Embed Size (px)
Citation preview

ORIGI NALARBEIT
FunktionelleMagnetresonanztomographie in derpsychopathologischen Forschung
Zusammenfassung: Psychiatrische Krankheitsbilder sinddurch psychopathologische Symptome charakterisiert, denenfunktionelle zentralnervöse Zustände entsprechen. Die funktionelle Magnetresonanztomographie (fMRT) wird seit wenigenJahren zur nichtinvasiven Abbildung zerebraler Aktivierungsmuster beim Menschen eingesetzt. Nach einer Darstellung der neurobiologischen Grundlagen und Voraussetzungen der Methodewerden die Ergebnisse unserer Arbeitsgruppe zu einer Reihe elementarer sensomotorischer Leistungen und höherer kognitiverFunktionen wie Arbeitsgedächtnis und semantische Repräsentationen referiert. Hieran schließt sich eine Diskussion der vorläufigen Ergebnisse von fMRT-Studien im psychopathologischenKontext, wobei Halluzinationen, Psychomotorik, Emotionen undZwang paradigmatisch für den gesamten Bereich psychiatrischerPhänomenologie stehen. Die Methodik der fMRT hat bestimmtephysikalische Grenzen und wirft praktische Probleme auf, diebeim Einsatz in der Psychiatrie zu berücksichtigen sind. Zu diesenProblembereichen gehören der Signalrauschabstand, die Methodik der Datenanalyse, die Bewegungsartefaktkontrolle und einbesseres Verständnis der neurovaskulären Kopplung. Aufgrundder raschen Entwicklung des Forschungsfeldes rückt die Kartierung höherer geistiger Leistungen und deren Pathologie in greifbare Nähe.
Functional Magnetic Resonance Imaging in Psychopathological Research: Mental disorders are characterised by psychopathological symptoms which correspond to functional brainstates. Functional magnetic resonance imaging (fMRI) is used forthe non-invasive study of cerebral activation patterns in man.First of all, the neurobiological principles and presuppositions ofthe method are outlined. Results from the Heidelberg imaginglab on several simple sensorimotor tasks as weil as higher cognitive functions, such as working and semantic memory, are thenpresented. Thereafter, results from preliminary fMRI studies ofpsychopathological symptoms are discussed, with emphasis onhallucinations, psychomotoric phenomena, emotions, as weil asobsessions and compulsions. Functional MRI is Iimited by thephysics underlying the method, as weil as by practical constraints regarding its use in conjunction with mentally iII patients. Within this framework, the problems of signal-to-noise
Fortsehr. Neural. Psychiat. 66 (1998) 241 - 258© Georg Thieme Verlag Stuttgart . New York
M. Spitzer', Th. Kammer2, M. E. Bellemann3
, G. Brix3,
B. Layer', Sabine Maier1, U. Kischka4
, F. Gückel 5
, Psychiatrische Universitätsklinik, Ulm2 Max-Planck-Institut für biologische Kybernetik, Tübingen3 Forschungsschwerpunkt Radiologische Diagnostik und Therapie,
Deutsches Krebsforschungszentrum (dkfz), Heidelberg4 Rehaklinik Rheinfelden, Schweiz5 Institut für Klinische Radiologie, Klinikum Mannheim, Klinische
Fakultät II der Universität Heidelberg
ratio, data analysis strategies, motion correction, and neurovascular coupling are considered. Because ofthe rapid developmentof the fjeld of fMRI, maps of higher cognitive functions and theirrespective pathology seem to be coming within easy reach.
• 1. Einleitung
Die funktionelle Magnetresonanztomographie (fMRT) wurdeAnfang der 90er Jahre am Massachusetts General Hospital(MGH) in Boston entwickelt [7,51] und ist heute weltweit eineder wichtigsten Methoden bei der Erforschung geistiger Funktionen. Die Methode ist für den Menschen nach dem heutigenKenntnisstand ungefährlich und bietet sich daher zur Untersuchung psychopathologischer Syndrome und Symptome an[70]. Die vorliegende Arbeit gibt anhand eigener Untersuchungen zunächst einen Überblick über die Methode, woran sichdie Darstellung einiger Ergebnisse zu höheren geistigen Leistungen und psychopathologischen Fragestellungen anschließt. Abschließend werden die Möglichkeiten und Grenzen der Methode für die psychopathologische Forschung kritisch diskutiert.
Funktionelle bildgebende Verfahren spielen in der Psychiatrieeine zunehmende Rolle. Diese Tatsache folgt bereits definitionsgemäß aus der gegenwärtigen Konzeptualisierung psychiatrischer Erkrankungen als Störungen der Funktion bestimmter neuronaler Informationsverarbeitungssysteme ohne zugleich bestehende makroanatomische Abnormitäten. Andersformuliert: Bei schizophrenen, depressiven, Zwangs- oderAngstsyndromen geht es nicht um anatomisch-strukturelleDefizite mit komplettem Ausfall bestimmter zentralnervöserFunktionen, sondern um gestörte Funktion bzw. das gestörteZusammenspiel von wahrscheinlich mehreren Funktionen.Verfahren, die solche Funktionszustände abzubilden in derLage sind, sollten daher der Erforschung psychiatrischer Erkrankungen dienlich und damit zukünftig für Diagnostik undTherapie relevant sein.
Funktionelle Zustände des Gehirns werden seit Berger [8] mitelektrophysiologischen Methoden wie der Elektroenzephalographie (EEG) und neuerdings der Magnetenzephalographie(MEG) in der Psychiatrie untersucht. Sie besitzen eine sehrhohe zeitliche Auflösung, sind jedoch in Hinblick auf ihreräumliche Auflösung limitiert (Abb. 1).
241
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.

242 Fortschr. Neural. Psychiat. 66 (1998) M. Spitzer et al.
Raum
-3 -2 -1 0 1 2 3 4 5 6 7Millisekunde Sekunde Minute Stunde Tag Ze~
Neben den elektrophysiologischen Methoden, die neuronaleFunktion direkt erfassen können, ist aufgrund der neurovaskulären Kopplung die Abbildung von Funktionszuständen einzelner Hirnareale auch durch Messung hämodynamischer Parameter möglich. Mit der Einführung computerisierterSchnittbildtechniken war die räumlich hochauflösende Abbildung funktioneller Zustände des ZNS mittels Einbringen radioaktiver Substanzen (Radionuklide) in den Körper möglichgeworden. Je nach Art des eingesetzten Nuklids lassen sichentweder ein Photon (single photon emission computerizedtomography, SPECT) oder zwei Photonen (Positronenemissionstomographie, PET) pro Kernzerfall nachweisen [68,83].
lokalen zerebralen Blutfluß und Blutvolumen bewirkt. DieserMechanismus der neurovaskulären Kopplung [65] führt zu einer Zunahme des oxygenierten Bluts in Bereichen vermehrterneuronaler Aktivität. Hierdurch wird der Sauerstoffbedarf derZellen deutlich überkompensiert, so daß es insgesamt in aktiven Bereichen zu einer Zunahme des Oxyhämoglobins kommt(Abb. 2). Im Magnetresonanztomographen führt das vermehrte Oxyhämoglobin zu einer Zunahme der lokalen Magnetfeldhomogenität, was zu einer Verminderung der Spindephasierung und dadurch zu einem Anstieg der Signalintensität inT2*-gewichteten MR-Bildern führt (Abb.3). Man benennt denbildgebenden Kontrast mit dem Akronym BOLD (Blood Oxygenation Level Dependent) [59].
Abb. 2 Neurophysiologische Grundlage der fMRT (BOLD-Kontrast).Oben ist schematisch ein Areal kortikaler Neuronen in Ruhe dargestellt.Im benachbarten Gefäß symbolisieren hellrote und dunkelrote Erythrozyten ein für den Ruhezustand charakteristisches Blutvolumen mit einer charakteristischen Relation von oxygeniertem und deoxygeniertemBlut. Unten sind die kortikalen Neuronen elektrisch aktiv. Damit verbundene vermehrte Stoffwechselprozesse führen aufgrund der regional sehrfein aufgelösten Autoregulation des zerebralen Kapillarsystemsin der Nachbarschaft der Neuronen zu einem vermehrten Blutfluß miteiner relativen Zunahme des oxygenierten Blutes (hellrot), verglichenmit dem deoxygenierten Blut (dunkelrot).
~~~~ mehr weniger--invasiv
(RT1
UEKP,MEG I PET
r ) ICT
l fMRTMR
OpticalImaging
CEinzelzellableitungen )
( Patch Clamp ) Licht-mikroskopie
-5
-1
-7
-4
m 0
cm -2
fJ.m -6
mm -3
Abb. 1 Methoden der Hirnforschung am Menschen zur Darstellungfunktioneller Zustände des ZNS. Auf der x-Achse ist die zeitliche Auflösung, auf der y-Achse die räumliche Auflösung der unterschiedlichenMethoden aufgetragen (modifiziert nach [16,31,48]). Aus der Abbildung geht hervor, daß es keine ideale Methode gibt, sondern vielmehrdie Methoden unterschiedlichen Erkenntnisansprüchen genügen. Diehöchste zeitliche Auflösung (im Bereich von Millisekunden) besitzen ereigniskorrelierte Potentiale (EKP) und Magnetfelder in der Magnetenzephalographie (MEG). Die räumliche Auflösung dieser Methoden wird injüngster Zeit besser, liegt aber dennoch noch nicht in Bereichen, in diedie funktionelle Magnetresonanztomographie (fMRT) vorstößt. Diezeitliche Auflösung der fMRT liegt je nach verwendeter Pulssequenz imBereich von 50 Millisekunden bis einigen Sekunden. Die räumliche Auflösung liegt bei etwa 1 -3 mm in der gemessenen Schicht bei einerSchichtdicke von 3 - 5 mm. Demgegenüber erreicht die Positronenemissionstomographie (PET) im günstigsten Fall eine räumliche Auflösung von 4 mm (in Abhängigkeit von Tracer und Gerät).
Im Gegensatz zu den genannten etablierten nuldearmedizinischen Techniken kann bei der fMRT auf den Einsatz ionisierender Strahlung verzichtet werden. Der daher nicht-invasiveAnsatz der fMRT beruht darauf, daß Oxyhämoglobin (Hb02)
und Desoxyhämoglobin (Hb) in den Erythrozyten unterschiedliche magnetische Eigenschaften besitzen: Oxyhämoglobin ist diamagnetisch, Desoxyhämoglobin ist paramagnetisch. Wenn ein kortikales Areal durch entsprechende Stimulation Eingangssignale erhält, nimmt die Aktivität der Neuronen in diesem Areal zu. Nach neuesten Befunden ist dieseZunahme aufgrund der internen Struktur von Neuronenverbänden und der daraus resultierenden Dynamik über weiteStrecken linear [88]. Der daraus resultierende vermehrte Sauerstoffverbrauch resultiert initial in einer sehr kurzzeitigenund lokalisierten [53] Verminderung des oxygenierten Hämoglobins im venösen Kapillarblut, die wiederum eine reflektorische Erweiterung zuführender Arteriolen mit zunehmendem
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.

Funktionelle Magnetresonanztomographie in der psychopathologischen Forschung Fortschr. Neurol. Psychiat. 66 (1998) 243
routinen, wie sie bei der Fehlersuche bei Computern verwendet werden, verglichen werden [93]. Da ein Computer im Regelfall sehr unterschiedliche Subsysteme benutzt, um eineAufgabe zu erfüllen, ist es bei einer Störung oft schwierig zuentscheiden, auf welche Fehlfunktion sie zurückgeht. Wennman den Computer dann dazu bringt, eine bestimmte Funktion immer wieder auszuführen (loop), kann diagnostiziertwerden, ob die betreffende Funktion störungsfrei ausgeführtwird. Analog wird in der Medizin etwa beim Unterberger Tretversuch eine bestimmte Bewegung immer wieder ausgeführt,um so eine Bewegungskomponente besser isolieren zu können.
Übertagerung
1 2 :J <4 , 11 T 11 8 10 11 12 1:1 '4 I~
'"'.,
AnaIcmJe (T,)Differenz
O':f'..~ ..
Abb. 3 Die Inspektion der nativen funktionellen T2 • -gewichteten MRBilder ist zur Beurteilung der Aktivierung unzureichend. Links sind zweiderartige Bilder unter Ruhebedingungen (oben) sowie unter visuellerStimulation (unten) gezeigt, die ohne weitergehende Bearbeitung keine Rückschlüsse auf kortikale Aktivierung erlauben. Diese wird nur dadurch sichtbar, daß man die Bilder unter Ruhebedingungen mit denenunter Aktivierung Bildpunkt für Bildpunkt vergleicht. Dies geschieht imeinfachsten Fall durch Subtraktion, heute in der Regel jedoch durch statistische Verfahren. Man erhält durch den Vergleich ein Bild, das nurdiejenigen Gehirnareale darstellt, deren Aktivität sich unter den veränderten Untersuchungsbedingungen (Betrachten von FlackerlichtversusRuhe) signifikant erhöht (Mitte). Diesem Bild unterlegt man ein herkömmliches strukturelles MR-Bild (Tl-gewichtet). Das hierdurch entstehende Gesamtbild zeigt die anatomischen Verhältnisse und zugleichdie funktionell bei einer bestimmten Aufgabe aktivierten Bereiche(rechts). Unten rechts ist das Aktivierungsprofil des mit dem PFeil rechtsoben markierten Bereichs über den Verlauf der gesamten Meßserie gezeigt. Zur Verbesserung des Signalrauschabstandes wurden drei Ruhezustände (auf der Abszisse mit schwarzen Balken markiert) mit zweiAktivierungszuständen (weiße Balken auf der Abszisse) abgewechselt.In jedem Zustand wurden drei Bilder akquiriert. Auf der Ordinate ist diekortikale Aktivierung in Prozent (bezogen auf die mittlere Aktivität inRuhe) aufgetragen. Der Kurvenverlauf zeigt einen regelhaften Anstiegder kortikalen Aktivierung bei visueller Stimulation.
Aklivierung (T2") Aktivierungsprofil In einem typischen fMRT-Experiment wird das Gehirn derVersuchsperson nacheinander in definierte Zustände versetzt,so daß sich die in diesen Zuständen gemessene Aktivierung inbestimmten Gehirnarealen vergleichen und eindeutig interpretieren läßt.
2. Methodik
Bei der Durchführung von funktionellen magnetresonanztomographischen Messungen ergaben sich sowohl in Hinblickauf die Datengenerierung als auch auf deren Analyse methodische Besonderheiten, die im folgenden anhand unseres eigenen experimentellen Ansatzes erläutert werden.
Allgemeiner Versuchsaufbau
Alle unten beschriebenen eigenen Untersuchungen wurden aneinem klinischen 1,5-Tesla-Ganzkörper-MR-System (Magnetom 63 SP®, Siemens AG, Erlangen), das mit einem konventionellen Gradientensystem ausgestattet ist, durchgeführt. Fürdie fMRT-Untersuchung wurde eine kommerziell erhältliche,zirkular polarisierte Kopfspule zum Senden und Empfangender Hochfrequenzsignale eingesetzt.
Die räumliche Auflösung der fMRT beträgt derzeit je nachMeßsequenz ca. 1-3 mm in der gemessenen Schicht [28](Abb.1). Die zeitliche Auflösung ist physiologisch durch dieReagibilität des Gefäßbettes limitiert: Man kann zwar mit denderzeit modernsten Geräten Bilder im Abstand von 50 ms generieren, diese Bilder besitzen jedoch in Hinblick auf die neuronale Aktivierung keinen entsprechend zeitlich hochaufgelösten Informationswert, da das kapillare Gefäßbett mit einerLatenz von etwa 500 ms und einem komplexen räumlich-zeitlichen Dispersionsmuster (sog. "hämodynamischer Filter") aufAktivitätsänderungen reagiert [23,38,53,57]. Hierbei handeltes sich gleichsam um die ersten nachweisbaren Anzeichen derReaktion, die exponentiell mit einer Zeitkonstante von etwa 3Sekunden ansteigt und nach etwa 5 Sekunden ihr Plateau erreicht hat (weswegen die zeitliche Auflösung der fMRT häufigin dieser Größenordnung in der Literatur angegeben wird [1,17,47,85 J). Das MR-Signal fällt nach der Stimulation exponentiell mit einer Zeitkonstante von etwa 5 Sekunden wieder ab[93].
Die visuelle Darbietung der Stimuli zur gezielten Aktivierungzerebraler Funktionszustände und die Erfassung der Antwortsignale der Probanden über Tastendruck erfolgte während derMR-Untersuchung über einen Stimulationscomputer (Macintosh Quadra® 660 AV, Apple Computer GmbH, München) mitangeschlossenem Videoprojektor (Vision® XG 3800 E, Sharp,Japan). Die Stimuli wurden auf eine Rückprojektionsleinwandprojiziert (Abb.4). Die Versuchspersonen betrachteten die Stimuli über einen Spiegel, der schwenkbar an der Kopfspulebefestigt war. Mit dieser Anordnung war ein Gesichtsfeld vonungefähr 24 0 in der horizontalen und 12 0 in der vertikalenRichtung realisierbar.
Bei Untersuchungen, die die Erfassung einer Reaktion bzw.einer motorischen Antwort erforderten, wurde entweder eineigens entwickelter Fußschalter eingesetzt oder ein Schaltkästchen mit vier Tasten verwendet. Beide waren durch einTelefonkabel mit der Tastatur des Stimulationscomputers verbunden. In Kontrollmessungen wurde zuvor sichergestellt,daß hierdurch keine Artefakte in den MR-Bildern verursachtwerden.
Die Untersuchung bestimmter Funktionen ist nur möglich,wenn diese unter experimentellen Bedingungen reliabel herbeigeführt werden können. Die hierzu in der funktionellenBildgebung verwendeten Stimulationsparadigmen könnenmit medizinischen diagnostischen Tests bzw. mit Diagnose-
Zur Reduktion von Bewegungsartefakten wurde auf eine bequeme Lagerung der Probanden geachtet: Eine weiche Unterlage (Wolldecke), Nackenpolster, bitemporale Kopfpolster sowie ggf. eine Knieunterlage zur Inklination des Beckens sorg-
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.
244 Fortschr. Neural. Psychiat. 66 (1998) M. Spitzer et al.
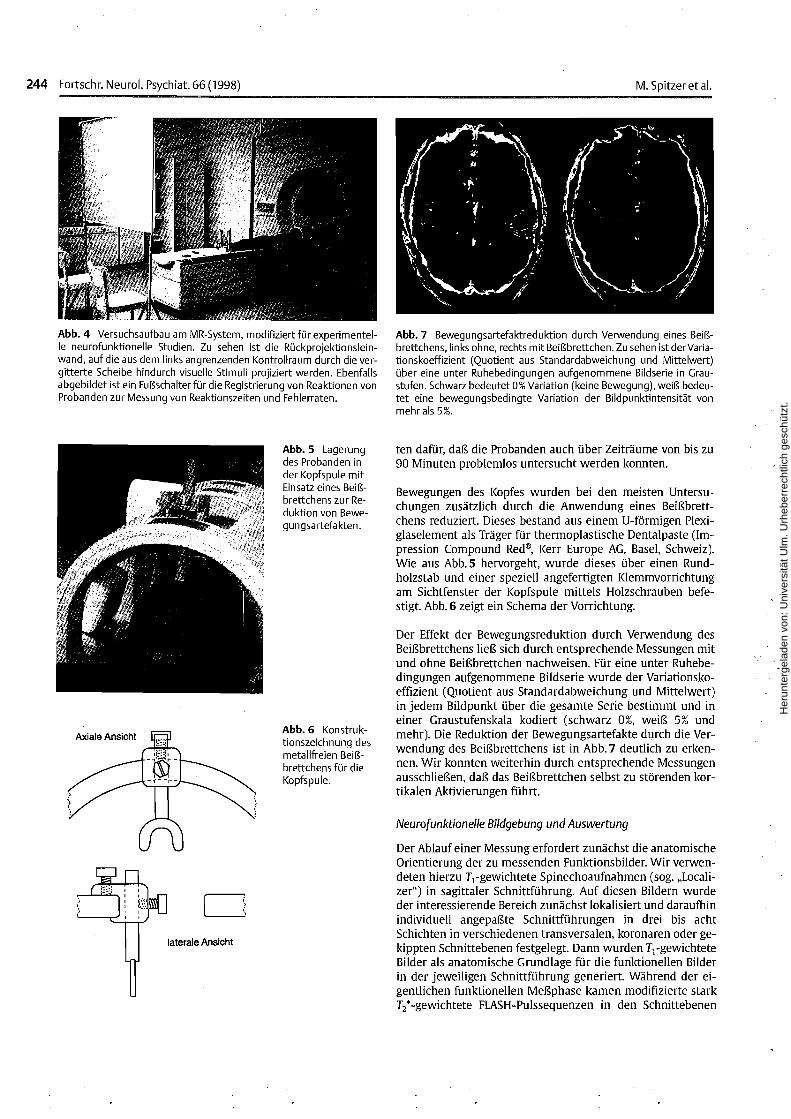
Abb.4 Versuchsaufbau am MR-System, modifiziert für experimentelle neurofunktionelle Studien. Zu sehen ist die Rückprojektionsleinwand, auf die aus dem links angrenzenden Kontrollraum durch die vergitterte Scheibe hindurch visuelle Stimuli projiziert werden. Ebenfallsabgebildet ist ein Fußschalter für die Registrierung von Reaktionen vonProbanden zur Messung von Reaktionszeiten und Fehlerraten.
Abb. 7 Bewegungsartefaktreduktion durch Verwendung eines Beißbrettchens, links ohne, rechts mit Beißbrettchen. Zu sehen ist derVariationskoeffizient (Quotient aus Standardabweichung und Mittelwert)über eine unter Ruhebedingungen aufgenommene Bildserie in Graustufen. Schwarz bedeutet 0% Variation (keine Bewegung), weiß bedeutet eine bewegungsbedingte Variation der Bildpunktintensität vonmehr als 5%.
Bewegungen des Kopfes wurden bei den meisten Untersuchungen zusätzlich durch die Anwendung eines Beißbrettchens reduziert. Dieses bestand aus einem U-förmigen Plexiglaselement als Träger für thermoplastische Dentalpaste (Impression Compound Red®, Kerr Europe AG, Basel, Schweiz).Wie aus Abb.5 hervorgeht, wurde dieses über einen Rundholzstab und einer speziell angefertigten Klemmvorrichtungam Sichtfenster der Kopfspule mittels Holzschrauben befestigt. Abb. 6 zeigt ein Schema der Vorrichtung.
Neurofunktionelle Bildgebung und Auswertung
Der Ablauf einer Messung erfordert zunächst die anatomischeOrientierung der zu messenden Funktionsbilder. Wir verwendeten hierzu Tcgewichtete Spinechoaufnahmen (sog. "Localizer") in sagittaler Schnittführung. Auf diesen Bildern wurdeder interessierende Bereich zunächst lokalisiert und daraufhinindividuell angepaßte Schnittführungen in drei bis achtSchichten in verschiedenen transversalen, koronaren oder gekippten Schnittebenen festgelegt. Dann wurden Tl-gewichteteBilder als anatomische Grundlage für die funktionellen Bilderin der jeweiligen Schnittführung generiert. Während der eigentlichen funktionellen Meßphase kamen modifizierte starkT2*-gewichtete FLASH-Pulssequenzen in den Schnittebenen
Der Effekt der Bewegungsreduktion durch Verwendung desBeißbrettchens ließ sich durch entsprechende Messungen mitund ohne Beißbrettchen nachweisen. Für eine unter Ruhebedingungen aufgenommene Bildserie wurde der Variationskoeffizient (Quotient aus Standardabweichung und Mittelwert)in jedem Bildpunkt über die gesamte Serie bestimmt und ineiner Graustufenskala kodiert (schwarz 0%, weiß 5% undmehr). Die Reduktion der Bewegungsartefakte durch die Verwendung des Beißbrettchens ist in Abb.7 deutlich zu erkennen. Wir konnten weiterhin durch entsprechende Messungenausschließen, daß das Beißbrettchen selbst zu störenden kortikalen Aktivierungen führt.
ten dafür, daß die Probanden auch über Zeiträume von bis zu90 Minuten problemlos untersucht werden konnten.
Abb. 5 Lagerungdes Probanden inder Kopfspule mitEinsatz eines Beißbrettchens zur Reduktion von Bewegungsartefakten.
Abb.6 Konstruktionszeichnung desmetallfreien Beißbrettchens für dieKopfspule.
olaterale Ansicht
Axiale Ansicht
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.

Funktionelle Magnetresonanztomographie in der psychopathalogischen Forschung Fortsehr. Neurol. Psychiat. 66 (1998) 245
Abb.8 Verringerung von Bewegungsartefakten durch Bildnachverarbeitung (Verschiebung und Drehung der Bilder einer Serie). Wie in Abb.7 ist in jedem Bildpunkt der Variationskoeffizient der Bildserie in einerGraustufenskala dargestellt. Links, ohne Bewegungskorrektur; rechts,mit nachträglicher Bewegungskorrektur (aus [43]. mit freundlicher Genehmigung von Elsevier Science, Amsterdam, Niederlande).
zum Einsatz [13]. Die Gesamtmeßzeit für die Untersuchungeines Probanden betrug je nach Fragestellung 60-90 min.
Die Analyse der Bilddatensätze erfolgte auf einer Workstationmit speziell entwickelter Software [13]. Um den störendenEinfluß residualer Kopfbewegungen auf die Datenanalyse weiter zu reduzieren, haben wir in einigen Studien zusätzlich zuden beschriebenen Maßnahmen zur Reduktion von Bewegungsartefakten während der Messung (Lagerung und Beißbrettehen) bei der Auswertung die einzelnen Bilder einer Bildserie so gedreht und verschoben (zweidimensionale affineTransformation), daß sie räumlich möglichst gut zur Deckungkommen (Abb.8).
In der fMRT wird die Aktivierung einzelner Gehirnarealegrundsätzlich durch Vergleich der Signale in Bildern aus zweiunterschiedlichen Funktionszuständen bestimmt. Geschahdies früher durch einfache voxelweise Differenzbildung, wurde in den vergangenen Jahren eine Vielzahl statistischer Vergleichsmethoden eingesetzt. Bis heute hat sich jedoch keineAuswertemethode als Standard herausgebildet.
Zur Detektion aktivierter Kortexareale verwendeten wir diestatistische Subtraktionstechnik mit Hilfe des Wilcoxon-Rangsummen-Tests. Hierbei wurde in einem nichtparametrischenVerfahren die Signifikanz der durch den Stimulus induziertenVerstärkung des MR-Signals gegenüber der Kontrollbedingungin jedem Bildpunkt der Bildmatrix ermittelt. Entsprechendden zugrundeliegenden physiologischen Abläufen und demBOLD-Kontrastmechanismus gingen nur positive induzierteSignaldifferenzen in die Auswertung ein [7,51,60]. Die in einem Bildelement auftretende signifikante Signalerhöhungwurde als relative Signaldifferenz d = (5-50 )/50 der Mittelwerteunter Stimulation (5) und unter der jeweiligen Kontrollbedingung (So) farbig kodiert und auf den entsprechenden Bildpunkt in den Tcgewichteten anatomischen Spinechobildernübertragen (Abb.3). Die von uns verwendeten fünf Abstufungen in der Farbskala entsprechen dabei relativen Signaldifferenzen d von 1 - 5%in Schritten von 1%. Die Orientierung allerAbbildungen folgt der radiologischen Schnittbildkonvention:Ansicht von unten, die linke Gehirnhälfte findet sich im Bildrechts.
Um isolierte signifikante Bildelemente (statistisches Rauschen) zu eliminieren, wurde zusätzlich gefordert, daß in einer 5 x 5-Umgebung eines Pixels mit signifikanter Signalände-
rung mindestens neun weitere signifikante Bildpunkte vorhanden sein müssen. Dieses Kompaktheitskriterium führte zueiner deutlichen Verringerung der Signalkontamination durchzufällig signifikante Pixel (statistischer Fehler erster Art) unddamit zu einer Verbesserung der Spezifität in den fMRT-Aktivierungskarten.
Bei der weiteren Auswertung wurden durch Inspektion derfMRT-Karten die aktivierten kortikalen Areale identifiziert. Zurgenaueren Analyse des Zeitverlaufs der MR-Signalintensitätwurden ausgewählte Kortexareale mit signifikanter Aktivierung manuell mit Hilfe der Region-of-lnterest-(ROl)-Technikmarkiert. Die mittlere Aktivitätsänderung in den selektiertenROls wurde berechnet und anschließend auf die mittlere Signalintensität des jeweiligen Kontrollparadigmas normiert(sog. Aktivierungsprofil; Abb.3). Die bei einzelnen Versuchspersonen auf diese Weise gewonnenen Daten zur Aktivierungunterschiedlicher Gehirnareale konnten dann auf verschiedene Weise gruppenstatistisch weiterverarbeitet werden (s. u.;Abb.19).
3. Experimentelle Untersuchungen primärersensomotorischer Areale
Im folgenden werden kurz paradigmatisch drei Studien zu primären sensorischen und motorischen Arealen vorgestellt.Dies hat zum einen historische Gründe, waren doch die erstenfMRT-Untersuchungen diesen einfach zu aktivierenden Gehirnregionen gewidmet. In systematischer Hinsicht vermittelndiese Studien einen Eindruck von der Leistungsfähigkeit undden Grenzen der Methodik. Für die praktische Arbeit unseresLabors waren diese Replikationsstudien zur Validierung derMeßmethoden und als Basis für die Weiterentwicklung derneuropsychologischen Stimulationsparadigmen unverzichtbar.
a) Visueller Kortex
Die ersten fMRT-Studien wurden am visuellen Kortex von Belliveau et al. [7] und Kwong et al. [51] durchgeführt. In diesenStudien war die Fragestellung weniger auf die kortikale Funktion gerichtet - elektrophysiologische Untersuchungen ausden 60er Jahren hatten die Aktivierung okzipitaler Areale beistroboskopischer Stimulation bereits eindeutig gezeigt [39] als vielmehr auf die Validierung der Meßmethodik anhandeines robusten neurobiologischen Modells. Entsprechend lages unserer Ansicht nach nahe, bei der Implementierung derfMRT-Methodik zunächst Stimulationsuntersuchungen des visuellen Kortex durchzuführen.
Zur Stimulation des visuellen Kortex wurde ein Schachbrettmuster mit Kontrastumkehr bei einer Frequenz von 4 Hz verwendet. Die Schnittführung wurde auf einer mittsagittalenAufnahme so geplant, daß der visuelle Kortex mit drei Schichten erfaßt werden konnte (Abb.9). Im Verlauf des Meßzyklus(12 Minuten Dauer) wechselten sich Ruhephasen (Augen geschlossen, dunkle Umgebung, 3mal 2 Minuten) mit visuellenStimulationsphasen (Schachbrettmuster, 2mal 2 Minuten) ab.Während der lOminütigen Meßzeit wurden in jeder Schicht15 Tz'-gewichtete Bilder akquiriert. Insgesamt wurden dreiProbanden untersucht.
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.
246 Fortschr.l\leurol. Psychiat. 66 (1998)
Abb. 9 Aktivierung des visuellen Kortex durch Kontrastwechselstimulation (alternierendes Schachbrett, 4 Hz). Oben ist die Schnittführungauf einer mittsagittalen Tl-gewichteten Aufnahme für drei Schichtendargestellt. Sie orientiert sich an der Unterkante des Lobus occipitalisund erfaßt vollständig den Sulcus calcarinus mit der Area striata (primärer visueller Kortex und angrenzende Areale). Unten ist die Aktivierunggegenüber Ruhe (Augen geschlossen) in zwei Schichten in Schrittenvon 1 -5% farbkodiert und projiziert auf Tl-gewichtete Bilder dargestellt. Die gute Übereinstimmung der unabhängig voneinander bestimmten Aktivierungsmuster in den beiden angrenzenden Schichtengibt einen Hinweis auf die Validität der Meßmethode.
Der statistische Vergleich der kortikalen Aktivität zwischenden Messungen unter visueller Stimulation und den Messungen in Ruhe ergab bei allen drei Probanden eine deutlicheZunahme der kortikalen Aktivierung in okzipitalen Hirnregionen unter visueller Stimulation.
Die Ergebnisse dieser Pilotstudie zeigten eindrücklich dieDurchführbarkeit von fMRT-Untersuchungen mittels konventioneller, im klinischen Routineeinsatz befindlicher l,5-TeslaMR-Tomographen. Es sei erwähnt, daß mittlerweile eine sehrgenaue "Kartierung" unterschiedlicher visueller Kortexareale(Vl, V2, V3 a, V3 b) mit spezieller MR-Technik (Echo planarimaging, EPI) sowie neu entwickelter Kartierungstechnikenvorgenommen wurde [67,72,86].
b) Motorkortex
Die motorische Repräsentation der verschiedenen Muskelgruppen von Kopf, Rumpf und Extremitäten im Bereich desprimär-motorischen Kortex (Area 4 nach Brodmann) ist durchPenfield [63) durch direkte kortikale Stimulation im Rahmenvon neurochirurgischen Eingriffen auf eindrucksvolle Art dargestellt worden. Zur Darstellung der funktionellen Aktivierung des Motorkortex wurden fünf Probanden aufgefordert,im Verlauf des Meßzyklus (20 Minuten Dauer) die Finger derrechten Hand gegen den Daumen zu bewegen. Dies geschahfür 2mal 4 Minuten, eingebettet in drei 4minütige Ruhepha-
M. Spitzer et al.
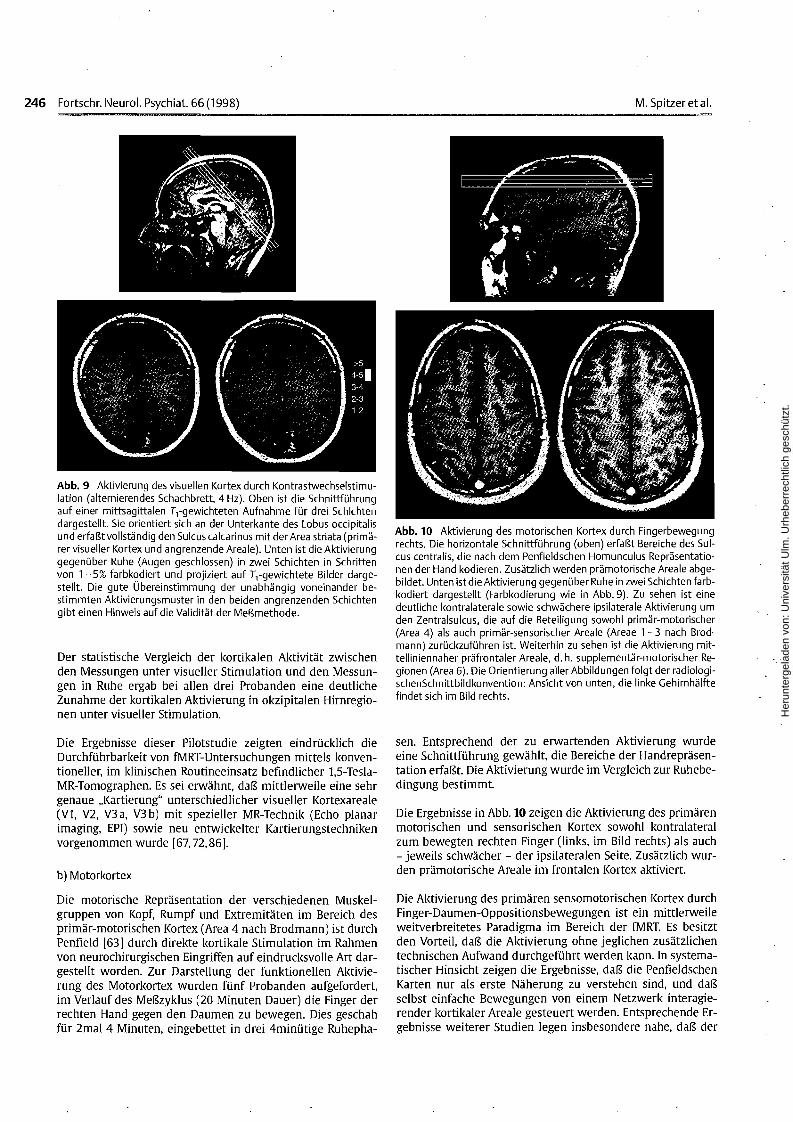
Abb. 10 Aktivierung des motorischen Kortex durch Fingerbewegungrechts. Die horizontale Schnittführung (oben) erfaßt Bereiche des Sulcus centralis, die nach dem Penfieldschen Homunculus Repräsentationen der Hand kodieren. Zusätzlich werden prämotorische Areale abgebildet. Unten ist die Aktivierung gegenüber Ruhe in zwei Schichten farbkodiert dargestellt (Farbkodierung wie in Abb.9). Zu sehen ist einedeutliche kontra laterale sowie schwächere ipsilaterale Aktivierung umden Zentralsulcus, die auf die Beteiligung sowohl primär-motorischer(Area 4) als auch primär-sensorischer Areale (Areae 1-3 nach Brodmann) zurückzuführen ist. Weiterhin zu sehen ist die Aktivierung mittelliniennaher präfrontaler Areale, d. h. supplementär-motorischer Regionen (Area 6). Die Orientierung aller Abbildungen folgt der radiologischenSchnittbildkonvention: Ansicht von unten, die linke Gehirnhälftefindet sich im Bild rechts.
sen. Entsprechend der zu erwartenden Aktivierung wurdeeine Schnittführung gewählt, die Bereiche der Handrepräsentation erfaßt. Die Aktivierung wurde im Vergleich zur Ruhebedingung bestimmt.
Die Ergebnisse in Abb. 10 zeigen die Aktivierung des primärenmotorischen und sensorischen Kortex sowohl kontralateralzum bewegten rechten Finger (links, im Bild rechts) als auch- jeweils schwächer - der ipsilateralen Seite. Zusätzlich wurden prärnotorische Areale im frontalen Kortex aktiviert.
Die Aktivierung des primären sensomotorischen Kortex durchFinger-Daumen-Oppositionsbewegungen ist ein mittlerweileweitverbreitetes Paradigma im Bereich der fMRT. Es besitztden Vorteil, daß die Aktivierung ohne jeglichen zusätzlichentechnischen Aufwand durchgeführt werden kann. In systematischer Hinsicht zeigen die Ergebnisse, daß die PenfieldschenKarten nur als erste Näherung zu verstehen sind, und daßselbst einfache Bewegungen von einem Netzwerk interagierender kortikaler Areale gesteuert werden. Entsprechende Ergebnisse weiterer Studien legen insbesondere nahe, daß der
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.
Funktionelle Magnetresonanztomographie in der psychopathologischen Forschung Fortschr. Neural. Psychiat. 66 (1998) 247
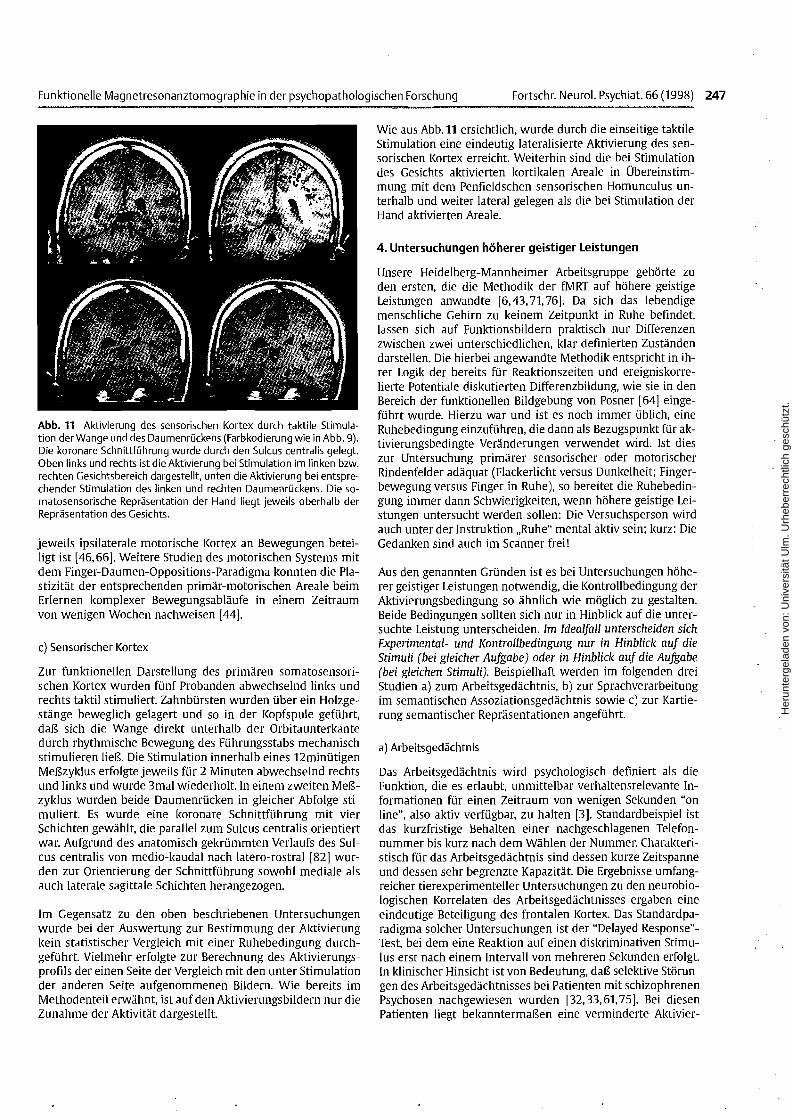
Abb. 11 Aktivierung des sensorischen Kortex durch taktile Stimulation der Wange und des Daumenrückens (Farbkodierung wie in Abb. 9).Die koronare Schnittführung wurde durch den Sulcus centralis gelegt.Oben links und rechts ist die Aktivierung bei Stimulation im linken bzw.rechten Gesichtsbereich dargestellt, unten die Aktivierung bei entsprechender Stimulation des linken und rechten Daumenrückens. Die somatosensorische Repräsentation der Hand liegt jeweils oberhalb derRepräsentation des Gesichts.
jeweils ipsilaterale motorische Kortex an Bewegungen beteiligt ist [46,66]. Weitere Studien des motorischen Systems mitdem Finger-Daumen-Oppositions-Paradigma konnten die Plastizität der entsprechenden primär-motorischen Areale beimErlernen komplexer Bewegungsabläufe in einem Zeitraumvon wenigen Wochen nachweisen [44].
c) Sensorischer Kortex
Zur funktionellen Darstellung des primären somatosensorischen Kortex wurden fünf Probanden abwechselnd links undrechts taktil stimuliert. Zahnbürsten wurden über ein Holzgestänge beweglich gelagert und so in der Kopfspule geführt,daß sich die Wange direkt unterhalb der Orbitaunterkantedurch rhythmische Bewegung des Führungsstabs mechanischstimulieren ließ. Die Stimulation innerhalb eines 12minütigenMeßzyklus erfolgte jeweils für 2 Minuten abwechselnd rechtsund links und wurde 3mal wiederholt. In einem zweiten Meßzyklus wurden beide Daumenrücken in gleicher Abfolge stimuliert. Es wurde eine koronare Schnittführung mit vierSchichten gewählt, die parallel zum Sulcus centralis orientiertwar. Aufgrund des anatomisch gekrümmten Verlaufs des Sulcus centralis von medio-kaudal nach latero-rostral [82] wurden zur Orientierung der Schnittführung sowohl mediale alsauch laterale sagittale Schichten herangezogen.
Im Gegensatz zu den oben beschriebenen Untersuchungenwurde bei der Auswertung zur Bestimmung der Aktivierungkein statistischer Vergleich mit einer Ruhebedingung durchgeführt. Vielmehr erfolgte zur Berechnung des Aktivierungsprofils der einen Seite der Vergleich mit den unter Stimulationder anderen Seite aufgenommenen Bildern. Wie bereits imMethodenteil erwähnt, ist auf den Aktivierungsbildern nur dieZunahme der Aktivität dargestellt.
Wie aus Abb. 11 ersichtlich, wurde durch die einseitige taktileStimulation eine eindeutig lateralisierte Aktivierung des sensorischen Kortex erreicht. Weiterhin sind die bei Stimulationdes Gesichts aktivierten kortikalen Areale in Übereinstimmung mit dem Penfieldschen sensorischen Homunculus unterhalb und weiter lateral gelegen als die bei Stimulation derHand aktivierten Areale.
4. Untersuchungen höherer geistiger leistungen
Unsere Heidelberg-Mannheimer Arbeitsgruppe gehörte zuden ersten, die die Methodik der fMRT auf höhere geistigeleistungen anwandte [6,43,71,76]. Da sich das lebendigemenschliche Gehirn zu keinem Zeitpunkt in Ruhe befindet,lassen sich auf Funktionsbildern praktisch nur Differenzenzwischen zwei unterschiedlichen, klar definierten Zuständendarstellen. Die hierbei angewandte Methodik entspricht in ihrer logik der bereits für Reaktionszeiten und ereigniskorrelierte Potentiale diskutierten Differenzbildung, wie sie in denBereich der funktionellen Bildgebung von Posner [64] eingeführt wurde. Hierzu war und ist es noch immer üblich, eineRuhebedingung einzuführen, die dann als Bezugspunkt für aktivierungsbedingte Veränderungen verwendet wird. Ist dieszur Untersuchung primärer sensorischer oder motorischerRindenfelder adäquat (Flackerlicht versus Dunkelheit; Fingerbewegung versus Finger in Ruhe), so bereitet die Ruhebedingung immer dann Schwierigkeiten, wenn höhere geistige leistungen untersucht werden sollen: Die Versuchsperson wirdauch unter der Instruktion "Ruhe" mental aktiv sein; kurz: DieGedanken sind auch im Scanner frei!
Aus den genannten Gründen ist es bei Untersuchungen höherer geistiger leistungen notwendig, die Kontrollbedingung derAktivierungsbedingung so ähnlich wie möglich zu gestalten.Beide Bedingungen sollten sich nur in Hinblick auf die untersuchte leistung unterscheiden. Im Idealfall unterscheiden sichExperimental- und Kontrollbedingung nur in Hinblick auf dieStimuli (bei gleicher Aufgabe) oder in Hinblick auf die Aufgabe(bei gleichen Stimuli). Beispielhaft werden im folgenden dreiStudien a) zum Arbeitsgedächtnis, b) zur Sprachverarbeitungim semantischen Assoziationsgedächtnis sowie c) zur Kartierung semantischer Repräsentationen angeführt.
a) Arbeitsgedächtnis
Das Arbeitsgedächtnis wird psychologisch definiert als dieFunktion, die es erlaubt, unmittelbar verhaltensrelevante Informationen für einen Zeitraum von wenigen Sekunden "online", also aktiv verfügbar, zu halten [3]. Standardbeispiel istdas kurzfristige Behalten einer nachgeschlagenen Telefonnummer bis kurz nach dem Wählen der Nummer. Charakteristisch für das Arbeitsgedächtnis sind dessen kurze Zeitspanneund dessen sehr begrenzte Kapazität. Die Ergebnisse umfangreicher tierexperimenteller Untersuchungen zu den neurobiologischen Korrelaten des Arbeitsgedächtnisses ergaben eineeindeutige Beteiligung des frontalen Kortex. Das Standardparadigma solcher Untersuchungen ist der "Delayed Response"Test, bei dem eine Reaktion auf einen diskriminativen Stimulus erst nach einem Intervall von mehreren Sekunden erfolgt.In klinischer Hinsicht ist von Bedeutung, daß selektive Störungen des Arbeitsgedächtnisses bei Patienten mit schizophrenenPsychosen nachgewiesen wurden [32,33,61,75]. Bei diesenPatienten liegt bekanntermaßen eine verminderte Aktivier-
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.
248 Fortsehr. Neural. Psychiat. 66 (1998)
*Kontrollbedingung
* * *'-------1
Aktivierungsbedingung
* *'-------1
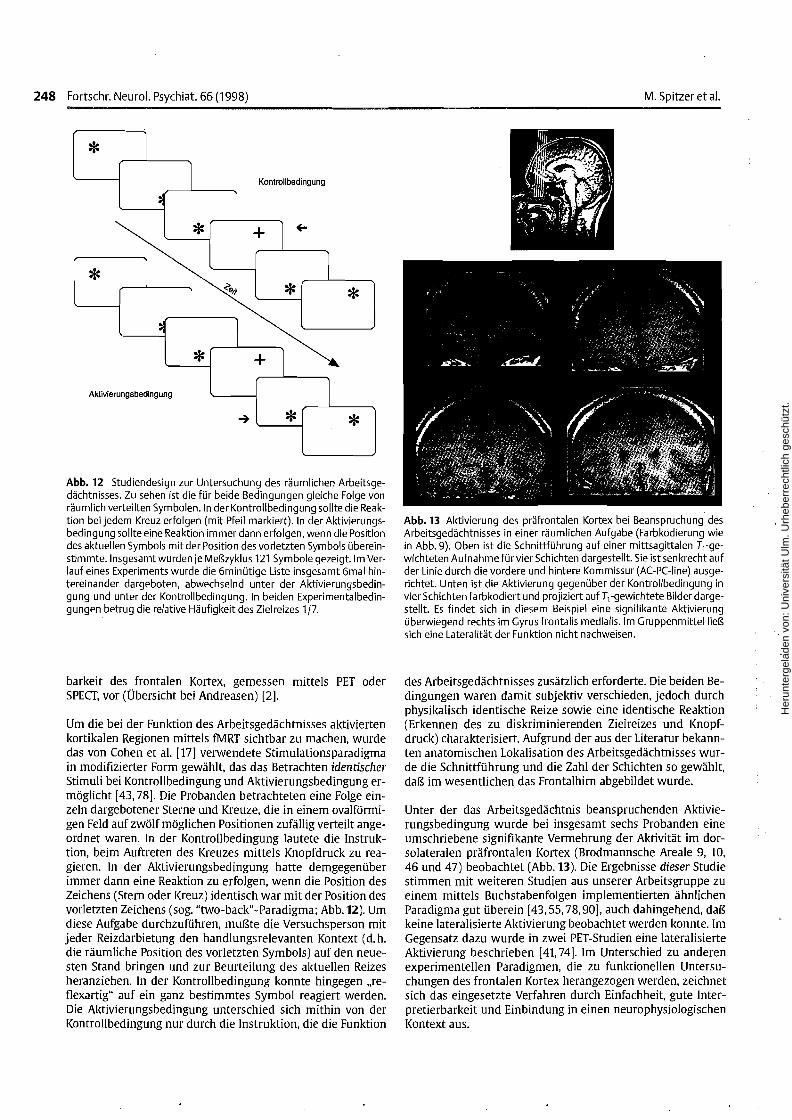
Abb. 12 Studiendesign zur Untersuchung des räumlichen Arbeitsgedächtnisses. Zu sehen ist die für beide Bedingungen gleiche Folge vonräumlich verteilten Symbolen. In der Kontrollbedingung sollte die Reaktion bei jedem Kreuz erfolgen (mit Pfeil markiert). In der Aktivierungsbedingung sollte eine Reaktion immer dann erfolgen, wenn die Positiondes aktuellen Symbols mit der Position des vorletzten Symbols übereinstimmte. Insgesamt wurden je Meßzyklus 121 Symbole gezeigt. Im Verlauf eines Experiments wurde die 6minütige Liste insgesamt 6mal hintereinander dargeboten, abwechselnd unter der Aktivierungsbedingung und unter der Kontrollbedingung. In beiden Experimentalbedingungen betrug die relative Häufigkeit des Ziel reizes 1/7.
barkeit des frontalen Kortex, gemessen mittels PET oderSPECT, vor (Übersicht bei Andreasen) [2].
Um die bei der Funktion des Arbeitsgedächtnisses aktiviertenkortikalen Regionen mittels fMRT sichtbar zu machen, wurdedas von eohen et al. [17] verwendete Stimulationsparadigmain modifizierter Form gewählt, das das Betrachten identischerStimuli bei Kontrollbedingung und Aktivierungsbedingung ermöglicht [43,78]. Die Probanden betrachteten eine Folge einzeln dargebotener Sterne und Kreuze, die in einem ovalförmigen Feld auf zwölf möglichen Positionen zufällig verteilt angeordnet waren. In der Kontrollbedingung lautete die Instruktion, beim Auftreten des Kreuzes mittels Knopfdruck zu reagieren. In der Aktivierungsbedingung hatte demgegenüberimmer dann eine Reaktion zu erfolgen, wenn die Position desZeichens (Stern oder Kreuz) identisch war mit der Position desvorletzten Zeichens (sog. "two-back"-Paradigma; Abb. 12). Umdiese Aufgabe durchzuführen, mußte die Versuchsperson mitjeder Reizdarbietung den handlungsrelevanten Kontext (d.h.die räumliche Position des vorletzten Symbols) auf den neuesten Stand bringen und zur Beurteilung des aktuellen Reizesheranziehen. In der Kontrollbedingung konnte hingegen "ref1exartig" auf ein ganz bestimmtes Symbol reagiert werden.Die Aktivierungsbedingung unterschied sich mithin von derKontrollbedingung nur durch die Instruktion, die die Funktion
M. Spitzer et al.
Abb. 13 Aktivierung des präfrontalen Kortex bei Beanspruchung desArbeitsgedächtnisses in einer räumlichen Aufgabe (Farbkodierung wiein Abb. 9). Oben ist die Schnittführung auf einer mittsagittalen Tl-gewichteten Aufnahme für vier Schichten dargestellt. Sie ist senkrecht aufder Linie durch die vordere und hintere Kommissur (AC-PC-Iine) ausgerichtet. Unten ist die Aktivierung gegenüber der Kontrollbedingung invier Schichten farbkodiert und projiziert auf Tl-gewichtete Bilder dargestellt. Es findet sich in diesem Beispiel eine signifikante Aktivierungüberwiegend rechts im Gyrus frontalis medialis. Im Gruppenmittelließsich eine Lateralität der Funktion nicht nachweisen. .
des Arbeitsgedächtnisses zusätzlich erforderte. Die beiden Bedingungen waren damit subjektiv verschieden, jedoch durchphysikalisch identische Reize sowie eine identische Reaktion(Erkennen des zu diskriminierenden Zielreizes und Knopfdruck) charakterisiert. Aufgrund der aus der Literatur bekannten anatomischen Lokalisation des Arbeitsgedächtnisses wurde die Schnittführung und die Zahl der Schichten so gewählt,daß im wesentlichen das Frontalhirn abgebildet wurde.
Unter der das Arbeitsgedächtnis beanspruchenden Aktivierungsbedingung wurde bei insgesamt sechs Probanden eineumschriebene signifikante Vermehrung der Aktivität im dorsolateralen präfrontalen Kortex (Brodmannsche Areale 9, 10,46 und 47) beobachtet (Abb. 13). Die Ergebnisse dieser Studiestimmen mit weiteren Studien aus unserer Arbeitsgruppe zueinem mittels Buchstabenfolgen implementierten ähnlichenParadigma gut überein [43,55,78,90], auch dahingehend, daßkeine lateralisierte Aktivierung beobachtet werden konnte. 1mGegensatz dazu wurde in zwei PET-Studien eine lateralisierteAktivierung beschrieben [41, 74]. Im Unterschied zu anderenexperimentellen Paradigmen, die zu funktionellen Untersuchungen des frontalen Kortex herangezogen werden, zeichnetsich das eingesetzte Verfahren durch Einfachheit, gute Interpretierbarkeit und Einbindung in einen neurophysiologischenKontext aus.
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.
Funktionelle Magnetresonanztomographie in der psychopathologischen Forschung Fortschr. Neurol. Psychiat. 66 (1998) 249
Tisch Bier WolkeStuhl Traube Hund
**** **** ******** **** ****
Tisch Bier WolkeStuhl Traube Hund
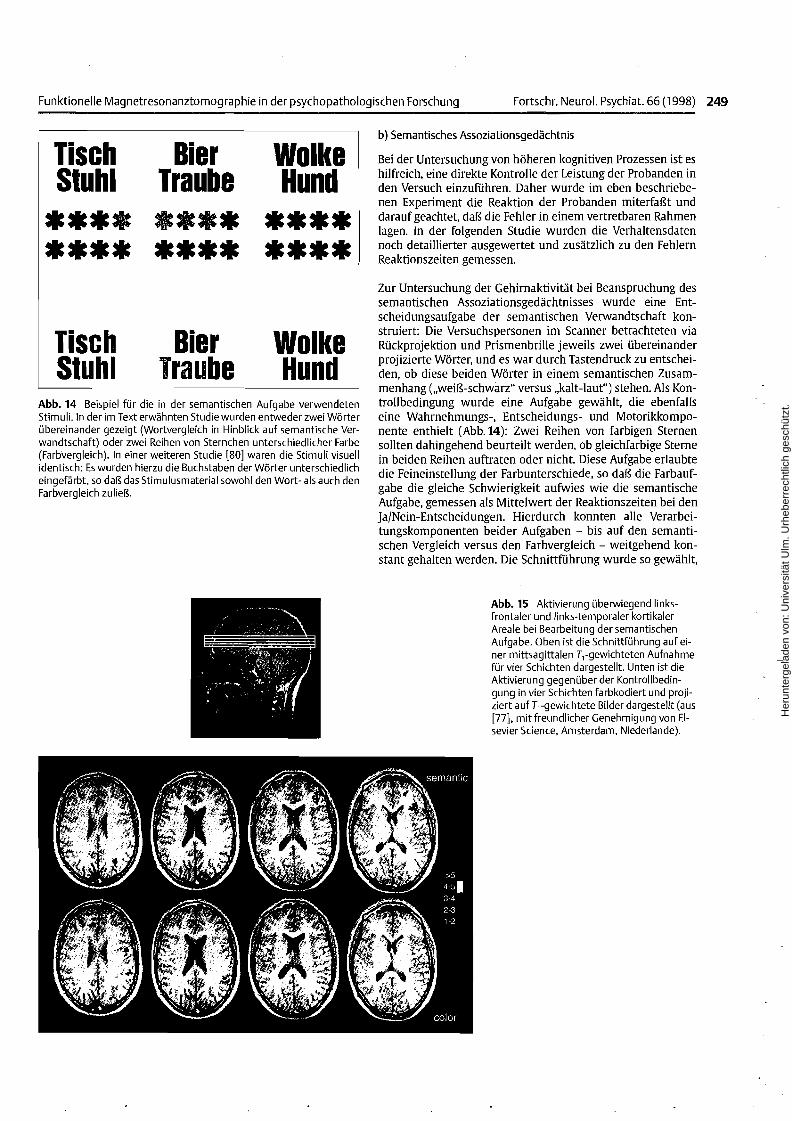
Abb. 14 Beispiel für die in der semantischen Aufgabe verwendetenStimuli. In der im Text erwähnten Studie wurden entweder zwei Wörterübereinander gezeigt (Wortvergleich in Hinblick auf semantische Verwandtschaft) oder zwei Reihen von Sternchen unterschiedlicher Farbe(Farbvergleich). In einer weiteren Studie [80] waren die Stimuli visuellidentisch: Es wurden hierzu die Buchstaben der Wörter unterschiedlicheingefärbt, so daß das Stimulusmaterial sowohl den Wort- als auch denFarbvergleich zuließ.
b) Semantisches Assoziationsgedächtnis
Bei der Untersuchung von höheren kognitiven Prozessen ist eshilfreich, eine direkte Kontrolle der Leistung der Probanden inden Versuch einzuführen. Daher wurde im eben beschriebenen Experiment die Reaktion der Probanden miterfaßt unddarauf geachtet, daß die Fehler in einem vertretbaren Rahmenlagen. In der folgenden Studie wurden die Verhaltensdatennoch detaillierter ausgewertet und zusätzlich zu den FehlernReaktionszeiten gemessen.
Zur Untersuchung der Gehirnaktivität bei Beanspruchung dessemantischen Assoziationsgedächtnisses wurde eine Entscheidungsaufgabe der semantischen Verwandtschaft konstruiert: Die Versuchspersonen im Scanner betrachteten viaRückprojektion und Prismenbrille jeweils zwei übereinanderprojizierte Wärter, und es war durch Tastendruck zu entscheiden, ob diese beiden Wärter in einem semantischen Zusammenhang ("weiß-schwarz" versus "kalt-laut") stehen. Als I<ontrollbedingung wurde eine Aufgabe gewählt, die ebenfallseine Wahrnehmungs-, Entscheidungs- und Motorikkomponente enthielt (Abb.14): Zwei Reihen von farbigen Sternensollten dahingehend beurteilt werden, ob gleichfarbige Sternein beiden Reihen auftraten oder nicht. Diese Aufgabe erlaubtedie Feineinstellung der Farbunterschiede, so daß die Farbaufgabe die gleiche Schwierigkeit aufwies wie die semantischeAufgabe, gemessen als Mittelwert der Reaktionszeiten bei denJa/Nein-Entscheidungen. Hierdurch konnten alle Verarbeitungskomponenten beider Aufgaben - bis auf den semantischen Vergleich versus den Farbvergleich - weitgehend konstant gehalten werden. Die Schnittführung wurde so gewählt,
Abb. 15 Aktivierung überwiegend linksfrontaler und links-temporaler kortikalerAreale bei Bearbeitung der semantischenAufgabe. Oben ist die Schnittführung auf einer mittsagittalen Tl-gewichteten Aufnahmefür vier Schichten dargestellt. Unten ist dieAktivierung gegenüber der Kontrollbedingung in vier Schichten farbkodiert und projiziert auf Tl-geWichtete Bilder dargestellt (aus[77], mit freundlicher Genehmigung von EIsevier Science, Amsterdam, Niederlande).
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.
250 Fortschr. Neurol. Psychiat. 66 (1998)
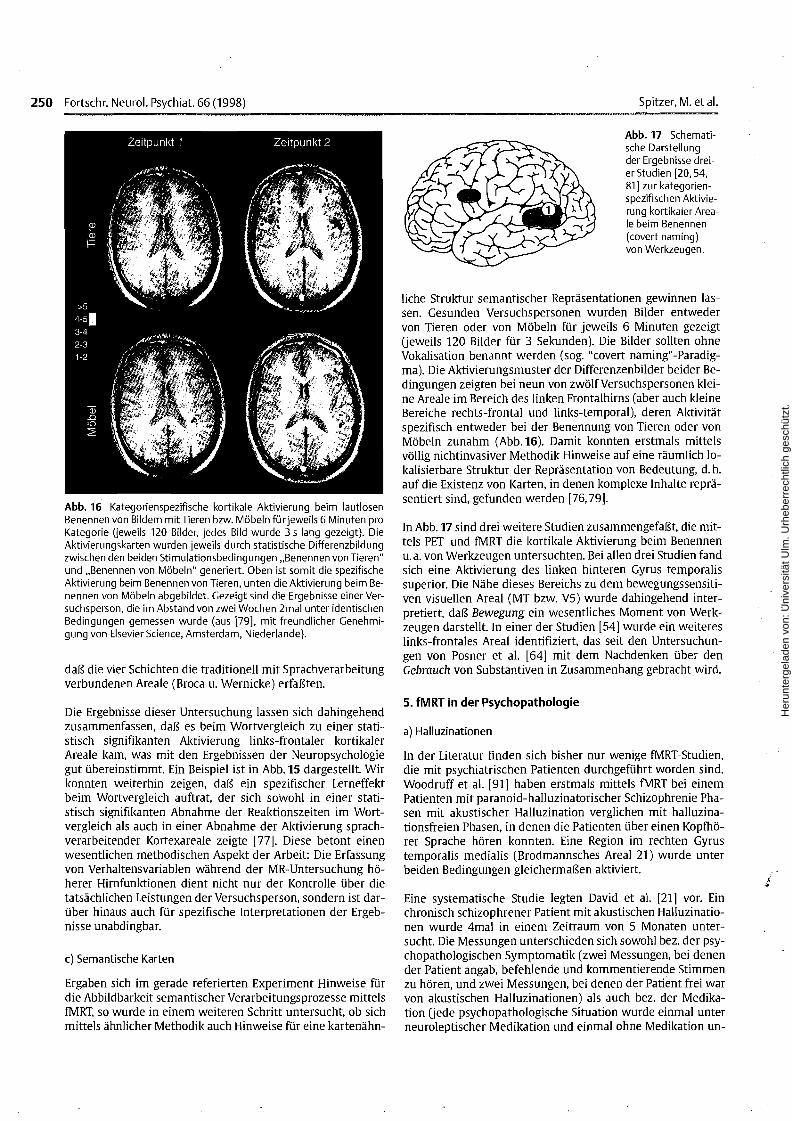
Abb. 16 Kategorienspezifische kortikale Aktivierung beim lautlosenBenennen von Bildern mitTieren bzw. Möbeln für jeweils 6 Minuten proKategorie (jeweils 120 Bilder, jedes Bild wurde 3 s lang gezeigt). DieAktivierungskarten wurden jeweils durch statistische Differenzbildungzwischen den beiden Stimulationsbedingungen "Benennen von Tieren"und "Benennen von Möbeln" generiert. Oben ist somit die spezifischeAktivierung beim Benennen von Tieren, unten die Aktivierung beim Benennen von Möbeln abgebildet. Gezeigt sind die Ergebnisse einer Versuchsperson, die im Abstand von zwei Wochen 2mal unter identischenBedingungen gemessen wurde (aus [79], mit freundlicher Genehmigung von Elsevier Science, Amsterdam, Niederlande).
daß die vier Schichten die traditionell mit Sprachverarbeitungverbundenen Areale (Broca u. Wernicke) erfaßten.
Die Ergebnisse dieser Untersuchung lassen sich dahingehendzusammenfassen, daß es beim Wortvergleich zu einer statistisch signifikanten Aktivierung links-frontaler kortikalerAreale kam, was mit den Ergebnissen der Neuropsychologiegut übereinstimmt. Ein Beispiel ist in Abb. 15 dargestellt. Wirkonnten weiterhin zeigen, daß ein spezifischer Lerneffektbeim Wortvergleich auftrat, der sich sowohl in einer statistisch signifikanten Abnahme der Reaktionszeiten im Wortvergleich als auch in einer Abnahme der Aktivierung sprachverarbeitender Kortexareale zeigte [77]. Diese betont einenwesentlichen methodischen Aspekt der Arbeit: Die Erfassungvon Verhaltensvariablen während der MR-Untersuchung höherer Hirnfunktionen dient nicht nur der Kontrolle über dietatsächlichen Leistungen der Versuchsperson, sondern ist darüber hinaus auch für spezifische Interpretationen der Ergebnisse unabdingbar.
c) Semantische Karten
Ergaben sich im gerade referierten Experiment Hinweise fürdie Abbildbarkeit semantischer Verarbeitungsprozesse mittelsfMRT, so wurde in einem weiteren Schritt untersucht, ob sichmittels ähnlicher Methodik auch Hinweise für eine kartenähn-
Spitzer, M. et al.
Abb. 17 Schematische Darstellungder Ergebnisse dreier Studien [20,54,81] zur kategorienspezifischen Aktivierung kortikaler Areale beim Benennen(covert naming)von Werkzeugen.
liche Struktur semantischer Repräsentationen gewinnen lassen. Gesunden Versuchspersonen wurden Bilder entwedervon Tieren oder von Möbeln für jeweils 6 Minuten gezeigtUeweils 120 Bilder für 3 Sekunden). Die Bilder sollten ohneVokalisation benannt werden (sog. "covert naming"-Paradigmal. Die Aktivierungsmuster der Differenzenbilder beider Bedingungen zeigten bei neun von zwölf Versuchspersonen kleine Areale im Bereich des linken Frontalhirns (aber auch kleineBereiche rechts-frontal und links-temporal), deren Aktivitätspezifisch entweder bei der Benennung von Tieren oder vonMöbeln zunahm (Abb.16). Damit konnten erstmals mittelsvöllig nichtinvasiver Methodik Hinweise auf eine räumlich 10kalisierbare Struktur der Repräsentation von Bedeutung, d. h.auf die Existenz von Karten, in denen komplexe Inhalte repräsentiert sind, gefunden werden [76,79].
In Abb.17 sind drei weitere Studien zusammengefaßt, die mittels PET und fMRT die kortikale Aktivierung beim Benennenu.a. von Werkzeugen untersuchten. Bei allen drei Studien fandsich eine Aktivierung des linken hinteren Gyrus temporalissuperior. Die Nähe dieses Bereichs zu dem bewegungssensitiven visuellen Areal (MT bzw. V5) wurde dahingehend interpretiert, daß Bewegung ein wesentliches Moment von Werkzeugen darstellt. In einer der Studien [54] wurde ein weitereslinks-frontales Areal identifiziert, das seit den Untersuchungen von Posner et al. [64] mit dem Nachdenken über denGebrauch von Substantiven in Zusammenhang gebracht wird.
5. fMRT in der Psychopathologie
a) Halluzinationen
In der Literatur finden sich bisher nur wenige fMRT-Studien,die mit psychiatrischen Patienten durchgeführt worden sind.Woodruff et al. [91] haben erstmals mittels fMRT bei einemPatienten mit paranoid-halluzinatorischer Schizophrenie Phasen mit akustischer Halluzination verglichen mit halluzinationsfreien Phasen, in denen die Patienten über einen Kopfhörer Sprache hören konnten. Eine Region im rechten Gyrustemporalis medialis (Brodmannsches Areal 21) wurde unterbeiden Bedingungen gleichermaßen aktiviert.
Eine systematische Studie legten David et al. [21] vor. Einchronisch schizophrener Patient mit akustischen Halluzinationen wurde 4mal in einem Zeitraum von 5 Monaten untersucht. Die Messungen unterschieden sich sowohl bez. der psychopathologischen Symptomatik (zwei Messungen, bei denender Patient angab, befehlende und kommentierende Stimmenzu hören, und zwei Messungen, bei denen der Patient frei warvon akustischen Halluzinationen) als auch bez. der Medikation Uede psychopathologische Situation wurde einmal unterneuroleptischer Medikation und einmal ohne Medikation un-
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.

Funktionelle Magnetresonanztomographie in der psychopathologischen Forschung Fortschr. Neurol. Psychiat. 66 (1998) 251
tersucht). Die Messungen bestanden aus einer kombiniertenvisuellen und akustischen Stimulation (Lichtblitze und Sprache) und wurden jeweils mit einer unspezifischen Ruhebedingung verglichen. Ausgewertet wurde jeweils die Anzahl deraktivierten Bildpunkte im visuellen und auditorischen Kortex.Dabei war in allen vier Messungen die Erregbarkeit des visuellen Kortex ähnlich, wohingegen sich die Erregbarkeit desauditorischen Kortex signifikant in Abhängigkeit von der psychopathologischen Situation unterschied. Unter der jeweilsidentischen Stimulation mit sprachlichem Material warenwährend Phasen mit akustischen Halluzinationen weniger Voxel aktiviert als während halluzinationsfreier Phasen. Dieserauf den ersten Blick kontraintuitive Befund wird von den Autoren folgendermaßen interpretiert: Akustische Halluzinationen verursachten (oder entstünden durch) neuronale Aktivität im auditorischen Kortex. Sie stünden im Wettbewerb mitakustischen Reizen um das gleiche neuronale Substrat. Daherkönne die (in allen vier Messungen gleich intensive) akustische Stimulation während Halluzinationen, die in der "Ruhemessung" ebenfalls den auditorischen Kortex aktivierten, insgesamt weniger zusätzliche Aktivität hervorrufen.
Diese Studie illustriert unserer Ansicht nach die grundsätzliche Problematik bei der Messung von Funktionszuständen imZusammenhang mit psychopathologischen Symptomen. DieKonzepte zur Genese dieser Symptome sind bez. des neuronalen Substrates noch recht vage, so daß die Ergebnisse funktioneller Messungen einen weiten Interpretationsspielraum offen lassen. Allerdings ist zu erwarten, daß in naher Zukunftweitere entsprechende Untersuchungen durchgeführt werden, so daß Replikationen oder Erweiterungen der bisherigenErgebnisse zu erwarten sind. Eine mittels PET durchgeführteStudie an fünf Patienten [73] zeigte beispielsweise, daß Halluzinationen mit einer Aktivität in überwiegend subkortikalenStrukturen (Basalganglien, Thalamus, Hippokampus) als auchim orbitofrontalen Kortex einhergehen.
b) Psychomotorik
Zu den wenigen bislang vorliegenden fMRT-Studien an schizophrenen Patienten gehört die Untersuchung von Wenz et al.[89] zur Aktivierung des Motorkortex bei schizophrenen Patienten im Vergleich zu normalen Kontrollen. Die Patienten(alle unter Medikation mit Clozapin) und Probanden mußteneinfache Fingerbewegungen abwechselnd rechts und linksdurchführen. Bei den Kontrollpersonen und den schizophrenen Patienten wurden sowohl kontralaterale als auch ipsilaterale sensomotorische und prärnotorische Areale aktiviert.Hierbei zeigten die Patienten insgesamt eine schwächere Aktivierung. Die ausnahmslos rechtshändigen gesunden Probanden zeigten eine charakteristische globale Zunahme der Aktivität bei linkshändiger Bewegung im Vergleich zu rechts. Beiden (ebenfalls nur rechtshändigen) Patienten hingegen fandsich eine Abnahme der Aktivität im Vergleich von Bewegunglinks und rechts. Die Autoren schlußfolgern, daß fMRT in derLage sei, gestörte interhemisphärische Interaktion bei schizophrenen Patienten aufzeigen zu können.
c) Emotionen
Grodd et al. [35] legten in einer Pilotstudie erste Ergebnissezur zerebralen Aktivierung bei verschiedenen definiertenEmotionszuständen vor. Den gesunden Probanden wurden
Portraits mit entweder traurigem, neutralem oder freudigemGesichtsausdruck vorgelegt. In der Aktivierungsbedingungsollten die Probanden versuchen, sich selbst so intensiv wiemöglich in den traurigen bzw. fröhlichen Gefühlszustand zuversetzen. Diese Zustände wurden in den Meßpausen mit einem standardisierten Fragebogen dokumentiert. Als Kontrollbedingung dienten sowohl unspezifische Ruhemessungen alsauch Messungen, bei denen die Probanden Namen zu den präsentierten Gesichtern zuordnen mußten. Es fand sich bei zehnvon zwölf Probanden eine umschriebene Aktivitätszunahmein der linken Amygdala während der traurigen Stimmungsinduktion. Die freudige Stimmung hingegen führte zu keinersignifikanten Aktivitätszunahme.
Bislang liegen nach unserer Kenntnis keine fMRT-Untersuchungen von Patienten mit affektiven Störungen vor.
d) Zwang
Breiter et al. [12] untersuchten zehn Patienten mit Zwangsstörungen und acht gesunde Probanden sowohl unter Kontrollbedingungen als auch unter einer Symptomprovokationsbedingung. Patienten mit Waschzwang wurden beispielsweiseentweder mit einem sauberen oder mit einem vermeintlichschmutzigen Tuch berührt, wodurch zwei Bedingungen geschaffen wurden, die sich somatosensorisch nicht, jedoch inHinblick auf die kognitive Zwangskomponente deutlich unterschieden. Auch Kontrollpersonen erlebten die Provokationsbedingung als sehr unangenehm, zeigten jedoch keine provokationsbedingte gesteigerte Aktivierung. Demgegenüber kames bei Patienten mit Zwangsstörungen unter der Provokationsbedingung zu einem signifikanten Anstieg der Aktivierung im Bereich des paralimbischen und limbischen Systemssowie der Basalganglien und des Temporallappens.
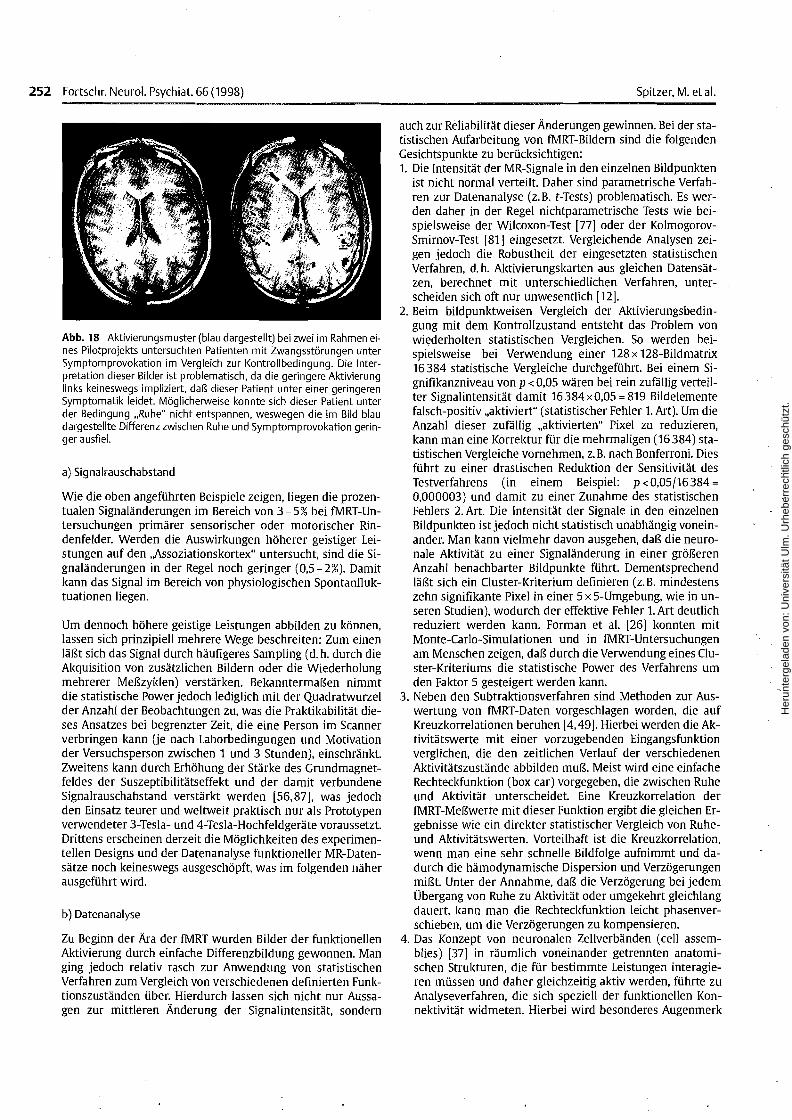
Erste eigene Untersuchungen zur fMRT bei Zwangspatientenzeigten, daß die Untersuchung von Zwangspatienten in derbeschriebenen Weise möglich ist und daß unter Provokationsbedingungen erhebliche Zwangsphänomene evozierbar sind.Auch zeigte sich, daß es durchaus möglich war, mit jedemeinzelnen Patienten die Provokationsbedingung vor der Untersuchung im Detail zu besprechen und das Evozieren derZwangssymptomatik gleichsam zu trainieren. Erste Auswertungen der Bilder ergaben zudem in einigen Fällen signifikante Aktivierungsmuster in frontalen und temporalen kortikalenArealen unter der Provokationsbedingung im Vergleich zurKontrollbedingung (Abb.18).
6. Probleme und mögliche Grenzen
Die Methodik der fMRT hat bestimmte physikalische Grenzenund wirft praktische Probleme auf, was ihren Einsatz, möglicherweise gerade in Hinblick auf psychiatrische Fragestellungen, limitiert. Im folgenden werden sechs Problembereichekurz angesprochen, die derzeit häufig diskutiert werden unddie sich mit den Schlagworten a) Signalrauschabstand, b) Datenanalyse, c) brain maps, d) brain-or-vein und e) Bewegungsartefaktkontrolle umschreiben lassen. Zudem sind wir derAuffassung, daß die derzeit stattfindenden umfangreichenMR-Untersuchungen das Problem von f) Zufallsbefunden ineiner bislang unterschätzten Weise aktualisieren werden.
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.
252 Fortschr. Neurol. Psychiat. 66 (1998)
Abb. 18 Aktivierungsmuster (blau dargestellt) bei zwei im Rahmen eines Pilotprojekts untersuchten Patienten mit Zwangsstörungen unterSymptomprovokation im Vergleich zur Kontrollbedingung. Die Interpretation dieser Bilder ist problematisch, da die geringere Aktivierunglinks keineswegs impliziert, daß dieser Patient unter einer geringerenSymptomatik leidet. Möglicherweise konnte sich dieser Patient unterder Bedingung "Ruhe" nicht entspannen, weswegen die im Bild blaudargestellte Differenz zwischen Ruhe und Symptomprovokation geringer ausfiel.
a) Signalrauschabstand
Wie die oben angeführten Beispiele zeigen, liegen die prozentualen SignaJänderungen im Bereich von 3 - 5% bei tMRT-Untersuchungen primärer sensorischer oder motorischer Rindenfelder. Werden die Auswirkungen höherer geistiger Leistungen auf den ,,Assoziationskortex" untersucht, sind die Signaländerungen in der Regel noch geringer (0,5 - 2%). Damitkann das Signal im Bereich von physiologischen Spontanfluktuationen liegen.
Um dennoch höhere geistige Leistungen abbilden zu können,lassen sich prinzipiell mehrere Wege beschreiten: Zum einenläßt sich das Signal durch häufigeres Sampling (d. h. durch dieAkquisition von zusätzlichen Bildern oder die Wiederholungmehrerer Meßzyklen) verstärken. Bekanntermaßen nimmtdie statistische Power jedoch lediglich mit der Quadratwurzelder Anzahl der Beobachtungen zu, was die Praktikabilität dieses Ansatzes bei begrenzter Zeit, die eine Person im Scannerverbringen kann Ue nach Laborbedingungen und Motivationder Versuchsperson zwischen 1 und 3 Stunden), einschränkt.Zweitens kann durch Erhöhung der Stärke des Grundmagnetfeldes der Suszeptibilitätseffekt und der damit verbundeneSignalrauschabstand verstärkt werden [56,87], was jedochden Einsatz teurer und weltweit praktisch nur als Prototypenverwendeter 3-Tesla- und 4-Tesla-Hochfeldgeräte voraussetzt.Drittens erscheinen derzeit die Möglichkeiten des experimentellen Designs und der Datenanalyse funktioneller MR-Datensätze noch keineswegs ausgeschöpft, was im folgenden näherausgeführt wird.
b) Datenanalyse
Zu Beginn der Ära der tMRT wurden Bilder der funktionellenAktivierung durch einfache Differenzbildung gewonnen. Manging jedoch relativ rasch zur Anwendung von statistischenVerfahren zum Vergleich von verschiedenen definierten Funktionszuständen über. Hierdurch lassen sich nicht nur Aussagen zur mittleren Änderung der Signalintensität, sondern
Spitzer, M. et al.
auch zur Reliabilität dieser Änderungen gewinnen. Bei der statistischen Aufarbeitung von tMRT-Bildern sind die folgendenGesichtspunkte zu berücksichtigen:1. Die Intensität der MR-Signale in den einzelnen Bildpunkten
ist nicht normal verteilt. Daher sind parametrische Verfahren zur Datenanalyse (z.B. t-Tests) problematisch. Es werden daher in der Regel nichtparametrische Tests wie beispielsweise der Wi1coxon-Test [77] oder der KolmogorovSmirnov-Test [81] eingesetzt. Vergleichende Analysen zeigen jedoch die Robustheit der eingesetzten statistischenVerfahren, d. h. Aktivierungskarten aus gleichen Datensätzen, berechnet mit unterschiedlichen Verfahren, unterscheiden sich oft nur unwesentlich [12].
2. Beim bildpunktweisen Vergleich der Aktivierungsbedingung mit dem Kontrollzustand entsteht das Problem vonwiederholten statistischen Vergleichen. So werden beispielsweise bei Verwendung einer 128 x 128-Bildmatrix16384 statistische Vergleiche durchgeführt. Bei einem Signifikanzniveau von p <0,05 wären bei rein zufällig verteilter Signalintensität damit 16384 x 0,05 = 819 Bildelementefalsch-positiv "aktiviert" (statistischer Fehler 1. Art). Um dieAnzahl dieser zufällig "aktivierten" Pixel zu reduzieren,kann man eine Korrektur für die mehrmaligen (16384) statistischen Vergleiche vornehmen, z. B. nach Bonferroni. Diesführt zu einer drastischen Reduktion der Sensitivität desTestverfahrens (in einem Beispiel: p < O,05{16 384 =0,000003) und damit zu einer Zunahme des statistischenFehlers 2. Art. Die Intensität der Signale in den einzelnenBildpunkten ist jedoch nicht statistisch unabhängig voneinander. Man kann vielmehr davon ausgehen, daß die neuronale Aktivität zu einer Signaländerung in einer größerenAnzahl benachbarter Bildpunkte führt. Dementsprechendläßt sich ein Cluster-Kriterium definieren (z.B. mindestenszehn signifikante Pixel in einer 5 x 5-Umgebung, wie in unseren Studien), wodurch der effektive Fehler 1. Art deutlichreduziert werden kann. Forman et al. [26] konnten mitMonte-Carlo-Simulationen und in tMRT-Untersuchungenam Menschen zeigen, daß durch die Verwendung eines Cluster-Kriteriums die statistische Power des Verfahrens umden Faktor 5 gesteigert werden kann.
3. Neben den Subtraktionsverfahren sind Methoden zur Auswertung von tMRT-Daten vorgeschlagen worden, die aufKreuzkorrelationen beruhen [4,49]. Hierbei werden die Aktivitätswerte mit einer vorzugebenden Eingangsfunktionverglichen, die den zeitlichen Verlauf der verschiedenenAktivitätszustände abbilden muß. Meist wird eine einfacheRechteckfunktion (box car) vorgegeben, die zwischen Ruheund Aktivität unterscheidet. Eine Kreuzkorrelation dertMRT-Meßwerte mit dieser Funktion ergibt die gleichen Ergebnisse wie ein direkter statistischer Vergleich von Ruheund Aktivitätswerten. Vorteilhaft ist die Kreuzkorrelation,wenn man eine sehr schnelle Bildfolge aufnimmt und dadurch die hämodynamische Dispersion und Verzögerungenmißt. Unter der Annahme, daß die Verzögerung bei jedemÜbergang von Ruhe zu Aktivität oder umgekehrt gleichlangdauert, kann man die Rechteckfunktion leicht phasenverschieben, um die Verzögerungen zu kompensieren.
4. Das Konzept von neuronalen Zellverbänden (cell assemblies) [37] in räumlich voneinander getrennten anatomischen Strukturen, die für bestimmte Leistungen interagieren müssen und daher gleichzeitig aktiv werden, führte zuAnalyseverfahren, die sich speziell der funktionellen Konnektivität widmeten. Hierbei wird besonderes Augenmerk
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.
Funktionelle Magnetresonanztomographie in der psychopathologischen Forschung Fortschr. Neurol. Psychiat. 66 (1998) 253
-2,5 -2 -1,5 -1 -0,5 ° 0,5 1 1,5 2 2.5links mittlere Aktivierung [%1 rechts
dischen Arbeiten von Tootell ein Verfahren beschrieben, weiches die gefaltete Kortexoberfläche auf eine zweidimensionaleFläche mit geringem Abbildungsfehler projiziert. Das Verfahren läßt sich intuitiv verstehen als mathematisches ,,Aufblasen" eines gefalteten Ballons, bis dieser eine glatte Oberflächeaufweist. Mittels solcher Karten läßt sich die Topographie kortikaler Repräsentationen am besten beurteilen und Z.B. Vergleiche zwischen unterschiedlichen Spezies durchführen.
Durch die hohe räumliche Auflösung der fMRT entsteht beider Kartierung höherer kognitiver Funktionen das folgendeProblem: Einerseits werden allgemeine, d. h. interindividuellreplizierbare topographische Karten kortikaler Repräsentationen gesucht. Andererseits ist jedoch davon auszugehen, daßdiese Karten durch Prozesse der Neuroplastizität, d.h. der Anpassung an individuelle Erfordernisse aus der Lebensgeschichte der Personen, individuell unterschiedliche Strukturen aufweisen und auch im Erwachsenenalter ständigem Wandel unterzogen sind (s. u.). Eine Lösung des Problems läßt sich nichtallgemein angeben. Sie muß vielmehr in Abhängigkeit von derjeweiligen Fragestellung im einzelnen erarbeitet werden.
1&&,
•lfiI
- ml
Ge
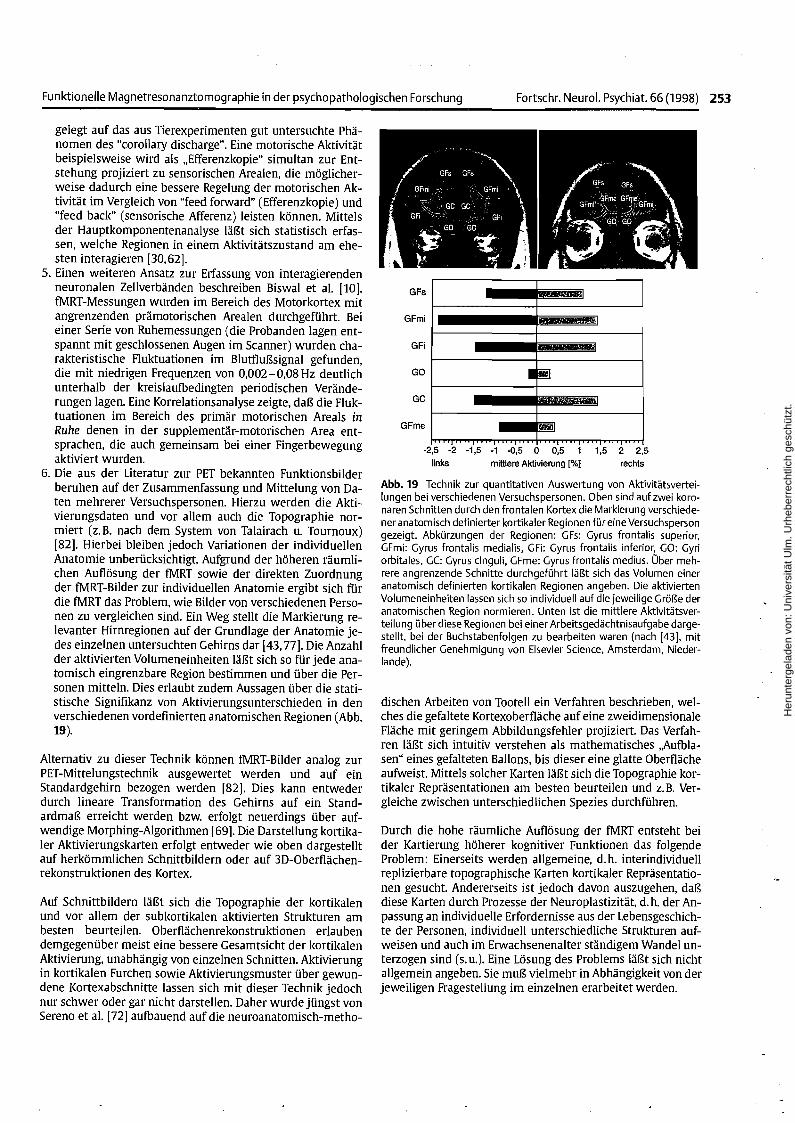
Abb. 19 Technik zur quantitativen Auswertung von Aktivitätsverteilungen bei verschiedenen Versuchspersonen. Oben sind auf zwei koronaren Schnitten durch den frontalen Kortex die Markierung verschiedener anatomisch definierter kortikaler Regionen für eine Versuchspersongezeigt. Abkürzungen der Regionen: GFs: Gyrus frontalis superior,GFmi: Gyrus frontalis medialis, GFi: Gyrus frontalis inferior, GO: Gyriorbitales, GC: Gyrus cinguli, GFme: Gyrus frontalis medius. Über mehrere angrenzende Schnitte durchgeführt läßt sich das Volumen eineranatomisch definierten kortikalen Regionen angeben. Die aktiviertenVolumeneinheiten lassen sich so individuell auf die jeweilige Größe deranatomischen Region normieren. Unten ist die mittlere Aktivitätsverteilung über diese Regionen bei einer Arbeitsgedächtnisaufgabe dargestellt, bei der Buchstabenfolgen zu bearbeiten waren (nach [43], mitfreundlicher Genehmigung von Elsevier Science, Amsterdam, Niederlande).
GFme
GFi
GFs
GFmi
GO
gelegt auf das aus Tierexperimenten gut untersuchte Phänomen des "corollary discharge". Eine motorische Aktivitätbeispielsweise wird als "Efferenzkopie" simultan zur Entstehung projiziert zu sensorischen Arealen, die möglicherweise dadurch eine bessere Regelung der motorischen Aktivität im Vergleich von "feed forward" (Efferenzkopie) und"feed back" (sensorische Afferenz) leisten können. Mittelsder Hauptkomponentenanalyse läßt sich statistisch erfassen, welche Regionen in einem Aktivitätszustand am ehesten interagieren [30,62].
5. Einen weiteren Ansatz zur Erfassung von interagierendenneuronalen Zellverbänden beschreiben Biswal et al. [10].fMRT-Messungen wurden im Bereich des Motorkortex mitangrenzenden prärnotorischen Arealen durchgeführt. Beieiner Serie von Ruhemessungen (die Probanden lagen entspannt mit geschlossenen Augen im Scanner) wurden charakteristische Fluktuationen im Blutflußsignal gefunden,die mit niedrigen Frequenzen von 0,002 - 0,08 Hz deutlichunterhalb der kreislaufbedingten periodischen Veränderungen lagen. Eine Korrelationsanalyse zeigte, daß die Fluktuationen im Bereich des primär motorischen Areals inRuhe denen in der supplementär-motorischen Area entsprachen, die auch gemeinsam bei einer Fingerbewegungaktiviert wurden.
6. Die aus der Literatur zur PET bekannten Funktionsbilderberuhen auf der Zusammenfassung und Mittelung von Daten mehrerer Versuchspersonen. Hierzu werden die Aktivierungsdaten und vor allem auch die Topographie normiert (z. B. nach dem System von Talairach u. Tournoux)[82]. Hierbei bleiben jedoch Variationen der individuellenAnatomie unberücksichtigt. Aufgrund der höheren räumlichen Auflösung der fMRT sowie der direkten Zuordnungder fMRT-Bilder zur individuellen Anatomie ergibt sich fürdie fMRT das Problem, wie Bilder von verschiedenen Personen zu vergleichen sind. Ein Weg stellt die Markierung relevanter Hirnregionen auf der Grundlage der Anatomie jedes einzelnen untersuchten Gehirns dar [43,77]. Die Anzahlder aktivierten Volumeneinheiten läßt sich so für jede anatomisch eingrenzbare Region bestimmen und über die Personen mitteln. Dies erlaubt zudem Aussagen über die statistische Signifikanz von Aktivierungsunterschieden in denverschiedenen vordefinierten anatomischen Regionen (Abb.19).
Alternativ zu dieser Technik können fMRT-Bilder analog zurPET-Mittelungstechnik ausgewertet werden und auf einStandardgehirn bezogen werden [82]. Dies kann entwederdurch lineare Transformation des Gehirns auf ein Standardmaß erreicht werden bzw. erfolgt neuerdings über aufwendige Morphing-Algorithmen [69]. Die Darstellung kortikaler Aktivierungskarten erfolgt entweder wie oben dargestelltauf herkömmlichen Schnittbildern oder auf 3D-Oberflächenrekonstruktionen des Kortex.
Auf Schnittbildern läßt sich die Topographie der kortikalenund vor allem der subkortikalen aktivierten Strukturen ambesten beurteilen. Oberflächenrekonstruktionen erlaubendemgegenüber meist eine bessere Gesamtsicht der kortikalenAktivierung, unabhängig von einzelnen Schnitten. Aktivierungin kortikalen Furchen sowie Aktivierungsmuster über gewundene Kortexabschnitte lassen sich mit dieser Technik jedochnur schwer oder gar nicht darstellen. Daher wurde jüngst vonSereno et al. [72] aufbauend auf die neuroanatomisch-metho-
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.

254 Fortschr. Neurol. Psychiat. 66 (1998)
c) Brain maps
Unter dem Stichwort der Gehirnkarten (brain maps) verbirgtsich eine Reihe z. T. sehr alter Fragen der Hirnforschung. Kortikale Areale lassen sich nach unterschiedlichen Kriterien differenzieren, was bereits um die Jahrhundertwende zur Publikation unterschiedlicher Einteilungssysteme des Kortex geführt hat. Die bekannteste Kartierung erfolgte durch Brodmann [14], der histologische Kriterien verwendete. Flechsig[25] unterschied kortikale Areale nach dem Zeitpunkt ihrerMyelinisierung. In einer aktuellen Erweiterung dieser Ansätzeverwenden Zilles et al. [96] zusätzlich biochemische Markerfür verschiedene neuronale Rezeptoren.
Die funktionelle Kartierung der Großhirnrinde erfolgte in derVergangenheit, d. h. vor der Möglichkeit nichtinvasiver Untersuchungen beim Menschen, vor allem tierexperimentell. Daneben wurden funktionelle Spezialisierungen von kortikalenArealen beim Menschen zunächst mittels Läsionsstudien (Broca, Wernicke) und später auch durch invasive Reizexperimente im Rahmen von neurochirurgischen Eingriffen ermittelt.Die Grenzen dieser Methoden können durch die jetzt zur Verfügung stehenden nichtinvasiven Verfahren überwunden werden und haben damit ein neues Forschungsfeld eröffnet.
Die funktionelle Kartierung der menschlichen Großhirnrindeist eine Aufgabe, die noch Jahrzehnte in Anspruch nehmenwird und durchaus mit der Kartierung des menschlichen Genoms vergleichbar ist [85]. Die Größenordnung dieser Aufgabe läßt sich unter Heranziehen einiger plausibler Zusatzannahmen abschätzen: Man kann zunächst davon ausgehen, daßdie Großhirnrinde schwach modular organisiert ist [50], d. h.daß eine bestimmte geistige Leistung durch das Zusammenwirken verschiedener Areale - möglicherweise um etwa einDutzend - bewirkt wird. In funktioneller Hinsicht lassen sichkortikale Areale dadurch unterscheiden, daß sie auf bestimmte Stimuli selektiv ansprechen bzw. bei bestimmten klar definierten Leistungen aktiv werden. Am besten untersucht ist indieser Hinsicht das visuelle System. Man kennt derzeit mindestens 32 unterschiedliche kortikale Areale, die für die Funktion des Sehens zuständig sind [24]. Diese Karten sind elektrophysiologisch durch Tierversuche an Primaten definiert undkonnten beim Menschen teilweise durch funktionelle Bildgebung mit recht guter Übereinstimmung ebenfalls nachgewiesen werden [72,95].
Die durchschnittliche Größe der visuellen Karten liegt bei Primaten bei 170 mm2• Beim Menschen sind diese Areale wahrscheinlich größer. So weist das Areal V1 beim Menschen diedoppelte Größe des Areals V1 beim Affen auf. Geht man voneiner Größe des menschlichen Kortex von 250000 mm2 aus,so läßt sich die Anzahl der hierin befindlichen und aufzudekkenden Karten unterschiedlicher Funktion mit 735 beziffern.Diese Zahl ist gEwiß nur das Ergebnis einer groben Abschätzung; sie liefert jedoch einen Anhaltspunkt für die Größenordnung des Problems.
Da wir weder wissen, wie redundant diese Areale sind, noch,wie höhere kognitive Leistungen durch ein Zusammenwirkendieser Karten zustande kommen, sind keine raschen Fortschritte bei der Aufklärung dieser Karten mittels funktionellerBildgebung zu erwarten. Um diese Karten selektiv zu aktivieren, sind gezielte kritische Stimuli notwendig, und diese wie-
Spitzer, M. et al.
derum sind bei höherstufigen Karten nicht trivial (es geht hiernicht mehr nur um Farbe oder Form oder Bewegung). Benötigtwerden daher neurophysiologische und neuropsychologischeModelle der zu untersuchenden Leistungen, da nur sie Anhaltspunkte und entsprechende Arbeitshypothesen liefernkönnen.
Ausgehend von neuropsychologischen Daten sowie von Netzwerkmodellen semantischer Speicherung in selbstorganisierenden Eigenschaftskarten wurde beispielsweise die Hypothese kategorienspezifischer Aktivierung in links-frontalenund temporalen Arealen aufgestellt und direkt überprüft (Abb.16u.17).
d) Brain-or-vein
Wie in der Einleitung dargestellt, beruhen die Signale derfMRT auf Blutflußänderungen im Kapillarbereich (BOLD-Kontrast). Da der Kontrast auf Unterschieden der magnetischenEigenschaften des Blutes beruht, das aus dem Kapillarbett venös drainiert wird, besteht die Gefahr der Abbildung größerervenöser Gefäße mit dieser Technik. Diese Gefäße sind u. U.einige Millimeter von dem Ort der kortikalen Aktivierung entfernt. Bei der Interpretation von mittels BaLD-Kontrast gewonnenen Aktivierungsbildern ist dies zu berücksichtigen.Venöse Strukturen lassen sich entweder aufgrund ihrer Formund Lage oder durch Überlagerung mit MR-angiographisch ermittelten Bildern identifizieren und von weitergehender topographischer Interpretation ausschließen [19,29,52].
Ganz allgemein ist die Beziehung von neuronaler Aktivitätund BaLD-Kontrast nicht vollständig aufgeklärt. Tierexperimentelle Untersuchungen von Malonek u. Grinvald [53] miteiner optischen Meßtechnik am eröffneten Schädel zeigen,daß eine visuelle Stimulation für wenige hundert Millisekunden eine Hypoxie hervorruft, die im visuellen Kortex sehr genau den Bereich der aktivierten kortikalen Säulen abbildet(ein Areal von 0,5 mm Durchmesser). Erst im Anschluß darankommt es zur überschießenden Reaktion des kapillaren Gefäßbettes mit Zunahme der Sauerstoffkonzentration in einemBereich von 3 - 5 Millimetern um die aktivierten kortikalenKolumnen. Dieser "overshoot" läßt sich 500 ms nach Beginndes sensorischen Stimulus erstmals nachweisen und hat seinMaximum nach etwa 5 Sekunden. Der initiale "undershoot"der Blutoxygenierung ist auch nachweisbar mit Hilfe der funktionellen MR-Spektroskopie [38,56]. Von der höheren räumlichen und zeitlichen Auslösung des "undershoot" wird jedochderzeit aufgrund technischer Einschränl<Ungen noch kein Gebrauch bei funktionellen Untersuchungen gemacht.
e) Kontrolle der Bewegungsartefakte
Aufgrund der hohen räumlichen Auflösung ist die fMRT besonders sensibel gegenüber Bewegungsartefakten. Die entsprechende Methodenkritik ging zuweilen soweit, daß die gesamte mittels fMRT gemessene kortikale Aktivierung als stimulationskorrelierter Bewegungsartefakt interpretiert wurde[36]. Diese Gefahr besteht insbesondere dann, wenn die Aktivierungsbedingung eine motorische Reaktion des Probandenbeinhaltet und sich die Bewegung, beispielsweise der Hand,auf den Kopf überträgt. Werden Differenzbilder gegen eineeinfache Ruhebedingung berechnet, so geht "task-correlatedmotion" in die Differenzbildung mit ein. Daher ist, wie oben
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.

Funktionelle Magnetresonanztomographie in der psychopathologischen Forschung Fortschr. Neurol. Psychiat. 66 (1998) 255
ausgeführt, bereits bei der Planung des Stimulationsparadigmas auf geeignete aktive Kontrollbedingungen zu achten. Zudem muß der Proband so bequem wie möglich gelagert werden. Bewegungsartefakte des Kopfes lassen sich dann durchgeeignete Hardware (Vakuumkissen, Beißbrettchen)und Software (Algorithmen zur Bewegungskorrektur) [92,94] reduzieren. Die hierzu eingesetzten Verfahren unterscheiden sichdurch ihre Komplexität und den damit verbundenen Rechenaufwand. Angestrebt werden dreidimensionale Korrekturenim sub-voxel-Bereich [36], die sowohl Translations- als auchRotationsbewegungen in allen Ebenen und um alle Achsenausgleichen. Hierzu können neben den MR-Daten auch zusätzliche Bewegungsmessungen, z. B. über Infrarotsignale,herangezogen werden. Darüber hinaus müssen aktivitätsunabhängige, mit kardiorespiratorischen Funktionen in Zusammenhang stehende Liquorpulsationen im gesamten Gehirnberücksichtigt werden [9].
Die Eliminination von Bewegungsartefakten ist nach unsererAnsicht eine entscheidende Voraussetzung für die Anwendungder fMRT in der psychopathologischen Forschung. Man mußdavon ausgehen, daß die gemessenen MR-Daten psychiatrischer Patienten in der Regel größere Bewegungsartefakte aufweisen als die Daten gesunder Probanden.
Bislang war nur von der Bewegung zwischen den Bildern einzelner Meßserien die Rede. Aufgrund der raschen Bildaufnahmezeiten der neuen MR-Tomographen spielt die während einer einzigen Aufnahme stattfindende Bewegung bei der Untersuchung gesunder Probanden keine wesentliche Rollemehr. Die heute überwiegend eingesetzte EPl-Technik ermöglicht die Akquisition eines Bildes innerhalb von 50 Millisekunden. Wie jeder Amateurfotograph weiß, sind bei ruhendemObjekt mit einer Belichtungszeit von 1/20 Sekunde gute Resultate zu erzielen, sich rasch bewegende Objekte können jedoch durchaus Probleme bereiten.
f) Zufallsbefunde
Alle Studien mit fMRT schließen eine anatomische Bildgebungzur Übersicht und Lokalisation der funktionell zu messendenRegionen mit ein. Dabei ist es jederzeit möglich, daß sowohlbei gesunden Probanden als auch bei psychiatrischen Patienten auffällige strukturelle Befunde erhoben werden. In unserem bisherigen Untersuchungskollektiv von 152 gesundenProbanden fanden sich insgesamt vier auffällige Befunde, vondenen einer klinische Relevanz hatte: Ein Gliom im Bereichdes Mesenzephalon hatte zu einem gerade noch kompensierten Hydrocephalus occ1usus geführt. Der Proband gab nachBefragen etwas gehäufte Kopfschmerzen in den vorangegangenen Wochen an. Sie waren aber für ihn nicht so eindrücklich, daß er einen Arzt konsultiert hätte. Eine umfangreichediagnostische Abklärung wurde eingeleitet. Die anderen auffälligen Befunde waren von harmloser Natur. Ein Proband hatte eine sehr große Arachnoidalzyste, die fast die ganze mittlere Schädelgrube rechts ausfüllte und den Temporalpol verdrängt hatte. Bei einem anderen Probanden fand sich eineunklare Struktur in der Kalotte hochparietal rechts, welchescheinbar Kontakt zur Dura hatte. Die Abklärung mit konventionellen Röntgenaufnahmen des Schädels ergab eine Osteose.Schließlich wies einer der Probanden als Normvariante einCavum septum pellucidum auf. In der Literatur finden sichunseres Wissens bisher keine systematischen Untersuchun-
gen solcher Zufallsbefunde an größeren Probandenkollektiven. Allerdings berichten auch andere Arbeitsgruppen von pathologischen Zufallsbefunden, manchmal mit der Konsequenzbesonders früher Diagnostik und dadurch bedingter besonders guter Therapiechancen. Insgesamt ist bei Studien angrößeren Probandenkollektiven grundsätzlich mit dem Auftreten von pathologischen Befunden zu rechnen. Daher solltendiese Untersuchungen stets von erfahrenen Neuroradiologenbegleitet werden.
7. Schlußfolgerungen und Ausblick
Die fMRT stellt eine wesentliche methodische Neuentwicklung zur nichtinvasiven Untersuchung höherer geistiger Leistungen dar. Entscheidend für die Anwendung auf Fragestellungen im Bereich der Pathologie solcher höherer geistigerLeistungen - der Psychopathologie - ist die Tatsache, daß aufder Ebene des einzelnen Individuums signifikante Ergebnisseerzielt werden können. Die Methode sollte demnach einenwesentlichen Beitrag in der psychiatrischen Forschung leistenkönnen. Ihre sich bereits jetzt abzeichnenden Möglichkeitensind erstaunlich: Untersuchungen zum Arbeitsgedächtnis mittels des oben beschriebenen Paradigmas (Abb. 12) aus der Arbeitsgruppe um Cohen [11,18] zeigen, daß die Aktivität despräfrontalen Kortex mit zunehmender Beanspruchung des Arbeitsgedächtnisses linear zunimmt. In unserer Studie zumWortvergleich in Hinblick auf semantische Verwandtschaft[77] (Abb. 14 u.15) nahm, wie oben ausgeführt, die kortikaleAktivierung mit dem Einfacherwerden der Aufgabe aufgrundselektiver Lerneffekte ab. just u. Carpenter [42] konnten unterschiedlich starke kortikale Aktivierungsmuster bei der Verarbeitung von Relativsätzen im Vergleich zu beigeordnetenHauptsätzen nachweisen, und O'Craven et al. [58] bestimmtenden Einfluß der selektiven Aufmerksamkeit auf die Verarbeitung visueller Bewegungsreize in bewegungssensitiven Arealen des extrastriatalen Kortex.
Eines der für die Psychiatrie bedeutsamsten Ergebnisse derHirnforschung der letzten jahre ist die Erkenntnis, daß auchnoch im ausgereiften, adulten Neokortex die Fähigkeit zur plastischen Reorganisation von landkartenförmig angeordnetenRepräsentationsorten besteht (Neuroplastizität) [44]. Es solltesich daher die Möglichkeit ergeben, erfahrungsabhängige Veränderungen der oben beschriebenen brain maps zu messen.Hiermit wäre man dann nicht mehr weit von einem neuronalen Korrelat dessen entfernt, was der Heidelberger Psychopathologe janzarik unter "Struktur" verstanden hat [40]. Einenersten Schritt in diese Richtung gehen Kim et al. [45] mit ihrerUntersuchung gesunder zweisprachiger Probanden, die entweder von früher Kindheit an zweisprachig aufgewachsen waren oder die zweite Fremdsprache erst später erlernten. Diefrontale Repräsentation beider Sprachen in einem Areal, dasungefähr dem Brocaschen Sprachzentrum entspricht, war beider erstgenannten Gruppe stark überlappend, bei der zweitenGruppe dagegen klar getrennt.
Auch die Auswirkungen von Neuromodulatoren wie beispielsweise Dopamin oder Serotonin auf definierte geistige Leistungen und damit einhergehende zerebrale Aktivierungsmustersind der fMRT zugänglich [80], wenn auch bislang hierzu vorallem PET-Studien vorliegen [22]. Hierdurch sollten Zusammenhänge zwischen hypo- oder hyperaktiven kortikalen Be-
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.

256 Fortsehr. Neurol. Psychiat. 66 (1998)
reichen, den psychiatrisch relevanten Neuromodulatoren undpsychopathologischen Phänomenen nachweisbar sein.
In methodischer Hinsicht ist zu bemerken, daß derzeit einezunehmende Standardisierung des experimentellen Setups [I,27,84] und der Auswertestrategien erfolgt, der Prozeß jedochkeineswegs abgeschlossen ist. Auch die Integration der fMRTmit anderen Methoden wie ereigniskorrelierten Potentialen(EKP), Magnetenzephalographie (MEG) oder der transkraniellen Magnetstimulation (TMS) ist derzeit Gegenstand intensiver Forschung [5].
Eine für psychopathologische Fragestellungen interessantemethodische Neuentwicklung stellt die "single trial"-Technikdar [15]. Wie oben dargestellt, wurden bei fMRT-Studien bisher sog. geblockte Versuchsdesigns eingesetzt. Die zu vergleichenden Bedingungen wurden in Blöcken von 30-360 s Dauerzusammengefaßt dargeboten. So betrug beispielsweise dieLänge eines Blockes in unserem Experiment zum Wortvergleich in Hinblick auf semantische Verwandtschaft (Abb. 14 u.15) 6 Minuten. Innerhalb dieser Zeit absolvierte die Versuchsperson eine größere Anzahl von Aufgaben der gleichen Kategorie. Ein solches Design ist hinsichtlich der psychologischenFragestellungen prinzipiell limitiert. So läßt sich beispielsweise die zerebrale Aktivität bei unterschiedlichen Entscheidungen nicht trennen, und es ist keine Auswertung der Aktivierung auf der Basis richtiger Entscheidungen im Vergleich zuFehlern durchführbar. Aus der psychologischen literatur istdarüber hinaus bekannt, daß die Effektstärken bei Blockdesigns geringer sind als bei randomisierten Designs. Mit derhohen zeitlichen Auflösung der fMRT ist es in jüngster Zeitmöglich geworden, Stimuli unterschiedlicher Kategorien randomisiert darzubieten und das MR-Signal nachträglich nachKategorien sortiert auszuwerten. In derartigen Auswertungenkann auch die Art der Reaktion auf eine einzelne Aufgabe berücksichtigt werden. Analog zur elektrophysiologischen Methode der ereigniskorrelierten Potentiale erfolgte auch in der"single trial"-Technik eine Mittelung des untersuchten Signalsüber mehrere Aufgaben gleicher Kategorie (sog. averaging).Das Abbild eines einzelnen mentalen Ergebnisses bleibt somitauch im Rahmen der fMRT ein Desiderat für die Zukunft [34].
In systematischer Hinsicht läßt sich die direkte Abbildungpsychopathologischer Symptome wie beispielsweise Zwängeoder Halluzinationen von der Abbildung kognitiver Grundfunktionen wie des Arbeitsgedächtnisses unterscheiden. Dieskann zu diagnostischen Zwecken im Sinne der Bildung vonUntergruppen sowie zur Beurteilung des klinischen Verlaufsbzw. des Therapieerfolgs eingesetzt werden. Das Erkenntnisinteresse geht im ersten Fall aus vom klinischen Bild und nähert sich der Hirnfunktion, wohingegen im zweiten Fall diedefinierte Hirnfunktion den Beginn des Argumentationsgangsmarkiert. Beide Forschungsrichtungen sollten langfristig zu einer funktionellen Neuropsychopathologie führen und damitzu einer genaueren Beschreibung und zu einem besseren Verständnis der Grundlagen psychiatrischen Handeins.
Danksagung
Diese Arbeit wurde durch die DFG (Projekt Nr. Sp. 364/1-3),durch einen Forschungspreis der DGPN-Duphar-Stiftung sowie durch einen NARSAD-Grant (Established Investigator's
Spitzer, M. et al.
Award der National Alliance for Research in Schizophrenia andDepression) gefördert.
literatur
1 Aine, c.j.: Aconceptual overview and critique offunctional neuroimaging techniques in humans: I. MRljFMRI and PET. CriticalRev. Neurobiol. 9 (1995) 229-309
2 Andreasen, N. c.: Schizophrenia. From mind to moleeule. American Psychiatrie Press, Washington, DC 1994
3 Baddeley, A: Working memory. Oxford University Press, Oxford1986
4 Bandettini, P. A, A jesmanowicz, E. C. Wong, j. S. Hyde: Processing strategies for time-course data sets in functional MRI ofthehuman brain. Magn. Reson. Med. 30 (1993) 161-173
5 Barinaga, M.: New imaging methods provide a better view intothe brain. Science 276 (1997) 1974-1976
6 Bellemann, M. E., M. Spitzer, G. Brix, T. Kammer, R. Loose, ASchwartz, F. Gückel: Neurofunktionelle MR-Bildgebung höhererkognitiver Leistungen des menschlichen Gehirns. Radiologe 35(1995) 272-282
7 Belliveau, j. w., D. N. Kennedy, C. McKinstry, B. R. Buchbinder, R.M. Weisskoff, M. S. Cohen, j. M. Vevea, T. j. Brady, B. R. Rosen:Functional mapping of the human visual cortex by magnetic resonance imaging. Science 254 (1991) 716-719
8 Berger, H.: Über das Elektroenzephalogramm des Menschen.Arch. Psychiat. Nervenkr. 87 (1929) 527 - 570
9 Biswal, B., E. A DeYoe, H. j. S.: Reduction of physiological fluctuations in fMRI using digital filters. Magn. Reson. Med. 35(1996) 107-113
10 BiswaI, B., F. Z. Yetkin, V. M. Haughton,j. S. Hyde: Functional connectivity in the motor cortex of resting human brain using echoplanar MRI. Magn. Reson. Med. 34 (1995) 537-541
11 Braver, T. S., j. D. Cohen, L. E. Nystrom, j. jonides, E. E. Smith, D.C. Noll: A parametric study of prefrontal cortex involvement inhuman working memory. Neuroimage 5 (1997) 49-62
12 Breiter, H. c., S. L. Rauch, I<. I<. Kwong, j. R. Baker, R. M. Weisskoff,D. N. Kennedy, A D.Kendrick, T. L. Davis, Ajiang, M.5. Cohen, C.E. Stern, j. W. Belliveau, L. Baer, R. L. O'Sullivan, C. R. Savage, M.A jenike, B. R. Rosen: Functional magnetic resonance imagingof symptom provocation in obsessive-compulsive disorder.Arch. Gen. Psychiatry 53 (1996) 595-606
13 Brix, G., F. GückeI, M. E. Bellemann, j. Röther, ASchwartz, H. j.Ostertag, W. j. Lorenz: Functional MR mapping of activated cortical areas. Nuklearmedizin 33 (1994) 200-205
14 Brodmann, 1<.: Vergleichende Localisationslehre der Großhirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues.Barth, Leipzig 1909
15 Buckner, R. L., P. A Bandettini, I<. M. O'Craven, R. L. Savoy, S. E.Petersen, M. E. Raichle, B. R. Rosen: Detection of cortical activation during averaged single trials of a cognitive task usingfunctional magnetic resonance imaging. Proc. Natl. Acad. Sei. 93,USA (1996) 14878-14883
16 Churchland, P. S., T. j. Sejnowski: Perspectives in cognitive neuroscience. Science 242 (1988) 741-745
17 Cohen, j. D., S. D. Forman, T. S. Braver, B. j. Casey, D. ServanSchreiber, D. C. Noll: Activation ofthe prefrontal cortex in a nonspatial working memory task with functional MRI. Hum. BrainMapp.l (1994) 293-304
18 Cohen, j. D., W. M. Perlstein, T. S. Braver, L. E. Nystrom, D. C. NoH,j.jonides, E. E. Smith: Temporal dynamics ofbrain activation during a working memory task. Nature 386 (1997) 604-608
19 Cohen, M. S., S. M. Kosslyn, H. C. Breiter, G. j. DiGirolamo, W. L.Thompson, A I<. Anderson, S. Y. Bookheimer, B. R. Rosen, j. W.Belliveau: Changes in corticaI activity during mental rotation. Amapping study using functional MRI. Brain 119 (1996) 89-100
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.

Funktionelle Magnetresonanztomographie in der psychopathologischen Forschung Fortschr. Neurol. Psychiat. 66 (1998) 257
20 Damasio, H., T. j. Grabowski, D. Tranei, R. D. Hichwa, A. R. Damasie: A neural basis for lexical retrieval. Nature 380 (1996) 499505
21 David, A S., P. W. R. Woodruff, R. Howard, J. D. C. Meilers, M.Brammer, E. Bullmore, I. Wright, C. Andrew, S. C. R. Williams:AuditOlY hallucinations inhibit exogenous activation of auditOlYassociation cortex. Neuroreport 7 (1996) 932-936
22 Dolan, R. j., P. Fleteher, C. D. Frith, K. j. Friston, R. S. j. Frackowiak,P. M. Grasby: Dopamineric modulation of impaired cognitive activation in the anterior cingulate cortex in schizophrenia. Nature378(1995)180-182
23 Ernst, T., j. Hennig: Observation of a fast response in functionalMR. Magn. Reson. Med. 32 (1994) 146-149
24 FeIleman, D. j., D. C. van Essen: Distributed hierarchical processing in the primate cerebral cortex. Cereb. Cortex 1 (1991) 1-47
25 Flechsig, P.: Anatomie des menschlichen Gehirns und Rückenmarks of myelogenetischer Grundlage. Thieme, Leipzig 1929
26 Forman, S. D.,j. D. Cohen, M. Fitzgerald, W. F. Eddy, M. AMintun,D. C. NoU: Improved assessment of significant activation infunctional magnetic resonance imaging (fMR1): use of a c1ustersize threshold. Magn. Reson. Med. 33 (1995) 636-647
27 Frackowiak, R. S. j., K. j. Friston, C. D. Frith, R. j. Dolan, j. C. Mazziotta: Human Brain Function. Academic Press, London 1997
28 Frahm, J" K. D. Merboldt, W. Hänicke: Functional MRI of humanbrain activation at high spatial resolution. Magn. Reson. Med. 29(1993) 139-144
29 Frahm, J" K. D. Merboldt, W. Hänicke, A Kleinschmidt, H. Boekker: Brain or vein - oxygenation or flow? On signal physiologyin functional MRI of human brain activation. NMR Biomed. 7(1994) 45-53
30 Friston, K. j.: Functional and effective connectivity in neuroimaging. A synthesis. Hum. Brain. Mapp. 2 (1994) 56-78
31 Gegenfurtner, K. R.: Visual neurobiology - colouring the cortex.Nature 388 (1997) 23-24
32 Goldman-Rakic, P. S.: Prefrontal cortical dysfunction in schizophrenia: The relevance of working memory. In: B. J. Carroll, j.E. Barrett (Eds.): Psychopathology and the brain. Raven Press,New York (1991) pp. 1 - 23
33 Goldman-Rakic, P. S.: Working memory dysfunction in schizophrenia.j. Neuropsychiatry Clin. Neurosci. 6 (1994) 348-357
34 Grabowski, T. J., A R. Damasio: Improving functional imagingtechniques: the dream of a single image for a single mentalevent. Proc. Natl. Acad. Sei. 93, USA (1996) 14302 -14303
35 Grodd, w., F. Schneider, U. Klose, T. Nägele: Funktionelle Kernspintomographie psychischer Funktionen am Beispiel experimentell induzierter Emotionen. Radiologe 35 (1995) 283-289
36 Hajnal, j. V., R. Mayer, A Oatridge, j. E. Schwieso, j. R. Young, G.M. Bydder: Artifacts due to stimulus correlated motion infunctional imaging of the brain. Magn. Reson. Med. 31 (1994)289-291
37 Hebb, D. 0.: The organization of behaviour. Wiley, New York1949
38 Hennig, j., T. Ernst, O. Speck, G. Deuschi, E. Feifel: Detection ofbrain activation using oxygenation sensitive functional spectroscopy. Magn. Reson. Med. 31 (1994) 85-90
39 Hubel, D. H.: Auge und Gehirn. Spektrum Akademischer Verlag,Heidelberg 1989
40 Janzarik, W.: Strukturdynamische Grundlagen der Psychiatrie.Enke, Stuttgart 1988
41 Jonides,j., E. E. Smith, R. A Koeppe, E. Awh, S. Minoshima, M. AMintun: Spatial working memory in humans as revealed by PET.Nature 363 (1993) 623-625
42 Just, M. A., P. A Carpenter, T. A Keller, W. F. Eddy, K. R. Thulborn:Brain activation modulated by sentence comprehension. Science274 (1996) 114-116
43 Kammer, T., M. E. Bellemann, F. Gückel, G. Brix, A Gass, H.Schlemmer, M. Spitzer: Functional MR imaging of the prefrontalcortex: Specific activation in a working memory task. Magn. Reson.lmaging 15 (1997) 879-889
44 Karni, A., G. Meyer, P. Jezzard, M. M. Adams, R. Turner, L. G. Ungerleider: Functional MRI evidence for adult motor cortex plasticity during mother skilliearning. Nature 377 (1995) 155-158
45 Kim, K. H. S., N. R. Relkin, K. M. Lee, j. Hirsch: Distinct corticalareas associated with native and second languages. Nature 388(1997) 171-174
46 Kim, S.-G., j. Ashe, K. Hendrich, j. M. Ellermann, H. Merkle, K.Ugurbil, A P. Georgopoulos: Functional magnetic resonanceimaging of motor cortex: Hemispheric asymmetry and handedness. Science 261 (1993) 615-617
47 Kim, S. G., W. Richter, K. Ugurbil: Limitations of temporal resolution in functional MRI. Magn. Reson. Med. 37 (1997) 631-636
48 Kischka, u., C. W. Walleseh, G. Wolf (Eds.): Methoden der Hirnforschung. Spektrum Akademischer Verlag, Heidelberg 1997
49 Kleinschmidt, A, W. Hänicke, M. Requardt, K. D. Merboldt, j.Frahm: Strategien der Datenanalyse in Hirnaktivierungsstudienmit funktioneller MR-Tomographie. Radiologe 35 (1995) 242251
50 Kosslyn, S. M., O. Koenig: Wet mind. The new cognitive neuroscience. Free Press, New York 1992
51 Kwong, K. K., j. W. Belliveau, D. A Chesler, I. E. Goldberg, R. M.Weisskoff, B. P. Poncelet, D. N. Kennedy, B. E. Hoppel, M. S. Cohen, R. Turner, H. M. Cheng, T. j. Brady, B. R. Rosen: Dynamicmagnetic resonance imaging of human brain activity during primary sensory stimulation. Proc. Natl. Acad. Sei. 89, USA (1992)5675-5679
52 Lee, A T., G. H. Glover, C. H. Meyer: Discrimination of large venous vessels in time-course spiral blood-oxygen-Ievel-dependent magnetic-resonance functional neuroimaging. Magn. Reson.Med.33 (1995)745-754
53 Malonek, D., A Grinvald: Interactions between electrical activityand cortical microcirculation revealed by imaging spectroscopy- implications for functional brain mapping. Science 272 (1996)551-554
54 Martin, A, C. L. Wiggs, L. G. Ungerleider, j. V. Haxby: Neural correlates of category-specific knowledge. Nature 379 (1996) 649652
55 Meilers, j. D. c., E. Bullmore, M. Brammer, S. C. R. Williams, C.Andrew, N. Sachs, C. Andrews, T. S. Cox, A Simmons, P. Woodruff,A S. David, R. Howard: Neural correlates of working memory ina visual letter monitoring task - an fMRI study. Neuroreport 7(1995) 109-112
56 Menon, R. S., S. Ogawa, X. Hu,j. P. Strupp, P. Anderson, K. Ugurbil:BOLD based functional MRI at 4 Tesla includes a capillary bedcontribution: echo-planar imaging correlates with previous optical imaging using intrinsic signals. Magn. Reson. Med. 33(1995) 453-459
57 Mitra, P. P., S. Ogawa, X. Hu, K. Ugurbil: The nature of spatiotemporal changes in cerebral hemodynamics as manifested infunctional magnetic resonance imaging. Magn. Reson. Med. 37(1997) 511-518
58 O'Craven, K. M., B. R. Rosen, K. K. Kwong, A Treisman, R. L. Savoy:Voluntary attention modulates fMRI activity in human MT-MST.Neuron 18 (1997) 591-598
59 Ogawa, S., T. M. Lee, A. R. Kay, D. W. Tank: Brain magnetic resonance imaging with contrast dependent on blood oxygenation.Proc. Natl. Acad. Sci. 87, USA (1990) 9868-9872
60 Ogawa, S., D. W. Tank, R. Menon, j. M. Ellermann, S. G. Kim, H.Merkle, K. Ugurbil: lntrinsic signal changes accompanying sensory stimulation: Functional brain mapping with magnetic resonance imaging. Proc. Natl. Acid. Sei. 89, USA (1992) 59515955
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.

258 Fortsehr. Neurol. Psychiat. 66 (1998)==~=~=,=.=============
Spitzer, M. et al.===~=~==="==-=
61 Park, S., P. S. Holzman: Schizophrenie show spatial worldng memory deficits. Arch. Gen. Psychiatry 49 (1992) 975-982
62 Paus, T., S. Marrett, K. Worsley, A Evans: Imaging motor-to-sensory discharges in the human brain - an experimental tool forthe assessment of functional connectivity. Neuroimage 4 (1996)78-86
63 Penfield, W., T. Rasmussen: The cerebral cortex of man: a c1inicalstudy of localization and function. Macmillan, New York 1950
64 Posner, M. 1., S. E. Petersen, P. T. Fox, M. E. Raichle: Localizationof cognitive operations in the human brain. Science 240 (1988)1627 -1631
65 Raichle, M. E.: Circulatory and metabolie correlates of brainfunction in normal humans. In: Plum, F., V. B. Mountcascle(Eds.): Handbook of Physiology, Section 1: The nervaus system,Volume 5: Higher functions ofthe brain. Oxford University Press,New York (1987) pp. 643-674
66 Raa, S. M., J. R. Binder, P. A Bandettini, T. A Hammeke, F. Z. Yetkin, A jesmanowicz, L. M. Usk, G. L. Morris, W. M. Mueller, L. D.Estkowski, E. C. Wong, V. M. Haughton,]. S. Hyde: Functional magnetic resonance imaging of complex human movements. Neurology 43 (1993) 2311-2318
67 Reppas,]. B., S. Niyogi, A. M. Dale, M. I. Sereno, R. B. H. Toatell:Representation of motion boundaries in retinotopic human vi~
sual cortical areas. Nature 388 (1997) 175-17968 Roland, P.: Brain activation. Wiley-Liss, New York 199469 Roland, P. E., K. Zilles: Brain atlases - a new research tooI. Trends
Neurosei. 17 (1994) 458-46770 Schneider, F., W. Grodd, H. ]. Machulla: Untersuchung psychi
scher Funktionen durch funktionelle Bildgebung mit Positronenemissionstomographie und Kernspintomographie. Nervenarzt 67 (1996) 721-729
71 Schwartz, A, G. Brix, F. Gückel, M. Bellemann, U. Kischka, M.Weisbrod, M. Spitzer: Functional MRI of higher cognitivefunction using task separation. Radiology 193 (P) (1994) 187
72 Sereno, M. 1., A. M. Dale,]. B. Reppas, K. K. Kwong,]. W. Belliveau,T.]. Brady, B. R Rosen, R B. H. Toatell : Borders of multiple visualareas in humans revealed by functional magnetic resonanceimaging. Science 268 (1995) 889-893
73 Silbersweig, D. A., E. Stern, C. Frith, C. Cahill, A Holmes, S. Grootoonk, ]. Seaward, P. McKenna, S. E. Chua, L. Schnorr, T. jones, R.S.]. Frackowiak: A functional neuroanatomy of hallucinations inschizophrenia. Nature 378 (1995) 176-179
74 Smith, E. E., ]. jonides, R. A Koeppe, E. Awh, E. H. Schumacher,S. Minoshima: Spatial versus object working memory: PET investigations.]. Cogn. Neurosei. 7 (1995) 337-356
75 Spitzer, M.: The psychopathology, neuropsychology, and neurobiology of associative and working memory in schizophrenia.Eur. Arch. Psychiatry Clin. Neurosei. 243 (1993) 5-70
76 Spitzer, M., M. E. Bellemann, F. Gückel, G. Brix, U. Kischka, T.Kammer, ASchwartz, S. Seyyedi, M. Weisbrod: Evidence formultiple distributed cortical map-Jike representations oflanguage categories from fMRI. Paper presented at the 3rd Proc.Soc. Magn. Reson. Med. Nice (1995) p. 1341
77 Spitzer, M., M. E. Bellemann, T. Kammer, F. Gückel, U. Kischka, S.Maier, ASchwartz, G. Brix: Functional MR imaging of semanticinformation processing and learning-related effects using psychometrically controlled stimulation paradigms. Cogn. BrainRes. 4 (1996) 149-161
78 Spitzer, M., T. Kammer, M. E. Bellemann, F. Gückel, M. Georgi, AGass, G. Brix: Funktionelle Magnetresonanztomographie bei Aktivierung des Arbeitsgedächtnisses. Röfo. Fortsehr. Röntgenstr.neuen bildgebend. Verf. 165 (1996) 52-58
79 Spitzer, M., U. Kischka, F. Gückel, M. E. Bellemann, T. Kammer, S.Seyyedi, M. Weisbrod, A. Schwatz, G. Brix: Functional magneticresonance imaging of category-specific cortical activation: Evidence for semantic maps. Cogn. Brain Res., in press, 1998
80 Spitzer, M., K. K. Kwong, W. Kennedy,]. R. Barker,]. W. Belliveau:Neuromodulatory effects on higher cognitive functions: an fMRIpilot study on language- vs. color-related decisions on the samestimuli. Hum. Brain. Mapp. Suppl. 1 (1995) 222
81 Spitzer, M., K. K. Kwong, W. Kennedy, B. R Rosen,]. W. Belliveau:Category-specific brain activation in fMRI during picture naming. Neuroreport 6 (1995) 2109-2112
82 Talairach, ]., P. Tournoux: Co-planar stereotaxie atlas of the human brain. Springer, New York 1988
83 Thatcher, R, M. Haller, T. Zeffiro, E. R. john, M. Huerta (Eds.):Functional neuroimaging. Technical Foundations. AcademicPress, San Diego, New York, Boston 1994
84 Thulborn, K. R., D. Davis, P. Erb, M. Strojwas, J. A. Sweeney: Clinical fMRI - implementation and experience. Neuroimage 4(1996) 101-107
85 Toga, A w., J. c. Mazziotta (Eds.): Brain mapping. The methods.Academic press, San Diego 1996
86 Toatel!, R. B. H., A M. Dale, M. I. Sereno, R Malach: New imagesfrom human visual cortex. Trends Neurosci. 19 (1996) 481-489
87 Turner, R, P. jezzard, H. Wen, K. K. Kwong, D. Le Bihan, T. Zeffiro,R. S. Balaban: Functional mapping of the human visual cortex at4 and 1.5 tesla using deoxygenation contrast EP!. Magn. Reson.Med. 29 (1993) 277-279
88 Vreeswijk, C. van, H. Sompolinsky: Chaos in neuronal networkswith balanced excitatory and inhibitory activity. Science 274(1996) 1724-1726
89 Wenz, F., L. R. Schad, M. V. Knopp, K. T. Baudendistel, F. Flömer,]. Schröder, G. van Kaiclc Functional magnetic resonance imaging at 1.5 T: Activation pattern in schizophrenie patients receiving neuroleptic medication Magn. Reson. Imaging 12 (1994)975-982
90 Wilson, F. A w., S. P. 6 Scalaidhe, P. s. Goldman-Rakic: Dissociation of object and spatial processing domains in primate prefrontal cortex. Science 260 (1993) 1955 -1957
91 Woodruff, P., M. Brammer,]. Meilers, I. Wright, E. Bullmore, S.Williams: Auditory hallucinations and perception of externalspeech. Lancet 346 (1995) 1035
92 Woods, R. P. w., S. R Cherry, J. c. Mazziotta: Rapid automatedalgorithm for aligning and reslicing PET images. J. Comput. Assist. Tomogr. 16 (1992) 620-633
93 Worden, M., W. Schneider: Cognitive task design for fMRI. Int.].Imag. Sys. Techno!. 6 (1995) 253-270
94 Zeffiro, T.: Clinical functional image analysis: artifact detectionand reduction. Neuroimage 4 (1996) S95-S100
95 Zeki, S.,]. D. G. Watson, c.]. Lueck, K.]. Friston, C. Kennard, R. S.]. Frackowiak: A direct demonstration of functional specialization in human visual cortex. J. Neurosei. 11 (1991) 641- 649
96 ZiIles, K., G. Schlaug, M. Matelli, G. Luppino, ASchleicher, M. Qu,A. Dabringhaus, R Seitz, P. E. Roland: Mapping of human andmacaque sensorimotor areas by integrating architectonic, transmitter receptor, MRI and PET data.]. Anat. 187 (1995) 515 - 537
Prof. Dr. Dr. M. Spitzer
Psychiatrische Universitätsklinik UlmLeimgrubenweg 12-14D-89070 Ulm
Her
unte
rgel
aden
von
: Uni
vers
ität U
lm. U
rheb
erre
chtli
ch g
esch
ützt
.





























