Embed Size (px)
Citation preview

Abteilung für Infektionsgenetik
Helmholtz-Zentrum für Infektionsforschung, Braunschweig
und
Tierärztliche Hochschule Hannover
In vivo Analyse der oralen Infektion mit biolumineszenten
Listeria monocytogenes im Mausmodell
INAUGURAL – DISSERTATION
Zur Erlangung des Grades einer
Doktorin der Naturwissenschaften
-Doctor rerum naturalium-
(Dr. rer. nat.)
durch die Tierärztliche Hochschule Hannover
von
Dipl. Biol. Silke Bergmann
aus Hohenmölsen
Hannover 2012

Wissenschaftliche Betreuung:
Prof. Dr. Klaus Schughart Abteilung für Infektionsgenetik Helmholtz-Zentrum für Infektionsforschung, Braunschweig und Tierärztliche Hochschule Hannover Prof. Dr. Peter Valentin-Weigand Institut für Mikrobiologie Tierärztliche Hochschule Hannover PD Dr. Eva Medina Abteilung für Infektionsimmunologie Helmholtz-Zentrum für Infektionsforschung, Braunschweig PD Dr. Andreas Lengeling The Roslin Institute and Royal (Dick) School of Veterinary Studies, Universität Edinburgh, Großbritannien
1. Gutachter: Prof. Dr. Klaus Schughart
2. Gutachter: PD Dr. Andreas Lengeling
mündliche Prüfung (Disputation) am: 05.11.2012

Inhaltsverzeichnis
i
Inhaltsverzeichnis
Inhaltsverzeichnis .............................................................................................................. i
Abkürzungsverzeichnis ...................................................................................................... vi
Tabellenverzeichnis .......................................................................................................... xi
Abbildungsverzeichnis ..................................................................................................... xii
Zusammenfassung ........................................................................................................... xv
Summary ....................................................................................................................... xvii
1 Einleitung .................................................................................................................. 1
1.1 Morphologie und Physiologie der Gattung Listeria ................................................ 1
1.2 Epidemiologie von Listeria monocytogenes ........................................................... 2
1.3 Humane Listeriose ................................................................................................ 4
1.4 Listeriose in domestizierten Tieren ....................................................................... 7
1.5 Wirtsspezifität und Wirtsinvasion ......................................................................... 9
1.5.1 Invasionsproteine und Speziesspezifität ............................................................................................. 9
1.5.2 Wirtsinvasion und intrazelluläre Lebensweise .................................................................................. 12
1.5.3 Überquerung epithelialer Barrieren im Wirt .................................................................................... 15
1.5.4 Grundlegende Mechanismen der Immunabwehr von Listeria monocytogenes ............................... 18
1.6 Studien der in vivo Infektion ................................................................................21
1.6.1 Tiermodelle zur Analyse listerialer Infektionsmechanismen ............................................................ 21
1.6.2 Mausmodelle zur Analyse listerialer Infektionsmechanismen ......................................................... 22
1.7 In vivo Imaging ....................................................................................................24
1.7.1 Das Prinzip der Biolumineszenz ........................................................................................................ 24
1.7.2 Die Technologie des in vivo Imaging ................................................................................................ 25
1.7.3 In vivo Imaging von Listeria monocytogenes Infektionen in der Maus ............................................. 26
1.8 Konzept und Fragestellung der vorliegenden Dissertation ....................................28

Inhaltsverzeichnis
ii
2 Ergebnisse ................................................................................................................31
2.1 Murinisierung der Listerienstämme Listeria monocytogenes 10403S Xen32
und LO28 .............................................................................................................31
2.1.1 Ergebnisse der Transformation ......................................................................................................... 32
2.1.2 Validierung der homologen Rekombination des murinisierten Internalin A .................................... 33
2.1.3 Analysen der Internalin A und Internalin B Expression in genetisch modifizierten
Listerienstämmen ............................................................................................................................. 35
2.1.4 Analyse der murinisierten Listerienstämme im Invasions-Assay ...................................................... 38
2.2 Vergleich der Wirtsspezifität und Virulenz des genetisch modifizierten
LmoXen32-mur mit dessen Wildtyp .....................................................................41
2.2.1 Überlebenszeiten nach der Infektion mit LmoXen32 und LmoXen32-mur in ausgewählten
Mausinzuchtstämmen....................................................................................................................... 41
2.2.2 Kinetik der in vivo Replikation von LmoXen32 und LmoXen32-mur in BALB/cJ Mäusen .................. 44
2.2.2.1 In vivo Imaging von LmoXen32 und LmoXen32-mur infizierten BALB/cJ Mäusen ..................... 45
2.2.2.2 Kinetik der bakteriellen Keimlast in den inneren Organen von LmoXen32 und
LmoXen32-mur infizierten BALB/cJ Mäusen .............................................................................. 48
2.2.2.3 Histopathologische Analyse von LmoXen32 und LmoXen32-mur infizierten BALB/cJ
Mäusen ....................................................................................................................................... 50
2.3 Vergleich der Wirtsspezifität und Virulenz des genetisch modifizierten
LmoEGDe-lux-mur mit dessen Wildtyp .................................................................52
2.3.1 Analyse der Stämme LmoEGDe-lux und LmoEGDe-lux-mur auf die Proteinexpression von
Internalin A und Internalin B sowie die Untersuchung der Invasionsrate in humanen und
murinen Colonkarzinom-Zellen. ........................................................................................................ 53
2.3.2 Charakterisierung und Vergleich der Virulenz verschiedener LmoEGDe Stämme in BALB/cJ
Mäusen ............................................................................................................................................. 55
2.3.3 Vergleich der Wirtsresistenz in ausgewählten Mausinzuchtstämmen nach der Infektion mit
LmoEGDe-lux und LmoEGDe-mur-lux ............................................................................................... 61
2.3.4 Kinetik der in vivo Replikation von LmoEGDe-lux und LmoEGDe-lux-mur in ausgewählten
Mausinzuchtstämmen....................................................................................................................... 69
2.3.4.1 In vivo Imaging von LmoEGDe-lux und LmoEGDe-mur-lux infizierten
Mausinzuchtstämmen ................................................................................................................ 69
2.3.4.2 Kinetik der bakteriellen Keimlast in den inneren Organen von LmoEGDe-lux und
LmoEGDe-lux-mur infizierten Mausinzuchtstämmen ................................................................ 73

Inhaltsverzeichnis
iii
2.3.4.3 Histopathologische Analyse von LmoEGDe-lux und LmoEGDe-mur-lux infizierten
Mausinzuchtstämmen ................................................................................................................ 81
2.3.4.4 Analyse der Chemokin-Zytokin-Produktion von LmoEGDe-lux und LmoEGDe-mur-lux
infizierten Mausinzuchtstämmen ............................................................................................... 82
2.4 Vergleich der Wirtsspezifität und Wirtsresistenz der Listerienstämme
LmoXen32-mur und LmoEGDe-mur-lux ................................................................85
2.5 Korrelation der bakteriellen Keimlast mit Biolumineszenzsignalen in den
inneren Organen von LmoXen32 und LmoXen32-mur infizierten
Mausinzuchtstämmen im Vergleich zu LmoEGDe-lux und LmoEGDe-lux-mur
infizierten Mausinzuchtstämmen .........................................................................86
2.6 Infektion des zentralen Nervensystems nach der Infektion mit LmoXen32,
LmoXen32-mur, LmoEGDe, LmoEGDe-lux und LmoEGDe-mur-lux .........................89
2.7 Interferonproduktion und Listerieninfektion ........................................................91
2.7.1 Charakterisierung des Einflusses der Interferoninduktion auf die Listerieninfektion ...................... 91
2.7.2 Charakterisierung der Listerieninfektion in Bezug auf die Interferon-β Produktion ........................ 94
3 Diskussion.................................................................................................................97
3.1 Der Infektionsverlauf nach einer Listerieninfektion ist von der Virulenz des
Listeriensubtypes abhängig ..................................................................................98
3.2 Die Murinisierung des Internalin A resultiert in einem invasiveren
Krankheitsverlauf .............................................................................................. 102
3.3 Der Infektionsverlauf nach einer Listerieninfektion ist abhängig vom
genetischen Hintergrund des Wirtes .................................................................. 108
3.4 Infektion des zentralen Nervensystems zeigt keine Reproduzierbarkeit und
ist nicht von der Murinisierung des Internalin A abhängig .................................. 117
3.5 Die Detektion des Biolumineszenzsignals in Abhängigkeit verschiedener
Faktoren ............................................................................................................ 121

Inhaltsverzeichnis
iv
3.6 Reaktionen des angeborenen Immunsystems auf die Infektion mit Listeria
monocytogenes ................................................................................................. 124
4 Ausblick .................................................................................................................. 128
5 Material und Methoden .......................................................................................... 130
5.1 Generierung genetisch veränderter Listeria monocytogenes Stämme ................. 130
5.1.1 Generierung elektrokompetenter Listerien .................................................................................... 130
5.1.2 Transformation und Selektion rekombinanter Listeria monocytogenes Stämme .......................... 131
5.2 Molekularbiologische Methoden ....................................................................... 134
5.2.1 Präparation von Plasmid-DNA ........................................................................................................ 134
5.2.2 Präparation genomischer bakterieller DNA .................................................................................... 134
5.2.3 PCR .................................................................................................................................................. 134
5.2.4 Gelelektrische Auftrennung von DNA-Fragmenten ........................................................................ 136
5.2.5 Sequenzierung ................................................................................................................................ 136
5.2.6 Proteinpräparation und Western-Blot-Analyse .............................................................................. 137
5.2.6.1 Proteinextraktion...................................................................................................................... 137
5.2.6.2 Western-Blot-Analyse ............................................................................................................... 138
5.3 Invasions-Assay ................................................................................................. 141
5.4 In vivo Mausinfektionsexperimente ................................................................... 142
5.4.1 Kultivierung und Präparation von Listeria monocytogenes ............................................................ 143
5.4.2 Orale Infektion ................................................................................................................................ 144
5.4.3 Intravenöse Infektion ...................................................................................................................... 145
5.4.4 Biolumineszenz-Imaging ................................................................................................................. 145
5.4.4.1 In vivo Detektion der bakteriellen Luziferase ........................................................................... 146
5.4.4.2 Ex vivo Detektion der bakteriellen Luziferase .......................................................................... 147
5.4.4.3 Detektion der IFN-β-Produktion ............................................................................................... 147
5.4.5 Bestimmung der bakteriellen Keimlast ........................................................................................... 148
5.5 Luminex-Assay................................................................................................... 148
5.6 Histologische und immunhistologische Färbung ................................................. 149
5.7 Statistische Auswertung .................................................................................... 150

Inhaltsverzeichnis
v
6 Literaturverzeichnis ................................................................................................ 151
7 Eidesstattliche Erklärung ......................................................................................... 174
8 Danksagung ............................................................................................................ 175

Abkürzungsverzeichnis
vi
Abkürzungsverzeichnis
A Adenin
ActA Aktin-bindendes Protein A
ADI Arginindeiminase
AS Aminosäure
A Ala Alanin C Cys Cystein D Asp Aspartat E Glu Glutamat F Phe Phenylalanin G Gly Glycin H His Histidin I Ile Isoleucin K Lys Lysin L Leu Leucin M Met Methionin N Asn Asparagin P Pro Prolin Q Gln Glutamin R Arg Arginin S Ser Serin T Thr Threonin V Val Valin W Trp Tryptophan X beliebige Aminosäure Y Tyr Tyrosin
APS Ammoniumperoxodisulfat
ATP Adenosintriphosphat
betl Glycin - Betaine Aufnahmesystem I
bp Basenpaar
BHI Brain-Heart-Infusion
BLI Biolumineszenz-Imaging
BiLe Gallensaft Exklusionsprotein (bile exclusion protein)
BSA Rinderserumalbumin (bovine serum albumin)
BSH Gallensaft Hydrolase (bile salt hydrolase)
C Cytosin
°C Grad Celsius

Abkürzungsverzeichnis
vii
CaCO3 Calciumcarbonat
CCD coupled cooled device
CCL2 (MCP-1 alt) monocyte chemoattractant protein
CDC Center for Disease Control and Prevention
CSF-1 Monozyten koloniestimulierender Faktor (colony stimulating
factor 1)
CFU koloniebildende Einheiten (colony forming unit)
cm Zentimeter
c-Met mesenchymal epithelial transition factor / hepatocyte growth
factor receptor (HGFR)
DMEM Dulbecco's Modified Eagle Medium
DNA Desoxyribonukleinsäure
dNTP Desoxynukleotidtriphosphatgemisch
EDTA Ethylendiamintetraazetat
EXIM Abteilung für Experimentelle Immunologie
FAE Follikel-assoziiertes Epithel
Fd Kapazität (µFD)
FKS Fötales Kälberserum (fetal calf serum)
FbpA Eisenbindendes Vorläuferprotein
flaA Flagellin A
x g Erdbeschleunigung; g = 9,81 m/s
g Gramm
G Guanin
GAD Glutamatedecarboxylase
gbu Glycin - Betaine Aufnahmesystem II
GM-CSF Granulozyten Monozyten koloniestimulierender Faktor
(granulocyte-macrophage colony-stimulating factor)
h Stunde
H2O2 Wasserstoffperoxid
HCI Salzsäure
hly Listeriolysin (Gen)

Abkürzungsverzeichnis
viii
i.p. intraperitoneal
i.v. intravenös
IFN-γ Interferon-gamma
IL Interleukin
Inl Internalin
Ipr1 Intracellular pathogen resistence 1
IR invertierte Sequenzwiederholung (inverted repeats)
K Kilo- (103)
kb Kilobasen
kDa kilo Dalton
KO knock-out
l Liter
LD50 mittlere lethale Dosis
LIPI-1 Listeria Pathogentätsinsel
LLO Listeriolysin (Protein)
Lmo Listeria monocytogenes
LRR leucinreiche Wiederholungen (Leucin-rich-repeats)
µ mikro- (10–6)
m milli- (10–3)
m Meter
M Molar (mol/l)
M-CSF Monozyten koloniestimulierender factor (macrophage colony
stimulating factor)
MALT Schleimhaut-assoziiertes lymphatisches Gewebe (mucosa
associated lymphoid tissue)
MAPK Mitogen-aktivierte Protein Kinase
MEM Minimal Eagle's Medium
MHC Haupthistokompatibilitätskomplex (major histocompability
complex)
mg Milligramm
MgCl2 Magnesiumchlorid

Abkürzungsverzeichnis
ix
min Minute
MIP Makrophagen inflammatorisches Protein (macrophage inflamma-
tory protein)
mLK mesenterische Lymphknoten
MLST Multi-Locus-Sequenz-Typisierung
MOI Multiplizität der Infektion (multiplicity of infection)
MOLI Abteilung für Molekulare Immunologie
Mol Mol (6,023 x 1023 Teilchen)
mpl/Mpl Metalloprotease
MyD88 myeloid differentiation primary response gene (88)
n nano- (10–9)
nM nano-molar (nmol/L)
NaCl Natriumchlorid
NaHCO3 natriumhydrogencarbonat
NaOH Natriumhydroxid
NALP3 NACHT, LRR and PYD domains-containing protein 3
NFκB nuclear factor “kappa-light-chain-enhancer” of activated B-cells
NLR NOD-like-Rezeptor
NOD Nukleotidbindungs- und Oligomerisierungsdomäne
OD Optische Dichte
opuC Carnitin Aufnahmesystem
p pico ( 10–12 )
PBS phospatgepufferte SalzLösung (Phosphate Buffered Saline)
PCR Polymerasekettenreaktion (polymerase chain reaction)
PrfA Pleiotropisches Aktivatorprotein
PLC-A Phospholipase A
PLC-B Phospholipase B
pL-PLc Phosphatidylinositolspezifische Phospholipase C
PSA Prophage LmoScott A
PVDF Polyvinylidenfluorid
QTL quantitative trait loci

Abkürzungsverzeichnis
x
rpm Umdrehungen je Minute
RT Raumtemperatur
SB Abteilung für Strukturbiologie
SDS Natriumdodecylsulfat
sec (s) Sekunde
SPF spezifisch Pathogen frei (specific pathogen free)
sr Steridian
T Thymin
TBE Tris-Borat-EDTA
TH-1, TH-2 T-Helfer-1, T-Helfer-2
TLR Toll-like Receptor
TGF transformierender wachstumsfaktor (transforming growth factor)
TNF Tumornekrosefaktor
Taq Thermus aquaticus
TE Tris-EDTA
Tris Tris (hydroxymethyl) aminoethan
U Einheit der Aktivität (Unit)
UV Ultraviolett
V Volt
Vip Lmo320, Virulenzprotein in Listeriavirulenz
Vol. Volumen
v/v Volumen pro Volumen
WT Wildtyp
w/v Gewicht pro Volumen
zLK zervikale Lymphknoten

Tabellenverzeichnis
xi
Tabellenverzeichnis
Tabelle 1-1: Die Arten der Gattung Listeria ............................................................................... 2
Tabelle 2-1: Zeitpunkt des Auftretens der Anzeichen einer Invasion des ZNS in
ausgewählten Mausinzuchtstämmen nach oraler Infektion mit Listeria monocytogenes ...... 90

Abbildungsverzeichnis
xii
Abbildungsverzeichnis
Abbildung 1-1: Infektionsweg von Listeria monocytogenes .................................................... 7
Abbildung 1-2: Speziesspezifität von Listeria monocytogenes ..............................................11
Abbildung 1-3: Infektionszyklus von Listeria monocytogenes ...............................................13
Abbildung 2-1: Darstellung der Sequenzen des DNA Abschnittes von inlA und der
entsprechenden codierenden Proteinsequenzen in den genetisch modifizierten
Listerienstämmen. ................................................................................................................34
Abbildung 2-2: Ergebnis der Analyse der Proteinexpression von Internalin A und
Internalin B in genetisch veränderten Listerienstämmen. ......................................................36
Abbildung 2-3: Aufnahme genetisch veränderter Listerienstämme in Caco-2 Zellen. ...........39
Abbildung 2-4: Aufnahme genetisch veränderter Listerienstämme in CT-26 Zellen. .............40
Abbildung 2-5: Überlebensraten und Verlauf der Körpergewichte ausgewählter
Mausinzuchtstämme nach oraler Infektion mit LmoXen32-mur. ............................................42
Abbildung 2-6: In vivo Imaging von oral infizierten Mausinzuchstämmen mit LmoXen32-
mur. ......................................................................................................................................43
Abbildung 2-7: Biolumineszenz-Imaging oral infizierter BALB/cJ Mäuse mit LmoXen32
bzw. LmoXen32-mur. ...........................................................................................................46
Abbildung 2-8: Verlauf des Körpergewichtes von BALB/cJ Mäusen nach oraler
Infektion mit 5x109 CFU LmoXen32 und LmoXen32-mur......................................................47
Abbildung 2-9: Bakterielle Keimlasten in den inneren Organe von BALB/cJ Mäusen
nach der oralen Infektion mit LmoXen32 oder LmoXen32-mur. ............................................49
Abbildung 2-10: Histopathologische und immunhistologische Analyse der Leber von
oral infizierten BALB/cJ Mäusen mit LmoXen32 und LmoXen32-mur. ..................................51
Abbildung 2-11: Ergebnis der Analyse der Proteinexpression von Internalin A und
Internalin B in genetisch veränderten Listerienstämmen. ......................................................54
Abbildung 2-12: Aufnahme genetisch veränderter Listerienstämme in Caco-2 und CT-
26 Zellen. .............................................................................................................................55
Abbildung 2-13: Überlebensraten und Verlauf des Körpergewichtes von BALB/cJ
Mäusen nach oraler Infektion mit LmoEGDe, LmoEGDe-lux und LmoEGDe-mur-lux. ..........56

Abbildungsverzeichnis
xiii
Abbildung 2-14: Biolumineszenz-Imaging oral infizierter BALB/cJ Mäuse mit
LmoEGDe-lux bzw. LmoEGDe-mur-lux. ...............................................................................58
Abbildung 2-15: Ex vivo Biolumineszenz-Imaging der inneren Organen oral infizierter
BALB/cJ mit LmoEGDe-lux und LmoEGDe-mur-lux an Tag vier nach der Infektion. .............59
Abbildung 2-16: Histopathologische und immunhistologische Analyse von Leber von
oral infizierten BALB/cJ mit 5x109 CFU LmoEGDe-lux bzw. LmoEGDe-mur-lux. ..................60
Abbildung 2-17: Überlebensraten von ausgewählten Mausinzuchtstämmen nach oraler
Infektion von 5x109 CFU LmoEGDe-lux und LmoEGDe-mur-lux. .........................................62
Abbildung 2-18: Verlauf des Körpergewichtes ausgewählter Mausinzuchtstämme nach
oraler Infektion mit 5x109 CFU LmoEGDe-lux oder LmoEGDe-mur-lux. ...............................64
Abbildung 2-19: Biolumineszenz-Imaging von mit 5x109 CFU LmoEGDe-lux oral
infizierten Mausinzuchstämmen. ..........................................................................................67
Abbildung 2-20: Biolumineszenz-Imaging von mit 5x109 CFU LmoEGDe-mur-lux oral
infizierten Mausinzuchstämmen. ..........................................................................................68
Abbildung 2-21: Biolumineszenz-Imaging von oral infizierten Mausinzuchstämmen mit
5x109 CFU LmoEGDe-lux bzw. LmoEGDe-mur-lux. .............................................................70
Abbildung 2-22: Verlauf des Körpergewichtes ausgewählter Mausinzuchtstämme nach
oraler Infektion mit 5x109 CFU LmoEGDe-lux bzw. LmoEGDe-mur-lux. ...............................72
Abbildung 2-23: Ex vivo BLI der inneren Organe ausgewählter Mausinzuchtstämme
nach der oralen Infektion mit 5x109 CFU LmoEGDe-lux bzw. LmoEGDe-mur-lux. ...............75
Abbildung 2-24: Bakterielle Keimlasten in den inneren Organe ausgewählter Maus-
inzuchtstämme nach der oralen Infektion mit 5x109 CFU LmoEGDe-lux bzw.
LmoEGDe-mur-lux. ..............................................................................................................77
Abbildung 2-25: Bakterielle Keimlast in den inneren Organen ausgewählter Maus-
inzuchtstämme am Tag fünf nach der oralen Infektion mit 5x109 CFU LmoEGDe-mur-
lux. .......................................................................................................................................80
Abbildung 2-26: Histopathologische und immunhistologische Analyse der Leber von
oral infizierten C57BL/6J und C3HeB/FeJ Mäusen mit 5x109 CFU LmoEGDe-mur-lux. ........82
Abbildung 2-27: Chemokin- und Zytokinproduktion ausgewählter Mausinzuchtstämme
nach der oraler Infektion mit 5x109 CFU LmoEGDe-lux bzw. LmoEGDe-mur-lux. ................84
Abbildung 2-28: Korrelation der bakteriellen Keimlast in den inneren Organe mit den
nachgewiesenen Biolumineszenzsignalen. ...........................................................................87
Abbildung 2-29: Korrelation der bakteriellen Keimlast in den inneren Organen mit den
nachgewiesenen Biolumineszenzsignalen. ...........................................................................88

Abbildungsverzeichnis
xiv
Abbildung 2-30: Histopathologie und Immunhistologie des Gehirns einer „kreiselnden“
129P2/Ola Maus nach der oralen Infektion mit 5x109 CFU LmoXen32-mur. .........................91
Abbildung 2-31: Überlebensrate und der Verlauf des Körpergewichtes von Ifi27l2a-KO
und C57BL/6N Mäusen nach intravenöser Infektion mit 2x104 CFU LmoEGDe-mur-lux. ......92
Abbildung 2-32: Bakterielle Keimlast in Leber und Milz von Ifi27l2a-KO und C57BL/6N
Mäusen nach intravenöser Infektion mit 2x104 CFU LmoEGDe-mur-lux. ..............................93
Abbildung 2-33: IFN-β Induktion nach oraler Infektion mit 5x109 CFU LmoEGDe-lux,
LmoEGDe-mur-lux, LO28 und LO28-mur in IFN-β-Reportermäusen BL6albino/IFN-
β+/Δβ-luc. .................................................................................................................................95
Abbildung 5-1: Transformationsstrategie zur Herstellung genetisch modifizierter
Listerienstämme. ................................................................................................................ 133
Abbildung 5-2: Prinzip des in vivo Imaging. ........................................................................ 146

Zusammenfassung
xv
Zusammenfassung
In vivo Analyse der oralen Infektion mit biolumineszenten
Listeria monocytogenes im Mausmodell
Silke Bergmann
Das Gram-positive Bakterium Listeria monocytogenes kann schwerwiegende
Infektionen in Mensch und Tier auslösen. Nach der oralen Aufnahme sind Listerien in
der Lage sowohl die Darmbarriere als auch die Blut-Hirn-Schranke und die
Plazentaschranke zu durchqueren.
Die Eintrittspforte für Listeria monocytogenes ist das intestinale Epithelium. Aufgrund
einer speziesspezifischen Interaktion des Oberflächenproteins Internalin A (InlA) mit
seinem Rezeptor E-Cadherin ist die Infektion muriner Epithelzellen und damit eine
Infektion der Maus über den oralen Infektionsweg nur schwer induzierbar. Zwei
Aminosäuresubstitutionen innerhalb des Internalin A erhöhen die Bindungsaffinität
zum murinem E-Cadherin und ermöglichen eine effizientere orale Infektion mit
Listeria monocytogenes im Mausmodell. Derart Internalin A modifizierte
Listerienstämme werden als murinisierte Listerienstämme bezeichnet.
In dieser Arbeit wurden verschiedene Mausinzuchtstämme (BALB/cJ, C57BL/6J,
A/JOlaHsd, C3HeB/FeJ) mit biolumineszenten Listerienstämmen oral infiziert, um
Infektionsverläufe in vivo nicht invasiv zu analysieren. Verglichen wurden
Infektionsverläufe mit dem biolumineszenten InlA-Wildtypstamm LmoEGDe-lux und
dem InlA-modifizierten, murinisierten Stamm LmoEGDe-mur-lux. Es konnte ein
invasiverer Krankheitsverlauf nach der Infektion mit den murinisierten Listerien im
Vergleich zu den isogenen Wildtypstämmen beobachtet werden. Dies resultierte in
einer reduzierten Überlebensrate und einer höheren bakteriellen Keimzahl in den
inneren Organen. Mit Hilfe des Biolumineszenz-Imaging konnten Unterschiede in der
Dissemination und Akkumulation von LmoEGDe-lux und LmoEGDe-mur-lux
festgestellt werden. Weiterhin konnte mit Hilfe von IFN-β-Luziferase-Reportermäusen

Zusammenfassung
xvi
gezeigt werden, dass LmoEGDe-mur-lux eine stärkere IFN-β-Synthese in
verschiedenen Geweben im Vergleich zu seinem isogenen Wildtypstamm induziert.
Einen Einfluss des murinisierten InlA auf die Überwindung der Blut-Hirn-Schranke
konnte nicht gezeigt werden.
Weiter wurde beobachtet, dass die untersuchten Mausinzuchtstämme
unterschiedliche Wirtsresistenzen gegenüber oral vermittelter Listeriose aufwiesen.
C57BL/6J Mäuse zeigten eine ausgeprägte Resistenz gegenüber der oralen Infektion
mit Listeria monocytogenes. Im Vergleich dazu erwiesen sich C3HeB/FeJ und
A/JOlaHsd Mäuse als äußerst empfindlich, was in einer geringeren Überlebensrate
und einer erhöhten bakteriellen Keimlast resultierte. Parallel zeigte die Analyse der
Chemokine und Zytokine bei den empfindlichen C3HeB/FeJ Mäusen eine gesteigerte
Produktion von IFN-γ, CCL2, IL-6, IL-10 und TNF-α im Vergleich zu den resistenten
C57BL/6J Mäusen.
Im Rahmen dieser Dissertation wurde das biolumineszente Listerieninfektionsmodell
auch genutzt, um die Wirtsresistenz von Interferon-α inducible protein 27 like 2A
Knock-out Mäusen (Ifi27l2a-KO) zu untersuchen. Im Vergleich zur Wildtypmauslinie
C57BL/6N resultierte die Infektion mit LmoEGDe-mur-lux in einer signifikant höheren
bakteriellen Keimlast in der Leber und der Milz von Ifi27l2a-KO Mäusen. Jedoch
konnten keine signifikanten Unterschiede in der Überlebensrate beider Mausstämme
gezeigt werden.

Summary
xvii
Summary
In vivo analysis of orally infected mice using bioluminescent
Listeria monocytogenes
Silke Bergmann
The gram-positive bacterium Listeria monocytogenes causes invasive, often fatal
infections in humans and animals. After oral ingestion Listeria monocytogenes can
cross the intestinal, blood-brain, and placental barrier.
The intestinal epithelium is the primary site for Listeria monocytogenes infection. Due
to a species-specific interaction of the bacterial surface protein Internalin A (InlA) with
its receptor E-Cadherin, an infection of murine epithelial-cells, and hence an infection
of mice via the oral route of infection is less effective. Substitution of two amino acids
in Internalin A increased the affinity of the protein to the murine E-Cadherin receptor.
Such modified Listeria strains designated as murinized Listeria strains have the
capability to efficiently pass through the murine intestinal barrier and to induce
Listeriosis in mice after oral inoculation.
In this PhD thesis work, different inbred strains of mice (BALB/cJ, C57BL/6J,
A/JOlaHsd, and C3HeB/FeJ) were intragastrically infected with bioluminescent
Listeria strains to study bacterial dissemination by non-invasive imaging in vivo. The
course of infection was compared between mice infected with the InlA-wildtype
LmoEGDe-lux Listeria strain and mice infected with InlA-modified murinized Listeria
strain LmoEGDe-mur-lux. Infections with LmoEGDe-mur-lux resulted independent of
the host genetic background in more severe Listeriosis disease symptoms as
compared to infections with the wildtype LmoEGDe-lux strain, and mice infected with
LmoEGDe-lux displayed reduced survival and higher bacterial loads in internal
organs. Differences in the systemic dissemination and accumulation of both
bioluminescent Listeria monocytogenes strains in internal organs were detected by
Bioluminescence-Imaging (BLI). By using a luciferase tagged IFN-β-reporter mouse it

Summary
xviii
was shown that infections with the LmoEGDe-mur-lux strain were associated with a
stronger induction of the IFN-β response in several tissues as compared to infections
with the LmoEGDe-lux wild-type strain. Importantly, an influence of the murinized
Internalin A on listerial invasion of the blood-brain was not detected.
Furthermore, the phenotyping of different inbred mouse strains after oral inoculation
with murinized and non-murinized Listeria strains revealed genetic differences in host
resistance to orally acquired Listeriosis. C57BL/6J mice showed a greater resistance
to orally acquired Listeriosis as compared to the other strains. C3HeB/FeJ and
A/JOlaHsd mice showed a more pronounced host susceptibility after oral infection,
which was reflected by a slower bacterial clearance and reduced survival.
Furthermore, the analysis of chemokines and cytokines in the serum after oral
infection with Listeria monocytogenes revealed higher concentrations of IFN-γ,
CCL2, IL-6, IL-10 and TNF-α in susceptible C3HeB/FeJ mice as compared to
resistant C57BL/6J.
The bioluminescent Listeria infection challenge model was also employed to study
the host response of Interferon-α inducible protein 27 like 2A knock-out (Ifi27l2a-KO)
mice. The infection of Ifi27l2a-KO mice with LmoEGDe-mur-lux resulted in a
significantly higher bacterial load in the liver and spleen as compared to C57BL/6N
wildtyp mice. However, there were no significant differences in the survival rate of
both mouse strains detectable.

Einleitung
1
1 Einleitung
In der wissenschaftlichen Forschung existieren eine Vielzahl von Tiermodellen. Sie
spielen eine große Rolle in den verschiedensten Bereichen der Grundlagen- und
angewandten Forschung wie zum Beispiel in der Genetik, der Verhaltensforschung,
der medizinischen und pharmazeutischen Forschung, für das Verständnis bioche-
mischer Prozesse sowie für das Verstehen der Interaktion zwischen Wirt und Patho-
gen während einer infektiösen Erkrankung.
Wie sich eine Krankheit entwickelt, hängt von verschiedenen inneren und äußeren
Einflüssen ab. Die genetische Prädisposition ist eine der wichtigsten intrinsischen
Faktoren. Um die Ursachen genetischer Krankheitsprädisposition zu erforschen, wird
im Tiermodell Maus häufig die genetische Diversität zwischen Mausinzuchtstämmen
für genetische Analysen genutzt (O`Brien und Woychik, 2003). Mit Hilfe von In-
zuchtstämmen lassen sich auch komplexe Genotyp - Phänotyp Beziehungen erklä-
ren (Buer und Balling, 2003; Lengeling et al., 2001). Auch die Wirtsspezifität und
Virulenz eines Krankheitserregers beeinflusst die Entwicklung einer Erkrankung. Ein
viel studiertes Pathogen für den Infektionsprozess, die angeborene und erworbene
Immunabwehr, die intrazelluläre Lebensweise und die Wirtsspezifität ist Listeria
monocytogenes.
1.1 Morphologie und Physiologie der Gattung Listeria
Listerien sind Gram-positive, nicht sporenbildende, feine Stäbchenbakterien mit ge-
ringem GC-Gehalt im Genom. Sie sind peritrich begeißelt und zeichnen sich durch
eine Beweglichkeit bei 20°C aus, sind jedoch nur eingeschränkt beweglich ab 37°C
(Leifson und Palen, 1955; Peel et al., 1988). Eine Besonderheit ist ihre intrazelluläre
Lebensweise. Außerdem leben Listerien fakultativ anaerob, sind anspruchslos und in
der Umwelt weit verbreitet. Diese Eigenschaften haben alle Mitglieder der Gattung
Listeria - Listeria monocytogenes, Listeria ivanovii, Listeria inocua, Listeria
welshimeri, Listeria seegligeri, Listeria grayi (Vazquez-Boland et al., 2001) sowie die
2010 neu beschriebenen Spezies Listeria marthii (Graves et al., 2010) und Listeria
rocourtiae (Leclercq et al., 2010) gemeinsam (Tabelle 1.1). Jedoch sind nur L.
Einleitung
2
monocytogenes und L. ivanovii von human- und tierpathogener Bedeutung (Glaser et
al., 2001). Sie können im Erdreich, im Wasser, auf Pflanzen und in Nahrungsmitteln
tierischen und pflanzlichen Ursprungs isoliert werden. Sie tolerieren extreme pH-
Werte (3,0 - 9,5), hohe Salzkonzentrationen (10%), einige Schwermetalle und
besitzen die Fähigkeit sich bei extremen Temperaturen (-0,1 - 45°C) zu vermehren
(Farber und Peterkin, 1991). Aufgrund dieser Eigenschaften stellen Listerien ein
großes Problem in der lebensmittelverarbeitenden Industrie dar. Der kulturelle
Nachweis erfolgt meist aerob auf Blut-Agarplatten. Es entstehen kleine graue
Kolonien, welche von einer leichten Hämolysezone umgeben sind. Dies ist Resultat
des Listeriolysin O (LLO), ein porenformendes Toxin, welches Erythrozyten lysieren
und Hämoglobin abbauen kann.
Tabelle 1-1: Die Arten der Gattung Listeria
Spezies Humanpathogen Serogruppen
L. monocytogenes ja 13
L. ivanovii (ja) 1
L. innocua nein (3)
L. seeligeri nein (4)
L. welshimeri nein 2
L. grayi nein
L. marthii (Graves et al., 2010) nein
L. rocourtiae (Leclercq et al., 2010) nein
1.2 Epidemiologie von Listeria monocytogenes
Listeria monocytogenes ist weltweit verbreitet und kann eine Listeriose bei Schafen
und Rindern auslösen. Es kann aber auch beim Menschen zu einer schweren loka-
len oder generalisierten Infektion führen (Swaminathan und Gerner-Smidt, 2007).
Erstmals wurde Listeria monocytogenes 1926 als Ursache einer Epidemie in Meer-
schweinchen und Kaninchen beschrieben (Murray et al., 1926). Rückblickend waren
einige Fälle von Listeriose in Menschen und Tieren schon in den 1920-ern bekannt.
Die ersten durch Listerien verseuchte Nahrungsmittel bedingte Infektionen im Men-
schen wurden 1979 mit 23 Fällen dokumentiert (Gellin und Broome, 1989). Ein weite-

Einleitung
3
res Beispiel für einen Ausbruch fand 1981 durch kontaminierten Krautsalat in den
Maritimprovinzen Kanadas statt (Schlech, 1983). Auch sind sporadische Fälle wie
zum Beispiel in Italien 1997, verursacht durch kontaminierten Maissalat, beschrieben
(Aureli et al. 2000). In Europa ist die Anzahl der Listerieninfektionen in den letzten
Jahren wieder angestiegen. Besonders bei den über 70-jährigen konnte ein Anstieg
beobachtet werden (Denny und McLauchlin, 2008; Epidemiologisches Bulletin, 2009
Robert-Koch-Institut (RKI)). Zu den weiteren diskutierten Hypothesen zählen der
demografische Wandel und der zunehmende Verzehr sogenannter „ready to eat“-
Produkte, welche einen günstigen Nährboden für Listerien darstellen. Meist werden
Listerien jedoch über kontaminierte Milchprodukte übertragen (Denny und
McLauchlin, 2008). Auch zeigten die in 2011 in den USA durch verseuchte
Cantaloupe-Melone hervorgerufenen Infektionen, dass Listerien immer wieder eine
wichtige Rolle bei epidemieartigen Ausbrüchen von Infektionskrankheiten spielen
(MMWR Morb Mortal Wkly Rep. 2011, Center for Disease Control and Prevention
(CDC)). Trotzdem sind Fälle von Listeriose selten, in Europa mit 0,3 Fällen auf
100.000 Einwohner (Denny und McLauchlin, 2008), bzw. 0,1-11,3 Fälle auf
1.000.000 Einwohner pro Jahr (Epidemiologisches Bulletin, 2006, RKI). Infektionen
betreffen meist Risikogruppen wie Schwangere, Patienten mit geschwächtem
Immunsystem, ältere Menschen und Neugeborene.
Neben dem oralen Übertragungsweg sind weitere Infektionswege möglich wie z.B.
Hautinfektionen nach intensivem Kontakt mit verseuchtem Wasser oder zu er-
krankten Tieren. Weiterhin können Übertragungen von Mensch zu Mensch als
intrauterine oder perinatale Infektionen erfolgen. Auch tragen 5% der Erwachsenen
symptomlos Listeria monocytogenes in ihrem Gastrointestinaltrakt und stellen somit
eine Infektionsquelle dar (Schuchat et al., 1991).
Listeria monocytogenes wird in verschiedene Serotypen unterteilt. Mit Hilfe der
Oberflächen- (O) und Flagellen- (H) Antigene konnten 13 unterschiedliche Serotypen
identifiziert werden (Donker-Voet et al., 1972; Paterson et al., 1940; Seeliger et al.,
1984), welche genetisch variieren. Nur 4 Serotypen, 1/2a, 1/2b, 1/2c und 4b, sind
human- und tierpathogen, wovon das Serovar 4b die Hauptquelle für epidemische
Ausbrüche darstellt (Gahan und Hill, 2005). Mit Hilfe weiterer Analysen wie

Einleitung
4
Pulsfeldgelelektrophorese, Ribotypisierung, DNA-Arrays und Multi-Locus-Sequenz-
Typisierungen (MLST) wurde gezeigt, dass Listeria monocytogenes Stämme in drei
separate Linien klassifiziert werden können (Ragon et al., 2008). Linie I enthält die
epidemischen Listeria monocytogenes Stämme mit höchster Pathogenität
(Serotypen: 4b, 1/2b, 3b, 4d, 4e und 7), Linie II die Typen, welche zu sporadischen
Erkrankungen im Menschen führen (Serotypen: 1/2a, 1/2c, 3a und 3c) und Linie III
enthält Typen niederer Pathogenität, welche bisher nicht aus humanen Proben
isoliert wurden (Serotypen: 4a und 4c) (Brosch et al., 1994; Graves et al., 1994;
Wiedmann et al., 1997b).
1.3 Humane Listeriose
Es existieren zwei pathogene Spezies der Gattung Listeria, Listeria monocytogenes
und Listeria ivanovii. Listeria monocytogenes ist human- und tierpathogen, wogegen
Listeria ivanovii hauptsächlich bei Wiederkäuern zu einer Listeriose führen kann.
Jedoch konnten auch Listeria ivanovii Infektionen beim Menschen beschrieben
werden (Guillet et al., 2010).
Die Infektion des Menschen mit Listeria monocytogenes erfolgt durch die Aufnahme
mit kontaminierten Nahrungsmitteln. Die Infektionsdosis kann von 10 bis 109 Colony
Forming Units (CFU) / mg Nahrungsmittel schwanken. Die Inkubationszeit liegt
zwischen 3-70 Tagen (Epidemiologisches Bulletin, 2006, RKI). Hauptansteckungs-
quelle sind hauptsächlich Rohmilchprodukte, Rohwürste, geräucherter Fisch und
vakuumverpackte, verzehrfertige Produkte. Trotz häufiger Exposition kommt es
selten zu einer beschriebenen Erkrankung, da im gesunden Menschen die Infektion
häufig symptomlos verläuft und eine höhere Infektionsdosis zum Auslösen einer
Erkrankung nötig ist. Selten können sich klinische Symptome einer nicht-invasiven
Gastroenteritis entwickeln, welche nach wenigen Tagen selbständig ausheilt. Risiko-
gruppen, für die Listeriose ein ernstes Problem darstellt, sind insbesondere schwan-
gere Frauen, Neugeborene, Ältere und immunsupprimierte Patienten z.B. nach
Organtransplantationen, Chemotherapie und AIDS Patienten. Hier ist eine geringere
Infektionsdosis ausreichend, um eine Erkrankung auszulösen.

Einleitung
5
Die häufigste Manifestationsform der Listeriose, bei nicht Schwangeren, betrifft das
zentrale Nervensystem, mit 55-70% der Fälle (Vazquez-Boland et al., 2001).
Bakterielle Meningitis und Enzephalitis resultieren in einer Mortalität von ca. 20%,
können aber im Falle von bestehenden schwächenden Vorerkrankungen eine
Sterblichkeit von 40-60% aufweisen (Lorber, 1997; Schuchat et al., 1997). Eine
weitere häufige Form der Listerieninfektion äußert sich in einer Sepsis (15-50% der
Fälle) und ist begleitet von einer Mortalitätsrate von bis zu 70% (Lorber, 1997).
Atypische Verläufe sind Pneumonie, Endokarditis und Hepatitis (Faber und Perterkin,
1991; Lorber 1997; McLauchlin, 1990). Weiterhin wird eine invasive Gastroenteritis
ebenfalls als Hauptmanifestationsform beschrieben, welche begleitet ist von Diarrhoe
und Fieber (Ho et al., 1986).
Bei der feto-maternalen und neonatalen Listeriose werden zwei Verlaufsformen
unterschieden: „early“ und „late onset“. Aufgrund einer physiologischen Suppression
der zellulär vermittelten Immunantwort während einer Schwangerschaft (Weinberg et
al., 1984) kann es zur Invasion der Plazenta und des Fetus kommen. In der Mutter
selbst verläuft die Infektion meist asymptomatisch. Die Infektion innerhalb der ersten
Schwangerschaftsmonate resultiert häufig in einer Früh- bzw. Totgeburt. Erfolgt eine
Infektion im letzten Drittel der Schwangerschaft entwickeln die Feten und Neu-
geborenen häufig schwerwiegende generalisierte Infektionen, wie Granulomatosis
infantiseptica („early onset“), verbunden mit einer Sterblichkeit von >50% (Klatt et al.,
1986; Schlech, 2000). Auch ist es möglich, dass sich die Neugeborenen während
des Geburtsvorgangs über infizierte maternale Exsudate mit Listeria monocytogenes
infizieren („late onset“) (Farber et al., 1991). Weiterhin ist bekannt, dass über
keimtragendes medizinisches Personal eine Ansteckung erfolgen kann. Symptome
sind häufig Meningitis, seltener Pneumonie oder Gastroenteritis. Die Mortalität liegt
hier bei ca. 5-10% (Lorber, 1997).
Seltener vorkommende Infektionen wie Dermatitis und Konjunktivitis können bei
Personen mit intensivem direktem Kontakt zu infizierten Tieren auftreten (McLauchlin
und Low, 1996).

Einleitung
6
Die Diversität der klinischen Symptome resultiert aus der Fähigkeit Listeria mono-
cytogenes drei Wirtsbarrieren zu durchqueren: die intestinale Barriere, die Plazenta-
schranke und die Blut-Hirn-Schranke (Vazquez-Boland et al., 2001). Nach Aufnahme
mit kontaminierten Lebensmitteln überleben Bakterien die Magensäure sowie allge-
meine Abwehrmechanismen und können die intestinale Barriere überqueren. Sie
infizieren sowohl die intestinalen Epithelzellen als auch die M-Zellen der Peyer´schen
Plaques. M-Zellen sind Bestandteile des Follikel-assoziierten Epithels (FAE). Sie
überdecken die Peyer-Plaques und spielen eine wichtige Rolle für die Funktion des
„Schleimhaut-assoziiertes lymphatisches Gewebe“ (MALT). M-Zellen nehmen Anti-
gene und Mikroorganismen über Endozytose auf und leiten sie an darunterliegende
lymphoide Zellen weiter, welche dann anschließend eine Immunantwort generieren
(Clark et al., 2000). Über das Blut- und Lymphsystem sowie die mesenterialen
Lymphknoten gelangen die Listerien zu Leber und Milz, den Hauptorganen der bak-
teriellen Kolonisation und Replikation (Vazquez-Boland et al., 2001). Über eine
erneute hämatogene Streuung gelangt Listeria monocytogenes zum Gehirn und zur
Plazenta und können auch dort die epithelialen Barrieren überqueren (Abbildung 1-
1).
Listeriose ist erfolgreich mit Antibiotika behandelbar. Auch sind bisher im klinischen
Bereich keine resistenten Stämme, im Gegensatz zu Isolaten aus Lebensmitteln,
aufgetreten. Die Dauer der Behandlung ist von der Wahl des Antibiotikums und den
klinischen Symptomen abhängig und kann bis zu sechs Wochen in Anspruch neh-
men. Gute Ergebnisse liefert eine Kombinationstherapie mit hoch dosiertem
Ampicillin und Gentamycin. Zusätzlich empfiehlt sich eine Gabe von Rifampicin zur
Inhibition des intrazellulären Lebenszyklus der Bakterien (Epidemilogisches Bulletin,
2006 (RKI)). Die beste Weise sich vor einer Listerieninfektion zu schützen, ist jedoch
die Vermeidung des Verzehrs kontaminierter Lebensmittel. Richtlinien in der
nahrungsmittelverarbeitenden Industrie sollen das Risiko einer Infektion reduzieren.
Trotz aller Verarbeitungsvorschriften sind Rohprodukte oder mangelnde Hygiene bei
der Verarbeitung potentiell kontaminierter Lebensmittel eine ständige Gefahr für
Listerieninfektionen. Besonders gefährdete Nahrungsmittel sind Rohmilchprodukte,
Weichkäse, rohes Fleisch, Räucherfisch und unsachgemäß gereinigtes Gemüse.

Einleitung
7
Ausnahmen bilden Äpfel, Karotten und Tomaten. Diese sind frei von Listerien
(Epidemiologisches Bulletin, 2006, RKI). Eine allgemeine Empfehlung zur Prävention
einer Listerieninfektion ist Fleisch vor dem Verzehr ausreichend zu erhitzen, pasteu-
risierte Milch und Milchprodukte zu verwenden und Gemüse ausgiebig zu waschen.
Abbildung 1-1: Infektionsweg von Listeria monocytogenes Nach Aufnahme über kontaminierte Nahrungsmittel überquert Listeria monocytogenes die intestinale Barriere und erreicht über das Lymph- und Blutsystem die Leber und Milz. Über eine erneute hämatogene Streuung erreichen die Listerien die Plazenta und das Gehirn und können sowohl die Plazentaschranke als auch die Blut-Hirn-Schranke überqueren (verändert nach Lecuit et al., 2007).
1.4 Listeriose in domestizierten Tieren
Listeria monocytogenes hat große Bedeutung in der Veterinärmedizin (Gray und
Killinger, 1966). Zudem tragen eine Vielzahl von Wild- und Nutztieren Listeria
monocytogenes asymptomatisch in ihrem Verdauungstrakt (Nightingale et al., 2004).
Listeriose, auch „circling disease“ oder „silage sickness“ genannt, betrifft meist
Wiederkäuer (Pfister et al., 2002; Wiedmann et al., 1997a). Auch sind Fälle von
Listeriose bei Kaninchen, Meerschweinchen, Schweinen, Hunden, Katzen und
Vögeln beschrieben wurden (Cooper, 1989; Decker et al., 1976; Long und Dukes,
1972; Murray, 1955). Ebenfalls ist Listeria ivanovii von Bedeutung eine Listeriose bei
Schafen auszulösen,
Bei Schafen und Rindern sind ca. 52% der Tiere mit Listeria monocytogenes infiziert
und alle Altersstufen sowie beide Geschlechter betroffen (Skovgaard und Morgen,
1988). Dennoch ist die Erkrankung selten. Tiere unter drei Jahren sind mehr

Einleitung
8
gefährdet als ältere. Jahreszeitlich häufen sich Fälle von Listeriose in den Winter-
und Frühlingsmonaten. Die Infektion erfolgt meist über kontaminiertes Futter wie
Silage oder verseuchte Weiden. Die Inkubationszeit variiert zwischen wenigen Tagen
bis zu mehreren Wochen (Low und Donachie, 1997). Nach der Aufnahme können
sich die Bakterien im Gastrointestinaltrakt anreichern und die intestinale Barriere
durchqueren. Weiterhin ist es möglich, dass Listeria monocytogenes über
Verletzungen der Mundschleimhaut eindringen kann und vom Rachenraum, entlang
der Hirnnerven, zum zentralen Nervensystem wandert und eine Meningoenzephalitis
verursacht (Barlow und McGorum, 1985).
Bei Wiederkäuern werden verschiedene Formen der Listeriose unterschieden. Adulte
Tiere erkranken gewöhnlich an der enzephalitischen Form. Hier kann die
Inkubationszeit zwei bis sechs Wochen andauern (Low und Donachie, 1997). Diese
ist anfangs gekennzeichnet durch Lethargie und Gewichtsverlust. Später folgen
neurologische Ausfallerscheinungen wie Gesichtslähmung, übermäßige
Speichelbildung, unkoordinierte Bewegungen und einer Torticollis. Innerhalb von 4 –
14 Tagen endet die Erkrankung tödlich, die Mortalität liegt bei Schafen bei 50% und
bei Rindern bei 71% (Czuprynski, 1993). Neugeborene oder junge Tiere erkranken
dagegen eher an der septischen oder viszeralen Form mit Fieber und Lethargie mit
späterer Todesfolge. Septische Verlaufsformen zeichnen sich durch eine
Inkubationszeit von zwei bis fünf Tagen aus und sind von der inokulierten
Listerienanzahl abhängig (Low und Renton, 1985). Ebenso kann es zu intrauterinen
Infektionen kommen, welche zu Frühgeburten und Aborten führen (Cooper und
Walker, 1998). Weitere Krankheitsbilder sind Keratokonjunktivitis oder Mastitis und
werden durch Fütterung mit kontaminierter Silage verursacht (Cooper und Walker,
1998). Die Diagnose wird meist über die klinischen Symptome gestellt. Die definitive
Diagnose erfolgt post-mortem mit Hilfe der histopathologischen Analyse. Die
Antibiotika der Wahl sind PenicillinG und Tetrazykline. Vorbeugende Maßnahmen um
eine Listeriose bei Nutztieren zu verhindern, liegt in der Fütterung von einer qualitativ
hochwertiger Silage mit geringem pH-Wert (≤5) (Fenlon, 1986).
Bei Vögeln, Hunden, Katzen, Kaninchen und Meerschweinen führt eine Infektion mit
Listeria monocytogenes zu Sepsis (Gray und Killinger, 1966). Vereinzelt kommt es

Einleitung
9
zum Auftreten einer Enzephalitis. Bei Kaninchen kann es zusätzlich zu Aborten oder
plötzlichem Tod kommen.
1.5 Wirtsspezifität und Wirtsinvasion
1.5.1 Invasionsproteine und Speziesspezifität
Die Aufnahme von Listeria monocytogenes über kontaminierte Nahrungsmittel macht
Listeriose zu einer wichtigen Zoonose-Erkrankung. Die Übertragung zwischen den
einzelnen Spezies sowie die Invasion und Replikation in der Zelle sind von der Inter-
aktion von Wirtsmolekülen mit Oberflächenproteinen des Pathogens abhängig. Die
Internaline von Listeria monocytogenes sind dafür von großer Bedeutung. Internaline
sind eine große Proteinfamilie, welche in zwei Genclustern auf dem Genom organi-
siert sind. Die Proteine sind gekennzeichnet über einen gleichen Aufbau. N-terminal
findet sich eine Signalsequenz, gefolgt von leucinreichen Wiederholungen (LRR-
Motiv). Weiter kann sich eine IR Region (inverted repeats) anschließen. Basierend
auf dem Aufbau und der bakteriellen Oberflächenbindungsdomäne werden die
Internaline in drei Familien unterteilt (Bierne et al., 2007; Cossart und Toledo-Anna,
2007): Internaline mit LPTXG- Motiv, wobei das X für jede beliebige Aminosäure
stehen kann, Internaline mit GW- oder WXL-Motiv und die dritte Gruppe sind
sekretierte Internaline (Hamon et al., 2006). Zu den LPTXG-Motiv Internalinen
gehören 19 Mitglieder darunter Internalin A (InlA), InlE, InlG, InlH, InlF, InlI und InlJ
(Sabet et al., 2005). Über dieses Motiv erfolgt eine kovalente Bindung an die
bakterielle Zellwand. Die zweite Klasse von Internalinen, GW oder WXL, beinhaltet
zwei Proteine, Internalin B (InlB) und Lmo0549. Über GW bzw. WXL Wiederholungen
erfolgt eine lockere Bindung des Proteins an die Lipoteichonsäuren der bakteriellen
Zellwand. In der dritten Gruppe der Internaline fehlt die zellwandverankernde
Domäne. Hierzu gehören vier Mitglieder darunter InlC, Lmo2445, Lmo2027 und
Lmo2470.
Die am besten charakterisierten Internaline sind InlA und InlB. Sie spielen die Haupt-
rolle bei der Invasion von Listeria monocytogenes in verschiedene Zelltypen. Der
Rezeptor für Internalin B ist die Rezeptor-Tyrosinkinase c-Met/HGF-R (mesenchymal

Einleitung
10
epithelial transition factor / hepatocyte growth factor Rezeptor) (Shen et al., 2000).
Dieser spielt eine Rolle während der Entwicklung, Geweberegeneration und Zell-
differenzierung und wird auf einer Vielzahl von Zellen exprimiert. InlB bindet an den
c-Met Rezeptor von Mensch und Maus (Met), erkennt aber nicht an das c-Met
Protein von Meerschweinchen und Kaninchen (Khelef et al., 2006). Internalin B kann
zusätzlich an die globuläre Kopfdomäne des Komplement C1q Moleküls (gC1qR)
binden (Braun et al., 2000) sowie über die GW-Domäne an Glukosaminoglykane
(Ebbes et al., 2011), was die InlB-c-Met Interaktion während der Zellinvasion
unterstützt. Internalin A spielt eine Rolle bei der Invasion von intestinalen
Epithelzellen, Endothelzellen und Hepatozyten. Es interagiert mit E-Cadherin, einem
calciumabhängigen Zell-Adhäsionsmolekül, welches während der Entwicklung, bei
der Polarisierung von Epithelien und in der Aufrechterhaltung der Gewebearchitektur
von Bedeutung ist (Mengaud et al., 1996b). E-Cadherin ist innerhalb der Familie der
Säugetiere hoch konserviert. Dabei sind 85% des humanen E-Cadherins mit dem
murinem E-Cadherin (Cdh1) identisch. Internalin A interagiert mit humanem E-
Cadherin, bindet aber auch E-Cadherin vom Kaninchen und vom Meerschweinchen,
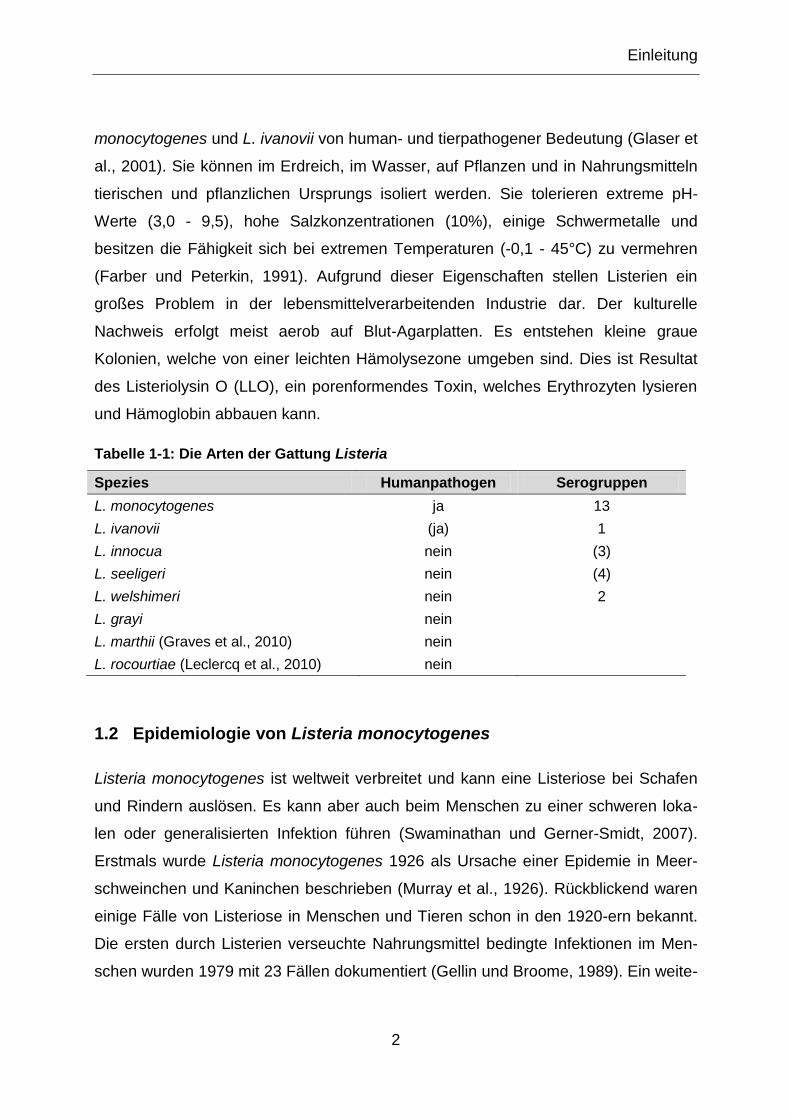
ist jedoch nicht fähig mit Maus-E-Cadherin zu interagieren. Daraus folgt, dass Mäuse
nicht zum natürlichen Wirt von Listeria monocytogenes gehören (Abbildung 1-2). Der
wichtigste Unterschied für die Speziesspezifität liegt in der Aminosäureposition 16
des E-Cadherins. Humanes E-Cadherin sowie das vom Meerschweinchen und
Kaninchen tragen an dieser Position ein Prolin. Innerhalb des murinen E-Cadherins
ist dagegen ein Glutamin positioniert (Lecuit et al., 1999). Wird Glutamin mit Prolin an
der Position 16 ausgetauscht, ist das Internalin A fähig mit diesem humanisierten
Maus-E-Cadherin zu interagieren (Lecuit et al., 1999; Lecuit et al., 2005). Dies
resultiert in einer verstärkten Interaktion des Rezeptors mit seinem Liganden
(Schubert et al., 2002). Ein anderer Weg liegt in der Murinisierung des Internalin A.
Zwei Aminosäuresubstitutionen erhöhen die Affinität des Internalin A zum E-
Cadherin. Zum einen ein Austausch von Serin zu Asparagin an der
Aminosäureposition 192 und zum anderen der Austausch von Tyrosin zu Serin an
der Position 369 (Wollert et al., 2007b). Über die Bestimmung der
Dissoziationskonstanten von Proteinkomplexen konnte gezeigt werden, dass der
Austausch von Serin zu Asparagin an Position 192 des Internalin A die Affinität zu
Einleitung
11
humanen E-Cadherin um den Faktor 40 erhöht. Erfolgt der zusätzliche Austausch
vom Tyrosin zu Serin an der Aminosäureposition 369 des InlA, so erhöht sich die
Bindungsaffinität zum E-Cadherin erneut um den Faktor 170 (in Summe 6700-facher
Anstieg der Bindungsaffinität). Durch diese beiden Modifikationen wird auch die
Affinität des Internalin A zu Maus-E-Cadherin erhöht. Die Bindungsaffinität der
Proteinkomplexe von InlA S192N-Y369S zu murinem E-Cadherin wird um ein
vierfaches erhöht und ist vergleichbar mit der Bindungsaffinität der Proteinkomplexe
von Wildtyp InlA zu humanem E-Cadherin (Wollert et al., 2007a).
Abbildung 1-2: Speziesspezifität von Listeria monocytogenes InlA und InlB bindet an Zelloberflächenrezeptoren (E-Cadherin und c-Met) und induziert die Invasion von Listeria monocytogenes in humane Zellen. Ein Aminosäureaustausch an Posi-tion 16 des E-Cadherins verhindert die Bindung von InlA an Maus-E-Cadherin. InlB kann nicht an c-Met von Meerschweinchen und Kaninchen binden (verändert nach Bonazzi et al., 2009).

Einleitung
12
1.5.2 Wirtsinvasion und intrazelluläre Lebensweise
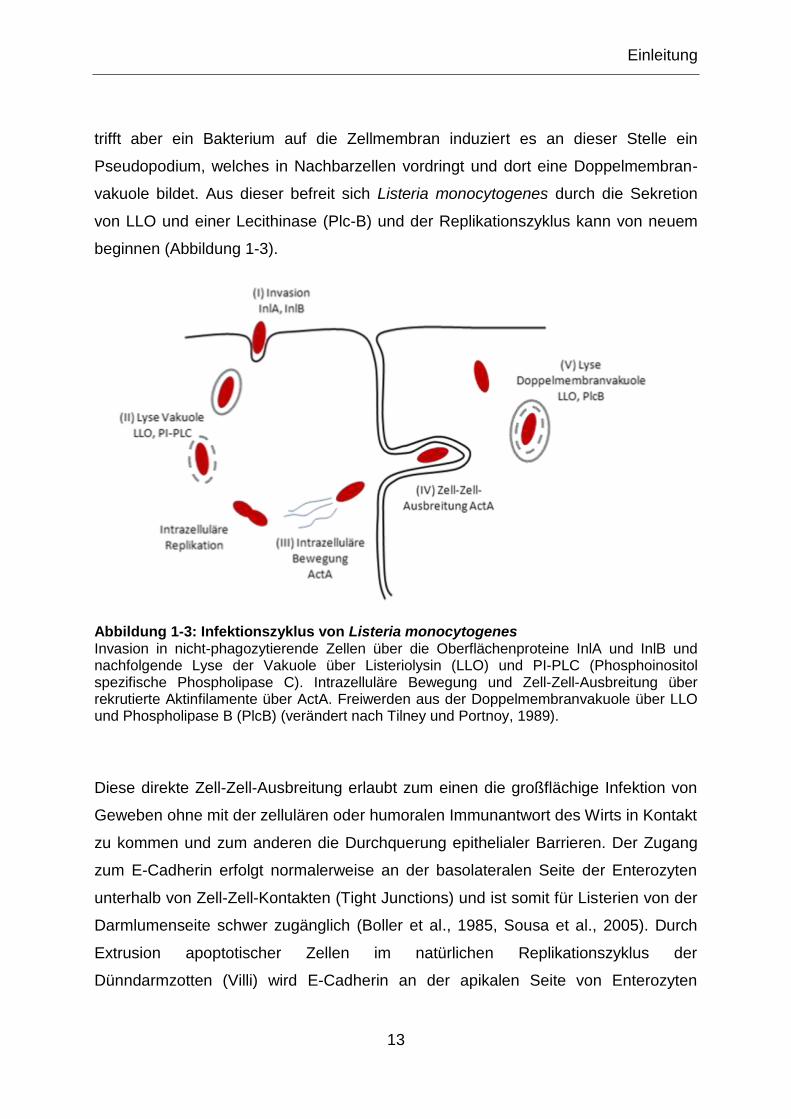
Invasive Bakterien haben verschiedene Strategien entwickelt um ihre eigene Auf-
nahme in nicht-phagozytierende Zellen zu induzieren, den Zipper- und Trigger-
Mechanismus. Die Aufnahme durch Trigger-Mechanismus benötigt keinen physio-
logischen Kontakt zur Wirtszelle. Bakterielle Induktoren werden sekretiert und indu-
zieren bei der Wirtszelle ein kräuseln der Membran, wobei das Pathogen in einer
Vakuole umschlossen wird. Listeria monocytogenes dringt in die Wirtszelle über
Zipper-Mechanismus ein. Invasionsproteine (InlA und InlB) binden an ihre Rezep-
tormoleküle und induzieren durch Umbau des Zytoskeletts Membranausstülpungen,
welche das Bakterium einschließen (Mostowy und Cossart, 2009).
Listeria monocytogenes kann über Makrophagen phagozytiert werden oder die
eigene Aufnahme in nicht-phagozytierende Zellen (Endothelzellen, intestinale
Epithelzellen, M-Zellen der Peyer’schen Plaque) induzieren. Bei letzterem spielen die
Oberflächenmoleküle Internalin A und Internalin B eine Schlüsselrolle (Pizzaro-Cerdà
et al., 2010). Internalin A erkennt und bindet über die LRR-Domäne an die Ekto-
domäne des E-Cadherins (Schubert et al., 2002). Dies führt zur Aktivierung von α-
und β-Catenin, welche dann mit dem Aktin des Zytoskeletts interagieren (Lecuit et
al., 2000) und durch Polarisierung und Umbau des Zytoskeletts eine Phagozytose-
vakuole bilden. Als zweites führt die Bindung des Internalin B an seinen Rezeptor c-
Met über GTPase-Aktivierung zur Polymerisierung und Verlängerung von Aktin-
molekülen und damit zur Ausbildung von Membranausstülpungen um das Bakterium
herum. Weitere Virulenzfaktoren, die bei der Invasion eine Rolle spielen, sind p60
und p104 (beides Adhäsionsproteine) sowie InlC, welches InlA und InlB bei der Inva-
sion in nicht-phagozytierende Zellen unterstützt (Bergmann et al., 2002, Rajabian et
al., 2009). Ist Listeria monocytogenes einmal phagozytiert, liegt es innerhalb eines
Phagolysosoms intrazellulär vor. Diese Vakuole wird durch Expression von Listerio-
lysin (LLO), Metalloprotease (Mpl) und einer Phosphatidylinositol-spezifischen Phos-
pholipase (PI-PLC) lysiert. Das Bakterium liegt dann frei im Zytoplasma und startet
mit der Replikation. Für die intrazelluläre Bewegung rekrutiert Listeria
monocytogenes Aktinfilamente und polymerisiert diese durch die Expression von
dem Aktin-bindenden Protein A (ActA). Die Bewegung von Listerien ist ungerichtet,
Einleitung
13
trifft aber ein Bakterium auf die Zellmembran induziert es an dieser Stelle ein
Pseudopodium, welches in Nachbarzellen vordringt und dort eine Doppelmembran-
vakuole bildet. Aus dieser befreit sich Listeria monocytogenes durch die Sekretion
von LLO und einer Lecithinase (Plc-B) und der Replikationszyklus kann von neuem
beginnen (Abbildung 1-3).
Abbildung 1-3: Infektionszyklus von Listeria monocytogenes Invasion in nicht-phagozytierende Zellen über die Oberflächenproteine InlA und InlB und nachfolgende Lyse der Vakuole über Listeriolysin (LLO) und PI-PLC (Phosphoinositol spezifische Phospholipase C). Intrazelluläre Bewegung und Zell-Zell-Ausbreitung über rekrutierte Aktinfilamente über ActA. Freiwerden aus der Doppelmembranvakuole über LLO und Phospholipase B (PlcB) (verändert nach Tilney und Portnoy, 1989).
Diese direkte Zell-Zell-Ausbreitung erlaubt zum einen die großflächige Infektion von
Geweben ohne mit der zellulären oder humoralen Immunantwort des Wirts in Kontakt
zu kommen und zum anderen die Durchquerung epithelialer Barrieren. Der Zugang
zum E-Cadherin erfolgt normalerweise an der basolateralen Seite der Enterozyten
unterhalb von Zell-Zell-Kontakten (Tight Junctions) und ist somit für Listerien von der
Darmlumenseite schwer zugänglich (Boller et al., 1985, Sousa et al., 2005). Durch
Extrusion apoptotischer Zellen im natürlichen Replikationszyklus der
Dünndarmzotten (Villi) wird E-Cadherin an der apikalen Seite von Enterozyten

Einleitung
14
kurzzeitig freigelegt und für Listeria monocytogenes zugängig (Pentecost et al., 2006;
Pentecost et al., 2010). Ebenso an Epithelfalten der Villi sowie um die schleim-
produzierenden Becherzellen kann E-Cadherin exprimiert sein (Nikitas et al., 2011).
Die meisten Gene, welche die verschiedenen Virulenzfaktoren kodieren, sind im
LIPI-1 (Listeria Pathogenitätsinsel) Cluster, einer 10 kb großen Region auf dem
Chromosom, vereint (Vazquez-Boland, et al., 2001). Hier werden die Gene prfA,
plcA, hly, mpl, actA und plcB kodiert. Dieses Gencluster fehlt bei nicht-pathogenen
Spezies (Gouin et al., 1994). Der inlAB Locus ist getrennt an einer anderen Region
des Chromosoms lokalisiert. Jedoch unterliegen alle bekannten Virulenzgene der
absoluten bzw. partiellen Kontrolle des PrfA Regulatorproteins (Pleiotropisches
Aktivatorprotein) (Mengaud et al., 1991). Befindet sich Listeria monocytogenes
außerhalb der Wirtszelle, liegt PrfA in geringer Konzentration vor. Nach der Adhäsion
von Listeria monocytogenes erfolgt eine verstärkte Expression von prfA. Auch der
Stressfaktor Sigma B wirkt wie PrfA regulierend (Kim et al., 2005), insbesondere bei
Einfluss von äußeren Faktoren wie z.B. Temperatur und pH-Wert.
Das Listeriolysin (LLO) von Listeria monocytogenes ist ein bedeutender Virulenz-
faktor und z.B. bei posttranslationalen Prozessen von infizierten Zellen beteiligt.
Diese posttranslationale Modifikation von Proteinen wird als SUMOylation bezeich-
net. Dabei erfolgt die kovalente Bindung des Zielproteins an ein Ubiquitin-ähnliches
Protein genannt SUMO (Small Ubiquitin-like Modifer). SUMOylation spielt eine Rolle
bei der Regulation der Transkription und der Stressantwort. Listeriolysin hat den
Effekt, dass es zu einer Degradation von SUMOylierten Proteinen in der Wirtszelle
führt und die bakterielle Infektion begünstigt (Ribet et al., 2010). Eine weitere Fähig-
keit des LLO ist die Histonmodifikation. In der Frühphase der Infektion induzieren
extrazellulär vorliegende Listerien die Dephosphorylierung von Serin 10 des H3 und
die Deacethylierung von H4 über sekretiertes LLO. Diese Modifikation resultiert in
der Repression der Expression verschiedener Gene, darunter auch Gene, die bei der
Immunantwort eine Rolle spielen (Hamon et al., 2007; Hamon et al., 2011). Ein
weiterer Virulenzmechanismus ist die Fähigkeit von Listeria monocytogenes sich der
Autophagozytose zu entziehen. Dies ist ein konservierter Prozess der Eukaryoten,
welcher erlaubt intrazelluläre Bestandteile innerhalb einer Doppelmembranvakuole

Einleitung
15
(Autophagosom) abzubauen oder zu recyceln. Über ActA wird die Umlagerung von
Listeria monocytogenes durch ubiquitinierte Proteine verhindert und dadurch die
Autophagosombildung unterdrückt (Birmingham et al., 2007; Yoshikawa et al.,
2009a; Yoshikawa et al., 2009b).
1.5.3 Überquerung epithelialer Barrieren im Wirt
Listeria monocytogenes besitzt die Fähigkeit aufgrund der intrazellulären Lebens-
weise epitheliale Barrieren zu durchqueren, die intestinale Barriere, die Plazenta-
schranke und die Blut-Hirn-Schranke. Der Gastrointestinaltrakt ist die Eintrittspforte,
wobei das Bakterium sich an verschiedene Lebensbedingungen im Organismus an-
passen muss. Bevor Listeria monocytogenes in den Darm übergeht, muss es dem
sauren Milieu des Magens widerstehen. Patienten welche Antazidum und H2-Blocker
einnehmen, haben ein größeres Risiko an einer Listeriose zu erkranken (Ho et al.,
1986; Schuchat et al., 1992). Dies zeigt, dass Magensäure einen Teil der Bakterien
abzutöten vermag. Jedoch ist Listeria monocytogenes in der Lage über die Expres-
sion der Glutamatdecarboxylase (GAD) (Cotter et al., 2001) und der Arginin-
deiminase (ADI) geringe pH-Werte zu tolerieren (Cotter und Hill, 2003). Beide
Systeme werden über den Stressfaktor Sigma B und den Hauptregulator der Viru-
lenzgene (PrfA) kontrolliert. Beim Übergang in den Dünndarm erfolgt aufgrund einer
hohen Salzkonzentration (0,3 M NaCl) ein Anstieg des osmotischen Drucks (Islam et
al., 1996). Durch die Expression von Osmoseregulatoren (betL, gbu und opuC)
erfolgt die Kompensation dieser erhöhten Salzkonzentration innerhalb von Listeria
monocytogenes (Sleator et al., 2003a; Sleator et al., 2003b) und somit die
Anpassung an die umgebenen Verhältnisse. Auch kann Listeria monocytogenes sek-
retiertem Gallensaft widerstehen. Durch die Mechanismen für Gallensaftresistenz,
BSH und BilE (Begley et al., 2002; Begley et al., 2005), ist es dem Bakterium möglich
innerhalb der Gallenblase zu überleben und sich zu vermehren (Hardy et al., 2004).
Die Resistenz gegenüber Gallensaft schwankt innerhalb der verschiedenen
Listerienstämme. So zeigte Listeria monocytogenes LO28 eine hohe Toleranz
gegenüber Gallensäure bis 5 M (Begley et al., 2002; Begley et al., 2005). Intestinales
Epithelium ist außerdem von einer Muzinschicht geschützt. Listeria monocytogenes
hat die Fähigkeit sich über die Oberflächenproteine InlB, InlC und InlJ an humanes

Einleitung
16
Muzin zu binden (Lindèn et al., 2008). Ebenso erhöhen die Flagellen die
Invasionseffizienz (Bigot et al., 2005; O´Neil und Marquis, 2006). Der Eintritt in die
intestinalen Epithelzellen erfolgt hauptsächlich über die InlA – E-Cadherin Interaktion.
InlB spielt keine direkte Rolle, kann aber die Funktion von InlA unterstützen (Disson
et al., 2008). Weitere für die Invasion unterstützende Faktoren sind das
Virulenzprotein in Listeriavirulenz (Vip), das eisenbindende Vorläuferprotein (FbpA)
und Internalin J (InlJ) (Cabanes et al., 2005; Dramsi et al., 2004; Lecuit et al., 2007).
Aufgrund der Speziesspezifität der Internalin A - E-Cadherin Interaktion sind Mäuse
resistent gegenüber einer oralen Listerieninfektion. Durch Generierung einer
transgenen Maus, welche humanes E-Cadherin auf der Oberfläche von Enterozyten
exprimiert, konnte die Rolle des InlA bei der Überquerung der intestinalen Barriere
bestätigt werden (Lecuit et al., 2001). Des Weiteren hat die Murinisierung des
Internalin A das invasive Potential von Listeria monocytogenes in oral infizierten
Mäusen erhöht (Wollert et al., 2007b). Listeria monocytogenes kann somit direkt in
Epithelzellen eintreten (InlA-abhängig) aber auch über die M-Zellen der Peyer´schen
Plaque (InlA-unabhängig) (MacDonald und Carter, 1980) die intestinale Barriere
durchqueren.
Listeria monocytogenes besitzt auch die Fähigkeit die Plazentaschranke zu passie-
ren. Während einer Schwangerschaft erhöht sich das Risiko einer Frau an einer
Listeriose zu erkranken um den Faktor 18 (Southwick und Purich, 1996). Dabei spielt
die physiologische Unterdrückung der zellulär vermittelten Immunantwort während
einer Schwangerschaft, durch die veränderte hormonelle Situation, eine entschei-
dende Rolle (Pung et al., 1985; Weinberg, 1984). Diese schützt den Fetus vor der
Abstoßung, erhöht aber auch die Anfälligkeit für intrauterine Infektionen (Redline et
al., 1988; Redline und Lu, 1987; Redline und Lu, 1988). Experimentell konnte gezeigt
werden, dass in der Endphase einer Schwangerschaft von Mäusen die T-Zell-
vermittelte Eliminierung von Listeria monocytogenes aus der Plazenta verzögert ist.
Dies korreliert mit einer Invasion der Plazenta und des fetalen Gewebes bedingt
durch eine geringere IFN-γ, IL-2 und TNF-α Produktion durch einen erhöhten Östro-
genspiegel (Nakane et al., 1985; Salem et al., 1999). Für die Überquerung der
Plazentaschranke spielen die Interaktionen der Oberflächenproteine InlA und InlB mit

Einleitung
17
ihren Rezeptoren E-Cadherin und c-Met eine entscheidende Rolle. In schwangeren
Meerschweinchen, welche für die InlB-c-Met-Interaktion defizient sind, konnte
demonstriert werden, dass die Durchquerung der plazentalen Barriere Internalin A
unabhängig erfolgen kann (Bakardjiev et al., 2004; Bakardjiev et al., 2005). Jedoch
sind für die effektive Invasion der humanen Plazenta beide intakte InlA- und InlB-
Wege nötig (Lecuit et al., 2004; Lecuit et al., 2009). Auch konnte in schwangeren
Mäusen, defiziente InlA – E-Cadherin – Interaktion, die Bedeutung des InlB für die
Invasion der Plazenta nach intravenöser und oraler Infektion mit Listeria
monocytogenes beschrieben werden (Abram et al., 1997; Abram et al., 2003;
LeMonnier et al, 2006; Poulsen et al., 2011). In in vitro Versuchen mit humanen
Trophoblasten, ein Modell mit intakten InlA und InlB Interaktionswegen, konnte
gezeigt werden, dass nach einer Infektion mit Listerienmutanten für InlA oder InlB
und für die Doppelmutante die Effizienz der Plazentainfektion verringert war (Lecuit
et al., 2004). Dies konnte in der Rennmaus (Gerbillinae), ein Tiermodell für intakte
InlA- und InlB-Wege, bestätigt werden (Disson et al., 2008).
Listeriose geht einher mit einer Infektion des zentralen Nervensystems (ZNS) beim
Menschen als auch bei Nutztieren. Dabei kann Listeria monocytogenes das Gehirn
auf vielfältige Weise erreichen. Beschrieben wurde die hämatogene Streuung der
Bakterien, welche dann direkt in die Endothelzellen der Blut-Hirn-Schranke
eindringen können (Drevets et al., 2004). Dabei wird eine Beteiligung von InlA und
InlB nicht ausgeschlossen. Obwohl Neuronen in der Regel N-Cadherin exprimieren,
welches nicht mit InlA interagiert, konnte exprimiertes E-Cadherin bei primären sen-
sorischen Mausneuronen nachgewiesen werden (Shimamura et al., 1992). InlA-
Mutanten sind ebenfalls weniger effizient bei der Infektion von kultivierten dorsalen
Wurzelganglion-Neuronen (Dons et al., 1999). Der InlB-Rezeptor c-Met wird auf einer
Reihe von Zellen des zentralen Nervensystems exprimiert (Jung et al., 1994; Maina
et al.; 1997). Es konnte gezeigt werden, dass über die InlB–c-Met–Interaktion Listeria
monocytogenes in humane mikrovaskuläre Hirnendothelzellen eindringen kann
(Greiffenberg et al., 1998). Eine Alternative zur Rezeptor-vermittelten Invasion ist die
Durchquerung der Blut-Hirn-Schranke innerhalb infizierter Makrophagen, dem
sogenannten „Trojanischen Pferd“-Modell (Drevets et al., 1993; Drevets, 1999;

Einleitung
18
Drevets et al., 2008; Drevets und Bronze, 2008). Es konnte in vitro nachgewiesen
werden, dass Listeria monocytogenes effizienter über Zell-Zell-Ausbreitung
Neuronen infizieren kann als über eine direkte Invasion (Dramsi et al., 1998). Auch
eine mögliche interne Infektionsquelle könnten persistierende infizierte Monozyten
aus dem Knochenmark sein. Nach intravenöser Infektion von BALB/c und CD1
Mäuse wurde beobachtet, dass listerieninfizierte Makrophagen im Knochenmark
persistieren können (Hardy et al., 2009). Eine weitere Möglichkeit das zentrale
Nervensystem zu infizieren, kann über den neuralen Transport von Listeria
monocytogenes erfolgen und wurde schon früh beschrieben (Asahi et al., 1952;
Dons et al., 2007). Der intraaxonale Transport von Listerien konnte in Schafen nach
der Infektion direkt in die Zahnpulpa bestätigt werden, als sechs von 21 Tieren
neurologische Zeichen entwickelten (Barlow und McGorum, 1985). In einem weiteren
Experiment wurden in die Kranialnerven oder in fasziale Muskeln von Mäusen
Listerien injiziert und klinische und histologische Zeichen einer Rhombenzephalitis
beobachtet (Antal et al., 2001). Eine mögliche natürliche Eintrittspforte für diese Art
der Infektion könnten, gerade bei Wiederkäuern, Verletzungen der Mundschleimhaut
sein, welche dann wiederholt durch kontaminierte Silage Listeria monocytogenes
ausgesetzt sind.
1.5.4 Grundlegende Mechanismen der Immunabwehr von Listeria
monocytogenes
Bevor Listeria monocytogenes in den Fokus geriet, über kontaminierte Nahrungs-
mittel Infektionen auszulösen, war es schon ein Modellorganismus zur Untersuchung
der angeborenen und erworbenen Immunantwort. Im Mausmodell konnte beschrie-
ben werden, dass das Pathogen innerhalb von Makrophagen überleben und sich
vermehren kann. Die primäre Infektion mit dem Bakterium induziert eine zellulär
vermittelte Immunantwort (Mackaness, 1962). Antikörper spielen dabei keine Rolle.
Von dieser Entdeckung ausgehend folgten weitere Studien, die den Mechanismus
der zellulär vermittelten Immunantwort weiter untersuchten (Mackaness, 1964;
Mackaness, 1969). Eine subletale Infektion der Maus löst eine starke Immunantwort
des angeborenen und erworbenen Immunsystems aus.

Einleitung
19
Die angeborene Immunantwort spielt in der frühen Phase der Listerieninfektion eine
entscheidende Rolle (Corr und O`Neill, 2009; Stavru et al., 2011). Gerade neutro-
phile Granulozyten sind wichtig, um die bakterielle Ausbreitung und Vermehrung
durch Produktion antimikrobieller Wirkstoffe zu kontrollieren (Rogers und Unanue
1993; Brinkmann et al., 2004). Sie werden schnell durch ausgestoßenes IL-6 zum
Infektionsherd rekrutiert und führen selbst durch die Produktion vom Monozyten
koloniestimulierenden Faktor (CSF-1) und dem Monozyten chemotaktischen Protein
(CCL2) zur Einwanderung von Makrophagen (Guleria und Pollard, 2001; Mandel und
Cheers, 1980). Als Antwort darauf produzieren diese TNF-α und IL-12, welche
Natürliche Killerzellen zur Freisetzung von Interferon-gamma (IFN-γ) anregen
(Havell, 1987; Tripp et al., 1993), was wiederum weitere Makrophagen aktiviert.
Durch die neutrophil bedingte Produktion von reaktiven Sauerstoffspezies erfolgt die
durch Makrophagen bedingte Eliminierung von Listeria monocytogenes.
Inflammatorische Zytokine sind wichtig für die Kommunikation der beteiligten
Immunzellen. Besonders IFN-γ spielt eine Hauptrolle. So zeigten sich IFN-γ-
Rezeptor-defiziente (Ifngr1-KO) Mäuse als besonders empfindlich für eine
Listerieninfektion (Huang et al., 1993). Auch ist bekannt, dass Listeria
monocytogenes Typ-I-Interferone, welche bei der antiviralen Immunantwort beteiligt
sind, induzieren kann. Jedoch wirken IFN-Typ-I begünstigend auf die Listerien wäh-
rend einer Infektion ein. IFN-I-Rezeptor defiziente Mäuse zeigten sich resistent
gegenüber einer Listerieninfektion (Auerbuch et al., 2004; Carrero et al., 2004;
O`Connell et al., 2004).
Die listerielle Infektion von Makrophagen aktiviert zwei Wege der Genexpression
(McCaffrey et al., 2004). Zum einen die Gene der Typ-I-Interferone, wofür Listeria
monocytogenes frei im Zytoplasma vorliegen muss und zum anderen erfolgt über
Toll-like-Rezeptoren (TLR) die Aktivierung der NF-κB Kaskade mit nachfolgender
Genexpression von Zytokinen. TLRs sind auf der Oberfläche von Zellen und auf der
Membran endozytotischer Vesikel zu finden. Sie erkennen diverse Strukturen von
Listeria monocytogenes wie Flagellin (TLR-5), DNA (TLR-9), Lipoteichonsäure und
Peptidoglykan (TLR-2). TLRs rekrutieren Adaptermoleküle wie z.B. MyD88 (über
TLR-2), welches wiederum über weitere Signalwege den Transkriptionsfaktors NF-κB

Einleitung
20
aktiviert (Edelson und Unanue, 2002). Neben den Toll-like-Rezeptoren spielen auch
NOD-like-Rezeptoren (NLR) eine Rolle, wovon einige die Funktion als intrazelluläre
Erkennungsmoleküle für Listeria monocytogenes übernehmen (Warren et al., 2008).
Die am besten untersuchten NLRs sind NOD2 und NALP3. NOD2 führt zur
Aktivierung von NF-κB und des MAPK Signalweges. NALP3 aktiviert Kaspase-1 und
führt zur Sekretion von IL-1β und IL-18 (Bohn et al., 1998; Mariathasan et al., 2006).
Die Zellen des erworbenen Immunsystems bestehen aus B- und T-Zellen. Die adap-
tive Immunantwort bei Listeria monocytogenes Infektionen ist schnell und T-Zell
bedingt. CD4- sowie CD8-positive T-Zellen vermitteln eine schützende Immunität.
Beiden Zelltypen wird spezifisch über Wirtsmechanismen Listeria monocytogenes
Antigene präsentiert (Pamer, 2004; Zenewiez und Shen, 2007). Auf jeder kern-
haltigen Zelle sind MHC-Klasse-I Moleküle exprimiert. Während der Infektion mit
Listerien werden Listerien-spezifische Peptide auf MHC-Klasse-I Moleküle geladen
und den CD8+ T-Zellen präsentiert (Lara-Tejero und Pamer, 2004). Die sich daraus
ergebende Immunität erfolgt zum einen über die Sekretion von IFN-γ und führt
dadurch zur Aktivierung von Makrophagen und zum anderen zur Lyse infizierter
Zellen und der Freisetzung intrazellulärer Bakterien, welche dann von aktivierten
Makrophagen eliminiert werden. Antigenpräsentierende Zellen wie Dendritische
Zellen, B-Zellen und Makrophagen tragen dagegen MHC-Klasse-II-Moleküle auf ihrer
Oberfläche und präsentieren Bakterienantigene den CD4+ T-Zellen (Lara-Tejero und
Pamer, 2004). Diese differenzieren sich weiter zu TH-1 Zellen, welche mit der
Sekretion von TH-1 Zytokinen zur Eliminierung von Listeria monocytogenes
beitragen (Hsieh et al., 1993). Des Weiteren spielen regulatorische CD4+ T-Zellen
eine wichtige Rolle bei der Kontrolle der CD8+ T-Zell Proliferation während einer
erneuten Listerieninfektion (Kursar et al., 2002). Die humoral vermittelte Immunität
durch B-Zellen ist von geringer Bedeutung. Nach einer Infektion mit Listerien erfolgt
nur eine schwache Antikörperproduktion gegen Listeriolysin. Diese schützt nicht vor
einer Reinfektion mit Listeria monocytogenes, kann aber in Interaktion mit
Makrophagen Listeriolysin neutralisieren (Edelson und Unanue, 2001).
Dendritische Zellen sind häufig Vermittler zwischen der angeborenen und erwor-
benen Immunantwort (Medzhitov, 2001). Sie induzieren eine starke CD4 und CD8 T-

Einleitung
21
Zell Antwort, welche dann auch eine stabile Population von listerienspezifischen Ge-
dächtnis-T-Zellen hervorbringt.
1.6 Studien der in vivo Infektion
1.6.1 Tiermodelle zur Analyse listerialer Infektionsmechanismen
Verschiedene Spezies sind empfänglich für die Infektion mit Listeria monocytogenes.
Nach der Entdeckung des Pathogen 1924, durch die Auslösung einer Monozytose
und bakteriellen Sepsis bei Labortieren, Meerschweinchen und Kaninchen (Murray et
al., 1926), konnte das Bakterium aus verschiedenen Wirten isoliert werden. So iso-
lierte nur ein Jahr später Pirie (Pirie, 1927) Listeria monocytogenes aus der Leber
von Rennmäusen. Die Listeriose als Erkrankung bei landwirtschaftlichen Nutzieren,
wie Schafen und Rindern, wurde ebenfalls schon früh als „circling disease“ beschrie-
ben (Gill, 1937; Mathews, 1928). Weiter folgten die Entdeckungen der perinatalen
Infektion beim Menschen sowie die Auslösung von Aborten bei Schafen und Rindern
als Folge einer Listerieninfektion (Burn, 1936; Biester et al., 1939; Biester und
Schwarte, 1940; Biester und Schwarte, 1941; Graham et al., 1943; Poppensiek,
1944). Verschiedene Versuchstiere wurden als Wirt für Listeria monocytogenes ver-
wendet. Kaninchen und Rennmäuse zeigten sich als sehr empfänglich für
Listerieninfektionen. Hier konnten nach der Infektion Sepsis und in schwangeren
Tieren Aborte festgestellt werden (Kautter et al., 1959; Kautter et al., 1963). Die In-
fektion von Ratten und Meerschweinchen über Aerosole zeigten letale Krankheits-
verläufe (Kautter et al., 1959). Die subkutane Infektion von Kaninchen führte zu
lokalen Abszessen (Bloom, 1928). Hamster (Nordland, 1960) und Ratten erwiesen
sich jedoch als resistent gegenüber einer Listerieninfektion (Gray, 1958) ebenso wie
Katzen und Hunde. Als Infektionsmodelle haben sich daher neben der Maus noch
Meerschweinchen, Kaninchen und Rennmäuse als geeignet erwiesen. Meer-
schweinchen und Kaninchen gehören zu den natürlichen Wirten (Murray et al.,
1926). Beim Meerschweinchen konnte zuerst die Invasion von Listeria mono-
cytogenes in nicht-phagozytierende Zellen, wie Enterozyten, gezeigt werden (Racz et
al., 1972). Aufgrund der speziesspezifischen InlA-E-Cadherin Interaktion sind sie

Einleitung
22
empfänglich für orale Infektionen. Ebenso können nach einer Infektion von schwan-
geren Tieren mit Listeria monocytogenes perinatale Infektionen und Aborte ausgelöst
werden. Jedoch zeigten sich Meerschweinchen resistenter eine effiziente Infektion
des ZNS zu generieren aufgrund des Mangel einer effizienten Interaktion von InlB mit
seinem Rezeptor c-Met (Khelef et al., 2006). Meerschweinchen und Kaninchen sind
als Model für die Überquerung der intestinalen Barriere sowie Plazentaschranke ge-
eignet (Bakardjiev et al., 2004). Die Rennmaus ist von Natur aus empfänglich für
beide Interaktionswege, InlA und InlB. Nach oraler Listerieninfektion konnte gezeigt
werden, dass beide Wege für die Überquerung der Plazentaschranke ein Rolle spie-
len (Disson et al., 2008; Disson et al., 2009). Auch gibt es Untersuchungen des
Infektionsprozesses von Listeria monocytogenes im Zebrafisch (Danio rerio). Die
intravenöse Injektion in die Larve resultiert in fatalen Infektionen (Levraud et al.,
2009). Untersuchungen der Listerieninfektion bei Insekten, z.B. Drosophila
melanogaster, erlaubt die Unterscheidung von pathogenen und nicht-pathogenen
Listerienstämmen (Jensen et al., 2007). Infektionen von Galleria mellonella dienten
der Charakterisierung der angeborenen Immunabwehr (Mukherjee et al., 2010).
1.6.2 Mausmodelle zur Analyse listerialer Infektionsmechanismen
Aufgrund einer ausgesprochenen Vielfalt diverser Mausstämme und der einfachen
und kostengünstigen Haltung als Labortier bleibt häufig die Maus das Versuchstier
der ersten Wahl. Frühe Experimente mit intraperitonealen Infektionen von Listeria
monocytogenes zeigten septische Verläufe (Seeliger und Plab, 1959). Zur Analyse
von ZNS Infektionen wurde die intrazerebrale Injektion etabliert (Kautter et al., 1959),
welche zu Meningitis und Enzephalitis führte. Weiter wurden Mäuse über Aerosole
infiziert, welches in einer hohen Sterblichkeit resultierte (Dediè, 1955). Das erste
Mausmodell zur Untersuchung der Immunantwort nach einer Listerieninfektion wurde
von Mackaness 1962 etabliert. Es wurde gezeigt, dass nach intravenöser Infektion
von Mäusen Listeria monocytogenes innerhalb von Makrophagen persistieren und
überleben kann. Außerdem induziert eine primäre Listerieninfektion eine schützende
zellulär vermittelte Immunität (Mackaness, 1962). Dies konnte über den Transfer
lymphoider Milzzellen einer mit Listeria monocytogenes infizierten Maus in eine
uninfizierte Maus demonstriert werden, welche vor einer nachfolgenden

Einleitung
23
Listerieninfektion geschützt war (Mackaness, 1969). Weitere Studien folgten, die die
Resistenz und Empfänglichkeit von Mausstämmen gegenüber intravenöser Listeria
monocytogenes Infektionen untersuchten (McKenzie und Cheers, 1978; Skamene
und Kongshavn, 1979). Sie zeigten, dass C57BL Mäuse und verwandte Unter-
stämme resistent gegenüber einer Listerieninfektion reagieren und andere Mausin-
zuchtstämme wie A/J, BALB/c, CBA, C3H und 129 sich empfindlich verhalten. Über
die Generierung rekombinanter Inzuchtlinien aus A/J und C57BL/6J konnten zwei
Regionen im Genom lokalisiert werden, welche das bakterielle Wachstum kontrollie-
ren (Gervais et al, 1984). Eine Defizienz für das C5 Komplementfaktorgen (Hc) in A/J
Mäusen erhöht ihre Empfindlichkeit gegenüber Infektionen mit Listeria mono-
cytogenes (Gervais et al, 1984). Die Suszeptibilität der A/J und Resistenz der
C57BL/6J Mäuse konnte auch nach intragastrischer Infektion bestätigt werden
(Czuprynski und Faith, 2002). Auch das Geschlecht der jeweiligen Versuchstiere hat
Einfluss auf den Ausgang einer Listerieninfektion. So konnte gezeigt werden, dass
weibliche Mäuse empfindlicher reagieren als männliche (Pasche et al., 2005). Die
Infektion von Mäusen mit Listeria monocytogenes kann intravenös, intraperitoneal,
subkutan und oral erfolgen. Jedoch sind für die orale Infektion von Mäusen hohe
Dosen erforderlich, um eine systemische Erkrankung auszulösen. Dies beruht auf
einer speziesspezifischen Bindung von Internalin A an E-Cadherin. (siehe Abschnitt
1.5.1.) Zur Untersuchung der Bedeutung der InlA-E-Cadherin-Interaktion bei der
Überquerung der intestinalen Barriere wurde eine transgene Maus generiert. Diese
exprimiert humanes E-Cadherin unter Kontrolle des Promotors für das intestinale
Fettsäurebindungsprotein neben murinem E-Cadherin auf den Enterozyten des
Dünndarms (Lecuit et al., 2001, Lecuit und Cossart, 2002). Mit Hilfe dieser Maus
konnte die Rolle des InlA bei der Überquerung der Darmbarriere geklärt werden.
Dieses Modell hat seine Grenzen, es kann nicht die Rolle des E-Cadherins bei ande-
ren Zelltypen, außer Enterozyten, und somit die Überquerung anderer epithelialer
Barriere beschreiben. Nachfolgend wurde die knock-in Maus „E16P“ generiert. Alle
Zellen dieser Maus exprimieren durch den Austausch von Glutamin zu Prolin an der
Position 16 des Maus-E-Cadherins humanes E-Cadherin anstelle von murinem E-
Cadherin (Disson et al., 2008). Weiter wurde die Rolle der InlA-E-Cadherin-Inter-
aktion bei der Überquerung der Plazentaschranke untersucht. Die Infektion von

Einleitung
24
schwangeren knock-in Mäusen zeigte eine Abhängigkeit von InlA und InlB (Disson et
al., 2008).
Die entgegengesetzte Möglichkeit zur humanen Maus ist die Murinisierung von
Listeria monocytogenes (Wollert et al., 2007b). Wie in Abschnitt 1.5.1 ausführlich
beschrieben, erhöht der Austausch der Aminosäuren S192N-Y369S innerhalb des
Internalins die Bindungsaffinität zu humanen und murinem E-Cadherin. Dieses Mo-
dell erlaubt die orale Infektion mit Listeria monocytogenes in den verschiedensten
Mausinzuchtstämmen und Maus-knock-out Linien.
1.7 In vivo Imaging
1.7.1 Das Prinzip der Biolumineszenz
Biolumineszenz ist ein lichtemittierender, chemischer Prozess und in einer Vielzahl
wirbelloser Tiere verbreitet. Verschiedene Reportergene wie die Glühwürmchen Lu-
ziferase (DeLuca und McElroy, 1974), Renilla Luziferase (Matthews et al., 1977) und
die Photorhabdus luminescens Luziferase (Hastings und Nealson, 1977) wurden
charakterisiert. Die am häufigsten für Biolumineszenz-Imaging verwendete Luzi-
ferase entstammt Photinus pyralis, dem Glühwürmchen (firefly Luziferase). Als Sub-
strat dient Luziferin, welches in Abhängigkeit von ATP und Sauerstoff und unter
Emission von Licht, mit einem Emissionsmaximum von 560 nm, zu Oxyluziferin oxi-
diert wird. Bei einigen Infektionsmodellen ist die Luziferase im Genom von Tieren
integriert und benötigt die Zugabe des Substrats von außen. Das emittierte Licht ist
von starker Intensität, jedoch ist die Lichtreaktion zeitlich limitiert und stark
sauerstoffabhängig. Fünf Gene in einem Operon codieren für Proteine die
gemeinsam an einem Biosyntheseweg beteiligt sind, der über enzymatische Katalyse
zur Freisetzung von Licht führt. Die Gene luxCDE codieren für eine
Fettsäurereduktase, welche in die Synthese von Fettaldehyd involviert ist. Dies dient
als Substrat für die Lichtreaktion, welche durch eine Luziferase, kodiert durch luxAB,
katalysiert wird. Reduziertes Flavinmononukleotid und langkettiges Fettsäurealdehyd
wird unter Emission von blau-grünem Licht (490 nm) oxidiert. Lux-Operons von
Bakterien codieren sowohl für das Enzym als auch für das Substrat für die

Einleitung
25
Lichtreaktion, Flavinmononukleotid und Sauerstoff sind in jedem aeroben Bakterium
enthalten. Dieses Reportersystem hat den Vorteil, dass es die Stabilität der
Lichtproduktion erhöht (Doyle et al., 2004).
1.7.2 Die Technologie des in vivo Imaging
Für in vivo Imaging sind verschiedene Systeme wie z.B. Computertomografie, Rönt-
genstrahlung, Ultraschall (MRT) und Positronen-Emissions-Tomografie (PET)
erhältlich. Biolumineszenz-Imaging ist eine weitere Alternative und dient im
Besonderen der Beobachtung infektiöser Prozesse. Mit der Isolierung und Charakte-
risierung diverser Luziferasen (DeLuca und McElroy, 1974; Gates und DeLuca, 1975;
Hastings und Nealson, 1977; Matthews et al., 1977; McElroy und DeLuca, 1983)
erfolgte auch deren Verwendung in der wissenschaftlichen Forschung. Erstmals
wurde E. coli 1985 mit dem Lux-Operon transformiert (Engebrecht et al., 1985). Für
die Markierung von Zellen und Geweben oder die Analyse von Genexpressionen und
der Untersuchung der Wirts-Pathogen-Interaktion wurden die Reportergene des
bakteriellen Luziferase Operons in das Genom von Bakterien, Zelllinien oder Tieren
integriert. Im Jahr 1995 wurde die photonische Detektion von mikrobiologischen
Krankheitserregern beschrieben (Contag et al., 1995). In den letzten Jahren wurde
diese Technik zur Herstellung rekombinanter Mikroorganismen, im Besonderen
Krankheitserreger, zur Untersuchung von Infektionsprozessen verwendet. Eine Viel-
zahl biolumineszenter Pathogene wie z.B. Salmonella enterica (Burns-Guydish et al.,
2005), Staphylococcus aureus (Francis et al., 2000), Streptococcus pneumonia
(Francis et al., 2001), Herpes-simplex-virus Typ I (Luker et al., 2002) und Listeria
monocytogenes (Hardy et al., 2004; Riedel et al., 2007; Monk et al., 2010) wurden
entwickelt, um den Verlauf einer Infektion im Versuchstier zu untersuchen.
Das in vivo Biolumineszenz-Imaging (BLI) ist eine nicht invasive, hochsensitive
Methode, welche Echtzeitaufnahmen zum Infektionsverlauf des ganzen lebenden
Tieres erlaubt (Contag et al., 2002; Contag et al., 2008). Die Methode wird ver-
wendet, um Pathogene in lebenden Organismen zu lokalisieren und ihre Ausbreitung
durch die inneren Organe des Versuchstiers zu dokumentieren. Das emittierte Licht
der Pathogene wird durch das Gewebe transmittiert und generiert Lichtpunkte auf

Einleitung
26
der Oberfläche des Tieres. Diese werden über eine CCD Kamera (cooled coupeld
device) detektiert. Aufgrund einer Kühlung der Kamera ist es möglich störendes
Wärmerauschen zu reduzieren. Das System analysiert die Werte und stellt mit Hilfe
von Falschfarben die Intensität des Lichtes in einem Bild dar. Die Daten werden als
Photonen/s/cm2/sr dargestellt, wobei sich sr (Steradian/Raumwinkel) auf die Pho-
tonen von einem festen Winkel der Sphäre beziehen. Die Sensitivität vom BLI ist von
verschiedenen Faktoren abhängig, von der Stärke des generierten Signals, der An-
zahl der Zellen, welche die Reportergene exprimieren, von der Effizienz der Re-
portergene und vom Gewebetyp sowie von der Gewebetiefe der biolumineszenten
Lichtquelle. Mittlerweile sind Biolumineszenz-Imaging Systeme mit weiteren
Methoden, z.B. CT, gekoppelt (CaliperLS).
1.7.3 In vivo Imaging von Listeria monocytogenes Infektionen in der Maus
Eine allgemeine Methode ist es Luziferasegene mit Hilfe von Transposons in das
Genom von Listerien zu integrieren. Dies erfordert daraufhin potenzielle Klone auf
ihre Biolumineszenzstärke zu untersuchen (Hardy et al., 2004). Jedoch kann die
Insertion des Lux-Operons andere Gene wie z.B. Virulenzgene inaktivieren. Zur
Beobachtung von Genexpressionen, wurde ein synthetisches Lux-Operon generiert,
welches über einen chromosomalen Integrationsvektor pPL2lux ins Genom von
Listerien integriert wurde (Riedel et al., 2007). Die Verwendung von BLI im Falle von
Listeria monocytogenes führte zu neuen Entdeckungen in dessen Pathogenese und
Wirts-Pathogen-Interaktion. Nach der Infektion von Mäusen mit biolumineszenten
Listeria monocytogenes in geringer und hoher Infektionsdosis konnte beobachtet
werden, dass diese innerhalb der Gallenblase persistieren und extrazellulär
replizieren (Hardy et al., 2004). Diese Beobachtung beschreibt eine Möglichkeit von
Listeria monocytogenes sich asymptomatisch und vom Immunsystem unerkannt im
Wirt zu vermehren. Über die Sekretion von Gallensäure kann sich das Pathogen in
den Darm ausbreiten und somit als Infektionsquelle für neue Wirte dienen oder zur
Reinfektion des alten Wirtes führen (Hardy et al., 2006). Eine andere Arbeit zeigte,
dass nach intravenöser Infektion von Listeria monocytogenes in Mäuse diese im
Knochenmark persistieren und eine Quelle für Infektionen des zentralen Nerven-
systems darstellen können (Hardy et al., 2009).

Einleitung
27
Einige verschiedene biolumineszente und auch fluoreszierende Listerienstämme sind
beschrieben worden, welche sich für ein Imaging von Infektionsprozessen in vivo
eignen (Andersen et al., 2006; Balestrino et al., 2010; Bron et al., 2008; Hardy et al.,
2004; Monk et al., 2010; Riedel et al., 2007). Zur Untersuchung der Invasion von
Listeria monocytogenes wurde der Lmo ScottA Stamm (Serovar 4b) mit verschie-
denen Fluoreszenzfarbstoffen markiert ohne die Virulenz der modifizierten Stämme
zu beeinflussen. Dies ermöglicht Co-Infektionen diverser markierter Listeria mono-
cytogenes Stämme und erlaubt die Visualisierung und Identifikation einzelner
Listerien auf zellulärer Ebene mit Hilfe der Fluoreszenzmikroskopie (Andersen et al.,
2006). Zur Analyse des Infektionsprozesses unter dem Einfluss verschiedener Viru-
lenzfaktoren wurden diverse Mutanten von LmoEGDe mit verschiedenen Fluores-
zenzmarkern markiert, welche durch Integrationsvektoren in das Genom von Listeria
monocytogenes integriert wurden (Balestrino et al., 2010). Hardy et al. verwendeten
für Analysen den Listerienstamm LmoXen32 (als Serovar 2c klassifiziert), welcher
durch genetische Modifikation ausgehend von Listeria monocytogenes 10403S
(Serovar 1/2a) hergestellt worden war. Listeria monocytogenes 10403S wurde mit
dem Plasmid pAUL-A Tn4001 luxABCDE KmR transformiert. Hier erfolgte die chro-
mosomale Integration der lux-Kassette in den Promotor des Flagellin A Gens (flaA)
(Hardy et al., 2004).
Eine Alternative sind die biolumineszenten Abkömmlinge des LmoEGDe. Mit Hilfe
des Integrationsvektors pPL2, welcher ein PSA (Prophage LmoScott A) Listerio-
phagen Integrase Gen und eine DNA-Bindungsstelle enthält, kann die zielgerichtete
Integration einer einzelnen DNA-Kopie in das tRNAArg-Gen von Listeria mono-
cytogenes erfolgen (Lauer et al., 2002). Der Vektor pPL2lux, ein Abkömmling von
pPL2, enthält das synthetische Operon luxABCDE. Dies kodiert zum einen für einen
Fettsäurereduktasekomplex (LuxCDE) und zum anderen für eine Luziferase (LuxAB)
zur Katalyse der Biolumineszenzreaktion. Bron et al. fusionierten luxABCDE mit
secA- und hlyA-Promotoren und integrierte mit Hilfe des pPL2lux Vektors eine ein-
zelne Kopie ins tRNAArg-Gens des Chromosoms von Listeria monocytogenes zur
Generierung von LmoEGDe-lux (Bron et al., 2006). Nachfolgend testeten Riedel et
al. mit Hilfe des pPL2lux Vektors verschiedene Promotoren auf deren Bio-

Einleitung
28
lumineszenzexpressionrate in Listeria monocytogenes (Riedel et al., 2007). Der Lis-
terienstamm LmoEGDe-mur (Monk et al., 2010) wurde über die Einbringung von
zwei Internalin A Modifikationen (S192N, Y369S) murinisiert. Durch Integration des
Lux-Operons mit Hilfe des Integrationsvektors pIMK2lux in die 3`Region der tRNAArg
erfolgte die Generierung des LmoEGDe-lux-mur, welcher die orale Infektion im
Mausmodell für die Untersuchung der Wirt-Pathogen-Interaktion erlaubt (Monk et al.,
2010). Die genetische Modifikation erfolgte mit Hilfe des pIMK Vektors, ein Derivat
des pPL2 Vektors. Dieser erlaubt zielgerichtet die chromosomale Mutagenese über
Integration und Rekombination des gewünschten DNA-Abschnittes (Lauer et al.,
2002; Monk et al., 2008).
1.8 Konzept und Fragestellung der vorliegenden Dissertation
Zahlreiche vorherige Arbeiten konnten die wichtige Funktion des Internalin A für die
Invasion von Listeria monocytogenes in intestinale Epithelzellen und das Durch-
brechen der Darmbarriere nach der oralen Infektion zeigen. Zu Beginn dieser Arbeit
zeigten Wollert et al. (Wollert et al., 2007b), dass durch gezielte genetische Modi-
fikation des inlA-Locus von Listeria monocytogenes ein sogenannter „murinisierter“
Listerienstamm generiert werden kann, der ein modifiziertes Internalin A Protein
exprimiert, welches mit hoher Affinität an den murinen E-Cadherin Rezeptor binden
kann. Dieser modifizierte Listerienstamm (Lmo-InlAm) exprimiert ein Internalin A
(InlAS192N-Y369S) indem im Vergleich zum Wildtyp nur zwei Aminosäuren ausgetauscht
worden sind. Dieser Listerienstamm zeigte in vivo im Vergleich zum Wildtyp eine
wesentliche höhere Invasionseffizienz bei der Überquerung der intestinalen
Wirtsbarriere (Wollert et al., 2007b). Dadurch wurde ein neues orales
Listerieninfektionsmodell in der Maus etabliert, mit welchem Pathomechanismen der
Listeriose nach der Infektion von Mäusen über den natürlichen Infektionsweg studiert
werden können. Die Arbeit von Wollert et al. hat offen gelassen ob das modifizierte
Internalin A auch für die Überquerung anderer epithelialer Barrieren im Wirt wichtig
ist.

Einleitung
29
Ziel meiner Arbeit war es, basierend auf der Arbeit von Wollert et al. ein Mausmodell
zu generieren in dem mit Hilfe von Biolumineszenz-Imaging die Ausbreitung Inter-
nalin A modifizierter Listerien nach oraler Infektion genauer analysiert werden kann.
Folgende Fragestellungen lagen der Arbeit zugrunde:
1) Zeigen murinisierte Lmo-InlAm Bakterien eine veränderte Dissemination im Orga-
nismus nach oraler Inokulation im Vergleich zum nicht modifizierten, isogenen
Wildtyp?
2) Eignet sich das Verfahren des in vivo Biolumineszenz-Imaging (BLI) um die Aus-
breitung Lux-Operon exprimierender Listerien im Wirtsorganismus der Maus nicht
invasiv zu analysieren? Wie sensitiv ist dieses Verfahren und welche Bakterien-
konzentrationen müssen in verschiedenen Organen erreicht werden um eine
Infektion über BLI zu delektieren?
3) Spielt das veränderte (murinisierte) Internalin A eine Rolle bei der Überquerung
der Blut-Hirn-Schranke? Vorangegangene Arbeiten haben spekuliert, dass die
Interaktion von Internalin A mit E-Cadherin auch für das Eindringen von Listerien
in das zentrale Nervensystem wichtig sein könnten. Der murinisierte Listerien-
stamm erlaubt es diese Fragestellung in verschiedenen Mausinzuchtstämmen
nachzugehen.
4) Spielt der genetische Hintergrund des Wirts eine Rolle für die Empfindlichkeit
gegenüber der oralen Listerieninfektion und Ausprägung der Listeriose? Dieser
Fragestellung kann durch gezielte Analyse verschiedener Mausinzuchtstämme
im oralen Listerieninfektionsmodell nachgegangen werden. Murinisierte Listerien-
stämme erlauben es Mausinzuchtstämme unabhängig von der Präsenz eines
transgenen, humanisierten E-Cadherin Alleles zu untersuchen. Verschiedene
Mausinzuchtstämme könnten auch unterschiedliche Resistenzen für die Trans-
mission der Listerien in das zentrale Nervensystem zeigen. Falls es hier Unter-
schiede zwischen Mausstämmen gibt, wäre ein resistenter und empfindlicher
Mausstamm für ein weiteres Studium von Mechanismen zerebraler Listeriose
und der damit verbundenen Meningoenzephalitis sehr wichtig.

Einleitung
30
Im Detail sollten die Mausinzuchtstämme BALB/cJ, C57BL/6J, C3HeB/FeJ und
A/JOlaHsd auf die Wirt-Pathogen-Interaktion untersucht werden, um Einblicke in die
genetisch kontrollierte Resistenz zu erhalten. Dabei soll zum einen die Virulenz der
Listerienstämme berücksichtigt werden und zum anderen der genetische Hintergrund
der verschiedenen Mausinzuchtlinien. Für die BLI-Analysen mit dem Xenogen IVIS
200 System wurden die Listerienstämme LmoXen32, LmoXen32-mur, LmoEGDe-lux
und LmoEGDe-lux-mur verwendet, wobei die Stämme LmoXen32-mur und
LmoEGDe-lux-mur die Internalin A S192N-Y369S optimierten Mutanten darstellen.
Der Stamm LmoXen32-mur wurde von mir selbst genetisch verändert werden, um
eine murinisierte Variante zu generieren. Die schon biolumineszenten Listerien-
stämme LmoEGDe-lux und LmoEGDe-lux-mur wurden von Dr. Gahan Cormac
(University College Cork, Cork, Irland) freundlicherweise für die Analysen zur Verfü-
gung gestellt.

Ergebnisse
31
2 Ergebnisse
2.1 Murinisierung der Listerienstämme Listeria monocytogenes 10403S
Xen32 und LO28
Im Rahmen dieser Dissertation wurde ein Mausmodell für das in vivo Bio-
lumineszenz-Imaging (BLI) oraler Infektionen mit Listeria monocytogenes etabliert.
Grundlage war die Generierung eines murinisierten und biolumineszenten Listerien-
stamms. Die Infektion mit Listeria monocytogenes beruht auf einer ausgeprägten
Speziesspezifität (Lecuit et al., 1999). Somit entsprechen Mäuse nicht dem natür-
lichen Wirt von Listerien und sind für die orale Infektion nicht geeignet. Die voran-
gegangene Arbeit von Wollert et al. zeigte, dass der Aminosäureaustausch an den
Positionen S192N-Y369S des Oberflächenproteins Internalin A von Listeria mono-
cytogenes die Bindungsaffinität zu seinem Rezeptor E-Cadherin erhöht (Wollert et
al., 2007b). Weiter resultierten diese Modifikationen in der Aminosäuresequenz
ebenfalls in einer Verstärkung der Bindungsaffinität zu Maus-E-Cadherin. Dies
erhöhte die Empfänglichkeit von Mäusen für die orale Infektion mit diesem genetisch
modifizierten Listerienstamm. Ausgehend vom kommerziell zu erwerbenden bio-
lumineszenten Listerienstamm LmoXen32 (CaliperLS; Hardy et al., 2004) erfolgte die
genetische Modifikation (Murinisierung) des Internalin A über den Austausch von
Wildtyp Internalin A mit murinisierten Internalin A (Codon 192: TCT → AAC; Codon
369: TAT → TCA).
Im Rahmen einer Kooperation mit der Abteilung für Molekulare Immunologie (MOLI,
Helmholtz-Zentrum für Infektionsforschung) erfolgte die Murinisierung des Listerien-
stamms LO28, Serotyp 1/2c (Vicente et al., 1985). Listeria monocytogenes LO28
exprimiert aufgrund einer „nonsense“ Mutation im inlA Gen ein verkürztes Internalin
A von 63 kilo Dalton (kDa) (Jonquieres et al., 1998). Als Folge dieser Mutation wird
Internalin A nicht auf der Zellwandoberfläche von LO28 exprimiert, sondern das
Protein unter Zellkulturbedingungen in den Zellüberstand sekretiert (Jonquieres et
al., 1998). Eine weitere Konsequenz des verkürzten Internalin A ist die geringere
Invasionsrate von LO28 in epitheliale Zelllinien im Vergleich zu anderen Listerien-

Ergebnisse
32
stämmen, welche funktionsfähiges Internalin A exprimieren (Roldgaard et al., 2009).
Jedoch konnte gezeigt werden, dass während der Infektion von Makrophagen mit
LO28 eine erhöhte IFN-β-Synthese induziert wurde im Vergleich zur Infektion mit
Listeria monocytogenes EGDe (Reutterer et al., 2008). Dieser Befund konnte in vivo
im Mausmodell bestätigt werden (Solodova et al., 2011). Um der Fragestellung
nachzugehen ob eine „Murinisierung“ des Internalin A von LO28 mit einer zusätzlich
erhöhten IFN-β-Produktion im Wirt verbunden ist, wurde auch ein murinisierter LO28
Stamm hergestellt (LO28-mur).
Die generierten Listerienstämme LmoXen32-mur und LO28-mur wurden auf die
genetische Veränderung des inlA-Locus, die Expression der Oberflächenproteine
Internalin A und Internalin B und auf die Invasion in humanen und murinen Zellen in
vitro analysiert. Dies diente als Voraussetzung für dann später erfolgte Analysen im
Mausmodell.
2.1.1 Ergebnisse der Transformation
Der Listerienstamm LO28 sowie der biolumineszente Stamm Listeria monocytogenes
Xen32 (CaliperLS) wurden mit dem Plasmid pAUL-A-inlAm-inlB (S192N: TCT →
AAC; Y369S: TAT → TCA) (freundlicherweise von Dr. Wolf-Dieter Schubert, Ab-
teilung der Strukturbiologie, Helmholtz-Zentrum für Infektionsforschung, zur Ver-
fügung gestellt) transformiert. Aufgrund eines aus Bacillus subtilis stammenden
temperatursensitiven Replikationsoperons (ts rep) (Gryczan et al., 1982; Youngman
et al., 1983) kann keine extrachromosomale Replikation des Plasmides bei 42°C
erfolgen (Chakraborty et al., 1992). Nach der Transformation des Plasmids in
Listerien integrieren homologe Sequenzen des Vektors über Rekombination in das
Genom von LmoXen32 und LO28 und es erfolgte der Austausch der DNA-Abschnitte
von Wildtyp InlA mit dem murinisierten InlA. Eine erfolgreiche Transformation konnte
mittels Erythromycin selektioniert werden. Mit Verlust der vektorcodierten
Antibiotikaresistenz wurde die homologe Rekombination angezeigt. Von den nach
der Transformation und induzierten Integration erhaltenen Klonen zeigten ca. 1/10
einen Verlust der Antibiotikaresistenz (siehe Material/Methoden 5.1.2). Um zu
analysieren ob in diesen Fällen eine homologe Rekombination erfolgte, wurden

Ergebnisse
33
jeweils acht Klone ausgewählt und auf Sequenzunterschiede der genomischen DNA
in den modifizierten Listerienstämmen untersucht.
2.1.2 Validierung der homologen Rekombination des murinisierten Internalin A
Jeweils acht Klone wurden auf die korrekte Integration von inlAm-inlB ins Genom von
LmoXen32-mur und LO28-mur über die Bestimmung der inlA-Sequenz untersucht.
Über Amplifizierung mittels PCR und anschließender Sequenzierung des kompletten
inlA-Locus konnte die homologe Rekombination und damit die gewünschte Verände-
rung des inlA Leserasters bestätigt werden. Ein Repräsentant der erfolgreich modi-
fizierten Listeria monocytogenes Stämme LmoXen32-mur und LO28-mur ist in Ab-
bildung 2-1 dargestellt.
Die Untersuchung der inlA-Sequenz, des in der vorangegangenen Arbeit von Wollert
et al. generierten murinisierten Listerienstamms LmoEGDe S192N-Y369S
(LmoEGDe-mur) (Wollert et al., 2007b) sowie dessen Wildtyp LmoEGDe, diente zum
Vergleich der DNA- und Proteinsequenz. An Condonposition 192 des murinisierten
Internalin A erfolgte die Substitution des Tripletts TCT zu AAC. Dies führte in der
Proteinsequenz zum Austausch der Aminosäure Serin zu Asparagin an der Position
192 (Abbildung 2-1 A). An Codonposition 369 des inlA erfolgte die Substitution des
Tripletts TAT zu TCA, welches innerhalb der Proteinsequenz an Position 369 zum
Austausch von Tyrosin zu Serin führte (Abbildung 2-1 B). Parallel erfolgte die
Analyse der Sequenz des kompletten inlA-Locus der Wildtypstämme LmoXen32 und
LO28 zur Überprüfung der korrekten Integration über homologe Rekombination von
inlAm-inlB ins Genom von LmoXen32-mur und LO28-mur.
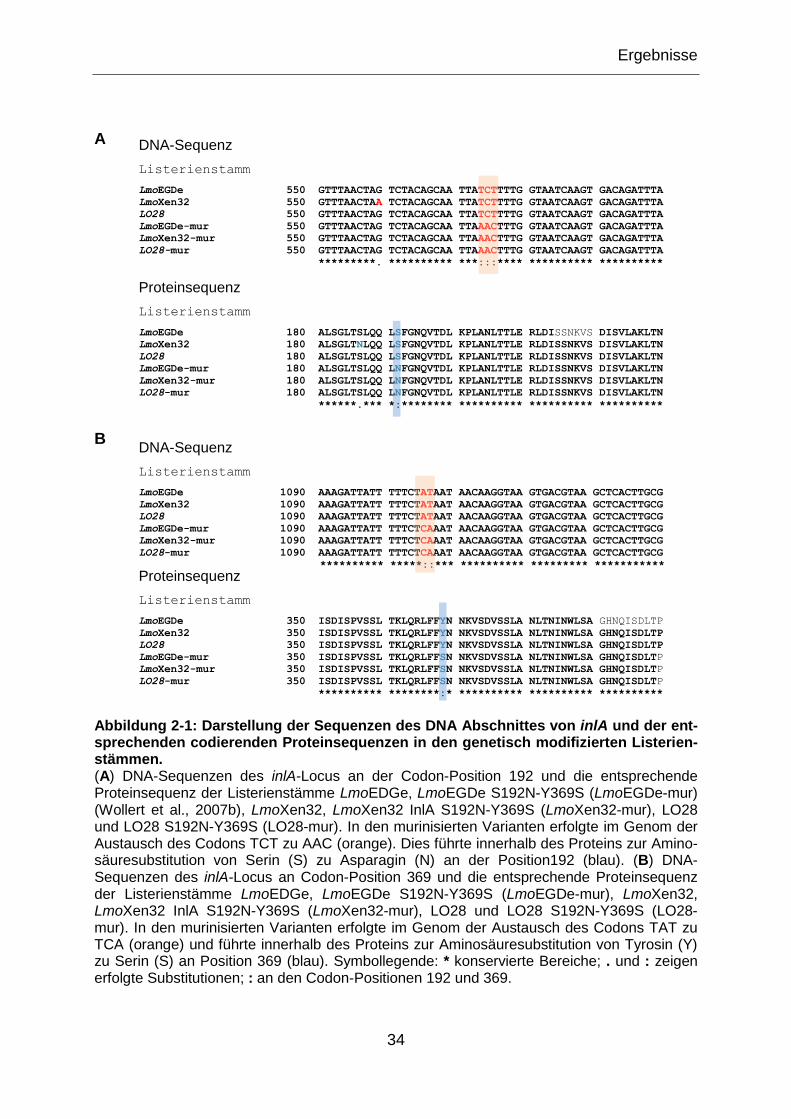
Ergebnisse
34
A
B
Abbildung 2-1: Darstellung der Sequenzen des DNA Abschnittes von inlA und der ent-sprechenden codierenden Proteinsequenzen in den genetisch modifizierten Listerien-stämmen. (A) DNA-Sequenzen des inlA-Locus an der Codon-Position 192 und die entsprechende Proteinsequenz der Listerienstämme LmoEDGe, LmoEGDe S192N-Y369S (LmoEGDe-mur) (Wollert et al., 2007b), LmoXen32, LmoXen32 InlA S192N-Y369S (LmoXen32-mur), LO28 und LO28 S192N-Y369S (LO28-mur). In den murinisierten Varianten erfolgte im Genom der Austausch des Codons TCT zu AAC (orange). Dies führte innerhalb des Proteins zur Amino-säuresubstitution von Serin (S) zu Asparagin (N) an der Position192 (blau). (B) DNA-Sequenzen des inlA-Locus an Codon-Position 369 und die entsprechende Proteinsequenz der Listerienstämme LmoEDGe, LmoEGDe S192N-Y369S (LmoEGDe-mur), LmoXen32, LmoXen32 InlA S192N-Y369S (LmoXen32-mur), LO28 und LO28 S192N-Y369S (LO28-mur). In den murinisierten Varianten erfolgte im Genom der Austausch des Codons TAT zu TCA (orange) und führte innerhalb des Proteins zur Aminosäuresubstitution von Tyrosin (Y) zu Serin (S) an Position 369 (blau). Symbollegende: * konservierte Bereiche; . und : zeigen erfolgte Substitutionen; : an den Codon-Positionen 192 und 369.
Proteinsequenz
Listerienstamm
LmoEGDe 180 ALSGLTSLQQ LSFGNQVTDL KPLANLTTLE RLDISSNKVS DISVLAKLTN LmoXen32 180 ALSGLTNLQQ LSFGNQVTDL KPLANLTTLE RLDISSNKVS DISVLAKLTN LO28 180 ALSGLTSLQQ LSFGNQVTDL KPLANLTTLE RLDISSNKVS DISVLAKLTN LmoEGDe-mur 180 ALSGLTSLQQ LNFGNQVTDL KPLANLTTLE RLDISSNKVS DISVLAKLTN LmoXen32-mur 180 ALSGLTSLQQ LNFGNQVTDL KPLANLTTLE RLDISSNKVS DISVLAKLTN LO28-mur 180 ALSGLTSLQQ LNFGNQVTDL KPLANLTTLE RLDISSNKVS DISVLAKLTN ******.*** *:******** ********** ********** **********
Proteinsequenz
Listerienstamm
LmoEGDe 350 ISDISPVSSL TKLQRLFFYN NKVSDVSSLA NLTNINWLSA GHNQISDLTP LmoXen32 350 ISDISPVSSL TKLQRLFFYN NKVSDVSSLA NLTNINWLSA GHNQISDLTP LO28 350 ISDISPVSSL TKLQRLFFYN NKVSDVSSLA NLTNINWLSA GHNQISDLTP LmoEGDe-mur 350 ISDISPVSSL TKLQRLFFSN NKVSDVSSLA NLTNINWLSA GHNQISDLTP LmoXen32-mur 350 ISDISPVSSL TKLQRLFFSN NKVSDVSSLA NLTNINWLSA GHNQISDLTP LO28-mur 350 ISDISPVSSL TKLQRLFFSN NKVSDVSSLA NLTNINWLSA GHNQISDLTP ********** ********:* ********** ********** **********
DNA-Sequenz
Listerienstamm
LmoEGDe 550 GTTTAACTAG TCTACAGCAA TTATCTTTTG GTAATCAAGT GACAGATTTA LmoXen32 550 GTTTAACTAA TCTACAGCAA TTATCTTTTG GTAATCAAGT GACAGATTTA LO28 550 GTTTAACTAG TCTACAGCAA TTATCTTTTG GTAATCAAGT GACAGATTTA LmoEGDe-mur 550 GTTTAACTAG TCTACAGCAA TTAAACTTTG GTAATCAAGT GACAGATTTA LmoXen32-mur 550 GTTTAACTAG TCTACAGCAA TTAAACTTTG GTAATCAAGT GACAGATTTA LO28-mur 550 GTTTAACTAG TCTACAGCAA TTAAACTTTG GTAATCAAGT GACAGATTTA *********. ********** ***:::**** ********** **********
DNA-Sequenz
Listerienstamm
LmoEGDe 1090 AAAGATTATT TTTCTATAAT AACAAGGTAA GTGACGTAA GCTCACTTGCG LmoXen32 1090 AAAGATTATT TTTCTATAAT AACAAGGTAA GTGACGTAA GCTCACTTGCG LO28 1090 AAAGATTATT TTTCTATAAT AACAAGGTAA GTGACGTAA GCTCACTTGCG LmoEGDe-mur 1090 AAAGATTATT TTTCTCAAAT AACAAGGTAA GTGACGTAA GCTCACTTGCG LmoXen32-mur 1090 AAAGATTATT TTTCTCAAAT AACAAGGTAA GTGACGTAA GCTCACTTGCG LO28-mur 1090 AAAGATTATT TTTCTCAAAT AACAAGGTAA GTGACGTAA GCTCACTTGCG ********** *****::*** ********** ********* ***********

Ergebnisse
35
2.1.3 Analysen der Internalin A und Internalin B Expression in genetisch modi-
fizierten Listerienstämmen
Die Oberflächenproteine InlA und InlB sind gemeinsam innerhalb eines Genclusters
im Genom von Listeria monocytogenes codiert und unterliegen beide der Kontrolle
des Regulatorproteins PrfA (de las Heras et al., 2011). Aufgrund der Hauptfunktion
von Internalin A und Internalin B bei der Invasion nicht-phagozytierender Zellen
(Seveau et al., 2007) ist die korrekte Expression beider Oberflächenproteine von
großer Bedeutung. Durch genetische Veränderung von inlA könnte die Expression
beider Invasionsproteine betroffen sein, da die Expression beider Gene von einem
gemeinsamen Operon gesteuert ist. Nachdem die korrekte Intergration des Plas-
mides pAUL-A-inlAm-inlB ins Genom von LmoXen32 und LO28 und damit die Gene-
rierung des genetisch modifizierten Listerienstamms LmoXen32-mur sowie LO28-
mur bestätigt werden konnte, erfolgte die Analyse der Expression der Oberflächen-
proteine InlA und InlB in den genetisch veränderten Listerienstämmen über die
Durchführung eines Western-Blots. Internalin A ist im Falle von LmoXen32 über eine
kovalente Bindung in der Zellwand verankert, während Internalin B nicht-kovalent
über Lipoteichonsäure an die bakteriellen Zellwand gebunden ist. Aus diesem Grund
erfolgte die Analyse von InlA in Trichloressigsäure präzipitierten Zellwandextrakt und
der Nachweis von InlB im Zellkulturüberstand. Der Nachweis beider Proteine erfolgte
über Western-Blot-Analyse mit monoklonalen Antikörpern gegen InlA und InlB (Ab-
bildung 2-2). Das Aktin-bindende Protein A (ActA) wird von einem anderen Operon
im Listeriengenom codiert. Seine Expression erfolgt unabhängig von inlA und inlB
(Vazquez-Boland et al., 2001) wird aber auch in vivo über das Regulatorprotein PrfA
kontrolliert (Mengaud et al., 1991). Daher erfolgte der Nachweis der Protein-
expression des Aktin-bindenden Protein A als Ladekontrolle.
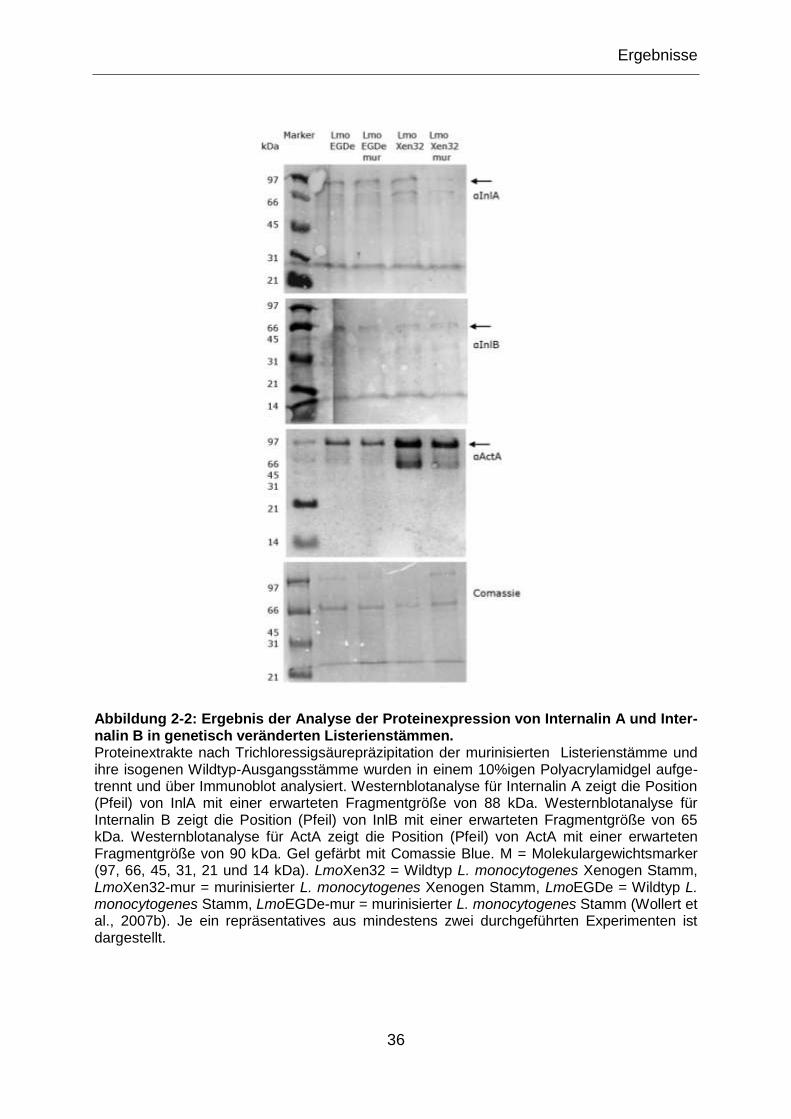
Ergebnisse
36
Abbildung 2-2: Ergebnis der Analyse der Proteinexpression von Internalin A und Inter-nalin B in genetisch veränderten Listerienstämmen. Proteinextrakte nach Trichloressigsäurepräzipitation der murinisierten Listerienstämme und ihre isogenen Wildtyp-Ausgangsstämme wurden in einem 10%igen Polyacrylamidgel aufge-trennt und über Immunoblot analysiert. Westernblotanalyse für Internalin A zeigt die Position (Pfeil) von InlA mit einer erwarteten Fragmentgröße von 88 kDa. Westernblotanalyse für Internalin B zeigt die Position (Pfeil) von InlB mit einer erwarteten Fragmentgröße von 65 kDa. Westernblotanalyse für ActA zeigt die Position (Pfeil) von ActA mit einer erwarteten Fragmentgröße von 90 kDa. Gel gefärbt mit Comassie Blue. M = Molekulargewichtsmarker (97, 66, 45, 31, 21 und 14 kDa). LmoXen32 = Wildtyp L. monocytogenes Xenogen Stamm, LmoXen32-mur = murinisierter L. monocytogenes Xenogen Stamm, LmoEGDe = Wildtyp L. monocytogenes Stamm, LmoEGDe-mur = murinisierter L. monocytogenes Stamm (Wollert et al., 2007b). Je ein repräsentatives aus mindestens zwei durchgeführten Experimenten ist dargestellt.

Ergebnisse
37
Zum einen wurde der murinisierte Listerienstamm LmoXen32-mur und der ent-
sprechende isogene Wildtyp LmoXen32 auf die Expression von InlA, InlB und ActA
untersucht und zum anderen erfolgte zusätzlich die Analyse der Proteinexpression
des in der vorangegangenen Arbeit von Wollert et al. generierten murinisierten
Listerienstamms LmoEGDe S192N-Y369S (LmoEGDe-mur) (Wollert et al., 2007b)
sowie dessen Wildtyp LmoEGDe im Vergleich. Die erwarteten Molekulargewichte
lagen bei InlA bei 88 kDa, bei InlB 65 kDa und bei ActA 90 kDa (Lingenau et al.,
1995). Die Analyse der Proteinexpression zeigte, dass sämtliche getesteten
Listerienstämme InlA in der Zellwand exprimieren. Jedoch ist InlA in den ver-
schiedenen Listerienstämmen unterschiedlich stark exprimiert. LmoXen32 und
LmoXen32-mur zeigten eine schwächere Expression als LmoEGDe und LmoEGDe-
mur. Ebenso konnte die Expression von InlB in den Zellüberständen aller Listerien-
stämmen nachgewiesen werden, zeigte aber ebenfalls eine schwächere Expression
in LmoXen32 und LmoXen32-mur im Vergleich zu den EGDe-Listerienstämmen. Als
Kontrolle für die Proteindegradation und die relativ geladene Proteinkonzentration
wurde ein Western-Blot für das Aktin-bindende Protein A durchgeführt. Die Ex-
pression des Proteins ActA erfolgt unabhängig von der Expression der Invasions-
proteine InlA und InlB. Jedoch unterliegen alle der Regulation durch PrfA (de las
Heras et al., 2011). Es wurde jedoch beobachtet, dass ActA in LmoXen32 und
LmoXen32-mur stärker exprimiert wurde. Die korrekte Expression beider Internaline
diente als Voraussetzung für den späteren in vivo Vergleich von murinisierten
Listerienstämmen mit ihrem isogenen Wildtyp im Mausmodell.
Die Expression der Proteine Internalin B und ActA konnte im generierten Listerien-
stamm LO28-mur sowie dessen Wildtyp LO28 ebenfalls nachgewiesen werden.
Während der Murinisierung des LO28 Stammes erfolgt die homologe Rekombination
des inlA-Locus und es kommt somit zum Austausch des verkürzten InlA mit einem
murinisierten funktionsfähigen InlA. Die Expression des Oberflächenproteins Inter-
nalin A konnte weder in LO28 noch LO28-mur nachgewiesen werden (Daten nicht
gezeigt).

Ergebnisse
38
2.1.4 Analyse der murinisierten Listerienstämme im Invasions-Assay
Wie bereits beschrieben, erhöht die Murinisierung des InlA die Bindungsaffinität zu
humanen und murinen E-Cadherin (Wollert et al., 2007b). Durch die Interaktion
beider Moleküle wird die Invasion in nicht-phagozytierende Zellen induziert. Zur
Analyse ob eine verstärkte Aufnahme der murinisierten Listerienstämme aufgrund
einer erhöhten Bindungsaffinität zum Rezeptor erfolgt, wurde ein Gentamycin-
Invasions-Assay (Elsinghorst, 1994) zum einen in Caco-2 Zellen (Abbildung 2-3) und
zum anderen in CT-26 Zellen (Abbildung 2-4) durchgeführt (siehe Material/Methoden
5.3). Die humane Colonkarzinom-Zelllinie Caco-2 zeigt unter Zellkulturbedingungen
ähnliche Eigenschaften wie Enterozyten (Rousset, 1986). Es erfolgt die für intestinale
Epithelzellen typische Polarisierung in eine apikale und basolaterale Domäne,
welche über die Expression von E-Cadherin auf der Zelloberfläche gekennzeichnet
ist. Somit sind Caco-2 Zellen für die Analyse der Internalisation von Listeria mono-
cytogenes geeignet (Mengaud et al., 1996b). Für die Untersuchung der Invasion in
murinen Zellen wurde die Colonkarzinom-Zelllinie CT-26 verwendet. CT-26 Zellen
wurden erstmals aus BALB/c Mäusen isoliert, welche zur Tumorinduktion
chemotherapeutisch behandelt wurden (Corbett et al., 1975). Sie haben das
Erscheinungsbild undifferenzierter Fibroblasten und zeigen unter Kulturbedingungen
keine Kontakt-Wachstumshemmung (Brattain et al., 1980). In Studien konnte jedoch
gezeigt werden, dass sie als Invasionsmodell für die Aufnahme von Listerien in nicht-
phagozytierende murine Zellen geeignet sind (Monk et al., 2010).
Untersucht wurde die Invasionsrate der murinisierten Listerienstämme LmoXen32-
mur und LO28-mur sowie von deren unmodifizierten Wildtyp-Stämmen LmoXen32
und LO28 in Caco-2 und CT-26 Zellen. Parallel erfolgte die Analyse der von Wollert
et al. generierten murinisierten Listerien LmoEGDe S192N-Y369S (LmoEGDe-mur)
(Wollert et al., 2007b) sowie dessen Wildtyps LmoEGDe im Invasions-Assay zum
Vergleich. Eine Stunde nach der Infektion mit 2x106 koloniebildenden Einheiten
(CFU) Listerien (MOI 10:1) erfolgte die Behandlung mit Gentamycin zur Abtötung
aller extrazellulär vorliegender Bakterien. Somit wurden nur die intrazellulär vorhan-
denen und replizierenden Listerien erfasst. Die Invasion der murinisierten Varianten
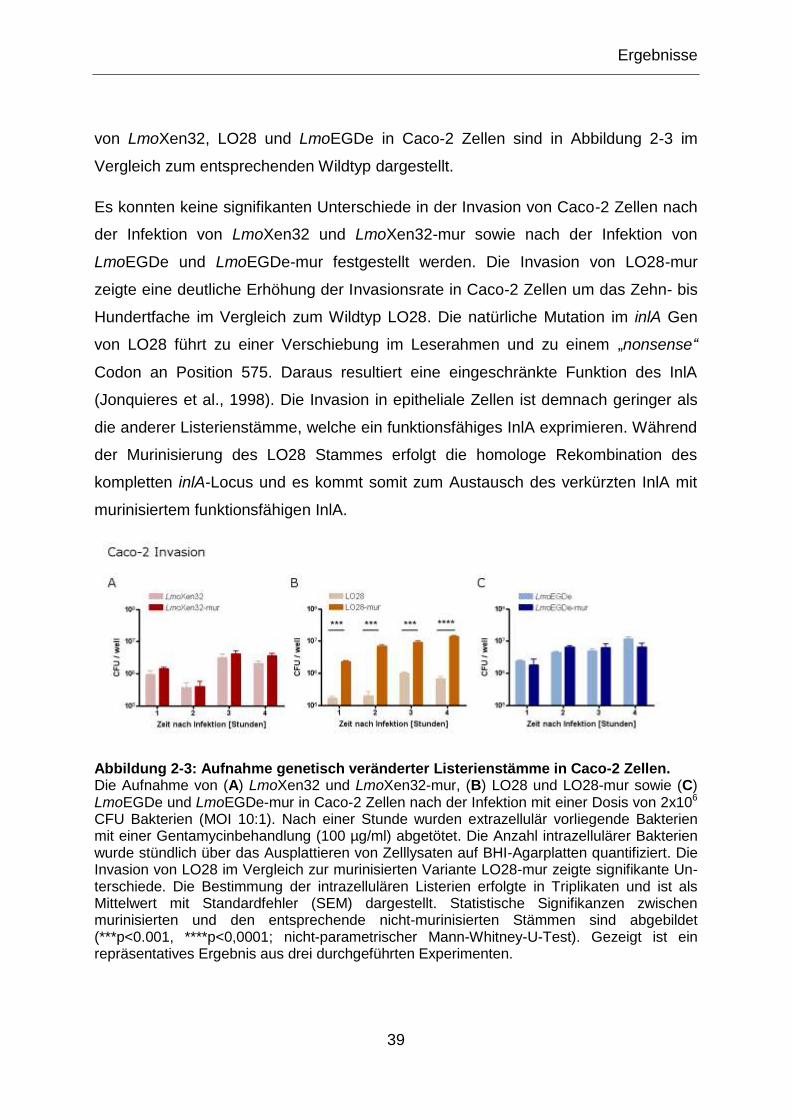
Ergebnisse
39
von LmoXen32, LO28 und LmoEGDe in Caco-2 Zellen sind in Abbildung 2-3 im
Vergleich zum entsprechenden Wildtyp dargestellt.
Es konnten keine signifikanten Unterschiede in der Invasion von Caco-2 Zellen nach
der Infektion von LmoXen32 und LmoXen32-mur sowie nach der Infektion von
LmoEGDe und LmoEGDe-mur festgestellt werden. Die Invasion von LO28-mur
zeigte eine deutliche Erhöhung der Invasionsrate in Caco-2 Zellen um das Zehn- bis
Hundertfache im Vergleich zum Wildtyp LO28. Die natürliche Mutation im inlA Gen
von LO28 führt zu einer Verschiebung im Leserahmen und zu einem „nonsense“
Codon an Position 575. Daraus resultiert eine eingeschränkte Funktion des InlA
(Jonquieres et al., 1998). Die Invasion in epitheliale Zellen ist demnach geringer als
die anderer Listerienstämme, welche ein funktionsfähiges InlA exprimieren. Während
der Murinisierung des LO28 Stammes erfolgt die homologe Rekombination des
kompletten inlA-Locus und es kommt somit zum Austausch des verkürzten InlA mit
murinisiertem funktionsfähigen InlA.
Abbildung 2-3: Aufnahme genetisch veränderter Listerienstämme in Caco-2 Zellen. Die Aufnahme von (A) LmoXen32 und LmoXen32-mur, (B) LO28 und LO28-mur sowie (C) LmoEGDe und LmoEGDe-mur in Caco-2 Zellen nach der Infektion mit einer Dosis von 2x106 CFU Bakterien (MOI 10:1). Nach einer Stunde wurden extrazellulär vorliegende Bakterien mit einer Gentamycinbehandlung (100 µg/ml) abgetötet. Die Anzahl intrazellulärer Bakterien wurde stündlich über das Ausplattieren von Zelllysaten auf BHI-Agarplatten quantifiziert. Die Invasion von LO28 im Vergleich zur murinisierten Variante LO28-mur zeigte signifikante Un-terschiede. Die Bestimmung der intrazellulären Listerien erfolgte in Triplikaten und ist als Mittelwert mit Standardfehler (SEM) dargestellt. Statistische Signifikanzen zwischen murinisierten und den entsprechende nicht-murinisierten Stämmen sind abgebildet (***p<0.001, ****p<0,0001; nicht-parametrischer Mann-Whitney-U-Test). Gezeigt ist ein repräsentatives Ergebnis aus drei durchgeführten Experimenten.
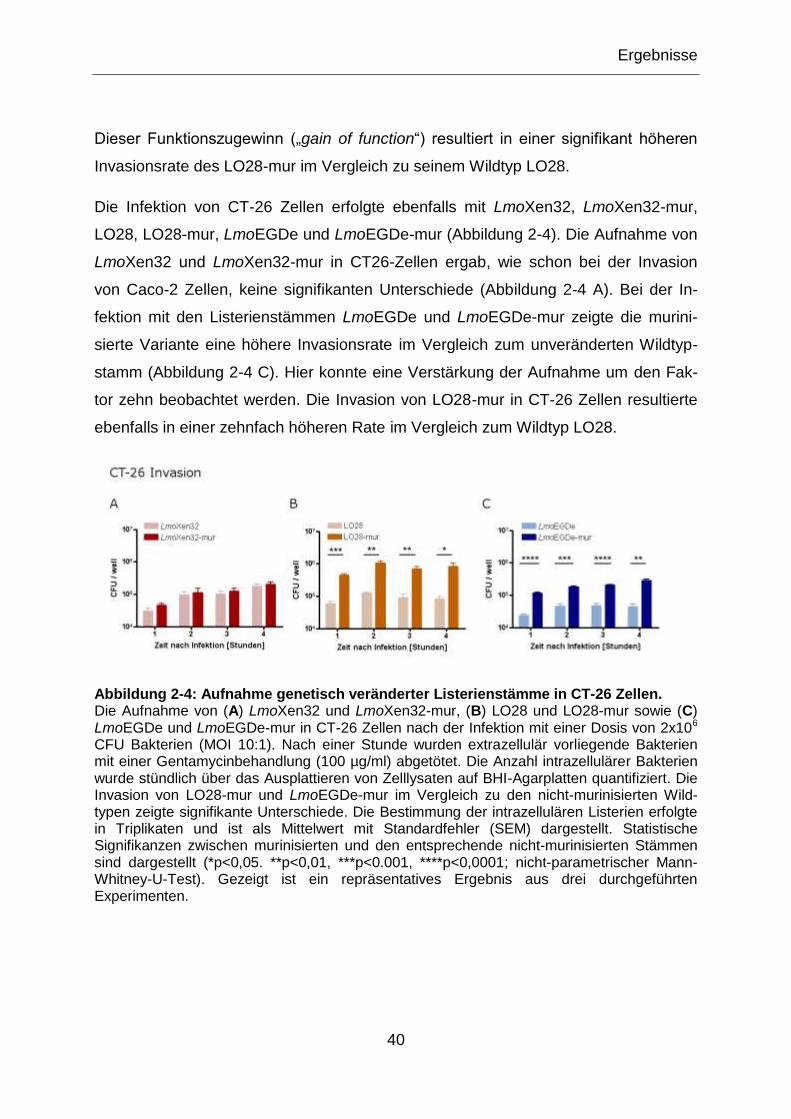
Ergebnisse
40
Dieser Funktionszugewinn („gain of function“) resultiert in einer signifikant höheren
Invasionsrate des LO28-mur im Vergleich zu seinem Wildtyp LO28.
Die Infektion von CT-26 Zellen erfolgte ebenfalls mit LmoXen32, LmoXen32-mur,
LO28, LO28-mur, LmoEGDe und LmoEGDe-mur (Abbildung 2-4). Die Aufnahme von
LmoXen32 und LmoXen32-mur in CT26-Zellen ergab, wie schon bei der Invasion
von Caco-2 Zellen, keine signifikanten Unterschiede (Abbildung 2-4 A). Bei der In-
fektion mit den Listerienstämmen LmoEGDe und LmoEGDe-mur zeigte die murini-
sierte Variante eine höhere Invasionsrate im Vergleich zum unveränderten Wildtyp-
stamm (Abbildung 2-4 C). Hier konnte eine Verstärkung der Aufnahme um den Fak-
tor zehn beobachtet werden. Die Invasion von LO28-mur in CT-26 Zellen resultierte
ebenfalls in einer zehnfach höheren Rate im Vergleich zum Wildtyp LO28.
Abbildung 2-4: Aufnahme genetisch veränderter Listerienstämme in CT-26 Zellen. Die Aufnahme von (A) LmoXen32 und LmoXen32-mur, (B) LO28 und LO28-mur sowie (C) LmoEGDe und LmoEGDe-mur in CT-26 Zellen nach der Infektion mit einer Dosis von 2x106 CFU Bakterien (MOI 10:1). Nach einer Stunde wurden extrazellulär vorliegende Bakterien mit einer Gentamycinbehandlung (100 µg/ml) abgetötet. Die Anzahl intrazellulärer Bakterien wurde stündlich über das Ausplattieren von Zelllysaten auf BHI-Agarplatten quantifiziert. Die Invasion von LO28-mur und LmoEGDe-mur im Vergleich zu den nicht-murinisierten Wild-typen zeigte signifikante Unterschiede. Die Bestimmung der intrazellulären Listerien erfolgte in Triplikaten und ist als Mittelwert mit Standardfehler (SEM) dargestellt. Statistische Signifikanzen zwischen murinisierten und den entsprechende nicht-murinisierten Stämmen sind dargestellt (*p<0,05. **p<0,01, ***p<0.001, ****p<0,0001; nicht-parametrischer Mann-Whitney-U-Test). Gezeigt ist ein repräsentatives Ergebnis aus drei durchgeführten Experimenten.

Ergebnisse
41
Es konnte gezeigt werden, dass die Invasionsrate des murinisierten Listerienstamms
LO28-mur in die humane Zelllinie Caco-2 sowie in die murine Colonkarzinom-Zelllinie
CT-26 im Vergleich zu LO28 signifikant erhöht sind. LmoEGDe-mur zeigte nur bei
der Invasion in murine Zellen einen deutlichen Unterschied im Vergleich zum
isogenen Wildtyp.
2.2 Vergleich der Wirtsspezifität und Virulenz des genetisch modi-
fizierten LmoXen32-mur mit dessen Wildtyp
Aufgrund der Anpassung des Internalin A an seinen murinen Rezeptor sollte jeder
Mausstamm für die orale Infektion mit murinisierten Listerien empfänglich sein. Zur
Beurteilung der Virulenz des generierten murinisierten Listerienstamms LmoXen32-
mur wurde im in vivo Mausmodell die Überlebensrate nach oraler Infektion in aus-
gewählten Mausinzuchtstämmen untersucht. Des Weiteren erfolgte die Bestimmung
der bakteriellen Keimlast in den inneren Organen nach der Infektion von LmoXen32-
mur für die Analyse der Invasionseffizienz und Replikation im Vergleich zum Wildtyp
LmoXen32.
2.2.1 Überlebenszeiten nach der Infektion mit LmoXen32 und LmoXen32-mur
in ausgewählten Mausinzuchtstämmen
Für die Bestimmung der Überlebensrate wurden die Mausinzuchtstämme C57BL/6J,
129P2/Ola, BALB/cJ und C3HeB/FeJ verwendet. In vorangegangenen Studien der
intravenösen und oralen Infektion mit Listeria monocytogenes konnte gezeigt wer-
den, dass sich C57BL/6J Mäuse resistent gegenüber einer Listerieninfektionen ver-
halten (Cheers et al., 1978; Cheers und McKenzie, 1978; Czuprynski et al., 2002,
Czuprynski et al., 2003; Iizawa et al., 1994). Dagegen zeigten sich andere Mausin-
zuchtstämme, darunter BALB/cJ und C3HeB/FeJ, empfänglicher für eine Infektion
mit Listeria monocytogenes (Archinal und Wilder, 1988; Cheers und Stanley, 1988:
Mainou-Fowler, 1988; Munder et al., 2005). Die Ursachen für eine gesteigerte Sus-
zeptibilität betreffen meist Komponenten des angeborenen und erworbenen Immun-
systems (Zenewicz und Shen, 2007).
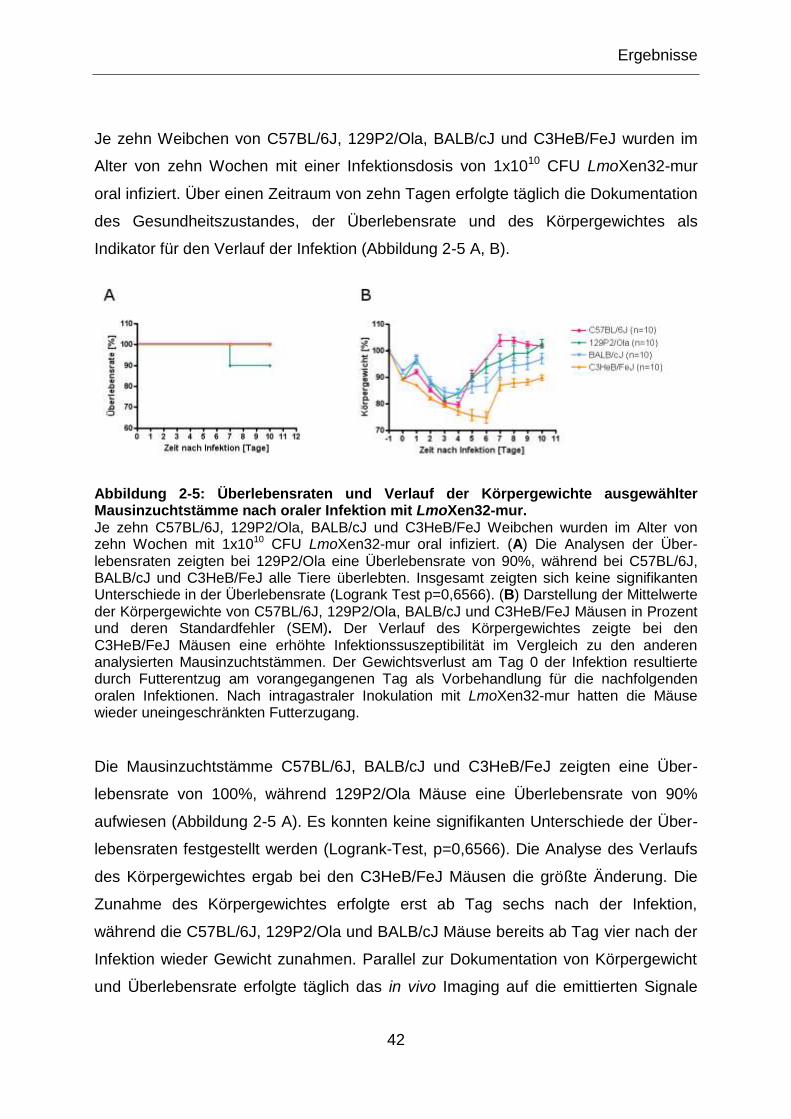
Ergebnisse
42
Je zehn Weibchen von C57BL/6J, 129P2/Ola, BALB/cJ und C3HeB/FeJ wurden im
Alter von zehn Wochen mit einer Infektionsdosis von 1x1010 CFU LmoXen32-mur
oral infiziert. Über einen Zeitraum von zehn Tagen erfolgte täglich die Dokumentation
des Gesundheitszustandes, der Überlebensrate und des Körpergewichtes als
Indikator für den Verlauf der Infektion (Abbildung 2-5 A, B).
Abbildung 2-5: Überlebensraten und Verlauf der Körpergewichte ausgewählter Mausinzuchtstämme nach oraler Infektion mit LmoXen32-mur. Je zehn C57BL/6J, 129P2/Ola, BALB/cJ und C3HeB/FeJ Weibchen wurden im Alter von zehn Wochen mit 1x1010 CFU LmoXen32-mur oral infiziert. (A) Die Analysen der Über-lebensraten zeigten bei 129P2/Ola eine Überlebensrate von 90%, während bei C57BL/6J, BALB/cJ und C3HeB/FeJ alle Tiere überlebten. Insgesamt zeigten sich keine signifikanten Unterschiede in der Überlebensrate (Logrank Test p=0,6566). (B) Darstellung der Mittelwerte der Körpergewichte von C57BL/6J, 129P2/Ola, BALB/cJ und C3HeB/FeJ Mäusen in Prozent und deren Standardfehler (SEM). Der Verlauf des Körpergewichtes zeigte bei den C3HeB/FeJ Mäusen eine erhöhte Infektionssuszeptibilität im Vergleich zu den anderen analysierten Mausinzuchtstämmen. Der Gewichtsverlust am Tag 0 der Infektion resultierte durch Futterentzug am vorangegangenen Tag als Vorbehandlung für die nachfolgenden oralen Infektionen. Nach intragastraler Inokulation mit LmoXen32-mur hatten die Mäuse wieder uneingeschränkten Futterzugang.
Die Mausinzuchtstämme C57BL/6J, BALB/cJ und C3HeB/FeJ zeigten eine Über-
lebensrate von 100%, während 129P2/Ola Mäuse eine Überlebensrate von 90%
aufwiesen (Abbildung 2-5 A). Es konnten keine signifikanten Unterschiede der Über-
lebensraten festgestellt werden (Logrank-Test, p=0,6566). Die Analyse des Verlaufs
des Körpergewichtes ergab bei den C3HeB/FeJ Mäusen die größte Änderung. Die
Zunahme des Körpergewichtes erfolgte erst ab Tag sechs nach der Infektion,
während die C57BL/6J, 129P2/Ola und BALB/cJ Mäuse bereits ab Tag vier nach der
Infektion wieder Gewicht zunahmen. Parallel zur Dokumentation von Körpergewicht
und Überlebensrate erfolgte täglich das in vivo Imaging auf die emittierten Signale
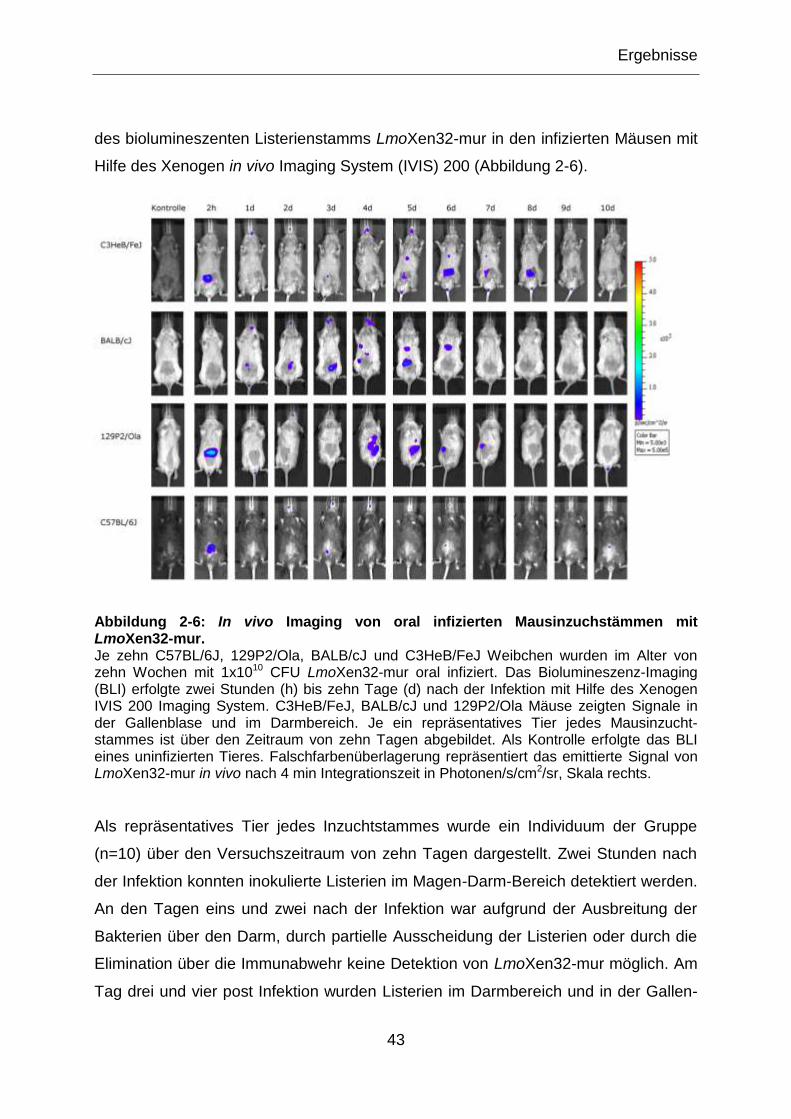
Ergebnisse
43
des biolumineszenten Listerienstamms LmoXen32-mur in den infizierten Mäusen mit
Hilfe des Xenogen in vivo Imaging System (IVIS) 200 (Abbildung 2-6).
Abbildung 2-6: In vivo Imaging von oral infizierten Mausinzuchstämmen mit LmoXen32-mur. Je zehn C57BL/6J, 129P2/Ola, BALB/cJ und C3HeB/FeJ Weibchen wurden im Alter von zehn Wochen mit 1x1010 CFU LmoXen32-mur oral infiziert. Das Biolumineszenz-Imaging (BLI) erfolgte zwei Stunden (h) bis zehn Tage (d) nach der Infektion mit Hilfe des Xenogen IVIS 200 Imaging System. C3HeB/FeJ, BALB/cJ und 129P2/Ola Mäuse zeigten Signale in der Gallenblase und im Darmbereich. Je ein repräsentatives Tier jedes Mausinzucht-stammes ist über den Zeitraum von zehn Tagen abgebildet. Als Kontrolle erfolgte das BLI eines uninfizierten Tieres. Falschfarbenüberlagerung repräsentiert das emittierte Signal von LmoXen32-mur in vivo nach 4 min Integrationszeit in Photonen/s/cm2/sr, Skala rechts.
Als repräsentatives Tier jedes Inzuchtstammes wurde ein Individuum der Gruppe
(n=10) über den Versuchszeitraum von zehn Tagen dargestellt. Zwei Stunden nach
der Infektion konnten inokulierte Listerien im Magen-Darm-Bereich detektiert werden.
An den Tagen eins und zwei nach der Infektion war aufgrund der Ausbreitung der
Bakterien über den Darm, durch partielle Ausscheidung der Listerien oder durch die
Elimination über die Immunabwehr keine Detektion von LmoXen32-mur möglich. Am
Tag drei und vier post Infektion wurden Listerien im Darmbereich und in der Gallen-

Ergebnisse
44
blase von BALB/cJ und 129P2/Ola Mäusen nachgewiesen. Ab Tag fünf nach der
Infektion konnte auch in der Gallenblase und im abdominalen Bereich der
C3HeB/FeJ Mäuse Listerien detektiert werden. Innerhalb dieser Organe waren die
Bakterien bis Tag sechs und sieben, in C3HeB/FeJ Mäusen bis zum Tag acht, nach
der Infektion detektierbar. In C57BL/6J Mäusen konnten ab dem Tag eins nach der
Infektion keine spezifischen Biolumineszenzsignale nachgewiesen werden. Durch
Einsetzen der adaptiven Immunantwort ab Tag sieben nach der Infektion erfolgte die
Rekonvaleszenz und die Biolumineszenz konnte nachfolgend nicht mehr detektiert
werden. Die einzelnen Inzuchtstämme wiesen Unterschiede in der Detektierbarkeit
der Biolumineszenzsignale von LmoXen32-mur auf. In unpigmentierten bzw. wenig
pigmentierten Mäusestämmen wie BALB/cJ und 129P2/Ola waren die
Biolumineszenzsignale der Listerien gut detektierbar. Dagegen war in pigmentierten
Mäusen, wie C3HeB/FeJ und C57BL/6J, aufgrund der dunklen Fellfärbung und der
Pigmentierung der Haut die Detektion der Biolumineszenz reduziert.
Die orale Infektion der Mausinzuchtstämme C57BL/6J, 129P2/Ola, BALB/cJ und
C3HeB/FeJ mit LmoXen32-mur resultierte in geringen Unterschieden in der Infek-
tionssuszeptibilität. C57BL/6J erwiesen sich als resistent, dagegen zeigten sich die
C3HeB/FeJ aufgrund eines starken Gewichtsverlustes und 129P2/Ola durch eine
geringere Überlebensrate empfindlicher für die Listerieninfektion. Jedoch resultierte
die LmoXen32-mur Infektion in einer geringen Pathologie mit geringerer Mortalitäts-
rate. Wie bei Hardy et al. beschrieben, war die Detektion der Biolumineszenzsignale
auf die Gallenblase und den Abdominalbereich begrenzt (Hardy et al., 2004). Somit
wird deutlich, dass aufgrund der attenuierten Virulenz des LmoXen32-mur ein Ver-
gleich der Wirtsresistenz ausgewählter Mausinzuchtstämme nur bedingt möglich ist,
da alle Mausstämme kaum von der Infektion betroffen waren und LmoXen32-mur gut
abwehren konnten.
2.2.2 Kinetik der in vivo Replikation von LmoXen32 und LmoXen32-mur in
BALB/cJ Mäusen
Die Analyse des BLI in ausgewählten Mausinzuchtstämmen ergab, dass LmoXen32-
mur ein attenuierter Listerienstamm ist und Unterschiede der Suszeptibilität

Ergebnisse
45
ausgewählter Mausinzuchtstämme im in vivo Modell schwer erkennbar sind.
Aufgrund dieser Beobachtung erfolgte die Analyse der bakteriellen Keimlast in den
inneren Organen von BALB/cJ Mäusen (Gruppengröße n=8) nach oraler Infektion mit
LmoXen32-mur im Vergleich zum Wildtypstamm LmoXen32. An den Tagen eins,
drei, fünf und sieben nach der Infektion wurden jeweils acht Tiere für die Analyse
verwendet. Parallel erfolgte die histopathologische Untersuchung der Organe Milz,
Leber und Dünndarm sowie täglich die Dokumentation der Änderungen im Verlauf
des Körpergewichtes und die Detektion der Biolumineszenz aller infizierten Tiere
über das in vivo Imaging.
2.2.2.1 In vivo Imaging von LmoXen32 und LmoXen32-mur infizierten BALB/cJ
Mäusen
Je 36 BALB/cJ Weibchen wurden im Alter von zehn Wochen mit einer Infektions-
dosis von 5x109 CFU LmoXen32 bzw. LmoXen32-mur oral infiziert. Über den Ver-
suchszeitraum von sieben Tagen erfolgte das tägliche Biolumineszenz-Imaging der
BALB/cJ Mäuse mit dem IVIS 200 System (Abbildung 2-7). Eine Stunde nach der
Infektion sind beide Listerienstämme im Magen der Tiere detektierbar. An den Tagen
eins und zwei nach der Infektion war die Detektion des Signals aufgrund der
Dissemination der Bakterien über den Darm und deren Ausscheidung reduziert. Am
Tag drei post Infektion konnten Listerien wieder im Abdominalbereich von
LmoXen32-mur infizierten Tieren nachgewiesen werden. Ab diesem Zeitpunkt sind
Unterschiede zwischen LmoXen32 und LmoXen32-mur erkennbar. LmoXen32 war
weniger detektierbar als die murinisierte Variante und die messbare Verbreitung
begrenzt sich nur auf den Darmbereich. Ab Tag drei nach der Infektion zeigten
LmoXen32-mur infizierte BALB/cJ Mäuse eine stärkere Biolumineszenz im
abdominalen Bereich, im Vergleich zu LmoXen32 infizierten Mäusen. Zusätzlich
konnten an Tag vier und fünf nach der Infektion mit LmoXen32-mur deutliche
Biolumineszenzsignale in der Gallenblase detektiert werden, was eine Akkumulation
und Replikation der Listerien innerhalb des Organs anzeigte. LmoXen32-mur war bei
einzelnen Tieren bis zum Tag sieben nach der Infektion in der Gallenblase und im
Darmbereich detektierbar.
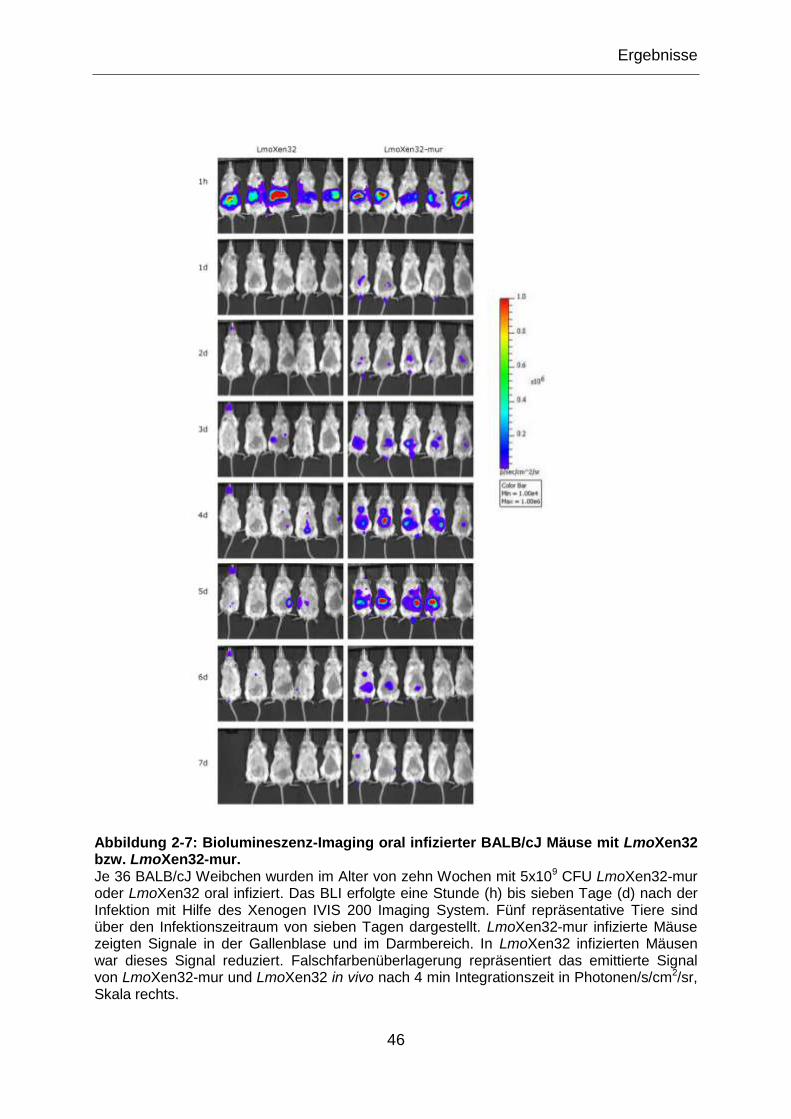
Ergebnisse
46
Abbildung 2-7: Biolumineszenz-Imaging oral infizierter BALB/cJ Mäuse mit LmoXen32 bzw. LmoXen32-mur. Je 36 BALB/cJ Weibchen wurden im Alter von zehn Wochen mit 5x109 CFU LmoXen32-mur oder LmoXen32 oral infiziert. Das BLI erfolgte eine Stunde (h) bis sieben Tage (d) nach der Infektion mit Hilfe des Xenogen IVIS 200 Imaging System. Fünf repräsentative Tiere sind über den Infektionszeitraum von sieben Tagen dargestellt. LmoXen32-mur infizierte Mäuse zeigten Signale in der Gallenblase und im Darmbereich. In LmoXen32 infizierten Mäusen war dieses Signal reduziert. Falschfarbenüberlagerung repräsentiert das emittierte Signal von LmoXen32-mur und LmoXen32 in vivo nach 4 min Integrationszeit in Photonen/s/cm2/sr, Skala rechts.
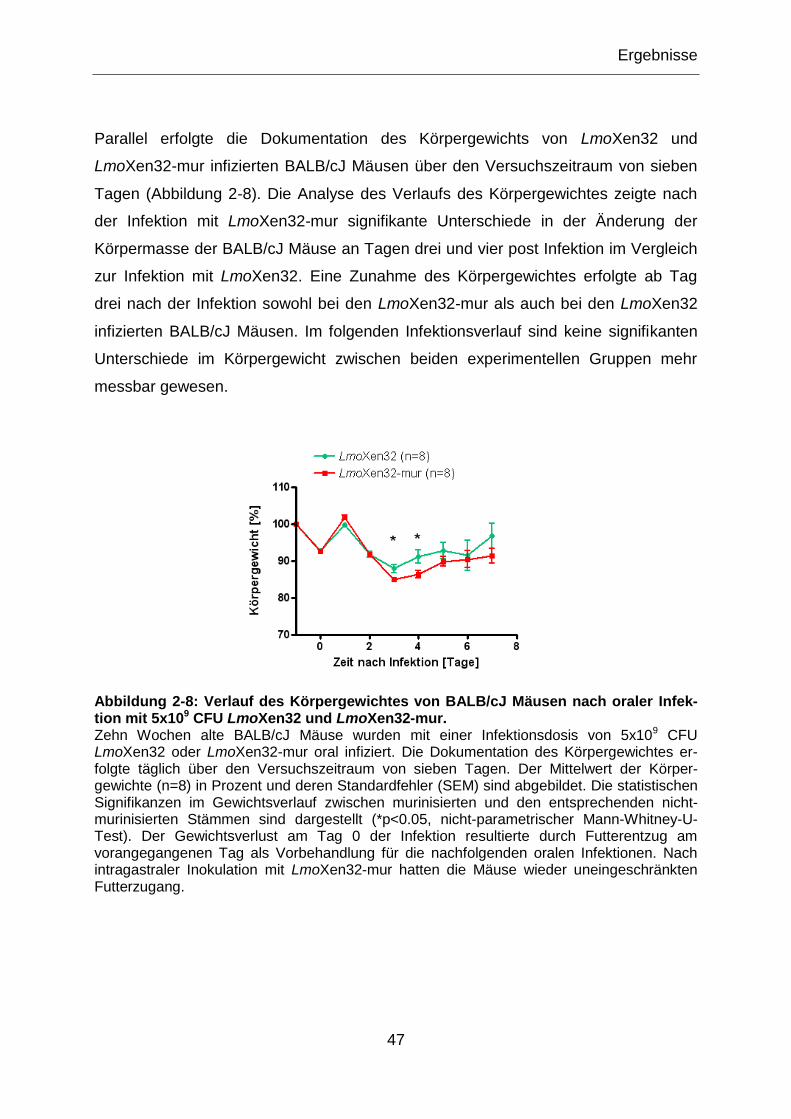
Ergebnisse
47
Parallel erfolgte die Dokumentation des Körpergewichts von LmoXen32 und
LmoXen32-mur infizierten BALB/cJ Mäusen über den Versuchszeitraum von sieben
Tagen (Abbildung 2-8). Die Analyse des Verlaufs des Körpergewichtes zeigte nach
der Infektion mit LmoXen32-mur signifikante Unterschiede in der Änderung der
Körpermasse der BALB/cJ Mäuse an Tagen drei und vier post Infektion im Vergleich
zur Infektion mit LmoXen32. Eine Zunahme des Körpergewichtes erfolgte ab Tag
drei nach der Infektion sowohl bei den LmoXen32-mur als auch bei den LmoXen32
infizierten BALB/cJ Mäusen. Im folgenden Infektionsverlauf sind keine signifikanten
Unterschiede im Körpergewicht zwischen beiden experimentellen Gruppen mehr
messbar gewesen.
Abbildung 2-8: Verlauf des Körpergewichtes von BALB/cJ Mäusen nach oraler Infek-tion mit 5x109 CFU LmoXen32 und LmoXen32-mur. Zehn Wochen alte BALB/cJ Mäuse wurden mit einer Infektionsdosis von 5x109 CFU LmoXen32 oder LmoXen32-mur oral infiziert. Die Dokumentation des Körpergewichtes er-folgte täglich über den Versuchszeitraum von sieben Tagen. Der Mittelwert der Körper-gewichte (n=8) in Prozent und deren Standardfehler (SEM) sind abgebildet. Die statistischen Signifikanzen im Gewichtsverlauf zwischen murinisierten und den entsprechenden nicht-murinisierten Stämmen sind dargestellt (*p<0.05, nicht-parametrischer Mann-Whitney-U-Test). Der Gewichtsverlust am Tag 0 der Infektion resultierte durch Futterentzug am vorangegangenen Tag als Vorbehandlung für die nachfolgenden oralen Infektionen. Nach intragastraler Inokulation mit LmoXen32-mur hatten die Mäuse wieder uneingeschränkten Futterzugang.

Ergebnisse
48
2.2.2.2 Kinetik der bakteriellen Keimlast in den inneren Organen von LmoXen32 und
LmoXen32-mur infizierten BALB/cJ Mäusen
Zur Untersuchung des Infektionsverlaufs und der Dissemination von Listeria mono-
cytogenes zu den inneren Organen wurden BALB/cJ Mäuse mit jeweils 5x109 CFU
LmoXen32 oder LmoXen32-mur oral infiziert. An den Tagen eins, drei, fünf und sie-
ben nach der Infektion wurden die Organe Milz, Leber, mesenterische Lymphknoten
(mLK), Dünndarm, Gallenblase und Gehirn entnommen und auf die bakterielle
Keimlast untersucht. Zur Bestimmung der Listerienanzahl wurden die Organe in PBS
homogenisiert und serielle Verdünnungen auf BHI-Agarplatten ausgestrichen. In
Abbildung 2-9 sind die Keimlasten der verschiedenen Organe für die Tage drei (A),
fünf (B) und sieben (C) dargestellt. Am Tag drei nach der Infektion zeigten sich nur
geringe Unterscheide in den Keimlasten von LmoXen32 und LmoXen32-mur in den
Organen Milz, Dünndarm, Gehirn und mLK. Signifikante Unterschiede konnten in der
Leber und der Gallenblase nachgewiesen werden. Die Keimlasten von LmoXen32-
mur waren um den Faktor zehn höher als die Keimlasten der LmoXen32 infizierten
Organe (Abbildung 2-9 A). Im Verlauf der Infektion sank die Listerienanzahl in den
inneren Organen Milz, Leber, mLK, Gehirn und Dünndarm von Tag drei auf Tag fünf
um den Faktor zehn, unabhängig vom Listerienstamm. In der Gallenblase erfolgte
dagegen die Zunahme der Listerienanzahl von Tag drei auf Tag fünf nach der
Infektion mit LmoXen32 und LmoXen32-mur um das Zehnfache. Am Tag fünf nach
der Infektion waren signifikante Unterschiede in der Keimlast der mesenterischen
Lymphknoten zwischen LmoXen32-mur und LmoXen32 infizierten Tieren erkennbar
(Abbildung 2-9 B). Alle anderen Organe wiesen keine Unterschiede in der
Listerienanzahl auf. Von Tag fünf auf Tag sieben nach der Infektion reduzierte sich
die Bakterienanzahl im Dünndarm sowie in der Milz und den mLK der LmoXen32
infizierten Mäuse um den Faktor 100. Dagegen konnte keine Reduktion der
bakteriellen Keimlast in den Organen Milz und mLK der LmoXen32-mur infizierten
Tieren festgestellt werden. So konnten am Tag sieben nach der Infektion signifikante
Unterschiede zwischen den Keimlasten beider Bakterienstämme in den mLK und in
der Milz beobachtet werden.
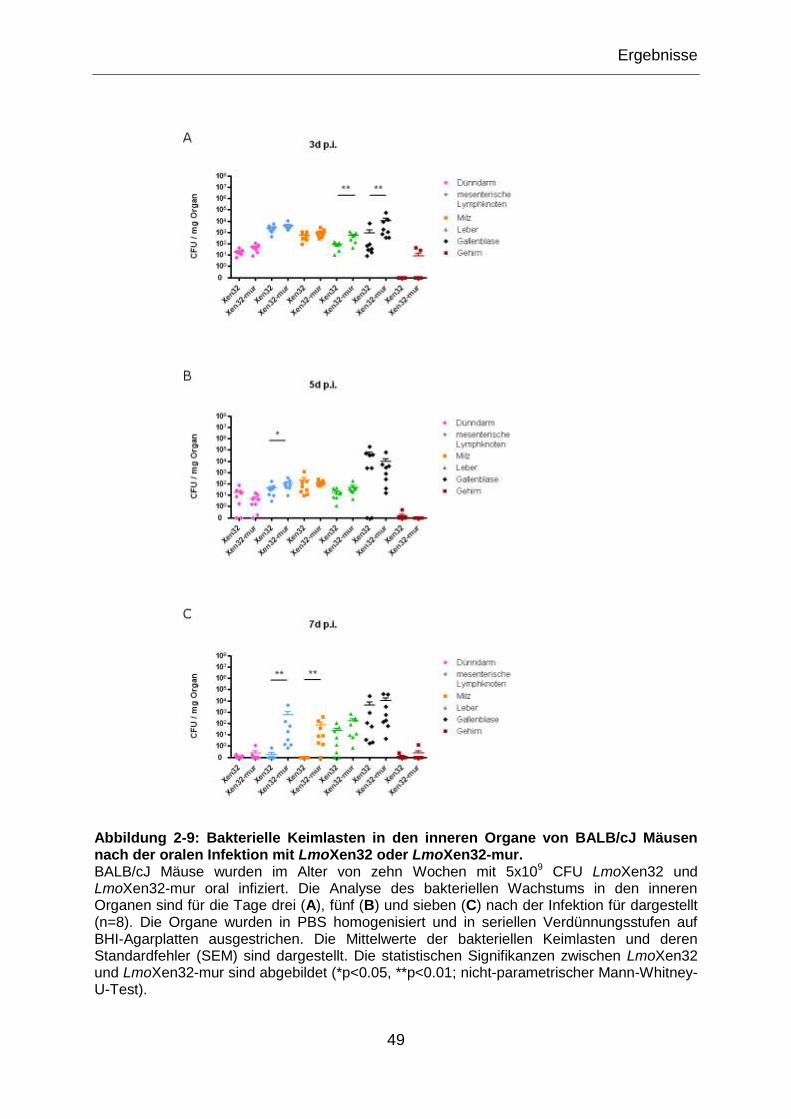
Ergebnisse
49
Abbildung 2-9: Bakterielle Keimlasten in den inneren Organe von BALB/cJ Mäusen nach der oralen Infektion mit LmoXen32 oder LmoXen32-mur. BALB/cJ Mäuse wurden im Alter von zehn Wochen mit 5x109 CFU LmoXen32 und LmoXen32-mur oral infiziert. Die Analyse des bakteriellen Wachstums in den inneren Organen sind für die Tage drei (A), fünf (B) und sieben (C) nach der Infektion für dargestellt (n=8). Die Organe wurden in PBS homogenisiert und in seriellen Verdünnungsstufen auf BHI-Agarplatten ausgestrichen. Die Mittelwerte der bakteriellen Keimlasten und deren Standardfehler (SEM) sind dargestellt. Die statistischen Signifikanzen zwischen LmoXen32 und LmoXen32-mur sind abgebildet (*p<0.05, **p<0.01; nicht-parametrischer Mann-Whitney-U-Test).

Ergebnisse
50
2.2.2.3 Histopathologische Analyse von LmoXen32 und LmoXen32-mur infizierten
BALB/cJ Mäusen
Neben der Analyse der bakteriellen Keimlast erfolgte parallel die histopathologische
und immunhistologische Untersuchung der Organe Milz, Leber und Dünndarm.
Hierzu wurden die Organe an Tag eins, drei, fünf und sieben nach der Infektion prä-
pariert, in Formaldehyd fixiert und mit Hämatoxylin-Eosin (HE) gefärbt. Weiterhin
wurde mit Hilfe immunhistologischer Verfahren versucht Listeria monocytogenes
innerhalb der Gewebe nachzuweisen. Abbildung 2-10 zeigt die histopathologische
Analyse der Leber an Tag fünf nach der oralen Infektion mit 5x109 CFU LmoXen32
und LmoXen32-mur. Sowohl die orale Infektion des Listerienstamms LmoXen32 als
auch von LmoXen32-mur führte zu moderaten Schädigungen des Gewebes mit
Nekrosen und Infiltrationen von Granulozyten und Makrophagen innerhalb des
Leberparenchyms. Der Vergleich der LmoXen32 und der LmoXen32-mur infizierten
Organen wies nur geringe Unterschiede im Schweregrad der Pathologie auf. Die
orale Infektion mit LmoXen32-mur führte vermehrt zu Infiltrationen von Granulozyten
und Makrophagen in der Leber als die Infektion mit LmoXen32. An Tag fünf post
Infektion konnte das größte Ausmaß der Schädigung des Gewebes, in Anzahl der
Nekrosen und Infiltration von Immunzellen, festgestellt werden. Ab Tag sieben nach
der Infektion war sowohl in den LmoXen32 als auch in den LmoXen32-mur infizierten
Organen eine Verringerung der entzündlichen Läsionen aufgrund der geringeren
Anzahl von Nekrosen und Zellinfiltrationen zu beobachten. Mit Hilfe der immun-
histologischen Analyse konnte der Nachweis von LmoXen32-mur und LmoXen32
innerhalb der entzündlichen Infiltrate des Leberparenchyms erbracht werden. Die
histopathologische Analyse der Milz an Tag fünf nach der Infektion führte ebenfalls
zum Nachweis inflammatorischer Läsionen mit Infiltrationen von Makrophagen und
Granulozyten sowohl in den LmoXen32 als auch in den LmoXen32-mur infizierten
Tieren (Daten nicht gezeigt).
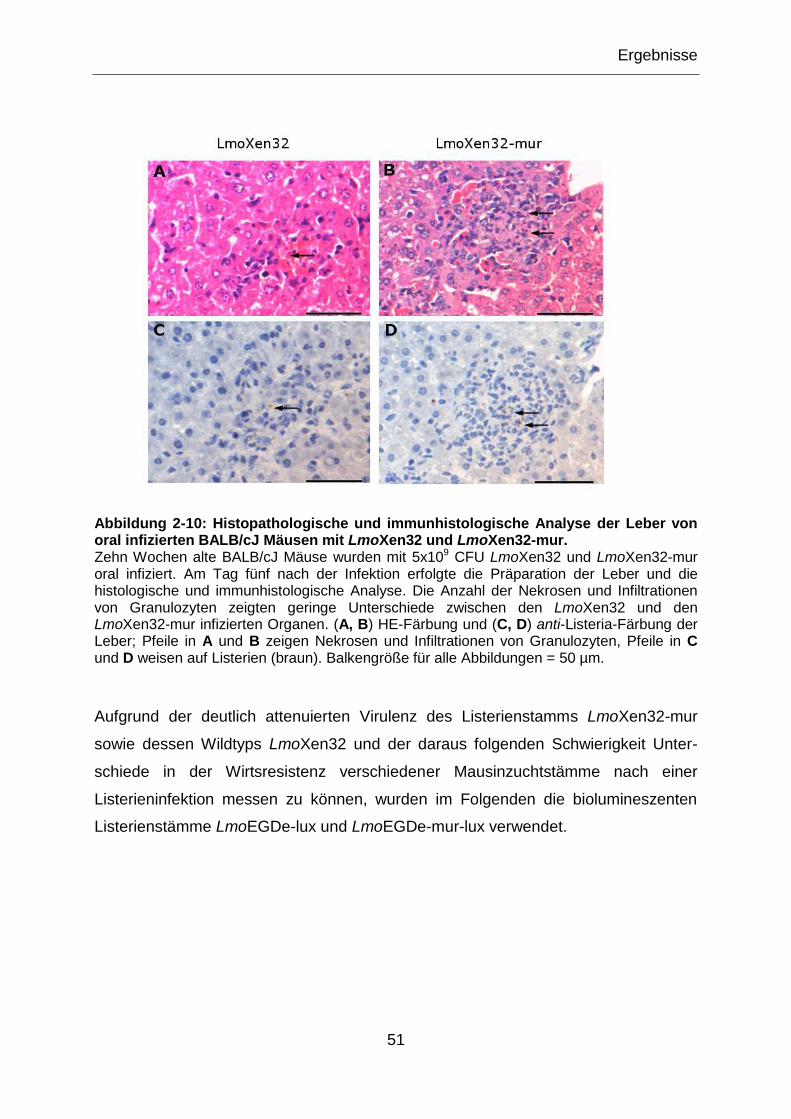
Ergebnisse
51
Abbildung 2-10: Histopathologische und immunhistologische Analyse der Leber von oral infizierten BALB/cJ Mäusen mit LmoXen32 und LmoXen32-mur. Zehn Wochen alte BALB/cJ Mäuse wurden mit 5x109 CFU LmoXen32 und LmoXen32-mur oral infiziert. Am Tag fünf nach der Infektion erfolgte die Präparation der Leber und die histologische und immunhistologische Analyse. Die Anzahl der Nekrosen und Infiltrationen von Granulozyten zeigten geringe Unterschiede zwischen den LmoXen32 und den LmoXen32-mur infizierten Organen. (A, B) HE-Färbung und (C, D) anti-Listeria-Färbung der Leber; Pfeile in A und B zeigen Nekrosen und Infiltrationen von Granulozyten, Pfeile in C und D weisen auf Listerien (braun). Balkengröße für alle Abbildungen = 50 µm.
Aufgrund der deutlich attenuierten Virulenz des Listerienstamms LmoXen32-mur
sowie dessen Wildtyps LmoXen32 und der daraus folgenden Schwierigkeit Unter-
schiede in der Wirtsresistenz verschiedener Mausinzuchtstämme nach einer
Listerieninfektion messen zu können, wurden im Folgenden die biolumineszenten
Listerienstämme LmoEGDe-lux und LmoEGDe-mur-lux verwendet.

Ergebnisse
52
2.3 Vergleich der Wirtsspezifität und Virulenz des genetisch
modifizierten LmoEGDe-lux-mur mit dessen Wildtyp
Die Listerienstämme LmoXen32 und LmoXen32-mur zeigten in den voran-
gegangenen Experimenten eine deutlich attenuierte Virulenz. Dies resultierte in einer
Überlebensrate von 100% in BALB/cJ, C3HeB/FeJ und C57BL/6J Mäusen nach
einer oralen Infektion mit 1x1010 CFU sowie bei BALB/cJ Mäusen in einer geringen
Invasion der inneren Organe. Der Stamm LmoXen32 ist ein Derivat des Lmo
10403S. Bei der Generierung von LmoXen32 erfolgte die Integration des Luziferase-
Operons mittels Transposonmutagenese in die Promotorregion des Flagellin A (flaA)
Gens. Dies resultierte in einer vierfach höheren LD50 des LmoXen32 als im Ver-
gleich zu seinem Ursprungsstamm Lmo 10403S (Hardy et al., 2004). Aufgrund dieser
deutlichen Attenuierung wurden die folgenden Experimente mit den Listerien-
stämmen LmoEGDe-lux und LmoEGDe-mur-lux durchgeführt. Diese Listerien-
stämme wurden freundlicherweise von Dr. Gahan Cormac bereitgestellt (Department
of Microbiology, University College Cork, Irland). Zur Generierung des biolumines-
zenten Listerienstamms LmoEGDe-lux erfolgte die Integration einer einzelnen Kopie
der luxABCDE Gene mit Hilfe des pPL2lux Vektors in die 3`Region des tRNAArg-
Gens des Chromosoms von Listeria monocytogenes (Bron et al., 2006; Riedel et al.,
2007). LmoEGDe-mur-lux wurde ausgehend von murinisierten Listerienstamm
LmoEGDe-mur (S192N-Y369S) über die Integration des Lux-Operons mit Hilfe des
Listeriophagen Integrationsvektors pIMK2lux generiert (Monk et al., 2010). Dies er-
möglicht die direkte Interaktion von Internalin A mit seinem murinem Rezeptor E-
Cadherin und erlaubt somit Untersuchung der Wirt-Pathogen-Interaktion im Maus-
modell nach oraler Infektion. Beide zur Verfügung gestellten Stämme, LmoEGDe-lux
und LmoEGDe-lux-mur, wurden auf die Expression der Oberflächenproteine Inter-
nalin A und Internalin B untersucht sowie auf das Invasionsverhalten in die humane
Colonkarzinom-Zelllinie Caco-2 und in die murinen Colonkarzinom-Zellen CT-26.
Dies diente als Vorrausetzung für die in vivo Analyse der oralen Infektion in
ausgewählten Mausinzuchtstämmen.

Ergebnisse
53
2.3.1 Analyse der Stämme LmoEGDe-lux und LmoEGDe-lux-mur auf die
Proteinexpression von Internalin A und Internalin B sowie die Unter-
suchung der Invasionsrate in humanen und murinen Colonkarzinom-
Zellen.
Wie bereits unter Punkt 2.1.3 beschrieben, erfolgte die Analyse der Expression der
Oberflächenproteine InlA und InlB in den genetisch veränderten Listerienstämmen
LmoEGDe-lux und LmoEGDe-mur-lux über die Durchführung eines Western-Blots.
Die kompletten Proteinlysate wurden auf einem Polyacrylamidgel aufgetragen und
über das Immunoblotverfahren mit monoklonalen Antikörpern gegen InlA und InlB
nachgewiesen (Abbildung 2-11). Als Ladekontrolle für die relativ geladenen Protein-
konzentration und die Proteindegradation diente wie bereits in Abschnitt 2.1.3 be-
schrieben der Nachweis des Aktin-bindenden Proteins A. Die erwarteten Mole-
kulargewichte lagen bei InlA bei 88 kDa, bei InlB 65 bei kDa und bei ActA bei 90 kDa
(Lingenau et al., 1995). Die Analyse der Proteinexpression zeigte, dass sämtliche
getesteten Listerienstämme InlA, InlB und ActA exprimieren.
Mit Hilfe des Gentamycin-Invasions-Assays wurde analysiert, ob eine verstärkte Auf-
nahme der murinisierten Listerienstämme aufgrund einer erhöhten Bindungsaffinität
zum Rezeptor in humane Caco-2 und murine CT-26 Zellen erfolgte. Untersucht
wurde die Invasionsrate des murinisierten Listerienstamms LmoEGDe-mur-lux und
von dessen unmodifizierten Wildtyps LmoEGDe-lux in Caco-2 und CT-26 Zellen
(Abbildung 2-12). Eine Stunde nach der Infektion mit 2x106 CFU Listerien erfolgte die
Behandlung mit Gentamycin zur Abtötung aller extrazellulär vorliegenden Bakterien.
Somit wurden nur die intrazellulär vorhandenen und replizierenden Listerien erfasst.
Die murinisierte Variante von LmoEGDe-lux zeigte im Caco-2 Invasions-Assay keine
signifikanten Unterschiede zu dem entsprechenden nicht-murinisierten Listerien-
stamm (Abbildung 2-12 A). Die Infektion von CT-26 Zellen ergab signifikante Unter-
schiede zwischen der Invasion von LmoEGDe-lux und LmoEGDe-mur-lux (Abbildung
2-12 B).
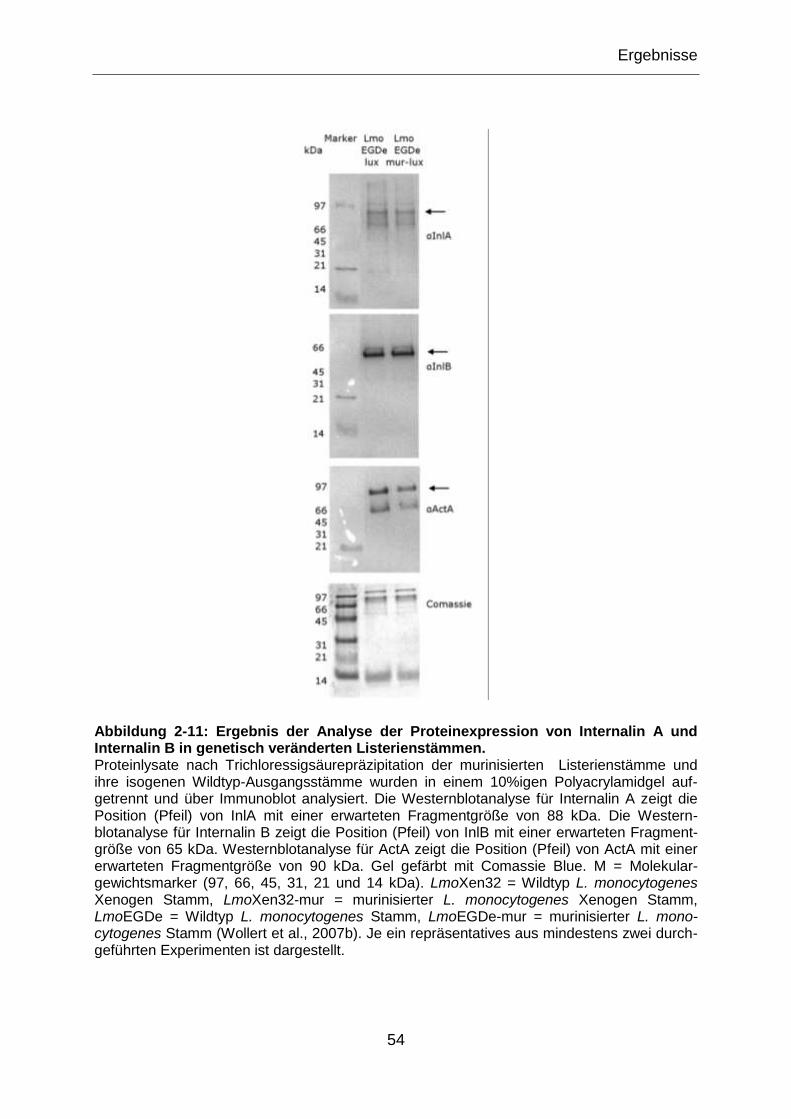
Ergebnisse
54
Abbildung 2-11: Ergebnis der Analyse der Proteinexpression von Internalin A und Internalin B in genetisch veränderten Listerienstämmen. Proteinlysate nach Trichloressigsäurepräzipitation der murinisierten Listerienstämme und ihre isogenen Wildtyp-Ausgangsstämme wurden in einem 10%igen Polyacrylamidgel auf-getrennt und über Immunoblot analysiert. Die Westernblotanalyse für Internalin A zeigt die Position (Pfeil) von InlA mit einer erwarteten Fragmentgröße von 88 kDa. Die Western-blotanalyse für Internalin B zeigt die Position (Pfeil) von InlB mit einer erwarteten Fragment-größe von 65 kDa. Westernblotanalyse für ActA zeigt die Position (Pfeil) von ActA mit einer erwarteten Fragmentgröße von 90 kDa. Gel gefärbt mit Comassie Blue. M = Molekular-gewichtsmarker (97, 66, 45, 31, 21 und 14 kDa). LmoXen32 = Wildtyp L. monocytogenes Xenogen Stamm, LmoXen32-mur = murinisierter L. monocytogenes Xenogen Stamm, LmoEGDe = Wildtyp L. monocytogenes Stamm, LmoEGDe-mur = murinisierter L. mono-cytogenes Stamm (Wollert et al., 2007b). Je ein repräsentatives aus mindestens zwei durch-geführten Experimenten ist dargestellt.
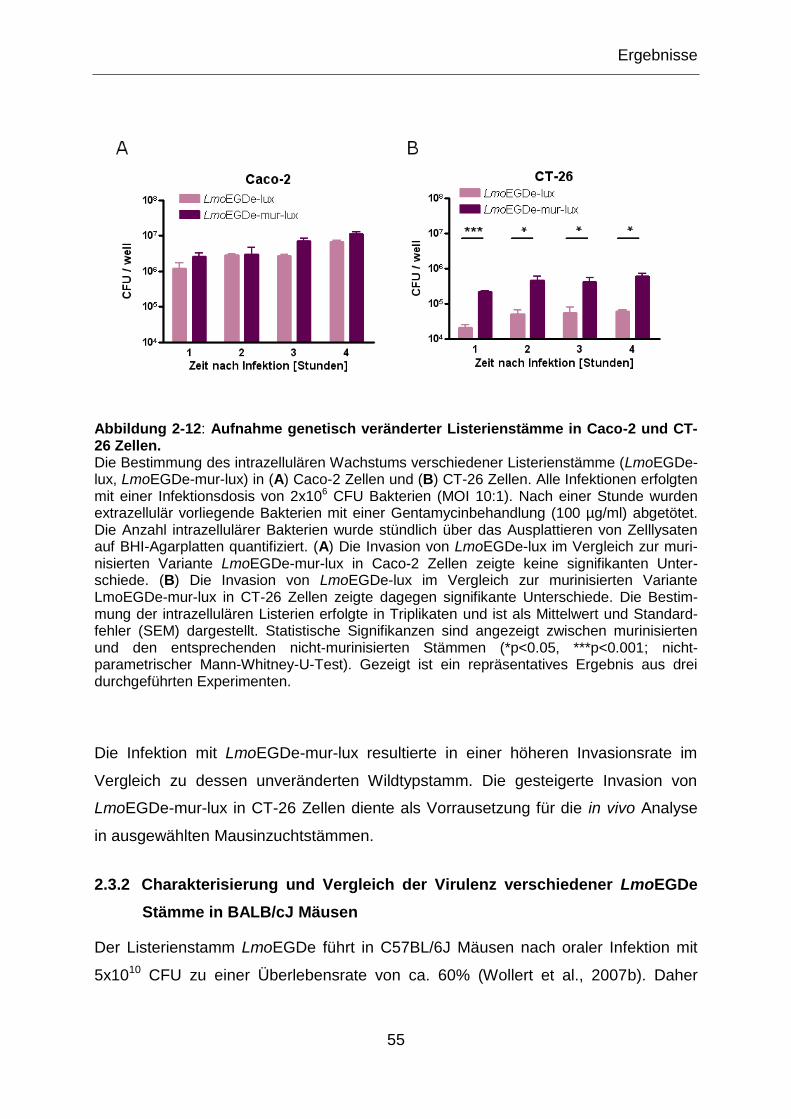
Ergebnisse
55
Abbildung 2-12: Aufnahme genetisch veränderter Listerienstämme in Caco-2 und CT-26 Zellen. Die Bestimmung des intrazellulären Wachstums verschiedener Listerienstämme (LmoEGDe-lux, LmoEGDe-mur-lux) in (A) Caco-2 Zellen und (B) CT-26 Zellen. Alle Infektionen erfolgten mit einer Infektionsdosis von 2x106 CFU Bakterien (MOI 10:1). Nach einer Stunde wurden extrazellulär vorliegende Bakterien mit einer Gentamycinbehandlung (100 µg/ml) abgetötet. Die Anzahl intrazellulärer Bakterien wurde stündlich über das Ausplattieren von Zelllysaten auf BHI-Agarplatten quantifiziert. (A) Die Invasion von LmoEGDe-lux im Vergleich zur muri-nisierten Variante LmoEGDe-mur-lux in Caco-2 Zellen zeigte keine signifikanten Unter-schiede. (B) Die Invasion von LmoEGDe-lux im Vergleich zur murinisierten Variante LmoEGDe-mur-lux in CT-26 Zellen zeigte dagegen signifikante Unterschiede. Die Bestim-mung der intrazellulären Listerien erfolgte in Triplikaten und ist als Mittelwert und Standard-fehler (SEM) dargestellt. Statistische Signifikanzen sind angezeigt zwischen murinisierten und den entsprechenden nicht-murinisierten Stämmen (*p<0.05, ***p<0.001; nicht-parametrischer Mann-Whitney-U-Test). Gezeigt ist ein repräsentatives Ergebnis aus drei durchgeführten Experimenten.
Die Infektion mit LmoEGDe-mur-lux resultierte in einer höheren Invasionsrate im
Vergleich zu dessen unveränderten Wildtypstamm. Die gesteigerte Invasion von
LmoEGDe-mur-lux in CT-26 Zellen diente als Vorrausetzung für die in vivo Analyse
in ausgewählten Mausinzuchtstämmen.
2.3.2 Charakterisierung und Vergleich der Virulenz verschiedener LmoEGDe
Stämme in BALB/cJ Mäusen
Der Listerienstamm LmoEGDe führt in C57BL/6J Mäusen nach oraler Infektion mit
5x1010 CFU zu einer Überlebensrate von ca. 60% (Wollert et al., 2007b). Daher
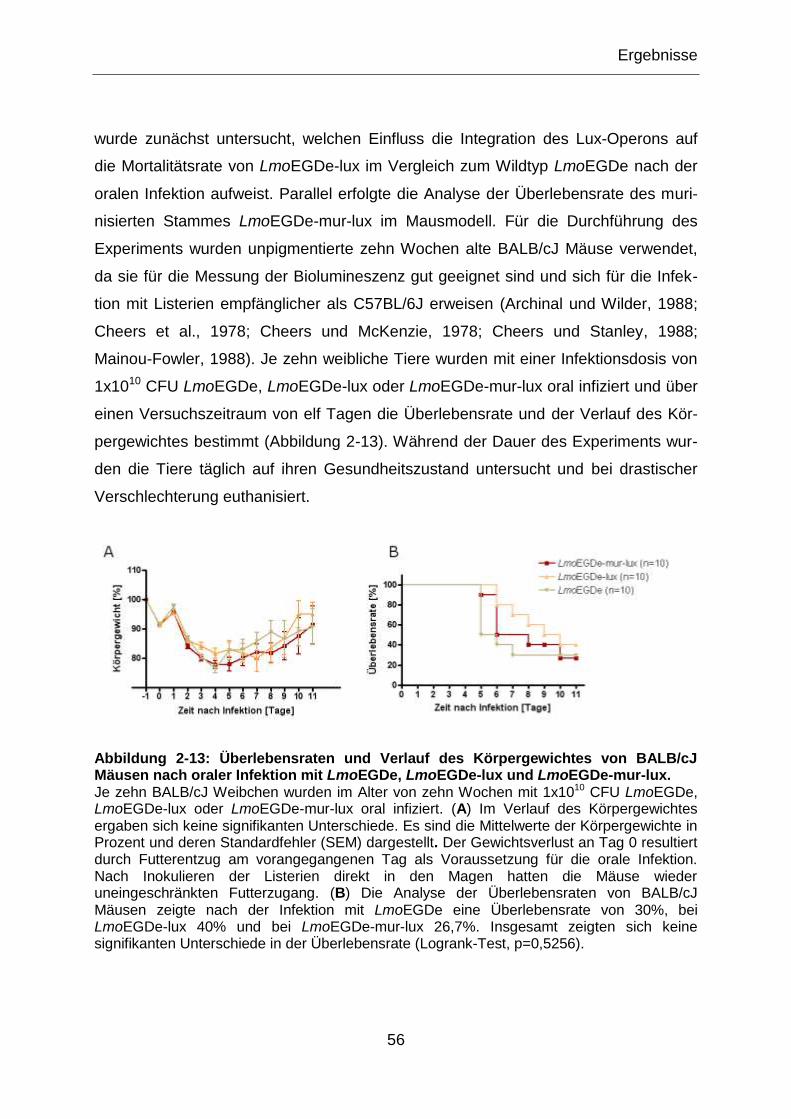
Ergebnisse
56
wurde zunächst untersucht, welchen Einfluss die Integration des Lux-Operons auf
die Mortalitätsrate von LmoEGDe-lux im Vergleich zum Wildtyp LmoEGDe nach der
oralen Infektion aufweist. Parallel erfolgte die Analyse der Überlebensrate des muri-
nisierten Stammes LmoEGDe-mur-lux im Mausmodell. Für die Durchführung des
Experiments wurden unpigmentierte zehn Wochen alte BALB/cJ Mäuse verwendet,
da sie für die Messung der Biolumineszenz gut geeignet sind und sich für die Infek-
tion mit Listerien empfänglicher als C57BL/6J erweisen (Archinal und Wilder, 1988;
Cheers et al., 1978; Cheers und McKenzie, 1978; Cheers und Stanley, 1988;
Mainou-Fowler, 1988). Je zehn weibliche Tiere wurden mit einer Infektionsdosis von
1x1010 CFU LmoEGDe, LmoEGDe-lux oder LmoEGDe-mur-lux oral infiziert und über
einen Versuchszeitraum von elf Tagen die Überlebensrate und der Verlauf des Kör-
pergewichtes bestimmt (Abbildung 2-13). Während der Dauer des Experiments wur-
den die Tiere täglich auf ihren Gesundheitszustand untersucht und bei drastischer
Verschlechterung euthanisiert.
Abbildung 2-13: Überlebensraten und Verlauf des Körpergewichtes von BALB/cJ Mäusen nach oraler Infektion mit LmoEGDe, LmoEGDe-lux und LmoEGDe-mur-lux. Je zehn BALB/cJ Weibchen wurden im Alter von zehn Wochen mit 1x1010 CFU LmoEGDe, LmoEGDe-lux oder LmoEGDe-mur-lux oral infiziert. (A) Im Verlauf des Körpergewichtes ergaben sich keine signifikanten Unterschiede. Es sind die Mittelwerte der Körpergewichte in Prozent und deren Standardfehler (SEM) dargestellt. Der Gewichtsverlust an Tag 0 resultiert durch Futterentzug am vorangegangenen Tag als Voraussetzung für die orale Infektion. Nach Inokulieren der Listerien direkt in den Magen hatten die Mäuse wieder uneingeschränkten Futterzugang. (B) Die Analyse der Überlebensraten von BALB/cJ Mäusen zeigte nach der Infektion mit LmoEGDe eine Überlebensrate von 30%, bei LmoEGDe-lux 40% und bei LmoEGDe-mur-lux 26,7%. Insgesamt zeigten sich keine signifikanten Unterschiede in der Überlebensrate (Logrank-Test, p=0,5256).

Ergebnisse
57
Die Resultate der Körpergewichtsänderung sind in Abbildung 2-13 A dargestellt und
zeigten keine signifikanten Unterschiede. Die Gewichtsabnahme der BALB/cJ Mäuse
infiziert mit LmoEGDe bzw. LmoEGDe-mur-lux erfolgte bis zum Tag vier nach der
Infektion und anschließend ab Tag fünf war die Gewichtszunahme bis zum Ende des
Beobachtungszeitraums zu beobachten. BALB/cJ Mäuse infiziert mit LmoEGDe-lux
zeigten dagegen einen Gewichtsverlust bis zum Tag sieben nach der Infektion
gefolgt von einer kontinuierlichen Gewichtszunahme bis zum Versuchsende. Die
Infektion mit LmoEGDe resultierte in einer Überlebensrate von 30%. Bis zum Tag
fünf nach der Infektion verstarb die Hälfte der mit LmoEGDe infizierten BALB/cJ. Die
Mäuse infiziert mit LmoEGDe-lux zeigten eine Überlebensrate von 40% und von den
LmoEGDe-mur-lux infizierten BALB/cJ überlebten 26,7% (Abbildung 2-13 B). Die
Auswertung der Überlebensrate ergab keine signifikanten Unterschiede (p=0,5256,
Logrank-Test).
Parallel zur Dokumentation von Körpergewicht und Überlebensrate erfolgte täglich
das in vivo Imaging auf die emittierten Signale der biolumineszenten Listerien-
stämme LmoEGDe-lux und LmoEGDe-mur-lux in den infizierten Mäuse mit Hilfe des
Xenogen IVIS (Abbildung 2-14). Als repräsentatives Tier jede Gruppe (n=10) wurde
ein Individuum über den gesamten Versuchszeitraum dargestellt. Eine Stunde nach
der oralen Infektion konnten inokulierte Listerien im Magen-Darm-Trakt detektiert
werden. Am Tag eins und zwei nach der Infektion war aufgrund der Ausbreitung der
Bakterien über den Darm, durch deren partielle Ausscheidung oder durch die Elimi-
nation über die Immunabwehr keine bzw. nur eine geringe Detektion von LmoEGDe-
lux und LmoEGDe-mur-lux möglich. Deutliche Unterschiede in der Messung der
Biolumineszenzsignale von LmoEGDe-lux und LmoEGDe-mur-lux waren im Laufe
des Versuches erkennbar. BALB/cJ Mäuse infiziert mit LmoEGDe-lux zeigten über
die Dauer des Versuches geringere Signale als die mit LmoEGDe-mur-lux infizierten
Tiere, ein Resultat, welches die Ergebnisse von Monk et al. bestätigt (Monk et al.,
2010). Biolumineszenzsignale in der Leber konnten ab Tag fünf nach der Infektion in
LmoEGDe-mur-lux Tieren detektiert werden. Das in Abbildung 2-14 dargestellte Tier
verstarb innerhalb von sechs Tagen. Eine Akkumulation in den inneren Organen von
LmoEGDe-lux infizierten Mäusen wurde nur selten beobachtet. Jedoch führt auf-
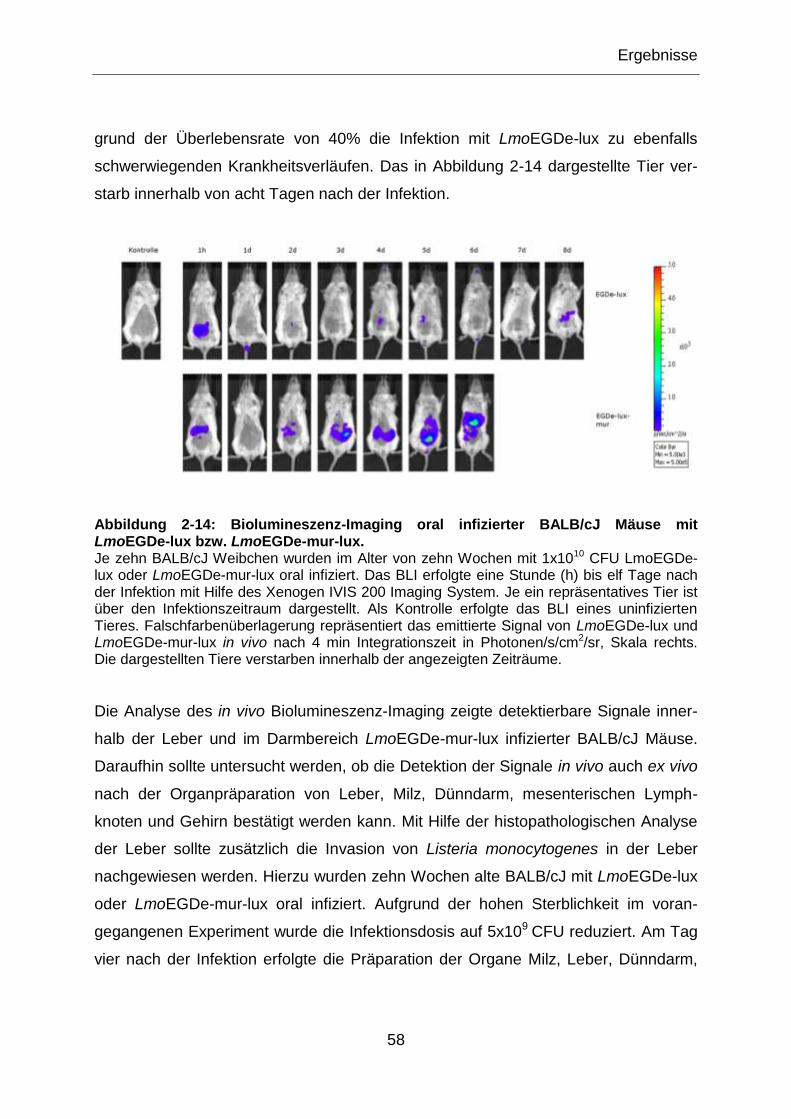
Ergebnisse
58
grund der Überlebensrate von 40% die Infektion mit LmoEGDe-lux zu ebenfalls
schwerwiegenden Krankheitsverläufen. Das in Abbildung 2-14 dargestellte Tier ver-
starb innerhalb von acht Tagen nach der Infektion.
Abbildung 2-14: Biolumineszenz-Imaging oral infizierter BALB/cJ Mäuse mit LmoEGDe-lux bzw. LmoEGDe-mur-lux. Je zehn BALB/cJ Weibchen wurden im Alter von zehn Wochen mit 1x1010 CFU LmoEGDe-lux oder LmoEGDe-mur-lux oral infiziert. Das BLI erfolgte eine Stunde (h) bis elf Tage nach der Infektion mit Hilfe des Xenogen IVIS 200 Imaging System. Je ein repräsentatives Tier ist über den Infektionszeitraum dargestellt. Als Kontrolle erfolgte das BLI eines uninfizierten Tieres. Falschfarbenüberlagerung repräsentiert das emittierte Signal von LmoEGDe-lux und LmoEGDe-mur-lux in vivo nach 4 min Integrationszeit in Photonen/s/cm2/sr, Skala rechts. Die dargestellten Tiere verstarben innerhalb der angezeigten Zeiträume.
Die Analyse des in vivo Biolumineszenz-Imaging zeigte detektierbare Signale inner-
halb der Leber und im Darmbereich LmoEGDe-mur-lux infizierter BALB/cJ Mäuse.
Daraufhin sollte untersucht werden, ob die Detektion der Signale in vivo auch ex vivo
nach der Organpräparation von Leber, Milz, Dünndarm, mesenterischen Lymph-
knoten und Gehirn bestätigt werden kann. Mit Hilfe der histopathologischen Analyse
der Leber sollte zusätzlich die Invasion von Listeria monocytogenes in der Leber
nachgewiesen werden. Hierzu wurden zehn Wochen alte BALB/cJ mit LmoEGDe-lux
oder LmoEGDe-mur-lux oral infiziert. Aufgrund der hohen Sterblichkeit im voran-
gegangenen Experiment wurde die Infektionsdosis auf 5x109 CFU reduziert. Am Tag
vier nach der Infektion erfolgte die Präparation der Organe Milz, Leber, Dünndarm,
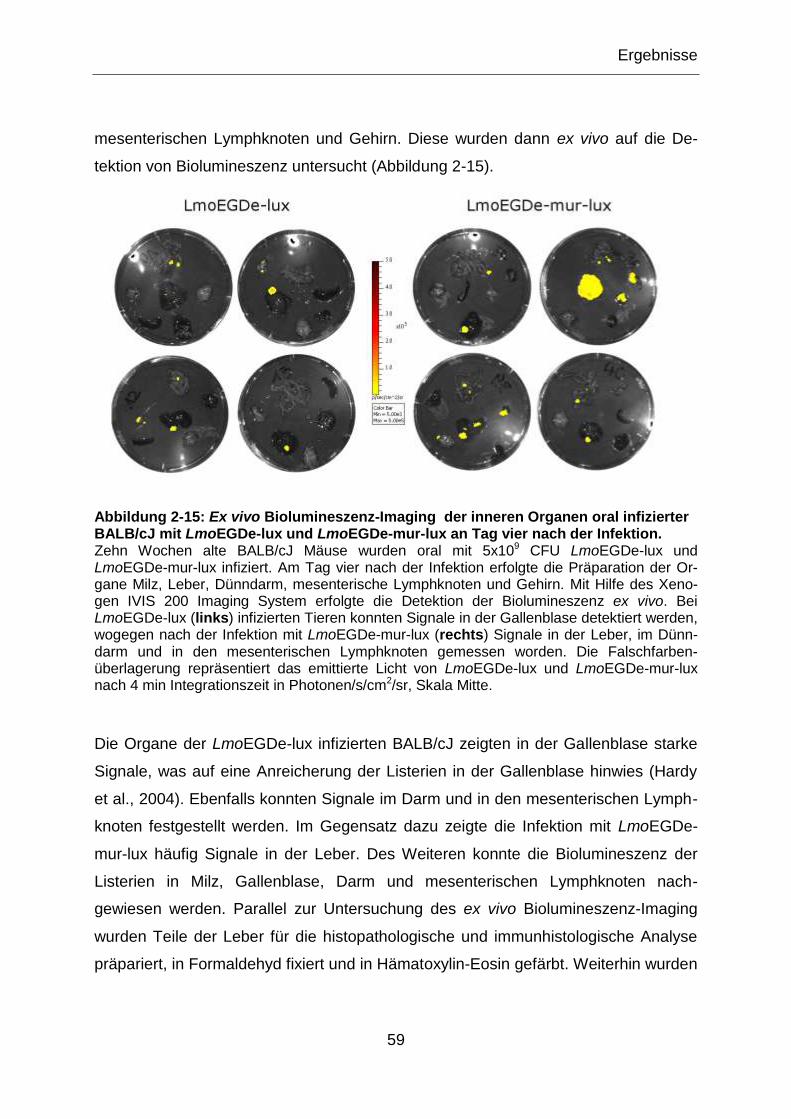
Ergebnisse
59
mesenterischen Lymphknoten und Gehirn. Diese wurden dann ex vivo auf die De-
tektion von Biolumineszenz untersucht (Abbildung 2-15).
Abbildung 2-15: Ex vivo Biolumineszenz-Imaging der inneren Organen oral infizierter BALB/cJ mit LmoEGDe-lux und LmoEGDe-mur-lux an Tag vier nach der Infektion. Zehn Wochen alte BALB/cJ Mäuse wurden oral mit 5x109 CFU LmoEGDe-lux und LmoEGDe-mur-lux infiziert. Am Tag vier nach der Infektion erfolgte die Präparation der Or-gane Milz, Leber, Dünndarm, mesenterische Lymphknoten und Gehirn. Mit Hilfe des Xeno-gen IVIS 200 Imaging System erfolgte die Detektion der Biolumineszenz ex vivo. Bei LmoEGDe-lux (links) infizierten Tieren konnten Signale in der Gallenblase detektiert werden, wogegen nach der Infektion mit LmoEGDe-mur-lux (rechts) Signale in der Leber, im Dünn-darm und in den mesenterischen Lymphknoten gemessen worden. Die Falschfarben-überlagerung repräsentiert das emittierte Licht von LmoEGDe-lux und LmoEGDe-mur-lux nach 4 min Integrationszeit in Photonen/s/cm2/sr, Skala Mitte.
Die Organe der LmoEGDe-lux infizierten BALB/cJ zeigten in der Gallenblase starke
Signale, was auf eine Anreicherung der Listerien in der Gallenblase hinwies (Hardy
et al., 2004). Ebenfalls konnten Signale im Darm und in den mesenterischen Lymph-
knoten festgestellt werden. Im Gegensatz dazu zeigte die Infektion mit LmoEGDe-
mur-lux häufig Signale in der Leber. Des Weiteren konnte die Biolumineszenz der
Listerien in Milz, Gallenblase, Darm und mesenterischen Lymphknoten nach-
gewiesen werden. Parallel zur Untersuchung des ex vivo Biolumineszenz-Imaging
wurden Teile der Leber für die histopathologische und immunhistologische Analyse
präpariert, in Formaldehyd fixiert und in Hämatoxylin-Eosin gefärbt. Weiterhin wurden
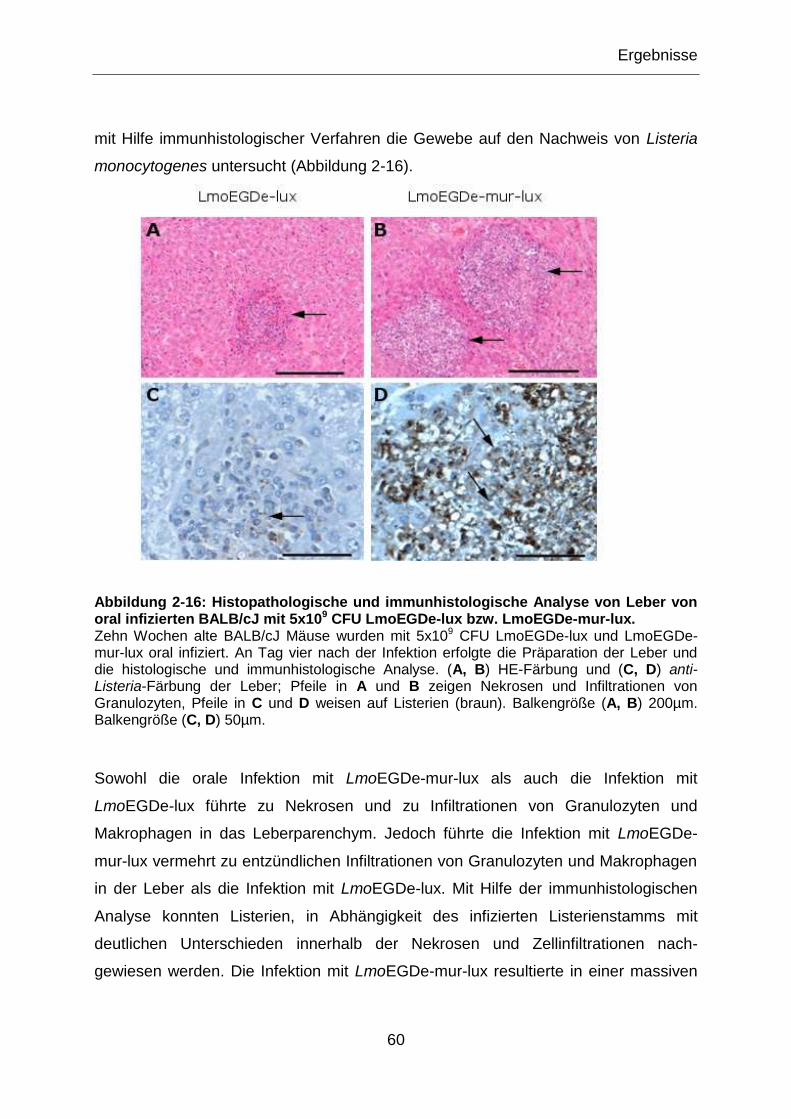
Ergebnisse
60
mit Hilfe immunhistologischer Verfahren die Gewebe auf den Nachweis von Listeria
monocytogenes untersucht (Abbildung 2-16).
Abbildung 2-16: Histopathologische und immunhistologische Analyse von Leber von oral infizierten BALB/cJ mit 5x109 CFU LmoEGDe-lux bzw. LmoEGDe-mur-lux. Zehn Wochen alte BALB/cJ Mäuse wurden mit 5x109 CFU LmoEGDe-lux und LmoEGDe-mur-lux oral infiziert. An Tag vier nach der Infektion erfolgte die Präparation der Leber und die histologische und immunhistologische Analyse. (A, B) HE-Färbung und (C, D) anti-Listeria-Färbung der Leber; Pfeile in A und B zeigen Nekrosen und Infiltrationen von Granulozyten, Pfeile in C und D weisen auf Listerien (braun). Balkengröße (A, B) 200µm. Balkengröße (C, D) 50µm.
Sowohl die orale Infektion mit LmoEGDe-mur-lux als auch die Infektion mit
LmoEGDe-lux führte zu Nekrosen und zu Infiltrationen von Granulozyten und
Makrophagen in das Leberparenchym. Jedoch führte die Infektion mit LmoEGDe-
mur-lux vermehrt zu entzündlichen Infiltrationen von Granulozyten und Makrophagen
in der Leber als die Infektion mit LmoEGDe-lux. Mit Hilfe der immunhistologischen
Analyse konnten Listerien, in Abhängigkeit des infizierten Listerienstamms mit
deutlichen Unterschieden innerhalb der Nekrosen und Zellinfiltrationen nach-
gewiesen werden. Die Infektion mit LmoEGDe-mur-lux resultierte in einer massiven

Ergebnisse
61
Invasion von Bakterien innerhalb des entzündlichen Gewebes, wogegen die Invasion
des Gewebes von Listerien in LmoEGDe-lux infizierten Mäusen deutlich geringer
ausfiel.
2.3.3 Vergleich der Wirtsresistenz in ausgewählten Mausinzuchtstämmen
nach der Infektion mit LmoEGDe-lux und LmoEGDe-mur-lux
Frühere Analysen zeigten eine genetische Regulation der angeborenen Resistenz
von Mäusen nach der intravenösen oder intraperitonealen Infektion mit Listeria
monocytogenes über den Hc (hämolytisches Komplement) Locus (kontrolliert die
Konzentration des Komplementfaktors C5) der sich auf dem Mauschromosom 2
kartiert (Cheers und McKenzie, 1978; Czuprynski et al., 1985; Gervais et al, 1984;
Skamene et al., 1979). Ein Mangel an Komplementfaktor C5 resultiert in einer er-
höhten Suszeptibilität von A/J Mäusen auf die Infektion mit Listeria monocytogenes
(Gervais et al, 1984; Ooi und Colten, 1979). Ebenso zeigten sich BALB/cJ und
C3HeB/FeJ Mäuse empfänglich für eine Listerieninfektion (Archinal und Wilder,
1988; Cheers und Stanley, 1988; Mainou-Fowler, 1988; Munder et al., 2005). Im Ge-
gensatz dazu konnte gezeigt werden, dass C57BL/6J Mäuse einen resistenten Phä-
notyp in Bezug auf die Infektion mit Listeria monocytogenes aufwiesen (Cheers et al.,
1978; Cheers und McKenzie, 1978; Czuprynski et al., 2002; Czuprynski et al., 2003;
Iizawa et al., 1994).
Wie im vorangegangenen Versuch gezeigt werden konnte, führte aufgrund der An-
passung des Internalin A an seinen murinen Rezeptor die orale Infektion von
BALB/cJ Mäusen mit LmoEGDe-mur-lux zu einem virulenteren Krankheitsgeschehen
mit Infiltrationen von Listeria monocytogenes in der Leber im Vergleich zu der
Infektion mit LmoEGDe-lux. Zur Beurteilung der Virulenz des murinisierten
Listerienstamms LmoEGDe-mur-lux und für den Vergleich der Wirtsresistenz
ausgewählter Mausinzuchtstämme erfolgte die Infektion von BALB/cJ, A/JOlaHsd,
C3HeB/FeJ und C57BL/6J Mäusen mit LmoEGDe-mur-lux im Vergleich zum
isogenen Wildtyp LmoEGDe-lux. Je zehn Weibchen von C57BL/6J, A/JOlaHsd,
BALB/cJ und C3HeB/FeJ wurden im Alter von zehn Wochen mit einer Infektionsdosis
von 5x109 CFU LmoEGDe-mur-lux bzw. LmoEGDe-lux oral infiziert. Über den
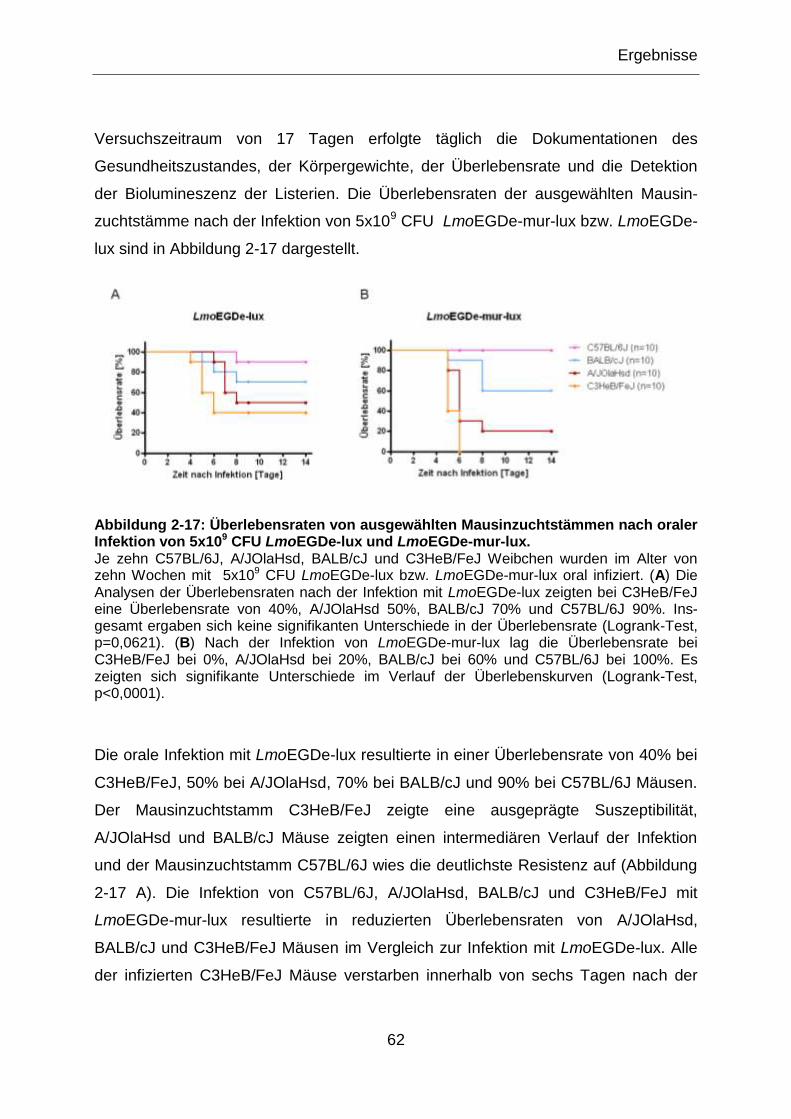
Ergebnisse
62
Versuchszeitraum von 17 Tagen erfolgte täglich die Dokumentationen des
Gesundheitszustandes, der Körpergewichte, der Überlebensrate und die Detektion
der Biolumineszenz der Listerien. Die Überlebensraten der ausgewählten Mausin-
zuchtstämme nach der Infektion von 5x109 CFU LmoEGDe-mur-lux bzw. LmoEGDe-
lux sind in Abbildung 2-17 dargestellt.
Abbildung 2-17: Überlebensraten von ausgewählten Mausinzuchtstämmen nach oraler Infektion von 5x109 CFU LmoEGDe-lux und LmoEGDe-mur-lux. Je zehn C57BL/6J, A/JOlaHsd, BALB/cJ und C3HeB/FeJ Weibchen wurden im Alter von zehn Wochen mit 5x109 CFU LmoEGDe-lux bzw. LmoEGDe-mur-lux oral infiziert. (A) Die Analysen der Überlebensraten nach der Infektion mit LmoEGDe-lux zeigten bei C3HeB/FeJ eine Überlebensrate von 40%, A/JOlaHsd 50%, BALB/cJ 70% und C57BL/6J 90%. Ins-gesamt ergaben sich keine signifikanten Unterschiede in der Überlebensrate (Logrank-Test, p=0,0621). (B) Nach der Infektion von LmoEGDe-mur-lux lag die Überlebensrate bei C3HeB/FeJ bei 0%, A/JOlaHsd bei 20%, BALB/cJ bei 60% und C57BL/6J bei 100%. Es zeigten sich signifikante Unterschiede im Verlauf der Überlebenskurven (Logrank-Test, p<0,0001).
Die orale Infektion mit LmoEGDe-lux resultierte in einer Überlebensrate von 40% bei
C3HeB/FeJ, 50% bei A/JOlaHsd, 70% bei BALB/cJ und 90% bei C57BL/6J Mäusen.
Der Mausinzuchtstamm C3HeB/FeJ zeigte eine ausgeprägte Suszeptibilität,
A/JOlaHsd und BALB/cJ Mäuse zeigten einen intermediären Verlauf der Infektion
und der Mausinzuchtstamm C57BL/6J wies die deutlichste Resistenz auf (Abbildung
2-17 A). Die Infektion von C57BL/6J, A/JOlaHsd, BALB/cJ und C3HeB/FeJ mit
LmoEGDe-mur-lux resultierte in reduzierten Überlebensraten von A/JOlaHsd,
BALB/cJ und C3HeB/FeJ Mäusen im Vergleich zur Infektion mit LmoEGDe-lux. Alle
der infizierten C3HeB/FeJ Mäuse verstarben innerhalb von sechs Tagen nach der

Ergebnisse
63
Infektion. Die A/JOlaHsd Mäuse wiesen eine Überlebensrate von 20% auf, die
BALB/cJ zeigten eine Überlebensrate von 60% und 100% der C57BL/6J überlebten.
Die C3HeB/FeJ erwiesen sich als empfindlichster Stamm, A/JOlaHsd und BALB/cJ
zeigten eine intermediäre Suszeptibilität und als Mausstamm mit der aus-
geprägtesten Resistenz erwies sich C57BL/6J (Abbildung 2-17 B). Die deutliche
Reduzierung der Überlebensraten von A/JOlaHsd, BALB/cJ und C3HeB/FeJ Mäusen
nach der Infektion mit dem murinisierten Listerienstamm LmoEGDe-mur-lux im
Vergleich zu LmoEGDe-lux ist das Resultat einer gesteigerten Virulenz von
LmoEGDe-mur-lux aufgrund der Murinisierung des Internalin A.
Parallel zur Bestimmung der Überlebensrate erfolgte die Dokumentation des Körper-
gewichts der mit LmoEGDe-lux bzw. LmoEGDe-mur-lux infizierten C57BL/6J,
A/JOlaHsd, BALB/cJ und C3HeB/FeJ Mäuse über den Versuchszeitraum von 14
Tagen (Abbildung 2-18). Die Analyse des Verlaufs des Körpergewichtes der
C3HeB/FeJ Mäusen (Abbildung 2-18 A) zeigte nach der Infektion mit LmoEGDe-lux
und LmoEGDe-mur-lux keinen signifikanten Unterschied. Die C3HeB/FeJ Mäuse
reagierten auf die Infektion mit LmoEGDe-lux und LmoEGDe-mur-lux gleichermaßen
mit Gewichtsverlust. Bis zum Tag sechs waren alle mit LmoEGDe-mur-lux infizierten
C3HeB/FeJ verstorben. Die mit LmoEGDe-lux infizierten Tiere zeigten eine Ge-
wichtsabnahme bis zum Tag sechs nach der Infektion gefolgt von einer kontinuier-
lichen Gewichtszunahme bis zum Versuchsende. Im Verlauf der Änderung des Kör-
pergewichtes von A/JOlaHsd Mäusen ergaben sich signifikante Unterscheide an Tag
drei, vier und fünf nach der Infektion mit LmoEGDe-lux und LmoEGDe-mur-lux
(Abbildung 2-18 B). Die Gewichtsabnahme der mit LmoEGDe-lux bzw. LmoEGDe-
mur-lux infizierten A/JOlaHsd erfolgte jeweils bis zum Tag sechs post Infektion. Im
Fall der LmoEGDe-lux infizierten Tiere konnte ab Tag sechs nach der Infektion eine
kontinuierliche Gewichtszunahme bis zum Versuchsende beobachtet werden. Die
LmoEGDe-mur-lux infizierten A/JOlaHsd Mäuse verzeichneten ab Tag sechs eine
Gewichtszunahme bis zum Tag elf nach der Infektion, gefolgt von einer erneuten
Gewichtsabnahme bis zum Tag 13. Ab Tag 13 nach der Infektion bis zum Ende des
Versuchszeitraums konnte eine Gewichtzunahme beobachtet werden. Im Verlauf der
Änderung des Körpergewichtes von BALB/cJ Mäusen ergab sich ein signifikanter
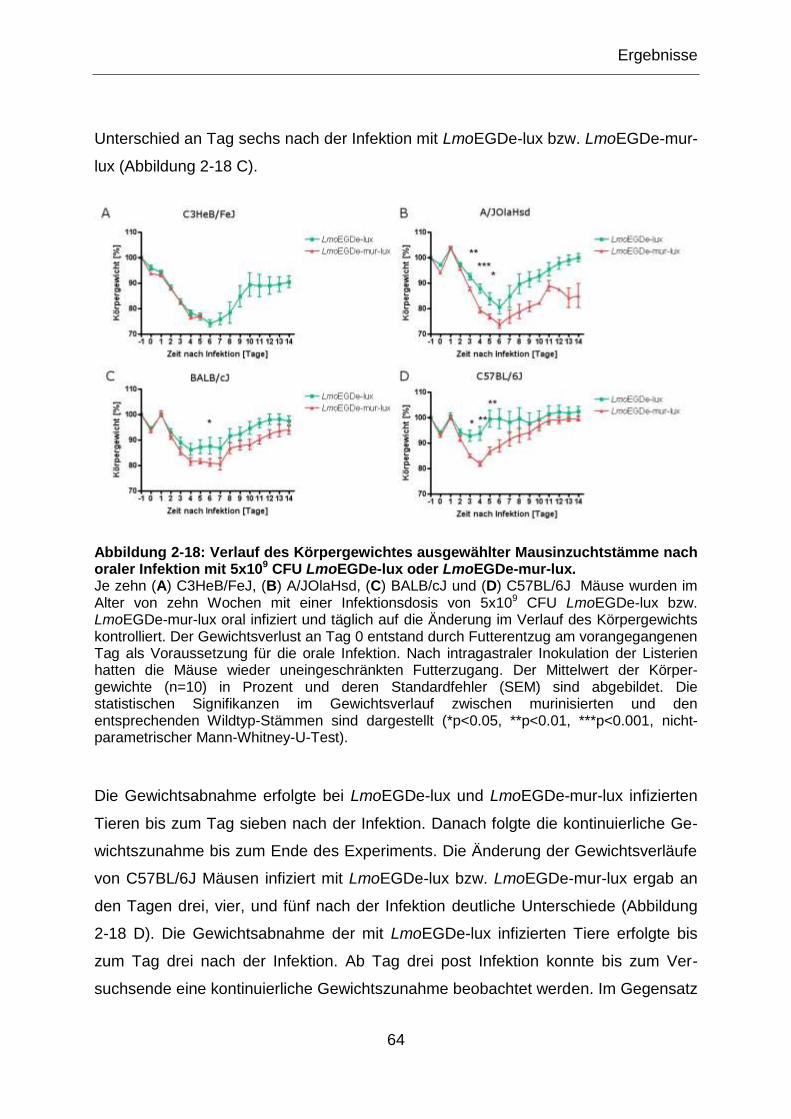
Ergebnisse
64
Unterschied an Tag sechs nach der Infektion mit LmoEGDe-lux bzw. LmoEGDe-mur-
lux (Abbildung 2-18 C).
Abbildung 2-18: Verlauf des Körpergewichtes ausgewählter Mausinzuchtstämme nach oraler Infektion mit 5x109 CFU LmoEGDe-lux oder LmoEGDe-mur-lux. Je zehn (A) C3HeB/FeJ, (B) A/JOlaHsd, (C) BALB/cJ und (D) C57BL/6J Mäuse wurden im Alter von zehn Wochen mit einer Infektionsdosis von 5x109 CFU LmoEGDe-lux bzw. LmoEGDe-mur-lux oral infiziert und täglich auf die Änderung im Verlauf des Körpergewichts kontrolliert. Der Gewichtsverlust an Tag 0 entstand durch Futterentzug am vorangegangenen Tag als Voraussetzung für die orale Infektion. Nach intragastraler Inokulation der Listerien hatten die Mäuse wieder uneingeschränkten Futterzugang. Der Mittelwert der Körper-gewichte (n=10) in Prozent und deren Standardfehler (SEM) sind abgebildet. Die statistischen Signifikanzen im Gewichtsverlauf zwischen murinisierten und den entsprechenden Wildtyp-Stämmen sind dargestellt (*p<0.05, **p<0.01, ***p<0.001, nicht-parametrischer Mann-Whitney-U-Test).
Die Gewichtsabnahme erfolgte bei LmoEGDe-lux und LmoEGDe-mur-lux infizierten
Tieren bis zum Tag sieben nach der Infektion. Danach folgte die kontinuierliche Ge-
wichtszunahme bis zum Ende des Experiments. Die Änderung der Gewichtsverläufe
von C57BL/6J Mäusen infiziert mit LmoEGDe-lux bzw. LmoEGDe-mur-lux ergab an
den Tagen drei, vier, und fünf nach der Infektion deutliche Unterschiede (Abbildung
2-18 D). Die Gewichtsabnahme der mit LmoEGDe-lux infizierten Tiere erfolgte bis
zum Tag drei nach der Infektion. Ab Tag drei post Infektion konnte bis zum Ver-
suchsende eine kontinuierliche Gewichtszunahme beobachtet werden. Im Gegensatz

Ergebnisse
65
zu den LmoEGDe-lux infizierten C57BL/6J erfolgte bei den LmoEGDe-mur-lux infi-
zierten C57BL/6J eine Gewichtsabnahme bis zum Tag vier nach der Infektion, ge-
folgt von der kontinuierlichen Gewichtszunahme bis zum Abschluss des Experi-
ments. Die orale Infektion von C57BL/6J, A/JOlaHsd, BALB/cJ Mäusen mit
LmoEGDe-mur-lux resultierte somit in einer größeren Änderung im Verlauf des Kör-
pergewichtes im Vergleich zur Infektion mit LmoEGDe-lux.
Über den Versuchszeitraum von 14 Tagen erfolgte das tägliche Biolumineszenz-
Imaging aller infizierten C57BL/6J, A/JOlaHsd, BALB/cJ und C3HeB/FeJ Mäuse zur
Analyse der Dissemination der Listerien in vivo. Eine repräsentative Gruppe von fünf
Tieren jedes Mausinzuchtstammes ist in Abbildung 2-19 (infiziert mit LmoEGDe-lux)
und Abbildung 2-20 (infiziert mit LmoEGDe-mur-lux) dargestellt. Eine Stunde nach
der intragastralen Inokulation von 5x109 CFU LmoEGDe-lux konnten Listerien
innerhalb des Magen-Darm-Traktes detektiert werden (Abbildung 2-19). Im Verlauf
der Infektion mit LmoEGDe-lux konnten die Listerien an Tag eins nach der Infektion
nur in C57BL/6J nachgewiesen werden. Bis zum Versuchsende war eine Detektion
von Listerien in C57BL/6J nicht mehr möglich. In den A/JOlaHsd, BALB/cJ und
C3HeB/FeJ Mäuse war eine Detektion der Listerien an Tag eins bis zum Tag zwei
nach der Infektion nicht messbar. Erst ab Tag drei nach der Infektion konnten
Listerien in der Gallenblase und im Abdominalbereich von einzelnen C3HeB/FeJ,
BALB/cJ und A/JOlaHsd Mäusen detektiert werden. Ab Tag vier nach der Infektion
konnte eine Zunahme des Biolumineszenzsignals im Darmbereich von A/JOlaHsd,
BALB/cJ und C3HeB/FeJ Mäusen beobachtet werden. Vereinzelt war, wie im Fall
von BALB/cJ und C3HeB/FeJ Mäusen, die Detektion von Listerien in der Leber mög-
lich. Ab Tag sechs nach der Infektion erfolgte eine kontinuierliche Reduzierung der
Detektion der Biolumineszenz in vivo, welche ab Tag acht nach der Infektion nicht
mehr nachgewiesen werden konnte.
Im Fall der intragastralen Inokulation von 5x109 CFU LmoEGDe-mur-lux waren
ebenfalls eine Stunde nach der Infektion Listerien innerhalb des Magen-Darm-
Traktes detektierbar (Abbildung 2-20). An den Tagen eins und zwei nach der Infek-
tion war die Detektion des Signals aufgrund der Dissemination der Bakterien über
den Darm und deren Ausscheidung reduziert. Am Tag zwei nach Infektion konnten

Ergebnisse
66
Listerien im Abdominalbereich von C3HeB/FeJ detektiert werden. Ab Tag drei nach
der Infektion waren Biolumineszenzsignale im abdominalen Bereich von BALB/cJ
und einzelnen A/JOlaHsd Mäusen messbar. Ein Anstieg der Biolumineszenz im
Darmbereich der A/JOlaHsd, BALB/cJ und C3HeB/FeJ Mäusen konnte ab Tag vier
nach der Infektion beobachtet werden. Zusätzlich konnte in allen C3HeB/FeJ
Mäusen sowie in je einer BALB/cJ und A/JOlaHsd Maus die Biolumineszenz der
Listerien innerhalb der Leber nachgewiesen werden. Bis zum Tag fünf nach der
Infektion konnte in BALB/cJ und A/JOlaHsd Mäusen die Persistenz der
Biolumineszenz innerhalb des abdominalen Bereiches festgestellt werden. Ab Tag
sechs nach der Infektion erfolgte kontinuierlich die Reduzierung der Detektion der
Biolumineszenz in vivo. Ab Tag acht nach der Infektion waren keine
Biolumineszenzsignale mehr nachweisbar. Im Vergleich der LmoEGDe-lux und
LmoEGDe-mur-lux infizierten Tieren waren Unterschiede in der Stärke der
Biolumineszenz erkennbar. Die Biolumineszenz war in LmoEGDe-mur-lux infizierten
Mäusen ausgeprägter nachweisbar. Auch sind Unterschiede in Lokalisation der
Signale erkennbar. Die Mausinzuchtstämme C3HeB/FeJ, A/JOlaHsd und BALB/cJ,
infiziert mit LmoEGDe-lux, zeigten eine Akkumulation von Signalen in der
Gallenblase und im Darmbereich. Bei zwei C3HeB/FeJ Tieren konnten
Biolumineszenzsignale auch in der Leber detektiert werden. Im Gegensatz dazu
konnte nach der Infektion mit LmoEGDe-mur-lux in der Leber und im Darmbereich
einer Vielzahl der Individuen der Mausinzuchtstämme C3HeB/FeJ, A/JOlaHsd und
BALB/cJ Biolumineszenz nachgewiesen werden. C57BL/6J Mäuse erwiesen sich als
sehr resistent sowohl bei der Infektion mit LmoEGDe-lux als auch LmoEGDe-mur-
lux. Jedoch sind C57BL/6J Mäuse aufgrund ihrer pigmentierten Haut und der
schwarzen Fellfarbe für die Messung der Biolumineszenz ungeeignet.
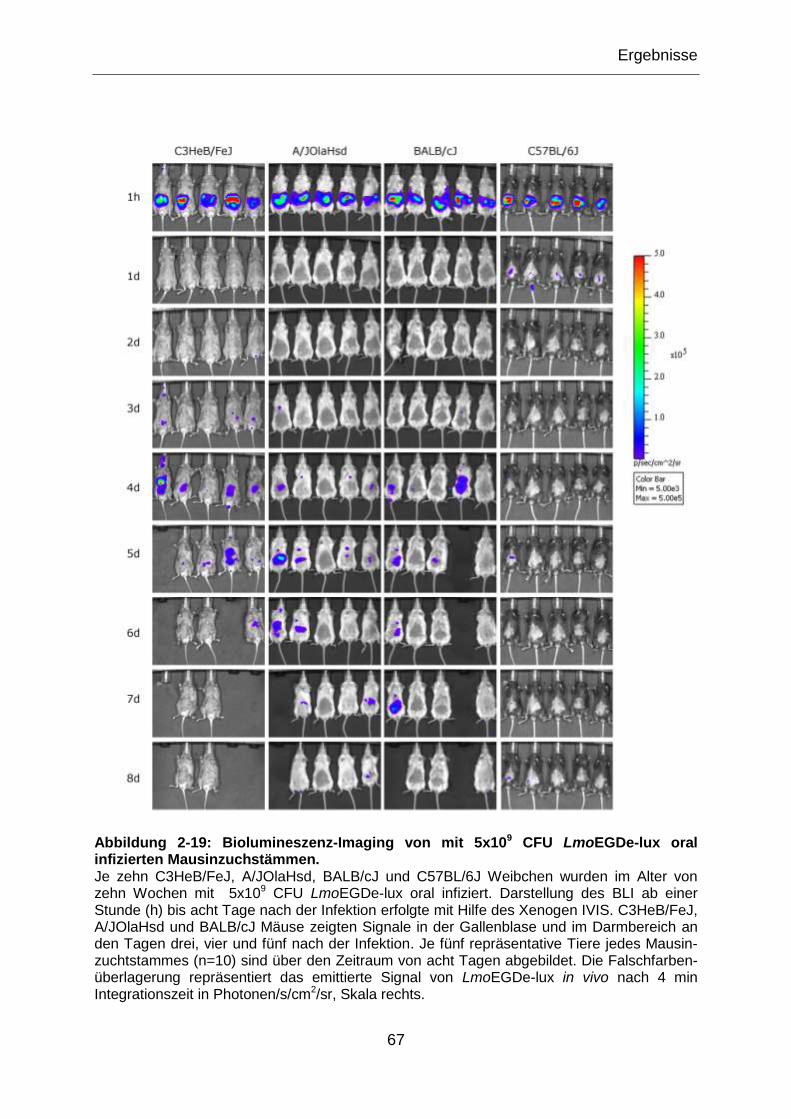
Ergebnisse
67
Abbildung 2-19: Biolumineszenz-Imaging von mit 5x109 CFU LmoEGDe-lux oral infizierten Mausinzuchstämmen. Je zehn C3HeB/FeJ, A/JOlaHsd, BALB/cJ und C57BL/6J Weibchen wurden im Alter von zehn Wochen mit 5x109 CFU LmoEGDe-lux oral infiziert. Darstellung des BLI ab einer Stunde (h) bis acht Tage nach der Infektion erfolgte mit Hilfe des Xenogen IVIS. C3HeB/FeJ, A/JOlaHsd und BALB/cJ Mäuse zeigten Signale in der Gallenblase und im Darmbereich an den Tagen drei, vier und fünf nach der Infektion. Je fünf repräsentative Tiere jedes Mausin-zuchtstammes (n=10) sind über den Zeitraum von acht Tagen abgebildet. Die Falschfarben-überlagerung repräsentiert das emittierte Signal von LmoEGDe-lux in vivo nach 4 min Integrationszeit in Photonen/s/cm2/sr, Skala rechts.
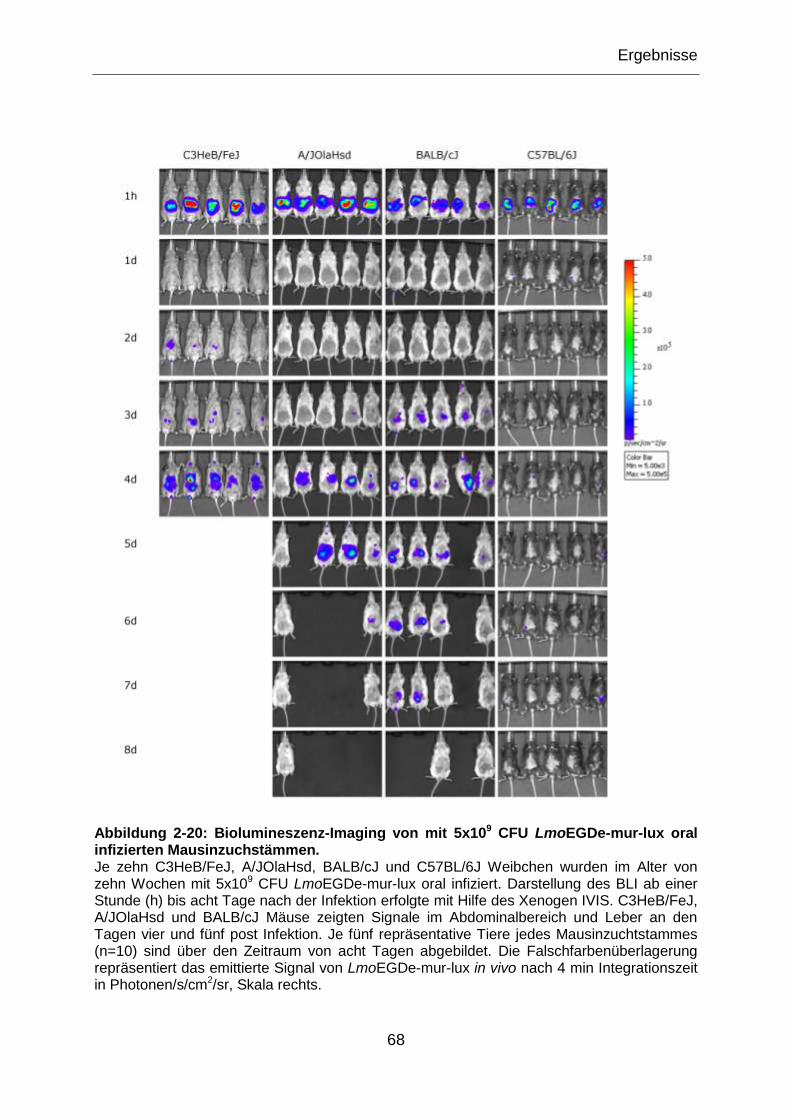
Ergebnisse
68
Abbildung 2-20: Biolumineszenz-Imaging von mit 5x109 CFU LmoEGDe-mur-lux oral infizierten Mausinzuchstämmen. Je zehn C3HeB/FeJ, A/JOlaHsd, BALB/cJ und C57BL/6J Weibchen wurden im Alter von zehn Wochen mit 5x109 CFU LmoEGDe-mur-lux oral infiziert. Darstellung des BLI ab einer Stunde (h) bis acht Tage nach der Infektion erfolgte mit Hilfe des Xenogen IVIS. C3HeB/FeJ, A/JOlaHsd und BALB/cJ Mäuse zeigten Signale im Abdominalbereich und Leber an den Tagen vier und fünf post Infektion. Je fünf repräsentative Tiere jedes Mausinzuchtstammes (n=10) sind über den Zeitraum von acht Tagen abgebildet. Die Falschfarbenüberlagerung repräsentiert das emittierte Signal von LmoEGDe-mur-lux in vivo nach 4 min Integrationszeit in Photonen/s/cm2/sr, Skala rechts.

Ergebnisse
69
2.3.4 Kinetik der in vivo Replikation von LmoEGDe-lux und LmoEGDe-lux-mur
in ausgewählten Mausinzuchtstämmen
Die Analyse der Überlebensrate sowie das Biolumineszenz-Imaging in ausgewählten
Mausinzuchtstämmen nach der Infektion von LmoEGDe-lux und LmoEGDe-mur-lux
zeigte Unterschiede in der Suszeptibilität von C3HeB/FeJ, A/JOlaHsd, BALB/cJ und
C57BL/6J Mäusen auf. Aufgrund dieser Beobachtung erfolgte die Untersuchung der
bakteriellen Keimlast in den inneren Organen Milz, Leber, Gallenblase, mesen-
terischen Lymphknoten, Dünndarm und Gehirn von C3HeB/FeJ, A/JOlaHsd,
BALB/cJ und C57BL/6J Mäusen nach oraler Infektion mit LmoEGDe-mur-lux im Ver-
gleich zum isogenen Wildtypstamm LmoEGDe-lux. An den Tagen eins, drei, fünf und
sieben nach der Infektion wurden jeweils acht Tiere für die Analyse verwendet.
Parallel erfolgte die histopathologische Untersuchung der Organe Milz, Leber und
Dünndarm. Ergänzend wurden die präparierten Organe ex vivo sowie täglich alle
infizierten Tiere in vivo auf die Detektierbarkeit von Biolumineszenzsignalen unter-
sucht. Weiter erfolgte täglich die Dokumentation der Änderungen im Verlauf des Kör-
pergewichtes.
2.3.4.1 In vivo Imaging von LmoEGDe-lux und LmoEGDe-mur-lux infizierten
Mausinzuchtstämmen
Je 40 Weibchen der Mausstämme C3HeB/FeJ, A/JOlaHsd, BALB/cJ und C57BL/6J
wurden im Alter von zehn Wochen mit einer Infektionsdosis von 5x109 CFU
LmoEGDe-lux bzw. LmoEGDe-mur-lux oral infiziert und über den Versuchszeitraum
von sieben Tagen erfolgte das tägliche Biolumineszenz-Imaging aller infizierten Tiere
mit Hilfe des IVIS 200 System (Abbildung 2-23). Eine Stunde nach der Infektion
waren beide Listerienstämme im Magen der Tiere detektierbar. Am Tag eins nach
der Infektion war keine Detektion der Signale in C3HeB/FeJ, A/JOlaHsd und
BALB/cJ Mäusen aufgrund der Dissemination der Bakterien über den Darm und
deren Ausscheidung möglich. Nur in C57BL/6J Mäusen konnte einen Tag nach der
Infektion Biolumineszenzsignale im Abdominalbereich nachgewiesen werden.
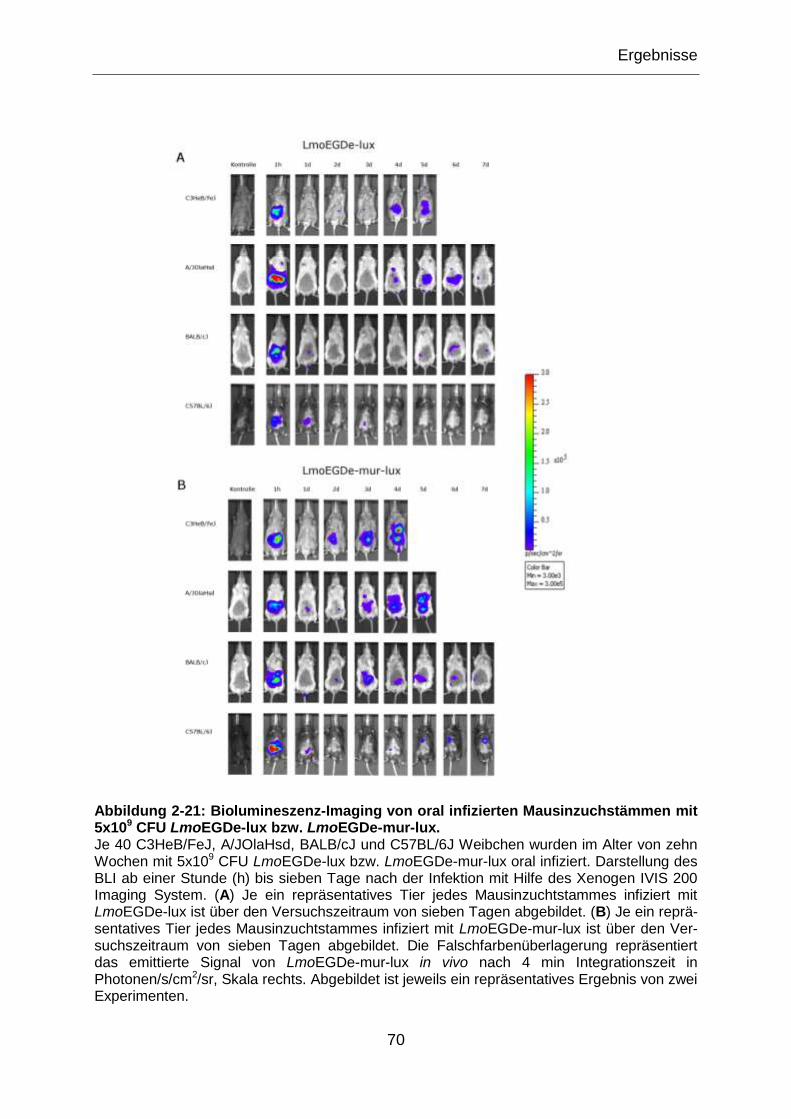
Ergebnisse
70
Abbildung 2-21: Biolumineszenz-Imaging von oral infizierten Mausinzuchstämmen mit 5x109 CFU LmoEGDe-lux bzw. LmoEGDe-mur-lux. Je 40 C3HeB/FeJ, A/JOlaHsd, BALB/cJ und C57BL/6J Weibchen wurden im Alter von zehn Wochen mit 5x109 CFU LmoEGDe-lux bzw. LmoEGDe-mur-lux oral infiziert. Darstellung des BLI ab einer Stunde (h) bis sieben Tage nach der Infektion mit Hilfe des Xenogen IVIS 200 Imaging System. (A) Je ein repräsentatives Tier jedes Mausinzuchtstammes infiziert mit LmoEGDe-lux ist über den Versuchszeitraum von sieben Tagen abgebildet. (B) Je ein reprä-sentatives Tier jedes Mausinzuchtstammes infiziert mit LmoEGDe-mur-lux ist über den Ver-suchszeitraum von sieben Tagen abgebildet. Die Falschfarbenüberlagerung repräsentiert das emittierte Signal von LmoEGDe-mur-lux in vivo nach 4 min Integrationszeit in Photonen/s/cm2/sr, Skala rechts. Abgebildet ist jeweils ein repräsentatives Ergebnis von zwei Experimenten.

Ergebnisse
71
Nach der Infektion mit LmoEGDe-lux wurden Signale im Darmbereich und in der
Gallenblase von C3HeB/FeJ und A/JOlaHsd Mäusen am Tag vier post Infektion
lokalisiert. Bei den infizierten C3HeB/FeJ Mäusen konnten zusätzlich noch Bio-
lumineszenzsignale in der Leber nachgewiesen werden. In infizierten C57BL/6J
Mäusen konnten keine spezifischen Signale in vivo detektiert werden. Auch bei
infizierten BALB/cJ Mäusen erfolgte die Detektion der biolumineszenten Listerien im
Darmbereich am Tag sechs nach der Infektion. Im Gegensatz zur Infektion mit
LmoEGDe-lux war die Detektion der Biolumineszenz in mit LmoEGDe-mur-lux
infizierten C3HeB/FeJ Mäusen im Abdominalbereich ab Tag zwei nach der Infektion
möglich. Ab Tag drei konnten die Listerien auch in Darmbereich der A/JOlaHsd und
BALB/cJ Mäusen nachgewiesen werden. Am Tag vier nach der Infektion erfolgte
zusätzlich die Detektion der Signale in der Leber von C3HeB/FeJ und A/JOlaHsd
Mäusen. In infizierten C57BL/6J Mäusen konnte die Detektion von biolumineszenten
Listerien innerhalb der Gallenblase ab Tag vier bis sieben nach der Infektion
beobachtet werden. Es wird deutlich, dass die Detektion der Biolumineszenzsignale
in LmoEGDe-lux infizierten im Vergleich zu LmoEGDe-mur-lux infizierten Tieren
verringert ist, was die Resultate von Monk et al. bestätigte (Monk et al., 2010).
Weiterhin wiesen Mausinzuchtstämme, welche empfindlich auf eine Infektion mit
Listerien reagieren (C3HeB/FeJ) ein ausgeprägteres Biolumineszenzsignal in der
Leber auf, während in resistenten Mausstämmen (C57BL/6J) eine Anreicherung in
der Gallenblase zu beobachten war.
Ergänzend zum in vivo Biolumineszenz-Imaging erfolgte die Messung der Verläufe
der Körpergewichte über den gesamten Zeitraum des Experimentes. In Abbildung 2-
22 sind die Änderungen im Gewichtsverlauf von LmoEGDe-lux infizierten im
Vergleich zu den LmoEGDe-mur-lux infizierten C57BL/6J, A/JOlaHsd, BALB/cJ und
C3HeB/FeJ Mäusen dargestellt. Die Analyse des Verlaufs des Körpergewichtes der
C3HeB/FeJ Mäuse (Abbildung 2-22 A) zeigte eine kontinuierliche Gewichtsabnahme
nach der Infektion mit LmoEGDe-lux und LmoEGDe-mur-lux bis zum Tag fünf nach
der Infektion. LmoEGDe-mur-lux infizierte C3HeB/FeJ verstarben bis zum Tag sechs
nach der Infektion. Am Tag drei nach der Infektion wurde ein signifikanter
Unterschied im Gewichtsverlauf LmoEGDe-lux und LmoEGDe-mur-lux infizierter
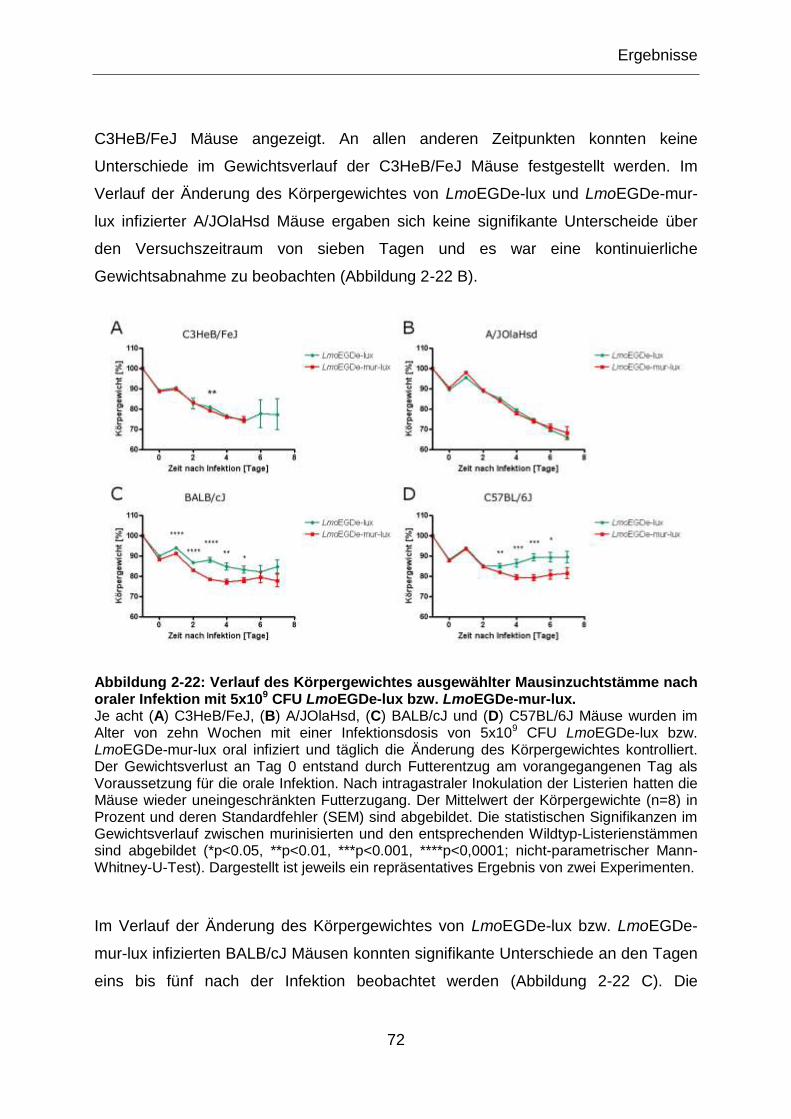
Ergebnisse
72
C3HeB/FeJ Mäuse angezeigt. An allen anderen Zeitpunkten konnten keine
Unterschiede im Gewichtsverlauf der C3HeB/FeJ Mäuse festgestellt werden. Im
Verlauf der Änderung des Körpergewichtes von LmoEGDe-lux und LmoEGDe-mur-
lux infizierter A/JOlaHsd Mäuse ergaben sich keine signifikante Unterscheide über
den Versuchszeitraum von sieben Tagen und es war eine kontinuierliche
Gewichtsabnahme zu beobachten (Abbildung 2-22 B).
Abbildung 2-22: Verlauf des Körpergewichtes ausgewählter Mausinzuchtstämme nach oraler Infektion mit 5x109 CFU LmoEGDe-lux bzw. LmoEGDe-mur-lux. Je acht (A) C3HeB/FeJ, (B) A/JOlaHsd, (C) BALB/cJ und (D) C57BL/6J Mäuse wurden im Alter von zehn Wochen mit einer Infektionsdosis von 5x109 CFU LmoEGDe-lux bzw. LmoEGDe-mur-lux oral infiziert und täglich die Änderung des Körpergewichtes kontrolliert. Der Gewichtsverlust an Tag 0 entstand durch Futterentzug am vorangegangenen Tag als Voraussetzung für die orale Infektion. Nach intragastraler Inokulation der Listerien hatten die Mäuse wieder uneingeschränkten Futterzugang. Der Mittelwert der Körpergewichte (n=8) in Prozent und deren Standardfehler (SEM) sind abgebildet. Die statistischen Signifikanzen im Gewichtsverlauf zwischen murinisierten und den entsprechenden Wildtyp-Listerienstämmen sind abgebildet (*p<0.05, **p<0.01, ***p<0.001, ****p<0,0001; nicht-parametrischer Mann-Whitney-U-Test). Dargestellt ist jeweils ein repräsentatives Ergebnis von zwei Experimenten.
Im Verlauf der Änderung des Körpergewichtes von LmoEGDe-lux bzw. LmoEGDe-
mur-lux infizierten BALB/cJ Mäusen konnten signifikante Unterschiede an den Tagen
eins bis fünf nach der Infektion beobachtet werden (Abbildung 2-22 C). Die

Ergebnisse
73
Gewichtsabnahme erfolgte bei LmoEGDe-mur-lux infizierten Tieren bis zum Tag
sieben nach der Infektion. Bei LmoEGDe-lux infizierten BALB/cJ Mäusen erfolgte die
Gewichtsabnahme bis zum Tag zwei nach der Infektion, gefolgt von einer Gewicht-
zunahme zum Tag drei und einer erneuten Gewichtsabnahme bis zum Tag sechs
nach der Infektion. Ab Tag sieben war eine erneute Gewichtszunahme zu beobach-
ten. Die Änderung der Gewichtsverläufe von C57BL/6J Mäusen infiziert mit
LmoEGDe-lux bzw. LmoEGDe-mur-lux zeigte an den Tagen drei, vier, fünf und
sechs nach der Infektion signifikante Unterschiede (Abbildung 2-22 D). Die
Gewichtsabnahme der mit LmoEGDe-lux infizierten Tiere erfolgte bis zum Tag zwei
nach der Infektion. Ab Tag drei post Infektion konnte bis zum Versuchsende eine
kontinuierliche Gewichtszunahme beobachtet werden. Im Gegensatz zu den
LmoEGDe-lux infizierten C57BL/6J erfolgte bei den LmoEGDe-mur-lux infizierten
C57BL/6J eine Gewichtsabnahme bis zum Tag fünf nach der Infektion, gefolgt von
der kontinuierlichen Gewichtszunahme bis zum Abschluss des Experiments.
2.3.4.2 Kinetik der bakteriellen Keimlast in den inneren Organen von LmoEGDe-lux
und LmoEGDe-lux-mur infizierten Mausinzuchtstämmen
Listeria monocytogenes besitzt die Fähigkeit die intestinale Barriere zu durchqueren.
Sie infizieren sowohl die intestinalen Epithelzellen als auch die M-Zellen der
Peyer´schen Plaques (Clark et al., 2000). Über das Blut- und Lymphsystem sowie
die mesenterialen Lymphknoten gelangen die Listerien zu Leber und Milz, den
Hauptorganen der bakteriellen Kolonisation und Replikation (Lecuit et al., 2001). Der
Nachweis detektierbarer Biolumineszenzsignale in infizierten C57BL/6J, A/JOlaHsd,
BALB/cJ und C3HeB/FeJ Mäusen in vivo weist auf die Akkumulation von Listerien in
den inneren Organen hin. Im weiteren Verlauf erfolgte die Detektion der
Biolumineszenzsignale auch ex vivo nach der Organpräparation von Leber, Milz,
Dünndarm, mesenterischen Lymphknoten und Gehirn. Zusätzlich wurde die
bakterielle Keimlast in den präparierten Organen Milz, Leber, Dünndarm, mLK,
Gallenblase und dem Gehirn bestimmt. Weiterhin sollte mit Hilfe der
histopathologischen Analyse der Leber die Invasion der Leber mit Listeria
monocytogenes beobachtet werden. An den Tagen eins, drei, fünf und sieben nach
der oralen Infektion mit 5x109 CFU LmoEGDe-lux und LmoEGDe-mur-lux wurden

Ergebnisse
74
jeweils acht Tiere von C57BL/6J, A/JOlaHsd, BALB/cJ und C3HeB/FeJ Mäusen für
die Analyse verwendet. Nach der Präparation der Organe wurden diese zunächst ex
vivo auf die Detektion von Biolumineszenzsignalen und somit auf das Vorkommen
von Listerien mit Hilfe des Xenogen IVIS untersucht (Abbildung 2-23). Erst am Tag
drei nach der oralen Infektion mit LmoEGDe-lux war ein deutliches Signal innerhalb
der mesenterischen Lymphknoten (mLK) infizierter A/JOlaHsd und C3HeB/FeJ
Mäuse sowie in den mLK LmoEGDe-mur-lux infizierter C57BL/6J, A/JOlaHsd,
BALB/cJ und C3HeB/FeJ Mäuse messbar. Während bei LmoEGDe-lux infizierten
C3HeB/FeJ Mäusen am Tag fünf nach der Infektion Signale in der Leber, Milz, mLK
sowie im Darm detektierbar waren, konnten bei den A/JOlaHsd und BALB/cJ
Mäusen Biolumineszenzsignale nur in mLK und in der Gallenblase nachgewiesen
werden. Die detektierbaren Biolumineszenzsignale am Tag sieben nach der Infektion
mit LmoEGDe-lux begrenzten sich bei A/JOlaHsd Mäusen auf die Gallenblase. Bei
den infizierten C57BL/6J Mäusen war keine Biolumineszenz innerhalb der Organe
über den gesamten Zeitraum nachweisbar. Im Dünndarm der LmoEGDe-mur-lux
infizierten C57BL/6J und A/JOlaHsd Mäuse konnten Signale am Tag eins nach der
Infektion nachgewiesen werden (Abbildung 2-23 B). Am Tag drei nach der oralen
Infektion mit LmoEGDe-mur-lux wurden Signale in den mLK aller analysierten
Mausinzuchtstämme beobachtet. Bis zum Tag fünf nach der Infektion mit LmoEGDe-
mur-lux konnten Biolumineszenzsignale in der Leber, im Dünndarm und in den mLK
von C3HeB/FeJ, A/JOlaHsd und BALB/cJ Mäusen detektiert werden. Im Falle der
C57BL/6J Mäuse begrenzte sich der Nachweis der Biolumineszenz auf die Gallen-
blase. Am Tag sieben nach der Infektion mit LmoEGDe-mur-lux konnten in den Gal-
lenblasen der A/JOlaHsd, BALB/cJ und C57BL/6J Mäusen Listerien nachgewiesen
werden. Der Vergleich von LmoEGDe-lux mit LmoEGDe-lux-mur zeigte Unterschiede
in der Biolumineszenzstärke besonders an Tag drei und fünf nach der Infektion auf.
LmoEGDe-lux-mur zeigte deutlichere Signale in den mLK an Tag drei nach der
Infektion in allen analysierten Mausinzuchtstämmen und in der Leber von
C3HeB/FeJ, A/JOlaHsd und BALB/cJ Mäusen an Tag fünf nach der Infektion. Auch
sind empfindliche und resistente Mausinzuchtstämme deutlich voneinander differen-
zierbar.
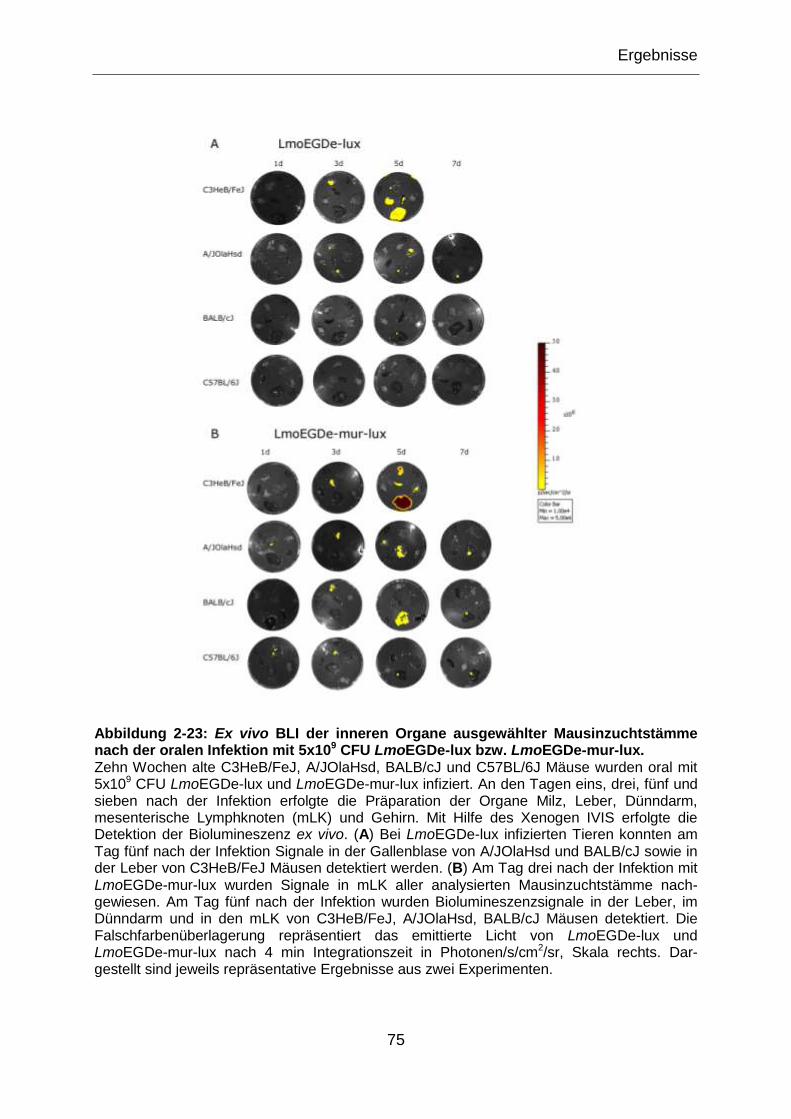
Ergebnisse
75
Abbildung 2-23: Ex vivo BLI der inneren Organe ausgewählter Mausinzuchtstämme nach der oralen Infektion mit 5x109 CFU LmoEGDe-lux bzw. LmoEGDe-mur-lux. Zehn Wochen alte C3HeB/FeJ, A/JOlaHsd, BALB/cJ und C57BL/6J Mäuse wurden oral mit 5x109 CFU LmoEGDe-lux und LmoEGDe-mur-lux infiziert. An den Tagen eins, drei, fünf und sieben nach der Infektion erfolgte die Präparation der Organe Milz, Leber, Dünndarm, mesenterische Lymphknoten (mLK) und Gehirn. Mit Hilfe des Xenogen IVIS erfolgte die Detektion der Biolumineszenz ex vivo. (A) Bei LmoEGDe-lux infizierten Tieren konnten am Tag fünf nach der Infektion Signale in der Gallenblase von A/JOlaHsd und BALB/cJ sowie in der Leber von C3HeB/FeJ Mäusen detektiert werden. (B) Am Tag drei nach der Infektion mit LmoEGDe-mur-lux wurden Signale in mLK aller analysierten Mausinzuchtstämme nach-gewiesen. Am Tag fünf nach der Infektion wurden Biolumineszenzsignale in der Leber, im Dünndarm und in den mLK von C3HeB/FeJ, A/JOlaHsd, BALB/cJ Mäusen detektiert. Die Falschfarbenüberlagerung repräsentiert das emittierte Licht von LmoEGDe-lux und LmoEGDe-mur-lux nach 4 min Integrationszeit in Photonen/s/cm2/sr, Skala rechts. Dar-gestellt sind jeweils repräsentative Ergebnisse aus zwei Experimenten.

Ergebnisse
76
Mit zunehmender Resistenz sinkt die Detektierbarkeit der Biolumineszenz. In den
inneren Organen der für die Listerieninfektion empfänglichen C3HeB/FeJ Mäuse
konnten starke Biolumineszenzsignale in der Leber, in der Milz und in den mLK
nachgewiesen werden. Dagegen waren in den resistenten C57BL/6J Mäusen nur
schwache Biolumineszenzsignale detektierbar.
Für die weitere Analyse der bakteriellen Keimlast wurden die präparierten Organe
Leber, Milz, Dünndarm, mLK und Gehirn in PBS homogenisiert und in Verdünnungs-
stufen auf BHI-Agarplatten ausgestrichen. Nach 24 Stunden wurden die gewach-
senen Kolonien gezählt und die bakterielle Keimlast pro Milligramm Organ berech-
net. In Abbildung 2-24 sind die bakteriellen Keimlasten der Organe Milz, Leber,
Dünndarm, mLK und Gehirn der Mausinzuchtstämme C57BL/6J, A/JOlaHsd,
BALB/cJ und C3HeB/FeJ an Tag drei und fünf nach der Infektion mit LmoEGDe-lux
und LmoEGDe-lux-mur dargestellt. Die bakterielle Keimlast in LmoEGDe-lux und
LmoEGDe-lux-mur infizierten C3HeB/FeJ Mäusen weiste nur an Tag drei nach der
Infektion einen signifikanten Unterschied in der Milz auf. In allen anderen analy-
sierten Organen war kein Unterschied in der Listerienanzahl zu beobachten. Im wei-
teren Infektionsverlauf war ein Anstieg der Bakterienanzahl in der Milz, der Gallen-
blase und im Gehirn von Tag drei auf Tag fünf um das Zehnfache, in der Leber sogar
um das Hundertfache, zu beobachten (Abbildung 2-24 A). Eine Änderung der Bakte-
rienlast im Dünndarm und in mLK war von Tag drei auf Tag fünf nach der Infektion
nicht nachzuweisen. Jedoch konnte ein signifikanter Unterschied der Listerienanzahl
an Tag fünf nach der Infektion, im Vergleich von LmoEGDe-lux zu LmoEGDe-lux-
mur, im Dünndarm bestimmt werden. Die Unterschiede der bakteriellen Keimlasten
in den analysierten Organe der C3HeB/FeJ Mäusen, nach der Infektion mit murini-
sierten und nicht-murinisierten Listerienstämmen, zeigten nur in Milz (3d pi) und
Dünndarm (5d pi) signifikante Unterschiede. C3HeB/FeJ Mäuse reagierten auf die
Infektion mit beiden Listerienvarianten sehr empfindlich. Im Fall der A/JOlaHsd
Mäuse zeigte eine Infektion mit LmoEGDe-lux-mur an Tag drei nach der Infektion nur
in der Leber einen signifikanten Unterschied in der Listerienanzahl im Vergleich zu
einer Infektion mit LmoEGDe-lux.
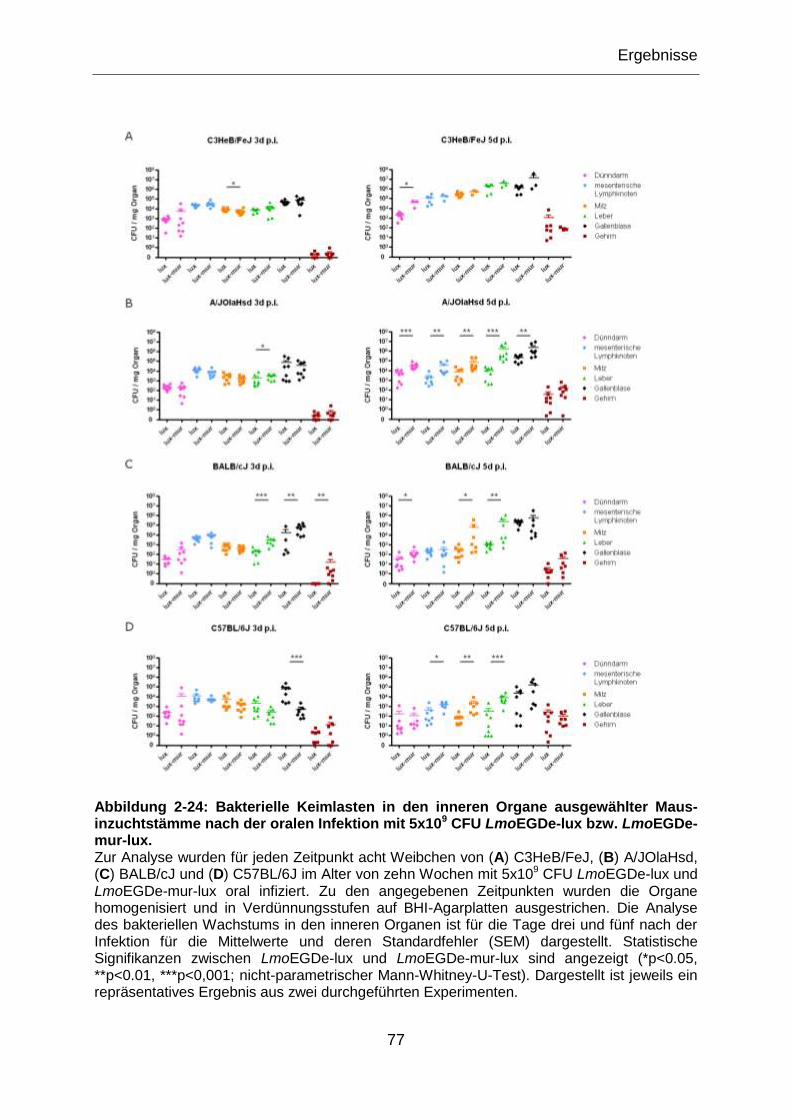
Ergebnisse
77
Abbildung 2-24: Bakterielle Keimlasten in den inneren Organe ausgewählter Maus-inzuchtstämme nach der oralen Infektion mit 5x109 CFU LmoEGDe-lux bzw. LmoEGDe-mur-lux. Zur Analyse wurden für jeden Zeitpunkt acht Weibchen von (A) C3HeB/FeJ, (B) A/JOlaHsd, (C) BALB/cJ und (D) C57BL/6J im Alter von zehn Wochen mit 5x109 CFU LmoEGDe-lux und LmoEGDe-mur-lux oral infiziert. Zu den angegebenen Zeitpunkten wurden die Organe homogenisiert und in Verdünnungsstufen auf BHI-Agarplatten ausgestrichen. Die Analyse des bakteriellen Wachstums in den inneren Organen ist für die Tage drei und fünf nach der Infektion für die Mittelwerte und deren Standardfehler (SEM) dargestellt. Statistische Signifikanzen zwischen LmoEGDe-lux und LmoEGDe-mur-lux sind angezeigt (*p<0.05, **p<0.01, ***p<0,001; nicht-parametrischer Mann-Whitney-U-Test). Dargestellt ist jeweils ein repräsentatives Ergebnis aus zwei durchgeführten Experimenten.

Ergebnisse
78
Im Verlauf der Infektion konnte ein Anstieg der Listerienanzahl in der Milz und in der
Gallenblase der LmoEGDe-lux-mur infizierten A/JOlaHsd Mäuse um das Zehnfache,
in der Leber, im Gehirn und im Dünndarm um das Hundertfache beobachtet werden.
Während die orale Infektion mit LmoEGDe-lux in einem geringen Anstieg der
Listerienanzahl in Milz, Leber, Dünndarm und Gehirn resultierte, sank die bakterielle
Keimlast in den mLK von Tag drei auf Tag fünf nach der Infektion um den Faktor
zehn. Die Unterschiede der bakteriellen Keimlasten in den analysierten Organe der
A/JOlaHsd Mäuse, im Vergleich der Infektion murinisierter mit nicht-murinisierten
Listerienstämme, zeigte an Tag fünf nach der Infektion signifikante Unterschiede in
der Milz, in der Leber, in der Gallenblase, im Dünndarm und in den mLK (Abbildung
2-24 B). Die bakterielle Keimlast in den analysierten Organen bei LmoEGDe-lux im
Vergleich zu LmoEGDe-lux-mur infizierten BALB/cJ Mäusen wies am Tag drei nach
der Infektion signifikante Unterschiede in der Leber, in der Gallenblase und im Gehirn
auf. Im weiteren Infektionsverlauf war ein Anstieg der Bakterienanzahl in der Gallen-
blase bei LmoEGDe-lux-mur infizierten BALB/cJ Mäusen, von Tag drei auf Tag fünf,
um das Zehnfache, in der Leber und der Milz sogar um das Hundertfache, zu
beobachten. Nach der Infektion mit LmoEGDe-lux konnte in BALB/cJ Mäusen eine
Zunahme der Listerienanzahl in der Gallenblase, von Tag drei auf Tag fünf nach der
Infektion, nachgewiesen werden. Die bakterielle Keimlast in den mLK reduzierte sich
von Tag drei auf Tag fünf, sowohl bei der Infektion mit LmoEGDe-lux als auch bei der
Infektion mit LmoEGDe-lux-mur um den Faktor zehn. Signifikante Unterschiede
konnten beim Vergleich der bakteriellen Keimlasten der LmoEGDe-lux zu LmoEGDe-
lux-mur infizierten BALB/cJ Mäusen, an Tag fünf nach der Infektion, in der Milz, in
der Leber und im Dünndarm nachgewiesen werden. Die Listerienanzahl in den
analysierten Organen von LmoEGDe-lux im Vergleich zu LmoEGDe-lux-mur infizier-
ten C57BL/6J Mäusen wies am Tag drei nach der Infektion einen signifikanten
Unterschied in der Gallenblase auf. Von Tag drei auf Tag fünf nach der Infektion war
ein Anstieg der Bakterienanzahl in der Leber und im Gehirn bei LmoEGDe-lux-mur
infizierten C57BL/6J Mäusen, um das Zehnfache, in der Gallenblase um das
Hundertfache, zu beobachten. Die Infektion mit LmoEGDe-lux führte in C57BL/6J
Mäusen von Tag drei auf Tag fünf nach der Infektion zu einer Zunahme der

Ergebnisse
79
Listerienanzahl im Gehirn um den Faktor zehn. Die bakterielle Keimlast in den mLK
reduzierte sich von Tag drei auf Tag fünf, sowohl bei der Infektion mit LmoEGDe-lux
als auch bei der Infektion mit LmoEGDe-lux-mur um den Faktor zehn. Ebenso sank
die Listerienanzahl in der Milz LmoEGDe-lux infizierter C57BL/6J sowie im Dünn-
darm LmoEGDe-lux-mur infizierter C57BL/6J Mäuse, von Tag drei auf Tag fünf, um
den Faktor hundert. Signifikante Unterschiede konnten beim Vergleich der bakteriel-
len Keimlasten der LmoEGDe-lux zu LmoEGDe-lux-mur infizierten C57BL/6J Mäuse,
am Tag fünf nach der Infektion, in der Milz, in der Leber und in den mLK nach-
gewiesen werden.
Die Analyse der bakteriellen Keimlasten zeigten die deutlichsten Änderungen der
Listerienanzahl in den untersuchten Organen von Tag drei auf Tag fünf nach der
Infektion. Für die Analyse der Wirtsresistenz von C3HeB/FeJ, A/JOlaHsd, BALB/cJ
und C57BL/6J wurden die Bakterienlasten der Organe der LmoEGDe-lux-mur
infizierten Mausinzuchtstämme am Tag fünf nach der Infektion miteinander ver-
glichen (Abbildung 2-25). Die C3HeB/FeJ Mäuse wiesen am Tag fünf nach der Infek-
tion mit LmoEGDe-lux-mur die höchste Anzahl an Listerien im Dünndarm, in den
mLK, in der Milz, in der Leber und in der Gallenblase auf, gefolgt von A/JOlaHsd und
BALB/cJ Mäusen. Dabei konnte eine hundertfach höhere Listerienanzahl in den
Organen der C3HeB/FeJ Mäuse im Vergleich zu C57BL/6J Mäuse festgestellt
werden. Die A/JOlaHsd Mäuse zeigten eine zehnfach höhere bakterielle Keimlast in
den mLK und der Gallenblase und eine bis zu hundertfach höhere Listerienanzahl im
Dünndarm, der Milz und der Leber im Vergleich zu C57BL/6J Mäusen. Die Infektion
der BALB/cJ Mäuse resultierte an Tag fünf nach der Infektion mit LmoEGDe-lux-mur
in einer zehnfach höheren Listerienanzahl in den mLK, der Leber und der Milz im
Vergleich zu C57BL/6J Mäusen. Die signifikanten Unterschiede in den Bakterien-
lasten von Dünndarm, mLK, Milz, Leber und Gallenblase zwischen den
ausgewählten Mausinzuchtstämmen sind in der Abbildung 2-25 dargestellt. Die
Anzahl der Listerien im Gehirn der C3HeB/FeJ, A/JOlaHsd, BALB/cJ und C57BL/6J
Mäuse zeigten keine signifikanten Unterschiede.
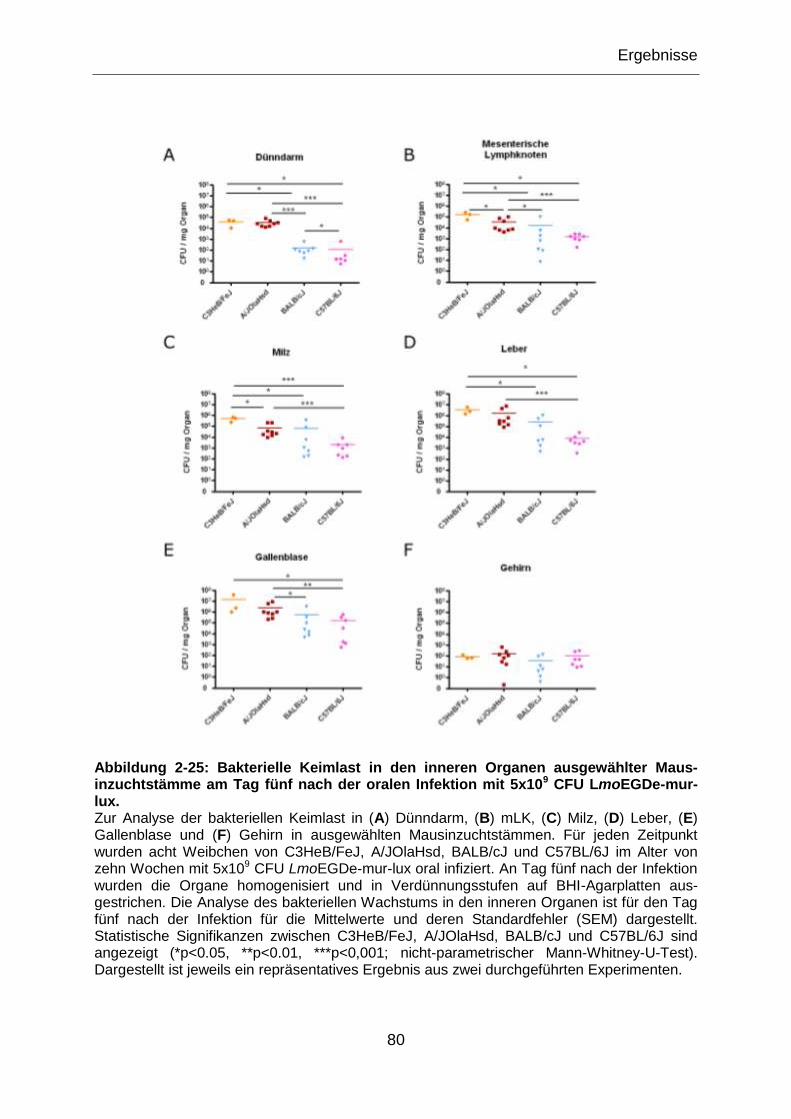
Ergebnisse
80
Abbildung 2-25: Bakterielle Keimlast in den inneren Organen ausgewählter Maus-inzuchtstämme am Tag fünf nach der oralen Infektion mit 5x109 CFU LmoEGDe-mur-lux. Zur Analyse der bakteriellen Keimlast in (A) Dünndarm, (B) mLK, (C) Milz, (D) Leber, (E) Gallenblase und (F) Gehirn in ausgewählten Mausinzuchtstämmen. Für jeden Zeitpunkt wurden acht Weibchen von C3HeB/FeJ, A/JOlaHsd, BALB/cJ und C57BL/6J im Alter von zehn Wochen mit 5x109 CFU LmoEGDe-mur-lux oral infiziert. An Tag fünf nach der Infektion wurden die Organe homogenisiert und in Verdünnungsstufen auf BHI-Agarplatten aus-gestrichen. Die Analyse des bakteriellen Wachstums in den inneren Organen ist für den Tag fünf nach der Infektion für die Mittelwerte und deren Standardfehler (SEM) dargestellt. Statistische Signifikanzen zwischen C3HeB/FeJ, A/JOlaHsd, BALB/cJ und C57BL/6J sind angezeigt (*p<0.05, **p<0.01, ***p<0,001; nicht-parametrischer Mann-Whitney-U-Test). Dargestellt ist jeweils ein repräsentatives Ergebnis aus zwei durchgeführten Experimenten.

Ergebnisse
81
Somit lässt sich die Suszeptibilität eines Mausstammes an der bakteriellen Keimlast
beurteilen. Je empfindlicher ein Mausstamm, desto höher die Bakterienlast in den
inneren Organen.
2.3.4.3 Histopathologische Analyse von LmoEGDe-lux und LmoEGDe-mur-lux
infizierten Mausinzuchtstämmen
Parallel zur Analyse der bakteriellen Keimlast erfolgte die histopathologische und
immunhistologische Analyse in den Organen Milz, Leber und Dünndarm. Die Organe
Milz, Leber und Dünndarm wurden an Tag eins, drei, fünf und sieben nach der Infek-
tion präpariert, in Formaldehyd fixiert und mit Hämatoxylin-Eosin gefärbt. Weiterhin
erfolgte mit Hilfe immunhistologischer Verfahren die Analyse zum Nachweis auf
Listeria monocytogenes innerhalb des Gewebes. Abbildung 2-26 zeigt die histo-
pathologische Analyse der Leber von C3HeB/FeJ und C57BL/6J am Tag fünf nach
der oralen Infektion mit 5x109 CFU LmoEGDe-mur-lux. Die orale Infektion von
C57BL/6J Mäusen mit dem Listerienstamm LmoEGDe-mur-lux führte zu moderaten
Schädigungen des Leberparenchyms mit vereinzelten Nekrosen und Infiltrationen
von Granulozyten und Makrophagen. Im Vergleich dazu zeigte die infizierte Leber
der C3HeB/FeJ Mäuse eine massivere Schädigung des Lebergewebes mit entzünd-
lichen Läsionen. Es konnten vermehrt Nekrosen und Infiltrationen von Granulozyten
und Makrophagen nachgewiesen werden. Mit Hilfe der immunhistologischen Analyse
sollte der Nachweis der Listerien innerhalb des Gewebes über die immun-
histologische Färbung unter Verwendung eines monoklonalen Antikörpers gegen
Listeria monocytogenes erfolgen (Abbildung 2-26 C und D). Listerien konnten
innerhalb der Infiltrate und Nekrosen des Leberparenchyms der infizierten
C3HeB/FeJ Mäuse reichlich nachgewiesen werden. Auch konnten in den
histologischen Präparaten der Leber von C57BL/6J Mäusen Listerien nachgewiesen
werden. Jedoch war die Anzahl der dokumentierten Bakterien in C57BL/6J Mäusen
deutlich geringer im Vergleich zu den nachgewiesenen Listerien in C3HeB/FeJ
Mäusen.
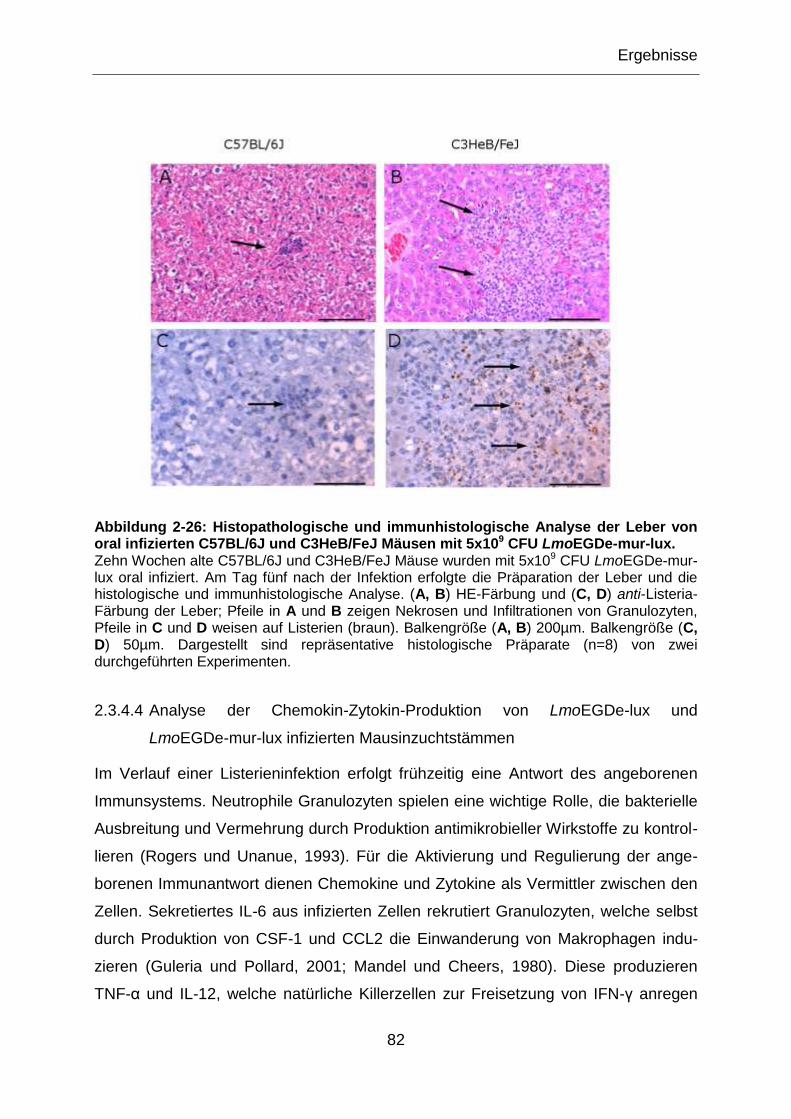
Ergebnisse
82
Abbildung 2-26: Histopathologische und immunhistologische Analyse der Leber von oral infizierten C57BL/6J und C3HeB/FeJ Mäusen mit 5x109 CFU LmoEGDe-mur-lux. Zehn Wochen alte C57BL/6J und C3HeB/FeJ Mäuse wurden mit 5x109 CFU LmoEGDe-mur-lux oral infiziert. Am Tag fünf nach der Infektion erfolgte die Präparation der Leber und die histologische und immunhistologische Analyse. (A, B) HE-Färbung und (C, D) anti-Listeria-Färbung der Leber; Pfeile in A und B zeigen Nekrosen und Infiltrationen von Granulozyten, Pfeile in C und D weisen auf Listerien (braun). Balkengröße (A, B) 200µm. Balkengröße (C, D) 50µm. Dargestellt sind repräsentative histologische Präparate (n=8) von zwei durchgeführten Experimenten.
2.3.4.4 Analyse der Chemokin-Zytokin-Produktion von LmoEGDe-lux und
LmoEGDe-mur-lux infizierten Mausinzuchtstämmen
Im Verlauf einer Listerieninfektion erfolgt frühzeitig eine Antwort des angeborenen
Immunsystems. Neutrophile Granulozyten spielen eine wichtige Rolle, die bakterielle
Ausbreitung und Vermehrung durch Produktion antimikrobieller Wirkstoffe zu kontrol-
lieren (Rogers und Unanue, 1993). Für die Aktivierung und Regulierung der ange-
borenen Immunantwort dienen Chemokine und Zytokine als Vermittler zwischen den
Zellen. Sekretiertes IL-6 aus infizierten Zellen rekrutiert Granulozyten, welche selbst
durch Produktion von CSF-1 und CCL2 die Einwanderung von Makrophagen indu-
zieren (Guleria und Pollard, 2001; Mandel und Cheers, 1980). Diese produzieren
TNF-α und IL-12, welche natürliche Killerzellen zur Freisetzung von IFN-γ anregen

Ergebnisse
83
(Havell, 1987; Tripp et al., 1993), was wiederum weitere Makrophagen aktiviert.
Durch die neutrophil bedingte Produktion von reaktiven Sauerstoffspezies erfolgt die
durch Makrophagen bedingte Eliminierung von Listeria monocytogenes.
Inflammatorische Zytokine sind wichtig für die Kommunikation der beteiligten Immun-
zellen. Zur Analyse des Effektes einer Infektion mit Listeria monocytogenes auf die
angeborene Immunabwehr wurden am Tag drei und fünf nach der Infektion mit
LmoEGDe-lux und LmoEGDe-mur-lux Serumproben von C3HeB/FeJ, A/JOlaHsd,
BALB/cJ und C57BL/6J Mäusen entnommen und die Konzentration verschiedener
Chemokine und Zytokine mit Hilfe eines Multi-Bead-Immunoassays untersucht.
Spektral kodierte Kügelchen dienen als Träger für die Antikörper gegen ausgewählte
Chemokine und Zytokine. Die unterschiedlichen spektralen Eigenschaften der Beads
erlauben die quantitative Analyse der chemotaktischen Stoffe. In Abbildung 2-27 sind
Konzentration von IFN-γ, IL-10, TNF-α, IL-6, CCL2, IL-5 und IL-1β in den
Mausinzuchtstämmen C3HeB/FeJ, A/JOlaHsd, BALB/cJ und C57BL/6J an Tag drei
und fünf nach der Infektion mit LmoEGDe-lux bzw. LmoEGDe-mur-lux dargestellt. In
C3HeB/FeJ resultierte sowohl die Infektion mit LmoEGDe-lux als auch mit
LmoEGDe-mur-lux in einen Anstieg der dargestellten Zytokine von Tag drei auf Tag
fünf nach der Infektion (Abbildung 2-27 A). Es konnten signifikante Unterschiede bei
der Konzentration von IFN-γ, TNF-α und CCL2 von Tag drei auf Tag fünf nach der
Infektion sowohl bei LmoEGDe-lux infizierten als auch bei LmoEGDe-mur-lux infi-
zierten C3HeB/FeJ Mäusen nachgewiesen werden. Ebenfalls konnte eine signifi-
kante Erhöhung von IL-10 und IL-6 bei LmoEGDe-mur-lux infizierten C3HeB/FeJ
Mäusen festgestellt werden. Bei IL-5 und IL-1β war dagegen nur ein geringer Anstieg
nachzuweisen. Die Infektion von A/JOlaHsd Mäusen mit LmoEGDe-lux bzw.
LmoEGDe-mur-lux resultierte ebenfalls in einem Anstieg von IFN-γ, TNF-α, IL-6,
CCL2, IL-5 und IL-1β von Tag drei auf Tag fünf nach der Infektion. Dabei konnte ein
signifikanter Unterschied von Tag drei auf Tag fünf nach der Infektion mit LmoEGDe-
mur-lux bei TNF-α, IL-6 und CCL2 beobachtet werden. Weiterhin zeigten sich Unter-
schiede in der Produktion der Chemokine und Zytokine in Abhängigkeit von der
Infektion mit LmoEGDe-lux und LmoEGDe-mur-lux.
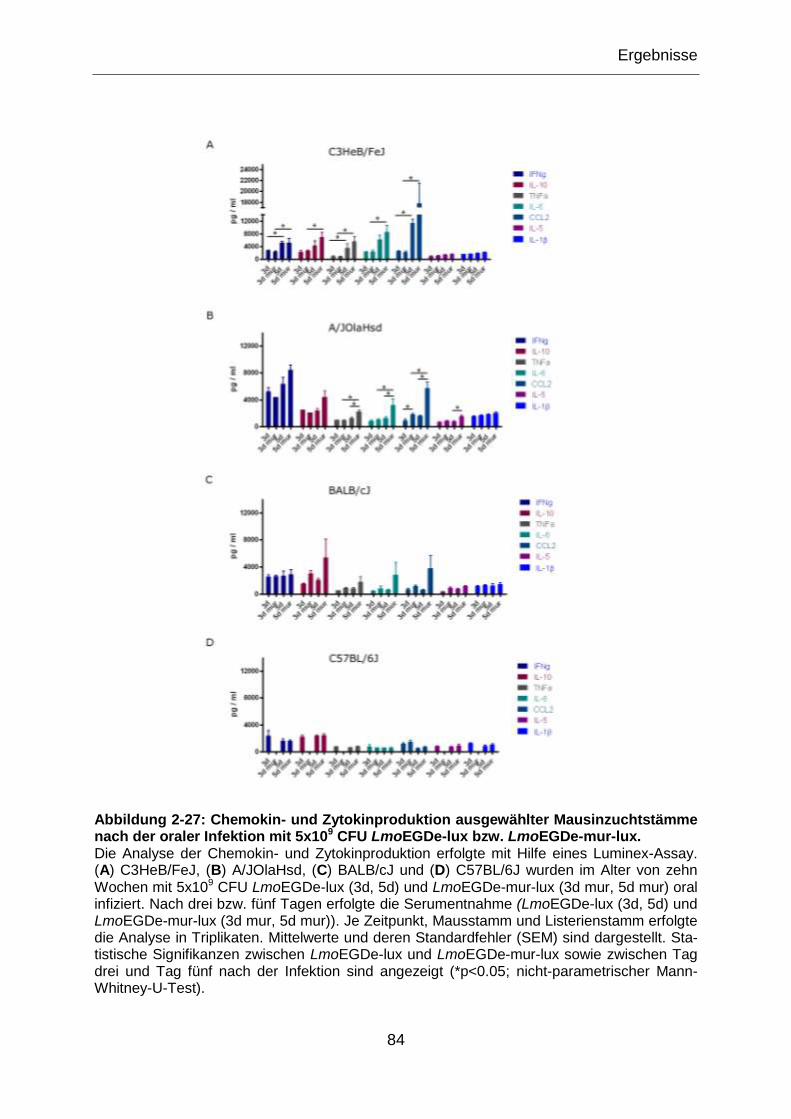
Ergebnisse
84
Abbildung 2-27: Chemokin- und Zytokinproduktion ausgewählter Mausinzuchtstämme nach der oraler Infektion mit 5x109 CFU LmoEGDe-lux bzw. LmoEGDe-mur-lux. Die Analyse der Chemokin- und Zytokinproduktion erfolgte mit Hilfe eines Luminex-Assay. (A) C3HeB/FeJ, (B) A/JOlaHsd, (C) BALB/cJ und (D) C57BL/6J wurden im Alter von zehn Wochen mit 5x109 CFU LmoEGDe-lux (3d, 5d) und LmoEGDe-mur-lux (3d mur, 5d mur) oral infiziert. Nach drei bzw. fünf Tagen erfolgte die Serumentnahme (LmoEGDe-lux (3d, 5d) und LmoEGDe-mur-lux (3d mur, 5d mur)). Je Zeitpunkt, Mausstamm und Listerienstamm erfolgte die Analyse in Triplikaten. Mittelwerte und deren Standardfehler (SEM) sind dargestellt. Sta-tistische Signifikanzen zwischen LmoEGDe-lux und LmoEGDe-mur-lux sowie zwischen Tag drei und Tag fünf nach der Infektion sind angezeigt (*p<0.05; nicht-parametrischer Mann-Whitney-U-Test).

Ergebnisse
85
So konnte am Tag drei nach der Infektion ein signifikanter Unterschied der CCL2
Synthese zwischen LmoEGDe-lux und LmoEGDe-mur-lux infizierten A/JOlaHsd
Mäusen festgestellt werden. Weiter resultierte die Infektion mit LmoEGDe-mur-lux
am Tag fünf nach der Infektion in einer signifikant höheren Produktion von TNF-α, IL-
6, CCL2 und IL-5 als im Vergleich zur Infektion mit LmoEGDe-lux (Abbildung 2-27 B).
In BALB/cJ Mäusen konnte sowohl nach der Infektion mit LmoEGDe-lux als auch mit
LmoEGDe-mur-lux eine Zunahme der Konzentration von IL-10, TNF-α, IL-6, IL-5 und
IL-1β von Tag drei auf Tag fünf nach der Infektion im Serum nachgewiesen werden.
Signifikante Unterschiede wurden jedoch nicht festgestellt. Ein Anstieg der Kon-
zentration von CCL2 konnte nur nach der Infektion von LmoEGDe-mur-lux
beobachtet werden. Dagegen konnte eine Zunahme der Konzentration von IFN-γ
nach der Infektion mit beiden Listerienvarianten nicht belegt werden (Abbildung 2-27
C). Nach der Infektion von C57BL/6J Mäusen mit LmoEGDe-lux bzw. LmoEGDe-
mur-lux waren nur geringe Mengen von Chemokinen und Zytokinen messbar, die
Konzentrationen von IFN-γ, IL-10, TNF-α, IL-5 und IL-1β lagen an Tag drei nach der
Infektion mit LmoEGDe-mur-lux unter der Nachweisgrenze. Es konnten bei
C57BL/6J Mäusen keine signifikanten Veränderungen in der Produktion der
chemotaktischen Stoffe nachgewiesen werden (Abbildung 2-27 D).
2.4 Vergleich der Wirtsspezifität und Wirtsresistenz der Listerien-
stämme LmoXen32-mur und LmoEGDe-mur-lux
Die vorangegangenen Infektionen mit LmoXen32-mur und LmoEGDe-mur-lux
resultierten in unterschiedlichen Ergebnissen der Wirtsresistenz von C3HeB/FeJ,
A/JOlaHsd, BALB/cJ und C57BL/6J Mäusen. Die Infektion mit den attenuierten
Listerienstämmen LmoXen32 und LmoXen32-mur resultierte in einer geringeren
Mortalitätsrate als die Infektionen mit LmoEGDe-lux und LmoEGDe-mur-lux (siehe
Abbildungen 2-5 und 2-17). Die Ergebnisse des in vivo Imaging zeigten, dass
LmoEGDe-mur-lux in der Leber und Abdominalbereich detektierbar war, wogegen
nach der Infektion mit LmoXen32-mur Biolumineszenzsignale in der Gallenblase
oder im Darm nachgewiesen werden konnte (Vergleich Abbildung 2-6 und Abbildung
2-20). Die Untersuchung der bakteriellen Keimlast ergab eine höhere Listerienanzahl

Ergebnisse
86
in den Organen LmoEGDe-mur-lux infizierter BALB/cJ Mäuse als in den Organen
LmoXen32-mur infizierter BALB/cJ Mäuse (Vergleich Abbildung 2-9 und Abbildung 2-
24). Auch konnten während der histopathologischen und immunhistologischen
Analyse der Leber von BALB/cJ Mäusen massive Infiltrationen von neutrophilen
Granulozyten, Makrophagen und Listerien nach der Infektion mit LmoEGDe-mur-lux
(Abbildung 2-16) nachgewiesen werden. Im Gegensatz dazu führte die Infektion mit
LmoXen32-mur zu moderaten Schädigungen des Gewebes mit weniger
entzündlichen Läsionen innerhalb des Leberparenchyms (Abbildung 2-10). Die
Ursache der deutlichen Attenuierung des Listerienstamms LmoXen32-mur liegt in der
genetischen Modifikation über die Integration des luxABCDE Operons in das Genom
von Listeria monocytogenes Lmo 10403S (Hardy et al., 2004). Diese Integration
resultierte in einer viermal höheren LD50 im Vergleich zum ausgehenden
Wildtypstamm Lmo 10403S (Hardy et al., 2004).
Die Infektion mit LmoEGDe-mur-lux zeigte deutliche Unterschiede im Verlauf der
Listerieninfektion der Mausinzuchtstämme C3HeB/FeJ, A/JOlaHsd, BALB/cJ und
C57BL/6J. Die Infektion mit LmoEGDe-mur-lux resultierte bei C3HeB/FeJ Mäuse in
einer ausgeprägten Suszeptibilität mit einer geringeren Überlebensrate (Abbildung 2-
17). Die Infektion von A/JOlaHsd und BALB/cJ Mäusen zeigte intermediäre Infek-
tionsverläufe. In C57BL/6J konnte eine deutliche Resistenz gegenüber einer
Listerieninfektion nachgewiesen werden. Die Infektion mit LmoEGDe-mur-lux erlaubt
die Abhängigkeit der genetischen Prädisposition im Mausmodell zu untersuchen.
2.5 Korrelation der bakteriellen Keimlast mit Biolumineszenzsignalen in
den inneren Organen von LmoXen32 und LmoXen32-mur infizierten
Mausinzuchtstämmen im Vergleich zu LmoEGDe-lux und LmoEGDe-
lux-mur infizierten Mausinzuchtstämmen
Das Biolumineszenz-Imaging erlaubt, den Infektionsverlauf in vivo zu verfolgen und
Aussagen über Dissemination und Krankheitsentwicklung nach einer oralen
Listerieninfektion zu treffen. Für die Analyse, erfolgte mit Hilfe des Xenogen IVIS, die
Messung der Signalstärke der Biolumineszenz in den inneren Organen. Diese
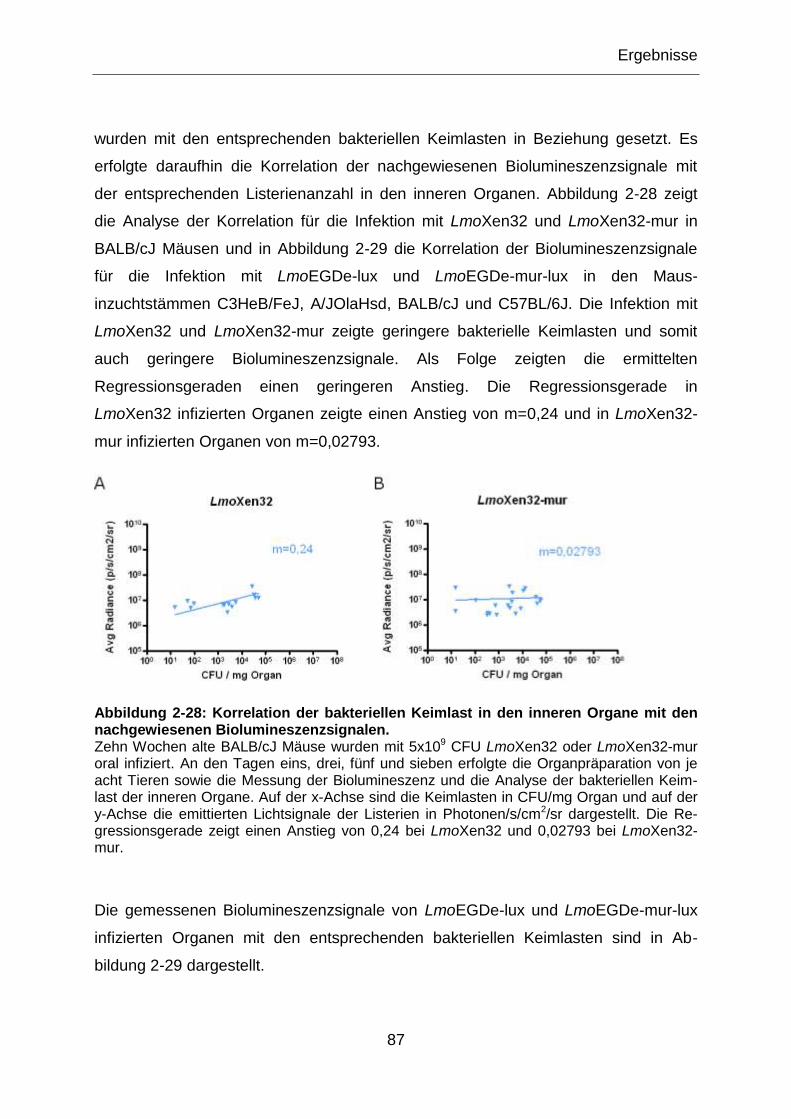
Ergebnisse
87
wurden mit den entsprechenden bakteriellen Keimlasten in Beziehung gesetzt. Es
erfolgte daraufhin die Korrelation der nachgewiesenen Biolumineszenzsignale mit
der entsprechenden Listerienanzahl in den inneren Organen. Abbildung 2-28 zeigt
die Analyse der Korrelation für die Infektion mit LmoXen32 und LmoXen32-mur in
BALB/cJ Mäusen und in Abbildung 2-29 die Korrelation der Biolumineszenzsignale
für die Infektion mit LmoEGDe-lux und LmoEGDe-mur-lux in den Maus-
inzuchtstämmen C3HeB/FeJ, A/JOlaHsd, BALB/cJ und C57BL/6J. Die Infektion mit
LmoXen32 und LmoXen32-mur zeigte geringere bakterielle Keimlasten und somit
auch geringere Biolumineszenzsignale. Als Folge zeigten die ermittelten
Regressionsgeraden einen geringeren Anstieg. Die Regressionsgerade in
LmoXen32 infizierten Organen zeigte einen Anstieg von m=0,24 und in LmoXen32-
mur infizierten Organen von m=0,02793.
Abbildung 2-28: Korrelation der bakteriellen Keimlast in den inneren Organe mit den nachgewiesenen Biolumineszenzsignalen. Zehn Wochen alte BALB/cJ Mäuse wurden mit 5x109 CFU LmoXen32 oder LmoXen32-mur oral infiziert. An den Tagen eins, drei, fünf und sieben erfolgte die Organpräparation von je acht Tieren sowie die Messung der Biolumineszenz und die Analyse der bakteriellen Keim-last der inneren Organe. Auf der x-Achse sind die Keimlasten in CFU/mg Organ und auf der y-Achse die emittierten Lichtsignale der Listerien in Photonen/s/cm2/sr dargestellt. Die Re-gressionsgerade zeigt einen Anstieg von 0,24 bei LmoXen32 und 0,02793 bei LmoXen32-mur.
Die gemessenen Biolumineszenzsignale von LmoEGDe-lux und LmoEGDe-mur-lux
infizierten Organen mit den entsprechenden bakteriellen Keimlasten sind in Ab-
bildung 2-29 dargestellt.
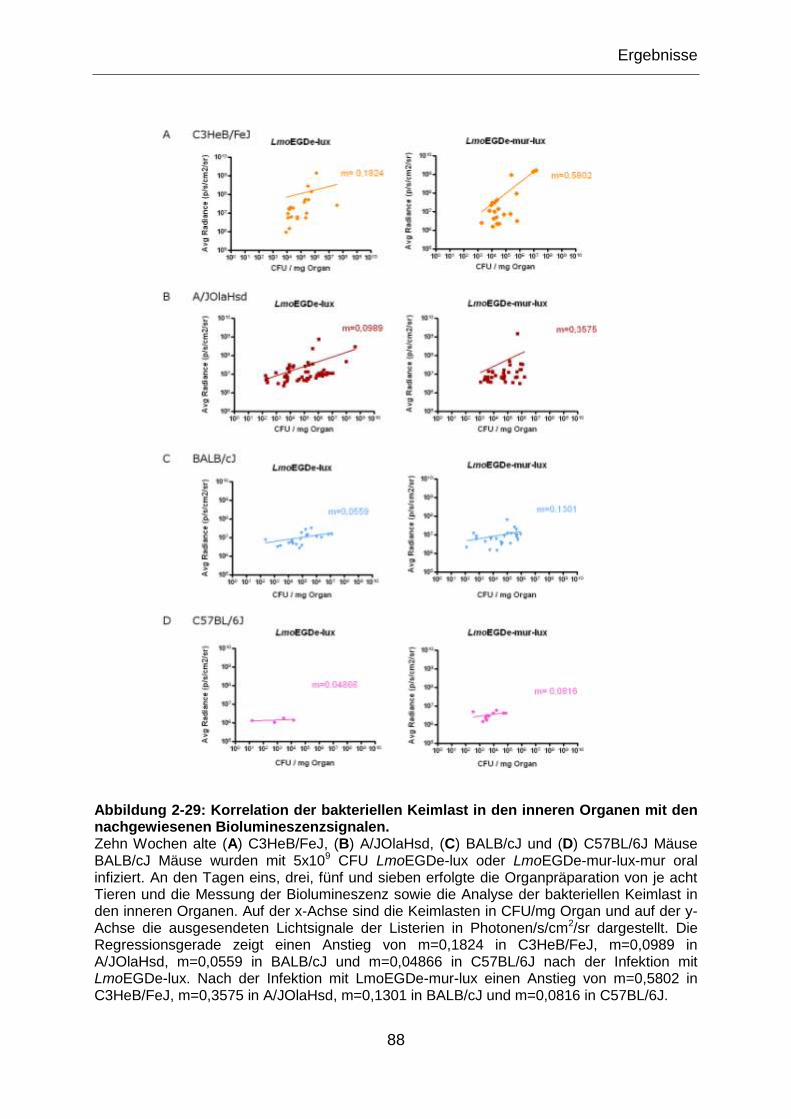
Ergebnisse
88
Abbildung 2-29: Korrelation der bakteriellen Keimlast in den inneren Organen mit den nachgewiesenen Biolumineszenzsignalen. Zehn Wochen alte (A) C3HeB/FeJ, (B) A/JOlaHsd, (C) BALB/cJ und (D) C57BL/6J Mäuse BALB/cJ Mäuse wurden mit 5x109 CFU LmoEGDe-lux oder LmoEGDe-mur-lux-mur oral infiziert. An den Tagen eins, drei, fünf und sieben erfolgte die Organpräparation von je acht Tieren und die Messung der Biolumineszenz sowie die Analyse der bakteriellen Keimlast in den inneren Organen. Auf der x-Achse sind die Keimlasten in CFU/mg Organ und auf der y-Achse die ausgesendeten Lichtsignale der Listerien in Photonen/s/cm2/sr dargestellt. Die Regressionsgerade zeigt einen Anstieg von m=0,1824 in C3HeB/FeJ, m=0,0989 in A/JOlaHsd, m=0,0559 in BALB/cJ und m=0,04866 in C57BL/6J nach der Infektion mit LmoEGDe-lux. Nach der Infektion mit LmoEGDe-mur-lux einen Anstieg von m=0,5802 in C3HeB/FeJ, m=0,3575 in A/JOlaHsd, m=0,1301 in BALB/cJ und m=0,0816 in C57BL/6J.

Ergebnisse
89
Die Korrelation der Biolumineszenzstärke mit der Listerienanzahl ergab Re-
gressionsgeraden mit einer größeren Steigung in LmoEGDe-mur-lux infizierten
Organen im Vergleich zu LmoEGDe-lux infizierten Organen. Die Regressions-
geraden nach der Infektion mit LmoEGDe-lux zeigten einen Anstieg von m=0,1824 in
C3HeB/FeJ, m=0,0989 in A/JOlaHsd, m=0,0559 in BALB/cJ und m=0,04866 in
C57BL/6J. Nach der Infektion mit LmoEGDe-mur-lux konnte ein Anstieg von
m=0,5802 in C3HeB/FeJ, m=0,3575 in A/JOlaHsd, m=0,1301 in BALB/cJ und
m=0,0816 in C57BL/6J Mäusen festgestellt werden. In den Mausstämmen
C3HeB/FeJ und A/JOlaHsd war eine Korrelation zwischen der Biolumineszenz und
der Listerienanzahl in den inneren Organen erkennbar. Ab einer Bakterienlast von
ca. 104 CFU/mg Organ war ein linearer Anstieg von bakterieller Keimlast zu der
Stärke des Biolumineszenzsignals nachweisbar. Bei BALB/cJ Mäusen zeigte sich ein
geringerer Anstieg der Regressionsgerade. Die Infektion von C57BL/6J Mäusen
resultierte aufgrund ihrer ausgeprägten Resistenz in einer geringeren Bakterienlast.
Dadurch waren nur wenige schwache Biolumineszenzsignale detektierbar und nur
ein geringer Anstieg erkennbar.
2.6 Infektion des zentralen Nervensystems nach der Infektion mit
LmoXen32, LmoXen32-mur, LmoEGDe, LmoEGDe-lux und
LmoEGDe-mur-lux
Listeria monocytogenes besitzt die Fähigkeit epitheliale Barrieren zu überqueren.
Neben der intestinalen Barriere und der Plazentaschranke besitzen sie auch die
Fähigkeit die Blut-Hirn-Schranke zu durchqueren, welches zu Meningitis und Enze-
phalitis führen kann. Während der vorangegangenen Experimente zur Analyse des
Infektionsprozesses in vivo nach oraler Infektion mit 5x109 CFU Listeria mono-
cytogenes zeigten acht Tiere neurologische Ausfallerscheinungen. Diese waren zu
Beginn gekennzeichnet durch Torticollis, später gefolgt von „kreiselnden“ Bewe-
gungen um die Längsachse und dem Verweilen am Käfigboden. Die Zeitpunkte des
Auftretens dieser Symptome konnten sieben bis acht Tage nach der oralen Infektion
beobachtet werden und sind in Tabelle 2-1 zusammengefasst.
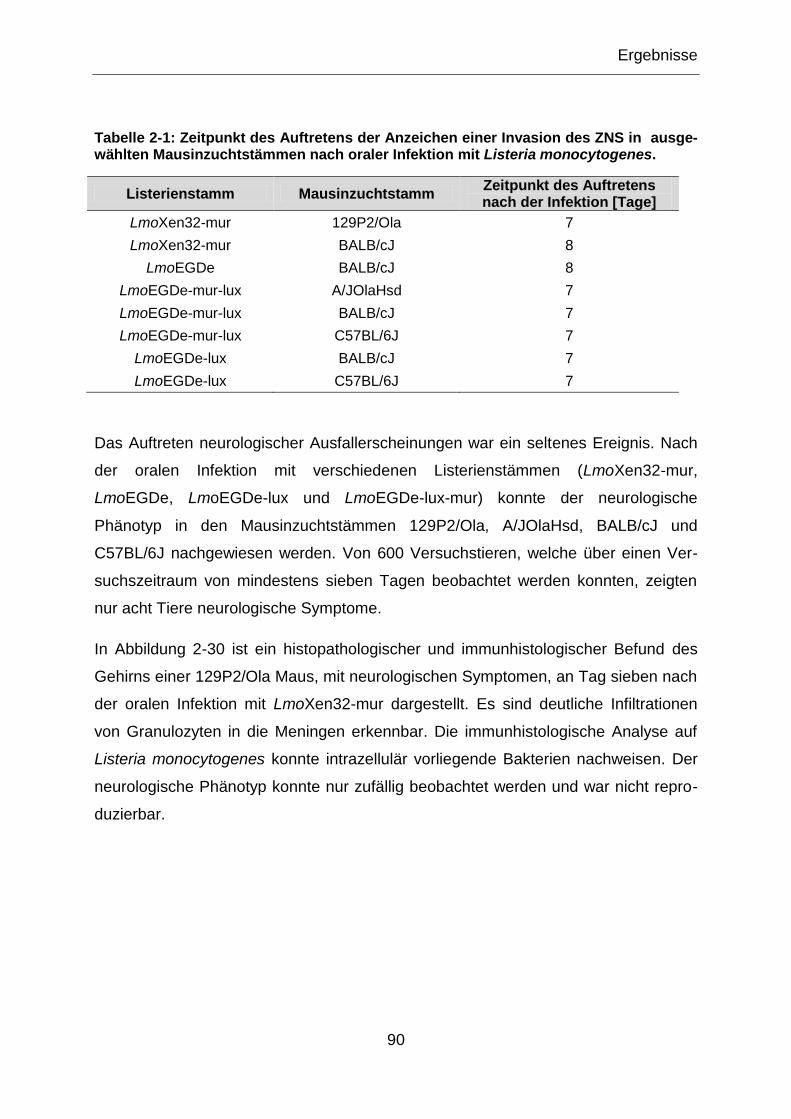
Ergebnisse
90
Tabelle 2-1: Zeitpunkt des Auftretens der Anzeichen einer Invasion des ZNS in ausge-wählten Mausinzuchtstämmen nach oraler Infektion mit Listeria monocytogenes.
Listerienstamm Mausinzuchtstamm Zeitpunkt des Auftretens nach der Infektion [Tage]
LmoXen32-mur 129P2/Ola 7
LmoXen32-mur BALB/cJ 8
LmoEGDe BALB/cJ 8
LmoEGDe-mur-lux A/JOlaHsd 7
LmoEGDe-mur-lux BALB/cJ 7
LmoEGDe-mur-lux C57BL/6J 7
LmoEGDe-lux BALB/cJ 7
LmoEGDe-lux C57BL/6J 7
Das Auftreten neurologischer Ausfallerscheinungen war ein seltenes Ereignis. Nach
der oralen Infektion mit verschiedenen Listerienstämmen (LmoXen32-mur,
LmoEGDe, LmoEGDe-lux und LmoEGDe-lux-mur) konnte der neurologische
Phänotyp in den Mausinzuchtstämmen 129P2/Ola, A/JOlaHsd, BALB/cJ und
C57BL/6J nachgewiesen werden. Von 600 Versuchstieren, welche über einen Ver-
suchszeitraum von mindestens sieben Tagen beobachtet werden konnten, zeigten
nur acht Tiere neurologische Symptome.
In Abbildung 2-30 ist ein histopathologischer und immunhistologischer Befund des
Gehirns einer 129P2/Ola Maus, mit neurologischen Symptomen, an Tag sieben nach
der oralen Infektion mit LmoXen32-mur dargestellt. Es sind deutliche Infiltrationen
von Granulozyten in die Meningen erkennbar. Die immunhistologische Analyse auf
Listeria monocytogenes konnte intrazellulär vorliegende Bakterien nachweisen. Der
neurologische Phänotyp konnte nur zufällig beobachtet werden und war nicht repro-
duzierbar.
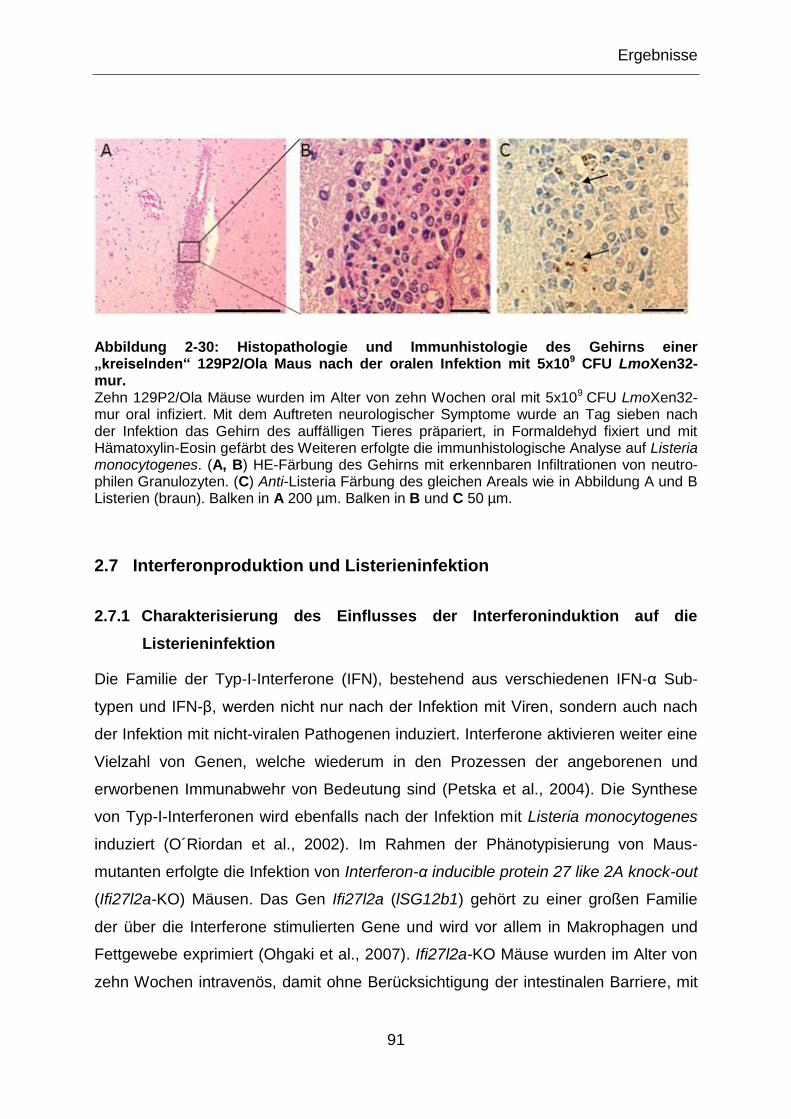
Ergebnisse
91
Abbildung 2-30: Histopathologie und Immunhistologie des Gehirns einer „kreiselnden“ 129P2/Ola Maus nach der oralen Infektion mit 5x109 CFU LmoXen32-mur. Zehn 129P2/Ola Mäuse wurden im Alter von zehn Wochen oral mit 5x109 CFU LmoXen32-mur oral infiziert. Mit dem Auftreten neurologischer Symptome wurde an Tag sieben nach der Infektion das Gehirn des auffälligen Tieres präpariert, in Formaldehyd fixiert und mit Hämatoxylin-Eosin gefärbt des Weiteren erfolgte die immunhistologische Analyse auf Listeria monocytogenes. (A, B) HE-Färbung des Gehirns mit erkennbaren Infiltrationen von neutro-philen Granulozyten. (C) Anti-Listeria Färbung des gleichen Areals wie in Abbildung A und B Listerien (braun). Balken in A 200 µm. Balken in B und C 50 µm.
2.7 Interferonproduktion und Listerieninfektion
2.7.1 Charakterisierung des Einflusses der Interferoninduktion auf die
Listerieninfektion
Die Familie der Typ-I-Interferone (IFN), bestehend aus verschiedenen IFN-α Sub-
typen und IFN-β, werden nicht nur nach der Infektion mit Viren, sondern auch nach
der Infektion mit nicht-viralen Pathogenen induziert. Interferone aktivieren weiter eine
Vielzahl von Genen, welche wiederum in den Prozessen der angeborenen und
erworbenen Immunabwehr von Bedeutung sind (Petska et al., 2004). Die Synthese
von Typ-I-Interferonen wird ebenfalls nach der Infektion mit Listeria monocytogenes
induziert (O´Riordan et al., 2002). Im Rahmen der Phänotypisierung von Maus-
mutanten erfolgte die Infektion von Interferon-α inducible protein 27 like 2A knock-out
(Ifi27l2a-KO) Mäusen. Das Gen Ifi27l2a (lSG12b1) gehört zu einer großen Familie
der über die Interferone stimulierten Gene und wird vor allem in Makrophagen und
Fettgewebe exprimiert (Ohgaki et al., 2007). Ifi27l2a-KO Mäuse wurden im Alter von
zehn Wochen intravenös, damit ohne Berücksichtigung der intestinalen Barriere, mit
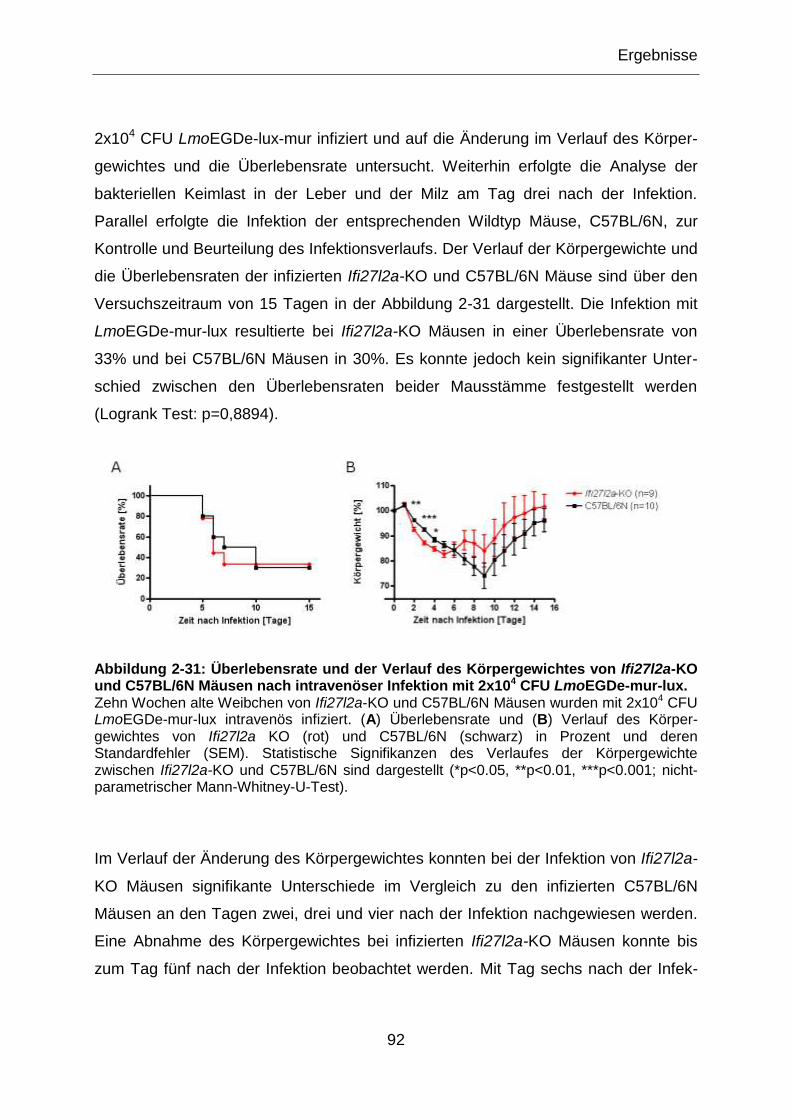
Ergebnisse
92
2x104 CFU LmoEGDe-lux-mur infiziert und auf die Änderung im Verlauf des Körper-
gewichtes und die Überlebensrate untersucht. Weiterhin erfolgte die Analyse der
bakteriellen Keimlast in der Leber und der Milz am Tag drei nach der Infektion.
Parallel erfolgte die Infektion der entsprechenden Wildtyp Mäuse, C57BL/6N, zur
Kontrolle und Beurteilung des Infektionsverlaufs. Der Verlauf der Körpergewichte und
die Überlebensraten der infizierten Ifi27l2a-KO und C57BL/6N Mäuse sind über den
Versuchszeitraum von 15 Tagen in der Abbildung 2-31 dargestellt. Die Infektion mit
LmoEGDe-mur-lux resultierte bei Ifi27l2a-KO Mäusen in einer Überlebensrate von
33% und bei C57BL/6N Mäusen in 30%. Es konnte jedoch kein signifikanter Unter-
schied zwischen den Überlebensraten beider Mausstämme festgestellt werden
(Logrank Test: p=0,8894).
Abbildung 2-31: Überlebensrate und der Verlauf des Körpergewichtes von Ifi27l2a-KO und C57BL/6N Mäusen nach intravenöser Infektion mit 2x104 CFU LmoEGDe-mur-lux. Zehn Wochen alte Weibchen von Ifi27l2a-KO und C57BL/6N Mäusen wurden mit 2x104 CFU LmoEGDe-mur-lux intravenös infiziert. (A) Überlebensrate und (B) Verlauf des Körper-gewichtes von Ifi27l2a KO (rot) und C57BL/6N (schwarz) in Prozent und deren Standardfehler (SEM). Statistische Signifikanzen des Verlaufes der Körpergewichte zwischen Ifi27l2a-KO und C57BL/6N sind dargestellt (*p<0.05, **p<0.01, ***p<0.001; nicht-parametrischer Mann-Whitney-U-Test).
Im Verlauf der Änderung des Körpergewichtes konnten bei der Infektion von Ifi27l2a-
KO Mäusen signifikante Unterschiede im Vergleich zu den infizierten C57BL/6N
Mäusen an den Tagen zwei, drei und vier nach der Infektion nachgewiesen werden.
Eine Abnahme des Körpergewichtes bei infizierten Ifi27l2a-KO Mäusen konnte bis
zum Tag fünf nach der Infektion beobachtet werden. Mit Tag sechs nach der Infek-
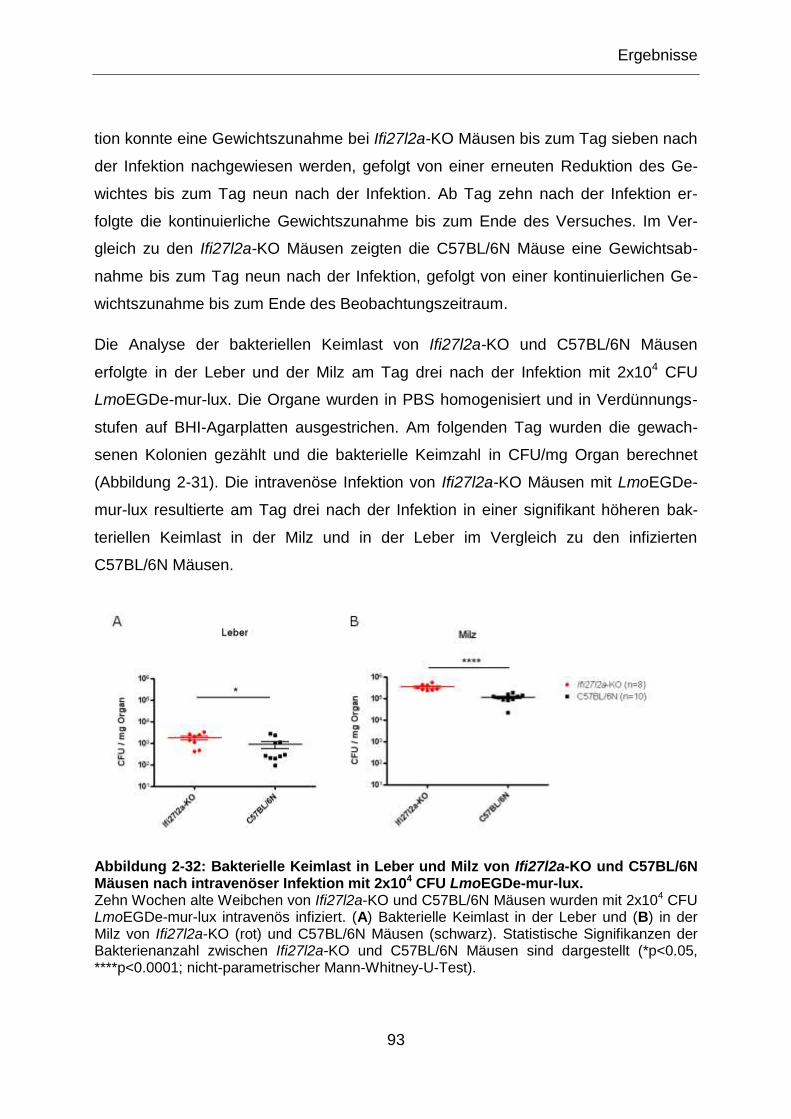
Ergebnisse
93
tion konnte eine Gewichtszunahme bei Ifi27l2a-KO Mäusen bis zum Tag sieben nach
der Infektion nachgewiesen werden, gefolgt von einer erneuten Reduktion des Ge-
wichtes bis zum Tag neun nach der Infektion. Ab Tag zehn nach der Infektion er-
folgte die kontinuierliche Gewichtszunahme bis zum Ende des Versuches. Im Ver-
gleich zu den Ifi27l2a-KO Mäusen zeigten die C57BL/6N Mäuse eine Gewichtsab-
nahme bis zum Tag neun nach der Infektion, gefolgt von einer kontinuierlichen Ge-
wichtszunahme bis zum Ende des Beobachtungszeitraum.
Die Analyse der bakteriellen Keimlast von Ifi27l2a-KO und C57BL/6N Mäusen
erfolgte in der Leber und der Milz am Tag drei nach der Infektion mit 2x104 CFU
LmoEGDe-mur-lux. Die Organe wurden in PBS homogenisiert und in Verdünnungs-
stufen auf BHI-Agarplatten ausgestrichen. Am folgenden Tag wurden die gewach-
senen Kolonien gezählt und die bakterielle Keimzahl in CFU/mg Organ berechnet
(Abbildung 2-31). Die intravenöse Infektion von Ifi27l2a-KO Mäusen mit LmoEGDe-
mur-lux resultierte am Tag drei nach der Infektion in einer signifikant höheren bak-
teriellen Keimlast in der Milz und in der Leber im Vergleich zu den infizierten
C57BL/6N Mäusen.
Abbildung 2-32: Bakterielle Keimlast in Leber und Milz von Ifi27l2a-KO und C57BL/6N Mäusen nach intravenöser Infektion mit 2x104 CFU LmoEGDe-mur-lux. Zehn Wochen alte Weibchen von Ifi27l2a-KO und C57BL/6N Mäusen wurden mit 2x104 CFU LmoEGDe-mur-lux intravenös infiziert. (A) Bakterielle Keimlast in der Leber und (B) in der Milz von Ifi27l2a-KO (rot) und C57BL/6N Mäusen (schwarz). Statistische Signifikanzen der Bakterienanzahl zwischen Ifi27l2a-KO und C57BL/6N Mäusen sind dargestellt (*p<0.05, ****p<0.0001; nicht-parametrischer Mann-Whitney-U-Test).

Ergebnisse
94
2.7.2 Charakterisierung der Listerieninfektion in Bezug auf die Interferon-β
Produktion
Interferon-β spielt bei der Infektion mit Viren und intrazellulären Mikroorganismen
eine wichtige Rolle für die Regulierung der Immunantwort (Stockinger und Decker,
2008; Stockinger et al., 2009). Mit Etablierung einer Interferon-β-Reportermaus war
es möglich, die gewebespezifische Interferon-β-Produktion aufzuzeigen. So konnte
die Visualisierung der IFN-β-Produktion in verschiedenen Geweben nach der
Infektion mit Influenza A oder La Crosse Virus in vivo beobachtet werden
(Lienenklaus et al., 2009). In dieser Maus ist ein Luziferase-Reportergen unter
Kontrolle des IFN-β-Promoters in den Interferon-β1 Locus (Ifnb1) über Gen-Targeting
eingebracht worden. Somit erfolgt eine Expression der Luziferase nach Induktion des
Ifnb1 Promoters in vivo innerhalb der Organe und kann durch die Injektion von D-
Luziferin visualisiert werden. Zur Analyse der IFN-β-Produktion nach der Infektion mit
Listeria monocytogenes unter Einfluss der Internalin A Murinisierung, wurden IFN-β-
Reportermäuse (BL6albino/-β+/Δβ-luc) mit LmoEGDe-lux, LmoEGDe-mur-lux, LO28
und LO28-mur oral infiziert. Die Listerienstämme EGDe und LO28 unterscheiden
sich in ihrer Fähigkeit eine IFN-β-Synthese zu induzieren. Der Stamm LO28 induziert
im Vergleich zu LmoEGDe eine stärkere IFN-β-Produktion (Reutterer et al., 2008).
Zur Analyse der IFN-β-Synthese nach einer Infektion mit Listeria monocytogenes
wurden je fünf Weibchen der IFN-β-Reportermäuse im Alter von zehn bis zwölf
Wochen mit 5x109 CFU LmoEGDe-lux, LmoEGDe-mur-lux, LO28 und LO28-mur oral
infiziert und täglich auf die IFN-β-Produktion untersucht (Abbildung 2-33). An Tag
eins nach der Infektion waren Signale in den zervikalen Lymphknoten (zLK) einzelner
Tiere unabhängig vom infizierten Listerienstamm erkennbar. Ab Tag zwei nach der
Infektion zeigten die LO28-mur infizierten Mäuse die deutlichste Induktion der IFN-β-
Produktion in den zervikalen Lymphknoten (zLK), im Darmbereich und teilweise in
der Leber.
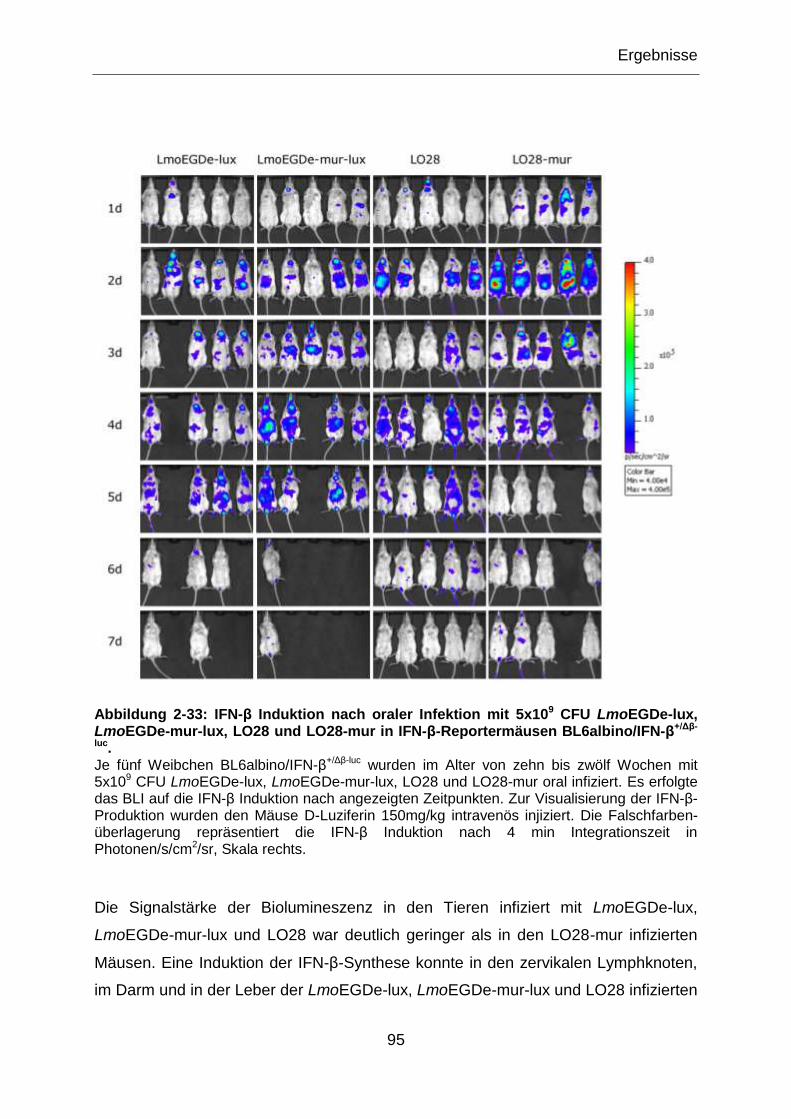
Ergebnisse
95
Abbildung 2-33: IFN-β Induktion nach oraler Infektion mit 5x109 CFU LmoEGDe-lux, LmoEGDe-mur-lux, LO28 und LO28-mur in IFN-β-Reportermäusen BL6albino/IFN-β+/Δβ-
luc. Je fünf Weibchen BL6albino/IFN-β+/Δβ-luc wurden im Alter von zehn bis zwölf Wochen mit 5x109 CFU LmoEGDe-lux, LmoEGDe-mur-lux, LO28 und LO28-mur oral infiziert. Es erfolgte das BLI auf die IFN-β Induktion nach angezeigten Zeitpunkten. Zur Visualisierung der IFN-β-Produktion wurden den Mäuse D-Luziferin 150mg/kg intravenös injiziert. Die Falschfarben-überlagerung repräsentiert die IFN-β Induktion nach 4 min Integrationszeit in Photonen/s/cm2/sr, Skala rechts.
Die Signalstärke der Biolumineszenz in den Tieren infiziert mit LmoEGDe-lux,
LmoEGDe-mur-lux und LO28 war deutlich geringer als in den LO28-mur infizierten
Mäusen. Eine Induktion der IFN-β-Synthese konnte in den zervikalen Lymphknoten,
im Darm und in der Leber der LmoEGDe-lux, LmoEGDe-mur-lux und LO28 infizierten

Ergebnisse
96
Tiere beobachtet werden. Am Tag drei nach der Infektion war eine Detektion der
Biolumineszenz in LmoEGDe-lux und LO28 infizierten Tieren in den zervikalen
Lymphknoten und vereinzelt im Darmbereich messbar. Ein Tier der mit LO28
infizierten Mäuse zeigte zusätzlich ein Signal in der Leber und in den inguinalen
Lymphknoten. Dagegen konnte bei den LmoEGDe-mur-lux und LO28-mur infizierten
Mäusen an Tag drei nach der Infektion zusätzlich Biolumineszenzsignale in der
Leber nachgewiesen werden. Am Tag vier nach der Infektion zeigten die Tiere
infiziert mit LmoEGDe-mur-lux die deutlichsten Biolumineszenzsignale, vor allem in
der Leber, im Abdominalbereich und in den zervikalen Lymphknoten. Im Vergleich
dazu konnten bei den LmoEGDe-lux infizierten Mäusen nur Signale in den zervikalen
Lymphknoten und vereinzelt im Darmbereich detektiert werden. Die Mäuse infiziert
mit LO28 zeigten an Tag vier nach der Infektion eine gesteigerte Induktion der IFN-β-
Synthese in der Leber, im Darmbereich und in den zervikalen sowie inguinalen
Lymphknoten im Vergleich zu Tag drei nach der Infektion. Bei den LO28-mur
infizierten Tieren verringerte sich dagegen die Detektierbarkeit der Biolumineszenz
von Tag drei auf Tag vier nach der Infektion und konnte in die zervikalen und
inguinalen Lymphknoten sowie vereinzelt in der Leber und im Darmbereich lokalisiert
werden. An Tag fünf konnte eine Zunahme der IFN-β-Produktion in den LmoEGDe-
lux infizierten Tieren von Tag vier nach der Infektion in den zLK und im Darmbereich
beobachtet werden. Ab Tag fünf reduzierte sich die IFN-β-Synthese in der Leber im
Darmbereich und in den zervikalen Lymphknoten der LmoEGDe-mur-lux, LO28 und
LO28-mur infizierten Mäusen. Ab Tag sechs nach der Infektion konnte nur noch ver-
einzelt in den LO28 infizierten Mäusen eine schwache Biolumineszenz nach-
gewiesen werden. Diese Ergebnisse bestätigen die Resultate von Reutterer et al.,
dass LO28 eine stärkere IFN-β-Synthese induziert als LmoEGDe (Reutterer et al.,
2008). Eine IFN-β-Produktion in der Milz, wie bei Solodova et al. nach intravenöser
Infektion beschrieben, konnte nicht nachgewiesen werden (Solodova et al., 2011).

Diskussion
97
3 Diskussion
Die Infektion des Menschen mit Listeria monocytogenes erfolgt über die orale Auf-
nahme von kontaminierten Lebensmitteln. Die Maus als Tiermodell ist jedoch für die
Infektion über den oralen Infektionsweg wenig empfänglich. Durch neue detaillierte
Beschreibungen der speziesabhängigen Interaktion des bakteriellen Oberflächen-
proteins Internalin A und seinem Rezeptor E-Cadherin sind in den letzten Jahren
große Fortschritte im Verständnis des Wirtstropismus erzielt worden. Neue Erkennt-
nisse, welche die molekularen Grundlagen bei der Überquerung der Darmbarriere
beschreiben und die bakterielle Invasion nach der oralen Infektion mit Listeria mono-
cytogenes ermöglichten, konnten gewonnen werden. Der entscheidendste Faktor für
die Speziesspezifität einer Listerieninfektion liegt in der Aminosäureposition 16 des
E-Cadherins. Innerhalb des Mäuse-E-Cadherins ist ein Glutamin an dieser Stelle
positioniert (Lecuit et al., 1999). Humanes E-Cadherin sowie das von Meerschwein-
chen und Kaninchen tragen dagegen an dieser Position ein Prolin. Erfolgt ein Aus-
tausch von Glutamin mit Prolin an der Position 16 des Maus-E-Cadherins, ist das
Internalin A fähig mit diesem humanisierten Rezeptor zu interagieren (Lecuit et al.,
1999; Lecuit et al., 2005). Eine Alternative bildet die Generierung eines murinisierten
Internalin A. Die Substitutionen zweier Aminosäuren resultieren in einer erhöhten
Affinität des Internalin A zum E-Cadherin. Zum einen der Austausch von Serin zu
Asparagin an der Aminosäureposition 192 und zum anderen der Austausch von
Tyrosin zu Serin an der Position 369 (Wollert et al., 2007a). Somit konnte diese
Speziesbarriere im Tiermodell durch Generierung transgener Mäuse bzw. genetisch
modifizierter Listerienstämme überbrückt werden (Disson et al., 2008; Lecuit et al.,
2002; Monk et al., 2010; Wollert et al., 2007b).
Über den Einsatz neuer bildgebender Verfahren, wie das auch in dieser Arbeit be-
nutzte in vivo Biolumineszenz-Imaging (BLI), kann die Ausbreitung und der
Infektionsverlauf nach der oralen Infektion mit biolumineszenten Listerien, im Tier-
modell, direkt verfolgt werden (Hardy et al., 2004; Riedel et al., 2007).

Diskussion
98
Ziel dieser Dissertation war die Etablierung eines in vivo Mausmodells für die orale
Infektion mit biolumineszenten Listeria monocytogenes Stämmen. Dabei sollte unter
Verwendung des in vivo BLI die Dissemination von Lux-Operon exprimierenden
Listerien im Wirtsorganismus der Maus nicht invasiv und unabhängig von der
Präsenz eines transgenen, humanisierten E-Cadherins analysiert werden. Weiter
stand die Fragestellung im Vordergrund, in wieweit der genetische Hintergrund des
Wirts den Krankheitsverlauf verschiedener Mausinzuchtstämme nach oraler Infektion
mit Listeria monocytogenes beeinflusst. Im Detail wurde erstmals die Wirtsresistenz
der Mausinzuchtstämme BALB/cJ, C57BL/6J, C3HeB/FeJ und A/JOlaHsd nach
oraler Infektion mit den Listerienstämmen LmoXen32, LmoXen32-mur, LmoEGDe-lux
und LmoEGDe-lux-mur analysiert. Die Analyse der Dissemination murinisierter
Listerien im Vergleich zu ihren isogenen Wildtypstämmen zu den inneren Organen
erfolgte über die Bestimmung der bakteriellen Keimlast. In diesem Zusammenhang
wurde auch untersucht, ob das veränderte (murinisierte) Internalin A einen Einfluss
auf die Überquerung der Blut-Hirn-Schranke hat und ob die Interaktion von Internalin
A mit E-Cadherin für das Eindringen von Listerien in das zentrale Nervensystem
wichtig sein könnte.
3.1 Der Infektionsverlauf nach einer Listerieninfektion ist von der
Virulenz des Listeriensubtypes abhängig
Das fakultativ intrazelluläre Bakterium Listeria monocytogenes ist in der Umwelt weit
verbreitet und kann schwerwiegende Infektionen auslösen. Innerhalb von Listeria
monocytogenes sind eine Reihe von Serotypen charakterisiert, welche sich
genetisch unterscheiden (Piffaretti et al., 1989) sowie unterschiedliche
Virulenzpotentiale aufzeigen eine Erkrankung auszulösen (Buchrieser et al., 1993;
Jacquet et al., 2004; Pohl et al., 2006). Über den Nachweis der Oberflächen-
Antigene und Flagellen-Antigene konnten 13 verschiedene Serotypen bestimmt
werden (Seeliger und Höhne, 1979), welche in drei phylogenetische Linien
unterschieden werden. Mit Hilfe der Multi-Locus-Enzym-Elektrophorese und
Pulsfeldgelelektrophorese konnten zwei Hauptlinien identifiziert werden (Brosch et
al., 1994; Graves et al., 1994; Piffaretti et al., 1989; Rasmussen et al., 1991). Eine

Diskussion
99
dritte Linie konnte über die Verwendung von Ribotypisierung und DNA-Arrays
charakterisiert werden (Doumith et al., 2004; Wiedmann et al., 1997b). Linie I
beinhaltet die Isolate der Serotypen 4b, 1/2b, 3b, 4d und 4e, Linie II die
Listerienisolate vom Serotyp 1/2a, 1/2c, 3a und 3c und in Linie III werden die Isolate
4a und 4c eingeordnet. Mittels Multi-Locus-Sequenz-Typisierung (MLST), eine
Methode die Sequenzvariationen in Genfragmenten von Haushaltsgenen analysiert,
können Allelprofile verschiedener Bakterienisolate bestimmt und für
Verwandtschaftsanalysen genutzt werden (Maiden et al., 1998; Salcedo et al., 2003).
Über die Verwendung von MLST zur Analyse der verschiedenen Listeria mono-
cytogenes Isolate wurde die phylogenetische Verwandtschaft verschiedener klonaler
Listerienkomplexe, der evolutionären Ursprung der Serotypen sowie Internalin A
Polymorphismen beschrieben (Ragon et al., 2008). Für epidemiologische Studien
von Listerien-Infektionsausbrüchen in Mensch und Tier ist es sehr wichtig, dass
Genotypen verschiedener Substämme mit dem pathologischen Erscheinungsbild und
den Schweregrad von Erkrankungen korreliert werden können (Buchrieser, 2007;
Chen et al., 2007).
Die im Verlaufe dieser Arbeit durchgeführten oralen Infektionen von BALB/cJ Mäuse
mit je einer Infektionsdosis von 5x109 CFU LmoXen32-mur bzw. LmoEGDe-lux-mur,
resultierten in Unterschieden verschiedener analysierter Parameter wie der Über-
lebensrate, dem Gewichtsverlauf nach der Infektion, der bakteriellen Keimlast, der
Histopathologie und dem BLI. Nach der Infektion mit LmoXen32-mur zeigte sich ein
stark abgeschwächter Infektionsverlauf in BALB/cJ Mäusen, was im Vergleich zu
LmoEGDe-mur-lux infizierten Tieren in einer geringeren Mortalitätsrate resultierte
(Abschnitt 2.4). Weiterhin zeigte sich die attenuierte Virulenz der Stämme LmoXen32
und LmoXen32-mur im Nachweis einer geringeren bakteriellen Keimlast in den
inneren Organe Leber, Milz, Dünndarm und mesenterischen Lymphknoten (mLK)
(Abschnitt 2.2.2 und Abschnitt 2.3.4). Die histopathologische Analyse ergab nach der
Infektion mit LmoXen32 und LmoXen32-mur nur moderate Schädigungen des
Leberparenchyms mit einer nachweislich geringeren Anzahl an entzündlichen
Läsionen und weniger Infiltrationen von Granulozyten im Vergleich zu einer Infektion
mit LmoEGDe-lux und LmoEGDe-mur-lux (Abschnitt 2.2.2 und Abschnitt 2.3.2).

Diskussion
100
Zudem zeigten LmoXen32-mur eine geringere Invasionseffizienz in die murinen
Kolonkarzinom CT-26 Zellen im Vergleich zu LmoEGDe-mur (Abschnitt 2.1.4).
Obwohl beide Stämme, LmoXen32 (Serotyp 1/2c; Derivat von Lmo 10403S, Serotyp
1/2a) und LmoEGDe-lux (LmoEGDe, Serotyp 1/2a) in die gleiche phylogenetische
Linie II eingeordnet werden, weisen sie jedoch unterschiedliche Virulenzen auf
(Ragon et al., 2008). Eine Ursache für diese Pathogenitätsunterschiede liegt in der
Integrationsstelle des Lux-Operons innerhalb des Listeria Genoms von LmoXen32
und LmoEGDe-lux. Die Integration des synthetischen Operons luxABCDE erfolgte
bei LmoEGDe-lux in die 3´Region eines tRNAArg Gens (lmot17) mit Hilfe des
Integrationsvektors pPL2lux (Bron et al., 2006; Riedel et al., 2007). Der Vektor
pPL2lux ist ein Derivat des Integrationsvektors pPL2. Dieser kann bei verschiedenen
Listeria monocytogenes Stämmen unabhängig von ihrem Serotyp eingesetzt werden
(Lauer et al., 2002). Der Vektor pPL2 enthält ein PSA (Phage aus LmoEGDe ScottA)
Listeriophagen Integrase-Gen sowie eine DNA-Erkennungssequenz für die
zielgerichtete Integration ins listerielle Chromosom. Innerhalb der 3`Region des
tRNAArg Gens (lmot17) sowie in der Listeriophagensequenz des pPL2 Vektors sind
17 bp eines Sequenzabschnitt identisch, über welche die Integration erfolgt. Nach
erfolgter Insertion ist ein Teil der tRNAArg Gensequenz unterbrochen. Diese wird
jedoch über die identische 17 bp lange PSA-Sequenz komplettiert und somit das
Leseraster und damit die volle Funktionsfähigkeit des tRNAArg Gens beibehalten
(Lauer et al., 2002). Des Weiteren codieren 67 Gene im Genom von Listeria
monocytogenes für tRNAs, davon fünf Gene für tRNAArg Moleküle (Glaser et al.,
2001). Die genetische Modifikation von LmoEGDe zu LmoEGDe-lux sollte daher
keinen Einfluss auf die Expression von tRNAArg haben und die Pathogenität von
LmoEGDe-lux nicht beeinträchtigen. Der Vorteil des auf Phagen basierenden
Integrationsvektors besteht in der Integration einer einzelnen Kopie ins Genom sowie
in dessen Stabilität in Abwesenheit von Selektionsmarkern.
LmoEGDe-mur-lux wurde ausgehend von murinisierten Listerienstamm LmoEGDe-
mur (Monk et al., 2010) über die Integration des Lux-Operons mit Hilfe des Listerio-
phagen Integrationsvektors pIMK2lux generiert. Das Plasmid pIMK2lux ist ein Derivat
des Integrationsvektors pPL2 (siehe oben), mit reduzierter Plasmidgröße und

Diskussion
101
Kanamycinresistenz (Monk et al., 2008). Wie schon bei der Generierung von
LmoEGDe-lux erfolgt die Integration des Operons luxABCDE ins tRNAArg Gen
(lmot17) von Listeria monocytogenes ohne Einschränkung der Genfunktion und sollte
die Virulenz von LmoEGDe-mur-lux ebenfalls nicht beeinflussen. Weiterhin konnte
gezeigt werden, dass die Luziferaseaktivität von LmoEGDe-lux (Integration mittels
pPL2lux) äquivalent zur Aktivität von LmoEGDe-mur-lux (Integration mittels
pIMK2lux) ist und somit beide Stämme eine vergleichbare Biolumineszenz-
signalstärke erzeugen (Monk et al., 2008).
Im Gegensatz zu den LmoEGDe Stämmen wurden bei der Generierung von
LmoXen32 das Luziferase-Operon mittels Transposonmutagenese zufällig in die
Promotorregion des Flagellin A (flaA) Gens integriert (Hardy et al., 2004). Diese
Modifikation resultierte im Vergleich zum Ursprungsstamm Lmo 10403S in einer
vierfach höheren LD50 des LmoXen32 im Mausinfektionsmodell. Bereits publizierte
Arbeiten konnten zeigen, dass die Expression von flaA in LmoEGDe bei 37°C verrin-
gert ist und Flagellenstrukturen nicht nachweisbar waren (Gründling et al., 2004).
Dagegen zeigten 2% von Lmo 10403S (Ausgangsstamm für LmoXen32) Flagellen
auf der Oberfläche und es konnte außerdem eine Expression von flaA Transkripten
bei Temperaturen von 37°C nachgewiesen werden (Gründling et al., 2004). Andere
Studien bestätigten eine Mobilität von Lmo 10403S bei höheren Temperaturen (Way
et al., 2004). Trotz Repression der Expression von flaA bei 37°C ist Flagellin A zu
Beginn einer Infektion während des extrazellulären Wachstums und der intra-
zellulären Infektion von Bedeutung (Peel et al., 1988). Bewegliche Listerien besitzen
bei der initialen Kolonisation von Dünndarm und Leber im Vergleich zu unbe-
weglichen Listerien einen Vorteil für die optimale Invasion in Zellen (O’Neil und
Marquis, 2006). Dabei spielt die Fähigkeit zur Mobilität eine entscheidende Rolle. Es
konnte gezeigt werden, dass flagellentragende aber unbewegliche Bakterien keine
verstärkte Adhärenz bzw. Invasion in humane Epithelzellen aufweisen als
flagellenlose Listerien (O’Neil und Marquis, 2006). Dies weist darauf hin, dass Fla-
gellen eine geringe Funktion als Adhäsine während der Invasion haben, jedoch die
Mobilität ein entscheidender Faktor für die Invasion in humane Zellkulturen ist. Somit
könnte die Integration des Lux-Operons in das flaA Gen von LmoXen32 zu einer

Diskussion
102
Funktionseinschränkung der Flagellen führen und die sich daraus resultierende
Mobilitätsminderung in einer geringeren Virulenz von LmoXen32 auswirken.
3.2 Die Murinisierung des Internalin A resultiert in einem invasiveren
Krankheitsverlauf
Die Fähigkeit epitheliale Barrieren zu überqueren, ist eine wichtige Virulenzeigen-
schaft von Listeria monocytogenes. Viele Studien konnten die Bedeutung der Ober-
flächenproteine InlA und InlB während der zellulären Invasion und der Passage
epithelialer Barrieren aufschlüsseln (Disson et al., 2008; Khelef et al., 2006).
Internalin A weist in Abhängigkeit seines Rezeptors E-Cadherin eine
Speziesspezifität auf, wie schon in Abschnitt 1.5.1 beschrieben (Lecuit et al., 1999).
Die Überquerung der intestinalen Barriere kann Internalin A abhängig über die
Invasion der Enterozyten erfolgen und Internalin A unabhängig über die M-Zellen der
Peyer´schen Plaque (Lecuit et al., 2001; Marco et al., 1997). Die InlA-abhängige
Überquerung der Darmbarriere erfolgt über die Interaktion mit E-Cadherin, welches
an der basolateralen Seite der intestinalen Epithelzellen exprimiert wird. Somit ist der
Zugang für Listeria monocytogenes zum E-Cadherin erschwert (Boller et al., 1985,
Sousa et al., 2005). Jedoch kann kurzzeitig E-Cadherin an der Darmlumenseite
zugängig sein. Während des Zellzyklus und natürlicher Apoptose der Enterozyten an
den Villi erfolgt die Extrusion abgestorbener Zellen und eine Exposition von E-
Cadherin an der apikalen Seite der Darmvilli (Pentecost et al., 2006; Pentecost et al.,
2010). Ebenso wurde beschrieben, dass an Epithelfalten der Villi sowie um
schleimproduzierende Becherzellen E-Cadherin frei zugängig ist und somit Listeria
monocytogenes die Interaktion von InlA mit E-Cadherin ermöglicht (Nikitas et al.,
2011).
In vorangegangen Arbeiten erfolgte die orale Infektion von Mäusen ohne Berück-
sichtigung einer intakten E-Cadherin - Internalin A - Interaktion (Czuprynski et al.,
2002; Czuprynski et al., 2003; Daniels et al., 2000; LeMonnier et al., 2006). Studien
unter Verwendung anderer Tiermodelle, wie dem Meerschweinchen, konnten die
Bedeutung des InlA während einer Infektion beschreiben. Aufgrund der nicht

Diskussion
103
funktionellen InlB-c-Met-Interaktion im Meerschweinchen konnten Mechanismen
listerialer Invasion in diesem Tiermodell jedoch nur begrenzt untersucht werden
(Bakardjiev et al., 2006). Mit der Murinisierung des listeriellen Internalin A durch den
Austausch der Aminosäuren Ser192Asn und Tyr369Ser konnte die Affinität des
Internalin A zu murinen E-Cadherin gesteigert werden (Wollert et al., 2007b). Zur
Etablierung eines oralen Infektionsmodells erfolgte in dieser Arbeit die ausführliche
Analyse der Infektionsprozesse von LmoEGDe-lux im Vergleich zu LmoEGDe-mur-
lux bei oral infizierten Mäusen. Mit Hilfe der erzielten Resultate lassen sich Aussagen
über die Verbreitung von Listerien in Abhängigkeit von funktionalem Internalin A
treffen. Die InlA-abhängige Invasion in Enterozyten und somit die Überquerung der
intestinalen Barriere ermöglicht den schnellen und direkten Zugang zum Lymph- und
Blutsystem und führt folglich zu einer schnelleren Dissemination des Pathogens in
die Leber. Bezugnehmend auf die Ergebnisse von Wollert et al. resultiert die
Murinisierung des Internalin A in einer 6700-fachen Erhöhung der Affinität der
Proteinkomplexe von Internalin A zu humanem E-Cadherin (Wollert et al., 2007b).
Dagegen zeigten die murinisierten Listerien in vitro nur eine Verdopplung der
Invasionsrate in die humanen intestinalen Colonkarzinom-Zellen (Caco-2) (Wollert et
al., 2007b).
Die Ergebnisse der vorliegenden Arbeit ergaben ebenfalls nur eine geringe
Steigerung der Invasionsrate der murinisierten Listerienstämme LmoXen32-mur,
LmoEGDe-mur und LmoEGDe-mur-lux im Vergleich zu ihren nicht-murinisierten
isogenen Wildtypstämmen LmoXen32, LmoEGDe und LmoEGDe-lux in Caco-2
Zellen (Abschnitt 2.1.4). Dagegen resultierte die Infektion von murinen
Colonkarzinom-Zellen (CT-26) in einer signifikanten Erhöhung und damit in einer
gesteigerten Invasionsrate der murinisierten Listerienstämme LmoEGDe-mur und
LmoEGDe-mur-lux, im Vergleich zu ihren isogenen Wildtypstämmen. Dies bestätigt
die gesteigerte Affinität des murinisierten Internalin A zu dem murinen E-Cadherin,
wie von Wollert et al. anhand der Dissoziationskonstanten der Proteinkomplexen
nachgewiesen werden konnte (Wollert et al., 2007b). Die Murinisierung des
Listerienstamms LO28 resultierte sowohl in humanen als auch in murinen Colon-
karzinom-Zellen in einer signifikanten Erhöhung der Invasionsrate. Aufgrund einer

Diskussion
104
„nonsense“ Mutation im inlA Gen exprimiert Listeria monocytogenes LO28 ein
verkürztes Internalin A (Jonquieres et al., 1998). Folglich resultierte die Infektion von
epithelialen Zelllinien in einer geringeren Invasionsrate von LO28 im Vergleich zu
anderen Listerienstämmen, welche funktionsfähiges Internalin A exprimieren
(Roldgaard et al., 2009). LO28 exprimiert InlA nicht auf der Zellwandoberfläche,
sondern sekretiert es unter Zellkulturbedingungen in den Zellüberstand (Jonquieres
et al., 1998). Durch die Murinisierung des LO28-Stammes erfolgt der Austausch des
verkürzten InlA mit murinisierten funktionsfähigen InlA über homologe Rekombination
des kompletten inlA-Locus. Dies führt zu einem Funktionszugewinn („gain of
function“), welcher in einer signifikant höheren Invasionsrate des LO28-mur resultiert
und im in vitro Invasions-Assay erkennbar war.
Im in vivo Mausmodell resultierte die Infektion ausgewählter Mausinzuchtstämme mit
LmoEGDe-mur-lux in einer höheren Mortalitätsrate als die Infektion mit LmoEGDe-
lux (Abschnitt 2.3.3). Über die Änderungen im Verlauf des Körpergewichtes konnten
signifikante Unterschiede zwischen murinisierten und den entsprechenden nicht-
murinisierten Listerien beobachtet werden, welche in einer größeren Änderung des
Gewichtsverlusts bei A/JOlaHsd, BALB/cJ und C57BL/6J Mäusen nach der oralen
Infektion mit LmoEGDe-mur-lux resultierte. Weiterhin zeigte die Analyse der
bakteriellen Keimlast der inneren Organe nach der Infektion mit murinisierten
Listerien eine zehn- bis hundertfach höhere Anzahl von Bakterien im Vergleich zu
der Infektion mit nicht-murinisierten Listerien (Abschnitt 2.3.4.2). In BALB/cJ Mäusen
konnte an Tag drei nach der Infektion mit LmoEGDe-mur-lux zudem eine signifikante
Erhöhung der Listerienanzahl in der Leber im Vergleich zu LmoEGDe-lux
nachgewiesen werden, was die Ergebnisse von Monk et al. sowie die publizierten
Resultate im humanisierten Mausmodell von Lecuit et al. bestätigte (Lecuit et al.,
2001; Monk et al., 2010). Der in dieser Arbeit durchgeführte immunhistologische
Nachweis von Listerien konnte nach der Infektion mit LmoEGDe-mur-lux im
Vergleich zu LmoEGDe-lux am Tag vier nach der Infektion vermehrte Infiltrationen
von Listerien und Granulozyten im Leberparenchym nachweisen (Abschnitt 2.3.2).
Diese Ergebnisse zeigten, dass Infektionen mit InlA-murinisierten Listerien in einem
verstärkten Krankheitsverlauf resultierten. Somit lässt sich weiter schlussfolgern,

Diskussion
105
dass die Murinisierung des Internalin A mit einer gesteigerten Invasion der Organe
einhergeht. Auch konnte mit Hilfe des in vivo Biolumineszenz-Imaging gezeigt
werden, dass die Infektion mit LmoEGDe-mur-lux in einer verstärkten Invasion der
inneren Organe, speziell in der Leber, resultierte (Abschnitt 2.3.3 und Abschnitt
2.3.4.3). Diese Ergebnisse sind vergleichbar mit den Resultaten von Monk et al. in
welchen ebenfalls eine Akkumulation von murinisierten Listerien in der Leber
festgestellt werden konnte (Monk et al., 2010). Aufgrund der Murinisierung des
Internalin A erfolgt die verstärkte Infektion von intestinalen Epithelzellen. Murinisierte
Listerien gelangen somit in einer größeren Anzahl über die Darmbarriere ins Blut-
und Lymphsystem und führen zu einer vermehrten Invasion der inneren Organe. In
oral infizierten Meerschweinchen konnte gezeigt werden, dass die Ausbreitung der
Listerien zur Leber direkt über den Dünndarm und weiter über das Pfortadersystem
erfolgen kann sowie indirekt über die mLK und die anschließende lymphatische und
hämatogene Dissemination (Melton-Witt et al., 2011). Somit ist die Anzahl der
Listerien, welche zu Leber gelangen prozentual höher, als im Vergleich zu den
weiteren inneren Organen Milz und Gehirn, welche nur über den indirekten
Disseminationsweg besiedelt werden. Jedoch zeigte die intravenöse Infektion von
C57BL/6J Mäusen, dass in der frühen Phase der Infektion Listeria monocytogenes
innerhalb von CD8α+ dendritische Zellen vom Darm zu der Milz transportiert werden
(Neuenhahn et al., 2006) und diese damit an der frühen Dissemination von Listerien
involviert sind.
All diese Resultate konnten die Bedeutung der Internalin A - E-Cadherin - Interaktion
während der Überquerung der intestinalen Barriere bestätigen. Für die weitere
Dissemination und die systemische Infektion im weiteren Krankheitsverlauf sind
weitere Faktoren von Bedeutung. Die Ausbreitung von Zelle zu Zelle erfolgt unter
Beteiligung weiterer Virulenzfaktoren. Internalin C (InlC) und das Aktin-bindende
Protein A (ActA) sind für die Zell-Zell-Ausbreitung von Wichtigkeit. So zeigten sich
jeweilige Deletionsmutanten ineffizient in der Ausbildung von
Membranausstülpungen zur Invasion benachbarter Zellen in vitro (Domann et al.,
1992; Rajabian et al., 2009). Auch unterstützt das Virulenzprotein Vip über die
Interaktion mit seinem Rezeptor Gp-96 („endoplasmatische Retikulum ansässiges

Diskussion
106
Chaperon“) die Invasion in Säugetierzellen (Cabenes et al., 2002). Listerielle
Deletionsmutanten für Vip zeigten eine zehnfach geringere Invasionsrate in Caco-2
Zellen als die isogenen Wildtypstämme. Ebenfalls spielt Phospholipase C (PlcB) bei
der Zell-Zell-Ausbreitung eine wichtige Rolle. Listeria monocytogenes befreit sich
durch die Sekretion von PlcB aus einer Doppelmembranvakuole als Voraussetzung
für die intrazelluläre Replikation. Nach intrazerebraler Injektion von Listeria
monocytogenes bei Mäusen konnte die Bedeutung von PlcB bei der Zell-Zell-
Ausbreitung demonstriert werden. Deletionsmutanten für PlcB wiesen im Vergleich
zum Wildtyp einen weniger virulenten Krankheitsverlauf auf (Schlüter et al., 1998).
Ein weiterer Faktor für die systemische Ausbreitung von Listerien ist InlB, welcher
über die Interaktion mit dem c-Met-Rezeptor die Internalisation in eine Vielzahl von
Säugetierzellen induziert (Braun et al., 1998).
Neben Internalin A vermittelter Invasion von Epithelzellen können Listerien InlA-
unabhängig über M-Zell-vermittelte Phagozytose die intestinale Barriere überqueren.
M-Zellen sind innerhalb des Follikel-assozierten Epithels (FAE) eingebettet, welches
die lymphoiden Follikel (Peyer´sche Plaque) der Darmmukosa überdeckt (Clark und
Jepson, 2003). Sie sind somit Bestandteile des „Mukosa-assoziiertes lymphatisches
Gewebe“ (MALT) (Kraehenbuhl und Neutra, 2000). M-Zellen sind auf transepithelia-
len Antigentransport spezialisiert. Sie sind charakterisiert über das Vorhandensein
unregelmäßiger Mikrovilli auf der Zelloberfläche sowie finden sich an der
basolateralen Seite zytoplasmatische Invaginationen, welche Lymphozyten und
Makrophagen enthalten (Clark und Jepson, 2003). Aufgrund der reduzierten Anzahl
der Mikrovilli und einer vorhandenen Glykokalyx auf der Zelloberfläche der M-Zellen
erfolgt die Exposition zu den im Darmlumen vorliegenden Antigenen. M-Zellen
können Makromoleküle und Mikroorganismen über Phagozytose, Endozytose und
rezeptorvermittelte Endozytose aufnehmen (Clark und Jepson, 2003). Nach
Aufnahme in die M-Zellen werden Antigene und Bakterien aktiv vom intestinalen
Lumen zu den lymphoiden Zellen der intraepithelialen Zellinvagination transportiert.
Der weitere Transport der Pathogene erfolgt innerhalb von Makrophagen zu
Antigenpräsentierenden Zellen im germinalen Zentrum der lymphoiden Follikel
(Sansonetti und Phalipon, 1999). Durch diesen Prozess kann die Induktion der

Diskussion
107
spezifischen und adaptiven Immunantwort erfolgen. Die Rolle der M-Zellen bei der
Überquerung der intestinalen Barriere ist noch nicht vollständig geklärt. So zeigten
Untersuchungen, dass Listerien auch in Abwesenheit von intakten Peyer´schen
Plaque das Intestinum durchqueren können (Havell et al., 1999). Einen Tag nach der
intragastralen Infektion von immundefizienten SCID Mäusen wurden Listerien in den
Mesenterien, der Milz und der Leber beobachtet (Havell et al., 1999). Jedoch konnte
bei Wollert et al. gezeigt werden, dass Listeria monocytogenes sehr wohl Internalin A
unabhängig die Darmbarriere durchqueren können. So wurden in den Peyer´schen
Plaque von C57BL/6J Mäusen nach oraler Infektion von nicht-murinisierten Listerien
gleichermaßen Listerien nachgewiesen wie nach der Infektion von murinisierten
Listerien (Wollert et al., 2007b).
Die in dieser Arbeit durchgeführten Analysen zur bakteriellen Keimlast bestätigen die
InlA-unabhängige Durchquerung der intestinalen Barriere, da nicht-murinisierte
Listerien bereits einen Tag nach der oralen Infektion in den inneren Organen Leber,
Milz, mLK, Dünndarm und Gehirn nachgewiesen werden konnten (Daten nicht
gezeigt). Dies deutet darauf hin, dass nicht nur Enterozyten als Haupteintrittspforte
dienen. Corr et al., konnten mit Hilfe eines in vitro Zellkulturmodells, welches die M-
Zell-Aktivität imitiert, nachweisen, dass Listeria monocytogenes vermehrt über M-
Zellen transportiert werden. In diesem Modell erfolgte eine Kombinationszellkultur
von murinen Peyer'schen Plaque Lymphozyten mit C2Bbe1 epithelialen Enterozyten
(Derivate von Caco-2 Zellen), welche daraufhin einen M-Zell-Phänotyp entwickelten
(Corr et al., 2006). Typische Eigenschaften der M-Zellen sind ein unregelmäßiger
Mikrovillisaum, die Präsenz der basolateralen Zellinvagination sowie die Expression
des aktinassoziierten Protein Villin. Die Infektion dieses Zellkulturmodells mit Listeria
monocytogenes zeigte, dass die Listerien vermehrt über diese M-Zellen InlA-
unabhängig transportiert werden als im Vergleich zu C2Bbe1 Monozellkulturen (Corr
et al., 2006).
Auch andere Pathogene nutzen die M-Zellen als Route für die intestinale Invasion.
Neuere Forschungsergebnisse konnten zeigen, dass auch andere Gram-negative
Pathogene wie Yersinia enterocolitica und pseudotuberculosis, Shigella flexneri und
verschiedene Salmonella Pathovare wie Salmonella enterica Serovar Typhimurium

Diskussion
108
M-Zellen als Eintrittspforte in den Organismus nutzen (Clark und Jepson, 2001;
Grützkau et al., 1990; Hanski et al., 1989; Jenson et al., 1998; Jepson und Clark,
2001; Sansonetti und Phalipon, 1999; Sansonetti, 2001). Die Invasion in M-Zellen
von Salmonella typhimurium und Shigella flexneri erfolgt über einen Trigger-
Mechanismus, vermittelt über das Typ-III-Sekretionssystem (Cossart und Sansonetti,
2004). Die Infektion mit Salmonellen führt innerhalb von 30 Minuten zu massiven
Änderungen in der Aktinpolymerisation des Zytoskeletts der M-Zellen und zur Auf-
nahme der Salmonellen. Dies hat letztendlich die Zerstörung der M-Zellen zur Folge
und resultiert in einer ausgeprägten Schädigung des FAE (Jensen et al., 1998).
Dagegen führt die Infektion mit Shigellen indirekt durch Induktion einer massiven
inflammatorischen Antwort zur Schädigung des intestinalen Epithels (Sansonetti et
al., 1996). Die Infektion muriner M-Zellen mit Yersinia pseudotuberculosis führt
dagegen nur zu geringen Veränderungen des M-Zell-Aktins. Die Invasion der M-
Zellen erfolgt über einen Zipper-ähnlichen Mechanismus an dem mehrere Yersinien
beteiligt sind, welche dann in einer Vielzahl intrazellulär im Zytoplasma vorliegen
(Clark und Jepson, 2001). In vitro Studien konnten zeigen, dass Yersinia Spezies
über das Zelloberflächenmolekül β1-Integrin binden und ihre eigene Aufnahme
induzieren (Isberg und Leong, 1990).
Zusammenfassend betrachtet, erlaubt die Murinisierung des Internalin A die Invasion
der murinen intestinalen Epithelzellen über die Interaktion mit E-Cadherin und somit
eine systemische Infektion im Mausmodell über die orale Infektionsroute verbunden
mit klinischen Symptomen einer Listeriose. Die in dieser Arbeit durchgeführten
Analysen bestätigen, dass Listeria monocytogenes InlA-abhängig über die Entero-
zyten und zusätzlich InlA-unabhängig über die M-Zellen die intestinale Barriere über-
queren kann.
3.3 Der Infektionsverlauf nach einer Listerieninfektion ist abhängig vom
genetischen Hintergrund des Wirtes
Infektionskrankheiten gehören weltweit, neben Herz-Kreislauf-Erkrankungen, immer
noch zu den Erkrankungen mit der höchsten Mortalität (Daten 2008, The World

Diskussion
109
Health Report, 2011). Welchen Verlauf eine Infektion nimmt, hängt zum einen von
verschiedenen äußeren Einflüssen ab wie zum Beispiel der Umwelt und dem Patho-
gen selbst, zum anderen spielen eine Reihe innerer Faktoren eine Rolle, wie die
genetisch bedingte Prädisposition eines Wirtes, an einer Infektion zu erkranken. Die
genetische Prädisposition ist eine der wichtigsten intrinsischen Faktoren und resul-
tiert in verschiedenen Variationen des Phänotyps einer Erkrankung. Die große
Herausforderung der Infektionsforschung liegt daher in der Etablierung eines
passenden Tiermodells für das Verständnis von Wirt-Pathogen-Beziehungen. Die
Maus ist häufig Tiermodell der ersten Wahl. Sie hat den Vorteil, dass eine Vielzahl an
gut charakterisierten Mausinzuchtstämmen existieren, sie eine kurze Generations-
folge aufweisen und die Haltung relativ unkompliziert erfolgen kann. Auch erlaubt die
große genetische Diversität von Mausinzuchtstämmen die Untersuchung komplexer
Genotyp - Phänotyp Beziehungen und die Analyse der genetischen Krankheits-
prädisposition im Tiermodell.
Im Rahmen dieser Arbeit konnten Unterschiede in den Resistenzen der Mausin-
zuchtstämme C3HeB/FeJ, A/JOlaHsd, BALB/cJ und C57BL/6J nach der oralen
Infektion mit Listeria monocytogenes aufgezeigt werden. Wie über die Analyse der
Überlebensrate erkennbar (Abschnitt 2.3.3) zeigten C3HeB/FeJ Mäuse eine ausge-
prägte Suszeptibilität gegenüber der oralen Infektion mit Listeria monocytogenes,
was in einer reduzierten Überlebensrate resultierte. Bei A/JOlaHsd und BALB/cJ
Mäusen konnte eine intermediäre Resistenz beobachtet werden. Dagegen erwiesen
sich C57BL/6J Mäuse als äußerst resistent und eine Überlebensrate von 90-100%
konnte beobachtet werden.
Die Ursachen einer gesteigerten Suszeptibilität gegenüber einer Listerieninfektion
liegen zum einen in der Eigenschaft von Listeria monocytogenes sich der
Immunantwort aufgrund der intrazellulären Lebensweise zu entziehen und zum
anderen in der Fähigkeit des Immunsystems intrazelluläre Listerien zu erkennen und
zu eliminieren. Somit resultiert der genetische Hintergrund von C3HeB/FeJ,
A/JOlaHsd, BALB/cJ und C57BL/6J Mäusen nicht nur in Variationen des
biochemischen und metabolischen Phänotyps (Champy et al., 2008), sondern hat

Diskussion
110
auch Einfluss auf die Immunantwort nach oraler Infektion mit Listeria mono-
cytogenes.
In der Vergangenheit konnten mit Hilfe von Listeria monocytogenes Infektionen
wichtige Erkenntnisse über die angeborene und erworbene Immunantwort erzielt
werden, wie der Nachweis der zellulär vermittelten Immunantwort nach erfolgter
Listerieninfektionen (Mackaness, 1962; Mackaness, 1969; Pamer, 2004). Die
Infektion verschiedener Mausinzuchtstämme mit Listeria monocytogenes resultierte
des Weiteren in unterschiedlicher Resistenz und Suszeptibilität in Abhängigkeit des
gewählten Mausinzuchtstammes (Cheers et al., 1978; Czuprynski et al., 1985;
Mainou-Fowler et al., 1988) und führte zur Identifizierung von Faktoren für die
genetische Kontrolle der Immunantwort im Verlauf einer Listerieninfektion (Cheers
und McKenzie, 1978; Gervais et al., 1984; Skaneme et al., 1979).
Die in dieser Arbeit durchgeführten Experimente zeigten, dass C57BL/6J Mäuse
resistent gegenüber einer oralen Infektion mit dem murinisierten Listerienstamm
LmoEGDe-mur-lux reagierten. Die ausgeprägte Resistenz von C57BL/6J Mäusen
konnte schon gegenüber der intravenösen oder intraperitonealen Listerieninfektion
beobachtet werden (Cheers et al,. 1978; Cheers und McKenzie, 1978; Czuprynski et
al., 1985; Mainou-Fowler et al., 1988). Wie im Abschnitt 2.3.3 ausführlich
beschrieben, zeigten C57BL/6J Mäuse nach der oralen Listerieninfektion die
geringste Veränderung im Körpergewichtsverlauf. Auch war mit Hilfe des BLI der
ausgewählten Mausinzuchtstämme erkennbar, dass die Infektion mit LmoEGDe-mur-
lux in den resistenteren C57BL/6J Mäusen zu einer Akkumulation der Listerien in der
Gallenblase führte. Dagegen zeigten A/JOlaHsd und BALB/cJ Mäuse nach der
Infektion mit LmoEGDe-mur-lux eine verstärkte Ansammlung der Listerien in der
Leber und den mLK (Abschnitt 2.3.4.2). A/JOlaHsd Mäuse reagierten zudem
empfindlicher als BALB/cJ Mäuse, was in einer geringeren Überlebensrate resul-
tierte. Frühere Arbeiten konnten zeigen, dass die intravenöse Infektion von
A/JOlaHsd Mäusen mit LmoEGDe in einer hundertfach höheren bakteriellen Keimlast
in der Milz und Leber resultierte sowie in einer drastisch reduzierten Anzahl der
peripheren Monozyten 24 Stunden nach der Infektion im Vergleich zu C57BL/6J
Mäusen (Sadarangani et al., 1980).

Diskussion
111
Die Infektion mit Listeria monocytogenes induziert frühzeitig eine Monozytose
(Murray et al., 1926). Molekulares Material von geringem Molekulargewicht, welches
mit der bakteriellen Zellwand assoziiert ist, wie Peptidoglykan und Lipoteinonsäuren,
führt 48 Stunden nach der Infektion zu einer drei- bis sechsfachen Erhöhung der
Monozyten im peripheren Blut (Galsworthy and Fewster, 1988; Paquet et al., 1986).
Diese Monozytose, aufgrund der inflammatorischen Immunantwort, vermittelt eine
Resistenz gegenüber der Listerieninfektion. Bei A/JOlaHsd Mäusen konnte jedoch
keine Erhöhung der Monozytenanzahl nach der intraperitonealen Injektion mit dem
eine Monozytose induzierenden Zellwandextrakt von Listeria monocytogenes
nachgewiesen werden (Galsworthy and Fewster, 1988).
Eine weitere mögliche Ursache für die gesteigerte Suszeptibilität der A/JOlaHsd
gegenüber der C57BL/6J Mäuse wird der reduzierten Menge an Gamma-Interferon
(IFN-γ) und des Granulozyten-Makrophagen Kolonie-stimulierenden Faktors (GM-
CSF) zugeschrieben (Iizawa et al., 1993). IFN-γ spielt eine Rolle bei der
angeborenen sowie erworbenen Immunantwort und dient der Makro-
phagenaktivierung. GM-CSF stimuliert die Reifung von Monozyten und
Granulozyten. In vitro konnte beobachtet werden, dass Makrophagen in Gegenwart
von IFN-γ effizienter bei der Eliminierung intrazellulärer Listerien reagieren (Portnoy
et al., 1989). Frühere Studien zeigten, dass eine verstärkte Infiltration von
neutrophilen Granulozyten und Monozyten im infizierten Gewebe die Resistenz
gegenüber Listeria monocytogenes erhöhen. Bei A/J Mäusen konnte diesbezüglich
eine geringere Infiltration von Phagozyten im infizierten Gewebe nach intra-
peritonealer Infektion mit LmoEGDe nachgewiesen werden, als im Vergleich zu
C57BL/6J Mäusen (Czuprynski et al., 1985). Die verminderte Sekretion von GM-SCF
und IFN-γ kann somit die Empfindlichkeit von A/JOlaHsd Mäusen gegenüber einer
Listerieninfektion erhöhen.
Weiterhin wurden A/J Mäuse im Gegensatz zu C57BL/6J Mäuse als defizient für die
C5 Komponente des Komplementsystems charakterisiert. Eine 2 bp Deletion des Hc
(hämolytisches Komplement) Gens auf Chromosom 2 resultiert in einer
Verschiebung des Leserasters und damit in einem defizienten Protein, was in einer
Störung der Rekrutierung von inflammatorischen Zellen führt (Cheers und McKenzie,

Diskussion
112
1978; Gervais et al., 1984; Czuprynski et al., 1985). Der C5-Komplementfaktor ist
Bestandteil des Membranangriffskomplexes und führt zusammen mit weiteren
Faktoren des Komplementsystems zur Formation einer Pore und zur anschließenden
Lyse der Bakterienzelle. Die C5-Defizienz kann daher in einer weniger effektiven
Immunantwort und einer gesteigerten Suszeptibilität der A/JOlaHsd Mäuse in der
fortgeschrittenen Phase der Infektion resultieren.
Somit konnte gezeigt werden, dass genetische Defekte von wichtigen Komponenten
des Immunsystems, wie die C5-Defizienz, die Reduktion von GM-CSF und IFN-γ
sowie eine reduzierte Monozytose zur erhöhten Suszeptibilität der A/JOlaHsd Mäuse
beitragen.
BALB/cJ Mäuse sind im Gegensatz zu A/JOlaHsd Mäusen C5 suffizient. Jedoch
ergab sich aus den hier durchgeführten Untersuchungen, dass BALB/cJ Mäuse, im
Vergleich zu C57BL/6J Mäusen, ebenfalls empfindlich auf eine Listerieninfektion
reagierten. So zeigten BALB/cJ Mäuse eine geringere Überlebensrate als C57BL/6J
Mäuse und größere Gewichtsverluste im Verlauf der Infektion (Abschnitt 2.3.3).
Ebenfalls konnte über BLI in vivo und ex vivo eine vermehrte Detektion von
murinisierten Listerien nach der oralen Infektion von LmoEGDe-mur-lux in der Leber
und im Darmbereich von BALB/cJ Mäusen im Vergleich zu C57BL/6J Mäusen
nachgewiesen werden (Abschnitt 2.3.3 und Abschnitt 2.3.4.2). Weiter zeigte sich die
Suszeptibilität der BALB/cJ Mäuse in einer erhöhten bakteriellen Keimlast in den
inneren Organen Dünndarm, mLK, Milz und Leber am Tag fünf nach der Infektion im
Vergleich zu C57BL/6J Mäusen (Abschnitt 2.3.4.2).
Die gesteigerte Suszeptibilität von BALB/cJ Mäusen konnte ebenfalls in früheren
Studien beobachtet werden. In BALB/cJ Mäusen war nach der intravenösen Infektion
mit verschiedenen Serotypen von Listeria monocytogenes (Serotyp 1/2a, 3b und 4b)
eine zehn- bis zwanzigfach geringere LD50 feststellbar als im Vergleich zu C57BL/6J
Mäusen (Mainou-Fowler et al., 1988). Eine genetische Kartierung von Quantitativen
Trait Loci (QTL) identifizierte in einer F2-Population der parentalen Inzuchtstämme
BALB/cJ und C57BL/6J zwei Listeria-Suszeptibilitätsloci auf den Chromosomen 5
und 13, welche die systemische Infektion von Listeria monocytogenes kontrollieren

Diskussion
113
und die Ursache der Suszeptibilität von BALB/cJ Mäusen kennzeichnen (Boyartchuk
et al., 2001).
Andere Studien zur Resistenz von Mausinzuchstämmen gegenüber einer Infektion
mit Listeria monocytogenes zeigten, dass Makrophagen von BALB/cJ Mäusen in
ihrer Fähigkeit das Wachstum von Listerien zu inhibieren, eingeschränkt sind
(Cheers et al., 1978). Nach der intravenösen Infektion mit Listeria monocytogenes
wurde eine höhere bakterielle Keimlast in der Leber und der Milz von BALB/cJ
Mäusen festgestellt als im Vergleich zu C57BL/6 Mäusen. Weiterhin resultierte die
Transplantation von Milzzellen aus intravenös infizierten BALB/cJ und C57BL/6
Mäusen in entsprechend syngenetische Mausstämme nur bei C57BL/6 Mäusen in
einer schützenden zellulär vermittelten Immunantwort (Cheers et al., 1978).
Die Infektion mit Listeria monocytogenes führt zu einer deutlichen Produktion von
IFN-γ, IL-10, TNF-α, IL-6 und CCL2. Besonders in den empfindlichen Mausstämmen
C3HeB/FeJ, A/JOlaHsd und BALB/cJ konnte eine Erhöhung an Tag drei und fünf
nach der oralen Infektion beobachtet werden (Abschnitt 2.3.4.4). Der Nachweis der
Chemokine und Zytokine weist darauf hin, dass mit zunehmender Suszeptibilität der
Mausinzuchstämme eine vermehrte Sekretion der chemotaktischen Stoffe erfolgt.
CCL2 wird von eingewanderten Granulozyten sekretiert, was die Immigration von
Makrophagen induziert (Guleria und Pollard 2001; Mandel und Cheers 1980). Diese
produzieren TNF-α und IL-12, welche natürliche Killerzellen zur Freisetzung von IFN-
γ anregen (Havell, 1987; Tripp et al., 1993). IFN-γ selbst ist von großer Bedeutung
und essentiell für die Resistenz gegenüber Listeria monocytogenes. So konnte
gezeigt werden, dass IFN-γ eine wichtige Funktion bei der Aktivierung von
Makrophagen einnimmt (Buchmeier und Schreiber, 1985). Die intraperitoneale
Infektion von C3HeB/FeJ Mäusen resultierte in einem Anstieg der IFN-γ Produktion
in der Milz an Tag sechs nach der Infektion. Die Neutralisierung des IFN-γ mit Hilfe
monoklonaler Antikörper verhinderte jedoch die Aktivierung von Makrophagen und
somit die Eliminierung der Listerien (Buchmeier und Schreiber, 1985). Auch erwiesen
sich IFN-γ-defiziente (GKO-/-BALB/cJ) Mäuse als besonders empfindlich für eine
intravenöse Listerieninfektion, was nach der Infektion in einer geringeren LD50 der
KO-Mäuse resultierte im Vergleich zu den Wildtyp BALB/cJ Mäusen (Harty und

Diskussion
114
Bevan, 1995). Die Sekretion von IL-6 führt im Besonderen zur Aktivierung von
Immunzellen und systemischen Inflammationsantworten (Jones, 2005). IL-10 wirkt
anti-inflammatorisch und inhibiert inflammatorisch bedingte Gewebeschäden (Moore
et al., 2001). In Studien der Sepsis und systemischen Infektion im Menschen konnten
ebenfalls erhöhte Konzentrationen von IFN-γ, IL-10, IL-6 und CCL2 im Serum
festgestellt werden (Murdoch und Finn, 2003; Tamayo et al., 2011). Schon 24
Stunden nach der gestellten Diagnose konnte der Anstieg der pro- und anti-
inflammatorischen Zytokine festgestellt werden, was darauf hinweist, dass sehr
frühzeitig im Verlauf einer Erkrankung die Immunantwort eingeleitet wird.
Für die in dieser Dissertation analysierten C3HeB/FeJ Mäuse konnte bereits in
früheren Arbeiten eine ausgeprägte Suszeptibilität nach einer intravenösen und
oralen Listerieninfektion gezeigt werden (Cheers und McKenzie, 1978; Park et al.,
2004). Auch nach der oralen Infektion sowohl mit murinisierten als auch mit nicht-
murinisierten Listeria monocytogenes konnte in dieser vorliegenden Arbeit eine
ausgeprägte Suszeptibilität nachgewiesen werden. Wie in Abschnitt 2.3.3 erkennbar,
resultierte die orale Listerieninfektion von C3HeB/FeJ Mäusen in einer stark
reduzierten Überlebensrate sowie in einem ausgeprägten Gewichtsverlust. Mit Hilfe
des BLI war in vivo und ex vivo eine vermehrte Ansammlung der Listerien in der
Leber der empfindlichen C3HeB/FeJ Mäuse zu beobachten (Abschnitt 2.3.4.2).
Weiterhin ergab die Analyse der bakteriellen Keimlast eine deutlich erhöhte Anzahl
von Listerien in den präparierten Organen Leber, Milz, Gallenblase, mLK und
Dünndarm. Auch konnten mit Hilfe der histopathologischen Analyse der Leber von
C57BL/6J und C3HeB/FeJ Mäusen vermehrt nekrotische Läsionen und Infiltrationen
von neutrophilen Granulozyten im Leberparenchym der C3HeB/FeJ Mäuse nach-
gewiesen werden (Abschnitt 2.3.4.3).
Frühere publizierte Arbeiten zeigten, dass C3HeB/FeJ Mäuse sehr empfindlich auf
die Infektion mit Mycobacterium tuberculosis reagierten (Kramnik et al., 2000;
Medina und North, 1998). Ein neuer Locus auf Chromosom 1 (Sst1) konnte dadurch
identifiziert werden, welcher die Suszeptibilität gegenüber Tuberkulose reguliert. So
reagierten C3HeB/FeJ Mäuse sehr empfindlich und C57BL/6J Mäuse resistent auf
die intravenöse Infektion mit Mycobacterium tuberculosis und Mycobacterium bovis

Diskussion
115
(Kramnik et al., 2000). Der Sst1 Wirtsresistenzlocus ist ebenfalls von Bedeutung die
angeborene Immunantwort nach einer Infektion mit Listeria monocytogenes zu
kontrollieren (Boyatrschuk et al., 2004). Über eine Generierung von kongenen
Mausstämmen, die das Sst1 C57BL/6J Resistenzallele im C3HeB/FeJ Rezipienten-
genom tragen, konnte gezeigt werden, dass dieser Locus zur Wirtsresistenz
gegenüber einer Listerieninfektion beiträgt. Der resistente Sst1 Locus der C57BL/6J
(B6) Mäuse korreliert des Weiteren mit einer höheren Produktion von reaktiven
Sauerstoffspezies in Sst1B6 Makrophagen, was daraufhin in der Eliminierung von
Listeria monocytogenes resultierte (Boyatrschuk et al., 2004). Innerhalb des Sst1
Locus konnte das Gen Ipr1 (Intracellular pathogen resistence 1) identifiziert werden,
welches die Resistenz gegenüber einer intravenösen Mycobacterium tuberculosis
Infektion erhöht (Pan et al., 2005). Ipr1 wird im Sst1 Resistenzallel exprimiert, auch in
Makrophagen von C57BL/6J Mäusen, jedoch aber nicht im Sst1 Locus von
C3HeB/FeJ Mäusen. Die Generierung einer Ipr1 transgenen C3HeB/FeJ Maus,
welche das Ipr1 Resistenzallel in gesamter Länge exprimiert, konnten statistisch
signifikante Unterschiede in den bakteriellen Keimlasten in der Lunge nach einer
Mycobacterium tuberculosis Infektion beobachtet werden. Ebenso konnte in vitro
gezeigt werden, dass Ipr1 transgene Makrophagen die Multiplikation von
Mycobacterium tuberculosis effizienter kontrollieren konnten (Pan et al., 2005). Ipr1
vermittelt zusätzlich die Resistenz von Makrophagen gegenüber weiteren
intrazellulären Pathogenen wie Listeria monocytogenes. Mit Hilfe der
Durchflußzytometrie konnte nach der Infektion von Sst1 sensitiven Makrophagen mit
Listeria monocytogenes Merkmale nekrotischer Zellen (Ethidiumbromidpos und
Annexin Vneg) erbracht werden, wogegen die Infektion von Irp1 transgenen
Makrophagen Merkmale einer Apoptose (Annexin Vpos und Ethidiumbromidneg)
aufwiesen (Pan et al., 2005). Somit trägt der empfindliche Sst1 Locus von
C3HeB/FeJ dazu bei, die Suszeptibilität gegenüber einer Infektion mit Listeria
monocytogenes zu erhöhen.
Ein weiterer genetischer Faktor, welcher für die Empfindlichkeit gegenüber einer
Listerieninfektion von Bedeutung ist, liegt im Genotyp des Haupthistokompatibilitäts-
komplex (MHC) Haplotyps. Der H-2 Locus von Mäusen kontrolliert während einer

Diskussion
116
Infektion die Eliminierung von Pathogenen, darunter auch von Bakterien (Nauciel et
al., 1990). Es konnten viele verschiedene Haplotypen identifiziert werden
(C3HeB/FeJ: H-2k, A/JOlaHsd: H-2a, BALB/cJ: H-2d und C57BL/6J: H-2b), welche in
der Resistenz oder Suszeptibilität gegenüber einer Listerieninfektion von Bedeutung
sind (Cheers und McKenzie, 1978). Die Auswirkungen des Haplotyps auf die Infek-
tion mit Gruppe A Streptokokken konnte auch in vivo beschrieben werden
(Goldmann et al., 2005). Mäuse des Haplotyps H-2k (BALB/k) und H-2d (BALB/c)
wurden intravenös mit Streptococcus pyogenes infiziert. Die Analyse der
Überlebensrate ergab eine Mortalitätsrate von 100% bei BALB/k Mäusen vergleich-
bar mit C3H/HeN (H-2k). Dagegen zeigten BALB/c (H-2d) eine Überlebensrate von
100%. Der Haplotyp des MHC-Locus hat somit entscheidenden Einfluss auf die aus-
geprägte Suszeptibilität gegenüber einer Infektion mit Gruppe A Streptokokken und
könnte auch Einfluss auf die Erregerabwehr von Listeria monocytogenes haben.
C3HeB/FeJ Mäuse (Haplotyp H-2k) zeigten nach der oralen Infektion mit Listeria
monocytogenes die ausgeprägteste Suszeptibilität. Dies lässt schlussfolgern, dass
der Haplotyp H-2k auch zu einer erhöhten Empfindlichkeit gegenüber einer
Listerieninfektion beitragen kann.
C3HeB/FeJ Mäuse weisen darüber hinaus eine erhöhte Empfindlichkeit gegenüber
einer Narkose mit Isofluran auf (Champy et al., 2008). Während des täglichen BLI
waren die Tiere der Isofluran-Narkose repetitiv exponiert, was weiteren Einfluss auf
den Verlauf einer Listerieninfektion haben könnte. Zudem wird Isofluran eine leber-
toxische Wirkung zugeschrieben. Dies könnte im Zusammenhang mit einer
Listerieninfektion und der damit verbundenen Invasion der Leber mit Listeria mono-
cytogenes zu einer zusätzlichen Funktionseinschränkung der Leber von C3HeB/FeJ
Mäusen zur Folge haben und sich auf den Infektionsverlauf zusätzlich negativ aus-
wirken.

Diskussion
117
3.4 Infektion des zentralen Nervensystems zeigt keine Reproduzier-
barkeit und ist nicht von der Murinisierung des Internalin A ab-
hängig
Die Fähigkeit von Listeria monocytogenes epitheliale Barrieren zu durchqueren,
ermöglicht auch die Invasion des zentralen Nervensystems (ZNS). Durch die
Überquerung der Blut-Hirn-Schranke kann es zur Auslösung einer Meningitis und
Enzephalitis kommen. Auf welchem Weg Listeria monocytogenes das Gehirn infiziert
ist noch nicht vollständig geklärt. Es liegen verschiedene Untersuchungen vor, die
beschreiben, dass Listerien die Blut-Hirn-Schranke intra- oder interzellulär durch-
queren können (Drevets et al., 2004). Weiterhin sind Listerien fähig, über infizierte
Makrophagen oder entlang von Neuronen ins Gehirn einzudringen (Antal et al., 2001;
Antal et al., 2005; Drevets, 1999; Drevets et al., 2001; Drevets et al., 2004; Huang et
al., 2001; Jin et al., 2001). Während einer Infektion mit Listeria monocytogenes
überqueren die Listerien zunächst die intestinale Barriere und gelangen über das
Blut- und Lymphsystem zur Leber und Milz (Vazquez-Boland et al., 2001). Aufgrund
der Replikation und starker Vermehrung der Listerien mit Zugang zu Blutgefäßen
erfolgt dann eine sekundäre Bakteriämie und die weitere Ausbreitung über den
Organismus zum Gehirn.
Die epitheliale Barriere der Blut-Hirn-Schranke besteht aus Endothelzellen, welche
von einer Basallamina umgeben sind. An diese lagern sich Astrozyten und Perizyten
an. Letztere sind mit den Endothelzellen fest über Zell-Zell-Kontakte verankert und
bilden eine physiologische Barriere (Staddon und Rubbin, 1996). Diese erlaubt
jedoch den kontrollierten Transport von Substanzen mittels Transzytose (Tuma und
Hubbard, 2003) und stellt eine Möglichkeit der Invasion für Listeria monocytogenes
dar. Somit könnte die Interaktion der Oberflächenproteine InlA und InlB mit ihren
Rezeptoren E-Cadherin und c-Met die Überquerung der Blut-Hirn-Schranke über den
rezeptorabhängigen Transport in Endothelzellen der Blut-Hirn-Barriere ermöglichen.
Es konnte gezeigt werden, dass E-Cadherin an primären sensorischen Neuronen
exprimiert ist und die Interaktion mit Internalin A erlaubt (Shimamura et al., 1992).
Ebenso wird der Rezeptor c-Met auf einer Reihe von Zellen des zentralen

Diskussion
118
Nervensystems exprimiert und ermöglicht die Interaktion mit Internalin B (Jung et al.,
1994; Maina et al., 1997). In vitro Arbeiten zeigten, dass über die InlB-c-Met-Inter-
aktion Listeria monocytogenes in humane mikrovaskuläre Hirnendothelzellen ein-
dringen kann (Greiffenberg et al., 1998). Die Infektion mit Listerienmutanten für InlA
und InlB ergab, dass die Deletion des InlB in einer Reduzierung der Invasionrate
resultierte. Die InlA-Deletion zeigte dagegen keine signifikante Reduktion der
Invasionsrate, was darauf hinweist, dass InlA für die Invasion in vaskuläre
Hirnendothelien keine Rolle spielt (Greiffenberg et al., 1998).
In dieser Arbeit wurden ausgewählte Mausinzuchtstämme (A/JOlaHsd, BALB/cJ,
C3HeB/FeJ, C57BL/6J und 129P2/Ola) mit murinisierten Listerienstämmen
(LmoXen32-mur, LmoEGDe-mur-lux) und ihren isogenen Wildtypstämmen
(LmoXen32, LmoEGDe-lux und LmoEGDe) oral infiziert. Sowohl die Infektion mit
murinisierten Listerienvarianten als auch die Infektion mit den nicht-murinisierten
Wildtypstämmen führte zu neurologischen Ausfallserscheinungen wie Torticollis und
„kreiselnde“ Bewegungen um die Längsachse. Das Auftreten dieser neurologischen
Ausfallerscheinung konnte als Invasion des ZNS durch Listeria monocytogenes
gewertet werden. Der Zeitpunkt des Auftretens der klinischen Symptome einer ZNS-
Infektion war am Tag sieben bis Tag acht nach der oralen Infektion zu beobachten.
Klinische Symptome konnten in den Mausinzuchtstämmen 129P2/Ola, A/JOlaHsd,
BALB/cJ und C57BL/6J nachgewiesen werden (Abschnitt 2.6). Die histologische
Analyse des Gehirns zeigte Infiltration von Granulozyten und Makrophagen im
Bereich der Meningen sowie konnte mit Hilfe der immunhistologischen Analyse der
Nachweis von Listeria monocytogenes innerhalb dieser Inflitrationen erbracht
werden. Der neurologische Phänotyp wurde insgesamt nur in acht von 600 Tieren,
welche über den Versuchszeitraum von über sieben Tagen beobachtet worden sind,
nachgewiesen. Drei Mäuse entwickelten nach der Infektion mit nicht-murinisierten
Listerienstämmen und fünf Mäuse nach der Infektion mit murinisierten
Listerienstämmen klinische Symptome einer ZNS Infektion. Das Auftreten der
neurologischen Symptome war somit nur in einzelnen Tieren zu beobachten und es
konnte keine Reproduzierbarkeit der Ergebnisse erreicht werden. Die gewonnenen
Ergebnisse bestätigen, dass eine intakte Internalin A - E-Cadherin Interaktion nicht

Diskussion
119
Voraussetzung für die Überquerung der Blut-Hirn-Barriere ist und die Murinisierung
des Internalin A keinen Einfluss dabei hat. Da neurologische Symptome sowohl bei
intakter InlA - E-Cadherin - Interaktion (murinisierte Listerien) als auch bei
ineffizienter InlA - E-Cadherin - Interaktion (nicht-murinisierte Listerien) beobachtet
werden konnten, lässt sich schlussfolgern, dass die Murinisierung des Internalin A
nicht zu einer verstärkten Invasion des ZNS beiträgt. Auch die Generierung der
knock-in Maus „E16P“, welche durch den Austausch von Glutamin zu Prolin an der
Position 16 des Maus-E-Cadherins humanes E-Cadherin anstelle von murinem E-
Cadherin exprimiert, konnte die Rolle des InlA bei der Überquerung Blut-Hirn-
Schranke ebenfalls nicht aufzeigen (Disson et al., 2008).
Weitere diskutierte Infektionswege für die Invasion des Gehirns und die Durch-
querung der Blut-Hirn-Schranke ist der Transport der Listerien innerhalb infizierter
Makrophagen. Dieser Infektionsweg ist auch als sogenannte „Trojanisches Pferd“
Hypothese beschrieben worden (Drevets, 1999; Drevets et al., 2001). In der
Frühphase der Infektion wurde der intrazelluläre Transport von Listeria mono-
cytogenes innerhalb von Makrophagen und dendritischen Zellen zur Milz beobachtet
(Neuenhahn et al., 2006; Neuenhahn et al., 2010; Pron et al., 2001).
Listerieninfizierte Makrophagen konnten im peripheren Blut und im Gefäßsystem des
Gehirns von infizierten Mäusen nachgewiesen werden (Drevets, 1999; Prats et al.,
1992). Die Isolierung dieser infizierten peripheren Leukozyten und die anschließende
intravenöse Injektion in Mäuse resultierte in einer Organinfektion von Leber, Milz und
Gehirn, was mit Hilfe der Analyse der bakteriellen Keimlast in diesen Organen
aufgezeigt werden konnte (Drevets, 1999). Infolge einer Infektion mit Listeria
monocytogenes mit anschließender systemischer Dissemination kann eine Invasion
des Knochenmark erfolgen. So konnte nach der intravenösen Infektion mit Listeria
monocytogenes infizierte myeloide Knochenmarkszellen nachgewiesen werden,
welche nach intravenöser Transplantation in C57BL/6J Mäuse zu einer Invasion des
Gehirns führten (Join-Lambert et al., 2005). Ebenso zeigten BALB/c Mäuse nach
oraler und CD1 Mäuse nach intravenöser Infektion mit biolumineszenten Listerien
eine Invasion des Knochenmarks, welche über die Detektion der Biolumineszenz in
der Tibia nachgewiesen werden konnte (Hardy et al., 2009). Über infizierte

Diskussion
120
Makrophagen kann zudem die weitere Zell-Zell-vermittelte Ausbreitung in
Hirnendothelien erfolgen. In vitro Arbeiten zeigten, dass Listeria monocytogenes über
Zell-Zell-vermittelte Dissemination Endothelien und Neuronen effizient infizieren kann
(Dramsi et al., 1998; Drevets, 1999). Ebenso weisen infizierte Phagozyten eine
ausgeprägte Adhärenz zu epithelialen Zellen auf, was eine Zell-zu-Zell Ausbreitung
begünstigt und intrazellulären Listerien die Infektion von endothelialen Zellen
ermöglicht (Drevets, 1999; Drevets et al., 2001; Greiffenberg et al., 1998). Somit
stellen infizierte Monozyten eine mögliche Infektionsquelle für die Invasion des ZNS
dar. Die weitere Ausbreitung der Listerien in tiefere Gewebeschichten des ZNS-
Parenchyms erfolgt über eine Zell-Zell-vermittelte Ausbreitung. Ein wichtiger
Virulenzfaktor für die weitere Zell-Zell-vermittelte Invasion ist die Phosphatidylcholin-
spezifische Phospholipase C (PlcB). Listerienmutanten, welche defizient für PlcB
sind, zeigten eine reduzierte Virulenz nach intrazerebrale Injektion in C57BL/6J
Mäusen, was in einer geringeren bakteriellen Keimlast und Mortalitätsrate resultierte
(Schlüter et al., 1998).
Eine weitere Alternative für die Invasion des zentralen Nervensystems liegt im
Transport von Listeria monocytogenes entlang von Neuronen. Schon früh wurde
beobachtet, dass die Injektion von Listerien in die Mundschleimhaut von Mäusen und
Ziegen zu klinischen Symptome einer Hirnstammenzephalitis führte (Asahi et al.,
1952). Die Präparation des Gehirns und der kranialen Nerven ergab Infiltrationen von
mononukleären Zellen entlang des Nervus trigeminus und im Hirnstamm. In Fällen
einer spontanen Enzephalitis in Schafen konnten ebenfalls intraaxonal Listerien
elektronenmikroskopisch nachgewiesen werden (Charlton, 1977; Otter und
Blakemore, 1989). Der intraaxonale Transport wurde bestätigt als sechs von
21Schafe, welche direkt in die Zahnpulpa mit Listerien infiziert wurden waren
neurologische Symptome entwickelten (Barlow und McGorum, 1985). Der
Zusammenhang zwischen einer neuronalen Infektion und dem Auftreten einer
Erkrankung des ZNS wurde im Mausmodell bestätigt. So führte die Injektion von
Listerien in die Kranialnerven oder in die faszialen Muskeln von Hsd/ICR Mäusen zu
klinischen Symptomen einer Rhombenzephalitis (Antal et al., 2001). Die Infektionen
von Neuronen mit extrazellulär vorliegenden Listeria monocytogenes ist jedoch in

Diskussion
121
vitro selten zu beobachten. So resultierte die Infektion von murinen fetalen
Gehirnzellen mit Listeria monocytogenes in einer geringeren Invasionsrate (3,5%) im
Vergleich zu der Infektion von Makrophagen (56,6%) (Peters und Hewicker-
Trautwein, 1994).
Ein nicht zu vernachlässigender Faktor für eine Infektion des ZNS mit Listeria mono-
cytogenes ist zusätzlich die Lebensweise des Wirtes. Gerade bei Wiederkäuern kann
die Nahrungsaufnahme von Silage zu Verletzungen der Mundschleimhaut führen,
welche dann Listerien aus kontaminierter Silage die Infektion der in der Mund-
schleimhaut vorliegenden Nerven ermöglichen. Im Menschen ist dieser neuronale
Infektionsweg nicht vollständig geklärt, da die Risikolebensmittel, wie Salate und
Käse, keine abrasiven Eigenschaften aufweisen und die Mundschleimhaut wenig
verletzen, ähnliche Voraussetzungen wie im Mausmodell mit murinisierten Listerien.
Somit lässt sich vermuten, dass die Infektion des Gehirns am wahrscheinlichsten
über infizierte Makrophagen und über eine anschließende Zell-Zell-vermittelte
Ausbreitung erfolgt. Dies bedarf jedoch weiterer Analysen und könnte unter
Verwendung der murinisierten Listerien LmoEDGe-mur-lux im Mausmodell zum
Beispiel mit Hilfe von Zweiphotonenmikroskopie im Detail erfolgen.
3.5 Die Detektion des Biolumineszenzsignals in Abhängigkeit verschie-
dener Faktoren
Die in vivo Analyse von Infektionsprozessen mit Hilfe des Biolumineszenz-Imaging
ermöglicht die Lokalisation von Pathogenen im lebenden Organismus und die
Beobachtung ihrer Dissemination zu den inneren Organen des Versuchstiers.
Basierend auf der Eigenschaft von Lichtphotonen durch Gewebe transmittiert zu
werden, wurden verschiedene Systeme etabliert, die emittiertes Licht aus einer
internen biologischen Quelle detektieren können.
Biolumineszenz ist ein lichtemittierender chemischer Prozess, welcher über Luzi-
ferasen katalysiert wird. Mit Hilfe verschiedener Luziferasen (DeLuca und McElroy,
1974; Hastings und Nealson 1977; Matthews et al., 1977) erfolgte die Generierung

Diskussion
122
von biolumineszenten Pathogenen wie Staphylococcus aureus (Francis et al., 2000),
Streptococcus pneumonia (Francis et al., 2001), Herpes-simplex-virus Typ I (Luker et
al., 2002), Salmonella enterica (Burns-Guydish et al., 2005) und Listeria
monocytogenes (Hardy et al, 2004; Monk et al., 2010; Riedel et al., 2007), um Infek-
tionsprozesse im Versuchstier direkt zu beobachten.
Das in vivo Biolumineszenz-Imaging erlaubt die nicht invasive und hochsensitive
Echtzeitaufnahme des Infektionsverlauf des ganzen lebenden Tieres nach der Infek-
tion mit biolumineszenten Pathogenen. Die Tiere werden in einer lichtdichten
Kammer platziert und das emittierte Licht über eine CCD Kamera (cooled coupeld
device) detektiert. Aufgrund einer Kühlung der Kamera kann störendes Wärme-
rauschen reduziert werden. Die Messung der Tiere erfolgt unter Narkose, um die
Biolumineszenzsignale anatomisch zuordnen zu können. Dabei werden die Mäuse
auf einer Wärmeplatte platziert, um sie vor einer nakosebedingten Auskühlung zu
schützen. Die Signalintensität der Biolumineszenz wird mit Hilfe des Systems in
Falschfarben dargestellt (Abschnitt 5.4.4).
Die in dieser Arbeit durchgeführten Analysen des in vivo Biolumineszenz-Imaging
ergaben nach der Infektion ausgewählter Mausinzuchstämme mit biolumineszenten
Listerien verschiedene Intensitäten. A/JOlaHsd und BALB/cJ Mäuse eigneten sich
aufgrund ihrer unpigmentierten Haut und Fellfarbe gut für die Detektion der
Biolumineszenz. Ebenso zeigten C3HeB/FeJ Mäuse intensive Bio-
lumineszenssignale, was über deren ausgeprägte Suszeptibilität zu erklären ist. Je
empfindlicher ein Mausstamm reagiert, desto höher ist die Invasions- und Repli-
kationsrate der Listerien in vivo und dementsprechend kann die Detektion inten-
sitätsstarker Biolumineszenzen erfolgen (siehe Abschnitt 2.3.3 und Abschnitt 3.3).
Ergebnisse der in dieser Arbeit durchgeführten Experimente ergaben direkt nach der
Infektion detektierbare biolumineszente Listerien im Magen-Darm-Trakt. An den
Tagen eins und zwei nach der Infektion war keine oder nur eine sehr geringe
Biolumineszenzen nachweisbar (Abschnitt 2.3.4.2). Die geringe Biolumineszenz im
Darmbereich in der frühen Phase der Infektion resultierte aus der Dissemination der
Listerien über den Darm und infolge dessen mit deren partiellen Ausscheidung.

Diskussion
123
Jedoch kann zusätzlich der physiologische Sauerstoffmangel innerhalb des Darms
die Biolumineszenzreaktion reduziert haben. Die Biolumineszenzreaktion ist
sauerstoffabhängig, wobei Luziferasen die Oxidation ihres spezifischen Substrates in
Abhängigkeit von Sauerstoff und ATP unter Freisetzung von Lichtphotonen
katalysieren (Greer und Szalay, 2002). So konnte nach der oralen Infektion von
BALB/c Mäusen mit luziferaseexprimierenden Salmonellen keine Biolumineszenz im
Dünndarm detektiert werden. Erst nach Injektion von Luft in den Darm konnten
Photonen in der Nähe der Injektionsstelle nachgewiesen werden (Contag et al.,
1995).
Weiterhin wird die Transmission von Licht durch das Gewebe über die Absorption
und Streuung des Lichtes abgeschwächt (Tuchin, 2000; Rice et al., 2001). Streuung
von Licht erfolgt an allen Zell- und Organellmembranen. Dies hat zur Folge, dass
emittiertes Licht abgelenkt und die räumliche Auflösung reduziert wird. Je tiefer im
Gewebe sich die Listerien befinden, desto größer ist die Streuung und geringer die
Detektion der Biolumineszenz. Die Absorption von Licht ist abhängig vom Zell- oder
Gewebetyp sowie von der Anwesenheit des Hämoglobins oder Melanins (Tromberg
et al., 2000). Hämoglobin absorbiert Licht in einer Wellenlänge von 400-600 nm.
Während der Biolumineszenzreaktion wird Licht mit einer Wellenlänge von 490 nm
emittiert somit kann speziell in den gut durchbluteten Organen wie Milz und Leber,
ebenfalls Zielorgane der Listerien (Lecuit et al., 2001), das emittierte Licht der
biolumineszenten Listerien abgeschwächt werden.
Weitere Faktoren, welche die Detektion der Biolumineszenz beeinflussen, sind die
Anzahl der biolumineszenten Zellen, welche die Reportergene exprimieren und die
Stärke des generierten Signals. Einen weiteren Einfluss auf die Detektion der
Biolumineszenz hat das Luziferase-Reportersystem. Mit der Integration des
kompletten Lux-Operons, welches sowohl für das Enzym als auch für das Substrat
der Lichtreaktion codiert, ins Genom von z.B. Bakterien, wird eine stabilere Licht-
produktion erreicht (Doyle et al., 2004). Bei anderen Reportersystemen, bei denen
nur die Luziferasegene ins Genom von Tieren integriert sind, muss die Zugabe des
Substrats für die Biolumineszenzreaktion zusätzlich von außen erfolgen. Die Injektion

Diskussion
124
des Substrates ermöglicht die Detektion intensitätsstarker Biolumineszenzen, jedoch
ist dieses Signal nur über einen kurzen Zeitraum stabil.
Wie diese Arbeit anschaulich demonstriert, ist eine bestimmte Anzahl von bio-
lumineszenten Listerien nötig, um spezifische Biolumineszenzsignale zu erhalten. Ab
einer Bakterienlast von 104 CFU / mg Organ war eine Korrelation der Bakterienlast
mit der Biolumineszenz erkennbar (Abschnitt 2.5). Hier liegt auch die Limitation des
BLI. Eine geringere Anzahl von Listerien ist nicht detektierbar. Wie die Analyse der
bakteriellen Keimlast zeigte, lag die Anzahl der Listerien in den inneren Organen
Milz, Leber, mLK, Dünndarm und Gehirn von C57BL/6J Mäusen an Tag drei und fünf
nach der Infektion zwischen 103-104 CFU / mg Organ. Es konnten jedoch in vivo und
ex vivo keine Biolumineszenzsignale in diesen Organen detektiert werden (Abschnitt
2.4.3.1 und Abschnitt 2.3.4.2). Spezifische Biolumineszenzsignale der Listerien
waren erst ab einer bakteriellen Keimlast größer als 104 CFU/mg Organ möglich.
Ebenfalls von wichtiger Bedeutung für die Analyse über BLI ist die Wahl des
Mausstamms. Dunkles Fell und eine dunkle Pigmentierung der Haut absorbieren
mehr Licht als Tiere mit weißem Fell und nicht pigmentierter Haut. Daher erwiesen
sich BALB/cJ und A/JOlaHsd Mäusen eher als geeignet für das in vivo Imaging als
C57BL/6J Mäuse. Die Detektion der Biolumineszenz kann zusätzlich über eine Rasur
des Fells erleichtert werden.
Ein großer Vorteil des in vivo Biolumineszenz-Imaging ist die Möglichkeit der wieder-
holten Messung individueller Tiere über den gewünschten Versuchszeitraum. Dieses
nicht invasive Verfahren ermöglicht die Anzahl der Versuchstiere zu reduzieren.
Jedoch sollten die Ergebnisse des BLI mit Endpunktprotokollen, wie der Bestimmung
der bakteriellen Keimlast, bestätigt werden.
3.6 Reaktionen des angeborenen Immunsystems auf die Infektion mit
Listeria monocytogenes
Die angeborene Immunantwort stellt die erste Verteidigungslinie nach der Infektion
mit pathogenen Mikroorganismen dar. Phagozytierende Zellen wie Makrophagen,

Diskussion
125
neutrophile Granulozyten und dendritische Zellen sind von Bedeutung, um die bakte-
rielle Ausbreitung und Vermehrung durch die Produktion antimikrobieller Wirkstoffe
zu hemmen (Rogers und Unanue, 1993). Intrazelluläre Bakterien wie Listeria mono-
cytogenes haben Strategien entwickelt, um innerhalb dieser Phagozyten zu über-
leben und zu replizieren (Flannagan et al., 2009; Ray et al., 2009). Dazu gehören die
Fähigkeit zur Lyse der Invasionsvakuole, die intrazelluläre Replikation und die Aus-
breitung von Zelle zu Zelle. Die Detektion von bakteriellen Komponenten über die
Zellen des Immunsystems führt zur Sekretion von inflammatorischen Zytokinen,
welche wichtig für die Kommunikation der beteiligten Immunzellen sind (Leber et al.,
2008). Listeria monocytogenes induziert die Synthese von Typ-I-Interferonen,
bestehend aus IFN-β und 13 verschiedenen IFN-α Subtypen (O´Riordan et al., 2002;
Perry et al., 2005; Petska et al., 2004). Interferone aktivieren weiter eine Vielzahl von
Genen, welche wiederum in den Prozessen der angeborenen und erworbenen
Immunabwehr von Bedeutung sind. Die Sekretion von IFN-αβ wirkt sich jedoch
begünstigend auf eine Listerieninfektion aus. Mäuse, welche für den Interferon-I-
Rezeptor defizient sind, zeigten sich resistent gegenüber der Infektion mit Listeria
monocytogenes (Auerbuch et al., 2004). So resultierte die intravenöse Infektion von
IFN-α/βR-/- Mäusen in einer geringeren bakteriellen Keimlast in der Leber und der
Milz als die Infektion von BALB/cJ Wildtypmäusen.
Im Rahmen der Phänotypisierung von Mausmutanten erfolgte in dieser Arbeit die
Infektion von Interferon-α inducible protein 27 like 2A knock-out (Ifi27l2a-KO)
Mäusen. Das Gen Ifi27l2a (lSG12b1) wird von Typ-I-IFN stimuliert und gehört zu
einer großen Familie der über Interferone aktivierten Gene und wird vor allem in
Makrophagen und im Fettgewebe exprimiert (Li et al., 2009, Ohgaki et al., 2007). Die
Analyse der Überlebensrate der Ifi27l2a-KO Mäuse im Vergleich zu den
entsprechenden Wildtypkontrollen, C57BL/6N, ergab keine signifikanten
Unterschiede (Abschnitt 2.7.1). Jedoch zeigte die Analyse der bakteriellen Keimlast
eine signifikant höhere Listerienanzahl in Ifi27l2a-KO Mäusen in der Leber und Milz
an Tag drei nach der intravenösen Infektion im Vergleich zu C57BL/6N Mäusen.
Ebenso zeigten die Kurvenverläufe der Körpergewichte signifikante Unterschiede
von Ifi27l2a-KO Mäusen im Vergleich zu den infizierten C57BL/6N Mäusen an den

Diskussion
126
Tagen zwei, drei und vier nach der Infektion. Die Ifi27l2a-KO Mäuse reagierten mit
größeren Gewichtsverlusten als die C57BL/6N Mäuse. Der Genverlust von Ifi27l2a
könnte dazu beitragen, dass in den ersten Tagen nach der Infektion ein größerer
Gewichtsverlust bei Ifi27l2a-KO Mäusen nachzuweisen war. Jedoch erfolgte die
Gewichtszunahme zu einem früheren Zeitpunkt in Ifi27l2a-KO Mäusen (Tag fünf
nach der Infektion) im Vergleich zu C57BL/6N (Tag neun nach der Infektion). Dies
könnte die Möglichkeit der Kompensation des Ifi27l2a-KO im Verlauf der Infektion
demonstrieren. Ifi27l2a ist nur ein Gen aus einer ganzen Gruppe von Genen, welche
über IFN stimuliert werden. Somit ist anzunehmen, dass die fehlende über Ifi27l2a
vermittelte Funktion über andere IFN-stimulierte Gene kompensiert wird. Zur
weiteren Analyse Ifi27l2a-KO wäre es hilfreich, die bakterielle Keimlast an zusätz-
lichen Zeitpunkten nach der Infektion zu untersuchen.
Der Nachweis von IFN-β war aufgrund fehlender effektiver Detektionssysteme in
vorangegangenen Arbeiten begrenzt möglich. Mit Hilfe der von Lienenklaus et al.,
etablierten Interferon-β-Reportermaus (Lienenklaus et al., 2009) konnte in dieser
Arbeit die Visualisierung der IFN-β-Produktion in verschiedenen Geweben nach der
Infektion mit Listeria monocytogenes in vivo beobachtet werden. Untersucht wurde
die IFN-β-Produktion nach der Infektion mit den murinisierten Listerienstämmen
LmoEGDe-mur-lux und LO28-mur, im Vergleich mit ihren isogenen Wildtypstämmen
LmoEGDe-lux und LO28 (Abschnitt 2.7.2). Die Listerienstämme EGDe und LO28
unterscheiden sich in ihrer Fähigkeit eine IFN-β-Synthese zu aktivieren. Der
Listerienstamm LO28 induziert im Vergleich zu LmoEGDe eine stärkere IFN-β-
Produktion (Reutterer et al., 2008).
Wie in dieser vorliegenden Arbeit gezeigt werden konnte, resultierte die
Murinisierung des Internalin A zusätzlich in einer Erhöhung der Interferon-β-Synthese
in den verschiedenen Geweben (Abbildung 2-33). So zeigte die Infektion mit LO28-
mur und LmoEGDe-mur-lux eine erhöhte IFN-β-Produktion als die Infektion mit den
isogenen Wildtypstämmen LO28 und LmoEGDe-lux. Weiterhin konnten am Tag zwei
nach der oralen Infektion mit Listeria monocytogenes die Produktion von IFN-β in
den zervikalen Lymphknoten, der Leber und im Darm nachgewiesen werden. Die
Detektion von Listerien nach oraler Infektion in den Lymphknoten wurde bereits in

Diskussion
127
früheren Arbeiten gezeigt (Tam und Wick, 2006). Ebenso konnten Listerien nach
intravenöser Infektion in den zervikalen und inguinalen Lymphknoten nachgewiesen
werden (Solodova et al., 2011). Wie bei Solodova et al., beschrieben, wurde nach
der intravenösen Infektion mit LO28 die IFN-β-Synthese in der Milz jedoch nicht in
der Leber beobachtet. (Solodova et al., 2011). In der vorliegenden Arbeit war eine
IFN-β-Produktion in der Milz nach oraler Infektion nicht nachweisbar. Hier scheint die
Infektionsroute eine wichtige Rolle zu spielen. Nach intravenöser Infektion erfolgt die
systemische Ausbreitung sofort und ist nicht wie nach der oralen Infektion von der
Passage durch die intestinale Barriere abhängig. Im Meerschweinchenmodell konnte
nachgewiesen werden, dass die Ausbreitung von Listerien zur Leber nach der oralen
Infektion zum einen direkt vom Dünndarm aus über die Pfortader erfolgen oder
indirekt über die mLK (Melton-Witt et al., 2011). Die Ausbreitung zur Milz erfolgt
dagegen nur über den indirekten Weg. Dies führte nach der oralen Infektion zu
einem höheren Prozentsatz von Listerien in der Leber im Vergleich zur Milz (Melton-
Witt et al., 2011). Dies könnte erklären, warum im murinisierten Listerienmodell in der
Maus nur eine IFN-β-Produktion in der Leber und nicht in der Milz detektiert wurde,
da die Invasion von einer höheren Anzahl Listerien in die Leber eine verstärkte IFN-
β-Synthese induziert.

Ausblick
128
4 Ausblick
Mit Hilfe der generierten murinisierten Listerienstämme wird die Induzierung einer
humanen Listeriose im Mausmodell möglich. Durch die zusätzliche Integration des
Lux-Operons in das Genom von Listeria monocytogenes kann der Krankheitsverlauf
zusätzlich in vivo verfolgt werden. So können individuelle Tiere über den gesamten
Versuchszeitraum täglich untersucht werden. Die nicht invasive Analyse der
Listerieninfektion über Biolumineszenz-Imaging erlaubt daher, die Menge der
eingesetzten Versuchstiere erheblich zu reduzieren.
Das in vivo Imaging von Listeria monocytogenes erbrachte in den letzten Jahren
neue Erkenntnisse über deren Pathophysiologie und Invasion. Die Beschreibung der
Fähigkeit extrazellulär in der Gallenblase zu replizieren, erbrachte neue Einsichten in
die Lebensweise von Listerien (Hardy et al., 2004).
Die Etablierung des murinisierten Listerienmodells erlaubt die Infektion jedes belie-
bigen Mausstammes und somit die Untersuchung der Listerieninfektion in Abhängig-
keit des genetischen Hintergrund des Wirtes. Es wird damit auch gezielt möglich
sein, knock-out- bzw. knock-in-Mausmutanten zu phänotypisieren. Die Auswirkung
einzelner Gene auf die angeborene und erworbene Immunantwort bei einer
Listerieninfektion über die natürliche orale Infektionsroute kann somit analysiert
werden. Zusätzlich kann mit Hilfe des in vivo Imaging die Dissemination der Listerien
über den Organismus verfolgt werden. Es könnten Erkenntnisse gewonnen werden,
welche weiteren bisher nicht identifizierten Virulenzfaktoren eine Rolle bei der Über-
querung epithelialer Barrieren wie der Plazentaschranke, der Blut-Hirn-Schranke und
der intestinalen Barriere sowie bei der Interaktion mit der Immunabwehr spielen.
Wie Listeria monocytogenes die Plazentaschranke überquert, ist noch nicht voll-
ständig verstanden und wird kontrovers diskutiert. Viele verschiedene Faktoren
spielen dabei eine Rolle. Zum einen erfolgt während der Schwangerschaft, infolge
einer veränderten hormonellen Situation, eine physiologische Suppression der T-
Zell-vermittelten Immunantwort (Pung et al., 1985; Weinberg, 1984). Diese schützt

Ausblick
129
zwar den Fetus vor der Abstoßung, erhöht aber auch die Anfälligkeit für intrauterine
Infektionen (Redline et al., 1988; Redline und Lu, 1987; Redline und Lu, 1988).
Weiterhin führt aufgrund einer geringeren IFN-γ, IL-2 und TNF-α Produktion, bedingt
durch einen erhöhten Östrogenspiegels während einer Schwangerschaft, die Infek-
tion mit Listerien zur Invasion der Plazenta und des fetalen Gewebes (Nakane et al.,
1985; Salem et al., 1999). Auch resultierte, nach der oralen Infektion von
schwangeren C57BL/6J Mäusen mit Listeria monocytogenes, diese physiologische
Immunsuppression in einer reduzierten Resistenz und führte zu schwerwiegenden
fetalen Infektionen mit Aborten (Poulsen et al., 2011). Die Oberflächenproteine InlA
und InlB und deren Interaktion mit ihren Rezeptoren E-Cadherin und c-Met sind bei
der Überquerung der Plazentaschranke mit involviert. Für die effektive Invasion der
humanen Plazenta mit Listeria monocytogenes sind beide intakten InlA- und InlB-
Wege nötig. Nach einer Infektion mit Deletionsmutanten für InlA oder InlB sowie für
die Doppelmutante war die Effizienz der Plazentainfektion deutlich verringert (Lecuit
et al., 2004). Die Etablierung der murinisierten Listerienvarianten (Monk et al., 2010;
Wollert et al., 2007b) könnte zu weiteren Untersuchungen der Rolle des Internalin A
beim Überqueren der Plazentaschranke im Mausmodell genutzt werden. Beide
Interaktionswege, InlA und InlB, sind funktionsfähig und die orale Infektion von
schwangeren Mäusen kann als Modell für die Infektion im Menschen hilfreich sein.
Eine weitere diskutierte Hypothese für die Überquerung der Plazentaschranke ist,
wie bei der Überquerung der Blut-Hirn-Schranke, die Invasion über infizierte
Phagozyten („Trojanisches Pferd“) (Bakardjiev et al., 2005; Bakardjiev et al., 2006;
LeMonnier et al., 2007). Listerien erreichen über infizierte Monozyten die Plazenta.
Durch eine nachfolgende Zell-Zell-Ausbreitung über die Trophoblasten können die
Listerien das fetale Gewebe erreichen. Dies könnte unter Verwendung der
murinisierten und biolumineszenten Listerien LmoEDGe-mur-lux im Mausmodell in
vivo und nicht invasiv mittels BLI erfolgen.
Zudem ist die Etablierung von Tiermodellen der initiale Schritt Krankheitsgeschehen
zu verstehen, welche am Ende mit der Entwicklung von wirksamen Therapeutika
einhergehen.

Material und Methoden
130
5 Material und Methoden
Die im Verlauf dieser Arbeit verwendeten Reagenzien und Chemikalien wurden,
soweit nicht an anderer Stelle erwähnt, von den Firmen Applichem GmbH, Becton &
Dickinson, BioRad, Gibco, Invitrogen, J.T. Baker, Macherey & Nagel, Merck, New
England Biolabs, PAA, Qiagen, Roche, Roth, Serva und Sigma bezogen. Die
Herstellung sowie Verwendung der verschiedenen Pufferlösungen und Medien sind
bei den entsprechenden Methoden vermerkt. Generell wurden alle Lösungen unter
Verwendung von Wasser aus einer Millipore-MilliQ-Anlage angesetzt. Bei Bedarf
erfolgte die Sterilisation der Lösungen durch 20-minütiges Autoklavieren bei 121°C
und 2 bar oder über Filtration. War steriles Arbeiten erforderlich wurden sterilisierte
Gefäße und Verbrauchsmaterialien verwendet. Soweit nicht an anderer Stelle
erwähnt, erfolgte die Nutzung in molekularbiologischen Laboren allgemein gebräuch-
lichen Gerätschaften und Materialien. Der Einsatz kommerzieller Kitsysteme sowie
deren Versuchsdurchführung erfolgte nach Vorschrift und wird bei Abweichungen im
Versuchsablauf an entsprechender Stelle der Methoden erwähnt.
5.1 Generierung genetisch veränderter Listeria monocytogenes Stämme
5.1.1 Generierung elektrokompetenter Listerien
Als Voraussetzung für die Transformation von Listeria monocytogenes mit dem
Integrationsvektor pAUL-A-inlAm-inlB wurden elektrokompetente Listerienzellen
benötigt. Die Stämme LO28 und LmoXen32 wurden über Nacht in einer Flüssigkultur
von Brain-Heart-Infusion (BHI) bei 37°C angezogen und am nächsten Morgen 1:50 in
500 ml BHI-Medium mit 0,5 M Sucrose überimpft. Es folgte die Inkubation bei 37°C
im Schüttelinkubator bei 120 Umdrehungen pro Minute (rpm) bis die optische Dichte
600 (OD600) bei 0,2 lag (ca. 3 h). Danach erfolgte die Zugabe von PenicillinG zur
Unterdrückung der Zellwandsynthese in der Endkonzentration 10 µg/ml und die
weitere Inkubation bei 37°C bis zur OD600 von 0,5-0,7. Die Zellen wurden bei 3.000 x
g und 4°C für 20 min abzentrifugiert und anschließend dreimal in einem sich redu-
zierendem Volumen von 1 mM HEPES mit 0,5 M Sucrose gewaschen. Nach dem

Material und Methoden
131
letzten Waschschritt erfolgte die erneute Zentrifugation und die Aufnahme des
Zellsediments in 500 µl eiskaltem 1 mM HEPES mit 0,5 M Sucrose und 15% Gly-
cerin. Aliquots von 50 µl wurden bei -70°C gelagert (Park und Stewart, 1990).
Materialen: LmoXen32 CaliperLS LO28 Abteilung für
Molekulare Immunologie (MOLI), HZI
Plasmid pAUL-A-inlAm-inlB Abteilung für
Strukturbiologie (SB), HZI
BHI (Brain-Heart-Infusion) - Medium Becton & Dickinson
BHI + 0,5 M Sucrose:
2 x BHI (500ml) Becton & Dickinson 1 M Sucrose (500ml) Invitrogen Penicillin G USB
Waschpuffer: 1 mM HEPES + 0,5 M Sucrose
1 mM HEPES Roth 0,5 mM Sucrose Invitrogen
Aufnahmepuffer: 1 mM HEPES + 0,5 M Sucrose + 15% Glycerin (v/v) in Aquadest
1 mM HEPES Roth 0,5 mM Sucrose Invitrogen 15% (v/v) Glycerin Roth
Geräte: Zentrifuge Eppendorf 5804R Photometer Gene Quant pro Amersham Bioscience
5.1.2 Transformation und Selektion rekombinanter Listeria monocytogenes
Stämme
Die elektrokompetenten Listerien (50 µl) wurden mit 1 µg Vektor-DNA (eluiert in
Trispuffer) pAUL-A-inlAm-inlB über Elektroporation (2,5 kV, 25 µFd und 100 Ohm)
transformiert. Danach erfolgte die Zugabe von 1 ml BHI-Medium mit 0,5 M Sucrose
und die Inkubation bei 37°C für drei Stunden. Anschließend wurden Aliquots auf BHI-
Agarplatten mit 2 µg/ml Erythromycin (Ery) ausplattiert und über Nacht bei
Raumtemperatur (RT) kultiviert (Abbildung 5-1 (1)). Gewachsene Klone wurden auf

Material und Methoden
132
die erfolgreiche Transformation über den Nachweis der Erythromycinresistenz
selektioniert. Die plasmidtragenden Klone wurden erneut auf BHI-Agarplatten 2 µg/ml
Ery ausgestrichen und bei 42°C über Nacht kultiviert. Bei 42°C kann keine
extrachromosomale Replikation des Plasmides erfolgen und er kommt zur Integration
des Plasmides ins listerielle Genom (Abbildung 5-1 (2)). Eine erneute Anzucht auf
BHI-Ery-Agarplatten der gewachsen Klone diente der erneuten Selektion der
plasmidtragenden Klone. Im Anschluss erfolgte die Analyse der Integration in die
chromosomale DNA von Listeria monocytogenes mittels Präparation der
genomischen DNA, PCR und Sequenzierung des murinisierten inlA-Locus (siehe
Abschnitt 5.2). Zur Induzierung des Rekombinationsereignisses und Reversion des
Vektors wurde eine einzelne Kolonie in BHI-Flüssigkultur ohne Selektionsmarker bei
25°C angezogen. Anschließend sind 10 µl der Übernachtkultur in 50 ml frisches BHI-
Medium überimpft wurden und erneut bei 25°C kultiviert. Dieser Versuchsschritt
wurde dreimal wiederholt. Verdünnungen wurden auf BHI-Platten ausplattiert und
über Nacht ohne Selektionsmarker inkubiert. Ein Kolonie-Lift-up Assay wurde
durchgeführt und die gewachsenen Klone mittels sterilem Whatman-Filter auf BHI-
Ery-Agarplatten (2 µg/ml) überimpft. Nach dem Wachstum über Nacht erfolgte der
Vergleich der Platten mit und ohne Erythromycin. Klone, welche ihre
Erythromycinresistenz verloren hatten, zeigten ein Integrationsereignis über
homologe Rekombination mit Reversion des Vektors an (Abbildung 5-1 (3)). Zur
Analyse des Rekombinationsereignisses erfolgte die Präparation der chromosomale
DNA von Listeria monocytogenes und anschließender Validierung des murinisierten
Internalin A mittels PCR, Sequenzierung des inlA-Locus und der Western-Blot-
Analyse (siehe Abschnitt 5.2). Die Transformationsdurchführung zur Herstellung
genetisch modifizierter Listerienstämme ist in Abbildung 5-1 dargestellt.
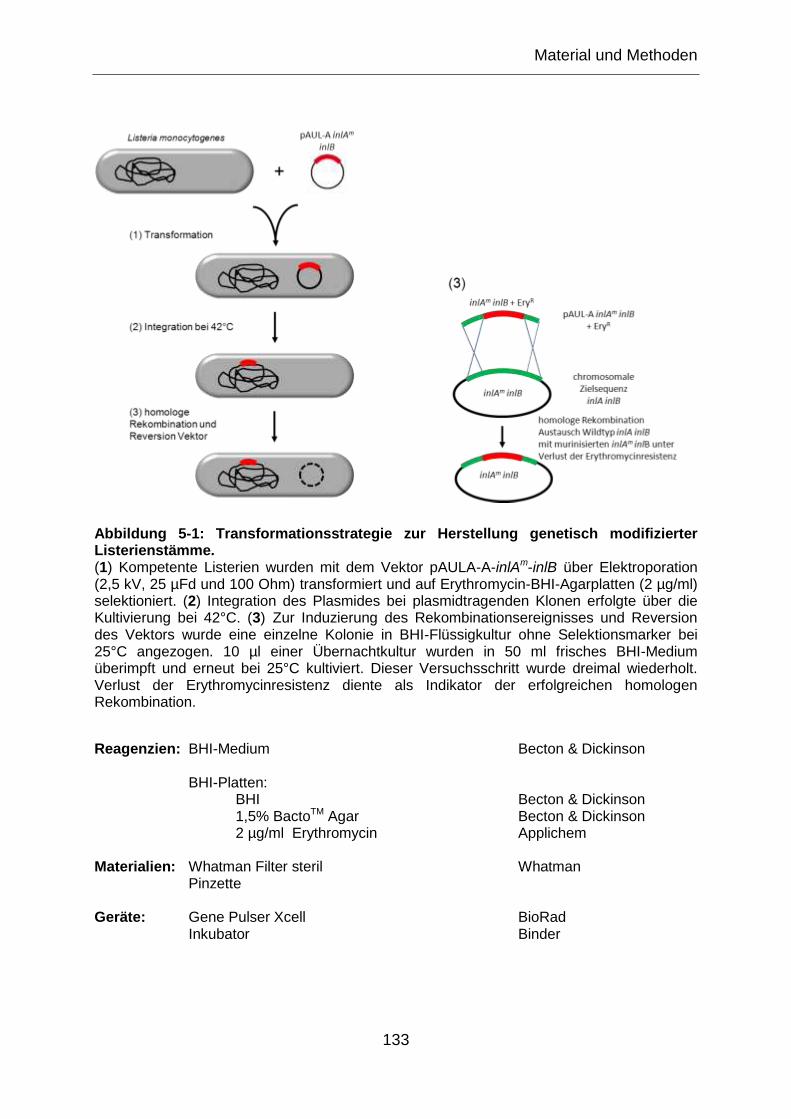
Material und Methoden
133
Abbildung 5-1: Transformationsstrategie zur Herstellung genetisch modifizierter Listerienstämme. (1) Kompetente Listerien wurden mit dem Vektor pAULA-A-inlAm-inlB über Elektroporation (2,5 kV, 25 µFd und 100 Ohm) transformiert und auf Erythromycin-BHI-Agarplatten (2 µg/ml) selektioniert. (2) Integration des Plasmides bei plasmidtragenden Klonen erfolgte über die Kultivierung bei 42°C. (3) Zur Induzierung des Rekombinationsereignisses und Reversion des Vektors wurde eine einzelne Kolonie in BHI-Flüssigkultur ohne Selektionsmarker bei 25°C angezogen. 10 µl einer Übernachtkultur wurden in 50 ml frisches BHI-Medium überimpft und erneut bei 25°C kultiviert. Dieser Versuchsschritt wurde dreimal wiederholt. Verlust der Erythromycinresistenz diente als Indikator der erfolgreichen homologen Rekombination.
Reagenzien: BHI-Medium Becton & Dickinson BHI-Platten: BHI Becton & Dickinson
1,5% BactoTM Agar Becton & Dickinson 2 µg/ml Erythromycin Applichem
Materialien: Whatman Filter steril Whatman
Pinzette Geräte: Gene Pulser Xcell BioRad Inkubator Binder

Material und Methoden
134
5.2 Molekularbiologische Methoden
5.2.1 Präparation von Plasmid-DNA
Die Präparation der Plasmid-DNA von transformierten Listerien erfolgte mit Hilfe des
Nucleo Spin® Plasmid Mini Kits von Macherey & Nagel. Dafür wurden 2 ml einer
Übernachtkultur 30 sec bei 13.000 x g zentrifugiert. Das Zellsediment wurde in Puffer
A1 mit 2 mg/ml Lysozym für 30 min bei 37°C inkubiert zur Lyse der Gram-positiven
Zellwand. Alle anschließenden Arbeitsschritte wurden gemäß des im Plasmid Mini Kit
mitgelieferten Versuchsprotokoll durchgeführt.
Materialien: Nucleo Spin® Plasmid Mini Kit Macherey & Nagel Lysozym Serva
5.2.2 Präparation genomischer bakterieller DNA
Die Präparation der genomischen DNA von Listeria monocytogenes erfolgte unter
Verwendung des Nucleo-Spin® Tissue Kit von Macherey & Nagel. Dafür wurden 2 ml
einer Übernachtkultur 5 min bei 8.000 x g zentrifugiert. Das Zellsediment wurde in
Lysepuffer mit 2 mg/ml Lysozym für 60 min bei 37°C inkubiert zur Lyse der Gram-
positiven Zellwand. Danach erfolgte die Zugabe von Proteinase K und die Inkubation
bei 56°C für 1-3 h. Alle anschließenden Arbeitsschritte erfolgten gemäß der im Kit
mitgelieferten Produkt- und Versuchsbeschreibung.
Materialien: Nucleo Spin® Tissue Kit Macherey & Nagel Lysepuffer: 20 mM Tris-HCl Sigma 2 mM EDTA Roth 1% Triton X-100 Applichem
2 mg/ml Lysozym Serva
5.2.3 PCR
Der Nachweis der homologen Rekombination des inlA-inlB Locus erfolgte über PCR.
Die PCR-Reaktionen wurden in dünnwandigen 0,2 ml Reaktionsgefäßen und auf Eis

Material und Methoden
135
durchgeführt. Die PCR-Bedingungen waren für alle durchgeführten PCR-Reaktionen
identisch.
Für die Amplifikation wurde folgender Reaktionsansatz verwendet:
H2O 1 x Reaktionspuffer (10 x, mit MgCl2) 10 mM dNTP-Mix 10 pmol/µl Primer 1 10 pmol/µl Primer 2 2 Unit (U) Taq Polymerase 200 ng DNA
Für die Amplifikation der Fragmente wurden folgende Oligonukleotide eingesetzt:
inlA int for 5`-GTG GAA CCG GTG ACG CAG-3´
inlA for 5`-ACG AGA TGA AGG ATA TCA CTA AAC G-3´
inlA 446 for 5`-TGT TCA ACA ATC AGA TAA CGG ATA TAG-3´
inlA 1356 rev 5´-TAC CGC CAT CGC TAA TAG TAG CTG-3´
inlB for 5`-AGC TAG ATG TGG TTT TCG GAC TAT ATC TAG-3´
inlB 3500 for 5`-ATG GCA TTT ACC AGA ATT TAC AAA TGA AG-3´
inlB 3670 rev 5`-AGT CGG AGG TTT AGG TGC AGT TAT CCG-3´
inlB rev 5`-CGC GGG AAT GCA GGC ATC TAC -3´
inlB rev-2 5`-GCA CCT ATA TTA GTC CGA TAT AGT TCT TG -3´
PCR-Bedingungen zur Amplifikation von genomischen DNA-Fragmenten:
95°C 5 min 95°C 1 min 58°C 1 min 35 Zyklen 72°C 1 min 72°C 10 min
Materialien: High Fidelity Taq Polymerase Roche 10 mM dNTP-Mix Invitrogen Primer 10mM MWG Operon Geräte: Thermocycler Biometra

Material und Methoden
136
5.2.4 Gelelektrische Auftrennung von DNA-Fragmenten
Die Auftrennung von PCR-Produkten erfolgte mittels Gelelektrophorese. Agarose-
gele und Puffer wurden mit TBE-Puffer hergestellt. Je nach Fragmentgröße sind 1-2
%ige Agarosegele mit 0,2 µg/ml Ethidiumbromid eingesetzt worden. Für die
Beladung des Geles wurden die Proben mit (6:1) Probenpuffer versetzt. Zusätzlich
wurde ein Größenmarker aufgetragen zur späteren Abschätzung der Fragment-
größe.
Die elektrophoretische Auftrennung erfolgte bei konstant gehaltener Spannung von 5
Volt / cm. Die Auswertung wurde mit Hilfe eines UV-Transilluminator Geldokumen-
tationsgerätes durchgeführt.
Materialien: TBE Puffer (10x): 89 mM Tris Sigma 89 mM Borsäure Roth 2 mM NaEDTA Roth Agarose Invitrogen Ethidiumbromid Serva Probenpuffer: in 1x TBE 50% Glycerin Roth 2% (w/v) Bromphenolblau Serva 2log DNA Leiter New England Biolabs Geräte: Gelelektrophorese Biometra Geldokumentationsgeräte Intas
5.2.5 Sequenzierung
Die Ausführung der Sequenzierungsreaktion erfolgte über den Service der Abteilung
Genomanalyse des Helmholtz-Zentrums für Infektionsforschung (HZI). Die zu
sequenzierenden Fragmente wurden aus dem Agarosegel präpariert und über
Gelextraktion aufgereinigt. Die aufgearbeiteten PCR-Produkte und entsprechende
Primer wurden an die Abteilung Genomanalyse, HZI weitergeleitet. Die erhaltenen
Sequenzen wurden dann mit Hilfe der Software MEGA 4.1 ausgewertet.

Material und Methoden
137
5.2.6 Proteinpräparation und Western-Blot-Analyse
5.2.6.1 Proteinextraktion
Die Expression der Oberflächenproteine Internalin A und Internalin B der murini-
sierten Listerienstämme wurden im Vergleich zu ihren isogenen Wildtyp-Stämmen
analysiert. Internalin A ist über eine kovalente Bindung in der Zellwand verankert,
während Internalin B nicht-kovalent über Lipoteichonsäuren an die bakterielle Zell-
wand gebunden ist. Aus diesem Grund erfolgte die separate Analyse von InlA im
Zellwandextrakt und der Nachweis von InlB im Zellkulturüberstand. Eine Übernacht-
kultur von Listeria monocytogenes wurde in 50 ml frisches BHI-Medium überimpft
und bei 37°C und 120 rpm bis zur OD600 von 1,0 kultiviert. Anschließend erfolgte die
Zentrifugation der Kultur bei 3500 x g für 30 min. Der Überstand diente für die
Extraktion der Proteine des Zellkulturüberstandes und das Zellsediment für die
Extraktion der Zellwandproteine. Der Überstand wurde in ein neues 50 ml Röhrchen
transferiert und 4,5 ml 100% Trichloressigsäure (w/v) und 4 ml Desoxycholat (1
mg/ml) zugegeben. Zur Präzipitation wurde der Überstand bei 4°C und über Nacht
inkubiert und dann am nächsten Tag bei 5500 x g für 60 min abzentrifugiert. Die
überstehende Flüssigkeit wurde entfernt und das Zellsediment in 2 ml Aceton
gewaschen, luftgetrocknet und in 200 µl SDS-Puffer resuspendiert. Anschließend
wurde das Proteinlysat mit Tris-HCl (1 M) pH 8.0 Puffer neutralisiert. Für die
Extraktion der Zellwandproteine wurde das nach der Zentrifugation der 50 ml
Flüssigkultur abgesetzte Zellsediment in 1,4 ml PBS mit 2% SDS resuspendiert und
eine Stunde bei 37°C und 300 rpm inkubiert. Anschließend erfolgte die Zentrifugation
der Zellen für 15 min bei 13000 x g. Der Überstand mit löslichen Bestandteilen wurde
in ein 2 ml Reaktionsgefäß transferiert. Durch Zugabe von 140 µl 100%
Trichloressigsäure (w/v) und 140 µl Desoxycholat (1 mg/ml) und der Inkubation über
Nacht bei 4°C erfolgte die Präzipitation der Proteine. Diese wurden am
darauffolgenden Tag 30 min bei 13000 x g abzentrifugiert, in 1 ml Aceton
gewaschen, luftgetrocknet und in 100 µl SDS-Puffer resuspendiert. Das Proteinlysat
wurde mit 1 M Tris-HCl pH 8.0 auf einen neutralen pH-Wert eingestellt.

Material und Methoden
138
Materialien: BHI-Medium Becton & Dickinson Trichloressigsäure (100% w/v) Roth
Desoxycholat Roth Aceton J.T. Baker
SDS-Puffer: 1% SDS Roth Tris-HCl pH 8.0: Tris Sigma HCl Merck
Listerienstämme: LmoEGDe (Serotyp 1/2a) Leibniz-Institut Deutsche Sammlung von Mikroorgansimen und
Zellkulturen GmbH (DSMZ)
LmoEGDe-mur Wollert et al., 2007 LmoXen32 Caliper LmoXen32-mur diese Arbeit LO28 MOLI, HZI LO28-mur diese Arbeit LmoEGDe-lux Riedel et al., 2007 LmoEGDe-mur-lux Monk et al., 2010
5.2.6.2 Western-Blot-Analyse
Die Bestimmung der Proteinkonzentration erfolgte unter Verwendung des Nanodrop
UV Spektrophotometer (Peqlab). Die Proteinextrakte wurden zunächst mit reduzie-
rendem Probenpuffer 1:1 versetzt und 5 min bei 95°C inkubiert. Anschließend
erfolgte die Auftrennung der Proteine über ein 10% SDS-Polyacrylamidgel in
Abhängigkeit von der Molekülgröße. Eine bessere Auftrennung wurde durch die
Verwendung eines zusätzlichen Sammelgels (5%) erreicht. Die Proteinproben und
ein Molekulargewichtsmarker wurden auf das Gel aufgetragen und bei 100 V für 90
min aufgetrennt.
Materialien: Probenpuffer 50 ml: 0,1 M Tris-HCl pH 6.8 Sigma
10% (w/v) Gycerin Roth 4% (w/v) SDS Roth 50 mg Bromphenolblau Serva 8% (v/v) β-Mercaptoethanol Sigma bei Bedarf frisch zugeben

Material und Methoden
139
Trenngel 10%: 10% Acrylamid-bis Roth
380 mM Tris pH 8.8 Sigma 0,1% (w/v) SDS Roth 0,1% (w/v) Ammoniumpersulfat Euroclone 0,04% (v/v) TEMED BioRad
Sammelgel: 5% Acrylamid-bis Roth
62 mM Tris pH 6.8 Sigma 0,1% (w/v) SDS Roth 0,1% (w/v) Ammoniumpersulfat Euroclone 0,1% (v/v) TEMED BioRad 10 x Laufpuffer pH 8.2:
250 mM Tris Sigma 2,5 M Glycin Roth 1% SDS Roth Molekulargewichtsmarker BioRad Geräte: Gelelektrophorese BioRad
Nach der gelelektrophoretischen Auftrennung wurden die Proteine für den Nachweis
der Expression von Internalin A (InlA), Internalin B (InlB) und dem Aktin-bindenden
Protein A (ActA) auf eine Immun-Blot Polyvinylidenfluorid (PVDF) Membran
transferiert. Dafür wurden die Polyacrylamidgele aus der Elektrophoreseapparatur
entfernt und auf, in Blotpuffer getränkte, Filterpapiere gelegt. Die im absoluten
Methanol aktivierte Immun-Blot PVDF Membran wurde luftblasenfrei auf das Gel
gebracht und diese erneut von einer Lage Filterpapier bedeckt. Der gesamte Aufbau
wurde in eine mit Transferpuffer gefüllte Blotkammer gebracht und die Proteine 3 h
bei 60 V elektrophoretisch auf die Membran transferiert. Nach dem Blotten wurde die
Membran zunächst zweimal in PBS-T (PBS mit 0,3% Tween 20) gewaschen und
anschließend in PBS-T mit 1% BSA zur Blockierung unspezifischer Bindungsstellen
2 h bei Raumtemperatur inkubiert. Es schlossen sich drei Waschschritte zu je 5 min
mit PBS-T und ein Waschschritt mit PBS für 5 min an. Danach erfolgte die Zugabe
der primären Antikörper (α-InlA, α-InlB und α-ActA) in der Verdünnung 1:1000 in PBS
und die Inkubation für 2 h bei Raumtemperatur bzw. über Nacht bei 4°C. Nicht
gebundene Antikörper würden über dreimaliges Waschen für 5 min mit PBS-T und
einem Waschschritt mit PBS für 5 min entfernt. Im Anschluss erfolgte die Zugabe des

Material und Methoden
140
Sekundärantikörpers (Anti-Maus Peroxidase konjugierter IgG(γ) polyklonal Anti-
körper) 1:2000 in PBS verdünnt und die Inkubation für 2 h bei Raumtemperatur.
Daraufhin wurde die Membran dreimal 5 min mit PBS-T und einmal mit PBS für 5
min gewaschen. Die Entwicklung der Membran erfolgte über die Umsetzung von
Diaminobenzidin unter Zugabe von Wasserstoffperoxid durch die Peroxidase zu
einer Braunfärbung.
Materialien: Transferpuffer: 1,25 mM Tris Sigma 192 mM Glycin Roth 20% (v/v) Methanol J.T. Baker PBS-T: PBS Gibco 0,3% (v/v) Tween 20 Sigma PBS-T BSA: PBS Gibco 0,3% (v/v) Tween 20 Sigma 1% (w/v) BSA Sigma
Antikörper: Anti-Internalin A (M160DbF11) Abteilung für Mouse monoclonal (IgG1) Experimentelle Immunologie
(EXIM), HZI Dr. Christian Erck
Anti-Internalin B (IC100F4) Lingnau et al., 1995 Mouse monoclonal (IgG1) EXIM, HZI
Anti-Lmo ActA (44g6s) Abnova Mouse monoclonal (IgG2b) Peroxidase-konjugiert Ziege KPL Anti-Maus IgG(γ) polyclonal Entwicklerlösung:
PBS Gibco 0,005% (w/v) Diaminobenzidin Sigma 0,0003% (v/v) Wasserstoffperoxid Merck Immun-Blot PVDF Membran BioRad Geräte: Blotkammer BioRad

Material und Methoden
141
5.3 Invasions-Assay
Zur Analyse ob eine verstärkte Aufnahme der murinisierten Listerienstämme
aufgrund einer erhöhten Bindungsaffinität vom murinisierten Internalin A zum
Rezeptor E-Cadherin erfolgt, wurde ein Gentamycin-Invasions-Assay durchgeführt
(Elsinghorst, 1994). Untersucht wurde die Aufnahme von LmoEGDe, LmoEGDe-mur,
LmoXen32, LmoXen32-mur, LO28, LO28-mur, LmoEGDe-lux und LmoEGDe-mur-lux
in humane Colonkarzinom-Zellen (Caco-2 Zellen) und in murine Colonkarzinom-
Zellen (CT-26 Zellen). Die Zellen wurden in MEM mit Earls´s Salz und fötales
Kälberserum (FKS), L-Glutamin, Natriumpyrovat und nichtessentieller Aminosäuren
bei 37°C und 7% CO2 kultiviert. Zwei Tage vor der Infektion wurden je 2 x 105 Zellen
pro Vertiefung einer 24-well-Platte ausgesät. Eine Übernachtkultur von Listeria
monocytogenes wurde 1:50 in frisches BHI-Medium überimpft und bei 37°C und 120
rpm bis zur mittleren Log-Wachstumsphase kultiviert. Die Listerien wurden zweimal
in MEM mit Earls´s Salz ohne FKS gewaschen und der konfluente Zellrasen mit 2 x
106 Bakterien pro Vertiefung infiziert. Die Platten wurden für 5 min bei 500 x g
zentrifugiert und 1 h bei 37°C und 7% CO2 inkubiert. Die Zellen wurden mit PBS
gewaschen und es erfolgte die Zugabe von Medium mit 100 µg/ml Gentamycin zur
Abtötung extrazellulär vorliegender Bakterien. Jeweils nach 1, 2, 3, und 4 h wurden
die Zellen mit PBS gewaschen und die Zellen unter Zugabe von sterilem H2O mit 0,2
% TritonX-100 lysiert. Serielle Verdünnungen wurden auf BHI-Platten ausplattiert und
für 24 h bei 37°C kultiviert. Gewachsenen Kolonien wurden gezählt und die Zellzahl
je „well“ berechnet. Die Bestimmung der Keimzahl erfolgte je Zeitpunkt in Triplikaten
und wurde dreimal wiederholt.
Materialien: Listerienstämme: LmoEGDe (Serotyp 1/2a) DSMZ LmoEGDe-mur Wollert et al., 2007 LmoXen32 Caliper LmoXen32-mur diese Arbeit LO28 MOLI, HZI LO28-mur diese Arbeit LmoEGDe-lux Riedel et al., 2007 LmoEGDe-mur-lux Monk et al., 2010
Zellen: Caco-2 ATCC HTB-37

Material und Methoden
142
CT-26 ATCC CRL-2638 Zellkulturmedium: MEM + Earle´s Gibco 20% FKS PAA 1% nicht-essentielle AS Gibco 2 mM Glutamin Gibco 1 mM Natriumpyrovat Gibco MEM + Earle´s Gibco 1% nicht-essentielle AS Gibco 2 mM Glutamin Gibco 1 mM Natriumpyrovat Gibco Lyse: H2O steril 0,2% Triton X-100 Applichem BHI-Platten: BHI Becton &Dickinson
1,5 % BactoTM Agar Becton &Dickinson 24-well-Platten TPP Geräte: Zentrifuge 5804R Eppendorf Zellkulturinkubator Heareus Inkubator Binder
5.4 In vivo Mausinfektionsexperimente
Alle Tierexperimente sowie die Tierhaltung und –zucht sind in den Einrichtungen des
Helmholtz-Zentrums für Infektionsforschung in Braunschweig unter SPF (specific
pathogen free) Bedingungen gemäß Tierversuchsantrag und der behördlichen
Genehmigung durchgeführt worden. Die Haltung der Tiere erfolgte mit Haltungsfutter
(sniff) auf Espe-Granulat-Einstreu mit Zugabe von Zellstoff als Nestbaumaterial.
Mausstämme: 129P2/Ola Harlan A/JOlaHsd Harlan BALB/cJ Janvier C3HeB/FeJ Charles River
C57BL/6J Janvier C57BL/6N Taconic Ifi27latm1(KOMP)Vlcg (Ifi27l2a-KO) Zucht, INFG BL6white/IFN-β+/Δβ-luc Zucht, MOLI, Dr. Stefan Lienenklaus

Material und Methoden
143
Listerienstämme: LmoXen32 CaliperLS LmoXen32-mur diese Arbeit
LO28 MOLI, HZI LO28-mur diese Arbeit LmoEGDe-lux Riedel et al., 2007 LmoEGDe-mur-lux Monk et al., 2010
5.4.1 Kultivierung und Präparation von Listeria monocytogenes
Von allen gentechnisch modifizierten oder bereitgestellten Listerienstämmen wurden
zunächst Glycerolstocks angefertigt. Dafür wurde von den Bakterien zunächst eine
Übernachtkultur in BHI-Medium angefertigt. Die Listerien wurden dann 1:10 in
frisches BHI-Medium überimpft und bis zu einer OD600 von 1,0 kultiviert. Hiervon
erfolgte die Präparation der Glycerolstocks. Dafür sind 430 µl Listerienkultur mit 570
µl sterilem Glycerin vermischt und anschließend bei -70°C gelagert worden.
Die Durchführung der Präparation der Listerien für die Infektion erfolgte nach
Vorschrift SOP (Standard Operating Procedure) LM-0007. Für die Infektion wurden
die Listerien aus einem aufgetauten Glycerolstock auf eine BHI-Platte ausgestrichen
und über Nacht bei 37°C kultiviert. Am nächsten Tag erfolgte die Anfertigung einer
Übernachtkultur von einer Kolonie in frischem BHI-Medium. Zur Animpfung der
Hauptkultur wurde die Übernachtkultur 1:10 in frisches BHI-Medium überimpft und
bis zum Erreichen der mittleren logarithmischen Wachstumsphase kultiviert. Die
Listerienkultur wurde für 5 min bei 1600 x g zentrifugiert, in vorgewärmten PBS
resuspendiert und erneut abzentrifugiert. Das Zellsediment wurde in 10 ml
vorgewärmten PBS vorsichtig resuspendiert und die Zellzahl über Auszählen
bestimmt.
Die Bestimmung der Listerienanzahl erfolgte nach dem Standardprotokoll SOP STD-
0002 über direktes Auszählen. Hierfür wurde von den resuspendierten Listerien eine
serielle Verdünnung angefertigt. Von einer Verdünnungsstufe ausgehend erfolgte die
Auszählung der Listerien in 5 x 16 Kleinquadraten einer THOMA Zählkammer (Tiefe
0,02 mm) mit vierhundertfacher Vergrößerung.
Die Berechnung erfolgte unter Nutzung der folgenden Formel:

Material und Methoden
144
Nachfolgend konnte die gewünschte Konzentration der Listeriensuspension mit PBS
eingestellt werden. Zur Bestimmung der tatsächlich verabreichten Infektionsdosis
wurden 50 µl der seriellen Verdünnungsstufen (1:10) auf BHI-Agarplatten aus-
plattiert, 24 h bei 37°C kultiviert, die gewachsenen Kolonien ausgezählt und die
CFU/ml berechnet.
Listerien: LmoXen32 CaliperLS LmoXen32-mur diese Arbeit LO28 MOLI, HZI LO28-mur diese Arbeit
LmoEGDe-lux Riedel et al., 2007 LmoEGDe-mur-lux Monk et al., 2010 Materialien: PBS Gibco
BHI-Medium: Becton & Dickinson
BHI-Platten: BHI Becton & Dickinson
1,5% BactoTM Agar Becton & Dickinson THOMA-Zählkammer (0,02) Karl Hecht - Assistent Geräte: Zentrifuge 5804R Eppendorf Zellkulturinkubator Heareus Inkubator Binder
5.4.2 Orale Infektion
Die Präparation der Listerien erfolgte wie in Punkt 5.4.1. beschrieben. Die
Durchführung der oralen Infektion erfolgte gemäß der Vorschrift SOP LM-0006. Nach
dem letzten Waschschritt wurden die Bakterien auf die gewünschte Infektionsdosis in
200 µl PBS verdünnt und 300 µl PBS mit 50 mg CaCO3 zugegeben (Lecuit et al.,
2001). Die Gesamtmenge des Inokulums betrug somit 500 µl und wurde den Mäusen
direkt über eine Schlundsonde (21G) in den Magen verabreicht. Als Voraussetzung
für die orale Infektion erfolgte ein Futterentzug in der vorangegangenen Nacht sowie
der Austausch des Trinkwassers mit gepuffertem Wasser (26 g NaHCO3 pro Liter

Material und Methoden
145
H2O, pH 8.0). Nach der Infektion hatten die Tiere wieder vollständigen Futterzugang.
Im weiteren Infektionsverlauf wurde täglich der Gesundheitszustand der Mäuse
protokolliert und bei einer drastischen Verschlechterung die Tiere gegebenenfalls
euthanisiert. Je nach Aufbau des Experimentes erfolgte zudem das tägliche Bio-
lumineszenz-Imaging (Abschnitt 5.4.4) bzw. die Präparation der Organe zur Bestim-
mung der bakteriellen Keimlast (Abschnitt 5.4.5).
Materialien: PBS Gibco CaCO3 Roth
NaHCO3 Roth
Knopfkanüle 1,0 x 40 mm (21G) Heiland 1 ml Insulinspritzen Therumo
5.4.3 Intravenöse Infektion
Die intravenöse Infektion erfolgte in Kooperation mit Dr. Bastian Pasche, Tierex-
perimentelle Einheit am Helmholtz-Zentrum für Infektionsforschung. Die Präparation
der Listerienzellen erfolgte wie in Punkt 5.4.1. beschrieben. Nach dem letzten
Waschschritt wurden die Bakterien auf die gewünschte Infektionsdosis in 100 µl PBS
verdünnt und in die laterale Schwanzvene verabreicht. Wie schon nach bei der
oralen Infektion beschrieben, erfolgte täglich die Protokollierung des Gesund-
heitszustandes, die Biolumineszenz-Messung und zu den gewünschten Zeitpunkten
die Präparation der Organe.
Materialien: 1 ml Einmalspritzen Braun Rotlichtquelle Philips
5.4.4 Biolumineszenz-Imaging
Das in vivo Biolumineszenz-Imaging (BLI) erlaubt eine nicht invasive Detektion von
emittierten Lichtsignalen. Photonen werden durch das Gewebe transmittiert und
generieren Lichtpunkte auf der Oberfläche des Tieres. Diese werden über eine CCD
Kamera (cooled coupeld device) detektiert und durch die Analyse des Systems wird
mit Hilfe von Falschfarben die Intensität des Lichtes in einem Bild dargestellt

Material und Methoden
146
(Abbildung 5-2). Die Daten werden als Photonen je Sekunde je Quadratzentimeter
und festem Raumwinkel (Photonen/s/cm2/sr) dargestellt.
Abbildung 5-2: Prinzip des in vivo Imaging. Photonen einer biolumineszenten Lichtquelle werden durch das Gewebe transmittiert und generieren ein diffuses Signal auf der Oberfläche des Tieres, welches über das Imaging System detektiert wird. Über die Analyse des Systems wird ein Bild generiert, welches unter Verwendung von Farben die Intensität des emittierten Lichtes darstellt.
5.4.4.1 In vivo Detektion der bakteriellen Luziferase
Für die Detektion der Biolumineszenz erfolgte eine Immobilisierung der Mäuse über
Narkotisierung mit Isofluran. Die Mäuse (max. 5) wurden in einer Narkosekammer
unter Verwendung eines Isofluranzerstäubers mit 2% des Anästhetikums in reinem
Sauerstoff narkotisiert. In die Narkosekammer strömten 2 l/min Sauerstoff, wobei die
Endkonzentration von Isofluran zur Einleitung der Narkose 6 ml/h betrug. Die narko-
tisierten Mäuse wurden in die Dunkelkammer des IVIS 200 Systems auf eine
Wärmeplatte gelegt, um sie vor einer narkosebedingten Auskühlung zu schützen.
Während der gesamten Biolumineszenzmessung wurde die Narkose unter der
Einleitung von 3 ml/h Isofluran und die Positionierung der Mäuse in Narkoseröhrchen
aufrechterhalten. Die Integrationszeit der Biolumineszenzmessung betrug 4 min
unter mittlerem „Binning“ (mittlere Sensitivität und mittlere Auflösung) und offener
Linse. Nach abgeschlossener Messung wurden die Mäuse wieder in einen Käfig
überführt. Die Auswertung erfolgte unter Verwendung der Living Image 3.2 Software
(CaliperLS).
Biolumineszente Lichtquelle
CCD
Imaging System

Material und Methoden
147
Material: Isofluran Abbott Animal Health Geräte: Xenogen IVIS 200 System CaliperLS Gas Anesthesia System Caliper LS Software: Living Image 3.2 Software CaliperLS
5.4.4.2 Ex vivo Detektion der bakteriellen Luziferase
Die für die Bestimmung der bakteriellen Keimlast wurden die präparierten Organe ex
vivo auf vorhandene Biolumineszenz untersucht. Die Organe wurden innerhalb von
Petrischalen in die Dunkelkammer des IVIS 200 Systems gelegt. Die Detektion der
Biolumineszenz erfolgte über eine Integrationszeit von 4 min, bei offener Linse und
mittleren „Binning“. Die Auswertung erfolgte mit der Living Image 3.2 Software
(CaliperLS).
Geräte: Xenogen IVIS 200 System CaliperLS Gas Anesthesia System Caliper LS Software: Living Image 3.2 Software CaliperLS
5.4.4.3 Detektion der IFN-β-Produktion
Das in vivo Imaging der IFN-β-Produktion erfolgte über die Luziferaseexpression als
Indikator. Den Mäusen wurde intravenös das Substrat D-Luziferin in PBS 150 mg/kg
Körpergewicht in die laterale Schwanzvene injiziert. Die Tiere wurden anschließend
mittels Isofluran-Narkose anästhesiert. Das in vivo Imaging erfolgte wie unter
Abschnitt 5.4.4.1 beschrieben.
Material: Isofluran Abbott Animal Health D-Luziferin 150 mg/kg in PBS Synchem 1 ml Einmalspritzen Braun Geräte: Xenogen IVIS 200 System CaliperLS Gas Anesthesia System Caliper LS Rotlichtquelle Philips Software: Living Image 3.2 Software CaliperLS

Material und Methoden
148
5.4.5 Bestimmung der bakteriellen Keimlast
Die Durchführung der Quantifizierung der Listerienanzahl erfolgte nach der SOP
Vorschrift LM-0003. Die Tiere wurden zu den entsprechenden Zeitpunkten mit CO2
(bei zusätzlicher Herzblutentnahme) oder durch zervikale Dislokation getötet und die
inneren Organe Milz, Leber, Dünndarm, mesenterische Lymphknoten und Gehirn
entnommen. Zunächst erfolgte die Detektion der Biolumineszenz ex vivo (siehe
Abschnitt 5.4.4). Der Dünndarm wurde über Ausstreichen entleert und 2 h in DMEM
mit 100 µg/ml Gentamycin zur Abtötung extrazellulär vorliegender Bakterien
inkubiert. Alle Organe wurden gewogen und in 5 ml vorgekühltem PBS (4°C) mit Hilfe
eines Stabhomogenisators zerkleinert. Zwischen der Homogenisierung der einzelnen
Organen erfolgte die Reinigung des Homogenisators in 2 x PBS, 1 x 70% Ethanol
und abschließend 1 x PBS für je 5 Sekunden (sec). Anschließend wurde von den
Homogenisaten eine serielle Verdünnungsreihe (1:10) angefertigt und 50 µl auf BHI-
Agarplatten ausgestrichen und 24 h bei 37°C kultiviert. Die gewachsenen Kolonien
wurden ausgezählt und die bakterielle Keimlast in Colony Forming Units (CFU) je
Milligramm Organ berechnet.
Materialien: PBS Gibco BHI-Platten: BHI Becton & Dickinson
BactoTM Agar Becton & Dickinson DMEM Gibco 100 µg/ml Gentamycin Serva Geräte: Waage Heareus
Stabhomogenisator Polytron Inkubator Binder
5.5 Luminex-Assay
Die Analyse der Chemokin- und Zytokinproduktion im Serum erfolgte mit Hilfe des
Invitrogen Multiplex-Bead-Immunoassay-Kits (Mouse Cytokine 20-Plex Panel).
Spektral kodierte Kügelchen dienen als Träger für die Antikörper gegen ausgewählte
Chemokine und Zytokine. Diese unterschiedlichen spektralen Eigenschaften der

Material und Methoden
149
Beads erlaubt die Unterscheidung und Quantifizierung der einzelnen chemotak-
tischen Stoffe.
Die Entnahme des Blutes zur Serumgewinnung erfolgte direkt aus dem Herzen der
zuvor mit CO2 getöteten Mäuse. Die entnommene Menge von 0,5 ml Blut wurde in
Serumröhrchen (Microtainer) überführt und mindestens 30 min bei Raumtemperatur
inkubiert. Anschließend erfolgte die Zentrifugation bei 13.000 x g für 2 min und die
Abnahme des Serums und Lagerung bei -70°C bis zur Versuchsdurchführung. Alle
weiteren Schritte sind anhand der Versuchsbeschreibung durchgeführt worden. Die
Messung und Auswertung erfolgte mit Hilfe des Luminex® xMAP® Systems.
Materialien: 1 ml Tuberkulinspritzen Braun Mircotainer Becton & Dickinson
Mouse Cytokine 20-Plex Panel Invitrogen Geräte: Ultraschallbad Elma Elmasonic Vortexer IKA Orbitalschüttler Polymax 1040 Heidolph Vakuumfiltrationseinheit BioRad Luminex® xMAP® Systems Invitrogen
5.6 Histologische und immunhistologische Färbung
Die Anfertigung der histologischen und immunhistologischen Präparate erfolgte über
die Histologieplattform des Helmholtz-Zentrums für Infektionsforschung. Die Prä-
paration der Organe erfolgte zu den gewünschten Zeitpunkten nach Tötung der
Mäuse über zervikale Dislokation bzw. CO2 Inhalation. Die Fixierung der Gewebs-
proben wurde für 24 h in 4% Formaldehyd durchgeführt. Anschließend erfolgte die
Entwässerung und Einbettung in Paraffin. Die Färbung der histologischen Schnitte
(Dicke 2 µm) erfolgte mit Hämatoxylin-Eosin. Für den immunhistologischen Nachweis
von Listeria monocytogenes wurden die Schnitte mit Hilfe des L. monocytogenes
Listeria-O-Antiserums (polyklonal Kaninchen) behandelt. Als Sekundärantikörper
wurde der Ziege-anti-Kaninchen-IgG genutzt. Die Gegenfärbung erfolgte mit Häma-
toxylin.
Materialien: L. monocytogenes Listeria-O-Antiserums Becton & Dickinson Ziege-anti-Kaninchen IgG (Biotin markiert) Thermo-Scientific

Material und Methoden
150
5.7 Statistische Auswertung
Die statistische Auswertung der Überlebensraten erfolgte über Kaplan-Meier und
Logrank Analyse. Die bakterielle Keimlast, Verlauf der Gewichtskurven und Kon-
zentration der Chemokine wurden als Mittelwert mit Standardfehler (SEM) dargestellt
und mit Hilfe des nicht-parametrischen Mann-Whitney-U-Tests analysiert. Die
Berechnungen erfolgten unter Nutzung des GraphPadPrism5 (GraphPad Software
Inc., San Diego, USA).

Literaturverzeichnis
151
6 Literaturverzeichnis
ABRAM, M., and DORIC, M. (1997). Primary Listeria monocytogenes infection in gestating mice. Folia Microbiol. (Praha) 42, 65–71.
ABRAM, M., SCHLÜTER, D., VUCKOVIC, D., WRABER, B., DORIC, M., and DECKERT, M.
(2003). Murine model of pregnancy‐associated Listeria monocytogenes infection. FEMS Immunology & Medical Microbiology 35, 177–182.
ANDERSEN, J.B., ROLDGAARD, B.B., LINDNER, A.B., CHRISTENSEN, B.B., and LICHT, T.R. (2006). Construction of a multiple fluorescence labelling system for use in co-invasion studies of Listeria monocytogenes. BMC Microbiol 6, 86.
ANTAL, E.A., LØBERG, E.M., BRACHT, P., MELBY, K.K., and MAEHLEN, J. (2001). Evidence for intraaxonal spread of Listeria monocytogenes from the periphery to the central nervous system. Brain Pathol. 11, 432–438.
ANTAL, E.A., LØBERG, E.M., DIETRICHS, E., and MAEHLEN, J. (2005). Neuropathological findings in 9 cases of listeria monocytogenes brain stem encephalitis. Brain Pathol. 15, 187–191.
ARCHINAL, W.A., and WILDER, M.S. (1988). Susceptibility of HRS/J mice to listeriosis: dynamics of infection. Infect. Immun. 56, 607–612.
ASAHI, O., and HOSODA, T. Studies on listeriosis in domestic animals. II. Studies on experimental listeriosis in goats. Med. Biol. (Japan) 100–103.
AUERBUCH, V., BROCKSTEDT, D.G., MEYER-MORSE, N., O’RIORDAN, M., and PORTNOY, D.A. (2004). Mice lacking the type I interferon receptor are resistant to Listeria monocytogenes. J. Exp. Med. 200, 527–533.
AURELI, P., FIORUCCI, G.C., CAROLI, D., MARCHIARO, G., NOVARA, O., LEONE, L., and SALMASO, S. (2000). An outbreak of febrile gastroenteritis associated with corn contaminated by Listeria monocytogenes. N. Engl. J. Med. 342, 1236–1241.
AUTENRIETH, I.B., BEER, M., BOHN, E., KAUFMANN, S.H., and HEESEMANN, J. (1994). Immune responses to Yersinia enterocolitica in susceptible BALB/c and resistant C57BL/6 mice: an essential role for gamma interferon. Infect. Immun. 62, 2590–2599.
BAKARDJIEV, A.I., STACY, B.A., FISHER, S.J., and PORTNOY, D.A. (2004). Listeriosis in the pregnant guinea pig: a model of vertical transmission. Infect. Immun 72, 489–497.
BAKARDJIEV, A.I., STACY, B.A. and PORTNOY, D.A. (2005). Growth of Listeria monocytogenes in the guinea pig placenta and role of cell-to-cell spread in fetal infection. J. Infect. Dis 191, 1889–1897.
BAKARDJIEV, A.I., THERIOT, J.A., and PORTNOY, D.A. (2006). Listeria monocytogenes traffics from maternal organs to the placenta and back. PLoS Pathog Jul;2(7):e80.
BALESTRINO, D., HAMON, M.A., DORTET, L., NAHORI, M.-A., PIZARRO-CERDA, J., ALIGNANI, D., DUSSURGET, O., COSSART, P., and TOLEDO-ARANA, A. (2010). Single-

Literaturverzeichnis
152
cell techniques using chromosomally tagged fluorescent bacteria to study Listeria monocytogenes infection processes. Appl. Environ. Microbiol 76, 3625–3636.
BARLOW, R.M., and McGORUM, B. (1985). Ovine listerial encephalitis: analysis, hypothesis and synthesis. Vet. Rec. 116, 233–236.
BEGLEY, M., GAHAN, C.G.M., and HILL, C. (2002). Bile stress response in Listeria monocytogenes LO28: adaptation, cross-protection, and identification of genetic loci involved in bile resistance. Appl. Environ. Microbiol. 68, 6005–6012.
BEGLEY, M., SLEATOR, R.D., GAHAN, C.G.M., and HILL, C. (2005). Contribution of Three Bile-Associated Loci, bsh, pva, and btlB, to Gastrointestinal Persistence and Bile Tolerance of Listeria monocytogenes. Infection and Immunity 73, 894–904.
BERGMANN, B., RAFFELSBAUER, D., KUHN, M., GOETZ, M., HOM, S., and GOEBEL, W. (2002). InlA- but not InlB-mediated internalization of Listeria monocytogenes by non-phagocytic mammalian cells needs the support of other internalins. Mol. Microbiol. 43, 557–570.
BIERNE, H., SABET, C., PERSONNIC, N., and COSSART, P. (2007). Internalins: a complex family of leucine-rich repeat-containing proteins in Listeria monocytogenes. Microbes and Infection 9, 1156–1166.
BIESTER, H.E., SCHWARTE, L.H., and EVELETH, D.F. (1939). Studies on a rapidly developing intestinal adenoma in a pig. Am. J. Pathol. 15, 385–389.2.
BIESTER, H.E., and SCHWARTE, L.H. (1940). Listerella infection in swine. J. Am. Vet. Med. Assoc. 96:339-342.
BIESTER, H.E., and SCHWARTE, L.H. (1941). Bovine listerellosis in Iowa with studies on a recovered case. North Am. Vet. 22:729-734.
BIGOT, A., PAGNIEZ, H., BOTTON, E., FREHEL, C., DUBAIL, I., JACQUET, C., CHARBIT, A., and RAYNAUD, C. (2005). Role of FliF and FliI of Listeria monocytogenes in flagellar assembly and pathogenicity. Infect. Immun. 73, 5530–5539.
BIRMINGHAM, C.L., CANADIEN, V., GOUIN, E., TROY, E.B., YOSHIMORI, T., COSSART, P., HIGGINS, D.E., and BRUMELL, J.H. (2007). Listeria monocytogenes evades killing by autophagy during colonization of host cells. Autophagy 3, 442–451.
BLOOM, W. (1928). The formation of abscesses in an infection with Bacterium monocytogenes. Arch. Pathol. 6:995-1007
BOHN, E., SING, A., ZUMBIHL, R., BIELFELDT, C., OKAMURA, H., KURIMOTO, M., HEESEMANN, J., and AUTENRIETH, I.B. (1998b). IL-18 (IFN-gamma-inducing factor) regulates early cytokine production in, and promotes resolution of, bacterial infection in mice. J. Immunol. 160, 299–307.
BOLLER, K., VESTWEBER, D., and KEMLER, R. (1985). Cell-adhesion molecule uvomorulin is localized in the intermediate junctions of adult intestinal epithelial cells. J. Cell Biol. 100, 327–332.

Literaturverzeichnis
153
BONAZZI, M., LECUIT, M., and COSSART, P. (2009). Listeria monocytogenes Internalin and E-Cadherin: From Bench to Bedside. Cold Spring Harbor Perspectives in Biology. Oct;1(4)
BOYARTCHUK, V.L., BROMAN, K.W., MOSHER, R.E., D’ORAZIO, S.E., STARNBACH, M.N., and DIETRICH, W.F. (2001). Multigenic control of Listeria monocytogenes susceptibility in mice. Nat. Genet. 27, 259–260.
BOYARTCHUK, V., ROJAS, M., YAN, B.-S., JOBE, O., HURT, N., DORFMAN, D.M., HIGGINS, D.E., DIETRICH, W.F., and KRAMNIK, I. (2004). The host resistance locus sst1 controls innate immunity to Listeria monocytogenes infection in immunodeficient mice. J. Immunol. 173, 5112–5120.
BRATTAIN, M.G., STROBEL-STEVENS, J., FINE, D., WEBB, M., and SARRIF, A.M. (1980). Establishment of mouse colonic carcinoma cell lines with different metastatic properties. Cancer Res. 40, 2142–2146.
BRAUN, L., and COSSART, P. (2000). Interactions between Listeria monocytogenes and host mammalian cells. Microbes and Infection 2, 803–811.
BRAUN, L., OHAYON, H., and COSSART, P. (1998). The InlB protein of Listeria monocytogenes is sufficient to promote entry into mammalian cells. Molecular Microbiology 27, 1077–1087.
BRINKMANN, V., REICHARD, U., GOOSMANN, C., FAULER, B., UHLEMANN, Y., WEISS, D.S., WEINRAUCH, Y., and ZYCHLINSKY, A. (2004). Neutrophil Extracellular Traps Kill Bacteria. Science 303, 1532–1535.
BRON, P.A., MONK, I.R., CORR, S.C., HILL, C., and GAHAN, C.G.M. (2006). Novel Luciferase Reporter System for In Vitro and Organ-Specific Monitoring of Differential Gene Expression in Listeria monocytogenes. Appl. Environ. Microbiol. 72, 2876–2884.
BROSCH, R., CHEN, J., and LUCHANSKY, J.B. (1994). Pulsed-field fingerprinting of listeriae: identification of genomic divisions for Listeria monocytogenes and their correlation with serovar. Appl. Environ. Microbiol. 60, 2584–2592.
BUCHMEIER, N.A., and SCHREIBER, R.D. (1985). Requirement of endogenous interferon-gamma production for resolution of Listeria monocytogenes infection. Proc. Natl. Acad. Sci. U.S.A. 82, 7404–7408.
BUCHRIESER, C. (2007). Biodiversity of the species Listeria monocytogenes and the genus Listeria. Microbes and Infection 9, 1147–1155.
BUCHRIESER, C., BROSCH, R., CATIMEL, B., and ROCOURT, J. (1993). Pulsed-field gel electrophoresis applied for comparing Listeria monocytogenes strains involved in outbreaks. Can. J. Microbiol. 39, 395–401.
BUER, D.H. and BALLING, R. (2003). Mice, microbes and models of Infection. Nat. Rev. Genet. 4, 195-205
BURN, C.G. (1936). Clinical and Pathological Features of an Infection Caused by a New Pathogen of the Genus Listerella. Am. J. Pathol. 12, 341–348.1.

Literaturverzeichnis
154
BURNS-GUYDISH, S.M., OLOMU, I.N., ZHAO, H., WONG, R.J., STEVENSON, D.K., and CONTAG, C.H. (2005). Monitoring age-related susceptibility of young mice to oral Salmonella enterica serovar Typhimurium infection using an in vivo murine model. Pediatr. Res. 58, 153–158.
CABANES, D., DEHOUX, P., DUSSURGET, O., FRANGEUL, L., and COSSART, P. (2002). Surface proteins and the pathogenic potential of Listeria monocytogenes. Trends Microbiol. 10, 238–245.
CABANES, D., SOUSA, S., CEBRIA, A., LECUIT, M., GARCIA-DEL PORTILLO, F., and COSSART, P. (2005). Gp96 is a receptor for a novel Listeria monocytogenes virulence factor, Vip, a surface protein. Embo J 24, 2827–2838.
CHAKRABORTY, T., LEIMEISTER-WÄCHTER, M., DOMANN, E., HARTL, M., GOEBEL, W., NICHTERLEIN, T., and NOTERMANS, S. (1992). Coordinate regulation of virulence genes in Listeria monocytogenes requires the product of the prfA gene. J Bacteriol 174, 568–574.
CHAMPY, M.-F., SELLOUM, M., ZEITLER, V., CARADEC, C., JUNG, B., ROUSSEAU, S., POUILLY, L., SORG, T., and AUWERX, J. (2008). Genetic background determines metabolic phenotypes in the mouse. Mamm. Genome 19, 318–331.
CHARLTON, K.M. (1977). Spontaneous listeric encephalitis in sheep. Electron microscopic studies. Vet. Pathol. 14, 429–434.
CHEERS, C., and MCKENZIE, I.F. (1978). Resistance and susceptibility of mice to bacterial infection: genetics of listeriosis. Infect. Immun. 19, 755–762.
CHEERS, C., MCKENZIE, I.F., PAVLOV, H., WAID, C., and York, J. (1978). Resistance and susceptibility of mice to bacterial infection: course of listeriosis in resistant or susceptible mice. Infect Immun 19, 763–770.
CHEERS, C., and STANLEY, E.R. (1988). Macrophage production during murine listeriosis: colony-stimulating factor 1 (CSF-1) and CSF-1-binding cells in genetically resistant and susceptible mice. Infect. Immun. 56, 2972–2978.
CHEN, Y., ZHANG, W., and KNABEL, S.J. (2007). Multi-virulence-locus sequence typing identifies single nucleotide polymorphisms which differentiate epidemic clones and outbreak strains of Listeria monocytogenes. J. Clin. Microbiol. 45, 835–846.
CLARK, M.A., HIRST, B.H., and JEPSON, M.A. (2000). Lectin-mediated mucosal delivery of drugs and microparticles. Advanced Drug Delivery Reviews 43, 207–223.
CLARK, M.A., and JEPSON, M.A. (2003). Intestinal M cells and their role in bacterial infection. International Journal of Medical Microbiology 293, 17–39.
CONTAG, C.H., and BACHMANN, M.H. (2002). Advances in in vivo bioluminescence imaging of gene expression. Annu Rev Biomed Eng 4, 235–260.
CONTAG, C.H., CONTAG, P.R., MULLINS, J.I., SPILMAN, S.D., STEVENSON, D.K., and BENARON, D.A. (1995). Photonic detection of bacterial pathogens in living hosts. Mol. Microbiol. 18, 593–603.

Literaturverzeichnis
155
CONTAG, C.H., and ROSS, B.D. (2002). It’s not just about anatomy: in vivo bioluminescence imaging as an eyepiece into biology. J Magn Reson Imaging 16, 378–387.
CONTAG, P.R. (2008). Bioluminescence imaging to evaluate infections and host response in vivo. Methods Mol. Biol. 415, 101–118.
COOPER, G.L. (1989). An encephalitic form of listeriosis in broiler chickens. Avian Dis. 33, 182–185.
COOPER, J., and WALKER, R.D. (1998). Listeriosis. Vet. Clin. North Am. Food Anim. Pract. 14, 113–125.
CORBETT, T.H., GRISWOLD, D.P., JR, ROBERTS, B.J., PECKHAM, J.C., and SCHABEL, F.M., Jr (1975). Tumor induction relationships in development of transplantable cancers of the colon in mice for chemotherapy assays, with a note on carcinogen structure. Cancer Res. 35, 2434–2439.
CORR, S., HILL, C., and GAHAN, C.G.M. (2006). An in vitro cell-culture model demonstrates internalin- and hemolysin-independent translocation of Listeria monocytogenes across M cells. Microb. Pathog. 41, 241–250.
CORR, S.C., and O’NEILL, L.A.J. (2009). Listeria monocytogenes infection in the face of innate immunity. Cellular Microbiology 11, 703–709.
COSSART, P., and TOLEDO-ARANA, A. (2008). Listeria monocytogenes, a unique model in infection biology: an overview. Microbes and Infection 10, 1041–1050.
COTTER, P.D., GAHAN, C.G., and HILL, C. (2001). A glutamate decarboxylase system protects Listeria monocytogenes in gastric fluid. Mol. Microbiol. 40, 465–475.
COTTER, P.D., and HILL, C. (2003). Surviving the acid test: responses of gram-positive bacteria to low pH. Microbiol. Mol. Biol. Rev. 67, 429–453.
CZUPRYNSKI, C.J., CANONO, B.P., HENSON, P.M., and CAMPBELL, P.A. (1985). Genetically determined resistance to listeriosis is associated with increased accumulation of inflammatory neutrophils and macrophages which have enhanced listericidal activity. Immunology 55, 511–518.
CZUPRYNSKI, C.J., and FAITH, N.G. (2002). Sodium Bicarbonate Enhances the Severity of Infection in Neutropenic Mice Orally Inoculated with Listeria monocytogenes EGD. Clinical and Diagnostic Laboratory Immunology 9, 477–481.
CZUPRYNSKI, C.J., FAITH, N.G., and STEINBERG, H. (2003). A/J Mice Are Susceptible and C57BL/6 Mice Are Resistant to Listeria monocytogenes Infection by Intragastric Inoculation. Infect Immun 71, 682–689.
DANIELS, J.J., AUTENRIETH, I.B., and GOEBEL, W. (2000). Interaction of Listeria monocytogenes with the intestinal epithelium. FEMS Microbiol. Lett. 190, 323–328.
DECKER, R.A., ROGERS, J.J., and LESAR, S. (1976). Listeriosis in a young cat. J. Am. Vet. Med. Assoc. 168, 1025.
DEDIÈ, K. (1955). Beitrag zur Epizootologie der Listeriose. Arch. Exptl. Veterinaermed. 9:251- 264.

Literaturverzeichnis
156
DELUCA, M., and MCELROY, W.D. (1974). Kinetics of the firefly luciferase catalyzed reactions. Biochemistry 13, 921–925.
DENNY, J., and MCLAUCHLIN, J. (2008). Human Listeria monocytogenes infections in Europe--an opportunity for improved European surveillance. Euro Surveill. 13.
VAN DEUREN, M., BRANDTZAEG, P., and VAN DER MEER, J.W.M. (2000). Update on Meningococcal Disease with Emphasis on Pathogenesis and Clinical Management. Clin. Microbiol. Rev. 13, 144–166.
DISSON, O., GRAYO, S., HUILLET, E., NIKITAS, G., LANGA-VIVES, F., DUSSURGET, O., RAGON, M., LE MONNIER, A., BABINET, C., and COSSART, P. (2008). Conjugated action of two species-specific invasion proteins for fetoplacental listeriosis. Nature 455, 1114–1118.
DISSON, O., NIKITAS, G., GRAYO, S., DUSSURGET, O., COSSART, P., and LECUIT, M. (2009). Modeling human listeriosis in natural and genetically engineered animals. Nat. Protocols 4, 799–810.
DOMANN, E., WEHLAND, J., ROHDE, M., PISTOR, S., HARTL, M., GOEBEL, W., LEIMEISTER-WÄCHTER, M., WUENSCHER, M., and CHAKRABORTY, T. (1992). A novel bacterial virulence gene in Listeria monocytogenes required for host cell microfilament interaction with homology to the proline-rich region of vinculin. Embo J. 11, 1981–1990.
DONKER-VOET, J. (1972). Proceedings: Listeria monocytogenes: some biochemical and serological aspects. Acta Microbiol Acad Sci Hung 19, 287–291.
DONS, L., JIN, Y., KRISTENSSON, K., and ROTTENBERG, M.E. (2007). Axonal transport of Listeria monocytogenes and nerve-cell-induced bacterial killing. J. Neurosci. Res. 85, 2529–2537.
DONS, L., WECLEWICZ, K., JIN, Y., BINDSEIL, E., OLSEN, J.E., and KRISTENSSON, K. (1999). Rat dorsal root ganglia neurons as a model for Listeria monocytogenes infections in culture. Med. Microbiol. Immunol. 188, 15–21.
DOUMITH, M., CAZALET, C., SIMOES, N., FRANGEUL, L., JACQUET, C., KUNST, F., MARTIN, P., COSSART, P., GLASER, P., and BUCHRIESER, C. (2004). New aspects regarding evolution and virulence of Listeria monocytogenes revealed by comparative genomics and DNA arrays. Infect. Immun. 72, 1072–1083.
DOYLE, T.C., BURNS, S.M., and CONTAG, C.H. (2004). In vivo bioluminescence imaging for integrated studies of infection. Cell. Microbiol. 6, 303–317.
DRAMSI, S., BOURDICHON, F., CABANES, D., LECUIT, M., FSIHI, H., and COSSART, P. (2004). FbpA, a novel multifunctional Listeria monocytogenes virulence factor. Molecular Microbiology 53, 639–649.
DRAMSI, S., LÉVI, S., TRILLER, A., and COSSART, P. (1998). Entry of Listeria monocytogenes into Neurons Occurs by Cell-to-Cell Spread: an In Vitro Study. Infection and Immunity 66, 4461 –4468.
DREVETS, D.A. (1999). Dissemination of Listeria monocytogenes by Infected Phagocytes. Infection and Immunity 67, 3512–3517.

Literaturverzeichnis
157
DREVETS, D.A., and BRONZE, M.S. (2008). Listeria monocytogenes: epidemiology, human disease, and mechanisms of brain invasion. FEMS Immunology & Medical Microbiology 53, 151–165.
DREVETS, D.A., JELINEK, T.A., and FREITAG, N.E. (2001). Listeria monocytogenes-infected phagocytes can initiate central nervous system infection in mice. Infection and Immunity 69, 1344 –1350.
DREVETS, D.A., LEENEN, P.J., and CAMPBELL, P.A. (1993). Complement receptor type 3 (CD11b/CD18) involvement is essential for killing of Listeria monocytogenes by mouse macrophages. J. Immunol. 151, 5431–5439.
DREVETS, D.A., LEENEN, P.J.M., and GREENFIELD, R.A. (2004). Invasion of the central nervous system by intracellular bacteria. Clinical Microbiology Reviews 17, 323–347.
DREVETS, D.A., SCHAWANG, J.E., DILLON, M.J., LERNER, M.R., BRONZE, M.S., and BRACKETT, D.J. (2008). Innate Responses to Systemic Infection by Intracellular Bacteria Trigger Recruitment of Ly-6Chigh Monocytes to the Brain. The Journal of Immunology 181, 529–536.
EBBES, M., BLEYMÜLLER, W.M., CERNESCU, M., NÖLKER, R., BRUTSCHY, B., and NIEMANN, H.H. (2011). Fold and function of the InlB B-repeat. J. Biol. Chem. 286, 15496–15506.
EDELSON, B.T., and UNANUE, E.R. (2001). Intracellular antibody neutralizes Listeria growth. Immunity 14, 503–512.
EDELSON, B.T., and UNANUE, E.R. (2002). MyD88-dependent but Toll-like receptor 2-independent innate immunity to Listeria: no role for either in macrophage listericidal activity. J. Immunol. 169, 3869–3875.
ELSINGHORST, E.A. (1994). Measurement of invasion by gentamicin resistance. Meth. Enzymol. 236, 405–420.
ENGEBRECHT, J., SIMON, M., and SILVERMAN, M. (1985). Measuring gene expression with light. Science 227, 1345–1347.
Epidemiologisches Bulletin. (2006). Listeriose (RKI).
Epidemiologisches Bulletin. (2010). Listeriose 2001 bis 2009 (RKI).
FARBER, J.M., and PETERKIN, P.I. (1991). Listeria monocytogenes, a food-borne pathogen. Microbiol. Rev. 55, 476–511.
Farber, J.M., Peterkin, P.I., Carter, A.O., Varughese, P.V., Ashton, F.E., and Ewan, E.P. (1991). Neonatal listeriosis due to cross-infection confirmed by isoenzyme typing and DNA fingerprinting. J. Infect. Dis. 163, 927–928.
FENLON, D.R. (1986). Rapid quantitative assessment of the distribution of Listeria in silage implicated in a suspected outbreak of listeriosis in calves. Vet. Rec. 118, 240–242.
FLANNAGAN, R.S., COSÍO, G., and GRINSTEIN, S. (2009). Antimicrobial mechanisms of phagocytes and bacterial evasion strategies. Nat. Rev. Microbiol. 7, 355–366.

Literaturverzeichnis
158
FRANCIS, K.P., JOH, D., BELLINGER-KAWAHARA, C., HAWKINSON, M.J., PURCHIO, T.F., and CONTAG, P.R. (2000). Monitoring bioluminescent Staphylococcus aureus infections in living mice using a novel luxABCDE construct. Infect. Immun 68, 3594–3600.
FRANCIS, K.P., YU, J., BELLINGER-KAWAHARA, C., JOH, D., HAWKINSON, M.J., XIAO, G., PURCHIO, T.F., CAPARON, M.G., LIPSITCH, M., and CONTAG, P.R. (2001). Visualizing pneumococcal infections in the lungs of live mice using bioluminescent Streptococcus pneumoniae transformed with a novel gram-positive lux transposon. Infect. Immun 69, 3350–3358.
GAHAN, C.G.., and HILL, C. (2005). Gastrointestinal phase of Listeria monocytogenes infection. Journal of Applied Microbiology 98, 1345–1353.
GATES, B.J., and DELUCA, M. (1975). The production of oxyluciferin during the firefly luciferase light reaction. Arch. Biochem. Biophys. 169, 616–621.
GELLIN, B.G., and BROOME, C.V. (1989). Listeriosis. Jama 261, 1313–1320.
GERVAIS, F., STEVENSON, M., and SKAMENE, E. (1984). Genetic control of resistance to Listeria monocytogenes: regulation of leukocyte inflammatory responses by the Hc locus. J. Immunol. 132, 2078–2083.
GILL, D.A. Ovine bacterial encephalitis (Circling Disease) and the bacterial genus Listerella. 1937 Australian Vet. J. 13:46-56.
GLASER, P., FRANGEUL, L., BUCHRIESER, C., RUSNIOK, C., AMEND, A., BAQUERO, F., BERCHE, P., BLOECKER, H., BRANDT, P., and CHAKRABORTY, T. (2001). Comparative genomics of Listeria species. Science 294, 849–852.
GALSWORTHY, S.B., and FEWSTER, D. (1988). Comparison of responsiveness to the monocytosis-producing activity of Listeria monocytogenes in mice genetically susceptible or resistant to listeriosis. Infection 16 Suppl 2, S118–122.
GOLDMANN, O., LENGELING, A., BÖSE, J., BLOECKER, H., GEFFERS, R., CHHATWAL, G.S., and MEDINA, E. (2005). The role of the MHC on resistance to group a streptococci in mice. J. Immunol. 175, 3862–3872.
GOUIN, E., ADIB-CONQUY, M., BALESTRINO, D., NAHORI, M.-A., VILLIERS, V., COLLAND, F., DRAMSI, S., DUSSURGET, O., and COSSART, P. (2010). The Listeria monocytogenes InlC protein interferes with innate immune responses by targeting the IκB kinase subunit IKKα. Proceedings of the National Academy of Sciences 107, 17333 –17338.
GRAHAM, R., LEVINE, N.D., and MORRILL, C.C. (1943). Listerellosis in domestic animals. Univ. Ill. Agr. Expt. Sta. Bull. 499.
GRAVES, L.M., HELSEL, L.O., STEIGERWALT, A.G., MOREY, R.E., DANESHVAR, M.I., ROOF, S.E., ORSI, R.H., FORTES, E.D., MILILLO, S.R. and DEN BAKKER, H.C. (2010). Listeria marthii sp. nov., isolated from the natural environment, Finger Lakes National Forest. Int. J. Syst. Evol. Microbiol. 60, 1280–1288.
GRAVES, L.M., SWAMINATHAN, B., REEVES, M.W., HUNTER, S.B., WEAVER, R.E., PLIKAYTIS, B.D., and SCHUCHAT, A. (1994). Comparison of ribotyping and multilocus enzyme electrophoresis for subtyping of Listeria monocytogenes isolates. J. Clin. Microbiol. 32, 2936–2943.

Literaturverzeichnis
159
GRAY, M.L. Discussions. (1958). In E. Roots and D. Strauch [ed.], Listeriosen. Beiheft I, Zentr. Veterinaermed. Paul Paray Verlag, Berlin.
GRAY, M.L., and KILLINGER, A.H. (1966). Listeria monocytogenes and listeric infections. Bacteriol Rev 30, 309–382.
GREER, L.F., 3rd, and SZALAY, A.A. (2002). Imaging of light emission from the expression of luciferases in living cells and organisms: a review. Luminescence 17, 43–74.
GREIFFENBERG, L., GOEBEL, W., KIM, K.S., WEIGLEIN, I., BUBERT, A., ENGELBRECHT, F., STINS, M., and KUHN, M. (1998). Interaction of Listeria monocytogenes with human brain microvascular endothelial cells: InlB-dependent invasion, long-term intracellular growth, and spread from macrophages to endothelial cells. Infect. Immun. 66, 5260–5267.
GRÜNDLING, A., BURRACK, L.S., BOUWER, H.G.A., and HIGGINS, D.E. (2004). Listeria monocytogenes regulates flagellar motility gene expression through MogR, a transcriptional repressor required for virulence. Proc. Natl. Acad. Sci. U.S.A. 101, 12318–12323.
GRÜTZKAU, A., HANSKI, C., HAHN, H., and RIECKEN, E.O. (1990). Involvement of M cells in the bacterial invasion of Peyer’s patches: a common mechanism shared by Yersinia enterocolitica and other enteroinvasive bacteria. Gut 31, 1011–1015.
GRYCZAN, T.J., HAHN, J., CONTENTE, S., and DUBNAU, D. (1982). Replication and incompatibility properties of plasmid pE194 in Bacillus subtilis. J. Bacteriol. 152, 722–735.
GUILLET, C., JOIN-LAMBERT, O., LE MONNIER, A., LECLERCQ, A., MECHAÏ, F., MAMZER-BRUNEEL, M.F., BIELECKA, M.K., SCORTTI, M., DISSON, O. and BERCHE, P. (2010). Human listeriosis caused by Listeria ivanovii. Emerging Infect. Dis. 16, 136–138.
GULERIA, I., and POLLARD, J.W. (2001). Aberrant macrophage and neutrophil population dynamics and impaired Th1 response to Listeria monocytogenes in colony-stimulating factor 1-deficient mice. Infect. Immun. 69, 1795–1807.
HAGBERG, L., BRILES, D.E., and EDÉN, C.S. (1985). Evidence for separate genetic defects in C3H/HeJ and C3HeB/FeJ mice, that affect susceptibility to gram-negative infections. J. Immunol. 134, 4118–4122.
HAMON, M., BIERNE, H., and COSSART, P. (2006). Listeria monocytogenes: a multifaceted model. Nat. Rev. Microbiol 4, 423–434.
HAMON, M.A., BATSCHÉ, E., RÉGNAULT, B., THAM, T.N., SEVEAU, S., MUCHARDT, C., and COSSART, P. (2007). Histone modifications induced by a family of bacterial toxins. Proc. Natl. Acad. Sci. U.S.A. 104, 13467–13472.
HAMON, M.A., and COSSART, P. (2011). K+ Efflux Is Required for Histone H3 Dephosphorylation by Listeria monocytogenes Listeriolysin O and Other Pore-Forming Toxins. Infection and Immunity 79, 2839 –2846.
HANSKI, C., KUTSCHKA, U., SCHMORANZER, H.P., NAUMANN, M., STALLMACH, A., HAHN, H., MENGE, H., and RIECKEN, E.O. (1989). Immunohistochemical and electron microscopic study of interaction of Yersinia enterocolitica serotype O8 with intestinal mucosa during experimental enteritis. Infect. Immun. 57, 673–678.

Literaturverzeichnis
160
HARDY, J., CHU, P., and CONTAG, C.H. (2009). Foci of Listeria monocytogenes persist in the bone marrow. Dis Model Mech 2, 39–46.
HARDY, J., FRANCIS, K.P., DEBOER, M., CHU, P., GIBBS, K., and CONTAG, C.H. (2004). Extracellular replication of Listeria monocytogenes in the murine gall bladder. Science 303, 851–853.
HARDY, J., MARGOLIS, J.J., and CONTAG, C.H. (2006). Induced biliary excretion of Listeria monocytogenes. Infect. Immun 74, 1819–1827.
HARTY, J.T., and BEVAN, M.J. (1995). Specific immunity to Listeria monocytogenes in the absence of IFN gamma. Immunity 3, 109–117.
HASTINGS, J.W., and NEALSON, K.H. (1977). Bacterial bioluminescence. Annu. Rev. Microbiol. 31, 549–595.
HAVELL, E.A. (1987). Production of tumor necrosis factor during murine listeriosis. J. Immunol. 139, 4225–4231.
HAVELL, E.A., BERETICH, G.R., JR, and CARTER, P.B. (1999). The mucosal phase of Listeria infection. Immunobiology 201, 164–177.
DE LAS HERAS, A., CAIN, R.J., BIELECKA, M.K., and VÁZQUEZ-BOLAND, J.A. (2011). Regulation of Listeria virulence: PrfA master and commander. Curr. Opin. Microbiol. 14, 118–127.
HO, J.L., SHANDS, K.N., FRIEDLAND, G., ECKIND, P., and FRASER, D.W. (1986). An outbreak of type 4b Listeria monocytogenes infection involving patients from eight Boston hospitals. Arch. Intern. Med. 146, 520–524.
HSIEH, C.S., MACATONIA, S.E., TRIPP, C.S., WOLF, S.F., O’GARRA, A., and MURPHY, K.M. (1993). Development of TH1 CD4+ T cells through IL-12 produced by Listeria-induced macrophages. Science 260, 547–549.
HUANG, S., HENDRIKS, W., ALTHAGE, A., HEMMI, S., BLUETHMANN, H., KAMIJO, R., VILCEK, J., ZINKERNAGEL, R.M., and AGUET, M. (1993). Immune response in mice that lack the interferon-gamma receptor. Science 259, 1742–1745.
IIZAWA, Y., WAGNER, R.D., and CZUPRYNSKI, C.J. (1993). Analysis of cytokine mRNA expression in Listeria-resistant C57BL/6 and Listeria-susceptible A/J mice during Listeria monocytogenes infection. Infect. Immun. 61, 3739–3744.
ISBERG, R.R., and LEONG, J.M. (1990). Multiple beta 1 chain integrins are receptors for invasin, a protein that promotes bacterial penetration into mammalian cells. Cell 60, 861–871.
ISLAM, S., MAHALANABIS, D., CHOWDHURY, A.K., SARKER ABDUL WAHED, S.A., and RAHMAN, A.S. (1996). Intestinal transport of water, sodium & glucose from an electrolyte solution with & without bicarbonate. Indian J. Med. Res. 104, 254–256.
JACQUET, C., DOUMITH, M., GORDON, J.I., MARTIN, P.M.V., COSSART, P., and LECUIT, M. (2004). A molecular marker for evaluating the pathogenic potential of foodborne Listeria monocytogenes. J. Infect. Dis 189, 2094–2100.

Literaturverzeichnis
161
JENSEN, R.L., PEDERSEN, K.S., LOESCHCKE, V., INGMER, H., and LEISNER, J.J. (2007). Limitations in the use of Drosophila melanogaster as a model host for gram-positive bacterial infection. Lett. Appl. Microbiol. 44, 218–223.
JENSEN, V.B., HARTY, J.T., and JONES, B.D. (1998). Interactions of the Invasive PathogensSalmonella typhimurium, Listeria monocytogenes, and Shigella flexneri with M Cells and Murine Peyer’s Patches. Infection and Immunity 66, 3758 –3766.
JEPSON, M.A., and CLARK, M.A. (2001). The role of M cells in Salmonella infection. Microbes Infect. 3, 1183–1190.
JIN, Y., DONS, L., KRISTENSSON, K., and ROTTENBERG, M.E. (2001). Neural route of cerebral Listeria monocytogenes murine infection: role of immune response mechanisms in controlling bacterial neuroinvasion. Infect. Immun 69, 1093–1100.
JOIN-LAMBERT, O.F., EZINE, S., LE MONNIER, A., JAUBERT, F., OKABE, M., BERCHE, P., and KAYAL, S. (2005). Listeria monocytogenes-infected bone marrow myeloid cells promote bacterial invasion of the central nervous system. Cell. Microbiol 7, 167–180.
JONES, S.A. (2005). Directing transition from innate to acquired immunity: defining a role for IL-6. J. Immunol. 175, 3463–3468.
JONQUIERES, R., BIERNE, H., MENGAUD, J., and COSSART, P. (1998). The inlA Gene of Listeria monocytogenes LO28 Harbors a Nonsense Mutation Resulting in Release of Internalin. Infect. Immun. 66, 3420–3422.
JUNG, W., CASTREN, E., ODENTHAL, M., VANDE WOUDE, G.F., ISHII, T., DIENES, H.P., LINDHOLM, D., and SCHIRMACHER, P. (1994). Expression and functional interaction of hepatocyte growth factor-scatter factor and its receptor c-met in mammalian brain. J. Cell Biol. 126, 485–494.
KAUTTER, D.A., SILVERMAN, S.J., and ROESSLER, W.G. (1959). Studies on the virulence of Listeria monocytogenes via the respiratory route for laboratory animals. Bacteriol. Proc., p. 95.
KAUTTER, D.A., SILVERMAN, S.J., ROESSLER, W.G. and DRAWDY, J.F. (1963). Virulence of Listeria monocytogenes for experimental animals. J. Infect. Dis. 112, 167–180.
KHELEF, N., LECUIT, M., BIERNE, H., and COSSART, P. (2006). Species specificity of the Listeria monocytogenes InlB protein. Cell. Microbiol 8, 457–470.
KIM, H., MARQUIS, H., and BOOR, K.J. (2005). σB contributes to Listeria monocytogenes invasion by controlling expression of inlA and inlB. Microbiology 151, 3215 –3222.
KLATT, E.C., PAVLOVA, Z., TEBERG, A.J., and YONEKURA, M.L. (1986). Epidemic perinatal listeriosis at autopsy. Hum. Pathol. 17, 1278–1281.
KRAEHENBUHL, J.P., and NEUTRA, M.R. (2000). Epithelial M cells: differentiation and function. Annu Rev Cell Dev Biol. 16, 301-32.
KRAMNIK, I., DIETRICH, W.F., DEMANT, P., and BLOOM, B.R. (2000). Genetic control of resistance to experimental infection with virulent Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. U.S.A. 97, 8560–8565.

Literaturverzeichnis
162
KURSAR, M., BONHAGEN, K., FENSTERLE, J., KÖHLER, A., HURWITZ, R., KAMRADT, T., KAUFMANN, S.H.E., and MITTRÜCKER, H.-W. (2002). Regulatory CD4+CD25+ T cells restrict memory CD8+ T cell responses. J. Exp. Med. 196, 1585–1592.
LARA-TEJERO, M., and PAMER, E.G. (2004). T cell responses to Listeria monocytogenes. Curr. Opin. Microbiol. 7, 45–50.
LAUER, P., CHOW, M.Y.N., LOESSNER, M.J., PORTNOY, D.A., and CALENDAR, R. (2002). Construction, characterization, and use of two Listeria monocytogenes site-specific phage integration vectors. J. Bacteriol. 184, 4177–4186.
LEBER, J.H., CRIMMINS, G.T., RAGHAVAN, S., MEYER-MORSE, N.P., COX, J.S., and PORTNOY, D.A. (2008). Distinct TLR- and NLR-Mediated Transcriptional Responses to an Intracellular Pathogen. PLoS Pathog 4 (1), e6.
LECLERCQ, A., CLERMONT, D., BIZET, C., GRIMONT, P.A.D., LE FLÈCHE-MATÉOS, A., ROCHE, S.M., BUCHRIESER, C., CADET-DANIEL, V., LE MONNIER, A. and LECUIT, M. (2010). Listeria rocourtiae sp. nov. Int. J. Syst. Evol. Microbiol. 60, 2210–2214.
LECUIT, M. (2005). Understanding how Listeria monocytogenes targets and crosses host barriers. Clin. Microbiol. Infect 11, 430–436.
LECUIT, M. (2007). Human listeriosis and animal models. Microbes Infect 9, 1216–1225.
LECUIT, M. (2009). Listeria monocytogenes crossing of the placental barrier. Arch Pediatr 16, 932–933.
LECUIT, M., and COSSART, P. (2002). Genetically-modified-animal models for human infections: the Listeria paradigm. Trends Mol Med 8, 537–542.
LECUIT, M., DRAMSI, S., GOTTARDI, C., FEDOR-CHAIKEN, M., GUMBINER, B., and COSSART, P. (1999). A single amino acid in E-Cadherin responsible for host specificity towards the human pathogen Listeria monocytogenes. Embo J 18, 3956–3963.
LECUIT, M., HURME, R., PIZARRO-CERDA, J., OHAYON, H., GEIGER, B., and COSSART, P. (2000). A role for alpha-and beta-catenins in bacterial uptake. Proc. Natl. Acad. Sci. U.S.A 97, 10008–10013.
LECUIT, M., NELSON, D.M., SMITH, S.D., KHUN, H., HUERRE, M., VACHER-LAVENU, M.-C., GORDON, J.I., and COSSART, P. (2004). Targeting and crossing of the human maternofetal barrier by Listeria monocytogenes: role of internalin interaction with trophoblast E-Cadherin. Proc. Natl. Acad. Sci. U.S.A 101, 6152–6157.
LECUIT, M., VANDORMAEL-POURNIN, S., LEFORT, J., HUERRE, M., GOUNON, P., DUPUY, C., BABINET, C., and COSSART, P. (2001). A transgenic model for listeriosis: role of internalin in crossing the intestinal barrier. Science 292, 1722–1725.
LEIFSON, E., and PALEN, M.I. (1955). Variations and spontaneous mutations in the genus Listeria in respect to flagellation and motility. J. Bacteriol. 70, 233–240.
LENGELING, A., PFEFFER, K. and BALLING, R. (2001). The battle of two genomes: genetics of bacterial host/pathogen interactions in mice. Mamm. Genome 12, 261-271.

Literaturverzeichnis
163
LEVRAUD, J.-P., DISSON, O., KISSA, K., BONNE, I., COSSART, P., HERBOMEL, P. and LECUIT, M. (2009). Real-time observation of listeria monocytogenes-phagocyte interactions in living zebrafish larvae. Infect. Immun 77, 3651–3660.
LI, B., SHIN, J., and LEE, K. (2009). Interferon-stimulated gene ISG12b1 inhibits adipogenic differentiation and mitochondrial biogenesis in 3T3-L1 cells. Endocrinology 150, 1217–1224.
LIENENKLAUS, S., CORNITESCU, M., ZIĘTARA, N., ŁYSZKIEWICZ, M., GEKARA, N., JABŁOŃSKA, J., EDENHOFER, F., RAJEWSKY, K., BRUDER, D., HAFNER, M. (2009). Novel Reporter Mouse Reveals Constitutive and Inflammatory Expression of IFN-β In Vivo. The Journal of Immunology 183, 3229 –3236.
LINDÉN, S.K., BIERNE, H., SABET, C., PNG, C.W., FLORIN, T.H., MCGUCKIN, M.A., and COSSART, P. (2008). Listeria monocytogenes internalins bind to the human intestinal mucin MUC2. Arch. Microbiol. 190, 101–104.
LINGNAU, A., DOMANN, E., HUDEL, M., BOCK, M., NICHTERLEIN, T., WEHLAND, J., and CHAKRABORTY, T. (1995). Expression of the Listeria monocytogenes EGD inlA and inlB genes, whose products mediate bacterial entry into tissue culture cell lines, by PrfA-dependent and -independent mechanisms. Infect. Immun 63, 3896–3903.
LONG, J.R., and DUKES, T.W. (1972). Listeriosis in newborn swine. Can. Vet. J. 13, 49–52.
LORBER, B. (1997). Listeriosis. Clin. Infect. Dis. 24, 1–9; quiz 10–11.
LORENZ, W.W., MCCANN, R.O., LONGIARU, M., and CORMIER, M.J. (1991). Isolation and expression of a cDNA encoding Renilla reniformis luciferase. Proc. Natl. Acad. Sci. U.S.A. 88, 4438–4442.
LOW, J.C., and DONACHIE, W. (1997). A review of Listeria monocytogenes and listeriosis. The Veterinary Journal 153, 9–29.
LOW, J.C., and RENTON, C.P. (1985). Septicaemia, encephalitis and abortions in a housed flock of sheep caused by Listeria monocytogenes type 1/2. Vet. Rec. 116, 147–150.
LUKER, G.D., BARDILL, J.P., PRIOR, J.L., PICA, C.M., PIWNICA-WORMS, D., and LEIB, D.A. (2002). Noninvasive bioluminescence imaging of herpes simplex virus type 1 infection and therapy in living mice. J. Virol. 76, 12149–12161.
MACDONALD, T.T., and CARTER, P.B. (1980). Cell-mediated immunity to intestinal infection. Infect. Immun. 28, 516–523.
MACKANESS, G.B. (1962). Cellular resistance to infection. J Exp Med 116, 381–406.
MACKANESS, G.B. (1964). The immunological basis of acquired cellular resistance. J Exp Med 120, 105–120.
MACKANESS, G.B. (1969). The influence of immunologically committed lymphoid cells on macrophage activity in vivo. J Exp Med 129, 973–992.
MAIDEN, M.C., BYGRAVES, J.A., FEIL, E., MORELLI, G., RUSSELL, J.E., URWIN, R., ZHANG, Q., ZHOU, J., ZURTH, K., CAUGANT, D.A. (1998). Multilocus sequence typing: a portable approach to the identification of clones within populations of pathogenic microorganisms. Proc. Natl. Acad. Sci. U.S.A. 95, 3140–3145.

Literaturverzeichnis
164
MAINA, F., HILTON, M.C., PONZETTO, C., DAVIES, A.M., and KLEIN, R. (1997). Met receptor signaling is required for sensory nerve development and HGF promotes axonal growth and survival of sensory neurons. Genes Dev. 11, 3341–3350.
MAINOU-FOWLER, T., MACGOWAN, A.P., and POSTLETHWAITE, R. (1988). Virulence of Listeria spp.: course of infection in resistant and susceptible mice. J. Med. Microbiol. 27, 131–140.
MANDEL, T.E., and CHEERS, C. (1980). Resistance and susceptibility of mice to bacterial infection: histopathology of listeriosis in resistant and susceptible strains. Infect. Immun. 30, 851–861.
MARCO, A.J., ALTIMIRA, J., PRATS, N., LÓPEZ, S., DOMINGUEZ, L., DOMINGO, M., and BRIONES, V. (1997). Penetration of Listeria monocytogenes in mice infected by the oral route. Microb. Pathog 23, 255–263.
MARIATHASAN, S., WEISS, D.S., NEWTON, K., MCBRIDE, J., O’ROURKE, K., ROOSE-GIRMA, M., LEE, W.P., WEINRAUCH, Y., MONACK, D.M., and DIXIT, V.M. (2006). Cryopyrin activates the inflammasome in response to toxins and ATP. Nature 440, 228–232.
MATHEWS, F.P. Encephalitis in calves. (1928). J. Am. Vet. Med. Assoc. 73:513-516.
Matthews, J.C., Hori, K., and Cormier, M.J. (1977). Purification and properties of Renilla reniformis luciferase. Biochemistry 16, 85–91.
MCCAFFREY, R.L., FAWCETT, P., O’RIORDAN, M., LEE, K.-D., HAVELL, E.A., BROWN, P.O., and PORTNOY, D.A. (2004). A specific gene expression program triggered by Gram-positive bacteria in the cytosol. Proc. Natl. Acad. Sci. U.S.A. 101, 11386–11391.
MCELROY, W.D., and DELUCA, M.A. (1983). Firefly and bacterial luminescence: basic science and applications. J. Appl. Biochem. 5, 197–209.
MCLAUCHLIN, J. (1990). Human listeriosis in Britain, 1967-85, a summary of 722 cases. 1. Listeriosis during pregnancy and in the newborn. Epidemiol. Infect. 104, 181–189.
MCLAUCHLIN, J., and LOW, J.C. (1994). Primary cutaneous listeriosis in adults: an occupational disease of veterinarians and farmers. Vet. Rec. 135, 615–617.
MEDINA, E., and NORTH, R.J. (1998). Resistance ranking of some common inbred mouse strains to Mycobacterium tuberculosis and relationship to major histocompatibility complex haplotype and Nramp1 genotype. Immunology 93, 270–274.
MEDZHITOV, R. (2001). Toll-like receptors and innate immunity. Nat. Rev. Immunol. 1, 135–145.
MEIGHEN, E.A. (1991). Molecular biology of bacterial bioluminescence. Microbiol. Rev. 55, 123–142.
MELTON-WITT, J.A., RAFELSKI, S.M., PORTNOY, D.A., and BAKARDJIEV, A.I. (2012). Oral infection with signature-tagged Listeria monocytogenes reveals organ-specific growth and dissemination routes in guinea pigs. Infect. Immun. 80, 720–732.

Literaturverzeichnis
165
MENGAUD, J., DRAMSI, S., GOUIN, E., VAZQUEZ-BOLAND, J.A., MILON, G., and COSSART, P. (1991). Pleiotropic control of Listeria monocytogenes virulence factors by a gene that is autoregulated. Mol. Microbiol. 5, 2273–2283.
MENGAUD, J., OHAYON, H., GOUNON, P., MEGE R-M, and COSSART, P. (1996). E-Cadherin is the receptor for internalin, a surface protein required for entry of L. monocytogenes into epithelial cells. Cell 84, 923–932.
MMWR. Morbidity and Mortality Weekly Report (2011). Multistate outbreak of listeriosis associated with Jensen Farms cantaloupe-United States, August-September 2011. MMWR Morb. Mortal. Wkly. Rep. 60, 1357–1358.
MONK, I., CASEY, P., HILL, C., and GAHAN, C. (2010). Directed evolution and targeted mutagenesis to murinize listeria monocytogenes internalin A for enhanced infectivity in the murine oral infection model. BMC Microbiology 10, 318.
MONK, I.R., GAHAN, C.G.M., and HILL, C. (2008). Tools for functional postgenomic analysis of listeria monocytogenes. Appl. Environ. Microbiol 74, 3921–3934.
LE MONNIER, A., AUTRET, N., JOIN-LAMBERT, O.F., JAUBERT, F., CHARBIT, A., BERCHE, P., and KAYAL, S. (2007). ActA is required for crossing of the fetoplacental barrier by Listeria monocytogenes. Infect. Immun 75, 950–957.
LE MONNIER, A., JOIN-LAMBERT, O.F., JAUBERT, F., BERCHE, P., and KAYAL, S. (2006). Invasion of the placenta during murine listeriosis. Infect. Immun 74, 663–672.
MOORE, K.W., DE WAAL MALEFYT, R., COFFMAN, R.L., and O’GARRA, A. (2001). Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 19, 683–765.
MOSTOWY, S., and COSSART, P. (2009). Cytoskeleton rearrangements during Listeria infection: Clathrin and septins as new players in the game. Cell Motility and the Cytoskeleton 66, 816–823.
MUKHERJEE, K., ALTINCICEK, B., HAIN, T., DOMANN, E., VILCINSKAS, A., and CHAKRABORTY, T. (2010). Galleria mellonella as a model system for studying Listeria pathogenesis. Appl. Environ. Microbiol. 76, 310–317.
MUNDER, A., ZELMER, A., SCHMIEDL, A., DITTMAR, K.E.J., ROHDE, M., DORSCH, M., OTTO, K., HEDRICH, H.-J., TÜMMLER, B., and WEISS, S. (2005). Murine pulmonary infection with Listeria monocytogenes: differential susceptibility of BALB/c, C57BL/6 and DBA/2 mice. Microbes Infect. 7, 600–611.
MURDOCH, C., and FINN, A. (2003). The role of chemokines in sepsis and septic shock. Contrib Microbiol 10, 38–57.
MURRAY, E., WEBB, R., and SWANN, M. (1926). A disease of rabbits characterized by large mononuclear leucocytosis, caused by a hitherto undescribed bacillus, Bacterium monocytogenes (n. sp.). J. Pathol. Bacteriol. 470–39.
MURRAY, E.G. (1955). A characterization of listeriosis in man and other animals. Can Med Assoc J 72, 99–103.

Literaturverzeichnis
166
NAKANE, A., MINAGAWA, T., and YASUDA, I. (1985). Induction of alpha/beta interferon and gamma interferon in mice infected with Listeria monocytogenes during pregnancy. Infect. Immun. 50, 877–880.
NAUCIEL, C., RONCO, E., and PLA, M. (1990). Influence of different regions of the H-2 complex on the rate of clearance of Salmonella typhimurium. Infect. Immun. 58, 573–574.
NEUENHAHN, M., KERKSIEK, K.M., NAUERTH, M., SUHRE, M.H., SCHIEMANN, M., GEBHARDT, F.E., STEMBERGER, C., PANTHEL, K., SCHRÖDER, S., and CHAKRABORTY, T. (2006). CD8α+ Dendritic Cells Are Required for Efficient Entry of Listeria monocytogenes into the Spleen. Immunity 25, 619–630.
NEUENHAHN, M., SCHIEMANN, M., and BUSCH, D.H. (2010). DCs in mouse models of intracellular bacterial infection. Methods Mol. Biol 595, 319–329.
NIGHTINGALE, K.K., SCHUKKEN, Y.H., NIGHTINGALE, C.R., FORTES, E.D., HO, A.J., HER, Z., GROHN, Y.T., MCDONOUGH, P.L., and WIEDMANN, M. (2004). Ecology and Transmission of Listeria monocytogenes Infecting Ruminants and in the Farm Environment. Appl Environ Microbiol 70, 4458–4467.
NIKITAS, G., DESCHAMPS, C., DISSON, O., NIAULT, T., COSSART, P., and LECUIT, M. (2011). Transcytosis of Listeria monocytogenes across the intestinal barrier upon specific targeting of goblet cell accessible E-Cadherin. The Journal of Experimental Medicine 208, 2263–2277.
NORDLAND, O.S. (1960). Host Parasite Relations In Initiation of Infection: III. Hyperglycemia without Stress in Experimental Infection with L. Monocytogenes. Can J Comp Med Vet Sci 24, 88–95.
O’BRIEN, T., and WOYCHIK, R. (2003). Our small relative. Nat. Genet. 33, 3–4.
O’CONNELL, R.M., SAHA, S.K., VAIDYA, S.A., BRUHN, K.W., MIRANDA, G.A., ZARNEGAR, B., PERRY, A.K., NGUYEN, B.O., LANE, T.F., and TANIGUCHI, T. (2004). Type I Interferon Production Enhances Susceptibility to Listeria monocytogenes Infection. The Journal of Experimental Medicine 200, 437–445.
O’NEIL, H.S., and MARQUIS, H. (2006). Listeria monocytogenes Flagella Are Used for Motility, Not as Adhesins, To Increase Host Cell Invasion. Infection and Immunity 74, 6675–6681.
O’RIORDAN, M., YI, C.H., GONZALES, R., LEE, K.-D., and PORTNOY, D.A. (2002). Innate recognition of bacteria by a macrophage cytosolic surveillance pathway. Proc. Natl. Acad. Sci. U.S.A. 99, 13861–13866.
OHGAKI, S., IIDA, K., YOKOO, T., WATANABE, K., KIHARA, R., SUZUKI, H., SHIMANO, H., TOYOSHIMA, H., and YAMADA, N. (2007). Identification of ISG12b as a putative interferon-inducible adipocytokine which is highly expressed in white adipose tissue. J. Atheroscler. Thromb. 14, 179–184.
OOI, Y.M., and COLTEN, H.R. (1979). Genetic defect in secretion of complement C5 in mice. Nature 282, 207–208.
OTTER, A., and BLAKEMORE, W.F. (1989). Observation on the presence of Listeria monocytogenes in axons. Acta Microbiol. Hung. 36, 125–131.

Literaturverzeichnis
167
PAMER, E.G. (2004). Immune responses to Listeria monocytogenes. Nat. Rev. Immunol. 4, 812–823.
PAN, H., YAN, B.-S., ROJAS, M., SHEBZUKHOV, Y.V., ZHOU, H., KOBZIK, L., HIGGINS, D.E., DALY, M.J., BLOOM, B.R., and KRAMNIK, I. (2005). Ipr1 gene mediates innate immunity to tuberculosis. Nature 434, 767–772.
PAQUET, A., JR, RAINES, K.M., and BROWNBACK, P.C. (1986). Immunopotentiating activities of cell walls, peptidoglycans, and teichoic acids from two strains of Listeria monocytogenes. Infect. Immun. 54, 170–176.
PARK, J.-H., PARK, Y.H., SEOK, S.H., CHO, S.A., KIM, D.J., LEE, H.Y., KIM, S.H., and PARK, J.H. (2004). Suppurative gastritis in BALB/c mice infected with Listeria monocytogenes via the intragastric route. J. Comp. Pathol. 130, 130–136.
PARK, S.F., and STEWART, G.S. (1990). High-efficiency transformation of Listeria monocytogenes by electroporation of penicillin-treated cells. Gene 94, 129–132.
PASCHE, B., KALAYDJIEV, S., FRANZ, T.J., KREMMER, E., GAILUS-DURNER, V., FUCHS, H., HRABÉ DE ANGELIS, M., LENGELING, A., and BUSCH, D.H. (2005). Sex-dependent susceptibility to Listeria monocytogenes infection is mediated by differential interleukin-10 production. Infect. Immun. 73, 5952–5960.
PATERSON, J.S. (1940). The antigenic structure of organisms of the genus Listerella. J. Pathol. Bacteriol. 427–436.
PEEL, M., DONACHIE, W., and SHAW, A. (1988). Temperature-dependent expression of flagella of Listeria monocytogenes studied by electron microscopy, SDS-PAGE and western blotting. J. Gen. Microbiol. 134, 2171–2178.
PENTECOST, M., KUMARAN, J., GHOSH, P., and AMIEVA, M.R. (2010). Listeria monocytogenes internalin B activates junctional endocytosis to accelerate intestinal invasion. PLoS Pathog 6(5):e1000900
PENTECOST, M., OTTO, G., THERIOT, J.A., and AMIEVA, M.R. (2006). Listeria monocytogenes invades the epithelial junctions at sites of cell extrusion. PLoS Pathog 2(1): e3.
PERRY, A.K., CHEN, G., ZHENG, D., TANG, H., and CHENG, G. (2005). The host type I interferon response to viral and bacterial infections. Cell Res 15, 407–422.
PESTKA, S., KRAUSE, C.D., and WALTER, M.R. (2004). Interferons, interferon-like cytokines, and their receptors. Immunol. Rev. 202, 8–32.
PETERS, M., and HEWICKER-TRAUTWEIN, M. (1994). Infection of murine fetal brain cell cultures with Listeria monocytogenes. Vet. Microbiol. 41, 19–28.
PETERS, M., and HEWICKER-TRAUTWEIN, M. (1996). Studies on the cell tropism of Listeria monocytogenes in ovine fetal brain cell cultures. Vet. Microbiol. 49, 169–179.
PFISTER, H., REMER, K.A., BRCIC, M., FATZER, R., CHRISTEN, S., LEIB, S., and JUNGI, T.W. (2002). Inducible nitric oxide synthase and nitrotyrosine in listeric encephalitis: a cross-species study in ruminants. Vet. Pathol. 39, 190–199.

Literaturverzeichnis
168
PIFFARETTI, J.C., KRESSEBUCH, H., AESCHBACHER, M., BILLE, J., BANNERMAN, E., MUSSER, J.M., SELANDER, R.K., and ROCOURT, J. (1989). Genetic characterization of clones of the bacterium Listeria monocytogenes causing epidemic disease. Proc. Natl. Acad. Sci. U.S.A. 86, 3818–3822.
PIRIE, J.H. (1927). A new disease of veld rodents. “Tiger River disease.”Pub. S. African Inst. Med. Research 163–186.
PIZARRO‐CERDÁ, J., BONAZZI, M., and COSSART, P. (2010). Clathrin‐mediated endocytosis: What works for small, also works for big. BioEssays 32, 496–504.
POHL, M.A., WIEDMANN, M., and NIGHTINGALE, K.K. (2006). Associations among Listeria monocytogenes genotypes and distinct clinical manifestations of listeriosis in cattle. Am. J. Vet. Res. 67, 616–626.
POPPENSIEK, G.C. Listerellosis-a case report. (1944). J. Am. Vet. Med. Assoc. 105:147-148
PORTNOY, D.A., SCHREIBER, R.D., CONNELLY, P., and TILNEY, L.G. (1989). Gamma interferon limits access of Listeria monocytogenes to the macrophage cytoplasm. J. Exp. Med. 170, 2141–2146.
POULSEN, K.P., FAITH, N.G., STEINBERG, H., and CZUPRYNSKI, C.J. (2011). Pregnancy reduces the genetic resistance of C57BL/6 mice to Listeria monocytogenes infection by intragastric inoculation. Microbial Pathogenesis 50, 360–366.
PRATS, N., BRIONES, V., BLANCO, M.M., ALTIMIRA, J., RAMOS, J.A., DOMÍNGUEZ, L., and MARCO, A. (1992). Choroiditis and meningitis in experimental murine infection with Listeria monocytogenes. Eur. J. Clin. Microbiol. Infect. Dis. 11, 744–747.
PRON, B., BOUMAILA, C., JAUBERT, F., BERCHE, P., MILON, G., GEISSMANN, F., and GAILLARD, J.L. (2001). Dendritic cells are early cellular targets of Listeria monocytogenes after intestinal delivery and are involved in bacterial spread in the host. Cell. Microbiol. 3, 331–340.
PUNG, O.J., TUCKER, A.N., VORE, S.J., and LUSTER, M.I. (1985). Influence of estrogen on host resistance: increased susceptibility of mice to Listeria monocytogenes correlates with depressed production of interleukin 2. Infect. Immun. 50, 91–96.
RÁCZ, P., TENNER, K., and MÉRÖ, E. (1972). Experimental Listeria enteritis. I. An electron microscopic study of the epithelial phase in experimental listeria infection. Lab. Invest 26, 694–700.
RAGON, M., WIRTH, T., HOLLANDT, F., LAVENIR, R., LECUIT, M., LE MONNIER, A., and BRISSE, S. (2008). A new perspective on Listeria monocytogenes evolution. PLoS Pathog 4, e1000146.
RAJABIAN, T., GAVICHERLA, B., HEISIG, M., MULLER-ALTROCK, S., GOEBEL, W., GRAY-OWEN, S.D., and IRETON, K. (2009). The bacterial virulence factor InlC perturbs apical cell junctions and promotes cell-to-cell spread of Listeria. Nat Cell Biol 11, 1212–1218.
RASMUSSEN, O.F., BECK, T., OLSEN, J.E., DONS, L., and ROSSEN, L. (1991). Listeria monocytogenes isolates can be classified into two major types according to the sequence of the listeriolysin gene. Infect. Immun. 59, 3945–3951.

Literaturverzeichnis
169
RAY, K., MARTEYN, B., SANSONETTI, P.J., and TANG, C.M. (2009). Life on the inside: the intracellular lifestyle of cytosolic bacteria. Nat Rev Micro 7, 333–340.
REDLINE, R., and LU, C. (1988). Specific defects in the anti-listerial immune response in discrete regions of the murine uterus and placenta account for susceptibility to infection. The Journal of Immunology 140, 3947 –3955.
REDLINE, R.W., and LU, C.Y. (1987). Role of local immunosuppression in murine fetoplacental listeriosis. J. Clin. Invest. 79, 1234–1241.
REDLINE, R.W., SHEA, C.M., PAPAIOANNOU, V.E., and LU, C.Y. (1988). Defective anti-listerial responses in deciduoma of pseudopregnant mice. Am. J. Pathol. 133, 485–497.
REUTTERER, B., STOCKINGER, S., PILZ, A., SOULAT, D., KASTNER, R., WESTERMAYER, S., RÜLICKE, T., MÜLLER, M., and DECKER, T. (2008). Type I IFN are host modulators of strain-specific Listeria monocytogenes virulence. Cell. Microbiol 10, 1116–1129.
RIBET, D., HAMON, M., GOUIN, E., NAHORI, M.-A., IMPENS, F., NEYRET-KAHN, H., GEVAERT, K., VANDEKERCKHOVE, J., DEJEAN, A., and COSSART, P. (2010). Listeria monocytogenes impairs SUMOylation for efficient infection. Nature 464, 1192–1195.
RICE, B.W., CABLE, M.D., and NELSON, M.B. (2001). In vivo imaging of light-emitting probes. J Biomed Opt, 432–440.
RIEDEL, C.U., MONK, I.R., CASEY, P.G., MORRISSEY, D., O’SULLIVAN, G.C., TANGNEY, M., HILL, C., and GAHAN, C.G.M. (2007). Improved Luciferase Tagging System for Listeria monocytogenes Allows Real-Time Monitoring In Vivo and In Vitro. Applied and Environmental Microbiology 73, 3091 –3094.
ROGERS, H.W., and UNANUE, E.R. (1993). Neutrophils are involved in acute, nonspecific resistance to Listeria monocytogenes in mice. Infect. Immun. 61, 5090–5096.
ROLDGAARD, B.B., ANDERSEN, J.B., HANSEN, T.B., CHRISTENSEN, B.B., and LICHT, T.R. (2009). Comparison of three Listeria monocytogenes strains in a guinea-pig model simulating food-borne exposure. FEMS Microbiol. Lett. 291, 88–94.
ROUSSET, M. (1986). The human colon carcinoma cell lines HT-29 and Caco-2: two in vitro models for the study of intestinal differentiation. Biochimie 68, 1035–1040.
SABET, C., LECUIT, M., CABANES, D., COSSART, P., and BIERNE, H. (2005). LPXTG protein InlJ, a newly identified internalin involved in Listeria monocytogenes virulence. Infect. Immun 73, 6912–6922.
SABET, C., TOLEDO-ARANA, A., PERSONNIC, N., LECUIT, M., DUBRAC, S., POUPEL, O., GOUIN, E., NAHORI, M.-A., COSSART, P., and BIERNE, H. (2008). The Listeria monocytogenes virulence factor InlJ is specifically expressed in vivo and behaves as an adhesin. Infect. Immun 76, 1368–1378.
SADARANGANI, C., SKAMENE, E., and KONGSHAVN, P.A. (1980). Cellular basis for genetically determined enhanced resistance of certain mouse strains to listeriosis. Infect. Immun. 28, 381–386.

Literaturverzeichnis
170
SALCEDO, C., ARREAZA, L., ALCALÁ, B., DE LA FUENTE, L., and VÁZQUEZ, J.A. (2003). Development of a multilocus sequence typing method for analysis of Listeria monocytogenes clones. J. Clin. Microbiol. 41, 757–762.
SALEM, M.L., MATSUZAKI, G., MADKOUR, G.A., and NOMOTO, K. (1999). Beta-estradiol-induced decrease in IL-12 and TNF-alpha expression suppresses macrophage functions in the course of Listeria monocytogenes infection in mice. Int. J. Immunopharmacol. 21, 481–497.
SANSONETTI, P.J. (2001). Microbes and microbial toxins: paradigms for microbial-mucosal interactions III. Shigellosis: from symptoms to molecular pathogenesis. Am. J. Physiol. Gastrointest. Liver Physiol. 280, G319–323.
SANSONETTI, P.J., and PHALIPON, A. (1999). M cells as ports of entry for enteroinvasive pathogens: mechanisms of interaction, consequences for the disease process. Semin. Immunol. 11, 193–203.
SCHLECH, W.F., 3rd (2000). Foodborne listeriosis. Clin. Infect. Dis 31, 770–775.
SCHLECH, W.F., 3RD, LAVIGNE, P.M., BORTOLUSSI, R.A., ALLEN, A.C., HALDANE, E.V., WORT, A.J., HIGHTOWER, A.W., JOHNSON, S.E., KING, S.H., NICHOLLS, E.S. (1983). Epidemic listeriosis--evidence for transmission by food. N. Engl. J. Med. 308, 203–206.
SCHLÜTER, D., DOMANN, E., BUCK, C., HAIN, T., HOF, H., CHAKRABORTY, T., and DECKERT-SCHLÜTER, M. (1998). Phosphatidylcholine-specific phospholipase C from Listeria monocytogenes is an important virulence factor in murine cerebral listeriosis. Infect. Immun. 66, 5930–5938.
SCHUBERT, W.D., URBANKE, C., ZIEHM, T., BEIER, V., MACHNER, M.P., DOMANN, E., WEHLAND, J., CHAKRABORTY, T., and HEINZ, D.W. (2002). Structure of internalin, a major invasion protein of Listeria monocytogenes, in complex with its human receptor E-Cadherin. Cell 111, 825–836.
SCHUCHAT, A., DEAVER, K.A., WENGER, J.D., PLIKAYTIS, B.D., MASCOLA, L., PINNER, R.W., REINGOLD, A.L., and BROOME, C.V. (1992). Role of foods in sporadic listeriosis. I. Case-control study of dietary risk factors. The Listeria Study Group. Jama 267, 2041–2045.
SCHUCHAT, A., ROBINSON, K., WENGER, J.D., HARRISON, L.H., FARLEY, M., REINGOLD, A.L., LEFKOWITZ, L., and PERKINS, B.A. (1997). Bacterial meningitis in the United States in 1995. Active Surveillance Team. N. Engl. J. Med. 337, 970–976.
SCHUCHAT, A., SWAMINATHAN, B., and BROOME, C.V. (1991). Epidemiology of human listeriosis. Clin. Microbiol. Rev. 4, 169–183.
SEELIGER, H.P. (1984). Modern taxonomy of the Listeria group relationship to its pathogenicity. Clin Invest Med 7, 217–221.
SEELIGER, H.P.R., and HÖHNE, K. (1979). Chapter II Serotyping of Listeria monocytogenes and Related Species. In Methods in Microbiology, (Academic Press), pp. 31–49.

Literaturverzeichnis
171
SEELIGER, H.P., and PLAB, M. (1959). [Experimental studies on the induction and therapy of subacute Listeria infection in white mice. Part I. Studies on the therapy of listerosis]. Arztl Forsch 13, 581–586.
SEVEAU, S., PIZARRO-CERDA, J., and COSSART, P. (2007). Molecular mechanisms exploited by Listeria monocytogenes during host cell invasion. Microbes Infect 9, 1167–1175.
SHEN, Y., NAUJOKAS, M., PARK, M., and IRETON, K. (2000). InIB-dependent internalization of Listeria is mediated by the Met receptor tyrosine kinase. Cell 103, 501–510.
SHIMAMURA, K., TAKAHASHI, T., and TAKEICHI, M. (1992). E-Cadherin expression in a particular subset of sensory neurons. Dev. Biol. 152, 242–254.
SKAMENE, E., and KONGSHAVN, P.A. (1979). Phenotypic expression of genetically controlled host resistance to Listeria monocytogenes. Infect. Immun. 25, 345–351.
SKAMENE, E., KONGSHAVN, P.A., and SACHS, D.H. (1979). Resistance to Listeria monocytogenes in mice: genetic control by genes that are not linked to the H-2 complex. J. Infect. Dis. 139, 228–231.
SKOVGAARD, N., and MORGEN, C.A. (1988). Detection of Listeria spp. in faeces from animals, in feeds, and in raw foods of animal origin. Int. J. Food Microbiol. 6, 229–242.
SLEATOR, R.D., GAHAN, C.G.M., and HILL, C. (2003a). A postgenomic appraisal of osmotolerance in Listeria monocytogenes. Appl. Environ. Microbiol. 69, 1–9.
SLEATOR, R.D., WOOD, J.M., and HILL, C. (2003b). Transcriptional regulation and posttranslational activity of the betaine transporter BetL in Listeria monocytogenes are controlled by environmental salinity. J. Bacteriol. 185, 7140–7144.
SOLODOVA, E., JABLONSKA, J., WEISS, S., and LIENENKLAUS, S. (2011). Production of IFN-β during Listeria monocytogenes Infection Is Restricted to Monocyte/Macrophage Lineage. PLoS ONE 6, e18543.
SOUSA, S., CABANES, D., ARCHAMBAUD, C., COLLAND, F., LEMICHEZ, E., POPOFF, M., BOISSON-DUPUIS, S., GOUIN, E., LECUIT, M., LEGRAIN, P. (2005). ARHGAP10 is necessary for alpha-catenin recruitment at adherens junctions and for Listeria invasion. Nat. Cell Biol. 7, 954–960.
SOUTHWICK, F.S., and PURICH, D.L. (1996). Intracellular pathogenesis of listeriosis. N. Engl. J. Med. 334, 770–776.
STADDON, J.M., and RUBIN, L.L. (1996). Cell adhesion, cell junctions and the blood-brain barrier. Curr. Opin. Neurobiol. 6, 622–627.
STAVRU, F., ARCHAMBAUD, C., and COSSART, P. (2011). Cell biology and immunology of Listeria monocytogenes infections: novel insights. Immunol. Rev 240, 160–184.
STOCKINGER, S., and DECKER, T. (2008). Novel functions of type I interferons revealed by infection studies with Listeria monocytogenes. Immunobiology 213, 889–897.
STOCKINGER, S., KASTNER, R., KERNBAUER, E., PILZ, A., WESTERMAYER, S., REUTTERER, B., SOULAT, D., STENGL, G., VOGL, C., FRENZ, T. (2009). Characterization

Literaturverzeichnis
172
of the Interferon-Producing Cell in Mice Infected with Listeria monocytogenes. PLoS Pathog 5, e1000355.
SWAMINATHAN, B., and GERNER-SMIDT, P. (2007). The epidemiology of human listeriosis. Microbes and Infection 9, 1236–1243.
TAM, M.A., and WICK, M.J. (2006). Differential expansion, activation and effector functions of conventional and plasmacytoid dendritic cells in mouse tissues transiently infected with Listeria monocytogenes. Cell. Microbiol. 8, 1172–1187.
TAMAYO, E., FERNÁNDEZ, A., ALMANSA, R., CARRASCO, E., HEREDIA, M., LAJO, C., GONCALVES, L., GÓMEZ-HERRERAS, J.I., DE LEJARAZU, R.O., and BERMEJO-MARTIN, J.F. (2011). Pro- and anti-inflammatory responses are regulated simultaneously from the first moments of septic shock. Eur. Cytokine Netw. 22, 82–87.
TILNEY, L.G., and PORTNOY, D.A. (1989). Actin filaments and the growth, movement, and spread of the intracellular bacterial parasite, Listeria monocytogenes. J. Cell Biol 109, 1597–1608.
TRIPP, C.S., WOLF, S.F., and UNANUE, E.R. (1993). Interleukin 12 and tumor necrosis factor alpha are costimulators of interferon gamma production by natural killer cells in severe combined immunodeficiency mice with listeriosis, and interleukin 10 is a physiologic antagonist. Proc. Natl. Acad. Sci. U.S.A. 90, 3725–3729.
TROMBERG, B.J., SHAH, N., LANNING, R., CERUSSI, A., ESPINOZA, J., PHAM, T., SVAASAND, L., and BUTLER, J. (2000). Non-invasive in vivo characterization of breast tumors using photon migration spectroscopy. Neoplasia 2, 26–40.
TUCHIN, V. (2000) Tissue Optics. Bellingham, WA: SPIE (International Society of Optical Engineering) Press.
TUMA, P.L., and HUBBARD, A.L. (2003). Transcytosis: crossing cellular barriers. Physiol. Rev. 83, 871–932.
VÁZQUEZ-BOLAND, J.A., KUHN, M., BERCHE, P., CHAKRABORTY, T., DOMÍNGUEZ-BERNAL, G., GOEBEL, W., GONZÁLEZ-ZORN, B., WEHLAND, J., and KREFT, J. (2001). Listeria pathogenesis and molecular virulence determinants. Clin. Microbiol. Rev 14, 584–640.
VEIGA, E., and COSSART, P. (2007). Listeria InlB Takes a Different Route to Met. Cell 130, 218–219.
VICENTE, M.F., BAQUERO, F., and PEREZ-DIAZ,, J. (1985). Cloning and expression of Listeria monocytogenes haemolysin in Escherichia coli. 30, 77–79.
WARREN, S.E., MAO, D.P., RODRIGUEZ, A.E., MIAO, E.A., and ADEREM, A. (2008). Multiple Nod-like receptors activate caspase 1 during Listeria monocytogenes infection. J. Immunol. 180, 7558–7564.
WAY, S.S., THOMPSON, L.J., LOPES, J.E., HAJJAR, A.M., KOLLMANN, T.R., FREITAG, N.E., and WILSON, C.B. (2004). Characterization of flagellin expression and its role in Listeria monocytogenes infection and immunity. Cell. Microbiol. 6, 235–242.

Literaturverzeichnis
173
WEINBERG, E.D. (1984). Pregnancy-associated depression of cell-mediated immunity. Rev. Infect. Dis. 6, 814–831.
DE WET, J.R., WOOD, K.V., HELINSKI, D.R., and DELUCA, M. (1985). Cloning of firefly luciferase cDNA and the expression of active luciferase in Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 82, 7870–7873.
WIEDMANN, M., ARVIK, T., BRUCE, J.L., NEUBAUER, J., DEL PIERO, F., SMITH, M.C., HURLEY, J., MOHAMMED, H.O., and BATT, C.A. (1997a). Investigation of a listeriosis epizootic in sheep in New York state. Am. J. Vet. Res. 58, 733–737.
WIEDMANN, M., BRUCE, J.L., KEATING, C., JOHNSON, A.E., MCDONOUGH, P.L., and BATT, C.A. (1997b). Ribotypes and virulence gene polymorphisms suggest three distinct Listeria monocytogenes lineages with differences in pathogenic potential. Infect. Immun. 65, 2707–2716.
WOLLERT, T., HEINZ, D.W., and SCHUBERT, W.-D. (2007a). Thermodynamically reengineering the listerial invasion complex InlA/E-Cadherin. Proc. Natl. Acad. Sci. U.S.A 104, 13960–13965.
WOLLERT, T., PASCHE, B., ROCHON, M., DEPPENMEIER, S., VAN DEN HEUVEL, J., GRUBER, A.D., HEINZ, D.W., LENGELING, A., and SCHUBERT, W.-D. (2007b). Extending the host range of Listeria monocytogenes by rational protein design. Cell 129, 891–902.
World Health Organization. (2011). The World Health Statistics 2011. World Health Organization, Geneva, Switzerland.
YOSHIKAWA, Y., OGAWA, M., HAIN, T., CHAKRABORTY, T., and SASAKAWA, C. (2009a). Listeria monocytogenes ActA is a key player in evading autophagic recognition. Autophagy 5, 1220–1221.
YOSHIKAWA, Y., OGAWA, M., HAIN, T., YOSHIDA, M., FUKUMATSU, M., KIM, M., MIMURO, H., NAKAGAWA, I., YANAGAWA, T., ISHII, T. (2009b). Listeria monocytogenes ActA-mediated escape from autophagic recognition. Nat. Cell Biol. 11, 1233–1240.
YOUNGMAN, P.J., PERKINS, J.B., and LOSICK, R. (1983). Genetic transposition and insertional mutagenesis in Bacillus subtilis with Streptococcus faecalis transposon Tn917. Proc. Natl. Acad. Sci. U.S.A. 80, 2305–2309.
ZENEWICZ, L.A., and SHEN, H. (2007). Innate and adaptive immune responses to Listeria monocytogenes: A short overview. Microbes Infect 9, 1208–1215.

Eidesstattliche Erklärung
174
7 Eidesstattliche Erklärung
Hiermit erkläre ich, dass ich die Dissertation:
„In vivo Analyse der oralen Infektion mit biolumineszenten Listeria
monocytogenes im Mausmodell“,
selbständig verfasst habe.
Ich habe keine entgeltliche Hilfe von Vermittlungs- bzw. Beratungsdiensten in
Anspruch genommen. Niemand hat von mir unmittelbar oder mittelbar entgeltliche
Leistungen für Arbeiten erhalten, die im Zusammenhang mit dem Inhalt der
vorgelegten Dissertation stehen.
Ich habe die Dissertation an folgenden Institutionen angefertigt:
Abteilung für Infektionsgenetik, Helmholtz-Zentrum für Infektionsforschung,
Braunschweig und Stiftung Tierärztliche Hochschule Hannover
Die Dissertation wurde bisher nicht für eine Prüfung oder Promotion oder für einen
ähnlichen Zweck zur Beurteilung eingereicht.
Ich versichere, dass ich die vorstehenden Angaben nach bestem Wissen vollständig
und der Wahrheit entsprechend gemacht habe.
................................................... Ort, Datum ..................................................... Unterschrift

Danksagung
175
8 Danksagung
Mein besonderer Dank gilt Dr. Andreas Lengeling für die Übernahme der Betreuung
meines Projektes. Seine ständige Unterstützung, sein überaus großes Interesse an
meiner Arbeit sowie sein Engagement und seine Diskussionsanstöße haben
entscheidend zum Fortschritt dieser Arbeit beigetragen.
Prof. Dr. Klaus Schughart danke ich für die Möglichkeit, meine Doktorarbeit in der
Abteilung Infektionsgenetik anfertigen zu können und für die ausgezeichneten
Arbeitsbedingungen, welche keine Wünsche offen gelassen haben. Für sein
Interesse an meiner Arbeit und seine Unterstützung bin ich ihm sehr dankbar.
Weiter möchte ich den Mitgliedern meiner Betreuungsgruppe, PD Dr. Eva Medina
und Prof. Dr. Peter Valentin-Weigand für Ihre Unterstützung und Anregungen
danken.
Ich danke Dr. Bastian Pasche für die Einführung in die Arbeit mit Listeria
monocytogenes sowie für seine Kooperation bei der intravenösen Infektionen.
Ich danke Dr. Heike Kollmus für den Beistand in der Organisation meines Projektes,
sowie Dr. Mahmoud Bahgat Riad für die fachliche Unterstützung im Bereich der
„Proteinnachweise“. Der ganzen Abteilung Infektionsgenetik gebührt mein Dank für
konstruktive Anregungen und das gute Arbeitsklima.
Im Besonderen möchte ich Sylvana Foth für die Hilfe bei organisatorischen Belangen
danken. Weiter den Doktoranden Leonie Dengler, Nora Mehnert, Sarah Leist und
Matthias Preuße für das gemeinsame Abendteuer PhD-Studium. Mein herzlichster
Dank geht an Christin Fricke, zum einen für ihre Versorgungskompetenz und
Organisation des Laboralltags sowie für die schöne Zeit im „social office“. Danke an
alle für die Zeit, die ich neben dem Laboralltag mit Euch verbringen durfte.
Dr. Stefan Lienenklaus möchte ich für die Einführung in das Biolumineszenz-Imaging
des IVIS 200 Systems danken sowie für die Bereitstellung der IFN-β-
Reportermäusen.

Danksagung
176
Dr. Gahan Cormac danke ich für die Bereitstellung der Listerienstämme LmoEGDe-
lux und LmoEGDe-mur-lux.
Bei Anna Rinkel möchte ich mich für die Anfertigung der histologischen Präparate
und Färbungen bedanken.
Weiter möchte ich den Tierpflegern für die Hege und Pflege unserer kleinsten
vierbeinigen Mitarbeiter danken, welche für diese Arbeit unerlässlich waren.
Meiner Familie danke ich für die große Unterstützung und die Motivationsschübe
während der Anfertigung dieser Arbeit. Ohne Euch wäre es nicht möglich gewesen
Studium und Doktorarbeit so „reibungslos“ hinter sich zu bringen.