Embed Size (px)
Citation preview

Institut für VirologieHELMUT-RUSKA-HAUS
Akkreditiert nach DIN EN ISO 15189 der Humboldt-Universität
Jahresbericht 2004Berichte des Instituts für Virologie, Band 14 (2004)

Schlaufenbildung eines DNA-Moleküls durchProteinbindung (Kraftfeldmikroskopie)Reich et al., J. Mol. Biol. 2004
Chimaere Viruspartikel mit inseriertenFremdepitopen als rekombinante Vakzine(Computermodell nach Kryoelektronenmikroskopie)Geldmacher et al., Virology 2004
Institut für VirologieHELMUT-RUSKA-HAUS
Direktor: Univ.-Prof. Dr. med. D. H. KrügerNationales Konsiliarlaboratorium für Hantaviren
Charité – Universitätsmedizin BerlinSchumannstr. 20/21
10117 BerlinPostadresse: 10098 Berlin
Tel. +49-30-450-52 50 92Fax +49-30-450-52 59 07
www.charite.de/virologie/
Abbildungen:
links:
rechts:

1
Inhalt
Seite
A. Vorwort 2
B. Kollegium des Instituts 4
C. Lehre 6
D. Vorstellung der Forschungsprojekte 7
E. Medizinische Versorgung 17
F. Publikationen 22
G. Vorträge, Poster, Abstractpublikationen 26
H. Verzeichnis der öffentlichen Institutskolloquien/Gastvorlesungen 35
I. Wissenschaftliche Ehrungen und Mitgliedschaft in wissenschaftlichen Gremien 37
Anlage 1 “61 years ago”: “Über das Virus der Varicellen und des Zoster” von Helmut Ruska
Anlage 2 Aus dem Vorlesungsverzeichnis 2004
Anlage 3 Akkreditierungsurkunde für die virologische Diagnostik
Anlage 4 Leistungsspektrum in der Krankenversorgung
Anlage 5 „Seminarsprecher des Jahres“ am Institut für Virologie
Anlage 6 Programm des Festkolloquiums aus Anlass des 80. Geburtstages von Herrn Professor Dr. Hans-Alfred Rosenthal

2
A. Vorwort Im 47. Jahr seiner Gründung kann das Institut eine positive Bilanz seiner Arbeit in Lehre,
Forschung und Krankenversorgung vorlegen. Von besonderem Wert ist die Einbeziehung des
Institutes in das in Gründung befindliche „Interdisziplinäre Zentrum für Infektionsforschung
und Immunität“ (ZIBI) der Humboldt-Universität, welches das entsprechende Forschungs-
potential am Standort bündeln und in Kooperation neue Förderquellen erschließen soll. Ein
erster schöner Erfolg des ZIBI ist die Bewilligung des gemeinsamen Graduiertenkollegs 1121
durch die DFG, in dem das Institut drei Graduaten betreuen wird. Auch in der neuen Förder-
periode des Sonderforschungsbereiches 421 ist das Institut wieder vertreten.
Aus dem Anlauf des Graduiertenkollegs werden für uns neue Aufgaben in der Aus- und
Weiterbildung erwachsen. Außerdem steht der neue Masterstudiengang „Molekulare Medi-
zin“ der Medizinischen Fakultät, in dem das Institut das Modul „Infektionen“ koordiniert, vor
seinem Start. Diese neuen Aktivitäten sind vor dem Hintergrund zu sehen, dass wir bereits in
den vergangenen Jahren mit unserem Engagement im Reformstudiengang Medizin sowie mit
der Einführung des Nebenfachs Virologie im Diplomstudiengang Biologie zusätzliche Ver-
pflichtungen übernommen haben, die über unsere „traditionellen“ und selbstverständlich
weitergeführten Lehraufgaben bei der Ausbildung der Studenten in den „Normalstudiengän-
gen“ Medizin, Zahnmedizin sowie Medizin- und Pflegepädagogik weit hinausgehen. Aber
auch die Koordinierung und Durchführung der Lehre für die Mediziner unter den Bedingun-
gen der neuen Approbationsordnung und der Fusion der Charité – Universitätsmedizin Berlin
war durchaus eine neue Herausforderung.
Im vergangenen Jahr konnte die Re-Akkreditierung unserer virusdiagnostischen Leistungen
und Laboratorien nach ISO 15189 erfolgreich bestanden werden. Insgesamt haben sich unsere
erbrachten Leistungen in der Krankenversorgung erneut gesteigert und entsprechen für 2004
einer Summe von fast 100 Millionen DKG-NT-Punkten. Hervorzuheben sind die Etablierung
und Nutzung neuer, selbst entwickelter Verfahren zum viralen Nukleinsäurenachweis, mit
denen wir der Charité größere Kostenersparnisse ermöglichen.

3
Allen Mitarbeitern und Studenten des Institutes, die durch ihre engagierte Arbeit diese Lei-
stungen ermöglicht haben, gilt herzlicher Dank. Genauso sind wir unseren Kooperationspart-
nern sehr verbunden.
Den Namenspatron unseres Hauses, Helmut Ruska (1908-1973), hätte es sicherlich gefreut zu
erfahren, wie die von ihm eingeführte Elektronenmikroskopie von uns heute gemeinsam mit
Kooperationspartnern in weiterentwickelter Form genutzt wird: Die Titelseite dieses Jahres-
berichtes wird durch Aufnahmen geziert, die mittels Kryoelektronenmikroskopie und Raster-
kraftmikroskopie gewonnen wurden. Zur Erinnerung an Helmut Ruska enthält dieser Jahres-
bericht eine Kopie seiner Arbeit, die 1942 in der damaligen Klinischen Wochenschrift
erschienen ist und erstmalig das Schicksal von Zellen nach Infektion mit Herpes- bzw.
Pockenviren vergleichend beschreibt – übrigens mit grundsätzlichen Schlussfolgerungen, die
heute noch gültig sind.
Berlin, im Februar 2005
Univ.-Prof. Dr. med. Detlev H. Krüger
Institutsdirektor
Ich danke Frau Christina Grübel für die technische Redaktion des Jahresberichtes.

4
B. Kollegium des Instituts Hochschullehrer und Arbeitsgruppenleiter Krüger, Detlev H., Univ.-Prof. Dr. med. Meisel, Helga, Dr. rer. nat. Prösch, Susanna, PD Dr. rer. nat. Rang, Andreas, Dr. rer. nat. (seit März 2004) Reuter, Monika, PD Dr. rer. nat. Schönrich, Günther, Univ.-Prof. Dr. med. Professoren mit Lehrauftrag Kurth, Reinhard, Prof. Dr. med. (Präsident des Robert Koch-Instituts Berlin) Scherneck, Siegfried, Prof. Dr. rer. nat. (Max Delbrück Centrum für Molekulare Medizin, Berlin) Mitarbeiter(innen) Albrecht, Inka, Dipl.-Biol. (bis Mai 2004) Auste, Brita Bergemann, Andrea Brehmer, Hildegard Bunn, Carmen, Dipl.-Biochem. Dauer, Karin Demakowski, Marina Dürrenfeld, Astrid Endres, Anne-Sophie, Dr. med. Friedrich, Ingrid Ganzer, Annemarie (bis Okt. 2004) Goßler, Bettina (seit Okt. 2004) Grade, Katrin Grübel, Christina Grünberg, Marina Gutzeit, Cindy, Dipl.-Biol. (seit Dez. 2004) Härle, Kathrin, Dr. med. (bis Jan. 2004) Hanemann, Jeannette Heider, Harald, Dr. med. Jacob, Christine, Dipl.-Biol. Jantschak, Joachim, Dr. rer. nat. Just, Monika Kaspari, Marion, Dipl.-Biol. (seit Sept. 2004) Kather, Angela, Dipl.-Biol. Kerger, Gabriele Kershaw, Olivia, Dr. med. vet. (bis Juni 2004) Kersten, Sigrid (Stellv. Leitende MTA)
Kleinkauf, Niels, Dr. med. Klempa, Boris, Dr. rer. nat. Koschke, Sylvia Kraus, Annette, Dipl.-Biol. (bis Okt. 2004) Kühnaß, Beate Lerch, Heike Mackeldanz, Petra Märschenz, Stefanie, Dipl.-Biol. Mertens, Christiane Möncke-Buchner, Elisabeth, Dipl.-Biol. Muske, Karin Noack, Ulrike Piehl, Dirk Pohl, Brigitte Priemer, Christina, Dipl.-Biol. Raftery, Martin, Dr. rer. nat. Scherneck, Ursula (Leitende MTA) Schilf, Rita Schories, Astrid Spingies, Christine Stein, Angela, Dr. med. Steinführer, Simone (z. Z. beurlaubt) Stern, Petra Stephan, Christine Tromp, Hannelore von Grüner, Annett (z. Z. beurlaubt) Wagenführ, Katja, Dipl.-Biol. (seit Okt. 2004) Woskobojnik, Ina

5
Gastdozenten und Gastmitarbeiter Baier, Michael, Dr. rer. nat., Robert-Koch-Institut Berlin Bannert, Norbert, Dr. rer. nat., Robert-Koch-Institut Berlin Denner, Joachim, Dr. rer. nat., Robert-Koch-Institut Berlin Hildt, Eberhard, Dr. rer. nat., Robert-Koch-Institut Berlin Niedrig, Matthias, PD Dr. rer. nat., Robert-Koch-Institut Berlin Tillner, Regina, Zentralbibliothek der Charité Ulrich, Rainer, PD Dr. rer. nat. (bis Jan. 2004) Studentische und wissenschaftliche Hilfskräfte Gille, Sascha (Juni-Dez. 2004) Kaspari, Marion (bis Aug. 2004) Kohl, Stefan (bis Okt. 2004) Kramski, Marit (Jan.-Nov. 2004) Lichtfuss, Gregor Meier, Johannes (bis März 2004)
Milatz, Susanne (seit April 2004) Puck, Clemens (seit Dez. 2004) Rothenberg, Maja (Jan.-Sept. 2004) Schmidt, Jonas Schmidt, Silvia (bis April 2004)
Auszubildende Garkin, Anastasia (bis Feb. 2004) Peuschel, Daniela (seit Sept. 2004)

6
C. Lehre Für die Studenten der Medizinischen Fakultät wird das Fach Virologie in den Studiengängen Humanmedizin, Zahnmedizin und Medizinpädagogik/Pflegepädagogik unterrichtet. Ähnlich wie in den vergangenen Jahren wurden im SS 2004 und WS 2004/2005 jeweils ca. 200 Stu-denten im Regelstudiengang Medizin, 60 Studenten im Reformstudiengang Medizin, ca. 60 Studenten der Zahnmedizin und ca. 20 Studenten der Medizinpädagogik/Pflegepädagogik unterrichtet und geprüft. Für die Studenten im 1. klinischen Semester des Regelstudiengangs Medizin erfolgte im SS 2004 erstmalig der Unterricht nach neuem Curriculum. Parallel wurden die Studenten des 2. klinischen Semesters noch nach altem Curriculum unterrichtet. Da der Unterricht für die Stu-denten im Regelstudiengang Medizin weiterhin parallel am Campus Mitte und Campus Ben-jamin Franklin durchgeführt wird, erfolgte in Vorbereitung des neuen Curriculums eine Abstimmung der Lehrinhalte für die Vorlesungen, Seminare und Praktika sowie die Erstel-lung eines einheitlichen Lernzielkatalogs. Das Praktikum im 2. klinischen Semester wird nun in Kombination Hygiene, Mikrobiologie, Virologie durchgeführt und schließt mit einer prak-tischen Prüfung (OSCE) der Studenten ab. Als Grundlage für das Praktikum wurden ein neues einheitliches Skript sowie entsprechende Prüfungsbögen für den OSCE erstellt. Sowohl das Praktikum als auch die OSCE-Prüfung wurden von den Studenten sehr positiv angenommen. Die im neuen Curriculum enthaltenen Seminare und Vorlesungen für Studenten im 3. und 4. klinischen Semester wurden übergangsweise auch für die Studenten angeboten, die noch nach dem alten Curriculum unterrichtet werden. Die Lehrveranstaltungen, insbesondere Praktika, für die Studenten im Reformstudiengang Medizin wurden in enger Zusammenarbeit mit den Blockverantwortlichen und Studenten ausgebaut, aktualisiert und intensiviert. Für die Praktika wurden den Studenten erweiterte Skripte zur Verfügung gestellt. Im WS 2003/2004 beteiligte sich die Virologie erstmalig an den OSCE-Prüfungen der Studenten des Reformstudienganges im Block Entzündung/Infektion. Mit großem Einsatz wurde auch die Ausbildung der Studenten des Diplomstudienganges Biologie im Fach Virologie durchgeführt. Dazu wurden drei Vorlesungsreihen, ein Komplex- und ein Arbeitsgruppenpraktikum sowie ein von den Studenten gestaltetes Oberseminar durchgeführt. Die ersten Biologiestudenten, die das Fach Virologie absolviert und anschlies-send Diplomanden am Institut wurden, konnten ihre Diplomarbeiten jetzt erfolgreich ab-schließen. Das Zentrum für Infektionsbiologie und Immunität (ZIBI) bot im WS 2004/2005 eine Ring-vorlesung „Infection Biology“ an, die von Hörern aus allen Berliner Universitäten ganz aus-gezeichnet angenommen wurde. An dieser Ringvorlesung hat sich das Institut aktiv beteiligt.

7
D. Übersichten zu den Forschungsprojekten D. 1. Förderung durch außenbegutachtete Drittmittel D. 1. 1. Untersuchungen zum Mechanismus der TNFα-abhängigen (Re)aktivierung
des humanen Cytomegalievirus und seiner therapeutischen Beeinflussbarkeit Projektleiter: Susanna Prösch, Detlev H. Krüger Förderung: Deutsche Forschungsgemeinschaft (Sonderforschungsbereich 421), Univer-
sitäre Forschungsförderung Durch verschiedene klinische Studien konnten wir und andere zeigen, dass Patienten mit systemischen Entzündungen ein deutlich erhöhtes Risiko für eine aktive Infektion oder auch Erkrankung mit dem humanen Cytomegalievirus haben. Als Ursache postulierten wir auf der Basis von experimentellen Daten, dass der Entzündungsmediator TNFα die Reaktivierung des Virus insbesondere in den latent infizierten monozytären Vorläuferzellen über die Aktivie-rung von zellulärem NF-κB stimuliert. Damit kommt es zu einer Hochregulation der sehr frü-hen Gene des Virus. Dieser Prozess kann durch die gegenwärtig zugelassenen Virostatika, die die virale DNA-Synthese hemmen, nicht unterdrückt werden. Ein Schwerpunkt unserer Untersuchungen ist daher die Entwicklung von neuen potentiellen Anti-HCMV-Mitteln, die die sehr frühe Genexpression von HCMV hemmen. Die Experimente führten zur Entdeckung des in-vitro-Anti-HCMV-Effektes von Proteasomeninhibitoren (PI). Erste Untersuchungen zum Wirkmechanismus der PI zeigten, dass sie direkt in die Regulation der sehr frühen Genexpression von HCMV eingreifen und dass dieser Effekt unabhängig von TNFα/NF-κB ist. Schwerpunkte unserer Forschungsarbeiten im vergangenen Jahr waren die Verifizierung des anti-CMV-Effektes von PI im Tiermodell und die Fortsetzung der Untersuchungen zum Mechanismus der anti-HCMV-Wirkung von Proteasomeninhibitoren. In Vorbereitung tier-experimenteller Untersuchungen wurde zunächst der Einfluss von verschiedenen PI auf die Replikation des Ratten-CMV (RCMV) in der Zellkultur untersucht. Hier zeigte sich, dass PI die Replikation des RCMV ebenso effektiv hemmen wie die des HCMV, womit die RCMV-infizierte Ratte als geeignetes Modell für in-vivo-Untersuchungen verifiziert wurde. Parallel wurden für die Auswertung von Tierexperimenten eine quantitative RCMV-DNA-real-time-PCR etabliert. In ersten Tierversuchen zeigte sich dann auch, dass der PI MG262 (nicht aber der PI MG132) die Replikation des RCMV im infizierten Tier hemmt. Diese Untersuchungen werden gegenwärtig fortgeführt. Im Zusammenhang mit dem potentiellen Einsatz von PI zur Unterdrückung der HCMV-Reaktivierung erhebt sich die Frage des Einflusses von PI auf die Immunabwehr gegenüber dem Virus. Um diese Frage experimentell angehen zu können, wurde ein in-vitro-Modell eta-bliert, in dem virusinfizierte humane Lungenfibroblasten als Antigen-präsentierende Zellen fungieren. Der Einfluss von PI auf die Aktivierung von CD8+T-Zellen kann somit nach Inku-bation mit PI-behandelten bzw. unbehandelten HCMV-infizierten Lungenfibroblasten gemes-sen werden. Erste Ergebnisse zeigen, dass der PI MG132 die CD8+-spezifische Immunant-wort gegenüber HCMV zwar hemmt, diese Hemmung aber nicht mehr als 50 % des Kon-trollwertes erreicht. Weitere Untersuchungen sollen jetzt zeigen, ob es in Gegenwart der PI zur gezielten Hemmung von T-Zellen kommt, die bestimmte Virusproteine erkennen.

8
Unsere Untersuchungen zum Mechanismus der PI-vermittelten Hemmung der HCMV-Repli-kation mittels verschiedener unabhängiger Methoden haben zunächst bestätigt, dass PI gezielt in die Synthese der sehr frühen Genprodukte eingreifen. Angriffspunkt ist die Synthese des absolut essentiellen Proteins IE2. Die Hemmung der IE2-Expression erfolgt bereits auf Ebene der Transkription. Gegenwärtige Untersuchungen konzentrieren sich jetzt auf die Identifizie-rung der Targets der PI. In Kooperation mit dem Institut für Biochemie (Prof. Kloetzel) sowie Industriekooperationen wurden weitere PI auf ihren antiviralen Effekt gegenüber HCMV und RCMV, aber auch HSV1 und 2 getestet. In Zusammenarbeit mit S. Vogt (Pädiatrie) und A. Zimmermann (RKI bzw. Uni Düsseldorf) wurde ein BACmid des RCMV-England charakterisiert. Hierbei handelt es sich um eine Virusmutante, in der die Sequenzen von bestimmten Proteinen deletiert wurden und mit deren Hilfe die Funktion dieser Proteine für die Replikation des Virus in vitro und in vivo aufgeklärt werden soll. Die phänotypische und genotypische Charakterisierung der Virusmutante wird demnächst abgeschlossen. Die Replikation des Virus soll dann in vivo untersucht werden. Kooperationen: H. D. Volk, C. Höflich, Institut für Med. Immunologie, Charité
H. Hengel, A. Zimmermann, A. Helenius, RKI, Berlin (jetzt Institut für Virologie, Universität Düsseldorf)
P.-M. Kloetzel, Institut für Biochemie, Charité Millennium Pharmaceuticals, Inc., Cambridge, MA, USA
D. 1. 2. Störungen der Funktion von Dendritischen Zellen nach Infektion mit Herpes-
viren Projektleiter: Günther Schönrich Förderung: Deutsche Forschungsgemeinschaft (Sonderforschungsbereich 421), Univer-
sitäre Forschungsförderung Dendritische Zellen (DC) bekamen ihren Namen aufgrund ihres baumartigen Erscheinungs-bildes. Trotz ihres Namens gehören sie nicht zum Nerven- sondern zum Immunsystem. Sie spielen die entscheidende Rolle bei der Induktion und Organisation der Immunabwehr eines Organismus. Ihre herausragende Stellung unter den Immunzellen beruht u. a. auf ihrer einzig-artigen Ausstattung zur Aufnahme, Verarbeitung und Präsentation von Antigenen. Außerdem sind DC genau dort lokalisiert, wo Organismen mit der Außenwelt kommunizieren: Haut und Schleimhaut von Lunge, Darm und Urogenitaltrakt. Damit sind sie die „Außenposten“ des Immunsystems, d. h. diejenigen Immunzellen, auf die eindringende Erreger (Bakterien, Para-siten und Viren) zuerst treffen. Die Qualität und Quantität einer Immunantwort und damit der Ausgang einer Infektion hängt ganz entscheidend von der Art und Weise ab, wie die DC mit den eindringenden Infektionskeimen interagieren. Das Verständnis der dabei sich abspielen-den molekularen Veränderungen trägt einerseits zur Aufklärung der Funktionsweise dieser wichtigen Immunzellen bei. Andererseits werden dadurch die Strategien aufgedeckt, die Infektionserreger anwenden, um sich trotz des Immunsystems im Organismus auszubreiten und dort zu persistieren (Immunevasion).

9
Herpesviren gehören zu den „erfolgreichsten“ Erregern überhaupt. Diese komplexen Viren sind weltweit verbreitet, infizieren den größten Teil der Menschheit und können zahlreiche Krankheiten hervorrufen. Dabei gilt der Grundsatz „einmal infiziert, immer infiziert“. Auf-grund ihrer Eigenschaften sind DC ein strategisch wichtiges Ziel von Herpesviren. Neben den klassischen, Peptid-bindenden Molekülen des Major Histocompatibility Complex (MHC) exprimieren DC auch sogenannte CD1-Moleküle auf der Zelloberfläche. Trotz ihrer MHC-ähnlichen Struktur weisen diese Antigen-präsentierenden Moleküle keinen Polymorphismus auf. Außerdem binden sie amphipathische Antigene (Glykolipide, Lipopeptide), deren hydro-phobe Bereiche von CD1-restringierten T-Zellen erkannt werden. Humane DC exprimieren 5 Isoformen von CD1-Molekülen, die aufgrund ihrer Homologien in Gruppe 1 (CD1a-c) und 2 (CD1d) eingeteilt werden. Außerdem existiert noch das CD1e-Molekül, welches keiner Gruppe zugeordnet werden kann und nicht auf die Zelloberfläche gelangt. Die einzelnen CD1-Moleküle re-zirkulieren zwischen der Zelloberfläche und ganz unterschiedlichen Zell-kompartimenten. Durch diese Aufgabenteilung wird der Zugang zu den potentiellen Anti-genen optimiert. CD1-restringierte T-Zellen regulieren sowohl die Immunantwort gegen intrazelluläre Erreger und Tumorzellen als auch die Balance zwischen Toleranz und Auto-immunität. In-vivo-Experimente mit Mäusen weisen darauf hin, dass CD1-Moleküle für die antivirale Immunabwehr relevant sind. In unseren Arbeiten konnten wir erstmalig zeigen, dass humanpathogene Herpesviren mit der Rezirkulation der CD1-Moleküle interferieren (Raftery et al., Publikation in Vorbereitung). Die bekannten, MHC-I-inhibierenden viralen Evasions-Proteine sind daran nicht beteiligt und verursachen überraschenderweise das Gegenteil: eine Hochregulation von CD1d, welches als zelluläres Stress-Signal eine frühe antivirale Reaktion des Immunsystems hervorruft (Raftery et al., Publikation in Vorbereitung). Weitere Untersuchungen sollen nun die virusassoziierten Mechanismen analysieren, welche für die Umprogrammierung der Rezirkulation von CD1-Molekülen verantwortlich sind. Außerdem sollen die molekularen Ereignisse geklärt werden, die zu der reziproken Koregulation von MHC-I- und CD1d-Molekülen führen. Die dabei gewonnenen Erkenntnisse werden helfen, ein bisher unbekanntes Feld der antiviralen Immunantwort und der viralen Immunevasion zu verstehen. Darüber hinaus werden die Viren als wertvolle Werkzeuge dienen, um neue Einblicke in die physiologische Regulation und Funktion der humanen CD1-Moleküle zu gewinnen. Kooperationen: U. Schaible, Max-Planck-Institut für Infektionsbiologie, Berlin
S. H. E. Kaufmann, Max-Planck-Institut für Infektionsbiologie, Berlin T. Giese, Institut für Immunologie, Ruprecht-Karls-Universität, Heidelberg
D. 1. 3. Immunevasion von Herpesviren durch Induktion von T-Zell-Apoptose Projektleiter: Günther Schönrich Förderung: Deutsche Forschungsgemeinschaft, Universitäre Forschungsförderung Das Herpes-simplex-Virus (HSV) ist weltweit verbreitet und eines der häufigsten Viren über-haupt. Fast jeder Mensch wird im Laufe seines Lebens von diesem Erreger befallen. Wie alle Mitglieder der Familie der Herpesviren persistiert HSV lebenslang im Organismus. Überra-schenderweise kann das Virus auch Dendritische Zellen (DC) infizieren, obwohl es dafür

10
bekannt ist, dass es sich hauptsächlich in Epithelzellen der Haut bzw. der Schleimhäute ver-mehrt. Durch die Virusinfektion verlieren die Immunzellen ihre Funktionsfähigkeit. Darüber hinaus wird ein Teil der infizierten DC im späteren Verlauf der Infektion in den sogenannten programmierten Zelltod (Apoptose) getrieben. Dies ist erstaunlich, da Immunzellen relativ resistent gegenüber Apoptose-Induktion sind. Deshalb nehmen wir an, dass das Virus inner-halb der Zellen die Sensibilität für Todessignale erhöht. Tatsächlich gelang es, wichtige Apoptose-regulierende Proteine zu identifizieren, deren Expression durch HSV moduliert wird (Müller et al., 2004). Die Menge des antiapoptotischen c-FLIP-Proteins wird stark reduziert, obwohl HSV eine verstärkte Herstellung der c-FLIP-mRNA induziert. Außerdem findet sich in HSV-infizierten Zellen eine größere Menge des proapoptotischen p53 Proteins. Möglicherweise erhöht ein bestimmter Polymorphismus des p53-Gens die Anfälligkeit für HSV-infizierte Apoptose. Weiterführende Analysen ergaben außerdem, dass das c-FLIP-Protein durch Proteasomen-abhängigen Abbau in infizierten Zel-len vermindert wird. Darüber hinaus zeigte es sich, dass HSV auch in anderen Zelltypen c-FLIP auf der Proteinebene herunterreguliert. Jetzt sollen die Mechanismen, die zum Abbau von c-FLIP und zur Hochregulation von p53 führen, weiter aufgedeckt werden. Außerdem soll analysiert werden, ob das Apoptose-blockierende c-FLIP auch vermindert in den Tod-induzierenden Signalkomplex (Death Inducing Signalling Complex, DISC) eingebaut wird. Kooperationen: P. Krammer, H. Walczak, Deutsches Krebsforschungszentrum, Heidelberg
Y. Samstag, Institut für Immunologie, Ruprecht-Karls-Universität, Heidel-berg
T. Giese, Institut für Immunologie, Ruprecht-Karls-Universität, Heidelberg I. Mohr, New York University School of Medicine, New York
D. 1. 4. Molekulare Diversität und Epidemiologie der Hantaviren Projektleiter: Detlev H. Krüger, Helga Meisel Förderung: Deutsche Forschungsgemeinschaft, Universitäre Forschungsförderung Wir wissen aus der Arbeit der Vorjahre, dass in Mitteleuropa mindestens drei verschiedene Hantavirustypen zirkulieren: Puumala, Dobrava und Tula. Vom Hantavirus Dobrava konnten wir eine neue genetische Linie (DOBV-Aa) sowohl im Tierreservoir (Apodemus agrarius) als auch im infizierten Patienten mit Nierenversagen nachweisen. Von der DOBV-Aa-Linie konnte nun erstmalig ein vermehrungsfähiges Virusisolat gewonnen und für weitergehende Untersuchungen eingesetzt werden. Seren von deutschen und slowakischen Patienten mit Nierenversagen infolge Hantavirus-infektionen wurden im Focusreduktionsneutralisationstest auf die Spezifität ihrer Antikörper untersucht. Dabei stellte sich heraus, dass die meisten deutschen Patienten höhere Antikör-pertiter gegen das einheimische DOBV-Aa-Isolat als gegen ein von Apodemus flavicollis stammendes Virusisolat (DOBV-Af) vom Balkan aufwiesen. Obwohl jetzt klar ist, dass DOBV-Aa in Mitteleuropa ein ebenso große Rolle als Krankheitserreger spielt wie das Puu-mala-Hantavirus, wird es interessant sein zu untersuchen, in welcher Form auch hier DOBV-Af vorkommt.

11
Im vergangenen Jahr konnte der molekularepidemiologische Überblick über die Hantavirus-stämme in Mitteleuropa weiter komplettiert werden. So gelang es, Nukleotidsequenzen fol-gender Hantaviren zu amplifizieren und phylogenetisch zu analysieren: Puumala (Nord-deutschland), Tula und DOBV-Af (Westslowakei), DOBV-Aa (Ostslowakei). Mit neuen Bioinformatik-Methoden zur phylogenetischen Nukleotidsequenz-Analyse wurde eine Gesamtuntersuchung aller Hantavirusstämme durchgeführt, die von echten Mäusen (Murinae) beherbergt werden. Die Ergebnisse werfen ein neues Licht auf das Auftreten genetischer Rekombinations- und Reassortmentprozesse in der Evolution dieser Hantaviren. Schließlich wurde im vergangenen Jahr von uns eine RT-PCR-Methode etabliert, die die quantitative Bestimmung der Kopienzahl von Hantavirusgenomen erlaubt und für die Virustypen Dobrava, Puumala, Hantaan, Seoul und Tula einsetzbar ist (LightCycler- und TaqMan Real-Time PCR-Formate). Kooperationen: M. Labuda, Intitut für Zoologie der Slowakischen Akademie der
Wissenschaften, Bratislava, Slowakei B. Hjelle, University of New Mexico at Albuquerque, NM, USA H. A. Schmidt, Institut für Bioinformatik, Universität Düsseldorf M. Schütt, Medizinische Klinik, Universitätsklinikum Lübeck M. Zeier, Sektion Nephrologie, Klinikum der Universität Heidelberg
D. 1. 5. Aufklärung von Struktur und Funktion der Restriktionsendonuklease EcoRII Projektleiterin: Monika Reuter Förderung: Deutsche Forschungsgemeinschaft, Universitäre Forschungsförderung Eine Restriktionsendonuklease ist ein Enzym, das DNA an einer definierten Nukleotid-sequenz spaltet. EcoRII war die erste Restriktionsendonuklease, für die von uns eine wichtige Besonderheit beschrieben wurde, nämlich dass ihre Enzymaktivität von der kooperativen Wechselwirkung mit zwei spezifischen DNA-Erkennungsorten abhängt. Die simultane Inter-aktion mit zwei Erkennungsorten ist inzwischen für zahlreiche Restriktionsendonukleasen nachgewiesen worden, wobei die Art und Weise, wie die Enzyme die beiden Erkennungsorte im aktiven Komplex zusammenbringen und wie sie anschließend die DNA spalten, sehr viel-fältig ist. Eine weitere Besonderheit der Restriktionsendonuklease EcoRII offenbarte sich durch die Aufklärung ihrer dreidimensionalen Struktur und durch die Tatsache, dass eine Verkürzung des Proteins zu einer drastischen Aktivierung der katalytischen Aktivität des Enzyms führt. Die Enzymaktivität von EcoRII wird sehr wahrscheinlich über einen Autoinhibitionsmecha-nismus – eine Art molekularen Schalter – reguliert. Eukaryotische Signaltransduktionspro-teine und Transkriptionsfaktoren zählen zu den sogenannten „Schalterproteinen“, für Restrik-tionsendonukleasen war diese Regulation aber bisher nicht bekannt. Die N-terminalen Domänen des dimeren EcoRII-Proteins reprimieren offenbar in direkter Weise das aktive Zentrum, das durch die beiden katalytischen Domänen gebildet wird (inak-tiver Zustand). Durch die Bindung eines Liganden oder Aktivators an die N-terminalen regu-latorischen Domänen – und das ist ausschließlich spezifische DNA – kommt es zu einer Kon-

12
formationsänderung, so dass die katalytischen Domänen frei werden und das aktive Zentrum formieren können, wo dann ein weiteres spezifisches DNA-Molekül gebunden und gespalten werden kann (aktivierter Zustand). Während wir zeigen konnten, dass die beiden katalytischen Domänen als dimere Struktur einen EcoRII-Erkennungsort binden, ist noch nicht klar, wie die beiden N-terminalen Domä-nen die Bindung des Aktivators bewerkstelligen. Binden beide Domänen gemeinsam das aktivierende DNA-Molekül, oder ist eine ausreichend? Analytische Ultrazentrifugationsexpe-rimente zur Bestimmung der molekularen Masse der N-terminalen Domäne im Komplex mit spezifischer DNA ergaben, dass ein N-Terminus ein DNA-Molekül bindet. Größere Kom-plexe wurden nicht beobachtet. Darüber hinaus schlussfolgern wir aus Gelshift-Experimenten mit der N-terminalen EcoRII-Domäne und spezifischer DNA in verschiedenen Konzentra-tionsbereichen, dass die N-terminale Domäne auch in dimerer Form nur ein DNA-Molekül bindet. Die Frage nach der Funktion der zweiten regulatorischen N-terminalen Domäne bleibt bisher offen. Kooperationen: L. Chen, X. E. Zhou, Dept. of Chemistry, University of Alabama in
Huntsville, USA V. Pingoud, A. Pingoud, Institut für Biochemie, Justus-Liebig-Universität
Gießen V. Siksnys, G. Tamulaitis, Institut für Biotechnologie, Vilnius, Litauen
D. 1. 6. DNA-Erkennung und -Katalyse durch Typ-III-Restriktionsendonukleasen Projektleiter: Detlev H. Krüger, Monika Reuter Förderung: Deutsche Forschungsgemeinschaft, Universitäre Forschungsförderung EcoP15I ist eine ATP-abhängige Typ-III-Restriktionsendonuklease, die etwa 25 bp in 3’-Richtung außerhalb ihrer spezifischen Erkennungssequenz doppelsträngige DNA schnei-det. Das ist der größte definierte Abstand zwischen Erkennungs- und Spaltort, der bisher bei Restriktionsendonukleasen gefunden wurde. Diese besondere Eigenschaft hat dazu geführt, dass im vergangenen Jahr EcoP15I erstmals als sogenanntes „tagging“-Enzym zur Trans-kriptomanalyse mittels SuperSAGE eingesetzt wurde. Während diese erste Anwendung mit überzeugendem Erfolg an mit Pilzen infizierten Reispflanzen durchgeführt werden konnte, haben wir begonnen, mit SuperSAGE das Genexpressionsprofil von virusinfizierten humanen Zellen zu charakterisieren. Um über größere Mengen der Restriktionsendonuklease EcoP15I für SuperSAGE zu verfü-gen, haben wir die codierenden Gene in einen Überexpressionsvektor kloniert, der das Protein gleichzeitig mit einem Affinitätsschwanz versieht, der eine schnelle Aufreinigung von EcoP15I ermöglicht. Die gentechnischen Veränderungen, die dazu notwendig waren, beein-flussen nicht die katalytische Aktivität von EcoP15I. Diese Voraussetzung macht nun erstmals die umfassende Struktur-Funktions-Analyse einer Typ-III-Restriktionsendonuklease möglich. Obwohl wir kürzlich im Rasterkraft-Mikroskop EcoP15I-induzierte DNA-Schlaufen darstellen konnten, die die Fähigkeit des Enzyms zur DNA-Translokation bestätigen, bleiben zahlreiche Fragen zum Reaktionsmechanismus offen.

13
Wir haben zunächst begonnen, mit verschiedenen experimentellen Herangehensweisen Informationen über die Struktur der beiden EcoP15I-Untereineinheiten in An- und Abwesen-heit von DNA zu erhalten. Kooperationen: R. Terauchi, H. Matsumura, Iwate Institut für Biotechnologie, Iwate, Japan
G. Kahl, P. Winter, Fachbereich Biologie der Goethe-Universität Frankfurt am Main
M. W. Linscheid, S. Pieper, Institut für Chemie der Humboldt-Universität zu Berlin
M. Mücke, D. Jeruzalmi, Dept. of Molecular and Cellular Biology, Harvard University, Cambridge, MA, USA
D. 2. Universitäre Forschungsförderung D. 2. 1. Rolle der Sequenzheterogenität des Hepatitis-B-Virus in der Pathogenese der
chronischen Hepatitiden unter Immunsuppression Projektleiterin: Helga Meisel In der Vergangenheit haben wir gezeigt, dass die Anreicherung und Persistenz von komple-xen Hepatitis-B-Virus (HBV)-Varianten mit einem bestimmten Mutationsmuster (Deletionen und Insertionen im regulatorischen Core-Promoter-Bereich, Deletionen in Nukleokapsidgen und Oberflächen-Regionen) signifikant mit der Entwicklung einer Leberzirrhose bei langzeit-immunsupprimierten Nierentransplantatempfängern mit chronischer Hepatitis B assoziiert ist. Das Ziel unserer Arbeiten ist es, die funktionellen Konsequenzen dieser komplexen Muta-tionsmuster zu charakterisieren, um die mögliche Rolle dieser Viren für die Pathogenese zu verstehen. Dazu wurden humane Hepatomazelllinien mit klonierter HBV-DNA von repräsentativen Virusvarianten aus unterschiedlichen Stadien der Lebererkrankung transfiziert. Da die Vari-anten zum größten Teil für ihre Vermehrung und Partikelbildung auf intakte Nukleokapsid- und Oberflächenproteine angewiesen sind, wurden Ko-Transfektionen mit HBV-Wildtyp durchgeführt. Dies entspricht auch der Situation im infizierten Patienten, wo HBV-Varianten zu jedem Zeitpunkt neben dem Wildtyp vorliegen. In den vergangenen zwei Jahren haben wir gezeigt, dass die im Verlauf der Entwicklung schwerer Lebererkrankung akkumulierenden komplexen Varianten in zwei verschiedenen Hepatoma-Zelllinien einen im Vergleich zum Wildtyp veränderten Phänotyp mit einer deut-lich verstärkten Replikation und intrazellulären Anreicherung gegenüber dem Wildtyp, einer stark veränderten Transkription sowie einer defekten Proteinexpression aufweisen. Durch Primer-Extension-Assays konnten wir zeigen, dass die Expression der Präcore-mRNA der Varianten stark vermindert und die der prägenomischen RNA erhöht ist, was weitgehend auf die Mutationen im regulatorischen Core-Promoter-Bereich zurückzuführen ist. Zur Zeit untersuchen wir, ob bei diesen HBV-Varianten das Spleißen, d. h. das Zurecht-schneiden der prägenomischen RNA nach bereits erfolgter Transkription, gegenüber ihrem Wildtyp verändert ist. Die ersten Ergebnisse weisen darauf hin, dass die Varianten im Gegen-

14
satz zum Wildtyp nicht mehr gespleißt werden, was wahrscheinlich auf eine Kombination bestimmter Mutationen im C-Gen und Oberflächengen zurückzuführen ist. Durch verglei-chende Analysen der komplexen Varianten mit Genomen, die nur die mutierten Core-Promo-ter- und Nukleokapsidgen-Abschnitte dieser Varianten in einem genetischen Wildtyp-Hinter-grund enthalten, haben wir gezeigt, dass beide Genombereiche im Wesentlichen für die ver-stärkte Replikation und Anreicherung der Varianten verantwortlich sind. Zusätzlich können jedoch Mutationen außerhalb von Core-Promoter und Nukleokapsidgen einen deutlichen positiven oder negativen Einfluss auf Replikation und Genexpression haben. Um zu klären, ob sich der Phänotyp der komplexen Varianten nicht nur bei einzelnen Vari-antengenomen sondern auch in der gesamten HBV-Population zeigt, die zu einem bestimmten Zeitpunkt während des Fortschreitens der Lebererkrankung im Patienten vorlag, wurden repräsentative Gemische von Variantengenomen mit Wildtyp ko-transfiziert. Es wurde deut-lich, dass auch die Variantenpopulation gegenüber dem Wildtyp verstärkt repliziert und sich anreichert. Weitere Untersuchungen wurden zur Wirkung des mutierten HBV-X-Proteins der Varianten auf NFκB-Aktivierung und Apoptose durchgeführt. Dabei zeigte sich ein Verlust der für Wildtyp-X bekannten NFκB-Aktivierung bei den trunkierten X-Proteinen, die bei den kom-plexen, mit Zirrhose assoziierten Varianten auftreten. Die Experimente zur Apoptose dauern an. Kooperationen: A. Brinckmann, N. Ruf, P. Nürnberg, Gene Mapping Centre, Max-
Delbrück-Centrum für Molekulare Medizin, Berlin Buch T. Heise, Heinrich-Pette-Institut, Hamburg R. Prange, Institut für Mikrobiologie und Hygiene, Johannes-Gutenberg-
Universität, Mainz G. Kristiansen, Institut für Pathologie, Charité
D. 2. 2. Immunpathogenese von Hantaviren Projektleiter: Günther Schönrich, Detlev H. Krüger Die Weltgesundheitsorganisation weist auf eine zunehmende gesundheitliche Bedrohung für den Menschen durch bestimmte Viren hin. Darunter befinden sich neben den Aids-, Ebola- und Marburg-Viren auch die Hantaviren. Hantaviren sind an Nagetiere adaptiert, die als natürliche Wirte fungieren und nicht krank werden. Wenn bestimmte Hantaviren jedoch auf den Menschen übertragen werden, dann können sie ein hämorrhagisches Fieber mit Nieren-versagen hervorrufen. Dem liegt ein Verlust der Barrierefunktion von Endothelzellen zu-grunde, welche die Blutgefäße auskleiden. Die zugrunde liegenden Pathomechanismen sind jedoch unklar. Möglicherweise eliminiert die adaptive Immunantwort der Infizierten nicht nur die Krank-heitserreger, sondern schädigt auch die Endothelfunktion im Sinne einer Immunpathogenese. Alternativ könnte das Virus auch direkt die Funktionsfähigkeit des endothelialen Zytoskeletts negativ beeinflussen. Pathogene Hantaviren können sich vermutlich effizient im menschli-chen Endothel ausbreiten, während die nicht-pathogenen dazu nur sehr schlecht in der Lage sind. Wir konnten die Ursache dieser unterschiedlichen Verhaltensweisen der pathogenen und nicht-pathogenen Hantaviren dingfest machen. Demnach kann ein pathogenes Hantavirus die

15
früh einsetzende angeborene Immunantwort der Endothelzellen verzögern (Kraus et al., 2004). Dabei spielt das IFN-β-induzierte MxA-Protein eine große Rolle, welches von der virusinfizierten Zelle als Abwehrstoff gegen bestimmte RNA-Viren gebildet wird, um deren Vermehrung in der Zelle zu verhindern. Pathogene Hantaviren können offensichtlich die Synthese dieses Proteins für eine gewisse Zeit verzögern. Dadurch schaffen sie sich ein Zeit-fenster, innerhalb dessen sie sich ungestört in den Endothelzellen vermehren können. Im Gegensatz dazu kann ein nicht-pathogenes Hantavirus das frühe Einsetzen der MxA-Bildung nicht verhindern, so dass seine Ausbreitung im Zellverband schon im Keim erstickt wird. Weitere Untersuchungen sollen nun das virale Protein identifizieren, welches bei pathogenen Hantaviren dafür verantwortlich ist, dass MxA verzögert gebildet wird. Zu diesem Zweck wird eine Analyse der viralen Transkripte in infizierten Zellen durchgeführt, um sogenannte Nichtstrukturproteine zu identifizieren. Danach soll herausgefunden werden, an welcher Stelle der vom IFN-Rezeptor ausgehenden Signalkette das hemmende virale Molekül angreift. Außerdem wollen wir klären, ob Hantaviren direkt zelluläre Proteine beeinflussen, welche die Dynamik des endothelialen Zytoskeletts kontrollieren. Schließlich werden wir prüfen, ob Hantaviren neben Dendritischen Zellen auch andere Zellen des Immunsystems infizieren können. Diese Untersuchungen werden zum Verständnis der Pathogenese des virusassoziier-ten Hämorrhagischen Fiebers mit Renalem Syndrom wesentlich beitragen. Aufbauend auf diesen Erkenntnissen könnte langfristig eine wirksame spezifische Therapie entwickelt werden. Kooperationen: S. Hippenstiel, N. Suttorp, Innere Medizin, Charité, Berlin
T. Giese, Institut für Immunologie, Ruprecht-Karls-Universität, Heidelberg Y. Samstag, Institut für Immunologie, Ruprecht-Karls-Universität, Heidel-
berg D. 2. 3. Charakterisierung antiviraler Aktivitäten gegen Hantaviren und potentieller
Escape-Mechanismen Projektleiter: Andreas Rang Hantaviren haben ein minussträngiges, segmentiertes RNA-Genom und gehören zur Familie der Bunyaviridae. Der natürliche Wirt der Hantaviren sind Nagetiere, in welchen das Virus keine pathologischen Effekte verursacht. Eine Übertragung auf den Menschen kann jedoch schwerwiegende Folgen haben und hämorrhagisches Fieber mit renalem oder pulmonalem Syndrom hervorrufen. Ob und welche Syndrome ausgelöst werden, ist im wesentlichen abhängig von der Hantavirus-Spezies, die einen Menschen infiziert. Welche viralen Faktoren für einen pathologischen oder asymptomatischen Infektionsverlauf entscheidend sind, ist bis-her weitgehend unbekannt. Die Virusinfektion führt in einer primären Reaktion zur Aktivie-rung angeborener Immunmechanismen, die einen ersten Schutzwall gegen die Ausbreitung des Virus im Wirt darstellen. Im Zuge dessen kommt es zur Ausschüttung von Interferon (IFN), das Zellen in einen sogenannten „antiviralen Status“ versetzt und eine Infektion u. a. von Hantaviren verhindern kann. Wir wollen klären, wie der IFN-induzierte Block gegen Hantaviren wirkt, und welche viralen Produkte bei dieser Hemmung betroffen sind. Zu diesem Themenkomplex gehört auch die Frage, ob verschiedene Hantaviren unterschiedlich effizient durch IFN gehemmt werden, und ob diese Unterschiede mit der Pathogenität der Hantaviren im Menschen korrelieren.

16
Unsere bisherigen Untersuchungen mit dem hochpathogenen Hantaanvirus haben gezeigt, dass die Produktion von infektiösem Virus in Vero-E6-Zellen durch IFNα ca. 30fach vermin-dert wird. Die Effizienz der Rezeptorbindung oder Aufnahme des Virus war dabei nicht sicht-bar beeinträchtigt. Ferner ergab sich nur eine etwa zweifache Reduktion der Menge an vira-lem Kapsidprotein. Wesentlich deutlicher war die Stärke der antiviralen Aktivität auf der Ebene der RNA. So war die Menge der viralen L-RNA ca. 25fach niedriger als die in unbe-handelten Kontrollzellen. Wir gehen deshalb davon aus, dass weder die Virusaufnahme in die Zelle noch das Kapsidprotein, sondern die Transkription, Replikation und/oder Stabilität der viralen RNA als wesentlicher Angriffspunkt der induzierten antiviralen Aktivität fungieren. In weiteren Untersuchungen sollen die Mechanismen genauer charakterisiert werden, die für die Reduktion der RNA-Menge verantwortlich sind.

17
E. Medizinische Versorgung E.1. Allgemeine Angaben Unsere diagnostischen Laboratorien sind seit dem 23. November 2003 nach DIN EN ISO 15189 akkreditiert. Im November 2004 erfolgte, im Rahmen der regulären Überwachung des Qualitätsmanagementsystems, eine erste fachgutachterliche Überprüfung, die uns erneut eine hohe fachliche Kompetenz in der Durchführung der virologischen Untersuchungen beschei-nigt hat. Die Zahl der Einsendungen lag 2004 wiederum bei mehr als 30.000. Zusammen mit der PCR-Testung von Blutspenden auf HAV, HBV, HCV, HIV und Parvo B19 erfolgte mit > 300.000 virusdiagnostischen Untersuchungen erneut eine Steigerung der virusdiagnostischen Leistungen. 2004 verzeichneten wir eine zunehmende Inanspruchnahme unserer 24-Stunden-Rufbereit-schaft, insbesondere zur Abklärung von Virusinfektionen des Zentralnervensystems, bei Nadelstichverletzungen und bei Organspenden. Dazu hat nicht unwesentlich die Umstellung der meisten unserer 22 akkreditierten qualitativen und quantitativen PCRs auf Real-Time-PCR (LightCycler) unter Mitführung von internen Kontrollen im Jahr 2004 beigetragen. E.2.Klinische Studien Eine große klinische Studie wurde mit der Zentralklinik Emil von Behring Heckeshorn in Berlin zur Rolle der Viren bei der akuten Exazerbation der chronischen Bronchitis durchge-führt, bei der verschiedene Körpersekrete auf Rhino-, Adeno-, Coxsackie-, Metapneumo-, RSV, Influenza- und Parainfluenzaviren mittels PCR und Virusanzucht parallel zur inflam-matorischen Antwort untersucht wurden. Die Untersuchungen sind weitgehend abgeschlos-sen. Die interessanten Ergebnisse werden im September 2005 auf dem European Respiratory Congress in Kopenhagen vorgestellt. Eine weitere große klinische Studie zum prognostischen Wert und Spezifität der E6/E7-mRNA-Bestimmung onkogener Humaner Papillomviren mittels einer Real-Time-NASBA-Technik (NORCHIP) im Vergleich zur HPV-DNA-Bestimmung zwecks Früherkennung des Zervixkarzinoms wurde an 350 zytologisch positiven und zytologisch negativen Zervixabstri-chen von drei großen gynäkologischen Zentren Deutschlands durchgeführt und wird z. Z. ausgewertet. E.3. Blutspenderscreening Eine Erweiterung des HCV-RNA-Blutspenderscreening um den Nachweis von HIV-RNA erfolgt ab Mai 2004 mit einer vom Paul-Ehrlich-Institut (PEI) bestätigten In-house-Methode. Die Leistungsfähigkeit unseres Verfahrens wurde durch erfolgreiche Teilnahme am HIV-Ringversuch, den das Paul-Ehrlich-Institut für die Überprüfung von In-house-Methoden für die Nukleinsäure-Amplifikationstechniken im Blutspendewesen organisiert, gezeigt.

18
Ab Juni 2004 werden von uns alle Plasmen sowie Erythrozyten- und Thrombozytenkonzen-trate zusätzlich zur HIV- und HCV-RNA auch auf HAV-RNA sowie HBV- und Parvovirus-DNA mittels einer In-house-PCR mit automatischer Probenaufarbeitung getestet. Die Häufigkeit der isoliert HCV-RNA-positiven Spender (die also PCR-positiv, aber Antikör-per-negativ waren) betrug bei dem von uns untersuchten Spenderklientel 2004 (wie im Jahr 2003) 0,002 %. Auch der Anteil an HCV-RNA/anti-HCV-positiven Spendern und an nur anti-HCV-positiven Spendern lag mit 0,055 % bzw. mit 0,032 % exakt bei den Vorjahreswerten. Von den durch die Infektionsserologie ermittelten anti-HCV-reaktiven Konserven bestätigten wir mit 66,2 % (verglichen mit 41,2 %) wesentlich mehr als im Vorjahr, was auf eine Umstellung des anti-HCV-Screeningtests in der Infektionsserologie zurückzuführen ist. Von den getesteten Blutspenden erwies sich ein Spender als HIV-RNA/anti-HIV-positiv (0,003 %). Drei Spenden, die im Screeningtest der Infektionsserologie anti-HIV-reaktiv waren, konnten von uns in weiteren ELISAs und Western-Blots nicht bestätigt werden. In der Parvovirus-PCR mit einer von uns festgelegten Ausschlussgrenze von 104 IE/ml erwies sich nur eine Spende als für Parvovirus B19 höher positiv mit einer Viruslast von > 1 Million IE/ml (Genotyp 1). Von den mit der HBV-PCR getesteten Konserven waren 0,007 % HBV-DNA- und HBsAg-positiv. Alle in der HAV-PCR untersuchten Konserven erwiesen sich als HAV-RNA-negativ. E.4. Konsiliarlaboratorium für Hantaviren 2004 haben wir kontinuierlich, aber zusätzlich gehäuft im Zusammenhang mit dem Hanta-virusausbruch vom Typ Puumala in Bayern, eine Vielzahl von Anfragen zur möglichen Übertragung, zum Tierreservoir, klinischen Verlauf, Diagnostik, Therapie und Prophylaxe erhalten. In Zusammenarbeit mit dem Bernhard-Nocht-Institut untersuchten wir bei 38 aus diesem Epidemiegebiet stammenden Patienten den Verlauf der serologischen Hantavirus-Marker mit verschiedenen diagnostischen Methoden, die interessante Ergebnisse hinsichtlich der Wertigkeit der verschiedenen Methoden zur Differentialdiagnose einer akuten Hantavirus-infektion geliefert haben. Außer den 38 mit Puumalavirus-infizierten Patienten (37 aus Bayern, einer aus Baden-Würt-temberg) haben wir die Verdachtsdiagnose einer Hantavirusinfektion bei 5 der insgesamt 139 Patienten bestätigen können, von denen wir Serumabnahmen erhalten haben. Bei diesen Pati-enten (drei aus der Umgebung von Lübeck an der Trave/Wakenitz, einem Gebiet, wo wir bereits in der Vergangenheit mehrere akute Hantavirusinfektionen vom Typ Dobrava gesehen haben, einer aus Schwedt und einer aus Chemnitz) lagen akute Hantavirusinfektionen vom Typ Dobrava vor. Die Dobravavirus-infizierten Patienten mit einem durchschnittlich 8-tägi-gen Klinikaufenthalt ohne Dialysepflichtigkeit zeigten das klinische Bild einer Nephropathia epidemica, z. T. assoziiert mit einer Begleithepatitis und Myokarditis, die Nierenbiopsie ergab milde Tubulusschäden der Nierenrinde.

19
Diese Ergebnisse zeigen wiederum, dass im nördlichen Teil Deutschlands fast ausschließlich akute Hantavirusinfektionen vom Virustyp Dobrava auftreten. E.5. HIV Diagnostik Seit spätestens 2002 wird in vielen westlichen Industriestaaten, wenngleich auf niedrigem Niveau, wieder eine Zunahme von HIV-Erstdiagnosen beobachtet. Ein Teil des beobachteten Anstiegs scheint allein durch die Änderung der Therapiestrategien (Verzögerung des Behandlungsbeginns) begründet zu sein. Darüber hinaus trägt aber wahrscheinlich auch ein verändertes Risikoverhalten, die Zunahme anderer sexuell übertragbarer Infektionen sowie die verlängerte Überlebensdauer von HIV-Infizierten zu dem Anstieg bei. Nach Berichten des RKI für 2004 ist dieser Trend in Deutschland am deutlichsten in den neuen Bundesländern inklusive Berlin, insbesondere in der Gruppe der homosexuellen Männer, zu beobachten. Allein aus den Einsendungen an unser Institut ergaben sich für 2004 50 HIV-Erstdiagnosen (2003: 46 Erstdiagnosen). Die genotypische HIV-Resistenztestung ist inzwischen fest implementierter Bestandteil in europäischen und internationalen Richtlinien zur antiretroviralen Therapie. Wie schon in den vorangegangenen Jahren beobachten wir unverändert eine Zunahme (22 % gegenüber 2003) der Inanspruchnahme dieser Untersuchung zur Resistenztestung von HI-Viren. Besonders erfreulich ist der zunehmende Einsatz der Resistenztestung auch bei noch Therapie-naiven Patienten. Europaweit liegt die Häufigkeit der Übertragung von Resistenzen bei 5 %-27 %. Nach Daten der Serokonverter-Studie des RKI liegt die Übertragungshäufigkeit auch in Deutschland seit 2001 auf einem hohen Niveau zwischen 16 % und 18 %. Unsere Daten erge-ben eine Häufigkeit von ca. 13 % (bei jedoch vergleichsweise geringen Fallzahlen). Entspre-chend findet sich in den aktuellen Deutsch-Österreichischen Leitlinien zur antiretroviralen Therapie (Stand Mai 2004) die Empfehlung zur Resistenztestung auch bei primären, kürzlich erworbenen Neuinfektionen und bei chronischen Infektionen, wenn beabsichtigt ist, eine The-rapie zu initiieren. Entsprechend der Bedeutung der genotypischen Resistenztestung für die HIV-Therapie, haben wir im Dezember 2004 in Zusammenarbeit mit dem Nationalen Referenzzentrum für Retro-viren (Erlangen) einen zweitägigen Workshop in der Charité durchgeführt. Hier wurden gemeinsam mit Virologen verschiedener deutscher Universitäten und Experten aus speziali-sierten Privatlaboratorien Probleme bei der Interpretation von HIV-Sequenzdaten, die für die Resistenztestung erhoben werden, diskutiert. Unter anderem wurden Vorschläge zur Verbes-serung und Vereinheitlichung bestehender Interpretationsalgorithmen erarbeitet. E.6. Interessante Fälle der Diagnostik im Jahre 2004 Primäre Virusinfektionen bei Patienten höheren Alters Hepatitis C Bei einem 77-jährigen Mann mit einer starken Transaminasenerhöhung wurde drei Monate nach einer Bypass-Operation und mehreren angiologischen Operationen eine anikterische akute Hepatitis diagnostiziert. In der ersten Serumabnahme waren eine hohe HCV-Last und anti-HCV-Antikörper nachweisbar. Der Immunoblot zeigte nur eine isolierte c33-Bande, was

20
am ehesten mit einer frischen HCV-Infektion vereinbar ist. Die Typisierung ergab den Geno-typ 1b. Die retrospektive Testung zweier ca. zwei Monate vor der Bypass-Operation abge-nommenen, in unserer Serumbank noch vorhandenen Serumproben ergab keinen Hinweis auf eine bereits zuvor bestehende HCV-Infektion. Es gibt keinerlei Anhaltspunkte für eine Dro-genanamnese. Damit ist nicht auszuschließen, dass die Infektion im Zusammenhang mit den Operationen erfolgte, möglicherweise (trotz Blutspenderscreening) durch die Gabe von Blut-produkten: Leider hat das Gesundheitsamt erst jetzt die Ermittlungen zur Auffindung der Infektionsquelle aufgenommen. HIV Eine 74-jährige Patientin wurde mit dem klinischen Verdacht einer Polymyalgia rheumatica mit Fieber, Myalgien und Kopfschmerzen in der Rheumatologie stationär aufgenommen. Nach klinischer Untersuchung wurde zusätzlich der Verdacht auf eine atypische Pneumonie geäußert. Die virologischen Untersuchungen bewiesen eine HIV1-Infektion mit einer Viruslast von > 100.000 cop/ml Plasma und einem voll ausgeprägten Antikörpermuster im Immunoblot. Bei einer CD4-Zahl von 60/µl und CD8 von 1700/µl befand sich die Patientin bereits im klini-schen Stadium AIDS. Die Resistenztestung ergab eine gegen die gängigen Medikamente voll empfindliche Rekombinante aus den Subtypen A und E (circulating recombinant form CRF01_AE). Unter HAART fiel die Viruslast schnell ab. Darüber hinaus wurde CMV-DNA in der Bronchiallavage, massiv erhöhte HSV1/2-IgM-Antikörper im Serum, benigne HPV-Typen im Genitalabstrich sowie Marker einer ausgeheil-ten Hepatitis B nachgewiesen. Anamnestisch hatte die Patientin keine Blutprodukte erhalten. Auf Befragung gab die Patien-tin mehrere Aufenthalte in Kenia mit entsprechenden Risikokontakten vor ca. zwei Jahren an. Epstein-Barr-Virus In diesem Zusammenhang sei ergänzend ein Fall aus dem Jahre 2001 erwähnt, wo bei einem 90-jährigen Mann mit dem typischen Bild einer schweren Mononukleose, eine EBV-Primär-infektion sowohl serologisch (EBV-IgM positiv, EBV-IgG positiv, EBNA1-IgG negativ; Vorproben aus der Serumbank EBV-Antikörper negativ) als auch molekularbiologisch (Leu-kozyten EBV-DNA PCR-positiv) diagnostiziert wurde. Als mögliche Infektionsquelle kommt in erster Linie eines der fünf Enkelkinder in Frage. Eine komplette Genesung nach prolon-giertem klinischem und serologischem Verlauf korrelierte mit dem späten Auftreten von EBV-IgG und EBNA-Antikörpern erst ca. sieben Monate nach klinischer Erkrankung. Erwähnenswert ist in diesem Zusammenhang auch der fehlende Antikörpernachweis gegen die Herpesviren HSV 1 und 2 sowie CMV bei diesem Patienten. Subakute Sklerosierende Panenzephalitis (SSPE) und Schwangerschaft Eine fulminant verlaufende SSPE nach einer Schwangerschaft bei einer türkischen Patientin hatten wir bereits schon einmal im Jahresbericht 1999 beschrieben. Einen ähnlichen Verlauf haben wir jetzt bei einer 23-jährigen Frau aus Saudi-Arabien beobachtet, die sechs Tage nach Entbindung eines gesunden Säuglings Symptome einer postpartalen Eklampsie entwickelte.

21
Sechs Monate später wurde die bereits apallische und im klinischen Endstadium befindliche Patientin nach Deutschland in die Charité verlegt. Die virologische Diagnose einer SSPE ergab sich aus dem einzig auffälligen Befund sehr hoher Masern-spezifischer IgG-Antikörper in Serum und Liquor mit einem isoliert erhöhten Masern-Antikörperindex von > 13, die durch den Nachweis überwiegend oligoklonaler Ban-den gegen das Masern-Nukleokapsidprotein vom virologischen Institut der Universität Würz-burg bestätigt wurde. Über eine Maserninfektion in früher Kindheit lagen keine Informationen vor. SSPE als Slow-Virus-Infektion manifestiert sich typischerweise im Kindes- bzw. Jugendalter. Das Risiko für die Entwicklung einer SSPE ist bei einer durchgemachten Maserninfektion vor dem 2. Lebensjahr deutlich erhöht. In der Literatur sind bisher nur etwas mehr als 50 Fälle von SSPE bei Erwachsenen über 18 Jahren (Beginn zwischen 20. und 35. Lebensjahr) beschrieben worden. Im Erwachsenenalter manifestiert sich eine SSPE anfangs häufig als kortikale Blindheit. Die sonst typische Myoklonie kann in den Hintergrund treten. Das klini-sche Bild ähnelt der Eklampsie. Während oder kurz nach einer Schwangerschaft verläuft die SSPE oft fulminant, was möglicherweise auf hormonelle und immunologische Veränderun-gen zurückzuführen ist.

22
F. Publikationen 2004 F.1. Original- und Übersichtsarbeiten in referierten Zeitschriften Donoso-Mantke O, Meyer R, Prösch S, Niedrig M: Frequent detection of viral nucleic acids in heart valve tissue. J. Clin. Microbiol. 42 (2004) 2298-2300 Drosten C, Nippraschk T, Manegold C, Meisel H, Brixner V, Roth WK, Apedjinou A, Günther S: Prevalence of hepatitis B virus DNA in anti-HBc-positive/HBsAg-negative sera correlates with HCV but not HIV serostatus. J. Clin. Virol. 29 (2004) 59-68 Ehlers I, Horke S, Reumann K, Rang A, Grosse F, Will H, Heise T: Functional characterization of the interaction between human La and hepatitis B virus RNA. J. Biol. Chem. 279 (2004) 43437-43447 Gedvilaite A, Zvirbliene A, Staniulis J, Sasnauskas K, Krüger DH, Ulrich R: Segments of Puumala hantavirus nucleocapsid protein inserted into chimeric polyomavirus-derived virus-like particles induce a strong immune response in mice. Viral Immunol. 17 (2004) 51-68 Geldmacher A, Schmaler M, Krüger DH, Ulrich R: Yeast-expressed hantavirus Dobrava nucleocapsid protein induces a strong, long-lasting, and highly cross-reactive immune response in mice. Viral Immunol. 17 (2004) 115-122 Geldmacher A, Skrastina D, Petrovskis I, Borisova G, Berriman JA, Roseman AM, Crowther RA, Fischer J, Musema S, Gelderblom HR, Lundkvist A, Renhofa R, Ose V, Krüger DH, Pumpens P, Ulrich R: An amino-terminal segment of hantavirus nucleocapsid protein presented on hepatitis B virus core particles induces a strong and highly cross-reactive antibody response in mice. Virology 323 (2004) 108-119 Klempa B, Schütt M, Auste B, Labuda M, Ulrich R, Meisel H, Krüger DH: First molecular identification of human Dobrava virus infection in Central Europe. J. Clin. Microbiol. 42 (2004) 1322-1325

23
Kraus AA, Raftery MJ, Giese T, Ulrich R, Zawatzky R, Hippenstiel S, Suttorp N, Krüger DH, Schönrich G: Differential antiviral response of endothelial cells after infection with pathogenic and non-pathogenic hantaviruses. J. Virol. 78 (2004) 6143-6150 Möncke-Buchner E, Mackeldanz P, Krüger DH, Reuter M: Overexpression and affinity chromatography purification of the Type III restriction endo-nuclease EcoP15I for use in transcriptome analysis. J. Biotechnol. 114 (2004) 99-106 Müller DB, Raftery MJ, Kather A, Giese T, Schönrich G: Induction of apoptosis and modulation of c-FLIPL and p53 in immature dendritic cells infected with herpes simplex virus. Eur. J. Immunol. 34 (2004) 941-951 Raftery MJ, Wieland D, Gronewald S, Kraus AA, Giese T, Schönrich G: Shaping phenotype, function, and survival of dendritic cells by cytomegalovirus-encoded IL-10. J. Immunol. 173 (2004) 3383-3391 Rang A, Linke B, Jansen B: Detection of RNA variants transcribed from the transgene in Roundup Ready soybean. Eur. Food Res. Technol. (in press, published online: 2 December 2004) Rasche FM, Uhel B, Ulrich R, Krüger DH, Karges W, Czock D, Hampl W, Keller F, Meisel H, von Müller L: Thrombocytopenia and acute renal failure in Puumala hantavirus infections. Emerg. Infect. Dis. 10 (2004) 1420-1425 Razanskiene A, Schmidt J, Geldmacher A, Ritzi A, Niedrig M, Lundkvist Å, Krüger DH, Meisel H, Sasnauskas K, Ulrich R: High yields of stable and highly pure nucleocapsid proteins of different hantaviruses can be generated in the yeast Saccharomyces cerevisiae. J. Biotechnol. 111 (2004) 319-333 Reich S, Gössl I, Reuter M, Rabe JP, Krüger DH: Scanning force microscopy of DNA translocation by the Type III restriction enzyme EcoP15I. J. Mol. Biol. 341 (2004) 337-343

24
Scholz M, Vogel JU, Hover G, Prösch S, Kotchetkov R, Cinatl J, Koch F, Doerr HW, Cinatl J Jr.: Thrombin induces Sp1-mediated antiviral effects in cytomegalovirus-infected human retinal pigment epithelial cells. Med. Microbiol. Immunol. 193 (2004) 195-203 Schroeder C, Heider H, Möncke-Buchner E, Lin TI: The influenza virus ion channel and maturation cofactor M2 is a cholesterol-binding protein. Eur. Biophys. J. Jun 25, 2004 (Epub ahead of print) Schütt M, Meisel H, Krüger DH, Ulrich R, Dalhoff K, Dodt C: Life-threatening Dobrava hantavirus infection with unusually extended pulmonary involve-ment. Clin. Nephrol. 62 (2004) 54-57 Seehofer D, Meisel H, Rayes N, Stein A, Langrehr JM, Settmacher U, Neuhaus P: Prospective evaluation of the clinical utility of different methods for the detection of human cytomegalovirus disease after liver transplantation. Am. J. Transplant. 4 (2004) 1331-1337 Tavakoli S, Schwerin W, Rohwer A, Hoffmann S, Weyer S, Weth R, Meisel H, Diepolder H, Geissler M, Galle PR, Löhr HF, Böcher WO: Phenotype and function of monocyte derived dendritic cells in chronic hepatitis B virus infection. J. Gen. Virol. 85 (2004) 2829-2836 Zhou XE, Wang Y, Reuter M, Mücke M, Krüger DH, Meehan EJ, Chen L: Crystal structure of Type IIE restriction endonuclease EcoRII reveals an autoinhibition mechanism by a novel effector-binding fold. J. Mol. Biol. 335 (2004) 307-319 F.2. Buchbeiträge Krüger DH, Feldmann H: Hantaviren. In: Diagnostik und Therapie von Viruskrankheiten: Leitlinien der Gesellschaft für Virologie. Kompendium für Studierende und Ärzte (Mertens T, Haller O, Klenk HD, Hrg.), 2. überarb. Auflage. Urban & Fischer, Elsevier GmbH, München, 2004, S. 83-88

25
Reuter M, Mücke M, Krüger DH: Structure and function of Type IIE restriction endonucleases: From a plasmid that restricts phage replication to a new molecular DNA recognition mechanism. In: Restriction Endonucleases (Pingoud A, ed). Springer, Berlin Heidelberg New York 2004, pp 261-295 F.3. Miscellaneous Brüvere R, Braun S, Zajakina A, Aleksejeva J, Saripo A, Meisel H, Pumpens P, Kozlovska T: Subcellular localisation of internally deleted HBV core proteins. Acta Universitatis Latviensis, Biology 676 (2004) 53-64 Endres AS, Meisel H: Follow-up of iatrogenic hepatitis C virus infections related to anti-Rh prophylaxis. Viral Hepatitis 12 (2004) 22 Kahl G, Winter P, Matsumura H, Reuter M, Krüger DH, Terauchi R: Molekulares Wechselspiel von Wirt und Pathogen: Simultane, genomweite Transkriptprofi-lierung zweier Organismen mit SuperSAGE. BioSpektrum 10 (2004) 511-513 Ulrich R, Meisel H, Schütt M, Schmidt J, Kunz A, Klempa B, Niedrig M, Kimmig P, Pauli G, Krüger DH, Koch J: Verbreitung von Hantavirusinfektionen in Deutschland. Bundesgesundheitsblatt Gesundheitsforschung Gesundheitsschutz. 47 (2004) 661-670 F.4. Patente Möncke-Buchner E, Mackeldanz P, Reuter M, Krüger DH (inventors): EcoP15I – process conditions and efficient method for its large-scale purification. European patent application EP 04090139.5, April 7, 2004

26
G. Vorträge, Poster, Abstractpublikationen 2004 G.1. Fachtagungen und Gasteinladungen Flunker G, Böhm M, Stein A, Meisel H, Kühl JS, Gürtler L, Seidel W: Genetic variability of human adenoviruses detected in immunosuppressed patients. Abstr. Jahrestagung der Gesellschaft für Virologie, Tübingen, März 2004, S. 115 Gedvilaite A, Zvirbliene A, Dorn D, Lawatscheck R, Pecher G, Özel M, Voronkova T, Schönrich G, Raftery M, Krüger DH, Sasnauskas K, Ulrich R: Hamster polyomavirus-derived virus-like particles represent a promising universal carrier for various vaccine development applications. Abstr. Jahrestagung der Gesellschaft für Virologie, Tübingen, März 2004, S. 426 Gedvilaite A, Zvirbliene A, Dorn D, Lawatscheck R, Pecher G, Özel M, Voronkova T, Schönrich G, Raftery M, Krüger DH, Sasnauskas K, Ulrich R: Hamster polyomavirus-derived virus-like particles represent a promising universal carrier for various vaccine development applications. Abstr. 6th International Conference on Hemorrhagic Fever with Renal Syndrome (HFRS), Hanta-virus Pulmonary Syndrome (HPS) and Hantaviruses, Seoul, June 2004, p. 148 Geldmacher A, Skrastina D, Borisova G, Petrovskis I, Krüger DH, Pumpens P, Ulrich R: A hantavirus nucleocapsid protein segment exposed on hepatitis B virus core particles is highly immunogenic in mice when applied without adjuvants or in the presence of pre-exi-sting anti-core antibodies. Abstr. Jahrestagung der Gesellschaft für Virologie, Tübingen, März 2004, S. 431 Geldmacher A, Skrastina D, Borisova G, Petrovskis I, Krüger DH, Pumpens P, Ulrich R: A hantavirus nucleocapsid protein segment exposed on hepatitis B virus core particles is highly immunogenic in mice when applied without adjuvants or in the presence of pre-exi-sting anti-core antibodies. Abstr. 6th International Conference on Hemorrhagic Fever with Renal Syndrome (HFRS), Hanta-virus Pulmonary Syndrome (HPS) and Hantaviruses, Seoul, June 2004, p. 149 Kather A, Kohl S, Müller DB, Raftery MJ, Schönrich G: Induction of cell apoptosis after infection with Herpes simplex virus. Forschungswochenende des Zentrums für Infektionsbiologie und Immunität der Humboldt-Universität, Zeuthen, Okt. 2004 Kather A, Müller DB, Raftery MJ, Bräutigam K, Johnen C, Kirchhoff S, Krammer PH, Schönrich G: Downregulation of c-FLIP protein in Herpes simplex virus infected cells. 12th Euroconference on Apoptosis, Chania, Greece, Sept. 2004

27
Kather A, Müller DB, Raftery MJ, Schönrich G: Downregulation of c-FLIPL Protein in Herpes Simplex Virus infected cells. Abstr. Jahrestagung der Gesellschaft für Virologie, Tübingen, März 2004, S. 320 Kather A, Schönrich G: Viral modulation of immune function. 7. Kolloquium des SFB 421, Teikyo University, Zeuthen, April 2004 Kleinkauf N, Klempa B, Schmitz T, Krüger DH, Meisel H: Consecutive horizontal and vertical transmission of HIV carrying the NNRTI resistance mutation K103N to a drug-naive mother and child. Abstr. Jahrestagung der Gesellschaft für Virologie, Tübingen, März 2004, S. 540 Klempa B, Krüger DH, Schmidt HA: Molecular evolution of Murinae-associated hantaviruses. Abstr. 6th International Conference on Hemorrhagic Fever with Renal Syndrome (HFRS), Hanta-virus Pulmonary Syndrome (HPS) and Hantaviruses, Seoul, June 2004, p. 33 Klempa B, Stanko M, Labuda M, Ulrich R, Meisel H, Krüger DH: Dobrava hantavirus as an emerging virus in Central Europe: Characterization of the first virus isolate. Abstr. Jahrestagung der Gesellschaft für Virologie, Tübingen, März 2004, S. 298 Klempa B, Stanko M, Labuda M, Ulrich R, Meisel H, Krüger DH: Dobrava hantavirus in Central Europe: Characterisation of the virus isolate from striped field mouse (Apodemus agrarius) captured in Slovakia. Abstr. 6th International Conference on Hemorrhagic Fever with Renal Syndrome (HFRS), Hanta-virus Pulmonary Syndrome (HPS) and Hantaviruses, Seoul, June 2004, p. 102 Koch J, Meisel H, Ulrich R, Wolbert A, Auste B, Razanskiene A, Sasnauskas K, Ziaja B, Niedrig M, Pauli G, Krüger DH: The seroprevalence for hantavirus infections analysed by a representative serum panel of the German population is about 1 %. Abstr. 6th International Conference on Hemorrhagic Fever with Renal Syndrome (HFRS), Hanta-virus Pulmonary Syndrome (HPS) and Hantaviruses, Seoul, June 2004, p. 143 Kraus AA, Raftery MJ, Giese T, Ulrich R, Zawatzky R, Hippenstiel S, Suttorp N, Krüger DH, Schönrich G: Differential antiviral response of endothelial cells after infection with pathogenic and non-pathogenic hantaviruses. Abstr. Jahrestagung der Gesellschaft für Virologie, Tübingen, März 2004, S. 52

28
Kraus AA, Raftery MJ, Giese T, Ulrich R, Zawatzky R, Schönrich G, Krüger DH: Differential antiviral response of endothelial cells after infection with pathogenic and non-pathogenic hantaviruses. Abstr. 6th International Conference on Hemorrhagic Fever with Renal Syndrome (HFRS), Hanta-virus Pulmonary Syndrome (HPS) and Hantaviruses, Seoul, June 2004, p. 43 Kraus AA, Raftery MJ, Ulrich R, Krüger DH, Schönrich G: Hantaviruses and the concept of immunopathogenesis. Abstr. 6th International Conference on Hemorrhagic Fever with Renal Syndrome (HFRS), Hanta-virus Pulmonary Syndrome (HPS) and Hantaviruses, Seoul, June 2004, p. 39 Krüger DH: Genetisches Reassortment und intragenische Rekombination in der Evolution der Hantaviren. Kolloquium am Friedrich-Loeffler-Institut für Medizinische Mikrobiologie, Universität Greifswald, Jan. 2004 Krüger DH: Transkriptom-Analyse durch Super-SAGE. 9. Kolloquium des SFB 421, Zeuthen, April 2004 Krüger DH: Emerging viruses - Molekulare Evolution und Pathogenese der Hantaviren. Infektiologisch-Mikrobiologisch-Virologisches Kolloquium der Universität Ulm, April 2004 Krüger DH: New hantaviruses – epidemiology and clinical relevance. ConVir 2004, 3rd European Conference on Viral Diseases, Regensburg, May 2004 Krüger DH: Emerging viruses – Molekulare Evolution und Pathogenese der Hantaviren. Seminar des Sonderforschungsbereiches 466, Universität Erlangen-Nürnberg, Erlangen, Juni 2004 Krüger DH: Structure of viruses and the interplay of virus and host cell. Ringvorlesung „Infection Biology“, Zentrum für Infektionsbiologie und Immunität der Humboldt-Universität (ZIBI), Berlin, 25. Okt. 2004 Krüger DH: Virale Zoonosen – Gefahren durch den Wirtswechsel vom Tier zum Menschen: HIV, Hanta-viren, Vogelgrippe. Sitzung der Berliner Medizinischen Gesellschaft, Berlin, 10.11.2004

29
Krüger DH: Emerging viruses – Molecular evolution and pathogenicity of hantaviruses. Kolloquienreihe „Infection Biology and Immunity“, Zentrum für Infektionsbiologie und Immunität der Humboldt-Universität (ZIBI), Berlin, 25. Nov. 2004 Krüger DH, Klempa B, Ulrich R, Schönrich R, Labuda M, Meisel H: Distribution of different pathogenic hantaviruses in Central Europe. Abstr. 6th International Conference on Hemorrhagic Fever with Renal Syndrome (HFRS), Hanta-virus Pulmonary Syndrome (HPS) and Hantaviruses, Seoul, June 2004, p. 62 Krüger DH, Reich S, Möncke-Buchner E, Mackeldanz P, Matsumura H, Terauchi R, Reuter M: Type III restriction endonuclease EcoP15I: Mechanism of DNA interaction and use in mole-cular diagnostics. Abstr. 5th New England Biolabs Meeting on Restriction/Modification, Bristol, Sept. 2004, p. 87 Krüger DH, Reuter M: An engineered enzyme, EcoRII-C, allows reliable detection of DNA cytosine methylation. Symposium der Deutschen Akademie der Naturforscher Leopoldina „DNA Methylation: Significance in Biology and Pathogenesis“. Weißenburg in Bayern, May 2004 Lichtfuss G, Märschenz S, Hildt E, Meisel H: Functional analysis of naturally occurring hepatitis B virus X protein mutants associated with liver cirrhosis in immunosuppressed patients. Abstr. International Hepatitis B Virus Meeting, Woods Hole, Massachusetts, USA, Oct. 2004, p. 81 Märschenz S, Endres AS, Brinckmann A, Nürnberg P, Krüger DH, Meisel H: Phenotype of complex hepatitis B virus variants associated with liver cirrhosis in immuno-suppressed patients. Abstr. International Hepatitis B Virus Meeting, Woods Hole, MA, October 2004, p. 80 Märschenz S, Endres AS, Kristiansen G, Krüger DH, Meisel H: Phenotypic characterization of complex hepatitis B virus variants. Abstr. Jahrestagung der Gesellschaft für Virologie, Tübingen, März 2004, S. 181 Matsumura H, Reich S, Fujisawa S, Ito A, Saitoh H, Winter P, Kahl G, Reuter M, Krüger DH, Terauchi R: SuperSAGE, a transcriptome tool for any eukaryotic organisms and direct application of 26 bp tags to functional genomics. Plant & Animal Genomes XII Conference, San Diego, CA, Jan. 2004

30
Matsumura H, Reich S, Ito A, Saitoh H, Kamoun S, Winter P, Kahl G, Reuter M, Krüger DH, Terauchi R: SuperSAGE: a universal functional genomics tool for eukaryotes. Plant & Animal Genomes XII Conference, San Diego, CA, Jan. 2004 Matsumura H, Reich S, Ito A, Saitoh H, Winter P, Kahl G, Reuter M, Krüger DH, Terauchi R: SuperSAGE: a powerful SAGE variant suitable for gene function analysis. Internat. Conference “SAGE 2004“, Boston, MA, Oct. 2004 Meier J, Lienicke U, Tschirch S, Wauer RR, Hamprecht K, Krüger DH, Prösch S: Human cytomegalovirus mother-to-child transmission in preterm infants: Mechanism of HCMV (re)activation during lactation. Abstr. Jahrestagung der Gesellschaft für Virologie, Tübingen, März 2004, S. 197 Meisel H: Diagnostik der Meningitis/Meningoencephalitis und Encephalitits aus der Sicht der Virologie. Infektiologischer Workshop „Schwerkranke und Infektionen; Sepsis, Pneumonia, Meningitis“ der Charité Berlin und der DRK-Kliniken, Berlin, Nov. 2004 Meisel H, Klempa B, Ulrich R, Haring B, Schütt M, Schwarz A, von Müller L, Krüger DH: Hantavirus infections in Germany: Hemorrhagic fever with renal syndrome cases due to Puumala, Dobrava, and Tula virus infections. Abstr. 6th International Conference on Hemorrhagic Fever with Renal Syndrome (HFRS), Hanta-virus Pulmonary Syndrome (HPS) and Hantaviruses, Seoul, June 2004, p. 58 Meisel H, Plentz A, Klempa B, Kershaw O, Endres AS, Liefeld L, Modrow S: Recurrent high parvovirus B19 (genotype 2) viremia in a renal transplant patient treated with high-dose intravenous immunoglobulin. Abstr. Jahrestagung der Gesellschaft für Virologie, Tübingen, März 2004, S. 116 Prösch S: HCMV reactivation during lactation and mother-to-child transmission in preterm infants. Jahrestagung der Niederländischen Gesellschaft für virologische Labordiagnostik, AZM, University of Maastricht, NL, Okt. 2004 Prösch S: Role of different cellular transcription factors for regulation of the IE1/2 enhancer/promoter and HCMV replication. 5th International Meeting on HCMV-related immunopathology „Oncomodulation“, Frankfurt/Main, Okt. 2004

31
Prösch S: Human Cytomegalovirus – mechanisms of pathogenesis. ZIBI-Forschungswochenende, Zeuthen, Okt. 2004 Prösch S: CMV-Pneumonie nach Transplantation: Aktuelle Prophylaxe- und Therapiestrategien. 19. Jahrestagung der Paul-Ehrlich-Gesellschaft, Bonn, Okt. 2004 Prösch S: Mechanismen der HCMV-(Re)aktivierung und Identifikation neuer potentieller Targets für die antivirale Therapie. Wiss. Vortrag im Sonderforschungsbereich 473, Universität Erlangen-Nürnberg, Erlangen, Okt. 2004 Prösch S, Zimmermann A, Wendt CEC, Wuttke R, Beutler T, Asadullah K, Krüger DH, Hengel H, Volk HD: Glucocorticoid stimulation of human cytomegalovirus replication: Identification of functio-nally active glucocorticoid rreceptor responsive elements (GRE) in the human cytomegalo-virus IE1/2 enhancer. Abstr. Jahrestagung der Gesellschaft für Virologie, Tübingen, März 2004, S. 244 Prösch S, Zimmermann A, Wendt CEC, Wuttke R, Beutler T, Asadullah K, Krüger DH, Hengel H, Volk HD: Identification and functional assessment of glucocorticoid receptor responsive elements (GRE) in the human cytomegalovirus IE1/2 enhancer. Abstr. 29th Annual International Herpesvirus Workshop, Reno, NV, July 2004, 1.32 Raftery M, Schönrich G: Viral modulation of CD1 molecules. Abstr. Jahrestagung der Gesellschaft für Virologie, Tübingen, März 2004, S. 34 Raftery M, Schönrich G: Viral modulation of CD1 molecules. 7. Kolloquium des SFB 421, Teikyo University, Zeuthen, April 2004 Raftery M, Schönrich G: The CD1 system reloaded: antiviral function and viral evasion of an ancient antigen presenta-tion system. Abstr. 3. Workshop Study Group „Immunobiology of viral infections“, Schloss Zeilitzheim, Sept. 2004, T-26

32
Razanskiene A, Petraityte R, Geldmacher A, Schmidt J, Ritzi A, Niedrig M, Lundkvist Å, Krüger DH, Meisel H, Sasnauskas K, Ulrich R: High yields of stable and highly pure nucleocapsid proteins of different hantaviruses can be generated in the yeast Saccharomyces cerevisiae. Abstr. 6th International Conference on Hemorrhagic Fever with Renal Syndrome (HFRS), Hanta-virus Pulmonary Syndrome (HPS) and Hantaviruses, Seoul, June 2004, p. 150 Reuter M, Möncke-Buchner E, Krüger DH: Transcriptome analysis of cells and interacting parasites by SuperSAGE. ZIBI-Forschungswochenende, Zeuthen, Okt. 2004 Reuter M, Mücke M, Zhou XE, Chen L, Krüger DH: Modular design of restriction endonuclease EcoRII and its functional implications. Abstr. 5th New England Biolabs Meeting on Restriction/Modification, Bristol, Sept. 2004, p. 33 Schmidt J: Detection of hantavirus-reactive sera in wild-trapped rodents from Thailand by newly developed immunological assays based on yeast-expressed Seoul virus nucleocapsid protein. Abstr. 15th European Students Conference, Berlin, Okt. 2004, S. 145 Schmidt J, Jandrig B, Khoprasert Y, Klempa B, Yoshimatsu K, Arikawa J, Meisel H, Niedrig M, Krüger DH, Jäkel T, Ulrich R: Detection of hantavirus-reactive sera in wild-trapped rodents from Thailand by newly develo-ped immunological assays based on yeast-expressed Seoul virus nucleocapsid protein. Abstr. 6th International Conference on Hemorrhagic Fever with Renal Syndrome (HFRS), Hanta-virus Pulmonary Syndrome (HPS) and Hantaviruses, Seoul, June 2004, p. 134 Schmidt J, Jandrig B, Khoprasert Y, Klempa B, Yoshimatsu K, Arikawa J, Meisel H, Niedrig M, Krüger DH, Jäkel T, Ulrich R: Detection of Thailand hantavirus-reactive sera in wild-trapped rodents from Thailand by new immunological assays based on yeast-expressed Seoul virus nucleocapsid protein. Abstr. Medizinische B-Schutz-Tagung, München, Okt. 2004, S. 104 Schmidt J, Meisel H, Hjelle B, Capria SG, Vial PG, Padula P, Krüger DH, Ulrich R: Development and evaluation of serological assays for the detection of human infections by New World hantaviruses imported to Europe. Abstr. Jahrestagung der Gesellschaft für Virologie, Tübingen, März 2004, S. 75 Schmidt J, Meisel H, Hjelle B, Capria SG, Vial PG, Padula P, Krüger DH, Ulrich R: Development and evaluation of serological assays for the detection of human infections by new world hantaviruses imported to Europe. Abstr. 6th International Conference on Hemorrhagic Fever with Renal Syndrome (HFRS), Hanta-virus Pulmonary Syndrome (HPS) and Hantaviruses, Seoul, June 2004, p. 123

33
Schmidt J, Meisel H, Hjelle B, Capria SG, Vial PG, Padula P, Krüger DH, Ulrich R: Development and evaluation of serological assays for the detection of human infections by new world hantaviruses imported to Europe. Abstr. Medizinische B-Schutz-Tagung, München, Okt. 2004, S. 105 Schmidt S, Endres AS, Märschenz S, Krüger DH, Meisel H: Phenotypic susceptibility of hepatitis B virus variants from immunosuppressed patients to nucleoside analogues. Abstr. Jahrestagung der Gesellschaft für Virologie, Tübingen, März 2004, S. 228 Schönrich G: Dendritische Zellen als Ziel der viralen Immunevasion. Vortrag Universitätsklinikum Kiel, April 2004 Schönrich G: Virale Strategien der Immunevasion. Vortrag Universitätsklinikum Mainz, Juni 2004 Schönrich G: Pathogenetische Relevanz von Immune Escape Mechanismen bei viralen Infektionen. Vortrag Universitätsklinikum Mainz, Juni 2004 Schönrich G: The role of DCs in the regulation of T cell-mediated immunity. Vortrag am Karolinska-Institut, Stockholm, Schweden, Sept. 2004 Schönrich G: DCs as a target of viral immune evasion strategies. Vortrag am Karolinska-Institut, Stockholm, Schweden, Sept. 2004 Schönrich G: Dendritic cells as a target for viral immune evasion strategies. Kolloquienreihe „Infection Biology and Immunity“, Zentrum für Infektionsbiologie und Immu-nität der Humboldt-Universität (ZIBI), Berlin, Okt. 2004 Ulrich R, Schmidt J, Razanskiene A, Tackmann K, Mattis R, Müller T, Klempa B, Jäkel T, Niedrig M, Pauli G, Koch J, Meisel H, Krüger DH, Conraths FJ: Diagnostik von zoonotischen Hantaviren bei Mensch und Maus. Abstr. 23. AVID-Tagung, Deutsche Veterinärmedizinische Gesellschaft, Arbeitskreis für veterinärmedizinische Infektionsdiagnostik, Kloster Banz, Sept. 2004, p. V4

34
Von Müller L, Rasche FM, Uhel B, Ulrich R, Krüger DH, Mertens T, Karges W, Czock D, Hampl W, Keller F, Meisel H: Thrombocytopenia is a predictor for acute renal failure in Puumala hantavirus infections. Abstr. Jahrestagung der Gesellschaft für Virologie, Tübingen, März 2004, S. 73 G.2. Öffentlichkeitsarbeit Krüger DH: Geflügelgrippe – Gefahr für den Menschen durch Wirtswechsel des Virus? Radiosendung, Südwestrundfunk, Feb. 2004 Krüger DH: Neue Viren - neue Gefahren? Wiss. Vortragsserie der Lehmanns Fachbuchhandlung für Wissenschaft & Literatur, Berlin, April 2004 Krüger DH: Situation nach einem Hantavirus-Ausbruch in der Gegend von Passau. TV-Sendung des Bayerischen Rundfunks, 26.09.2004
35
H. Öffentliche Institutskolloquien/Gastvorlesungen des Jahres 2004
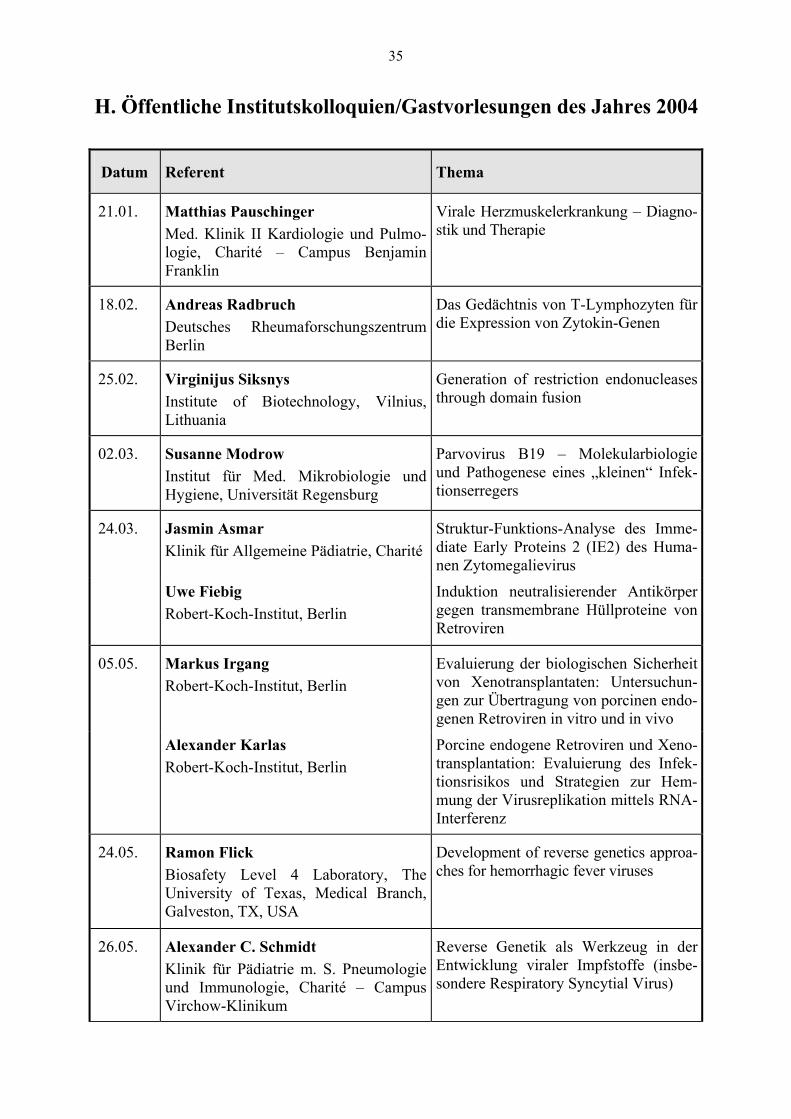
Datum Referent Thema
21.01. Matthias Pauschinger Med. Klinik II Kardiologie und Pulmo-logie, Charité – Campus Benjamin Franklin
Virale Herzmuskelerkrankung – Diagno-stik und Therapie
18.02. Andreas Radbruch Deutsches Rheumaforschungszentrum Berlin
Das Gedächtnis von T-Lymphozyten für die Expression von Zytokin-Genen
25.02. Virginijus Siksnys Institute of Biotechnology, Vilnius, Lithuania
Generation of restriction endonucleases through domain fusion
02.03. Susanne Modrow Institut für Med. Mikrobiologie und Hygiene, Universität Regensburg
Parvovirus B19 – Molekularbiologie und Pathogenese eines „kleinen“ Infek-tionserregers
24.03. Jasmin Asmar Klinik für Allgemeine Pädiatrie, Charité
Struktur-Funktions-Analyse des Imme-diate Early Proteins 2 (IE2) des Huma-nen Zytomegalievirus
Uwe Fiebig Robert-Koch-Institut, Berlin
Induktion neutralisierender Antikörper gegen transmembrane Hüllproteine von Retroviren
05.05. Markus Irgang Robert-Koch-Institut, Berlin
Evaluierung der biologischen Sicherheit von Xenotransplantaten: Untersuchun-gen zur Übertragung von porcinen endo-genen Retroviren in vitro und in vivo
Alexander Karlas Robert-Koch-Institut, Berlin
Porcine endogene Retroviren und Xeno-transplantation: Evaluierung des Infek-tionsrisikos und Strategien zur Hem-mung der Virusreplikation mittels RNA-Interferenz
24.05. Ramon Flick Biosafety Level 4 Laboratory, The University of Texas, Medical Branch, Galveston, TX, USA
Development of reverse genetics approa-ches for hemorrhagic fever viruses
26.05. Alexander C. Schmidt Klinik für Pädiatrie m. S. Pneumologie und Immunologie, Charité – Campus Virchow-Klinikum
Reverse Genetik als Werkzeug in der Entwicklung viraler Impfstoffe (insbe-sondere Respiratory Syncytial Virus)
36
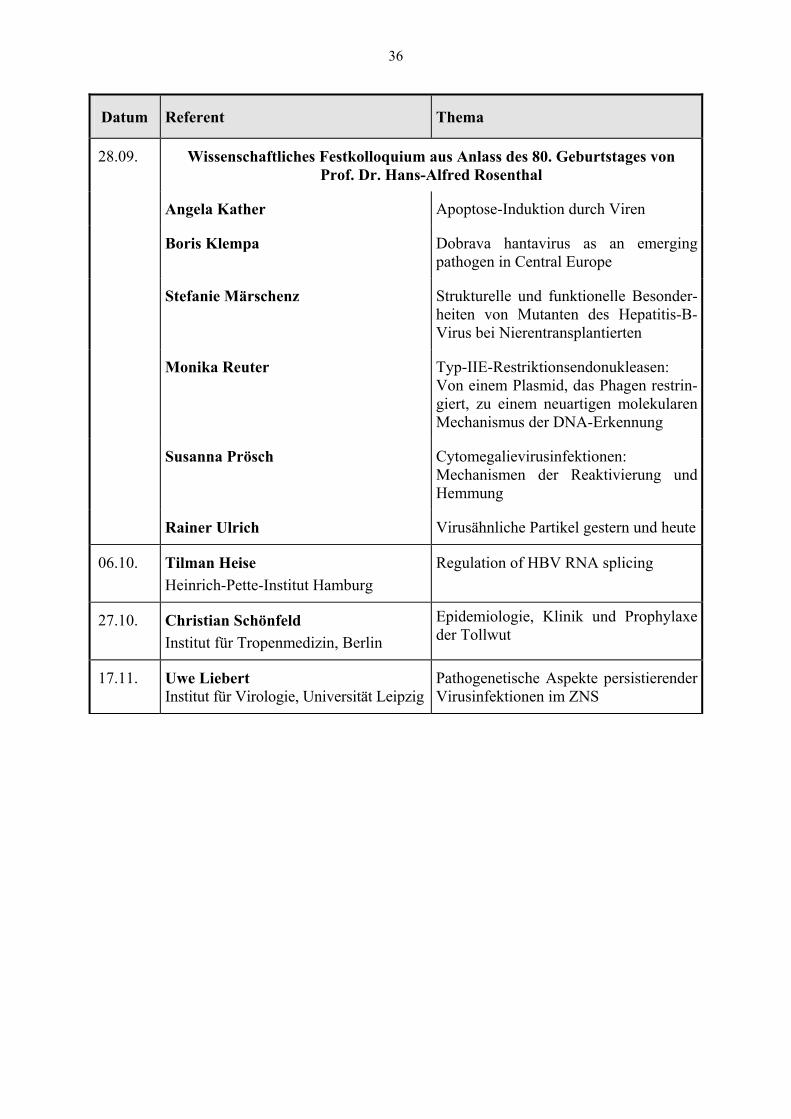
Datum Referent Thema
28.09. Wissenschaftliches Festkolloquium aus Anlass des 80. Geburtstages von Prof. Dr. Hans-Alfred Rosenthal
Angela Kather Apoptose-Induktion durch Viren
Boris Klempa Dobrava hantavirus as an emerging pathogen in Central Europe
Stefanie Märschenz Strukturelle und funktionelle Besonder-heiten von Mutanten des Hepatitis-B-Virus bei Nierentransplantierten
Monika Reuter Typ-IIE-Restriktionsendonukleasen: Von einem Plasmid, das Phagen restrin-giert, zu einem neuartigen molekularen Mechanismus der DNA-Erkennung
Susanna Prösch Cytomegalievirusinfektionen: Mechanismen der Reaktivierung und Hemmung
Rainer Ulrich Virusähnliche Partikel gestern und heute
06.10. Tilman Heise Heinrich-Pette-Institut Hamburg
Regulation of HBV RNA splicing
27.10. Christian Schönfeld Institut für Tropenmedizin, Berlin
Epidemiologie, Klinik und Prophylaxe der Tollwut
17.11. Uwe Liebert Institut für Virologie, Universität Leipzig
Pathogenetische Aspekte persistierender Virusinfektionen im ZNS

37
I. Wissenschaftliche Ehrungen und Mitgliedschaft in wissen-schaftlichen Gremien 2004
Detlev H. Krüger
• Mitglied des Wissenschaftlichen Beirates des Paul-Ehrlich-Institutes (Bundesinsti-tut für Sera und Impfstoffe), Langen
• Member of the International Advisory Board, Institute of Biotechnology/European Biotechnology Centre of Excellence, Vilnius, Lithuania
• Mitglied des Beirates und verschiedener Kommissionen der Gesellschaft für Viro-logie
• Mitglied des Beirates der Paul-Ehrlich-Gesellschaft für Chemotherapie
• Member of Board, International Society for Hantaviruses and Hantavirus Diseases
• Stellv. Mitglied des Arbeitskreises Blut beim Bundesgesundheitsministerium
• Mitglied des Diagnostikausschusses der Deutschen Vereinigung zur Bekämpfung der Viruskrankheiten (DVV)
• Mitglied des Weiterbildungsausschusses der Ärztekammer Berlin
• Mitglied des Wissenschaftlichen Beirates des Biotechnologie-Parkes Luckenwalde/ Land Brandenburg
• Mitglied des Kompetenzzentrums für hochkontagiöse lebensbedrohliche Krankhei-ten bei der Senatorin für Gesundheit, Soziales und Verbraucherschutz des Landes Berlin
Jonas Schmidt
• Zondek Prize of the Grand Jury, 15th European Students Conference (ESC), Berlin, Okt. 2004
• ESC Session Award „Infectious Diseases“, 15th European Students Conference
(ESC), Berlin, Okt. 2004 Günther Schönrich
• Leiter der Arbeitsgemeinschaft „Immunbiologie von Virusinfektionen“ der Gesellschaft für Virologie

Anlage 1
„61 years ago“
Ruska H: Über das Virus der Varicellen und des Zoster Klinische Wochenschrift 22 (1943) 703-704

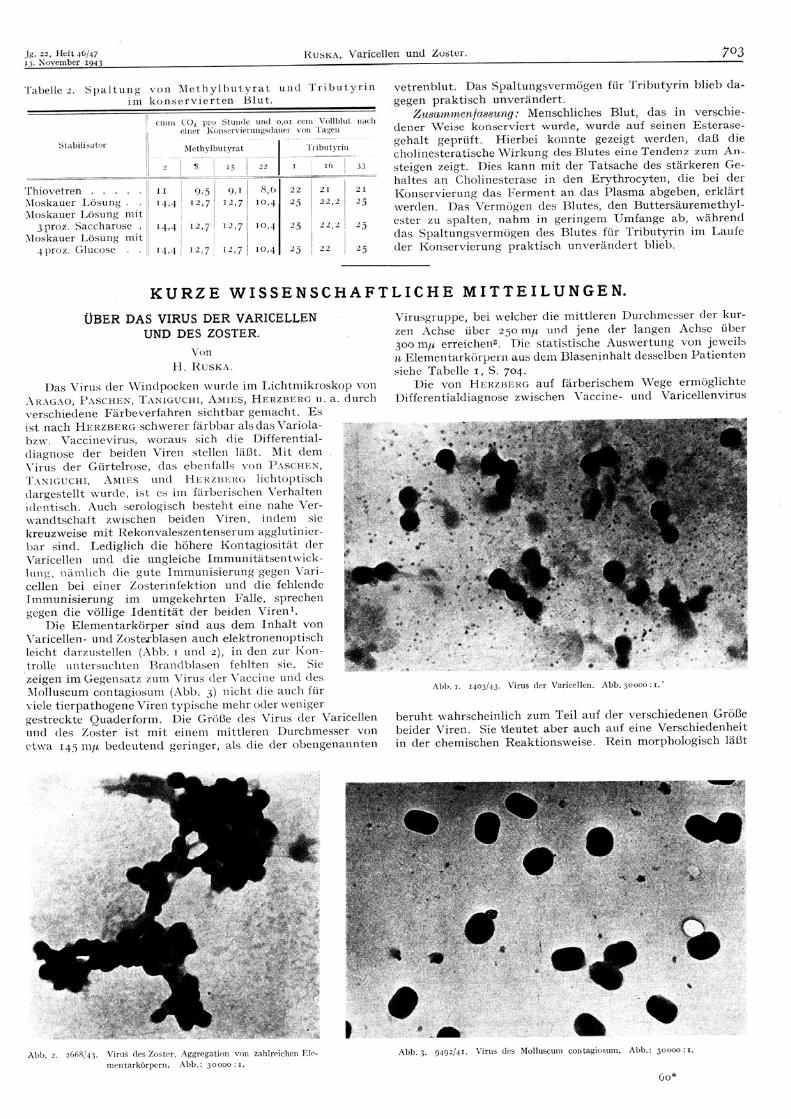


Anlage 2/S. 1
Anlage 2 Auszug aus dem Vorlesungsverzeichnis der Humboldt-Universität Sommersemester 2004 40 079 Medizinische Mikrobiologie/Virologie (1. klin. Sem. HM) VL Di.
Mi. 12:00-13:30 12:00-13:30
wöch. wöch.
DOR 96 DOR 96
D. H. Krüger
40 114 Mikrobiologie/Virologie (2. klin. Sem. HM) KU n. V. n. V. 14tgl. DOR 94 S. Prösch u. a.
40 115 Mikrobiologie/Virologie (4. Stdj. ZM) VL Do. 7:30-9:00 wöch. DOR 94 D. H. Krüger
40 116 Allgemeine und molekulare Virologie (DB) VL n. V. n. V. n. V. L-ANA, 1 M. Reuter u. a.
40 117 Mikrobiologie/Virologie (MP/PP; BF) VL n. V. n. V. mtl. ZI 5 H. Heider u. a.
Probleme der medizinischen und molekularen Virologie (4. Stdj. DB) SE Mi. 16:00-18:00 14tgl. S-VI D. H. Krüger u. a. Reformstudiengang Medizin 40 210 Schwangerschaft/Geburt/Neugeborenes (6. Sem.) SE Fr. 9:15-15:45 S 20-TÄF G. Schönrich
40 211 Säugling/Kleinkind (6. Sem.) SE Di. 9:15-15:45 S 20-TÄF Fr. 9:15-15:45
A. Stein, D. H. Krüger
Wintersemester 2004/2005 Hygiene, Mikrobiologie, Virologie (5., 6. Sem., HM)
CCM/CVK, CBF CUB 052 VL s. Aush. s. Aush. wöch. s. Aush. D. H. KrügerCUB 053 PR s. Aush. s. Aush. wöch. s. Aush. S. Prösch u. a.
Hygiene, Mikrobiologie, Virologie (6. Sem., HM) CCM/CVK, CBF
CUB 073 VL s. Aush. s. Aush. wöch. s. Aush. D. H. Krüger

Anlage 2/S. 2
Hygiene, Mikrobiologie, Virologie (7. Sem., HM) CCM/CVK, CBF
CUB 096 VL s. Aush. s. Aush. wöch. s. Aush. D. H. KrügerCUB 097 SE s. Aush. s. Aush. wöch. s. Aush. D. H. Krüger u. a.
Hygiene, Mikrobiologie, Virologie (8. Sem., HM) CCM/CVK, CBF
CUB 131 VL s. Aush. s. Aush. wöch. s. Aush. D. H. Krüger
Mikrobiologie/Virologie (3. Stdj. MP/PP-BF) CCM
CUB 215 VL n. V. n. V. mtl. ZI 5 H. Heider u. a.
Mikrobiologie/Virologie (4. Stdj. ZM) CUB 266 VL Do. 07:30-09:00 wöch. DOR 94 D. H. Krüger
Allgemeine und Molekulare Virologie (DB) CUB 373 VL n. V. n. V. n. V. L-ANA, 1 M. Reuter u. a.
Komplexpraktikum „Grundlagen der molekularen Virologie“ KU Dauer: zwei Wochen S-VI H. Meisel u. a.
Probleme der medizinischen und molekularen Virologie (4. Stdj. DB) SE Mi. 16:00-18:00 14tgl. S-VI D. H. Krüger u. a.
Ringvorlesung „Infection Biology“ VL Mo. 18:00-20:00 wöch. MPI D. H. Krüger
G. Schönrich u. a. Reformstudiengang Medizin Entzündung/Abwehr (3. Sem.)
CCM CUB 318 SE Di. 9:15-15:45 S 20-TÄF D. H. Krüger
G. SchönrichS. Prösch
PR Fr. 9:15-15:45 S 20-TÄF G. SchönrichH. Heider, S. Prösch
Haut (5. Sem.) CCM
CUB 322 PR Fr. 9:15-15:45 S 20-TÄF N. Kleinkauf, I. Nindl
Schulkind (7. Sem.) CCM
CUB 335 SE Fr. 9:15-12:45 S 20-TÄF G. Schönrich

Anlage 3

Anlage 4 Leistungsspektrum in der Krankenversorgung
4a Untersuchungsbogen
4b Ergänzende Informationen
4c Anforderungsschein Konsiliarlaboratorium für Hantaviren
4d Anforderungsschein Arbeitsmedizinischer Dienst

Anlage 4a/S. 1
Anlage 4a
Institut für Virologie EINSENDER Direktor: Prof. Dr. med. D. H. Krüger Charité - Universitätsmedizin Berlin Campus Charité Mitte, Schumannstr. 20/21, 10117 Berlin (Campus-interne Adresse: Helmut-Ruska-Haus, Rahel-Hirsch-Weg 3) Telefon (030) 450 525 084, 450 525 160 Fax (030) 450 525 908 Rufbereitschaft nachts und am Wochenende: Diensthabender Arzt über 0173-2852736, MTA über 0173-2852806 Virologische Untersuchungen (für Hinweise zu Diagnostik, Probenentnahme und -transport beachten Sie bitte unser Leistungsverzeichnis, im Inter- bzw. Intranet unter www.charite.de/virologie)
Patientenetikett oder Patientenname: Vorname: Geburtsdatum: Geschlecht weibl. männl. Kostenträger ambulant stationär Kasse privat
Untersuchungsmaterial: Nativblut/Serum Urin Nabelschnurbl. EDTA/Citrat-Blut Stuhl Fruchtwasser Liquor Sputum Nasenspülw. BAL Trachealsekret Rachenspülw. Abstrich: Biopsie: Sonstiges:
Entnahmedatum / -zeit: .................... / .....................
cito Nadelstichverletzung: Opfer Index Schwangerschaft (.............. . Woche)
tel. Absprache mit diensthabendem Arzt der Virologie: mit Fr./Hr.: am / um:
Sicherstellung eines unverzüglichen Probentransportes
Angabe einer Telefonnummer zur Befundmitteilung: Tel: Pieper:
Klinische Angaben (bitte unbedingt ausfüllen) Aktuelle Symptomatik / Verdachtsdiagnose:
Transplantation → Organ:
vor Tx Tx am:
Immunsuppression:
seit:
Innerhalb der letzten 6 Wochen:
Immunglobulingabe Blut- o. Blutproduktgabe
Antivirale Chemotherapie:
........................................ seit: ..................
........................................ seit: ..................
Datum Unterschrift des einsendenden Arztes Telefon-Nr. für Rückfragen Labornummer (event. auch Fax-Nr.)
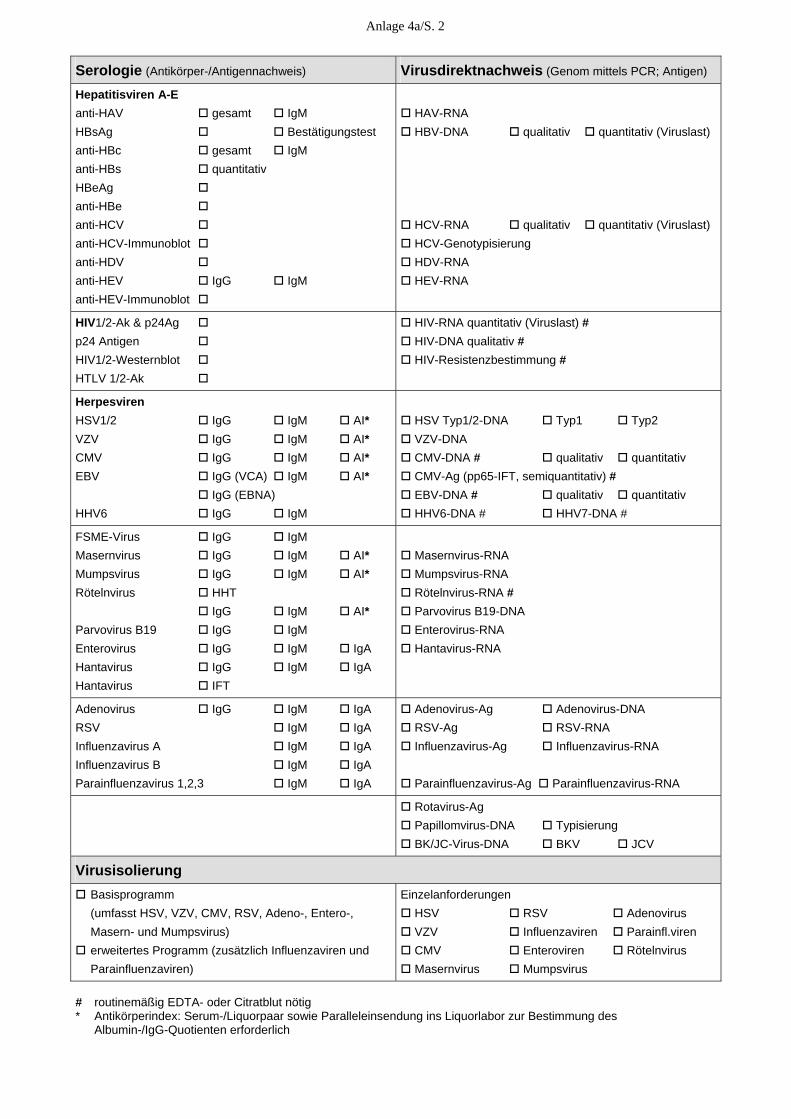
Anlage 4a/S. 2
Serologie (Antikörper-/Antigennachweis) Virusdirektnachweis (Genom mittels PCR; Antigen) Hepatitisviren A-E anti-HAV gesamt IgM HBsAg Bestätigungstest anti-HBc gesamt IgM anti-HBs quantitativ HBeAg anti-HBe anti-HCV anti-HCV-Immunoblot anti-HDV anti-HEV IgG IgM anti-HEV-Immunoblot
HAV-RNA HBV-DNA qualitativ quantitativ (Viruslast)
HCV-RNA qualitativ quantitativ (Viruslast) HCV-Genotypisierung HDV-RNA HEV-RNA
HIV1/2-Ak & p24Ag p24 Antigen HIV1/2-Westernblot HTLV 1/2-Ak
HIV-RNA quantitativ (Viruslast) # HIV-DNA qualitativ # HIV-Resistenzbestimmung #
Herpesviren HSV1/2 IgG IgM AI* VZV IgG IgM AI* CMV IgG IgM AI* EBV IgG (VCA) IgM AI* IgG (EBNA) HHV6 IgG IgM
HSV Typ1/2-DNA Typ1 Typ2 VZV-DNA CMV-DNA # qualitativ quantitativ CMV-Ag (pp65-IFT, semiquantitativ) # EBV-DNA # qualitativ quantitativ HHV6-DNA # HHV7-DNA #
FSME-Virus IgG IgM Masernvirus IgG IgM AI* Mumpsvirus IgG IgM AI* Rötelnvirus HHT IgG IgM AI* Parvovirus B19 IgG IgM Enterovirus IgG IgM IgA Hantavirus IgG IgM IgA Hantavirus IFT
Masernvirus-RNA Mumpsvirus-RNA Rötelnvirus-RNA # Parvovirus B19-DNA Enterovirus-RNA Hantavirus-RNA
Adenovirus IgG IgM IgA RSV IgM IgA Influenzavirus A IgM IgA Influenzavirus B IgM IgA Parainfluenzavirus 1,2,3 IgM IgA
Adenovirus-Ag Adenovirus-DNA RSV-Ag RSV-RNA Influenzavirus-Ag Influenzavirus-RNA
Parainfluenzavirus-Ag Parainfluenzavirus-RNA
Rotavirus-Ag Papillomvirus-DNA Typisierung BK/JC-Virus-DNA BKV JCV
Virusisolierung
Basisprogramm (umfasst HSV, VZV, CMV, RSV, Adeno-, Entero-, Masern- und Mumpsvirus)
erweitertes Programm (zusätzlich Influenzaviren und Parainfluenzaviren)
Einzelanforderungen HSV RSV Adenovirus VZV Influenzaviren Parainfl.viren CMV Enteroviren Rötelnvirus Masernvirus Mumpsvirus
# routinemäßig EDTA- oder Citratblut nötig * Antikörperindex: Serum-/Liquorpaar sowie Paralleleinsendung ins Liquorlabor zur Bestimmung des Albumin-/IgG-Quotienten erforderlich

Anlage 4b/S. 1
Anlage 4b
Institut für Virologie Direktor: Prof. Dr. med. D. H. Krüger Charité - Universitätsmedizin Berlin Campus Charité Mitte, Schumannstr. 20/21, 10117 Berlin (Campus-interne Adresse: Helmut-Ruska-Haus, Rahel-Hirsch-Weg 3) Telefon (030) 450 525 084, 450 525 160 Fax (030) 450 525 908 Rufbereitschaft nachts und am Wochenende: Com-Center über 450 577 044, diensthabender Arzt über 0173-2852736, MTA über 0173-2852806
Ergänzende Informationen zum Anforderungsschein für virologische Untersuchungen Transport: Der Transport des Untersuchungsmaterials ins Labor sollte schnellstmöglich erfolgen. Bei Verzögerung ist das Material bei 4 °C aufzubewahren. Material für die Virusanzucht sowie Citrat- und EDTA-Blut dürfen nicht eingefroren werden! Virustransportmedium für Abstriche, Biopsiematerial etc. kann durch die Virologie bereitgestellt werden. Ist aktuell kein Virustransportmedium zur Hand, können 0,5 ml sterile isotone NaCl-Lösung verwendet werden. Keine Bakterietten mit Stichagar für die Virologie verwenden! Dringende Untersuchungen: Besonders dringende Anforderungen telefonisch anmelden, auf dem Schein als “CITO!” markieren Ansprechpartner und Telefonnummer für die Befundübermittlung angeben Material direkt in die Virologie transportieren Abkürzungen: Abstr Abstrichmaterial (zellreich) Ag Antigen AI virusspezifischer Antikörperindex Ak Antikörper BAL Bronchiallavage BKV BK-Virus (Polyomaviren) CB Citratblut (grüne Monovette) CMV Cytomegalievirus EBV Epstein Barr Virus VCA EBV - Kapsid-Antigen EBNA EBV - nukleäres Antigen EDTA EDTA-Blut (rote Monovette) Fw Fruchtwasser Gewebe Biopsie, Autopsie HAV Hepatitis A Virus HBV Hepatitis B Virus HCV Hepatitis C Virus HDV Hepatitis D Virus HEV Hepatitis E Virus
HHT Hämagglutinationhemmtest HHV6 Humanes Herpesvirus Typ 6 HHV7 Humanes Herpesvirus Typ 7 HSV Herpes simplex Virus IFT Immunfluoreszenztest JCV JC-Virus (Polyomaviren) Km Knochenmark Leb Lebergewebe (Biopsie, Autopsie) Liq Liquor cerebrospinalis NPS Nasopharynxsekret PCR Polymerasekettenreaktion Ra.-Spülw. Rachenspülwasser o. Nasopharynxsekret RSV Respiratorisches Synzytial Virus Ser Serum (weiße Monovette) TS Trachealekret Tx Transplantation VZV Varizella zoster Virus
Informationen zu den für Einzelanforderungen benötigten Materialien: (Routine-Materialien sind fett dargestellt)
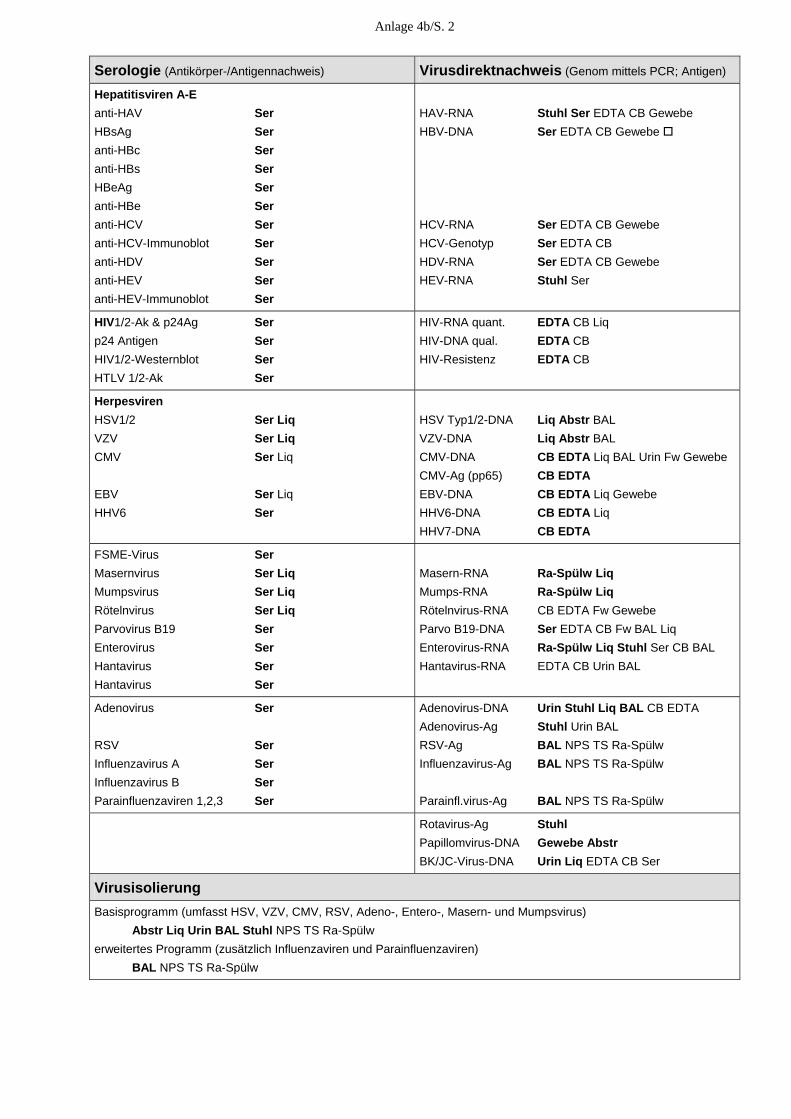
Anlage 4b/S. 2
Serologie (Antikörper-/Antigennachweis) Virusdirektnachweis (Genom mittels PCR; Antigen) Hepatitisviren A-E anti-HAV Ser HBsAg Ser anti-HBc Ser anti-HBs Ser HBeAg Ser anti-HBe Ser anti-HCV Ser anti-HCV-Immunoblot Ser anti-HDV Ser anti-HEV Ser anti-HEV-Immunoblot Ser
HAV-RNA Stuhl Ser EDTA CB Gewebe HBV-DNA Ser EDTA CB Gewebe HCV-RNA Ser EDTA CB Gewebe HCV-Genotyp Ser EDTA CB HDV-RNA Ser EDTA CB Gewebe HEV-RNA Stuhl Ser
HIV1/2-Ak & p24Ag Ser p24 Antigen Ser HIV1/2-Westernblot Ser HTLV 1/2-Ak Ser
HIV-RNA quant. EDTA CB Liq HIV-DNA qual. EDTA CB HIV-Resistenz EDTA CB
Herpesviren HSV1/2 Ser Liq VZV Ser Liq CMV Ser Liq EBV Ser Liq HHV6 Ser
HSV Typ1/2-DNA Liq Abstr BAL VZV-DNA Liq Abstr BAL CMV-DNA CB EDTA Liq BAL Urin Fw Gewebe CMV-Ag (pp65) CB EDTA EBV-DNA CB EDTA Liq Gewebe HHV6-DNA CB EDTA Liq HHV7-DNA CB EDTA
FSME-Virus Ser Masernvirus Ser Liq Mumpsvirus Ser Liq Rötelnvirus Ser Liq Parvovirus B19 Ser Enterovirus Ser Hantavirus Ser Hantavirus Ser
Masern-RNA Ra-Spülw Liq Mumps-RNA Ra-Spülw Liq Rötelnvirus-RNA CB EDTA Fw Gewebe Parvo B19-DNA Ser EDTA CB Fw BAL Liq Enterovirus-RNA Ra-Spülw Liq Stuhl Ser CB BAL Hantavirus-RNA EDTA CB Urin BAL
Adenovirus Ser RSV Ser Influenzavirus A Ser Influenzavirus B Ser Parainfluenzaviren 1,2,3 Ser
Adenovirus-DNA Urin Stuhl Liq BAL CB EDTA Adenovirus-Ag Stuhl Urin BAL RSV-Ag BAL NPS TS Ra-Spülw Influenzavirus-Ag BAL NPS TS Ra-Spülw Parainfl.virus-Ag BAL NPS TS Ra-Spülw
Rotavirus-Ag Stuhl Papillomvirus-DNA Gewebe Abstr BK/JC-Virus-DNA Urin Liq EDTA CB Ser
Virusisolierung
Basisprogramm (umfasst HSV, VZV, CMV, RSV, Adeno-, Entero-, Masern- und Mumpsvirus) Abstr Liq Urin BAL Stuhl NPS TS Ra-Spülw erweitertes Programm (zusätzlich Influenzaviren und Parainfluenzaviren) BAL NPS TS Ra-Spülw

Anlage 4c/S. 1
Anlage 4c
Institut für VIROLOGIE EINSENDER Direktor: Prof. Dr. med. D. H. Krüger Charité - Universitätsmedizin Berlin Campus Charité Mitte, Schumannstr. 20/21, 10117 Berlin (Campus-interne Adresse: Helmut-Ruska-Haus, Rahel-Hirsch-Weg 3)
(030) 450 525 084, 450 525 160 Fax (030) 450 525 908
Konsiliarlaboratorium für Hantavirus-Diagnostik ____________________________________________________________________________________________________________________________________________________________________________________________________________________________________________
Patientenetikett oder Patientenname: Vorname: Geburtsdatum: Geschlecht o weibl. o männl. Entnahmedatum: ..................................
Untersuchungsmaterial: o Nativblut/Serum o BAL o EDTA-/Citratblut (PBL) o Liquor o Urin o Sonstiges: Kostenträger o ambulant o stationär o Kasse o privat (Adresse angeben)
..................................................................................
.................................................................................. Aktuelle Symptomatik / Verdachtsdiagnose: .............................................................................................................................. seit o 1-6 Tagen .............................................................................................................................. o 1-2 Wochen .............................................................................................................................. o 3-4 Wochen o Verdacht auf akute Hantavirus-Infektion: o Kontrolle des Immunstatus (nur IgG Ak) o anamnest. Kontakt zu Nagetieren oder deren Exkrementen o intensiver Naturkontakt in ländlicher Gegend o weitere Erkrankungsfälle im Umfeld bekannt o Fieber o Muskelschmerzen o Kopfschmerzen o Rückenschmerzen
o Nierenfunktionsverschlechterung o Kreatinin im Serum erhöht o Oligurie o Polyurie o dialysepflichtig
o Exanthem o Thrombozytopenie o Hämorrhagische Manifestationen
o Respiratorische Symptomatik o Pneumonie
o Transaminasenerhöhung o Hepatosplenomegalie
Untersuchungsanforderungen: Serologie Screening-ELISA o gesamt IgG (ELISA) o gesamt o HTN o PUU o DOB IgM (ELISA) o gesamt o HTN o PUU o DOB IgA (ELISA) o gesamt o HTN o PUU o DOB Antikörper (IFT) o gesamt o HTN o PUU o DOB Antikörper (Blot) o gesamt o HTN o PUU o DOB Typisierung (cFRNT) o gesamt o HTN o PUU o DOB
Virusdirektnachweis Hantavirus-RNA (PCR) o
Datum Unterschrift des einsendenden Arztes Telefonnr. für Rückfragen Labornummer (event. auch Fax-Nr.) .................... ....................................................................... ...............................................................
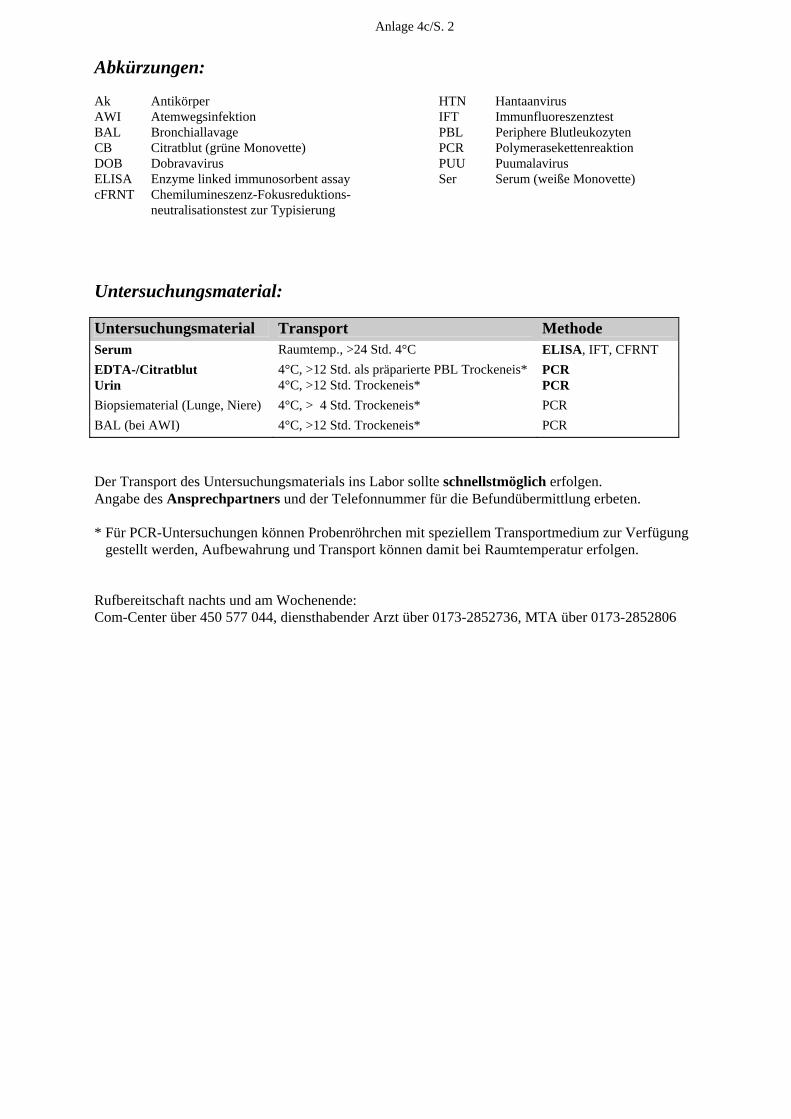
Anlage 4c/S. 2
Abkürzungen: Ak Antikörper AWI Atemwegsinfektion BAL Bronchiallavage CB Citratblut (grüne Monovette) DOB Dobravavirus ELISA Enzyme linked immunosorbent assay cFRNT Chemilumineszenz-Fokusreduktions- neutralisationstest zur Typisierung
HTN Hantaanvirus IFT Immunfluoreszenztest PBL Periphere Blutleukozyten PCR Polymerasekettenreaktion PUU Puumalavirus Ser Serum (weiße Monovette)
Untersuchungsmaterial: Untersuchungsmaterial Transport Methode Serum Raumtemp., >24 Std. 4°C ELISA, IFT, CFRNT EDTA-/Citratblut 4°C, >12 Std. als präparierte PBL Trockeneis* PCR Urin 4°C, >12 Std. Trockeneis* PCR Biopsiematerial (Lunge, Niere) 4°C, > 4 Std. Trockeneis* PCR BAL (bei AWI) 4°C, >12 Std. Trockeneis* PCR
Der Transport des Untersuchungsmaterials ins Labor sollte schnellstmöglich erfolgen. Angabe des Ansprechpartners und der Telefonnummer für die Befundübermittlung erbeten. * Für PCR-Untersuchungen können Probenröhrchen mit speziellem Transportmedium zur Verfügung gestellt werden, Aufbewahrung und Transport können damit bei Raumtemperatur erfolgen. Rufbereitschaft nachts und am Wochenende: Com-Center über 450 577 044, diensthabender Arzt über 0173-2852736, MTA über 0173-2852806

Anlage 4d/S. 1
Anlage 4d
Institut für VIROLOGIE EINSENDER: Direktor: Prof. Dr. med. D. H. Krüger Arbeitsmedizinisches Zentrum Charité - Universitätsmedizin Berlin Campus Mitte Campus Charité Mitte, Schumannstr. 20/21, 10117 Berlin Betriebsärzte MPA-PA -570 723 (Campus-interne Adresse: Helmut-Ruska-Haus, Rahel-Hirsch-Weg 3) D-Arzt MAC-KON -531 000 Telefon (030) 450 525 084, 450 525 160 Fax (030) 450 525 908 Campus Virchow Betriebsärzte WPA-PA -570 700/702 D-Arzt WUC-RET -552 000 Anforderungsschein (Betriebsambulatorium) Name, Vorname bzw. Patientencode: Geburtsdatum: Geschlecht: männl. weibl.
Untersuchungsmaterial: Serum/Venenblut Citrat-/EDTA-Blut sonstiges: ................. Entnahmedatum / -zeit: .................... / ..................... Kostenträger = BA
Untersuchungsanforderungen:
Serologischer Status:
Hepatitis A (anti-HAV gesamt) Hepatitis B vor Impfung (HBsAg, anti-HBs, anti-HBc) → bei HBs-Positivität HBeAg, anti-HBe, HBV-PCR Hepatitis B nach Impfung (anti-HBs) → Anzahl bisheriger HepB-Impfungen: ....................... HBsAg bestätigt positiv (HBV-PCR quantitativ) → bei Negativität HBV-PCR qualitativ Hepatitis C-Screening (anti-HCV) → bei Positivität Immunoblot, HCV-PCR Anti-HCV gesichert positiv (HCV-PCR qualitativ) → bei Positivität HCV-PCR quantitativ HIV (anti-HIV/p24 Ag Kombitest) → bei Positivität (Immunoblot, ggf. p24Ag)
Röteln (Röteln-HHT, ggf. Röteln-IgG ELISA) Cytomegalie (CMV-IgM, -IgG) Masern (Masern-IgG) Parvovirus B19 (Parvo-IgM, -IgG) Mumps (Mumps-IgG) Epstein-Barr-Virus (EBV-IgM, -IgG) Varizella zoster (VZV-IgG) Adenovirus (Adenovirus-IgG)
Nadelstichverletzung:
Verletzte Person Hepatitis B-Impftiter (anti-HBs) cito Indexpatient ist ...................................................
Hepatitis B - Status (HBsAg, anti-HBc, anti-HBs) → bei Positivität weitere Abklärung Hepatitis C - Status (anti-HCV) → bei Positivität weitere Abklärung HIV - Status (anti-HIV und p24 Ag) → bei Positivität weitere Abklärung
Indexpatient HBsAg cito anti-HIV cito Verletzt wurde ...................................................... anti-HCV bei Positivität (Immunoblot, HCV-PCR) Telefonische Mitteilung der „cito“-Befunde erbeten an (Name, Tel.): ..........................................................................
Weitere detaillierte Untersuchungsanforderungen siehe Rückseite .................... ............................................................. .................................................................... Datum Unterschrift des einsendenden Arztes Telefon-Nr. für Rückfragen Labornummer (event. auch Fax-Nr.)
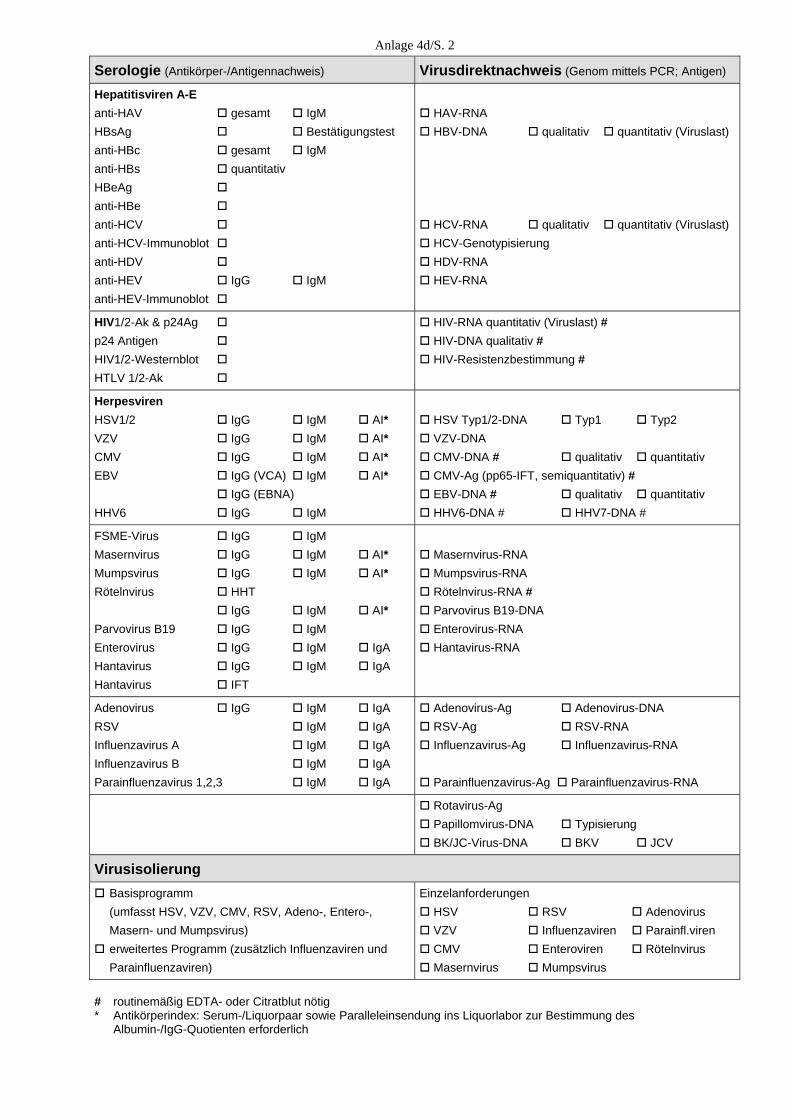
Anlage 4d/S. 2
Serologie (Antikörper-/Antigennachweis) Virusdirektnachweis (Genom mittels PCR; Antigen) Hepatitisviren A-E anti-HAV gesamt IgM HBsAg Bestätigungstest anti-HBc gesamt IgM anti-HBs quantitativ HBeAg anti-HBe anti-HCV anti-HCV-Immunoblot anti-HDV anti-HEV IgG IgM anti-HEV-Immunoblot
HAV-RNA HBV-DNA qualitativ quantitativ (Viruslast)
HCV-RNA qualitativ quantitativ (Viruslast) HCV-Genotypisierung HDV-RNA HEV-RNA
HIV1/2-Ak & p24Ag p24 Antigen HIV1/2-Westernblot HTLV 1/2-Ak
HIV-RNA quantitativ (Viruslast) # HIV-DNA qualitativ # HIV-Resistenzbestimmung #
Herpesviren HSV1/2 IgG IgM AI* VZV IgG IgM AI* CMV IgG IgM AI* EBV IgG (VCA) IgM AI* IgG (EBNA) HHV6 IgG IgM
HSV Typ1/2-DNA Typ1 Typ2 VZV-DNA CMV-DNA # qualitativ quantitativ CMV-Ag (pp65-IFT, semiquantitativ) # EBV-DNA # qualitativ quantitativ HHV6-DNA # HHV7-DNA #
FSME-Virus IgG IgM Masernvirus IgG IgM AI* Mumpsvirus IgG IgM AI* Rötelnvirus HHT IgG IgM AI* Parvovirus B19 IgG IgM Enterovirus IgG IgM IgA Hantavirus IgG IgM IgA Hantavirus IFT
Masernvirus-RNA Mumpsvirus-RNA Rötelnvirus-RNA # Parvovirus B19-DNA Enterovirus-RNA Hantavirus-RNA
Adenovirus IgG IgM IgA RSV IgM IgA Influenzavirus A IgM IgA Influenzavirus B IgM IgA Parainfluenzavirus 1,2,3 IgM IgA
Adenovirus-Ag Adenovirus-DNA RSV-Ag RSV-RNA Influenzavirus-Ag Influenzavirus-RNA
Parainfluenzavirus-Ag Parainfluenzavirus-RNA
Rotavirus-Ag Papillomvirus-DNA Typisierung BK/JC-Virus-DNA BKV JCV
Virusisolierung
Basisprogramm (umfasst HSV, VZV, CMV, RSV, Adeno-, Entero-, Masern- und Mumpsvirus)
erweitertes Programm (zusätzlich Influenzaviren und Parainfluenzaviren)
Einzelanforderungen HSV RSV Adenovirus VZV Influenzaviren Parainfl.viren CMV Enteroviren Rötelnvirus Masernvirus Mumpsvirus
# routinemäßig EDTA- oder Citratblut nötig * Antikörperindex: Serum-/Liquorpaar sowie Paralleleinsendung ins Liquorlabor zur Bestimmung des Albumin-/IgG-Quotienten erforderlich

Anlage 5
„Seminarsprecher des Jahres“ des Institutes für Virologie
1990 W. H. Gerlich, Göttingen/Gießen
1991 H. Gelderblom, Berlin
1992 A. S. von Kekulé, Martinsried
1993 R. Kurth, Langen
1994 U. Heinemann, Berlin-Buch
1995 T. Mettenleiter, Insel Riems
1996 Å. Lundkvist, Stockholm
1997 L. Gürtler, München
1998 A. Kage, Berlin
1999 T. F. Meyer, Tübingen/Berlin
2000 K. Hamprecht, Tübingen
2001 L. Gürtler, Greifswald
2002 J. Sinclair, Cambridge
2003 S. Becker, Marburg
2004 A. Radbruch, Berlin
Seit 1990 wählen die akademischen Mitarbeiter des Institutes den Gastdozenten der öffentlichen Institutskolloquien/Gastvorlesungen, der als „Seminarsprecher des Jahres“ geehrt wird.





























