Embed Size (px)
Citation preview

Aus dem Institut für Prophylaxe und Epidemiologie der Kreislaufkrankheiten,
Klinikum Innenstadt, Ludwig-Maximilians-Universität München Vorstand: Prof. Dr. med. P.C. Weber
Funktion und Regulation der
Tyrosinkinase Syk in Thrombozyten
Dissertation zum Erwerb des Doktorgrades der Humanbiologie
an der Medizinischen Fakultät der Ludwig-Maximilians-Universität zu München
vorgelegt von Petra M. Maschberger
aus Altötting
2004

2
Mit Genehmigung der Medizinischen Fakultät der Universität München
Berichterstatter: Prof. Dr. med. W. Siess Mitberichterstatter: Prof. Dr. B. Walzog Priv.Doz. Dr. M. Kretzler Dekan: Prof. Dr. med. Dr. h.c. K. Peter Tag der mündlichen Prüfung: 21.10.2004

3
Meiner Familie gewidmet

4
Gliederung 1. Einleitung 7
1.1 Klinische Relevanz der Untersuchungen 7
1.2 Thrombozyten, oxidativ modifiziertes LDL, Thrombin und kardiovaskuläre
Erkrankungen 7
1.3 Tyrosinkinase Syk 9
1.4 Syk-Aktivierung in Thrombozyten 13
1.5 Fragestellung 17
2. Material und Methoden 18
2.1 Material 18
2.1.1 Geräte 18
2.1.2 Chemikalien, Enzyme, Antikörper, Fluoreszenzfarbstoffe, Reagenzien 18
2.1.3 Bakterien- und Hefestämme, Vektoren, cDNA-Banken 20
2.2 Methoden 23
2.2.1 Isolation von humanen Thrombozyten, Maus-Thrombozyten und Maus-
Megakaryozyten 23
2.2.2 Messung des shape change und der Aggregation 25
2.2.3 Milde Oxidation von LDL 25
2.2.4 Immunpräzipitation von Proteinen aus Thrombozyten-Lysaten (Syk, FAK,
PLCγ2) 26
2.2.5 Isolierung des Thrombozyten-Aktinzytoskeletts 27
2.2.6 Pull-down Assay aus Thrombozytenlysaten mit der Tandem-SH2-Domäne
von Syk 27
2.2.7 Messung der Proteintyrosinphosphorylierung mittels Western Blot 28
2.2.7.1 Diskontinuierliche SDS-Gelelektrophorese in Polyacrylamidgelen 28
2.2.7.2 Proteintransfer aus SDS-Polyacrylamidgelen auf Nitrocellulose-
Membranen 29
2.2.7.3 Immunologischer Nachweis der Proteine 30

5
2.2.7.4 Proteinfärbung mit Ponceau S 31
2.2.8 Densitometrische Auswertung des Western Blots 31
2.2.9 Transiente Transfektion von CHO-Zellen 31
2.2.10 Untersuchungen mittels konfokaler Laserfluoreszenzmikroskopie an
ausgebreiteten humanen und Mäuse-Thrombozyten, und CHO-Zellen 32
2.2.11 Untersuchungen an knock-out Mäusen 35
2.2.12 Yeast Two-Hybrid System 35
3. Ergebnisse 38
3.1 Methodik 38
3.1.1 Expression von Syk in Thrombozyten, aber nicht in Endothelzellen 38
3.1.2 Messung der Syk-Tyrosinphosporylierung nach Immunpräzipitation 39
3.1.3 Expression von Syk bei verschiedenen Probanden 43
3.2 Tyrosinphosphorylierung von Syk während des shape change 44
3.2.1 Aktivierung nach Stimulation des Thrombin-Rezeptors PAR-1 und nach
mox-LDL (milde oxidiertes LDL) 44
3.2.2 Integrin αIIbβ3 unabhängige Tyrosinphosphorylierung von Syk während des
shape change 48
3.2.2.1 Thrombasthenia Glanzmann 48
3.2.2.2 RGDS 51
3.2.3 Regulation 51
3.2.3.1 Proteinkinase C 51
3.2.3.2 Calcium 52
3.2.3.3 Piceatannol und weitere Proteintyrosinkinase-Hemmer, Iloprost 55
3.3 Funktion der Syk-Tyrosinphosphorylierung für den shape change 57
3.3.1 Untersuchungen an mit EGTA vorbehandelten Thrombozyten 57
3.3.2 Einfluß von allgemeinen und spezifischen Inhibitoren der Src-Kinasen auf Syk-
Tyrosinphosphorylierung und shape change 60
3.3.3 Untersuchungen an Syk(-/-)-Mausthrombozyten 63

6
3.4 Integrin αIIbβ3 abhängige Tyrosinphosphorylierung von Syk 65
3.4.1 Transfizierte CHO-Zellen als Modellsystem für einen ITAM unabhängigen
Signalweg der Syk-Aktivierung 65
3.4.2 Thrombozyten 68
3.4.2.1 Thrombozyten-Aggregation: FcRγ-Ketten Tyrosinphosphorylierung,
Syk- und PLCγ2-Aktivierung 70
3.4.2.2 Dissoziation des Integrins αIIbβ3: FcRγ-Ketten Tyrosinphospho-
rylierung, Syk- und PLCγ2-Aktivierung 81
3.4.2.3 Ausbreitung 85
3.5 Yeast Two-Hybrid System zur Identifikation von Interaktionspartnern von Syk 93
4. Diskussion 95
4.1 Integrin αIIbβ3 unabhängige Tyrosinphosphorylierung von Syk während des
shape change 95
4.2 Integrin αIIbβ3 abhängige Tyrosinphosphorylierung von Syk 102
4.3 Perspektive 106
5. Zusammenfassung 107 6. Literaturverzeichnis 109 7. Abkürzungsverzeichnis 124 Anhang Eigene Veröffentlichungen
Danksagung
Lebenslauf

7
1. Einleitung
1.1 Klinische Relevanz der Untersuchungen
Kardio- und cerebrovaskuläre Erkrankungen sind in westlichen Industrieländern führend für
Mortalität und Morbidität. Ein fundamentales Verständnis der pathogenetischen
Mechanismen ermöglicht die Entwicklung neuer Ansätze in Diagnostik, Prävention und
Therapie entsprechender Krankheitsbilder. Bei der Pathogenese der Arteriosklerose sind
verschiedene Zelltypen beteiligt: Thrombozyten, Endothelzellen, Monozyten/Makrophagen
und glatte Muskelzellen, die miteinander und mit Stoffwechselfaktoren interagieren.
Kommunikation zwischen den verschiedenen Zellen wird durch biologische Signale
vermittelt. Zelluläre Interaktionen können hierbei als Vorgänge betrachtet werden, die sich in
zwei Schritten vollziehen. Zunächst bindet ein extrazelluläres Molekül an einen spezifischen
Rezeptor auf der Zelle und versetzt ihn in einen aktiven Zustand. Dann stimuliert der
Rezeptor intrazelluläre biochemische Mechanismen, die zu einer Zellantwort führen.
Die Tyrosinkinase Syk wird bei der Stimulation des B-Zellrezeptors auf Lymphozyten und
verschiedener Fc-Rezeptoren auf Lymphozyten und Monozyten durch
Tyrosinphosphorylierung aktiviert. Auch in Thrombozyten wurde die Aktivierung von Syk
nach Stimulation mit diversen physiologischen Agonisten z.B. Thrombin, PAF und Kollagen
beobachtet. Die Rolle dieser Tyrosinkinase für die Thrombozytenfunktion und thrombozytäre
Signaltransduktion, sowie deren Regulation sind noch weitgehend ungeklärt.
1.2 Thrombozyten, oxidativ modifiziertes LDL, Thrombin und
kardiovaskuläre Erkrankungen
Thrombozyten sind neben ihrer physiologischen Bedeutung in der Hämostase auch in einer
Vielzahl von pathologischen Prozessen involviert. In arteriosklerotischen Blutgefäßen spielen
sie bei der Bildung von Thromben mit der Folge des Myokardinfarkts und Apoplex eine
entscheidende Rolle. Seit Virchow wurden viele Anstrengungen unternommen, um die
Pathogenese der Arteriosklerose zu erklären. Die 1976 entwickelte response to injury
Hypothese lieferte ein wichtiges Konzept, um diesen komplizierten Prozeß zu verstehen.

8
Diese Hypothese involviert eine Aktivierung von Thrombozyten und Monozyten nach einer
Verletzung des Endothels (Ross und Glomset, 1976; Ross, 1986; Ross, 1993). Das Endothel
wird in der Weise geschädigt, daß es für viele Blutproteine, auch das LDL durchlässig wird.
Außerdem können Monozyten an aktiviertes Endothel adhärieren, in die Gefäßintima
immigrieren und dort zu Makrophagen differenzieren. Diese nehmen oxidiertes LDL auf und
werden dadurch zu Schaumzellen, die in der Regel die Gefäßwand nicht mehr verlassen.
Es wird angenommen, daß nicht LDL, sondern vor allem oxidativ modifiziertes LDL den
pathogenetischen Faktor bei der Arteriosklerose darstellt. Weiche lipid-reiche Plaques, welche
u.a. LDL und oxidiertes LDL enthalten, sind vulnerabel und legen nach Ruptur thrombogene
Substanzen und LDL-Partikel frei (Fernandez-Ortiz et al., 1994). Dadurch können
zirkulierende Plättchen innerhalb von Sekunden aktiviert und zur Aggregation gebracht
werden, was zu einer intravaskulären Thrombusbildung führt. Der entstandene Thrombus
kann einen Schlaganfall oder Myokardinfarkt hervorrufen (Falk et al., 1995). In vitro aktiviert
oxidiertes, jedoch nicht natives LDL Thrombozyten, wobei milde oxidiertes LDL (mox-LDL)
aktiver ist als stark oxidiertes LDL (ox-LDL), und im Gegensatz zu stark oxidiertem LDL zur
Thrombozyten-Aggregation führt (Meraji et al., 1992; Weidtmann et al., 1995).
Mox-LDL könnte auch im Kreislauf anwesend sein. Eine Fraktion von LDL, welche aus
elektronegativen, schwach oxidierten LDL-Partikeln geringer Dichte besteht, wurde im
peripheren Blut gefunden (Avogaro et al., 1991; Sevanian et al., 1997). Eine Interaktion der
Plättchen mit zirkulierendem mox-LDL könnte die verstärkte Neigung der Plättchen zur
Aggregation erklären, welche häufig bei Patienten mit kardiovaskulären Erkrankungen
beobachtet wird. Tatsächlich wurde bei Patienten mit koronaren Herzerkrankungen oder
Arteriosklerose ein Vorherrschen von zirkulierenden LDL-Partikeln geringer Dichte
beschrieben (Slyper et al., 1994). Außerdem werden erhöhte Konzentrationen von ox-LDL im
Blut von Patienten mit instabiler Angina pectoris gefunden (Holvoet et al., 1998). Die
Interaktion von zirkulierendem mox-LDL mit Thrombozyten könnte die intravaskuläre
Thrombusbildung an hämodynamisch kritischen Stenosen von Koronararterien und
Carotisarterien begünstigen.
Während ox-LDL oft toxische Effekte auf Zellen hat (Hughes et al., 1994; Coffey et al.,
1995), scheint mox-LDL die Funktion von Zellen der Gefäßwand und der Plättchen durch
Stimulation von spezifischen Rezeptoren und Signaltransduktionswegen zu verändern
(Parhami et al., 1993; Berliner et al., 1995; Weidtmann et al., 1995).

9
Thrombin ist nicht nur der stärkste physiologische Thrombozyten-Stimulus, sondern aktiviert
auch andere vaskuläre Zellen, wie z. B. Endothelzellen, und spielt bei den der Arteriosklerose
zugrunde liegenden hämostatischen, thrombotischen und entzündlichen Prozessen eine
zentrale Rolle (Coughlin et al., 2000). Thrombin wird durch Aktivierung des
Gerinnungssystems gebildet und aktiviert Thrombozyten und andere Zellen über die Bindung
an spezifische G-Protein gekoppelte Rezeptoren (protease-activated receptors, PARs). Die
Aufklärung der molekularen Mechanismen der Thrombozytenaktivierung durch mox-LDL
und Thrombin könnte neue präventive und therapeutische Strategien bei atherosklerotischen
kardiovaskulären Erkrankungen liefern.
1.3 Tyrosinkinase Syk
Der Begriff Kinase geht auf den griechischen Ausdruck kinein zurück, was Bewegen
bedeutet, also Übertragung einer energiereichen Phosphatgruppe von Adenosintriphosphat
(ATP) auf bestimmte Aminosäuren. Diese dynamische Proteinphosphorylierung stellt einen
wichtigen Signalübertragungsmechanismus in der Zelle dar. Vor allem werden hierbei die
drei Aminosäuren Serin, Threonin und Tyrosin phosphoryliert. Die Phosphorylierung durch
Tyrosinkinasen macht weniger als 0,01% der totalen intrazellulären Proteinphosphorylierung
aus.
Die Tyrosinkinase Syk (spleen tyrosine kinase) wurde zuerst in Schweine-Splenozyten
beschrieben (Chan et al., 1991; Taniguchi et al., 1991; Chan et al., 1992; Yamada et al.,
1993). Syk ist stark konserviert zwischen Mensch und Schwein. Das für das humane Syk
kodierende Gen wurde auf dem Chromosom 9, Bande q22 lokalisiert (Law et al., 1994).
Außer Syk in B-Lymphozyten und Mastzellen wurde eine zu Syk homologe
Proteintyrosinkinase, ZAP-70, in T-Lymphozyten beschrieben. Eine autosomal rezessive
Erkrankung des Immunsystems, bei der ZAP-70 nicht gebildet wird, gibt Hinweise auf die
Signaltransduktion bei der T-Lymphozytenentwicklung (Perlmutter et al., 1994). Bei ZAP-70
handelt es sich um ein Phosphotyrosin-enthaltendes Protein mit Molekülmasse 70 kD,
welches mit den ζ-Ketten des T-Zell-Antigen-Rezeptor Komplexes während der T-
Zellaktivierung assoziiert. Daher stammt auch der Name: zeta-associated protein-70 (Chan et
al., 1992). ZAP-70 negative Patienten entwickelten zwar eine normale Anzahl an
zirkulierenden Lymphozyten, jedoch führte eine T-Zellrezeptorstimulierung nicht zur

10
Proliferation (Arpaia et al., 1994). Desweiteren war bei Patienten mit einer ZAP-70 Mutation
zu beobachten, daß bei der T-Zellproduktion nur CD4+, aber nicht CD8+ Zellen entstanden,
was auf eine fehlerhafte Reifung der Thymuszellen hinweist.
Syk wird vor allem in hämatopoetischen Zellen exprimiert. Hier wird Syk nach Aktivierung
des B-Zellrezeptors auf Lymphozyten und verschiedener Fc-Rezeptoren auf Lymphozyten
und Monozyten stimuliert. Außerdem ist Syk in der Lymphozytendifferenzierung und in der
Mastzell-Degranulation involviert (Turner et al., 1995). Desweiteren wird Syk nach
Ligandenbindung von β1 Integrinen in Neutrophilen und β2 Integrinen in monozytären Zellen
aktiviert (Lin et al., 1995; Yan et al., 1997).
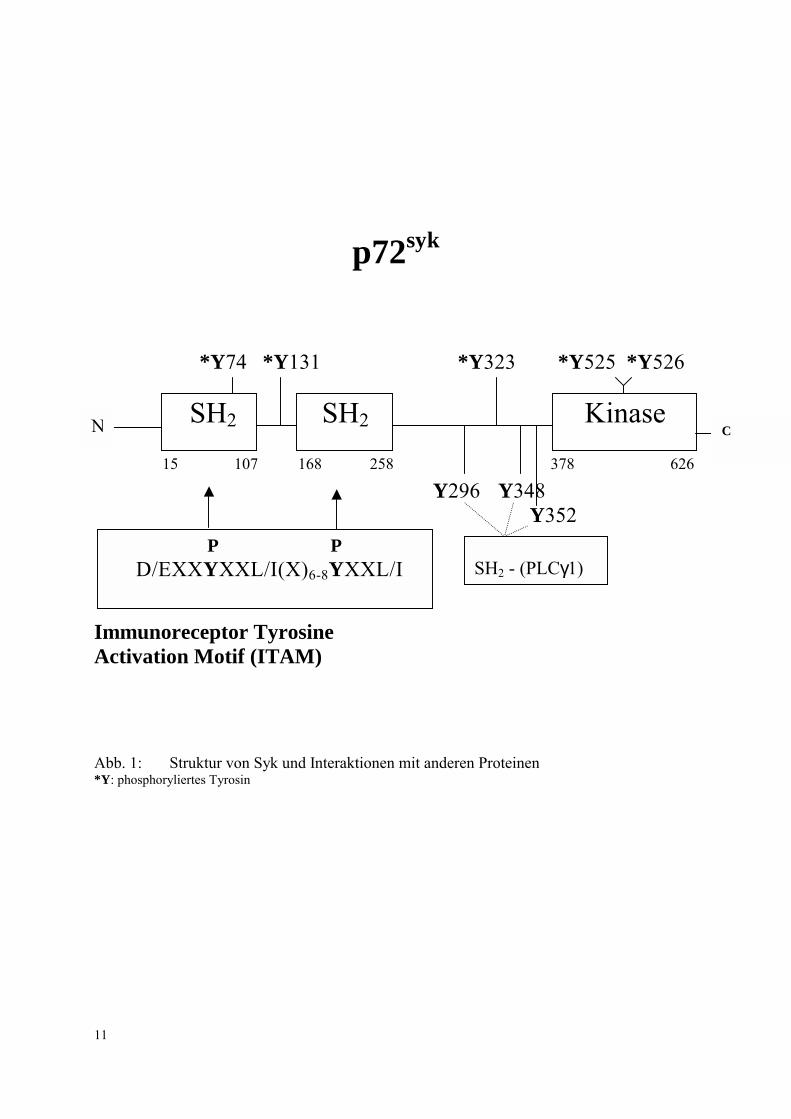
Syk ist eine Proteintyrosinkinase, die tandemartig angeordnet zwei aminoterminale SH2-
Domänen, zwei Spacer-Regionen, die Kinase-Domäne und den C-Terminus enthält (Abb. 1)
(Chan et al., 1992). Die Aktivierung des Proteins Syk erfolgt vermutlich über ein doppelt
phosphoryliertes Immunoreceptor Tyrosine Activation Motiv (ITAM) (Kurosaki et al., 1995),
das einen Bereich von etwa 18 � 20 Aminosäuren umfasst (Rowley et al., 1995). In
Thrombozyten kommen zwei ITAM-enthaltende Proteine vor, der FcγRIIA Rezeptor (Chacko
et al., 1994) und die FcRγ Untereinheit (Gibbins et al., 1996; Asselin et al., 1997), wobei die
letztere bei der Thrombozytenaktivierung durch Kollagen eine wesentliche Rolle spielt
(Asselin et al., 1997; Ichinohe et al., 1997; Poole et al., 1997). Die Assoziation der Syk
Tandem-SH2-Domäne mit tyrosinphosphoryliertem ITAM (s. Abb. 1) stimuliert die Syk-
Autophosphorylierung. Vor allem die hierbei in der katalytischen Domäne erfolgende
Phosphorylierung von Tyrosin 525 und Tyrosin 526 ist für die Aktivierung der Kinase
verantwortlich (Kimura et al., 1996; El-Hillal et al., 1997).
11
p72syk
*Y74 *Y131 *Y323 *Y525 *Y526
15 107 168 258 378 626
Y296 Y348 Y352 Immunoreceptor Tyrosine Activation Motif (ITAM) Abb. 1: Struktur von Syk und Interaktionen mit anderen Proteinen *Y: phosphoryliertes Tyrosin
SH2 SH2 Kinase
P P D/EXXYXXL/I(X)6-8YXXL/I SH2 - (PLCγ1)
C N

12
Phospholipase Cγ1 stellt ein Substrat von Syk dar, dessen SH2-Domäne mit konservierten
Phosphotyrosinresten in der Linkerregion von Syk interagiert (Abb. 1) (Law et al., 1996).
PLCγ1, ein Isoenzym der Phosphoinositide-spezifischen Phospholipase C-Familie, nimmt
eine zentrale Rolle in Signaltransduktionswegen ein. Die Tyrosinphosphorylierung von
PLCγ1 führt zur Aktivierung des Enzyms und zur Bildung von Inosit-1,4,5-Trisphosphat und
1,2-Diacylglycerid, welches Ca2+-Mobilisierung und die Aktivierung der Proteinkinase C
induziert.
Mit dem Yeast Two-Hybrid System konnte die Gruppe um Altman Vav1 (guanine nucleotide
exchange factor, GEF, für die Rho/Rac Familie der GTPasen) als Interaktionspartner
downstream von Syk ermitteln (Deckert et al., 1996). Es gibt drei verschiedene Vav Proteine:
Vav1 wird vor allem im hämatopoetischen System, Vav2 und Vav3 werden ubiquitärer
exprimiert. Vav1 ist ein durch ein Proto-Onkogen kodiertes Protein. Vav1 assoziiert mit Syk
bzw. ZAP-70 in B- und T-Zellen. Es handelt sich hierbei um eine direkte Interaktion, welche
die katalytische Aktivität von Syk erfordert, die SH2-Domäne von Vav1 und spezifische
Tyrosinreste in der Linkerdomäne von Syk. Vav1 wird von Syk-Kinasen in vivo und in vitro
phosphoryliert. Syk und Vav1 induzieren zusammen die Aktivierung von JNK (Jun N-
terminal kinase), ERK2 (extracellular-signal-regulated kinase 2), der Kinase Akt sowie die
Tyrosinphosphorylierung des Oncoproteins Cbl (Miranti et al., 1998). Weiterhin sind die
Proteine der Vav-Familie an Prozessen der Zytoskelett-Reorganisation beteiligt (Hornstein et
al., 2004).
Cbl konnte als negativer Regulator downstream von der Tyrosinkinase Syk identifiziert
werden. Cbl and Syk bilden einen Komplex, in dem die N-terminale Hälfte von Syk mit
seiner Tandem-SH2-Domäne an die N-terminale Domäne von Cbl (AS: 1-655) bindet (Ota et
al., 1997). Die Überexpression von Cbl führte zu einer erniedrigten Tyrosinphosphorylierung
von Syk. Cbl kann somit als endogener Regulator der Signaltransduktion und der
Zellaktivierung dienen.
Sidorenko et al. zeigten, daß Syk und Lyn aus humanen B-Lymphozyten direkt interagieren
können. Syk und Lyn wurden aus reifen und aktivierten B-Zellen coimmunpräzipitiert und
gel-gereinigtes Syk und Lyn reassoziierten in vitro, was ihre direkte Assoziation zeigt
(Sidorenko et al., 1995). Die Lck-Kinase, eine weitere Proteintyrosinkinase der Src-Familie,
konnte via Yeast Two-Hybrid System als Interaktionspartner identifiziert werden (Couture et

13
al., 1996). Die Tyrosinreste Tyr518 und Tyr519 der Autophosphorylierungsstellen von Syk
binden an die SH2-Domäne von Lck während der Thrombozyten-Aggregation.
1.4 Syk-Aktivierung in Thrombozyten
Viele zelluläre Proteine werden während der Thrombozytenaktivierung an Tyrosin
phosphoryliert. Verschiedene Mechanismen regulieren diesen Prozeß. Die Aktivierung von
Thrombozyten über G-Protein gekoppelte Rezeptoren führt zum Formwandel von diskoid zu
sphärisch und der Ausbildung von Pseudopodien, dem sogenannten shape change. Dieser
stellt die früheste Antwort während der Thrombozytenaktivierung dar und kann unabhängig
von einem Anstieg der zytosolischen Ca2+-Konzentration und einer Sekretion des
Granulainhalts erfolgen (Siess et al., 1989; Rasmussen et al., 1993). Während des
Formwandels kommt es zu einer vermehrten Tyrosinphosphorylierung von Proteinen mit der
Molekülmasse von 62 kDa, 68 kDa und 130 kDa. Da durch den Tyrosinkinasehemmer
Genistein die Stimulierung der Tyrosinphosphorylierung dieser Proteine und der Formwandel
inhibiert werden, führte dies zu der Annahme, daß die Aktivierung von Tyrosinkinasen eine
wichtige Rolle für die frühe Phase der Thrombozyten-Aktivierung spielt (Negrescu et al.,
1995).
Nach dem Formwandel kommt es während der physiologischen Thrombozyten-Aktivierung
zur Aggregation. Der Aggregation liegt die Bindung von Fibrinogen an das Integrin αIIbβ3
zugrunde. Integrine sind nicht-kovalente αβ Heterodimere. Jede Untereinheit besteht aus
einer relativ großen NH2-terminalen extrazellulären Domäne, einer einzelnen
Membrandomäne und einer kurzen C-terminalen zytoplasmatischen Domäne. Ruhende
Thrombozyten enthalten etwa 80000 Oberflächenkopien des Integrins αIIbβ3 und einen
zusätzlichen Pool an Integrin αIIbβ3 in den Membranen der α-Granula und dem offenen
kanikulären System (Wagner et al., 1996). Die Aktivierung von Thrombozyten über das
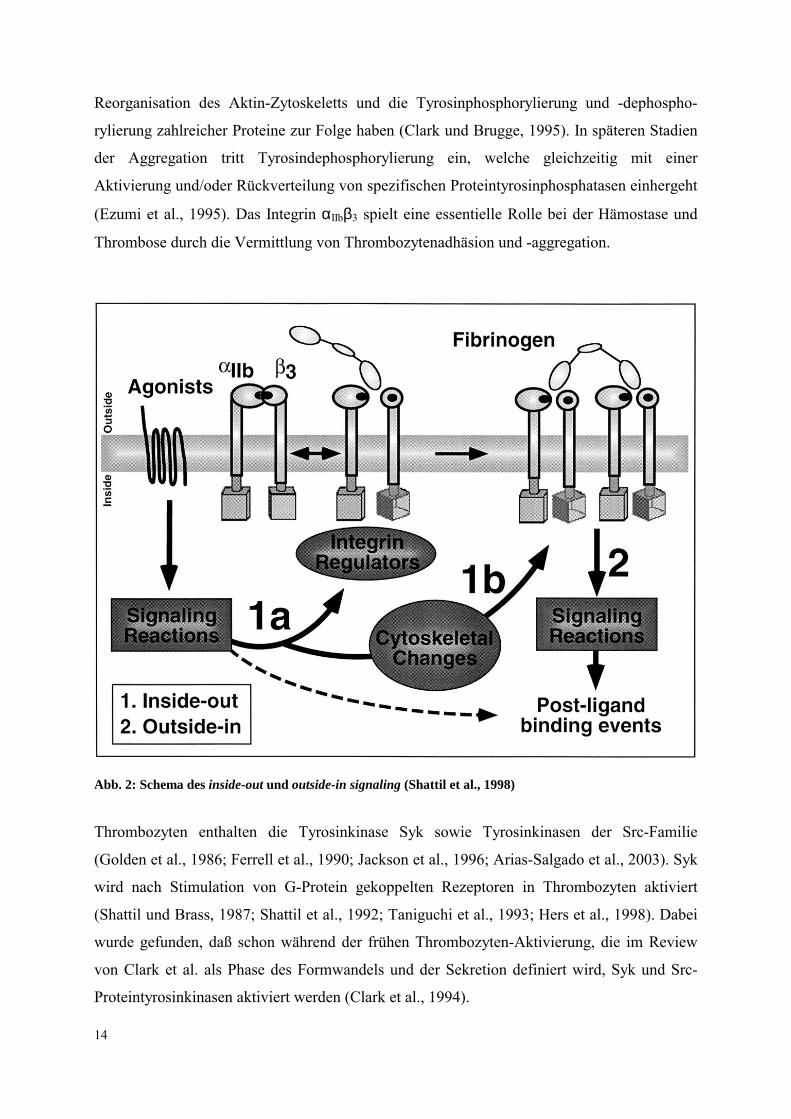
Integrin αIIbβ3 läuft in zwei Phasen ab (s. Abb. 2). Zum einen werden die
Thrombozytenfunktionen durch einen Prozeß reguliert, der als inside-out signaling bekannt
ist. Hierbei erhöhen bzw. erniedrigen Plättchenagonisten bzw. bestimmte Plättchenhemmer
(Prostazyklin) die Affinität des Integrins αIIbβ3 für adhesive Liganden wie Fibrinogen (Shattil
und Ginsberg, 1997; Shattil et al., 1998). Die Bindung von Fibrinogen an das αIIbβ3-Integrin
verursacht ein clustering der αIIbβ3-Integrine und führt zu outside-in Signalen, die die
14
Reorganisation des Aktin-Zytoskeletts und die Tyrosinphosphorylierung und -dephospho-
rylierung zahlreicher Proteine zur Folge haben (Clark und Brugge, 1995). In späteren Stadien
der Aggregation tritt Tyrosindephosphorylierung ein, welche gleichzeitig mit einer
Aktivierung und/oder Rückverteilung von spezifischen Proteintyrosinphosphatasen einhergeht
(Ezumi et al., 1995). Das Integrin αIIbβ3 spielt eine essentielle Rolle bei der Hämostase und
Thrombose durch die Vermittlung von Thrombozytenadhäsion und -aggregation.
Abb. 2: Schema des inside-out und outside-in signaling (Shattil et al., 1998)
Thrombozyten enthalten die Tyrosinkinase Syk sowie Tyrosinkinasen der Src-Familie
(Golden et al., 1986; Ferrell et al., 1990; Jackson et al., 1996; Arias-Salgado et al., 2003). Syk
wird nach Stimulation von G-Protein gekoppelten Rezeptoren in Thrombozyten aktiviert
(Shattil und Brass, 1987; Shattil et al., 1992; Taniguchi et al., 1993; Hers et al., 1998). Dabei
wurde gefunden, daß schon während der frühen Thrombozyten-Aktivierung, die im Review
von Clark et al. als Phase des Formwandels und der Sekretion definiert wird, Syk und Src-
Proteintyrosinkinasen aktiviert werden (Clark et al., 1994).

15
Während der Thrombozyten-Aggregation, ausgelöst durch Aktivatoren von G-Protein
gekoppelten Rezeptoren wie PAF (Rezaul et al., 1994), Thrombin und Thromboxan-Analoga,
wird die Syk-Tyrosinphosphorylierung weiter erhöht (Clark et al., 1994). Untersuchungen von
der Gruppe von Shattil zeigten, daß Syk außer während der Aggregation auch während der
Thrombozytenausbreitung (spreading) vermehrt tyrosinphosphoryliert wird und dies eine
Folge der Aktivierung des Integrins αIIbβ3 durch Fibrinogen ist (Miranti et al., 1998). Die
Behandlung mit Antikörpern, welche die Fibrinogenbindung an das Integrin αIIbβ3 blockiert,
reduzierte die Tyrosinphosphorylierung von Syk um 40% (Clark et al., 1994). Weiterführende
Untersuchungen an transfizierten CHO-Zellen zeigten, daß Syk ein proximales Glied bei der
Integrin αIIbβ3 vermittelten Signaltransduktion darstellt. Dabei sind die zytoplasmatischen
Enden sowohl von der αIIb- als auch der β3-Untereinheit für die Syk-Aktivierung erforderlich
(Gao et al., 1997). Woodside et al. beschreiben eine direkte Bindung von Syk an das
zytoplasmatische Ende des β3-Integrins (Woodside et al., 2001). Außerdem ist Syk an einer
Stimulus-induzierten Aktivierung des Integrins αIIbβ3 in Mausthrombozyten beteiligt (Law et
al., 1999). Allerdings enthalten diese Integrine in ihren zytoplasmatischen Bereichen keine
ITAMs, d.h. es wurde vorgeschlagen, daß die Integrin αIIbβ3-abhängige Syk-Aktivierung
ITAM-unabhängig verläuft (Gao et al., 1997). Außer der Ligandenbindung an das Integrin
αIIbβ3 führt die Dissoziation des Integrins αIIbβ3 durch EGTA zur Tyrosinphosphorylierung
von Syk und zu einem Anstieg der Syk-Kinaseaktivität (Negrescu und Siess, 1996). Diese
Untersuchungen schlagen somit fundamentale Unterschiede in der Aktivierung von Syk durch
das Integrin αIIbβ3 und durch Immunrezeptoren vor.
Außer nach Stimulation von G-Protein gekoppelten Rezeptoren wird Syk nach Kollagen-
Aktivierung von Thrombozyten stark aktiviert. Am Modell der Syk knock-out Maus konnte
von der Gruppe um S.P. Watson eine zentrale Rolle für Syk in der Kollagen-stimulierten
thrombozytären Signaltransduktion gezeigt werden. Bei der Adhäsion von Thrombozyten an
subendotheliales Kollagen wird eine Signalkaskade ausgelöst, die vor allem durch die
Bindung von Kollagen an das Glykoprotein VI (GPVI) auf der Thrombozytenmembran
vermittelt wird. Die Bindung von Kollagen an GPVI führt unter Beteiligung von Src-
Tyrosinkinasen zur Phosphorylierung des ITAM innerhalb der FcRγ-Kette, die eng mit GPVI
assoziiert ist. Das tyrosinphosphorylierte ITAM bindet an die Tandem-SH2-Domäne der
Tyrosinkinase Syk. Die folgende Aktivierung von Syk führt zur Tyrosinphosphorylierung und
Aktivierung der Phospholipase Cγ2 (Gibbins et al., 1996; Watson and Gibbins, 1998; Watson,
1999). Die Frage, welche Tyrosinkinasen ITAM innerhalb der FcRγ-Kette an Tyrosin

16
hosphorylieren, wurde weitgehend geklärt. Es sind dies Tyrosinkinasen der Src-Familie. Src
ist in Thrombozyten stark exprimiert und repräsentiert 0,2 - 0,4% des gesamten
Plättchenproteins (Golden und Brugge, 1989). Vor allem Fyn und Lyn assoziieren in
Kollagen-aktivierten Thrombozyten mit tyrosinphosphorylierten Proteinen, wozu auch die
FcRγ-Kette gehört (Briddon et al., 1999). Auch nach Thrombozyten-Aktivierung mit
Thrombin werden Tyrosinkinasen der Src-Familie (Src, Fyn Yes und Lyn) aktiviert (Clark et
al., 1994). Inwiefern für die Aktivierung der Syk-Tyrosinkinase in Thrombin-aktivierten
Thrombozyten Tyrosinkinasen der Src-Familie eine Rolle spielen, ist unklar. Während also
Fibrinogen über die Bindung an das Integrin αIIbβ3 eine ITAM-unabhängige Syk-Aktivierung
induziert, kommt es nach einer Stimulation mit Kollagen über die Bindung an GPVI zu einer
ITAM-abhängigen Syk-Stimulation.

17
1.5 Fragestellung
In der vorliegenden Arbeit wurde im Bereich der Signaltransduktion in Thrombozyten die
Funktion und Regulation der Tyrosinkinase Syk während Formwandel, Aggregation und
Ausbreitung untersucht.
Als Thrombozytenstimuli wurden vorwiegend mox-LDL und Thrombin bzw. das Heptapeptid
YFLLRNP, das den Thrombinrezeptor PAR-1 partiell aktiviert, verwendet. Die Aktivierung
der Thrombozyten durch mox-LDL ist von pathophysiologischer Bedeutung für die
Atherothrombose bei Patienten mit kardiovaskulären Erkrankungen. Da in der Arbeitsgruppe
gefunden wurde, daß mox-LDL Lysophosphatidsäure (LPA) enthält und Plättchen über die
Aktivierung von LPA-Rezeptoren stimuliert werden, wurde die frühe Aktivierung und
Regulation der Tyrosinkinase Syk auch nach Aktivierung der Thrombozyten durch LPA
untersucht.
Da unklar war, ob Syk während der frühen Phase der Thrombozytenaktivierung, d.h. während
des Formwandels (ohne Sekretion von Inhaltsstoffen aus den Granula) aktiviert wird, wurden
hierzu Untersuchungen durchgeführt. Weiterhin wurden Experimente zur Regulation der Syk-
Aktivierung während des shape change und zur Funktion von Syk für den Formwandel
unternommen.
Außerdem wurde die Funktion und Regulation von Syk während der
Thrombozytenaggregation und -ausbreitung (spreading) untersucht. Hier sollte mit
biochemischen und zellbiologischen Methoden die Signaltransduktionskette ausgehend von
der Aktivierung des Integrins αIIbβ3 bis hin zur Aktivierung von Syk und der Phospholipase
Cγ2 aufgeklärt werden (�Integrin αIIbβ3 outside-in signaling�).

18
2. Material und Methoden
2.1 Material
2.1.1 Geräte
Autoklav (Bioclav, Schütt); Brutschrank (WTCbinder, Tuttlingen); BiofugeA Tischzentrifuge
(Heraeus Sepatech, Osterode); Lumi-Aggregometer, Platelet Ionized Calcium Aggregometer
(Chronolog, NOBIS Labordiagnostica GmbH, Endingen); 2-Kanal-LABOR-Aggregometer
(Fresenius, Bad Homburg); Lichtmikroskop D65323 (Leitz, Wetzlar); Multiphor II LKB
Novablot (Pharmacia Biotech, Freiburg); Omnifuge 2. ORS (Heraeus Instruments GmbH,
Biometra, Göttingen); Protean II xi Cell (Bio-Rad, München); Speed Vac Univapo 150 H
(Uni Equip, Martinsried); Spektralphotometer Uvikon 930 (Kontron Instruments);
Thermocycler UNO-Thermoblock (Biometra); Thermomixer 5436 (Eppendorf, Hamburg);
Tischzentrifuge MR 1812 (Jouan GmbH, Unterhaching).
2.1.2 Chemikalien, Enzyme, Antikörper, Fluoreszenzfarbstoffe, Reagenzien
Trypton und Agar für Bakteriennährmedien wurden von Difco (Detroit), Hefeextrakt von
Merck (Darmstadt) bezogen. Medien und anderweitige Reagenzien stammten von den Firmen
Gibco/BRL (Eggenstein), PharMingen (San Diego), Seromed (Berlin) und Dianova
(Hamburg).
Soweit nicht im Text anders erwähnt, wurden alle weiteren Chemikalien in p.a.-Qualität von
den Firmen Merck (Darmstadt), Sigma (Deisenhofen), Serva (Heidelberg) oder ICN
(Eschwege) bezogen.
Es wurde mit Nucleinsäure-modifizierenden Enzymen und Restriktionsenzymen mit den
dazugehörigen Inkubationspuffern sowie Proteaseinhibitoren von Boehringer Mannheim,
Pharmacia oder ICN gearbeitet.

19
Desweiteren wurden folgende Antikörper eingesetzt:
Anti-β3 monoklonal, cat # I19620 (Transduction Laboratories)
Anti-p125Fak/Protein A-Agarose monoklonal, cat # 16-109 (UBI, Lake Placid, NY/USA)
Anti-γ-subunit (TCR, Fc Receptors) polyklonal, cat # 06-727 (UBI, Lake Placid, NY/USA)
Anti-Phosphotyrosine PY20 monoklonal, cat # 05-321 (UBI, Lake Placid, NY/USA)
Anti-Phosphotyrosine 4G10 monoklonal (UBI, Lake Placid, NY/USA)
Anti-PLCγ2 (C-19) polyklonal, cat # 406 (Santa Cruz Biotechnology Inc., CA/USA)
Anti-Syk (4D10) monoklonal, cat # sc-1240 (Santa Cruz Biotechnology Inc., CA/USA)
Anti-Syk (LR, frei oder als Agarose-Konjugat) polyklonal, cat # sc-573 (Santa Cruz
Biotechnology Inc., CA/USA)
Anti-Syk (C-20) polyklonal, cat # sc-929 (Santa Cruz Biotechnology Inc., CA/USA)
Anti-Vinculin, cat # 63-780 (ICN Biomedicals, Inc., Ohio/USA)
Anti-Maus IgG, an HRP gebunden (Amersham)
Anti-Kaninchen IgG, an HRP gebunden (Amersham)
Kontroll-Kaninchen IgG, kovalent an Agarose gebunden (Sigma, St. Louis, MO/USA)
Fluoreszenzfarbstoffe:
FITC anti-Maus (New Alexa� 488, A-11001, Molecular Probes)
Rhodamine phalloidin (Molecular Probes Inc., Eugene, OR/USA)
TRITC anti-Maus (New Alexa� 568, A-11004, Molecular Probes)
TRITC anti-Kaninchen (New Alexa� 568, A-11011, Molecular Probes)
Weitere Substanzen:
Apyrase (Sigma A-6535, Deisenhofen)
BAPTA-AM (5,5-Dimethyl-bis-(o-aminophenoxy)ethan-N,N,N´,N´-tetra-(acetoxymethyl)-
ester) (Molecular Probes Inc., Eugene, OR/USA)
Kollagen (Horn-Chemie, München)
FR-1 (Novabiochem)
Genistein (Calbiochem)
Oleoyl-LPA (Sigma)
PD173956 (Parke Davis, Geschenk von Dr. A.J. Kraker, MI/USA)
Piceatannol (Biomol, USA)

20
PP1 (Calbiochem-Novabiochem Corp., San Diego, CA/USA)
RGDS (Sigma)
Rho-Kinase Hemmer Y-27632 (Geschenk von Dr. A. Yoshimura, Yoshitomi Pharmaceutical
Industries, Saitama, Japan)
Ro-31-8220 (Roche; Geschenk von Dr. P. Presek, Pharmakologisches Institut, Universität
Gießen)
Thrombin human (Boehringer Mannheim)
Tyrphostin A1 (Calbiochem)
Tyrphostin A23 (Calbiochem)
Tyrphostin A47 (Calbiochem)
YFLLRNP (Bachem Ltd., Bubendorf, Schweiz)
2.1.3 Bakterien- und Hefestämme, Vektoren, cDNA-Banken
Bakterien-Stämme:
E.coli DH5αTM Kompetente Zellen (Gibco BRL, Life Technologies)
Genotyp: F-, φ80dlacZ∆M15 ∆(lacZYA-argF)U169 deoR recA1 endA1 hsdR17 (rK-,mK
+)
phoA supE44 λ- thi-1 gyrA96 relA1
E.coli Max-EfficiencyTM HB101Kompetente Zellen (Gibco BRL, Life Technologies)
Genotyp: F- mcrB mrr hsdS20 (rB-, mB
-) recA13 supE44 ara14 galK2 lacY1 proA2
rpsL20(Smr) xy15λ-leu mtl1
KC8 Chemisch Kompetente Zellen (Clontech)
DH5α/pAS2-1
DH5α/pAS2-1/p72 (Syk full-length)
DH5α/pAS2-1/Syk kinase dead
DH5α/pGBT9
DH5α/pGBT9/3a

21
DH5α/pGBT9/Syk full-length
DH5α/pGBT9/Syk kinase dead
28F4/pACT
Kompetente Zellen E.coli DH5αTM und Max-EfficiencyTM HB101 wurden von Gibco-BRL
Life Technologies, USA bezogen. Für Dauerkulturen wurden Bakterien bzw. Hefestämme mit
15 % final sterilem Glycerin versetzt, gut vermischt und schnell bei �70°C eingefroren.
Hefe-Stämme:
Name Genotyp Referenz
S.c. Y190 MATa ade2-101 gal4-∆ gal80-∆ his3 leu2-3, 112 trp1-∆901
ura3-52
LYS2::GAL1-HIS3
URA3::GAL1-lacZ
(Durfee et al.,
1993)
Y190/pAS2-1
Y190/pAS2-1/Syk full-length
Y190/pAS2-1/Syk kinase dead
Y190/pAS2-1/Syk full-length, pGAD10/Knochenmarks-cDNA-Bank; clone 26
Y190/pGBT9
Y190/pGBT9/2b
Y190/pGBT9/3a
Y190/pGBT9/Syk full-length
Y190/pGBT9/Syk kinase dead
Y190/pGBT9/GP3a/Endonexin, (+)-Kontrolle für β-Galaktosidasetest
S.c. CG-1945

22
Vektoren:
PAcG2T Baculovirus Transfer Vektor
pAS2-1: DNA-BD Vektor (DNA-bindende Domäne; bait-Plasmid)
pAS2-1/Syk full-length
pAS2-1/Syk kinase dead
pUC 9 DNA
clone 28 (Endonexin)
pGBT9
pGBT9/3a
pGBT9/2b
pGBT9/Syk full-length
pGBT9/Syk kinase dead
cDNA-Banken:
(in E.coli DH5αTM)
DH5α/pGAD10/Knochenmarks-cDNA-Bank (pGAD10: Vektor mit DNA-aktivierender
Domäne, DNA-AD, prey-Plasmid)
DH5α/pGAD10/Fötale Leberzellen-cDNA-Bank

23
2.2 Methoden
2.2.1 Isolation von humanen Thrombozyten, Maus-Thrombozyten und
Maus-Megakaryozyten
Isolation von humanen Thrombozyten
Die Blutentnahme erfolgte aus der Kubitalvene von gesunden Probanden (Alter: 20-50 Jahre)
in 1/10 Volumen Natriumcitratlösung (3,13%, Fa. Fresenius, Bad Homburg). Das Blut wurde
bei Raumtemperatur 20 min bei 180xg zentrifugiert und anschließend das Plättchen-reiche
Plasma (PRP) abpipettiert. Nach Inkubation des PRP mit 1 mM Acetylsalicylsäure und 0,3
U/ml Apyrase für 15 min bei 37°C, wurden 9 mM Citronensäure und 5 mM EDTA (pH 7,4)
zugegeben und bei 800xg (20 min, RT) zentrifugiert.
Das Thrombozytenpellet wurde anschließend in Puffer B (10% des Vollblut-Volumens) mit
0,6 U/ml Apyrase resuspendiert.
Puffer B:
20 mM Hepes, 138 mM NaCl, 2,9 mM KCl, 1 mM MgCl2, 0,36 mM NaH2PO4, 1 mM EGTA,
pH 6,2
Vor dem erneuten Zentrifugationsschritt bei 800xg, 20 min, RT wurde eine Probe mit
THROMBO PLUS Spezialzähllösung (Fa. Sarstedt, Nümbrecht) verdünnt und die
Thrombozyten-Zahl in der Neubauer-Zählkammer bestimmt.
Das Zellpellet wurde schließlich in entsprechendem Volumen Puffer C mit 0,6 U/ml Apyrase
und 5 mM Glucose aufgenommen. Hierbei ergeben 200 ml Blut etwa 10 ml
Thrombozytensuspension mit einer Konzentration von 800000 Zellen/µl.
Puffer C:
20 mM Hepes, 138 mM NaCl, 2,9 mM KCl, 1 mM MgCl2, 0,36 mM NaH2PO4, pH 7,4

24
Isolation von Maus-Thrombozyten und -Megakaryozyten
Maus-Thrombozyten
Die Präparation der Maus-Thrombozyten erfolgte am Department of Pharmacology,
University of Oxford, UK (Dr. S.P. Watson) und wurde folgendermaßen durchgeführt:
1. Blutabnahme (etwa 1 ml Blut) im Verhältnis 1:10 in ACD (Acid-Citrate-Dextrose:
Na-Citrat 25 g/l, Glucose 20 g/l, Citronensäure 15 g/l) durch Herzpunktion.
2. Abzentrifugieren des PRP (platelet rich plasma) bei 120xg, 10 min, RT.
3. PRP abpipettieren; Zugabe von 0,3 U/ml Apyrase (und je nach Versuchsanordnung
evtl. 1 mM Aspirin), Inkubation für 15 min bei 37°C.
4. Zugabe von 9 mM Citronensäure und 5 mM EDTA, pH 7,4; Zentrifugation bei
1000xg für 10min bei RT.
5. Zugabe von Puffer B (Volumen: 10% des Blutvolumens), der 3 U/ml Apyrase enthält
und Resuspension.
6. Zentrifugation bei 1000xg für 10 min bei RT.
7. Zugabe von vorgewärmtem Puffer C (37°C), der 0,6 U/ml Apyrase und 5 mM
Glucose enthält; Zellzahlzählung mittels Zählkammer oder Zell-Counter und
Einstellung der finalen Thrombozytenkonzentration.
8. Thrombozyten vor Versuchsbeginn 30 min ruhen lassen.
Maus-Megakaryozyten
Ebenso wurden Maus-Megakaryozyten am Department of Pharmacology, University of
Oxford untersucht.
Hank´s Puffer:
143 mM NaCl, 5,6 mM KCl, 10 mM Hepes
Am Versuchstag wurden dem Hank´s Puffer 10 mM final Glucose zugegeben. Nach 20 � 30
min O2-Begasung wurden 0,4% BSA zugefügt und der pH-Wert auf 7,22 eingestellt. Pro
Maus wurden zwei 1 ml Gefäße mit Hank´s Puffer und 0,2 U/ml Apyrase final vorbereitet.

25
Dem verbleibenden Puffer wurden 2 mM MgCl2 und 0,2 mM CaCl2 zugegeben.
Die Mäuse (4-8 Wochen alt) wurden mittels cervikaler Dislokation getötet und die
Femoralknochen extrahiert, wobei das Knochenmark vorsichtig durch eine Spritzennadel (23)
in Hank´s Puffer gewaschen wurde. Anschließend wurde Prostazyklin (1 µg/ml) zugesetzt
und die Zellen nach einem 8-minütigen Zentrifugationsschritt bei 500 rpm in frischem Puffer
resuspendiert.
2.2.2 Messung des shape change und der Aggregation
Aliquots der Thrombozytensuspension (Thrombozytenkonzentration: i.d.R. 800000/µl;
Volumen: 0,5 - 1 ml) wurden in Aggregometerküvetten überführt und bei 37°C mit oder ohne
entsprechende Hemmsubstanzen vorinkubiert (Zeit: 5 - 20 min). Nach Zugabe verschiedener
Thrombozyten-Stimuli (mox-LDL, dem partiellen Thrombinrezeptor-Agonisten YFLLRNP,
Thrombin, Kollagen) wurden der shape change bzw. die Aggregation der Thrombozyten in
einem 2-Kanal-LABOR-Aggregometer oder in einem Lumi-Aggregometer gemessen.
Desweiteren erfolgte die Abnahme entsprechender Proben und Überführung in vorgelegten 2x
Laemmli-Puffer (s. 2.2.7) zur Bestimmung der Proteintyrosinphosphorylierung in den
Thrombozyten, bzw. nach Überführung in Lysis-Puffer (s. 2.2.4) zur Immunpräzipitation und
Bestimmung der Tyrosinphosphorylierung der Proteine Syk, FAK und PLCγ2.
2.2.3 Milde Oxidation von LDL
Die LDL-Präparation erfolgte entsprechend dem Protokoll von (Schulz et al., 1995) und
(Weidtmann et al., 1995). Hierbei wurde von 400 ml Blut ausgegangen. LDL (0,5 - 5 mg/ml)
wurde in einem N2-gesättigten Puffer (pH 7,4) dialysiert, welcher NaCl (150 mM),
Chloramphenicol (0,15 mM) und EDTA (1 mM) enthielt und bei 4°C unter Lichtabschluß und
N2 gelagert wurde. Vor der Oxidation wurde EDTA aus dem LDL-Puffer entfernt. Dies
erfolgte über Entsalzungssäulen (Econo-Pac 10 DG, Bio-Rad Laboratories GmbH, München).
Mit Hilfe von Konzentratoren (Centricon 100, Fa. Amicon, Beverly, MA, USA) wurde auf
eine Proteinkonzentration von 10-20 mg/ml in PBS eingeengt. Der Proteingehalt wurde nach
einer modifizierten Lowry Methode mit Rinderserumalbumin als Standard bestimmt

26
(Markwell et al., 1978). Die Oxidation des LDL (etwa 0,3 - 0,5 ml pro Eppendorff-Cup)
erfolgte durch Zugabe von 640 µM CuSO4 bei 37°C unter leichtem Schütteln (niedrigste
Stufe) für 18 - 20 h in einem Eppendorff Thermomixer. Das Eppendorff-Cup blieb hierbei
geöffnet und wurde mit Parafilm, welcher mit einer Nadel perforiert worden war, abgedeckt.
2.2.4 Immunpräzipitation von Proteinen aus Thrombozyten-Lysaten (Syk,
FAK, PLCγγγγ2)
Die Thrombozytensuspension (0,6 ml; 800000 Zellen/µl) in der Aggregationsküvette wurde
vor oder nach Stimulation mit 0,6 ml 2x eiskaltem Lysis-Puffer A versetzt und für 30 min auf
Eis lysiert.
Lysis-Puffer A 2x:
2% Triton X-100, 2 mM EGTA, 100 mM Hepes, 150 mM NaCl, pH 7,4
vor Gebrauch frisch zugeben: Proteasehemmercocktail �Complete Mini EDTA-free�
(Boehringer Mannheim, #1836153, 1 Tbl. in 5 ml), 2 mM PMSF, 2 mM Na3VO4
Die folgenden Schritte wurden entweder im Kühlraum durchgeführt oder die Proben auf Eis
gelagert. Die Lysate wurden durch Zentrifugation bei 13.000xg für 10 min bei 4°C von
unlöslichen Bestandteilen getrennt. Der Überstand (1,2 ml) wurde mit den entsprechenden an
Agarose gekoppelten Antikörper-Konjugaten bei 4°C für 2 h oder 12 h inkubiert. Alternativ
erfolgte bei einigen Experimenten eine Inkubation mit dem Antikörper für 1 h bei 4°C,
gefolgt von einer einstündigen Inkubation mit CL4B Protein A-Sepharose (2,5 mg = 50 µl)
bei 4°C. Bei letzterer Methode wurde ein Preclear-Schritt mit Kaninchen IgG (1 µg) und
CL4B Protein A-Sepharose vorgeschaltet.
Verwendete Antikörpermengen für die Immunpräzipitation:
*): an Agarose gekoppeltes Antikörper-Konjugat
Syk-IP FAK-IP PLCγ-2-IP
1. 2.*) *)
15 µg 10 µg 20 µg 1 µg
Die Immunpräzipitate wurden durch Zentrifugation bei 13.000xg für 10 min bei 4°C
abgetrennt. Dann erfolgten 3 Waschschritte mit 1 ml kalten 1x Lysis-Puffer A, 1 x Waschen

27
mit kaltem 10 mM HEPES, 500 mM NaCl (pH 8,0) und 1 x Waschen mit kaltem 10 mM
HEPES (pH 8,0). Schließlich wurden die Immunpräzipitate für 3 min bei 95°C in 100 µl 2x
Laemmli-Puffer gekocht, bevor die Proben gelelektrophoretisch aufgetrennt und über Western
Blot nachgewiesen wurden.
Ein zu einem späteren Zeitpunkt durchgeführter Vergleich verschiedener Lysis-Puffer für die
Syk-Immunpräzipitation zeigte sehr gute Resultate bei der Verwendung des Lysis-Puffers C,
der für die Pull-down Assays eingesetzt wurde (s. 2.2.6), wobei das Detergens NP-40 gegen
Triton X-100 ausgetauscht wurde.
2.2.5 Isolierung des Thrombozyten-Aktinzytoskeletts
Die Zytoskelettfraktionen wurden nach einer Modifikation der Methode von (Jennings et al.,
1981) präpariert. Thrombozytensuspensionen (0,6 ml) wurden vor oder zu verschiedenen
Zeitpunkten nach Stimulation durch Zugabe gleicher Volumina eiskaltem 2x Triton Lysis-
Puffer B (pH 7,5) lysiert.
Triton Lysis-Puffer B 2x (pH 7,5):
100 mM Tris-HCl, 2% Triton X-100, 10 mM EGTA, 10 mM EDTA, 2 mM Na3VO4, 20
µg/ml Leupeptin, 2 mM PMSF, 25 µM Pepstatin A und 20 µg/ml Aprotinin.
Die Proben (1,2 ml) werden für 30 min auf Eis gestellt und anschließend bei 15.600xg für 15
min bei 4°C zentrifugiert. Die Pellets wurden einmal mit Triton X-100 (1%) Lysis-Puffer B
gewaschen und in 100 µl Gelelektrophorese-Auftragspuffer resuspendiert.
2.2.6 Pull-down Assay aus Thrombozytenlysaten mit der Tandem-SH2-
Domäne von Syk
Ein GST-Fusionsprotein, welches die Tandem-SH2-Domäne von Syk (Geschenk von Dr. S.P.
Watson, Institute of Pharmacology, Oxford) enthielt, wurde hierfür an Glutathione
Sepharose 4B (Pharmacia Biotech, Uppsala, Schweden) gebunden. Die Thrombozyten-
Suspension (0,35 ml) wurde mit gleicher Menge Lysis-Puffer C versetzt:

28
Lysis-Puffer C (Pull-down Assay) 2x:
2% Nonidet P-40, 2 mM EGTA, 2 mM EDTA, 300 mM NaCl, 20 mM Tris, pH 7,5
vor Gebrauch frisch zugeben: Complete Mini EDTA-free (Boehringer Mannheim, #1836153,
1 Tbl. in 5 ml), 5 mM Na3VO4, 8 mM Pefablock
Zunächst wurde ein Preclear-Schritt mit 5 µg GST-Sepharose für 0,7 ml Thrombozyten-Lysat
für 1 h bei 4°C durchgeführt. Nach einer anschließenden Zentrifugation (13.000 rpm, 5 min)
wurde der Überstand mit je 10 µg Fusionsprotein versetzt und 2,5 h bei 4°C inkubiert. Das
Pellet wurde mit 1 ml 1x Lysis-Puffer C (+Protease-, Phosphatasehemmer) und 4x mit 1 ml
TBS-T gewaschen. Nach den Waschschritten wurde wie in 2.2.4 beschrieben weiterverfahren.
2.2.7 Messung der Proteintyrosinphosphorylierung mittels Western Blot
Das Western Blot-Verfahren verbindet die gelelektrophoretische Analyse mit der Spezifität
der immunologischen Reaktion.
2.2.7.1 Diskontinuierliche SDS-Gelelektrophorese in Polyacrylamidgelen
Puffer:
Elektrodenpuffer: 0,6% (w/v) Tris-Base, 0,1% (w/v) SDS, 2,88% (w/v) Glycerin,
pH 8,3 � 8,4
Laemmli-Puffer: 250 mM Tris/HCl, pH 6,8, 20% (w/v) SDS, 87% (w/v) Glycerin, 1% (w/v)
Bromphenolblau
Elektrophoresen wurden in 8 � 12%igen Polyacrylamidgelen (200 mm x 200 mm x 1 mm)
bzw. 5 � 20% Gradientengelen durchgeführt. Die Auftrennung erfolgte in einer
Gelelektrophoreseapparatur (Protean II xi Cell, Bio-Rad) mit dem von (Laemmli, 1970)
beschriebenen diskontinuierlichen Puffersystem. Außerdem wurden auch mit Tricin-Gelen
nach Schägger für die Auftrennung niedermolekularer Peptide (1-15 kDa) gearbeitet
(Schägger und v. Jagow, 1987). Die Herstellung der Gele (20 x 20 cm) und die Durchführung

29
der Elektrophoresen (vertikal) erfolgte wie in genannter Literatur beschrieben.
Der Probenpuffer wurde mit Mercaptoethanol oder DTT versetzt, um die Proteine zu
reduzieren. Die Proben wurden 1:1 mit Probenpuffer versetzt und vor dem Auftragen auf das
Gel 5 min bei 95°C im Thermomixer erhitzt. Zur Ermittlung der Molekülmassen der Proteine
wurden käufliche Molekulargewichtsmarker (Full Range RainbowTM RPN 800, Amersham
LIFE SCIENCE, Buckinghamshire, UK) mit auf das Gel aufgetragen. Die Proteinauftrennung
erfolgte bei 10 mA über Nacht unter kontinuierlicher Kühlung, gefolgt von einer einstündigen
Fokussierung bei 35 mA.
2.2.7.2 Proteintransfer aus SDS-Polyacrylamidgelen auf Nitrocellulose-
Membranen
Anodenpuffer 1:
0,3 M Tris, 10% Methanol, pH 10,4
Anodenpuffer 2:
25 mM Tris, 10% Methanol, pH 10,4
Kathodenpuffer:
25 mM Tris base, 40 mM 6-Amino-n-Capronsäure, 10% Methanol, pH 9,4
Der Proteintransfer wurde mit einer Semidry-Blot-Apparatur (LKB 2117 Multiphor II,
semidry) mit Graphitelektroden durchgeführt. Für den Transfer wurde folgender Sandwich-
Aufbau verwendet:
• 4 Lagen 3 MM Chromatographie-Papier (Munktell-Filterpapier, Pharmacia, Freiburg) in
Anodenpuffer 1 equilibriert
• 4 Lagen in Anodenpuffer 2
• Nitrocellulose (Hybond C-Super, Supported Pure Nitrocellulose, 0,45 µm, Amersham)
• Polyacrylamidgel
• 7 Lagen in Kathodenpuffer
Blotbedingungen: 1 mA/cm² für 2 h bei RT.

30
2.2.7.3 Immunologischer Nachweis der Proteine
10x TBS-T:
1,5 M NaCl, 0,25% (v/v) Tween 20, 100 mM Tris-Base, pH 7,6
Blocklösung:
5% (w/v) Magermilchpulver (Bio-Rad, Hercules, CA, USA) in 1x TBS-T
Vor der Antikörpernachweisreaktion wurden freie Bindungsstellen auf der Membran mit der
Magermilchlösung (40 ml für 12,5 cm x 16 cm Membran) für 1 h bei RT blockiert. Nach
mehreren Waschschritten mit TBS-T (2x kurz, 1x 15 min, 2x 5 min) erfolgte die Inkubation
mit dem 1. Antikörper für 1 h bei RT. Nach erneutem Waschen wurde die Reaktion mit dem
entsprechenden 2. Antikörper (anti-mouse Ig (m) bzw. anti-rabbit Ig (r), HRP linked whole
antibody, Fa. Amersham) durchgeführt.
Verwendete Antikörperkonzentrationen (Verdünnungen):
1. AK PY20
1:2000
4G10(PY)
1:2000
Syk
1:1000
FAK
1:2000
FcRγ-Kette
1:1000
PLCγ-2
1:1000
2. AK 1:15000 (m) 1:15000 (m) 1:2500 (r) 1:15000 (m) 1:2500 (r) 1:2500 (r)
Danach wurde nochmals gewaschen und anschließend die Chemilumineszenzreaktion
(SuperSignal Chemiluminescent Substrate Luminol/Enhancer, Fa. Pierce, Rockford, IL,
USA) entsprechend der Angaben des Herstellers durchgeführt. Emittiertes Licht der
Wellenlänge 466 nm wurde mit geeigneten Filmen (HyperfilmTMECLTM, Fa. Amersham)
detektiert.
Bei den Anti-Phosphotyrosinimmunoblots von Syk, FAK und PLCγ2 Immunpräzipitaten
wurden mittels stripping (2% SDS, 62,5 mM Tris-HCl, pH 6,8, 100 mM Mercaptoethanol,
Inkubation 10 min bei 50°C) die Anti-Phosphotyrosin-Antikörper von der Membran entfernt.
Anschließend wurde der Blot mit spezifischen Antikörpern inkubiert, mit dem Ziel, die
Menge der immunpräzipitierten Proteine densitometrisch zu quantifizieren.

31
2.2.7.4 Proteinfärbung mit Ponceau S
Mit dem roten Farbstoff Ponceau S (Sigma Diagnostics, St. Louis, MO, USA) können die
Proteine nach Transfer auf Nitrocellulose visualisiert werden.
Die Membran wird hierfür für etwa 10 min mit verdünnter Lösung Ponceau S inkubiert.
Anschließend wird die Membran mit PBS mehrmals gewaschen, bis das Proteinbandenmuster
gut zu erkennen ist. Nach Ergebnisdokumentation über Scannen oder Kopieren kann die
Membran nach erneuten PBS-Waschschritten mit der Magermilchlösung behandelt werden
(2.2.7.3).
2.2.8 Densitometrische Auswertung des Western Blots
Die quantitative Auswertung der Schwärzung der Banden auf dem Film erfolgte
densitometrisch mit der Software GelScanTMXL von Pharmacia LKB Biotechnology (Scanner
SHARP JX-325, Pharmacia Biotech). Gemessen wurde die Absorption der spezifischen
gesamten Bande in Bezug auf einen Hintergrund-Parameter. Die Absorption pro
Flächeneinheit wurde nach Integration in Absorptionseinheiten x mm2 angegeben.
2.2.9 Transiente Transfektion von CHO-Zellen
Diese Untersuchungen wurden von mir in der Arbeitsgruppe von Dr. S.J. Shattil (Department
of Vascular Biology, Scripps Research Institute, La Jolla, USA) durchgeführt. Die mit αIIbβ3-
Integrin stabil transfizierten CHO-Zellen wurden auf eine 100 mm Petrischale aufgebracht, so
daß sie am Tag der Transfektion mit dem Syk-Plasmid zu 40-60% konfluent waren. Am Tag
der Transfektion wurden die Zellen mit Dulbecco�s Modified Eagle Medium (DMEM)
gewaschen, um fötales Rinderserum aus dem Kulturmedium zu entfernen. Die Syk-Plasmid-
DNA wurde via Maxi-prep (Qiagen, Santa Clarita, CA) gereinigt. Eine Gesamtmenge von 4
µg Plasmid-DNA wurde zu 200 µl DMEM gegeben und mit 20 µl Lipofectamin-Lösung für
15 min in Polystyrene-Röhrchen inkubiert. LipofectinTM (GIBCO BRL, Gaithersburg, MD,

32
USA), ein kationisches Lipid, geht spontan Wechselwirkungen mit der rekombinierten DNA
ein und bildet einen Lipid-DNA-Komplex. Hierdurch wird die Aufnahme der DNA in die
Zellen ermöglicht. Anschließend wurde die Mischung mit 3,8 ml DMEM verdünnt und die
Zellen wurden damit überschichtet.
Nach einer Inkubation von 6 h bei 37°C, 5% CO2 erfolgte ein Mediumswechsel zu DMEM
mit 10% FCS, 1% nichtessentiellen Aminosäuren, 2 mM L-Glutamine, 100 U/ml Penicillin
und 100 µg/ml Streptomycin (komplettes Wachstumsmedium). Einen Tag später wurde noch
einmal frisches komplettes Wachstumsmedium auf die Zellen gegeben, bevor diese dann
wiederum einen Tag später für das jeweilige Experiment eingesetzt wurden.
2.2.10 Untersuchungen mittels konfokaler Laserfluoreszenzmikroskopie an
ausgebreiteten humanen und Mäuse-Thrombozyten, und CHO-Zellen
Die Immunfluoreszenz dient dazu, die Lokalisation von Proteinen innerhalb der Zelle zu
untersuchen. Spezifische Antikörper gegen das zu untersuchende Protein (Antigen) werden
durch die Bindung an Zweit-Antikörper, die mit einem Fluoreszenzfarbstoff markiert sind,
sichtbar gemacht. Das Antigen in der Probe muß hierbei in eine unlösliche Form gebracht
werden. Dazu dient eine chemische Vernetzung von Proteinen oder eine Präzipitation durch
organische Lösungsmittel. Die Permeabilisierung mit Hilfe von Detergenzien oder
organischen Lösungsmitteln macht ein Eindringen der Antikörper in die Probe möglich. Die
meisten Filter für die Immunfluoreszenz sind auf die Standardfluorophore Fluorescein und
Rhodamin abgestimmt.
Physikalische Daten der Fluorophore
Fluorophor Fluorescein Rhodamin
Excitation 495 nm 552 nm
Emission 525 nm 570 nm
Farbe grün rot
Verwendete Materialien und Reagenzien:
• Glasobjektträger und Deckgläschen
• PBS:

33
KH2PO4 6 mM, Na2HPO4 16 mM, NaCl 140 mM, pH 7,4
• Fibrinogen (Sigma) bzw. Polylysin (Sigma)
• Coating Puffer:
NaCl 150 mM, Na2HPO4 50 mM, NaH2PO4 50 mM, pH 8,0
• Rinderserumalbumin (BSA)
• 3,7% Formaldehyd/PBS (methanol-freies Formaldehyd, Polysciences)
• 0,2% Triton X-100/PBS
• 10% steriles Normal-Goat-Serum/PBS
• Primärantikörper, Rhodamin-Phalloidin (zur Anfärbung)
• Fluorescein (FITC)- bzw. Rhodamin (TRITC)-konjugierte Zweitantikörper (Anti-Maus,
Anti-Kaninchen) (Alexa Dyes, Molecular Probes)
• Mounting Medium (Immuno-Fluore Mounting Medium, ICN Biochemicals)
• Konfokales Laserfluoreszenzmikroskop Leika DM IRB, Leika software TCSNT version
1.5.451
Methode
1. Coverslips mit Seife für 5 min waschen, anschließend 5 Waschschritte (5x) mit H20.
2. Coverslips mit 20 µg/ml Fibrinogen (bzw. Polylysin) in Coating-Puffer bedecken und
für 2 Stunden bei 37°C oder über Nacht bei 4°C inkubieren.
3. 1x mit PBS waschen.
4. Mit 20 mg/ml BSA für 2 Stunden bei Raumtemperatur (RT) blocken.
5. 2x mit 2 ml PBS waschen.
6. 500 µl Probe auf die Fibrinogen-gecoateten Deckgläser geben (Thrombozyten: 2 x 108
Zellen/ml; CHO-Zellen: 2 x 105 Zellen/ml) und für 60 min bei 37°C inkubieren. Für
Thrombozyten wurde alternativ auch spreading auf Polylysin-gecoateten Coverslips
durchgeführt.
7. 2x mit PBS waschen und mit 3,7 % Formaldehyd/PBS, 10 min, RT fixieren.
8. 2x mit PBS waschen. (Bei diesem Schritt kann das Experiment unterbrochen werden;
PBS dazugeben und bei 4°C lagern.)
9. Fixierte Zellen mit 0,2% Triton X-100/PBS für 5 min, RT, permeabilisieren. 2x mit

34
PBS waschen.
10. Unspezifische Bindungsstellen mit 10% sterilem Normal-Goat-Serum/PBS für 30 min
bei RT blockieren.
11. Primärantikörper mit Blocklösung verdünnen (1:100) und vor Gebrauch bei 14000
rpm für 10 min zentrifugieren. Die Coverslips werden 45 min bei 37°C auf 100 µl
Antikörperlösung, die auf Parafilm vorgelegt werden, inkubiert. Für die Aktinfärbung
wird Rhodamin-Phalloidin in einer Verdünnung von 1:25 eingesetzt.
12. 5x für 2 min in PBS waschen.
13. Sekundärantikörper in Blocklösung verdünnen (1:200). Bei 37°C für 45 min
inkubieren. 5x waschen.
14. Deckgläser 1x mit H2O waschen.
15. Einen kleinen Tropfen Mounting Medium (Immuno Fluore Mounting Medium, ICN
Biomedicals, Aurora, OH, USA) auf einen sauberen Glasobjektträger geben und
vorsichtig die Deckgläschen auf das Medium geben. Die Ränder werden mit
Nagellack versiegelt.
16. Im Dunkeln bei 4°C lagern.
17. Die Proben werden mit konfokaler Laserimmunfluoreszenzmikroskopie ausgewertet.
Verwendet werden: Leika diaplan Fluoreszenzmikroskop, Biorad MRC 1024
Scanning Confocal Laser Microscop Software.
Als Kontrolle für die Immunfluoreszenzmikroskopie wurde das entsprechende
Immunglobulin anstelle des spezifischen Primärantikörpers eingesetzt.
Um die Proteintyrosinphosphorylierung während des spreading zu untersuchen, wurden
Parallelansätze von ausgebreiteten Thrombozyten (600 µl) mit 100 µl kochendem Laemmli-
Puffer bedeckt, mit einem Scrabber vom Coverslip gelöst und via Western Blot die
Proteintyrosinphosphorylierung gemessen. Hierfür wurden 8-Wellplatten (12,5 cm x 7,5 cm)
und große Coverslips (3,5 cm x 3 cm) eingesetzt.

35
2.2.11 Untersuchungen an knock-out Mäusen
Die Arbeiten mit Wildtyp und knock-out Mäusen erfolgten zum größten Teil in der
Arbeitsgruppe von Dr. S.P. Watson am Institute of Pharmacology in Oxford/UK. Die BALB/c
Mäuse wurden nach einer Bestrahlung mit den entsprechenden defizienten fötalen
Leberzellen rekonstituiert. Als Kontrolle dienten BALB/c Wildtyp-Mäuse.
2.2.12 Yeast Two-Hybrid System
Das Yeast Two-Hybrid System wurde von Fields et al. entwickelt. Es handelt sich um eine
hefegenetische Methode, die es ermöglicht, Protein-Proteininteraktionen in vivo
nachzuweisen (Fields et al., 1989). Als Maß hierfür wird die Transkription eines Reportergens
genutzt. Das verwendete System enthält den eukaryotischen Transkriptionsfaktor GAL4, der
in der Hefe den Galaktose-Metabolismus reguliert. Viele eukaryotische
Transkriptionsfaktoren, so auch GAL4, bestehen aus zwei physikalisch getrennten modularen
Domänen: aus einer DNA-bindenden Domäne und einer die Transkription aktivierenden
Domäne.
Die Transkription geeigneter Reportergene (LacZ, HIS3) ermöglicht den Nachweis der
Interaktion zwischen den zwei Proteinen. Über den β-Galaktosidase Assay mit X-Gal als
Substrat wurde die aktivierbare Transkription des LacZ Gens gemessen. Das zweite
Reportergen HIS3 ermöglicht eine gezielte Selektion positiver Klone.
Das Yeast Two-Hybrid System wurde verwendet, um Interaktionspartner des Proteins Syk zu
identifizieren. Hierfür wurde der Hefestamm S.c. Y190 zunächst mit dem bait-Vektor pAS2-
1/Syk transformiert und in einem zweiten Schritt mit der jeweiligen cDNA-Bank (250µg)
cotransformiert. In dem verwendeten Yeast Two-Hybrid System kann das bait-Plasmid die
TRP-Deletion, das prey-Plasmid die LEU-Deletion von Y190 ersetzen. Die transformierten
Zellen wurden auf Drop-out Medium plattiert, welchem Leucin und Tryptophan fehlte (LEU-
TRP-), um somit beide Plasmide positiv zu selektionieren. Mit Hilfe des β-Galaktosidasetests
wurden die Kolonien auf eine two-hybrid Interaktion überprüft. Bei einer positiven
Interaktion, die zur Aktivierung des HIS3 Reportergens führt, wachsen die Zellen dann auch
auf dem 3-fach Drop-out Medium (LEU-TRP-HIS-).

36
Klonierung der Plasmide:
Syk/DNA-BD Hybride: Die Syk-DNA wurde aus dem PAcG2T Baculovirus Transfer Vektor
(Geschenk Dr. S.P. Watson, Oxford) isoliert und mit dem bait-Vektor pAS2-1 ligiert (Syk
full-length). Desweiteren wurde eine Kinase-inaktive Mutante (Geschenk Dr. S.J. Shattil, La
Jolla) in den bait-Vektor pAS2-1 eingefügt (Syk kinase dead).
Im folgenden werden die grundlegenden Arbeitsmethoden mit E.coli erklärt (ausführliche
Protokolle hierzu s. Clontech Handbuch):
Bakterienanzucht
LB-Medium:
1% (w/v) Bacto-Tryptone, 0,5% (w/v) Bacto-Hefeextrakt, 1% (w/v) NaCl, pH 7,2, eingestellt
mit NaOH.
LB-Agar:
LB-Medium mit 1,25% (w/v) Bacto-Agar
Für transformierte E.coli-Zellen DH5α wurde dem Medium 150 µl/ml Ampicillin (Hoechst)
zugesetzt.
Alle E.coli-Stämme wurden bei 37°C in Vollmedium vermehrt. Aufbewahrt wurden sämtliche
Bakterienstämme in LB-Medium mit 20% (v/v) Glycerin bei �80°C oder bei 4°C auf LB-
Agar-Platten.
Isolierung von Plasmid-DNA
Die Glycerinkultur der jeweiligen Klone wurde aus -80°C kurz angetaut und eine kleine
Menge davon mit einer sterilen Impföse auf einer LB-Agar-Platte ausgestrichen. Nach einer
Trocknungszeit von 5 - 10 min bei Raumtemperatur wurde die Agarplatte über Nacht bei
37°C inkubiert. Am darauffolgenden Tag wurden 1 - 3 ml LB-Medium mit einer von der
Agar-Platte gepickten Einzelkolonie angeimpft und die E.coli-Zellen anschließend 4 - 5 h bei
37°C und 200 rpm vermehrt. Aliquots aus diesen E.coli-Kulturen wurden danach zum
Beimpfen von 10 ml bzw. 50 ml Übernacht-Kulturen verwendet.

37
Als Ausgangsmaterial für die Isolierung von Plasmid-DNA dienten die Übernacht-Kulturen.
Die Isolierung und Reinigung von supercoiled Plasmid-DNA aus E.coli Zellen wurde mit
Hilfe der Plasmid-Isolierungskits der Firma Qiagen durchgeführt. Die Herstellung des
Zellysats basiert auf einer optimierten alkalischen Lyse gemäß der Methode von Birnboim
und Doly (Birnboim und Doly, 1979) und die Reinigung der Plasmid-DNA auf einer
Chromatographie in Anionenaustauschersäulen. Im Einzelnen wurde dem Protokoll des
Herstellers gefolgt. Die Reinheit der isolierten Plasmid-DNA war für alle durchzuführenden
Reaktionen ausreichend.
Die hefegenetischen Arbeiten wurden wie folgt durchgeführt:
Lithiumacetat-Transformation von Hefe
nach (Schiestl et al., 1993)
Nachweis des in Hefe exprimierten Proteins
Um die Proteinexpression in der Hefe zu kontrollieren, wurden Proteinextrakte hergestellt
(Puffer und Vorgehensweise s. Clontech Handbuch), diese über SDS-PAGE getrennt und über
einen Westernblot die Expression des rekombinanten Proteins überprüft.
Überprüfen der HIS+, ββββ-GAL+ Kolonien
(ausführliches Protokoll s. Clontech Handbuch)
Die im Blau/Weiß-Screening gefundenen positiven Kolonien müssen auf ihre Echtheit
überprüft werden.
Hierzu wird die DNA aus der Hefe isoliert (Hoffman und Winston, 1987) und der E.coli
Stamm HB101 transformiert. Nach dem Ausplattieren werden von den erhaltenen Kolonien
so viele wie möglich auf ihre Interaktion nach erneuter Plasmidisolation und Transformation
in den Hefestamm Y190/pAS2-1/Syk full-length getestet. Entsprechende Positiv- und
Negativkontrollen wurden angefertigt.

38
3. Ergebnisse
3.1 Methodik
Es wurden einige Vorversuche zur Prüfung der Spezifität des eingesetzten polyklonalen
Antikörpers und zur Optimierung der Immunpräzipitation von Syk in Thrombozyten
durchgeführt. Für die Isolierung der Thrombozyten waren mehrmalige Wasch- und
Zentrifugationsschritte notwendig. Um die Bildung und Wirkung von Thromboxan A2 und
ADP während der Thrombozytenpräparation zu unterbinden, wurde das PRP mit Aspirin
inkubiert. Der Wasch- und Resuspensionspuffer enthielt Apyrase, welches ADP inaktiviert.
3.1.1. Expression von Syk in Thrombozyten, aber nicht in Endothelzellen
Um das Protein Syk spezifisch in einem Lysat und damit Gemisch verschiedenster Proteine
nachzuweisen, wurde die Methode des Western Blots eingesetzt. Hierbei wurden die Proteine
in einem ersten Schritt via SDS-Gelelektrophorese nach ihrer Größe aufgetrennt, auf eine
Nitrocellulosemembran übertragen und schließlich das gesuchte Protein im Immunoblot
spezifisch nachgewiesen. Im nachfolgend beschriebenen Experiment wurden Lysate von
Thrombozyten und Endothelzellen untersucht. Für das Immunostaining wurde ein
polyklonaler anti-Syk Antikörper verwendet. Die Blots (s. Abb. 3) zeigten in den
Thrombozytenlysaten eine klare Bande in der Höhe von 72 kDa, welche das Protein Syk
repräsentiert. In den Endothelzellysaten war eine Doppelbande in der Höhe von etwa 75 kDa
zu beobachten. Der Kontrollblot mit Kaninchen IgG zeigte jedoch in den Endothelzellen
außer weiteren unspezifischen Banden die gleiche Doppelbande; in Thrombozyten waren
keine unspezifischen Banden erkennbar. Syk ist somit spezifisch in humanen Thrombozyten
exprimiert, nicht jedoch in humanen Endothelzellen.

39
1
2 41 3
anti-Syk IgGKaninchen
97
66
45
1 2 3 4
Kaninchen IgG
Endothelz.
Marker
TZ TZEndothelz.
A B
Syk
Abb. 3: Expression von Syk in Thrombozyten, aber nicht in Endothelzellen
Es wurden konfluente Endothelzellen- (Spuren 1, 2; 2,5 x 106 Endothelzellen = 1 mg Protein) und
Thrombozytensuspensionen (Spuren 3, 4; 5 x 108 Thrombozyten = 1 mg Protein) im Verhältnis 1:1 mit 2x
Laemmli-Puffer lysiert (jeweils Doppelwerte aus dem gleichen Versuchsansatz). Nach Auftrennung über SDS-
Gelelektrophorese und Übertragen der Proteine auf eine Nitrocellulose-Membran wurde ein anti-Syk Blot (A)
(polyklonales Kaninchen anti-Syk IgG, Verdünnung 1:2500) durchgeführt. Nach stripping der Membran zur
Entfernung gebundener Antikörper wurde die Membran mit der gleichen Konzentration an unspezifischem
Kaninchen IgG (B) (40 ng/ml) inkubiert, um die Spezifität des Syk-Immunoblots zu prüfen.
3.1.2 Messung der Syk-Tyrosinphosphorylierung nach Immunpräzipita-
tion
Das Zusammenspiel von Kinasen und Phosphatasen und der durch sie erfolgende Transfer
von Phosphatgruppen spielt eine entscheidende Rolle bei der Signaltransduktion. Da nur ein
kleiner Prozentsatz der Proteine bei der Aktivierung tyrosinphosphoryliert wird, bedient man

40
sich der Immunpräzipitation als Methode zur Anreicherung des erwünschten Proteins.
Die folgenden Abbildungen 4 bis 6 zeigen die Resultate von Optimierungsversuchen
verschiedener Parameter zum Nachweis von tyrosinphosphoryliertem Syk. Es wurden die
Einflüsse von Spender, verwendetem Lysatvolumen, Thrombozytenkonzentration,
Antikörpermenge und Protein A-Sepharose Menge untersucht. In Abb. 4 war auffallend, daß
der Anteil an tyrosinphosphoryliertem Syk von Spender 8 sehr viel niedriger war als der von
Spender 11 bei etwa gleicher Menge an immunpräzipitiertem Syk. Während daher im Fall
Spender 11 eine Lysatmenge von 300 µl und eine Thrombozytenkonzentration von 8 x 108 /
ml ausreichen würde, müßte bei Spender 8 viel mehr Zellmaterial eingesetzt werden, um
verwertbare Ergebnisse zu erhalten. Eine weitere Variationsmöglichkeit besteht in der
Verwendung von Protein A-Sepharose oder Agarose als Kopplungspartner für den Syk-
Antikörper sowie in der Antikörpermenge. Bei letzterer wurde zwischen 5 und 40 µg pro 0,6
ml Lysatvolumen variiert. Unter Verwendung von Protein A-Sepharose gekoppeltem Syk-
Antikörper im Vergleich zu an Agarose gekoppelten Syk-Antikörper konnte eine etwa 2 - 3-
fach höhere Syk-Immunpräzipitation festgestellt werden (Abb. 5). Ähnliche Unterschiede
waren im anti-Phosphotyrosin Blot der entsprechenden Syk-Immunpräzipitate zu beobachten.
Im Gegensatz zur Protein A-Sepharose ist der kommerzielle Syk-Antikörper kovalent an die
Agarose gebunden. Innerhalb einer Optimierungsreihe wurde gefunden, daß bei Verwendung
des an Protein A-Sepharose gekoppelten Antikörpers die Antikörpermenge von 10 µg für ein
gut verwertbares Signal ausreichte. Desweiteren ergab sich, daß ein Lysatvolumen von 0,6
ml, eine Antikörpermenge von 10 µg und 2,5 µg Protein A-Sepharose die besten Ergebnisse
erbrachte. Bereits ein Lysatvolumen von 0,3 ml und eine Antikörpermenge von 5 µg lieferten
verwertbare Resultate (Abb. 6).
Bei der Auswertung der Syk-Tyrosinphosphorylierung wurde stets die immunpräzipitierte
Syk-Menge mitgemessen und es wurde der Quotient Syk-PY/Syk von jeder Probe gebildet,
um Schwankungen der immunpräzipitierten Menge an Syk zu berücksichtigen. Die Menge
von immunpräzipitiertem Syk lag in einem Bereich von 15 - 60 % der Gesamtmenge von Syk
in Thrombozyten.

41
anti-Phosphotyrosin anti-Syk
Syk
Konz.[x 10 /ml]8
Lysatvol.
Spender Sp 11 Sp 11Sp 8 Sp 8
300 µl 300 µl300 µl 300 µl600 µl 600 µl600 µl 600 µl
8 88 88 88 816 1616 1616 1616 16
A
Abb. 4: Vergleich der Quantität von immunpräzipitiertem Syk und der Stärke der Syk-
Tyrosinphosphorylierung in Abhängigkeit von Blutspender, Thrombozytenkonzentration und
Lysatmenge
Von zwei Spendern (Sp 11 und Sp 8) wurden Thrombozytensuspensionen der angegebenen
Thrombozytenkonzentration präpariert und Syk wurde mit einem spezifischen an Agarose gekoppelten anti-Syk
Kaninchen IgG immunpräzipitiert. Von 75 % der Syk-Immunpräzipitate wurde ein Anti-Phosphotyrosin-Blot
(A), von den übrigen 25 % ein anti-Syk Blot (B) angefertigt.

42
AK-Menge [µg]
Syk
Protein A-Seph. Agarose
anti-Syk
5 10 20 40 5 40
Abb. 5: Optimierung der Syk-Immunpräzipitation durch Variation der Antikörpermenge bzw. Vergleich
von Protein A-Sepharose und Agarose gekoppeltem Antikörper
Suspensionen von gewaschenen Thrombozyten wurden mit 2x Lysis-Puffer A lysiert. Entweder wurde Syk mit
anti-Syk IgG gekoppelter Agarose oder mit anti-Syk IgG und anschließender Zugabe von Protein A-Sepharose
immunpräzipitiert. Es wurden Antikörperkonzentrationen zwischen 5 µg und 40 µg eingesetzt. Von den Syk-
Immunpräzipitaten wurde ein anti-Syk Blot angefertigt.
A B
anti-Phosphotyrosin anti-Syk
Lysatvolumen
AK-Menge [µg]
Prot.A-Seph. [mg]
Syk
600 µl 600 µl300 µl 300 µl
10 1010 105 52,5 2,52,5 2,55 5
5 52,5 2,51,2 1,21,2 1,20,6 0,60,6 0,6
Abb. 6: Optimierung der Syk-Immunpräzipitation durch Variation des Lysat-Volumens, der
Antikörpermenge und der Protein A-Sepharose Menge
Durch Variation von Lysatmenge, Menge an Syk-Antikörper und Menge an Protein A-Sepharose wurde die
optimale Kombination für eine hohe Proteinausbeute bei möglichst geringem Materialverbrauch ermittelt. (A)
zeigt den Anti-Phosphotyrosin-Blot, (B) den anti-Syk Blot.

43
3.1.3 Expression von Syk bei verschiedenen Probanden
Die Expression von Syk wurde in humanen Thrombozyten sowohl bei verschiedenen
Probanden als auch bei den gleichen Probanden an verschiedenen Tagen gemessen, um
eventuelle inter- und intraindividuelle Schwankungen zu ermitteln. Es waren deutliche
interindividuelle und z.T. auch intraindividuelle Variationen der Expression von Syk zu
beobachten. In Tab. 1 sind die Resultate eines Vergleichs von 11 Blutspendern aufgelistet,
wobei die Testperson Sp 1 an 3 Tagen (a, b, c) untersucht wurde. Bei Testperson 1 am
Versuchstag a wurden die höchsten Werte ermittelt, welche 100% gesetzt wurden und auf
welche sämtliche andere Werte bezogen wurden. Spender 7 liegt mit einem Durchschnittswert
von 31% des Maximums am niedrigsten in der Höhe der Syk-Expression. Die Ergebnisse für
die 3 Testtage von Spender 1 liegen zwischen 43% und 100%.
Zusammenfassend kann die Schwankung der Methode als gering bezeichnet werden, was aus
dem Mittelwert +/- Standardabweichung der Mehrfachbestimmungen, die bei vielen Spendern
durchgeführt wurden, zu erkennen ist. Es bestehen jedoch interindividuelle Variationen der
Syk-Expression und auch intraindividuelle Schwankungen der Syk-Expression (s. Spender 1).
Tab. 1: Vergleich der Syk-Expression in Thrombozyten von verschiedenen Probanden
Spender Syk-Expression in Thrombozyten [%]
Sp 1a 100
Sp 1b 44,7 ± 16,2; n = 3
Sp 1c 43,0 ± 10,6; n = 3
Sp 2 88 / 76
Sp 3 55
Sp 4 77,7 ± 7,8; n = 3
Sp 5 60 / 56
Sp 6 48 / 54
Sp 7 22 / 40
Sp 8 43,7 ± 9,5; n = 3
Sp 9 39,7 ± 4,0; n = 3
Sp 10 46 / 60
Sp 11 57 / 43
Von verschiedenen Probanden (Sp 1-11) wurden Thrombozytenlysate (von 4 x 107 Zellen) über Gelelektrophorese aufgetrennt, und in einem Syk-Immunoblot wurde die Expression von Syk gemessen. Die Ergebnisse sind Doppelwerte bzw. Mittelwerte ± SD von Mehrfachbestimmungen eines Versuchs bezogen auf einen internen Standard (Proband 1am Versuchstag a = 1a = 100 %). Von Proband 1 liegen außerdem Ergebnisse von 2 weiteren Versuchstagen vor, welche mit den Nummern 1b und 1c bezeichnet wurden.

44
3.2 Tyrosinphosphorylierung von Syk während des shape change
3.2.1 Aktivierung nach Stimulation des Thrombin-Rezeptors PAR-1 und
nach mox-LDL (milde oxidiertes LDL)
YFLLRNP, ein Heptapeptid, ist ein partieller Thrombinrezeptoragonist, der in einer
Konzentration von 300 µM einen shape change induziert, aber zu keiner detektierbaren
Sekretion von ATP aus dichten Granula führt (Rasmussen et al., 1993). Als weiterer Agonist
wurde mox-LDL untersucht, das in Aspirin-vorbehandelten Thrombozyten nur einen
Formwandel, aber keine ATP-Sekretion oder Aggregation induziert (Weidtmann, 1996;
Baumann-Siemons, 2000). Untersuchungen aus der Arbeitsgruppe haben gezeigt, daß mox-
LDL Lysophosphatidsäure enthält, welche für den Thrombozytenformwandel induziert durch
mox-LDL verantwortlich ist, und diesen über die Aktivierung von LPA-Rezeptoren induziert
(Siess et al., 1999). Um die Bildung von Mikroaggregaten zu verhindern, wurden die
Suspensionen von gewaschenen Thrombozyten mit RGDS (2 mM) für 2 min vorbehandelt.
Dosis-Wirkungs-Kurven zeigten, daß der shape change durch geringe Konzentrationen von
mox-LDL von nur 10-15 µg/ml induziert werden konnte. Es wurde gefunden, daß die Zugabe
von YFLLRNP oder mox-LDL die Tyrosinphosphorylierung von Syk während des shape
change stimulierte (Abb. 7A, B). Die Tyrosinphosphorylierung von Syk nahm nach
YFLLRNP (300 µM, 30 s) um das 5,4±1,7-fache (Mittelwert ± SD, n = 8, P<0,0001) und
nach mox-LDL (0,5 mg/ml, 30 s) um das 2,5±1,0-fache (Mittelwert ± SD, n = 21, P<0,0001)
zu. Die relativ hohe Standardabweichung ist auf inter- und intraindividuelle Schwankungen
zurückzuführen (s. 3.1.3) sowie in geringem Maß auch auf Abweichungen, die methodischen
Ursprungs sind (s. 3.1.2).
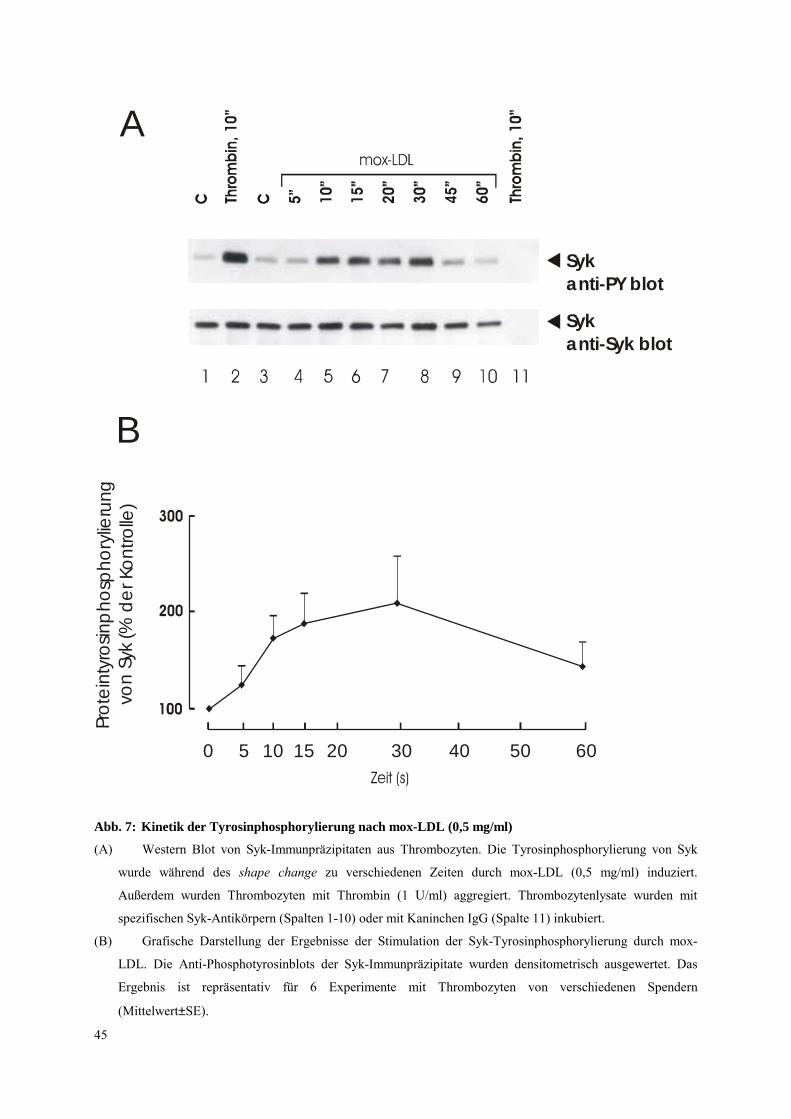
In Kinetiken der Syk-Tyrosinphosphorylierung (Abb. 7A, B) wurde gefunden, daß die
Stimulierung der Tyrosinphosphorylierung 10 s nach Zugabe von mox-LDL zu beobachten
war. Die maximalen Werte (2-3-fach) wurden 15-30 s nach Zugabe von mox-LDL (s. Abb.
7B) erreicht und nahmen dann wieder ab. Nach Stimulation der Thrombozyten mit mox-LDL
zeigten der shape change und der Anstieg der Tyrosinphosphorylierung von Syk ähnliche
Dosiswirkungskurven (s. Abb. 8). Diese Korrelation könnte darauf hinweisen, daß die
Tyrosinphosphorylierung der Tyrosinkinase Syk für den shape change wichtig ist.
45
B
A
Sykanti-PY blot
Sykanti-Syk blot
0 5 10 15 20 30 40 50 60
Pro
tein
t yro
sinp
hosp
hory
lier u
ng
von
S yk
(% d
er K
on t
rolle
)
Abb. 7: Kinetik der Tyrosinphosphorylierung nach mox-LDL (0,5 mg/ml)
(A) Western Blot von Syk-Immunpräzipitaten aus Thrombozyten. Die Tyrosinphosphorylierung von Syk
wurde während des shape change zu verschiedenen Zeiten durch mox-LDL (0,5 mg/ml) induziert.
Außerdem wurden Thrombozyten mit Thrombin (1 U/ml) aggregiert. Thrombozytenlysate wurden mit
spezifischen Syk-Antikörpern (Spalten 1-10) oder mit Kaninchen IgG (Spalte 11) inkubiert.
(B) Grafische Darstellung der Ergebnisse der Stimulation der Syk-Tyrosinphosphorylierung durch mox-
LDL. Die Anti-Phosphotyrosinblots der Syk-Immunpräzipitate wurden densitometrisch ausgewertet. Das
Ergebnis ist repräsentativ für 6 Experimente mit Thrombozyten von verschiedenen Spendern
(Mittelwert±SE).
46
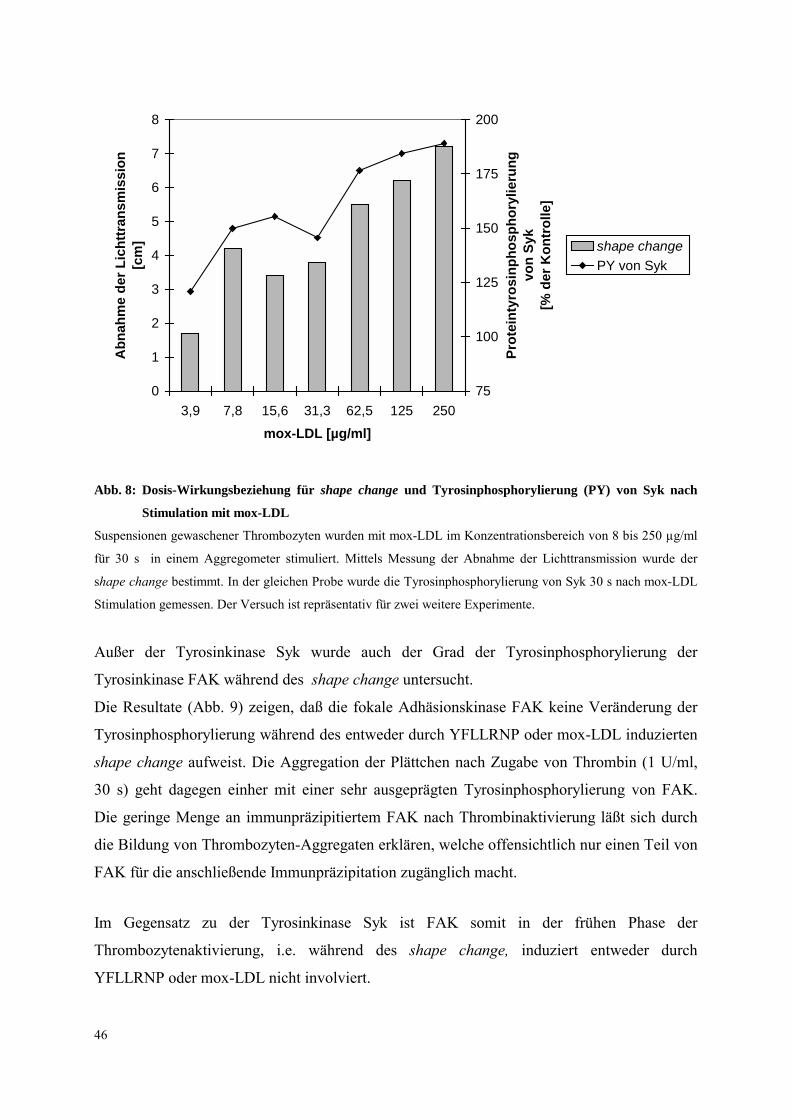
Abb. 8: Dosis-Wirkungsbeziehung für shape change und Tyrosinphosphorylierung (PY) von Syk nach
Stimulation mit mox-LDL
Suspensionen gewaschener Thrombozyten wurden mit mox-LDL im Konzentrationsbereich von 8 bis 250 µg/ml
für 30 s in einem Aggregometer stimuliert. Mittels Messung der Abnahme der Lichttransmission wurde der
shape change bestimmt. In der gleichen Probe wurde die Tyrosinphosphorylierung von Syk 30 s nach mox-LDL
Stimulation gemessen. Der Versuch ist repräsentativ für zwei weitere Experimente.
Außer der Tyrosinkinase Syk wurde auch der Grad der Tyrosinphosphorylierung der
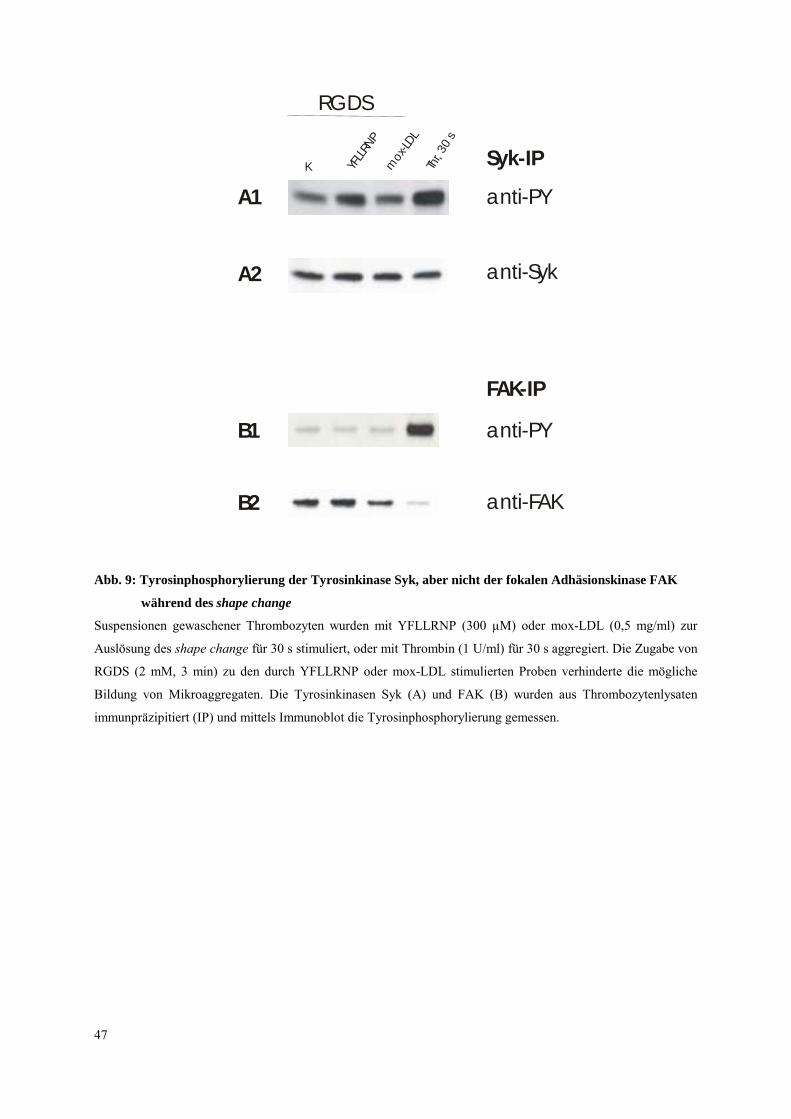
Tyrosinkinase FAK während des shape change untersucht.
Die Resultate (Abb. 9) zeigen, daß die fokale Adhäsionskinase FAK keine Veränderung der
Tyrosinphosphorylierung während des entweder durch YFLLRNP oder mox-LDL induzierten
shape change aufweist. Die Aggregation der Plättchen nach Zugabe von Thrombin (1 U/ml,
30 s) geht dagegen einher mit einer sehr ausgeprägten Tyrosinphosphorylierung von FAK.
Die geringe Menge an immunpräzipitiertem FAK nach Thrombinaktivierung läßt sich durch
die Bildung von Thrombozyten-Aggregaten erklären, welche offensichtlich nur einen Teil von
FAK für die anschließende Immunpräzipitation zugänglich macht.
Im Gegensatz zu der Tyrosinkinase Syk ist FAK somit in der frühen Phase der
Thrombozytenaktivierung, i.e. während des shape change, induziert entweder durch
YFLLRNP oder mox-LDL nicht involviert.
0
1
2
3
4
5
6
7
8
3,9 7,8 15,6 31,3 62,5 125 250mox-LDL [µg/ml]
Abn
ahm
e de
r Lic
httr
ansm
issi
on
[cm
]
75
100
125
150
175
200
Prot
eint
yros
inph
osph
oryl
ieru
ng
von
Syk
[% d
er K
ontr
olle
]
shape changePY von Syk
47
A1
A2
B1
B2
Syk-IP
FAK-IP
anti-PY
anti-Syk
anti-PY
anti-FAK
RGDS
K YFLL
RNP
mox
-LDL
Thr,
30 s
Abb. 9: Tyrosinphosphorylierung der Tyrosinkinase Syk, aber nicht der fokalen Adhäsionskinase FAK
während des shape change
Suspensionen gewaschener Thrombozyten wurden mit YFLLRNP (300 µM) oder mox-LDL (0,5 mg/ml) zur
Auslösung des shape change für 30 s stimuliert, oder mit Thrombin (1 U/ml) für 30 s aggregiert. Die Zugabe von
RGDS (2 mM, 3 min) zu den durch YFLLRNP oder mox-LDL stimulierten Proben verhinderte die mögliche
Bildung von Mikroaggregaten. Die Tyrosinkinasen Syk (A) und FAK (B) wurden aus Thrombozytenlysaten
immunpräzipitiert (IP) und mittels Immunoblot die Tyrosinphosphorylierung gemessen.

48
3.2.2 Integrin ααααIIbββββ3 unabhängige Tyrosinphosphorylierung von Syk
während des shape change
3.2.2.1 Thrombasthenia Glanzmann
Bei Patienten mit Thrombasthenia Glanzmann ist die Expression des Integrins αIIbβ3 in
Thrombozyten stark reduziert. Untersuchungen an Thrombozyten dieser Patienten wurden
durchgeführt, (a) um zu bestätigen, daß Syk während des shape change unabhängig von dem
Integrin αIIbβ3 aktiviert wird, und (b) um herauszufinden, ob bei der initialen
Thrombozytenaktivierung durch mox-LDL das Integrin αIIbβ3 eine Rolle spielt. Frühere
Untersuchungen wiesen darauf hin, daß natives LDL an das Integrin αIIbβ3 (Koller et al.,
1989, Hackeng et al., 1999) bindet. Es wurde daher die Frage untersucht, ob das Integrin
αIIbβ3 bei der Thrombozyten-Aktivierung durch mox-LDL beteiligt ist. Es wurden drei
Patienten (A,B,C) mit Thrombasthenia Glanzmann untersucht.
Es wurde zunächst in Dosis-Wirkungskurven (Abb. 10) die Aktivierbarkeit (shape change)
von thrombasthenischen Plättchen mit der von Kontrollplättchen nach mox-LDL (0,5 mg/ml,
30 s) verglichen. Die EC50 betrug bei den Kontrollplättchen 0,063 mg/ml, während sie bei
Plättchen des thrombasthenischen Patienten A bei 0,015 mg/ml lag. Letztere reagierten somit
sogar sensitiver auf mox-LDL, was jedoch auch vom Blutspender abhängen kann.
Desweiteren wurde bei allen drei Patienten A, B und C die Tyrosinphosphorylierung der
Proteine P62, P68, P130 und von Syk (keine Syk-IP bei Patient A, da Aktivierung zu
schwach) nach mox-LDL (0,5 mg/ml, 30 s) mit der von Kontroll-Thrombozyten verglichen
(Abb. 11, Daten Patient B). Bei allen Patienten war ein ähnliches Protein-
Tyrosinphosphorylierungsmuster während des shape change zu finden, was darauf schließen
läßt, daß das Integrin αIIbβ3 bei der initialen Thrombozytenaktivierung durch mox-LDL
nicht beteiligt ist. Weiterhin wurde die Tyrosinphosphorylierung von Syk in
thrombasthenischen Plättchen (Patient B) nach Aktivierung mit YFLLRNP (300 µm, 20 s)
gemessen (Abb. 11). Auch hier war das Phosphorylierungsmuster vergleichbar mit dem der
Kontrollplättchen. Die Ergebnisse zeigen, daß die Tyrosinphosphorylierung von Syk während
des shape change unabhängig vom Integrin αIIbβ3 abläuft. Weiterhin ist das Integrin αIIbβ3
für den Gestaltwandel in der frühen Phase der Thrombozytenaktivierung nicht erforderlich.
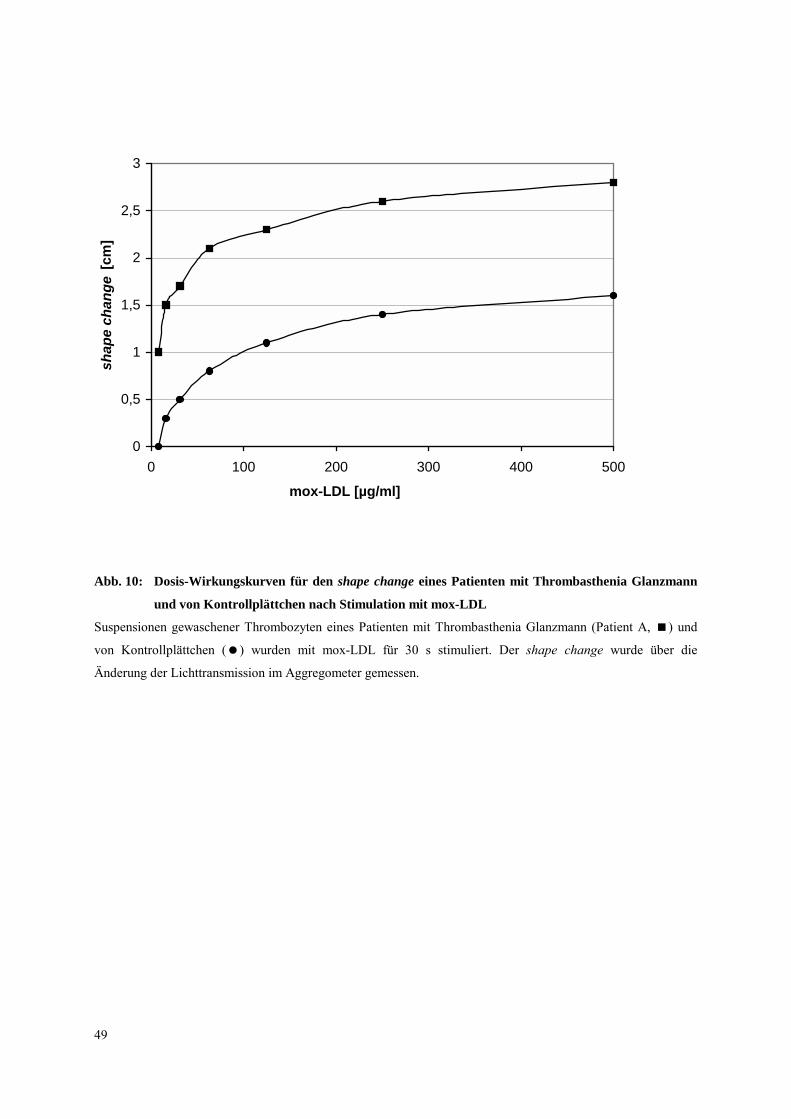
49
Abb. 10: Dosis-Wirkungskurven für den shape change eines Patienten mit Thrombasthenia Glanzmann
und von Kontrollplättchen nach Stimulation mit mox-LDL
Suspensionen gewaschener Thrombozyten eines Patienten mit Thrombasthenia Glanzmann (Patient A, �) und
von Kontrollplättchen (�) wurden mit mox-LDL für 30 s stimuliert. Der shape change wurde über die
Änderung der Lichttransmission im Aggregometer gemessen.
0
0,5
1
1,5
2
2,5
3
0 100 200 300 400 500
mox-LDL [µg/ml]
shap
e ch
ange
[cm
]
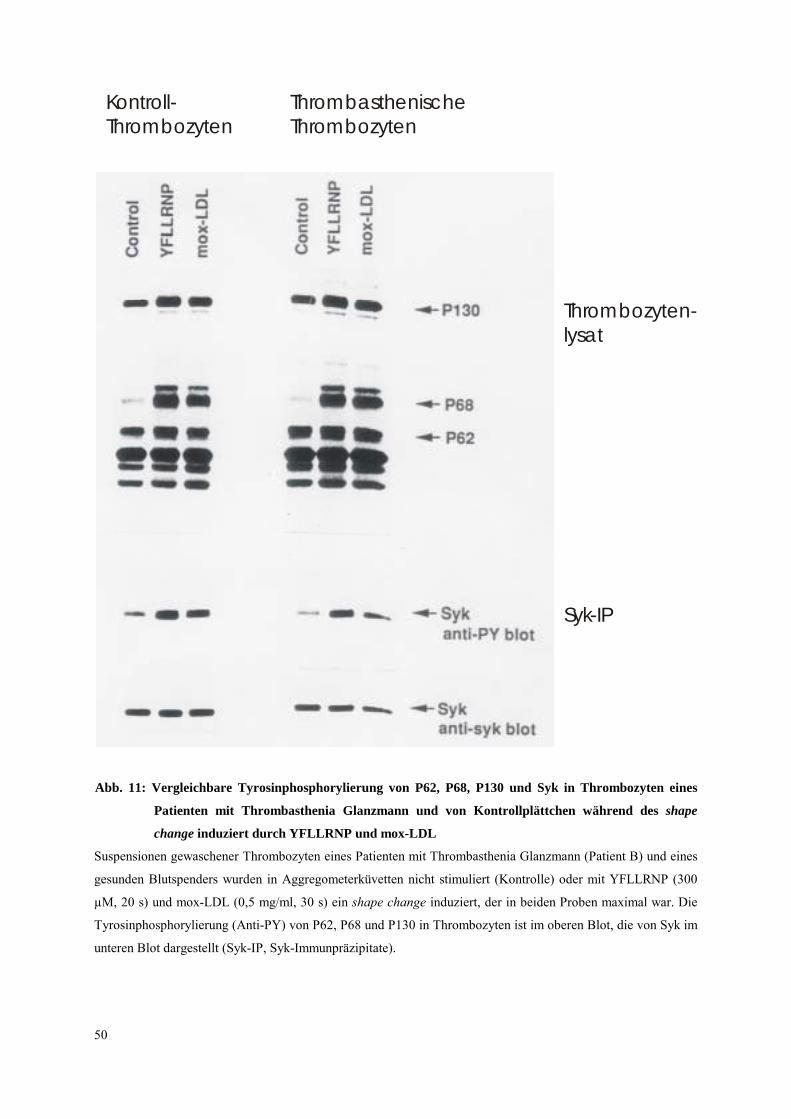
50
Kontroll-Thrombozyten
ThrombasthenischeThrombozyten
Thrombozyten-lysat
Syk-IP
Abb. 11: Vergleichbare Tyrosinphosphorylierung von P62, P68, P130 und Syk in Thrombozyten eines
Patienten mit Thrombasthenia Glanzmann und von Kontrollplättchen während des shape
change induziert durch YFLLRNP und mox-LDL
Suspensionen gewaschener Thrombozyten eines Patienten mit Thrombasthenia Glanzmann (Patient B) und eines
gesunden Blutspenders wurden in Aggregometerküvetten nicht stimuliert (Kontrolle) oder mit YFLLRNP (300
µM, 20 s) und mox-LDL (0,5 mg/ml, 30 s) ein shape change induziert, der in beiden Proben maximal war. Die
Tyrosinphosphorylierung (Anti-PY) von P62, P68 und P130 in Thrombozyten ist im oberen Blot, die von Syk im
unteren Blot dargestellt (Syk-IP, Syk-Immunpräzipitate).

51
3.2.2.2 RGDS
Das Tetrapeptid RGDS verhindert die Bindung von Fibrinogen, Fibronectin und vWF an das
Integrin αIIbβ3. Der shape change und die begleitende Tyrosinphosphorylierung der Proteine
P62, P68, P130 und Syk induziert durch YFLLRNP (300 µm) oder mox-LDL (0,5 mg/ml)
blieb unbeeinflußt von einer RGDS (2 mM, 3 min) Vorbehandlung der Thrombozyten. Dies
verdeutlicht weiter die Integrin αIIbβ3-unabhängige Stimulation der
Tyrosinphosphorylierung von Syk und der Proteine P62, P68 und P130 während des shape
change (Resultate nicht gezeigt).
3.2.3 Regulation
3.2.3.1 Proteinkinase C Um den Einfluß der Proteinkinase C auf den shape change und die assoziierte
Tyrosinphosphorylierung der Proteine P62, P68 und P130 der Thrombozyten zu untersuchen,
wurden die Thrombozytensuspensionen vor der Stimulation mit mox-LDL (0,5 mg/ml, 30 s)
bzw. YFLLRNP (300 µM, 30 s) mit dem Proteinkinase C-Inhibitor Ro-31-8220 (0,2 µM, 20
min). Unter diesen Bedingungen war die Thrombozyten-Aggregation induziert durch den
Phorbolester Phorbol 12,13-dibutyrat (200 nM) komplett gehemmt, was auf eine effektive und
komplette Hemmung der Proteinkinase C schließen läßt. Der Anstieg der
Tyrosinphosphorylierung von Syk während des shape change nach Stimulation mit
YFLLRNP betrug 240 ± 14,1%, nach mox-LDL 244 ± 197,8% (n = 3), bezogen auf die
Kontrolle vorinkubiert mit Ro-31-8220. Im Vergleich dazu die Werte ohne Vorinkubation mit
Ro-31-8220 s. 3.2.1. Da durch den Inhibitor Ro-31-8220 keine Hemmeffekte auftraten, läßt
dies auf eine Unabhängigkeit des Syk-Tyrosinphosphorylierung von der Proteinkinase C
schließen.

52
3.2.3.2 Calcium
Es wurde der Einfluß der Regulation von Syk durch zytosolisches Ca2+ untersucht. BAPTA-
AM cheliert zytosolisches Ca2+, wobei die kombinierte Behandlung von BAPTA-AM (20
µM, 20 min) und EGTA (2 mM, 1 min) das zytosolische Ca2+ in ruhenden Thrombozyten
erniedrigt (Bauer et al., 1999). Nach dieser Vorbehandlung der Thrombozyten war der shape
change nach mox-LDL und YFLLRNP verzögert und reduziert. Die BAPTA-AM und EGTA-
Behandlung erniedrigte die basale Tyrosinphosphorylierung von Syk um 64,8% ± 10,0 (n =
4). Eine Stimulation mit YFLLRNP (300 µM) führte zu keiner Erhöhung der Syk-
Tyrosinphosphorylierung. Aus diesen Befunden kann geschlossen werden, daß für die Syk-
Tyrosinphosphorylierung in ruhenden Thrombozyten und nach Stimulation mit YFLLRNP
die normale zytosolische Ca2+-Konzentration (80 - 100 nM) erforderlich ist.
Während des shape change findet eine starke Neuorganisation des Zytoskeletts statt (Siess,
1989). Ruhende diskoide Thrombozyten enthalten 3 � 5% tyrosinphosphoryliertes P62, 6 �
9% P68 und 10 � 20% P130 im Aktin-reichen Zytoskelett. Während des shape change
induziert durch mox-LDL, steigt der Gehalt an tyrosinphosphoryliertem P62, P68 und P130
auf das 2,9-fache, 7,2-fache und 2,4-fache an (Tab. 2). Ein möglicher Mechanismus für die
erhöhte Tyrosinphosphorylierung der Zytoskelettproteine ist die Translokation von
Proteintyrosinkinasen zum Zytoskelett. Es wurde daher die zelluläre Verteilung von mehreren
Tyrosinkinasen in ruhenden Thrombozyten und während des shape change induziert durch
mox-LDL untersucht. Unstimulierte Plättchen enthielten 4 � 7% Fyn, 3 � 6% Src und 0,7 �
1,4% Syk im Zytoskelett. Während des shape change induziert durch mox-LDL stieg die
Assoziation von Fyn mit dem Zytoskelett stärker (4,8-fach) an als die von Src (1,5-fach) und
Syk (3,1-fach). Die Translokation von aktiven Tyrosinkinasen zum Zytoskelett könnte
wichtig sein für die Phosphorylierung von spezifischen Zielproteinen im Zytoskelett.
Desweiteren wurde der Effekt der BAPTA-AM/EGTA Inkubation untersucht. Dies hatte
kaum einen Effekt auf die Zytoskelettassoziation der Proteintyrosinkinasen Src, Fyn und Syk
nach Stimulation mit mox-LDL. Sie führte jedoch zu einer Abnahme der
tyrosinphosphorylierten Proteine P62, P68 und P130 im Zytoskelett von nicht-stimulierten
Zellen. Die Stimulation der Thrombozyten mit mox-LDL erhöhte jedoch auch in BAPTA-
AM/EGTA-vorbehandelten Zellen die Proteintyrosinphosphorylierung in ganzen Zellen
(Daten nicht gezeigt) und im Zytoskelett (Tab. 2). Es scheint also, daß eine normale basale
Höhe des zytosolischen Ca2+ (80 � 100 nM) erforderlich ist für die Tyrosinphosphorylierung

53
von P62, P68 und P130 in ruhenden Thrombozyten. In mit mox-LDL stimulierten
Thrombozyten hingegen oder nach Thrombin-Rezeptoraktivierung ist die Stimulation der
Tyrosinphosphorylierung von P62, P68 und P130 nicht von einem Anstieg des zytosolischen
Ca2+ abhängig. Dies ist im Gegensatz zu den Ergebnissen der Syk-Tyrosinphosphorylierung.
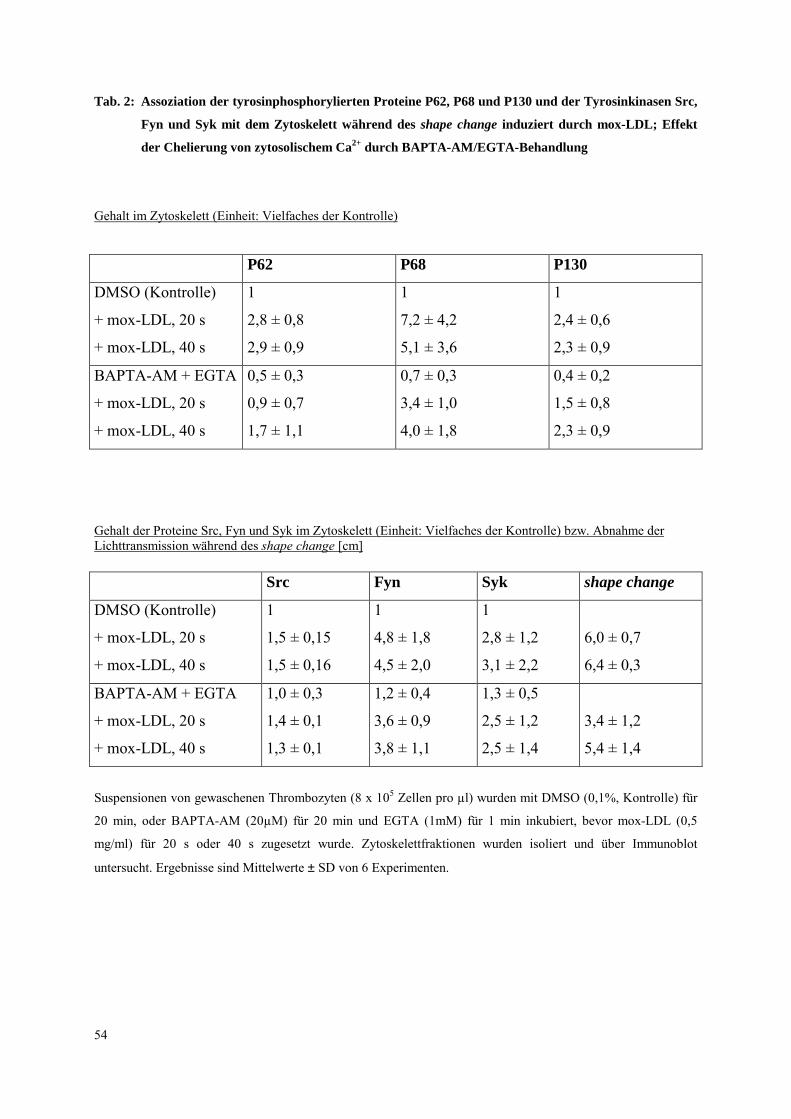
54
Tab. 2: Assoziation der tyrosinphosphorylierten Proteine P62, P68 und P130 und der Tyrosinkinasen Src,
Fyn und Syk mit dem Zytoskelett während des shape change induziert durch mox-LDL; Effekt
der Chelierung von zytosolischem Ca2+ durch BAPTA-AM/EGTA-Behandlung
Gehalt im Zytoskelett (Einheit: Vielfaches der Kontrolle)
P62 P68 P130
DMSO (Kontrolle)
+ mox-LDL, 20 s
+ mox-LDL, 40 s
1
2,8 ± 0,8
2,9 ± 0,9
1
7,2 ± 4,2
5,1 ± 3,6
1
2,4 ± 0,6
2,3 ± 0,9
BAPTA-AM + EGTA
+ mox-LDL, 20 s
+ mox-LDL, 40 s
0,5 ± 0,3
0,9 ± 0,7
1,7 ± 1,1
0,7 ± 0,3
3,4 ± 1,0
4,0 ± 1,8
0,4 ± 0,2
1,5 ± 0,8
2,3 ± 0,9
Gehalt der Proteine Src, Fyn und Syk im Zytoskelett (Einheit: Vielfaches der Kontrolle) bzw. Abnahme der Lichttransmission während des shape change [cm] Src Fyn Syk shape change
DMSO (Kontrolle)
+ mox-LDL, 20 s
+ mox-LDL, 40 s
1
1,5 ± 0,15
1,5 ± 0,16
1
4,8 ± 1,8
4,5 ± 2,0
1
2,8 ± 1,2
3,1 ± 2,2
6,0 ± 0,7
6,4 ± 0,3
BAPTA-AM + EGTA
+ mox-LDL, 20 s
+ mox-LDL, 40 s
1,0 ± 0,3
1,4 ± 0,1
1,3 ± 0,1
1,2 ± 0,4
3,6 ± 0,9
3,8 ± 1,1
1,3 ± 0,5
2,5 ± 1,2
2,5 ± 1,4
3,4 ± 1,2
5,4 ± 1,4
Suspensionen von gewaschenen Thrombozyten (8 x 105 Zellen pro µl) wurden mit DMSO (0,1%, Kontrolle) für
20 min, oder BAPTA-AM (20µM) für 20 min und EGTA (1mM) für 1 min inkubiert, bevor mox-LDL (0,5
mg/ml) für 20 s oder 40 s zugesetzt wurde. Zytoskelettfraktionen wurden isoliert und über Immunoblot
untersucht. Ergebnisse sind Mittelwerte ± SD von 6 Experimenten.

55
3.2.3.3 Piceatannol und weitere Proteintyrosinkinase-Hemmer, Iloprost
Desweiteren wurde die Wirkung des in der Literatur als Syk-selektiv beschriebenen
Hemmstoffs Piceatannol untersucht (Keely und Parise, 1996). Es wurde gefunden, daß
Piceatannol die Stimulation der Tyrosinphosphorylierung der Proteine P62, P68 und P130
während des shape change induziert durch mox-LDL hemmt. Die dosisabhängige Reduktion
der Tyrosinphosphorylierung der Proteine P62, P68 und P130 nach Auftrennung über SDS-
PAGE und Western Blot Analyse ist in Abb. 12 dargestellt. Die Reduktion der
Tyrosinphosphorylierung erfolgte analog zur Abnahme des shape change. Vor allem für P68
ist die Abnahme der Tyrosinphosphorylierung mit ansteigender Konzentration an Piceatannol
ausgeprägt. In parallel dazu durchgeführten Syk-Immunpräzipitations-Experimenten (n = 3)
wurde untersucht, ob Piceatannol die Tyrosinphosphorylierung von Syk hemmt. Es wurde
jedoch keine Hemmung der Syk-Tyrosinphosphorylierung festgestellt (Daten nicht gezeigt).
Die in der Literatur beschriebene Syk-selektive Hemmwirkung von Piceatannol konnte somit
nach Stimulation der Thrombozyten mit mox-LDL nicht gefunden werden.
P130
P68
P62
K KKKK KK KK KK Kmox-LDL
mox-LDL
mox-LDL
mox-LDL
mox-LDL
mox-LDL
Piceatannol15 µg/ml 30 µg/ml
DMSO DMSO Piceatannol45 µg/ml 60 µg/ml0,3% 0,6%
Abb. 12: Dosisabhängige Reduktion der Tyrosinphosphorylierung in Thrombozytenlysaten durch
Piceatannol
Suspensionen gewaschener Thrombozyten wurden mit DMSO (0,3%, bzw. 0,6%) oder ansteigenden
Konzentrationen von Piceatannol (15 - 60 µg/ml in DMSO) 10 min bei 37°C inkubiert und anschließend mit
mox-LDL (0,5 mg/ml, 30 s) stimuliert. Gezeigt ist ein Antiphosphotyrosin-Blot der Thrombozytenlysate
(repräsentativ für 3 Experimente).

56
Desweiteren wurden verschiedene generelle Tyrosinkinase-Hemmer auf ihren Effekt auf die
Tyrosinphosphorylierung von Syk während des shape change untersucht. Negrescu et al.
konnten zeigen, daß die Proteintyrosinkinase-Inhibitoren Genistein und Tyrphostin A23 den
YFLLRNP induzierten shape change sowie die Erhöhung der Tyrosinphosphorylierung der
Proteine P62, P68 und P130 hemmen (Negrescu et al., 1995). Eine Vorinkubation der
gewaschenen Thrombozytensuspension mit Tyrphostin A23 (250 µM, 20 min) hemmte den
shape change induziert durch YFLLRNP und führte nur zu einer geringen Erhöhung der Syk-
Tyrosinphosphorylierung (1,2-fach) im Vergleich zu 4,3-fach in den unbehandelten Proben,
insgesamt zu einer Reduktion um 96%. Eine Vorbehandlung der gewaschenen Thrombozyten
mit Genistein (150 µM, 3 min) zeigte nach Stimulation mit YFLLRNP eine Reduktion der
Tyrosinphosphorylierung von Syk um 62,0 ± 1,9% (n = 3). Die inaktiven Strukturanaloge
Daidzein (für Genistein) und Tyrphostin A1 (für Tyrphostin A23) zeigten keinen Effekt auf
den shape change und die Tyrosinphosphorylierung von P62, P68, P130 und Syk nach
Stimulation mit YFLLRNP.
Das Prostazyklin-Analog Iloprost erhöht den cAMP-Spiegel um das 5 - 7-fache, verhindert
den Plättchen shape change und inhibierte die durch YFLLRNP induzierte
Tyrosinphosphorylierung von P62, P68 und P130 (Negrescu et al., 1995). Gewaschene
Thrombozyten wurden mit Iloprost (2 nM, 2 min) vorinkubiert und anschließend mit
YFLLRNP bzw. mox-LDL stimuliert. Neben einer Hemmung des shape change war auch die
Stimulation der Tyrosinphosporylierung der Tyrosinkinase Syk reduziert, nach YFLLRNP um
74,7 ± 17,5% (n = 3) und nach mox-LDL um 39,3 ± 26,1% (n = 3). Ohne Zusatz von Iloprost
war der Anstieg der Tyrosinphosphorylierung nach YFLLRNP 3,4-fach und nach mox-LDL
2,7-fach.

57
3.3 Funktion der Syk-Tyrosinphosphorylierung für den shape change
3.3.1 Untersuchungen an mit EGTA vorbehandelten Thrombozyten
Die Behandlung von Thrombozyten mit EGTA (5 mM, 20 min) führt zur Dissoziation des
Integrins αIIbβ3, einem Anstieg der Tyrosinphosphorylierung der Proteine P62, P68, P130
und zur Syk-Stimulierung, ohne daß die Thrombozyten aktiviert werden (Negrescu und Siess,
1996). Nach EGTA-Vorbehandlung kann ein unveränderter shape change durch YFLLRNP
bzw. mox-LDL ausgelöst werden (Negrescu und Siess, 1996). In der folgenden Abbildung
(Abb. 13) sind die Veränderungen der Tyrosinphosphorylierung von Syk nach EGTA-
Behandlung und einer anschließenden Stimulation durch YFLLRNP zusammengestellt.
Die EGTA-Vorinkubation allein führte zu einer nahezu 5-fachen Erhöhung der
Tyrosinphosphorylierung von Syk. Nach einer auf eine EGTA-Vorinkubation folgenden
Stimulation mit YFLLRNP wurde ein shape change induziert, es war jedoch kein weiterer
signifikanter Anstieg der Tyrosinphosphorylierung von P62, P68 und P130 (Daten nicht
gezeigt) sowie von Syk festzustellen (Abb. 13).
Diese Ergebnisse zeigen, daß Syk zwar aktiviert wird, aber funktionell für den shape change
keine Rolle spielt.
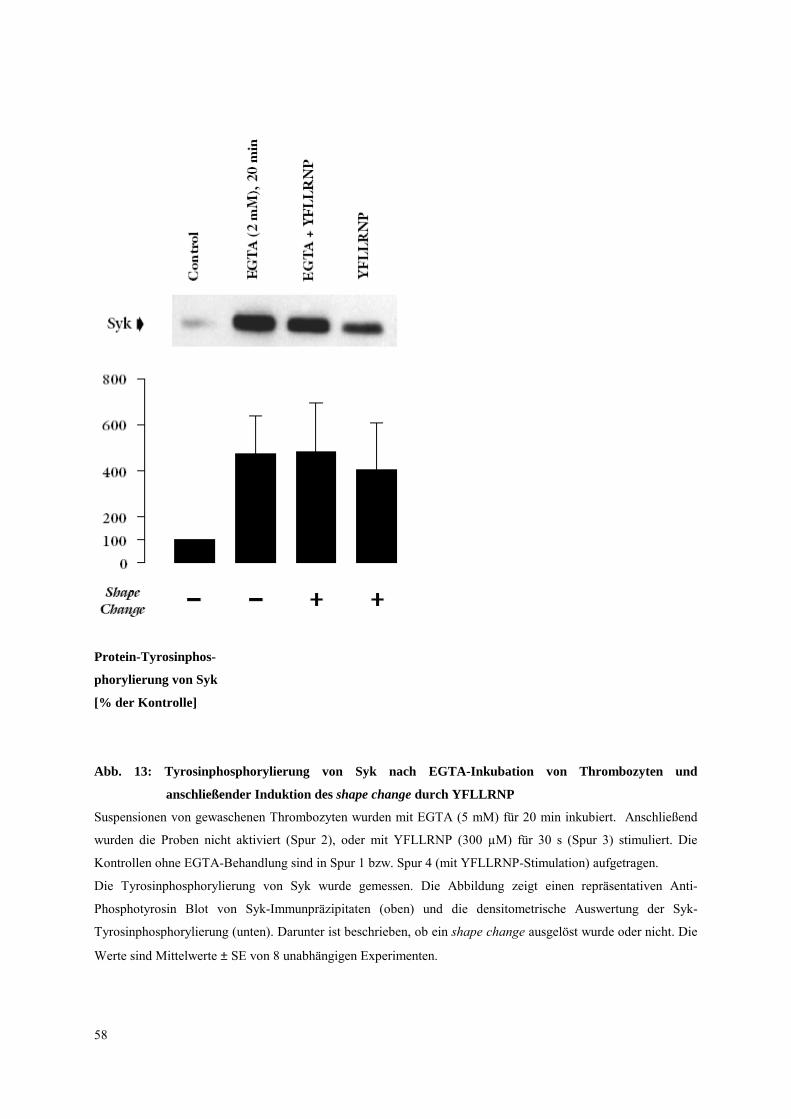
58
Protein-Tyrosinphos-
phorylierung von Syk
[% der Kontrolle]
Abb. 13: Tyrosinphosphorylierung von Syk nach EGTA-Inkubation von Thrombozyten und
anschließender Induktion des shape change durch YFLLRNP
Suspensionen von gewaschenen Thrombozyten wurden mit EGTA (5 mM) für 20 min inkubiert. Anschließend
wurden die Proben nicht aktiviert (Spur 2), oder mit YFLLRNP (300 µM) für 30 s (Spur 3) stimuliert. Die
Kontrollen ohne EGTA-Behandlung sind in Spur 1 bzw. Spur 4 (mit YFLLRNP-Stimulation) aufgetragen.
Die Tyrosinphosphorylierung von Syk wurde gemessen. Die Abbildung zeigt einen repräsentativen Anti-
Phosphotyrosin Blot von Syk-Immunpräzipitaten (oben) und die densitometrische Auswertung der Syk-
Tyrosinphosphorylierung (unten). Darunter ist beschrieben, ob ein shape change ausgelöst wurde oder nicht. Die
Werte sind Mittelwerte ± SE von 8 unabhängigen Experimenten.

59
Da die durch EGTA induzierte Tyrosinphosphorylierung von Syk Schwankungen unterlag,
die eventuell durch die Gegenwart von Spuren von Plasma in der Suspension gewaschener
Thrombozyten verursacht worden sein könnten, wurde die Wirkung von Plasma auf die
Tyrosinphosphorylierung von Syk, P62, P68 und P130 untersucht. Es wurde vermutet, daß
Plasma eine gewisse Schutzfunktion auf das Integrin αIIbβ3 ausübt.
Nach EGTA-Behandlung war zunächst in allen Proben (Plasma 0 - 2%) eine Stimulation von
P62, P68 und Syk festzustellen (Abb. 14). Plasma bewirkte vor allem im
Konzentrationsbereich von 0,5% - 2% eine Verminderung der Tyrosinphosphorylierung einer
Bande unterhalb von P68 (s. Abb. 14 Pfeil) als auch eine leichte Abschwächung der Syk-
Tyrosinphosphorylierung nach EGTA-Behandlung. Die Verminderung der Bande unterhalb
von P68 ist wahrscheinlich durch eine Interferenz des Western Blots mit Albumin erklärbar.
Albumin hat ebenfalls eine Molekülmasse von etwa 68 kD wie P68 und es ist wahrscheinlich,
daß Albumin die tyrosinphosphorylierte Bande unterhalb von P68 auf dem Blot überlagert
und eine Bindung des Antikörpers stört.
anti-Syk
anti-PYSyk
Syk
Plasma [%]
Plasma [%]
Syk-IP
anti-PY
P130
P68
P62
Kontrolle EGTA
0
0
0
0
0,25
0,25
0,25
0,25
0,5
0,5
0,5
0,5
1
1
1
1
2
2
2
2
Abb. 14: Einfluß von Plasma im Thrombozytensuspensionsmedium auf die basale und durch EGTA stimulierte Tyrosinphosphorylierung in Thrombozyten
Suspensionen gewaschener humaner Thrombozyten wurden in Gegenwart von Plasma (0,25 - 2%) mit oder ohne
5 mM EGTA für 20 min bei 37 °C inkubiert und die Tyrosinphosphorylierung von P62, P68, P130 bzw. von Syk
gemessen.

60
3.3.2 Einfluß von allgemeinen und spezifischen Inhibitoren der Src-
Kinasen auf Syk-Tyrosinphosphorylierung und shape change
Es wurden zwei strukturell nicht verwandte spezifische Inhibitoren von Src-Kinasen PP1 und
PD173956 (Hanke et al., 1996) eingesetzt. Mit den selektiven Src-Kinase Inhibitoren PP1 und
PD173956 wurden Untersuchungen zur Rolle der Tyrosinkinase Syk für den shape change
durchgeführt. Wie Bauer et al. fanden, wird der shape change induziert durch
Thrombinrezeptoraktivierung mit YFLLRNP oder ADP (gemessen im Aggregometer) durch
diese Inhibitoren nicht gehemmt (Bauer et al., 2001).
Die folgenden Färbungen von F-Aktin in humanen Thrombozyten (s. Abb. 15) zeigen, daß
auch der morphologisch gemessene Formwandel durch die Vorbehandlung der Thrombozyten
mit Src-Kinase Inhibitoren unbeeinflußt blieb. Deutlich ist zunächst die Verkleinerung der
diskoiden Thrombozyten, die unregelmäßige Oberfläche, einzelne Pseudopodien und die
Zunahme der Intensität der Fluoreszenz nach Stimulation der Thrombozyten mit YFLLRNP,
was auf eine Zunahme und Umverteilung von F-Aktin während des Formwandels
zurückzuführen ist (vgl. Abb. 15A, B). Diese morphologischen Veränderungen waren durch
Vorbehandlung mit PP1 und PD 173956 unbeeinflußt (Abb. 15C, D).
Bei einer Stimulation der Thrombozyten mit mox-LDL bzw. LPA war der shape change nach
Zusatz der Src-Kinase Inhibitoren nicht gehemmt (Abb. 16A). Es wurde jedoch gefunden, daß
die Tyrosinphosphorylierung der Proteine P62, P68 und P130 (Abb. 16B) sowie von Syk
(Abb. 16C) nach entsprechender Vorinkubation mit PP1 bzw. PD173956 komplett gehemmt
war.
Trotz vollständiger Hemmung der Stimulation der Tyrosinphosphorylierung von Syk und der
Proteine P62, P68 und P130 durch die Src-Kinase Inhibitoren PP1 und PD173956 kommt es
zu einer vollständigen Ausbildung des shape change, induziert durch YFLLRNP, mox-LDL
oder LPA. Syk wird zwar während des shape change vermehrt an Tyrosin phosphoryliert,
diese Tyrosinphosphorylierung ist jedoch nicht essentiell für die Ausbildung dieser frühen
Thrombozytenantwort.
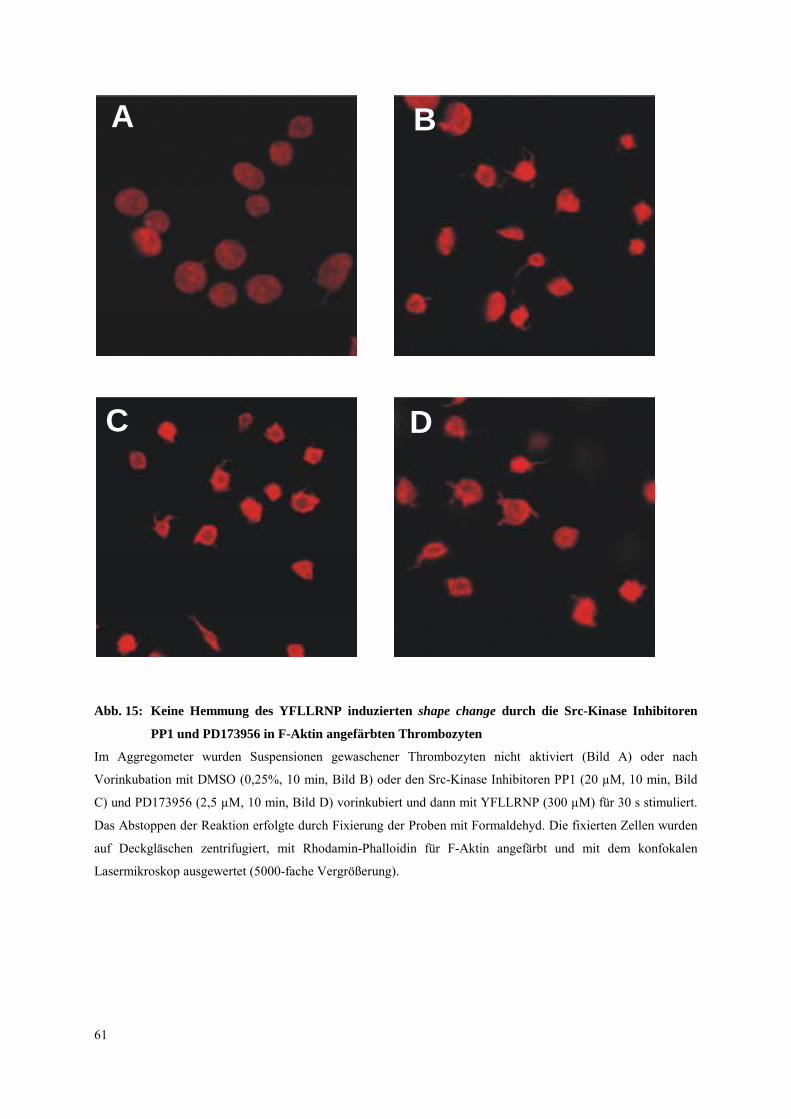
61
A B
C D
Abb. 15: Keine Hemmung des YFLLRNP induzierten shape change durch die Src-Kinase Inhibitoren
PP1 und PD173956 in F-Aktin angefärbten Thrombozyten
Im Aggregometer wurden Suspensionen gewaschener Thrombozyten nicht aktiviert (Bild A) oder nach
Vorinkubation mit DMSO (0,25%, 10 min, Bild B) oder den Src-Kinase Inhibitoren PP1 (20 µM, 10 min, Bild
C) und PD173956 (2,5 µM, 10 min, Bild D) vorinkubiert und dann mit YFLLRNP (300 µM) für 30 s stimuliert.
Das Abstoppen der Reaktion erfolgte durch Fixierung der Proben mit Formaldehyd. Die fixierten Zellen wurden
auf Deckgläschen zentrifugiert, mit Rhodamin-Phalloidin für F-Aktin angefärbt und mit dem konfokalen
Lasermikroskop ausgewertet (5000-fache Vergrößerung).
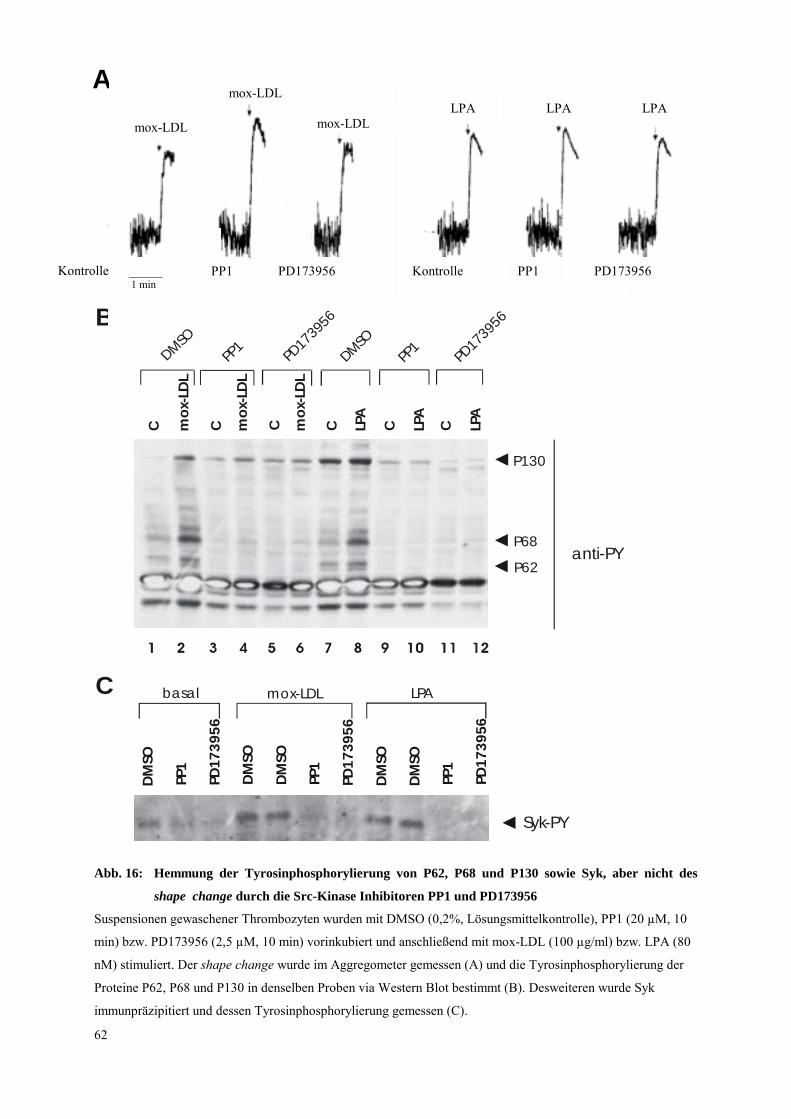
62
DMSO
mo
x-LD
L
C
basal
DM
SO
PP1
B
C
Abb. 16: Hemmung
shape chan
Suspensionen gewasche
min) bzw. PD173956 (
nM) stimuliert. Der sha
Proteine P62, P68 und
immunpräzipitiert und
A
mox-LDL
P
mox-LDL
PP1
PD
mo
x-LD
L
CC
mox-
DM
SO
DM
SO
PD17
3956
der Tyrosinph
ge durch die S
ner Thrombozy
2,5 µM, 10 min
pe change wurd
P130 in denselb
dessen Tyrosinp
mox-LDLLPA LPA LPA
P1 PD1
73956 Kontrolle PP1 PD173956 Kontrolle _____ 1 minanti-PY
P130
P68
P62
DMSOPP
11739
56
PD17
3956
LPA
LPA
LPA
mo
x-LD
L
C CC
LDL LPA
DM
SO
DM
SO
PP1
PP1
PD17
3956
PD17
3956
Syk-PY
osphorylierung von P62, P68 und P130 sowie Syk, aber nicht des
rc-Kinase Inhibitoren PP1 und PD173956
ten wurden mit DMSO (0,2%, Lösungsmittelkontrolle), PP1 (20 µM, 10
) vorinkubiert und anschließend mit mox-LDL (100 µg/ml) bzw. LPA (80
e im Aggregometer gemessen (A) und die Tyrosinphosphorylierung der
en Proben via Western Blot bestimmt (B). Desweiteren wurde Syk
hosphorylierung gemessen (C).

63
3.3.3 Untersuchungen an Syk(-/-)-Maus-Thrombozyten
Um die funktionelle Rolle von Syk für den shape change definitiv aufzuklären, wurden
Untersuchungen an Syk-defizienten Maus-Thrombozyten durchgeführt. Es wurde gefunden,
daß in Syk-defizienten Maus-Thrombozyten ein normaler shape change auslösbar war und die
Stimulation der Tyrosinphosphorylierung der Proteine P62, P68 und P130 vergleichbar war
mit den Ergebnissen in Wildtypkontrollen (Abb. 17 und Bauer et al., 2001). Die Stimulation
der Thrombozyten erfolgte mit dem Thromboxan A2-Rezeptor-Agonisten U46619 (100 nM).
Die beschriebenen Resultate zeigen, daß Syk zwar aktiviert wird, aber nicht funktionell
während des shape change involviert ist. Darüberhinaus lassen die Experimente mit den
Inhibitoren PP1 und PD173956 den Schluß zu, daß Syk downstream von Src-Kinasen
aktiviert wird. Weiterhin zeigen Ergebnisse mit den Syk knock-out Maus-Thrombozyten, daß
die Tyrosinkinase Syk nicht an der Tyrosinphosphorylierung der Proteine P62, P68 und P130
beteiligt ist.
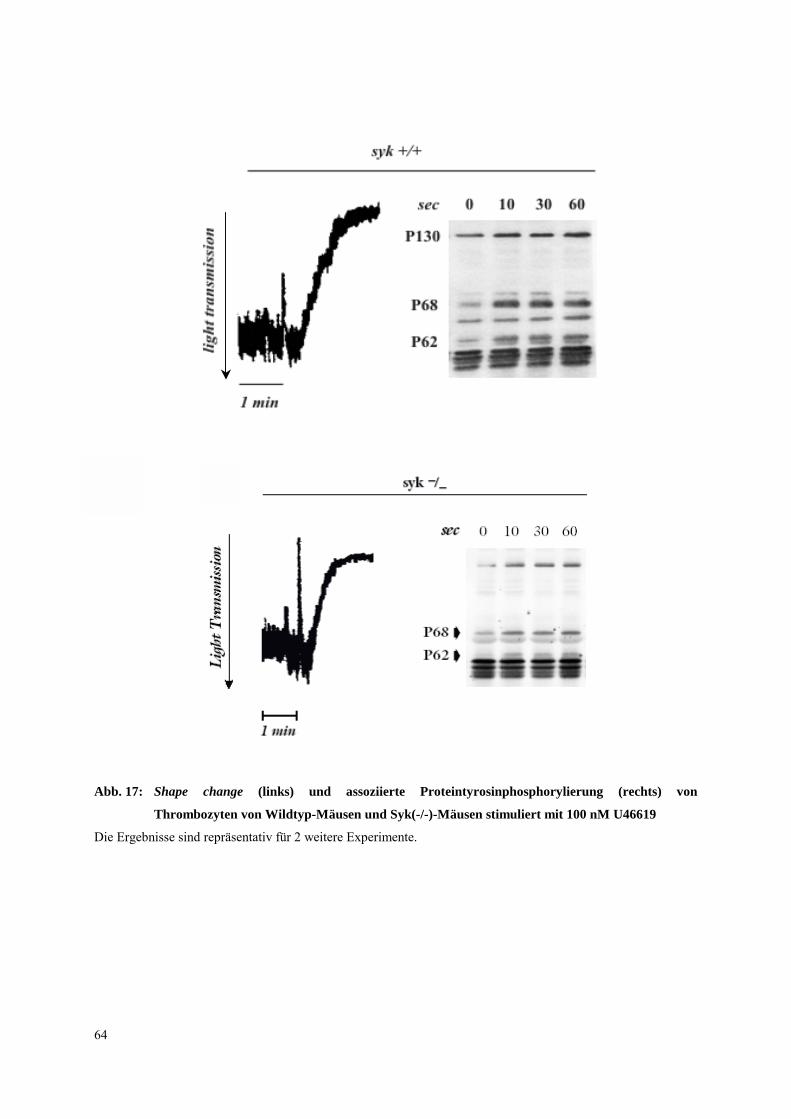
64
Abb. 17: Shape change (links) und assoziierte Proteintyrosinphosphorylierung (rechts) von
Thrombozyten von Wildtyp-Mäusen und Syk(-/-)-Mäusen stimuliert mit 100 nM U46619
Die Ergebnisse sind repräsentativ für 2 weitere Experimente.

65
3.4 Integrin ααααIIbββββ3 abhängige Tyrosinphosphorylierung von Syk
3.4.1 Transfizierte CHO-Zellen als Modellsystem für einen ITAM unabhängigen Signalweg der Syk-Aktivierung
CHO-Zellen dienten als Modellsystem, um die Lokalisation der Tyrosinkinase Syk zu
verschiedenen Zeiten während der Zellausbreitung zu untersuchen. In diesem Zelltyp konnte
ein ITAM unabhängiger Signalweg beschrieben werden (Gao et al., 1997). CHO-Zellen
(Zellinie A5) waren stabil mit dem Integrin αIIbβ3 und transient mit der Tyrosinkinase Syk
transfiziert. In Abhängigkeit von der Zeit breiteten sich die CHO-Zellen auf Fibrinogen-
beschichteten Deckgläschen nach Inkubation bei 37°C aus. In
immunfluoreszenzmikroskopischen Untersuchungen wurde die Lokalisation des Proteins Syk
und eventuelle Colokalisationen mit anderen Proteinen zu verschiedenen Zeitpunkten des
spreading bestimmt.
Abb. 18 zeigt die CHO-Zellen in einem frühen Stadium des spreading. Nach 15 min konnte
Syk im Zytosol, aber vor allem auch an der Zellmembran detektiert werden, wobei es sowohl
mit F-Aktin als auch dem Integrin αIIbβ3 colokalisierte. Da es an den Randbereichen von
Zellen häufig zu Artefakterscheinungen aufgrund der Zellabrundungen kommt, wurde ein
Kontrollversuch mit GFP (green fluorescent protein) transfizierten Zellen durchgeführt. Wie
erwartet war das GFP über das gesamte Zytosol verteilt und das Überlagerungsbild zeigte
keine Colokalisation an der Zellmembran (Abb. 19).
Nach vollständiger Ausbreitung der CHO-Zellen (60 min bei 37°C) kommt es zur Ausbildung
von Streßfasern (Färbung von F-Aktin mit Rhodamin-Phalloidin) und fokalen
Adhäsionsstellen, an denen sich das Integrin αIIbβ3 konzentriert (Antikörper gegen das αIIb)
(Abb. 20). Das Protein Syk ist wieder über die gesamte Zelle verteilt und es sind keine
Colokalisationen mit dem Integrin αIIbβ3 mehr zu beobachten. Die Resultate deuten also auf
eine frühe Beteiligung der Tyrosinkinase Syk bei der αIIbβ3-vermittelten Zellausbreitung hin,
wobei das Protein zunächst an der Zellmembran konzentriert und später wieder diffus über die
Zelle verteilt ist.
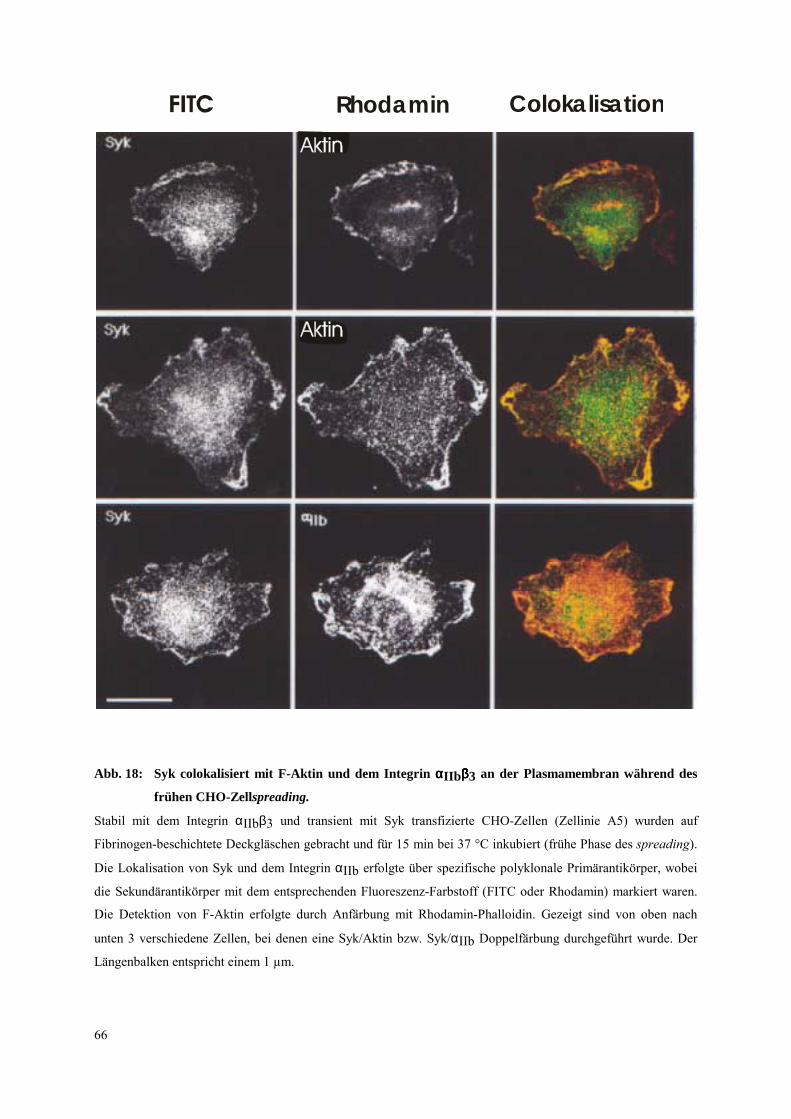
66
Rhodamin Colokalisation
Abb. 18: Syk colokalisiert mit F-Aktin und dem Integrin ααααIIbββββ3 an der Plasmamembran während des
frühen CHO-Zellspreading.
Stabil mit dem Integrin αIIbβ3 und transient mit Syk transfizierte CHO-Zellen (Zellinie A5) wurden auf
Fibrinogen-beschichtete Deckgläschen gebracht und für 15 min bei 37 °C inkubiert (frühe Phase des spreading).
Die Lokalisation von Syk und dem Integrin αIIb erfolgte über spezifische polyklonale Primärantikörper, wobei
die Sekundärantikörper mit dem entsprechenden Fluoreszenz-Farbstoff (FITC oder Rhodamin) markiert waren.
Die Detektion von F-Aktin erfolgte durch Anfärbung mit Rhodamin-Phalloidin. Gezeigt sind von oben nach
unten 3 verschiedene Zellen, bei denen eine Syk/Aktin bzw. Syk/αIIb Doppelfärbung durchgeführt wurde. Der
Längenbalken entspricht einem 1 µm.
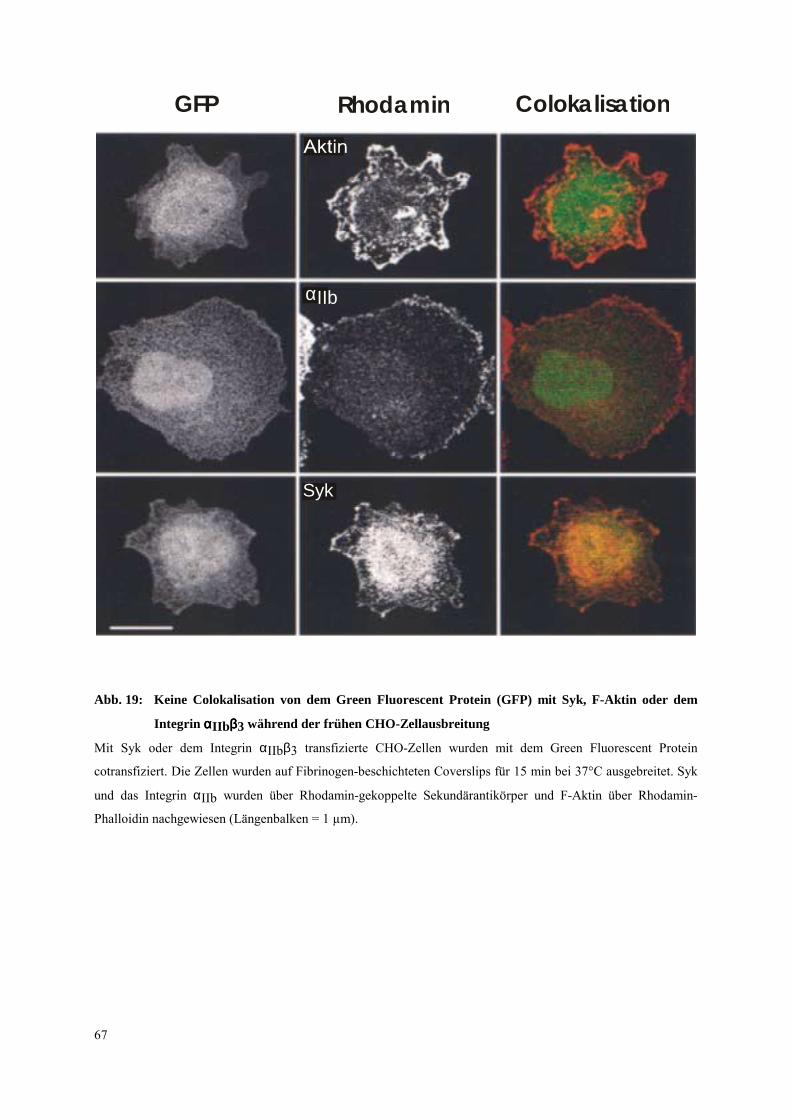
67
GFP Rhodamin Colokalisation
αIIb
Syk
Abb. 19: Keine Colokalisation von dem Green Fluorescent Protein (GFP) mit Syk, F-Aktin oder dem
Integrin ααααIIbββββ3 während der frühen CHO-Zellausbreitung
Mit Syk oder dem Integrin αIIbβ3 transfizierte CHO-Zellen wurden mit dem Green Fluorescent Protein
cotransfiziert. Die Zellen wurden auf Fibrinogen-beschichteten Coverslips für 15 min bei 37°C ausgebreitet. Syk
und das Integrin αIIb wurden über Rhodamin-gekoppelte Sekundärantikörper und F-Aktin über Rhodamin-
Phalloidin nachgewiesen (Längenbalken = 1 µm).
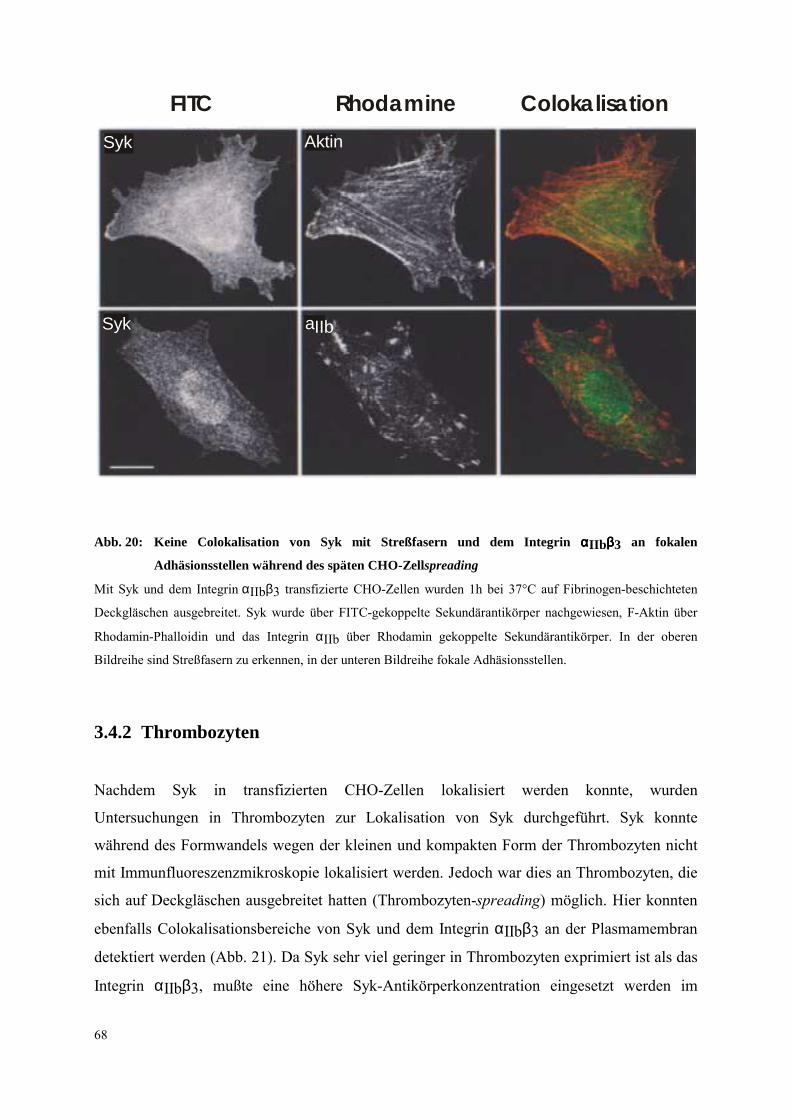
68
FITC Rhodamine Colokalisation
Sk
Syk
Aktin
aIIb
Syk
Abb. 20: Keine Colokalisation von Syk mit Streßfasern und dem Integrin ααααIIbββββ3 an fokalen
Adhäsionsstellen während des späten CHO-Zellspreading
Mit Syk und dem Integrin αIIbβ3 transfizierte CHO-Zellen wurden 1h bei 37°C auf Fibrinogen-beschichteten
Deckgläschen ausgebreitet. Syk wurde über FITC-gekoppelte Sekundärantikörper nachgewiesen, F-Aktin über
Rhodamin-Phalloidin und das Integrin αIIb über Rhodamin gekoppelte Sekundärantikörper. In der oberen
Bildreihe sind Streßfasern zu erkennen, in der unteren Bildreihe fokale Adhäsionsstellen.
3.4.2 Thrombozyten
Nachdem Syk in transfizierten CHO-Zellen lokalisiert werden konnte, wurden
Untersuchungen in Thrombozyten zur Lokalisation von Syk durchgeführt. Syk konnte
während des Formwandels wegen der kleinen und kompakten Form der Thrombozyten nicht
mit Immunfluoreszenzmikroskopie lokalisiert werden. Jedoch war dies an Thrombozyten, die
sich auf Deckgläschen ausgebreitet hatten (Thrombozyten-spreading) möglich. Hier konnten
ebenfalls Colokalisationsbereiche von Syk und dem Integrin αIIbβ3 an der Plasmamembran
detektiert werden (Abb. 21). Da Syk sehr viel geringer in Thrombozyten exprimiert ist als das
Integrin αIIbβ3, mußte eine höhere Syk-Antikörperkonzentration eingesetzt werden im

69
Vergleich zur Antikörperkonzentration gegen das Integrin αIIbβ3. Um nachzuweisen, daß die
Syk-Anfärbung spezifisch war, wurden Kontrollen mit IgG (0,5 µg) bzw. beim anti-Syk (C-
20) Peptid-Antikörper mit dem entsprechenden Peptid (5 µg) durchgeführt, welche kein
Signal bzw. nur ein schwaches Hintergrundsignal aufwiesen.
α βΙΙb 3 Syk
Abb. 21: Colokalisation des Integrins ααααIIbββββ3 und von Syk in ausgebreiteten Thrombozyten
Thrombozyten wurden auf Poly-L-Lysin-beschichtete Deckgläschen zentrifugiert und bei 37°C für 15 min
inkubiert. Die Lokalisation des Integrin αIIbβ3 und von Syk wurde über Immunfluoreszenz ermittelt. Es sind
Doppelfärbungen des Integrins αIIbβ3 und von Syk gezeigt. Die Pfeile zeigen beispielhaft Bereiche der
Colokalisation beider Proteine an der Zellmembran.

70
3.4.2.1 Thrombozyten-Aggregation: FcRγγγγ-Ketten Tyrosinphosphorylierung,
Syk- und PLCγγγγ2-Aktivierung
Um der Frage der Regulation von Syk nach Aktivierung des Integrins αIIbβ3 nachzugehen,
werden Untersuchungen an aggregierenden Thrombozyten durchgeführt. Die
Thrombozytenaggregation wird über die Bindung von Fibrinogen an das Integrin αIIbβ3
vermittelt, welche zu outside-in Signalen wie Tyrosinphosphorylierung und Aktivierung der
Tyrosinkinase Syk führt. Eine Auftrennung von Syk-Immunpräzipitaten von Thrombin-
aggregierten Thrombozyten auf einem Gradientengel zeigte, daß die ITAM-enthaltende
FcRγ�Kette vermehrt tyrosinphosphoryliert wird und mit Syk assoziiert. Dabei zeigte die
tyrosinphosphorylierte FcRγ-Kette eine Doppelbande bei etwa 15 kDa Molekülmasse.
Identifiziert wurde die FcRγ-Kette durch stripping der Membran und durch einen Immunoblot
mit einem polyklonalen Antikörper gegen die FcRγ-Kette. Es wurde gefunden, daß bei der
Verwendung des EDTA enthaltenden Lysis-Puffers für die Syk-Immunpräzipitation, der für
die Pull-down Assays mit dem GST-Syk Tandem-SH2 Fusionsprotein verwendet wurde
(2.2.6), die Tyrosinphosphorylierung deutlich besser nachweisbar war. Die
Tyrosinphosphorylierung von Syk und der assoziierten FcRγ-Kette war 10 s nach Induktion
der Aggregation durch Thrombin detektierbar und war nach 30 s maximal (Abb. 22). Als
Positivkontrolle diente Kollagen, ein starker Stimulus der Tyrosinphosphorylierung von Syk
und der FcRγ-Kette (Gibbins et al., 1996).
Nach einer Vorbehandlung der Blutplättchen mit RGDS, welches die Bindung von Fibrinogen
an das Integrin αIIbβ3 und die Thrombin-induzierte Aggregation hemmt, konnte keine
Tyrosinphosphorylierung der FcRγ-Kette in den Syk-Immunpräzipitaten beobachtet werden
(Abb. 22). Ebenfalls kam es unter diesen Bedingungen bei Pull-down Assays zu keiner
Zunahme an phosphorylierter FcRγ-Kette (Zugabe von RGDS und kein Rühren; Daten nicht
gezeigt). Syk war jedoch tyrosinphosphoryliert, allerdings schwächer als in den aggregierten
Proben (s. Abb. 23, Spur 2 im Vergleich zu Spuren 6 bis 8). Die Anwesenheit der
tyrosinphospho-rylierten FcRγ-Kette in den Syk-Immunpräzipitaten war also abhängig von
der Bindung von Fibrinogen an das Integrin αIIbβ3 und der Thrombozytenaggregation.
Die Stimulus-induzierte Assoziation von Syk und der FcRγ-Kette in aggregierenden
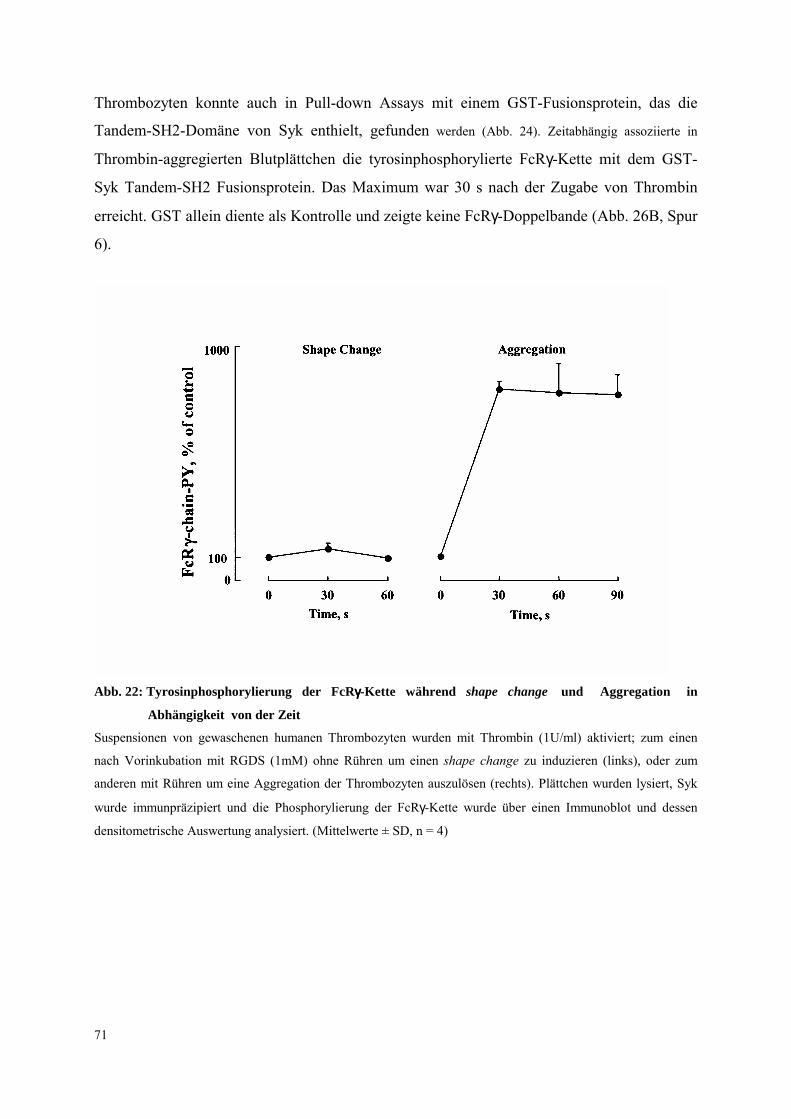
71
Thrombozyten konnte auch in Pull-down Assays mit einem GST-Fusionsprotein, das die
Tandem-SH2-Domäne von Syk enthielt, gefunden werden (Abb. 24). Zeitabhängig assoziierte in
Thrombin-aggregierten Blutplättchen die tyrosinphosphorylierte FcRγ-Kette mit dem GST-
Syk Tandem-SH2 Fusionsprotein. Das Maximum war 30 s nach der Zugabe von Thrombin
erreicht. GST allein diente als Kontrolle und zeigte keine FcRγ-Doppelbande (Abb. 26B, Spur
6).
Abb. 22: Tyrosinphosphorylierung der FcRγγγγ-Kette während shape change und Aggregation in
Abhängigkeit von der Zeit
Suspensionen von gewaschenen humanen Thrombozyten wurden mit Thrombin (1U/ml) aktiviert; zum einen
nach Vorinkubation mit RGDS (1mM) ohne Rühren um einen shape change zu induzieren (links), oder zum
anderen mit Rühren um eine Aggregation der Thrombozyten auszulösen (rechts). Plättchen wurden lysiert, Syk
wurde immunpräzipiert und die Phosphorylierung der FcRγ-Kette wurde über einen Immunoblot und dessen
densitometrische Auswertung analysiert. (Mittelwerte ± SD, n = 4)
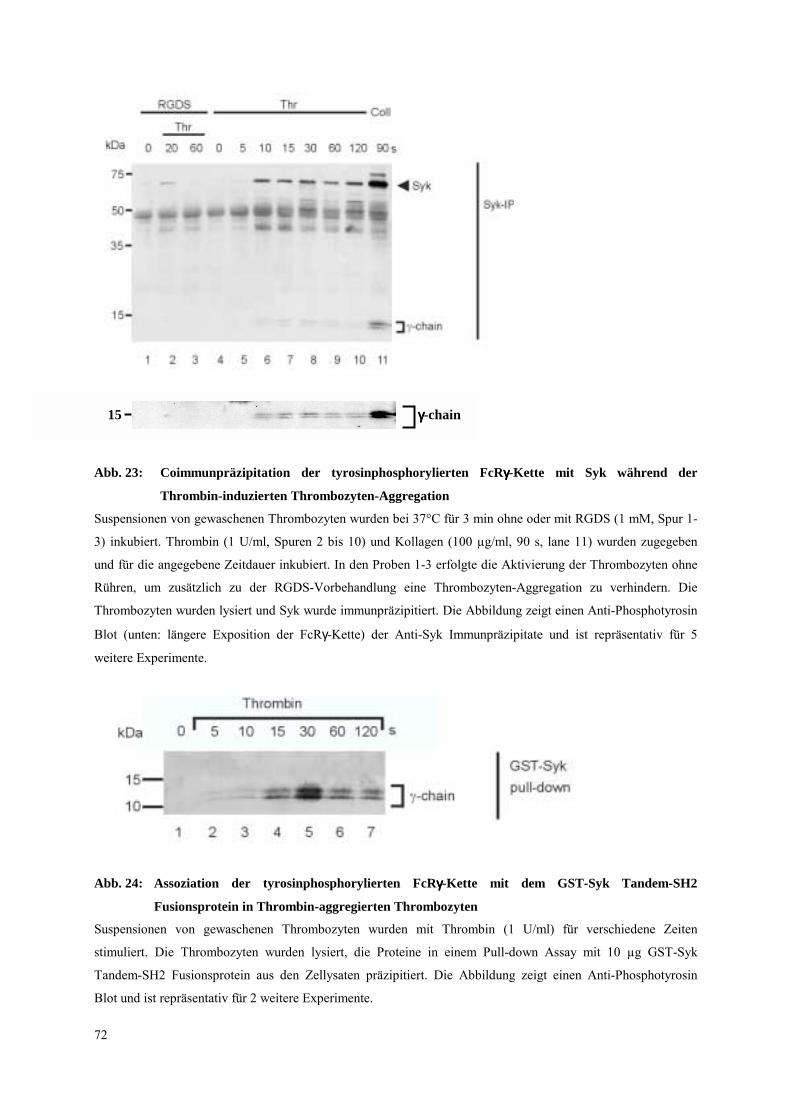
72
Abb. 23: Coimmunpräzipitation der tyrosinphosphorylierten FcRγγγγ-Kette mit Syk während der
Thrombin-induzierten Thrombozyten-Aggregation
Suspensionen von gewaschenen Thrombozyten wurden bei 37°C für 3 min ohne oder mit RGDS (1 mM, Spur 1-
3) inkubiert. Thrombin (1 U/ml, Spuren 2 bis 10) und Kollagen (100 µg/ml, 90 s, lane 11) wurden zugegeben
und für die angegebene Zeitdauer inkubiert. In den Proben 1-3 erfolgte die Aktivierung der Thrombozyten ohne
Rühren, um zusätzlich zu der RGDS-Vorbehandlung eine Thrombozyten-Aggregation zu verhindern. Die
Thrombozyten wurden lysiert und Syk wurde immunpräzipitiert. Die Abbildung zeigt einen Anti-Phosphotyrosin
Blot (unten: längere Exposition der FcRγ-Kette) der Anti-Syk Immunpräzipitate und ist repräsentativ für 5
weitere Experimente.
Abb. 24: Assoziation der tyrosinphosphorylierten FcRγγγγ-Kette mit dem GST-Syk Tandem-SH2
Fusionsprotein in Thrombin-aggregierten Thrombozyten
Suspensionen von gewaschenen Thrombozyten wurden mit Thrombin (1 U/ml) für verschiedene Zeiten
stimuliert. Die Thrombozyten wurden lysiert, die Proteine in einem Pull-down Assay mit 10 µg GST-Syk
Tandem-SH2 Fusionsprotein aus den Zellysaten präzipitiert. Die Abbildung zeigt einen Anti-Phosphotyrosin
Blot und ist repräsentativ für 2 weitere Experimente.
15 γγγγ-chain
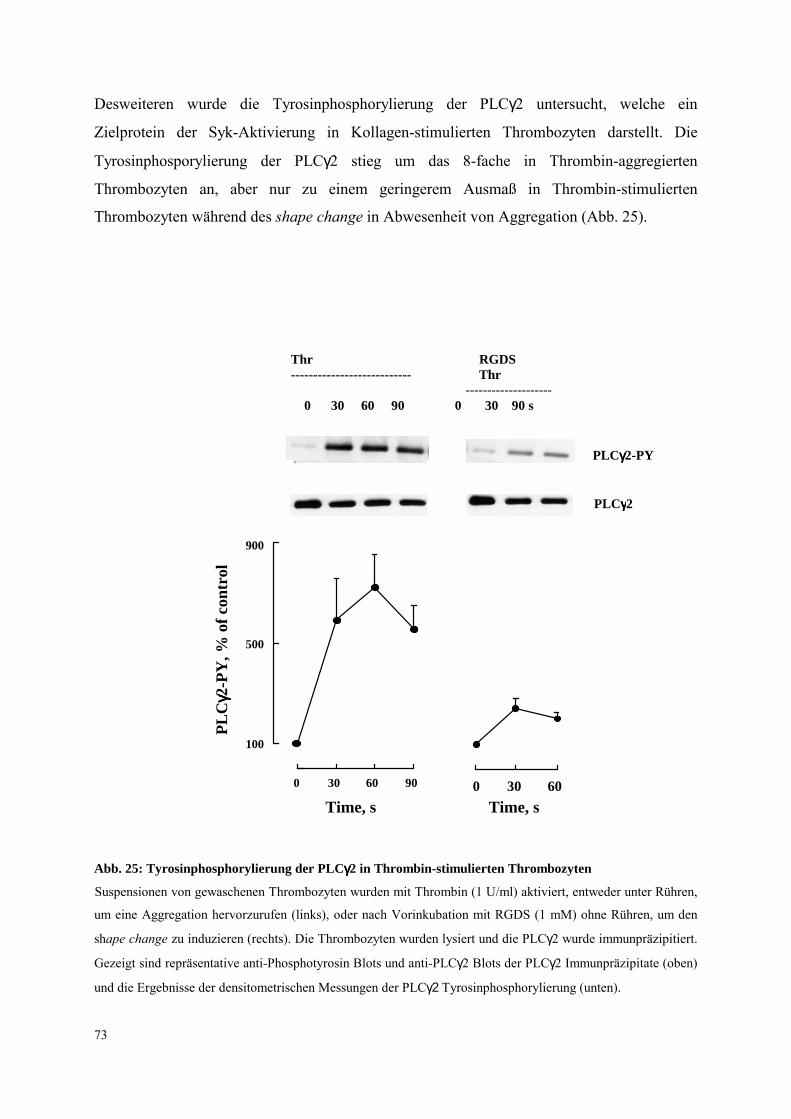
73
Desweiteren wurde die Tyrosinphosphorylierung der PLCγ2 untersucht, welche ein
Zielprotein der Syk-Aktivierung in Kollagen-stimulierten Thrombozyten darstellt. Die
Tyrosinphosporylierung der PLCγ2 stieg um das 8-fache in Thrombin-aggregierten
Thrombozyten an, aber nur zu einem geringerem Ausmaß in Thrombin-stimulierten
Thrombozyten während des shape change in Abwesenheit von Aggregation (Abb. 25).
Abb. 25: Tyrosinphosphorylierung der PLCγγγγ2 in Thrombin-stimulierten Thrombozyten
Suspensionen von gewaschenen Thrombozyten wurden mit Thrombin (1 U/ml) aktiviert, entweder unter Rühren,
um eine Aggregation hervorzurufen (links), oder nach Vorinkubation mit RGDS (1 mM) ohne Rühren, um den
shape change zu induzieren (rechts). Die Thrombozyten wurden lysiert und die PLCγ2 wurde immunpräzipitiert.
Gezeigt sind repräsentative anti-Phosphotyrosin Blots und anti-PLCγ2 Blots der PLCγ2 Immunpräzipitate (oben)
und die Ergebnisse der densitometrischen Messungen der PLCγ2 Tyrosinphosphorylierung (unten).
0 30 60 90 0 30 90 s
Thr RGDSThr
PLC γγγγ2-PY
PLC γγγγ2
9060300
100
500
900
Time, s
PLC
γγ γγ2-P
Y, %
of c
ontr
ol
60300Time, s
PLCγγγγ2-PY
PLCγγγγ2
Thr RGDS --------------------------- Thr -------------------- 0 30 60 90 0 30 90 s

74
Um die Tyrosinkinasen zu identifizieren, welche verantwortlich sind für die
Tyrosinphosporylierung der FcRγ-Kette und der Syk-Aktivierung, wurden die spezifischen
Inhibitoren der Src-Kinasen PP1 und PD173956 verwendet. Es wurde gefunden, daß diese
zwei Inhibitoren die Tyrosinphosphorylierung der FcRγ-Kette in Thrombin-stimulierten
Thrombozyten hemmten. Die Hemmung der Tyrosinphosphorylierung der FcRγ-Kette war in
Syk-Immunpräzipitaten und in GST-Syk Tandem-SH2 Pull-down Assays nachweisbar
(Abb. 26A, B). Außerdem hemmten die Src-Kinase Inhibitoren die Assoziation der FcRγ-
Kette mit der Syk Tandem-SH2-Domäne (Abb. 26B unten), was darauf schließen läßt, daß die
Tyrosinphosphorylierung der FcRγ-Kette erforderlich ist für die Assoziation mit der Tandem-
SH2-Domäne von Syk. Weiterhin war die Tyrosinphosphorylierung von Syk ebenfalls
inhibiert (Abb. 26A).
Zusammengefaßt zeigen diese Ergebnisse, daß die Aktivierung des Integrins αIIbβ3 durch
Fibrinogen in Thrombin-aggregierten Thrombozyten zur Src-Kinasen vermittelten
Tyrosinphosphorylierung der FcRγ-Kette führt, welche dann mit der Tandem-SH2-Domäne
von Syk assoziiert. Anschließend kommt es zu einer vermehrten Syk-Tyrosinphospho-
rylierung, Syk-Aktivierung und Tyrosinphosphorylierung der PLCγ2. Dagegen wird während
des shape change ohne Aggregation die Tyrosinphosphorylierung von Syk und der PLCγ2 nur
gering stimuliert, und zwar durch einen Signalweg, der unabhängig von der FcRγ-Kette ist.
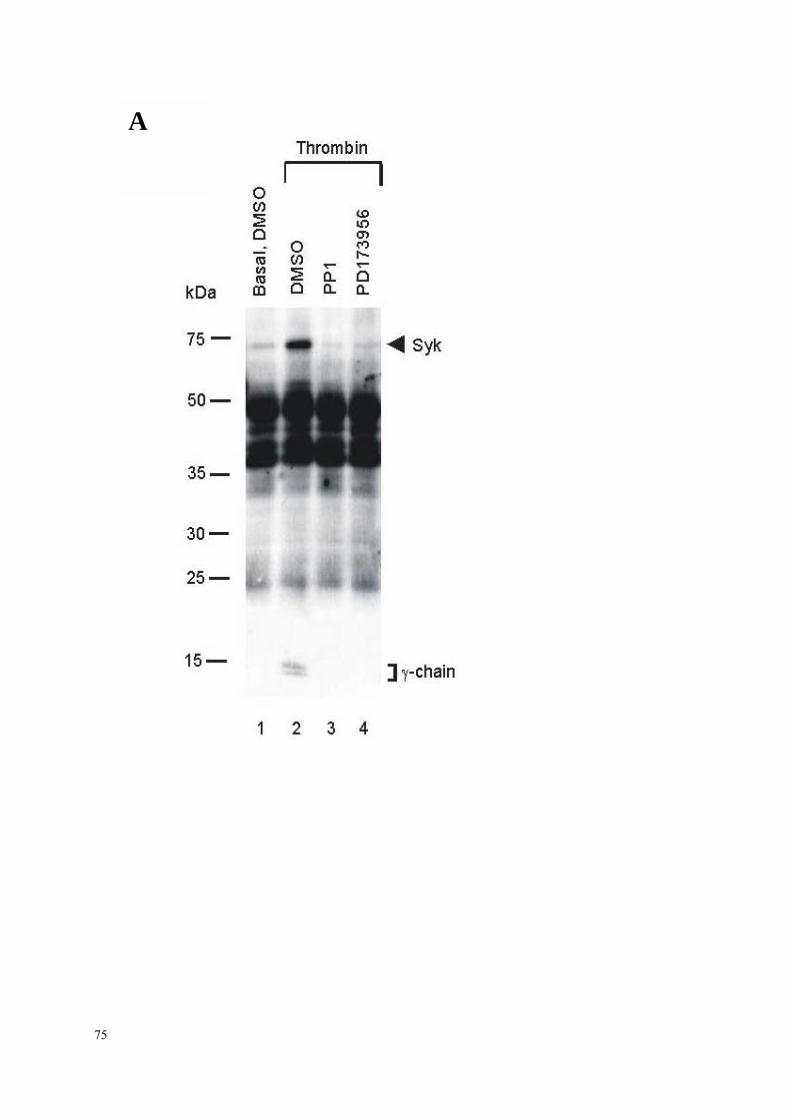
75
A
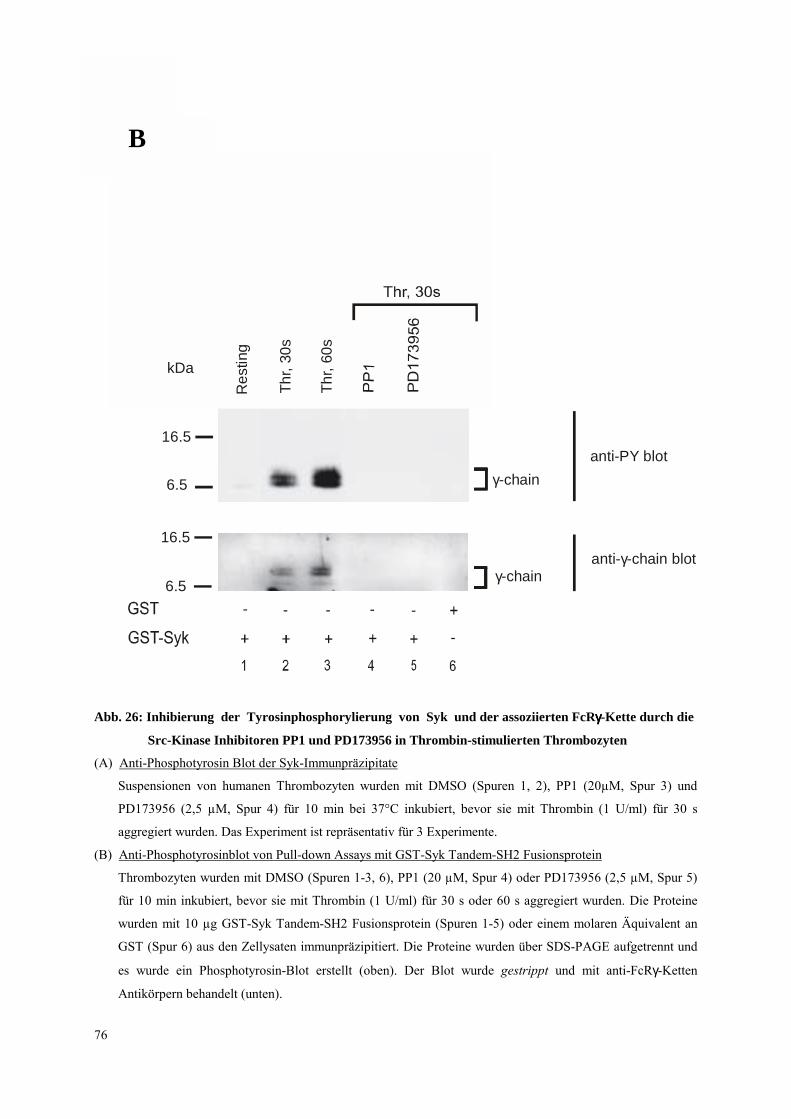
76
Fig. 3B
47.5
anti-PY blotγ-chain
γ-chainanti- -chain blotγ
16.5
16.5
6.5
6.5
32.5
-
- - -- -
kDa
Res
ting
Thr,
30s
Thr,
60s
Abb. 26: Inhibierung der Tyrosinphosphorylierung von Syk und der assoziierten FcRγγγγ-Kette durch die
Src-Kinase Inhibitoren PP1 und PD173956 in Thrombin-stimulierten Thrombozyten
(A) Anti-Phosphotyrosin Blot der Syk-Immunpräzipitate
Suspensionen von humanen Thrombozyten wurden mit DMSO (Spuren 1, 2), PP1 (20µM, Spur 3) und
PD173956 (2,5 µM, Spur 4) für 10 min bei 37°C inkubiert, bevor sie mit Thrombin (1 U/ml) für 30 s
aggregiert wurden. Das Experiment ist repräsentativ für 3 Experimente.
(B) Anti-Phosphotyrosinblot von Pull-down Assays mit GST-Syk Tandem-SH2 Fusionsprotein
Thrombozyten wurden mit DMSO (Spuren 1-3, 6), PP1 (20 µM, Spur 4) oder PD173956 (2,5 µM, Spur 5)
für 10 min inkubiert, bevor sie mit Thrombin (1 U/ml) für 30 s oder 60 s aggregiert wurden. Die Proteine
wurden mit 10 µg GST-Syk Tandem-SH2 Fusionsprotein (Spuren 1-5) oder einem molaren Äquivalent an
GST (Spur 6) aus den Zellysaten immunpräzipitiert. Die Proteine wurden über SDS-PAGE aufgetrennt und
es wurde ein Phosphotyrosin-Blot erstellt (oben). Der Blot wurde gestrippt und mit anti-FcRγ-Ketten
Antikörpern behandelt (unten).
B

77
Um die funktionelle Relevanz des beschriebenen Signalwegs zu untersuchen, wurden
Aggregationsstudien mit den Src-Familie Kinase spezifischen Inhibitoren PP1 und PD173956
durchgeführt.
Die Src-Kinase Inhibitoren hatten eine hemmende Wirkung auf die Thrombin-induzierte
Aggregation. Sowohl PP1 (20 µM) als auch PD173956 (2,5 µM) hemmten die Aggregation
nach niedrigen Thrombin-Konzentrationen (0,05-0,15 U/ml) (Abb. 27). Die Hemmung betrug
40 ± 11% nach Vorinkubation der Plättchen mit 20 µM PP1 und 71 ± 16% nach
Vorbehandlung mit 2,5 µM PD 173956 (Mittelwert ± SD, n=4 Experimente mit
Thrombozyten verschiedener Spender). Der shape change, wie bereits in 3.3.2 ausführlich
beschrieben, war unbeeinflußt von der Inkubation mit den Src-Kinase Inhibitoren PP1 und
PD173956. Wurden hohe Konzentrationen an Thrombin eingesetzt, verhinderten PP1 und
PD173956 die Plättchenaggregation nicht (Abb. 27B).
Die Ergebnisse legen nahe, daß das Integrin αIIbβ3 outside-in signaling zu einer durch die
Src-Familie Kinasen vermittelten Tyrosinphosphorylierung der FcRγ-Kette und deren
Assoziation mit Syk führt. Dadurch kommt es zur Aktivierung von Syk und zur
Tyrosinphosphorylierung der PLCγ2. Dieser Signalweg scheint eine Rolle bei der
Thrombozytenaggregation zu spielen, welche durch niedrige, nicht aber hohe Thrombin-
Konzentrationen ausgelöst wird.
Diese Schlußfolgerung wird durch Studien an Thrombozyten von Syk und FcRγ-Ketten
defizienten Mäusen unterstützt. In drei Experimenten wurden die Aggregationkurven nach
Stimulation mit unterschiedlichen Konzentrationen an Thrombin (0,01 � 0,3 U/ml) in FcRγ-Ketten (-/-)
und Wildtyp-Maus-Thrombozyten verglichen. Tabelle 3 zeigt die Ergebnisse in einer
Übersicht. In Abb. 28 A sind die Aggregationskurven von Versuch 3 abgebildet. Die FcRγ-
Ketten(-/-) und auch Syk (+/-) (Abb. 28B) Maus-Thrombozyten zeigten eine verzögerte und
verringerte Aggregation nach Stimulation mit niedrigen im Vergleich zu höheren
Konzentrationen an Thrombin.
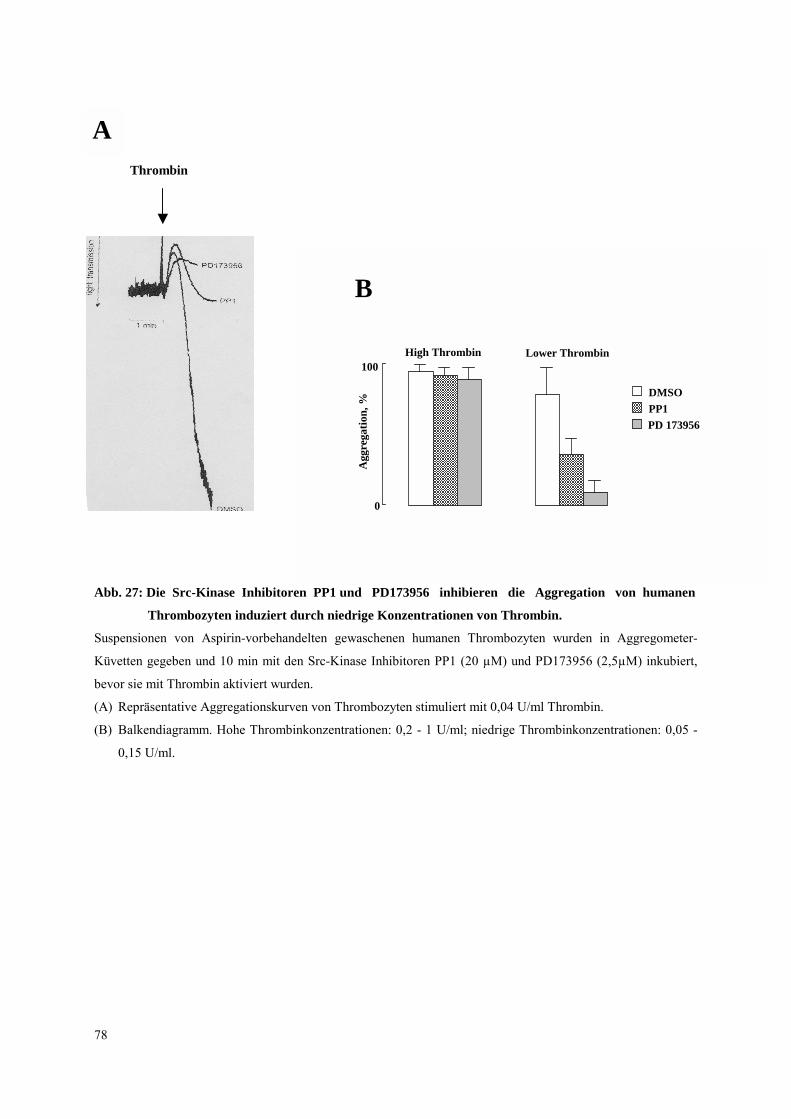
78
Thrombin
Abb. 27: Die Src-Kinase Inhibitoren PP1 und PD173956 inhibieren die Aggregation von humanen
Thrombozyten induziert durch niedrige Konzentrationen von Thrombin.
Suspensionen von Aspirin-vorbehandelten gewaschenen humanen Thrombozyten wurden in Aggregometer-
Küvetten gegeben und 10 min mit den Src-Kinase Inhibitoren PP1 (20 µM) und PD173956 (2,5µM) inkubiert,
bevor sie mit Thrombin aktiviert wurden.
(A) Repräsentative Aggregationskurven von Thrombozyten stimuliert mit 0,04 U/ml Thrombin.
(B) Balkendiagramm. Hohe Thrombinkonzentrationen: 0,2 - 1 U/ml; niedrige Thrombinkonzentrationen: 0,05 -
0,15 U/ml.
0
100
DMSO
Agg
rega
tion,
%High Thrombin Lower Thrombin
PP1,20 µMPD 173956
A
B
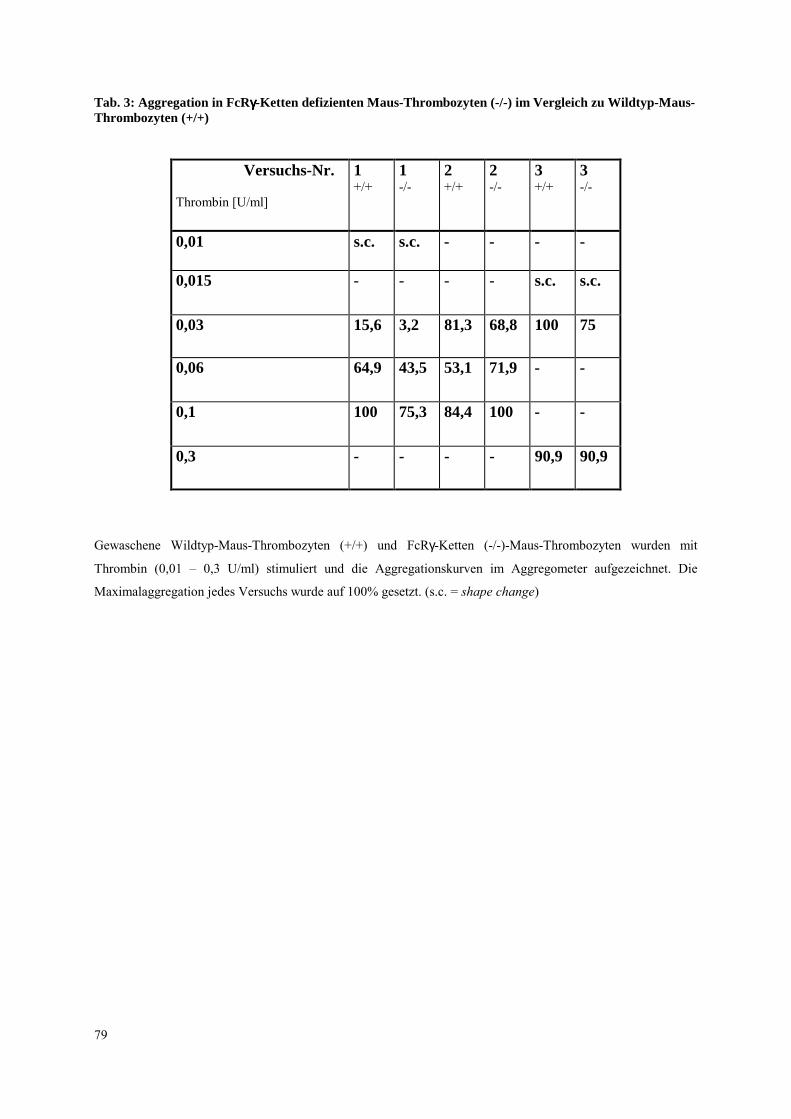
79
Tab. 3: Aggregation in FcRγγγγ-Ketten defizienten Maus-Thrombozyten (-/-) im Vergleich zu Wildtyp-Maus- Thrombozyten (+/+)
Versuchs-Nr. Thrombin [U/ml]
1 +/+
1 -/-
2 +/+
2 -/-
3 +/+
3 -/-
0,01
s.c. s.c. - - - -
0,015 - - - - s.c. s.c.
0,03 15,6 3,2 81,3 68,8 100 75
0,06 64,9 43,5 53,1 71,9 - -
0,1 100 75,3 84,4 100 - -
0,3 - - - - 90,9 90,9
Gewaschene Wildtyp-Maus-Thrombozyten (+/+) und FcRγ-Ketten (-/-)-Maus-Thrombozyten wurden mit
Thrombin (0,01 � 0,3 U/ml) stimuliert und die Aggregationskurven im Aggregometer aufgezeichnet. Die
Maximalaggregation jedes Versuchs wurde auf 100% gesetzt. (s.c. = shape change)

80
Abb. 28: Erniedrigung der thrombin-induzierten Aggregation von defizienten Maus-Thrombozyten ohne
FcRγγγγ-Kette (A) oder Syk (B)
(A) Shape change und Aggregation von Thrombozyten, welche aus Wildtyp- und FcRγ-Ketten (-/-) Mäusen
isoliert wurden, wurden durch verschiedene Konzentration von Thrombin (0,015 U/ml bis 0,3 U/ml)
induziert. Die Kurven der FcRγ-Ketten knock-out Maus-Thrombozyten sind mit (*) gekennzeichnet.
(B) Thrombozyten wurden aus Wildtyp(C5)- und Syk-defizienten(X2, Syk+/-)-Mäusen isoliert und die
Aggregation der Thrombozyten durch verschiedene Konzentrationen an Thrombin (0,05 U/ml bis 0,3 U/ml)
induziert.
A
B 0,05 0,1 0,3 U/ml Thrombin
0,015 0,03 0,3 U/ml Thrombin

81
3.4.2.2 Dissoziation des Integrins ααααIIbββββ3: FcRγγγγ-Ketten Tyrosinphospho-
rylierung, Syk- und PLCγγγγ2-Aktivierung
Da die Dissoziation des Integrins αIIbβ3 durch EGTA-Behandlung der Thrombozyten
(5 mM, 20 min) die Tyrosinphosphorylierung von Syk stimuliert (s. 3.3.1 und Negrescu und
Siess, 1996), war die Frage, ob hier auch die FcRγ-Ketten Tyrosinphosphorylierung involviert
ist. Es wurde gefunden, daß nach EGTA-Behandlung der Thrombozyten (5 mM, 20 min) die
FcRγ-Kette tyrosinphosphoryliert wird und mit Syk coimmunpräzipitiert (Abb. 29). Dabei
war bei Verwendung des EDTA enthaltenden Lysispuffers C (s. 2.2.6) mit Triton X-100
anstelle von NP-40 für die Syk-Immunpräzipitation die Tyrosinphosphorylierung der FcRγ-
Kette deutlich besser nachweisbar.
Desweiteren bewirkte die durch EGTA induzierte Dissoziation des Integrins αIIbβ3 eine
Stimulation der Tyrosinphosphorylierung der PLCγ2 (Abb. 30).
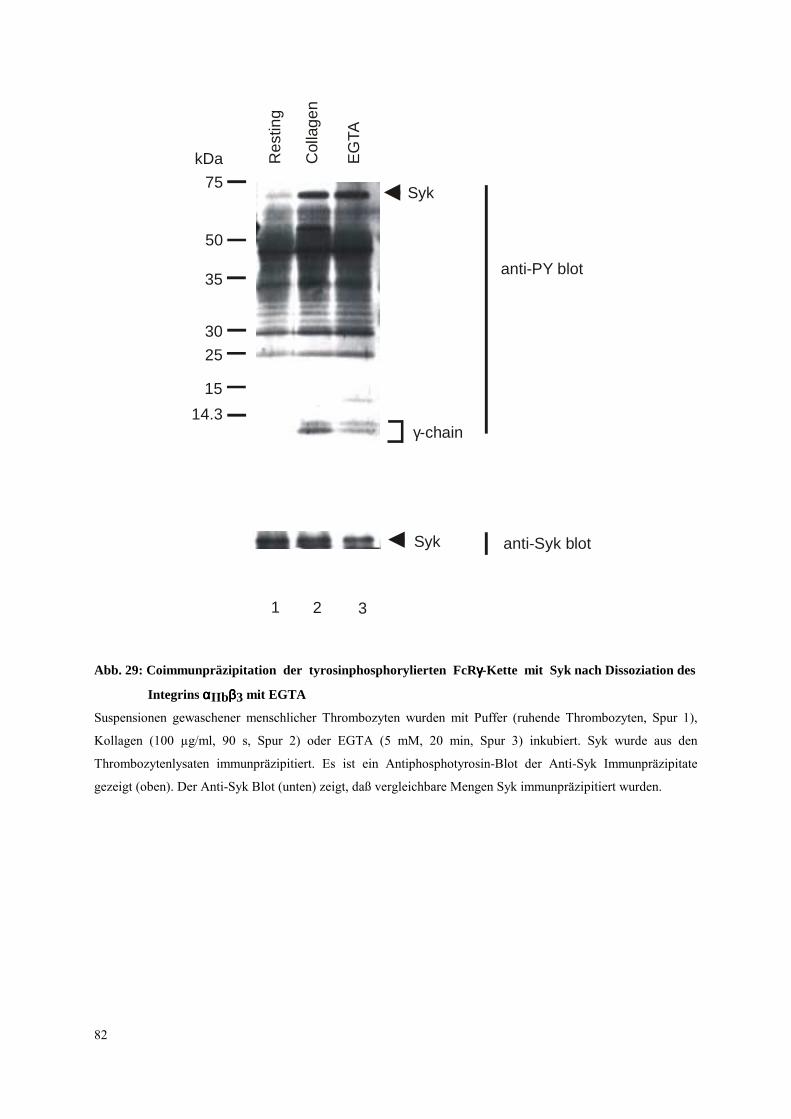
82
Abb. 29: Coimmunpräzipitation der tyrosinphosphorylierten FcRγγγγ-Kette mit Syk nach Dissoziation des
Integrins ααααIIbββββ3 mit EGTA
Suspensionen gewaschener menschlicher Thrombozyten wurden mit Puffer (ruhende Thrombozyten, Spur 1),
Kollagen (100 µg/ml, 90 s, Spur 2) oder EGTA (5 mM, 20 min, Spur 3) inkubiert. Syk wurde aus den
Thrombozytenlysaten immunpräzipitiert. Es ist ein Antiphosphotyrosin-Blot der Anti-Syk Immunpräzipitate
gezeigt (oben). Der Anti-Syk Blot (unten) zeigt, daß vergleichbare Mengen Syk immunpräzipitiert wurden.
Res
ting
Col
lage
n
EGTA
Syk anti-Syk blot
anti-PY blot
γ-chain
Syk
kDa75
50
35
3025
1514.3
1 2 3
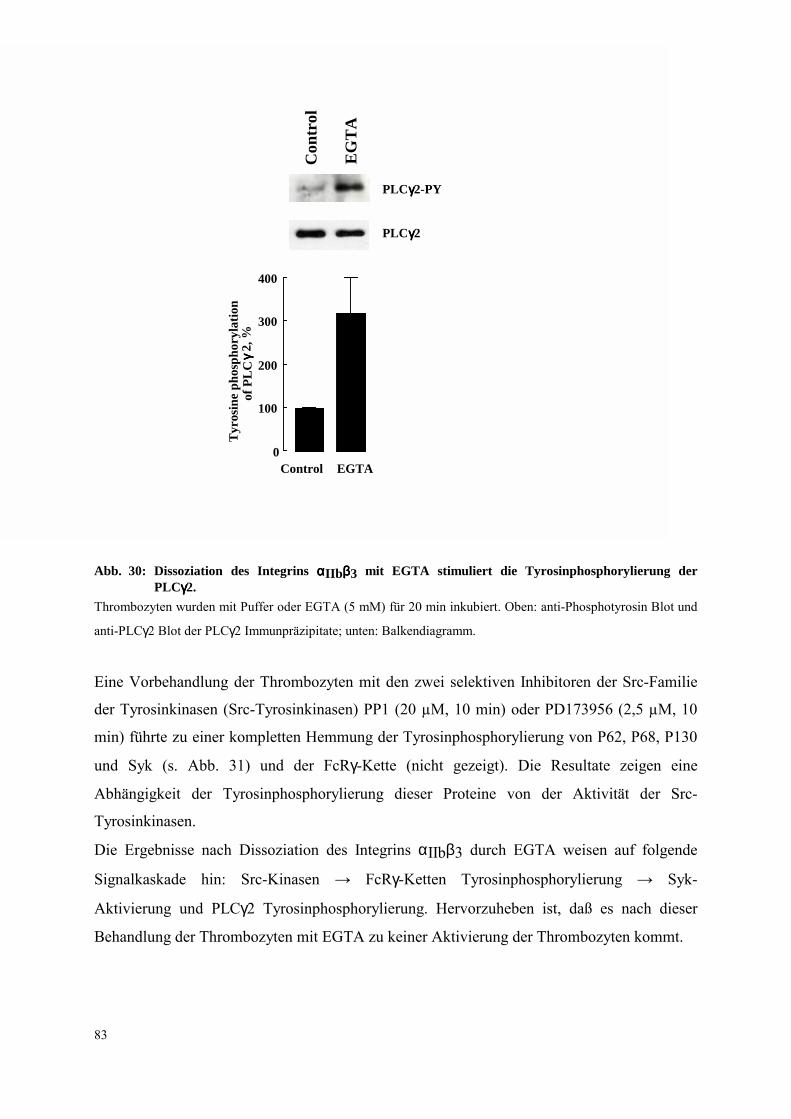
83
Abb. 30: Dissoziation des Integrins ααααIIbββββ3 mit EGTA stimuliert die Tyrosinphosphorylierung der PLCγγγγ2.
Thrombozyten wurden mit Puffer oder EGTA (5 mM) für 20 min inkubiert. Oben: anti-Phosphotyrosin Blot und
anti-PLCγ2 Blot der PLCγ2 Immunpräzipitate; unten: Balkendiagramm.
Eine Vorbehandlung der Thrombozyten mit den zwei selektiven Inhibitoren der Src-Familie
der Tyrosinkinasen (Src-Tyrosinkinasen) PP1 (20 µM, 10 min) oder PD173956 (2,5 µM, 10
min) führte zu einer kompletten Hemmung der Tyrosinphosphorylierung von P62, P68, P130
und Syk (s. Abb. 31) und der FcRγ-Kette (nicht gezeigt). Die Resultate zeigen eine
Abhängigkeit der Tyrosinphosphorylierung dieser Proteine von der Aktivität der Src-
Tyrosinkinasen.
Die Ergebnisse nach Dissoziation des Integrins αIIbβ3 durch EGTA weisen auf folgende
Signalkaskade hin: Src-Kinasen → FcRγ-Ketten Tyrosinphosphorylierung → Syk-
Aktivierung und PLCγ2 Tyrosinphosphorylierung. Hervorzuheben ist, daß es nach dieser
Behandlung der Thrombozyten mit EGTA zu keiner Aktivierung der Thrombozyten kommt.
Control EGTA0
100
200
300
400
Con
trol
EG
TA
Tyro
sine
phos
phor
ylat
ion
of
PL
Cγ γ γ γ
2, %
PLCγγγγ2
PLCγγγγ2-PY
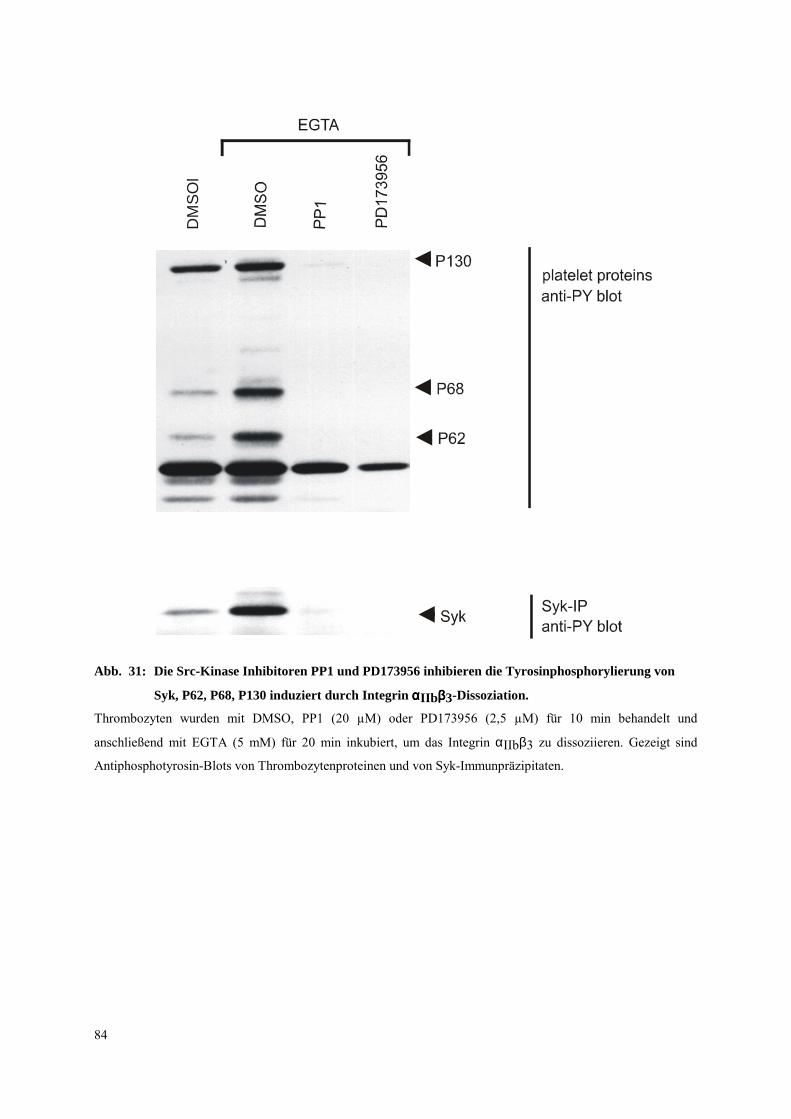
84
Abb. 31: Die Src-Kinase Inhibitoren PP1 und PD173956 inhibieren die Tyrosinphosphorylierung von
Syk, P62, P68, P130 induziert durch Integrin ααααIIbββββ3-Dissoziation.
Thrombozyten wurden mit DMSO, PP1 (20 µM) oder PD173956 (2,5 µM) für 10 min behandelt und
anschließend mit EGTA (5 mM) für 20 min inkubiert, um das Integrin αIIbβ3 zu dissoziieren. Gezeigt sind
Antiphosphotyrosin-Blots von Thrombozytenproteinen und von Syk-Immunpräzipitaten.
DM
SOl

85
3.4.2.3 Ausbreitung
Spreading von humanen Thrombozyten
Thrombozyten, die durch Zentrifugation auf Glas aktiviert und anschließend bei 37°C
inkubiert werden, breiten sich aus (spreading). Dabei ist die Bindung von Fibrinogen an das
Integrin αIIbβ3 essentiell für diesen Prozeß.
Nach Vorbehandlung der Thrombozyten mit dem Fibrinogen-Rezeptor Antagonisten RGDS
war eine deutliche Hemmung des spreading zu erkennen (Abb. 32A, C; F-Aktin-Färbung).
Auch zeigten Versuche mit Thrombozyten von Patienten mit Thrombasthenia Glanzmann,
welchen das Integrin αIIbβ3 fehlt, kein spreading (Abb. 32B, Phasenkontrast). Hier breiteten
sich diese Thrombozyten nicht aus, sondern verblieben im shape change Stadium, zeigten
also kugelige, sphärische Gestalt und Pseudopodien.
Es wurde mittels spezifischer Inhibitoren (Tab. 4) untersucht, ob für die Regulation der
Thrombozyten-Ausbreitung Tyrosinkinasen wichtig sind. Weiterhin wurde die Wirkung von
Hemmern der Proteinkinase C und Rho-Kinase getestet. Dabei wurden sowohl das spreading
fluoreszenzmikroskopisch als auch die Protein-Tyrosinphosphorylierung während des
spreading biochemisch gemessen. Als Kontrolle dienten unstimulierte Thrombozyten in
Suspension.

86
A
CC
A
Abb. 32: Hemmung des spreading bei Thrombasthenia Glanzmann Thrombozyten bzw. nach RGDS-
Behandlung
Suspensionen von gewaschenen humanen Thrombozyten eines normalen Blutspenders (A, Aktinfärbung) und
von einem Patienten mit Thrombasthenia Glanzmann (B, Phasenkontrast-Bild) wurden auf Poly-L-Lysin
beschichtete Deckgläschen zentrifugiert (250 x g, 5 min) und für 15 min bei 37°C inkubiert. C: Vorinkubation
der Thrombozyten für 3 min mit RGDS (1mM) (Aktinfärbung).
87
Tab. 4: Agentien, deren Effekt auf das spreading von humanen Thrombozyten untersucht wurde, und die
jeweils im Experiment eingesetzte Konzentration und Einwirkzeit
Agentien Eigenschaften Konzentration Zeit
FR-1 Fibrinogenrezeptor Antagonist 15 µM 3 min
PD 173956 Src-Kinase Hemmer 2,5 µM 10 min
PP1 Src-Kinase Hemmer 20 µM 10 min
Ro-31-8220 Protein Kinase C Hemmer 0,2 µM, 2 µM 20 min
BAPTA-AM
EGTA Hemmung des zytosolischen Ca2+ 20 µM
1 mM
20 min
1 min
Y-27632 Rho-Kinase Hemmer 20 µM 30 min
Genistein allgemeiner Tyrosinkinasehemmer 150 µM 3 min
Die verschiedenen Hemmsubstanzen wurden mit Suspensionen gewaschener Thrombozyten
(400000 Zellen / µl) wie in der Tabelle 4 beschrieben bei 37°C inkubiert. Vor dem spreading-
Versuch wurden Proben abgenommen und die Tyrosinphosphorylierung der
Thrombozytenproteine gemessen. Anschließend wurde die Thrombozytensuspension auf eine
Konzentration von 15000 / µl verdünnt und auf mit Poly-L-Lysin-beschichtete Deckgläschen
(24 x 32mm) 5 min zentrifugiert.
Den Einfluß mancher der verschiedenen in Tab. 4 beschriebenen Agentien auf die
Morphologie der Thrombozyten während des spreading ist in Abb. 33 gezeigt. Die
Proteintyrosinphosphorylierung ist in Abb. 34 dargestellt.
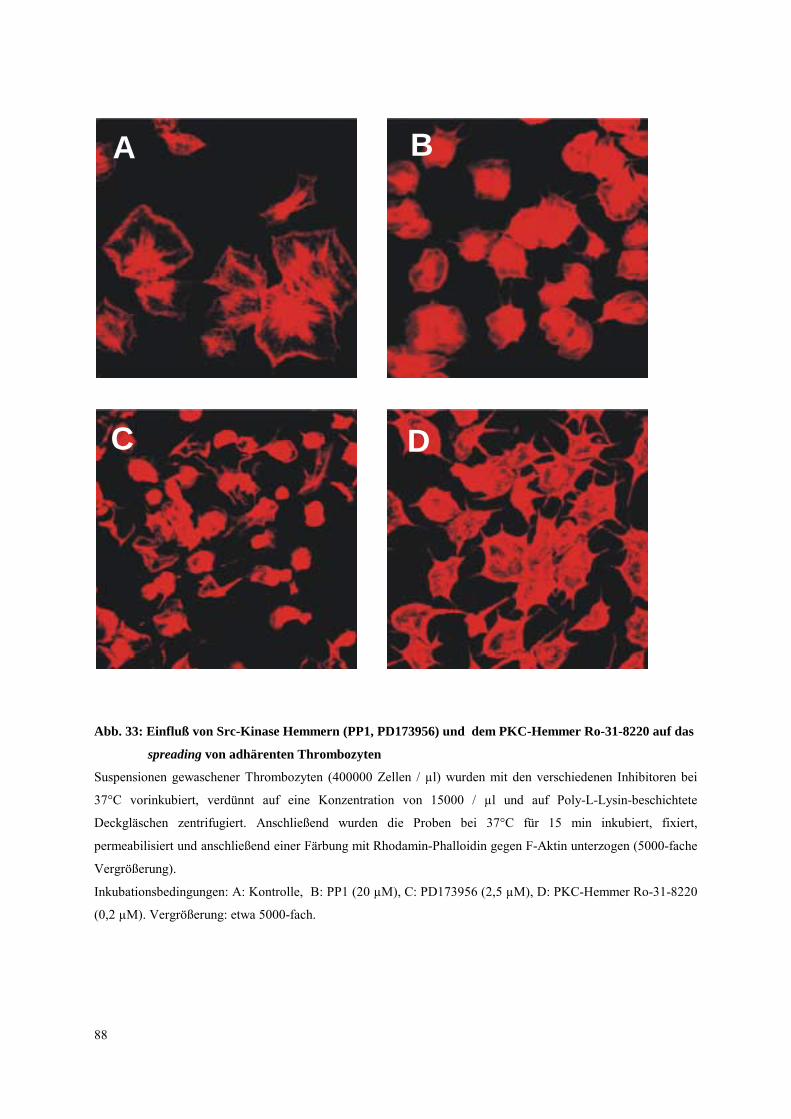
88
A
C D
B
Abb. 33: Einfluß von Src-Kinase Hemmern (PP1, PD173956) und dem PKC-Hemmer Ro-31-8220 auf das
spreading von adhärenten Thrombozyten
Suspensionen gewaschener Thrombozyten (400000 Zellen / µl) wurden mit den verschiedenen Inhibitoren bei
37°C vorinkubiert, verdünnt auf eine Konzentration von 15000 / µl und auf Poly-L-Lysin-beschichtete
Deckgläschen zentrifugiert. Anschließend wurden die Proben bei 37°C für 15 min inkubiert, fixiert,
permeabilisiert und anschließend einer Färbung mit Rhodamin-Phalloidin gegen F-Aktin unterzogen (5000-fache
Vergrößerung).
Inkubationsbedingungen: A: Kontrolle, B: PP1 (20 µM), C: PD173956 (2,5 µM), D: PKC-Hemmer Ro-31-8220
(0,2 µM). Vergrößerung: etwa 5000-fach.
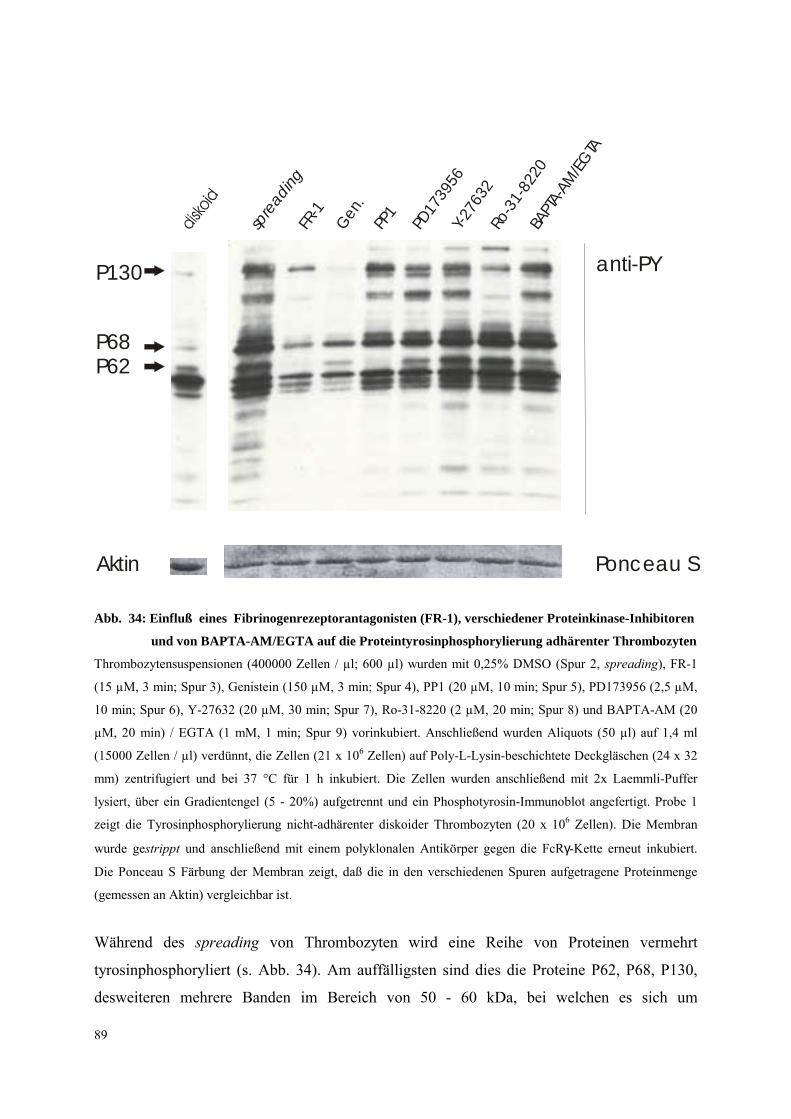
89
anti-PY
Ponceau SAktin
P130
P68P62
spre
adin
gFR
-1
Gen
.PP
1
PD17
3956
Y-27
632
Ro-3
1-82
20BA
PTA-
AM/E
GTA
Abb. 34: Einfluß eines Fibrinogenrezeptorantagonisten (FR-1), verschiedener Proteinkinase-Inhibitoren
und von BAPTA-AM/EGTA auf die Proteintyrosinphosphorylierung adhärenter Thrombozyten
Thrombozytensuspensionen (400000 Zellen / µl; 600 µl) wurden mit 0,25% DMSO (Spur 2, spreading), FR-1
(15 µM, 3 min; Spur 3), Genistein (150 µM, 3 min; Spur 4), PP1 (20 µM, 10 min; Spur 5), PD173956 (2,5 µM,
10 min; Spur 6), Y-27632 (20 µM, 30 min; Spur 7), Ro-31-8220 (2 µM, 20 min; Spur 8) und BAPTA-AM (20
µM, 20 min) / EGTA (1 mM, 1 min; Spur 9) vorinkubiert. Anschließend wurden Aliquots (50 µl) auf 1,4 ml
(15000 Zellen / µl) verdünnt, die Zellen (21 x 106 Zellen) auf Poly-L-Lysin-beschichtete Deckgläschen (24 x 32
mm) zentrifugiert und bei 37 °C für 1 h inkubiert. Die Zellen wurden anschließend mit 2x Laemmli-Puffer
lysiert, über ein Gradientengel (5 - 20%) aufgetrennt und ein Phosphotyrosin-Immunoblot angefertigt. Probe 1
zeigt die Tyrosinphosphorylierung nicht-adhärenter diskoider Thrombozyten (20 x 106 Zellen). Die Membran
wurde gestrippt und anschließend mit einem polyklonalen Antikörper gegen die FcRγ-Kette erneut inkubiert.
Die Ponceau S Färbung der Membran zeigt, daß die in den verschiedenen Spuren aufgetragene Proteinmenge
(gemessen an Aktin) vergleichbar ist.
Während des spreading von Thrombozyten wird eine Reihe von Proteinen vermehrt
tyrosinphosphoryliert (s. Abb. 34). Am auffälligsten sind dies die Proteine P62, P68, P130,
desweiteren mehrere Banden im Bereich von 50 - 60 kDa, bei welchen es sich um

90
Tyrosinkinasen der Src-Familie handelt, sowie eine Doppelbande mit einem
Molekulargewicht von ca. 95 kDa. Die fokale Adhesionskinase FAK konnte mit einem
monoklonalen Antikörper identifiziert werden. Sie läuft direkt unterhalb der P130 Bande.
Desweiteren war auch eine gesteigerte Tyrosinphosphorylierung der FcRγ-Kette (15 kDa
Molekülmasse) während des spreading zu beobachten (Daten nicht gezeigt).
Die ausgebreiteten Kontrollthrombozyten zeigten ein deutliches spreading. Es sind
�Radspeichen�-förmige Strukturen zu erkennen, die von einem mehr oder weniger
kreisförmigen Saum umgeben sind (s. Aktin-Färbung Abb. 33A). Die diversen Inhibitoren
hatten folgende Einflüsse auf das spreading und die damit verbundene Stimulation der
Protein-Tyrosinphosphorylierung: Der Fibrinogenrezeptor-Antagonist FR-1 verhinderte durch
Hemmung des Integrins αIIbβ3 das spreading, nicht jedoch den Gestaltwandel (Ausbildung
von Pseudopodien) der adhärenten Thrombozyten (nicht gezeigt), und reduzierte drastisch die
Tyrosinphosphorylierung. Das Integrin αIIbβ3 ist somit essentiell für das spreading von
Thrombozyten.
Der Tyrosinkinaseinhibitor Genistein hatte einen starken Hemmeffekt auf die Ausbreitung der
Thrombozyten (nicht gezeigt) und auf die durch das spreading hervorgerufene
Tyrosinphosphorylierung. Auffallend war die komplette Hemmung der
Tyrosinphosphorylierung von P130. Eine Vorinkubation mit den Src-Kinase Inhibitoren PP1
und PD173956 zeigte morphologisch eine partielle Hemmung der Thrombozytenausbreitung
(Abb. 33B, C). Parallel dazu war eine Reduktion der Tyrosinphosphorylierung zu beobachten.
Auffallend war die komplette Hemmung der Tyrosinphosphorylierung des Proteins P62 nach
Behandlung der Thrombozyten mit PP1.
Nach einer Vorinkubation mit dem Rho-Kinase Hemmer Y-27632 fiel bei den
morphologischen Untersuchungen auf, daß häufiger Spätstadien der Ausbreitung zu finden
waren (Aktinverdichtungen, häufig in Form von kreisförmigen Strukturen am Zellrand; Daten
nicht gezeigt), was auf eine Beschleunigung des spreading durch Y-27632 hindeutet. Die
Tyrosinphosphorylierung blieb unbeeinflußt.
Desweiteren wurde der Einfluß des PKC-Hemmers Ro-31-8220 untersucht. Das spreading
nach einer Vorbehandlung mit Ro-31-8220 war nicht so stark ausgeprägt wie bei den
Kontrollthrombozyten (Abb. 33D). Beim Tyrosinphosphorylierungsmuster nach dem
spreading fiel auf, daß die fokale Adhäsionskinase FAK bei 125 kDa und die beschriebene
Doppelbande bei 95 kDa nicht mehr phosphoryliert waren.
Nach BAPTA-AM (20 µM, 20 min), EGTA (1mM, 1 min) waren keine signifikanten

91
Änderungen beim spreading als auch bei der Tyrosinphosphorylierung zu erkennen.
Zusammengefaßt zeigen die Ergebnisse, daß Src-Kinasen für das spreading erforderlich sind.
Desweiteren scheint auch die Proteinkinase C wichtig zu sein, wovon in der weiteren
Signalkette auch die Tyrosinphosphorylierung der fokalen Adhäsionkinase FAK abhängt.
Nach Hemmung der Rho-Kinase und nach Erniedrigung des Ca2+-Spiegels durch BAPTA-
AM, EGTA-Behandlung waren keine signifikanten Unterschiede zu erkennen.
Maus-Thrombozyten
Zur Klärung der Rolle der Src-Kinasen Fyn und Lyn sowie der Tyrosinkinase Syk für das
spreading, wurden Fyn(-/-)-und FcRγ-Ketten(-/-)-Maus-Thrombozyten untersucht. Diese
waren über eine Kooperation mit dem Institute of Pharmacology, University of Oxford, UK,
zugänglich.
Um bei Maus-Thrombozyten ein vergleichbares spreading-Verhalten und einen Anstieg der
Tyrosinphosphorylierung wie bei humanen Thrombozyten beobachten zu können, mußten die
Versuchsbedingungen modifiziert werden. Dafür wurden die Thrombozyten im wesentlichen
nicht mit Aspirin vorbehandelt. Desweiteren wurden die Zentrifugationszeiten erniedrigt und
die Drehzahlen modifiziert (2.2.1). Die gewaschenen Thrombozyten wurden auf mit
Fibrinogen beschichtete Deckgläschen aufzentrifugiert und anschließend für 15 min bei 37°C
inkubiert.
Während des spreading konnte dann in Maus-Thrombozyten ebenso wie in humanen
Thrombozyten eine ähnliche Erhöhung des Musters der Proteintyrosinphosphorylierung
beobachtet werden (s. Abb. 35).
92
Maus-TZ Humane TZK S K S
PYkDa
105
75
50
35
30
Abb. 35: Tyrosinphosphorylierung von Maus-Thrombozyten im Vergleich zu humanen Thrombozyten
Die spreading-Experimente wurden für die Maus-Thrombozyten optimiert, um eine starke Ausbreitung der
Maus-Thrombozyten zu sehen (2.2.1). Die Abbildung zeigt das Tyrosinphosphorylierungsmuster nach
Auftrennung des Thrombozytenproteine über ein Gradientengel (5 � 20%). Es werden ruhende Thrombozyten
(Kontrolle) und ausgebreitete Maus-Thrombozyten (spreading) den entsprechenden Resultaten von humanen
Thrombozyten gegenübergestellt.
Wurden nun Maus-Wildtypthrombozyten mit Fyn(-/-)-Thrombozyten in ihrer Morphologie
und ihrem Tyrosinphosphorylierungsmuster während des spreading verglichen, so waren
keine signifikanten Unterschiede festzustellen (Daten nicht gezeigt), was darauf schließen
läßt, daß Fyn, eine Kinase der Src-Familie, zumindest für das spreading von murinen
Thrombozyten nicht von entscheidender Bedeutung ist bzw. in ihrer Funktion von anderen
Kinasen ersetzt werden kann.
Ebenso verhielt es sich bei spreading Versuchen, die mit FcRγ-Ketten(-/-)-Maus-
Thrombozyten durchgeführt wurden. Auch hier waren die Morphologie und die Änderung der
Tyrosinphosphorylierung vergleichbar mit den Befunden in Wildtyp-Maus-Thrombozyten
(Daten nicht gezeigt).

93
3.5 Yeast Two-Hybrid System zur Identifikation von Interaktions-partnern
von Syk
Das Yeast Two-Hybrid System wurde eingesetzt, um Interaktionspartner des Proteins Syk zu identifizieren.
Die Syk kodierende Sequenz lag in Baculovirus (PAcG2T Baculovirus Transfer Vector, humanes Syk kloniert
als BamH1 Fragment; Geschenk Dr. S.P. Watson, Oxford) vor. Syk (wild type, full length) konnte erfolgreich in
den bait-Vektor kloniert und im Hefestamm S. cerevisiae Y190 exprimiert werden (Abb. 36).
Transformationen1 2
pAS2-1
pAS2-1
pAS2-1+Syk
1 2 3 4
Abb. 36: Expression von Syk im Hefestamm S. cerevisiae Y190
Syk wurde in den DNA-bindenden Vektor pAS2-1 kloniert (bait). Es erfolgte eine chemische Transformation
des Hefestamms S. cerevisiae Y190 mit dem Vektor und nach Inkubation der Nachweis von Syk mittels Western
Blot. Dargestellt sind die Ergebnisse von zwei verschiedenen Transformationen. Bei Transformation 2 sind die
Proteinextrakte von 4 verschiedenen Transformanten aufgetragen worden.
Anschließend wurde ein Screen mit einer menschlichen Knochenmarks-cDNA Bank (250 µg;
Zahl der unabhängigen Klone: 3,0 x 106) durchgeführt. Der Screen mit dem bait, pAS2-1, das
Syk full-length enthielt, und der menschlichen Knochenmarks-cDNA Bank lieferte einen
positiven Klon, welcher beim β-Galaktosidasetest blau wurde. Als Kontrolle dienten hierbei
die Interaktionspartner GP3a und Endonexin. Eine Isolation des prey Plasmids und
anschließende erneute Transformation konnte den positiven Befund jedoch nicht bestätigen.
Desweiteren wurde ein Screen des bait Vektors pGBT9/Syk full-length in Y190 mit der
menschlichen Knochenmarks-cDNA Bank (250 µg) unternommen. Auch dieser Versuch
lieferte kein positives Ergebnis. Schließlich wurde eben genannter bait Vektor mit einer
Mischung aus zwei cDNA-Banken gescreent: 200 µg menschliche Knochenmarks-cDNA-
Bank und 175 µg fötale Leberzellen cDNA-Bank (2,0 x 106 unabhängige Klone). Dieser
Screen lieferte zwei blaue Kolonien, die jedoch auch nicht verifiziert werden konnten.

94
Desweiteren wurde die Subklonierung der Kinase-inaktiven Syk-Mutante in den Vektor
pGBT9 durchgeführt. Da die Umsetzung der Substrate durch die Kinase in der Regel sehr
schnell erfolgt, reicht der Kontakt der interagierenden Proteine möglicherweise nicht aus, um die
Transkription der β-Galaktosidase im Yeast Two-Hybrid System anzuschalten. Diese
Schwierigkeit wird mit der Kinase-inaktiven Mutante umgangen. Sie enthält eine Mutation in
der Kinase-Domäne des Proteins. Das Substrat wird zwar gebunden, aber nicht umgesetzt,
woraus eine längere Kontaktzeit resultiert.

95
4. Diskussion
4.1 Integrin ααααIIbββββ3 unabhängige Tyrosinphosphorylierung von Syk während
des shape change
In der vorliegenden Arbeit wurden die Aktivierung von Syk und dessen Regulation während
des Formwandels, der Ausbreitung und Aggregation von humanen Thrombozyten untersucht.
Dabei wurden Unterschiede gefunden, die in der Folge einzeln diskutiert werden.
Die Untersuchungen wurden an isolierten Thrombozyten durchgeführt. Die Expression der
Tyrosinkinase Syk mittels Western Blot war in humanen Thrombozyten gut nachweisbar,
während Syk in humanen Endothelzellen nicht gefunden wurde. Entgegen der beschriebenen
Befunde konnte die Expression von Syk in Endothelzellen von der Arbeitsgruppe um
Yamamura (Yanagi et al., 2001a) nachgewiesen werden. In dieser Untersuchung war Syk
sowohl in HUVECs (human umbilical vein endothelial cells) als auch in BAECs (bovine
aortic endothelial cells), wenn auch relativ schwach im Vergleich zu Wildtyp-
Mausleberzellen, zu finden. Unter den in 3.1.1 beschriebenen Bedingungen mit dem
verwendeten Antikörper war es nicht möglich, Syk in Endothelzellen zu detektieren. Eine
Expression von Syk wurde auch in weiteren nicht-hämatopoetischen Zellen gezeigt, so z.B. in
Fibroblasten, Epithelzellen, Brustgewebe, Hepatozyten und neuronalen Zellen. Es konnte
gezeigt werden, daß Syk funktionell wichtig ist für diese Zelltypen (Yanagi et al., 2001b).
Tsuchida et al. berichten über die Rolle von Syk in menschlichen Hepatozyten durch dessen
Beteiligung an einem Signalweg, der zur MAP-Kinase Aktivierung durch G-Protein
gekoppelte Rezeptoren führt (Tsuchida et al., 2000). Yanagi et al. vermuten, daß Syk in
Endothelzellen eine wichtige regulatorische Funktion für die Organisation des Zytoskeletts
hat (Yanagi et al., 2001a). Außerdem ist Syk bei der Differenzierung von embryonalen
Karzinom-P19-Zellen im Maushirn beteiligt (Tsujumura et al., 2001).
Während des shape change in humanen Thrombozyten ausgelöst durch das Heptapeptid
YFLLRNP oder durch mox-LDL werden die Proteine P62, P68 und P130
tyrosinphosphoryliert (Negrescu et al., 1995; Baumann-Siemons, 2000). In der vorliegenden

96
Arbeit wurde gezeigt, daß während des Formwandels, ausgelöst durch YFLLRNP und mox-
LDL, auch Syk vermehrt an Tyrosin phosphoryliert wird.
Um Syk von möglichen anderen tyrosinphosphorylierten Proteinen im Bereich 68 - 72 kDa
abzugrenzen, war eine Immunpräzipitation von Syk notwendig. Die Immunpräzipitation
zeigte tatsächlich, daß die im Western Blot von Thrombozytenlysaten bei 68 kDa laufende
tyrosinphosphorylierte Proteinbande nicht mit Syk identisch ist. Da die Stimulation der
Tyrosinphosphorylierung von Syk nur nach vorheriger Immunpräzipitation nachweisbar war,
war es wichtig, die Methode der Syk-Immunpräzipitation zu optimieren (s. 3.1.2). In diesen
methodischen Versuchen wurden inter- und intraindividuelle Schwankungen der Expression
von Syk festgestellt. Weiterhin wurden beim Vergleich von Experimenten mit Thrombozyten
verschiedener Spender erhebliche Unterschiede der Syk-Tyrosinphosphorylierung in
ruhenden Thrombozyten beobachtet (Abb. 4). Da die Syk-Tyrosinphosphorylierung in
ruhenden Thrombozyten nicht konstant war, fiel die Stimulation der Syk-
Tyrosinphosphorylierung während des shape change unterschiedlich aus. Schließlich war der
Anstieg der Syk-Tyrosinphosphorylierung vor allem nach Stimulation mit mox-LDL sehr
variabel. Dies könnte möglicherweise auf den unterschiedlichen Oxidationszustand von mox-
LDL zurückzuführen sein und/oder auf die unterschiedliche Antwort der
Thrombozytenpräparationen auf mox-LDL. Nach Thrombin-Rezeptor Aktivierung mittels
YFLLRNP nahm die Syk-Tyrosinphosphorylierung um das 5,4-fache zu, während nach mox-
LDL ein Anstieg um das 2,5-fache zu verzeichnen war (s. 3.2.1). Möglicherweise werden
nach Thrombin-Rezeptor-Aktivierung mehr Signalwege als nach Stimulation mit mox-LDL
aktiviert, so daß daraus eine stärkere Syk-Tyrosinphosphorylierung resultiert.
Auch Clark et al. stellen eine Aktivierung von Syk während der frühen Aktivierung von
Thrombozyten fest (Clark et al., 1994). Wichtig ist zu beachten, daß in diesen Experimenten
der shape change mit der Sekretion von Granula-Inhaltsstoffen assoziiert war, während in den
Studien unserer Arbeitsgruppe allein der shape change (ohne Sekretion) untersucht wurde.
Wie in der vorliegenden Arbeit beschrieben auch Gallet et al. die Aktivierung von Syk
während des shape change ohne Sekretion (Gallet et al., 1999). Desweiteren fand diese
Arbeitsgruppe eine Assoziation von Syk mit dem Zytoskelettprotein Cortactin nach
Thromboxan A2-Rezeptoraktivierung. Die Syk-Kinaseaktivierung und Tyrosinphospho-
rylierung von Cortactin korrelierten in ihrer Kinetik mit dem shape change. Die Autoren
schlossen aus diesen Ergebnissen, daß Cortactin möglicherweise ein Substrat von Syk

97
darstellt und wichtig für die Zytoskelettreorganisation während des shape change ist. Diese
Schlußfolgerung unterstützend beschrieben Maruyama et al. in der humanen leukämischen
Zellinie K562 nach Aktivierung die Translokation von Syk vom Zytosol zum Zytoskelett und
die Tyrosinphosphorylierung von Cortactin durch Syk. Cortactin ist ein an F-Aktin bindendes
Protein, das für die Reorganisation von Mikrofilamenten wichtig zu sein scheint (Maruyama
et al., 1996).
Überraschenderweise zeigten die eigenen Versuche, daß Syk keine Rolle für die dem shape
change zugrundeliegenden Zytoskelettveränderungen spielt. Syk(-/-) Maus-Thrombozyten
zeigten bei den Lichttransmissionsmessungen im Aggregometer einen den Wildtyp-Maus-
Thrombozyten vergleichbaren shape change und einen vergleichbaren Anstieg der
Tyrosinphosphorylierung von P62, P68 und P130 während des Formwandels. Syk ist also
nicht für den shape change und den Anstieg der Tyrosinphosphorylierung von P62, P68 und
P130 während des Formwandels verantwortlich. Die Untersuchungen an Syk(-/-) Mäusen
wurden durch Experimente mit humanen Thrombozyten unterstützt. Die Inkubation der
Thrombozyten mit EGTA, welches das Integrin αIIbβ3 dissoziiert, aktivierte Syk, ohne eine
Plättchenaktivierung zu induzieren (Negrescu und Siess, 1996 und Abb. 13 der vorliegenden
Arbeit). Eine weitere Zugabe von YFLLRNP führte zum shape change, der aber keinen
weiteren Anstieg der Tyrosinphosphorylierung von Syk verursachte. Studien mit spezifischen
Src-Kinase Hemmern und mit Fyn und Lyn defizienten Maus-Thrombozyten zeigten, daß die
Tyrosinkinasen der Src-Familie und die Phosphorylierung ihrer Substrate auch nicht
funktionell in den shape change involviert sind (Bauer et al., 2001). Aus diesen
Untersuchungen ist zu schließen, daß zwar Syk wie die Tyrosinkinasen der Src-Familie
während des shape change aktiviert werden, daß aber Syk und die Src-Kinasen keine Rolle
für die Zytoskelettveränderungen während des shape change spielen.
Die Versuche mit dem vermeintlich spezifischen Hemmer der Syk-Tyrosinkinase Piceatannol
zeigten eine dosisabhängige Reduzierung der Tyrosinphosphorylierung der Proteine P62, P68
und P130, aber keine Hemmung der Tyrosinphosphorylierung von Syk. In einer früheren
Untersuchung fanden Keely und Parise in Thrombozyten eine Hemmung der Syk-
Tyrosinphosphorylierung und Kinaseaktivität durch Piceatannol, aber keinen Effekt auf die
Src-Kinase Aktivität (Keely und Parise, 1996). Wie in der vorliegenden Arbeit beschrieben
Law et al. ebenfalls Probleme beim Einsatz von Piceatannol als Syk-selektiven Hemmer.
Auch andere Kinasen wurden gehemmt, sogar bei sehr geringen Konzentrationen an

98
Piceatannol (Law et al., 1999). Da Piceatannol also nicht selektiv für die Tyrosinkinase Syk
ist, ist es unbrauchbar für Funktionsuntersuchungen in Thrombozyten. Frühere
Untersuchungen der Inhibierung des shape change durch �Breitband�-Tyrosinkinase-
Inhibitoren wie Genistein oder Tyrphostin A23 (Negrescu et al., 1995), die in der
vorliegenden Arbeit bestätigt wurden (s. 3.2.3.3), zeigten, daß möglicherweise eine nicht-
identifizierte Tyrosinkinase beim shape change beteiligt ist.
Was die Regulation der Syk-Aktivierung während des shape change betrifft, führten die
Untersuchungen zu folgenden Ergebnissen: Da die beiden strukturell nicht verwandten Src-
Kinase Hemmstoffe PP1 und PD173956 die Aktivierung von Syk während des shape change
induziert durch mox-LDL bzw. LPA komplett gehemmt haben (s. Abb. 16), liegt die
Vermutung nahe, daß die Syk-Aktivierung sekundär zur Aktivierung der Kinasen der Src-
Familie erfolgt. Eine Aktivierungskaskade von zunächst Src-Kinasen und anschließend Syk
wurde bereits für die Kollagen-induzierte Signaltransduktion in Thrombozyten beschrieben
(Poole et al., 1997; Briddon und Watson, 1999). Beim Kollagen-vermittelten Signalweg
kommt es zur Tyrosinphosphorylierung der FcRγ-Kette durch Src-Familie Kinasen und zur
Assoziation der FcRγ-Kette mit der Tandem-SH2-Domäne von Syk. Letzteres führt zur
Tyrosinphosphorylierung und Aktivierung von Syk. Während des shape change wurde die
FcRγ-Kette jedoch nicht vermehrt an Tyrosin phosphoryliert, was
Coimmunpräzipitationsstudien zeigten (s. 3.4.2.1). Daraus ist zu schließen, daß bei der Src-
Kinase-abhängigen Stimulation von Syk während des shape change die FcRγ-Kette nicht
beteiligt ist.
YFLLRNP, welches zu einer Thrombinrezeptor-Aktivierung führt, aber weder Ca2+ erhöht
noch die Proteinkinase C aktiviert (Rasmussen et al., 1993; Bauer et al., 1999), führte zur
Syk-Aktivierung während des shape change. Gleichermaßen induzieren in der Gegenwart von
EGTA mox-LDL und niedrige Konzentrationen von LPA keine Erhöhung des zytosolischen
Ca2+ in Thrombozyten, aber eine Syk-Aktivierung (Abb. 16 und Maschberger et al., 2000).
Daraus ist zu schließen, daß weder ein Anstieg des zytosolischen Ca2+ noch die Proteinkinase
C für die Syk-Aktivierung während des shape change verantwortlich sind. Dies unterstützend
zeigten die Ergebnisse mit dem PKC-Hemmer Ro-31-8220, daß dieser während des shape
change sowohl nach Stimulation mit YFLLRNP als auch mit mox-LDL die Syk-
Tyrosinphosphorylierung nicht hemmte.

99
Untersuchungen zur Regulation der Syk-Aktivierung durch zytosolisches Ca2+ ergaben, daß
nach einer Vorbehandlung der Thrombozyten mit BAPTA-AM/EGTA, welches das basale
zytosolische Ca2+ erniedrigt, der shape change, induziert durch mox-LDL und YFLLRNP,
jedoch verzögert abläuft und reduziert ist. Desweiteren war die basale
Tyrosinphosphorylierung von Syk erniedrigt und es kam auch nach Aktivierung zu keiner
Erhöhung. Dies zeigt, daß normale basale Werte von zytosolischem Ca2+ (80 - 100 nM)
erforderlich sind für eine Tyrosinphosphorylierung von Syk sowohl in ruhenden
Thrombozyten als auch nach Stimulation mit mox-LDL und YFLLRNP. Im Gegensatz zu
diesen Ergebnissen induzierte mox-LDL auch nach einer Behandlung mit BAPTA-
AM/EGTA die Translokation von tyrosinphosphoryliertem P62, P68, P130, Src, Fyn und Syk
zum Zytoskelett (s. 3.2.3.2). Die durch mox-LDL induzierte Translokation von
tyrosinphosphorylierten Proteinen und Tyrosinkinasen zum Zytoskelett und der durch Src-
Kinasen vermittelte Anstieg der Proteintyrosinphosphorylierung sind somit unabhängig von
zytosolischem Ca2+. In einer früheren Studie wurde ebenfalls gefolgert, daß die
Proteintyrosinphosphorylierung nicht sekundär zur Ca2+-Mobilisierung nach
Plättchenstimulation mit LPA erfolgt (Torti et al., 1996). Ebenfalls wie die Syk-
Tyrosinphosphorylierung erfordert der Rho/Rho-Kinase Signaltransduktionsweg, welcher
über die Phosphorylierung der myosin light chain (MLC) zum shape change führt, für eine
optimale Aktivität die Basalkonzentration von zytosolischem Ca2+ in ruhenden
Thrombozyten. Er wird jedoch nicht durch den Anstieg des zytosolischen Ca2+ während des
shape change reguliert (Bauer et al., 1999; Bauer, 2003).
Weiterhin zeigt die Arbeit, daß die Syk-Aktivierung während des shape change von dem
Integrin αIIbβ3 unabhängig ist. Dies ist zu erwarten, weil während des shape change kein
Integrin αIIbβ3 outside-in signaling erfolgt. Jedoch war zu Beginn der Arbeit die Frage offen,
ob für die Wirkung von mox-LDL auf die Thrombozyten das Integrin αIIbβ3 wichtig ist.
Andere Arbeitsgruppen hatten gefunden, daß in Thrombozyten LDL und ox-LDL mit hoher
Affinität an spezifische Stellen binden, welche sich vom klassischen LDL-Rezeptor
unterscheiden (Pedreno et al., 1992; Pedreno et al., 1994), und hatten die Glycoproteine IIb
und IIIa (das Integrin αIIbβ3) als Bindestellen für LDL auf Plättchenmembranen, aber nicht
auf intakten Plättchen identifiziert (Koller et al., 1989; Pedreno et al., 1997). Eine neuere
Studie zeigte, daß das Integrin αIIbβ3 bei der Thrombozytensensitisierung durch LDL beteiligt
ist (Hackeng et al., 1999). In der vorliegenden Arbeit wurden Plättchen von Patienten mit

100
Thrombasthenia Glanzmann, denen das Integrin αIIbβ3 fehlt, untersucht (s. 3.2.2). In diesen
Thrombozyten war die Stimulation der Tyrosinphosphorylierung von P62, P68, P130 und von
Syk während des shape change vergleichbar zu der in Kontrollthrombozyten. Desweiteren
war auch der shape change nach mox-LDL in Thrombasthenischen und normalen Plättchen
ähnlich. Die gleichen Ergebnisse erhielt man auch nach Vorbehandlung von normalen
humanen Thrombozyten mit RGDS, einem Fibrinogenrezeptor Antagonisten (s. 3.2.2).
Daraus ist zu schließen, daß mox-LDL nicht über das Integrin αIIbβ3 den Thrombozyten-
shape change, die Proteintyrosinphosphorylierung und Syk stimuliert.
Untersuchungen der Arbeitsgruppe hatten gezeigt, daß mox-LDL über die Aktivierung des
LPA-Rezeptors den shape change induziert. Vergleicht man die Wirkung von mox-LDL mit
der von LPA, der aktiven Substanz in mox-LDL (Siess et al., 1999), so wurde festgestellt, daß
mox-LDL und LPA einen ähnlichen Anstieg der Syk-Tyrosinphosphorylierung während des
shape change hervorrufen (Abb. 16). Nach niedrigen Konzentrationen an LPA werden Src-
und Syk-Tyrosinkinasen während des shape change aktiviert, während nach hohen LPA
Konzentrationen das zytosolische Ca2+ erhöht wird (Maschberger et al., 2000). Mox-LDL und
hohe Konzentrationen an LPA induzieren hauptsächlich einen Ca2+-Einstrom über die
Plasmamembran und relativ wenig Ca2+-Mobilisierung aus intrazellulären Speichern, ein
Befund, der ausschließlich für diese Stimuli zutrifft und für andere Agonisten nicht
beobachtet wurde (Siess, 1989; Siess et al., 1999). Yang et al. fanden, daß LPA und
Sphingosin 1-phosphat (S1-P) die Tyrosinphosphorylierung von Thrombozytenproteinen,
darunter auch Syk, stimulieren und zur Syk-Aktivierung führen. Zu bemerken ist, daß hier
sehr große Mengen an LPA, nämlich 40 µM zur Stimulation eingesetzt wurden (Yang et al.,
1996) im Vergleich zu nur 80 nM in unseren Versuchen. Auch Torti et al. zeigten die schnelle
Tyrosinphosphorylierung von mehreren intrazellulären Proteinen inklusive Syk nach
Plättchenaktivierung mit LPA (Torti et al., 1996). In keiner dieser Studien wurde die
Aktivierung von Syk zu spezifischen Thrombozytenantworten korreliert.
Aus der Sicht des heutigen Standes der Forschung ist unklar, ob der Plättchen-LPA-Rezeptor
zu den drei Siebentransmembrandomänen-Rezeptoren für LPA (LPA1, LPA2, LPA3) gehört
(Goetzl und An, 1998). Es liegt nahe zu spekulieren, daß die Aktivierung der zwei
voneinander unabhängigen Signalwege, Src-Kinase und Syk-Aktvierung einerseits und
zytosolische Ca2+-Erhöhung andererseits, durch die Aktivierung von zwei verschiedenen
LPA-Rezeptoren vermittelt wird. Alternativ könnte ein LPA-Rezeptor verschiedene
Signaltransduktionswege aktivieren. So wurde gezeigt, daß ein LPA-Rezeptor an mehrere

101
heterotrimere G-Proteine wie Gi und Gq koppeln kann (Moolenaar et al., 1997). Analog zu
Arbeiten in anderen Zelltypen könnten in Thrombozyten die Tyrosinkinasen Src und Syk
durch Gi-Aktivierung stimuliert werden, während die Gq-Aktivierung zu einer Ca2+-Antwort
führt. Tatsächlich wurde eine Gi-vermittelte Src-Kinase und Syk-Aktivierung in anderen
Zelltypen wie Fibroblasten (Hordijk et al., 1994) und Avian B-Zellen (DT40 Zellen) (Wan et
al., 1996) beschrieben. In Thrombozyten von Gq-defizienten Mäusen konnte die Aktivierung
von Src-Familientyrosinkinasen und Syk, unabhängig von einem zytosolischen Ca2+-Anstieg,
beobachtet werden (Klages et al., 1999).
Welche mögliche Bedeutung hat nun die Syk Aktivierung während des shape change? Mittels
des monoklonalen Antikörpers PAC-1, der spezifische Epitope der aktiven Konformation des
Integrins αIIbβ3 auf der Plättchenmembran erkennt, konnte festgestellt werden, daß während
des shape change ein beträchtlicher Teil des Integrins αIIbβ3 aktiviert wird. Desweiteren
wurde gefunden, daß die Bindung von PAC-1 während des durch YFLLRNP induzierten
shape change nach Vorbehandlung mit den Src-selektiven Hemmern vollständig unterdrückt
war (Bauer et al., 2001 und Abb. 16). Diese Src-selektiven Hemmer inhibierten die
Proteintyrosinphosphorylierung und die Aktivierung von Syk während des shape change
(Bauer et al., 2001). Neben den Src-Kinasen, könnte also auch die Kinase Syk am Integrin
αIIbβ3 inside-out signaling beteiligt sein. Dies unterstützend konnte gezeigt werden, daß Syk
bei der Stimulus-induzierten Aktivierung des Integrins αIIbβ3 in Maus-Thrombozyten beteiligt
ist. Versuche in Syk(-/-) Maus-Thrombozyten, in denen die Aktivierbarkeit des Integrins
αIIbβ3 über die Bindung von FITC-gekoppeltem Fibrinogen gemessen wurde, zeigten, daß die
Fibrinogenbindung nach Stimulation mit ADP ± Epinephrin im Vergleich zu
Kontrollthrombozyten erniedrigt war (Law et al., 1999).
Zusammenfassend zeigen die Resultate, daß die Aktivierung von Syk während des raschen
Formwandels der Thrombozyten über einen Mechanismus, der abhängig von der Aktivierung
von Src-Kinasen, aber ohne Beteiligung des Integrins αIIbβ3 und unabhängig von der ITAM-
Tyrosinphosphorylierung der FcRγ-Kette erfolgt. Für den shape change selbst hat Syk jedoch
keine Funktion, es könnte jedoch für die Aktivierung des Integrins αIIbβ3 wichtig sein, welche
für die sich an den Formwandel anschließende Thrombozyten-Aggregation bzw. -Ausbreitung
(spreading) wichtig ist.

102
4.2 Integrin ααααIIbββββ3 abhängige Tyrosinphosphorylierung von Syk
Das Integrin αIIbβ3 vermittelt die Thrombozytenaggregation und das spreading in vaskulären
Wunden durch Interaktionen mit löslichen oder Matrix-assoziierten Liganden. Diese
Funktionen kommen bei der Hämostase und bei der arteriellen Thrombose zum Tragen.
Die Untersuchungen der Arbeit zeigten, daß die Aktivierung des Integrins αIIbβ3 in Thrombin-
aggregierten Thrombozyten die Stimulierung eines ähnlichen Signalwegs bewirkt wie die
Aktivierung des Glykoproteins VI durch Kollagen: es wurde eine Tyrosinphosphorylierung
der FcRγ-Kette durch Src-Familie Kinasen gefunden. Diese bewirkt eine Assoziation der
FcRγ-Kette mit der Tandem-SH2-Domäne von Syk, was zur Tyrosinphosphorylierung und
Aktivierung von Syk führt, und in der Folge zur Tyrosinphosphorylierung der Phospholipase
Cγ2. RGDS, welches die Bindung von Fibrinogen und anderen Liganden an das Integrin
αIIbβ3 blockiert, inhibierte die Tyrosinphosphorylierung der FcRγ-Kette komplett, ebenso
deren Assoziation mit Syk, und reduzierte drastisch die Tyrosinphosphorylierung von Syk
und PLCγ2 in Thrombin-stimulierten Thrombozyten. Zwei strukturell unterschiedliche
spezifische Hemmer der Tyrosinkinasen der Src-Familie (PP1 und PD173956) führten zu
einer Reduktion der Tyrosinphosphorylierung von Syk und einer kompletten Hemmung der
Tyrosinphosphorylierung der FcRγ-Kette in Thrombin-aggregierten Thrombozyten. Die
Bedeutung der FcRγ-Kette für die Stimulation der Syk-Tyrosinphosphorylierung in
Thrombin-aggregierten Thrombozyten wird durch eine Studie von Poole et al. unterstützt.
Diese fanden, daß die Syk-Tyrosinphosphorylierung in FcRγ-Ketten-defizienten Maus-
Thrombozyten stark reduziert war, wenn sie mit niedrigen Thrombinkonzentrationen
aggregiert wurden (Poole et al., 1997).
Es zeigte sich weiter, daß dieser durch das Integrin αIIbβ3 vermittelte Signalweg funktionell
für die Thrombozytenaggregation induziert durch niedrige Thrombinkonzentrationen relevant
ist. Diese Schlußfolgerungen beruhen auf den Ergebnissen mit den Src-Kinase Inhibitoren
PP1 und PD173956 und den Studien an FcRγ-Ketten- und Syk-defizienten
Mausthrombozyten, welche eine Verminderung der Plättchenaggregation nach Stimulation
mit niedrigen Thrombinkonzentrationen zeigten. Die Inhibierung der
Thrombozytenaggregation war nach der Vorbehandlung der Thrombozyten mit den Src-
Kinase Inhibitoren stärker als in FcRγ-Ketten- oder Syk-defizienten Mäusen. Das liegt

103
möglicherweise daran, daß Src-Kinasen auch unabhängig von Syk bei dem Integrin αIIbβ3
inside-out signaling und Integrin αIIbβ3 outside-in signaling beteiligt sind. Src Kinasen wie
Src selbst und Lyn sind konstitutiv mit dem Integrin αIIbβ3 in Thrombozyten assoziiert
(Dorahy et al., 1995; Obergfell et al., 2002). Eine derartige Assoziation könnte die Integrin
αIIbβ3 vermittelte Aktivierung von Src-Kinasen bewirken sowie die anschließende
Tyrosinphosphorylierung der FcRγ-Kette.
Im Unterschied zu älteren Veröffentlichungen (Blake et al., 1994; Poole et al., 1997) ergaben
die Untersuchungen der Arbeit eine eindeutige Tyrosinphosphorylierung der PLCγ2 in
Thrombin-stimulierten Thrombozyten. Die unterschiedlichen Ergebnisse könnten durch die
unterschiedlichen experimentellen Bedingungen erklärt werden. In den genannten Studien
wurden die Experimente ohne Aggregation durchgeführt (kein Rühren), was in der
vorliegenden Arbeit auch zu keiner Tyrosinphosphorylierung der FcRγ-Kette führte. Die
Thrombozytenaggregation war Voraussetzung für einen Anstieg der Tyrosinphosphorylierung
der FcRγ-Kette. Wonerow et al. berichten über eine Integrin αIIbβ3-vermittelte Stimulation der
PLCγ2 Tyrosinphosphorylierung in Thrombozyten, welche auf Fibrinogen adhärierten
(Wonerow et al., 2002). Die Kinasen (Kinase), welche die PLCγ2 in stimulierten
Thrombozyten tyrosinphosphorylieren, sind nicht bekannt. In Kollagen-stimulierten
Thrombozyten scheint Syk die verantwortliche Kinase zu sein, da die PLCγ2 in Syk-
defizienten Mausthrombozyten nach Stimulation mit Kollagen nicht tyrosinphosphoryliert
wird (Poole et al., 1997). Jedoch kann Syk im Gegensatz zu Src-Kinasen die PLCγ2 nicht in
vitro tyrosinphosphorylieren (Rodriguez et al., 2001).
Der gleiche Signalweg wie nach Aktivierung des Integrins αIIbβ3 in Thrombin-aggregierten
Thrombozyten wurde durch eine längere Behandlung der Thrombozyten mit EGTA
beobachtet. Die durch das EGTA induzierte Integrin αIIbβ3-Dissoziation bewirkte ebenfalls
eine Src-Kinasen vermittelte FcRγ-Ketten Tyrosinphosphorylierung, deren Assoziation mit
tyrosinphosphoryliertem Syk sowie eine vermehrte Tyrosinphosphorylierung von PLCγ2.
Allerdings wird durch die Integrin αIIbβ3-Dissoziation keine Thrombozytenaktivierung
verursacht (Negrescu und Siess, 1996). Die Aktivierung dieses Signalwegs allein ist also noch
nicht ausreichend, um eine physiologische Antwort zu induzieren. Ein silent pathway von der
Tyrosinphosphorylierung der FcRγ-Kette zu Syk und der PLCγ2 wurde kürzlich auch in
Thrombozyten nach Aktivierung des Glykoproteins Ib von einigen, aber nicht allen

104
Arbeitsgruppen beschrieben (Falati et al., 1999, Wu et al., 2001, Marshall et al., 2002). Auch
in Lymphozyten wurde ein silent pathway der Tyrosinphosphorylierung der PLCγ2
beobachtet (Fluckiger et al., 1998). Erklärt wird die Tyrosinphosphorylierung der PLCγ2 in
Abwesenheit einer Zellaktivierung durch eine Studie, die zeigt, daß die
Tyrosinphosphorylierung nur bestimmter, spezifischer Tyrosinreste der PLCγ2 notwendig ist,
um eine Aktivierung des Enzyms zu verursachen (Ozdener et al., 2002).
Die Beobachtungen in Thrombozyten stehen im Gegensatz zu den Ergebnissen in CHO-
Zellen, welche stabil mit dem Integrin αIIbβ3 und transient mit Syk transfiziert wurden (Gao et
al., 1997). In diesem Zellmodell wurde eine von ITAMs unabhängige Aktivierung von Syk
durch das Integrin αIIbβ3 gefunden. Der Unterschied der beiden Studien liegt vor allem an den
zwei verschiedenen Zelltypen, die untersucht wurden: zum einen Thrombozyten mit der
physiologischen Expression an Integrin αIIbβ3, FcRγ-Kette und Syk gegenüber den
transfizierten CHO-Zellen mit möglicherweise überexprimierter Menge an Integrin αIIbβ3 und
Syk, und unbekannten Mengen an ITAM enthaltenden Proteinen. Im Modellsystem CHO-
Zellen konnte klar gezeigt werden, daß Syk nach Integrin αIIbβ3-Stimulierung über einen
ITAM-unabhängigen Weg aktiviert werden kann. Möglicherweise hängt die Regulation der
Syk-Aktivierung in stimulierten Thrombozyten vom Ausmaß der Integrin αIIbβ3-Aktivierung
ab: eine schwache Aktivierung nach Stimulation der Aggregation mit niedrigen Thrombin-
Konzentrationen führt zu einer funktionell relevanten ITAM-abhängigen Aktivierung von
Syk, während eine starke Aktivierung zur Syk-Aktivierung (z.B. spreading, siehe Diskussion
im folgenden) unabhängig von tyrosinphosphorylierten ITAMs in der FcRγ-Kette führen
könnte.
Das spreading, welches im Anschluß an den shape change erfolgt, ist essentiell für die
Adhäsion von Thrombozyten an geschädigtes Endothel. Während der shape change
unabhängig vom Integrin αIIbβ3 erfolgt, erfordert das spreading die Anwesenheit und das
signaling über das Integrin αIIbβ3. Die Ergebnisse der Arbeit und anderer Arbeitsgruppen
zeigten, daß das spreading Src-Familie Kinasen und die Tyrosinkinase Syk involviert.
Zellbiologische Analysen in der vorliegenden Arbeit ergaben, daß Syk in adhärierenden,
ausgebreiteten Thrombozyten sowohl im Zytosol als auch an der Plasmamembran lokalisiert
war. Hier war teilweise eine Colokalisation mit dem Integrin αIIbβ3 erkennbar.
Untersuchungen von auf Fibrinogen ausgebreiteten CHO-Zellen zeigten eine Colokalisation

105
von Syk und Integrin αIIbβ3 in der frühen Phase, jedoch nicht in der Spätphase des spreading
In aggregierenden und adhärierenden, ausgebreitenen Thrombozyten sowie in CHO-Zellen ist
Syk ein proximales Glied in der Integrin αIIbβ3 abhängigen Signaltransduktionskette.
Außerdem zeigte sich in eigenen Versuchen eine vermehrte Tyrosinphosphorylierung der
FcRγ-Kette während des spreading von humanen Thrombozyten, was eine anschließende
Assoziation an die Tandem-SH2-Dömane von Syk und damit eine Syk-Aktivierung
ermöglichen kann. Bei der Vorbehandlung von humanen Thrombozyten mit den Src-Kinase
Inhibitoren PP1 und PD173956 waren sowohl das spreading partiell gehemmt als auch die
Proteintyrosinphosphorylierung reduziert. Die Src-Kinasen sind somit für die
Thrombozytenausbreitung erforderlich. In mehreren erst kürzlich veröffentlichten Artikeln
wurden Ergebnisse zum Integrin αIIbβ3-vermittelten Signalweg in Thrombozyten, die sich auf
immobilisiertem Fibrinogen ausbreiteten (spreading), dargestellt. Unter diesen Bedingungen,
unter welchen das Integrin αIIbβ3 outside-in signaling maximal stimuliert ist, wurde ein
Signalweg gefunden, welcher die Aktivierung der Kinase Src, welche konstitutiv an das β3-
Integrin gebunden ist, bewirkt. Anschließend wird Syk an das β3�Integrin gebunden und
aktiviert (Obergfell et al., 2002). Tatsächlich kann die N-terminale SH2-Domäne von Syk
direkt an den zytoplasmatischen Teil des β3-Integrins binden (Woodside et al., 2001;
Woodside et al., 2002). Allerdings waren in Untersuchungen von Law et al. keine
Unterschiede in der Adhäsion von Syk-defizienten zu Wildtyp-Mausthrombozyten an
immobilisiertes Fibrinogen feststellbar, woraus zu schließen ist, daß die Tyrosinkinase Syk
für das spreading nicht erforderlich ist (Law et al., 1999).
Mit dem Yeast Two-Hybrid System sollten Interaktionspartner der Tyrosinkinase Syk
ermittelt werden. Das Projekt wurde in den Labors von S.J. Shattil, La Jolla, USA begonnen.
Das Protein Syk konnte in Hefe exprimiert werden und es wurden eine Knochenmarks-
cDNA-Bank sowie eine fötale Leberzellen-cDNA-Bank nach möglichen Interaktionspartnern
gescreent. Es konnten jedoch keine positiven Klone identifiziert werden.
Zusammenfassend kann festgestellt werden, daß die Resultate zum ersten Mal zeigen, daß
Integrine nach Aktivierung über einen ITAM-abhängigen pathway Signale an die Syk-Kinase
und PLCγ2 weiterleiten, welcher dem Signalweg von Immunrezeptoren in Lymphozyten und
Monozyten, und dem des Kollagenrezeptors in Thrombozyten ähnlich ist. Die Aktivierung
von Syk nach Integrin-Aktivierung wurde auch in anderen hämatopoetischen Zellen

106
(Lymphozyten, Monozyten, Myeloidzellen und Neutrophilen) beschrieben. Hier wurde eine
Funktion von Syk bei der Adhäsion, dem spreading, bei zytoskelettalen Veränderungen und
für die NF-kappa B Aktivierung gefunden (Lin et al., 1995; Gotoh et al., 1997; Yan et al.,
1997; Miller et al., 1999; Stupack et al., 1999; Yan et al., 1999). Es ist möglich, daß die
Adhäsion von Matrixproteinen und die Aktivierung von β1-Integrinen und β2-Integrinen in
diesen Zellen zur Syk-Aktivierung über einen Mechanismus führt, welcher von der
Tyrosinphosphorylierung von ITAMs abhängig ist.
4.3 Perspektive
Oxidativ modifiziertes LDL, vor allem mm-LDL oder mox-LDL, könnte über ähnliche
Mechanismen, wie in der vorliegenden Arbeit für Thrombozyten gezeigt wurde, auch andere
Zellen wie Monozyten, Endothelzellen oder glatte Muskelzellen stimulieren, welche bei der
Pathogenese der Arteriosklerose eine Rolle spielen. Die Beobachtung, daß Genistein den
shape change und die Stimulation der Tyrosinphosphorylierung von Syk induziert durch mox-
LDL hemmte, könnte eine mögliche Relevanz für eine diätetische Prävention
arteriosklerotischer Erkrankungen haben. Genistein ist ein natürlicher �Breitband�-Inhibitor
von Tyrosinkinasen und gehört zu den Isoflavonoiden, die vor allem in Soja in hoher
Konzentration vorhanden sind. So wurde bei Untersuchungen von Japanern nach sojareicher
Diät eine 30-fach höhere Ausscheidung von Genistein im Vergleich zu Menschen mit
Normalkost nachgewiesen (Gooderham et al., 1996). Genistein ist ein kompetitiver Inhibitor
von Adenosintriphosphat (ATP) in Kinasereaktionen und wirkt nicht-kompetitiv mit dem
Proteinsubstrat. Da die ATP-Bindungsdomäne unter allen Tyrosinkinasen sehr stark
konserviert ist, wirkt Genistein nicht auf eine spezifische Tyrosinkinase. Eine Alternative zu
Genistein stellen synthetische Tyrphostine (tyrosine phosphorylation inhibitors) wie z.B.
Tyrphostin A23 und A47 dar. Dies sind Tyrosinkinase-Hemmer, die in Konkurrenz mit dem
Proteinsubstrat treten. Inhibitoren von Tyrosinkinasen wie das natürlich vorkommende
Isoflavonoid Genistein oder synthetische Tyrphostine könnten die pathologische
Überstimulation von Zellen durch oxidativ modifiziertes LDL erniedrigen.

107
5. Zusammenfassung Oxidativ modifiziertes low-density lipoprotein (LDL) und Thrombozyten spielen eine zentrale
Rolle in der Pathogenese kardiovaskulärer Erkrankungen. Die Thrombozytenaktivierung führt
abhängig von der Art der Stimulation zu einer raschen Formänderung (shape change),
Ausbreitung (spreading) oder zur Aggregation begleitet von einem Anstieg der
Tyrosinphosphorylierung von Tyrosinkinasen und deren Aktivierung. Die Untersuchung der
Regulation und Aktivierung der Tyrosinkinase Syk stand in der vorliegenden Arbeit im
Mittelpunkt.
Milde oxidiertes LDL (mox-LDL) führt zum shape change. Dabei werden zwei unabhängige
Signalwege, zum einen die Tyrosinphosphorylierung der Proteine P62, P68 und P130 sowie
der Tyrosinkinase Syk und zum anderen die zytosolische Ca2+-Erhöhung durch einen
erhöhten Ca2+-Einstrom stimuliert. Desweiteren kommt es zur Translokation von Fyn und Syk
zum Aktin-reichen Zytoskelett. Wie in der Diskussion ausgeführt, erfolgt die Stimulation
dieser Signalwege über die Aktivierung des LPA-Rezeptors, mit LPA als der aktiven
Substanz im mox-LDL. Das Integrin αIIbβ3 war in die Stimulation dieser frühen
Signaltransduktionswege durch mox-LDL nicht involviert. Die Stimulation der
Proteintyrosinphosphorylierung und Syk-Aktivierung wurde durch die
Tyrosinkinaseinhibitoren Genistein und Tyrphostin A23 und zwei spezifische Inhibitoren der
Src-Familie Kinasen PP1 und PD173956 gehemmt. Im Gegensatz zu PP1 und PD173956
unterdrückten Genistein und Tyrphostin A23 auch den shape change durch mox-LDL. Die
Aktivierung von Syk während des raschen Formwandels der Thrombozyten erfolgt über einen
Mechanismus, der abhängig von der Aktivierung von Src-Tyrosinkinasen, aber unabhängig
vom Integrin αIIbβ3 und von der ITAM-Tyrosinphosphorylierung der FcRγ-Kette ist. Für die
Zytoskelettveränderungen, die dem shape change zugrunde liegen, spielt die Syk-Aktivierung
keine Rolle. Die Syk-Aktivierung während des Formwandels könnte jedoch für die
Aktivierung des Integrins αIIbβ3 wichtig sein, welche für die anschließende Thrombozyten-
Aggregation bzw. -Ausbreitung (spreading) wichtig ist.
Die Aktivierungsmechanismen von Syk während shape change und Aggregation waren
unterschiedlich. In Thrombin-aggregierten Thrombozyten war die Syk-Aktivierung von dem
Integrin αIIbβ3 abhängig. Hier zeigten die Untersuchungen der Arbeit, daß die Aktivierung des

108
Integrins αIIbβ3 die Stimulierung eines ähnlichen Signalwegs bewirkt wie die Aktivierung des
Glykoproteins VI durch Kollagen: Tyrosinphosphorylierung der FcRγ-Kette durch Src-
Familie Kinasen → Assoziation der FcRγ-Kette mit der Tandem-SH2-Domäne von Syk →
Tyrosinphosphorylierung und Aktivierung von Syk → Tyrosinphosphorylierung der
Phospholipase Cγ2. Es zeigte sich weiter, daß dieser durch das Integrin αIIbβ3 vermittelte
Signalweg funktionell für die Thrombozytenaggregation, induziert durch niedrige
Thrombinkonzentrationen, relevant ist. Diese Schlußfolgerungen beruhen auf den
Ergebnissen mit den Src-Kinase Inhibitoren PP1 und PD173956 und den Studien an FcRγ-
Ketten- und Syk-defizienten Maus-Thrombozyten, welche eine Verminderung der
Plättchenaggregation nach Stimulation mit niedrigen Thrombinkonzentrationen zeigten. Der
gleiche Signalweg wie nach Aktivierung des Integrins αIIbβ3 in Thrombin-aggregierten
Thrombozyten wurde durch Dissoziation des Integrins αIIbβ3 mittels EGTA beobachtet.
Allerdings bewirkte die Integrin αIIbβ3-Dissoziation keine Thrombozytenaktivierung, woraus
zu schliessen ist, daß die Stimulierung dieses Signalwegs (Src-Kinasen Aktivierung →
Tyrosinphosphorylierung der FcRγ-Kette → Syk- und PLCγ2-Tyrosinphosphorylierung) nicht
ausreicht, um Thrombozyten physiologisch zu aktivieren.
Die Resultate zeigen zum ersten Mal, daß Integrine nach Aktivierung über einen ITAM-
abhängigen pathway Signale an die Syk-Kinase und PLCγ2 weiterleiten, welcher dem
Signalweg von Immunrezeptoren in Lymphozyten und Monozyten, und dem des
Kollagenrezeptors in Thrombozyten ähnlich ist.

109
6. Literaturverzeichnis
Arias-Salgado, E. G., Lizano, S., Sarkar, S., Brugge, J. S., Ginsberg, M. H. and Shattil, S. J.
(2003) Src kinase activation by direct interaction with the integrin beta cytoplasmic domain.
Proc.Natl.Acad.Sci U.S.A 100, 13298-13302.
Arpaia, E., Shahar, M., Dadi, H., Cohen, A. and Roifman, C. M. (1994) Defective T cell
receptor signaling and CD8+ thymic selection in humans lacking zap-70 kinase. Cell 76, 947-
958.
Asselin, J., Gibbins, J. M., Achison, M., Lee, Y. H., Morton, L. F., Farndale, R. W., Barnes,
M. J. and Watson, S. P. (1997) A collagen-like peptide stimulates tyrosine phosphorylation of
syk and phospholipase Cγ2 in platelets independent of the integrin α2β1. Blood 89, 1235-
1242.
Avogaro, P., Cazzolato, G. and Bittolo-Bon, G. (1991) Some questions concerning a small,
more electronegative LDL circulating in human plasma. Atherosclerosis 91, 163-171.
Bauer, M., Retzer, M., Wilde, J. I., Maschberger, P., Essler, M., Aepfelbacher, M., Watson, S.
P. and Siess, W. (1999) Dichotomous regulation of myosin phosphorylation and shape change
by Rho-kinase and calcium in intact human platelets. Blood 94, 1665-1672.
Bauer, M., Maschberger, P., Quek, L., Briddon, S. J., Dash, D., Weiss, M., Watson, S. P. and
Siess, W. (2001) Genetic and pharmacological analyses of involvement of Src-family, Syk
and Btk tyrosine kinases in platelet shape change. Src-kinases mediate integrin alphaIIb beta3
inside-out signalling during shape change. Thromb.Haemost. 85, 331-340.
Bauer, M. (2003) Funktion der Kinasen der Src-Familie, der Tyrosinkinase Btk, sowie der
Rho-abhängigen Kinase für die frühe Plättchenaktivierung. Dissertation, LMU München.
Baumann-Siemons, J. (2000) Milde oxidiertes LDL stimuliert die
Proteintyrosinphosphorylierung während des Gestaltwandels menschlicher Thrombozyten.
Dissertation, LMU München.

110
Berliner, J. A., Navab, M., Fogelman, A. M., Frank, J. S., Demer, L. L., Edwards, P. A.,
Watson, A. D. and Lusis, A. J. (1995) Atherosclerosis: basic mechanisms. Oxidation,
inflammation, and genetics. Circulation 91, 2488-2496.
Birnboim, H. C. and Doly, J. (1979) A rapid alkaline extraction procedure for screening
recombinant plasmid DNA. Nucleic Acids Res 7, 1513-1523.
Blake, R. A., Schieven, G. L. and Watson, S. P. (1994) Collagen stimulates tyrosine
phosphorylation of phospholipase C-gamma 2 but not phospholipase C-gamma 1 in human
platelets. FEBS Lett. 353, 212-216.
Bobe, R., Wilde, J. I., Maschberger, P., Venkateswarlu, K., Cullen, P. J., Siess, W. and
Watson, S. P. (2001) Phosphatidylinositol 3-kinase-dependent translocation of phospholipase
Cgamma2 in mouse megakaryocytes is independent of Bruton tyrosine kinase translocation.
Blood 97, 678-684.
BORN, G. V. and CROSS, M. J. (1963) Effect of adenosine diphosphate on the concentration
of platelets in circulating blood. Nature 197:974-6., 974-976.
Briddon, S. J. and Watson, S. P. (1999) Evidence for the involvement of p59fyn and p53/56lyn
in collagen receptor signalling in human platelets. Biochem.J. 338, 203-209.
Cantrell, D. (1998) The real LAT steps forward. Trends Cell Biol. 8, 180-182.
Chacko, G. W., Duchemin, A. M., Coggeshall, K. M., Osborne, J. M., Brandt, J. T. and
Anderson, C. L. (1994) Clustering of the platelet Fc gamma receptor induces noncovalent
association with the tyrosine kinase p72syk. J.Biol.Chem. 269, 32435-32440.
Chan, A. C., Pritchard, E. T., Gerrard, J. M., Man, R. Y. and Choy, P. C. (1982) Biphasic
modulation of platelet phospholipase A2 activity and platelet aggregation by mepacrine
(quinacrine). Biochim.Biophys.Acta 713, 170-172.
Chan, A. C., Irving, B. A., Fraser, J. D. and Weiss, A. (1991) The zeta chain is associated
with a tyrosine kinase and upon T-cell antigen receptor stimulation associates with ZAP-70, a

111
70-kDa tyrosine phosphoprotein. Proc.Natl.Acad.Sci U.S.A 88, 9166-9170.
Chan, A. C., Iwashima, M., Turck, C. W. and Weiss, A. (1992) ZAP-70: a 70 kd protein-
tyrosine kinase that associates with the TCR zeta chain. Cell 71, 649-662.
Clark, E. A., Shattil, S. J., Ginsberg, M. H., Bolen, J. and Brugge, J. S. (1994) Regulation of
the protein tyrosine kinase pp72syk by platelet agonists and the integrin alpha IIb beta 3.
J.Biol.Chem. 269, 28859-28864.
Clark, E. A. and Brugge, J. S. (1995) Integrins and signal transduction pathways: the road
taken. Science 268, 233-239.
Coffey, M. D., Cole, R. A., Colles, S. M. and Chisolm, G. M. (1995) In vitro cell injury by
oxidized low density lipoprotein involves lipid hydroperoxide-induced formation of alkoxyl,
lipid, and peroxyl radicals. J.Clin.Invest. 96, 1866-1873.
Coughlin, S. R. (2000) Thrombin signalling and protease-activated receptors. Nature 407,
258-264.
Couture, C., Deckert, M., Williams, S., Russo, F. O., Altman, A. and Mustelin, T. (1996)
Identification of the site in the Syk protein tyrosine kinase that binds the SH2 domain of Lck.
J Biol.Chem 271, 24294-24299.
Deckert, M., Tartare-Deckert, S., Couture, C., Mustelin, T. and Altman, A. (1996) Functional
and physical interactions of Syk family kinases with the Vav proto-oncogene product.
Immunity 5, 591-604.
Dorahy, D. J., Berndt, M. C. and Burns, G. F. (1995) Capture by chemical crosslinkers
provides evidence that integrin αIIbβ3 forms a complex with protein tyrosine kinases in intact
platelets. Biochem.J. 309, 481-490.
Durfee, T., Becherer, K., Chen, P. L., Yeh, S. H., Yang, Y., Kilburn, A. E., Lee, W. H. and
Elledge, S. J. (1993) The retinoblastoma protein associates with the protein phosphatase type
1 catalytic subunit. Genes Dev. 7, 555-569.

112
El Hillal, O., Kurosaki, T., Yamamura, H., Kinet, J. P. and Scharenberg, A. M. (1997) syk
kinase activation by a src kinase-initiated activation loop phosphorylation chain reaction.
Proc.Natl.Acad.Sci U.S.A 94, 1919-1924.
Ezumi, Y., Takayama, H. and Okuma, M. (1995) Differential regulation of protein-tyrosine
phosphatases by integrin alpha IIb beta 3 through cytoskeletal reorganization and tyrosine
phosphorylation in human platelets. J.Biol.Chem. 270, 11927-11934.
Falati, S., Edmead, C. E. and Poole, A. W. (1999) Glycoprotein Ib-V-IX, a receptor for von
Willebrand factor, couples physically and functionally to the Fc receptor gamma-chain, Fyn,
and Lyn to activate human platelets. Blood 94, 1648-1656.
Falk, E., Shah, P. K. and Fuster, V. (1995) Coronary plaque disruption. Circulation 92, 657-
671.
Fernandez-Ortiz, A., Badimon, J. J., Falk, E., Fuster, V., Meyer, B., Mailhac, A., Weng, D.,
Shah, P. K. and Badimon, L. (1994) Characterization of the relative thrombogenicity of
atherosclerotic plaque components: implications for consequences of plaque rupture.
J.Am.Coll.Cardiol. 23, 1562-1569.
Ferrell, J. E., Jr., Noble, J. A., Martin, G. S., Jacques, Y. V. and Bainton, D. F. (1990)
Intracellular localization of pp60c-src in human platelets. Oncogene 5, 1033-1036.
Fields, S. and Song, O. (1989) A novel genetic system to detect protein-protein interactions.
Nature 20;340, 245-246.
Fluckiger, A. C., Li, Z., Kato, R. M., Wahl, M. I., Ochs, H. D., Longnecker, R., Kinet, J. P.,
Witte, O. N., Scharenberg, A. M. and Rawlings, D. J. (1998) Btk/Tec kinases regulate
sustained increases in intracellular Ca2+ following B-cell receptor activation. EMBO J. 17,
1973-1985.
Gallet, C., Rosa, J. P., Habib, A., Lebret, M., Levy-Toledano, S. and Maclouf, J. (1999)
Tyrosine phosphorylation of cortactin associated with Syk accompanies thromboxane
analogue-induced platelet shape change. J.Biol.Chem. 274, 23610-23616.

113
Gao, J., Zoller, K. E., Ginsberg, M. H., Brugge, J. S. and Shattil, S. J. (1997) Regulation of
the pp72syk protein tyrosine kinase by platelet integrin alpha IIb beta 3. EMBO J 16, 6414-
6425.
Gibbins, J., Asselin, J., Farndale, R., Barnes, M., Law, C. L. and Watson, S. P. (1996)
Tyrosine phosphorylation of the Fc receptor gamma-chain in collagen-stimulated platelets.
J.Biol.Chem. 271, 18095-18099.
Goetzl, E. J. and An, S. (1998) Diversity of cellular receptors and functions for the
lysophospholipid growth factors lysophosphatidic acid and sphingosine 1-phosphate. FASEB
J. 12, 1589-1598.
Golden, A., Nemeth, S. P. and Brugge, J. S. (1986) Blood platelets express high levels of the
pp60c-src-specific tyrosine kinase activity. Proc.Natl.Acad.Sci U.S.A 83, 852-856.
Golden, A. and Brugge, J. S. (1989) Thrombin treatment induces rapid changes in tyrosine
phosphorylation in platelets. Proc.Natl.Acad.Sci U.S.A 86, 901-905.
Gooderham, M. H., Adlercreutz, H., Ojala, S. T., Wahala, K. and Holub, B. J. (1996) A soy
protein isolate rich in genistein and daidzein and its effects on plasma isoflavone
concentrations, platelet aggregation, blood lipids and fatty acid composition of plasma
phospholipid in normal men. J Nutr 126, 2000-2006.
Gotoh, A., Takahira, H., Geahlen, R. L. and Broxmeyer, H. E. (1997) Cross-linking of
integrins induces tyrosine phosphorylation of the proto-oncogene product Vav and the protein
tyrosine kinase Syk in human factor-dependent myeloid cells. Cell Growth Differ. 8, 721-729.
Hackeng, C. M., Huigsloot, M., Pladet, M. W., Nieuwenhuis, H. K., van Rijn HJ and
Akkerman, J. W. (1999) Low-density lipoprotein enhances platelet secretion via integrin-
αIIbβ3-mediated signaling. Arterioscler.Thromb.Vasc.Biol. 19, 239-247.
Hanke, J. H., Gardner, J. P., Dow, R. L., Changelian, P. S., Brissette, W. H., Weringer, E. J.,
Pollok, B. A. and Connelly, P. A. (1996) Discovery of a novel, potent, and Src family-
selective tyrosine kinase inhibitor. Study of Lck- and FynT-dependent T cell activation.

114
J.Biol.Chem. 271, 695-701.
Hers, I., Donath, J., van Willigen, G. and Akkerman, J. W. (1998) Differential involvement of
tyrosine and serine/threonine kinases in platelet integrin alphaIIbbeta3 exposure.
Arterioscler.Thromb.Vasc.Biol. 18, 404-414.
Hoffman, C. S. and Winston, F. (1987) A ten-minute DNA preparation from yeast efficiently
releases autonomous plasmids for transformation of Escherichia coli. Gene 57, 267-272.
Holvoet, P., Vanhaecke, J., Janssens, S., Van de Werf, F. and Collen, D. (1998) Oxidized
LDL and malondialdehyde-modified LDL in patients with acute coronary syndromes and
stable coronary artery disease. Circulation 98, 1487-1494.
Hordijk, P. L., Verlaan, I., van Corven, E. J. and Moolenaar, W. H. (1994) Protein tyrosine
phosphorylation induced by lysophosphatidic acid in Rat-1 fibroblasts. Evidence that
phosphorylation of map kinase is mediated by the Gi-p21ras pathway. J.Biol.Chem. 269, 645-
651.
Hornstein, I., Alcover, A. and Katzav, S. (2004) Vav proteins, masters of the world of
cytoskeleton organization. Cell Signal. 16, 1-11.
Hughes, H., Mathews, B., Lenz, M. L. and Guyton, J. R. (1994) Cytotoxicity of oxidized LDL
to porcine aortic smooth muscle cells is associated with the oxysterols 7-ketocholesterol and
7-hydroxycholesterol. Arterioscler.Thromb. 14, 1177-1185.
Ichinohe, T., Takayama, H., Ezumi, Y., Arai, M., Yamamoto, N., Takahashi, H. and Okuma,
M. (1997) Collagen-stimulated activation of Syk but not c-Src is severely compromised in
human platelets lacking membrane glycoprotein VI. J Biol Chem 272, 63-68.
Jackson, S. P., Schoenwaelder, S. M., Yuan, Y., Salem, H. H. and Cooray, P. (1996) Non-
receptor protein tyrosine kinases and phosphatases in human platelets. Thromb.Haemost. 76,
640-650.
Jennings, L. K., Fox, J. E., Edwards, H. H. and Phillips, D. R. (1981) Changes in the
cytoskeletal structure of human platelets following thrombin activation. J.Biol.Chem. 256,

115
6927-6932.
Keely, P. J. and Parise, L. V. (1996) The alpha2beta1 integrin is a necessary co-receptor for
collagen-induced activation of Syk and the subsequent phosphorylation of phospholipase
Cgamma2 in platelets. J Biol.Chem 271, 26668-26676.
Kimura, T., Sakamoto, H., Appella, E. and Siraganian, R. P. (1996) Conformational changes
induced in the protein tyrosine kinase p72syk by tyrosine phosphorylation or by binding of
phosphorylated immunoreceptor tyrosine-based activation motif peptides. Mol.Cell Biol. 16,
1471-1478.
Klages, B., Brandt, U., Simon, M. I., Schultz, G. and Offermanns, S. (1999) Activation of
G12/G13 results in shape change and Rho/Rho-kinase- mediated myosin light chain
phosphorylation in mouse platelets. J.Cell Biol. 144, 745-754.
Koller, E., Koller, F. and Binder, B. R. (1989) Purification and identification of the
lipoprotein-binding proteins from human blood platelet membrane. J.Biol.Chem. 264, 12412-
12418.
Kurosaki, T., Johnson, S. A., Pao, L., Sada, K., Yamamura, H. and Cambier, J. C. (1995) Role
of the Syk autophosphorylation site and SH2 domains in B cell antigen receptor signaling. J
Exp Med. 182, 1815-1823.
Laemmli, U. K. (1970) Cleavage of structural proteins during the assembly of the head of
bacteriophage T4. Nature 227, 680-685.
Law, C. L., Sidorenko, S. P., Chandran, K. A., Draves, K. E., Chan, A. C., Weiss, A.,
Edelhoff, S., Disteche, C. M. and Clark, E. A. (1994) Molecular cloning of human Syk. A B
cell protein-tyrosine kinase associated with the surface immunoglobulin M-B cell receptor
complex. J Biol.Chem 269, 12310-12319.
Law, C. L., Chandran, K. A., Sidorenko, S. P. and Clark, E. A. (1996) Phospholipase C-
gamma1 interacts with conserved phosphotyrosyl residues in the linker region of Syk and is a
substrate for Syk. Mol.Cell Biol. 16, 1305-1315.

116
Law, D. A., Nannizzi-Alaimo, L., Ministri, K., Hughes, P. E., Forsyth, J., Turner, M., Shattil,
S. J., Ginsberg, M. H., Tybulewicz, V. L. and Phillips, D. R. (1999) Genetic and
pharmacological analyses of Syk function in alphaIIbbeta3 signaling in platelets. Blood 93,
2645-2652.
Leong, L., Hughes, P. E., Schwartz, M. A., Ginsberg, M. H. and Shattil, S. J. (1995) Integrin
signaling: roles for the cytoplasmic tails of alpha IIb beta 3 in the tyrosine phosphorylation of
pp125FAK. J.Cell Sci. 108, 3817-3825.
Lin, T. H., Rosales, C., Mondal, K., Bolen, J. B., Haskill, S. and Juliano, R. L. (1995)
Integrin-mediated tyrosine phosphorylation and cytokine message induction in monocytic
cells. A possible signaling role for the Syk tyrosine kinase. J.Biol.Chem. 270, 16189-16197.
Markwell, M. A., Haas, S. M., Bieber, L. L. and Tolbert, N. E. (1978) A modification of the
Lowry procedure to simplify protein determination in membrane and lipoprotein samples.
Anal.Biochem. 87, 206-210.
Marshall, S. J., Asazuma, N., Best, D., Wonerow, P., Salmon, G., Andrews, R. K. and
Watson, S. P. (2002) Glycoprotein IIb-IIIa-dependent aggregation by glycoprotein Ibalpha is
reinforced by a Src family kinase inhibitor (PP1)-sensitive signalling pathway. Biochem.J.
361, 297-305.
Maruyama, S., Kurosaki, T., Sada, K., Yamanashi, Y., Yamamoto, T. and Yamamura, H.
(1996) Physical and functional association of cortactin with Syk in human leukemic cell line
K562. J Biol.Chem 271, 6631-6635.
Maschberger, P., Bauer, M., Baumann-Siemons, J., Zangl, K. J., Negrescu, E. V., Reininger,
A. J. and Siess, W. (2000) Mildly oxidized low density lipoprotein rapidly stimulates via
activation of the lysophosphatidic acid receptor Src family and Syk tyrosine kinases and Ca2+
influx in human platelets. J.Biol.Chem. 275, 19159-19166.
Meraji, S., Moore, C. E., Skinner, V. O. and Bruckdorfer, K. R. (1992) The importance of
oxidation or glycosylation of low-density lipoproteins in relation to platelet activation.

117
Platelets 3, 155-162.
Miller, L. A., Hong, J. J., Kinch, M. S., Harrison, M. L. and Geahlen, R. L. (1999) The
engagement of beta1 integrins on promonocytic cells promotes phosphorylation of Syk and
formation of a protein complex containing Lyn and beta1 integrin. Eur.J.Immunol. 29, 1426-
1434.
Miranti, C. K., Leng, L., Maschberger, P., Brugge, J. S. and Shattil, S. J. (1998) Identification
of a novel integrin signaling pathway involving the kinase Syk and the guanine nucleotide
exchange factor Vav1. Curr.Biol. 8, 1289-1299.
Moolenaar, W. H., Kranenburg, O., Postma, F. R. and Zondag, G. C. (1997)
Lysophosphatidic acid: G-protein signalling and cellular responses. Curr.Opin.Cell Biol. 9,
168-173.
Negrescu, E. V., Luber de Quintana, K. and Siess, W. (1995) Platelet shape change induced
by thrombin receptor activation. Rapid stimulation of tyrosine phosphorylation of novel
protein substrates through an integrin- and Ca2+-independent mechanism. J.Biol.Chem. 270,
1057-1061.
Negrescu, E. V. and Siess, W. (1996) Dissociation of the αIIbβ3-integrin by EGTA stimulates
the tyrosine kinase pp72syk without inducing platelet activation. J.Biol.Chem. 271, 26547-
26553.
Obergfell, A., Eto, K., Mocsai, A., Buensuceso, C., Moores, S. L., Brugge, J. S., Lowell, C.
A. and Shattil, S. J. (2002) Coordinate interactions of Csk, Src, and Syk kinases with
[alpha]IIb[beta]3 initiate integrin signaling to the cytoskeleton. J.Cell Biol. 157, 265-275.
Ota, Y. and Samelson, L. E. (1997) The product of the proto-oncogene c-cbl: a negative
regulator of the Syk tyrosine kinase. Science 276, 418-420.
Ozdener, F., Dangelmaier, C., Ashby, B., Kunapuli, S. P. and Daniel, J. L. (2002) Activation
of phospholipase Cgamma2 by tyrosine phosphorylation. Mol.Pharmacol 62, 672-679.
Parhami, F., Fang, Z. T., Fogelman, A. M., Andalibi, A., Territo, M. C. and Berliner, J. A.

118
(1993) Minimally modified low density lipoprotein-induced inflammatory responses in
endothelial cells are mediated by cyclic adenosine monophosphate. J.Clin.Invest. 92, 471-478.
Pedreno, J., de Castellarnau, C., Cullare, C., Sanchez, J., Gomez-Gerique J., Ordonez-Lianos,
J. and Gonzalez-Sastre, F. (1992) LDL binding sites on platelets differ from the "classical"
receptor of nucleated cells. Arterioscler.Thromb. 12, 1353-1362.
Pedreno, J., de Castellarnau, C., Cullare, C., Ortin, R., Sanchez, J. L., Llopart, R. and
Gonzalez-Sastre F. (1994) Platelet LDL receptor recognizes with the same apparent affinity
both oxidized and native LDL. Evidence that the receptor-ligand complexes are not
internalized. Arterioscler.Thromb. 14, 401-408.
Pedreno, J., Fernandez, R., Cullare, C., Barcelo, A., Elorza, M. A. and de Castellarnau, C.
(1997) Platelet integrin alpha IIb beta 3 (GPIIb-IIIa) is not implicated in the binding of LDL
to intact resting platelets. Arterioscler.Thromb.Vasc.Biol. 17, 156-163.
Perlmutter, R. M. (1994) Immunodeficiency. Zapping T-cell responses. Nature 370, 249-250.
Poole, A., Gibbins, J. M., Turner, M., van Vugt, M. J., van de Winkel, J. G., Saito, T.,
Tybulewicz, V. L. and Watson, S. P. (1997) The Fc receptor γ-chain and the tyrosine kinase
Syk are essential for activation of mouse platelets by collagen. EMBO J. 16, 2333-2341.
Rasmussen, U. B., Gachet, C., Schlesinger, Y., Hanau, D., Ohlmann, P., Van-Obberghen-
Schilling, E., Pouyssegur, J., Cazenave, J. P. and Pavirani, A. (1993) A peptide ligand of the
human thrombin receptor antagonizes alpha-thrombin and partially activates platelets.
J.Biol.Chem. 268, 14322-14328.
Rezaul, K., Yanagi, S., Sada, K., Taniguchi, T. and Yamamura, H. (1994) Protein-tyrosine
kinase p72syk is activated by platelet activating factor in platelets. Thromb.Haemost. 72, 937-
941.
Rodriguez, R., Matsuda, M., Perisic, O., Bravo, J., Paul, A., Jones, N. P., Light, Y., Swann,
K., Williams, R. L. and Katan, M. (2001) Tyrosine residues in phospholipase Cgamma 2
essential for the enzyme function in B-cell signaling. J.Biol.Chem. 276, 47982-47992.

119
Ross, R. and Glomset, J. A. (1976) The pathogenesis of atherosclerosis (first of two parts).
N.Engl.J Med. 295, 369-377.
Ross, R. and Glomset, J. A. (1976) The pathogenesis of atherosclerosis (second of two parts).
N.Engl.J Med. 295, 420-425.
Ross, R. (1986) The pathogenesis of atherosclerosis-an update. N.Engl.J Med. 20;314, 488-
500.
Ross, R. (1993) The pathogenesis of atherosclerosis: a perspective for the 1990s. Nature 362,
801-809.
Ross, R. (1999) Atherosclerosis-an inflammatory disease. N.Engl.J.Med. 340, 115-126.
Rowley, R. B., Burkhardt, A. L., Chao, H. G., Matsueda, G. R. and Bolen, J. B. (1995) Syk
protein-tyrosine kinase is regulated by tyrosine-phosphorylated Ig alpha/Ig beta
immunoreceptor tyrosine activation motif binding and autophosphorylation. J Biol Chem 270,
11590-11594.
Schagger, H. and von Jagow, G. (1987) Tricine-sodium dodecyl sulfate-polyacrylamide gel
electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal.Biochem.
166, 368-379.
Schiestl, R. H., Dominska, M. and Petes, T. D. (1993) Transformation of Saccharomyces
cerevisiae with nonhomologous DNA: illegitimate integration of transforming DNA into
yeast chromosomes and in vivo ligation of transforming DNA to mitochondrial DNA
sequences. Mol Cell Biol 13, 2697-2705.
Schulz, T., Schiffl, H., Scheithe, R., Hrboticky, N. and Lorenz, R. (1995) Preserved
antioxidative defense of lipoproteins in renal failure and during hemodialysis. Am J Kidney
Dis. 25, 564-571.
Sevanian, A., Bittolo, B. G., Cazzolato, G., Hodis, H., Hwang, J., Zamburlini, A., Maiorino,
M. and Ursini, F. (1997) LDL- is a lipid hydroperoxide-enriched circulating lipoprotein.

120
J.Lipid Res. 38, 419-428.
Shattil, S. J. and Brass, L. F. (1987) Induction of the fibrinogen receptor on human platelets
by intracellular mediators. J Biol.Chem 262, 992-1000.
Shattil, S. J., Cunningham, M., Wiedmer, T., Zhao, J., Sims, P. J. and Brass, L. F. (1992)
Regulation of glycoprotein IIb-IIIa receptor function studied with platelets permeabilized by
the pore-forming complement proteins C5b-9. J Biol.Chem 267, 18424-18431.
Shattil, S. J., O'Toole, T., Eigenthaler, M., Thon, V., Williams, M., Babior, B. M. and
Ginsberg, M. H. (1995) Beta 3-endonexin, a novel polypeptide that interacts specifically with
the cytoplasmic tail of the integrin beta 3 subunit. J Cell Biol. 131, 807-816.
Shattil, S. J. and Ginsberg, M. H. (1997) Perspectives series: cell adhesion in vascular
biology. Integrin signaling in vascular biology. J Clin Invest 100, 1-5.
Shattil, S. J., Kashiwagi, H. and Pampori, N. (1998) Integrin signaling: the platelet paradigm.
Blood 91, 2645-2657.
Sidorenko, S. P., Law, C. L., Chandran, K. A. and Clark, E. A. (1995) Human spleen tyrosine
kinase p72Syk associates with the Src-family kinase p53/56Lyn and a 120-kDa
phosphoprotein. Proc.Natl.Acad.Sci U.S.A 92, 359-363.
Siess, W. (1989) Molecular mechanisms of platelet activation. Physiol Rev. 69, 58-178.
Siess, W. (1990) Platelets in the pathogenesis of atherosclerosis. Adv.Exp.Med.Biol. 273, 119-
127.
Siess, W., Zangl, K. J., Essler, M., Bauer, M., Brandl, R., Corrinth, C., Bittman, R., Tigyi, G.
and Aepfelbacher, M. (1999) Lysophosphatidic acid mediates the rapid activation of platelets
and endothelial cells by mildly oxidized low density lipoprotein and accumulates in human
atherosclerotic lesions. Proc.Natl.Acad.Sci.U.S.A. 96, 6931-6936.
Slyper, A. H. (1994) Low-density lipoprotein density and atherosclerosis. Unraveling the

121
connection [see comments]. JAMA 272, 305-308.
Stupack, D. G., Li, E., Silletti, S. A., Kehler, J. A., Geahlen, R. L., Hahn, K., Nemerow, G. R.
and Cheresh, D. A. (1999) Matrix valency regulates integrin-mediated lymphoid adhesion via
Syk kinase. J.Cell Biol. 144, 777-788.
Taniguchi, T., Kobayashi, T., Kondo, J., Takahashi, K., Nakamura, H., Suzuki, J., Nagai, K.,
Yamada, T., Nakamura, S. and Yamamura, H. (1991) Molecular cloning of a porcine gene
syk that encodes a 72-kDa protein-tyrosine kinase showing high susceptibility to proteolysis.
J Biol.Chem 266, 15790-15796.
Taniguchi, T., Kitagawa, H., Yasue, S., Yanagi, S., Sakai, K., Asahi, M., Ohta, S., Takeuchi,
F., Nakamura, S. and Yamamura, H. (1993) Protein-tyrosine kinase p72syk is activated by
thrombin and is negatively regulated through Ca2+ mobilization in platelets. J.Biol.Chem. 268,
2277-2279.
Torti, M., Tolnai, F. E., Bertoni, A., Sinigaglia, F. and Balduini, C. (1996) Activation of
human platelets by lysophosphatidic acid. Blood Coagul.Fibrinolysis 7, 253-255.
Tsuchida, S., Yanagi, S., Inatome, R., Ding, J., Hermann, P., Tsujimura, T., Matsui, N. and
Yamamura, H. (2000) Purification of a 72-kDa protein-tyrosine kinase from rat liver and its
identification as Syk: involvement of Syk in signaling events of hepatocytes. J
Biochem.(Tokyo) 127, 321-327.
Tsujimura, T., Yanagi, S., Inatome, R., Takano, T., Ishihara, I., Mitsui, N., Takahashi, S. and
Yamamura, H. (2001) Syk protein-tyrosine kinase is involved in neuron-like differentiation of
embryonal carcinoma P19 cells. FEBS Lett. 489, 129-133.
Turner, M., Mee, P. J., Costello, P. S., Williams, O., Price, A. A., Duddy, L. P., Furlong, M.
T., Geahlen, R. L. and Tybulewicz, V. J. (1995) Perinatal lethality and blocked B-cell
development in mice lacking the tyrosine kinase Syk. Nature 378, 298-302.
Wagner, C. L., Mascelli, M. A., Neblock, D. S., Weisman, H. F., Coller, B. S. and Jordan, R.
E. (1996) Analysis of GPIIb/IIIa receptor number by quantification of 7E3 binding to human

122
platelets. Blood 88, 907-914.
Wan, Y., Kurosaki, T. and Huang, X. Y. (1996) Tyrosine kinases in activation of the MAP
kinase cascade by G-protein- coupled receptors. Nature 380, 541-544.
Watson, S. P. and Gibbins, J. (1998) Collagen receptor signalling in platelets: extending the
role of the ITAM. Immunol Today 19, 260-264.
Watson, S. P. (1999) Collagen receptor signaling in platelets and megakaryocytes.
Thromb.Haemost. 82, 365-376.
Weidtmann, A. (1996) Wirkungen von nativem und unterschiedlich stark oxidiertem LDL auf
Thrombozyten. Dissertation, LMU München.
Weidtmann, A., Scheithe, R., Hrboticky, N., Pietsch, A., Lorenz, R. and Siess, W. (1995)
Mildly oxidized LDL induces platelet aggregation through activation of phospholipase A2.
Arterioscler.Thromb.Vasc.Biol. 15, 1131-1138.
Wonerow, P., Obergfell, A., Wilde, J. I., Bobe, R., Asazuma, N., Brdicka, T., Leo, A.,
Schraven, B., Horejsi, V., Shattil, S. J. and Watson, S. P. (2002) Differential role of
glycolipid-enriched membrane domains in glycoprotein VI- and integrin-mediated
phospholipase Cgamma2 regulation in platelets. Biochem.J. 364, 755-765.
Woodside, D. G., Obergfell, A., Leng, L., Wilsbacher, J. L., Miranti, C. K., Brugge, J. S.,
Shattil, S. J. and Ginsberg, M. H. (2001) Activation of Syk protein tyrosine kinase through
interaction with integrin beta cytoplasmic domains. Curr.Biol. 11, 1799-1804.
Woodside, D. G., Obergfell, A., Talapatra, A., Calderwood, D. A., Shattil, S. J. and Ginsberg,
M. H. (2002) The N-terminal SH2 domains of Syk and ZAP-70 mediate phosphotyrosine-
independent binding to integrin beta cytoplasmic domains. J.Biol.Chem.
Wu, Y., Suzuki-Inoue, K., Satoh, K., Asazuma, N., Yatomi, Y., Berndt, M. C. and Ozaki, Y.
(2001) Role of Fc receptor gamma-chain in platelet glycoprotein Ib-mediated signaling. Blood
97, 3836-3845.

123
Yamada, T., Taniguchi, T., Yang, C., Yasue, S., Saito, H. and Yamamura, H. (1993)
Association with B-cell-antigen receptor with protein-tyrosine kinase p72syk and activation
by engagement of membrane IgM. Eur.J Biochem. 213, 455-459.
Yan, S. R., Huang, M. and Berton, G. (1997) Signaling by adhesion in human neutrophils:
activation of the p72syk tyrosine kinase and formation of protein complexes containing
p72syk and Src family kinases in neutrophils spreading over fibrinogen. J.Immunol. 158,
1902-1910.
Yan, S. R. and Novak, M. J. (1999) Beta2 integrin-dependent phosphorylation of protein-
tyrosine kinase Pyk2 stimulated by tumor necrosis factor alpha and fMLP in human
neutrophils adherent to fibrinogen. FEBS Lett. 451, 33-38.
Yanaga, F., Poole, A., Asselin, J., Blake, R., Schieven, G. L., Clark, E. A., Law, C. L. and
Watson, S. P. (1995) Syk interacts with tyrosine-phosphorylated proteins in human platelets
activated by collagen and cross-linking of the Fc gamma-IIA receptor [published erratum
appears in Biochem J 1996 Feb 1;313(Pt 3):1047]. Biochem.J. 311, 471-478.
Yanagi, S., Inatome, R., Ding, J., Kitaguchi, H., Tybulewicz, V. L. and Yamamura, H.
(2001a) Syk expression in endothelial cells and their morphologic defects in embryonic Syk-
deficient mice. Blood 98, 2869-2871.
Yanagi, S., Inatome, R., Takano, T. and Yamamura, H. (2001b) Syk expression and novel
function in a wide variety of tissues. Biochem.Biophys.Res Commun. 288, 495-498.
Yang, L., Yatomi, Y., Hisano, N., Qi, R., Asazuma, N., Satoh, K., Igarashi, Y., Ozaki, Y. and
Kume, S. (1996) Activation of protein-tyrosine kinase Syk in human platelets stimulated with
lysophosphatidic acid or sphingosine 1-phosphate. Biochem.Biophys.Res.Commun. 229, 440-
444.

124
7. Abkürzungsverzeichnis
A Absorption
ADP Adenosindiphosphat
AK Antikörper
APS Ammoniumpersulfat
AT Amidotriazol
BAECs bovine aortic endothelial cells
Bp Basenpaare
BSA bovine serum albumin
Ca2+ Calcium
cAMP cyclisches Adenosin 3�, 5�-monophosphat
cGMP cyclisches Guanosin 3�, 5�-monophosphat
Da Dalton
DMSO Dimethylsulfoxid
DTT Dithiothreitol
ECL enhanced chemiluminiscence
EDTA Ethylendiamintetraacetat
EGTA Ethylenglycoltetraacetat
EtOH Ethanol
FKS (FCS) Fötales Kälberserum (fetal calf serum)
Hepes 4-(2-Hydroxyethyl)-1-piperazinethansulfonat
HMG-CoA Hydroxy-methyl-glutaryl-CoA
HRP horseradish peroxidase
HUVEC human umbilical vein endothelial cells
Ig Immunglobulin
Ilo Iloprost
IP Immunpräzipitation
ITAM immunoreceptor tyrosine activation motif
LB Luria-Bertani (-Medium)
LDL low density lipoprotein
LiAc Lithiumacetat
LPA Lysophosphatidsäure

125
MCS multiple cloning site
MG Molekulargewicht
MLC myosin light chain
PAF platelet-activating factor
PAGE Polyacrylamid-Gelelektrophorese
PBS phosphate buffered saline
PEG Polyethylenglykol
PMSF Phenylmethylsulfonyl-Fluorid
PPP platelet poor plasma
PRP platelet rich plasma
PY Tyrosinphosphorylierung (Y = Tyrosin)
NO Stickstoffmonoxid
rpm rotations per minute
RT Raumtemperatur
S.c. Saccharomyces cerevisiae
SDS Sodiumdodecylsulfate
S1-P Sphingosin 1-phosphat
ss-DNA single-stranded DNA
TBE Tris-Borat-EDTA
TEMED N,N,N´,N´-Tetramethylethylendiamin
TRAP Thrombin-Rezeptor aktivierendes Protein
TRIS Tris-(hydroxymethyl)-aminomethan
Ü/N über Nacht

126
Anhang
Eigene Veröffentlichungen
1. Originalarbeiten
Miranti, C., Leng, L., Maschberger, P., Brugge, J.S., Shattil, S.J.: Identification of a novel
integrin signaling pathway involving the kinase Syk and the guanine nucleotide exchange
factor Vav1. Curr Biol 8(24):1289-1299(1998).
Bauer, M., Retzer, M., Wilde, J.I., Maschberger, P., Essler, M., Aepfelbacher, M., Watson, S.,
Siess, W.: Dichotomous regulation of myosin phosphorylation and shape change by Rho-
kinase and calcium in intact human platelets. Blood 94(5):1665-72(1999)
Maschberger, P., Bauer, M., Baumann-Siemons, J., Zangl, K.J., Negrescu, E.V., Reininger,
A.J., Siess, W.: Mildly oxidized LDL stimulates via activation of the lysophosphatidic acid
receptor rapidly Src-family and Syk tyrosine kinases, and Ca2+-influx in human platelets. J.
Biol.Chem. 275(25):19159-66(2000).
Bauer, M., Maschberger, P., Quek L., Briddon S.J., Dash D., Weiss M., Watson S.P., Siess,
W.: Genetic and pharmacological analyses of involvement of Src-family, Syk and Btk
tyrosine kinases in platelet shape change. Src-kinases mediate integrin alphaIIb beta3 inside-
out signalling during shape change. Thromb. Haemost. 85(2):331-40(2001).
Bobe, R., Wilde J.I., Maschberger, P., Essler M., Aepfelbacher M., Watson, S.P., Siess, W.:
Phosphatidylinositol 3-kinase-dependent translocation of phospholipase Cgamma2 in mouse
megakaryocytes is independent of Bruton tyrosine kinase translocation. Blood 97(3):678-
84(2001).
Maschberger, P., Watson, S.P., Siess, W.: Integrin αIIbβ3 activation stimulates the Src-kinases
mediated tyrosine phosphorylation of the Fc receptor γ-chain, Syk activation and
phospholipase Cγ2 tyrosine phosphorylation in human platelets. Manuskript in Vorbereitung

127
2. Buchbeiträge
Hato, T. Maschberger, P., Shattil, S.J.: Integrin signaling in platelets. Method Book: Signaling
through Cell Adhesion Molecules. CRC Press (2000).
3. Kongreßbeiträge, Abstracts, Vorträge
a) Vorträge
Maschberger, P., Negrescu, E.V., Baumann, J., Reininger, A.J., Siess, W.: Activation of the
protein tyrosine kinase p72Syk but not p125FAK during platelet shape change. 6th Erfurt
conference on Platelets organised on behalf of the European Platelet Group, Erfurt, 12.-
15.05.1996;
Platelets (1996) 7/92
Maschberger, P.: Activation of Syk during platelet shape change induced by mox-LDL and
YFLLRNP. 1st Platelet Signal Transduction Symposium Munich-Oxford, München, 28.-
29.09.1996
Maschberger, P.: Signaltransduction in platelets: Regulation and function of the tyrosine
kinase p72Syk during the initial platelet activation. Seminar, Department of Vascular Biology,
Scripps Research Institute, La Jolla, USA, 04.12.1997
Maschberger, P.: Regulation of the tyrosine kinase p72Syk during shape change, spreading and
aggregation of human platelets. Seminar, Department of Pharmacology, University of Oxford,
UK, 01.05.1998
Maschberger, P., Bauer, M., Negrescu, E.V., Shattil, S.J., Siess, W.: Regulation of the
tyrosine kinase p72Syk during platelet shape change, spreading and aggregation of human
platelets. ETRO-Euroconference, Paris, 14.-15.05.1998
Siess, W., Bauer, M., Bauer, K., Maschberger, P., Briddon, S.J., Watson, S.P.: Regulation of
platelet shape change and spreading. 1st UK Platelet Meeting, Oxford, UK, 23.-24.06.1998

128
Siess, W., Bauer, M., Maschberger, P., Retzer, M., Essler, M., Aepfelbacher, M., Briddon,
S.J., Wilde, J.I., Quek, L.S., Watson, S.P.: Regulation of platelet shape change and platelet
spreading. Fourth International Dahlem Symposium on Cellular Signal Recognition and
Transduction, Berlin, 08.-10.10.1998
b) Poster
Maschberger, P., Baumann, J., Negrescu, E.V., Siess, W.: Role of the tyrosine kinase Syk in
the initial platelet response induced by mildly oxidized LDL. Poster, 6th International
Lipoprotein Symposium, Hamburg, 29.05. - 01.06.1997
Bauer, M., Retzer, M., Wilde, J.I., Maschberger, P., Essler, M., Aepfelbacher, M., Watson,
S.P., Siess, W.: Rho kinase and calcium regulate myosin phosphorylation and shape change in
intact human platelets independently of each other. XIV. Plättchensymposium, Schloß
Oppurg, 15.-17.10.1999;
Platelets (2000) 11/111

129
DanksagungDanksagungDanksagungDanksagung Herrn Prof. Dr. P.C. Weber danke ich für die Möglichkeit, an seinem Institut diese interessante Arbeit durchzuführen. Mein besonderer Dank gilt Herrn Prof. Dr. W. Siess für seine engagierte Förderung der Arbeit, für viele wertvolle Anregungen und Diskussionen und seine Unterstützung bei der Planung und Durchführung der Auslandsaufenthalte. Bei Dr. E.V. Negrescu, Dr. M. Bauer, Dr. M. Otte, Dr. C. Johannes und allen anderen Kolleginnen und Kollegen im Labor bedanke ich mich für die anregenden Diskussionen, die Hilfsbereitschaft und die gute Zusammenarbeit. Frau C. Meister und Frau U. Wielert danke ich für die vielen praktischen Hilfestellungen und die nette Arbeitsatmosphäre. Herrn Prof. Dr. S.J. Shattil danke ich für die äußerst lehrreiche und interessante Zeit in seinem Labor am Scripps Research Institute in La Jolla, USA. Herrn Dr. S.P. Watson, Institute of Pharmacology, Oxford, UK danke ich für die Möglichkeit, einige Experimente mit k.o.-Mäusen in seinem Labor durchzuführen. Herrn Prof. Dr. Dr. J. Heesemann und Herrn Dr. K.H. Trebesius, Max-von-Pettenkofer Institut danke ich für die Möglichkeit der Benutzung des konfokalen Lasermikroskops und die Hilfestellung. Bei Herrn Prof. F. Hofmann, Frau Dr. C. Werner und Herrn Dr. M. Sausbier, Institut für Pharmakologie, TU München sowie bei Prof. Dr. J. van de Winkel und Dr. M. van Vugt, Utrecht, Niederlande bedanke ich mich für das Bereitstellen von Mausblutproben. Frau Dr. B. Kehrel, Universität Münster danke ich für das Zusenden von Blutproben von Thrombasthenia Glanzmann Patienten.

130
Lebenslauf I. Persönliche Daten Name Petra Margit Maschberger Geburtsdatum 14. April 1971 Geburtsort Altötting II. Ausbildung und berufliche Tätigkeit 09.1977 - 07.1981 Grundschule Marktl 09.1981 - 06.1990 König-Karlmann Gymnasium Altötting 10.1990 - 09.1995 Studium der Biotechnologie an der Technischen Universität Berlin Abschluß: Dipl.-Ing. Fachrichtung Biotechnologie 11.1995 - 05.1999 Promotion am Institut für Kreislaufforschung, Graduiertenkolleg Vaskuläre Biologie, Prof. W. Siess, LMU München WS 99/00 - WS 02/03 Studium der Pharmazie an der LMU München WS 99/00 - SS 02 Mitarbeit in Renner´schen Apotheke, München 06.2003 - 11.2003 1. Praktisches Halbjahr: Renner´sche Apotheke, München 04.2004 - 09.2004 2. Praktisches Halbjahr: Fujisawa Deutschland GmbH,
München; Bereich Herz/Kreislauf III. Auszeichnungen und Auslandsaufenthalte 11.07.96 Erwin-Stephan-Preis der TU Berlin 07.1997 - 12.1997 Auslandsaufenthalt am Scripps Research Institute, La Jolla,
USA 06.04. - 03.05.1998, 07.12. - 14.12.1998, Auslandsaufenthalte am Institute of Pharmacology, 17.03. - 20.03.1999 University of Oxford, UK

![Gurū Nānak Gurmukhi & Transliteration für deutsche ... srw guxw ky gwh ] suixAY syK pIr pwiqswh ] suixAY ADy pwvih rwhu ] suixAY hwQ hovY Asgwhu ] nwnk Bgqw sdw ivgwsu ] suixAY](https://img.pdfslide.org/doc/110x75/5af994667f8b9a19548cd241/guru-nanak-gurmukhi-transliteration-fr-deutsche-srw-guxw-ky-gwh-suixay-syk.jpg)