Embed Size (px)
Citation preview

Zbl. Bakt. 274, 259-280 (1990) © Gustav Fischer Verlag, StuttgartINew York
Deutsche Gesellschaft fiir Hygiene und Mikrobiologie
Kommission zur Erarbeitung von Verfahrensrichtlinien fur die mikrobiologische Diagnostik (DGHM-Verfahrensrichtlinien, Beitrag 2.6)
Isolierung und Identifizierung von Clostridien
GISELA SCHALLEHN
I. Allgemeines
Die Gattung Clostridium umfagt nach Bergey's Manual of Systematic Bacteriology (2) 83 Spezies. Clostridien sind anaerobe, in wenigen Fallen aerotolerante, relativ groge sporenbildende grampositive Stabchenbakterien, die in alteren Kulturen haufig gramnegativ werden. Die Sporen liegen mittel- bis endstandig. Einige Clostridium-Spezies (wie z. B. Clostridium perfringens) bilden selten Sporen, auger in speziellen Nahrmedien. Die spindelformige Auftreibung des Bakterienleibes, die zur Namensgebung fiihrte, ist zwar haufig vorhanden, aber nicht Voraussetzung zur Einordnung in das Genus. Die Stamme der meisten Arten sind beweglich aufgrund peritricher Begeigelung. Unbeweglich sind C. barati, C. innocuum, C. perfringens, C. ramosum.
Clostridien kommen im Darm von Mensch und Tier, im Meer- und Seewasser, Erdboden, Staub, an der Kleidung und im Untersuchungsmaterial haufig in Mischflora vor (8). Nurwenige Spezies sind menschenpathogen (6, 18,24). Darunter sind 1. Erreger von schweren WundinfektionenlIntoxikationen: Tetanus, GasbrandlGas
adem (29). Es sind keine Seuchenerreger, da der Ausbruch der Krankheit an eine vorangegangene Verletzung gebunden ist f)der durch endogene Infektionen (c. perfringens, c. septicum) erfolgt (20). Die Voraussetzungen sind vor allem bei Erdarbeiten, Verkehrsunfallen und im Krieg oder bei intestinalen Operationen bzw. malignen Erkrankungen gegeben.
2. Erreger von Nahrungsmittelvergiftungen: Botulismus (Intoxikation) (23), Clostridium perfringens Typ A-Enteritis (24).
3. Erreger der Antibiotika-induzierten pseudomembranosen Colitis (PMC), die durch Clostridium difficile hervorgerufen wird (15).
4. Die anderen Clostridium-Spezies, die im klinischen Untersuchungsmaterial nachgewiesen werden, wurden friiher in der Regel als apathogen angesehen. Aufgrund mehrerer Isolierungen aus Entziindungsprozessen oder aus Blutkulturen bei Sepsis, nimmt man jedoch heute an, dag zumindest einzelne Arten pathogenetische Eigenschaften besitzen und besonders bei Personen mit lokaler oder allgemeiner Immunschwache von klinischer Bedeutung sind (21).

260 G. Schallehn
II. Gewinnung und Transport des Untersuchungsmaterials
Fiir die Diagnose von Tetanus und Botulismus ist Patientenserum zwecks Toxinnachweis geeignet. Weiterhin kommen bei Tetanus Wundmaterial und bei Botulismus Lebensmittel sowie Erbrochenes in Betracht (s. Tab. 1).
Bei anaeroben Wundinfektionen/Sepsis ist die kulturelle Untersuchung von nekrotischem Operationsmaterial, Wundabstrichen, Blut wichtig. Gewebematerial und Wundabstriche sollen in geeigneten Transportmedien wie z. B. Port-a-cul® (Becton Dickinson), Pus, Exsudate oder Blut in hoher Schicht, z. B. Einwegspritzen, auf deren Nadel ein Gummistopfen gestochen wird, transportiert werden (Tab. 1; s. auch DGHM -Richtlinien: 2.7 Isolierung und Identifizierung von gramnegativen sporenlosen Anaerobiern. 2.10 Isolierung und Identifizierung von grampositiven sporenlosen Anaerobiern). Der Nachweis von C. difficile-Cytotoxin und C. perfringens-Enterotoxin erfolgt aus Stuhlproben, die entsprechend der Diagnostik enteraler Infektionen (z. B. Salmonellosen) entnommen werden.
III. Gang der Untersuchung
Medien fur die Zuchtung und Isolierung
Fur die Anzucht und Isolierung von Clostridien aus Untersuchungsmaterial werden gewebehaltige Flussigmedien, wie Leberbouillon nach Tarozzi, Hirn-Kreide-Bouillon nach Rosenow oder Hackfleischbouillon verwendet. Thioglycolatbouillon verliert bei aerober Aufbewahrung schon innerhalb von 8 h durch Oxydation des Thioglycolats irreversibel an reduzierender Wirksamkeit. Deshalb eignet sie sich nicht fur dieZuchtung anspruchsvoller Anaerobier (24).
Die flussigen Niihrmedien sollen vor der Verwendung 15 min ausgekocht werden und nach dem Beimpfen entweder mit flussigem sterilem Paraffin (Merck 7162) iiberschichtet oder anaerob bebrutet werden. Ein geringer Agarzusatz (0,1-0,5%) in den Bouillons verlangsamt das Wiedereindringen des Sauerstoffs aus der Atmosphare (9).
Ais feste Niihrboden eignen sich supplementierter Blutagar wie Glucose-Hefeextrakt-Cystein-Blutagar, Columbia-Agar und Blut-Schaedler-Agar (Becton Dickinson, bioMerieux). Ais aerobes Differentialmedium dient Blutagar.
Eine Vorreduzierung der Agarplatten im Anaerobiertopf verbessert das Wachstum und damit die Isolierungsrate anaerober Bakterien.
Selektivmedien sind zwar im Handel, sie sind jedoch fur die medizinische Diagnostik wenig geeignet, da durch die Empfindlichkeit der meisten Clostridien gegen die Hemmstoffe die Keimzahl stark reduziert wird.
Kulturatmosphiire und -temperatur
Die Bebrutung erfolgt in anaerober Atmosphare, d.h. in geeigneten Anaerobiertopfen, GasPak® (BBL) oder Anaerocult® A I (Merck) oder Anaerobierbrutschrank bei 35-37°C. Fur die Selektion von C. perfringens z. B. aus Stuhlproben eignet sich eine T emperatur von 46°C.
Die Regeneration der GasPak-Katalysatoren® (nach jeder Verwendung) erfolgt bei 160°C (2 Std.). Die Anaerobiertopfe werden mit Hilfe von Redox-Indikatoren (BBL, Oxoid, Merck) auf Funktionstuchtigkeit uberpruft.
lsolierung und ldentifizierung von Clostridien 261
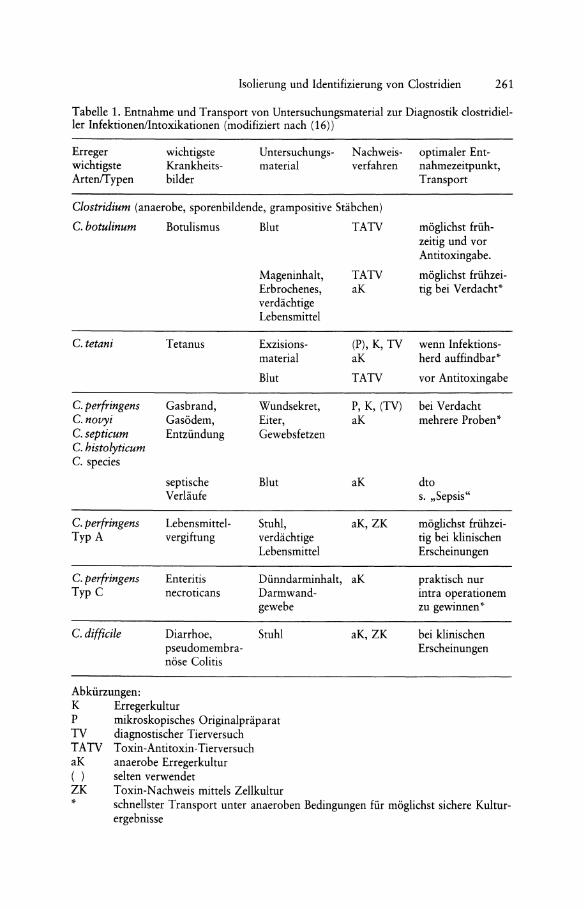
Tabelle 1. Entnahme und Transport von Untersuchungsmaterial zur Diagnostik clostridieller InfektionenlIntoxikationen (modifiziert nach (16))
Erreger wichtigste ArtenfT ypen
wichtigste Krankheitsbilder
Untersuchungsmaterial
Nachweis- optimaler Ent-verfahren nahmezeitpunkt,
Transport
Clostridium (anaerobe, sporenbildende, grampositive Stabchen)
C. botulinum Botulismus Blut TATV moglichst friih-zeitig und vor Antitoxingabe.
Mageninhalt, TATV moglichst friihzei-Erbrochenes, aK tig bei Verdacht* verdachtige Lebensmittel
C. tetani Tetanus Exzisions- (P), K, TV wenn lnfektions-material aK herd auffindbar*
Blut TATV vor Antitoxingabe
C. perfringens Gasbrand, Wundsekret, P, K, (TV) bei Verdacht C. novyi Gasodem, Eiter, aK mehrere Proben * C.septicum Entziindung Gewebsfetzen C. histolyticum C. species
septische Blut aK dto Verlaufe s. "Sepsis"
C. perfringens Lebensmittel- Stuhl, aK,ZK moglichst friihzei-TypA vergiftung verdachtige tig bei klinischen
Lebensmittel Erscheinungen
C. perfringens Enteritis Diinndarminhalt, aK praktisch nur Typ C necroticans Darmwand- intra operationem
gewebe zu gewinnen *
C. difficile Diarrhoe, Stuhl aK,ZK bei klinischen pseudomembra- Erscheinungen nose Colitis
Abkiirzungen: K Erregerkultur P mikroskopisches Originalpraparat TV diagnostischer Tierversuch TATV Toxin-Antitoxin-Tierversuch aK anaerobe Erregerkultur ( ) selten verwendet ZK Toxin-Nachweis mittels Zellkultur * schnellster Transport unter anaeroben Bedingungen fiir moglichst sichere Kultur
ergebnisse

262 G. Schallehn
Zuchtung, Isolierung und Identifizierung
Empfohlen wird neben einem Original-Grampraparat der Ansatz von Direktkulturen auf supplementiertem Blutagar anaerob und Blutagar aerob, sowie Anreicherungskulturen, die dann nach erfolgtem Wachstum zur Bakterienisolierung ebenfalls auf diese festen Medien ausgeimpft werden. Da die meisten Clostridium-Spezies relativ schnell wachsen, kann die Ablesung der Kulturplatten nach einer anaeroben Bebrutung von 1-3 Tagen bei 37"C erfolgen.
Auf Glucose-Hefeextrakt-Cystein-Blutagar oder Columbia-Agar wachsen die verschiedenen Clostridium-Spezies niihrboden- und stammabhiingig in unterschiedlich grogen, runden bis elliptischen, z. T. gefransten Kolonien, auf feuchten Agarplatten z. T. geschwiirmt, auch oft als Rasen, so dag charakteristische Kolonieformen nur bei wenigen Spezies diagnostisch genutzt werden k6nnen. Geringes aerobes Wachstum findet sich bei C. histolyticum, C. ramosum, C. tertium.
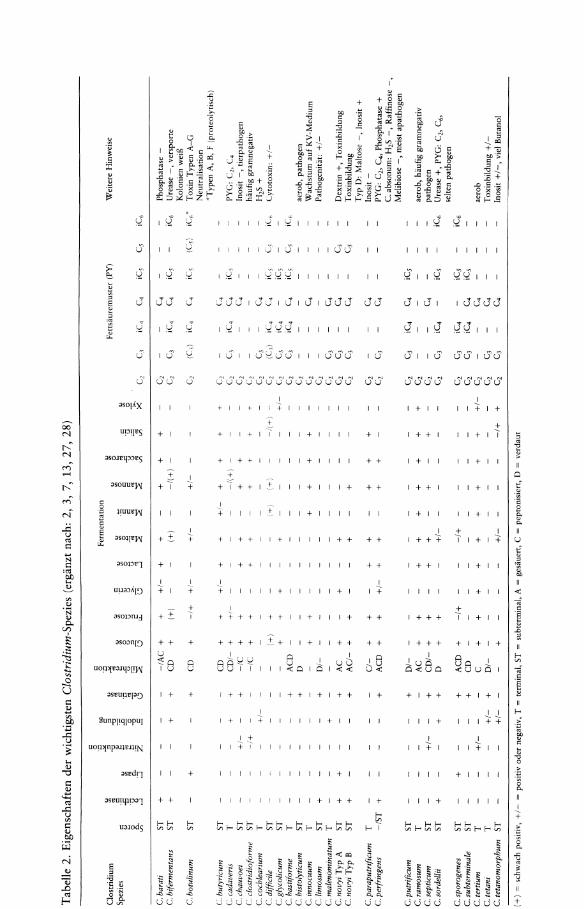
Von verdiichtigen Kolonien wird ein Grampriiparat angefertigt. Grampositive oder gramlabile Stiibchen mit Sporenbildung werden in ein gewebehaltiges Flussigmedium isoliert und die Kultur danach biochemisch gemiig Tabelle 2 und gaschromatographisch gepruft.
Nachweis von Sporenbildung
Einige Clostridium-Stiimme sporulieren sehr schlecht. Keine Sporen werden vor all em bei C. perfringens, selten bei C. ramosum und C. clostridioforme gefunden.
Werden keine Sporen im mikroskopischen Priiparat gesehen, kann zur Feststellung von Sporen in dem Material bzw. der Kultur auf den Hitzetest zuruckgegriffen werden: ca. 1-2 ml der Probe werden 10 Minuten auf 80°C im Wasserbad erhitzt (die Erhitzungszeit beginnt erst dann, wenn das Thermometer in der Kontrollflussigkeit 80°C anzeigt!). Die erhitzte Kulturprobe wird danach in die entsprechenden Kulturmedien ubertragen.
Biochemische Identifizierung
Da Clostridien verschiedene Enzyme teils saccharolytischer, teils proteolytischer und lipolytischer Art bilden, empfiehlt es sich, neb en Kohlenhydraten auch Gelatine, Milch und Eigelb als Testsubstrate zu verwenden (s. Tab. 2).
Die flussigen T estmedien in R6hrchen (2,5 ml) werden mit 0,1 ml einer gut gewachsenen (24-48 h) Kultur (Tarozzi-, Rosenow- oder Hackfleischbouillon) mittels Pasteurpipette oder Tuberkulinspritze tief beimpft. Die Agarplatten (Eigelbagar, Reinheitskontrollen) werden wie ublich fraktioniert ausgestrichen. Die Bebrutung erfolgt mindestens 3-5 Tage anaerob bei 37°C.
Der Nachweis der Kohlenhydratspaltung erfolgt durch Messung des pH-Wertes mit einem pH-Meter oder mit pH-Papier (Merck, Art.-Nr. 9555, 9556, 9557). Allgemein wird gemiig Holdeman et al. (7) ein pH-Wert von 5,5-6,0 als schwach sauer, pH unter 5,5 als stark sauer gewertet. Der pH-Wert von vorreduzierter Bouillon, die Arabinose, Ribose oder Xylose enthiilt, liegt nach 1-2tiigiger Bebrutung in einer COrAtmosphiire bei pH 5,9, so dag die Testkulturen in diesen Medien erst unter pH 5,7 als positiv gewertet werden k6nnen. Der pH der Harnstoffbouillon erreicht einen sehr alkalischen Wert (uber pH 8) bei Vorliegen einer Urease-bildenden Kultur.
Gelatinase-Bildung ist kenntlich an der Verflussigung der Gelatine (kalt stellen!).
Tab
elle
2.
Eig
ensc
haft
en d
er w
icht
igst
en C
lost
ridi
um-S
pezi
es (
erga
nzt
nach
: 2,
3,
7, 1
3, 2
7, 2
8)
Clo
stri
dium
Sp
ezie
s
C. b
arat
i C
. bif
erm
enta
ns
C.b
otul
inum
! ST
ST
ST
c. b
utyr
icum
ST
C
. cad
aver
is
T
C. c
hauv
oei
ST
C. c
lost
ridi
ofor
me
ST
C
. coc
hiea
rium
T
C
. diff
icile
S
T
C. g
lyco
licu
m
ST
C. h
asti
form
e T
C
. his
toly
ticu
m
ST
C. i
nn
ocu
um
T
!! ~ ] +
+
C.l
imos
um
ST
+ C
. mal
enom
inat
um
T
C. n
ovyi
Typ
A
ST
+
C. n
ovyi
Typ
B
ST
+
T
C. p
arap
utri
ficu
m
C. p
er/r
inge
ns
-1S
T
+
C. p
utri
ficu
m
C.r
amos
um
c.se
ptic
um
C.s
orde
llii
C. s
poro
gene
s C
. sub
term
inal
e C
.tert
ium
ST
T
S
T
ST
ST
S
T
T
C. t
etan
i T
C
. tet
anom
orph
um
ST
+
~ 0.
;.:J +
+
+
·8 1 ~ +1-
-1+
+1-
+1-
j +
~ ·s os ~ + +
+ +
+1-
+ +
+
+ +
+
+
+ + +
+ + + +
+1-
+
+1-
.g ~ o£ ~
" 8 G
-lA
C
+
CD
+
CD
+
CD
+
C
D/-
+
-/C
+
-I
C
+
AC
D
D
D/-
(+)
+ +
AC
+
AC
I-+
C/-
+
AC
D
+
D/-
AC
+
CD
/-+
D
+
AC
D
+ C
D
C
+ D
/-+
J 1
j +
+
1-+
(+
)
-1+
+
1-
+
+1-
+(-
+
+
+
+ +
+
+
+
+
+
+1-
+
+
+
-1+
+
+
+
+
+
+ +
+ + +
Fer
men
tati
on
" ~
~ .~
~
~
+
1+)
+1-
+ +
+
+
+
+
+
+
+1-
(+)
+
+
+
+
+1-
-1+
+ +
+1-
!! ~
~ ~
~
+
+
-/1+
)
+1-
+ +
-/(+
)
+
+
+
+
I+i
+
+
+ + +
+
+
+
+ +
+ +
.§ "" ~
!! ~
+ +
+
+
+
-/1+
) - +
1-
+ +
+
+ +
C2
C2
C2
C2
C,
C2
C2
C2
C2
C,
C2
C2
C,
C2
C,
C2
C2
C2
C2
C2
C2
C2
C2
C2
C2
C2
+
+1-
C2
C2
-1+
+
C
2
(+)
= sc
hwac
h po
siti
v, +
/-=
po
siti
v o
der
neg
ativ
, T
=
term
inal
, ST
=
subt
erm
inal
, A
=
gesa
uert
, C
=
pept
onis
iert
, D
=
verd
aut
Fet
tsau
rem
uste
r (P
Y)
C3
iC4
C3
iC.
(C,)
iC
.
C,
C3
IC3
)
C3
C3
C3
C3
C3
C3
C3
C3
iC4
iC4
iC4
iC4
iC.
iC.
iC.
c.
C.
C.
C.
C4
C.
C4
C.
C4
C4
C4
C4
C4
C4
C4
C4
C4
C4
C3
C3
iC.
C.
C3
C3
C.
C.
C.
iCs
iCs
iCs
ies
iCs
iCs
iCs
iCs
iCs
iCs
iCs
Cs
Wei
tere
Hin
wei
se
iC6
Pho
spha
tase
-iC
6 U
reas
e -,
vers
port
e K
olon
ien
wei
B
(C,)
iC
,*
Tox
in T
ypen
A-G
Cs
Cs
Cs
Cs
iC6
iC6
Neu
tral
isat
ion
*Typ
en A
, B
, F
(pro
teol
ytis
ch)
PY
G:
C"
C.
Inos
it -
, ti
erpa
thog
en
hauf
ig g
ram
nega
tiv
H2S
+
Cyt
otox
in:
+ /-
aero
b, p
atho
gen
Wac
hstu
m a
uf
KV
-Med
ium
P
atho
geni
t.t:
+1-
Dex
trin
+,
Tox
inbi
ldun
g T
oxin
bild
ung
Typ
D:
Mal
tose
-,
Inos
it +
In
osit
-P
YG
: C
z, C
., P
hosp
hata
se +
C
. abs
onum
: H
2S -
, R
affi
nose
-,
Mel
ibio
se -
, rn
eist
apa
thog
en
aero
b, h
aufi
g gr
amne
gati
v pa
thog
en
iC6
Ure
ase
+,
PYG
: C
b C
6,
selt
en p
atho
gen
iC6
aero
b T
oxin
bild
ung
+/
Inos
it +
1-,
viel
But
anol

264 G. Schallehn
Milch kann gesauert (A = acid), peptonisiert (C = clotted), verdaut (D = digested) werden.
Mit Eigelb (Egg Yolk)-Agar werden Lecithinase und Lipase nachgewiesen. Lecithinase (Phospholipase C) spaltet Lecithin in Phosphorylcholin und wasserunl6s1iches Diglycerid. Dadurch entsteht eine Prazipitationszone urn die Kolonie.
Die Lecithinase kann durch spezifische Antiseren gehemmt werden (Nagler-Reaktion): Dazu werden 0,05 ml Antitoxin auf einer Halfte der Eigelbagarplatte ausgespatelt. Senkrecht dazu wird die zu testende Kultur als Strich ausgeimpft. Nach anaerober Bebriitung wird die Hemmung der Lecithinase durch das Antitoxin sichtbar (3).
Lipase spaltet Fettsauren aus dem Lecithin, erkenntlich an einem Perlmuttglanz auf den Kolonien.
Der Nachweis von Phosphatase erfolgt mit einem Tropfreagenz aus a-Naphthyl-lphosphat und Diazonium-o-dianisidin. Durch die bakterielle Phosphatase wird die Phosphatgruppe des a-Naphthyl-1-phosphats abgespalten. Das freie a-Naphthyl verbindet sich dann mit dem Diazonium-o-dianisidin zu einem rotbraunen Azofarbstoff (19).
Die gaschromatographische Bestimmung der Stoffwechselprodukte (Fettsauren, Alkohole) erfolgt aus einer 3-5tagigen an aero ben Pepton-Hefeextrakt-Bouillon-Kultur (mit und ohne Glucose: PYG/PY) (7, 27). Die Kultureinsaat betragt ca. 1 ml/9 ml Bouillon. Die Bebriitung erfolgt anaerob bei 37°C.
Zum AusschluR von Kontaminationen der Tiefenkulturen werden aus jedem Kulturr6hrchen vor der Extraktion eine aerobe und anaerobe Kulturplatte angelegt und der pH-Wert der Kulturbouillon gemessen (indirekte Wachstumskontrollej Hinweis auf Kontamination bei asaccharolytischen Anaerobiern) (27).
a) Etherextraktion der fliichtigen Fettsauren und Alkohole Die Kulturbouillon wird mit 2-3 Tropfen 50%iger H2S04 auf pH 1-2 gebracht.
Danach werden ca. 3 ml Diethylether (Vorsicht! Explosiv!) zugegeben und die R6hrchen ca. 1 min lang geschuttelt (Vortex), bis eine milchige Emulsion entsteht. Nach Zentrifugieren wird die obere Etherschicht mit einer Pasteurpipette und Gummiball abgehoben und in kleine R6hrchen iiberpipettiert. 1 [11 des Etherextraktes wird in den Gaschromatographen injiziert.
b) Veresterung und Extraktion der nicht-fliichtigen Fettsauren 1 ml bewachsenes Kulturmedium wird in einem Schraubkappenrohrchen mit 2 ml
Methanol und 0,4 ml 50%iger H2S04 versetzt, gut verschlossen und 30 min bei 60°C im Wasserbad erhitzt. Nach dem Abkiihlen werden 1 ml H20 und 0,5 ml Chloroform zugesetzt und die Methylester durch Schiitteln extrahiert. Nach dem Zentrifugieren (5 min - 1500 x g) wird 1 [11 aus der Chloroformphase in die Saule des Gaschromatographen injiziert.
Die Identifizierung der Fettsauren und Alkohole erfolgt anhand einer Eichkurve. MeRbedingungen: Saule (Stahl oder Glas) 1,5-1,8 m, 10% SP 1200 + 1 % H3P04 auf Chromosorb W/AW; Injektionstemperatur 180°C, Ofentemperatur 130°C, Detektortemperatur 180°C. Isothermer Betrieb.
Kommerzielle [dentifizierungs-Kits
Die Schnellmethoden und miniaturisierten Systeme, die in den letzten Jahren fur die Identifizierung anaerober Bakterien entwickelt wurden (27), arbeiten iiberwiegend mit chromogenen Substraten zum Nachweis der Enzymaktivitat (Esterasen, Aminopeptidasen, a- und 0-Glucosidasen, alkalische Phosphatase u. a.). Das z. Zt. vielleicht beste System dieser Art ist das API ATB 32 A-Kit (bioMerieux), der innerhalb von 4 h bei

Isolierung und Identifizierung von Clostridien 265
aerober Inkubation der Teste eine Identifizierung der Bakterien mittels eines Codebuches erlaubt. Leider fuhrt auch dieses System besonders fur den unerfahrenen Mikrobiologen zu Fehlinterpretationen und falschen Identifizierungen u. a. bei C. difficile.
Pathogenitats- und Toxinteste (Tierversuche) Zu den pathogenenltoxinbildenden Clostridium-Spezies, die bei Erkrankungen des
Menschen eine Rolle spielen, gehoren C. botulinum, C. difficile, C. histolyticum, C. limosum, C. novyi Typ A, Typ B, C. perfringens Typ A, Typ C, C. sordellii (einige Stiimme), C. tetani. C. chauvoei und C. haemolyticum (c. novyi Typ D) gelten nur als tierpathogen.
Zur Information uber die von den einzelnen Spezies gebildeten Toxine und gewebeschiidigenden Enzyme sei auf die Literatur verwiesen (6, 10, 18,23,24,25).
Fur die Pathogenitatsteste (3, 7, 10, 25) - falls erforderlich - werden 20- bzw. 48-stundige Tarozzi- oder Hackfleischbouillon-Kulturen der zu prufenden ClostridiumStiimme benutzt. Empfiinglich fur Gasodemerreger sind vor allem Meerschweinchen.
Die Infektion der Meerschweinchen mit einem Korpergewicht (KG) von 250-500 g erfolgt durch intramuskuliire Injektion der Kultur in den linken Oberschenkel, wobei die Dosis immer auf g/KG berechnet wird und maximal 0,02 mllg betragen sollte. Tastbefunde von der Impfstelle bzw. die Sektionsbefunde der gestorbenen Tiere werden protokolliert zur Beurteilung der Pathogenitiit des getesteten Stammes.
Fur den Toxintest werden Miiuse benutzt (3, 7). Den Tieren von ca. 20-30 g KG werden 0,4-0,5 ml der Bouillonkultur, Kulturfiltrate bzw. Patientenserum (bei Verdacht auf Tetanus, Botulismus) i.p. oder i.v. (Schwanzvene) injiziert. Die Tiere werden uber einen Zeitraum von 72 h beobachtet.
Zur Neutralisation des Toxins mischt man die Probe vor der Injektion mit spezifischem Antitoxin in einem Reagenzglas und inkubiert die Mischung 30 min bei Zimmertemperatur oder 35 cC, oder injiziert erst Antitoxin und dann die Probe an getrennten Stellen. Antiseren fur die meisten Clostridien sind erhiiltlich bei Wellcome. Die Verwendung erfolgt wie vom Hersteller empfohlen.
Die Bestimmung der Toxin-Typen ist fur C. perfringens nicht erforderlich (7), da fast aile Stiimme aus menschlichem Untersuchungsmaterial dem Typ A zuzuordnen sind, abgesehen von der sehr seltenen Enteritis necroticans (Typ C).
C. novyi Typ A, B und D (c. haemolyticum) unterscheiden sich biochemisch (s. Tab. 2).
Die Kulturbedingungen (Bebrutungszeit, Medium, Temperatur) fur eine optimale Toxinproduktion der Stiimme weichen oft von den normalen Routinebedingungen ab (24, 25). So sollten Stiimme von C. novyi und C. tetani mindestens 48 h bei 3rC bebriitet werden, da das letale Toxin in der Zelle akkumuliert und erst in der Lyse frei wird (17). C. perfringens-Kulturen verlieren dagegen durch zu lange Kultivierung ihre Pathogenitiit, deshalb sind nur 5-20 h alte Kulturen fur Tierversuche (falls erforderlich) zu verwenden (25).
Nachweis von EnterotoxinlCytotoxinen mittels Zellkultur (11, 22) Zellkulturen: Vero (African green monkey kidney)-Zellen.
Humane embryonale Lungenfibroblasten (HEF) Wachstumsmedium: Eagles Grundlosung in Earl'scher Salzlosung (Boehringer,
Mannheim) mit Zusatz von 10% Kiilberserum, 5% fetalem Kiilberserum sowie 200 IE Penicillin G und 100 I-tg Streptomycin/ml

266 G. Schallehn
Die Monolayerkulturen werden in Mikrotiterplatten mit 96 Vertiefungen (Nunc, Wiesbaden) 48 h bei 3JOC und 5% COz-Atmosphare angezuchtet. Nach Ausbildung eines vollstandigen Zellrasens werden 50 bzw. 100 III des auf Toxin zu testenden Sterilfiltrats auf die Zellkulturen (50 bzw. 100 III Wachstumsmedium) gegeben und die Mikrotiterplatten fur weitere 24-72 h bei 37°C inkubiert. In den meisten Fallen ist eine 36-48stundige Inkubationszeit fur die Ausbildung der morphologischen Veranderungen in den Zellen ausreichend. Bei sehr hohem Cytotoxingehalt der Kulturfiltrate kann es innerhalb von 12-24 h zur kompletten Cytolyse kommen. Von diesen Filtraten werden daraufhin nur Testmengen von 10 und 20 III verwendet. Die Durchmusterung der einzelnen Cavitaten der Mikrotiterplatte erfolgt mit einem Umkehrmikroskop (Leitz, Wetzlar) bei einer VergroRerung von 10 x 4.
Anmerkung zur Identifizierung der Erreger von Wundinfektionen
Die wichtigsten Clostridium-Spezies, die im menschlichen Untersuchungsmaterial gelegentlich bis haufig gefunden werden, sind in Tabelle 2 aufgefuhrt. Weiterhin enthalt die Tabelle die saccharolytischen und proteolytischen Eigenschaften, die gebildeten Stoffwechselprodukte und Besonderheiten. Auf zusatzliche, nicht in der Tabelle angegebene Eigenschaften und besondere Merkmale soli im folgenden hingewiesen werden.
Werden im Gramprdparat plumpe grampositive Stab chen ohne Sporenbildung gefunden, kann dieser Befund ein wichtiger Hinweis fur den Arzt auf das Vorliegen einer C. perfringens-lnfektion (Gasbrand?) sein und als Schnelltest zur Abgrenzung von klinisch ahnlichen Gasodemen durch andere Erreger z. B. E. coli, Streptokokken etc. dienen.
C. perfringens bildet auf supplementierten Blutagarplatten glatte, rauhe oder gefranste Kolonien (26), die von einer Doppelzonenhamolyse umgeben sind. 1 Tropfen Phosphatase-Reagenz (19) {arbt die Kolonien in wenigen Minuten dunkelrotbraun (Schnelltest mit groRer Genauigkeit). In Leber- oder Hackfleischbouillon erfolgt ein schnelles Wachstum (3-5 h) mit Trubung und starker Gasbildung.
Sporenbildung kann in Ellner- oder Duncan-Strang-Medium (4) durch ein- bis mehrtagige Bebrutung der Kultur bei 3JOC erreicht werden (Kontroll-Grampraparat!). Die Lecithinase-Bildung wird auf Eigelbagar sichtbar und kann durch C. perfringensAntitoxin gehemmt werden (Nagler-Reaktion s. vorn). Die Lecithinasen von C. absonum, C. barati, C. bifermentans und C. sordellii werden durch C. perfringensAntitoxin nur teilweise gehemmt, nicht gehemmt werden die von C. novyi Typ A, B und D.
Tierversuch: Injiziert man 0,5 ml bis 1 ml einer gut gewachsenen Leberbouillonkultur von C. perfringens intramuskular einem Meerschweinchen, so stirbt das Tier innerhalb von 1-2 Tagen an einem Gasbrand, sofern die Clostridien virulent sind. Typische Sektionsbefunde (10, 18): Myonekrose an der Impfstelle, die Muskulatur ist zundrig zerfallen, oft aniimisch, die Bauchmuskulatur ist ebenfalls nekrotisiert und die Bauchhaut oft durch das gebildete Gas abgehoben. 1m Unterhautzellgewebe findet sich meist viel blutig-serose Flussigkeit, die fettig und durch die verflussigte Muskulatur trub bis schlcimig sein kann. Aus dem zerfallenen Gewebe, der Odemflussigkeit und den Organen (Leber, Herz) konnen C. perfringens-Bakterien mikroskopisch und kulturell nachgewiesen werden.
Die Anzucht von C. novyi ist auRert schwierig und nur mit vorreduzierten Medien unter strikt anaeroben Bedingungen erfolgreich. Auf vorreduzierten supplementierten

IsoIierung und Identifizierung von Clostridien 267
Blutagarplatten bilden Typ A runde bis elliptische Kolonien mit unregelmiHsigem Rand, Typ B flache durchscheinende Kolonien mit lockigen Auslaufern. Die Hamolysezone unter der Kolonie ist bei Typ B starker ausgepragt als bei Typ A. In Leber- oder Hackfleischbouillon erfolgt maBiges Wachstum mit Gasbildung haufig nur im unteren, starker anaeroben Teil des Rohrchens.
Die Stab chen von C. novyi Typ A wirken im Grampraparat schlanker (111m) und langer (2-10 11m) als C. perfringens. Auffallend groBe Stabchen bildet Typ B (bis 22 11m lang). Die ovalen subterminalen Sporen treiben den Bazillenleib nur geringfiigig auf.
Tierversuch: Das mit C. novyi Typ A oder B infizierte Meerschweinchen stirbt innerhalb von 1-2 Tagen an einer Toxinamie. Charakteristischer Sektionsbefund ist das meist farblose, jedoch manchmal blutige, gallertige bdem im Unterhautzellgewebe, das sich von der Impfstelle (Beinmuskulatur) iiber das Abdomen ausbreitet. Die Muskeln an der Impfstelle sind blaBrot und feucht, aber im Gegensatz zu C. perfringens nicht nekrotisch zerfallen.
C. septicum ist maBig sauerstoffempfindlich und wachst optimal bei 37°C - 40°C auf supplementierten Blutagarplatten in zarten durchscheinenden Kolonien mit sehr feinen Auslaufern oder Rasen und Hamolyse. Die Leber- und Hackfleischbouillonkulturen sind getriibt mit Gasbildung. Die Stabchen sind peritrich begeiBelt und stark beweglich. Haufig wachsen sie in Oberflachen- und Fliissigkulturen in langen Faden oder Ketten. Durch die subterminalen ovalen Sporen werden die Bazillenleiber spindelformig aufgetrieben.
Tierversuch: Nach i.m. Injektion von 0,2-0,5 ml einer 20-stiindigen C. septicumLeberbouillonkultur stirbt das Meerschweinchen meist innerhalb von 24 h an einer Toxin- und Bazillamie. Yom Ort der Infektion bis iiber das Abdomen breitet sich im Unterhautzellgewebe ein blutig-seroses bdem aus, weniger sulzig als bei C. novyi, Gasbildung ist haufig vorhanden. 1m Abklatschpraparat von der Leberoberflache sind C. septicum-Bazillen in Kettenbildung nachweisbar (c. chauvoei bildet keine Ketten). Zu beachten ist, daB C. septicum invasiv ist und post mortem yom Darm in das Gewebe einwandern und somit falschlich als Krankheitserreger ermittelt werden kann. Eine differentialdiagnostische Abgrenzung zu C. chauvoei ist am besten fluoreszenzmikroskopisch moglich (25).
Die Isolierung von C. histolyticum bietet wenig Schwierigkeiten, da es anaerob bis aerotolerant ist und oft sogar auf Blutagarplatten aerob in sehr kleinen Kolonien wachst. Auf anaerob bei 37°C bebriiteten supplementierten Blutagarplatten sind die Kolonien klein, rund, mit Hamolyse. In Leberbouillon erfolgt ein iippiges Wachstum mit teilweisem proteolytischem Abbau der Leberstiickchen und Schwarzung durch H2S-Bildung. C. histolyticum ist saccharolytisch wenig aktiv, bildet dafiir aber eine Reihe von Proteinasen.
Tierversuch: Bei Verdacht auf C. histolyticum sollten dem Meerschweinchen nur geringe Mengen (0,1-0,5 ml) Kultur in die Oberschenkelmuskulatur injiziert werden, da die Erkrankung sehr stiirmisch verlauft. Die Muskulatur an der Impfstelle und die dariiber befindliche Haut werden peptisch verdaut, so daB nur ein strukturloser, hamorrhagischer Gewebebrei iibrigbleibt. Haufig wird das Bein im Oberschenkel amputiert. Der Tod des Tieres tritt meist nach 2-3 Tagen ein.
Auch einige Stamme von C.limosum bilden Toxine, die zum Tode des Meerschweinchens innerhalb von 48 h fiihren. Die pathologisch-anatomischen Veranderungen

268 G. Schallehn
iihneln denen eines C. perfringens-Gasbrandes, vermutlich aufgrund der gebildeten Lecithinase: Die Muskulatur der Impfstelle (Oberschenkel) und die Bauchmuskulatur sind faserig zundrig zerfallen, oft aniimisch. Geringe Mengen von blutig-seroser Fliissigkeit finden sich im Unterhautzellgewebe.
Nur sehr wenige Stiimme von C. sordellii sind so pathogen, da~ sie zum Tode des Versuchstieres fuhren. Sie erzeugen nach i.m. Injektion eine Schwellung des Beines (Impfstelle) des Meerschweinchens und ein ausgedehntes sulziges, leicht rotliches Odem im Unterhautzellgewebe ahnlich dem C. novyi-Odem. Die Muskulatur ist hamorrhagisch, jedoch heller als bei C. septicum-Infektionen (10,22).
Auf supplementierten Blutagarplatten bildet C. tetani zarte, £lache, farblose Kolonien mit gefranstem Rand und Ausliiufern. Einige Stamme breiten sich in feinem Rasen iiber die ganze Nahrbodenoberflache aus; andere wachsen in sehr kleinen runden Kolonien, selten mit Auslaufern. Hamolyse tritt meist nur unter Einzelkolonien auf. In Leber- oder Hackfleischbouillon erfolgt langsames Wachstum mit ma~iger Gasbildung.
Die Stabchen des C. tetani sind schlank und in jungen Kulturen grampositiv. Die Sporen entwickeln sich innerhalb von 48-72 h. Sie sind rund, auftreibend, liegen terminal und geben dem Stabchen ein trommelschlagerartiges Aussehen. Die mikroskopische Diagnose aus gefarbten Praparaten ist nicht zuverlassig, da auch andere Clostridien (wie C. cochlearium) endstiindige Sporen bilden und einige Stamme von C. tetani die typischen "T rommelschliiger" nicht ausbilden.
Tierversuch: Gewebestiickchen als Untersuchungsmaterial wird einer Maus in der Niihe der Schwanzwurzel in eine Hauttasche implantiert. Patientenserum oder Kulturfliissigkeit wird in Mengen zwischen 0,1-0,5 ml den Mausen i.p. injiziert. Nach 1-3 Tagen gehen die Tiere unter den typischen Erscheinungen des Tetanus zugrunde. Charakteristisch ist die "Robbenstellung" mit krampfartig nach hinten gestreckten Beinen. Zur Kontrolle muB einer weiteren Maus vor dem Test Tetanus-Antitoxin injiziert werden. Das Tier soll uberleben.
Untersuchungen bei Verdacht auf Lebensmittelvergiftungen
1. Botulismus
C. botulinum bildet in anaeroben proteinhaltigen Medien, z. B. in Nahrungsmitteln, ein Toxin, das die Ursache fur eine Nahrungsmittelvergiftung, den Botulismus, ist (23). Nicht selten kann das Toxin (Typ B) bis zu 4 Wochen nach der Toxinaufnahme im Blut des Patienten nachgewiesen werden. Die Letalitat liegt zwischen 25 und 70%. Der Tod tritt meist durch Atemlahmung ein.
Aufgrund der serologischen Spezifitat des gebildeten Neurotoxins wird C. botulinum in 7 toxinogene Typen eingeteilt. Hauptsachlich 3 dieser Typen (A, B und E) kommen als Giftbildner in Lebensmitteln vor und sind fur den Botulismus des Menschen bedeutend. Die Typen Ch C2 und D rufen bei Tieren (Rindern, Gefliigel) Botulismuserkrankungen hervor. Fiir Menschen scheinen die Toxine dieser Typen bei oraler Aufnahme unschiidlich zu sein. Typ G (c. argentinense) wurde aus dem Boden in Argentinien und aus menschlichem Untersuchungsmaterial in der Schweiz isoliert.
Die einzelnen T ypen von C. botulinum bilden auf supplementierten Blutagarplatten verschiedene Kolonieformen (glatt, rauh, opak oder durchscheinend) mit irreguliirem Rand. Hamolyse wird bei einigen Stammen ausgebildet. Die Leber- oder Hackfleisch-

Isolierung und Identifizierung von Clostridien 269
bouillonkultur ist getriibt und zeigt Gasbildung. Die Typen A und B wachsen bei niedrigem pH von 5, Typ E nicht (23).
Die Stiibchen des C. botulinum sind gerade bis leicht gebogen. Die Sporen sind oval, subterminal und geben dem Stiibchen ein "tennisschliigerartiges" Aussehen.
Zur Laboratoriumsdiagnose ist der Nachweis von Toxin in Patientenserum, Erbrochenem, Nahrungsmitteln, Leichenmaterial (mittels Miiusetest) und/oder Isolierung des Erregers aus den Nahrungsmitteln wichtig (3), die Speziallabors vorbehalten ist. Zur erfolgreichen Isolierung des Erregers miissen die verdiichtig erscheinenden Bezirke im Nahrungsmittel (erkenntlich an Zersetzung, Verfiirbung oder bei Fleisch in der Niihe des Knochens) ausgewiihlt werden, da die Keime nicht gleichmiigig in festen Lebensmitteln verteilt sind. Nach Zerkleinerung oder Zentrifugation werden die Proben in Hackfleischbouillon und auf supplementierten Blutagarplatten verimpft und anaerob mehrere Tage bei 37°C bebriitet. Die gewachsenen Kolonien werden mikroskopisch und biochemisch identifiziert, sowie auf Toxinbildung gepriift. Zur Feststellung des Types wird die Neutralisation mit typspezifischen Antiseren (A-G) entsprechend dem beschriebenen Tierversuchsschema durchgefiihrt.
Tierversuch: 0,5 ml eines moglichst friihzeitig entnommenen Patientenserums oder eines Extraktes der Untersuchungsprobe werden einer weigen Maus i.p. injiziert, die 2. Maus erhiilt 0,5 ml Testmaterial und 0,5 ml polyvalentes C. botulinum-Antitoxin. Es empfiehlt sich, wegen des unbekannten T oxingehaltes unterschiedliche Mengen der Testprobe zu untersuchen, urn die charakteristischen Symptome (" Wespentaille") besser beobachten zu konnen, was bei einem zu schnellen T od nicht moglich ist. War im Testmaterial Botulinumtoxin vorhanden, wird das ungeschiitzte Tier unter den typischen Symptomen sterben, die mit Antitoxin behandelte Maus mug am Leben bleiben. Die Krankheitssymptome zeigen sich in allgemeiner Muskelschlaffheit, wespentaillenartiger Einziehung der Bauchwandungen, motorischen Liihmungen der Beine und Atemliihmung, die zum T ode fiihrt.
2. Clostridium perfringens Typ A-Enteritis
Bestimmte Stiimme von C. perfringens Typ A kommen als Erreger von Nahrungsmittelvergiftungen beim Menschen vor (24). Durch Aufnahme von C. perfringens infizierten Nahrungsmitteln (ca. 106_107 vegetative C. perfringens-Zellen/g Nahrungsmittel) tritt in 8-10 h nach der Mahlzeit eine Enteritis auf mit Obelkeit, profusem Durchfall von 1-2 Tagen und krampfartigen Bauchschmerzen. Erbrechen und Fieber fehlen meistens. Eine Therapie ist im allgemeinen nicht erforderlich. Ausgelost wird die Intoxikation durch ein hitzelabiles Enterotoxin, das von C. perfringens erst wiihrend der Versporung im Darm gebildet und freigesetzt wird. Deshalb ist eine Priifung auf vorhandenes Toxin in den verdiichtigen Nahrungsmitteln ohne Erfolg.
Der kulturelle Nachweis von C. perfringens im Nahrungsmittel und Stuhl erfolgt mit den iiblichen Methoden.
Beurteilung des positiven Befundes: Da bei etwa 70% der Menschen C. perfringens im Darm nachweisbar ist (ca. 103 bis 104 Kolonie-bildende Einheiten/g Faeces) (24), ist die Isolierung von C. perfringens aus dem Stuhl erkrankter Personen kein Hinweis auf eine Nahrungsmittelvergiftung durch diesen Erreger, solange nicht quantitative Untersuchungen vorgenommen werden (Anstieg auf 107_108 KE/g Faeces).
Erst die Isolierung von serologisch gleichen C. perfringens-Stiimmen aus dem Stuhl des Erkrankten und aus dem betreffenden Nahrungsmittel- was in Speziallabors durch Objektglas-Agglutination mit spezifischen Antiseren festgestellt werden kann - und/

270 G. Schallehn
oder der Nachweis, daB diese Stamme Enterotoxin bilden, kann die bakteriologische Diagnose unterstiitzen.
Enterotoxin-Nachweis mittels Vero (African green monkey kidney)-Zellkultur: 20 h alte C. perfringens-Kulturen in Hackfleischbouillon werden in Thioglycolatme
dium (ohne Indikator 135 C, BBL) iiberimpft (1 : 10), 20 min bei 75°C zur Abtotung der vegetativen Zellen und zur Stimulierung der Enterotoxinbildung erhitzt und dann iiber Nacht (ca. 20 h) bei 37°C anaerob im GasPak®-Topf bebriitet. Diese Kulturen werden danach in Duncan-Strang-Bouillon (1: 10) iiberimpft, in der sie wahrend der 20-stiindigen Bebriitung bei 37"C versporen und Enterotoxin bilden sollen (4). Die erhaltenen Kulturen werden zentrifugiert (30 min 4200 X g), die Dberstande sterilfiltiert (Millipore® 0,45 f,lm) und bis zur Verwendung bei 4°C aufbewahrt.
Ais Zellkulturen werden 2 Tage alte Vero-Zellen (50 f,ll) in Mikrotiterplatten verwendet, die nach Beimpfung mit 5 f,ll (Endverd. 1: 10) bis 50 f,ll (Endverd. 1 : 2) des toxinhaltigen Kulturfiltrats 24-48 h bei 37°C im COrBrutschrank inkubiert und dann auf cytopathische Veranderungen durchgesehen werden. 1m positiven Fall werden die langlich-ovalen Vero-Zellen durch das Enterotoxin aus dem Zellverband gelost und erscheinen abgerundet und ungeordnet (11).
Als Kontrollen dienen unbeimpfte Zellkulturen, das Duncan-Strong-Medium, ein sicher C. perfringens-Enterotoxin-haltiges Filtrat und Anti-Enterotoxin-Serum (Wellcome).
Fiir die Beurteilung des cytopathischen Effekts und zur Unterscheidung starker, schwacher bzw. negativer Enterotoxinbildung erwies sich die Konzentration von 10 f,ll / 50 f,ll Zellen als besonders geeignet (unveroffentlicht). Der cytopathische Effekt tritt nicht auf, wenn vor der Beschickung der Vero-Zellen 10 f,ll Anti-Enterotoxin-Serum in den Testansatz gegeben wird.
Dieser Enterotoxinnachweis mittels Zellkulturen ist relativ einfach durchfiihrbar und gut reproduzierbar. Es muB bei der Beurteilung der Ergebnisse jedoch beachtet werden, daB etwa ein Drittel der C. perfringens-Stamme allgemein Enterotoxin-Bildungsvermogen besitzen (unveroffentlicht).
Untersuchungen auf Clostridium difficile
Clostridium difficile wird gelegentlich in eitrigen Gewebsinfektionen gefunden, vor allem aber als Erreger von Diarrhoe, Colitis und pseudomembranoser Colitis nach Antibiotikatherapie des Patienten (s. auch DGHM-Richtlinien 1.3: Die bakterielle Diagnose enteraler Infektionen). Ausgelost werden diese Erkrankungen durch die von C. difficile gebildeten 2 Toxine, dem Enterotoxin (Toxin A) und dem Cytotoxin (Toxin B), wobei das Enterotoxin (Toxin A) auch eine, jedoch etwa tausendfach schwachere Cytotoxizitat besitzt. Es wurden bisher keine C. difficile-Stamme gefunden, die nur eines der beiden Toxine bildeten, so daB man annimmt, daB fiir die Toxin-Bildung der gleiche Genbereich verantwortlich ist (15).
Die mikrobiologische Diagnostik einer C. difficile-Erkrankung kann aus dem Stuhl auf zweierlei Art erfolgen: 1. durch den kulturellen Nachweis des Erregers; 2. durch den Nachweis der Toxine. Der serologische Nachweis von Antikorpern im Patientenserum eignet sich nicht fiir die Diagnose einer pseudomembranosen Colitis, da die Bildung von Antikorpern von Patient zu Patient sehr unterschiedlich erfolgt und der Antikorpertiter keine Relation zur Schwere der Erkrankung bietet (15).

Isolierung und Identifizierung von Clostridien 271
1. Kultureller Nachweis
a) Die Isolierung von C. difficile aus Stuhlproben erfolgt am besten iiber Selektivmedien, die Cefoxitin (16 !tg/ml) und Cycloserin (500 !tg/ml) enthalten, wie das CCFMedium nach George et al. (5), das in etwas abgewandelter Form von Oxoid und anderen Firmen kauflich erworben werden kann.
Auf CCF-Agar wiichst C. difficile nach 40-48 h anaerober Bebriitung bei 37°C in 2-5 mm grolSen, gelblich cremigen Kolonien, leicht erhaben, mit fransigem, gelegentlich auch glattem Rand und gelbgriiner Fluoreszenz unter UV-Licht.
Beurteilung: Die Selektivitiit dieses Niihrbodens fiir C. difficile ist nicht besonders grolS: Neben zahlreichen grampositiven und -negativen Stiibchenbakterien, Kokken und SprolSpilzen wachsen auger C. difficile andere Clostridium-Spezies wie C. cochlearium, C. innocuum, C. perfringens, C. ramosum und C. tertium auf CCFA (unveroffentlicht). Auch die von George et al. (5) als Identifizierungskriterien angegebene CCFA-Verfiirbung nach gelb und die griinliche Fluoreszenz der C. difficile-Kolonien unter UV -Licht erweisen sich als relativ unspezifisch. Der CCF A kann demnach nur zur Anreicherung und Vorselektierung von C. difficile dienen, fiir die Identifizierung miissen jedoch weitere biochemische Teste und der Nachweis der Toxinbildung angeschlossen werden.
b) Identifizierung: C. difficile wiichst nach 40-48 h Bebriitung bei 37°C auf supplementierten Blutagarplatten unter anaeroben Bedingungen in 2-5 mm grolSen, grau durchscheinenden, nicht hiimolysierenden Kolonien, die hiiufig ein erhabenes Zentrum und fransigen Randsaum aufweisen. Seine fermentativen Eigenschaften sind nicht sehr groK Es baut die in Tabelle 2 angegebenen Kohlendydrate sowie Melezitose und Rhamnose abo Der End-pH erreicht dabei meist nur 5,8, so daIS diese schwache Reaktion leicht iibersehen werden kann. Da die konventionelle Identifizierung der Clostridien 5-10 Tage dauert, sollen hier 2 Schnellmethoden zur Identifizierung von C. difficile angefiihrt werden, die recht brauchbar erscheinen:
Gaschromatographischer Nachweis von Capronsiiure und p-Kresol, die von C. difficile aus Norleucin und Tyrosin (NT-Bouillon) gebildet werden (14): Von der Vorkultur in Tarozzi-, Rosenow- oder Hackfleischbouillon wird 1 ml in 9 ml NT-Bouillon iiberfiihrt. Nach 24-48stiindiger Bebriitung bei 37°C im GasPak®-Topf werden die fliichtigen Fettsauren im Kulturiiberstand nach Ansauerung auf pH 2 mit 2,5 ml Ether ausgeschiittelt. 1 !Jl des Etherextraktes wird anschlielSend gaschromatographisch auf fliichtige Fettsiiuren und p-Kresol analysiert.
Beurteilung: AulSer C. difficile bildete keine der 37 gepriiften Clostridium-Spezies pKresol (unveroffentlicht). Somit kann dieser Test als sehr spezifisch eingestuft werden und erlaubt auch ohne weitere biochemische Differenzierung die Identifizierung von C. difficile und den AusschlulS anderer Clostridien. Die Dauer des Tests betriigt ca. 2-3 Tage.
Latex-Agglutinationstest (Freka® MicroScreen® C. difficile, Fresenius) Mit diesem Agglutinationstest konnen C. difficile-Bakterien nach ihrer Anziichtung
aus dem Patientenstuhl sowohl von der Selektivagarplatte als auch aus der Anreicherungskultur in Cefoxitin-Cycloserin-Bouillon (ca. 20 h bei 37°C) identifiziert werden.
Testprinzip: Latex-Partikel sind mit spezifischen IgG-Antikorpern gegen C. difficileZellwandantigene gekoppelt. 1 Tropfen dieses Reagenz wird auf dem Objekttriiger entweder mit 1 Tropfen der Selektivbouillon, die den Organismus enthiilt, oder mit einer Suspension der Kuhur von einem festen Niihrmedium gemischt. Innerhalb von 2 min agglutinieren die Latex-Partikel zu grolSen, sichtbaren Klumpen. Fehlt diese Agglu-
19 Zbl. Bakt. 274/2

272 G. Schallehn
tination, so ist C. difficile als Erreger auszuschlieRen. Zur Kontrolle wird das C. difficile-Reagenz mit 1 Tropfen physiologischer NaCI-Losung vermischt und auf Agglutination uberpruft. Eine Autoagglutination der Testkultur soli ebenfalls mit physiologischer NaCI-Losung ausgeschlossen werden.
Beurteilung: Wird keine Selektivbouillon verwendet, muB damit gerechnet werden, dag auBer c. difficile auch C. bifermentans, C. glycolium (1), c. ramosum, C. sordellii und C. clostridioforme agglutinieren (unveroffentlichte Befunde).
Von der Firma Fresenius wird auf die starke Kreuzreaktion von C. bifermentans und C. sordellii mit C. difficile-Antigen hingewiesen. Die Fa. Fresenius empfiehlt deshalb eine Subkultur auf CCF-Agar und eine erneute Testung der Kolonien mit dem Reagenz. Da auBer C. difficile nur C. ramosum gegen Cefoxitin resistent ist, wachsen die anderen 4 Clostridium-Spezies nicht auf CCF-Agar. Die Spezifitiit des Testes wird erhOht, wenn gleich Selektivmedium fur die Anzucht von C. difficile verwendet wird.
Der Latex-Agglutinationstest ergibt im negativen Fall ein eindeutiges Ergebnis, im positiven Fall ist ein zusiitzlicher Cytotoxin-Nachweis anzuraten, urn nicht-toxinogene C. difficile-Stiimme und andere kreuz-agglutinierende Clostridien auszuschlieBen. Da die Reaktionen sehr gut ablesbar sind, scheint der Test als Alternative zum ToxinNachweis aus dem Stuhl mittels Zellkultur durchaus geeignet.
2. Toxin-Nachweis
Der Toxin-Nachweis aus Stuhlproben kann mit verschiedenen Methoden erfolgen: a) uber die Zellkultur aus humanen embryonalen Lungenfibroblasten; b) mit einem kiiuflichen Latex-Agglutinationstest der Fa. Marion Scientific, U.S.A.
(Becton Dickinson). In beiden Fallen geht man von Stuhlaufschwemmungen aus (1 erbsgroRes Stuck/2,5 ml Peptonbouillon).
a) Fur die Testung auf Zellkulturen wird der Dberstand der Stuhlaufschwemmung sterilfiltriert (Millipore® 0,45 flm) und in Mengen von 100 f.tl auf die gleiche Menge humaner embryonaler Lungenfibroblasten-(HEF)-Kultur in Mikrotiterplatten getestet. Nach 24-72stundiger Bebrutung bei 37°C im COrBrutschrank werden die Zellkulturen auf cytopathische Veriinderungen durchgesehen. 1m positiven Fall kommt es zur Abrundung der Lungenfibroblasten (22). Als Kontrollen dienen ein C. difficile-Toxinfiltrat, sowie eine Neutralisation mit C. difficile- oder C. sordellii-Antitoxin (Wellcome) in nicht eindeutigen Fiillen.
b) Latex-Agglutinationstest (Culturette Brand CDT™, Marion Scientific). Prinzip: Latex-Partikel sind mit C. difficile-spezifischen Antikorpern beladen, die mit C. difficile-Toxin/Antigenen im Dberstand der Stuhlaufschwemmung agglutinieren. Entgegen der Vorschrift der Firma kann mit 1110 der Testvolumina gearbeitet werden. 5 fll Latex-Reagenz werden mit 10 fll Stuhlfiltrat vermischt und zusammen mit den gleichzeitig angelegten "positiv"- und "negativ"-Kontrollen durch Schwenken des Objekttriigers (3 min) agglutiniert. Die Beurteilung der Agglutination erfolgt gegen eine Lichtquelle.
Beurteilung: Der Toxin-Nachweis mit dem Latex-Agglutinationstest ist zwar bequemer und schneller durchfuhrbar als mit der Zellkultur, er ist aber in etwa 16% "falsch" negativ bzw. in 6% "falsch" positiv im Vergleich mit den Ergebnissen der Zellkultur (unveroffentlicht), was auch durch neuere Daten in der Literatur zum Ausdruck kommt (12, 15).
Bei der Beurteilung der positiven Befunde ist folgendes zu bedenken (15): Das Vorkommen von C. difficile im Stuhl ist abhiingig yom Alter des PatienteniMenschen: Bei

Isolierung und Identifizierung von Clostridien 273
NeugeborenenlSiiuglingen ab der 2. Woche bis zum Kleinkindesalter von 1-2 Jahren findet sich C. difficile in 7-48% der Stuhlproben, in neonatalen Intensivstationen sogar in 21-78% der Stuhlproben. Trotz des Nachweises von Toxin im Stuhl in unterschiedlicher Titerhohe sind die KleinkinderlSiiuglinge oft symptomlos. Eine pseudomembranose Colitis ist bei Kindern unter 2 Jahren noch nie beobachtet worden. Ab 2 Jahre bis ins Erwachsenenalter findet sich nur bei 4% der Personen C. difficile im Stuhl. Das heiGt, daG der'C. difficile-Nachweis bei Siiuglingen und Kleinkindern nicht iiberbewertet wer:den darf.
Aufbewahrung und Versand der Clostridium-Kulturen .
Fiir die Aufbewahrung der Clostridien eignen sich versporte Kulturen in Leber-, Hirnkreide- oder Hackfleischbouillon bei 4 °C oder bei -20°C. Die Lyophilisierung kann leicht zum Verlust der Toxizitiit (Bakteriophagen) oder Pathogenitiit fiihren. Schlecht versporende Clostridien wie Z. B. C. perfringens, C. ramosum etc. konnen in Ellner- oder Duncan-Strong-Medium zur Sporulation gebracht und dann eingefroren werden.
Fiir den Versand eignen sich sterile Wattetupfer, die durch Eintauchen in die Bouillonkultur mit der Bakteriensuspension getriinkt und in ein Transportmedium (Port-acul®-Rohrchen) iiberfiihrt werden.
Anhang
Niihrmedien und Reagenzien
Leberbouillon nach T arozzi
Kalbs- oder Rinderleber Aqua dest. im Dampftopf 2 h kochen.
500 g 1000 ml
Leber in Stiicke schneiden und in Rohrchen fiillen, hierauf 6 ml Bouillon nach folgendem Rezept fiillen und 15 min bei 1,1 atii autoklavieren.
Leberwasser 1000,0 ml NaCI 3,0 g Na2HP04 2,0 g Pepton (Difco) 10,0 g 1m Dampftopf ca. 2 h kochen, filtrieren pH 7,8.
Hirnkreide-Bouillon nach Rosenow (27)
Pepton (tryptisch) 10,0 g Rindfleischextrakt 3,0 g Glucose 2,0 g NaCI 2,0 g Cystein-Hydrochlorid 0,3 g Indikator nach Andrade 10,0 ml Aqua dest. 1000,0 ml Marmorpulver (CaC03) 25,0 g

274 G. Schallehn
Indikator nach Andrade: saures Fuchsin Aqua dest.
0,5 g 100,0 ml
Aqua dest., Pepton, NaCl, Rindfleischextrakt und Cystein mischen und einige Minuten kochen lassen, pH auf 7,2 einstellen. Danach 10 min kochen lassen und filtrieren. Dann den Indikator nach Andrade zugeben, danach die Glucose. Bei richtig eingestelltern pH nimmt die Fliissigkeit eine rosa Fiirbung an. In Rohrchen abfiillen, in die zuvor gegeben wurde: a) ein Stiickchen CaC03,
b) ein etwa haselnuggroges Stiick Rinder- oder Schafshirn (erhiirtet durch Gefrieren, ohne Hirnhiiute und oberfliichliche Gefiige).
1m Autoklav bei 115°C sterilisieren. Rosenow-Medium eignet sich zur Kultivierung aller, auch sehr anspruchsvoller Bak
terien. Rosenow-Medium mit Kalbshirn ist inzwischen auch in Rohrchen abgefiillt im Handel (Institut PasteurlFresenius).
Hackf/eischbouillon
500 g Rinderhack in 1 1 Aqua dest. zerquetschen, zur Entfernung des Fettes 15 min im Dampftopf kochen, in kaltem Wasser abkiihlen, das Fett mit einem Mulltuch abschOpfen, pH 8,0 mit NaOH einstellen. Diesen Vorgang zweimal wiederholen. Das Hackfleisch in einem dicken Mulltuch gut ausdriicken, broseln und zum kurzen Trocknen in einer Schale verteilen. Bouillon auf 1000 ml mit Aqua dest. auffiillen und die folgenden Substanzen hinzufiigen:
Hefeextrakt NaCl Na2H P04 x 2H20 Pepton (Difco) Glucose Trypton (Difco) 10s1. Stiirke (Merck)
im Wasserbad los en.
10,0 g 5,0 g 1,0 g 5,0 g 2,Og
10,0 g 1,0 g
Der endgiiltige pH-Wert der Bouillon solI 7,5 betragen. Abfiillen der Bouillon: Hackbrosel (ca. 3 cm3) in Rohrchen geben, ca. 7 ml Bouillon auffiillen und mit Glasstab die Hackbrosellocker aufriihren. Autoklavieren bei 120°C,
Hackfleischbouillon (Cooked Meat Medium) ist im Handel erhaltlich (Difco, BBL). Aufgrund der unterschiedlichen Zusammensetzung dieser Medien kann das Wachstum und auch die T oxinbildung der Clostridien verschieden stark sein.
Glucose-Hefeextrakt-Cystein-Blutagar (27)
Pepton tryptisch verdaut 10,0 g NaCl 5,0 g Fleischextrakt (Difco) 2,0 g Hefeextrakt (Difco) 5,0 g Cystein-HCl 0,3 g G~oo~ ~Og

Bacto-Agar (Difco) dest. H20 pH 7,2 -7,4
Isolierung und Identifizierung von Clostridien 275
20,0 g 1000,0 ml
Aufkochen, autoklavieren, vor Gebrauch 10% defibriniertes Schaf- oder Pferdeblut zusetzen. Giegen. Gleichfalls geeignet ist Schaedler-Agar (Becton Dicksinson, bioMerieux), dem die entsprechende Blutmenge zugesetzt wird.
KanamycinlVancomycin-Blutagar (KV-Agar) (27)
Schaedler-Agar (BBL) Kanamycin Vancomycin Schafblut
100,0 ~g/ml 7,5 ~glml 5,0 vol. %
Den Schaedler-Agar und das Kanamycin zusammengeben und autoklavieren; auf 50°C abkiihlen, Vancomycin und Schafblut hinzufiigen, in Platten giegen.
Anmerkung (27): Das KV-Medium ist in hohem Mage selektiv fiir BacteroidesKeime. Aminoglykosid-hochresistente Enterobacteriaceae wachsen zwar gelegentlich ebenfalls an, lassen sich aber durch aerobes Wachstum auf der Kontrollplatte, anhand des sonstigen Resistenzverhaltens u.a. ausschliegen. Selten sind auch Clostridien zum Wachstum auf KV-Medium befahigt. Die Differentialdiagnose wird in solchen Fallen dadurch erschwert, dag Clostridien auf KV-Medium haufig gramnegativ erscheinen.
Basalmedium fur den Nachweis der biochemischen Reaktionen (27)
Pepton (tryptisch) 10,0 g NaCI 5,0 g Rindfleischextrakt 2,0 g (Liebig oder Difco) Hefeextrakt (Liebig oder Difco) Cystein-Hydrochlorid Bacto-Agar (Difco) Bromkresolpurpurlosung (1,6%ig) Aqua dest. pH 7,4
5,0 g
0,3 g 0,5 g 2,0 ml
1000,0 ml
Die C-Quellen werden 1 %ig zugesetzt. Die C-Quelle sollte bei den Pentosen getrennt autoklaviert werden. Fiir Indol- und HzS-Nachweis sowie Nitratreduktion wird Basalmedium ohne Bromkresolpurpur verwendet.
Fiir den Indoltest werden 0,5 g Tryptophan, fiir die HzS-Bildung 0,2 g FeS04, 0, 3 g NazS03 und 0,1 g NaZSZOl zugesetzt. Fiir die Nitratreduktion werden zum Basalmedium 0,2 g Glucose und 5,0 g NaN03 gegeben (alle Angaben gil-Medium).
Die Medien werden zu 2,5 ml in diinne Rohrchen abgefiillt. Zur Feststellung der Gasbildung konnen Durham-Rohrchen verwendet werden.
Fiir den Urease-Nachweis werden dem Basalmedium (2,5 ml) vor der Verwendung 0,15 ml einer 66%igen Harnstofflosung (Merck 8487) zugesetzt.

276 G. Schallehn
Niihrgelatine (Oxoid): 0,3% Fleischextrakt "Lab Lemco", 0,5% Pepton, 0,5% NaCl, 15% Gelatine, Aqua dest.; pH 6,8.
Milch: Vollmilch wird zu ca. 2,5 mllRohrchen abgefiillt und mit fliissigem Paraffin iiberschichtet. Sterilisation: 0,2-0,3 atii, 105°C 15 min.
Eigelb (Egg Yolk)-Agar
Blood-Agar-Base Oxoid NaCI Leberextrakt (Oxoid) Aqua dest. pH 7,4
4,0 g 1,0 g 0,5 g
100,0 ml
Autoklavieren, auf 50°C abkiihlen und Zusatz von 10% Eigelb-Emulsion (Oxoid).
Pepton-Hefeextrakt-Bouillon (PYGIPY) (27)
parallel mit und ohne Glucose, in Rohrchen zu 9 ml
Pepton, tryptisch (Merck) Fleischextrakt (Liebig) Hefeextrakt (Oxoid) Cystein-HCI (Merck) NaCI Glucose Bacto-Agar (Difco) Aqua dest. pH 7,4
Ellner-Medium
Bacto-Pepton (Difco) Hefeextrakt (Difco) losliche Starke (Merck) MgS04
KH2P04
Na2HP04 X 7 H20 Aqua dest.
15,0 g 2,0 g
10,0 g 0,3 g 5,0 g
10,0 g 0,5 g
1000,0 ml
1,0 g 0,3 g 0,3 g 0,01 g 0,15 g 5,0 g
100,0 ml
pH 7,8 einstellen, zu je 4 oder 8 ml in diinnen Schraubkappenrohrchen autoklavieren.
Duncan-Strang-Medium (4)
Hefeextrakt (Oxoid) Proteose-Pep ton (Difco) losliche Starke (Merck) Natriumthioglycolat Na2HP04 x 2 H20 Aqua dest. pH 7,2 - 7,5
0,4 g 1,5 g 0,4 g 0,1 g 0,7 g
100,0 ml

lsolierung und ldentifizierung von Clostridien 277
Cycloserin-Cefoxitin-Fructose-Agar (CCFA) (5)
Clostridium-difficile-Selektivagar (Oxoid) Clostridium-difficile-Agar-Basis (Best.-Nr. CM 601) Zusammensetzung (g je Liter)
Proteose-Pepton Dinatriumhydrogenphosphat Kaliumdihydrogenphosphat Magnesiumsulfat Natriumchlorid Fructose Agar pH 7,4 ±
40,Og 5,0 g 1,0 g 0,1 g 2,0 g 6,0 g
15,Og
Clostridium-difficile-Supplement (Best.-Nr. SR 96) Zusammensetzung (per Flaschchen) D-Cycloserin 250,0 mg Cefoxitin 8,0 mg
Zubereitung 34,5 g der Basis werden in 500 ml destilliertem Wasser aufgelost und vorsichtig bis zur volligen Losung gekocht. Die Sterilisation erfolgt bei 121°C 15 min. Dem auf 50°C abgekiihlten Basismedium wird der Inhalt des Fliischchens Clostridium-difficile-Supplement aseptisch zugefiigt. Der Inhalt eines Flaschchens wird mit 2 ml sterilem destilliertem Wasser aufgelost. AnschlieBend wird dem Basissupplementgemisch 5-10% defibriniertes Pferdeblut (Best.-Nr. FSR 1056) zugesetzt. Gut mischen und in Petrischalen gieBen.
Norleucin-Tyrosin-(NT) Bouillon (14)
Trypticase Hefeextrakt NaCl Natriumsulfit Salzlosung (s.u.) Aqua dest. pH 7,6 L-Norleucin L-Tyrosin
Salzlosung:
CaCl2
MgS04 K2HP04 KH2P04
NaHC03
NaCl
0,5 g 0,5 g 0,25 g 0,01 g 4,0 ml
96,0 ml
0,2 g 0,2 g
0,2 gil 0,2 gil 1,0 gil 1,0 gil
10,0 gil 2,0 gil

278 G. Schallehn
CaCh und MgS04 in 300 ml Aqua dest. losen, 500 ml Aqua dest. zufiigen, unter standigem Riihren die anderen Salze hinzufiigen. Wenn alles gelost ist, 200 ml Aqua dest. dazugeben. Aufbewahrung bei 4°C.
Phosphatase-Reagenz (19)
a-Naphthyl-(1)-phosphat 0,2 g (Merck 6815) Diazonium-o-dianisidin 0,4 g (Echtblausalz B, Merck 3191)
Die Substanzen nacheinander in 10 ml 0,2 M Natriumcitratpuffer (pH 4,5) oder Natriumacetatpuffer (pH 4,5) losen. Die Losung 1 h bei 4°C stehen lassen und dann durch einen Papierfilter filtrieren. Das Reagenz ist 2-3 Wochen verwendbar, wenn es bei - 20°C bis +4°C aufbewahrt wird. Die Losung vor Gebrauch moglichst auf 37°C anwarmen. Ein paar Tropfen von dem Reagenz werden direkt auf die zu testende Kolonie auf das Niihrmedium getropft. Bei positiver Reaktion verfiirbt sich die Bakterienmasse 1-3 min nach dem Auftropfen des Reagenzes braunrot oder blau. Gelbbraune Verfarbung oder Fehlen derselben gelten als negative Reaktion.
Clostridien Antiseren (Pferd) zum Toxinnachweis (Wellcome)
Best.-Nr.
RP 04 RP 05 RP 06 RP 07 RP 08 RP 09
RP 00
Produkt PackungsgroRe
Clostridium perfringens Typ A 10 ml Clostridium perfringens Typ B Clostridium perfringens Typ C Clostridium perfringens Typ D Clostridium perfringens Typ E Clostridium perfringens-Kontrollserum (normales pferdeserum) Clostridium sordellii
Clostridien Antiseren (Kaninchen) fur 1FT (FITC markiert) Wellcome
Best.-Nr.
RP 78 RP 80 RP 81
Produkt
Clostridium chauvoei Clostridium novyi Clostridium septicum
PackungsgroRe
2 ml
Danksagung. Fiir die hilfreiche und kritische Durchsicht des Manuskripts danke ich Herrn Professor Dr. Gotz Linzenmeier, Essen.
Literatur
1. Brazier, J. S. and J. Hooker: Cross reactivity of Clostridium glycolicum with latex particle reagent for the identification of Clostridium difficile. Lett. App!. Microbio!. 8 (1989) 199-202.

Isolierung und Identifizierung von Clostridien 279
2. Cato, E. P., W. L. George, and S. M. Finegold: Genus Clostridium. Prazmowski 1880, 23AL, pp. 1141-1200. In: Sneath, P. H. A., N. S. Mair, M. E. Sharpe, and J. G. Holt (eds.), Bergey's Manual of Systematic Bacteriology, vol. 2. The Williams & Wilkins Co., Baltimore (1986)
3. Dowell, V. R. and T. M. Hawkins: Laboratory methods in anaerobic bacteriology: CDC Laboratory Manual. Center for Disease Control, Atlanta USA (1978)
4. Duncan, C. 1. and D. H. Strong: Improved medium for sporulation of Clostridium perfringens. Appl. Microbiol. 16 (1986) 82-89.
5. George, W. L., V. L. Sutter, D. Citron, and S. M. Finegold: Selective and differential medium for isolation of Clostridium difficile. J. Clin. Microbiol. 9 (1979) 214-219.
6. Hatheway, Ch. 1.: Toxigenic Clostridia. Clin. Microbiol. Rev. 3 (1990) 66-98. 7. Holdeman, L. V., E. P. Cato, and W. E. C. Moore: Anaerobe Laboratory Manual,
V.P.I. 4th ed. HVA Anaerobe Laboratory, Blacksburg (1977) 8. Jung, E. M. und G. Schallehn: Vorkommen und Verbreitung von Gasodemerregern.
Zbl. Bakt. Hyg., I. Abt. Ref. 254 (1977) 1-54 9. Linzenmeier, G.: Zur Methodik der Anaerobenziichtung. Uber die Eignung des Indika
torprinzips zur Wertbemessung anaerober Nahrmedien. I. Mitteilung: Messung des Redoxpotentials unter anaeroben Verhaltnissen. II. Mitteilung: Die Reoxydation. Arch. Hyg. 143 (1959)537-560,561-586
10. Linzenmeier, G.: Experimentelle Infektionen durch anaerobe Sporenbildner (Clostridien) der Gasbrandgruppe. In: Handbuch der experimentellen Pharmakologie Vol. XVI, Infektionen II, S. 244-291, Hrsg. O. Eichler. Springer, Berlin-Heidelberg-New York (1966)
11. McDonel, J. L. and B. A. McClane: Highly sensitive assay of Clostridium perfringens enterotoxin that uses inhibition of plating efficiency of vero cells grown in culture. J. Clin. Microbiol. 13 (1981) 940-946
12. Miles, B. L., J. A. Siders, and S. D. Allen: Evaluation of a commercial latex test for Clostridium difficile for reactivity with C. difficile and cross-reactions with other bacteria. J. Clin. Microbiol. 26 (1988) 2452-2455
13. Niculescu, E.-R., G. Kottenhoff und R. Malottke: Beitrag zur Differenzierung von Clostridien. Zbl. Bakt. Hyg. A 259 (1985) 307-316
14. Nunez-Montiel, O. L., F. S. Thompson, and V. R. Dowell jr.: Norleucin-Tyrosine broth for rapid identification of Clostridium difficile by gas-liquid chromatography. J. Clin. Microbiol. 17 (1983) 382-385
15. Rolfe, R. D. and S. M. Finegold: Clostridium difficile: Its role in intestinal disease. Academic Press, New York-London-Sydney-Toronto (1988)
16. Schaal, K. P.: Entnahme und Transport von Untersuchungsmaterial zur mikrobiologischen, parasitologischen und serologischen Diagnostik von Infektionskrankheiten. Der Krankenhausarzt 49 (1976) 1-71
17. Schallehn, G.: Zur Toxigenese von Clostridium novyi Typ A. 1. Mitt.: alpha-Toxinbildung in vitro. Zbl. Bakt. Hyg., I. Abt. Orig. A 232 (1975) 91-99
18. Schallehn, G.: Die Sporenbildner. Erkrankungen durch Sporenbildner. In: Lehrbuch der Medizinischen Mikrobiologie, 6. Aufl., S. 432-451, Hrsg. H. Brandis und G. Pulverer. G. Fischer, Stuttgart-New York (1988)
19. Schallehn, G. und H. Brandis: Phosphatase-Reagenz zur schnellen Identifizierung von Clostridium perfringens. Zbl. Bakt. Hyg., I. Abt. Orig. A 225 (1973) 343-345
20. Schallehn, G. und H. Brandis: Die Bedeutung von Clostridium septicum als Gasodemerreger. Immun. Infekt. 13 (1985) 3-8
21. Schallehn, G. und H. Brandis: Vorkommen und Bedeutung von Clostridien im Untersuchungsmaterial von stationar behandelten Patienten aus fUnf verschiedenen Gebieten der Bundesrepublik Deutschland. Wehrmed. Mschr. 31 (1987) 91-109
22. Schallehn, G. und M. H. Wolff: Morphologische Veranderungen humaner embryonaler Lungenfibroblasten durch Cytotoxine verschiedener Clostridium-Spezies. Zbl. Bakt. Hyg. A 267 (1988) 367-378

280 G. Schallehn
23. Smith, L. DS.: Botulism: The organism, its toxins, the disease. Charles C. Thomas, Springfield, IL USA (1977)
24. Smith, L. DS. and B. L. Williams: The pathogenic anaerobic bacteria, 3rd ed. Charles C. Thomas, Springfield, IL USA (1984)
25. Sterne, M. and I. Batty: Pathogenic Clostridia. Butterworths, London-Boston (1975) 26. Wagner, B. und G. Schallehn: Zur Koloniemorphologie von Clostridium perfringens
Iyp A. Zbl. Bakt. Hyg., I. Abt. Orig. A 251 (1982) 537-544 27. Werner, H.: Anaerobier-Infektionen, Pathogenese, Klinik, Therapie, Diagnostik, 2.
Aufl. Thieme, Stuttgart-New York (1985) 28. Wilde, E., H. Hippe, N. Tosunoglu, G. Schallehn, K. Herwig, and G. Gottschalk:
Clostridium tetanomorphum sp. nov., nom. rev .. Int. J. System. Bact. 39 (1989) 127-134
29. Zeissler, j., C. Krauspe und L. Rassfeld-Sternberg: Die Gas6deme des Menschen. Dr. D. Steinkopf, Darmstadt, Bd. I (1958), Bd. II und III (1960)
Prof. Dr. Gisela Schallehn, Institut fur Medizinische Mikrobiologie und Immunologie der Rheinischen Friedrich-Wilhelms-Universitat Bonn, Sigmund-Freud-Str. 25, D-5300 Bonn 1-Venusberg





























