40), stehe bei solchen Organisationen „die Grundlagenforschung
um des Wissens willen im Mittelpunkt“. Vordergründig fordern sie die Ergebnisoffenheit
des Forschens; im Hintergrund stehen die Erkenntnisse im Wesentlichen bereits fest.
Der Zweck der Schöpfungs-Forschung ist also vornehmlich Apologetik:
„Schöpfungsforschung lässt sich zwar nicht direkt evangelistisch einsetzen, doch erfüllt
sie die Funktion als ‚Vorbereitungs-‘‚ und ‚Unterstützungswerkzeug‘ zu einer Evangelisa-
tion und danach. Gerade Neubekehrte haben viele Fragen an den christlichen Glauben
und brauchen in dieser Hinsicht auch fundierte Hilfe. Schöpfungsforschung bringt die Bi-
bel in konkreten Bezug zur materiellen Welt und hilft suchenden und fragenden Men-
schen, Glaubenshindernisse aus dem Weg zu räumen.“20
Fragende Menschen, die nach bestmöglich gesichertem Wissen suchen, sollten sich
keinen Illusionen hingeben: Der Design-Ansatz ist in seiner aktuellen Form kein
brauchbares Erkenntnisinstrument. Ja, schlimmer: Als Instrument religiöser Missionie-
rung konterkariert er das wissenschaftliche Ethos der freien Suche nach Wahrheit.
I. Dank
Verschiedene Hinweise, die das argumentative Profil dieser Arbeit schärfen, verdanke
ich dem Biologen und Wissenschaftsphilosophen Dr. Martin MAHNER, Gründungsmit-
glied der GWUP und Leiter des Zentrums für Wissenschaft und kritisches Denken. Für
kritisches Gegenlesen und hilfreiche Verbesserungs-Vorschläge danke ich Prof. Dr.
Andreas BEYER, Dr. Hansjörg HEMMINGER und Steffen MÜNZBERG.
20 www.wort-und-wissen.de/disk/d07/5/d07-5.pdf

- 58 -
J. Literatur
BEHE, M. (1996) Darwin's black box. The biochemical challenge to evolution. Free Press, New York.
BEHE, M. (2008) The Edge of evolution. The search for the limits of Darwinism. Free
Press, New York.
BEYER, A. (2018) T-urf 13 revisited: Ein "intelligent designtes“ Protein – ohne Designer. Über die zufällige Entstehung einer nicht reduzierbar komplexen Struktur.
www.ag-evolutionsbiologie.net/html/2018/t-urf13-design-ohne-designer.html.
BODYŁ, A. et al. (2009) Early steps in plastid evolution: current ideas and controver-sies. Bioessays 31, S. 1219–1232.
BÖLTER, B. et al. (1998) Origin of a chloroplast protein importer. Proceedings of the National Academy of Sciences of the USA 95, S. 15831–15836.
BRIGANDT, I. (2013) Intelligent Design and the nature of science. Philosophical and
pedagogical points. In: KAMPOURAKIS, K. (Ed.) The philosophy of biology. History, philosophy and theory of the life sciences. Springer Spektrum Verlag, Heidelberg, S. 205–238.
BRINGMANN, A. & REICHENBACH, A. (2001) Role of Muller cells in retinal degenerations. Frontiers in Bioscience 6, E72–92.
BURDA, H. & BEGALL, S. (Hg., 2013) Evolution. Ein Lese-Lehrbuch, 2. Auflage. Springer
Spektrum Verlag, Heidelberg.
CARROLL, R.T. (2015) Argument from design. In: The Sceptic's dictionary. A collection of strange beliefs, amusing deceptions, and dangerous delusions.
skepdic.com/design.html Zugr. a. 03.10.2018.
COVELLO, P. S. & GRAY, M. W. (1992) Silent mitochondrial and active nuclear genes for subunit 2 of cytochrome c oxidase (cox2) in soybean: evidence for RNA-mediated
gene transfer. The EMBO Journal 11, S. 3815–3820.
DALEY, D. O. et al. (2002) Intracellular gene transfer: Reduced hydrophobicity facili-tates gene transfer for subunit 2 of cytochrome c oxidase. Proceedings of the Na-
tional Academy of Sciences of the USA 99, S. 10510–10515.
DE KRAKER, J.-W.; GERSHENZON, J. (2011) From amino acid to glucosinolate biosynthe-sis: Protein sequence changes in the evolution of methylthioalkylmalate synthase
in Arabidopsis. The Plant Cell 23, S. 38–53.
DEMBSKI, W. A. (2003) Evolution's logic of credulity. www.arn.org/docs/dembski/wd_logic_credulity.htm. Zugr. a. 31.08.2018.
DENTON, M. (2016) Evolution: still a theory in crisis. Discovery Institute Press, Seattle.
DISCOVERY INSTITUTE (2012) Intelligent Design in action: archaeology. evolu-tionnews.org/2012/05/id_in_action_ar/. Version vom 15.05.2012.

- 59 -
DRAPER, P. (2002) Irreducible Complexity and Darwinian Gradualism. A reply to Mi-
chael J. Behe. Faith and Philosophy 22, S. 3–21.
DRENDEL, K. (2016) The irreducible complexity of the universe. pemiwad-er.blog/2016/01/28/the-irreducible-complexity-of-the-universe. Version vom
28.01.2016.
FANI, R. (2012) The origin and evolution of metabolic pathways: why and how did pri-mordial cells construct metabolic routes? Evolution: Education and Outreach 5, S.
367–381.
FRANZE, K. et al. (2007) Müller cells are living optical fibers in the vertebrate retina. PNAS 104, S. 8287–8292.
FRANZE, K. & GROSCHE, J. (2008) Müller-Zellen in einem anderen Licht. Biospektrum 14, S. 701–703.
HAFER, A. (2015) No data required. Why Intelligent Design is not science. The Ameri-
can Biology Teacher 77, S. 507–513.
HALL, B. K. (2012) Parallelism, deep homology, and evo-devo. Evolution & Develop-ment 14, S. 29–33.
HAUGHT, J. F. (2004) Darwin, design, and divine providence. In: DEMBSKI, W. A. & RUSE, M. (Hg.) Debating design. From Darwin to DNA. Cambridge University Press, Cambridge, S. 229–244.
HEILIG, C. (2011) Anonymes oder Spezifisches Design? In: HEILIG, C. & KANY, J. (Hg.) Die Ursprungsfrage: Beiträge zum Status teleologischer Antwortversuche in der Naturwissenschaft. LIT-Verlag, Münster, S. 73-125.
HEILIG, C. (2015) Argumentation und Plausibilität: Das Design-Argument im deutsch-sprachigen Raum im Licht von Bayes' Theorem. Freiburger Zeitschrift für Philo-sophie und Theologie 62, S. 368–388.
HEMMINGER, H. (2019) Warum „intelligentes Design“ keine Naturwissenschaft ist. Eine Einführung für Nichtbiologen. www.theologie-naturwissenschaften.de/startseite/leitartikelarchiv/intelligent-design/
INABA, T. & SCHNELL, D. J. (2008) Protein trafficking to plastids: one theme, many va-riations. Biochemical Journal 413, S. 15–28.
JACOBS, G. H. et al. (1999) Human cone pigment expressed in transgenic mice yields
altered vision. Journal of Neuroscience 19, S. 3258–3265.
JUNKER, R. (2005) Argumente gegen Design. www.genesisnet.info/index.php?Sprache=de&Artikel=1641&l=2. Version vom
05.08.2005.
JUNKER, R. (2014) Brauchen wir eine neue Evolutionstheorie? www.genesisnet.info/index.php?News=221. Version vom 09.12.2014.
JUNKER, T. (2004) Evolution und die Marmeladetheorie des Erdkerns. Materialien zur Zeit 33, S. 10–13.

- 60 -
JUNKER, T. (2009) Kreationisten erklären die Evolution. Das „kritische Lehrbuch“ von
Reinhard Junker und Siegfried Scherer. In: NEUKAMM, M. (Hg.), S. 321–338.
JUNKER, R. & SCHERER, S. (2006) Evolution. Ein kritisches Lehrbuch, 6. Auflage. Weyel-Verlag, Gießen.
JUNKER, R. & SCHERER, S. (2013) Evolution. Ein kritisches Lehrbuch, 7. Auflage. Weyel-Verlag, Gießen.
KIRSCHNER, M. W. & GERHART, J. C. (2007) Die Lösung von Darwins Dilemma. Wie die
Evolution komplexes Leben schafft. Rowohlt-Verlag.
KOJONEN, E. V. R. (2016) The Intelligent Design debate and the temptation of scien-tism. Routledge, London.
KOVACHEVA, S. et al. (2005) In vivo studies on the roles of Tic110, Tic40 and Hsp93 during chloroplast protein import. The Plant Journal 41, S. 412–428.
LIGHT, S. & KRAULIS, P. (2004) Network analysis of metabolic enzyme evolution in
Escherichia coli. BMC Bioinformatics 5, 15. doi: 10.1186/1471-2105-5-15.
LÖNNIG, W.-E. (1993) Artbegriff, Evolution und Schöpfung. Grundsätzliche Einwände zur Artbildung durch Intelligent Design. www.weloennig.de/AesIV4.html. Zugr. a.
15.07.2018.
LORENZEN, S. (1988) Die Bedeutung synergetischer Modelle für das Verständnis der Makroevolution. Eclogae Geologicae Helvetiae 81, S. 927–933.
LUSKIN, C. (2006) National Geographic evolution article discusses evidence that sup-ports Intelligent Design (Part III). evolu-tionnews.org/2006/11/national_geographic_evolution.
MAHNER, M. (1986) Kreationismus. Inhalt und Struktur antievolutionistischer Argumen-tation. Pädagogisches Zentrum, Berlin.
MAHNER, M. (2007) Intelligent Design und der teleologische Gottesbeweis. In: KUT-
SCHERA, U. (Hg.): Kreationismus in Deutschland. Lit-Verlag, Münster, S. 340-351.
MAHNER, M. & BUNGE, M. (2000) Philosophische Grundlagen der Biologie. Springer-Verlag, Berlin.
MARTIN, W. et al. (2002) Evolutionary analysis of Arabidopsis, cyanobacterial, and chloroplast genomes reveals plastid phylogeny and thousands of cyanobacterial genes in the nucleus. Proceedings of the National Academy of Sciences of the
USA 99, S. 12246–12251.
MATT et al. (2014) Nichts Neues durch Evolution? In: dies, Schöpfung – oder wie es wirklich war, S. 53–65. tinyurl.com/yblbttg6. Version vom 15.04.2014.
MENUGE, A. J. L. (2004) Agents under fire: Materialism and the rationality of science. Rowman & Littlefield Publishers, Lanham.
MILLER, K. R. (2007) Finding Darwin’s god. A scientist's search for common ground
between god and evolution. Harper Perennial, New York.

- 61 -
MONTEIRO, A. & PODLAHA, O. (2009) Wings, horns, and butterfly eyespots: How do
complex traits evolve? PLoS Biology 7, e1000037.
MÜLLER, W. & HASSEL, M. (2018) Entwicklungs- und Reproduktionsbiologie des Men-schen und bedeutender Modellorganismen, 6. Auflage. Springer Spektrum Ver-
lag, Heidelberg.
MYERS, P. Z. (2010) More creationst misconceptions about the eye. pan-dasthumb.org/archives/2010/06/more-creationis.html. Version vom 07.06.2010.
NEUKAMM, M. (2009) Evolution im Fadenkreuz des Kreationismus. Darwins religiöse Gegner und ihre Argumentation. Vandenhoeck & Ruprecht, Göttingen.
NEUKAMM, M. (2014) Darwin heute. Evolution als Leitbild in den modernen Wissen-
schaften. Wissenschaftliche Buchgesellschaft, Darmstadt.
NEUKAMM, M.; BEYER, A. & PEITZ, H.-H. (2013) Zur Evolution des „Bakterienmotors“. www.ag-evolutionsbiologie.net/pdf/2013/Die-Evolution-bakterieller-Flagellen.pdf
NOWITZKI, U. (2002) Studien zum Gentransfer aus Organellen nach Endosymbiose anhand der Chloroplast-Cytosol Isoenzyme von Glucose-6-phosphat Isomerase aus Spinacia oleracea und der 3-Phosphoglycerat Kinase aus Euglena gracilis.
Dissertation, TU Braunschweig. d-nb.info/964591677/34. Zugr. a. 20.10.2018.
ORR, H. A. (1996) Darwin v. Intelligent Design. Boston Review 21, S. 28–31.
PROFFITT, F. (2004) In defense of Darwin and a former icon of evolution. Science 304,
S. 1894–1895.
RAMMERSTORFER, M. (2003) Eine Theorie zur Signalerkennung und ihre (möglichen) Folgen. members.aon.at/evolution/IDTkonseq.htm. Version vom 02.05.2003.
RAMMERSTORFER, M. (2004) Nervus laryngeus recurrens - suboptimal? mem-bers.aon.at/evolution/NLrecurrens.pdf.
RAMMERSTORFER, M. (2006) Nur eine Illusion? Biologie und Design. Tectum Wissen-
schaftsverlag, Marburg.
RHONHEIMER, M. (2007) Neodarwinistische Evolutionstheorie, Intelligent Design und die Frage nach dem Schöpfer. Imago Hominis 14, S. 47–81.
REUMANN, S. et al. (2005) Evolution of the general protein import pathway of plastids (review). Molecular Membrane Biology 22, S. 73–86.
ROBINSON, K. (1996) Darwin's Black Box. Irreducible Complexity or Irreproducible Irre-
ducibility? www.talkorigins.org/faqs/behe/review.html. Version vom 11.12.1996.
SCHMIDT-SALOMON, M. (2005) Existiert Gott? Erster Beitrag zur Debatte mit dem ameri-kanischen Kreationisten Dr. William Lane Craig. www.giordano-bruno-
stiftung.de/sites/gbs/files/download/existmss1.pdf. Version vom 26.04.2005.
SCHERER, S. (2010) Die Entstehung des bakteriellen Rotationsmotors ist unbekannt. www.evolutionslehrbuch.info/teil-4/kapitel-09-04-r01.pdf. Version vom 04/2010.
SCHUSTER, P. (2014) Evolution der Moleküle. Von der Evolution im Reagenzglas zur Erzeugung maßgeschneiderter Moleküle. In: NEUKAMM (Hg.), S. 133–170.

- 62 -
SOBER, E. (2008) Evidence and evolution: The logic behind the science. Cambridge
University Press, Cambridge.
SOLL, J. & SCHLEIFF, E. (2004) Protein import into chloroplasts. Nature Reviews Mo-lecular Cell Biology 5, S. 198–208.
STEGEMANN, S. & BOCK, R. (2006) Experimental reconstruction of functional gene transfer from the tobacco plastid genome to the nucleus. The Plant Cell 18, S. 2869–2878.
THORNHILL, R. H. & USSERY, D. W. (2000) A classification of possible routes of Darwini-an Evolution. Journal of Theoretical Biology 203, S. 111–116.
TOEPFER, G. (2004) Zweckbegriff und Organismus. Königshausen & Neumann, Würz-
burg.
TONKIN, C. J. et al. (2008) Evolution of malaria parasite plastid targeting sequences. PNAS 105, S. 4781–4785.
TROTTER, M. V.; WEISSMAN, D. B.; PETERSON, G. I.; PECK, K. M. & MASEL, J. (2014) Cryptic genetic variation can make “Irreducible Complexity” a common mode of adaptation in sexual populations. Evolution 68, S. 3357–3367.
ULLRICH, H. (2014) „Fehlkonstruktion Auge“: Am Ende nur ein Scherz? Studium Integ-rale Journal 21, S. 114–115.
WIDENMEYER, M. & JUNKER, R. (2016) Der Kern des Design-Arguments in der Biologie
und warum die Kritiker daran scheitern. www.wort-und-wissen.de/artikel/a22/a22.pdf. Version vom 27.07.2016.
WOOD, T. (2010) Is design an inference? toddcwood.blogspot.com/2010/02/is-design-
inference.html. Version vom 25.02.2010.
WORT UND WISSEN (2008) Die Endosymbiontenhypothese. evolutionslehr-buch.info/index2.php?artikel=teil-5/kapitel-10-03.html. Version vom 22.03.2011.
© AG Evolutionsbiologie. Letzte Aktualisierung: 01.08.2019