Embed Size (px)
Citation preview

Aus der Klinik für Dermatologie
der Universität zu Lübeck
Direktor: Prof. Dr. med. Detlef Zillikens
Immunpathologische Veränderungen in der Effektorphase des allergischen Kontaktekzems im
Menschen
Inauguraldissertation
zur
Erlangung der Doktorwürde der Universität zu Lübeck
-Aus der Medizinischen Fakultät-
vorgelegt von Christine Bangert
aus Mosbach
Lübeck 2009

1. Berichterstatter: Prof. Dr. Jürgen Grabbe
2. Berichterstatter: PD Dr. Helmut Haas
Tag der mündlichen Prüfung: 14.01.2010
zum Druck genehmigt. Lübeck, den 14.01.2010
gez. Prof. Dr. med. Werner Solbach
-Dekan der Medizinischen Fakultät -

INHALTSVERZEICHNIS
1 EINLEITUNG 1
1.1. Definition und Klinik des Kontaktekzems 1
1.2 Die Histologie des allergischen Kontaktekzems 5
1.3 Haptene 7 1.4. Die Epikutantestung 10 1.5. Die Pathogenese des Kontaktekzems 12 1.5.1. Dendritische Zellen 13 1.5.2. T-Zellen im Kontaktekzem 18 1.5.3. Die Sensibilisierungsphase 20 1.5.4. Die Effektorphase 21
2 ZIELSETZUNG 25
3 MATERIAL UND METHODEN 26
3.1 Material 3.1.1 Patienten 26 3.1.2 Biopsien 28 3.1.3 Blutproben 28 3.1.4 Antikörper 29 3.1.5 Chemikalien 31 3.1.6 Verbrauchsmaterial 32 3.1.7 Geräte 33 3.1.8 Puffer 33
3.2 Methoden 3.2.1 Anfertigung von Gefrierschnitten 34 3.2.2 Immunhistochemie 34 3.2.3 Immunfluoreszenz 35 3.2.4 Auswertung der Immunfluoreszenz 36 3.2.5 Gewinnung von PBMC 37 3.2.6 Herstellung von Einzelzellsuspensionen 37 3.2.7. Zellzählung in Suspension 37 3.2.8. Durchflußzytometrie 38 3.2.8.1 Durchführung und Darstellung 38 3.2.8.2 Phänotypische Analysen von Oberflächenmarkern 38

4 ERGEBNISSE 40
4.1. Anzahl und Verteilung der Leukozyten 40 4.2. T-Zellen im ECT 42 4.2.1. Typisierung der T-Zellen im dermalen Infiltrat 42 4.2.2. Charakterisierung der CD4+/CD25+ T-Zellsubpopulation 45 4.3. DZ Populationen im ECT 50 4.3.1. LZ im ECT 50 4.3.2. DDZ im ECT 52 4.3.3. Maturationsverhalten von LZ und DDZ im ECT versus NH 53 4.3.4. PDZ im Infiltrat der ECT Reaktionen 55 4.3.5. NK-Zellen und pDZ 655 4.4. Vergleich des Zellinfiltrats in ECT Reaktionen induziert durch Nickel oder Duftstoff 69 4.4.1. T-Zell-Infiltrat 69 4.4.2. DZ-Populationen 69 4.4.3. Identifizierung weiterer Leukozyten 70 4.4.4. Reaktion eines Nicht-Allergikers auf 24Std Nickelprovokation 70
5 DISKUSSION 74
6 ZUSAMMENFASSUNG 84
7 LITERATURVERZEICHNIS 86
8 DANKSAGUNG 97
9 LEBENSLAUF 98
10 ANHANG 103

ABKÜRZUNGSVERZEICHNIS APZ Antigen-präsentierende Zelle
CLA kutanes Lymphozyten-assoziiertes Antigen
DNFB Dinitrofluorbenzol
DDZ dermale dendritische Zelle
DZ dendritische Zelle
ECT Epikutantest
GM-CSF Granulozyten Macrophagen-Kolonie Stimulierender Faktor
HEV ‚high endothelial venules’
IDEZ inflammatorisch-dendritische epidermale Zellen
IDDZ inflammatorisch-dendritische dermale Zellen
IFN Interferon
IL Interleukin
KE Kontaktekzem
LZ Langerhans Zelle
LFA-1 Lymphozytenfunktion-assoziiertes Antigen-1
LK Lymphknoten
MHC ‘Major Histocompatibility Complex’
Ni Nickel
NH Normalhaut
NK-Zelle natürliche Killerzelle
PBMC periphere Blut-mononukleäre Zellen
pDZ plasmazytoide dendritische Zelle
Std Stunde
reg T-Zellen regulatorische T-Zellen
TGF ‚Transforming-Growth-Faktor’
TNF Tumor-Nekrose-Faktor
TZR T-Zellrezeptor

Einleitung Ch. Bangert
- 1 -
1. EINLEITUNG
1.1. Definition und Klinik des Kontaktekzems
Das Ekzem ist eine durch eine typische (krankheitsspezifische) Gewebsreaktion
charakterisierte Intoleranzreaktion der Haut auf meist äußerlich einwirkende, nicht-
infektiöse Noxen.
Klinisch-histologisch läßt sich die akute Ekzemreaktion in 4 Stadien einteilen: Im
Stadium erythematosum kommt es zur scharf begrenzten Rötung des betroffenen
Areals mit Ausbildung eines intraepidermalen, interzellulären Ödems (Spongiose), das
die Hautoberfläche matt erscheinen lässt. Das anschließende Stadium vesiculosum
zeichnet sich durch spongiotische Bläschenbildung aus, die von heftigem Juckreiz
begleitet wird. Durch Platzen der wasserklaren Bläschen entsteht eine nässende
Läsion, der sich nach Eintrocknen des Exsudats das Krustenstadium anschließt
(Stadium crustosum). Falls es zu keiner weiteren Noxeneinwirkung kommt, heilt das
Ekzem mit Schuppenbildung ab (Stadium squamosum). Bei andauerndem oder
wiederholtem Kontakt mit der auslösenden Substanz zeigt das klinische Bild
verschiedene Morphen (Erythem, Bläschen, Krusten, Schuppen und entzündliche
Knötchen), die von Kratzexkoriationen überlagert sein können. Im chronischen Stadium
des Ekzems kommt es zum Abklingen der akuten Entzündung und zur Lichenifizierung
der Haut [1], [2].
Ekzeme gehören mit 3 bis 20% Prävalenz zu den am häufigsten vorkommenden
Hautkrankheiten weltweit. Bedingt durch die große Vielfalt der ekzematösen Reaktion
(z.B. seborrhoisches, atopisches, nummuläres (mikrobielles), dyshidrotisches Ekzem,
Stauungsekzem) wird die Wahrscheinlichkeit, im Laufe des Lebens mindestens einmal
an einem Ekzem zu erkranken, auf etwa 100% geschätzt.
Zu den sehr häufigen Erscheinungsformen der ekzematösen Reaktion zählen das
allergische und toxische Kontaktekzem (KE).
Das toxische KE entsteht bereits bei Erstkontakt mit toxisch wirksamen Substanzen
(Alkalien, Säuren, organische und anorganische Öle, Lösungsmittel etc.) und ist
meistens an besonders exponierten Körperstellen (Unterarme und Hände) lokalisiert.
Ein klassisches Beispiel für ein toxisch-irritatives KE ist das sogenannte
‚Hausfrauenekzem’ bei Personen, die ohne Handschuhe in häufigen Kontakt mit
Wasser, Waschlaugen und Detergentien kommen. Durch das wiederholte Schwellen
der Hornschicht entwickelt Wasser einen eigenen irritierenden Einfluß, durch den die
inflammatorische Wirkung toxischer Substanzen verstärkt werden kann [1], [2].

Einleitung Ch. Bangert
- 2 -
Das allergische KE hingegen entsteht bei Kontakt der Haut mit chemisch
hochreaktiven, kleinmolekularen Substanzen, genannt Haptene, über eine allergische
Typ-IV Reaktion (Abb 1.1, 1.2). Diese allergische Reaktion wird in zwei Phasen
unterteilt [3]: in der Sensibilisierungs phase kommt es zur Ausbildung von
haptenspezifischen Effektor T-Zellen, die dann 48-72 Stunden (Std) nach erneutem
Kontakt mit dem entsprechenden Hapten in der Effektor phase die eigentliche
ekzematöse Reaktion auslösen (siehe auch 1.5. ‚Die Pathogenese des
Kontaktekzems’).
Im Gegensatz zum toxischen KE kommt es nur bei einem Teil der exponierten
Personen zu einer Ekzemreaktion.
Einen bevorzugten Befall eines bestimmten Körperteils gibt es nicht, da Haptene an
unterschiedlichen Stellen mit der Haut in Kontakt kommen (Abb. 1.1., 1.2.).
Obwohl es etwa 5-10x seltener als das toxische KE auftritt, so spielt das allergische KE
doch eine wesentlich bedeutsamere Rolle als Berufskrankheit. Da eine Chronifizierung
des KEs nur durch Haptenkarenz zu vermeiden ist, können betroffene Personen in
letzter Konsequenz zur Aufgabe ihres ursprünglich gewählten Berufes gezwungen sein
und sich dadurch plötzlich mit sozial und wirtschaftlich schwer lösbaren Problemen
konfrontiert sehen [2], [4].
Außer im Berufsleben kann es auch in der Freizeit zu relevanter Allergenexposition
kommen, die, je nach Hapten, unterschiedlichste Substanzklassen betrifft
(Körperpflege und Kosmetik, Tragen von Schmuck, Kontakt mit Textilien, Schuhwerk,
Salben oder Desinfektionsmittel).
Die Therapiemöglichkeiten des KEs sind derzeit noch limitiert. Im akuten Ekzemschub
gelten lokaltherapeutische Maßnahmen mit z.B. steroidhaltigen Produkten sowie
pflegenden Externa als Mittel der Wahl, eine längerfristige Heilung kann jedoch nur
durch die strikte Meidung des Haptens erzielt werden.
Einleitung Ch. Bangert
- 3 -
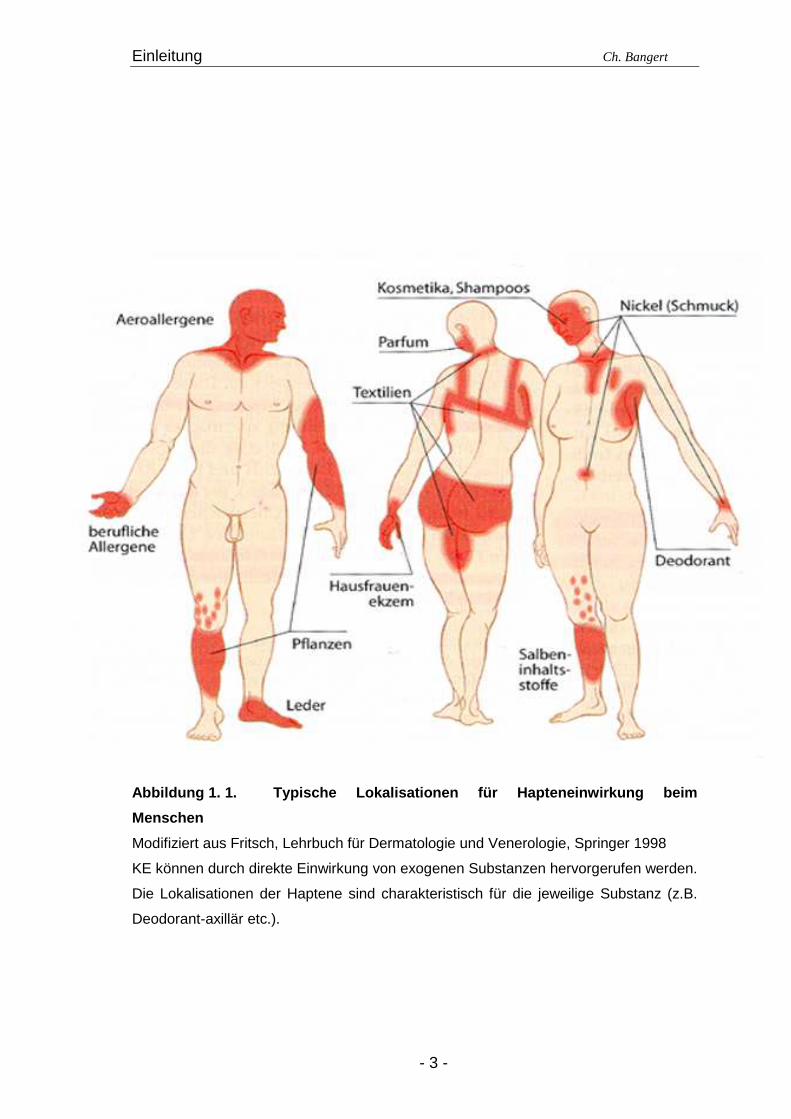
Abbildung 1. 1. Typische Lokalisationen für Haptene inwirkung beim
Menschen
Modifiziert aus Fritsch, Lehrbuch für Dermatologie und Venerologie, Springer 1998
KE können durch direkte Einwirkung von exogenen Substanzen hervorgerufen werden.
Die Lokalisationen der Haptene sind charakteristisch für die jeweilige Substanz (z.B.
Deodorant-axillär etc.).

Einleitung Ch. Bangert
- 4 -
Abbildung 1.2. Allergische Kontaktdermatitis ausgel öst durch Nickel
Modifiziert aus Johannes Ring, Allergy in Practice, Kapitel 5: Allergic Diseases (and
Differential Diagnosis), Springer 2005
Das obere Bild zeigt eine Patientin mit einem Kopfschmuck aus Metall, der nach
längerem Tragen eine allergische Kontaktdermatitis mit typischer lokal begrenzter
Rötung und Bläschen auslöste (unteres Bild).

Einleitung Ch. Bangert
- 5 -
1.2 Die Histologie des allergischen Kontaktekzems
Die Histologie des akuten allergischen KE zeigt üblicherweise eine Überlappung mit
anderen Arten von akuter Dermatitis und ist deswegen nicht als einziges
diagnostisches Kriterium zu sehen, sondern eher als unterstützende Maßnahme bei
Verdacht auf das Vorliegen eines KE.
Die klassischen histologischen Veränderungen der kontaktallergischen Reaktion
zeigen im frühen Stadium ein Ödem in den unteren Schichten der Dermis, gefolgt von
spongiotischer Bläschenbildung auf verschiedenen epidermalen Ebenen [5] (Abb. 1.3).
Diese Bläschenbildung wird durch die Ödem-bedingte Dehnung der interzellulären
Räume und die damit verbundene Auflösung der Haftung mittels Desmosomen
verursacht. Meistens finden sich die Vesikel intraepidermal, allerdings können sie sich
bei sehr starker Entzündung auch bis subepidermal ausbreiten. Im Allgemeinen kommt
es nur zu einer geringen Akanthose der Epidermis [6].
Die obere Dermis zeigt je nach Ausmaß des Ekzems ein deutlich leukozytäres,
mononukleares Infiltrat, das vor allem um den oberen Gefäßplexus lokalisiert ist. Auch
eosinophile Granulozyten sind vorhanden und können mittels H&E (Hämatoxylin
&Eosin) Färbung gut dargestellt werden.
Im Unterschied zu der oben beschriebenen akuten Form des Ekzems findet sich in
chronischen Läsionen kaum Spongiose der Epidermis. Statt dessen steht die
epidermale Akanthose im Vordergrund, die einer psoriasiformen Hyperplasie der
Epidermis ähneln kann [5]. Häufig zeigt sich eine dicke, hyperkeratotische Hornschicht
mit minimaler Parakeratose. In der papillären Dermis kann es zu einer milden
Fibrosierung kommen.

Einleitung Ch. Bangert
- 6 -
Abbildung 1.3. Die Histologie des KE
Modifiziert aus David Weedon, Skin Pathology Second Edition, Churchill Livingstone
2002
Die H&E Färbung zeigt spongiotische Vesikulation in der Epidermis (oberes Bild, Pfeil).
Im dichten inflammatorischen Infiltrat der papillären Dermis finden sich u.a. eosinophile
Granulozyten, die durch ihre intensive rote Farbe auffallen (unteres Bild, Pfeil).

Einleitung Ch. Bangert
- 7 -
1.3. Haptene
Der Begriff ‚Hapten’ wurde erstmals 1935 von Karl Landsteiner für niedermolekulare
Chemikalien eingeführt, die im Tierversuch nach kovalenter Bindung an Proteine
Hapten-spezifische Antikörper induziert haben [3].
Obwohl Ursprung und Natur dieser kleinen allergenen, elektrophilen Substanzen sehr
unterschiedlich sein können, stimmen sie in einigen Eigenschaften überein: sie sind
niedermolekular (MG < 500 Dalton), und können daher leicht die Hautbarriere
überschreiten, in die normalerweise größere Moleküle nicht eindringen können [7]. Der
Besitz lipophiler Reste erleichtert den meisten Haptenen zusätzlich das Überwinden
der Barriereschicht der Epidermis.
Aufgrund der geringen Größe besitzen Haptene selbst keine immunogenen
Eigenschaften, sondern brauchen die (kovalente) Bindung an körpereigene
Trägerpeptide wie z.B. Serumalbumin, epidermale Proteine oder in den ‚Major
Histocompatibility Complex’ (MHC)-Gruben präformierte Peptide, um eine immunogene
Wirkung durch T-Zellantworten zu erlangen [8]. Die Bindung an Proteine kann sehr
verschieden sein: bei Bindung an ein lösliches Trägerprotein wird das Hapten indirekt
von CD4+ T-Zellen über MHCII auf Antigen-präsentierenden Zellen (APZ) erkannt, da
das lösliche Protein vor Präsentation seiner Peptide prozessiert wird. Alternativ können
Haptene aber auch direkt an MHC-assoziierte Moleküle binden und dann entweder
über MHC Klasse I von CD8+ T-Zellen oder über MHC Klasse II von CD4+ T-Zellen
erkannt werden [8].
Andere Haptene, die z.B. von instabilen chemischen Substanzen abstammen, können
durch einen zusätzlichen Metabolisierungsschritt in einen Stoff mit allergener Wirkung
umgewandelt werden. Metallsalze, beispielsweise, gehen mit kutanen Proteinen keine
kovalente Bindung ein, sondern formen lediglich Komplexe über schwache chemische
Interaktionen.
Bislang wurden etwa 3000 Haptene entdeckt (Tab. 1.1), wobei nur ein kleiner Teil
davon häufig in unserer Umwelt zu finden ist. Ihre chemische Struktur reicht von
einfachen Metallionen, zum Beispiel Nickel (Ni), bis zu komplexen aromatischen
Ringen in Duftstoffen.
In der industrialisierten Welt zählt Ni zu den häufigsten Auslösern einer Kontaktallergie
[9]. Dieses Hapten wurde in vielen Studien zur Klärung der Struktur des Antigens sowie
zur Identifizierung der Ni-Protein-Bindungsstelle an T-Zellen untersucht [10]; [11]; [12].
Anders als klassische Haptene formen Ni-Ionen keine kovalenten Brücken mit
Proteinen, sondern werden in reversiblen Komplexen strukturiert [13].

Einleitung Ch. Bangert
- 8 -
Ni kann von T-Zellen sowohl direkt als auch indirekt erkannt werden, da nicht alle Ni-
spezifischen T-Zellklone in der Lage sind, das Hapten direkt auf Aldehyd-fixierten APZ
zu erkennen. Dies bedeutet, dass Ni vor seiner immunogenen Wirkung auf T-Zellen
zumindest zum Teil prozessiert werden muß [14].
Als Protein, das für den Transport des Ni-Ions von der äußeren Hautoberfläche zu den
APZ in der Epidermis [z.B. Langerhans Zellen (LZ)], verantwortlich sein könnte, wurde
aufgrund seiner bekannten Bindungskapazität an Ni das humane Serumalbumin in
Betracht gezogen. In einer rezenten Arbeit konnte tatsächlich gezeigt werden, dass
humanes Serumalbumin -Ni Komplexe, in Anwesenheit geeigneter APZ, Ni-reaktive T-
Zellen stimulieren können [12].
Im Gegensatz zu dem gut erforschten Ni-Ion sind experimentelle Daten zum
Mechanismus der Haptenpräsentation des Duftstoffmix eher begrenzt. Duftstoffe
kommen in einer Vielzahl von Kosmetik- und Haushaltsprodukten vor, eine Exposition
ist schwer vermeidbar. Der klassische Duftstoffmix, der auch bei Allergietestungen
verwendet wird, besteht aus Eugenol, Isoeugenol, Geraniol, Eichenmoos, a-Amyl
Zimtaldehyd, Zimtaldehyd, Zimtalkohol und Hydroxycitronella. Die häufigsten
allergenen Komponenten des Mix sind Isoeugenol, Eichenmoos, Hydroxycitronella und
Eugenol. Spezifische T-Zellantworten auf Eugenol und Isoeugenol konnten anhand von
Lymphozyten- Transformationstestungen nachgewiesen werden [15], so dass eine rein
irritative Reaktion auf Duftstoffe ausgeschlossen werden kann. Auch ist bekannt, dass
Zimtalkohol und Zimtaldehyd ihre allergene Wirkung durch ein gemeinsam
metaboliertes Hapten ausüben [16].
Es ist jedoch besonders wichtig zu erwähnen, dass nicht alle kleinen, elektrophilen
Moleküle nach Proteinbindung zwangsläufig bei jeder Person als Allergen wirken. Zu
den prädisponierenden Faktoren, die zur Entwicklung eines allergischen KE beitragen
können, zählen neben der Notwendigkeit einer kritischen Haptenmenge eine
bestehende Vorschädigung der Haut und vor allem auch der Histokompatibilitätstyp
der betreffenden Person.

Einleitung Ch. Bangert
- 9 -
Tabelle 1.1. Häufige Kontaktallergene und ihr Vorkommen (modifiziert nach Fritsch,
Lehrbuch für Dermatologie und Venerologie, Springer 1998)
Nickelsulfat: Metalle, Modeschmuck, Katalysatoren, Farben, Euromünzen
Kobaltsulfat: Zement, Galvanisation, Öle, Kühlmittel, Lacke
Duftsstoffe: Parfüms, Kosmetika
Perubalsam: Salben, Kosmetika, Lokalanästhetika
Wollwachsalkohole: Salbengrundlagen, Kosmetika, Skiwachse, Möbelpolitur
Neomycin: antibiotischer Zusatz in Lokaltherapeutika
Procain, Benzocain: Lokalanästhetika, antipruriginöse und analgesierende Externa
Kaliumdichromat: Zement, Antioxidans, Farben, Öle, Gerbstoffe, Streichhölzer
Thiomersal (Sulfonamide): Lokaltherapeutika, Augentropfen
Sorbitansesquioleat: Emulgator in Lokaltherapeutika, Kosmetika
Acrylate: Prothesen, Zahntechnik, Lacke
Kolophonium: adhäsives Harz, in Wachsen
Para-Phenylendiamin: dunkle Farbstoffe in Textilien, Druckerschwärze

Einleitung Ch. Bangert
- 10 -
1.4 Die Epikutantestung
Die gängige Testmethode zur Erfassung von Typ IV- Sensibilisierungen erfolgt durch
die Epikutantestung. Josef Jadassohn, der im Jahre 1895 erstmalig eine
kontaktallergische Reaktion auf Quecksilber beschrieb, brachte den Test im Jahre
1896 auf [17]. Im Falle einer korrekten Durchführung und Interpretation mit genauen
Regeln und Grundlagen stellt der Epikutantest (ECT) bis heute eine wissenschaftlich
anerkannte Untersuchungsmethode zur Diagnosestellung des KE dar.
Die Prinzipien der Testung sind in Tabelle 1.2. aufgeführt. Die zu testenden
Substanzen werden in eine „Finn chamber“ (kleine Aluminiumschälchen) gefüllt und
mittels Pflaster auf dem Rücken des Patienten fixiert (Abb. 1.4.). Der Vorteil des ECT
besteht in dem relativ kleinen einzelnen Testareal, wodurch eine große Anzahl
Substanzen gleichzeitig getestet werden kann, und zusätzlich selbst bei massiver
allergischer Reaktion auf eine zu testende Substanz keine körperliche
Beeinträchtigung des Patienten entsteht.
Die Beurteilung des ECT erfordert eine gewisse Erfahrung des Untersuchers, der die
Reaktionen nach einer bestimmten Skala einteilt:
Negativ (0)
Positiv +/-: mildes Erythem
Positiv + : Erythem mit leichter Infiltration
Positiv ++ : Erythem mit Papeln und kleinen Bläschen
Positiv +++ : Erythem, Blasen und Erosionen
Die Typ IV Allergie zeigt ihr Reaktionsmaximum nach 2-3 Tagen. Prinzipiell gilt, dass
ein allergisch bedingtes KE eine Zunahme der Reaktionsstärke von 48 Std nach 72 Std
zeigt, während ein irritatives KE eine „decrescendo“ Reaktion zwischen erster und
zweiter Ablesung aufweist.

Einleitung Ch. Bangert
- 11 -
Tabelle 1.2. Prinzipien der Epikutantestung (modifiziert nach: Robert L. Rietschel,
Fisher’s Contact Dermatitis, Fifth Edition)
1. Nur bekannte Substanzen werden anhand von standardisierten Konzentrationen
getestet. Falls Unsicherheiten bestehen, wird eine offene Testung mit
entsprechenden Kontrollen durchgeführt. Industrielle Substanzen unbekannter
Konzentration sollten nicht getestet werden.
2. Die Testung darf nur an Lokalisationen durchgeführt werden, die keine Zeichen einer
akuten Dermatitis aufweisen.
3. Der Patient wird ersucht, das Testpflaster für 48 Std nicht zu entfernen.
4. Der Patient wird während der Testzeit ersucht, nicht zu duschen, den Rücken
trocken zu halten und Sport zu vermeiden. Schwere körperliche und
schweißtreibende Arbeit sollte ebenfalls vermieden werden.
5. Nach 48 Std werden die Substanzen entfernt und eine Erstablesung durchgeführt.
Weitere Ablesungen sollten zwischen 72 und 120 Std nach Aufbringen der
Substanzen vorgenommen werden.
6. Irritative und allergische Reaktionen können in vielen Fällen schwer voneinander
unterschieden werden. In der Regel verursachen allergische Reaktionen mehr
Juckreiz als irritative Reaktionen.
Einleitung Ch. Bangert
- 12 -
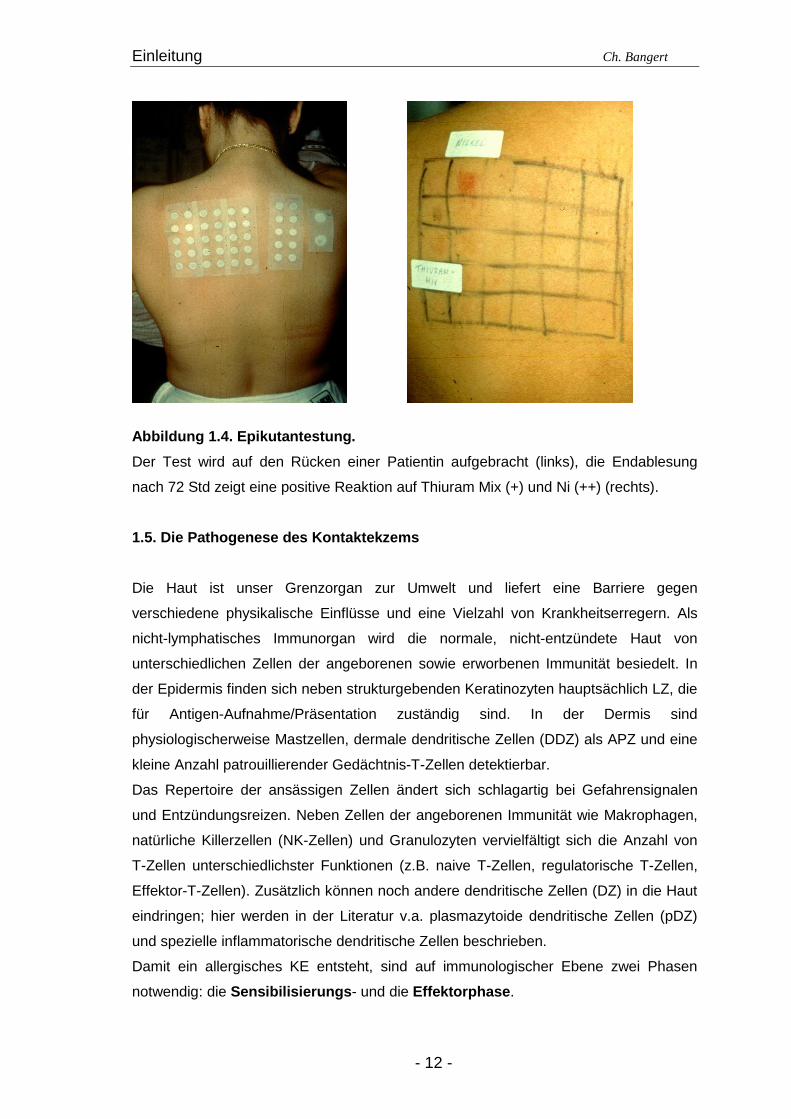
Abbildung 1.4. Epikutantestung.
Der Test wird auf den Rücken einer Patientin aufgebracht (links), die Endablesung
nach 72 Std zeigt eine positive Reaktion auf Thiuram Mix (+) und Ni (++) (rechts).
1.5. Die Pathogenese des Kontaktekzems
Die Haut ist unser Grenzorgan zur Umwelt und liefert eine Barriere gegen
verschiedene physikalische Einflüsse und eine Vielzahl von Krankheitserregern. Als
nicht-lymphatisches Immunorgan wird die normale, nicht-entzündete Haut von
unterschiedlichen Zellen der angeborenen sowie erworbenen Immunität besiedelt. In
der Epidermis finden sich neben strukturgebenden Keratinozyten hauptsächlich LZ, die
für Antigen-Aufnahme/Präsentation zuständig sind. In der Dermis sind
physiologischerweise Mastzellen, dermale dendritische Zellen (DDZ) als APZ und eine
kleine Anzahl patrouillierender Gedächtnis-T-Zellen detektierbar.
Das Repertoire der ansässigen Zellen ändert sich schlagartig bei Gefahrensignalen
und Entzündungsreizen. Neben Zellen der angeborenen Immunität wie Makrophagen,
natürliche Killerzellen (NK-Zellen) und Granulozyten vervielfältigt sich die Anzahl von
T-Zellen unterschiedlichster Funktionen (z.B. naive T-Zellen, regulatorische T-Zellen,
Effektor-T-Zellen). Zusätzlich können noch andere dendritische Zellen (DZ) in die Haut
eindringen; hier werden in der Literatur v.a. plasmazytoide dendritische Zellen (pDZ)
und spezielle inflammatorische dendritische Zellen beschrieben.
Damit ein allergisches KE entsteht, sind auf immunologischer Ebene zwei Phasen
notwendig: die Sensibilisierungs - und die Effektorphase .

Einleitung Ch. Bangert
- 13 -
In der klinisch stummen Sensibilisierungsphase wird ein eindringendes Hapten von
APZ erstmals den T-Zellen als ‚Gefahr’ präsentiert, woraufhin diese zur klonalen
Proliferation angeregt werden.
Die Effektorphase resultiert 48-72 Std nach erneutem Haptenkontakt im klinischen Bild
eines Ekzems.
Der ausführliche Ablauf dieser beiden Phasen nach derzeit gültigem Konzept sowie die
möglichen Zellpopulationen, die an der Pathogenese des allergischen KE beteiligt sind,
werden im Folgenden genauer erörtert.
1.5.1. Dendritische Zellen
Langerhans Zellen
Paul Langerhans beschrieb 1868 mittels der damals genutzten Goldchlorid-
Impregnationstechnik erstmals eine dendritisch geformte Zelle in der menschlichen
Epidermis [18]. Die so identifizierte LZ war die erste DZ, die überhaupt detektiert
wurde. Trotz dieser frühen Entdeckung blieb die wahre Identität dieser berühmten Zelle
noch mehr als ein Jahrhundert verborgen.
Zunächst wurde die LZ irrtümlich den Nervenzellen zugeordnet, später Melanozyten,
die ihre Fähigkeit zur Pigmentbildung bereits verloren hatten [19]. Birbeck et al gelang
Anfang der 60er Jahre sowohl die elektronenmikroskopische Darstellung von
Tennisschläger-ähnlich geformten Zellorganellen in den LZ, seither als Birbeck Granula
bezeichnet, als auch die Detektion der Zellen in der Melanozyten-freien Vitiligo [20].
Zusätzlich konnte in den darauffolgenden Jahren die Fähigkeit der ATPase (CD39)
Expression von LZ festgestellt werden [21], [22].
Die Verbindung zum Immunsystem wurde in den 70er Jahren hergestellt, als MHCII
Moleküle, Fc- und Komplementrezeptoren auf LZ entdeckt wurden [23], [24], [25].
Diesen ersten Berichten folgte bald der Nachweis der Antigenpräsentation der LZ
sowohl in Meerschweinchen und Mäusen als auch im Menschen [26], [27], [28], [29].
Heute zählt die LZ zu den gut charakterisierten Zellen, die noch immer die
Aufmerksamkeit vieler Wissenschaftler fesselt.
Die Dichte der LZ in der normalen menschlichen Haut variiert je nach Körperteil. So
findet sich z.B. an Kopf, Gesicht oder Stamm eine hohe Anzahl von LZ (zwischen 600-
1000/mm2), während Hand- und Fußsohlen lediglich ~200/mm2 aufweisen.
Die Vorläuferzelle der LZ ist die sogenannte CD34+ hämatopoetische Vorläuferzelle
und stammt aus dem Knochenmark [30], [31]. Zur Entwicklung der LZ aus dieser Zelle

Einleitung Ch. Bangert
- 14 -
werden die Zytokine GM-CSF, Interleukin (IL)-3, Tumor-Nekrose-Faktor (TNF)-α und
‚Transforming Growth Factor’ (TGF)-β1 benötigt [31], [32].
Die Besiedelung der Epidermis während der Ontogenese ist mit dem stufenweisen
Erwerb immunologisch wichtiger Moleküle wie CD45, CD1a, MHCII Komplex (HLA-
DR/DQ/DP), ATPase und Langerin (CD207) verbunden [33], [34]. Aufgrund der
Expression von Antigenen wie CD11b oder CD33 wird die LZ außerdem einem
myeloiden Ursprung zugeordnet [35], [36]. In letzter Zeit durchgeführte in vitro
Experimente lassen vermuten, dass die Differenzierung der LZ auch aus einer CD14+
myeloid-monozytären Zelle erfolgen kann [37].
Das an der Zelloberfläche der LZ exprimierte Antigen Langerin ist ein Kalzium-
abhängiges C-Typ Lektinmolekül der Gruppe II (Familie der Asialoglycoproteine), das
an der Entstehung der charakteristischen Birbeck Granula beteiligt ist [38]. Aufgrund
seines einzigartigen Vorkommens auf LZ eignet sich das Molekül hervorragend zur
Charakterisierung dieser epidermalen DZ.
C-Typ Lektine stellen ‚pattern recognition receptors’ (PRR) dar, die eine große Anzahl
unterschiedlicher Antigene/Mikroorganismen wie HIV, Hepatitis C Virus, CMV, EBV,
Helicobacter pylori etc. binden können [39], [40], [41]. Die genaue Identifizierung der
Mikroorganismen, die an Langerin binden, ist allerdings noch nicht gelungen. Auch ist
unklar, welche Funktion Langerin bei der Entwicklung der LZ tatsächlich einnimmt, da
Mäuse, bei denen das Langerin Gen entfernt wird, keinen deutlichen Funktionsverlust
der LZ aufweisen [42].
LZ sind äußerst mobile DZ, die ein phagozytiertes Antigen via afferentem Lymphgefäß
in den Lymphknoten (LK) transportieren, wo es den T-Zellen präsentiert wird und so
eine Immunantwort auslösen kann.
Die proinflammatorischen Zytokine TNF-α und IL-1β, die von LZ-benachbarten
Keratinozyten nach unterschiedlicher Provokation produziert werden, bilden den ersten
Impuls zur Auswanderung der Zellen [43]. Dann folgt eine Kaskade von Ereignissen
(Stimulierung von Chemokinrezeptoren, Abregulierung von haftenden Integrinen,
Expression von Matrix-Metalloproteinasen), die schließlich zur kompletten Emigration
der epidermalen DZ aus der Haut führt [44], [45], [46].
Welche Rolle schreibt man nun dieser immunologisch äußerst vielfältigen Zelle im KE
zu?
LZ nehmen nach dem Kontakt, der zur Sensibilisierung führt, das Hapten auf,
prozessieren es und wandern danach zum nächsten regionären LK, um es dort den T-
Zellen zu präsentieren. In Studien konnte gezeigt werden, dass das Zytokin IL-1β und

Einleitung Ch. Bangert
- 15 -
das Chemokin Osteopontin (OPN) eine wichtige Rolle bei der Migration der LZ und
somit auch in der Sensibilisierungsphase des KE spielen [47], [48], [49]. In Mäusen
ohne IL-1β oder OPN wird die Auswanderung der LZ aus der Epidermis verhindert.
Eine bei diesen Tieren durchgeführte Sensibilisierung mit dem Kontaktallergen
Trinitrophenyl führt bei erneuter Haptenapplikation zu einer deutlich abgeschwächten
Ekzemreaktion, was auf eine bedeutende Rolle der LZ in der Induktionsphase des KE
deutet [47], [48].
Ob die LZ in der Effektorphase des humanen KE nun eher eine Effektorrolle einimmt
oder einen regulierenden Einfluß auf T-Zellen ausübt, ist nicht bekannt [51].
Ursprünglich wurde dieser DZ eine Antigen(Hapten)-präsentierende Funktion
zugedacht, welche die T-Zell-mediierte Entzündung des Ekzems provoziert [52]. Später
zeigte sich jedoch, dass eine Depletion der LZ nach Sensibilisierung in der Maus zu
einer verstärkten, und nicht wie erwartet, zu einer abgeschwächten Ekzemreaktion
führt [50].
Dermale (myeloide) dendritische Zellen
DDZ wurden mehr als 120 Jahre später beschrieben als LZ [36], [53]. Trotz intensiver
Forschung sind DDZ noch immer weniger gut charakterisiert als ihre epidermalen
Partner. Dies ist zum Großteil der Tatsache zu verdanken, dass bisher kein
unverwechselbarer Marker vergleichbar z.B. Langerin auf LZ identifiziert wurde und es
daher schwer ist, DDZ aus dem Blut oder der Haut zu isolieren. Meistens muß auf
Mausmodelle oder in vitro generierte DZ zurückgegriffen werden.
DDZ haben durch die ihnen eigenen, sogenannten C-Typ Lektine DC-SIGN (CD209)
und Mannose Rezeptor (CD206), ähnlich wie LZ über Langerin, die Fähigkeit
Mikroorganismen, Zellfragmente oder auch einzelne Moleküle zu phagozytieren.
Neben der PRR-assoziierten Aufnahme von Mikroorganismen, können DDZ Rezeptor-
mediierte Endozytose und Makropinozytose durchführen und sind damit in der Lage,
lösliches Protein zu prozessieren und zu präsentieren [54].
Die Wanderung von DDZ kann ebenfalls über inflammatorische Zytokine/Chemokine
gelenkt werden. Alleinige Daten über das Migrationsverhalten der DDZ- im Unterschied
zu LZ- wurden in einer rezenten Studie von Kissenpfennig et al mittels in vivo-
Mikroskopie an Mäusen geliefert [55]. Die Autoren konnten zeigen, dass DDZ nach
Haptenapplikation bereits nach 24 Std im ableitenden LK zu finden sind und die größte
Anzahl der Hapten-tragenden DDZ nach 48 Std erreicht wird. Antigen-beladene LZ

Einleitung Ch. Bangert
- 16 -
hingegen migrieren deutlich langsamer und zeigen ihren Zahlengipfel erst nach 4
Tagen. Auch wandern DDZ im Unterschied zu LZ eher zu B-Zell nahen Arealen im LK
(äußerer Paracortex), während sich LZ bei T-Zell Arealen (innerer Paracortex)
ansiedeln.
Humane DDZ können je nach der Expression von CD1a und CD14 in unterschiedliche
Subpopulationen unterteilt werden [53]; i) Faktor XIIIa++/CD1a++/CD4+/DC-SIGN-
/CD14- und ii) Faktor XIIIa++/CD1a+/-/CD4+/DC-SIGN+/CD14+. Letztere sind
schwächere allogene Stimulatoren und dadurch mit Makrophagen vergleichbar.
Inwieweit Makrophagen und DDZ einen gemeinsamen Differenzierungsweg aufweisen,
ist bislang noch nicht im Detail geklärt. Charakteristisch für DDZ, aber nicht exklusiv, ist
außerdem die Expression von klassischen (HLA-DQ/DR) und nicht-klassischen MHC
Molekülen (CD1a, CD1c, CD1d), dem Scavenger Rezeptor CD36, Anti-
koagulationsfaktor XIIIa sowie die Adhäsionsmoleküle CD11b/CD18 und CD11c/CD18
[53]; [56]. Die genannten Moleküle können jedoch unter gewissen Umständen
ebenfalls von Makrophagen exprimiert werden, weswegen besonders in entzündlichen
Hauterkrankungen mit dichtem leukozytärem Infiltrat eine eindeutige Zuordnung der
DDZ schwierig ist.
Eine der DDZ ähnliche Zellpopulation wurde unter entzündlichen Bedingungen im
Infiltrat atopischer Dermatitis beschrieben: die sogenannten inflammatorischen
epidermalen dendritischen Zellen (IDEZ) [57]. Diese DZ zeichnet sich durch die
Präsenz des hochaffinen IgE Rezeptors (FcεRI) aus sowie durch die hohe Expression
von MHCII, CD1c und CD36, und die niedrige Präsenz von CD1a und CD1b. Rezente
in vitro Studien zeigen, dass IDEZ, nach Stimulation von FcεRI, bei T-Zellen die
Produktion von IFN-γ anregen und dadurch die typische T2 Antwort in der atopischen
Dermatitis zugunsten einer T1 Reaktion verändern können [58].
Zusätzlich besitzen IDEZ, ähnlich wie DDZ, die Fähigkeit zur Rezeptor-mediierten
Endozytose, da sie den Mannose Rezeptor CD206 tragen [59].
Das Vorkommen von IDEZ im allergischen KE wurde bisher nicht beschrieben.
Aufgrund ihrer Antigen-präsentierenden Funktion ist aber eine Aufnahme und
Präsentation der Haptene prinzipiell möglich und somit eine entzündungsverstärkende
Rolle im KE denkbar.

Einleitung Ch. Bangert
- 17 -
Plasmazytoide dendritische Zellen
PDZ wurden erstmals im Jahre 1958 beschrieben, als man ‚Plasmazellen’ ohne B-
oder Plasmazellmarker in der T-Zellzone von humanen LK entdeckte und sie
dementsprechend plasmazytoide T-Zellen taufte [60]. 20 Jahre später wurde von
Trinchieri et al eine Zellart entdeckt, die auf virale Stimulation mit der Produktion von
Zytokin IFN-α reagierte, das wiederum zu einer deutlichen Stimulation des
angeborenen Immunsystems führt [61].
Es dauerte weitere 20 Jahre bis die Verbindung zwischen diesen beiden Zellarten
hergestellt werden konnte; im Jahre 1999 wiesen zwei unterschiedliche Gruppen nach,
dass es sich bei den plasmazytoiden T-Zellen und Interferon-produzierenden Zellen
um die gleiche Zellart, nämlich um pDZ handelt [62]; [63].
PDZ lassen sich anhand ihres Phänotyps leicht charakterisieren: sie tragen
typischerweise CD4, CD123, MHCII und CD45RA an ihrer Oberfläche. Ein für diese
Zellart exklusives C-Typ Lektin ist ‚Blood Dendritic Cell-Antigen’ (BDCA)-2, das eine
wesentliche Rolle bei der Antigenpräsentation einnimmt [64]. Die Blockade von BDCA-
2 führt zur eingeschränkten Typ I IFN-Produktion von pDZ. Ein weiteres, bisher
ebenfalls auf keiner anderen DZ beschriebenes Oberflächenantigen ist BDCA-4
(Neuropilin-1). Bisher konnte keine wesentliche Funktion des BDCA-4 auf pDZ
festgestellt werden, so dass dieses Molekül zur Identifizierung dieser Zellen aus dem
Blut herangezogen wird, ohne dadurch deren Funktion zu beeinflussen [65].
Da pDZ im Menschen in der fetalen Leber, im Thymus und Knochenmark gefunden
wurden, wird derzeit eine Entwicklung der pDZ aus hämatopoetischen Stammzellen
angenommen [66].
Unter physiologischen Bedingungen findet man pDZ im Erwachsenen v.a. in der T-
Zellzone der LK, in der Nähe von ‚high endothelial venules’ (HEV) [67]. Die Bindung an
diese Venolen wird über CD62L (L-Selektin) gesteuert, von dem pDZ reichlich an ihrer
Oberfläche besitzen [63].
Während pDZ in normaler humaner Haut nicht vorhanden sind, wurden sie in
entzündlichen Hauterkrankungen beschrieben [68]. Das bekannteste Beispiel einer
entzündlichen Hauterkrankung mit hohem Anteil von Haut-infiltrierenden pDZ ist der
Lupus erythematodes (LE). Bei der systemischen Verlaufsform dieser Erkrankung sind
pDZ in der Zirkulation deutlich erniedrigt [69], aber eine hohe IFN-α Produktion der
aktivierten pDZ im Gewebe nachweisbar [68].

Einleitung Ch. Bangert
- 18 -
Auch in soliden Tumoren wie Mamma-Karzinomen oder Melanomen konnten pDZ
bereits detektiert werden [70], [71].
Trotz intensiver Forschung ist die volle Funktion der Zellen noch nicht bekannt und
weitere Studien sind notwendig, um die vielen offenen Fragen betreffend Ursprung und
Krankheitsbeteiligung dieser außergewöhnlichen DZ zu beantworten.
1.5.2. T-Zellen im Kontaktekzem
Das allergische KE ist eine T-Zell-mediierte Erkrankung und gehört zu dem Kreis der
Hypersensitivitätsreaktionen vom verzögerten Typ. Inwieweit die Ausprägung des
allergischen KEs durch die Aktivität von T-Zellen vom Typ 1 (T1) oder vom Typ 2 (T2)
bzw. von CD4+ oder CD8+ T-Zellen vermittelt wird, ist derzeit noch größtenteils unklar
[72], [73], [74], [75]. Typisch für eine T1 Antwort ist die Produktion von IFN- γ, während
bei der T2 Antwort die Zytokine IL-4 und IL-5 dominieren.
Immunhistochemische Analysen des zellulären Infiltrats ergaben ein deutliches
Überwiegen der CD4+ gegenüber den CD8+ Zellen [76].
Außerdem zeigten Untersuchungen von zwei unabhängigen Gruppen [73], [74] bei Ni-
Allergikern, dass aus peripherem Blut kultivierte Hapten-spezifische CD4+ Zellen ein
Zytokinmuster wie Typ 1 T-Zellen (IFN-γ und IL-2) aufweisen. Aus diesen Gründen
wurde der CD4+ T-Helfer Zelle lange die Rolle der Effektorzelle zugewiesen.
Untersuchungen im Mausmodell des allergischen KE sowie einige Studien des
humanen KE ergaben jedoch auch einige Hinweise auf eine pathogenetische
Beteiligung der CD8+ T-Zellen [77], [78], [79], [80].
Ein Indiz dafür, dass die Ekzemreaktion zumindest bei Ni-Allergikern nicht
ausschließlich auf die Präsenz von T1-Zellen als Effektorzellen zurückgeführt werden
kann, ist das Vorkommen von Ni-spezifischen CD4+ T-Zellen auch bei nicht-
allergischen Personen. Ni-spezifische CD8+ T-Zell Antworten konnten jedoch nur bei
allergischen Patienten detektiert werden [78], [81]. Überdies produzieren Ni–
spezifische CD4+ Zellklone von Allergikern eine bedeutend geringere Menge des
entzündungshemmenden Zytokins IL-10, als die Klone nicht-allergischer Individuen
[82]. Das könnte bedeuten, dass ein nicht-allergischer Zustand nicht notwendigerweise
durch einen Mangel an Antigen entsteht, wie vordergründig anzunehmen ist, sondern
dass bei Nicht-Allergikern, im Gegensatz zu Allergikern, massive
entzündungshemmende und gegenregulatorische Mechanismen in Kraft treten.
Zudem fanden Werfel et al. dass, anders als im peripheren Blut, die Mehrheit der aus
der inflammierten Haut stammenden CD4+ und CD8+ T-Lymphozyten ein T0 (IL-10)

Einleitung Ch. Bangert
- 19 -
oder ein T2 (IL-4, IL-5 und IL-13) Zytokinmuster aufweisen [83], was ebenfalls als
Argument gegen die T1 Theorie gewertet wird. Tatsächlich verdichteten sich in den
letzten Jahren die Hinweise auf das Vorliegen von entzündungsauslösenden Effektor-
CD8+ T-Zellen und entzündungshemmenden reg CD4+ T-Zellen (Abb 1.5.) [84].
Humane regulatorische T-Zellen (reg T-Zellen) wurden erst vor einigen Jahren und
deutlich später als das murine Pendant entdeckt [85], [86]. Diese Zellen tragen häufig
die Oberflächenmarker CD4 und CD25, wobei diese Antigene nicht einzigartig für reg
T-Zellen sind, sondern auch auf Effektor T-Zellen zu finden sind. Dadurch wird die
phänotypische Charakterisierung der Zellen erschwert und die endgültige
Identifizierung meist mittels funktioneller Versuche vorgenommen. Die am häufigsten
angewendete Methode ist das in vitro Ko-Kultur-Suppressions System, bei dem
während polyklonaler Stimulation potentielle CD4+CD25+ reg T-Zellen zu CD4+CD25-
T-Zellen hinzugefügt werden. Die reg T-Zellen verhindern die Proliferation der
CD4+CD25- T-Zellen und können anhand dessen als solche identifiziert werden [87].
Erst vor wenigen Jahren wurde ein zusätzliches Antigen detektiert, welches von reg T-
Zellen exprimiert wird und die Identifizierung der Zellen deutlich erleichtert: FOXP3,
das zu der ‚Forkhead’ Familie der DNA Transkriptionsfaktoren zählt [88]. FOXP3 wird
spezifisch mit der regulatorisch/suppressiven Aktivität von CD4+ T-Zellen in
Verbindung gebracht [89].
Eine weitere T-Zellpopulation, die im murinen KE nachgewiesen werden konnte, trägt
den γδ T-Zellrezeptor (TZR). Kutane γδ T-Zellen können von Hapten-tragenden
Keratinozyten aktiviert werden [90], wobei diese Aktivierung nicht auf MHC I oder MHC
II beschränkt ist [91]. In Folge kann diese Subpopulation, die vor allem in Haut und
Lymphorganen zu finden ist, zu T-Zellen mit unterschiedlichem Phänotyp ausreifen.
Askenase et al hat den γδ T-Zellen eine Rolle in der Entwicklung des murinen KE
zugeschrieben [92]. Eine andere Studie konnte jedoch zeigen, dass γδ T-Zellen die
Entwicklung von CD8+ Effektor-T-Zellen in der Sensibilisierungsphase hemmen und
somit eher eine entzündungsregulierende Wirkung ausüben [93].
Ob allerdings diese T-Zellen im humanen KE vorhanden sind und welche Funktion sie
innehaben, ist bisher ungeklärt.
1.5.3. Die Sensibilisierungsphase (afferenter Schen kel der Immunantwort)
Bei dem die Sensibilisierung auslösenden Erstkontakt mit dem entsprechenden Hapten
kommt es nach dem Eindringen durch die Hautoberfläche zur Phagozytose des

Einleitung Ch. Bangert
- 20 -
Haptens durch APZ der Haut, wie z.B. epidermale LZ und DDZ. Im Anschluß wandern
diese APZ innerhalb von 24 Std in die regionalen LK und präsentieren
Hapten/Peptidkomplexe den dort ansässigen naiven T-Zellen [94], [95] (Abb. 1.5.).
Während dieses Vorgangs werden APZ von einem ruhenden in einen aktivierten
Zustand versetzt, der vor allem durch die Ausschüttung proinflammatorischer Zytokine
[IL-1β, TNF-α, Granulozyten-Makrophagen Kolonie-stimulierender Faktor (GM-CSF)]
von Keratinozyten und dermalen Fibroblasten bedingt ist. Die Aktivierung der LZ/DDZ
kann zusätzlich auch durch das Haptenmolekül selbst beeinflusst werden, da in vitro
Experimente von DZ, die unterschiedlichen Haptenmolekülen ausgesetzt wurden,
Hinweise auf ein solches Phänomen liefern [96], [97].
Aktivierte DZ produzieren ebenfalls Zytokine (IL-1β, IL-6, IL-12) und es kommt zur
erhöhten Expression von MHCI und MHCII sowie von Kostimulations- und
Adhäsionsmolekülen (z.B. CD80, CD86, CD11b/CD18).
Mausexperimente zeigten, dass die Aktivierung und Migration der LZ in Abhängigkeit
von Haptenmolekülen ausschließlich bei der allergischen Reaktion beobachtet wird,
hingegen nicht bei der toxischen bzw. irritativen Immunantwort [48], [98], [99].
Die MHCI- oder II-restringierte Antigenpräsentation im LK führt schließlich zur
Differenzierung von naiven T-Zellen in Hapten-spezifische Effektorzellen und
gleichzeitig zur klonalen Proliferation. Die aktivierten Hapten-spezifischen T-Zellen
regulieren den Chemokinrezeptor CCR7 auf ihrer Oberfläche hinunter, ein Merkmal,
das die Zellen den peripheren Gedächtnis (Memory) T-Zellen zuordnet. Zu der
Aufgabe der peripheren Memory T-Zellen gehört das ‚Patroullieren’ im peripheren
Gewebe, v.a. der Haut [100].
Bei ihrer Umwandlung zu Effektorzellen erwerben die T-Zellen ein wichtiges
Adhäsionsmolekül, das kutane Lymphozyten-assoziierte Antigen (CLA), welches für
die spätere Migration der Zellen in die Haut verantwortlich ist. Ein weiteres Merkmal
der Haut-infiltrierenden Hapten-spezifischen T-Zellen ist die Expression von CCR4, der
ihnen über die Bindung an seinen von Endothelzellen produzierten Liganden CCL17
‚Thymus and Activation Regulated Chemokine’ (TARC) ermöglicht, am Endothel zu
adhärieren und in Folge über CLA in die Haut einzuwandern. Hierbei werden die zur
Adhäsion benötigten Rezeptoren ‚Very Late Antigen’ (VLA)-4 und ‚Lymphocyte
function-associated antigen’ (LFA)-1 auf T-Zellen hinaufreguliert [101]. Nach etwa 7-10
Tagen ist eine ausreichende Anzahl Effektorzellen gebildet, die immunologisch ein KE
auslösen kann. Die CLA+/CCR4+ T-Zellen verharren in der klinisch unauffälligen Haut
und sind bereit für den nächsten Antigenkontakt.

Einleitung Ch. Bangert
- 21 -
1.5.4. Die Effektorphase (efferenter Schenkel der I mmunantwort)
Nachdem sich die Sensibilisierung ereignet hat, löst ein erneuter Kontakt mit dem
entsprechenden Hapten eine Reihe von Ereignissen aus (Abb. 1.5.), die letztendlich in
dem klinischen Bild eines allergischen KE resultieren.
Das Immunsystem reagiert allergen-spezifisch, mit dem Ziel, das Hapten durch eine
entsprechende Entzündungsreaktion zu eliminieren.
Zu Beginn der Effektorphase produzieren Haut-ansässige Leukozyten wie
Keratinozyten (KZ) oder DZ proinflammatorische Zytokine wie TNF-α und IL-1β [102].
Frühe Ereignisse beinhalten auch Mastzell-Degranulation und Vasodilatation, gefolgt
vom Eindringen der Zellen des angeborenen Immunsystems, wie z.B. von neutrophilen
Granulozyten und Natürlichen Killer (NK)-Zellen. Die Mastzellen scheinen eine Rolle in
der Verstärkung der Immunantwort zu spielen, da sie, aktiviert von CD8+ T-Zellen,
TNF-α und ‚Macrophage Inflammatory Protein’ (MIP-2/CXCL8) ausschütten, die
wiederum für die Rekrutierung von CXCR1- und 2-tragenden Neutrophilen wichtig sind
[103].
Im nächsten Schritt kommt es zusätzlich zur schnellen Einwanderung von
CCR4+/CLA+ aktivierten Hapten-spezifischen T-Lymphozyten sowie von unspezifisch
rekrutierten Lymphozyten in das betroffene Hautareal [83], [104].
Hapten-spezifische T-Zellen stellen lediglich etwa 1% der gesamten Population
infiltrierender T-Zellen dar, trotzdem ist die Anzahl dieser in der Haut akkumulierenden,
spezifischen T-Lymphozyten 10-100 mal höher als deren Gesamtmenge im
Blutkreislauf zu diesem Zeitpunkt.
Sowohl Hapten-spezifische als auch nicht-spezifische T-Zellen produzieren Zytokine,
sobald sie den Ort der Entzündung erreichen. TNF-β übt einen zytotoxischen Effekt auf
das Gewebe aus und induziert die Expression des Adhäsionsmoleküls ‚Intercellular
Adhesion Molecule’ (ICAM)-1 (CD54) auf Gefäßendothelzellen und basalen
Keratinozyten innerhalb von 48 Std nach Haptenkontakt. Das wiederum verstärkt die
Bindung der T-Zellen über LFA-1 an ICAM-1, und sie gelangen dadurch nicht nur in die
Haut, sondern auch von der Dermis in die Epidermis [105].
CD8+ zytotoxische T-Zellen spielen in der Effektorphase des KE eine wichtige Rolle:
neben der Produktion von Interferon (IFN)-γ wird die Entzündung durch deren
Zytotoxizität anhand von FAS/FASL sowie Perforin/Granzyme gesteuert [77] (Abb
1.5.). Die Ziele dieser zytotoxischen Angriffe sind überwiegend Keratinozyten, die

Einleitung Ch. Bangert
- 22 -
dadurch in Apoptose gehen [106]; [107], [108]. Das erleichtert dem Hapten das
Eindringen in die Epidermis, was wiederum die Entzündung verstärkt [109]. Durch IFN-
γ wird eine Vielzahl anderer Zytokine von hautansässigen und durch die Haut
zirkulierenden Leukozyten gebildet, wie ‚IFN-γ Inducible Protein’ (IP)-10 (CXCL-10),
‚Monokine-Induced by IFN-γ’ (MIG/CXCL-9), IL-1, IL-6, GM-CSF und CXCL-8. Zytokine
wie GM-CSF aktivieren Makrophagen/Monozyten und DZ und erhöhen dadurch die
Ausschüttung weiterer Entzündungsmediatoren, die erneut Proliferation und
Aktivierung der T-Zellen verstärken.
Die Effektorphase hat nach 24-72 Std ihren Höhepunkt, danach kommt es zur
Regulierung der Immunantwort. Verantwortlich für die ‚Gegenregulation’ des
Immunsystems könnten die sogenannten CD4+CD25+ regulatorischen T-Zellen (reg T-
Zellen) sein, die über einen bislang noch nicht vollständig geklärten Mechanismus die
weitere Expansion der CD8+ T-Zellen blockieren [110], [111], [112] (Abb 1.5.).
Einleitung Ch. Bangert
- 23 -
Dermal dendritic cells
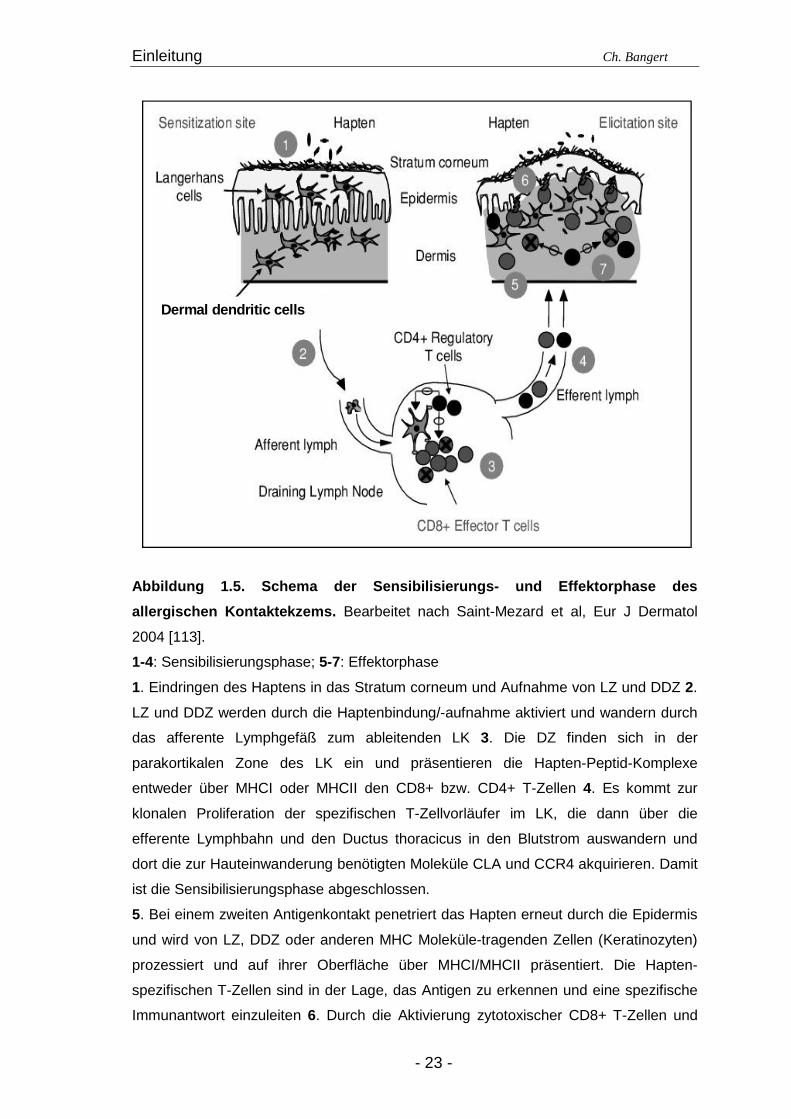
Abbildung 1.5. Schema der Sensibilisierungs- und Ef fektorphase des
allergischen Kontaktekzems. Bearbeitet nach Saint-Mezard et al, Eur J Dermatol
2004 [113].
1-4: Sensibilisierungsphase; 5-7: Effektorphase
1. Eindringen des Haptens in das Stratum corneum und Aufnahme von LZ und DDZ 2.
LZ und DDZ werden durch die Haptenbindung/-aufnahme aktiviert und wandern durch
das afferente Lymphgefäß zum ableitenden LK 3. Die DZ finden sich in der
parakortikalen Zone des LK ein und präsentieren die Hapten-Peptid-Komplexe
entweder über MHCI oder MHCII den CD8+ bzw. CD4+ T-Zellen 4. Es kommt zur
klonalen Proliferation der spezifischen T-Zellvorläufer im LK, die dann über die
efferente Lymphbahn und den Ductus thoracicus in den Blutstrom auswandern und
dort die zur Hauteinwanderung benötigten Moleküle CLA und CCR4 akquirieren. Damit
ist die Sensibilisierungsphase abgeschlossen.
5. Bei einem zweiten Antigenkontakt penetriert das Hapten erneut durch die Epidermis
und wird von LZ, DDZ oder anderen MHC Moleküle-tragenden Zellen (Keratinozyten)
prozessiert und auf ihrer Oberfläche über MHCI/MHCII präsentiert. Die Hapten-
spezifischen T-Zellen sind in der Lage, das Antigen zu erkennen und eine spezifische
Immunantwort einzuleiten 6. Durch die Aktivierung zytotoxischer CD8+ T-Zellen und

Einleitung Ch. Bangert
- 24 -
die Apoptose von Keratinozyten wird die Entzündungsreaktion eingeleitet, die durch die
Ausschüttung proinflammatorischer Zytokine und Chemokine zustande kommt 7. Die
Ausschüttung dieser Chemokine ist für die anschließende Rekrutierung von
zusätzlichen T-Zellen (einschließlich reg T-Zellen) verantwortlich. Die Klinik zeigt nach
48-72 Std die maximale Ausprägung des KEs.

Zielsetzung Ch. Bangert
- 25 -
2. ZIELSETZUNG
Bisher wurde nur lückenhaft untersucht, welche Zellen tatsächlich an der Effektorphase
des allergischen KE beteiligt sind. Das Auslösen dieser Phase erfordert vermutlich
intensiven Kontakt zwischen T-Zellen und APZ, die sowohl in der Haut residieren als
auch vom Blut eingewandert sein können. Um welche APZ es sich jedoch handelt, die
eine Hapten-spezifische Antwort auslöst, ist bislang unklar. Obwohl LZ häufig als
zentrale Verstärker der Effektorphase angesehen werden [52], zeigen andere
Ergebnisse, dass eine Depletion dieser Zellen nach der Sensibilisierungsphase eher
eine verstärkte als eine abgeschwächte Immunantwort bei erneuter Hapten-
Provokation zur Folge hat [50].
Auch ist nach wie vor unbekannt, weswegen die Erkrankung nur bei bestimmten
Personen und nicht (wie das toxische KE) bei allen Menschen auftritt. Neben dem
Mangel an Sensibilisierung wird bei Nicht-Allergikern vor allem das Vorkommen von
Hapten-spezifischen regulatorischen T-Zellen diskutiert, die eine allergische Reaktion
unterdrücken können [78]. Bislang ist nicht restlos geklärt, ob CD4+ oder CD8+ T-
Zellen die Immunantwort der Effektorphase auslösen [78], [79], [81], [83], [114], [115].
Diese gegensätzlichen Ergebnisse hinsichtlich der Natur der unmittelbar involvierten
Zellen in der Effektorphase weisen daraufhin, daß das allergische KE ein wesentlich
komplexeres Ereignis darstellt als bisher angenommen. Um einen umfassenden
Überblick über das Geschehen in dieser Erkrankung zu erhalten, haben wir in der
vorliegenden Studie das leukozytäre Infiltrat von ECT zu unterschiedlichen Zeitpunkten
phänotypisch analysiert und richten unser Augenmerk besonders auf APZ und T-
Zellen.
Zusätzlich wird der Einfluß von Ni und Duftstoffen auf die Qualität des entzündlichen
Infiltrats bestimmt. Im Besonderen interessiert uns, ob phänotypische Unterschiede
hinsichtlich der Verteilung von Effektor-, reg T-Zell-Populationen und/oder APZ durch
diese chemisch differenten Haptene induziert werden können.
Durch die erhaltenen Informationen über Kinetik, Ausmaß und Art der entzündlichen
Immunantwort soll ein weiterer wichtiger Grundstein für das Verständnis der
Immunpathologie im humanen KE gelegt und dadurch eine gezielte Forschung zur
Aufklärung der Immunpathogenese erleichtert werden.

Methoden Ch. Bangert
- 26 -
3. MATERIAL UND METHODEN 3.1 Material 3.1.1 Patienten Nach Anerkennung des Studienprotokolls durch die Medizinische Universität Wien und
Zustimmung der lokalen Ethikkommission wurde die Studie nach den Richtlinien der
Deklaration von Helsinki durchgeführt. Alle Patienten hatten zuvor ihr schriftliches
Einverständnis zur Studienteilnahme gegeben.
5 Patienten mit positivem ECT für Ni, 5 Patienten mit positivem ECT für Duftstoff sowie
5 gesunde Kontrollpersonen wurden für die Analyse des zellulären Infiltrats läsionaler
Haut in die Studie aufgenommen (Tab. 3.1.1.). Weitere 3 Patienten mit positivem ECT
für Ni wurden zur Analyse der Infiltrationskinetik von Leukozyten in die Haut
eingeschlossen.
5 Patienten mit positivem ECT für mindestens 3 verschiedene Haptene wurden
zusätzlich für die durchflußzytometrische Charakterisierung von pDZ in läsionaler Haut
und peripherem Blut aufgenommen.
Folgende Ausschlusskriterien wurden vor Einschluß der Patienten beachtet:
1) Orale Verabreichung von systemischen Steroiden oder
Immunsuppressiva bis 2 Wochen vor der Gewebsentnahme
2) PUVA, UVB Bestrahlung bis 4 Wochen vor der Gewebsentnahme
3) Applikation von kortikosteroidhaltigen Externa bis 2 Wochen vor
Entnahme der Gewebsprobe
4) Immunsuppression aufgrund einer Erkrankung (z.B. HIV-Infektion) oder
medikamentös induziert (bei onkologischen Patienten oder im Zuge einer
Transplantation)
5) Atopische Disposition

Methoden Ch. Bangert
- 27 -
Tabelle 3.1.1. Probandenübersicht
Probanden Alter Geschlecht Allergen Entzündungs- Nummer (Jahre) grad
1 47 W Nickel 2 2 43 W Nickel 3 3 44 W Nickel 1 4 48 W Nickel 2 5 46 W Nickel 2 6 58 M Duftstoff Mix 3 7 76 M Duftstoff Mix 2 8 48 W Duftstoff Mix 1 9 22 W Duftstoff Mix 2 10 27 W Duftstoff Mix 1
Probanden Alter Geschlecht Nummer (Jahre)
1 61 W 2 55 M 3 47 W 4 38 W 5 53 M
Normalhaut (Kontrollgruppe)
Epikutantestungen (72 Std) Std)

Methoden Ch. Bangert
- 28 -
3.1.2 Biopsien
Die Haptene (Ni bzw. Duftstoff) wurden auf ein Areal am Rücken der Patienten mittels
standardisiertem ECT für 48 Std aufgetragen. Nach 48 sowie 72 Std wurde die
allergische Reaktion nach den Kriterien der ‚International Contact Dermatitis Research
Group’ beurteilt und entsprechend dem Status der Entzündung von Stufe 1 bis 3
eingeteilt, wobei Stufe 1 die am wenigsten positive Reaktion klassifizierte. Zur
Charakterisierung des zellulären Hautinfiltrats wurde den Patienten 72 Std nach Beginn
der Haptenauftragung eine 4 mm im Durchmesser große Stanzbiopsie entnommen.
Um die Infiltrationskinetik der Leukozyten in der allergischen Reaktion zu beurteilen,
wurden bei 3 Patienten zum Zeitpunkt 0, nach 24, 48 und 72 Std eine 4 mm große
Probeexcision durchgeführt. Bei 2 Patienten wurde zusätzlich eine Biopsie nach 6 Std
entnommen. Die Analyse mittels Durchflußzytometrie erforderte die Exzision von 3x6
mm großen Stanzbiopsien bei 5 Patienten.
Allen Patienten wurde vor der Entnahme subkutan ein Lokalanästhetikum in das zu
biopsierende Hautareal injiziert, um den Eingriff möglichst schmerzfrei zu gestalten.
Eine Allergie auf Lokalanästhetika wurde zuvor anamnestisch ausgeschlossen. Nach
Entnahme der Biopsien wurde die Wunde mit 1-3 Stichen vernäht. Den Patienten
wurde geraten, schwere körperliche Belastungen sowie sportliche Aktivitäten für 14
Tage zu unterlassen und die Wunde trocken und sauber zu halten. Nach 2 Wochen
wurden die Nähte entfernt. Alle Patienten haben die Entnahme ohne Nebenwirkungen
oder Komplikationen vertragen.
3.1.3 Blutproben
Zur durchflußzytometrischen Analyse der pDZ im peripheren Blut wurde 5 Patienten
mit positiven ECT 100ml Vollblut in heparinisierten Röhrchen entnommen. Die
Blutprobe wurde parallel zur Entnahme der Hautbiopsien vorgenommen. Bei allen
Patienten lag zum Zeitpunkt der Blutentnahme keine akute infektiöse Erkrankung vor.

Methoden Ch. Bangert
- 29 -
3.1.4. Antikörper
Antikörper für immunhistochemische Analysen
Antigen Klon Firma
CD1a M2120; mIgG2b PeliCluster; CLB, Niederlande
CD1c M241;mIgG1 Ancell, Bayport, MN, USA
CD3 UCHT1; mIgG1 DAKO, Glostrup, Dänemark
CD4 SK 3; mIgG1 Becton Dickinson (BD)
Biosciences, San Jose, CA, US
CD8 C8/144B; mIgG1 DAKO
CD14 MФP9; mIgG2b BD
CD15 VIM16; rIgM Otto Majdic, Immunologie, AKH,
Wien, Österreich
CD25 2A3; mIgG1 BD
CD45RA L48; mIgG1 BD
CD45 HLE-2; mIgG1 BD
CD56 biot My31; mIgG1 BD
CD62L SK11; mIgG2a BD
CD83 HB15e; mIgG1 BD PharMingen, San Jose, CA,
USA
CD94 HP-3D9; mIgG1 BD PharMingen
CD117 95C3; mIgG1 An der Grub GmbH, Österreich
CDw123 biot 9F5; mIgG1 BD PharMingen
CxCR3 49801.111; mIgG1 R&DSystems GmbH,
Biomedica, Wien
ECP EG1; mIgG1 Kabi Pharmacia Diagnostics,
Schweden
FcεRIα-chain 15.1; mIgG1 (Ref. Wang et al, J Exp Med
1992)
Foxp3 biot 236A/E7, mIgG1 eBioscience, San Diego, CA,
USA

Methoden Ch. Bangert
- 30 -
Antigen Klon Firma
γδ -TCR 5.A6.E9; mIgG1 Endogen, Woburn, MA, USA
Ki-67 Ki-S5; mIgG1 DAKO
Langerin (CD207) DCGM4; mIgG1 Immunotech, Coulter, Marseille,
Frankreich
PNAd MECA-79; rIgM BD PharMingen
Sekundärantikörper
Bezeichnung Firma
Streptavidin TRITC Jackson Immuno Research
Laboratories, West Baltimore,
Pennsylvania, USA
Streptavidin Alexa Fluor 488 Molecular Probes, Eugene, OR,
USA
Streptavidin Cy5 Jackson Immuno Research
Laboratories
Rhodamine (TRITC) goat-anti-mouse IgG Jackson Immuno Research
Laboratories
Alexa Fluor 488 goat anti-mouse IgG Molecular Probes
Oregon Green labelled goat anti-FITC Molecular Probes
Antikörper für die Durchflußzytometrie
Antigen Klon Firma
BDCA-2 FITC AC144; mIgG1 Miltenyi Biotech GmbH,
Bergisch-Gladbach, Deutschland
CD4 FITC RPA-T4; mIgG1 BD PharMingen
CD4 PerCP RPA-T4; mIgG1 BD
CD3 FITC SK-7; mIgG1 BD

Methoden Ch. Bangert
- 31 -
CD3 PerCP SK7; mIgG1 BD
CD25 FITC M-A251; mIgG1 BD PharMingen
CD45RA FITC ALB11; mIgG1 Immunotech
CD62L FITC Dreg 56; mIgG1 BD PharMingen
CD83 FITC HB15a; mIgG2b Immunotech
CD86 FITC HA5.2B7; mIgG2b Immunotech
CD123 PE 9F5, mIgG1 BD PharMingen
CLA FITC HECA-452; rIgM BD PharMingen
CTLA-4 PE BNI3; mIgG2a BD PharMingen
MHCII FITC L243; mIgG2a BD
MHCII PerCP L243; mIgG2a BD
3.1.5. Chemikalien
Chemikalie Abkürzung Firma
3-Amino-9-Äthyl-Carbazol AEC Sigma-Aldrich St.Louis,
MO, USA
Acetonum purum Fluka, Buchs, Schweiz
Aethanol rein, 96% Anstaltsapotheke, AKH,
Wien, Österreich
Aethanol 70% Anstaltsapotheke, AKH
Aquamount Eindeckmedium BDH Laboratories Supplies,
Poole, UK
Bovines Serum Albumin BSA Sigma-Aldrich
Ficoll-Hypaque Amersham Biosciences AB,
Uppsala, Schweden
Humanes Immunglobulin Octagam Octapharma
Pharmazeutika, Wien,
Österreich
Hämatoxylin Merck
Normales Mausserum NMS DAKO
Nickelsulfat NiSO4 Sigma-Aldrich
PBS GIBCO, Invitrogen GmbH,
Lofer, Österreich
PBS (x10) GIBCO
RPMI 1640 GIBCO

Methoden Ch. Bangert
- 32 -
Tissue-Tek Einbettmedium OCT Sakura Finetek Europe
B.V., Zoeterwoude,
Niederlande
Trypanblau Sigma-Aldrich
Wasser H2O Anstaltsapotheke, AKH,
Wien
Wasserstoffperoxid 30% H2O2 MERCK-Schuchardt,
Hohenbrunn, Deutschland
Vectashield Mounting Medium Vector Laboratories,
Burlingame, CA, USA
Vectastain Elite ABC Kit Vector Laboratories
(Mouse anti-human)
3.1.6. Verbrauchsmaterial
15ml, 50ml Röhrchen Fa. Greiner, Frickenhausen,
Deutschland
Deckgläser 25x40mm Fa.Menzel-Gläser,
Braunschweig, Deutschland
Einbettschälchen ‚Cryomold Standard’ Sakura Finetek Europe B.V.
(25mmx20mmx5mm)
Objektträger ‚ChemMateTM Capillary Gap DAKO
Microscope Slides (100µm)’
PS (Polystyrol)-Röhrchen (0,6ml) Fa. Greiner
Sterilfilter (EasyflowTM) BD Falcon, BD Biosciences
Zellsieb 40µm, steril BD Falcon, BD Biosciences
Steriles Einmalskalpell Größe 10 Kehr Surgical Private
Limited, Indien
Sterile Spritze, 2ml Chirana T.injecta, Slowakei

Methoden Ch. Bangert
- 33 -
3.1.7. Geräte
Durchflusszytometer (FACScalibur) BD Biosciences
Durchlichtmikroskop Standard BHSU Olympus Life and Material Science
Europa GmbH, Hamburg,
Deutschland
Fluoreszenzmikroskop Axiophot Zeiss, Oberkochen, Deutschland
Laser Scanning Microscope (LSM) 510 Zeiss
Sterilbank (Lamina Air Flow) Fa. Heraeus, Osterode,
Deutschland
Zentrifuge (J-6B) Fa. Beckman, München,
Deutschland
Zentrifuge (Omnifuge 2.0 RS) Fa. Heraeus, Osterode
Zentrifuge (Biofuge pico) Fa. Heraeus, Osterode
Innova 4000, Incubator shaker P. Haack Laborausrüstung,
Wien, Österreich
3.1.8. Puffer
PBS-BSA-Puffer 2% Rinderserum-Albumin (BSA)
in PBS-Puffer

Methoden Ch. Bangert
- 34 -
3.2 Methoden
3.2.1. Anfertigung von Gefrierschnitten
Die von den Patienten entnommenen Biopsien werden direkt nach der Entnahme bis
zum Zeitpunkt des Einbettens in RPMI 1640 Medium aufbewahrt. Um die Hautproben
möglichst trocken einzubetten, werden mit einer Kompresse zunächst vorsichtig Blut
und RPMI entfernt. Im Anschluß werden die Biopsien in eine mit Tissue Tek
Einbettmedium gefüllte Kammer gebettet und mit flüssigem Stickstoff tiefgefroren. Bis
zur weiteren Verarbeitung werden sie bei –80°C gela gert.
Zum Anfertigen der Gefrierschnitte werden die Biopsien für etwa 30 Minuten bei -20°C
aufbewahrt und im Anschluß in 5µm dicke Schnitte verarbeitet. Die Schnitte werden
auf beschichtete Objektträger aufgetragen und für 20 Minuten luftgetrocknet. Danach
werden die Schnitte in eiskaltem Aceton bei 4°C für 10 Minuten fixiert und bis zur
weiteren Verarbeitung bei -20°C aufbewahrt.
3.2.2. Immunhistochemie
Zur Anfertigung von immunhistochemischen Färbungen werden die gefrorenen
Schnitte zunächst beschriftet und in PBS für ca. 10 Minuten aufgetaut. Zwischenzeitlich
werden die unmarkierten Antikörper in der jeweils entsprechenden Konzentration in 2%
BSA-PBS Puffer verdünnt. Zur Blockade unspezifischer Bindung des Zweitantikörpers
mischt man zusätzlich 1% normales Pferdeserum in den Puffer. Die gemischte Lösung
wird auf die Schnitte pipettiert und für 1 Stunde bei Raumtemperatur oder über Nacht
auf 4°C in einer feuchten Färbekammer inkubiert.
Nach der Inkubationszeit werden die Schnitte für 5 Minuten in 1xPBS gewaschen und
im Anschluß für 15 Minuten in 1% H202 zur Neutralisierung der endogenen Peroxidasen
aufbewahrt. Nach erneutem Waschen mit PBS werden 0,5% des Zweitantikörpers für
45 Minuten in einer feuchten Kammer aufgebracht.
Vor dem dritten und letzten Inkubationsschritt werden die Schnitte wiederum 3x5
Minuten in PBS gewaschen. Der 1%ige Avidin-Biotin-Komplex mit integrierter
Peroxidase wird für 30 Minuten bei Raumtemperatur auf den Schnitten in der feuchten
Kammer belassen, und die Schnitte werden im Anschluß mit AEC Lösung entwickelt.
Zellen, die den Antikörper binden, färben sich rot. Die Gegenfärbung wird mit
Hämatoxylin durchgeführt.

Methoden Ch. Bangert
- 35 -
3.2.3. Immunfluoreszenz
Die Liste der verwendeten Antikörper ist unter 3.1.4. aufgeführt.
Die Einzelfärbungen werden folgendermaßen vorgenommen:
Unmarkierte oder biotinylierte Antikörper werden in 2% BSA-PBS in der gewünschten
Konzentration verdünnt und die Schnitte damit über Nacht auf 4°C in einer feuchten
Kammer inkubiert. Nach dem Waschen mit PBS wird der Erstantikörper mittels
Zweitantikörper sichtbar gemacht und für 45 Minuten aufgebracht. Als
Sekundärantikörper wird je nach Färbekombination entweder Alexa 488- oder
Rhodamin-goat anti-mouse IgG, Streptavidin Alexa 488 oder Streptavidin TRITC
verwendet. Nach der Inkubation werden die Schnitte gewaschen, in 96% Äthanol
plaziert und direkt mit ‚Vectashield Mounting Medium’ eingedeckt.
Um das Zellinfiltrat besser analysieren zu können, sind auch Zwei- oder
Dreifachfärbungen notwendig.
Der erste Schritt der Doppelfärbungen erfolgt wie oben beschrieben. Die Schnitte
werden nach der Inkubation mit dem Sekundärantikörper jedoch nicht eingedeckt,
sondern für 20 Minuten mit 1-3% normalem Mausserum inkubiert, um freie
Bindungsstellen zu blockieren und somit den unspezifischen Hintergrund der Färbung
zu reduzieren. Danach wird der zweite Antikörper, diesmal mit Fluorescein (FITC)
direkt markiert oder biotinyliert, für mindestens 6 Std bei 4°C in der feuchten Kammer
aufgetragen.
Da FITC-markierte Antikörper dazu neigen, schnell zu verblassen, wird ein weiterer
Färbeschritt mit einem Oregon Green anti-FITC Antikörper angeschlossen (30 Minuten
bei Raumtemperatur). Biotinylierte Antikörper können mit Streptavidin TRITC für 45
Minuten entwickelt werden.
Für Dreifachfärbungen wird ein unmarkierter Antikörper mit TRITC ‚goat-anti-mouse
IgG’ entwickelt, die Schnitte mit normalem Mausserum inkubiert, ein direkt FITC-
markierter als auch ein biotinylierter Antikörper für 60 Minuten bei Raumtemperatur
aufgetragen und die Schnitte im Anschluß gleichzeitig mit Streptavidin Cy5 und Oregon
Green anti-FITC für 45 Minuten bei Raumtemperatur inkubiert.
In allen Färbeexperimenten wird analog eine Isotypkontrolle durchgeführt, die mit den
gewünschten Antikörpern hinsichtlich Konzentration und Typ übereinstimmt.

Methoden Ch. Bangert
- 36 -
3.2.4. Auswertung der Immunfluoreszenz
Die Schnitte werden von 2 unabhängigen Untersuchern an einem Fluoreszenz
Mikroskop in 400x Vergrößerung verblindet ausgezählt. Das Fluoreszenz Mikroskop ist
mit einem Doppelfilter für TRITC (Rot) und FITC (Grün) ausgestattet, der doppelt
gefärbte Zellen in einer gelben Farbe erscheinen lässt und somit klar erkennbar macht.
Die infiltrierenden Leukozyten werden für Epidermis und Dermis getrennt ausgezählt.
Die markierten Zellen werden pro Gesichtsfeld (GF) ausgezählt und als Anzahl pro mm
Basalmembran Epidermis oder pro mm2 Dermis angegeben.
Um die Anzahl der Zellen auf mm Epidermis umrechnen zu können, wird zunächst der
Durchmesser des verwendeten GF errechnet.
Hier gilt, dass die Sehfeldzahl durch die Maßstabszahl des verwendeten Objektivs
geteilt wird, um den Durchmesser des GF zu erhalten. Die Sehfeldzahl des
verwendeten Okulars 10x/25 des Fluoreszenzmikroskops Axiophot der Firma Zeiss
beträgt 25. Die Maßstabzahl des Objektivs ist 40, so dass der Durchmesser 0,625mm
beträgt (Durchmesser des GF= 25/40 (Sehfeldzahl/Maßstabzahl)=0,625mm)). Die
Länge der GF wurde daraufhin auf 1mm berechnet, um die gesuchte Zellzahl pro mm
Epidermis zu erhalten (z.B. 3 GF= 3*0,625mm=1,875mm. 300 Zellen/1,875mm=160
Zellen/mm)
Um die Anzahl der Zellen/mm2 Dermis zu errechnen, wird zunächst die Fläche des
Kreises bzw. des GF ermittelt (Kreisfläche=r2*π d.h. Fläche des
GF=(0,3125mm)2*π=0,097656mm2*3,141593=0,306796mm2).
Die gerundete Zahl und somit das Ergebnis der Fläche des Gesichtsfelds betragen
0,31mm2. Die Anzahl der Zellen pro GF wird schließlich mit 3,2258 multipliziert, um das
Ergebnis pro mm2 Dermis angeben zu können.
Nur Zellen, die deutlich positiv sind, werden in die Wertung aufgenommen.
Statistische Signifikanz wird mit dem Students T-Test bestimmt.
Um Dreifachfärbungen visuell dokumentieren zu können, wird konfokale
Lasermikroskopie (LSM510) angewendet.

Methoden Ch. Bangert
- 37 -
3.2.5. Gewinnung von mononukleären Zellen aus dem p eripheren Blut (PBMC)
Die Isolierung von mononukleären Leukozyten aus Patientenblut erfolgt über eine
Dichtegradientenzentrifugation. Heparinisiertes Vollblut wird zunächst im Verhältnis 1:2
mit PBS-Puffer verdünnt und dann mit 10ml Ficoll-Trennlösung, Dichte 1,077, in 50ml-
Röhrchen unterschichtet.
Nach 30min Zentrifugation bei 1600 rpm (550g) wird die Interphase, die die
mononukleären Zellen als Ring über der Ficoll-Lösung enthält, mittels einer
Einmalpipette abgenommen. Die so isolierte Fraktion wird in 50ml PBS-Puffer bei 1600
rpm gewaschen und anschließend mit 50ml PBS-Puffer bei geringerer Drehzahl (1200
rpm) zentrifugiert, um die leichteren Thrombozyten entfernen zu können. Abschließend
werden die verbleibenden mononukleären Zellen in PBS-Puffer resuspendiert und
gezählt.
3.2.6. Herstellung von Einzelzellsuspensionen aus d er Haut
Für die Einzelzellsuspensionen der Haut werden 3x4mm Biopsien von 5 verschiedenen
Patienten mit positivem ECT auf mindestens 3 verschiedene Haptene entnommen. Die
Biopsien werden mittels sterilem Sklapell in winzige Stückchen zerteilt und danach in
50ml Röhrchen, gefüllt mit RPMI 1640 Medium, für 2 Std in einem Zellschüttler bei
37°C inkubiert. Aufgrund der dadurch erzielten Lock erung des Gewebes wird die
Migration der Zellen in das Medium unterstützt.
Die kleinen Biopsiestückchen werden im Anschluß mitsamt Medium in ein Zellsieb mit
der Größe 40µm geschüttet, das auf 50ml Röhrchen aufgesetzt wird. Die Haut wird
dann mit dem Stempel einer 2ml Plastikspritze zerrieben und die Zellen so aus dem
Gewebe in eine Suspension überführt. Das verbleibende Gewebe, hauptsächlich
bestehend aus Kollagenfasern, wird entsorgt.
Die Zellsuspension wird im Anschluß gewaschen, zentrifugiert und die Zellen gezählt.
3.2.7. Zellzählung in Suspension
20µl der Zellsuspension werden mit 20µl Trypanblau zur Vitalitätsbestimmung verdünnt
und im Anschluß ausgezählt. Zellen, die sich durch Trypanblau verfärbt haben,
verfügen über keine intakte Zellwand mehr und werden nicht mitgezählt.
Die gesamte Suspension wird in eine Neubauer-Zählkammer (Kammertiefe 0,1mm)
pipettiert. Unter dem Mikroskop werden alle Zellen in den vorhandenen 20 Quadraten
(Gesamtfläche: 1mm2) ausgezählt und das Zählergebnis mit dem Verdünnungsfaktor

Methoden Ch. Bangert
- 38 -
sowie dem Faktor 104 für das Kammervolumen/ml multipliziert (siehe Formel). Die
gesamte Zellzahl ergibt sich aus der Multiplikation mit dem Suspensionsvolumen.
Ausgezählte Zellen *2 (Verdünnung) * 104 (Faktor für Kammervolumen) * x (ml
Suspension) = Gesamtzellzahl
3.2.8. Durchflusszytometrie
3.2.8.1. Durchführung und Darstellung
Die Durchflußzytometrie bzw. englisch: FACS-Analyse („fluorescent activated cell
sorting“) dient zur Identifizierung von zellspezifischen Antigenen. Hierfür werden die
Zellen über ein Schlauchsystem aufgesaugt und mit Druck in einen sogenannten
Mantelstrahl (Sheath) injiziert. Die Zellen werden so beschleunigt und hintereinander
aufgereiht, dass sie einzeln in die Meßkammer gelangen. Dort wird jede Zelle vom
Laserstrahl getroffen. Das gestreute Licht wird bei 180° (FSC = forward light scatter)
und 90° (SSC = sideward light scatter) gemessen. Di e entstehende Streuung ist
zelltypspezifisch und abhängig von der Größe der Zellen (FSC) sowie deren
Granularität (SSC). So entstehen in der Dot Plot Grafik abgegrenzte Punktwolken für
Granulozyten, Monozyten, Lymphozyten oder auch Erythrozyten.
Bei der Fluoreszenzanalyse mittels antigenspezifischer Antikörper werden die
einzelnen Zellen durch einen Laser angeregt und an elektronischen Detektoren
vorbeigeführt, die die Fluoreszenzintensität der Zelle registrieren. Die jeweils zu
analysierenden Zellpopulationen können über ein „Gate“ eingegrenzt und somit von
den anderen Zelltypen für die Analyse getrennt werden.
3.2.8.2. Phänotypische Analysen von Oberflächenmark ern
Zur Analyse der Proteinexpression auf der Zelloberfläche werden die Zellen zunächst
mit spezifischen Antikörpern gegen das zu untersuchende Antigen inkubiert. Die
verwendeten Antikörper sind unter 3.1.4. aufgeführt. Die Analysen werden mit FITC-,
PE- oder PerCP-konjugierten Antikörpern als direkte Dreifarbenfluoreszenz
durchgeführt.
Pro Ansatz werden etwa 3x105 aus dem Blut oder aus der Haut isolierte Zellen in etwa
50µl PBS pipettiert. Zur Blockade unspezifischer Bindungen der Antikörper an Fcγ-
Rezeptoren der Zellen werden pro Ansatz 100µg/ml humanes Immunglobulin
zugegeben. Anschließend werden die mit Fluoreszenzfarbstoffen konjugierten
Antikörper in der vom Hersteller empfohlenen Dosierung hinzugefügt. Die Inkubation

Methoden Ch. Bangert
- 39 -
erfolgt daraufhin für 15min auf Eis. Nach dreimaligem Waschen mit je 150µl PBS
werden die Zellen im FACS analysiert.
Je Ansatz werden 10.000 eingegrenzte Zellen mit Hilfe des Computerprogramms
‚CellQuest®’ ausgewertet. Um bei der Analyse von pDZ aus peripheren
Blutlymphozyten die T-Zellen auszugrenzen, wird ein für DZ spezifisches ‚Gate’
gesetzt. Ebenso werden zur besseren Analyse der reg T-Zellen aus der Haut die DZ
anhand ihrer Größe und Granularität mit einem ‚Gate’ ausgeschlossen.
Die Autofluoreszenz wird durch Negativkontrollen mit dem entsprechenden Isotyp
bestimmt. Die prozentuale Verteilung negativer und positiver Zellen wird anschließend
im Dot Plot ermittelt.
Die Expressionsdichte des entsprechenden Antigens wird durch die Höhe der
Fluoreszenzintensität (MFI = mittlere Fluoreszenzintensität) repräsentiert. Bei der
Auswertung mit der CellQuest® Software wird der Dot Plot in Quadranten unterteilt
sowie die Verteilung der Zellen und die MFI des jeweiligen Quadranten ermittelt. Die
jeweilige Prozentzahl der positiv gefärbten Zellen wird im rechten oberen Quadranten
der Dot Plots angegeben.

Ergebnisse Ch. Bangert
- 40 -
4. ERGEBNISSE
4.1. Anzahl und Verteilung der Leukozyten
In dieser Studie haben wir versucht, einen detaillierten Überblick über die
verschiedenen Zellarten zu geben, welche maßgeblich an der ekzematösen Reaktion
in der Effektorphase des humanen KE beteiligt sind. Es wurden Biopsien sowohl aus
positiven 72 Std ECT (n=10) als auch von Normalhaut (NH) (n=5) entnommen, und wir
führten mittels Immunfluoreszenz eine ausführliche Analyse der infiltrierenden
Zellpopulationen durch. Die Resultate sind in der Tabelle 4.1. zusammen gefasst. Die
Färbung der NH mit dem Pan-Leukozyten Marker CD45 zeigte nur wenig positive
Zellen in Epidermis und Dermis. Die genaue Analyse der gesunden Biopsien ergab,
dass diese CD45+ Zellen in der Epidermis hauptsächlich aus CD1a+ DZ und in der
Dermis vor allem aus CD3+ T-Zellen, CD14+ Makrophagen, CD1c+ DZ und CD117+
(c-kit) Mastzellen bestehen.
Im Gegensatz dazu fanden wir in den ECT ein massives leukozytäres CD45+ Infiltrat in
beiden Hautschichten. Verglichen mit der NH war die Anzahl der Zellen beinahe
doppelt so hoch in der Epidermis und verfünffacht in der Dermis. Zu einem großen Teil
bestand das Infiltrat aus T-Zellen oder Mitgliedern der DZ Familie. Es fand sich aber
auch ein beträchtlicher Anstieg der Zellzahl von Mastzellen, CD15+ neutrophilen und
EG1+ eosinophilen Granulozyten und Makrophagen. Ebenso stieg auch die Anzahl
von CD56+ oder CD94+ NK-Zellen.

Ergebnisse Ch. Bangert
- 41 -
Tabelle 4.1. Anzahl der Leukozyten in NH und 72 Std ECT Läsionena
Zellart Antigen (e) Epidermis (positive Zellen/mm) Dermis (positive Zellen/mm 2)
NH 72 Std ECT NH 72 Std ECT
Leukozyten CD45+ 14,1 (12,1-17,7) 30,8 (21,5-50,6) 86 (64,7-97,6) 435,3 (240,6-858)
T-Zellen CD3+ 2,7 (0-3,5) 20,9 (3,7-43,2) 39,5 (2,4-58,4) 513,1 (109,3-896)
DN T-Zellen b CD3+/CD4-/CD8- 0 (0) 0,4 (0-6,1) 0 (0-1) 14,4 (7,6-24,7)
Helfer T-Zellen
CD3+/CD4+ 2,3 (0-2,9) 16,8 (3,2-29,2) 20,4 (2,4-45,2) 396,7 (83-696,8)
Zytotoxische T-Zellen CD3+/CD8+ 0,6 (0,3-0,9) 6,7 (0,5-13) 8,6 (3,2-44,4) 133,9 (26,3-363,4)
γδ T−Zellen γδ-TCR+ 0 (0) 1 (0-6,4) 2,2 (0-3,7) 11,9 (2,2-36,6)
Teff / Tregc CD4+/CD25+ 0 (0) 2,2 (1,6-11,2) 3,2 (1,3-5,2) 21 (9,7-53,8)
NK T-Zellen CD94+/CD3+ 0 (0) 0 (0-3,1) 0,7 (0-1,6) 10,1 (3,2-19,4)
NK-Zellen CD94+ 0 (0) 1,9 (0-5,2) 2,4 (0,4-8,2) 39,7 (12,2-217,6)
NK-Zellen CD56+ 0 (0) 0,2 (0-5,3) 7,7 (3,7-19,7) 23,1 (8,1-115,1)
pDZ CD123+/CD45RA+ 0 (0) 3,5 (0-6,5) 1,4 (0-2,8) 39,8 (14,6-85,8)
LZ Langerin+ 27,7 (18,2-32,8) 18 (13,3-21,5) 1,1 (0,9-3,2) 17,5 (1,1-28,1)
LZ CD1a+ 18,7 (14,1-24,3) 12,1 (4,1-20,3) 13,5 (10,9-31,4) 44,5 (20,6-95,7)
DDZ CD1c+ 4,45 (1,6-7,7) 16,6 (3-26,3) 24 (11,6-49) 97,1 (53-164,5)
IDEZ/IDDZd CD1a+/CD1c+ 0,5 (0-2,4) 5,2 (1,4-8,2) 2,3 (1,1-20,4) 18,9 (4,8-43,5)
Mature LZ CD1a+/CD83+ 0,7 (0-1,6) 3,5 (1-6,4) 0,9 (0-1,8) 14,9 (9-40,9)
Mature DDZ CD1c+/CD83+ 1 (0,2-2) 1,4 (0,8-2,3) 5,9 (1,6-17,2) 11 (3,7-14,8)
Mastzellen CD117+(c-Kit) 0 (0) 0 (0) 18,5 (6-40,3) 31,1 (14,7-54,5)
Makrophagen CD14+ 0 (0-1) 1,9 (0-3,8) 15,2 (9,7-25,8) 30,6 (14,8-51)
Neutrophile CD15+ 0 (0) 0 (0-6,4) 0 (0) 29,8 (4,8-167,7)
Eosinophile EG1+ 0 (0) 0 (0) 0 (0) 21,5 (1,1-37,6)
a Die Zellen wurden mit den angegebenen Markern visualisiert und mittels IF analysiert.
Die Zahlen zeigen den Mittelwert und die Spannbreite (in Klammern) der Zellen, die in
10 ECT Reaktionen und 5 NH ermittelt wurden.
b Doppelt-negative T-Zellen
c Effektor T-Zellen / regulatorische T-Zellen
d Inflammatorische epidermale/dermale dendritische Zellen (IDEZ/IDDZ)

Ergebnisse Ch. Bangert
- 42 -
4.2. T-Zellen im ECT
4.2.1. Typisierung der T-Zellen im dermalen Infiltr at Die überwältigende Mehrheit der infiltrierenden Zellen der Epidermis und Dermis in
ECT Läsionen besteht nach 72 Std aus CD3+ T-Zellen. Zwei Drittel dieser Zellen
zählen zu den CD4+ Helfer T-Zellen während das verbleibende Drittel den CD8+
zytotoxischen T-Zellen zuzuordnen ist (Abbildung 4.2.1.1.). Wir konnten einen geringen
Anteil CD3+/CD4-/CD8- T-Zellen detektieren, aber keine CD3+/CD4+/CD8+ Zellen
(Abbildung 4.2.1.2.; Tabelle 4.1.).
In einem Maus Modell für Kontakthypersensitivität konnte gezeigt werden, dass γδ-T-
Zellen eine regulierende Rolle einnehmen können [93]. Wir versuchten, diese Zellen
auch im humanen ECT zu charakterisieren und verglichen deren Anzahl mit der Anzahl
der häufiger vorkommenden αβ-Rezeptor tragenden T-Zellen. Obwohl die Mehrheit der
dermalen infiltrierenden T-Zellen den αβ TZR trägt, zeigt doch eine kleine, gut
abgrenzbare Population den γδ Rezeptor (Tabelle 4.1.).
Um herauszufinden, ob die T-Zellen in der Haut proliferieren oder ob der Anstieg der
CD3+ Zellen durch einen Zustrom aus dem Blut bedingt ist, führten wir
Doppelfärbungen mit dem Proliferationsmarker Ki-67 und CD3 durch (Daten s.
Anhang). Keine der T-Zellen reagierte mit dem Antikörper Ki-67, so dass wir davon
ausgehen, dass in ECT Läsionen die T-Zellen aus dem Blut in die Läsion rekrutiert
werden.

Ergebnisse Ch. Bangert
- 43 -
Abbildung 4.2.1.1. Doppelfärbungen durch IF zur Charakterisierung der T-
Zellpopulationen im dermalen Infiltrat in 72 Std bestehenden ECT Reaktionen
(Originalvergrößerung: 200x). T-Helfer Zellen wurden durch anti-CD3 (FITC, links
oben)/anti-CD4 (TRITC, Mitte oben) charakterisiert, zytotoxische T-Zellen durch anti-
CD3 (FITC, links unten)/anti-CD8 (TRITC, Mitte unten). Zellen, die jeweils beide
Zielantigene tragen, erscheinen gelb (rechts oben bzw. unten). Die Mehrzahl der Zellen
ist der Subpopulation der CD4+ T-Zellen zuzuordnen.
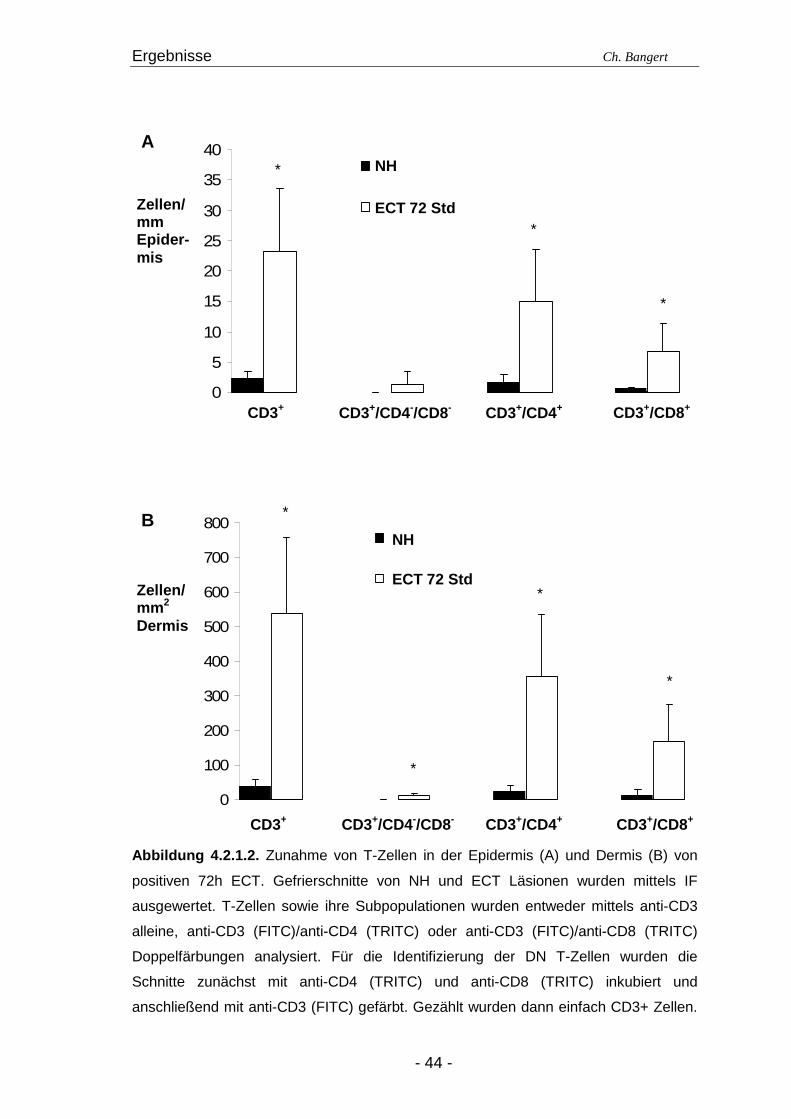
Ergebnisse Ch. Bangert
- 44 -
Abbildung 4.2.1.2. Zunahme von T-Zellen in der Epidermis (A) und Dermis (B) von
positiven 72h ECT. Gefrierschnitte von NH und ECT Läsionen wurden mittels IF
ausgewertet. T-Zellen sowie ihre Subpopulationen wurden entweder mittels anti-CD3
alleine, anti-CD3 (FITC)/anti-CD4 (TRITC) oder anti-CD3 (FITC)/anti-CD8 (TRITC)
Doppelfärbungen analysiert. Für die Identifizierung der DN T-Zellen wurden die
Schnitte zunächst mit anti-CD4 (TRITC) und anti-CD8 (TRITC) inkubiert und
anschließend mit anti-CD3 (FITC) gefärbt. Gezählt wurden dann einfach CD3+ Zellen.
0
5
10
15
20
25
30
35
40
Zellen/mm Epider-mis
Zellen/mm2 Dermis
CD3+ CD3+/CD4-/CD8- CD3+/CD4+ CD3+/CD8+
0
100
200
300
400
500
600
700
800
CD3+ CD3+/CD4-/CD8- CD3+/CD4+ CD3+/CD8+
∗
∗
∗
∗
∗
∗
∗
NH
ECT 72 Std
NH
ECT 72 Std
A
B

Ergebnisse Ch. Bangert
- 45 -
Insgesamt wurden nur Zellen ausgewertet, die eine deutlich positive Reaktion zeigten.
Die Daten zeigen den Mittelwert der T-Zellanzahl ± Standardabweichung in 10 ECT
Läsionen und 5 Biopsien von NH. Statistische Signifikanz wurde mit Students T-Test
ermittelt (٭p<0,012).
4.2.2. Charakterisierung der CD4+/CD25+ T-Zellsubpo pulation
Ungefähr 5% aller dermalen CD4+ Zellen reagierte mit Antikörpern gegen die IL-2-
Rezeptor α−Kette (CD25). Sowohl CD4 als auch CD25 können von aktivierten DZ,
Makrophagen oder reg T-Zellen exprimiert werden. Wir wollten wissen, ob im Infiltrat
der ECT Läsionen auch letztere vorhanden sind und führten deswegen eine FACS
Analyse von Zellsuspensionen durch, die wir aus Biopsien von 72 Std ECT gewannen.
Wir charakterisierten die T-Zellen im FACS anhand ihrer typischen Morphologie und
Größe. DZ oder Makrophagen konnten wir ausschließen.
Diese Untersuchungen zeigten eine CD4+/CD25+ T-Zellpopulation (ca. 30% der
Gesamtzellzahl), die zusätzlich das Antigen CTLA-4 aufwiesen. Die überwiegende
Mehrzahl der CD4+/CD25- T-Zellen hingegen zeigte keine gleichzeitige Expression
von CTLA-4 (Abb. 4.2.2.1.). Auch in situ konnte anhand von konfokaler
Lasermikroskopie mittels Dreifach- Immunfluoreszenzfärbungen eine beträchtliche
Anzahl von Zellen mit den phänotypischen Zeichen von reg T-Zellen nachgewiesen
und somit das Ergebnis der FACS Färbungen bestätigt werden (Abb. 4.2.2.2.). Eine
weitere Analyse mittels IF zeigte zusätzlich das Vorkommen des Moleküls Foxp3 auf
den CD4+/CD25+ T-Zellen (Daten s. Anhang).
Um Aufschluss über den zeitlichen Ablauf der Einwanderung von T-Zellen in die Haut
zu erhalten, untersuchten wir ECT Reaktionen, die wir zum Zeitpunkt 0, nach 24, 48
und 72 Std entnahmen. Bereits 24 Std nach Auftragen des Haptens beobachteten wir
eine beträchtliche Anzahl von CD4+ T-Zellen, die allerdings nur sehr wenig Reaktivität
gegen CD25 aufzeigten (Abb.4.2.2.3.). Allerdings stieg nach 48 Std Haptenkontakt der
relative Anteil von CD25+ Zellen an der CD4+ Zellpopulation deutlich an. Dies könnte
entweder bedeuten, dass die Subpopulation der CD4+/CD25+ T-Zellen verzögert aus
dem Blut rekrutiert wird oder dass sich die bereits eingewanderten CD25- T-Zellen
nach 48 Std in CD25+ Zellen umgewandelt hatten.
Diese CD25+/CD4+ T-Zelluntergruppe kann sowohl reg T-Zellen als auch Effektor T-
Zellen darstellen [85]. Derzeit kann noch nicht anhand eines bestimmten
Oberflächenmoleküls die eindeutige Zuordnung der Zellen in die eine oder andere

Ergebnisse Ch. Bangert
- 46 -
Gruppe erfolgen. Es gibt aber einige Antigene, die typischerweise auf reg T-Zellen zu
finden sind, wenn auch nicht ausschließlich. Zu diesen Molekülen zählen u.a. CTLA-4
und Foxp3.
Die von uns definierten Zellen wiesen diese Oberflächenstrukturen auf. Im humanen
KE könnten diese Zellen somit als reg T-Zellen zur Beendigung der Immunantwort
beitragen oder auch als Effektor- T-Zellen die Reaktion verstärken. Leider konnten wir
aufgrund der äußerst limitierten Zellzahlen, die wir aus den entnommenen 4mm ECT
Biopsien erhielten, keine funktionellen Untersuchungen durchführen, so dass die letzte
Bestätigung der regulatorischen Funktion dieser Zellen vorerst fehlt.
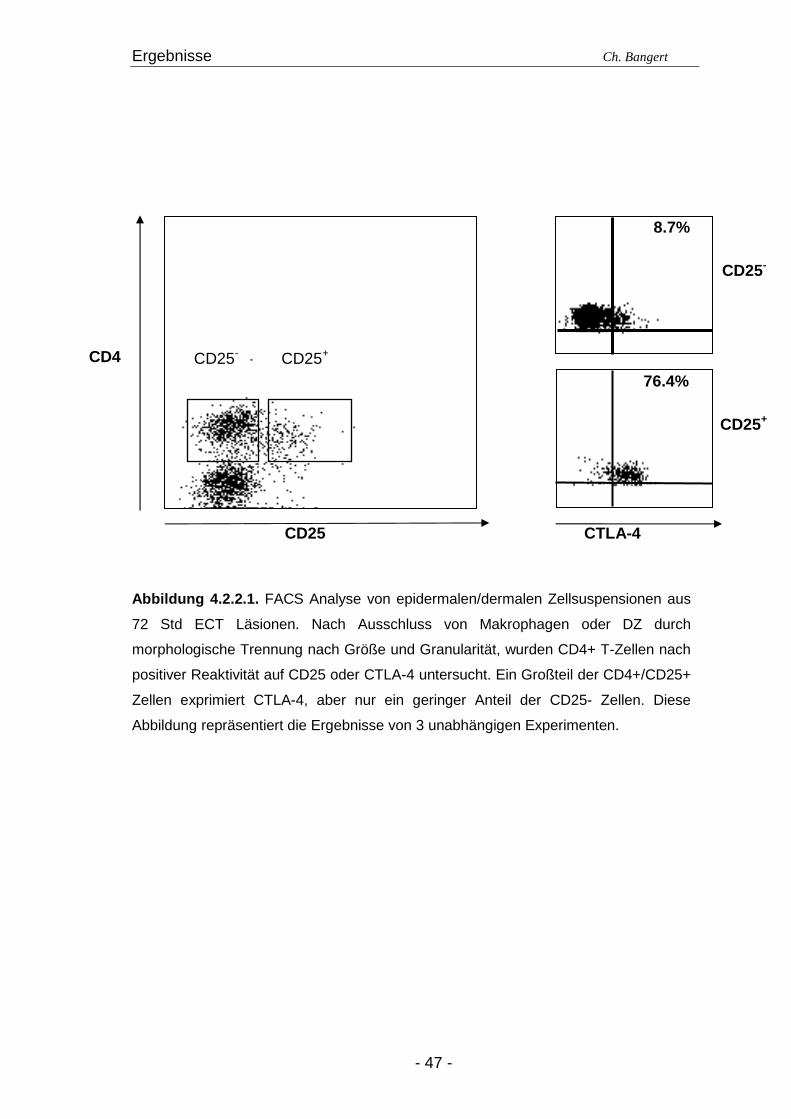
Ergebnisse Ch. Bangert
- 47 -
Abbildung 4.2.2.1. FACS Analyse von epidermalen/dermalen Zellsuspensionen aus
72 Std ECT Läsionen. Nach Ausschluss von Makrophagen oder DZ durch
morphologische Trennung nach Größe und Granularität, wurden CD4+ T-Zellen nach
positiver Reaktivität auf CD25 oder CTLA-4 untersucht. Ein Großteil der CD4+/CD25+
Zellen exprimiert CTLA-4, aber nur ein geringer Anteil der CD25- Zellen. Diese
Abbildung repräsentiert die Ergebnisse von 3 unabhängigen Experimenten.
CD25- CD25+
76.4%
8.7%
CD4
CD25 CTLA-4
CD25-
CD25+

Ergebnisse Ch. Bangert
- 48 -
Abbildung 4.2.2.2. In situ IF Färbungen mit CD4-FITC, CD25-TRITC
und CTLA-4-Cy5 in ECT Läsionen. In den typischen Infiltrathaufen der
Dermis finden sich viele CD4+ Zellen, die mehrfach auch CTLA-4 und
CD25 tragen. Das rechte untere Bild zeigt die Überlappung der
Färbungen, dreifach positive Zellen sind durch Pfeile gekennzeichnet
(Originalvergrößerung: 600x).

Ergebnisse Ch. Bangert
- 49 -
Abbildung 4.2.2.3. Zeitabfolge des Erscheinens von CD4+/CD3+ T-Zellen (oberes Bild)
und CD4+/CD25+ T-Zellen (unteres Bild) in das Infiltrat von positiven ECT. Die Biopsien
wurden zu vier unterschiedlichen Zeitpunkten nach Auftragung des Haptens
entnommen (Zeitpunkt 0, nach 24, 48 und 72 Std). Anhand von Doppelfärbungen in der
Immunfluoreszenz wurden entweder CD4/CD3 oder CD4/CD25 positive Zellen in der
Epidermis (Zellen/mm Basalmembran) und Dermis (Zellen/mm2) ausgezählt. In der
Epidermis gibt es nur sehr wenige T-Zellen zu jedem Zeitpunkt. Interessanterweise
findet sich nach 24 Std in der Dermis lediglich eine Zunahme von CD4+/CD3+ T-Zellen,
aber keine Vermehrung der CD4+/CD25+ Zellen. Nach 48 Std steigt der Anteil dieser
Zellen im Infiltrat jedoch deutlich an und bleibt auch noch nach 72 Std detektierbar.
0
5
10
15
20
25
30
35
40
Zellen/mm 2 Dermis
NH ECT 24 Std ECT 48 Std ECT 72 Std
CD4+/ CD25+ Zellen
0
50
100
150
200
250
300
CD4+/CD3+
Zellen
Zellen/mm Epidermis
Zellen/mm 2 Dermis
Zellen/mm Epidermis
NH ECT 24 Std ECT 48 Std ECT 72 Std

Ergebnisse Ch. Bangert
- 50 -
4.3. DZ Populationen im ECT
4.3.1. LZ im ECT
Zur Charakterisierung der LZ in NH und läsionaler Haut führten wir Färbungen mit anti-
CD1a und anti-Langerin durch (Abbildung 4.3.1.1.). Unter physiologischen Umständen
zeigte sich eine hohe Anzahl von CD1a+ und Langerin+ DZ in der Epidermis.
Demgegenüber wies die allergische Haut 72 Std nach Haptenapplikation eine deutliche
Veränderung auf: CD1a+ Zellen sind in der Epidermis deutlich reduziert und dafür in
der Dermis erhöht; Langerin+ LZ ebenfalls in der Epidermis vermindert, aber in der
Dermis nur leicht erhöht. Die Gesamtzahl der CD1a+ DZ ist im ECT verglichen mit NH
höher (Abbildung 4.3.1.2.).
Abbildung 4.3.1.1. Doppelfärbungen der LZ in 72 Std ECT Reaktionen mittels
Immunfluoreszenz anhand von CD1a-FITC (links) und Langerin-TRITC (Mitte). Die
Überlappung der beiden Antikörper zeigt sich in gelb (rechts). Die Zellen befinden sich
überwiegend in der Epidermis, wie durch die Pfeile angedeutet. Die weiße Linie markiert die
Basalmembran (Originalvergrößerung: 400x).
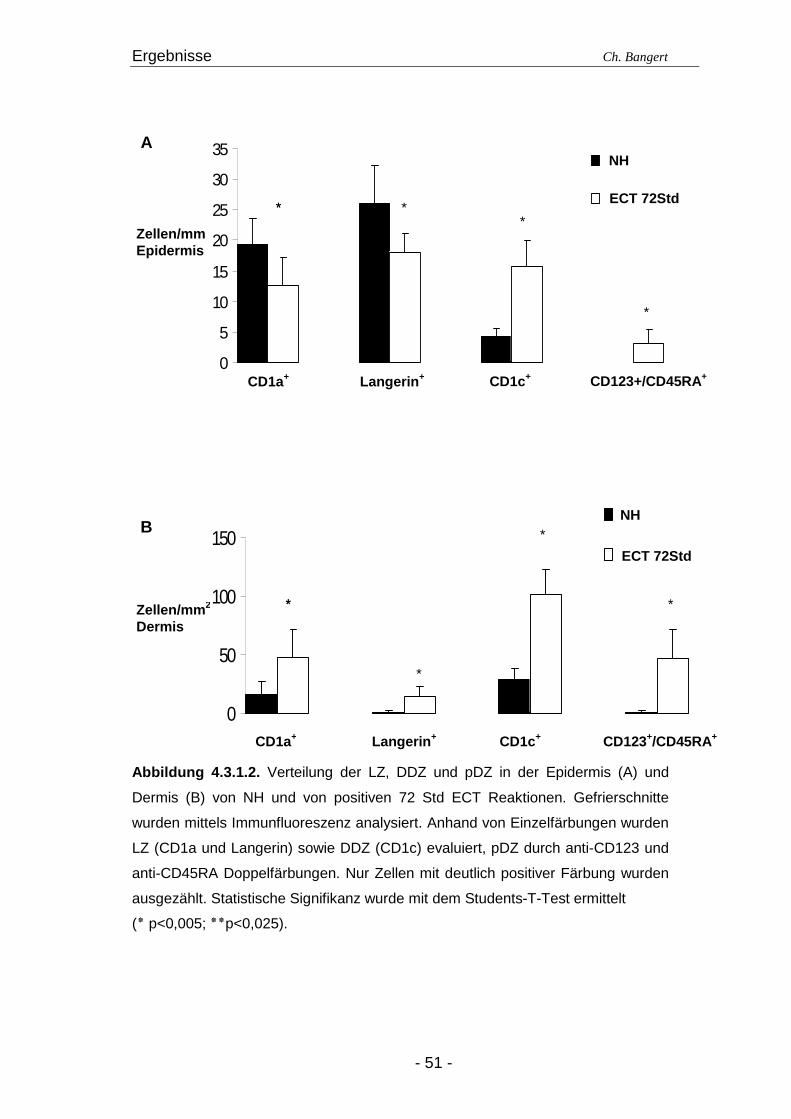
Ergebnisse Ch. Bangert
- 51 -
Abbildung 4.3.1.2. Verteilung der LZ, DDZ und pDZ in der Epidermis (A) und
Dermis (B) von NH und von positiven 72 Std ECT Reaktionen. Gefrierschnitte
wurden mittels Immunfluoreszenz analysiert. Anhand von Einzelfärbungen wurden
LZ (CD1a und Langerin) sowie DDZ (CD1c) evaluiert, pDZ durch anti-CD123 und
anti-CD45RA Doppelfärbungen. Nur Zellen mit deutlich positiver Färbung wurden
ausgezählt. Statistische Signifikanz wurde mit dem Students-T-Test ermittelt
.(p<0,025٭٭ ;p<0,005 ٭)
0
5
10
15
20
25
30
35
0
50
100
150
A
CD1a+ Langerin + CD1c+ CD123+/CD45RA+
CD1a+ Langerin + CD1c+ CD123+/CD45RA+
B
∗ ∗
∗
∗∗
∗∗ ∗
∗
∗
NH
ECT 72Std
NH
ECT 72Std
Zellen/mm Epidermis
Zellen/mm 2 Dermis
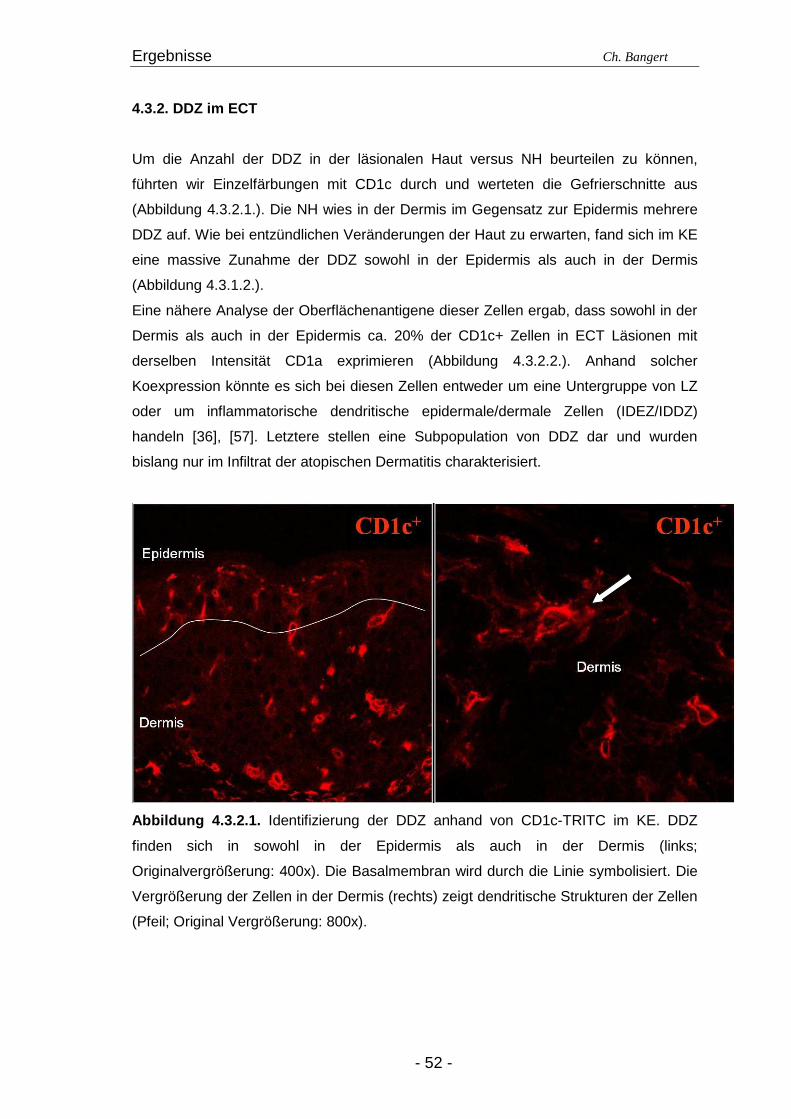
Ergebnisse Ch. Bangert
- 52 -
4.3.2. DDZ im ECT
Um die Anzahl der DDZ in der läsionalen Haut versus NH beurteilen zu können,
führten wir Einzelfärbungen mit CD1c durch und werteten die Gefrierschnitte aus
(Abbildung 4.3.2.1.). Die NH wies in der Dermis im Gegensatz zur Epidermis mehrere
DDZ auf. Wie bei entzündlichen Veränderungen der Haut zu erwarten, fand sich im KE
eine massive Zunahme der DDZ sowohl in der Epidermis als auch in der Dermis
(Abbildung 4.3.1.2.).
Eine nähere Analyse der Oberflächenantigene dieser Zellen ergab, dass sowohl in der
Dermis als auch in der Epidermis ca. 20% der CD1c+ Zellen in ECT Läsionen mit
derselben Intensität CD1a exprimieren (Abbildung 4.3.2.2.). Anhand solcher
Koexpression könnte es sich bei diesen Zellen entweder um eine Untergruppe von LZ
oder um inflammatorische dendritische epidermale/dermale Zellen (IDEZ/IDDZ)
handeln [36], [57]. Letztere stellen eine Subpopulation von DDZ dar und wurden
bislang nur im Infiltrat der atopischen Dermatitis charakterisiert.
Abbildung 4.3.2.1. Identifizierung der DDZ anhand von CD1c-TRITC im KE. DDZ
finden sich in sowohl in der Epidermis als auch in der Dermis (links;
Originalvergrößerung: 400x). Die Basalmembran wird durch die Linie symbolisiert. Die
Vergrößerung der Zellen in der Dermis (rechts) zeigt dendritische Strukturen der Zellen
(Pfeil; Original Vergrößerung: 800x).
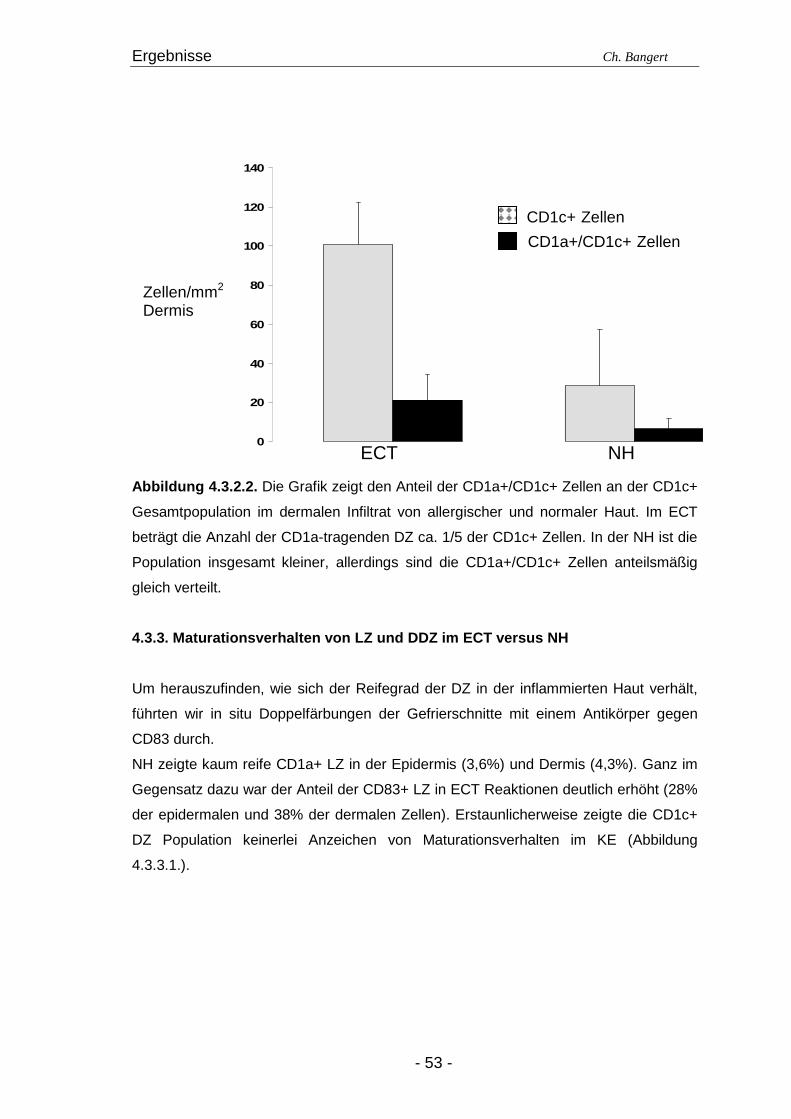
Ergebnisse Ch. Bangert
- 53 -
Abbildung 4.3.2.2. Die Grafik zeigt den Anteil der CD1a+/CD1c+ Zellen an der CD1c+
Gesamtpopulation im dermalen Infiltrat von allergischer und normaler Haut. Im ECT
beträgt die Anzahl der CD1a-tragenden DZ ca. 1/5 der CD1c+ Zellen. In der NH ist die
Population insgesamt kleiner, allerdings sind die CD1a+/CD1c+ Zellen anteilsmäßig
gleich verteilt.
4.3.3. Maturationsverhalten von LZ und DDZ im ECT v ersus NH
Um herauszufinden, wie sich der Reifegrad der DZ in der inflammierten Haut verhält,
führten wir in situ Doppelfärbungen der Gefrierschnitte mit einem Antikörper gegen
CD83 durch.
NH zeigte kaum reife CD1a+ LZ in der Epidermis (3,6%) und Dermis (4,3%). Ganz im
Gegensatz dazu war der Anteil der CD83+ LZ in ECT Reaktionen deutlich erhöht (28%
der epidermalen und 38% der dermalen Zellen). Erstaunlicherweise zeigte die CD1c+
DZ Population keinerlei Anzeichen von Maturationsverhalten im KE (Abbildung
4.3.3.1.).
ECT NH
Zellen/mm2 Dermis
0
20
40
60
80
100
120
140
CD1a+/CD1c+ Zellen CD1c+ Zellen
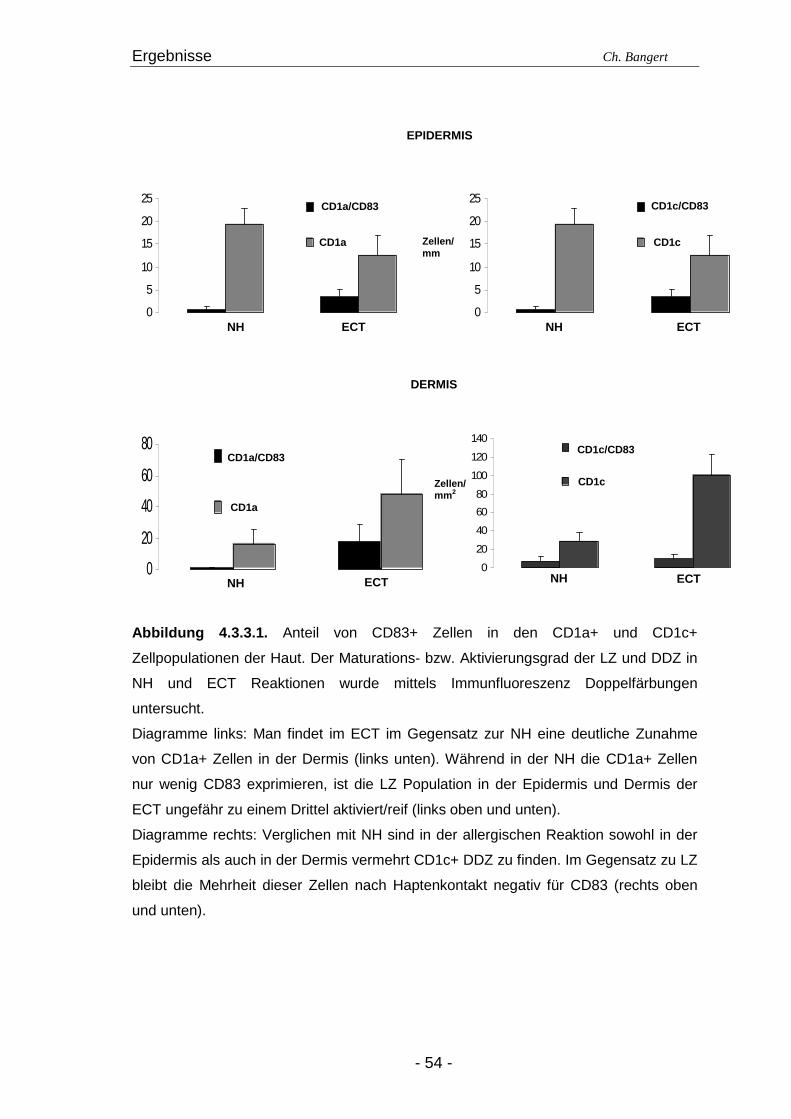
Ergebnisse Ch. Bangert
- 54 -
Abbildung 4.3.3.1. Anteil von CD83+ Zellen in den CD1a+ und CD1c+
Zellpopulationen der Haut. Der Maturations- bzw. Aktivierungsgrad der LZ und DDZ in
NH und ECT Reaktionen wurde mittels Immunfluoreszenz Doppelfärbungen
untersucht.
Diagramme links: Man findet im ECT im Gegensatz zur NH eine deutliche Zunahme
von CD1a+ Zellen in der Dermis (links unten). Während in der NH die CD1a+ Zellen
nur wenig CD83 exprimieren, ist die LZ Population in der Epidermis und Dermis der
ECT ungefähr zu einem Drittel aktiviert/reif (links oben und unten).
Diagramme rechts: Verglichen mit NH sind in der allergischen Reaktion sowohl in der
Epidermis als auch in der Dermis vermehrt CD1c+ DDZ zu finden. Im Gegensatz zu LZ
bleibt die Mehrheit dieser Zellen nach Haptenkontakt negativ für CD83 (rechts oben
und unten).
0
5
10
15
20
25
0
5
10
15
20
25
EPIDERMIS
DERMIS
NH NH ECT ECT
CD1c/CD83
CD1c
CD1a/CD83
CD1a Zellen/ mm
0
20
40
60
80
NH ECT
CD1a/CD83
CD1a
0
20
40
60
80
100
120
140
NH ECT
CD1c/CD83
CD1c Zellen/ mm 2

Ergebnisse Ch. Bangert
- 55 -
4.3.4. PDZ im Infiltrat der ECT Reaktionen
Interessanterweise konnten wir in den läsionalen Biopsien der ECT die erst vor einigen
Jahren identifizierte pDZ entdecken [63]. Zu unserer Überraschung war der Anteil
dieser Zellen an der gesamten infiltrierenden DZ Population im ECT relativ hoch
(Abbildung 4.3.1.2.).
Charakterisierung des Phänotyps der pDZ in situ
Im Menschen kann die pDZ anhand mehrerer verschiedener Oberflächenantigene
identifiziert werden. Zu diesen zählen: CD123, CD45RA, BDCA-2, CD68, CXCR3 und
CD62L [63], [64], [68], [116]. Um mehr über den Phänotyp dieser Zelle in NH und ECT
Läsionen zu erfahren, führten wir mittels Immunfluoreszenz Doppel- und
Dreifachfärbungen an Gefrierschnitten mit den aufgezählten Markern durch
(Abbildungen 4.3.4.1., 4.3.4.2.). In der NH konnten kaum pDZ detektiert werden. Die
Epidermis war überhaupt frei von pDZ, und in der Dermis waren einzelne Zellen mit
Reaktivität gegen CD123/CD45RA, CD123/BDCA-2 und CD123/CD68 zu finden
(Abbildung 4.3.4.3.). Läsionale Haut hingegen enthielt pDZ sowohl im epidermalen als
auch im dermalen Anteil der Haut. Während CD123+ Zellen eine vergleichbare
Koexpression von CD45RA (Mittelwert: 3,5 Zellen/mm Epidermis und 39,8/mm2
Dermis), BDCA-2 (1,6 Zellen/mm Epidermis und 32,8/mm2 Dermis) und CD68 (2,3
Zellen/mm Epidermis und 24/mm2 Dermis) aufwiesen, konnte CD62L (L-Selectin)
lediglich auf einem Drittel der pDZ nachgewiesen werden (0,4 Zellen/mm Epidermis
und 8,5/mm2 Dermis) (Abbildung 4.3.4.3.). Da wir eine lokale Entstehung der pDZ
anhand von Doppelfärbungen mit dem Proliferationsmarker Ki-67 ausschließen
konnten, wäre es denkbar, dass unsere Ergebnisse aus einer Einwanderung dieser
Zellen von dem Blut in die Haut resultieren. Diese Einwanderung könnte mit dem
Verlust von CD62L einhergehen.

Ergebnisse Ch. Bangert
- 56 -
Abbildung 4.3.4.1. In situ Doppelfärbungen von pDZ im dermalen Infiltrat
von ECT Biopsien, die 72 Std nach Haptenkontakt entnommen wurden.
Charakterisierung anhand von CD123-TRITC/ BDCA-2-FITC (oben,
Vergrößerung: 800x), CD68-TRITC/ CD123-FITC (Mitte, Vergrößerung:
400x) und CD45RA-TRITC/ CD123-FITC (unten, Vergrößerung: 400x). Die
Bilder auf der rechten Seite zeigen die Überlappung der auf der Zelle
vorhandenen Antigene jeweils in gelb.
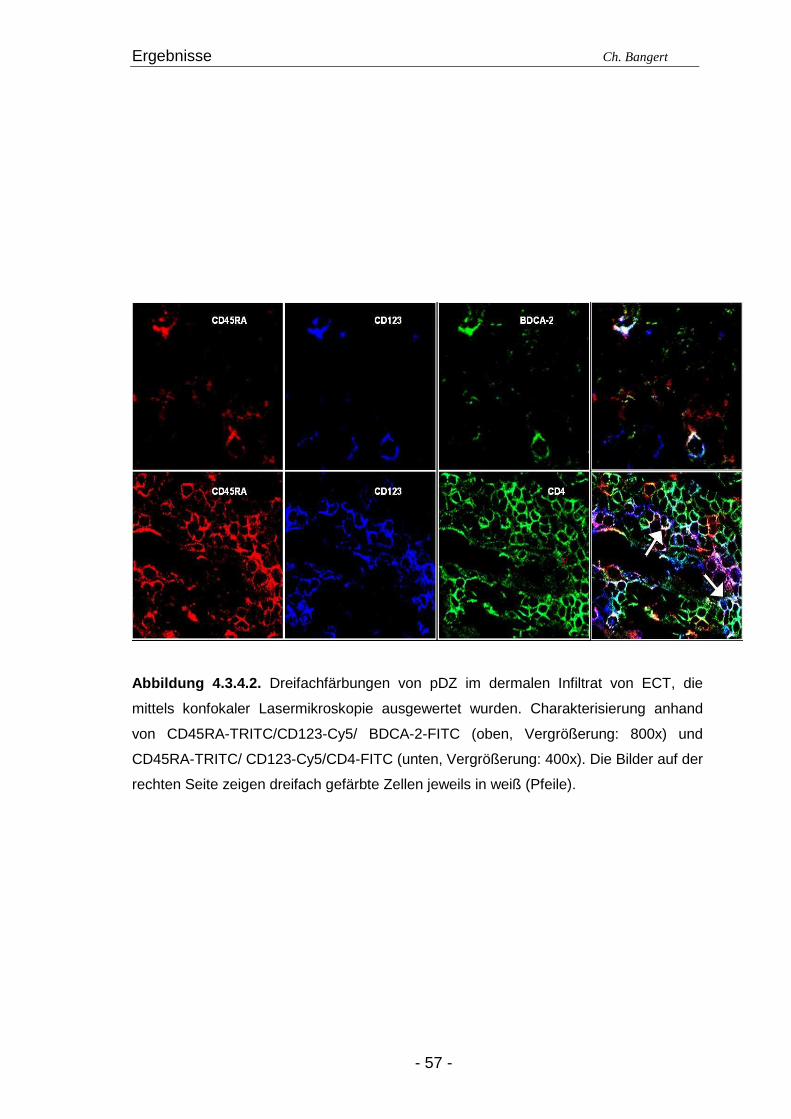
Ergebnisse Ch. Bangert
- 57 -
Abbildung 4.3.4.2. Dreifachfärbungen von pDZ im dermalen Infiltrat von ECT, die
mittels konfokaler Lasermikroskopie ausgewertet wurden. Charakterisierung anhand
von CD45RA-TRITC/CD123-Cy5/ BDCA-2-FITC (oben, Vergrößerung: 800x) und
CD45RA-TRITC/ CD123-Cy5/CD4-FITC (unten, Vergrößerung: 400x). Die Bilder auf der
rechten Seite zeigen dreifach gefärbte Zellen jeweils in weiß (Pfeile).
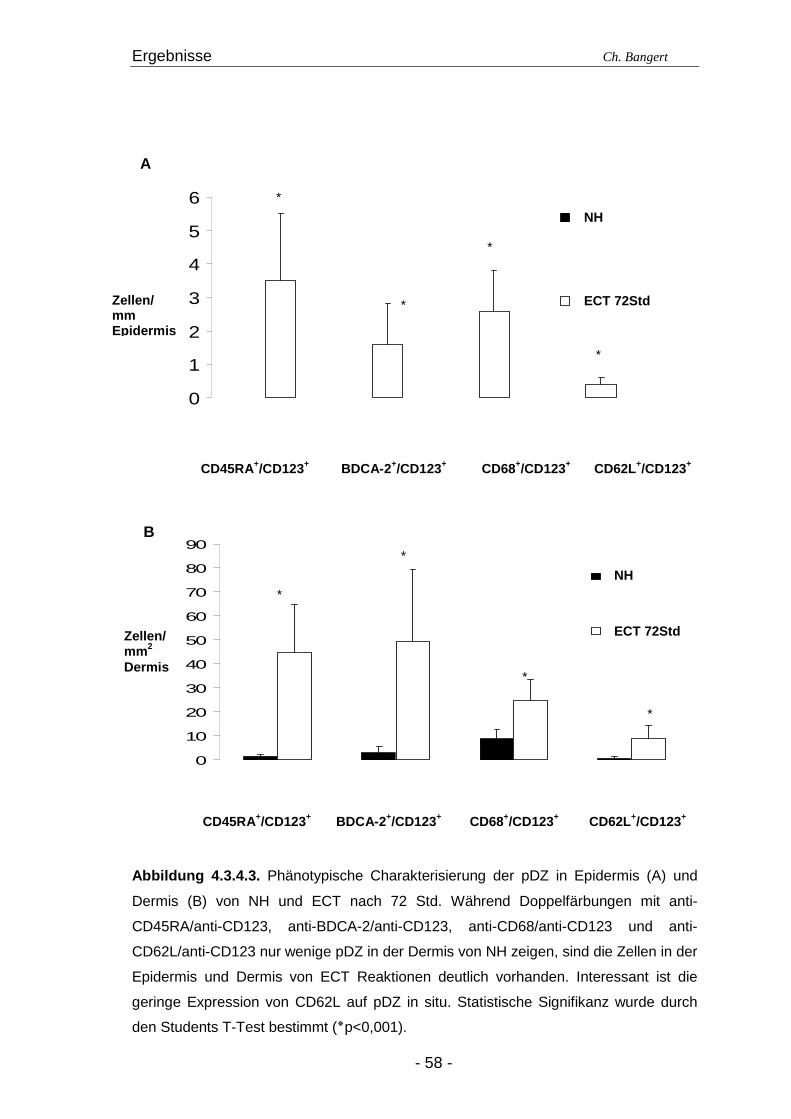
Ergebnisse Ch. Bangert
- 58 -
Abbildung 4.3.4.3. Phänotypische Charakterisierung der pDZ in Epidermis (A) und
Dermis (B) von NH und ECT nach 72 Std. Während Doppelfärbungen mit anti-
CD45RA/anti-CD123, anti-BDCA-2/anti-CD123, anti-CD68/anti-CD123 und anti-
CD62L/anti-CD123 nur wenige pDZ in der Dermis von NH zeigen, sind die Zellen in der
Epidermis und Dermis von ECT Reaktionen deutlich vorhanden. Interessant ist die
geringe Expression von CD62L auf pDZ in situ. Statistische Signifikanz wurde durch
den Students T-Test bestimmt (٭p<0,001).
A
CD45RA+/CD123+ BDCA-2+/CD123+ CD68+/CD123+ CD62L+/CD123+
0
1
2
3
4
5
6
∗
∗ NH
ECT 72Std
∗
∗
B
0
10
20
30
40
50
60
70
80
90∗
NH
ECT 72Std
∗
∗
∗
CD45RA+/CD123+ BDCA-2+/CD123+ CD68+/CD123+ CD62L+/CD123+
Zellen/ mm Epidermis
Zellen/mm 2 Dermis

Ergebnisse Ch. Bangert
- 59 -
Vergleich des Phänotyps in läsionaler Haut und Blut
Um die in der läsionalen Haut vorhandenen pDZ noch besser charakterisieren zu
können, führten wir an Einzelzellsuspensionen, die aus Biopsien der ECT-Reaktionen
angefertigt wurden, durchflußzytometrische Analysen durch und verglichen diese
Ergebnisse mit dem peripheren Blut des jeweiligen Patienten (Abbildung 4.3.4.4.).
Sowohl in der Haut als auch im Blut wiesen ungefähr 90% der definierten
CD123+/CD45RA+ Zellpopulation ebenso CD4 und MHCII an der Oberfläche auf. Bei
der Analyse der CD123+/MHCII+ Zellen zeigte sich, dass diese pDZ aus Haut und Blut
ein homogenes Expressionsmuster von BDCA-2 besitzen. PDZ aus dem Blut zeigten
keine Zeichen der Maturation, da keine Reaktivität gegen CD83 oder CD86 zu finden
war. Ganz im Gegensatz dazu sind ca. 50% der pDZ aus der Haut aktiviert/gereift, wie
die Expression von CD83 und CD86 vermuten lässt (Abbildung 4.3.4.4.).
Migrationsmoleküle auf pDZ
Weiterhin suchten wir nach Mechanismen, welche die Adhäsion der pDZ an
Endothelzellen und die Einwanderung aus dem peripheren Blut möglich machen
könnten.
Tatsächlich zeigten 80% der PDZ aus der Haut und 95% der PDZ aus dem Blut eine
verstärkte CLA Expression (cutaneous lymphocyte-associated antigen). Die Adhäsion
der CLA+ pDZ könnte über die Bindung an E-Selectin vollzogen werden, da bekannt
ist, dass dieser Ligand auf Venen in entzündlichen Hauterkrankungen wie dem KE
deutlich vermehrt vorkommt [117], [118].
Die im Blut zirkulierenden pDZ gelangen über die Bindung von CD62L an PNAd
(peripheral lymph node addressin) auf Endothelzellen von HEV (high endothelial
venules) in den LK [63].
Tatsächlich zeigte sich in unserem Experiment, dass beinahe 100% der pDZ aus dem
Blut das Oberflächenantigen CD62L trugen. Demgegenüber war das Molekül nur auf
etwa 50-70% der pDZ aus der Haut vorhanden (Abbildung 4.3.4.4.). Diese Abnahme
von CD62L in der Haut ist mit unseren in situ Daten gut vereinbar (Abbildung 4.3.4.3).
Um herauszufinden, ob pDZ tatsächlich die Möglichkeit haben, die ECT Läsionen über
CD62L zu betreten, untersuchten wir, ob PNAd auf dem Gefäßendothel in der Haut
vorhanden ist. Wir konnten zeigen, dass ein kleiner Anteil Endothelzellen in den nach
72 Std entnommenen ECT-Biopsien auf die immunhistochemische Färbung mit MECA-
79 positiv reagierte (Abbildung 4.3.4.5.).

Ergebnisse Ch. Bangert
- 60 -
Ein weiteres Signal zur Einwanderung in die Haut kann durch Chemokine vermittelt
werden, die von verschiedenen entzündlichen, infiltrierenden Zellen produziert werden
können [119]. Auf einer Untergruppe der pDZ konnten wir den Chemokinrezeptor
CXCR3 nachweisen, der die Zellen für eine Anlockung über Botenstoffe wie CXCL9
(MIG), CXCL10 (IP-10) oder CXCL11 (ITAC) empfänglich macht (Abbildung 4.3.4.6.).
Zusammenfassend zeigen unsere Ergebnisse, dass eine beträchtliche Anzahl
CD62L+/CLA+ pDZ in ECT Läsionen einwandern und dort auch teilweise maturieren.

Ergebnisse Ch. Bangert
- 61 -

Ergebnisse Ch. Bangert
- 62 -
Abbildung 4.3.4.4. FACS Analyse von Einzelzellsuspensionen aus Blut und Haut. A)
PDZ aus Haut und Blut exprimieren CD123/CD45RA/CD4/MHCII auf ihrer Oberfläche.
CD123+/CD45RA- sowie CD123+/CD45RA+ Populationen wurden ausgewählt und auf
ihre Expression von CD4 und MHCII hin begutachtet. Sowohl in der Haut als auch im
Blut zeigte die CD45RA+ Population, ganz im Gegensatz zur CD45RA- Gruppe,
deutlich MHCII und CD4 auf der Oberfläche.
B) Vergleich der Adhäsionsmoleküle und Maturationsmarker auf CD123+/MHCII+
Zellen in der Haut und im Blut. Während CLA auf pDZ aus beiden Milieus eine ähnliche
Reaktivität aufweist, haben pDZ aus der Haut zu etwa 30% den Ligand für E-Selektin,
nämlich CD62L, auf ihrer Oberfläche verloren. Im Vergleich zum Blut findet sich in der
Haut eine eindeutige Oberflächenexpression der Maturations-/Aktivierungsmoleküle
CD83 und CD86 auf pDZ. Die Abbildung repräsentiert eines von 5 ähnlichen
Experimenten.

Ergebnisse Ch. Bangert
- 63 -
Abbildung 4.3.4.5. Immunhistochemische Färbungen von humanen ECT Reaktionen
(A und B) sowie von NH (C) mit anti-PNAd (MECA-79).
A) Übersichtsaufnahme; Vergrößerung 100x. Im Infiltrat zeigen sich rot gefärbte,
MECA-79+ Zellen. B) Vergrößerter Ausschnitt von A) (200x).
C) In der NH zeigt das Gefäßendothel keine Reaktivität mit dem Liganden für CD62L,
anti-PNAd (Vergrößerung 100x).

Ergebnisse Ch. Bangert
- 64 -
Abbildung 4.3.4.6. Dreifachfärbungen von pDZ im dermalen Infiltrat von ECT
wurden mittels Immunfluoreszenz und konfokaler Lasermikroskopie ausgewertet.
Die Bilder zeigen dreifach positiv gefärbte Zellen jeweils in weiß (kleine Pfeile).
Doppelt gefärbte Zellen mit anti-CD123-Cy5 und anti-MHCII-FITC erscheinen
türkis und stellen Endothelzellen der Gefäße dar (große Pfeile). Links: Die
CD45RA+ (TRITC)/CD123+ (Cy5)/ MHCII+ (FITC) Zellen sind in der Nähe von
CD123+ Gefäßen (großer Pfeil), die man auch anhand der Morphologie
identifizieren kann (Vergrößerung: 400x). Rechts: CD123+ (Cy5)/MHCII+ (FITC)
pDZ tragen CXCR3 (TRITC) (Vergrößerung: 400x). Der große Pfeil deutet auch
hier auf das Lumen eines Gefäßes.

Ergebnisse Ch. Bangert
- 65 -
4.3.5. NK-Zellen und pDZ im ECT Infiltrat
Kinetik
Um zu untersuchen, wie schnell pDZ das KE infiltrieren und wieder verschwinden,
entnahmen wir Biopsien von ECT Reaktionen zu vier unterschiedlichen Zeitpunkten.
Die Auswertung der CD123+/CD45RA+ Zellen nach 24 Std Haptenkontakt zeigte eine
deutliche Zunahme der Zellzahl (0,7/mm Epidermis und 17,4/mm2 Dermis), die nach 48
Std gipfelte (1,2/mm Epidermis und 28,2/mm2 Dermis) und danach wieder abnahm
(Abbildung 4.3.5.1.). Auch Analysen von einem frühen Zeitpunkt (nach 6 Std) ergaben,
verglichen mit NH, bereits einen deutlichen Anstieg von pDZ im Infiltrat.
Die Tatsache, dass pDZ als Zellen des angeborenen Immunsystems bereits früh in der
Effektorphase ins KE rekrutiert werden, sowie Berichte über eine Beteiligung von
Proteinen des angeborenen Immunsystems an der Pathogenese der
Hypersensitivitätsreaktion [120, 121], verleitete uns dazu, nach Zellen dieser Immunität
zu suchen.
Wir identifizierten NK-Zellen anhand von CD56 oder CD94 im Infiltrat der seit 72 Std
bestehenden kontaktallergischen Reaktionen (Tabelle 4.1.). Weiters interessierte uns
der zeitliche Ablauf der Einwanderung dieser Zellen und die zuvor beschriebenen
Biopsien wurden auf CD56+ NK-Zellen untersucht.
Es fand sich eine Migrationskinetik ähnlich der von pDZ mit einer Zunahme der NK-
Zellen bereits nach 24 Std (13,7/mm2 Dermis), einem Gipfel nach 48 Std (21,8/mm2)
und einer Abnahme der Zellzahl nach 72 Std (Abbildung 4.3.5.1.).
Nachbarschaft
PDZ können bei entsprechender Stimulation große Mengen an IFN-α ausschütten,
welches wiederum ein potentieller Aktivator von NK-Zellen ist. Um herauszufinden, ob
dieses Szenarium im KE eintreten könnte, untersuchten wir, ob eine räumliche Nähe
von pDZ und NK- Zellen detektierbar ist.
Bei der Analyse des Infiltrats zeigte sich tatsächlich häufig eine Nachbarschaft dieser
Zellen (Abbildung 4.3.5.2.).
Da CD56 als Oberflächenmolekül ebenfalls auf NK-T-Zellen vorkommt, prüften wir als
nächstes, wie hoch der Anteil von CD56+/CD3+ NK-T-Zellen in den Läsionen ist.
Zu unserer Überraschung exprimierten keine der CD56+ Zellen den T-Zellrezeptor
(Abbildung 4.3.5.3.). Hingegen fanden wir einige CD94+/CD3+ NK-T-Zellen, ein Viertel
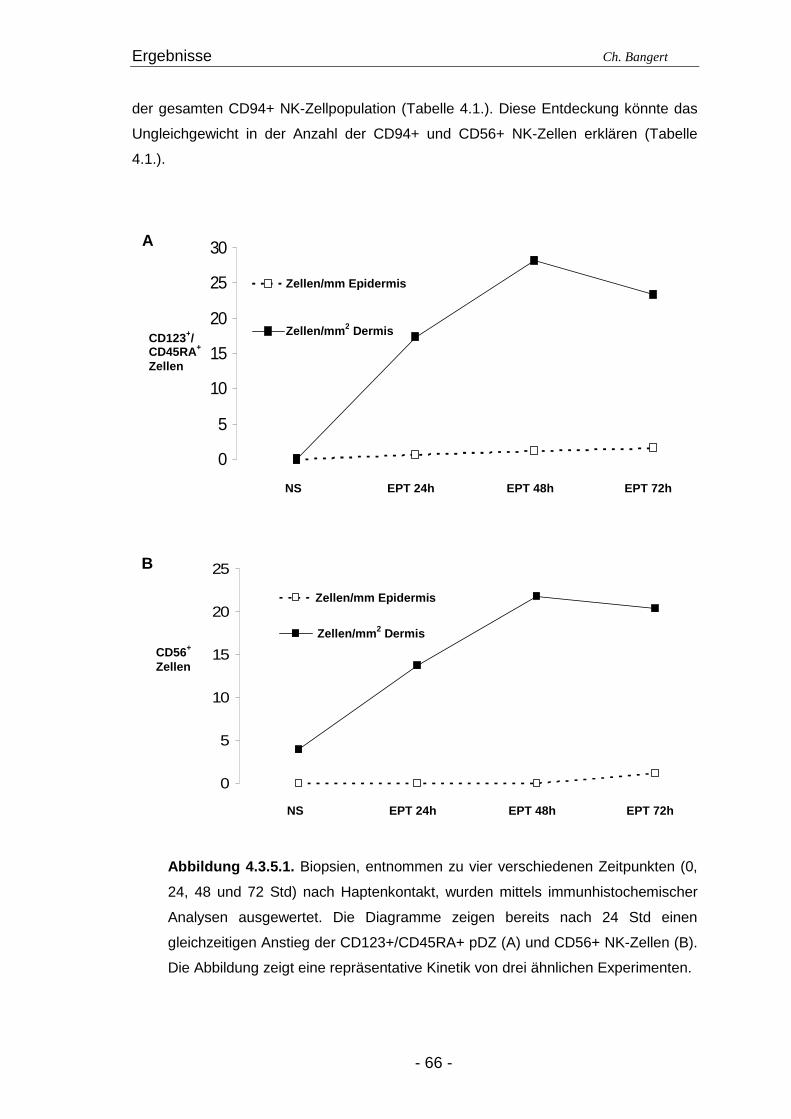
Ergebnisse Ch. Bangert
- 66 -
der gesamten CD94+ NK-Zellpopulation (Tabelle 4.1.). Diese Entdeckung könnte das
Ungleichgewicht in der Anzahl der CD94+ und CD56+ NK-Zellen erklären (Tabelle
4.1.).
Abbildung 4.3.5.1. Biopsien, entnommen zu vier verschiedenen Zeitpunkten (0,
24, 48 und 72 Std) nach Haptenkontakt, wurden mittels immunhistochemischer
Analysen ausgewertet. Die Diagramme zeigen bereits nach 24 Std einen
gleichzeitigen Anstieg der CD123+/CD45RA+ pDZ (A) und CD56+ NK-Zellen (B).
Die Abbildung zeigt eine repräsentative Kinetik von drei ähnlichen Experimenten.
A
B
0
5
10
15
20
25
Zellen/mm Epidermis
Zellen/mm 2 Dermis
NS EPT 24h EPT 48h EPT 72h
CD56+ Zellen
0
5
10
15
20
25
30
Zellen/mm Epidermis
Zellen/mm 2 Dermis
NS EPT 24h EPT 48h EPT 72h
CD123+/ CD45RA+ Zellen
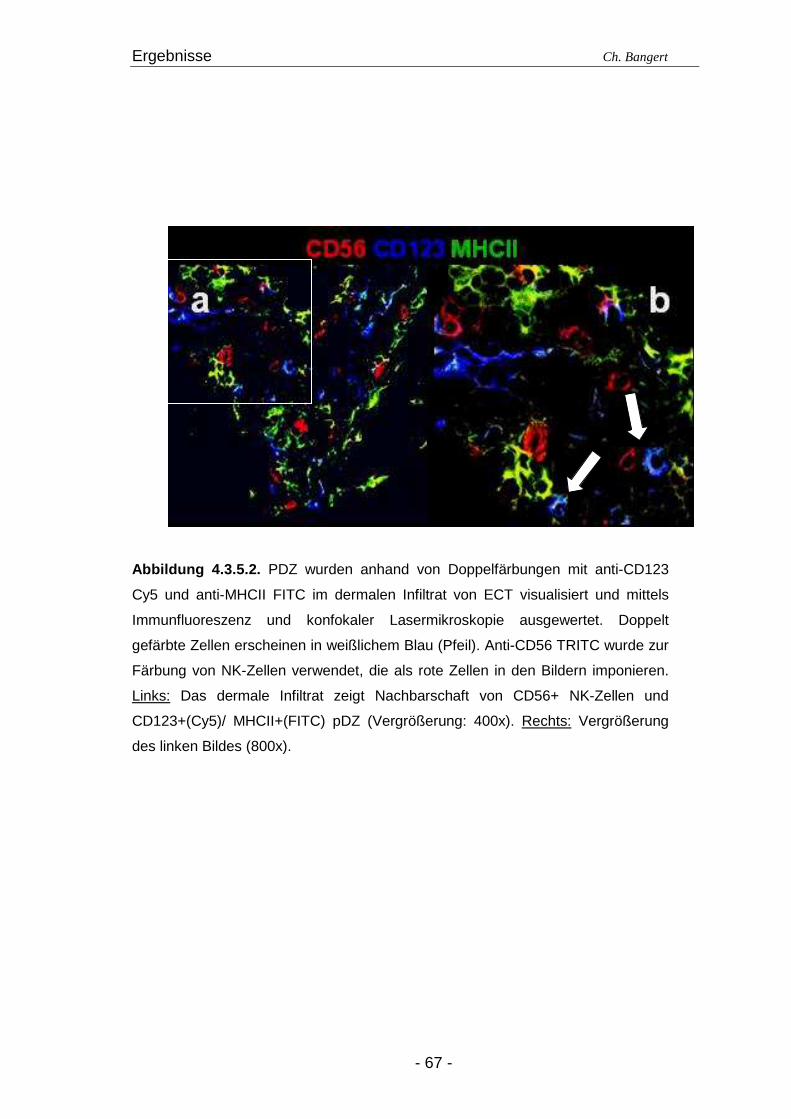
Ergebnisse Ch. Bangert
- 67 -
Abbildung 4.3.5.2. PDZ wurden anhand von Doppelfärbungen mit anti-CD123
Cy5 und anti-MHCII FITC im dermalen Infiltrat von ECT visualisiert und mittels
Immunfluoreszenz und konfokaler Lasermikroskopie ausgewertet. Doppelt
gefärbte Zellen erscheinen in weißlichem Blau (Pfeil). Anti-CD56 TRITC wurde zur
Färbung von NK-Zellen verwendet, die als rote Zellen in den Bildern imponieren.
Links: Das dermale Infiltrat zeigt Nachbarschaft von CD56+ NK-Zellen und
CD123+(Cy5)/ MHCII+(FITC) pDZ (Vergrößerung: 400x). Rechts: Vergrößerung
des linken Bildes (800x).

Ergebnisse Ch. Bangert
- 68 -
A
Abbildung 4.3.5.3. Doppelfärbungen mit anti-CD56 (TRITC) (links) und anti-CD3
(FITC) (Mitte) im dermalen Infiltrat von ECT Reaktionen zeigen keine
Überlappung der Oberflächenantigene auf den Zellen (Bilder rechts). Analysen
wurden mittels konfokaler Lasermikroskopie durchgeführt.

Ergebnisse Ch. Bangert
- 69 -
4.4. Vergleich des Zellinfiltrats in ECT Reaktionen induziert durch Nickel und
Duftstoff
4.4.1. T-Zell-Infiltrat
Bisher ist unbekannt, ob unterschiedliche Haptene eine Auswirkung auf die
Zusammensetzung des Zellinfiltrats in ECT Reaktionen, die nach 72 Std entnommen
wurden, haben.
Um herauszufinden, ob die Quantität und Qualität von T-Zellen in Läsionen, die durch
Ni oder Duftstoff hervorgerufen wurden, Unterschiede aufzeigt, analysierten wir jeweils
fünf Biopsien von den entsprechenden Kontaktallergikern mittels IF (Abbildung
4.4.1.1).
Es zeigte sich, dass die Anzahl der CD3+ T-Zellen in beiden Reaktionen vergleichbar
ist (Abbildung 4.4.1.1). Nickelallergiker wiesen eine leicht höhere absolute Zellzahl auf,
bedingt durch die etwas stärkeren ECT Reaktionen (siehe auch Material und
Methoden, Tabelle 3.1.1.).
Auch die einzelnen T-Zellpopulationen zeigten keine Unterschiede im Verhältnis
zueinander. Dominierend waren CD4+ T-Zellen, gefolgt von CD8+ T-Zellen. Hingegen
bildeten CD25+/CD4+ T-Zellen, CD94+/CD3+ NK-T-Zellen und γδ-Rezeptor tragende
T-Zellen nur einen kleinen Anteil des gesamten CD3+ Zellinfiltrats.
Statistische Signifikanz zwischen den Ni- und Duftstoff-induzierten Reaktionen bestand
nicht.
4.4.2. DZ-Populationen
Eine weitere zu identifizierende große Zellbevölkerungsgruppe ist die der DZ.
Unterschiede im Verhältnis der LZ, DDZ oder pDZ untereinander könnten Aufschluss
über eine unterschiedliche Art der Antigenpräsentation der Haptene geben.
Wie schon zuvor beschrieben, charakterisierten wir DDZ mittels anti-CD1c, LZ mit anti-
Langerin, pDZ mit anti-BDCA-2/anti-CD123 und IDEZ/IDDZ mit anti-CD1c/anti-CD1a in
ECT Reaktionen von Ni- und Duftstoffallergikern sowie in NH (Abbildung 4.4.2.1.).
Die Anzahl der Langerin+ LZ in der Epidermis der NH war höher als in der Epidermis
von ECT Reaktionen. PDZ und IDEZ/IDDZ waren in der NH praktisch nicht vorhanden.
Unterschiede hinsichtlich Verteilung und Anzahl von DZ in den ECT Reaktionen auf die
beiden Haptene waren nicht zu finden.

Ergebnisse Ch. Bangert
- 70 -
4.4.3. Identifizierung weiterer Leukozyten
Um einen vollständigen Überblick über die infiltrierenden Zellpopulationen zu erhalten,
charakterisierten wir auch Granulozyten (CD15), Makrophagen (CD14), NK-Zellen
(CD94) und Mastzellen (CD117) in den Läsionen und in der NH (Abbildung 4.4.3.1.).
Auch diese Zellarten betreffend kam es zu keinerlei signifikanten Unterschieden in der
Verteilung der Populationen im Ni- oder Duftstoff-induzierten KE. Lediglich die Anzahl
der CD94+ NK-Zellen war im Ni-ECT gegenüber dem Duftstoff-ECT erhöht, was aber
wiederum am wahrscheinlichsten auf die allgemein stärkere Reaktion auf Ni
zurückzuführen ist (Abbildung 4.4.3.1.).
Interessant war, dass Mastzellen zumindest nicht durch ihre Quantität an der
Pathologie des KE beteiligt zu sein scheinen, da die Anzahl der CD117+ Mastzellen in
der NH und im ECT vergleichbar hoch ist.
Weiterhin konnten wir zeigen, dass etwa die Hälfte aller CD14+ Makrophagen in den
Läsionen auch eine Oberflächenexpression von CD1c aufweist. Dies könnte ein
Hinweis auf den positiven Aktivierungsstatus der Zellen sein, oder es handelt sich um
die in humaner Haut beschriebenen Vorläuferzellen, die je nach Stimulationsmodus in
LZ oder DDZ transformiert werden können (TGF-β1: LZ; IL-4+GM-CSF: DDZ) [37],
[122] (Abbildung 4.4.3.1.).
4.4.4. Reaktion eines Nicht-Allergikers auf 24Std N ickelprovokation
Wir untersuchten weiterhin, ob bei einem Nicht-Allergiker nach Applikation von Ni
zumindest eine frühzeitige Immunreaktion ausgelöst wird, die sich in Form von
Einwanderung der Zellen des angeborenen Immunsystems wie NK-Zellen oder pDZ
zeigen würde. Die zelluläre Analyse ergab ein Infiltrat, welches mit dem Infiltrat in der
NH vergleichbar ist.
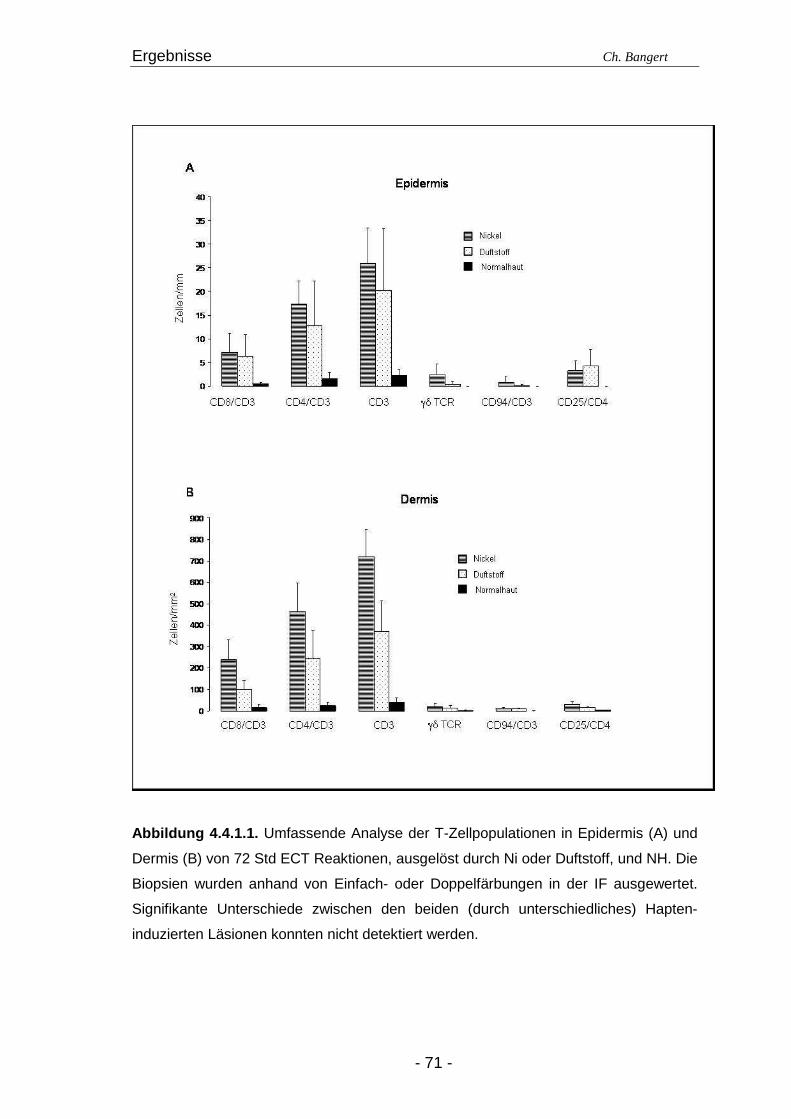
Ergebnisse Ch. Bangert
- 71 -
Abbildung 4.4.1.1. Umfassende Analyse der T-Zellpopulationen in Epidermis (A) und
Dermis (B) von 72 Std ECT Reaktionen, ausgelöst durch Ni oder Duftstoff, und NH. Die
Biopsien wurden anhand von Einfach- oder Doppelfärbungen in der IF ausgewertet.
Signifikante Unterschiede zwischen den beiden (durch unterschiedliches) Hapten-
induzierten Läsionen konnten nicht detektiert werden.
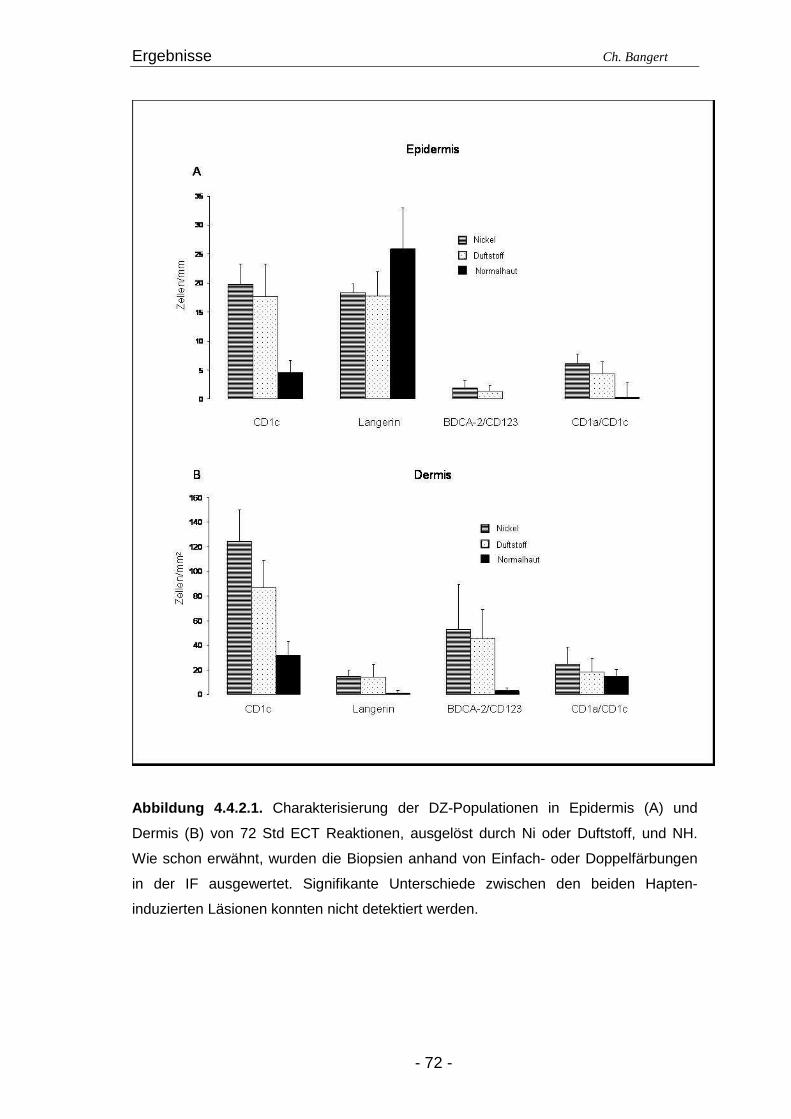
Ergebnisse Ch. Bangert
- 72 -
Abbildung 4.4.2.1. Charakterisierung der DZ-Populationen in Epidermis (A) und
Dermis (B) von 72 Std ECT Reaktionen, ausgelöst durch Ni oder Duftstoff, und NH.
Wie schon erwähnt, wurden die Biopsien anhand von Einfach- oder Doppelfärbungen
in der IF ausgewertet. Signifikante Unterschiede zwischen den beiden Hapten-
induzierten Läsionen konnten nicht detektiert werden.

Ergebnisse Ch. Bangert
- 73 -
Abbildung 4.4.3.1. Identifizierung weiterer Zellpopulationen in Epidermis (A) und
Dermis (B) von 72 Std ECT Reaktionen, ausgelöst durch Ni oder Duftstoff, und NH.
Auch hier wurden die Biopsien anhand von Einfach- oder Doppelfärbungen in der IF
ausgewertet. Signifikante Unterschiede zwischen den beiden Hapten-induzierten
Läsionen konnten nicht detektiert werden.

Diskussion Ch. Bangert
- 74 -
5. DISKUSSION
Viele Studien, die sich bisher mit dem humanen KE beschäftigt haben, konzentrierten
sich auf die Sensibilisierungsphase. Während einige Kenntnis über die dieser Reaktion
zugrunde liegenden zellulären und molekularen Ereignisse existiert, ist das Wissen
über Mechanismen der Effektorphase des KE begrenzt [74], [81]. Um mehr über
infiltrierende Zellpopulationen, insbesondere über APZ, die während dieses Prozesses
die T-Zell Antwort beeinflussen, zu erfahren, haben wir in der vorliegenden Studie das
leukozytäre Infiltrat von ECT phänotypisch analysiert. Hierbei richteten wir unser
Augenmerk besonders auf dendritische und T-Zellen. In diesem Zusammenhang
beschreiben wir das Vorkommen von pDZ in ECT Läsionen und vergleichen deren
Anzahl und Aktivierungszustand mit LZ und DDZ.
LZ und DDZ (Abb 5.1.)
Während der LZ in der Sensibilisierungsphase der Kontaktallergie eine maßgebliche
Rolle zugeschrieben wird [52], ist ihre Funktion in der Effektorphase, falls sie eine
solche innehaben, noch immer umstritten [49], [50]. Basierend auf der Expression von
Langerin und CD1a verglichen wir die Anzahl von LZ, die in NH und ECT Läsionen zu
finden sind. Auffallend war eine deutliche Verminderung der LZ in der Epidermis,
während es in der Dermis zu einem Anstieg der Zellzahl kam. Diese Daten sind
übereinstimmend mit Berichten in der Literatur. Die LZ sind in der Epidermis über
Adhäsionsmoleküle mit Keratinozyten verbunden. Bei entzündlichen Prozessen bzw.
auch nach Haptenapplikation schütten Keratinozyten u.a. die Zytokine IL-1β und TNF-α
aus, die LZ zur Hinabregulierung ihrer Adhäsionsmoleküle und in Folge zur Migration
aus der Epidermis veranlassen können [49], [123], [124].
Die Mehrheit der LZ zeigte CD83 an ihrer Oberfläche, ein Molekül, das während des
Maturationsprozesses von LZ bzw. DDZ zu finden ist. Dieser Maturationsvorgang
könnte auf die Haptenprovokation zurückzuführen sein, da einige in vitro Experimente
gezeigt haben, dass mit Haptenmolekülen inkubierte DZ/LZ in einen aktivierten
Zustand versetzt wurden [96], [97].
Es sollte betont werden, dass die Anzahl der CD1a+ Zellen jene der Langerin+ LZ in
der Dermis der ECT Läsionen deutlich überstieg. Die Frage, welcher Population diese
CD1a+Langerin- Zellen angehören, lässt sich mit drei möglichen Theorien
beantworten.

Diskussion Ch. Bangert
- 75 -
Sie könnten
1. LZ Vorläufer Zellen aus dem peripheren Blut
2. CD1a+/CD1c+ IDEZ/IDDZ
3. mature LZ nach Haptenprovokation auf dem Weg aus der Epidermis in den LK
darstellen [57].
Da wir nachweisen konnten, dass ca. 1/5 der CD1c+ Zellen auch CD1a in der Dermis
der ECT Läsionen exprimieren, könnte es sich tatsächlich um IDDZ handeln.
Antigen kann ebenso von DDZ aufgenommen und präsentiert werden. Bei DDZ
handelt es sich um eine Subpopulation von myeloiden DZ, die CD1c an ihrer
Oberfläche haben [36], [125]. Wir fanden ein dichtes CD1c+ Infiltrat in der Dermis von
ECT Reaktionen, die in ihrer Mehrheit kein CD83 an der Oberfläche exprimierten, d.h.
in einem immaturen, nicht-aktivierten Zustand waren. Basierend auf diesen
Entdeckungen stellten wir die Hypothese auf, dass LZ nach Antigenkontakt maturieren
und im Anschluss den Hapten-spezifischen Effektor T-Zellen das Allergen
präsentieren. Dadurch wird die Immunantwort der Effektorphase gestartet. Die unreifen
DDZ hingegen könnten eher eine regulierende Rolle spielen, in der sie reg T-Zellen
aktivieren, welche dann wiederum etwas später für die Beendigung der Immunantwort
verantwortlich sind [126].
Die Daten von Kissenpfennig et al widersprechen dieser These. Mittels eines
Mausmodells, in dem LZ durch an Langerin gebundenes Diphterietoxin vor
Sensibilisierung mit Dinitrofluorbenzol (DNFB) eliminiert wurden, konnte die Gruppe
zeigen, dass die Hypersensitivitätsreaktion nach erneutem DNFB Kontakt trotz Fehlen
der LZ in keinster Weise abgeschwächt wird [55]. Depletiert man bei Diphterietoxin
unbehandelten Mäusen 6 Wochen nach der Sensibilisierung mit DNFB und 3 Tage vor
erneutem DNFB Kontakt die LZ, kann die Immunantwort trotzdem ohne
Beeinträchtigung stattfinden. Dieses letzte Experiment zeigt, dass nach Ausbildung der
Hapten-spezifischen Gedächtnis T-Zellen keine LZ zur Auslösung der Effektorphase
notwendig sind [55].
Ob allerdings diese in der Maus generierten Daten direkt auf das humane KE
übertragbar sind, bleibt (wie immer) fraglich. Auch ist nach wie vor denkbar, dass
mature epidermale oder dermale LZ im Menschen zur Amplifizierung der
Immunantwort beitragen; d.h. sie verstärken lediglich durch Haptenpräsentation die
Effektorphase. Sollten sie nicht vorhanden sein, könnte das KE zwar milder, aber
dennoch stattfinden. Der (Gegen)Beweis bleibt aus, da hier im humanen System das
Vergleichsmodell fehlt.
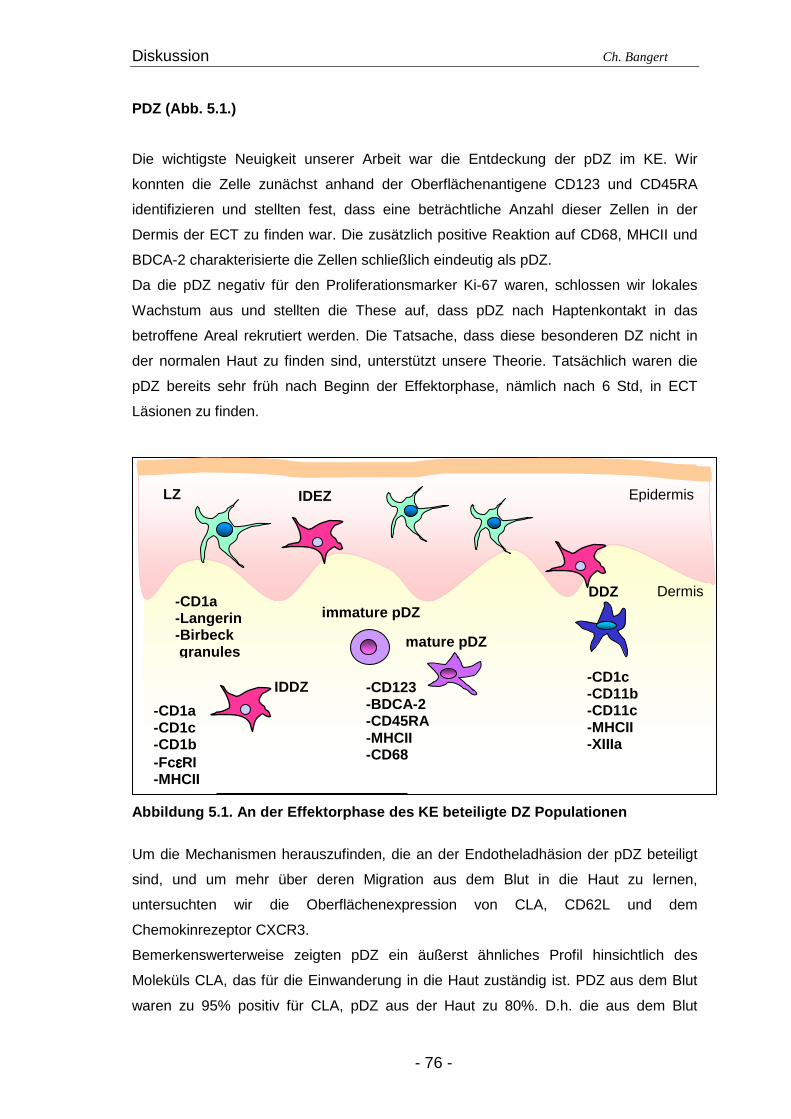
Diskussion Ch. Bangert
- 76 -
PDZ (Abb. 5.1.)
Die wichtigste Neuigkeit unserer Arbeit war die Entdeckung der pDZ im KE. Wir
konnten die Zelle zunächst anhand der Oberflächenantigene CD123 und CD45RA
identifizieren und stellten fest, dass eine beträchtliche Anzahl dieser Zellen in der
Dermis der ECT zu finden war. Die zusätzlich positive Reaktion auf CD68, MHCII und
BDCA-2 charakterisierte die Zellen schließlich eindeutig als pDZ.
Da die pDZ negativ für den Proliferationsmarker Ki-67 waren, schlossen wir lokales
Wachstum aus und stellten die These auf, dass pDZ nach Haptenkontakt in das
betroffene Areal rekrutiert werden. Die Tatsache, dass diese besonderen DZ nicht in
der normalen Haut zu finden sind, unterstützt unsere Theorie. Tatsächlich waren die
pDZ bereits sehr früh nach Beginn der Effektorphase, nämlich nach 6 Std, in ECT
Läsionen zu finden.
Abbildung 5.1. An der Effektorphase des KE beteilig te DZ Populationen
Um die Mechanismen herauszufinden, die an der Endotheladhäsion der pDZ beteiligt
sind, und um mehr über deren Migration aus dem Blut in die Haut zu lernen,
untersuchten wir die Oberflächenexpression von CLA, CD62L und dem
Chemokinrezeptor CXCR3.
Bemerkenswerterweise zeigten pDZ ein äußerst ähnliches Profil hinsichtlich des
Moleküls CLA, das für die Einwanderung in die Haut zuständig ist. PDZ aus dem Blut
waren zu 95% positiv für CLA, pDZ aus der Haut zu 80%. D.h. die aus dem Blut
-CD123 -BDCA-2 -CD45RA -MHCII -CD68
-CD1c -CD11b -CD11c -MHCII -XIIIa
-CD1a -Langerin -Birbeck granules
immature pDZ
mature pDZ
DDZ
LZ
IDDZ
IDEZ
Dermis
Epidermis
-CD1a -CD1c -CD1b -FcεεεεRI -MHCII

Diskussion Ch. Bangert
- 77 -
entnommenen pDZ haben unter inflammatorischen Bedingungen die Möglichkeit, die
Haut zu betreten, indem sie an E-Selektin auf den Endothelzellen der Blutgefäße
binden und schließlich den Zielort in der Haut über Interaktion mit Chemokinen
erreichen. Zusätzlich tragen die peripher isolierten pDZ von Nickelallergikern zu 100%
CD62L, ein Selektin, das an HEV bindet und somit den Eintritt in den LK erleichtert.
Interessanterweise haben pDZ in der Haut eine deutlich reduzierte
Oberflächenexpression von CD62L.
Dieses Phänomen wurde bereits im LK beobachtet, wo pDZ nach dem Eintritt in den
LK via HEV kein CD62L mehr aufweisen [63].
Eine weitere Beobachtung zeigt, dass ein Teil der infiltrierenden pDZ im ECT den
Chemokinrezeptor CXCR3 trägt. Die Liganden dieses Rezeptors, die Chemokine
CXCL9 (MIG), CXCL10 (IP-10) und CXCL11 (ITAC) werden u.a. von Keratinozyten
gebildet und sind in entzündlicher Haut verstärkt zu finden. Die Bindung von CXCR3 an
die entsprechenden Liganden könnte ein weiterer Mechanismus der DZ sein, um in die
Haut zu gelangen.
Tatsächlich wurde bereits in einer anderen Studie gezeigt, dass pDZ die Fähigkeit
haben über die Bindung an CXCR3/CXCR4 zu emigrieren [127].
Wie kommen pDZ also in die Haut? Unsere Daten zeigen, dass pDZ prinzipiell
mehrere Möglichkeiten haben, in die Läsionen einzudringen. Adhäsion an
Endothelzellen via Interaktionen von CLA/E-Selektin und CD62L/PNAd, wie zuvor
beschrieben wurde [117], [128] ist denkbar. Unser Nachweis von PNAd und E-Selektin
in inflammierter Haut unterstützt diese Theorie.
In früheren Studien wurde bereits gezeigt, dass pDZ, ähnlich wie LZ und DDZ,
durchaus fähig sind, potente T-Zellantworten zu provozieren [129], [130]. Da es
Beweise gibt, dass die Maturations-/Aktivierungszustand der DZ wichtig für die
entstehende Art der T-Zellantwort ist [131], untersuchten wir auch das Maturationsprofil
der aus Haut und Blut isolierten pDZ.
Anhand durchflußzytometrischer Analysen zeigte sich, dass 50% der Haut-
infiltrierenden pDZ die Maturations- bzw. Aktivierungsantigene CD83 und CD86 an
ihrer Oberfläche tragen. Die entsprechenden Zellen aus dem Blut hingegen wiesen
keinerlei Anzeichen von Maturation oder Aktivierung auf. Dies bedeutet, dass pDZ,
nachdem sie das Blut verlassen haben, zumindest zum Teil Maturationssignalen
ausgesetzt sind und sich dadurch die Fähigkeit erwerben, T-Zellen zu stimulieren. Ob
diese Stimuli allerdings bereits auf dem Weg in die Haut oder erst in der Läsion gesetzt
werden, ist derzeit nicht beurteilbar. Da die Zellen bereits nach 6 Std in der Haut zu

Diskussion Ch. Bangert
- 78 -
finden sind, unsere Analyse der Einzelzellsuspensionen aber auf der Untersuchung
einer 72 Std ECT Läsion beruht, könnten die 50% der aktivierten pDZ diesen Zustand
durchaus in der Haut erworben haben.
Humane in vitro Studien wiesen nach, dass pDZ je nach Art der Stimulation eine T1
oder T2 Antwort auslösen können. Wenn pDZ mit viralen Produkten stimuliert werden,
tendieren T-Zellen eher zu einer T1 Antwort, um den Eindringling möglichst effektiv
bekämpfen zu können. Im Falle einer IL-3/CD40L (Ligand) Aktivierung, wird eher eine
T2 Antwort ausgebildet [132], [133].
Es ist vorstellbar, dass pDZ im KE mit IL-3, welches von Mastzellen produziert wird,
und mit CD40L in Berührung kommen, welches auf T-Zellen vorhanden ist. Eine
weitere wichtige Studie zeigte, dass in der Maus pDZ, die mit CD40L stimuliert werden,
eine regulatorische CD8+ T-Zellantwort auslösen können [134].
Welche Rolle die pDZ in den ECT Läsionen einnehmen, sei es im immaturen oder
auch maturen Zustand, muss zum jetzigen Zeitpunkt offen bleiben.
T-Zellen
Die von uns analysierten CD3+ Zellen exprimieren vorwiegend den αβ TZR, was auch
zuvor in anderen bereits Studien gezeigt wurde [76], [135]. Tatsächlich zeigen nur
wenige der infiltrierenden T-Zellen den γδ TZR.
Wir fanden ein massives T-Zellinfiltrat in den ECT-Läsionen, welches eindeutig von
CD4+ T-Zellen dominiert wurde. Die CD4+:CD8+ T-Zellratio betrug etwa 2:1. Aufgrund
dieser zahlenmäßigen Überlegenheit der CD4 T-Zellen ließe sich leicht schlussfolgern,
dass die CD4 T-Zellen zu den Effektorzellen und KE-auslösenden Zellen zählen. Bis
vor etwa 10 Jahren war dies das vorherrschende Erklärungsmodell für Beteiligung der
T-Zellen in der Überempfindlichkeitsreaktion. Allerdings konnte mittlerweile in einigen
Studien gezeigt werden, dass vermutlich trotz der geringeren Quantität die CD8+ T-
Zellen im KE die Effektoren darstellen, während CD4+ Zellen die Immunreaktion
regulieren könnten [78], [77], [84].
Interessanterweise entdeckten wir tatsächlich eine CD4+ T-Zellpopulation, die
regulatorische T-Zellmarker, nämlich CD25 und CTLA-4, in der läsionalen Haut
aufwies. Diese Population konnten wir in der normalen Haut nicht finden. Nähere
Analysen zeigten, dass auch FOXP3+ Zellen im ECT vorhanden sind, ein weiterer
wichtiger Hinweis auf das Vorliegen einer reg T-Zellpopulation.
Diese Zellen konnten wir relativ spät im Entzündungsverlauf in der Haut detektieren: 24

Diskussion Ch. Bangert
- 79 -
Std nach Haptenprovokation zeigten sich vorwiegend CD4+/CD25- T-Zellen, nach 48
Std jedoch waren CD4+/CD25+ Zellen vorhanden. Dies könnte entweder bedeuten,
dass diese Zellen tatsächlich später aus dem Blut in die Haut rekrutiert werden oder
dass sie sich aus der CD4+/CD25- Population, die schon früh in den Läsionen
nachweisbar ist, umwandeln. Eine in situ Proliferation der CD4+ T-Zellen findet nicht
statt, wie wir anhand von Ki-67 Färbungen nachweisen konnten.
Es ist nicht eindeutig zu sagen, ob diese CD4+/CD25+ T-Zellen Effektorzellen oder
eher die bereits beschriebenen Allergen-spezifischen reg T-Zellen darstellen [78]. Eine
weitere Möglichkeit wäre, dass diese Zellen unspezifische (nicht-Hapten spezifische)
Suppressor T-Zellen sind, wie bereits zuvor in anderen Erkrankungen beschrieben
wurde [85], [136].
Damit könnten diese CD4+/CD25+ T-Zellen die Fähigkeit haben, die allergen-
spezifische Entzündung zu beenden.
Zellen des angeborenen Immunsystems in ECT Läsionen
Neben den Zellen des erworbenen Immunsystems spielen die Zellen der angeborenen
Immunität eine quantitative Rolle im KE. Wir charakterisierten eine unterschiedlich
große Anzahl von eosinophilen und neutrophilen Granulozyten, NK-Zellen, Mastzellen
und Makrophagen in der Dermis der ECT Läsionen (72 Std nach Haptenprovokation).
Die Anzahl der Zellen war durchaus vergleichbar mit der Zahl der infiltrierenden pDZ
oder LZ. Die Funktion dieser Zellen muss komplett offen bleiben: spekulativ könnte es
eine für die Pathogenese des KE ‚unwichtige’ reflektorische Alarmreaktion des
Organismus sein, der lediglich auf die proinflammatorischen Zytokine reagiert, die von
den Keratinozyten als sofortige Reaktion auf die Haptenprovokation ausgeschüttet
werden. Als Folge könnten die von uns beobachteten Zellen der angeborenen
Immunabwehr in die Haut gelockt werden [102], [137]. Interessant wäre in diesem
Zusammenhang die zelluläre Analyse eines chronischen KE, da es sich hier um einen
langandauernden Prozess handelt und nicht, wie beim ECT, um eine akute Reaktion
des Immunsystems.
NK-Zellen
Aufgrund des deutlichen Vorkommens von NK-Zellen in den Läsionen, untersuchten
wir das Einwanderungsverhalten dieser bei Bedarf wichtigen Effektorzellen in den ECT.
Es zeigte sich, dass NK-Zellen, ähnlich wie pDZ, sehr früh in den Läsionen zu finden
sind (nach 6 Std). Außerdem auffällig war die Tatsache, dass sich pDZ und NK-

Diskussion Ch. Bangert
- 80 -
Zellen innerhalb des entzündlichen Infiltrats in direkter Nachbarschaft befinden. Diese
Situation lässt auf eine Kommunikation zwischen diesen beiden Zellarten in der
Effektorphase des KE tippen. Tatsächlich wurde bereits in der atopischen Dermatitis
ein sogenannter ‚Crosstalk’ zwischen NK-Zellen und DZ nachgewiesen [138].
Durch ihre bekanntermaßen große Produktion von IFN-α können pDZ nicht nur einen
autokrinen Bogen erzeugen, der ihr eigenes Überleben verlängert [133], sondern auch
zur Aktivierung und Stimulation von NK-Zellen beitragen [139].
Vergleich des Infiltrats nach unterschiedlicher Hap tenapplikation
Von Relevanz war die Frage, ob dieses von uns charakterisierte Infiltrat spezifisch für
ein bestimmtes Hapten ist, oder ob die Auslösung des KE immer nach einem ähnlichen
Schema abläuft.
Wir untersuchten zwei unterschiedliche Gruppen von Kontaktallergikern, die entweder
eine Überempfindlichkeit auf Ni oder auf Duftstoff aufwiesen.
Nach genauer und umfassender Analyse der infiltrierenden Zellpopulationen zeigte
sich, dass die Immunreaktion in ECT Läsionen nicht von der Art des Haptens abhängig
ist.
Unterschiede hinsichtlich Bindung der Haptene an Peptide oder MHC-assoziierte
Bindungsstellen an den LZ lösen somit im Endeffekt keine Unterschiede der
Immunantwort auf zellulärer Ebene aus.
Wir untersuchten weiterhin, ob bei einem Nicht-Allergiker nach Applikation von Ni eine
frühzeitige Immunreaktion ausgelöst wird, die sich in Form von Einwanderung der
Zellen des angeborenen Immunsystems wie NK-Zellen oder pDZ zeigen würde. Dies
war nicht der Fall, was bedeutet, dass entweder ein starkes Irritans (z.B. Dodecyl-
Sulfat) [140], [141] oder eine bereits erfolgte Sensibilisierung auf ein Hapten notwendig
sind, um das von uns untersuchte Infiltrat auszulösen. Unsere Versuchsperson war
erwiesenermaßen bereits zuvor mit Ni in Kontakt gekommen, doch eine
Sensibilisierung wurde verhindert. Auf welcher Ebene die Sensibilisierung mit einem
Hapten verhindert wird bzw. ob eine Sensibilisierung überhaupt verhindert werden
muss, ist bislang unklar. Hapten-spezifische reg T-Zellen, die eine Immunantwort
unterdrücken können, findet man bei Nicht-Allergikern [82]. Diese Zellen sind aber
beim ersten Hautkontakt des Haptens noch nicht in der Haut zu finden und können
auch nicht die Akutreaktion der Keratinozyten auf ein Hapten beeinflussen. Es könnte
aber sein, dass diese Zellen beim Gesunden nach Kontakt mit einem beliebigen
Hapten und entsprechender Antigen-Präsentation im regionären LK proliferieren und
bereits im LK eine weitere Immunreaktion unterdrücken.

Diskussion Ch. Bangert
- 81 -
Hypothese/Fazit
Nach unserem Wissen ist dies die erste Studie, die eine detaillierte und umfassende
Analyse über die zelluläre Komposition und Kinetik des inflammatorischen Infiltrats im
KE liefert.
Anhand der detaillierten deskriptiven Analyse des zellulären Infiltrats in ECT Läsionen
können wir die folgende Hypothese zum Ablauf/zur Pathogenese der Effektorphase
aufstellen:
Eine nach Sensibilisierung erneut durchgeführte Haptenapplikation, entweder mit Ni
oder Duftstoff, führt (wahrscheinlich nach der Ausschüttung proinflammatorischer
Zytokine durch Keratinozyten) zur raschen Einwanderung von Zellen der angeborenen
Immunität wie NK-Zellen und auch pDZ. Beide Zellarten können in normaler Haut nicht
gefunden werden. PDZ tragen in ECT Läsionen verschiedene Oberflächenmoleküle
(CXCR3, CLA, CD62L), die ihnen unter diesen entzündlichen Bedingungen die
Bindung an Endothelzellen oder HEVs ermöglicht und somit den Zutritt in die Haut.
Eine Entwicklung aus Haut ansässigen Vorläuferzellen ist unwahrscheinlich, da die
pDZ nicht in der Haut proliferieren. In ECT Läsionen gehören sie der zweitgrößten
Gruppierung von DZ an und haben rein quantitativ das Potential, entscheidend zum
Ablauf der Immunreaktion beizutragen. Durch ihren frühen Eintritt in die Haut und
durch ihre Fähigkeit TNF-α und IFN-α zu produzieren, sichern sie ihr eigenes
Überleben und aktivieren gleichzeitig NK-Zellen, die in ihrer direkten Nachbarschaft im
KE-Infiltrat zu finden sind.
Etwa die Hälfte der pDZ sind in den ECT Läsionen in einem reifen Zustand,
wahrscheinlich durch die Stimulation mit CD40L (von T-Zellen) und IL-3 (von ebenfalls
vermehrten Mastzellen). Aufgrund der Expression von MHCII und BDCA-2 ist eine
Haptenaufnahme im immaturen Zustand denkbar, der nach Maturation der Zellen die
Antigenpräsentation der T-Zellen (in der Haut oder im LK) folgt. Die pDZ erreichen
ihren quantitativen Höhepunkt nach 72 Std im Infiltrat.
Die von den pDZ aktivierten NK-Zellen schütten ihrerseits IFN-γ aus, das zur weiteren
Amplifizierung der Immunreaktion beiträgt.
Die in der Haut ansässigen DZ, wie LZ und DDZ, verändern sich ebenfalls nach
Haptenapplikation. Stimuliert durch die Keratinozyten-Zytokine IL-1β oder TNF-
α regulieren LZ ihre Adhäsionsmoleküle an der Zelloberfläche hinunter und beginnen,
die Epidermis zu verlassen.
Eine rezente Studie zeigte, dass in vergrößerten regionären LK, die von Patienten mit
chronisch entzündlichen Hautkrankheiten entnommen wurden, viele LZ, die man in

Diskussion Ch. Bangert
- 82 -
einem afferenten Lymphgefäß oder auch in der T-Zellzone der LK fand, in einem
unreifen Zustand sind, da nur wenige CD1a+Langerin+ Zellen das Maturationsmolekül
CD83 tragen [142]. Bis zu dieser Studie waren Maturation und Migration der LZ ein
aneinander gekoppeltes Ereignis.
Im dermalen Infiltrat der ECT Läsionen zeigen die Hälfte der LZ eine positive
Reaktivität für CD83. Es ist denkbar, dass einige LZ bereits durch den lokalen Kontakt
mit CD40L, während oder vor der T-Zell-Haptenpräsentation, zu reifen Zellen
gewachsen sind. Vielleicht verbleiben diese LZ in der Dermis, um dort Hapten zu
präsentieren, während sich die immaturen LZ auf den Weg in den LK machen. Kürzlich
erschienene Berichte legen den Schluss nahe, dass LZ im KE weder eine essentielle
Rolle in der Sensibilisierungsphase noch in der Effektorphase des murinen KE spielen
[55].
Aufgrund ihrer Quantität und der epidermalen Haptennähe, ist eine komplett passive
Rolle der LZ im humanen KE jedoch schwer vorstellbar. Denkbar ist, dass diese Zellen
an der Amplifizierung der Effektorphase durch dermale Antigenpräsentation beitragen.
Die größte DZ Population bilden die DDZ. Diese Zellen bleiben vorwiegend in einem
immaturen, CD83- Zustand und sind vor allem in der Dermis anzutreffen.
Mit der Literatur durchaus vereinbar ist die These [55], [142], dass die DDZ unreif zu
den LK wandern (beladen mit dem Haptenmolekül), um dort den T-Zellen das Antigen
zu präsentieren. Denkbar wäre auch, dass durch den vorwiegend immaturen Status
dieser Zellen eher Hapten-spezifische reg T-Zellen ausgebildet oder nur noch zur
Proliferation angeregt werden, die sich dann auf den Weg in die Haut machen, um die
Immunreaktion zu beenden.
Eine weitere DZ Population, die wir charakterisieren konnten, sind IDEZ/IDDZ. Diese
Zellen sind CD1a+/CD1c+ und bislang lediglich in der atopischen Dermatitis
beschrieben [57]. Allerdings ist nicht sicher, ob es sich bei unseren Zellen tatsächlich
um IDEZ handelt, da wir auf die Oberflächenmoleküle CD1a/CD1c beschränkt waren
und diese Merkmale z.B. auch auf aktivierten LZ zu finden sind. Die Frage nach der
tatsächlichen Anwesenheit von IDEZ muss demnach offen bleiben.
Makrophagen zählen ebenfalls zu Zellen der angeborenen Immunität und sind in ECT
Läsionen nach 72 Std überraschenderweise nicht sehr zahlreich vertreten. Weiterhin
weist etwa die Hälfte aller CD14+ Makrophagen in den Läsionen auch eine
Oberflächenexpression von CD1c auf. Es ist gut vorstellbar, dass es sich um die in
humaner Haut beschriebenen DZ Vorläuferzellen handelt, die je nach
Stimulationsmodus in LZ oder DDZ transformiert werden können (TGF-β1: LZ; IL-
4+GM-CSF: DDZ) [37], [122].

Diskussion Ch. Bangert
- 83 -
Schlussendlich spielen T-Zellen eine wesentliche Rolle im KE. CLA+ T-Zellen
patrouillieren die Haut auf der Suche nach Gefahrensignalen. Nach Haptenapplikation
beginnt eine Kaskade von Ereignissen, die im Endeffekt alle das Ziel haben, die T-
Zellen zu aktivieren und somit eine erworbene Immunreaktion in Gang zu setzen, die
auf Basis von Gedächtnis T-Zellen abläuft.
CD4+/CD3+ T-Zellen gelangen bereits wenige Std nach Haptenapplikation in die
‚Gefahrenzone’. Eine lokale Proliferation der Zellen findet nicht statt, sondern sie
werden über CLA/E-Selektin Bindung in die inflammierte Haut gelockt. Erst über 24 Std
später sind zusätzlich CD4+/CD25+ Zellen in der Haut vorhanden. Falls die
CD4+/CD3+ T-Zellen in der Haut von DZ das Hapten präsentiert bekommen, könnten
sie sich im Anschluss in CD4+/CD25+ reg T-Zellen umwandeln. Diese reg T-Zellen
sind wahrscheinlich Hapten-spezifisch und tragen zur Beendigung der Immunantwort
bei.
Zahlenmäßig unterlegen sind die CD8+/CD3+ T-Zellen. Diese Zellen stellen die
Effektorzellen im KE dar [77]. Hapten-spezifische CD8+ T-Zellantworten können nur
bei Allergikern ausgelöst werden. Durch ihre Zytotoxizität werden Keratinozyten in
Apoptose getrieben [108] und die Immunreaktion aggraviert. Auch CD8+/CD3+ T-
Zellen sind bereits nach 24 Std in den ECT Läsionen detektierbar. Eine 2006
veröffentlichte Studie in einem Mausmodell für Kontakthypersensitivität konnte zeigen,
dass es nach Sensibilisierung der Mäuse mit DNFB zur Ausbildung von CD8+
Effektorzellen kommt, die IL-17 produzieren. Neutralisiert man dieses Zytokin, kommt
es zur Unterdrückung des KE [143].
Th17-Zellen spielen eine wichtige Rolle in unterschiedlichen allergisch bedingten oder
Autoimmunerkrankungen, die man bislang T1 oder T2 Zellen zugeschrieben hat. Die
Daten von He et al deuten auf eine Rolle dieser Zellen auch im allergischen KE hin.
Ob die CD8+ T-Zellen im humanen KE ebenfalls IL-17 produzieren, muss leider
unbeantwortet bleiben. In den letzten 20 Jahren haben sich viele Theorien zur
Entstehung des allergischen KE gebildet. Einige konnten im Laufe der Zeit bestätigt
werden, andere wurden verworfen.
Ob die von uns aufgestellte Hypothese der Wahrheit nahe kommt, kann nur in einigen
Folgestudien, die auf das von uns akquirierte Wissen aufbauen, evaluiert werden.

Zusammenfassung Ch. Bangert
- 84 -
6. ZUSAMMENFASSUNG
In der Effektorphase des allergischen KE reagiert das Immunsystem allergen-
spezifisch, mit dem Ziel, das auslösende Hapten durch eine entsprechende
Entzündungsreaktion zu eliminieren. In der frühen Effektorphase dominiert das
Eindringen von Zellen des angeborenen Immunsystems, wie z.B. neutrophilen
Granulozyten und NK-Zellen. Erst im Anschluss kommt es zur Einwanderung von
CLA+, aktivierten Hapten-spezifischen T- Lymphozyten sowie von unspezifisch
rekrutierten Lymphozyten in das betroffene Hautareal. Eine wichtige Rolle in dieser
Phase wird den dendritischen Zellen durch ihre Möglichkeit der Antigenpräsentation
und Verstärkung der Entzündungsreaktion zugedacht.
Während in der Sensibilisierungsphase des allergischen KE der LZ die Rolle der
maßgeblich beteiligten APZ zugewiesen werden konnte, ist die Natur einer solchen
Zelle in der Effektorphase bislang noch nicht eindeutig identifiziert.
Um herauszufinden, ob unterschiedliche Haptene Einfluss auf die quantitative und
qualitative Zusammensetzung des entzündlichen Infiltrats im KE haben, analysierten
wir sowohl Biopsien von positiven ECT auf Ni (n=5) und Duftstoff (n=5) nach 72 Std als
auch von NH (n=5) und stellten Vergleiche hinsichtlich des Phänotyps und der Menge
der infiltrierenden Leukozyten an. Des Weiteren konzentrierten wir uns auf die
Charakterisierung des Infiltrats im ECT zu unterschiedlichen Zeitpunkten und richteten
hierbei unser Augenmerk vor allem auf T-Zellen und DZ, um Hinweise auf die Natur
der zentral involvierten APZ zu erhalten.
Im Gegensatz zur NH zeigten die positiven ECT auf Ni und Duftstoff einen deutlichen
Anstieg der Leukozytenzahl in der Epidermis und Dermis. Detaillierte numerische und
phänotypische Vergleiche der Zusammensetzung des entzündlichen Infiltrats in Ni-
bzw. Duftstoff-Allergikern ergaben keine signifikanten Unterschiede, so dass man
einen einheitlichen Wirkmechanismus vermuten kann, den die Haptene nach Kontakt
in der Effektorphase auslösen. Die meisten der infiltrierenden Leukozyten
repräsentierten T-Zellen oder DZ, allerdings gab es auch ein vermehrtes Vorkommen
von Mastzellen, neutrophilen sowie eosinophilen Granulozyten und NK-Zellen.
Die überwiegende Mehrheit in den positiven Testungen waren T-Zellen, wobei zwei
Drittel aus CD4+ und ein Drittel aus CD8+ T-Zellen bestanden. Fünf Prozent der T-
Zellen zeigten interessanterweise eine positive Reaktivität für CD4/CD25/CTLA-4 und
repräsentierten damit den Phänotyp von aktivierten oder reg T-Zellen. Die kinetischen

Zusammenfassung Ch. Bangert
- 85 -
Analysen der ECT nach 24, 48 und 72 Std zeigten, dass beinahe die Hälfte der
gesamten CD4+/CD3+ T-Zellen bereits innerhalb von 24 Std einwandern, während
CD4+/CD25+ T-Zellen erst nach 24 Std im Infiltrat erscheinen. Dies könnte bedeuten,
dass CD4+/CD25+ Zellen sich entweder aus den bereits eingewanderten Zellen
entwickeln oder tatsächlich zu einem späteren Zeitpunkt in die Läsion einwandern.
Die dominante Zellpopulation unter den APZ in der Dermis läsionaler Haut waren
CD1c+ DDZ. LZ zeigten eine Abnahme in der Epidermis und eine Zunahme in der
Dermis von ECT im Vergleich zur NH. Dies lässt sich mit der Wanderung der LZ zum
regionären LK nach ihrer Haptenaufnahme erklären. Überraschenderweise
identifizierten wir eine große Anzahl CD123+/BDCA-2+/CD45RA+ pDZ in der Dermis
läsionaler Haut. Während sie in der NH nicht detektierbar waren, zeigte sich bereits 6
Std nach Haptenkontakt eine Infiltration der Zellen in die Dermis. Eine lokale
Proliferation von pDZ konnten wir durch Färbungen mit einem Proliferationsmarker (Ki-
67) ausschließen.
Durchaus interessant war auch die Tatsache, dass NK-Zellen als Zellen des
angeborenen Immunsystems ebenfalls bereits nach 6 Std in ECT zu finden und häufig
in der Nähe von pDZ angesiedelt waren. Eine Interaktion dieser beiden Zellarten wäre
denkbar, da pDZ IFN-α und TNF-α produzieren, dadurch NK-Zellen aktiviert und zur
Produktion von IFN-γ angeregt werden können.
Unsere Studie gibt eine umfassende und detaillierte Übersicht über das entzündliche
Infiltrat und dessen Kinetik in der Effektorphase des KE. Dieses Wissen bildet die Basis
für weitere Untersuchungen, in denen die Funktion der unterschiedlichen Zellarten
ermittelt werden sollte, um so endgültig die Mechanismen verstehen zu können, die für
das Entstehen und die Auflösung des KE verantwortlich sind.

Literatur Ch.Bangert
- 86 -
7. LITERATURVERZEICHNIS
1. Fritsch, P., Kontaktekzem. Lehrbuch für Dermatologie und Venerologie. 1998,
Berlin Springer.
2. Belsito, D.V., Allergic Contact Dermatitis. Fitzpatrick's Dermatology in General
Medicine. Vol. 1. 1999, New York: I. M. Freedberg, A. Z. Eisen, K. Wolff et al. .
1464-1480.
3. Landsteiner, K., Jacobs, J., Studies on the sensitization of animals with simple
chemical compounds. The Journal of Experimental Medicine, 1935. 61: p. 643-
656.
4. Ring, J., Allergy in Practice. Allergic Diseases (and Differential Diagnosis). 2005
Berlin: Springer.
5. White, C.R., Jr., Histopathology of exogenous and systemic contact eczema.
Semin Dermatol, 1990. 9(3): p. 226-9.
6. Phelps, R.G., M.K. Miller, and F. Singh, The varieties of "eczema":
clinicopathologic correlation. Clin Dermatol, 2003. 21(2): p. 95-100.
7. Eisen, H.N., L. Orris, and S. Belman, Elicitation of delayed allergic skin
reactions with haptens; the dependence of elicitation on hapten combination
with protein. J Exp Med, 1952. 95(5): p. 473-87.
8. Kapsenberg, M.L., Chemicals and proteins as allergens and adjuvants. Toxicol
Lett, 1996. 86(2-3): p. 79-83.
9. Schafer, T., et al., Epidemiology of contact allergy in adults. Allergy, 2001.
56(12): p. 1192-6.
10. Budinger, L., et al., Preferential usage of TCR-Vbeta17 by peripheral and
cutaneous T cells in nickel-induced contact dermatitis. J Immunol, 2001.
167(10): p. 6038-44.
11. Lu, L., et al., Components of the ligand for a Ni++ reactive human T cell clone. J
Exp Med, 2003. 197(5): p. 567-74.
12. Thierse, H.J., et al., Metal-protein complex-mediated transport and delivery of
Ni2+ to TCR/MHC contact sites in nickel-specific human T cell activation. J
Immunol, 2004. 172(3): p. 1926-34.
13. Thierse, H.J., et al., T cell receptor (TCR) interaction with haptens: metal ions
as non-classical haptens. Toxicology, 2005. 209(2): p. 101-7.
14. Moulon, C., J. Vollmer, and H.U. Weltzien, Characterization of processing
requirements and metal cross-reactivities in T cell clones from patients with
allergic contact dermatitis to nickel. Eur J Immunol, 1995. 25(12): p. 3308-15.

Literatur Ch.Bangert
- 87 -
15. Sieben, S., et al., Characterization of T cell responses to fragrances. Toxicol
Appl Pharmacol, 2001. 172(3): p. 172-8.
16. Buckley, D.A., et al., Simultaneous sensitivity to fragrances. Br J Dermatol,
2006. 154(5): p. 885-8.
17. Jadassohn, J., Verhandl Dtsch Derm Gesellschaft, 1896. 5. Kongreß
18. Langerhans, P., Über die Nerven der menschlichen Haut. Virchows Arch, 1868.
44: p. 325-337.
19. Wolff, K., The langerhans cell. Curr Probl Dermatol, 1972. 4: p. 79-145.
20. Seiji, M., T.B. Fitzpatrick, and M.S. Birbeck, The melanosome: a distinctive
subcellular particle of mammalian melanocytes and the site of melanogenesis. J
Invest Dermatol, 1961. 36: p. 243-52.
21. Wolff, K., [Histological observations on normal human skin following enzyme-
histochemical studies with adenosine triphosphate as substrate.]. Arch Klin Exp
Dermatol, 1963. 216: p. 1-17.
22. Mustakallio, K., Adenosine triphosphatase activity in neural elements of human
epidermis. Exp Cell Res, 1962. 28: p. 449-51.
23. Rowden, G., M.G. Lewis, and A.K. Sullivan, Ia antigen expression on human
epidermal Langerhans cells. Nature, 1977. 268(5617): p. 247-8.
24. Klareskog, L., et al., Epidermal Langerhans cells express Ia antigens. Nature,
1977. 268(5617): p. 248-50.
25. Stingl, G., et al., Epidermal Langerhans cells bear Fc and C3 receptors. Nature,
1977. 268(5617): p. 245-6.
26. Stingl, G., et al., Immunologic functions of Ia-bearing epidermal Langerhans
cells. J Immunol, 1978. 121(5): p. 2005-13.
27. Green, I., et al., Antigen presentation and allogeneic stimulation by Langerhans
cells. J Invest Dermatol, 1980. 75(1): p. 44-5.
28. Braathen, L.R. and E. Thorsby, Studies on human epidermal Langerhans cells.
I. Allo-activating and antigen-presenting capacity. Scand J Immunol, 1980.
11(4): p. 401-8.
29. Bjercke, S., et al., Enriched epidermal Langerhans cells are potent antigen-
presenting cells for T cells. J Invest Dermatol, 1984. 83(4): p. 286-9.
30. Volc-Platzer, B., et al., Cytogenetic identification of allogeneic epidermal
Langerhans cells in a bone-marrow-graft recipient. N Engl J Med, 1984.
310(17): p. 1123-4.
31. Caux, C., et al., GM-CSF and TNF-alpha cooperate in the generation of
dendritic Langerhans cells. Nature, 1992. 360(6401): p. 258-61.

Literatur Ch.Bangert
- 88 -
32. Caux, C., et al., CD34+ hematopoietic progenitors from human cord blood
differentiate along two independent dendritic cell pathways in response to GM-
CSF+TNF alpha. J Exp Med, 1996. 184(2): p. 695-706.
33. Elbe, A., et al., Maturational steps of bone marrow-derived dendritic murine
epidermal cells. Phenotypic and functional studies on Langerhans cells and
Thy-1+ dendritic epidermal cells in the perinatal period. J Immunol, 1989.
143(8): p. 2431-8.
34. Foster, C.A. and K.A. Holbrook, Ontogeny of Langerhans cells in human
embryonic and fetal skin: cell densities and phenotypic expression relative to
epidermal growth. Am J Anat, 1989. 184(2): p. 157-64.
35. Schuler, G. and R.M. Steinman, Murine epidermal Langerhans cells mature into
potent immunostimulatory dendritic cells in vitro. J Exp Med, 1985. 161(3): p.
526-46.
36. Lenz, A., et al., Human and murine dermis contain dendritic cells. Isolation by
means of a novel method and phenotypical and functional characterization. J
Clin Invest, 1993. 92(6): p. 2587-96.
37. Jaksits, S., et al., CD34+ cell-derived CD14+ precursor cells develop into
Langerhans cells in a TGF-beta 1-dependent manner. J Immunol, 1999. 163(9):
p. 4869-77.
38. Valladeau, J., et al., Langerin, a novel C-type lectin specific to Langerhans
cells, is an endocytic receptor that induces the formation of Birbeck granules.
Immunity, 2000. 12(1): p. 71-81.
39. Geijtenbeek, T.B., et al., Identification of DC-SIGN, a novel dendritic cell-
specific ICAM-3 receptor that supports primary immune responses. Cell, 2000.
100(5): p. 575-85.
40. Figdor, C.G., Y. van Kooyk, and G.J. Adema, C-type lectin receptors on
dendritic cells and Langerhans cells. Nat Rev Immunol, 2002. 2(2): p. 77-84.
41. van Kooyk, Y. and T.B. Geijtenbeek, DC-SIGN: escape mechanism for
pathogens. Nat Rev Immunol, 2003. 3(9): p. 697-709.
42. Kissenpfennig, A., et al., Disruption of the langerin/CD207 gene abolishes
Birbeck granules without a marked loss of Langerhans cell function. Mol Cell
Biol, 2005. 25(1): p. 88-99.
43. Roake, J.A., et al., Dendritic cell loss from nonlymphoid tissues after systemic
administration of lipopolysaccharide, tumor necrosis factor, and interleukin 1. J
Exp Med, 1995. 181(6): p. 2237-47.
44. Kucharzik, T., et al., CCR6 expression distinguishes mouse myeloid and
lymphoid dendritic cell subsets: demonstration using a CCR6 EGFP knock-in
mouse. Eur J Immunol, 2002. 32(1): p. 104-12.

Literatur Ch.Bangert
- 89 -
45. Schwarzenberger, K. and M.C. Udey, Contact allergens and epidermal
proinflammatory cytokines modulate Langerhans cell E-cadherin expression in
situ. J Invest Dermatol, 1996. 106(3): p. 553-8.
46. Ratzinger, G., et al., Matrix metalloproteinases 9 and 2 are necessary for the
migration of Langerhans cells and dermal dendritic cells from human and
murine skin. J Immunol, 2002. 168(9): p. 4361-71.
47. Weiss, J.M., et al., Osteopontin is involved in the initiation of cutaneous contact
hypersensitivity by inducing Langerhans and dendritic cell migration to lymph
nodes. J Exp Med, 2001. 194(9): p. 1219-29.
48. Shornick, L.P., et al., Mice deficient in IL-1beta manifest impaired contact
hypersensitivity to trinitrochlorobenzone. J Exp Med, 1996. 183(4): p. 1427-36.
49. Shornick, L.P., A.K. Bisarya, and D.D. Chaplin, IL-1beta is essential for
langerhans cell activation and antigen delivery to the lymph nodes during
contact sensitization: evidence for a dermal source of IL-1beta. Cell Immunol,
2001. 211(2): p. 105-12.
50. Grabbe, S., et al., Removal of the majority of epidermal Langerhans cells by
topical or systemic steroid application enhances the effector phase of murine
contact hypersensitivity. J Immunol, 1995. 155(9): p. 4207-17.
51. Silberberg, I., R.L. Baer, and S.A. Rosenthal, The role of Langerhans cells in
allergic contact hypersensitivity. A review of findings in man and guinea pigs. J
Invest Dermatol, 1976. 66(4): p. 210-7.
52. Romani, N. and G. Schuler, The immunologic properties of epidermal
Langerhans cells as a part of the dendritic cell system. Springer Semin
Immunopathol, 1992. 13(3-4): p. 265-79.
53. Nestle, F.O., et al., Characterization of dermal dendritic cells obtained from
normal human skin reveals phenotypic and functionally distinctive subsets. J
Immunol, 1993. 151(11): p. 6535-45.
54. Nestle, F.O., et al., Human dermal dendritic cells process and present soluble
protein antigens. J Invest Dermatol, 1998. 110(5): p. 762-6.
55. Kissenpfennig, A., et al., Dynamics and function of Langerhans cells in vivo:
dermal dendritic cells colonize lymph node areas distinct from slower migrating
Langerhans cells. Immunity, 2005. 22(5): p. 643-54.
56. Valladeau, J. and S. Saeland, Cutaneous dendritic cells. Semin Immunol, 2005.
17(4): p. 273-83.
57. Wollenberg, A., et al., Immunomorphological and ultrastructural characterization
of Langerhans cells and a novel, inflammatory dendritic epidermal cell (IDEC)
population in lesional skin of atopic eczema. J Invest Dermatol, 1996. 106(3): p.
446-53.

Literatur Ch.Bangert
- 90 -
58. Novak, N., et al., FcepsilonRI engagement of Langerhans cell-like dendritic
cells and inflammatory dendritic epidermal cell-like dendritic cells induces
chemotactic signals and different T-cell phenotypes in vitro. J Allergy Clin
Immunol, 2004. 113(5): p. 949-57.
59. Wollenberg, A., et al., Expression and function of the mannose receptor CD206
on epidermal dendritic cells in inflammatory skin diseases. J Invest Dermatol,
2002. 118(2): p. 327-34.
60. Lennert, K. and W. Remmele, [Karyometric research on lymph node cells in
man. I. Germinoblasts, lymphoblasts & lymphocytes.]. Acta Haematol, 1958.
19(2): p. 99-113.
61. Trinchieri, G., et al., Anti-viral activity induced by culturing lymphocytes with
tumor-derived or virus-transformed cells. Identification of the anti-viral activity as
interferon and characterization of the human effector lymphocyte subpopulation.
J Exp Med, 1978. 147(5): p. 1299-1313.
62. Siegal, F.P., et al., The nature of the principal type 1 interferon-producing cells
in human blood. Science, 1999. 284(5421): p. 1835-7.
63. Cella, M., et al., Plasmacytoid monocytes migrate to inflamed lymph nodes and
produce large amounts of type I interferon. Nat Med, 1999. 5(8): p. 919-23.
64. Dzionek, A., et al., BDCA-2, a novel plasmacytoid dendritic cell-specific type II
C-type lectin, mediates antigen capture and is a potent inhibitor of interferon
alpha/beta induction. J Exp Med, 2001. 194(12): p. 1823-34.
65. Dzionek, A., et al., Plasmacytoid dendritic cells: from specific surface markers
to specific cellular functions. Hum Immunol, 2002. 63(12): p. 1133-48.
66. Blom, B., et al., Generation of interferon alpha-producing predendritic cell (Pre-
DC)2 from human CD34(+) hematopoietic stem cells. J Exp Med, 2000.
192(12): p. 1785-96.
67. Facchetti, F., et al., The plasmacytoid monocyte/interferon producing cells.
Virchows Arch, 2003. 443(6): p. 703-17.
68. Farkas, L., et al., Plasmacytoid dendritic cells (natural interferon- alpha/beta-
producing cells) accumulate in cutaneous lupus erythematosus lesions. Am J
Pathol, 2001. 159(1): p. 237-43.
69. Preble, O.T., et al., Systemic lupus erythematosus: presence in human serum
of an unusual acid-labile leukocyte interferon. Science, 1982. 216(4544): p.
429-31.
70. Bell, D., et al., In breast carcinoma tissue, immature dendritic cells reside within
the tumor, whereas mature dendritic cells are located in peritumoral areas. J
Exp Med, 1999. 190(10): p. 1417-26.

Literatur Ch.Bangert
- 91 -
71. Vermi, W., et al., Recruitment of immature plasmacytoid dendritic cells
(plasmacytoid monocytes) and myeloid dendritic cells in primary cutaneous
melanomas. J Pathol, 2003. 200(2): p. 255-68.
72. Kapsenberg, M.L., et al., Nickel-specific T lymphocyte clones derived from
allergic nickel-contact dermatitis lesions in man: heterogeneity based on
requirement of dendritic antigen-presenting cell subsets. Eur J Immunol, 1987.
17(6): p. 861-5.
73. Kapsenberg, M.L., et al., Th1 lymphokine production profiles of nickel-specific
CD4+T-lymphocyte clones from nickel contact allergic and non-allergic
individuals. J Invest Dermatol, 1992. 98(1): p. 59-63.
74. Sinigaglia, F., et al., Isolation and characterization of Ni-specific T cell clones
from patients with Ni-contact dermatitis. J Immunol, 1985. 135(6): p. 3929-32.
75. Cher, D.J. and T.R. Mosmann, Two types of murine helper T cell clone. II.
Delayed-type hypersensitivity is mediated by TH1 clones. J Immunol, 1987.
138(11): p. 3688-94.
76. Brasch, J., J. Burgard, and W. Sterry, Common pathogenetic pathways in
allergic and irritant contact dermatitis. J Invest Dermatol, 1992. 98(2): p. 166-70.
77. Kehren, J., et al., Cytotoxicity is mandatory for CD8(+) T cell-mediated contact
hypersensitivity. J Exp Med, 1999. 189(5): p. 779-86.
78. Cavani, A., et al., Patients with allergic contact dermatitis to nickel and
nonallergic individuals display different nickel-specific T cell responses.
Evidence for the presence of effector CD8+ and regulatory CD4+ T cells. J
Invest Dermatol, 1998. 111(4): p. 621-8.
79. Xu, H., N.A. DiIulio, and R.L. Fairchild, T cell populations primed by hapten
sensitization in contact sensitivity are distinguished by polarized patterns of
cytokine production: interferon gamma-producing (Tc1) effector CD8+ T cells
and interleukin (Il) 4/Il-10-producing (Th2) negative regulatory CD4+ T cells. J
Exp Med, 1996. 183(3): p. 1001-12.
80. Kolesaric, A., G. Stingl, and A. Elbe-Burger, MHC class I+/II- dendritic cells
induce hapten-specific immune responses in vitro and in vivo. J Invest
Dermatol, 1997. 109(4): p. 580-5.
81. Probst, P., D. Kuntzlin, and B. Fleischer, TH2-type infiltrating T cells in nickel-
induced contact dermatitis. Cell Immunol, 1995. 165(1): p. 134-40.
82. Cavani, A., et al., Human CD4+ T lymphocytes with remarkable regulatory
functions on dendritic cells and nickel-specific Th1 immune responses. J Invest
Dermatol, 2000. 114(2): p. 295-302.

Literatur Ch.Bangert
- 92 -
83. Werfel, T., et al., Dichotomy of blood- and skin-derived IL-4-producing allergen-
specific T cells and restricted V beta repertoire in nickel-mediated contact
dermatitis. J Immunol, 1997. 158(5): p. 2500-5.
84. Saint-Mezard, P., et al., The role of CD4+ and CD8+ T cells in contact
hypersensitivity and allergic contact dermatitis. Eur J Dermatol, 2004. 14(3): p.
131-8.
85. Jonuleit, H., et al., Identification and functional characterization of human
CD4(+)CD25(+) T cells with regulatory properties isolated from peripheral
blood. J Exp Med, 2001. 193(11): p. 1285-94.
86. Dieckmann, D., et al., Ex vivo isolation and characterization of CD4(+)CD25(+)
T cells with regulatory properties from human blood. J Exp Med, 2001. 193(11):
p. 1303-10.
87. Thornton, A.M. and E.M. Shevach, CD4+CD25+ immunoregulatory T cells
suppress polyclonal T cell activation in vitro by inhibiting interleukin 2
production. J Exp Med, 1998. 188(2): p. 287-96.
88. Fontenot, J.D., M.A. Gavin, and A.Y. Rudensky, Foxp3 programs the
development and function of CD4+CD25+ regulatory T cells. Nat Immunol,
2003. 4(4): p. 330-6.
89. Walker, M.R., et al., Induction of FoxP3 and acquisition of T regulatory activity
by stimulated human CD4+CD25- T cells. J Clin Invest, 2003. 112(9): p. 1437-
43.
90. Huber, H., et al., Activation of murine epidermal TCR-gamma delta+ T cells by
keratinocytes treated with contact sensitizers. J Immunol, 1995. 155(6): p.
2888-94.
91. Schild, H., et al., The nature of major histocompatibility complex recognition by
gamma delta T cells. Cell, 1994. 76(1): p. 29-37.
92. Askenase, P.W., et al., Gamma delta T cells in normal spleen assist immunized
alpha beta T cells in the adoptive cell transfer of contact sensitivity. Effect of
Bordetella pertussis, cyclophosphamide, and antibodies to determinants on
suppressor cells. J Immunol, 1995. 154(8): p. 3644-53.
93. Guan, H., et al., GammadeltaT cells regulate the development of hapten-
specific CD8+ effector T cells in contact hypersensitivity responses. J Invest
Dermatol, 2002. 119(1): p. 137-42.
94. Cruz, P.D., Jr., Antigen-presenting cells in the induction of contact
hypersensitivity in mice: evidence that Langerhans cells are sufficient but not
required. J Invest Dermatol, 1990. 94(3): p. 386-8.
95. Hauser, C., Cultured epidermal Langerhans cells activate effector T cells for
contact sensitivity. J Invest Dermatol, 1990. 95(4): p. 436-40.

Literatur Ch.Bangert
- 93 -
96. Becker, D. and J. Knop, Mechanism in allergic contact dermatitis. Exp
Dermatol, 1993. 2(2): p. 63-9.
97. Singh, S., et al., The effects of dexamethasone, cyclosporine, and vitamin D(3)
on the activation of dendritic cells stimulated by haptens. Arch Dermatol Res,
1999. 291(10): p. 548-54.
98. Enk, A.H. and S.I. Katz, Early molecular events in the induction phase of
contact sensitivity. Proc Natl Acad Sci U S A, 1992. 89(4): p. 1398-402.
99. Zheng, H., et al., Resistance to fever induction and impaired acute-phase
response in interleukin-1 beta-deficient mice. Immunity, 1995. 3(1): p. 9-19.
100. Sallusto, F., et al., Two subsets of memory T lymphocytes with distinct homing
potentials and effector functions. Nature, 1999. 401(6754): p. 708-12.
101. Santamaria, L.F., et al., Allergen specificity and endothelial transmigration of T
cells in allergic contact dermatitis and atopic dermatitis are associated with the
cutaneous lymphocyte antigen. Int Arch Allergy Immunol, 1995. 107(1-3): p.
359-62.
102. Barker, J.N., et al., Keratinocytes as initiators of inflammation. Lancet, 1991.
337(8735): p. 211-4.
103. Biedermann, T., et al., Mast cells control neutrophil recruitment during T cell-
mediated delayed-type hypersensitivity reactions through tumor necrosis factor
and macrophage inflammatory protein 2. J Exp Med, 2000. 192(10): p. 1441-52.
104. Kalish, R.S., Recent developments in the pathogenesis of allergic contact
dermatitis. Arch Dermatol, 1991. 127(10): p. 1558-63.
105. Lewis, R.E., et al., Intercellular adhesion molecule expression in the evolving
human cutaneous delayed hypersensitivity reaction. J Invest Dermatol, 1989.
93(5): p. 672-7.
106. Traidl, C., et al., Disparate cytotoxic activity of nickel-specific CD8+ and CD4+ T
cell subsets against keratinocytes. J Immunol, 2000. 165(6): p. 3058-64.
107. Akiba, H., et al., Skin inflammation during contact hypersensitivity is mediated
by early recruitment of CD8+ T cytotoxic 1 cells inducing keratinocyte
apoptosis. J Immunol, 2002. 168(6): p. 3079-87.
108. Trautmann, A., et al., T cell-mediated Fas-induced keratinocyte apoptosis plays
a key pathogenetic role in eczematous dermatitis. J Clin Invest, 2000. 106(1): p.
25-35.
109. Trautmann, A., et al., New insights into the role of T cells in atopic dermatitis
and allergic contact dermatitis. Trends Immunol, 2001. 22(10): p. 530-2.
110. Gorbachev, A.V. and R.L. Fairchild, CD4+ T cells regulate CD8+ T cell-
mediated cutaneous immune responses by restricting effector T cell

Literatur Ch.Bangert
- 94 -
development through a Fas ligand-dependent mechanism. J Immunol, 2004.
172(4): p. 2286-95.
111. Dubois, B., et al., CD4+CD25+ T cells as key regulators of immune responses.
Eur J Dermatol, 2003. 13(2): p. 111-6.
112. Dubois, B., et al., Innate CD4+CD25+ regulatory T cells are required for oral
tolerance and inhibition of CD8+ T cells mediating skin inflammation. Blood,
2003. 102(9): p. 3295-301.
113. Saint-Mezard, P., et al., Allergic contact dermatitis. Eur J Dermatol, 2004. 14(5):
p. 284-95.
114. Wang, B., et al., CD4+ Th1 and CD8+ type 1 cytotoxic T cells both play a
crucial role in the full development of contact hypersensitivity. J Immunol, 2000.
165(12): p. 6783-90.
115. Gocinski, B.L. and R.E. Tigelaar, Roles of CD4+ and CD8+ T cells in murine
contact sensitivity revealed by in vivo monoclonal antibody depletion. J
Immunol, 1990. 144(11): p. 4121-8.
116. Wollenberg, A., et al., Plasmacytoid dendritic cells: a new cutaneous dendritic
cell subset with distinct role in inflammatory skin diseases. J Invest Dermatol,
2002. 119(5): p. 1096-102.
117. Picker, L.J., et al., ELAM-1 is an adhesion molecule for skin-homing T cells.
Nature, 1991. 349(6312): p. 796-9.
118. Harari, O.A., et al., Endothelial cell E- and P-selectin up-regulation in murine
contact sensitivity is prolonged by distinct mechanisms occurring in sequence. J
Immunol, 1999. 163(12): p. 6860-6.
119. Sebastiani, S., et al., The role of chemokines in allergic contact dermatitis. Arch
Dermatol Res, 2002. 293(11): p. 552-9.
120. Tsuji, R.F., et al., Required early complement activation in contact sensitivity
with generation of local C5-dependent chemotactic activity, and late T cell
interferon gamma: a possible initiating role of B cells. J Exp Med, 1997. 186(7):
p. 1015-26.
121. Tsuji, R.F., et al., Early local generation of C5a initiates the elicitation of contact
sensitivity by leading to early T cell recruitment. J Immunol, 2000. 165(3): p.
1588-98.
122. Larregina, A.T., et al., Dermal-resident CD14+ cells differentiate into
Langerhans cells. Nat Immunol, 2001. 2(12): p. 1151-8.
123. Cumberbatch, M., et al., Epidermal Langerhans cell migration and sensitisation
to chemical allergens. Apmis, 2003. 111(7-8): p. 797-804.
124. Cumberbatch, M., et al., Tumour necrosis factor-alpha induces Langerhans cell
migration in humans. Br J Dermatol, 1999. 141(2): p. 192-200.

Literatur Ch.Bangert
- 95 -
125. Cella, M., F. Sallusto, and A. Lanzavecchia, Origin, maturation and antigen
presenting function of dendritic cells. Curr Opin Immunol, 1997. 9(1): p. 10-6.
126. Jonuleit, H., et al., Dendritic cells as a tool to induce anergic and regulatory T
cells. Trends Immunol, 2001. 22(7): p. 394-400.
127. Krug, A., et al., IFN-producing cells respond to CXCR3 ligands in the presence
of CXCL12 and secrete inflammatory chemokines upon activation. J Immunol,
2002. 169(11): p. 6079-83.
128. Lechleitner, S., et al., Peripheral lymph node addressins are expressed on skin
endothelial cells. J Invest Dermatol, 1999. 113(3): p. 410-4.
129. Kohrgruber, N., et al., Survival, maturation, and function of CD11c- and CD11c+
peripheral blood dendritic cells are differentially regulated by cytokines. J
Immunol, 1999. 163(6): p. 3250-9.
130. Cella, M., et al., Plasmacytoid dendritic cells activated by influenza virus and
CD40L drive a potent TH1 polarization. Nat Immunol, 2000. 1(4): p. 305-10.
131. Steinman, R.M., D. Hawiger, and M.C. Nussenzweig, Tolerogenic dendritic
cells. Annu Rev Immunol, 2003. 21: p. 685-711.
132. Rissoan, M.C., et al., Reciprocal control of T helper cell and dendritic cell
differentiation. Science, 1999. 283(5405): p. 1183-6.
133. Kadowaki, N., et al., Natural interferon alpha/beta-producing cells link innate
and adaptive immunity. J Exp Med, 2000. 192(2): p. 219-26.
134. Gilliet, M. and Y.J. Liu, Generation of human CD8 T regulatory cells by CD40
ligand-activated plasmacytoid dendritic cells. J Exp Med, 2002. 195(6): p. 695-
704.
135. Sterry, W., et al., Dominance of memory over naive T cells in contact dermatitis
is due to differential tissue immigration. Br J Dermatol, 1990. 123(1): p. 59-64.
136. Shevach, E.M., CD4+ CD25+ suppressor T cells: more questions than answers.
Nat Rev Immunol, 2002. 2(6): p. 389-400.
137. Luger, T.A. and T. Schwarz, Evidence for an epidermal cytokine network. J
Invest Dermatol, 1990. 95(6 Suppl): p. 100S-104S.
138. Buentke, E., et al., Natural killer and dendritic cell contact in lesional atopic
dermatitis skin--Malassezia-influenced cell interaction. J Invest Dermatol, 2002.
119(4): p. 850-7.
139. Moretta, A., Natural killer cells and dendritic cells: rendezvous in abused
tissues. Nat Rev Immunol, 2002. 2(12): p. 957-64.
140. Skoog, M.L., Measurement and differentiation of the cellular infiltrate in
experimental toxic contact dermatitis. Acta Derm Venereol, 1980. 60(3): p. 239-
44.

Literatur Ch.Bangert
- 96 -
141. Sjogren, F. and C. Anderson, The spectrum of inflammatory cell response to
dimethyl sulfoxide. Contact Dermatitis, 2000. 42(4): p. 216-21.
142. Geissmann, F., et al., Accumulation of immature Langerhans cells in human
lymph nodes draining chronically inflamed skin. J Exp Med, 2002. 196(4): p.
417-30.
143. He, D., et al., CD8+ IL-17-producing T cells are important in effector functions
for the elicitation of contact hypersensitivity responses. J Immunol, 2006.
177(10): p. 6852-8.

Danksagung Ch. Bangert
- 97 -
8. DANKSAGUNG Mein Dank richtet sich in erster Linie an Herrn Prof. Dr. Georg Stingl, Vorstand der
Abteilung für Immundermatologie und Infektiöse Hautkrankheiten an der Klinik für
Dermatologie an der Medizinischen Universität Wien für die Ideengebung und
Überlassung des Themas sowie die hervorragende wissenschaftliche Betreuung,
Unterstützung, Freundschaft und große Geduld während der Anfertigung der
vorliegenden Arbeit.
Besonders danke ich auch Frau Prof. Dr. Tamara Kopp, Abteilung für
Immundermatologie, Klinik für Dermatologie, Medizinischen Universität Wien, für die
kompetente wissenschaftliche Betreuung und Ideengebung, die Lehre
unterschiedlichster wissenschaftlicher Techniken, die Hilfe bei der statistischen
Auswertung und vor allem auch für ihre konstante Freundschaft und Kollegialität
während der letzten Jahre.
Außerdem bedanke ich mich bei Herrn Prof. Dr. Jürgen Grabbe für die freundliche
Übernahme der Dissertationsbetreuung.
Für die sehr gute Zusammenarbeit möchte ich Herrn Dr. Georg Stary und Frau Sabine
Kohlhofer, Abteilung für Immundermatologie, Klinik für Dermatologie, Medizinischen
Universität Wien, danken.
Weiterhin war der wissenschaftliche Austausch mit Dr. Ernst Kriehuber und Prof. Dieter
Maurer, Abteilung für Immundermatologie, Klinik für Dermatologie, Medizinischen
Universität Wien, bei der Fertigstellung dieser Arbeit von großer Bedeutung.
Für die Darstellung und Anfertigungen der Abbildungen sowie für die kompetente
Beratung bei auftretenden Problemen im EDV Bereich möchte ich Herrn Andreas
Ebner danken.
Meinen Eltern danke ich besonders für die seelische Unterstützung und das allzeit
vorhandene Verständnis.

Lebenslauf Ch. Bangert
- 98 -
9. LEBENSLAUF Persönliche Daten:
Name Christine Bangert
Geburtsort Mosbach, Deutschland
Nationalität Deutsch
Schulische Ausbildung:
1980-1984 Müller-Guttenbrunn Grundschule, Mosbach
1984-1993 Nikolaus-Kistner Gymnasium, Mosbach
10.05.1993 Abitur
Studium:
09/93-11/2000 Studium der Humanmedizin an der Medizinischen
Universität zu Lübeck
14.11.2000 Beendigung des Studiums mit dem 3. Staatsexamen
bis 09/2001 Promotionssemester
04/2002-09/2003 AiP (Arzt im Praktikum), Klinik für Dermatologie,
Medizinische Universität Wien, Österreich
05.09.2003 Approbation
Berufliche Ausbildung:
02/2001-09/2003: wissenschaftliche Mitarbeiterin, Klinik für Dermatologie, AKH,
Medizinische Universität Wien, Österreich
seit 09/2003 Assistenzärztin, Klinik für Dermatologie, AKH, Medizinische
Universität Wien, Österreich
Publikationen:
Bangert C , Friedl J, Stary G, Stingl G, Kopp T: Immunopathologic features of allergic
contact dermatitis in man: participation of plasmacytoid dendritic cells in the
pathogenesis of the disease? (J. Invest Dermatol, 121:1409-1418; 2003)

Lebenslauf Ch. Bangert
- 99 -
Stary G, Bangert C , Stingl G, Kopp T: Dendritic cells in atopic dermatitis: expression of
FcepsilonRI on two distinct inflammation-associated subsets. (Int Arch Allergy
Immunol, 138:278-290; 2005)
Streit M, Ecker RC, Osterreicher K, Steiner GE, Bischof H, Bangert C , Kopp T,
Rogojanu R: 3D parallel coordinate systems-A new data visualization method in the
context of microscopy-based multicolor tissue cytometry. (Cytometry A 69:601-611;
2006)
Stary G, Bangert C , Tauber M, Strohal R, Kopp T, Stingl G: Tumoricidal activity of
TLR7/8 –activated inflammatory dendritic cells. (J Ex Med, 204:1441-1451; 2007)
Stary G, Meyer T, Bangert C , Kohrgruber N, Gmeinhart B, Kirnbauer R, Jantschitsch
C, Rieger A, Stary A, Geusau A: New chlamydia trachomatis L2 strains identified in a
recent outbreak of lymphogranuloma venereum in Vienna, Austria. (Sex Transm Dis,
35: 377-382; 2008)
C. Bangert , B. E. Strober, M. Cork, J.-P. Ortonne, T. Luger, T. Bieber, A. Ferguson,
R.C. Ecker, T.Kopp, S. Weise-Riccardi, A. Guettner, G. Stingl: Clinical and cytological
effects of pimecrolimus cream 1% after resolution of active atopic dermatitis lesions by
topical corticosteroids: a randomized trial (eingereicht im März 2009)
Buchkapitel:
Fitzpatrick’s Dermatology in General Medicine, 7th Edition; Klaus Wolff, Lowell A.
Goldsmith, Stephen I. Katz, Barbara A. Gilchrest, Amy S. Paller, David J. Leffell
Part 2. Disorders presenting in skin and mucous membranes
Section 4, Chapter 10: Innate and adaptive immunity in the skin
Robert L.Modlin, Jenny Kim, Dieter Maurer, Christine Bangert, Georg Stingl
Präsentationen
C. Bangert , G. Stingl, T. Kopp: CD123+/CD4+ cells – a predominant dendritic cell
population in allergic contact dermatitis (Poster, 32nd Annual Meeting der ESDR,
September 2002, Genf, Schweiz)

Lebenslauf Ch. Bangert
- 100 -
C. Bangert , G. Stingl, T. Kopp: CD123+/CD4+ cells – a predominant population in
allergic contact dermatitis (Poster, 7th International Symposium on Dendritic Cells,
September 2002, Bamberg, Deutschland)
C. Bangert , G. Stingl, T. Kopp: Detection of BDCA-2+CD123+MHCII+ plasmacytoid
dendritic cells in allergic contact dermatitis. (Poster, XXX. Jahrestagung der ADF,
Februar 2003, Frankfurt, Deutschland)
C. Bangert , G. Stingl, T. Kopp: Dendritic cells in allergic contact dermatitis-the
widening spectrum. (Poster, 4. gemeinsamer Jahrestag der ESDR, JSID und SID, Mai
2003, Miami Beach Florida, U.S.A.)
C. Bangert, G. Stary, G. Stingl, T. Kopp: Preferential occurrence of different
inflammatory dendritic cell subsets in contact hypersensitivity and atopic dermatitis
(Poster, 8th International Workshop on Langerhans Cells, September 2003, Tokyo,
Japan)
C. Bangert, S. Graffi, S. Altrichter, G. Stingl, T. Kopp: Plasmacytoid dendritic cells
induce hapten-specific T-cell proliferation (Poster, 65th Meeting of the SID, April 2004,
Providence, Rhode Island, USA)
C. Bangert, S. Graffi, S. Altrichter, G. Stingl, T. Kopp: Mature, but not immature
plasmacytoid dendritic cells induce hapten-specific T-cell proliferation (Poster, 34th
Annual ESDR Meeting, September 2004, Wien, Österreich)
C. Bangert, S. Altrichter, G. Stary, G. Stingl, T. Kopp: Presence of lytic proteins in
myeloid and plasmacytoid dendritic cells (Poster, 9th International Workshop on
Langerhans Cells, September 2005, Funchal, Madeira)
N. Kohrgruber, C. Bangert , P. Mastan, G. Stingl, A. Rieger: Bazilläre Angiomatose bei
einem HIV-infizierten Patienten (Vortrag, Jahrestagung 2005 der Österreichischen
Gesellschaft für Dermatologie und Venerologie, November 2005, Wien, Österreich)
C. Bangert, S. Altrichter, G. Stary, G. Stingl, T. Kopp: Presence of lytic proteins in
myeloid and plasmacytoid dendritic cells (Poster, 35th Annual ESDR Meeting,
September 2005, Tübingen, Deutschland)

Lebenslauf Ch. Bangert
- 101 -
C. Bangert, S. Altrichter, G. Stary, G. Stingl, T. Kopp: Presence of lytic proteins in
myeloid and plasmacytoid dendritic cells (Poster, XXXIII. Jahrestagung der ADF, März
2006, Aachen, Deutschland)
C. Bangert , S. Schlehner, D. Mechtcheriakova, G. Stingl, T. Kopp. TLR-mediated
inflammatory responses in keratinocytes: expression of TSLP and IL-31 (Poster, 36th
Annual ESDR Meeting, September 2006, Paris, Frankreich)
C. Bangert , S. Kohlhofer, R. Kunstfeld, N. Selenko-Gebauer, F. Karlhofer, G. Stingl, T.
Kopp. Effective treatment of recalcitrant disseminated granuloma annulare with the
TNF-α-inhibitor infliximab (Vortrag, Jahrestagung 2006 der Österreichischen
Gesellschaft für Dermatologie und Venerologie, November 2006, Salzburg, Österreich)
C. Bangert , B. Strober, M. Cork, T. Luger, T. Bieber, S. Weise-Riccardi, A. Guettner,
A.G. Schmidt, T. Hultsch, G. Stingl: The effects of pimecrolimus 1% cream on the
recurrence of atopic dermatitis exacerbation following treatment with a mid-potent
topical corticosteroid (Poster, 16th Congress of EADV, June 2007, Wien, Österreich)
C. Bangert , S. Kohlhofer, R. Kunstfeld, N. Selenko-Gebauer, F. Karlhofer, G. Stingl, T.
Kopp: TNF-α-bearing macrophages: a target for TNF-α inhibitors in non-infectious
granulomatous diseases? (Poster, 37th Annual ESDR Meeting, September 2007,
Zürich, Schweiz)
Preise:
ESDR Travel Award (4. gemeinsamer Jahrestag der ESDR, JSID, SID, Miami Beach,
FL, USA, Mai 2003)
Theodor-Billroth Preis der Ärztekammer für Wien 2004
ESDR Poster Preis 2007 (37th Annual ESDR Meeting, September 2007, Zürich,
Schweiz)
Anhang Ch.Bangert
- 102 -
10. ANHANG
Abbildung I. Isotyp zu Ergebnisse S.68 Abbildung 4.3.5.3.: Aufnahme mit
konfokalem Lasermikroskop. Die dargestellte Isotyopkontrolle repräsentiert alle IgG1
Isotypkontrollen in TRITC/FITC Doppelfärbungen, die in der vorliegenden Arbeit
gezeigt wurden.
Ki-67 CD3 Überlappung
Überlappung CD3 Ki-67
Anhang Ch.Bangert
- 103 -
Abbildung II zu Ergebnisse Seite 42: 4.2.1. Typisierung der T-Zellen im dermalen
Infiltrat. Doppelfärbungen mit dem Proliferationsmarker Ki-67 (rot) und CD3 (grün)
zeigen keine doppelt gefärbten Zellen, d.h. keineÜberlappung (gelb).
Abbildung III A zu Ergebnisse S.45: 4.2.2. Charakterisierung der CD4+/CD25+ T-
Zellsubpopulation. Dreifachfärbungen mit CD25 (TRITC, rot), CD4 (FITC, grün) und
FOXP3 (Cy5, blau) im dermalen Infiltrat von ECT Läsionen nach 72 Std zeigen
dreifach positive reg T-Zellen in einer weißlichen Farbe (Pfeil).
Die einzelnen Isotypkontrollen für die angegebenen Antikörper werden in den
darauffolgenden Abbildungen dargestellt (III B-D). Es wurde immer ein Antikörper
durch den Isotyp ersetzt und die Einstellungen des konfokalen Lasermikroskops an die
Kontrolle angepaßt.
Alle in der vorliegenden Arbeit durchgeführten Dreifachfärbungen wurden mit dem
entsprechenden Kontrollantikörper durchgeführt. Die Abbildungen sind repräsentativ
für alle gefärbten und analysierten IgG1 Isotypkontrollen bei Dreifachfärbungen.
CD25 CD4
FOXP3 Überlappung

Anhang Ch.Bangert
- 104 -
Abbildung III B: mIgG1 Isotyp für CD4 FITC
Abbildung III C: mIgG1 Isotyp für FOXP3 Cy5
CD25
FOXP3
Überlappung
mIgG1
CD25
mIgG1 Überlappung
CD4

Anhang Ch.Bangert
- 105 -
Abbildung III D: mIgG1 Isotyp für CD25 TRITC
mIgG1 CD4
FOXP3 Überlappunggg
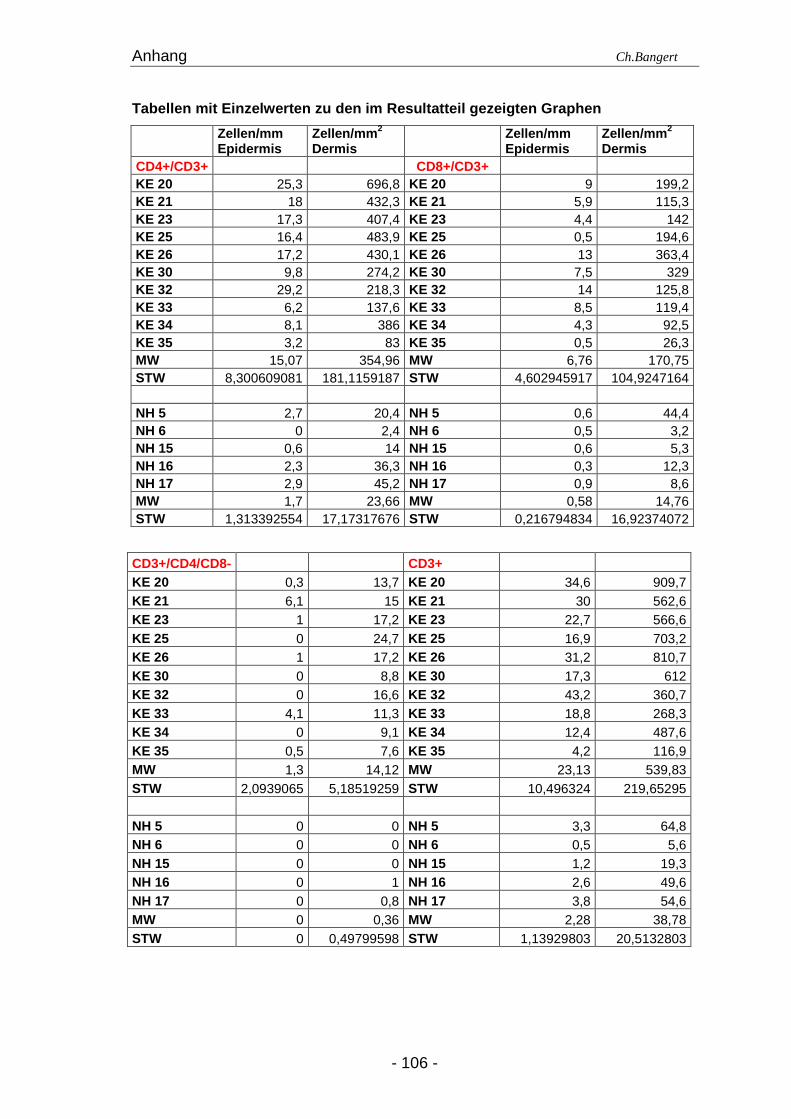
Anhang Ch.Bangert
- 106 -
Tabellen mit Einzelwerten zu den im Resultatteil ge zeigten Graphen
Zellen/mm Epidermis
Zellen/mm 2 Dermis
Zellen/mm Epidermis
Zellen/mm 2 Dermis
CD4+/CD3+ CD8+/CD3+ KE 20 25,3 696,8 KE 20 9 199,2 KE 21 18 432,3 KE 21 5,9 115,3 KE 23 17,3 407,4 KE 23 4,4 142 KE 25 16,4 483,9 KE 25 0,5 194,6 KE 26 17,2 430,1 KE 26 13 363,4 KE 30 9,8 274,2 KE 30 7,5 329 KE 32 29,2 218,3 KE 32 14 125,8 KE 33 6,2 137,6 KE 33 8,5 119,4 KE 34 8,1 386 KE 34 4,3 92,5 KE 35 3,2 83 KE 35 0,5 26,3 MW 15,07 354,96 MW 6,76 170,75 STW 8,300609081 181,1159187 STW 4,602945917 104,9247164 NH 5 2,7 20,4 NH 5 0,6 44,4 NH 6 0 2,4 NH 6 0,5 3,2 NH 15 0,6 14 NH 15 0,6 5,3 NH 16 2,3 36,3 NH 16 0,3 12,3 NH 17 2,9 45,2 NH 17 0,9 8,6 MW 1,7 23,66 MW 0,58 14,76 STW 1,313392554 17,17317676 STW 0,216794834 16,92374072
CD3+/CD4/CD8- CD3+ KE 20 0,3 13,7 KE 20 34,6 909,7 KE 21 6,1 15 KE 21 30 562,6 KE 23 1 17,2 KE 23 22,7 566,6 KE 25 0 24,7 KE 25 16,9 703,2 KE 26 1 17,2 KE 26 31,2 810,7 KE 30 0 8,8 KE 30 17,3 612 KE 32 0 16,6 KE 32 43,2 360,7 KE 33 4,1 11,3 KE 33 18,8 268,3 KE 34 0 9,1 KE 34 12,4 487,6 KE 35 0,5 7,6 KE 35 4,2 116,9 MW 1,3 14,12 MW 23,13 539,83 STW 2,0939065 5,18519259 STW 10,496324 219,65295 NH 5 0 0 NH 5 3,3 64,8 NH 6 0 0 NH 6 0,5 5,6 NH 15 0 0 NH 15 1,2 19,3 NH 16 0 1 NH 16 2,6 49,6 NH 17 0 0,8 NH 17 3,8 54,6 MW 0 0,36 MW 2,28 38,78 STW 0 0,49799598 STW 1,13929803 20,5132803
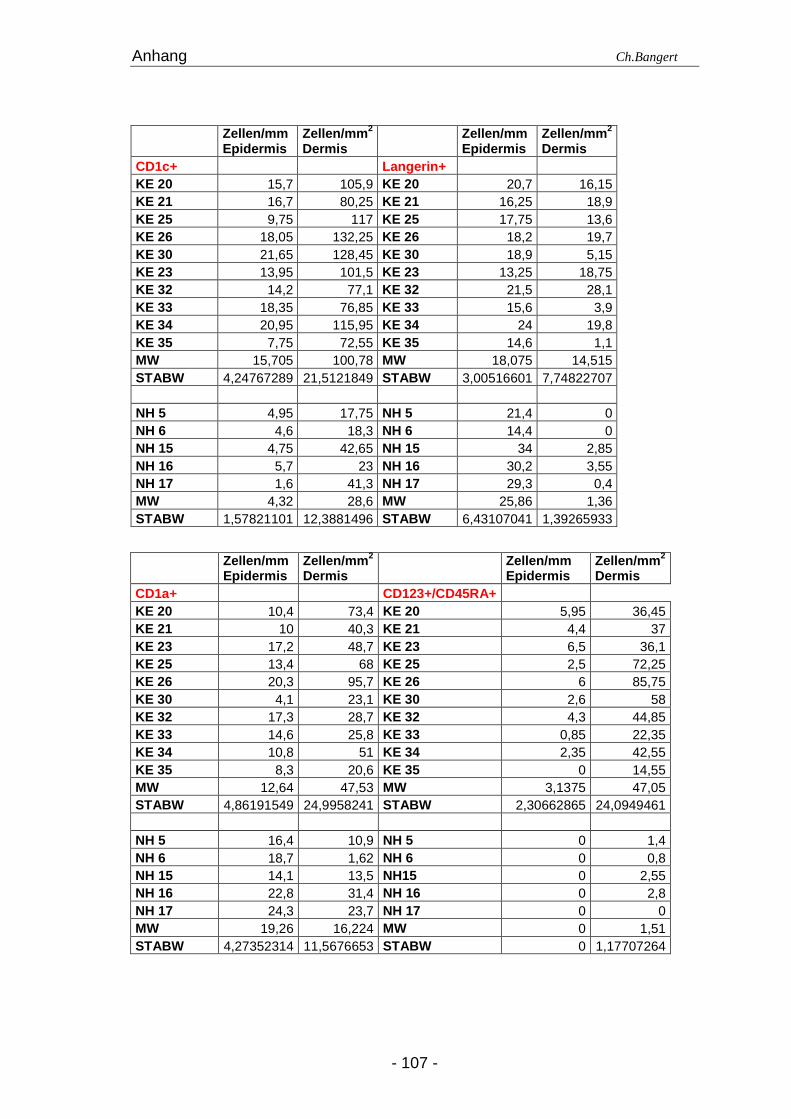
Anhang Ch.Bangert
- 107 -
Zellen/mm Epidermis
Zellen/mm 2 Dermis
Zellen/mm Epidermis
Zellen/mm 2 Dermis
CD1c+ Langerin+ KE 20 15,7 105,9 KE 20 20,7 16,15 KE 21 16,7 80,25 KE 21 16,25 18,9 KE 25 9,75 117 KE 25 17,75 13,6 KE 26 18,05 132,25 KE 26 18,2 19,7 KE 30 21,65 128,45 KE 30 18,9 5,15 KE 23 13,95 101,5 KE 23 13,25 18,75 KE 32 14,2 77,1 KE 32 21,5 28,1 KE 33 18,35 76,85 KE 33 15,6 3,9 KE 34 20,95 115,95 KE 34 24 19,8 KE 35 7,75 72,55 KE 35 14,6 1,1 MW 15,705 100,78 MW 18,075 14,515 STABW 4,24767289 21,5121849 STABW 3,00516601 7,74822707 NH 5 4,95 17,75 NH 5 21,4 0 NH 6 4,6 18,3 NH 6 14,4 0 NH 15 4,75 42,65 NH 15 34 2,85 NH 16 5,7 23 NH 16 30,2 3,55 NH 17 1,6 41,3 NH 17 29,3 0,4 MW 4,32 28,6 MW 25,86 1,36 STABW 1,57821101 12,3881496 STABW 6,43107041 1,39265933
Zellen/mm Epidermis
Zellen/mm 2 Dermis
Zellen/mm Epidermis
Zellen/mm 2 Dermis
CD1a+ CD123+/CD45RA+ KE 20 10,4 73,4 KE 20 5,95 36,45 KE 21 10 40,3 KE 21 4,4 37 KE 23 17,2 48,7 KE 23 6,5 36,1 KE 25 13,4 68 KE 25 2,5 72,25 KE 26 20,3 95,7 KE 26 6 85,75 KE 30 4,1 23,1 KE 30 2,6 58 KE 32 17,3 28,7 KE 32 4,3 44,85 KE 33 14,6 25,8 KE 33 0,85 22,35 KE 34 10,8 51 KE 34 2,35 42,55 KE 35 8,3 20,6 KE 35 0 14,55 MW 12,64 47,53 MW 3,1375 47,05 STABW 4,86191549 24,9958241 STABW 2,30662865 24,0949461 NH 5 16,4 10,9 NH 5 0 1,4 NH 6 18,7 1,62 NH 6 0 0,8 NH 15 14,1 13,5 NH15 0 2,55 NH 16 22,8 31,4 NH 16 0 2,8 NH 17 24,3 23,7 NH 17 0 0 MW 19,26 16,224 MW 0 1,51 STABW 4,27352314 11,5676653 STABW 0 1,17707264
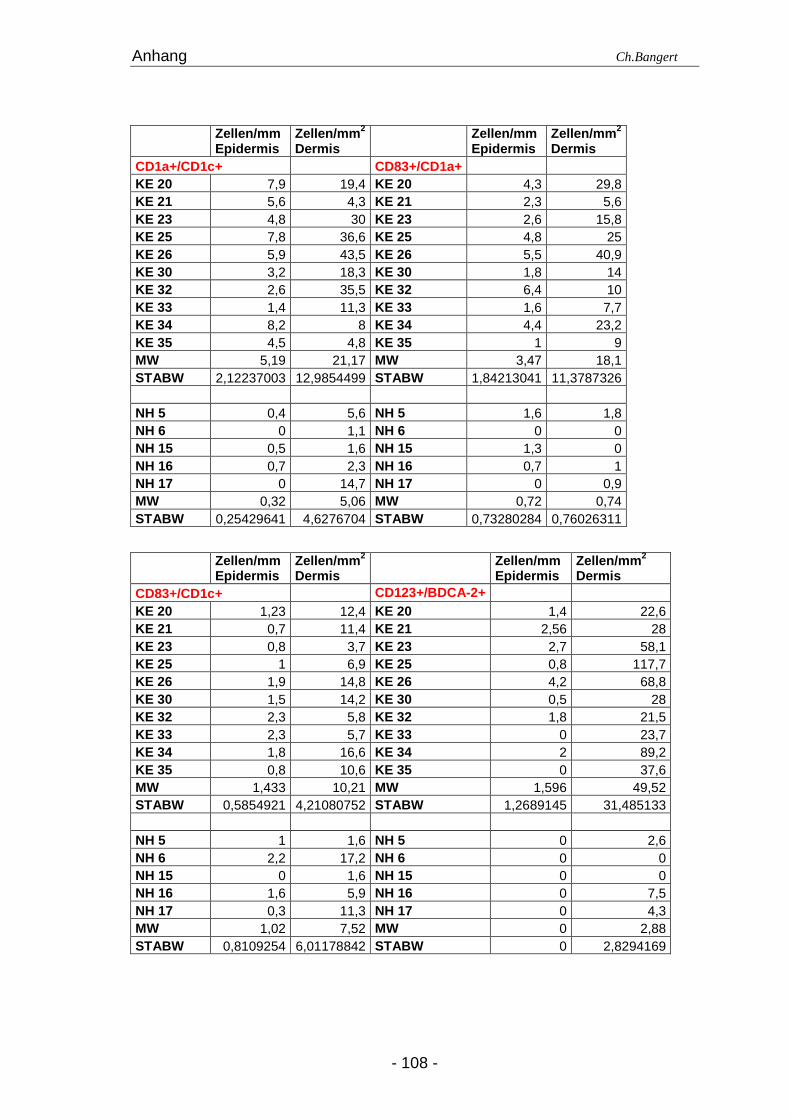
Anhang Ch.Bangert
- 108 -
Zellen/mm Epidermis
Zellen/mm 2 Dermis
Zellen/mm Epidermis
Zellen/mm 2 Dermis
CD1a+/CD1c+ CD83+/CD1a+ KE 20 7,9 19,4 KE 20 4,3 29,8 KE 21 5,6 4,3 KE 21 2,3 5,6 KE 23 4,8 30 KE 23 2,6 15,8 KE 25 7,8 36,6 KE 25 4,8 25 KE 26 5,9 43,5 KE 26 5,5 40,9 KE 30 3,2 18,3 KE 30 1,8 14 KE 32 2,6 35,5 KE 32 6,4 10 KE 33 1,4 11,3 KE 33 1,6 7,7 KE 34 8,2 8 KE 34 4,4 23,2 KE 35 4,5 4,8 KE 35 1 9 MW 5,19 21,17 MW 3,47 18,1 STABW 2,12237003 12,9854499 STABW 1,84213041 11,3787326 NH 5 0,4 5,6 NH 5 1,6 1,8 NH 6 0 1,1 NH 6 0 0 NH 15 0,5 1,6 NH 15 1,3 0 NH 16 0,7 2,3 NH 16 0,7 1 NH 17 0 14,7 NH 17 0 0,9 MW 0,32 5,06 MW 0,72 0,74 STABW 0,25429641 4,6276704 STABW 0,73280284 0,76026311
Zellen/mm Epidermis
Zellen/mm 2 Dermis
Zellen/mm Epidermis
Zellen/mm 2 Dermis
CD83+/CD1c+ CD123+/BDCA-2+ KE 20 1,23 12,4 KE 20 1,4 22,6 KE 21 0,7 11,4 KE 21 2,56 28 KE 23 0,8 3,7 KE 23 2,7 58,1 KE 25 1 6,9 KE 25 0,8 117,7 KE 26 1,9 14,8 KE 26 4,2 68,8 KE 30 1,5 14,2 KE 30 0,5 28 KE 32 2,3 5,8 KE 32 1,8 21,5 KE 33 2,3 5,7 KE 33 0 23,7 KE 34 1,8 16,6 KE 34 2 89,2 KE 35 0,8 10,6 KE 35 0 37,6 MW 1,433 10,21 MW 1,596 49,52 STABW 0,5854921 4,21080752 STABW 1,2689145 31,485133 NH 5 1 1,6 NH 5 0 2,6 NH 6 2,2 17,2 NH 6 0 0 NH 15 0 1,6 NH 15 0 0 NH 16 1,6 5,9 NH 16 0 7,5 NH 17 0,3 11,3 NH 17 0 4,3 MW 1,02 7,52 MW 0 2,88 STABW 0,8109254 6,01178842 STABW 0 2,8294169
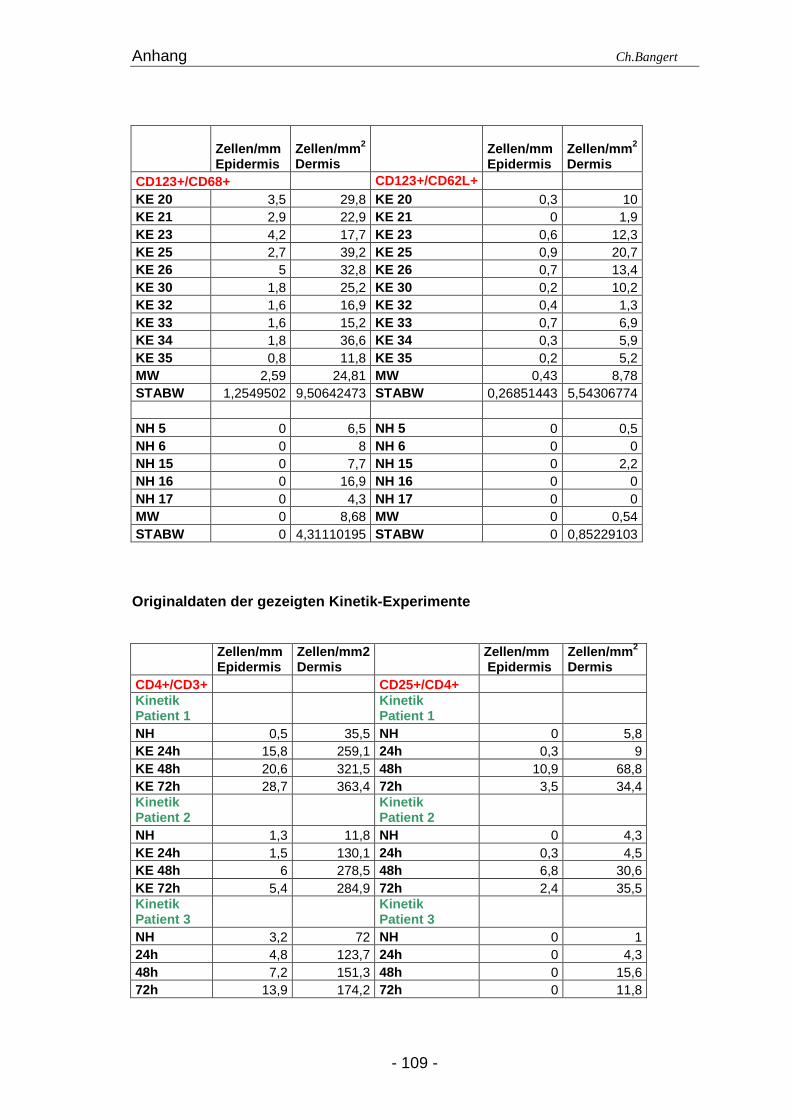
Anhang Ch.Bangert
- 109 -
Zellen/mm Epidermis
Zellen/mm 2 Dermis
Zellen/mm Epidermis
Zellen/mm 2 Dermis
CD123+/CD68+ CD123+/CD62L+ KE 20 3,5 29,8 KE 20 0,3 10 KE 21 2,9 22,9 KE 21 0 1,9 KE 23 4,2 17,7 KE 23 0,6 12,3 KE 25 2,7 39,2 KE 25 0,9 20,7 KE 26 5 32,8 KE 26 0,7 13,4 KE 30 1,8 25,2 KE 30 0,2 10,2 KE 32 1,6 16,9 KE 32 0,4 1,3 KE 33 1,6 15,2 KE 33 0,7 6,9 KE 34 1,8 36,6 KE 34 0,3 5,9 KE 35 0,8 11,8 KE 35 0,2 5,2 MW 2,59 24,81 MW 0,43 8,78 STABW 1,2549502 9,50642473 STABW 0,26851443 5,54306774 NH 5 0 6,5 NH 5 0 0,5 NH 6 0 8 NH 6 0 0 NH 15 0 7,7 NH 15 0 2,2 NH 16 0 16,9 NH 16 0 0 NH 17 0 4,3 NH 17 0 0 MW 0 8,68 MW 0 0,54 STABW 0 4,31110195 STABW 0 0,85229103
Originaldaten der gezeigten Kinetik-Experimente
Zellen/mm Epidermis
Zellen/mm2 Dermis
Zellen/mm Epidermis
Zellen/mm 2 Dermis
CD4+/CD3+ CD25+/CD4+ Kinetik Patient 1
Kinetik Patient 1
NH 0,5 35,5 NH 0 5,8 KE 24h 15,8 259,1 24h 0,3 9 KE 48h 20,6 321,5 48h 10,9 68,8 KE 72h 28,7 363,4 72h 3,5 34,4 Kinetik Patient 2
Kinetik Patient 2
NH 1,3 11,8 NH 0 4,3 KE 24h 1,5 130,1 24h 0,3 4,5 KE 48h 6 278,5 48h 6,8 30,6 KE 72h 5,4 284,9 72h 2,4 35,5 Kinetik Patient 3
Kinetik Patient 3
NH 3,2 72 NH 0 1 24h 4,8 123,7 24h 0 4,3 48h 7,2 151,3 48h 0 15,6 72h 13,9 174,2 72h 0 11,8

Anhang Ch.Bangert
- 110 -
Zellen/mm Epidermis
Zellen/mm 2 Dermis
Zellen/mm Epidermis
Zellen/mm 2 Dermis
CD56+ CD45RA+/CD123+ Kinetik Patient 1
Kinetik Patient 1
NH 0 7,9 NH 0 4 KE 24h 1,3 21,2 KE 24h 1,6 19,4 KE 48h 2,5 36,6 KE 48h 4,3 28,4 KE 72h 0,8 51,6 KE 72h 4,4 46,3 Kinetik Patient 2
Kinetik Patient 2
NH 0 3,9 NH 0 0 KE 24h 0 13,7 KE 24h 0,7 17,4 KE 48h 0 21,8 KE 48h 1,2 28,2 KE 72h 1,2 20,4 KE 72h 1,6 23,3 Kinetik Patient 3
Kinetik Patient 3
NH 0 9,2 NH 0 1,1 24h 0 20,4 24h 0 22,6 48h 0 24,4 48h 0 26,7 72h 0,5 30,1 72h 0 20,4