Embed Size (px)
Citation preview

RUHR-UNIVERSITÄT
FAKULTÄT FÜR
LEHRSTUHL
BOCHUM
CHEMIE UND BIOCHEMIE
PHYSIKALISCHE CHEMIE II
Konfokale Raman-Mikrospektroskopie zur
Untersuchung plasmabehandelter DNA und
einzelner, lebender humaner Spermatozoen
unter physiologieähnlichen Bedingungen
Dissertation
zur Erlangung des Doktorgrades der Naturwissenschaften
Dr. rer. nat.
der Fakultät für Chemie und Biochemie
der Ruhr-Universität Bochum
vorgelegt von
Eugen Edengeiser
Juni 2014


Eidesstattliche Erklärung
Die vorliegende Arbeit wurde im Zeitraum Oktober 2010 bis Juni 2014 unter der
Leitung von Prof. Dr. Martina Havenith am Lehrstuhl für Physikalische Chemie II an
der Fakultät für Chemie und Biochemie der Ruhr-Universität Bochum angefertigt.
Hiermit erkläre ich, dass ich die vorliegende Dissertation selbstständig verfasst,
und keine unzulässigen Hilfsmittel verwendet habe. Zusätzlich erkläre ich, dass mit
dieser Dissertation weder in vorliegender noch in anderer Form ein Promotionsver-
such an einer anderen Fakultät oder Hochschule unternommen wurde.
Eugen Edengeiser, Juni 2014
Betreuende Hochschullehrerin: Prof. Dr. Martina Havenith
Referentin: Prof. Dr. Martina Havenith
Korreferent: Prof. Dr. Christian Herrmann


Danksagung
Die Fertigstellung dieser Arbeit ist für mich ein Rückblick auf den zurückgelegten
Pfad und damit verbunden die Frage nicht wie der Ziel- sondern der Startpunkt
erreicht worden ist. Für immer verbunden mit der vorliegenden Dissertation, die im
Zeitraum von Oktober 2010 bis Juni 2014 entstanden ist, sind viele Menschen, die
mir geholfen haben diesen Weg erfolgreich zu bewältigen:
Zuallererst gebührt Martina Havenith ein „Danke!“ dafür, dass sie mir die Freiheit
gegeben hat vom Grundstudium an verschiedenste Projekte zu verfolgen. Die
Kombination aus Freiheit und Orientierungshilfe zurück zum Wesentlichen wird mir
immer positiv in Erinnerung bleiben.
Christian Herrmann dafür, dass er sich bereit erklärt hat, das Zweitgutachten
der vorliegenden Arbeit zu übernehmen.
Meinen Eltern dafür, dass sie mit Mitte 20 Familie und Freunde zurückgelassen
haben, um mit mir ein neues Abenteuer zu beginnen. Ihr Mut, ihr Optimismus und
Durchhaltevermögen waren und sind eine schier unendliche Quelle der Motivation.
Erik Bründermann, der viel mit meinem oben erwähnten Startpunkt in der Phy-
sikalischen Chemie II zu tun hat (in Form einer Diskussion um einen Bericht zum
Physikalisch-chemischen Grundpraktikum im Sommer 2007). Für das Begleiten
meines wissenschaftlichen Werdegangs mit steter konstruktiver Kritik, Unterstützung
und wertvollen Feedbackloops. Keine Frage, die er nicht mit ein oder zwei Sätzen
beantworten kann. Herzlichen Dank! Auch für die Durchsicht der Arbeit, letztlich bin
ich jedoch allein verantwortlich für Fehler in dieser Dissertation.
Gerhard Schwaab für seine Unterstützung und die wertvollen Diskussionen.
Julia Bandow, Jan Lackmann und Simon Schneider für die ausgezeichnete Ko-
operation mit der mikrobiellen Antibiotikaforschung im Bereich Plasma-behandelter
DNA.
Ebenso Eike Spielberg, Anja Mudring, Van-Son Dang und Anjana Devi für die
gemeinsamen Projekte im Bereich Anorganische Chemie.

Steffen Büning und Simon Ebbinghaus für die geduldige Unterstützung bei den
Fluoreszenz-Messungen.
Jeder der Konrad „Konna“ Meister kennt, kann zweifellos bestätigen, dass er
ein unvergleichlich guter Ratgeber in allen Belangen ist. Danke!
Für die vielfältige Unterstützung der Informations- und Koordinationszentrale
der Physikalischen Chemie II Sylke Kohlpoth, Sabine Weiß und besonders Ulla
Knieper.
Dem technischen Team Dennis Riemenschneider, Christian Fester, Solveig
Förster und Hannelore Sommer.
Neben der Zeit, die wir gemeinsam im Büro und manchmal auch im Labor
verbracht haben, herzlichen Dank an Konna, Jessi und Peppe für die „musika-
lischen“ Abende im Bandkeller, die dazu geführt haben, eine Menge interessanter
Menschen kennenzulernen.
Dem gesamten Lehrstuhl der PC II sowie im Speziellen den Mikroskopikern
um Diedrich Schmidt, Giuseppe Di Florio, Konstantin Kartaschew, Christina
Klinkhammer, Meike Mischo und Fouad Ballout für die tolle Atmosphäre!
Christoph, Jan, Sebastian, Eric, Nils und vor allem Dominic für die jahrelange
Freundschaft und die erheiternden Abende nicht nur im TS!
Letztlich gilt mein besonderer Dank Inga, Nadine, Steffi und Daniel mit denen
ich Seite an Seite das gesamte Studium und mehr meistern durfte.


Veröffentlichungen
Der Autor hat zu folgenden Publikationen beigetragen:
• Effects of the Effluent of a Microscale Atmospheric Pressure Plasma Jet
Operated with He/O2 Gas on Bovine Serum Albumin.
J.-W. Lackmann, E. Edengeiser, S. Schneider, J. Benedikt, M. Havenith, J.E.
Bandow. (Accepted in Plasma Medicine)
• (C4C1py)[Cu(SCN)2]: Coordination Polymer and Liquid.
E.T. Spielberg, E. Edengeiser, B. Mallick, M. Havenith, A.-V. Mudring. Che-
mistry - A European Journal 2014, 20, 5338-5345.
(DOI 10.1002/chem.201302777)
• Photons and particles emitted from cold atmospheric-pressure plasma inacti-
vate bacteria and biomolecules independently and synergistically.
J.-W. Lackmann, S. Schneider, E. Edengeiser, F. Jarzina, S. Brinckmann, E.
Steinborn, M. Havenith, J. Benedikt, J.E. Bandow. Journal of the Royal Society
Interface 2013, 10, 20130591. (DOI 10.1098/rsif.2013.0591)
• Electrical and optical properties of TiO2 thin films prepared by plasma-en-
hanced atomic layer deposition.
V.-S. Dang, H. Parala, J.H. Kim, K. Xu, N.B. Srinivasan, E. Edengeiser, M.
Havenith, A.D. Wieck, T. de los Arcos, R.A. Fischer, A. Devi. physica status
solidi (a), 2013, 2, 416-424. (DOI 10.1002/pssa.201330115)
Konferenzbeiträge (als Erstautor)
• Synergetic effects found in cold atmospheric plasma treatment of biomacro-
molecules explored with Raman spectroscopy.
E. Edengeiser, J.-W. Lackmann, S. Schneider, F. Jarzina, S. Brinckmann, E.

Steinborn, E. Bründermann, J. Benedikt, J.E. Bandow, M. Havenith. [Poster]
Bunsentagung, Karlsruhe, 09.05. - 11.05.2013
• Chemical Micro- and Nanomicroscopy Applications for ANKA-IR2 Nanoscope.
E. Edengeiser, M. Mischo, E. Bründermann, D.A. Schmidt, J. Steinmann, B.
Gasharova, Y.-L. Mathis, D. Moss, M. Havenith. [Poster]
ANKA/KNMF Users’ Meeting, Karlsruhe, 10.10. - 11.10.2012 - 1. Preis - Best
Poster
Konferenz-Veröffentlichungen
• The ANKA-IR2 nanoscope and micro- and nanospectroscopy applications.
E. Bründermann, D.A. Schmidt, B. Gasharova, Y.-L. Mathis, D. Moss, J. Stein-
mann, E. Edengeiser, M. Mischo, M. Havenith.
Proceedings of: 38th International Conference on Infrared, Millimeter, and Tera-
hertz Waves (IRMMW-THz), 2013 (DOI 10.1109/IRMMW-THz.2013.6665734)


Einleitung
Die vorliegende Dissertation behandelt konfokale Raman-Spektroskopie und -Mikro-
spektroskopie als neuartiges analytisches Werkzeug, um zwei physikochemische
Fragestellungen anhand biologischer Proben zu adressieren:
• Wie wird Desoxyribonukleinsäure durch Behandlung mit einer nicht-thermi-
schen, unter Atmosphärendruck arbeitenden Plasma-Quelle inaktiviert?
• Können einzelne, lebende humane Spermatozoen chemisch evaluiert werden
auf eine nicht-invasive Art und Weise unter physiologieähnlichen Bedingun-
gen?
Diese Arbeit wurde zur Beantwortung dieser Fragestellungen in 5 Kapitel unterteilt.
• Kapitel 1 liefert eine theoretische Betrachtung der Grundlagen der hier ausge-
arbeiteten Experimente.
• Kapitel 2 dient der Beschreibung der in dieser Arbeit verwendeten Materialien
und Methoden, insbesondere des Aufbaus des in dieser Arbeit hauptsächlich
verwendeten konfokalen Raman-Mikrospektroskops.
• Kapitel 3 befasst sich mit der Fragestellung, wie DNA durch Plasmabehandlung
inaktiviert werden kann.
• Kapitel 4 betrachtet die konfokale Raman-Mikrospektroskopie immobilisierter,
lebender Spermatozoen unter physiologieähnlichen Bedingungen.
• Kapitel 5 soll anhand zweier Anwendungsbeispiele im Bereich der anorgani-
schen Chemie die Interdisziplinarität der Raman-Spektroskopie aufzeigen.


Zusammenfassung
Nicht-thermische Atmosphärendruck-Plasmen werden im Bereich Wundheilung
evaluiert und gewinnen an Bedeutung in Desinfizierungsprozessen speziell in Anbe-
tracht multiresistenter Bakterien. Obwohl das Potential für technische Anwendungen
offensichtlich scheint, ist viel weniger über die zugrundeliegenden Mechanismen der
Inaktivierung von Bakterien und der Wundheilung bekannt. In dieser Arbeit wurde der
Effekt von reaktiven Partikeln und UV Photonen untersucht, die beide Bestandteile
von Plasma darstellen, um den Beitrag der einzelnen Komponente zu spezifizieren.
Unter Verwendung der X-Jet Technologie wurden Plasma-generierte reaktive Partikel
und Photonen, entstehend in der Plasma-Quelle, voneinander separiert, was eine
Untersuchung der einzelnen Effekte auf DNA Oligomere auf molekularer Ebene mög-
lich macht. Mit nicht-invasiver Raman-Spektroskopie konnte gezeigt werden, dass
Plasma-generierte Partikel und emittierte Photonen unterschiedliche Modifikationen
in DNA-Einzel- und -Doppelsträngen einführt. Überraschenderweise führte eine
Behandlung mit einer Kombination aus Partikeln und Photonen nicht ausschließlich
zu kumulativen Effekten sondern zu synergistischen Effekten beider Komponenten.
Profilometrie in Kombination mit Raman-Spektroskopie behandelter DNA Doppel-
stränge bestätigt, dass Ätzen allein nicht den Hauptbeitrag der beobachteten DNA
Schäden in vitro liefert.
Konfokale Raman-Mikrospektroskopie wurde zum ersten mal verwendet, um che-
mische, farbkodierte Bilder einzelner, lebender und immobilisierter menschlicher
Spermatozoen unter physiologieähnlichen Bedingungen zu erstellen. Dafür wurde
eine neue Kombination von Spermienaufreinigung, Messsubstrat und Substratbe-
schichtung entwickelt. Die Auswirkung der Exposition von Spermatozoen durch
Laserstrahlung während der Durchführung von Raman-Messungen mit unterschied-
licher Laserleistung wurde untersucht. Fluoreszenz-Mikroskopie zeigte, dass sich
30 mW Laserleistung für 160 s als schädlich für die Zellmembran der untersuchten
Spermien erwiesen. Eine Bestrahlung mit einer Leistung von 15 mW über die selbe

Zeit resultierte in keinen beobachtbaren Schäden an der Zellmembran, was die
untersuchten Zellen chemisch nicht unterscheidbar macht von nicht bestrahlten
Zellen. Mehrere einzelne Spermatozoen wurden vollständig automatisiert chemisch
untersucht. Die farbkodierten Raman-Bilder wurden in Echtzeit mit einer Messzeit
von ca. 1 Zelle pro 60 s erstellt. Als Sicherheitsfaktor wurde die Mess- und damit
Laserbestrahlungszeit während der automatisierten Messungen um 60% verringert
verglichen mit den Parametern, die sich als nicht schädlich herausstellten. Dieser
neuartige Ansatz zeigt, dass konfokale Raman-Mikrospektroskopie in der Lage ist,
chemische Informationen einzelner und lebender menschlicher Spermatozoen zu
sammeln. Mit den besonders deutlich separierten Zellkompartimenten stellen Sper-
matozoen ein besonderes Modellsystem für konfokale Raman-Mikrospektroskopie
dar, die allgemein gut zur Untersuchung vieler Zelltypen geeignet ist. Dies hebt das
Potential der Technik im Bereich der künstlichen Befruchtung hervor.


Abkürzungsverzeichnis
µAPPJ . . . . . . . . . . microscale atmospheric-pressure plasma jet
(C4C1py) . . . . . . . 1-Butyl-4-methyl-pyridinium
a. dest. . . . . . . . . . aqua destillata
ACES . . . . . . . . . . . N-(2-Acetamido)-2-aminoethansulfonsäure
ART . . . . . . . . . . . . Assisted reproductive technology
bb . . . . . . . . . . . . . . backbone
CAP . . . . . . . . . . . . Cold atmospheric-pressure plasma
CCD . . . . . . . . . . . . Charge-coupled device
cNB . . . . . . . . . . . . coupled nucleobases
CRM . . . . . . . . . . . . Confocal Raman microspectroscopy
FRET . . . . . . . . . . . Fluoreszenz-Resonanz-Energie-Transfer
Laser . . . . . . . . . . . Light amplification by stimulated emission of radiation
PDB . . . . . . . . . . . . Protein database
PI . . . . . . . . . . . . . . . Phosphidiumiodid
PMT . . . . . . . . . . . . Photo multiplier tube
RF . . . . . . . . . . . . . . Radiofrequenz
RMS . . . . . . . . . . . . Root mean square
S/N . . . . . . . . . . . . . Signal to noise
sccm . . . . . . . . . . . Standard-Kubikzentimeter pro Minute
slm . . . . . . . . . . . . . Standard-Liter pro Minute
STED . . . . . . . . . . . Stimulated emission depletion
STORM . . . . . . . . . Stochastic optical reconstruction microscopy
WHO . . . . . . . . . . . World Health Organization


Inhaltsverzeichnis
Abkürzungsverzeichnis 16
1 Theoretische Grundlagen & Einführung 1
1.1 Raman-Spektroskopie . . . . . . . . . . . . . . . . . . . . . . . . . . 1
1.2 Optische und konfokale Mikroskopie . . . . . . . . . . . . . . . . . . 8
1.3 Konfokale Raman-Mikrospektroskopie . . . . . . . . . . . . . . . . . 10
1.4 Fluoreszenz-Mikroskopie . . . . . . . . . . . . . . . . . . . . . . . . 10
1.5 Desoxyribonukleinsäure . . . . . . . . . . . . . . . . . . . . . . . . . 13
2 Material und Methoden 16
2.1 Konfokale Raman-Mikrospektroskopie . . . . . . . . . . . . . . . . . 16
2.1.1 Konfiguration für Messungen mit 488 nm Anregungswellenlänge 17
2.1.2 Konfiguration für Messungen mit 532 nm Anregungswellenlänge 18
2.1.3 Konfiguration für Messungen bei 785 nm Anregungswellenlänge 18
2.2 Plasma-Quelle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
2.3 Proben . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
2.3.1 Synthetische DNA Homo-Oligomere . . . . . . . . . . . . . . 18
2.3.2 Ätzraten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
2.3.3 Lebende humane Spermatozoen . . . . . . . . . . . . . . . . 19
2.3.4 Beschichtung von CaF2-Substrat mit Concanavalin A Typ IV . 19
2.3.5 Percoll-Dichtegradienten-Zentrifugation . . . . . . . . . . . . 19
2.3.6 Automatisierte konfokale Raman-Mikrospektroskopie . . . . . 20
2.3.7 Vorbehandlung der aufgenommen Datensätze . . . . . . . . 20
2.3.8 LIVE/DEAD Sperm Viability Kit zur Fluoreszenz-Markierung
von Spermatozoen . . . . . . . . . . . . . . . . . . . . . . . 20
2.4 Fluoreszenz-Mikroskopie . . . . . . . . . . . . . . . . . . . . . . . . 21
i

3 Effekte der Behandlung von DNA mit einem nicht-thermischen Atmos-
phärendruck-Plasma 23
3.1 Einführung und Motivation . . . . . . . . . . . . . . . . . . . . . . . 23
3.2 Ergebnisse . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
3.3 Diskussion und Ausblick . . . . . . . . . . . . . . . . . . . . . . . . 35
4 Konfokale Raman-Mikrospektroskopie immobilisierter, lebender Sper-
matozoen unter physiologieähnlichen Bedingungen 38
4.1 Einführung und Motivation . . . . . . . . . . . . . . . . . . . . . . . 38
4.2 Ergebnisse . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
4.3 Diskussion und Ausblick . . . . . . . . . . . . . . . . . . . . . . . . 54
5 Anwendungen konfokaler Raman-Spektroskopie 58
5.1 Konfokale Raman-Spektroskopie von TiO2-
Dünnschichten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 59
5.2 Temperaturabhängige Raman-Spektroskopie von
(C4C1py)[Cu(SCN)2] . . . . . . . . . . . . . . . . . . . . . . . . . . 61
6 Abbildungs- & Tabellenverzeichnis 66
7 Literaturverzeichnis 76
ii


The important thing in science
is not so much to obtain new facts
as to discover new ways
of thinking about them
Sir William Lawrence Bragg


1Theoretische Grundlagen & Einführung
Nicht-invasive und nicht-destruktive Methoden zur Charakterisierung kleiner Mole-
küle, von Zellen bis hin zu Geweben werden zunehmend sowohl in akademischen
als auch industriellen Umgebungen geschätzt. Schwingungsspektroskopische Tech-
niken zu denen Raman-Spektroskopie gezählt wird, liefern einen Ansatz wie die
oben formulierten Ziele mit einer hohen Sensitivität erreicht werden können. Die-
ses Kapitel dient der Einführung in die Raman-Spektroskopie und beginnt folglich
mit einer theoretischen Beschreibung der zugrundeliegenden Prinzipien, die den
Abhandlungen in [1–3] folgt. Dabei wird sowohl auf das klassische als auch auf
das quantenmechanische Bild eingegangen. Anschließend wird beschrieben wie
diese Technik mit Hilfe der Kombination mit klassischer konfokaler Lichtmikroskopie
zu konfokaler „Raman-Mikrospektroskopie“ erweitert wurde. Als komplementäre
Messtechnik wird auf Fluoreszenz-Mikroskopie eingegangen.
1.1 Raman-Spektroskopie
Der Beobachtung Arthur Comptons 1923 folgend, dass die Energie von Röntgen-
strahlen, die an Elektronen gestreut worden sind, abnehmen kann (Compton-Effekt),
postulierten Raman und Krishnan 1928 einen ähnlichen Vorgang für sichtbares Licht,
welches an Materie gestreut wird [4, 5].
Der Raman-Effekt wird beschrieben als Wechselwirkung elektromagnetischer
1
Raman-Spektroskopie 1
Strahlung mit einer Probe, in der entweder ein Schwingungsquantum angeregt
(Stokes-Streuung) oder ausgelöscht wird (Anti-Stokes-Streuung). Der Großteil der
Photonen wird elastisch gestreut (Rayleigh-Streuung). Folglich besitzt die Streu-
strahlung die gleiche Energie wie die Anregungsstrahlung. Wird ein Photon auf
inelastische Art und Weise gestreut, äußert sich dies darin, dass sich die Energie
des gestreuten Photons von der des Anregungsphotons unterscheidet. Inelastische
Stöße zeichnen sich durch unterschiedliche Anfangs- und Endzustände aus und sind
also immer mit einer Änderung der Energie verbunden. Ist der Anfangszustand der
Schwingungsgrundzustand und der Endzustand ein angeregtes Schwingungsniveau,
wird dies als Stokes-Streuung bezeichnet. Ist der Anfangszustand energetisch höher
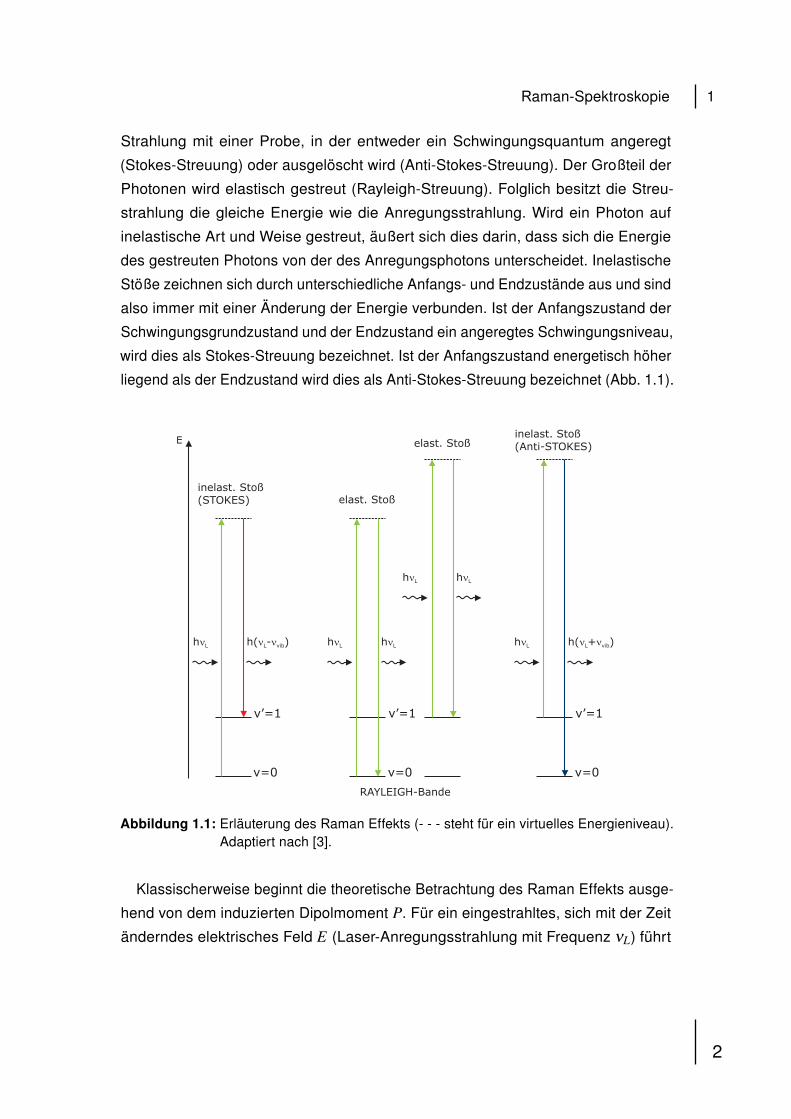
liegend als der Endzustand wird dies als Anti-Stokes-Streuung bezeichnet (Abb. 1.1).
v=0
v’=1
v=0
v’=1
v=0
v’=1
h�L h( - )� �L vib h�L h�Lh�L
h�L h�L
h( + )� �L vib
inelast. Stoß
(STOKES) elast. Stoß
inelast. Stoß
(Anti-STOKES)
RAYLEIGH-Bande
elast. StoßE
Abbildung 1.1: Erläuterung des Raman Effekts (- - - steht für ein virtuelles Energieniveau).Adaptiert nach [3].
Klassischerweise beginnt die theoretische Betrachtung des Raman Effekts ausge-
hend von dem induzierten Dipolmoment P. Für ein eingestrahltes, sich mit der Zeit
änderndes elektrisches Feld E (Laser-Anregungsstrahlung mit Frequenz νL) führt
2

Raman-Spektroskopie 1
die Polarisierbarkeit α zum induzierten Dipolmoment [3]:
P = α ·E = α ·E0 cos(2πνLt). (1.1)
Die molekülabhängige Größe α gibt an, wie leicht die Elektronenwolke des Moleküls
verformt werden kann. Schwingt das betrachtete Molekül mit einer Frequenz νvib
folgt daraus eine Modulierung der Polarisierbarkeit α (Gleichung 1.3) entlang der
Normalkoordinate q (Gleichung 1.2) der Schwingung (beides aus [3]).
q = q0 · cos(2πνvibt), (1.2)
α ≈ α0 +
(∂α
∂q
)q = α0 +α1q. (1.3)
Gleichung 1.3 gilt dabei für kleine interatomare Auslenkungen nach Taylorreihen
Entwicklung und α0 gibt die mittlere Polarisierbarkeit in der Gleichgewichtslage an.
Gleichung 1.3 in Gleichung 1.1 eingesetzt ergibt [3]:
P = (α0 +α1q0 cos(2πνvibt))E0 cos(2πνLt) (1.4)
= α0E0 cos(2πνLt)+α1q0E0 cos(2πνvibt)cos(2πνLt). (1.5)
Gleichung 1.5 lässt sich mit Hilfe trigonometrischer Additionstheoreme vereinfachen
zu [3]:
P = α0E0 cos(2πνLt)+12
α1q0E0[cos(2π(νL +νvib)t)+ cos(2π(νL−νvib)t)]. (1.6)
Daraus folgt, dass das gestreute Licht sowohl Anteile mit der Frequenz νL der Laser
-Anregungsstrahlung (erster Term, Rayleigh) aufweist, als auch die Frequenzen
νL +νvib (zweiter Term, Anti-Stokes) und νL−νvib (dritter Term, Stokes) detektiert
werden können. Des Weiteren muss als Auswahlregel gelten [3]:(∂α
∂q
)q 6= 0. (1.7)
Die Polarisierbarkeit des Moleküls muss sich für ein Raman-Signal im Verlauf der
Schwingung ändern. In einer quantenmechanischen Betrachtung wird das Matrix
Element zwischen Anfangszustand i (initial) und Endzustand f (final) untersucht. Die
Quantenübergänge sind wiederum von der Polarisation P aus einem eingestrahlten,
3

Raman-Spektroskopie 1
elektrischen Feld E abhängig. Wir schreiben [1]:
Pf i = 〈 f |P|i〉= 〈 f |ε0χP|i〉. (1.8)
Die Polarisation α wird in dieser Betrachtung als Suszeptibilität (Fähigkeit zur elektri-
schen Polarisierung) im Feld E behandelt. Unter der Annahme, dass die Wellenlänge
des eingestrahlten Lichtes viel größer ist, als die interatomaren Abstände wird das
elektrische Feld E als konstant betrachtet und Gleichung 1.8 kann geschrieben
werden als [1]
[χmn] f i = 〈 f |χmn|i〉, (1.9)
mit χmn als Übergangs-Suszeptibilität, die probenabhängig ist.
Für die Wellenfunktionen, die das vibronische System (Vibrations- und elektroni-
sche Energie) beschreiben, wird die adiabatische Annäherung verwendet, in der
die Wellenfunktion ψ(r,R) aus einer Wellenfunktion für die Elektronen mit den Ko-
ordinaten r und einer Wellenfunktion für die Atome mit den Koordinaten R besteht
[1]:
ψ(r,R) = ζ (r,R)ρ(R). (1.10)
Für [χmn] f i ergibt sich [1]:
[χmn] f i =∫
ρ∗f (R)ζ
∗f (r,R)χmnζi(r,R)ρi(R)drdR. (1.11)
Unter Betrachtung der Integration über die Elektronen-Koordinaten r und der
Annahme, dass die elektronischen Zustände des Anfangs- und Endzustands gleich
sind, ergibt sich der elektronische Teil χmn(R) der Übergangs-Suszeptibilität. Die
Einführung von Normalkoordinaten qk und Betrachtung nur des linearen Terms
resultiert in [1]:
[χmn] f i = (χmn)0〈ρ f k|ρik〉+∑k(∂ χmn
∂qk)0〈ρ f k|qk|ρik〉, (1.12)
mit der Klammerschreibweise als gesamte Schwingungs-Wellenfunktionen aus dem
Integral aus Gleichung 1.11. Die Erwartungswerte sind [1]:
〈ρ f k|ρik〉=
0 für ρ f k 6= ρik
1 für ρ f k = ρik
(1.13)
4

Raman-Spektroskopie 1
und [1]
〈ρ f k|qk|ρik〉=
0 für ρ f k = ρik√
ρik +1√
h2qk
für ρ f k = ρik +1√
ρik
√h
2qkfür ρ f k = ρik−1
(1.14)
Wegen der Orthogonalität der Wellenfunktionen können alle Erwartungswerte aus
Gleichung 1.12 in der Form von Gleichung 1.13 und Gleichung 1.14 ausgedrückt
werden. Das bedeutet, dass sich im ersten Term in Gleichung 1.12 der Quantenzu-
stand für ρ f k = ρik nicht geändert hat (Rayleigh-Streuung). Der zweite Term steht
für Raman-Streuung und ist ungleich 0 wenn ρik = ρik±1 und führt zur Übergangs-
Suszeptibilität in der Form [1]:
[χmn]ρik+1,ρik =√
ρik +1
√h
2qk
(∂ χmn
∂qk
)0
(1.15)
und [1]
[χmn]ρik−1,ρik =√
ρik
√h
2qk
(∂ χmn
∂qk
)0. (1.16)
Aus Gleichung 1.15 und Gleichung 1.16 werden die Stokes- beziehungsweise Anti-
Stokes-Streuprozesse ersichtlich. Ähnlich zu Gleichung 1.7 ergibt sich nun eine
Auswahlregel [1] (∂ χmn
∂qk
)6= 0 (1.17)
In einem Raman-Spektrum wird die Intensität des gestreuten Lichts auf der Y-Achse
gegen den Energieunterschied zwischen Anregungslicht und gestreutem Licht auf
der X-Achse aufgetragen. Die Lage einer Raman-Bande wird üblicherweise in Wellen-
zahlen (1 cm−1) angegeben. Sie steht folgendermaßen mit dem Energieunterschied
relativ zum Anregungslicht in Zusammenhang [6]:
ν =1λ0− 1
λg(1.18)
λ0 und λg Wellenlänge des Anregungslichts bzw. des gestreuten Lichts in cm.
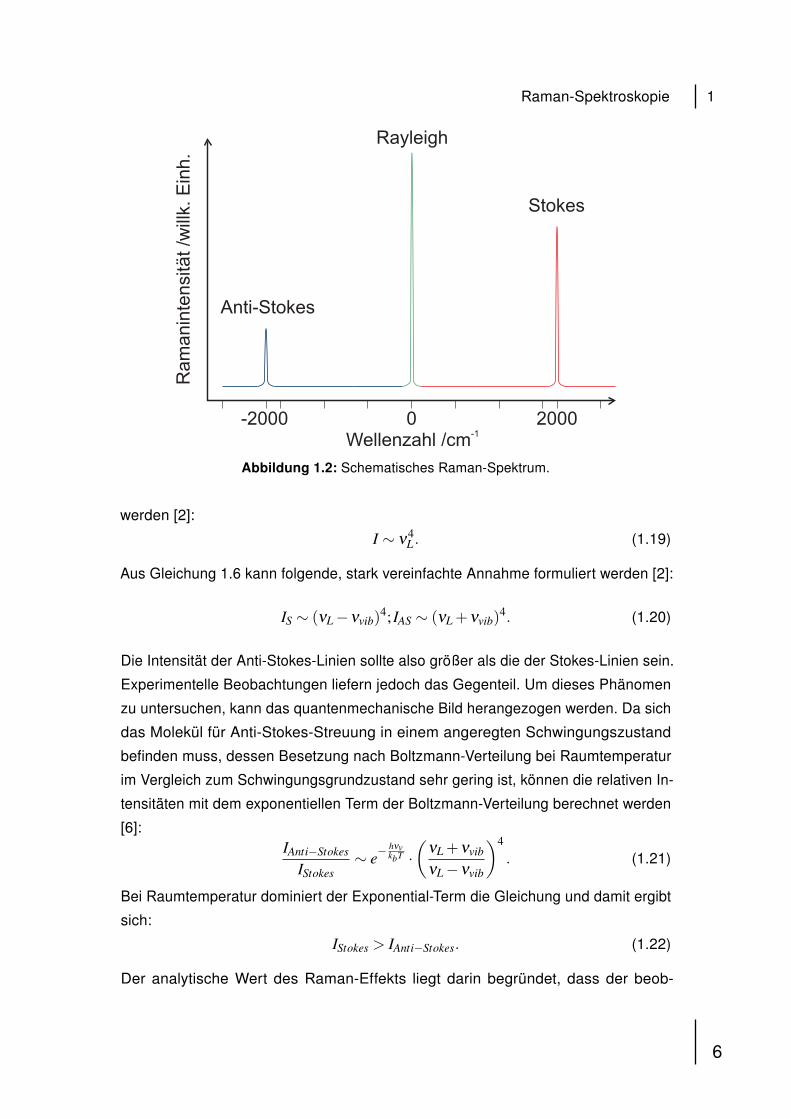
Abbildung 1.2 zeigt ein schematisches Raman-Spektrum. Charakteristisch ist,
dass es symmetrisch zur Rayleigh-Linie aufgebaut ist und dass die Anti-Stokes-Linie
eine geringere Intensität aufweist als die Stokes-Linie. Aus der der Untersuchung der
Strahlenstärke eines Dipols im Fernfeld kann die Intensität I des gestreuten Lichts
als proportional zur vierten Potenz der Frequenz des Anregungslichts betrachtet
5
Raman-Spektroskopie 1
0-2000 2000
Rayleigh
Stokes
Anti-Stokes
Ram
anin
tensität /w
illk. E
inh.
Wellenzahl /cm-1
Abbildung 1.2: Schematisches Raman-Spektrum.
werden [2]:
I ∼ ν4L . (1.19)
Aus Gleichung 1.6 kann folgende, stark vereinfachte Annahme formuliert werden [2]:
IS ∼ (νL−νvib)4; IAS ∼ (νL +νvib)
4. (1.20)
Die Intensität der Anti-Stokes-Linien sollte also größer als die der Stokes-Linien sein.
Experimentelle Beobachtungen liefern jedoch das Gegenteil. Um dieses Phänomen
zu untersuchen, kann das quantenmechanische Bild herangezogen werden. Da sich
das Molekül für Anti-Stokes-Streuung in einem angeregten Schwingungszustand
befinden muss, dessen Besetzung nach Boltzmann-Verteilung bei Raumtemperatur
im Vergleich zum Schwingungsgrundzustand sehr gering ist, können die relativen In-
tensitäten mit dem exponentiellen Term der Boltzmann-Verteilung berechnet werden
[6]:IAnti−Stokes
IStokes∼ e−
hνvkbT ·
(νL +νvib
νL−νvib
)4
. (1.21)
Bei Raumtemperatur dominiert der Exponential-Term die Gleichung und damit ergibt
sich:
IStokes > IAnti−Stokes. (1.22)
Der analytische Wert des Raman-Effekts liegt darin begründet, dass der beob-
6

Raman-Spektroskopie 1
achtete Energieunterschied des Anregungsphotons und des inelastisch gestreuten
Photons für die im Streuprozess beteiligten Atome und chemischen Strukturen cha-
rakteristisch ist. Raman-Spektroskopie stellt daher eine komplementäre Technik zu
der Infrarot-Spektroskopie dar, bei der sich das elektrische Dipolmoment bei der
Schwingung ändern muss. So ist mit der Raman-Spektroskopie zum Beispiel die
symmetrische Streckschwingung von CO2 beobachtbar.
Aus diesen Überlegungen ergibt sich, dass der Raman-Effekt nicht-invasiv genutzt
werden kann. Proben müssen in der Regel nicht vorbehandelt werden und können
ohne Weiteres als Feststoff oder in Lösung chemisch evaluiert werden, da Wasser
nur intensitätsschwache Banden zeigt, ganz im Gegensatz zur starken Absorption
von Wasser in der Infrarot-Spektroskopie. Tabelle 1.1 gibt an, welche Eigenschaften
von Stoffen in festem Aggregatzustand aber auch in Lösung über die Analyse der
charakteristischen Raman-Banden untersucht werden können:
Tabelle 1.1: Auswahl von mit Raman-Spektroskopie messbaren Eigenschaften.
Eigenschaften Ramanbande Literatur
Identifikation Frequenz [7–9]Orientierung Polarisitonsabhängigkeit [10–12]Kristallinität Breite [13–15]
Menge Intensität [16–18]
Die Hauptverwendung der Raman-Spektroskopie liegt in der Analyse der „chemi-
schen Fingerabdrücke“, also der charakteristischen Bandenmuster in den Raman-
Spektren mit denen sich chemische Strukturen eindeutig nachweisen lassen. Die
Orientierung von Fasern oder anderen orientierungsabhängigen Strukturen lässt
sich durch polarisationsabhängige Banden und Messungen visualisieren. Kristal-
linität verschiedener Stoffe geht in der Regel mit Banden einher, die in der Breite
(zum Beispiel Halbwertsbreite) variieren. Die Intensität bestimmter Banden lässt
sich häufig dazu verwenden, um semi-quantitative Abschätzungen über einen zu
untersuchenden Stoff anzustellen.
7

Optische und konfokale Mikroskopie 1
1.2 Optische und konfokale Mikroskopie
Die zentrale Technik auf der die heute angewandten Mikroskopiearten basieren,
stellt die optische Mikroskopie dar. Mitte des 17. Jahrhunderts berichtete Robert
Hooke in „Micrographia“ [19] seine Beobachtungen, die er bei der Betrachtung von
Korkschnitten in seinem Lichtmikroskop gesammelt hatte. Die „Wände“, die er in
Pflanzen gesehen hatte, führten zur Prägung des Begriffs „Zelle“.
Die Vergrößerung in einem optischen Mikroskop geschieht mit Hilfe von sichtba-
rem Licht, dessen Strahlengang ein Linsensystem passiert. Im Bereich der Zellbiolo-
gie wird klassische Lichtmikroskopie routinemäßig zur Beurteilung der Qualität von
Zellkulturen eingesetzt oder dient der Beobachtung morphologischer Phänomene
zum Beispiel während der Zellteilung unter verschiedenen Bedingungen [20].
Gut zweihundert Jahre später im späten zwanzigsten Jahrhundert änderte sich
die Form der Bildgebung durch die Erfindung des konfokalen Mikroskops (Patent US
3013467 A) fundamental. Eine Probe im klassischen Licht-Mikroskop muss in seiner
Gesamtheit gleichmäßig von Licht bestrahlt und abgebildet werden. Im konfokalen
Mikroskop wird fast ausschließlich nur Licht aus der Brennebene detektiert. Um ein
Gesamtbild zu erhalten, muss ein ausgewählter Bereich Punkt für Punkt abgerastert
werden. Abbildung 1.3 zeigt den prinzipiellen Aufbau eines konfokalen Mikroskops.
Abbildung 1.3: Schematischer Aufbau und Strahlengang eines Konfokalmikroskops. Adap-tiert nach [6].
In einem Konfokal-Mikroskop wird eine Lichtquelle (in der Regel ein Laser) über
8

Optische und konfokale Mikroskopie 1
ein Objektiv in die Probe fokussiert. In der vorliegenden Arbeit wurde ein frequenzver-
doppelter Nd:YAG Festkörperlaser, der Licht einer Wellenlänge von 532 nm emittiert
oder single-frequency Diodenlaser mit einer Wellenlänge von 488 nm beziehungs-
weise 785 nm. Das Streulicht aus der Brennebene der Probe (grüner Strahl) wird
über einen Strahlteiler und ein weiteres Objektiv auf eine Lochblende fokussiert und
auf einen Detektor weitergeleitet. Der Durchmesser der Lochblende wird so gewählt,
dass fast ausschließlich nur Licht aus der Brennebene durch die Lochblende treten
kann und zum Detektor weitergeleitet wird (Konfokalität). Abb. 1.3 zeigt, dass Licht,
das nicht aus der Brennebene stammt (gestrichelte rote Linie) nicht die Lochblende
passiert. So kann ein Bild einer Probe in allen drei Dimensionen erstellt werden und
die laterale Auflösung um einen Faktor bis zu√
2 verbessert werden [21].
Die Auflösung ist begrenzt durch Beugungs-Effekte. Das Rayleigh-Kriterium gibt
dabei den kleinsten Abstand an, bei dem zwei Punktobjekte noch separat detektiert
werden können (beide aus [6]):
∆x =1.22 · f ·λ
D(1.23)
mit Auflösung ∆x, Brennweite f des Objektivs und Durchmesser des Objektivs D.
∆x =0.61 ·λ
NA=
0.61 ·λnsin
(θ
2
) . (1.24)
NA des verwendeten Wassereintauch-Objektivs ist in Wasser 1 (1.0W). Die Auf-
lösung ist also von der Wellenlänge des verwendeten Lichts und der numerischen
Apertur des Objektivs abhängig. Bei Verwendung von Licht einer Wellenlänge von
532 nm als Anregungswellenlänge ergibt sich aus Gleichung 1.24 eine laterale Auflö-
sung des in dieser Arbeit verwendeten Mikroskops von etwa 325 nm. Die Auflösung
in Richtung der optischen Achse beträgt etwa 750 nm.
Zeigt eine Probe Fluoreszenz (also die Emission von Photonen aus einem elektro-
nisch angeregten Zustand des untersuchten Moleküls), führt dies zu einem intensiven
und breiten Hintergrundsignal. Da der klassische Fluoreszenz-Mechanismus um Grö-
ßenordnungen stärker ist als der relativ schwache Raman Effekt, dominiert dieser
üblicherweise die Raman-Banden im Spektrum. Möglichkeiten dem entgegenzuwir-
ken, sind das zu untersuchende Molekül auszubleichen (Abnahme der Fluoreszenz
mit der Zeit der Bestrahlung mit dem Anregungslicht) oder die Fluoreszenz durch
Zugabe eines Quenchers zu unterdrücken.
9

Konfokale Raman-Mikrospektroskopie 1
1.3 Konfokale Raman-Mikrospektroskopie
Obwohl der Raman-Effekt bereits 1928 postuliert worden ist, war die Entdeckung
und Entwicklung zweier kritischer Komponenten notwendig, um Raman-Experimente
durchführen zu können, wie sie heute routinemäßig in industriellen und akade-
mischen Laboren realisiert werden. Die Quelle für die für Raman-Spektroskopie
benötigte monochromatische elektromagnetische Strahlung mit hoher Intensität
stellte lange eine Quecksilberdampflampe dar, bis Theodore Maiman 1960 eine
neuartige Quelle für Licht einer bestimmten Wellenlänge mit hoher Intensität und
Stabilität, den ersten erfolgreichen Laser (light amplification by stimulated emission
of radiation, Laser), erfand [22].
Die zweite kritische Komponente, die ersetzt wurde, befand sich im Detektorteil der
Raman-Instrumente. Die üblich verwendeten Photomultiplier Tubes, PMT konnten
ersetzt werden mit CCD-Kameras (Charge-coupled device) [23]. Im PMT konnte stets
nur eine Bande von Interesse beobachtet werden, die über den Ausgangsschlitz im
Spektrometer eingestellt werden konnte. Als Detektor ersetzte die aus dem Halbleiter
Silizium bestehenden CCD-Kameras den Ausgangsschlitz und machten damit den
Übergang von Einkanal- zu Mehrkanalmessungen möglich. Die Kombination mit
konfokaler Mikroskopie ermöglichte es Raman-Spektren Bildpunkt für Bildpunkt mit
hoher Genauigkeit in allen drei Dimensionen einer Probe zu messen und führte
letztendlich zur Technik der „Konfokalen Raman-Mikrospektroskopie“.
1.4 Fluoreszenz-Mikroskopie
Die am weitesten verbreitete mikroskopische Technik zur Untersuchung zellulärer
Vorgänge ist die Fluoreszenz-Mikroskopie. Einerseits ist es von Vorteil, dass bereits
eine Bandbreite an verschiedensten hochspezifischen Fluoreszenzsonden existiert
[24, 25], andererseits bringt es den Nachteil mit sich, dass die zu untersuchenden
Proben mit genau diesen Fluoreszenzsonden gelabelt werden und an die Moleküle
binden müssen, was die physikochemischen Eigenschaften, gerade von relativ zur
Fluoreszenz-Sonde kleinen Molekülen ändern könnte [26].
Fluoreszenz basiert im Gegensatz zum Raman-Effekt auf der Absorption und
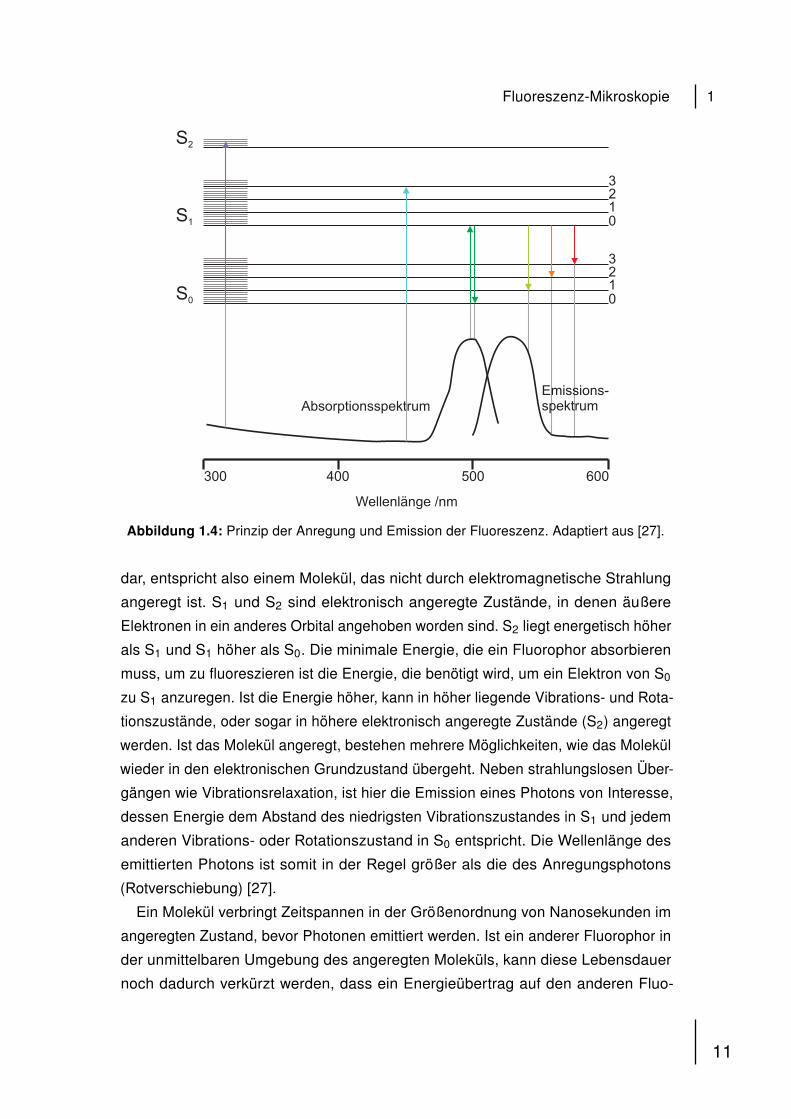
Emission von Photonen. Das Jablonski-Diagramm in Abb. 1.4 zeigt das zugrundelie-
gende Prinzip des Anregungs- und des Emissionsprozesses.
Links sind die Singulettzustände aufgetragen, in denen die Elektronen mit entge-
gengesetztem Spin gepaart vorliegen. S0 stellt den elektronischen Grundzustand
10
Fluoreszenz-Mikroskopie 1
S2
AbsorptionsspektrumEmissions-spektrum
Wellenlänge /nm
300 400 500 600
0123
0123
S1
S0
Abbildung 1.4: Prinzip der Anregung und Emission der Fluoreszenz. Adaptiert aus [27].
dar, entspricht also einem Molekül, das nicht durch elektromagnetische Strahlung
angeregt ist. S1 und S2 sind elektronisch angeregte Zustände, in denen äußere
Elektronen in ein anderes Orbital angehoben worden sind. S2 liegt energetisch höher
als S1 und S1 höher als S0. Die minimale Energie, die ein Fluorophor absorbieren
muss, um zu fluoreszieren ist die Energie, die benötigt wird, um ein Elektron von S0
zu S1 anzuregen. Ist die Energie höher, kann in höher liegende Vibrations- und Rota-
tionszustände, oder sogar in höhere elektronisch angeregte Zustände (S2) angeregt
werden. Ist das Molekül angeregt, bestehen mehrere Möglichkeiten, wie das Molekül
wieder in den elektronischen Grundzustand übergeht. Neben strahlungslosen Über-
gängen wie Vibrationsrelaxation, ist hier die Emission eines Photons von Interesse,
dessen Energie dem Abstand des niedrigsten Vibrationszustandes in S1 und jedem
anderen Vibrations- oder Rotationszustand in S0 entspricht. Die Wellenlänge des
emittierten Photons ist somit in der Regel größer als die des Anregungsphotons
(Rotverschiebung) [27].
Ein Molekül verbringt Zeitspannen in der Größenordnung von Nanosekunden im
angeregten Zustand, bevor Photonen emittiert werden. Ist ein anderer Fluorophor in
der unmittelbaren Umgebung des angeregten Moleküls, kann diese Lebensdauer
noch dadurch verkürzt werden, dass ein Energieübertrag auf den anderen Fluo-
11

Fluoreszenz-Mikroskopie 1
rophor stattfindet und dieses elektronisch anregt (Fluoreszenz-Energie-Resonanz-
Transfer, FRET). Bedingung ist, dass das Emissions-Spektrum des Energie-Donors
und das Absorptions-Spektrum des Akzeptors einen spektralen Überlapp aufwei-
sen. Der strahlungslose Energieübertrag resultiert in der Emission eines Photons
größerer Wellenlänge durch den Akzeptor. Der Energietransfer findet über Dipol-
Dipol-Wechselwirkungen statt mit der Wechselwirkungsenergie EDD [3]:
EDD =1
R3n2L( ~µD ~µA−
3R2 ( ~µD~R)(µAR)) (1.25)
mit dem Brechungsindex nL, dem Dipolmoment des Donors ~µD bzw. dem Dipol-
moment des Akzeptors ~µA und dem Abstand zwischen Donor und Akzeptor ~R.
Während Dipol-Dipol-Wechselwirkungen also generalisiert mit 1R3 abnehmen, ist
die Geschwindigkeitskonstante ktrans dem Quadrat der Wechselwirkungsenergie
proportional [3]:
ktrans =1
n4LR6 µ
2Dµ
2AK2 (1.26)
mit einer Zusammenfassung der Vektortherme zu K2. Unter Berücksichtigung der
Quantenausbeute Q, der Lebensdauer des Donors ohne Akzeptor τD und des
spektralen Überlapps J ergibt sich [3]:
ktrans = konst.(
K2JQn4
LR6τD
). (1.27)
Zusammenfassung der Konstanten zum Förster-Radius R0 folgt [3]:
ktrans =1
τD
(R0
R
)6
. (1.28)
Zur Bestimmung des Abstandes zwischen Donor und Akzeptor wird die Effizienz
des Energietransfers Etrans gemessen [3]:
Etrans =1
1+(
RR0
)6 . (1.29)
Ist R = R0 werden 50% der absorbierten Energie vom Donor auf den Akzeptor über-
tragen. R0 liegt häufig zwischen 2.5 nm und 5 nm [3]. Die starke Distanzabhängigkeit
des FRET-Prozesses erlaubt eine Untersuchung der intermolekularen Abstände
innerhalb einer oder mehrerer Proben in Dimensionen weit unter dem Beugungslimit
der Lichtmikroskopie [27].
12

Desoxyribonukleinsäure 1
Die Technik der klassischen konfokalen Fluoreszenz-Mikroskopie wurde seit Mitte
der 1990er Jahre konsequent um Methoden, die es ermöglichen, das Beugungslimit
und die damit verbundene laterale Auflösung von ungefähr 300 nm und axiale Auf-
lösung von 800 nm zu umgehen oder zu durchbrechen, erweitert. Die Referenzen
[28, 29] und [30] geben eine gute Übersicht über die damit verbundenen Metho-
den der 4π-, STORM (stochastic optical reconstruction microscopy)- und STED
(stimulated emission depletion)-Mikroskopie.
1.5 Desoxyribonukleinsäure
Mit „We wish to suggest a structure for the salt of deoxyribose nucleic acid (DNA)“ be-
richteten Watson und Crick 1953 von der doppelhelikalen Struktur der DNA [31]. DNA
besteht aus zwei Strängen aus sich wiederholenden Nukleotiden. Eine Nukleotidein-
heit besteht dabei aus einer Phosphatgruppe, einer Zuckerstruktur (Desoxyribose)
sowie üblicherweise einer von vier Basen (Abb. 1.5). In der Doppelhelix formen
Adenin und Thymin als Basenpaare zwei Wasserstoffbrückenbindungen miteinander,
Guanin und Cytosin drei. In Zellen, die sich nicht im Mitosestadium befinden, liegt
die DNA als Chromatin, einer hochkondensierten Faser aus Protein und DNA, vor.
13

Desoxyribonukleinsäure 1
Abbildung 1.5: Schematischer Aufbau von DNA, dargestellt im Solvens Wasser (rote Punk-te): Die beiden aüßeren Bänder symbolisieren die beiden Zucker-PhosphatRückgrate, während die davon horizontal abgehenden Stufen Basenpaaredarstellen, die die Stränge über Wasserstoffbrückenbindungen zusammen-halten. Struktur aus PDB-Eintrag 1BNA [32].
14


2Material und Methoden
2.1 Konfokale Raman-Mikrospektroskopie
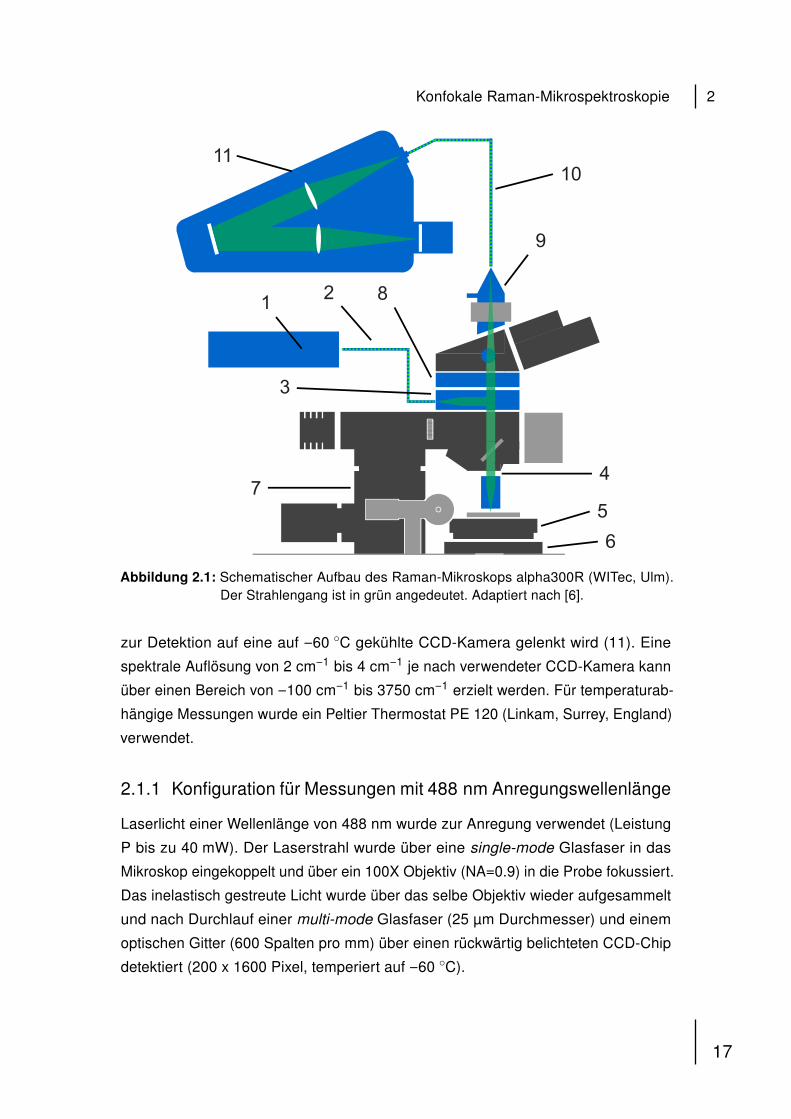
Abbildung 2.1 zeigt den schematischen Aufbau zusammen mit dem angedeuteten
Strahlengang des in dieser Arbeit verwendeten konfokalen Raman- Mikrospek-
troskops alpha300 RAS (WITec, Ulm). Als punktförmige Lichtquelle dient ein fre-
quenzverdoppelter Nd:YAG Festkörperlaser, der Licht einer Wellenlänge von 532 nm
emittiert oder ein single-frequency Diodenlaser mit einer Wellenlänge von 785 nm
(1). Die Laserstrahlung wird über eine wellenlängenspezifische single-mode-Glasfa-
ser (2) in das Mikroskop eingekoppelt (3) und anschließend durch das Mikroskop-
Objektiv (4) in die Probe fokussiert. Für Messungen in wässrigen Lösungen wurde
ein Wassereintauchobjektiv verwendet (60x/NA=1.0W, Arbeitsabstand = 2.0 mm,
Nikon, Chiyoda, Japan). Die Probe befindet sich auf einem Objekttisch (5), der
über eine piezoelektronische Einheit (6) in x- und y-Richtung bewegt werden kann.
Ein Schrittmotor (7) erlaubt durch Bewegung des Mikroskops eine Fokussierung in
z-Richtung. Das Streulicht wird durch das gleiche Objektiv wieder aufgenommen
und durchläuft einen holographischen Filter (8), der die Rayleigh-Strahlung, d.h.
reflektiertes Laserlicht reduziert, bevor es in eine multi-mode-Glasfaser fokussiert
wird (9+10). Der Kern der multi-mode-Glasfaser liefert dabei die Lochblende und
ermöglicht eine konfokale Messung. Die aus der multi-mode-Glasfaser austretende
Strahlung wird anschließend in einem Gitterspektrometer dispergiert bevor der Strahl
16
Konfokale Raman-Mikrospektroskopie 2
12
3
4
5
6
7
8
9
10
11
Abbildung 2.1: Schematischer Aufbau des Raman-Mikroskops alpha300R (WITec, Ulm).Der Strahlengang ist in grün angedeutet. Adaptiert nach [6].
zur Detektion auf eine auf −60 ◦C gekühlte CCD-Kamera gelenkt wird (11). Eine
spektrale Auflösung von 2 cm−1 bis 4 cm−1 je nach verwendeter CCD-Kamera kann
über einen Bereich von −100 cm−1 bis 3750 cm−1 erzielt werden. Für temperaturab-
hängige Messungen wurde ein Peltier Thermostat PE 120 (Linkam, Surrey, England)
verwendet.
2.1.1 Konfiguration für Messungen mit 488 nm Anregungswellenlänge
Laserlicht einer Wellenlänge von 488 nm wurde zur Anregung verwendet (Leistung
P bis zu 40 mW). Der Laserstrahl wurde über eine single-mode Glasfaser in das
Mikroskop eingekoppelt und über ein 100X Objektiv (NA=0.9) in die Probe fokussiert.
Das inelastisch gestreute Licht wurde über das selbe Objektiv wieder aufgesammelt
und nach Durchlauf einer multi-mode Glasfaser (25 µm Durchmesser) und einem
optischen Gitter (600 Spalten pro mm) über einen rückwärtig belichteten CCD-Chip
detektiert (200 x 1600 Pixel, temperiert auf −60 ◦C).
17

Plasma-Quelle 2
2.1.2 Konfiguration für Messungen mit 532 nm Anregungswellenlänge
Laserlicht einer Wellenlänge von 532 nm wurde zur Anregung verwendet (Leistung
P bis zu 30 mW). Der Laserstrahl wurde über eine single-mode Glasfaser in das
Mikroskop eingekoppelt und über ein 60X Objektiv (NA=1.0W) in die Probe fokussiert.
Das inelastisch gestreute Licht wurde über das selbe Objektiv wieder aufgesammelt
und nach Durchlauf einer multi-mode Glasfaser (50 µm Durchmesser) und einem
optischen Gitter (600 Spalten pro mm) über einen rückwärtig belichteten CCD-Chip
detektiert (200 x 1600 Pixel, temperiert auf −60 ◦C).
2.1.3 Konfiguration für Messungen bei 785 nm Anregungswellenlänge
Laserlicht einer Wellenlänge von 785 nm wurde zur Anregung verwendet (Leistung
P bis zu 100 mW). Der Laserstrahl wurde über eine single-mode Glasfaser in das
Mikroskop eingekoppelt und über ein 100X Objektiv (NA=0.9) in die Probe fokussiert.
Das inelastisch gestreute Licht wurde über das selbe Objektiv wieder aufgesammelt
und nach Durchlauf einer multi-mode Glasfaser (100 µm Durchmesser) und einem
optischen Gitter (300 Spalten pro mm) über einen rückwärtig belichteten CCD-Chip
detektiert (128 x 1024 Pixel, temperiert auf −60 ◦C).
2.2 Plasma-Quelle
Plasma-Jets wurden mit 1.4 slm He mit 8.4 sccm O2 in einer He-Atmosphäre betrie-
ben, gesteuert durch 13.56 MHz Radiofrequenz (RF)-Spannung bei einer root mean
square (RMS) Spannung von URMS ∼ 230 V. Zur Ablenkung von Partikeln wurde ein
He-Fluss von 5.6 slm verwendet. Eine doppelt gewundene Düsengeometrie wurde
vor dem Partikel-Jet verwendet, um V(UV) Photonen zu blockieren (Abb. 3.1).
2.3 Proben
2.3.1 Synthetische DNA Homo-Oligomere
Synthetische DNA Homo-Oligomere (Thermo Fischer Scientific, Ulm) bestehend
aus 18 Nukleotiden (Desoxyribose-Adenin18 (dA18), Desoxyribose-Thymin18 (dT18),
Desoxyribose-Guanin18 (dG18), Desoxyribose-Cytosin18 (dC18)) wurden als Ein-
zelstrang verwendet oder hybridisiert, um Doppelstränge zu bilden (dA18:dT18,
dG18:dC18). Proben (1 mM in destilliertem Wasser (aqua destillata, a. dest.)) wurden
18

Proben 2
auf einem Glasobjektträger getrocknet und nach Behandlung für 30 min mit einem
der Jets gemessen. Kontrollproben wurden mit He/O2-Gas behandelt.
2.3.2 Ätzraten
Zur Bestimmung von Ätzraten wurden 60 µL von dA18, dG18, dC18 und dT18 (1 mM
in a. dest.) auf Silizium-Wafern (Siegert Consulting, Aachen) getrocknet. Profile
wurden vor und nach der Behandlung mit den unterschiedlichen Jets (Dauer: 600 s)
mit einem DEKTAK 6M Profilometer (Veeco, New York, USA) gemessen und Pro-
benhöhen bestimmt.
2.3.3 Lebende humane Spermatozoen
Für konfokale Raman-Mikrospektroskopie wurden lebende Spermatozoen eines
gesunden, menschlichen Spenders verwendet. Das so erhaltene Ejakulat wurde vor
weiterer Verwendung für 30 min bei 35 ◦C temperiert (swim up).
2.3.4 Beschichtung von CaF2-Substrat mit Concanavalin A Typ IV
Der verwendete CaF2 Objektträger (Korth, Kiel) wurde zunächst mit a. dest. und
anschließend mit 70%igem Ethanol gereinigt. Concanavalin A Typ IV (Sigma-Aldrich,
Seelze) wurde in 50 mM ACES (Sigma-Aldrich, Seelze) zu einer Konzentration von
0.9 M aufgenommen. Nach Beschichtung des Substrats mit der Proteinlösung wurde
das Substrat für 2 h bei Raumtemperatur getrocknet. Im folgenden Waschschritt
wurde in umgekehrter Reihenfolge mit 70%igem Ethanol und a. dest. gewaschen.
Anschließend wurde wieder für 2 h bei Raumtemperatur getrocknet. Für Verwendung
am selben Tag wurde das Substrat für 2 h bei 37 ◦C temperiert. Eine Lagerung des
Substrats ist bei 4 ◦C möglich.
2.3.5 Percoll-Dichtegradienten-Zentrifugation
Percoll-Dichtegradienten-Zentrifugation wurde ausgehend von einer isotonischen
Percoll-Lösung (Percoll in 10% 1.5 M NaCl)( beides Sigma-Aldrich, Seelze) durch-
geführt. Aus dieser wurden 55%ige bzw. 80%ige Percoll-Lösung in Ham’s nutrient
mixture F12 (Sigma-Aldrich, Seelze) hergestellt und auf 35 ◦C temperiert. Pro 1 mL
Ejakulat wurden in einem 15 mL Falcon-Tube Zentrifugenröhrchen (VWR, Darm-
stadt) 2 mL 80%ige Percoll-Lösung mit 2 mL 55%iger Percoll-Lösung überschichtet.
Darauf wurde unter Vermeidung von Schlierenbildung 1 mL Ejakulat geschichtet
19

Proben 2
und für 30 min bei Raumtemperatur bei 500 g zentrifugiert. In einem Waschschritt
wurde danach der Überstand verworfen und das Pellet mit 5 mL Ringer-Lösung
(8.6 g NaCl, 0.3 g KCl, 0.32 g CaCl2 in 1 L a. dest., alles Sigma-Aldrich, Seelze),
gespült. Anschließend wurde für 15 min bei Raumtemperatur bei 500 g zentrifugiert.
Nach erneutem Verwerfen des Überstands wurden 2 mL Ringer-Lösung zum Pellet
gegeben und auf die gewünschte Konzentration verdünnt.
2.3.6 Automatisierte konfokale Raman-Mikrospektroskopie
Konfokale Raman-Mikrospektroskopie wurde unter der Konfiguration wie sie in
Abschnitt 2.1.2 beschrieben ist, durchgeführt. 1 mL der isolierten Spermatozoen-
Lösung wurde auf das CaF2 aufgetragen, mit vorgewärmter Ringer-Lösung (35 ◦C
überschichtet und sofort im Mikroskop betrachtet. Für automatisierte Messungen wur-
de das „Sample Raster“ Feature von WITec Control 1.56 (2010) verwendet. Punkte in
der Probe wurden manuell ausgewählt, in denen automatisierte Messungen mit vor-
eingestellter Geometrie und Integrationszeit pro Spektrum durchgeführt wurden. Die
Summenfilteranalyse integriert über die Intensitäten ausgewählter Raman-Banden
in den gemessenen Spektren und erstellt farbkodierte Raman-Bilder in Echtzeit.
2.3.7 Vorbehandlung der aufgenommen Datensätze
Raman-Spektrendatensätze wurden mit WITec Project 2.10 (2012) analysiert. Vor
Datenanalyse wurden die Spektren von Störbeiträgen kosmischer Strahlung befreit.
Für eine Hintergrundkorrektur wurden Bereiche im Raman-Spektrum ausgewählt, in
denen keine Raman-Banden zu erkennen sind und die Intensitätswerte in diesen
Bereichen über je nach verwendeter CCD-Kamera über 6 cm−1 bzw. 12 cm−1 gemit-
telt. Andere Bereiche wurden linear interpoliert, resultierend in einem Hintergrund,
der abgezogen werden konnte.
2.3.8 LIVE/DEAD Sperm Viability Kit zur Fluoreszenz-Markierung von
Spermatozoen
Eine 20 µM Stammlösung von SYBR-14 (Molecular Probes, Leiden, Niederlan-
de) in DMSO (Sigma-Aldrich, Seelze) wurde verwendet. Eine Endkonzentration
von 100 nM wurde zu den immobilisierten Spermatozoen gegeben und für 10 min
inkubiert. Für eine Fluoreszenz-Doppelmarkierung lebendiger und chemisch geschä-
digter Zellen wurde SYBR-14 entfernt und 12 µM Propidiumiodid (PI) (Molecular
20

Fluoreszenz-Mikroskopie 2
Probes, Leiden, Niederlande) zu den Zellen gegeben und für 10 min inkubiert.
2.4 Fluoreszenz-Mikroskopie
Nach Waschen der Zellen mit Ringer-Lösung wurden die Spermatozoen in einem
Axio Observer.Z1 Fluoreszenz-Mikroskop (Zeiss, Jena) beobachtet. SYBR-14 in
den Zellen wurde mit Licht einer Wellenlänge von 470 nm oder 555 nm angeregt.
Fluoreszenz-Bilder wurden mit Hilfe eines 10X Plan-Neofluar Objektivs (Zeiss, Jena)
und den Filter Sets 38HE und 63HE (Zeiss, Jena) aufgenommen.
21


3Effekte der Behandlung von DNA mit
einem nicht-thermischen
Atmosphärendruck-Plasma
3.1 Einführung und Motivation
Plasma kann beschrieben werden als partiell ionisiertes Gas, welches freie Elektro-
nen, Ionen und neutrale Partikel enthält. Thermische Plasmen unterscheiden sich
von nicht-thermischen Plasmen in dem Sinne, dass ihre Elektronentemperatur signi-
fikant höher ist, als die Gastemperatur (ungefähr Raumtemperatur). Radiofrequenz-
gesteuerte kalte Atmosphärendruck-Plasma-Jets (microscale atmospheric pressure
plasma, µAPPJ) sind bekannte Beispiele nicht-thermischer Plasmen. Das Gas, wel-
ches für diesen Plasma-Jet verwendet wurde, ist He-Gas mit einem Zusatz von
0.6% O2(v/v) resultierend in O Atomen, metastabilem O2 sowie O3 als reaktive
Sauerstoffspezies [33].
Neue Entwicklungen im Bereich der kalten Atmosphärendruck-Plasmen (cold
atmospheric pressure plasma, CAP) haben zu einem verstärkten Interesse in deren
medizinischen Anwendungen, das heißt in deren desinfizierenden Eigenschaften,
geführt [34]. CAPs sind in der Lage Mikroorganismen zu inaktivieren und Biomakro-
moleküle, wie Proteine und DNA chemisch zu modifizieren [35–38].
23
Einführung und Motivation 3
Trotz des klinischen Erfolgs der Plasma-Medizin ist wenig über die Mechanismen,
die zu diesen Modifikationen führen, bekannt. Gegenstand der Forschung ist es, Ein-
sicht in die Inaktivierungs-Mechanismen von Plasmen in vitro und in vivo zu erhalten.
Die vorliegende Arbeit konzentriert sich auf die Auswirkung der separierten Kom-
ponenten einer spezifischen Plasmaquelle auf die Struktur des Bio-Makromoleküls
DNA in vitro.
Als Plasmaquelle wurde ein µAPPJ verwendet, der in [33] beschrieben und
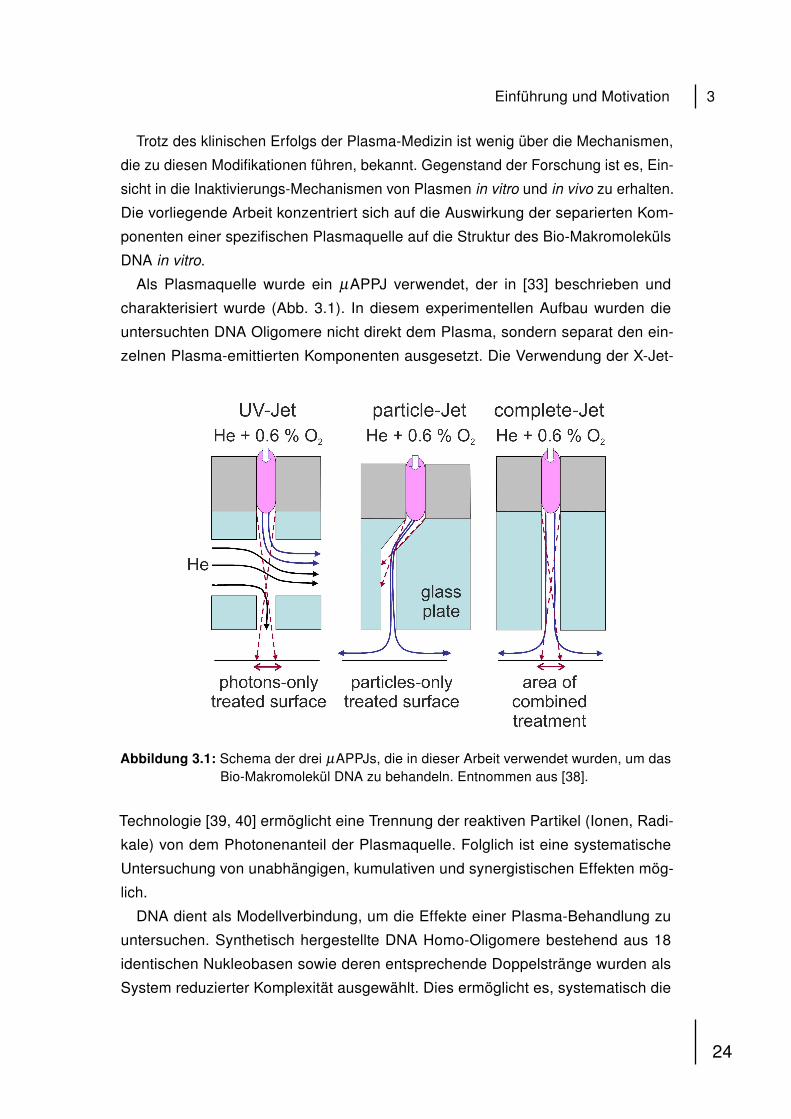
charakterisiert wurde (Abb. 3.1). In diesem experimentellen Aufbau wurden die
untersuchten DNA Oligomere nicht direkt dem Plasma, sondern separat den ein-
zelnen Plasma-emittierten Komponenten ausgesetzt. Die Verwendung der X-Jet-
Abbildung 3.1: Schema der drei µAPPJs, die in dieser Arbeit verwendet wurden, um dasBio-Makromolekül DNA zu behandeln. Entnommen aus [38].
Technologie [39, 40] ermöglicht eine Trennung der reaktiven Partikel (Ionen, Radi-
kale) von dem Photonenanteil der Plasmaquelle. Folglich ist eine systematische
Untersuchung von unabhängigen, kumulativen und synergistischen Effekten mög-
lich.
DNA dient als Modellverbindung, um die Effekte einer Plasma-Behandlung zu
untersuchen. Synthetisch hergestellte DNA Homo-Oligomere bestehend aus 18
identischen Nukleobasen sowie deren entsprechende Doppelstränge wurden als
System reduzierter Komplexität ausgewählt. Dies ermöglicht es, systematisch die
24

Einführung und Motivation 3
Effekte einer Plasma-Behandlung zu untersuchen, speziell im Hinblick auf jede
der vier Nukleobasen und das Desoxyribose-Phosphat-Rückgrat. Homo-Oligomere
wurden zusätzlich deshalb ausgewählt, um einen möglichen stabilisierenden Effekt
von gestapelten Nukleobasen über Wasserstoffbrückenbindungen zu untersuchen.
Während manche Modifikationen exklusiv unter den hier beschriebenen in vitro
Behandlungsbedingungen stattfinden, besteht die Möglichkeit, dass andere Effekte
nur in vivo in einer funktionierenden Zellumgebung beobachtet werden können [38].
Raman-Spektroskopie wurde verwendet, um die Plasma-induzierten Effekte auf
die DNA-Struktur auf eine nicht-destruktive, nicht-invasive Art und Weise zu klassifi-
zieren. Des Weiteren erlaubt Raman-Spektroskopie eine Korrelation mit Ätzeffekten,
indem die Gesamtintensität der gemessenen Raman-Spektren mit den Ergebnissen
aus Profilometrie-Experimenten verglichen wird. Die vorliegende Arbeit stellt einen
Schritt in Richtung Verständnis der Plasma-induzierten molekularen Prozesse an
biologischen Materialien dar.
25

Ergebnisse 3
3.2 Ergebnisse
Konfokale Raman-Spektroskopie wurde verwendet um Modifikationen von DNA
Homo-Oligomeren, bestehend aus 18 identischen Nukleobasen (dG18, dC18, dA18,
dT18) zu untersuchen, welche durch die emittierten Komponenten eines kalten
Atmosphärendruck-Plasmas eingeführt worden sind. Einzelpunkt Raman-Spektren
wurden sowohl von Kontrollproben als auch von DNA Homo-Oligomeren gemessen,
die zuvor mit den spezifischen Plasma-Jets (UV-Jet, Partikel-Jet, Komplett-Jet, 3.1)
für jeweils 30 min behandelt wurden. Die so erhaltenen Raman-Spektren wurden
durch Vergleich mit den Referenzen [41–46] analysiert und sind zum besseren
Vergleich auf der Y-Achse gegeneinander versetzt. Abbildung 3.9 zeigt die Auf-
stellung der 16 Raman-Spektren der unbehandelten Kontrollen der Einzelstrang
Homo-Oligomere und den -Oligomeren behandelt mit den spezifischen Plasma-
Jets. Jedes Spektrum wurde durch 24 Wiederholungen für jeweils 2.5 s erzeugt
(Gesamtintegrationszeit: 60 s, Anregungswellenlänge λ = 785 nm).
Vier unterschiedliche Arten von Banden wurden in den Spektren aller Oligonu-
kleotide annotiert:
1. Nukleobasen-spezifische Banden (üblicherweise im Bereich von 300 cm−1 bis
1600 cm−1) bezeichnet als Ax, Tx, Gx, Cx
2. v(PO2), eine hochstabile Struktur des Phosphat-Rückgrats für Referenzie-
rungszwecke (1092 cm−1)
3. Eine Bande entstehend aus der Schwingung gekoppelter Nukleobasen (cou-
pled nucleobases, cNB; 925 cm−1)
4. Eine Bande bei 1440 cm−1 generiert aus dem Desoxyribose-Teil der DNA
(backbone, bb) .
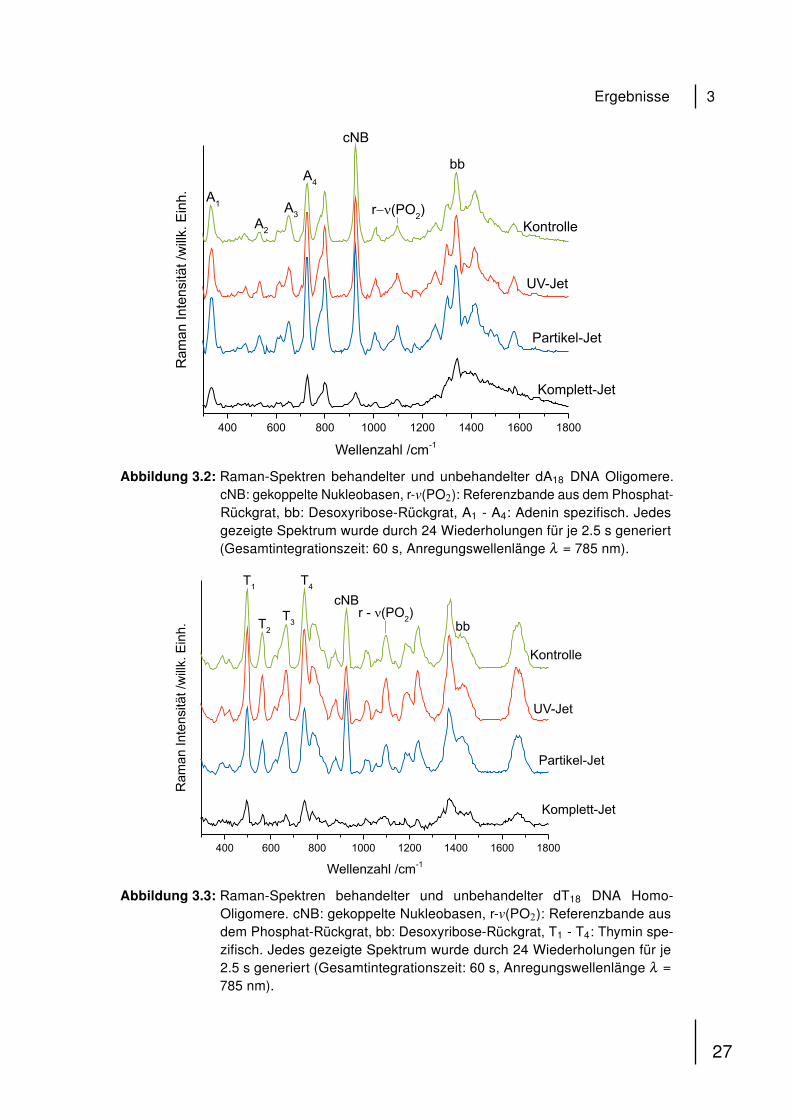
Bei dem Vergleich der Spektren von dA18 behandelt mit dem UV-Jet oder dem
Partikel-Jet allein wurden Änderungen in relativen Bandenintensitäten der Nukleo-
basen-Banden (bezeichnet mit A1 - A4) verglichen mit der Referenz-Bande bei
1092 cm−1 beobachtet (Abb. 3.2). Die relative Intensität der Bande der gekoppelten
Nukleobasen cNB nimmt auf eine ähnliche Art und Weise nach separater UV- und
Partikel-Behandlung ab. Im Gegensatz dazu konnte ein starker Abfall der relativen
Intensität der meisten Banden nach 30 minütiger Behandlung mit dem Komplett-Jet
beobachtet werden. Der deutlichste Effekt ist für die cNB-Bande sichtbar.
26
Ergebnisse 3
400 600 800 1000 1200 1400 1600 1800
r��(PO2)
bbA
4
A3
A2 Kontrolle
UV-Jet
Partikel-Jet
Ram
an
Inte
nsität/w
illk.E
inh.
Wellenzahl /cm-1
Komplett-Jet
cNB
A1
Abbildung 3.2: Raman-Spektren behandelter und unbehandelter dA18 DNA Oligomere.cNB: gekoppelte Nukleobasen, r-v(PO2): Referenzbande aus dem Phosphat-Rückgrat, bb: Desoxyribose-Rückgrat, A1 - A4: Adenin spezifisch. Jedesgezeigte Spektrum wurde durch 24 Wiederholungen für je 2.5 s generiert(Gesamtintegrationszeit: 60 s, Anregungswellenlänge λ = 785 nm).
400 600 800 1000 1200 1400 1600 1800
bb
cNBr - �(PO
2)T
3T2
Ram
an
Inte
nsität/w
illk.E
inh.
Wellenzahl /cm-1
T1
T4
Kontrolle
UV-Jet
Partikel-Jet
Komplett-Jet
Abbildung 3.3: Raman-Spektren behandelter und unbehandelter dT18 DNA Homo-Oligomere. cNB: gekoppelte Nukleobasen, r-v(PO2): Referenzbande ausdem Phosphat-Rückgrat, bb: Desoxyribose-Rückgrat, T1 - T4: Thymin spe-zifisch. Jedes gezeigte Spektrum wurde durch 24 Wiederholungen für je2.5 s generiert (Gesamtintegrationszeit: 60 s, Anregungswellenlänge λ =785 nm).
27

Ergebnisse 3
Die strukturellen Modifikationen, die durch die Behandlung mit den unterschied-
lichen Plasma-Jets in dT18 eingeführt wurden, sind in Abb. 3.3 gezeigt. Während
einige der relativen Bandenintensitäten einem ähnlichen Trend folgen wie in dA18
abgebildet (Abb. 3.2) - wie der komplette Verlust der cNB Bande bei 925 cm−1 nach
Komplett-Jet Behandlung - steigt die relative Intensität der cNB Bande nach Behand-
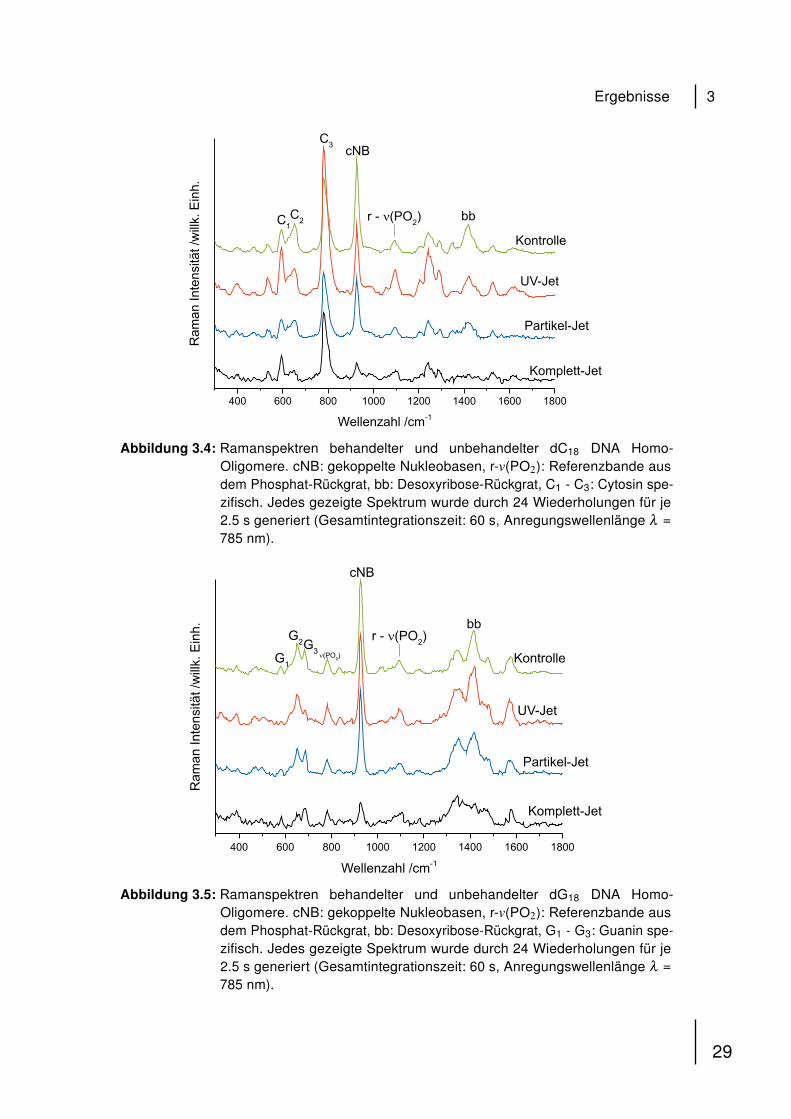
lung mit dem Partikel-Jet an. Abbildung 3.4 zeigt die Raman-Spektren von dC18.
Eine Behandlung mit dem UV-Jet resultiert in Modifikationen der relativen Banden-
intensitäten der Nukleobasen spezifischen Banden C1 und C3. Modifikationen der
Zuckerrückgrat Banden konnten nach Partikel-Jet Behandlung beobachtet werden,
resultierend in einer Abnahme der relativen Bandenintensität bei 1440 cm−1. Wieder
ist der Verlust der relativen Intensität der cNB Bande bei 925 cm−1 nach Komplett-Jet
Behandlung besonders ausgeprägt. Ähnliche Effekte wurden für dG18 gefunden
(Abb. 3.5). Ergebnisse werden hier zum Vergleich gezeigt. [38]. Die Bestimmung der
Ätzeffekte für die verschiedenen Jets soll die beobachtete Abnahme der Gesamt-
signalintensität speziell nach Komplett-Jet Behandlung ergänzen. Eine mögliche
Ursache für die beschriebene Abnahme liefert Ätzen, das heißt Plasma-induziertes
Entfernen von Probenmaterial. Beschleunigte Plasma-generierte Spezies intera-
gieren mit der Probe durch das Entfernen von Atomen (und deshalb Substanz)
aus der Probe. Es wurde bereits untersucht, dass der X-Jet dazu in der Lage ist,
Proteinproben zu Ätzen [39]. Hier wurde das Entfernen von Homo-Oligonukleotiden
untersucht. Die Ätzraten unter Berücksichtigung der Trocknungseffekte sind in Ta-
belle 3.1 dargestellt.
Tabelle 3.1: Ätzraten des X-Jets in (nmmin−1) von DNA Homo-Oligonukleotide.
UV-Jet Partikel-Jet Komplett-Jet
Beobachtete Ätzrate 280 380 660Trocknungseffekt 220 220 220
Tatsächliche Ätzrate 60 160 440
Der UV-Jet zeigt mit 60 nmmin−1 den geringsten Ätzeffekt. Mit 160 nmmin−1
ätzt der Partikel-Jet effektiver. Der Komplett-Jet liefert den größten Ätzbeitrag mit
440 nmmin−1. Trocknungseffekte von 220 nmmin−1 wurden zur Berechnung der
genannten Werte abgezogen. Es konnten keine signifikanten Unterschiede für die
vier unterschiedlichen Oligomere beobachtet werden.
Die durch Profilometrie bestimmten Ätzeffekte wurden anschließend mit den
Raman Signalintensitäten verglichen, die über das Integral (algebraische Sum-
28
Ergebnisse 3
400 600 800 1000 1200 1400 1600 1800
C3
C2C
1bb
Ram
an
Inte
nsität/w
illk.E
inh.
Wellenzahl /cm-1
cNB
r - �(PO2)
Kontrolle
UV-Jet
Partikel-Jet
Komplett-Jet
Abbildung 3.4: Ramanspektren behandelter und unbehandelter dC18 DNA Homo-Oligomere. cNB: gekoppelte Nukleobasen, r-v(PO2): Referenzbande ausdem Phosphat-Rückgrat, bb: Desoxyribose-Rückgrat, C1 - C3: Cytosin spe-zifisch. Jedes gezeigte Spektrum wurde durch 24 Wiederholungen für je2.5 s generiert (Gesamtintegrationszeit: 60 s, Anregungswellenlänge λ =785 nm).
400 600 800 1000 1200 1400 1600 1800
�(PO2)
bb
cNB
G3
G2
Ram
an
Inte
nsität/w
illk.E
inh.
Wellenzahl /cm-1
G1
r - �(PO2)
Kontrolle
UV-Jet
Partikel-Jet
Komplett-Jet
Abbildung 3.5: Ramanspektren behandelter und unbehandelter dG18 DNA Homo-Oligomere. cNB: gekoppelte Nukleobasen, r-v(PO2): Referenzbande ausdem Phosphat-Rückgrat, bb: Desoxyribose-Rückgrat, G1 - G3: Guanin spe-zifisch. Jedes gezeigte Spektrum wurde durch 24 Wiederholungen für je2.5 s generiert (Gesamtintegrationszeit: 60 s, Anregungswellenlänge λ =785 nm).
29

Ergebnisse 3
me der Trapeze) jedes gemessenen Raman-Spektrums von xi (300 cm−1) bis x f
(1800 cm−1) berechnet wurden [47]:
∫ x f
xi
f (x)dx≈f−1
∑i=1
(xi+1− xi)12[ f (xi+1)+ f (xi)] (3.1)
Für einen akkurateren Vergleich wurden die Werte der Integrale der Kontroll-
spektren auf 1 gesetzt und alle anderen Spektren dazu normiert. Abbildung 3.6
zeigt die Ergebnisse der berechneten Integrale für jedes der Spektren abgebildet
in Abb. 3.2 bis 3.5. Die X- und Y-Achse zeigen die Ergebnisse einer Behandlung
mit Kontroll-Gas oder einem der drei Jets. Symbole repräsentieren dabei die vier
Einzelstrang DNA Homo-Oligomere (dA18, dT18, dG18, dC18). Die Behandlung mit
dem UV-Jet resultiert in einem Integral von 1.25±0.15 verglichen zur jeweiligen
gasbehandelten Kontrolle. Partikel-Jet Behandlung zeigt ein kleineres Integral mit
einem größeren Änderungsbereich von 1.0±0.2 über die vier Homo-Oligomere,
während Komplett-Jet Behandlung in einer deutlichen Abnahme des berechneten
Integrals resultiert: 0.45±0.25.
0,0
0,5
1,0
1,5
2,0
0,0 0,5 1,0 1,5 2,00,0
0,5
1,0
1,5
2,0
0,00,51,01,52,0
Partikel-Jet
UV-Jet
Kontrolle
Kompett-Jet
dA18
dT18
dG18
dC18
Abbildung 3.6: Ein Sterndiagramm bildet das Integral (algebraische Summe der Trapeze,Glg. 3.1) von 300 cm−1 bis 1800 cm−1 ab. Auf jeder Achse sind die Werteder Integrale der Spektren nach Behandlung mit Helium-Gas (Kontrolle)oder einem der drei Jets aufgetragen. Die Symbole repräsentieren dabeidie vier DNA Einzelstrang Homo-Oligomere (dA18, dT18, dG18, dC18). DieWerte der Integrale der Kontrollspektren wurden auf 1 normiert.
30

Ergebnisse 3
Für die nachfolgende Analyse wurde der Wert von I(cNB)/I(v(PO2)) berechnet.
Dazu wurde der Wert der maximalen Intensität der cNB-Bande jedes Spektrums divi-
diert durch die jeweils korrespondierende Referenz-Bande I(v(PO2)). I(cNB)/I(v(PO2))
wurde in einem Sterndiagramm aufgetragen, in dem jede Achse das Ergebnis für die
Kontrollprobe beziehungsweise für die Behandlung mit dem UV-, dem Partikel- oder
dem Komplett-Jet widerspiegelt (Abb. 3.7). Unterschiedliche Symbole repräsentieren
die vier DNA Einzelstrang Homo-Oligomere (dA18, dT18, dG18, dC18). Eine starke
Abnahme von I(cNB)/I(v(PO2)) kann nach Komplett-Jet Behandlung für alle vier
untersuchten DNA Homo-Oligomere beobachtet werden. In dT18 und dG18 nimmt
die Intensität von I(cNB)/I(v(PO2)) nach Partikel-Jet Behandlung zu. Änderungen
in der Nukleobasen-Geometrie resultieren folglich entweder in einer Ab- oder einer
Zunahme der relativen Bandenintensität.
0,0
0,5
1,0
1,5
2,0
0,0 0,5 1,0 1,5 2,00,0
0,5
1,0
1,5
2,0
0,00,51,01,52,0
I(cNB)/I(�(PO2))
Partikel-Jet
UV-Jet
Kontrolle
Komplett-Jet
dA18
dT18
dG18
dC18
Abbildung 3.7: Ein Sterndiagramm bildet I(cNB)/I(v(PO2)) ab. Auf jeder Achse istI(cNB)/I(v(PO2)) nach Behandlung entweder mit Helium-Gas (Kontrolle)oder einem der drei Jets aufgetragen. Die Symbole repräsentieren dabei dievier DNA Einzelstrang Homo-Oligomere (dA18, dT18, dG18, dC18). Um einenbesseren Vergleich der Änderungen nach Behandlung mit einem der dreiJets zu gewährleisten wurde I(cNB)/I(v(PO2)) der Kontrollspektren ebenfallsauf 1 normiert.
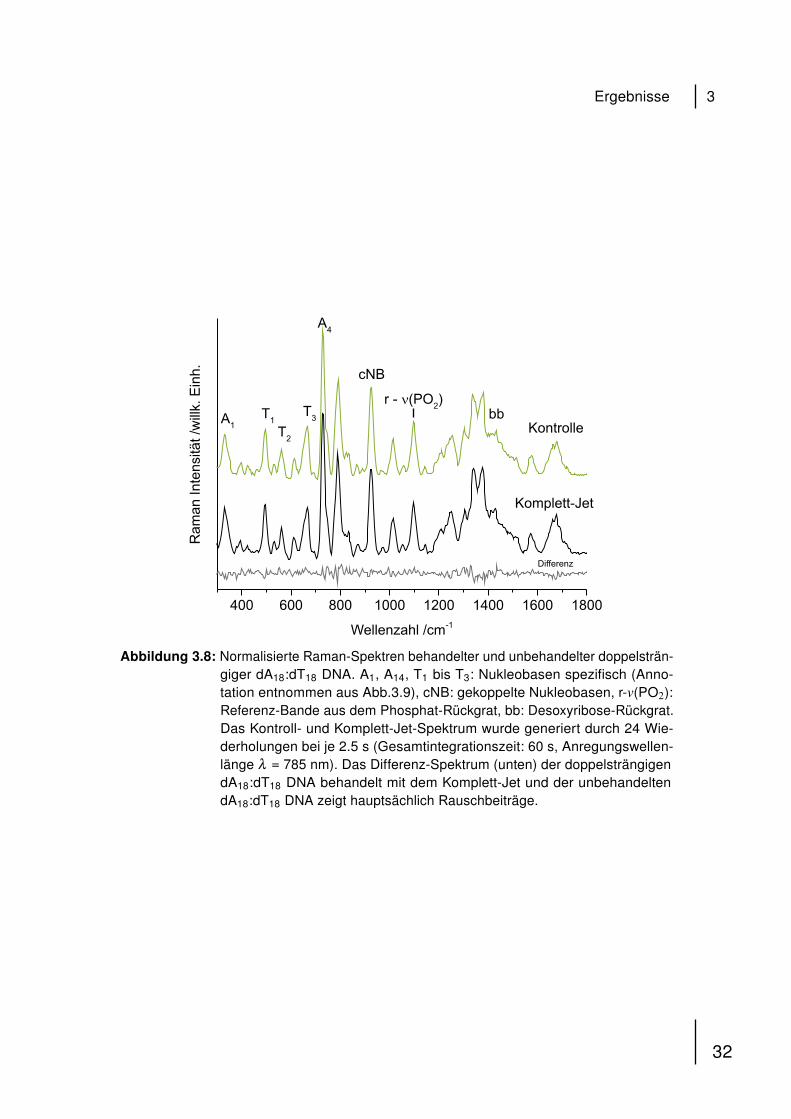
Um mögliche stabilisierende Effekte doppelsträngiger DNA zu untersuchen, wurde
in dieser Arbeit ebenfalls hybridisiertes dA18:dT18 auf gleiche Art und Weise mit dem
Komplett-Jet behandelt wie die korrespondierenden Einzelstränge A18 und T18. Die
Ergebnisse sind in Abb. 3.8 dargestellt.
31
Ergebnisse 3
400 600 800 1000 1200 1400 1600 1800
r - �(PO2)
Differenz
T2
cNB
T3A
1bb
A4
Ram
an
Inte
nsität/w
illk.E
inh.
Wellenzahl /cm-1
Kontrolle
Komplett-Jet
T1
Abbildung 3.8: Normalisierte Raman-Spektren behandelter und unbehandelter doppelsträn-giger dA18:dT18 DNA. A1, A14, T1 bis T3: Nukleobasen spezifisch (Anno-tation entnommen aus Abb.3.9), cNB: gekoppelte Nukleobasen, r-v(PO2):Referenz-Bande aus dem Phosphat-Rückgrat, bb: Desoxyribose-Rückgrat.Das Kontroll- und Komplett-Jet-Spektrum wurde generiert durch 24 Wie-derholungen bei je 2.5 s (Gesamtintegrationszeit: 60 s, Anregungswellen-länge λ = 785 nm). Das Differenz-Spektrum (unten) der doppelsträngigendA18:dT18 DNA behandelt mit dem Komplett-Jet und der unbehandeltendA18:dT18 DNA zeigt hauptsächlich Rauschbeiträge.
32

Ergebnisse 3
Sowohl das Kontroll- als auch das Spektrum nach Behandlung mit dem Komplett-
Jet zeigen Beiträge der Nukleobasen Adenin und Thymin im Bereich von 300 cm−1
bis 1700 cm−1. Die cNB-Bande zeigt eine schwache Intensitätsabnahme, während
andere spektrale Charakteristika annähernd unverändert nach Komplett-Jet Be-
handlung bleiben. Dies ist illustriert im Differenzspektrum (IKontrolle - IKomplett-Jet), das
hauptsächlich Rauschen aufweist.
33
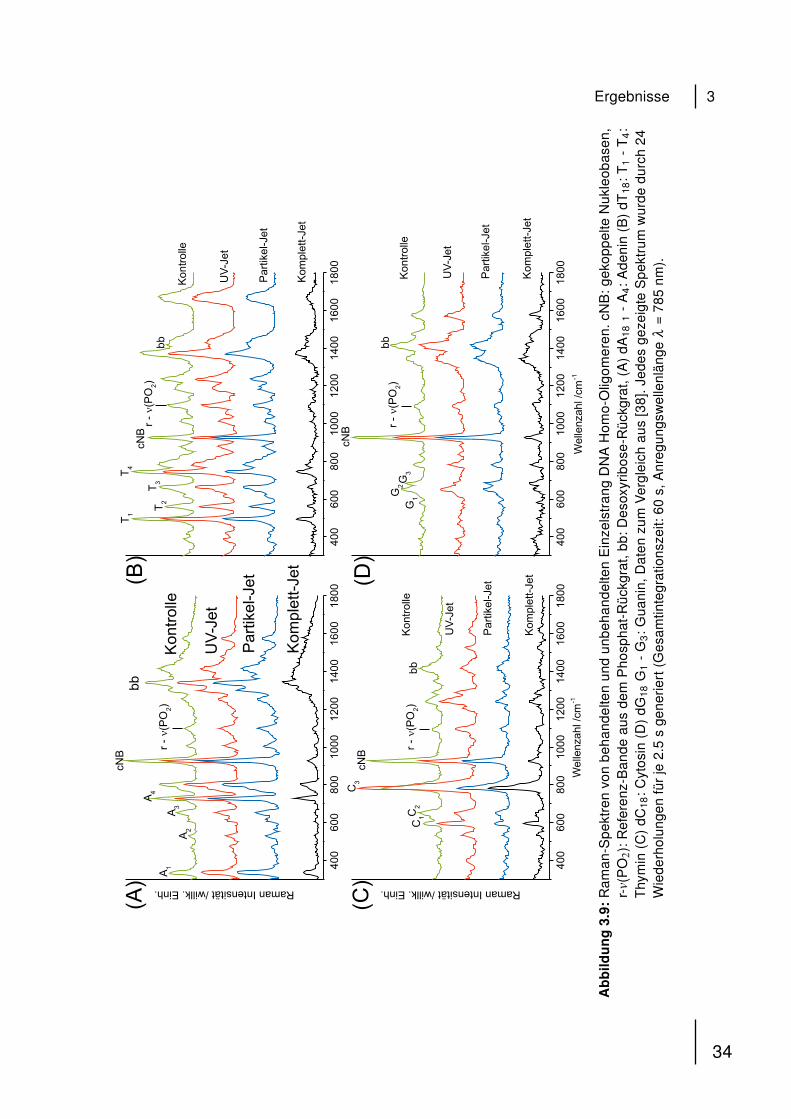
Ergebnisse 3
40
06
00
80
01
00
01
20
01
40
01
60
01
80
04
00
60
08
00
10
00
12
00
14
00
16
00
18
00
40
06
00
80
01
00
01
20
01
40
01
60
01
80
04
00
60
08
00
10
00
12
00
14
00
16
00
18
00
r -
�(P
O2)
(C)
(B)
A4
A3
A2
RamanIntensität/willk.Einh.
cN
B
A1
(A)
r -
�(P
O2)
Kontr
olle
UV
-Jet
Part
ikel-Jet
Kom
ple
tt-J
et
bb
bb
cN
BT
3T
2
T1
T4
r -
�(P
O2)
Kontr
olle
UV
-Je
t
Part
ikel-Jet
Kom
ple
tt-J
et
(D)
cN
BC
3
C2
C1
bb
RamanIntensität/willk.Einh.
We
llenzahl /c
m-1
cN
B
Kontr
olle
UV
-Je
t
Pa
rtik
el-Je
t
Ko
mp
lett
-Je
t
Kontr
olle
UV
-Je
t
Part
ikel-Jet
Ko
mp
lett
-Je
t
We
llenzahl /c
m-1
r -
�(P
O2)
G1
G2G
3
bb
Ab
bild
un
g3.
9:R
aman
-Spe
ktre
nvo
nbe
hand
elte
nun
dun
beha
ndel
ten
Ein
zels
tran
gD
NA
Hom
o-O
ligom
eren
.cN
B:g
ekop
pelte
Nuk
leob
asen
,r-
v(P
O2)
:Ref
eren
z-B
ande
aus
dem
Pho
spha
t-R
ückg
rat,
bb:D
esox
yrib
ose-
Rüc
kgra
t,(A
)dA
181
-A
4:A
deni
n(B
)dT
18:T
1-
T4:
Thy
min
(C)
dC18
:Cyt
osin
(D)
dG18
G1
-G
3:G
uani
n,D
aten
zum
Ver
glei
chau
s[3
8].J
edes
geze
igte
Spe
ktru
mw
urde
durc
h24
Wie
derh
olun
gen
für
je2.
5s
gene
riert
(Ges
amtin
tegr
atio
nsze
it:60
s,A
nreg
ungs
wel
lenl
änge
λ=
785
nm).
34

Diskussion und Ausblick 3
3.3 Diskussion und Ausblick
Ein Ziel dieser Arbeit war es Effekte der Plasma-emittierten Komponenten auf ein
in vitro DNA-Modellsystem zu untersuchen. Die Verwendung des X-Jets, der eine
Separierung reaktiver Partikel von Photonen vereinfacht, erlaubte dabei eine Unter-
suchung separater, kumulativer und synergistischer Mechanismen mit konfokaler
Raman-Spektroskopie.
Die Diskussion der molekularen Mechanismen und Effekte der individuellen Plas-
ma Komponenten basiert auf den bereits beschriebenen Mechanismen, nament-
lich Ätzen durch Ionen sowie Rückgrat-Modifikationen und Vernetzung von DNA
[38, 39]. Ätzeffekte können als chemische Oxidationsprozesse betrachtet werden.
Die Plasma-emittierten reaktiven Spezies führen zu Bindungsbrüchen, speziell in
Kohlenwasserstoffen. Diese offenen Bindungsstellen werden weiter modifiziert zu
molekularen Fragmenten und leicht flüchtigen Strukturen [48]. Es wurde gezeigt,
dass UV-Jet Behandlung einer amorphen Kohlenstoffschicht nicht zu Ätzeffekten
führt, während Ätzen für den Partikel- und den Komplett-Jet beobachtet wurde [38].
Ein ähnlicher Trend kann beobachtet werden, wenn die Integrale der gemessenen
Raman-Spektren mit Ätzraten bestimmt für DNA verglichen werden.
Wenn DNA Einzelstränge bestehend aus 18 identischen Nukleotiden (A,T,G oder
C) mit dem Komplett-Jet behandelt wurden, konnte ein synergistischer Effekt zwi-
schen UV Photonen und Partikeln für alle vier Typen untersuchter Einzelstrang DNA
beobachtet werden. Der in dieser Arbeit beobachtete synergistische Effekt der kom-
binierten (V)UV Photonen und reaktiven Partikel kann verstanden werden, wenn die
cNB-Bande bei 925 cm−1 analysiert wird. Behandlung allein mit dem UV- oder dem
Partikel-Jet führt kleinere Änderungen in die gekoppelte Nukleobasen-Geometrie ein,
mit der größten - wenn auch absolut gesehen kleinen - Änderung für dT18. Während
das Intensitätsverhältnis I(cNB)/I(v(PO2)) nach Behandlung mit dem Partikel-Jet
zunahm, wurde ein kompletter Verlust der cNB-Bande nach Komplett-Jet Behand-
lung beobachtet. Kleine Änderungen in der gekoppelten Nukleobasen-Geometrie
wurden in DNA Einzelsträngen in vitro sowohl durch den UV- als auch durch den
Partikel-Jet induziert. Die Kombination aus V(UV) Photonen und Partikeln führt
überraschenderweise zu einem nicht-kumulativen, synergistischen Effekt, der sich
durch den vollständigen Verlust der cNB Bande in dT18 und dG18 und dem nahezu
vollständigen Verlust in dC18 und dA18 äußert. Gekoppelte Nukleobasen- Geometrie-
und Nukleobasen/Rückgrat-Modifikationen gehen Hand in Hand, resultierend im
beobachteten synergistischen Mechanismus.
35

Diskussion und Ausblick 3
Werden die Ätzraten des Komplett-, UV- und Partikel-Jets des µAPPJ von DNA
verglichen mit den Integralen der Spektren (Abb. 3.6) und der Abnahme der cNB-
Bande bei 925 cm−1 (Abb. 3.7), kann ausgeschlossen werden, dass Ätzen der aus-
schließliche Mechanismus für die Inaktivierung der DNA ist. Die erhöhte strukturelle
und chemische Stabilität von Doppelstrang-DNA resultiert in verringerten chemi-
schen Modifikationen durch die Plasma-emittierten Komponenten. Das Spektrum der
Doppelstrang-DNA ist bis auf eine relative schwache Abnahme der cNB-Bande an-
nähernd unbeeinflusst von der Plasma Behandlung, obwohl Ätzen des Komplett-Jets
von DNA beobachtet wurde.
In komplexeren biologischen Systemen, wie eukaryotischen und prokaryotischen
Zellen, umgibt die Zellmembran beziehungsweise Zellwand die in der Zelle befindli-
chen Nukleinsäuren. Studien an B. subtilis behandelt mit der selben Plasma-Quelle
wie in dieser Arbeit, zeigten, dass nach 30 min Behandlung mit dem Komplett-Jet
Ätzeffekte in einem Radius von 3 mm um die zentrale Jet-Achse [38] beobachtet
werden konnten. Das Bakterienwachstum war jedoch auf einer weit größeren Fläche
der gesamten Petrischale (Radius 43.5 mm) inhibiert. Behandlung mit dem Partikel-
Jet zeigte ähnliche Ergebnisse, während nach Behandlung mit dem UV-Jet keine
Ätzeffekte und eine Inhibitionszone mit einem kleineren Radius von nur 1.5 mm be-
obachtet wurden. Zusätzlich wurde B. subtilis in Flüssigkultur in Umgebungsluft für
30 min mit dem Komplett-Jet behandelt. Die Expression von Stressindikator-Genen
für DNA (recA) zeigte eindeutig eine auf DNA-Schädigungen folgende Stressantwort
der Zellen [38]. Diese Ergebnisse unterstützen die in dieser Arbeit aufgestellte Hy-
pothese, dass Ätzen nicht der Hauptmechanismus für die beobachteten Effekte des
Komplett-Jets ist, sondern dass die Kombination aus (V)UV Photonen und reaktiven
Spezies auf synergistische Art und Weise, zum Beispiel durch eine chemische Akti-
vierung der reaktiven Partikel durch die (V)UV Photonen, chemische Modifikationen
in DNA einführen kann und damit in der Lage ist, diese zu inaktivieren.
Diese Ergebnisse stellen ein solides Fundament für zukünftige Studien von Effek-
ten in flüssiger und damit komplexerer Umgebung sowohl in vitro als auch in vivo
dar. DNA Schäden können in komplexeren biologischen Proben (bösartige Säuger-
zellen und -gewebe) untersucht werden. Die Verwendung anderer Plasma-Quellen
mit einer erhöhten Varianz reaktiver Spezies und Wechselwirkungen (dielektrische
Barriereentladung, X-Jet in Umgebungsluft) kann systematisch in Betracht gezogen
werden. Des Weiteren kann Chemie in flüssigen Proben analysiert werden.
36


4Konfokale Raman-Mikrospektroskopie
immobilisierter, lebender
Spermatozoen unter
physiologieähnlichen Bedingungen
4.1 Einführung und Motivation
Unfruchtbarkeit betrifft 3% bis 17% der Paare in industrialisierten Ländern und wurde
als öffentliches Gesundheitsproblem von der Weltgesundheitsorganisation (World
Health Organization, WHO) eingestuft [49]. In über 30% der Fälle von Infertilität
wurde der männliche Partner als alleinige oder maßgeblich beitragende Ursache
identifiziert [50]. Ergebnisse aus Studien, die nicht nur exklusiv in entwickelten
Ländern durchgeführt wurden, unterstützen die Theorie, dass aus verschiedensten
Gründen eine Abnahme in männlicher Fruchtbarkeit stattfindet [51]. In einer Studi-
enpopulation von 26609 französischen Männern nahm die Spermienkonzentration
bei Männern für ein Durchschnittsalter von 35 Jahren von 73.6 Millionen Spermien
pro ml 1989 auf einen Wert von 49.9 Millionen Spermien pro ml im Jahr 2005 ab
[52]. Dieser Wert liegt unter dem WHO-Kriterium von 55 Millionen Spermien pro ml
resultierend in verlängerten Zeiten für eine erfolgreiche Befruchtung [53].
38

Einführung und Motivation 4
Abbildung 4.1 zeigt den schematischen Aufbau der männlichen Keimzelle, des
Spermatozoons. Dem flachen, scheibenförmigen Kopfteil mit haploidem Zellkern
(DNA) folgt der Mittelteil, in dem sich die zur Fortbewegung durch die Geissel nötigen
Mitochondrien befinden [54, 55].
KopfMittelteilGeissel
Mitochondrien
Abbildung 4.1: Schematischer Aufbau eines humanen Spermatozoons. Der hochkonden-sierte haploide Zellkern befindet sich im Kopf. An den Mittelteil, der Mit-ochondrien beinhaltet, schließt sich die Geissel an, die der Fortbewegungder Zelle dient.
Zentral für die gesamte Disziplin der Abschätzung männlicher Fruchtbarkeit ist das
Konzept der Verwendung standardisierter Laboranalysen humaner Spermatozoen,
wie die regelmäßig von der WHO herausgegebenen, global eingesetzten Standards,
um den morphologischen Status von Spermatozoen zu beurteilen [53]. Aber nicht
nur konventionelle Parameter wie Spermien-Morphologie, Motilität und Konzentration
müssen betrachtet werden, wenn männliche Fruchtbarkeit quantifiziert wird, sondern
auch biochemische Modifikationen wie fragmentierte nukleäre DNA, gerade wenn
Beides nicht offensichtlich miteinander verknüpft auftaucht. Das bedeutet, dass
Spermien, die zunächst eine „normale“ Morphologie und Motilität wie von den WHO-
Kriterien beschrieben aufweisen, trotzdem nicht in der Lage sein können, eine
Schwangerschaft zu erzielen (Idiopathie) [56].
Während ein Arsenal an Methoden vorhanden ist, unterschiedliche chemische
Eigenschaften von Spermatozoen zu untersuchen, ist keine davon komplett nicht-
invasiv oder nicht-schädigend für die untersuchte Probe, was eine weitere Verwen-
dung der untersuchten Zellen für künstliche Befruchtung (assisted reproductive tech-
nology, ART) augenblicklich ausschließt [57, 58]. Daraus schlussfolgernd wird eine
Methode benötigt, mit der es möglich ist, den biochemischen Status eines einzelnen,
lebenden Spermiums im klinischen Rahmen zu evaluieren, um einen bestmöglichen
Ausgang bezüglich Fruchtbarkeit in ART zu gewährleisten. Dieses Problem wurde
39

Einführung und Motivation 4
kürzlich in zwei Reviews von Sanchez et al. und Jasensky et al. [59–61] beschrieben,
in denen eine Aktualisierung bezüglich des klinischen Wertes, aktuelle Anforde-
rungen und Perspektiven verschiedener Aspekte der Spermien-Analyse adressiert
wurde. Verdeutlicht wurde noch einmal die Notwendigkeit neuartiger bildgebender
Verfahren, um Gameten und Embryos nicht-invasiv für ART selektieren zu können.
Konfokale Raman-Mikrospektroskopie (confocal Raman microspectroscopy, CRM)
ist eine nicht-invasive Methode mit hoher räumlicher Auflösung im Sub-Mikrometer
Regime, die eine chemische Beurteilung kleiner Moleküle, verschiedener Arten
von Zellen und ganzen Geweben ermöglicht. Die Referenzen [62–64] geben eine
gute Übersicht über CRM im Gebiet zellulärer Mikroskopie. Die erste Studie von
Raman-Spektroskopie, die verwendet wurde, um Spermien zu untersuchen kann
in [65] gefunden werden, in dem Kubasek et al. erfolgreich das Raman-Spektrum
aufgereinigter Spermien DNA aus Lachs aufnehmen konnte. Dieser Arbeit folgend,
erschienen mehrere Studien, die die Realisierbarkeit von CRM als wertvolles analyti-
sches Werkzeug für die Untersuchung von Spermatozoen herausarbeiteten.
Huser et al. entfernte Membranen und Geisseln von menschlichen Spermatozoen,
um Unterschiede in den Spektren von Chromatin zu untersuchen [66]. Darauf
folgend konnte Meister et al. [67] den schädlichen Einfluss von UV Photonen auf die
unterschiedlichen Organellen von Spermien in luftgetrockneten Spermien-Schmieren
beobachten. Ebenfalls luftgetrocknete Spermien-Schmiere wurden von Mallidis et al.
analysiert, resultierend in der Fähigkeit UV-bestrahlte Proben von nicht behandelten
Zellen über einen Shift der DNA Phosphat-Rückgrat-Bande von 1095 cm−1 nach
1045 cm−1 zu unterscheiden [68]. Multivariate Analysemethoden wurden angewandt,
um zu zeigen, dass Samenplasma von normalen und abnormalen Spermien mittels
CRM unterschieden werden können [69]. Künstlich induzierte oxidative Schäden
in Spermien sind unter Hinzunahme von Fourier-Transform-Infrarot-Spektroskopie
und Durchflusszytometrie durch die Frequenzshifts der Banden im Bereich DNA
Phosphat-Rückgrat elaboriert worden [70]. Multivariate Datenanalyse zusammen
mit einem neuartigen Substrat erlaubte wiederum eine Trennung von normalen und
abnormalen Spermatozoen [71, 72].
Jede der genannten Studien analysierte chemische Schäden an Spermien mit
Raman-Spektroskopie als (luftgetrocknete) Schmiere oder Zellkomponenten-Extrak-
te, was wiederum die Notwendigkeit neuer Routinen und Methoden verdeutlicht, um
einzelne, lebende Spermatozoen unter physiologischen Konditionen nicht-invasiv
chemisch zu evaluieren.
Einzelne, lebende Spermatozoen konnten bisher nicht unter Bedingungen, die
40

Einführung und Motivation 4
für deren Physiologie relevant wären, chemisch evaluiert werden. Ebenfalls unbe-
kannt ist der Einfluss der Laserbestrahlung von Spermatozoen während Raman-
Experimenten mit dafür typischen Parametern.
In dieser Arbeit wurden erfolgreich lebende menschliche Spermatozoen unter
physiologieähnlichen Bedingungen mit CRM chemisch untersucht und eine neue
Methode entwickelt, die eine schnelle Messung einzelner, immobilisierter Spermato-
zoen ermöglicht. Des Weiteren wurde das Risiko bewertet, Spermien der Strahlung
des Anregungslasers während Raman-Messungen auszusetzen.
41

Ergebnisse 4
4.2 Ergebnisse
Bevor in diesem Abschnitt auf die Ergebnisse automatisierter CRM immobilisierter,
lebender Spermatozoen unter physiologieähnlichen Bedingungen eingegangen wird,
folgt zunächst eine Beschreibung wie die im Folgenden gezeigten Bilder aus den
gemessenen hyperspektralen Datensätzen erstellt wurden. Für die Bilderzeugung
wurde die univariate Methode der Summenfilteranalyse angewandt. In der univaria-
ten Analyse wird ein einzelnes Spektrum dafür benutzt, um einen damit verbundenen
Datenpunkt (Pixel) im Bild zu erzeugen. Jedem gemessenen Spektrum in einem
Datensatz wird also ein Pixel im daraus erzeugten Bild zugeordnet [21]. Die hier ver-
wendete Summenfilteranalyse stellt ein Beispiel einer solchen univariaten Methode
zur Bildgebung dar, auf die hier kurz eingegangen werden soll.
Die Summenfilteranalyse basiert auf der Integration der Intensitäten einer spezifi-
schen Bande von Interesse im Raman-Spektrum. Der Summenfilter summiert also
alle Intensitätswerte I(v) im Bereich der Wellenzahlen vn1 und vn1 auf:
Summe =n2
∑i=n1
I(vi) (4.1)
Der Vorteil dieser Art der Analyse ist die Umwandlung multidimensionaler Da-
tensätze zu einem einzigen Ergebnis in Form eines summierten Intensitätswertes.
Durch diese Methode wird also jedes einzelne gemessene Spektrum in einen sum-
mierten Intensitätswert der ausgewählten Schwingungsbande umgewandelt. Wird
diesem Intensitätswert eine Farbe auf einer Farbskala zugewiesen, kann Pixel für
Pixel ein farbkodiertes Bild aus den gemessenen Spektren erstellt werden. Das
Prinzip dieser Art der Datenanalyse ist in Abbildung 4.2 dargestellt.
Abbildung 4.3 (A) zeigt eine optische Aufnahme von mehreren lebenden, im-
mobilisierten Spermatozoen auf CaF2 bei physiologieähnlichen Bedingungen. Die
Spermatozoen weisen eine normale Morphologie auf. Die Raman Bilder in Abb. 4.3
(B) - (S), erstellt durch Summenfilteranalyse, zeigen drei voneinander unterscheidba-
re Bereiche: Die komplette Zelle wurde durch Integration der Intensitäten im Bereich
von verschiedenen Kohlenwasserstoffschwingungen von 2800 cm−1 bis 3000 cm−1
visualisiert (Abb. 4.3 (B) - (G)). Der Kopfteil der Spermatozoen konnte durch Ver-
wendung des Frequenzbereiches von 1090 cm−1 bis 1100 cm−1 dargestellt werden
(Abb. 4.3 (H) - (M)). Das Mittelstück, positioniert direkt hinter dem Kopf, erscheint
bei Verwendung des Summenfilters von 745 cm−1 bis 755 cm−1 (Abb. 4.3 (N) - (S)).
Die Durchschnittsspektren (siehe Gleichung 4.2 verschiedener Bereiche der Zelle
42
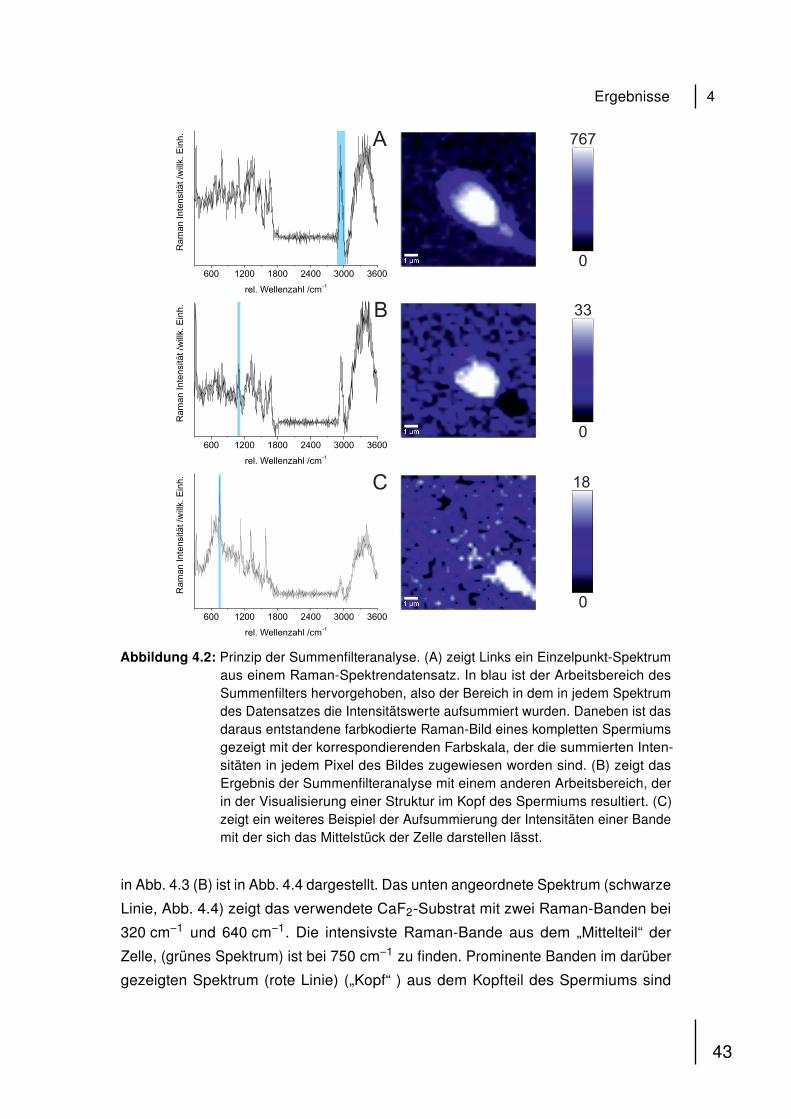
Ergebnisse 4
A
B
C
767
0
33
18
0
0
600 1200 1800 2400 3000 3600
Ra
ma
nIn
ten
sitä
t/w
illk
.E
inh
.
rel. Wellenzahl /cm-1
600 1200 1800 2400 3000 3600
Ra
ma
nIn
ten
sitä
t/w
illk
.E
inh
.
rel. Wellenzahl /cm-1
600 1200 1800 2400 3000 3600
Ra
ma
nIn
ten
sitä
t/w
illk
.E
inh
.
rel. Wellenzahl /cm-1
Abbildung 4.2: Prinzip der Summenfilteranalyse. (A) zeigt Links ein Einzelpunkt-Spektrumaus einem Raman-Spektrendatensatz. In blau ist der Arbeitsbereich desSummenfilters hervorgehoben, also der Bereich in dem in jedem Spektrumdes Datensatzes die Intensitätswerte aufsummiert wurden. Daneben ist dasdaraus entstandene farbkodierte Raman-Bild eines kompletten Spermiumsgezeigt mit der korrespondierenden Farbskala, der die summierten Inten-sitäten in jedem Pixel des Bildes zugewiesen worden sind. (B) zeigt dasErgebnis der Summenfilteranalyse mit einem anderen Arbeitsbereich, derin der Visualisierung einer Struktur im Kopf des Spermiums resultiert. (C)zeigt ein weiteres Beispiel der Aufsummierung der Intensitäten einer Bandemit der sich das Mittelstück der Zelle darstellen lässt.
in Abb. 4.3 (B) ist in Abb. 4.4 dargestellt. Das unten angeordnete Spektrum (schwarze
Linie, Abb. 4.4) zeigt das verwendete CaF2-Substrat mit zwei Raman-Banden bei
320 cm−1 und 640 cm−1. Die intensivste Raman-Bande aus dem „Mittelteil“ der
Zelle, (grünes Spektrum) ist bei 750 cm−1 zu finden. Prominente Banden im darüber
gezeigten Spektrum (rote Linie) („Kopf“ ) aus dem Kopfteil des Spermiums sind
43
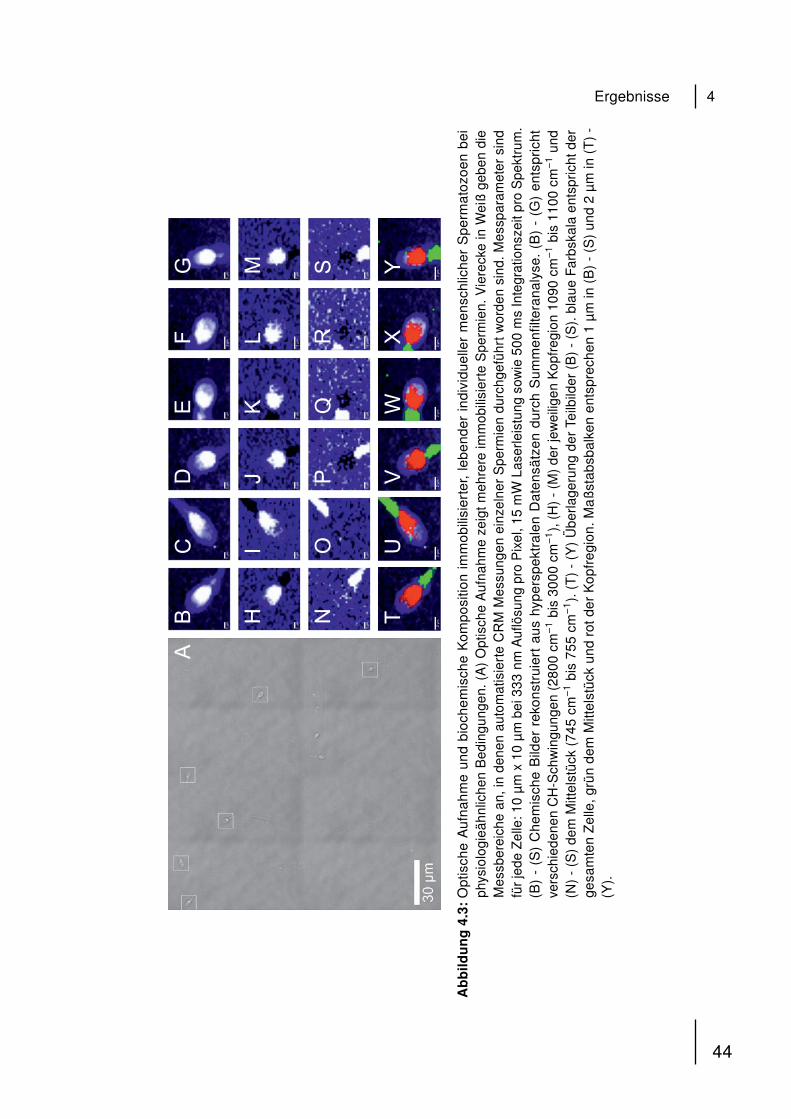
Ergebnisse 4
AB
CD
EF
G
HI
JK
LM
NO
PQ
RS
TU
VW
XY
Ab
bild
un
g4.
3:O
ptis
che
Auf
nahm
eun
dbi
oche
mis
che
Kom
posi
tion
imm
obili
sier
ter,
lebe
nder
indi
vidu
elle
rm
ensc
hlic
her
Spe
rmat
ozoe
nbe
iph
ysio
logi
eähn
liche
nB
edin
gung
en.(
A)
Opt
isch
eA
ufna
hme
zeig
tmeh
rere
imm
obili
sier
teS
perm
ien.
Vie
reck
ein
Wei
ßge
ben
die
Mes
sber
eich
ean
,in
dene
nau
tom
atis
iert
eC
RM
Mes
sung
enei
nzel
ner
Spe
rmie
ndu
rchg
efüh
rtw
orde
nsi
nd.M
essp
aram
eter
sind
für
jede
Zel
le:1
0µ
mx
10µm
bei3
33nm
Aufl
ösun
gpr
oP
ixel
,15
mW
Lase
rleis
tung
sow
ie50
0m
sIn
tegr
atio
nsze
itpr
oS
pekt
rum
.(B
)-
(S)
Che
mis
che
Bild
erre
kons
trui
ert
aus
hype
rspe
ktra
len
Dat
ensä
tzen
durc
hS
umm
enfil
tera
naly
se.
(B)
-(G
)en
tspr
icht
vers
chie
dene
nC
H-S
chw
ingu
ngen
(280
0cm
−1
bis
3000
cm−
1),
(H)
-(M
)de
rje
wei
ligen
Kop
freg
ion
1090
cm−
1bi
s11
00cm
−1
und
(N)
-(S
)de
mM
ittel
stüc
k(7
45cm
−1
bis
755
cm−
1).
(T)
-(Y
)Ü
berla
geru
ngde
rTe
ilbild
er(B
)-
(S).
blau
eFa
rbsk
ala
ents
pric
htde
rge
sam
ten
Zel
le,g
rün
dem
Mitt
elst
ück
und
rotd
erK
opfr
egio
n.M
aßst
absb
alke
nen
tspr
eche
n1
µmin
(B)
-(S
)un
d2
µmin
(T)
-(Y
).
44
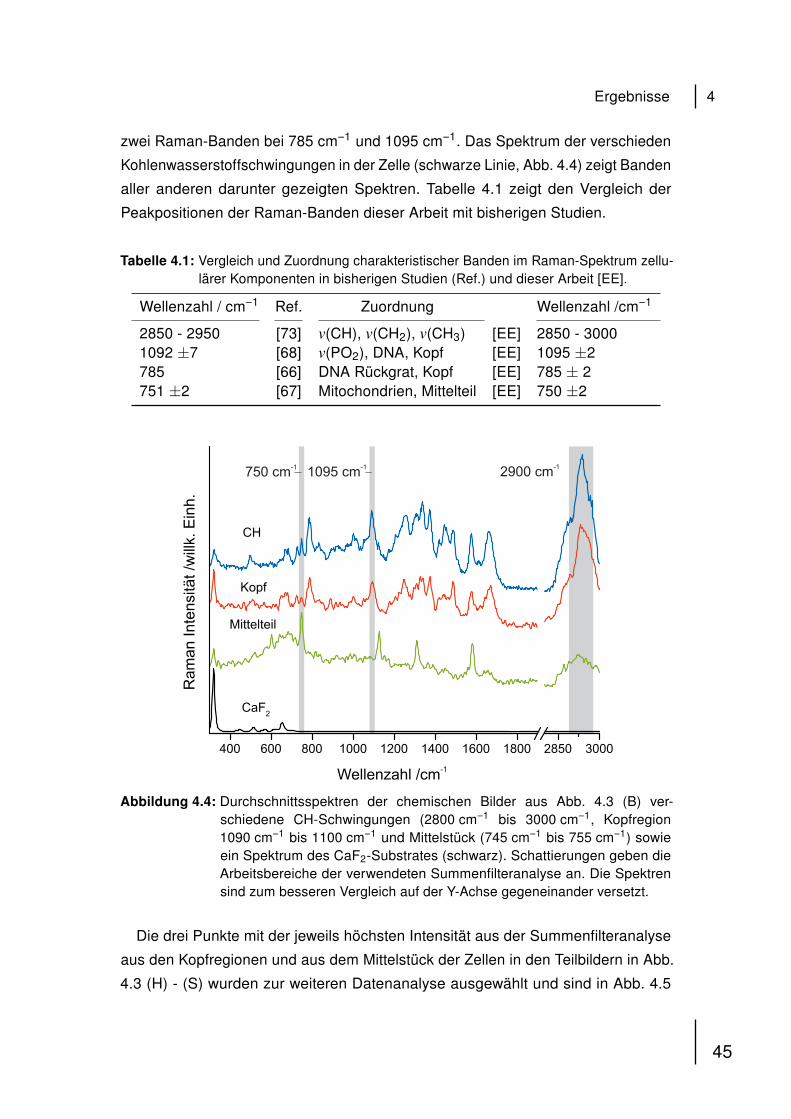
Ergebnisse 4
zwei Raman-Banden bei 785 cm−1 und 1095 cm−1. Das Spektrum der verschieden
Kohlenwasserstoffschwingungen in der Zelle (schwarze Linie, Abb. 4.4) zeigt Banden
aller anderen darunter gezeigten Spektren. Tabelle 4.1 zeigt den Vergleich der
Peakpositionen der Raman-Banden dieser Arbeit mit bisherigen Studien.
Tabelle 4.1: Vergleich und Zuordnung charakteristischer Banden im Raman-Spektrum zellu-lärer Komponenten in bisherigen Studien (Ref.) und dieser Arbeit [EE].
Wellenzahl / cm−1 Ref. Zuordnung Wellenzahl /cm−1
2850 - 2950 [73] v(CH), v(CH2), v(CH3) [EE] 2850 - 30001092 ±7 [68] v(PO2), DNA, Kopf [EE] 1095 ±2785 [66] DNA Rückgrat, Kopf [EE] 785 ± 2751 ±2 [67] Mitochondrien, Mittelteil [EE] 750 ±2
750 cm-1
1095 cm-1
2900 cm-1
Wellenzahl /cm-1
400 600 800 1000 1200 1400 1600 1800 2850 3000
Ram
an Inte
nsität /w
illk. E
inh.
CH
Kopf
Mittelteil
CaF2
Abbildung 4.4: Durchschnittsspektren der chemischen Bilder aus Abb. 4.3 (B) ver-schiedene CH-Schwingungen (2800 cm−1 bis 3000 cm−1, Kopfregion1090 cm−1 bis 1100 cm−1 und Mittelstück (745 cm−1 bis 755 cm−1) sowieein Spektrum des CaF2-Substrates (schwarz). Schattierungen geben dieArbeitsbereiche der verwendeten Summenfilteranalyse an. Die Spektrensind zum besseren Vergleich auf der Y-Achse gegeneinander versetzt.
Die drei Punkte mit der jeweils höchsten Intensität aus der Summenfilteranalyse
aus den Kopfregionen und aus dem Mittelstück der Zellen in den Teilbildern in Abb.
4.3 (H) - (S) wurden zur weiteren Datenanalyse ausgewählt und sind in Abb. 4.5
45
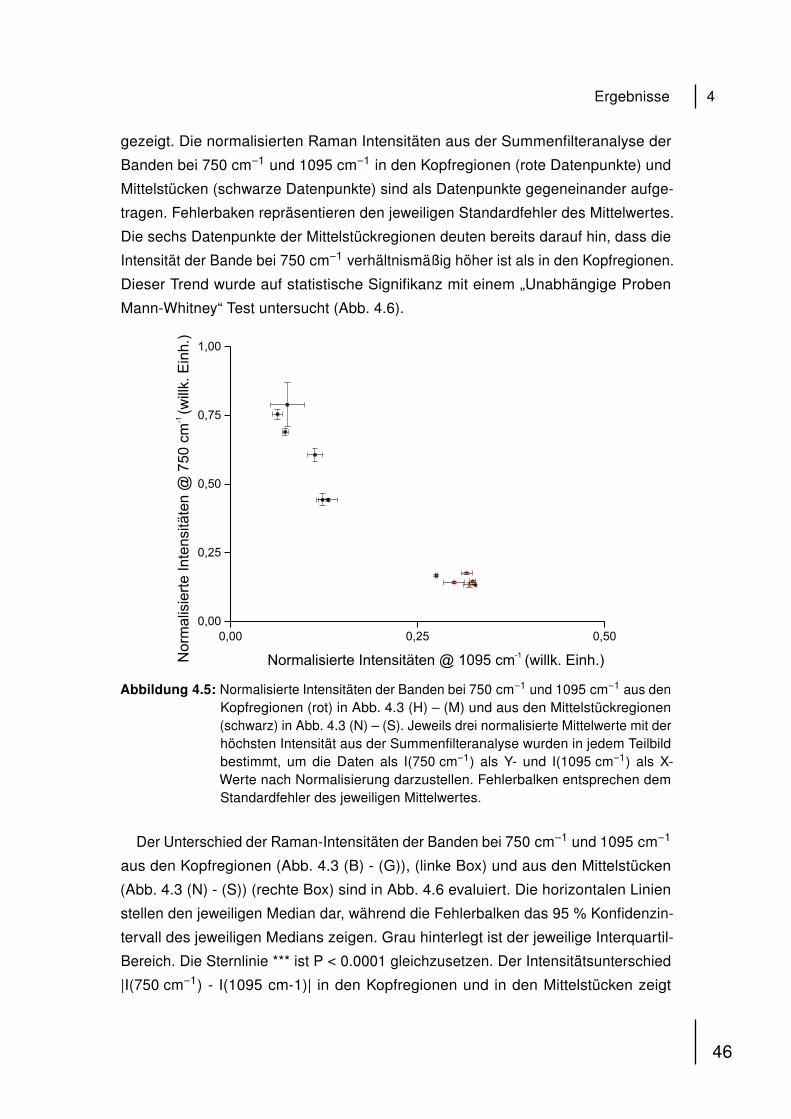
Ergebnisse 4
gezeigt. Die normalisierten Raman Intensitäten aus der Summenfilteranalyse der
Banden bei 750 cm−1 und 1095 cm−1 in den Kopfregionen (rote Datenpunkte) und
Mittelstücken (schwarze Datenpunkte) sind als Datenpunkte gegeneinander aufge-
tragen. Fehlerbaken repräsentieren den jeweiligen Standardfehler des Mittelwertes.
Die sechs Datenpunkte der Mittelstückregionen deuten bereits darauf hin, dass die
Intensität der Bande bei 750 cm−1 verhältnismäßig höher ist als in den Kopfregionen.
Dieser Trend wurde auf statistische Signifikanz mit einem „Unabhängige Proben
Mann-Whitney“ Test untersucht (Abb. 4.6).
0,00 0,25 0,50
0,00
0,25
0,50
0,75
1,00
Normalisierte Intensitäten @ 1095 cm-1
(willk. Einh.)Norm
alis
iert
e Inte
nsitäte
n @
750 c
m-1
(will
k. E
inh.)
Abbildung 4.5: Normalisierte Intensitäten der Banden bei 750 cm−1 und 1095 cm−1 aus denKopfregionen (rot) in Abb. 4.3 (H) – (M) und aus den Mittelstückregionen(schwarz) in Abb. 4.3 (N) – (S). Jeweils drei normalisierte Mittelwerte mit derhöchsten Intensität aus der Summenfilteranalyse wurden in jedem Teilbildbestimmt, um die Daten als I(750 cm−1) als Y- und I(1095 cm−1) als X-Werte nach Normalisierung darzustellen. Fehlerbalken entsprechen demStandardfehler des jeweiligen Mittelwertes.
Der Unterschied der Raman-Intensitäten der Banden bei 750 cm−1 und 1095 cm−1
aus den Kopfregionen (Abb. 4.3 (B) - (G)), (linke Box) und aus den Mittelstücken
(Abb. 4.3 (N) - (S)) (rechte Box) sind in Abb. 4.6 evaluiert. Die horizontalen Linien
stellen den jeweiligen Median dar, während die Fehlerbalken das 95 % Konfidenzin-
tervall des jeweiligen Medians zeigen. Grau hinterlegt ist der jeweilige Interquartil-
Bereich. Die Sternlinie *** ist P < 0.0001 gleichzusetzen. Der Intensitätsunterschied
|I(750 cm−1) - I(1095 cm-1)| in den Kopfregionen und in den Mittelstücken zeigt
46
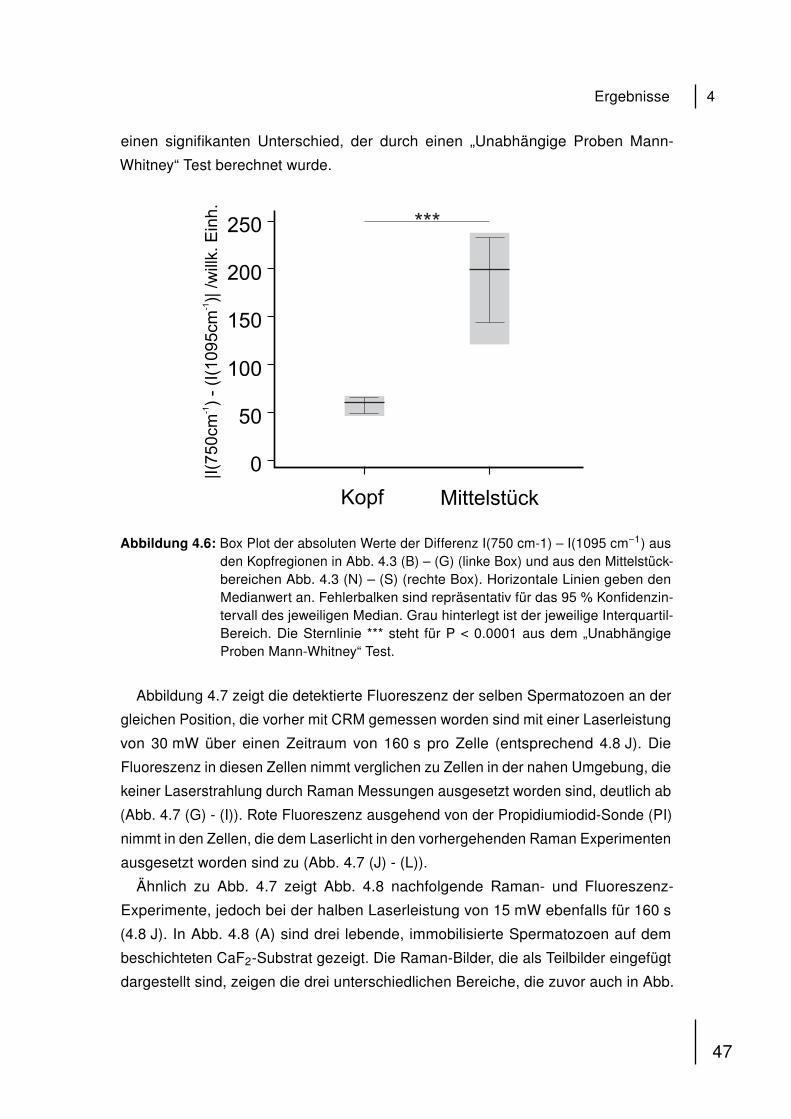
Ergebnisse 4
einen signifikanten Unterschied, der durch einen „Unabhängige Proben Mann-
Whitney“ Test berechnet wurde.
0
50
100
150
200
250
Kopf Mittelstück
***|I(7
50
cm
) -
(I(1
09
5cm
)| /
will
k.
Ein
h.
-1-1
Abbildung 4.6: Box Plot der absoluten Werte der Differenz I(750 cm-1) – I(1095 cm−1) ausden Kopfregionen in Abb. 4.3 (B) – (G) (linke Box) und aus den Mittelstück-bereichen Abb. 4.3 (N) – (S) (rechte Box). Horizontale Linien geben denMedianwert an. Fehlerbalken sind repräsentativ für das 95 % Konfidenzin-tervall des jeweiligen Median. Grau hinterlegt ist der jeweilige Interquartil-Bereich. Die Sternlinie *** steht für P < 0.0001 aus dem „UnabhängigeProben Mann-Whitney“ Test.
Abbildung 4.7 zeigt die detektierte Fluoreszenz der selben Spermatozoen an der
gleichen Position, die vorher mit CRM gemessen worden sind mit einer Laserleistung
von 30 mW über einen Zeitraum von 160 s pro Zelle (entsprechend 4.8 J). Die
Fluoreszenz in diesen Zellen nimmt verglichen zu Zellen in der nahen Umgebung, die
keiner Laserstrahlung durch Raman Messungen ausgesetzt worden sind, deutlich ab
(Abb. 4.7 (G) - (I)). Rote Fluoreszenz ausgehend von der Propidiumiodid-Sonde (PI)
nimmt in den Zellen, die dem Laserlicht in den vorhergehenden Raman Experimenten
ausgesetzt worden sind zu (Abb. 4.7 (J) - (L)).
Ähnlich zu Abb. 4.7 zeigt Abb. 4.8 nachfolgende Raman- und Fluoreszenz-
Experimente, jedoch bei der halben Laserleistung von 15 mW ebenfalls für 160 s
(4.8 J). In Abb. 4.8 (A) sind drei lebende, immobilisierte Spermatozoen auf dem
beschichteten CaF2-Substrat gezeigt. Die Raman-Bilder, die als Teilbilder eingefügt
dargestellt sind, zeigen die drei unterschiedlichen Bereiche, die zuvor auch in Abb.
47

Ergebnisse 4
A B C
D E F G H I J K L
Abbildung 4.7: Detektierte Fluoreszenzsignale der selben Spermien an der selben Positionnach Laserbestrahlung mit 30 mW. (A) Optisches Bild, das immobilisierteSpermien zeigt (Sichtfeld zum Vergleich zu Abb. 5.5). Vierecke in rot, grünund blau geben die Messbereiche an, in denen automatisierte CRM von ein-zelnen Spermien mit einer Laserleistung von 30 mW durchgeführt wordensind. (B) Spermien mit intakter Membran fluoreszieren hellgrün. (C) Zellenmit beschädigter Zellmembran fluoreszieren rot. (D) - (F) Digitale Zoomsder drei Spermatozoen von Interesse aus (A). (G) - (I) Digitale Zooms derSpermatozoen von Interesse aus (B). Das detektierte Fluoreszenzsignalder drei Spermien, die Laserstrahlung aus CRM ausgesetzt worden sind, istdeutlich geringer verglichen mit den Zellen in der Umgebung, die keinem La-serlicht ausgesetzt wurden. (J) - (L) Digitale Zooms der Spermatozoen vonInteresse aus (C). Rote Fluoreszenz konnte für die Spermien detektiert wer-den, die vorher mit CRM gemessen und damit Laserstrahlung ausgesetztworden sind.
4.3 gefunden worden sind (von links nach rechts: gesamte Zelle, Kopfregion und
Mittelstück). In Abb. 4.8 (B) ist ein ähnliches Sichtfeld dargestellt wie in Abb. 4.8 (A),
aufgenommen mit dem verwendeten Fluoreszenz-Mikroskop. Abb. 4.8 (C) zeigt das
SYBR-14 Fluoreszenzsignal in dem Bereich der Spermatozoen, der vorher schon
mit CRM gemessen worden ist (1), neben anderen Zellen (2),(3), die vorher nicht
durch Laserstrahlung aus CRM chemisch untersucht worden sind. Abb. 4.8 (D) zeigt
keine Fluoreszenz der PI-Sonde.
In Abb. 4.9 sind die farbkodierten Raman Bilder von 35 einzelnen Spermatozoen
gezeigt, die vollständig automatisiert gemessen worden sind. Die Farbskala ent-
spricht jener aus Abbildung 4.3. Jedes Teilbild einer Zelle ist aus 400 Datenpunkten,
die einzelnen Raman-Spektren entsprechen, zusammengesetzt. Die Geometrie der
Bilder beträgt dabei 13 µm x 13 µm mit einer Messzeit pro Spektrum von 0.15 s.
Diese Parameter resultieren in einem Durchsatz von ungefähr einer Zelle und einem
chemischen Bild pro 60 s mit einer lateralen Auflösung von 650 nm pro Pixel.
Um das erzielte Signal-zu-Rausch Verhältnis in jedem Teilbild von Abb. 4.9 zu
berechnen, wurde zunächst für jedes Teilbild der Punkt mit der höchsten Intensität
aus der Summenfilteranalyse für die Kohlenwasserstoffschwingungen bestimmt. In
48

Ergebnisse 4
A B C D
1
2
3
1
23
Abbildung 4.8: Raman-chemische Bilder und detektierte Fluoreszenz aus SYBR 14 undPI des selben Spermiums an der selben Position. (A) Optisches Bild,das immobilisierte Zellen darstellt. Viereck in Grün gibt den Messbereichan, in dem eine automatisierte CRM Messung mit einer Laserleistungvon 15 mW durchgeführt worden ist. Teilbilder sind die Raman chemi-schen Bilder rekonstruiert aus Summenfilteranalyse der Zelle markiertin (A) mit (1). Von links nach Rechts: Verschiedene CH-Schwingungen(2800 cm−1 bis 3000 cm−1), Kopfregion (1090 cm−1 bis 1100 cm−1) unddem Mittelstück (745 cm−1 bis 755 cm−1). Maßstabsbalken sind 2 µm. (C)Detektierte Fluoreszenz von SYBR-14. Das Spermatozoon (1), welchesvorher mit CRM gemessen worden ist erscheint als heller Punkt nebenanderen Zellen in der Umgebung (2),(3) die nicht vorher Laserstrahlung ausCRM Messungen ausgesetzt worden sind. (D) Kein Fluoreszenzsignal derPI-Sonde wurde detektiert.
jedem der so erhaltenen Raman-Spektren wurde daraufhin das Signal-zu-Rausch
(signal to noise, S/N) Verhältnis über Gleichung 4.2 berechnet. Die gemittelte In-
tensität in jedem Spektrum bei vB (spektraler Bereich ohne Signale, entsprechend
dem Hintergrundsignal, 2597 cm−1 bis 2603 cm−1) wurde dafür von der gemittelten
Intensität bei vCH (Kohlenwasserstoffbande, 2937 cm−1 bis 2943 cm−1) abgezogen
und durch das „Rauschen“ SB dividiert:
S/N =I(vCH)− I(vB)
SB(4.2)
mit
SB =
√1
N−1
N
∑i=1
(I(vi)− IB)2 (4.3)
und
IB =1N
N
∑i=1
I(vi) (4.4)
wobei vi zwischen 2597 cm−1 bis 2603 cm−1 variiert.
Abbildung 4.10 zeigt das S/N für jedes Teilbild in Abb. 4.9 und zeigt, dass das S/N
in einer laufenden automatisierten Messung von Teilbild zu Teilbild um einen Faktor
bis zu 2 schwanken kann.
Dieser Effekt wurde in mehreren unabhängigen automatisierten Messungen be-
49
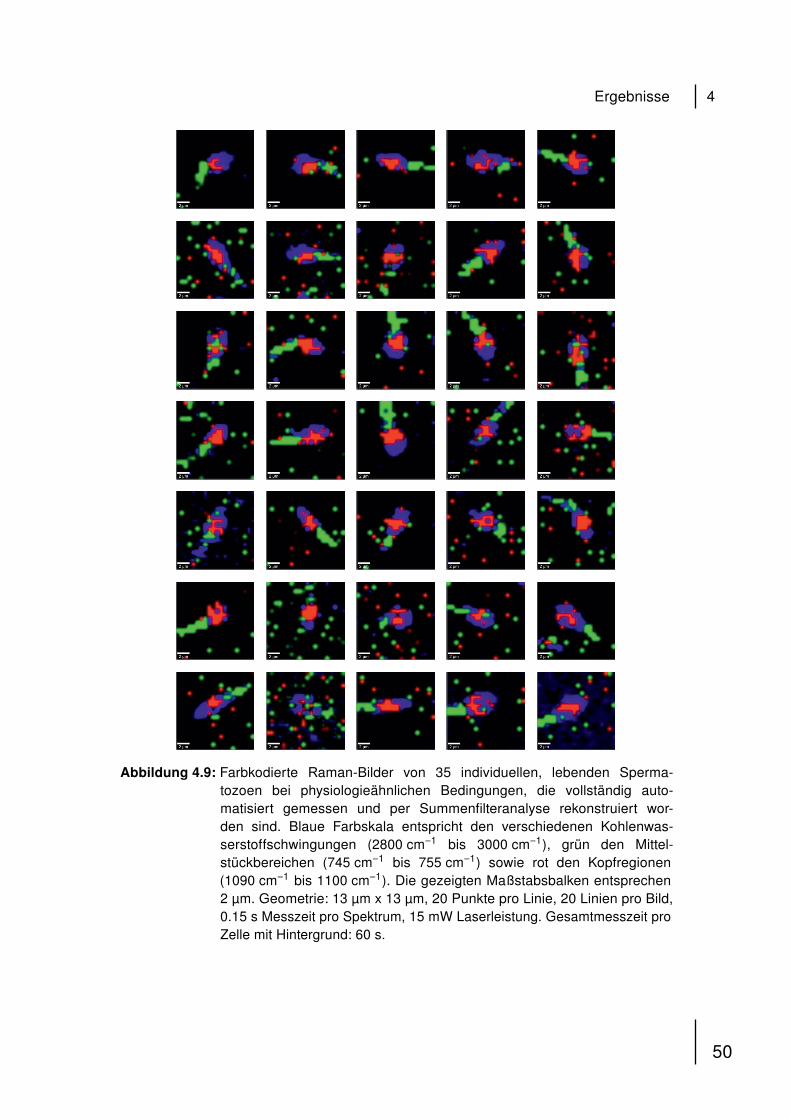
Ergebnisse 4
Abbildung 4.9: Farbkodierte Raman-Bilder von 35 individuellen, lebenden Sperma-tozoen bei physiologieähnlichen Bedingungen, die vollständig auto-matisiert gemessen und per Summenfilteranalyse rekonstruiert wor-den sind. Blaue Farbskala entspricht den verschiedenen Kohlenwas-serstoffschwingungen (2800 cm−1 bis 3000 cm−1), grün den Mittel-stückbereichen (745 cm−1 bis 755 cm−1) sowie rot den Kopfregionen(1090 cm−1 bis 1100 cm−1). Die gezeigten Maßstabsbalken entsprechen2 µm. Geometrie: 13 µm x 13 µm, 20 Punkte pro Linie, 20 Linien pro Bild,0.15 s Messzeit pro Spektrum, 15 mW Laserleistung. Gesamtmesszeit proZelle mit Hintergrund: 60 s.
50
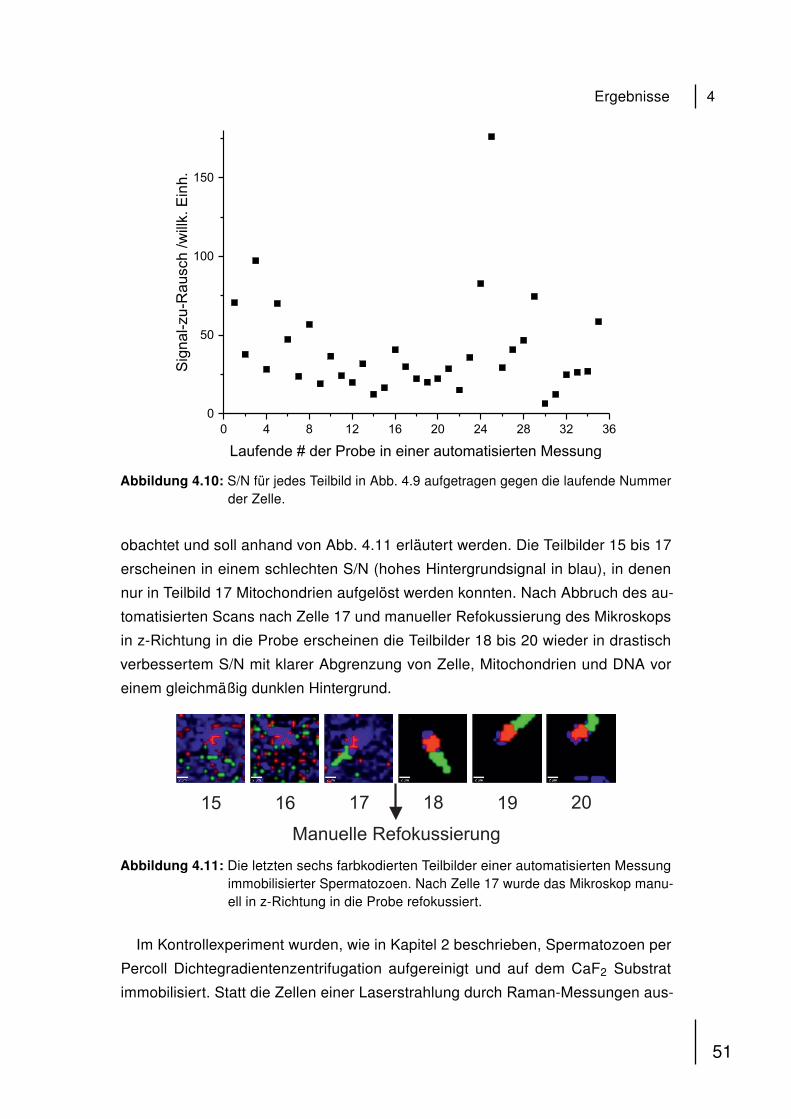
Ergebnisse 4
0 4 8 12 16 20 24 28 32 36
0
50
100
150S
ign
al-
zu
-Ra
usch
/will
k.E
inh
.
Laufende # der Probe in einer automatisierten Messung
Abbildung 4.10: S/N für jedes Teilbild in Abb. 4.9 aufgetragen gegen die laufende Nummerder Zelle.
obachtet und soll anhand von Abb. 4.11 erläutert werden. Die Teilbilder 15 bis 17
erscheinen in einem schlechten S/N (hohes Hintergrundsignal in blau), in denen
nur in Teilbild 17 Mitochondrien aufgelöst werden konnten. Nach Abbruch des au-
tomatisierten Scans nach Zelle 17 und manueller Refokussierung des Mikroskops
in z-Richtung in die Probe erscheinen die Teilbilder 18 bis 20 wieder in drastisch
verbessertem S/N mit klarer Abgrenzung von Zelle, Mitochondrien und DNA vor
einem gleichmäßig dunklen Hintergrund.
15 16 17 18 19 20
Manuelle Refokussierung
Abbildung 4.11: Die letzten sechs farbkodierten Teilbilder einer automatisierten Messungimmobilisierter Spermatozoen. Nach Zelle 17 wurde das Mikroskop manu-ell in z-Richtung in die Probe refokussiert.
Im Kontrollexperiment wurden, wie in Kapitel 2 beschrieben, Spermatozoen per
Percoll Dichtegradientenzentrifugation aufgereinigt und auf dem CaF2 Substrat
immobilisiert. Statt die Zellen einer Laserstrahlung durch Raman-Messungen aus-
51

Ergebnisse 4
zusetzen wurden die Spermatozoen für 4 h bei 35 ◦C temperiert und anschließend
analog zu den bereits gezeigten Zellen in Abb. 4.7 und Abb. 4.8 einem Viabilitätstest
per LIVE/DEAD Sperm Viability Kit unterzogen. In Abb. 4.12 ist das Ergebnis in zwei
Teilbildern dargestellt. Abbildung 4.12 (A) zeigt die SYBR-14 Fluoreszenz nach 4 h
Lagerung bei 35 ◦C, während Teilabbildung (B) die Fluoreszenz der PI-Sonde zeigt.
In Teilabbildung (A) ist ein Fluoreszenz-Signal zu erkennen, während in Teilabbildung
(B) kein Fluoreszenz-Signal der PI-Sonde detektiert werden konnte.
A B
Abbildung 4.12: Detektierte SYBR-14 und PI Fluoreszenzsignale von Spermatozoen nachPercoll-Dichtegradienten-Zentrifugation, Immobilisierung und Lagerung für4 h bei 35 ◦C. (A) Fluoreszenzsignal des SYBR-14/DNA Komplexes zeigtmehrere helle Punkte. (B) Ein Fluoreszenz-Signal des PI/SYYBR-14/DNAKomplexes wurde nicht detektiert.
Im zweiten Kontrollversuch wurde die Motilität der Geissel der zu immobilisie-
renden Spermatozoen bei verschiedenen Zeitpunkten nach Transfer auf das CaF2
Substrat beobachtet. Abb. 4.13 zeigt die Position der Geissel zum Zeitpunkt t = 0 s
kurz nach Transfer auf das CaF2 Substrat. Während der Kopf sofort immobilisert ist,
variiert die Position der Geissel in einer Zeitskala von Sekunden bis zur kompletten
Immobilisierung.
52
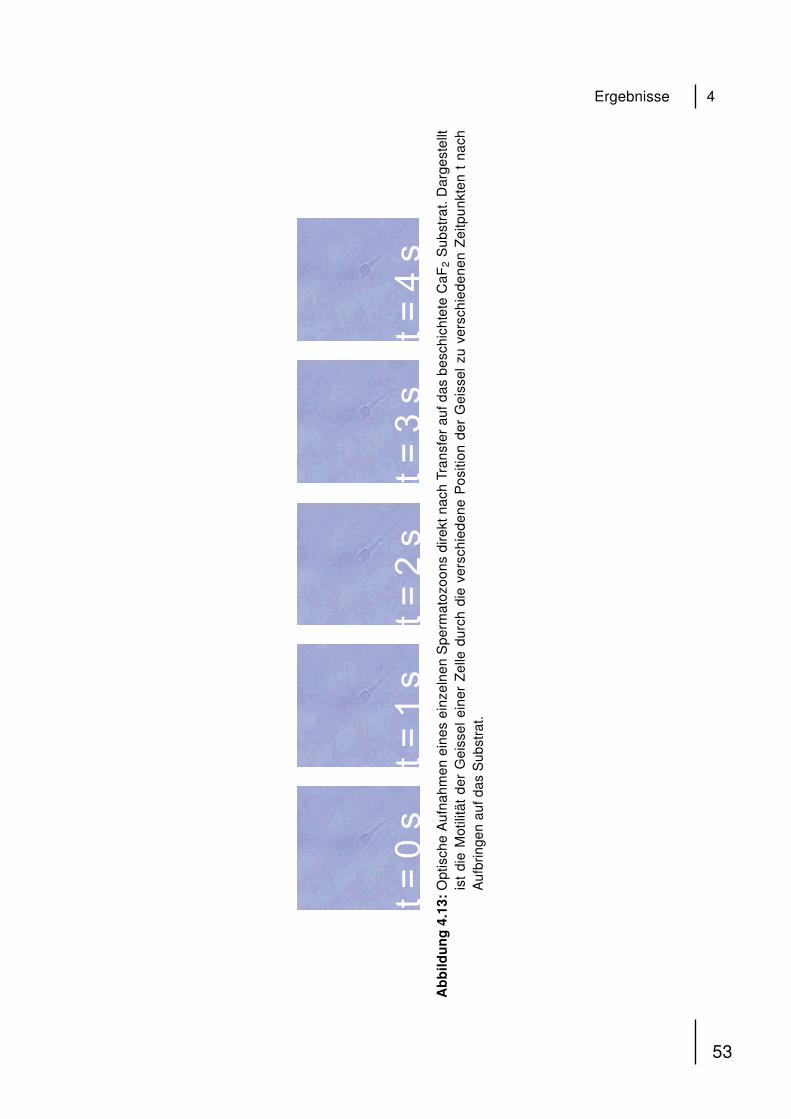
Ergebnisse 4
t =
0 s
t =
1 s
t =
2 s
t =
3 s
t =
4 s
Ab
bild
un
g4.
13:
Opt
isch
eA
ufna
hmen
eine
sei
nzel
nen
Spe
rmat
ozoo
nsdi
rekt
nach
Tran
sfer
aufd
asbe
schi
chte
teC
aF2
Sub
stra
t.D
arge
stel
ltis
tdi
eM
otili
tät
der
Gei
ssel
eine
rZ
elle
durc
hdi
eve
rsch
iede
neP
ositi
onde
rG
eiss
elzu
vers
chie
dene
nZ
eitp
unkt
ent
nach
Auf
brin
gen
aufd
asS
ubst
rat.
53

Diskussion und Ausblick 4
4.3 Diskussion und Ausblick
Die hyperspektralen Datensätze, die verwendet wurden, um die chemischen Bilder
in Abb. 4.3 durch Summenfilteranalyse zu erzeugen zeigen mehrere prominente
Banden unterschiedlicher chemischer Spezies (siehe Durchschnitts-Spektren in Abb.
4.4). Die Spektren, die in den Kopfregionen in Abb. 4.3 (H) - (M) aufgenommen
worden sind, zeigen Banden, die durch Vibrationen des DNA Phosphat Rückgrates
entstehen (1095 cm−1 [41]). Eine Bande bei 750 cm−1 kann sowohl im Mittelstück
als auch in den Kopfregionen der untersuchten lebenden Zellen gefunden werden
(Abb. 4.3 (N) - (S)).
Um zu evaluieren, ob die in dieser Arbeit verwendete Summenfilteranalyse in der
Lage ist eine mögliche Ambiguität mit statistischer Signifikanz aufzulösen, wurden
in jeder Region in jedem Teilbild die drei Pixel mit der höchsten Intensität der
Bande bei 750 cm−1 bzw. bei 1095 cm−1 ausgewählt. Werden die normalisierten
Intensitäten beider Banden in den Kopf- und in den Mittelstückregionen miteinander
verglichen, indem sie gegeneinander aufgetragen werden, ist bereits deutlich, dass
die Intensität der Bande bei 750 cm−1 verglichen zu der Bande bei 1095 cm−1 höher
in den Mittelstückbereichen ist als in den Kopfregionen (Abb. 4.5).
Dieser beobachtete Unterschied wurde auf statistische Signifikanz untersucht,
indem die absoluten Werte von I(750 cm−1) - I(1095 cm−1) in den Kopf- und Mit-
telstückbereichen in Abb. 4.3 (H) - (S) berechnet und über einen „Unabhängige
Proben Mann-Whitney“ Test miteinander verglichen wurden. Die Ergebnisse, die
in dem Box-Whisker Plot in Abb. 4.6 dargestellt sind, zeigen mit einer statistischen
Signifikanz von P < 0.0001, dass die Peak-zu-Peak Intensitätsdifferenz beider Ban-
den zu Gunsten der Bande bei 750 cm−1 höher in den Mittelstückbereichen ist, als
in den Kopfregionen der untersuchten Zellen. Als Schlussfolgerung dieses Ergeb-
nisses kann angenommen werden, dass obwohl die Bande bei 750 cm−1 sowohl
in den Spektren der Kopf- als auch der Mittelstücke der Spermatozoen gefunden
werden kann, der Ursprung der Bande von vollständig unterschiedlichen chemi-
schen Strukturen herrühren muss, die in den beiden untersuchten Bereichen der
Spermatozoen auftauchen. Die Bande bei 750 cm−1 wird folglich in dieser Arbeit
mitochondrialem Cytochrom c zugeordnet, während der Ursprung der Bande in den
Kopfbereichen der Nukleobase Thymin zugewiesen wird [41]. Diese Zuweisung ist
in guter Übereinstimmung mit früheren Studien, in denen CRM verwendet wurde,
um mitochondriale Cytochome zu untersuchen, zum Beispiel in HeLa-Zellen oder
Kardiomyozyten ([74, 75]). Die in dieser Arbeit verwendete Summenfilteranalyse
54

Diskussion und Ausblick 4
erlaubt die Visualisierung von Mitochondrien über die Integration der Bande bei
750 cm−1 in einer automatisierten Routine mit statistischer Signifikanz.
Spermatozoen inklusive ihrer DNA und der enthaltenen Mitochondrien können
mit CRM bei physiologieähnlichen Bedingungen per Summenfilteranalyse visuali-
siert werden. Der folgende Abschnitt behandelt die Diskussion möglicher negativer
Folgen, Spermatozoen Laserlicht in Raman-Messungen auszusetzen, zum Beispiel
Membranschädigungen oder nukleäre DNA Fragmentierung.
Viabilität und damit Fruchtbarkeit von Spermatozoen wurde per LIVE/DEAD Sperm
Viability Kit direkt im Anschluss an Raman-Messungen untersucht. Grundsätzlich
sind die Spermatozoenmembranen durchlässig für den Fluoreszenzfarbstoff SYBR-
14, was in einem detektierbaren (grünen) Fluoreszenzsignal in unbeschädigten
Zellen resultiert. Die Zugabe von PI führt zu drastisch reduzierter Fluoreszenz von
SYBR-14 in beschädigten Zellen, was letztlich zu einem rotorangenen Fluoreszenzsi-
gnal führt. PI ist in der Lage nur in Spermatozoen einzudringen, deren Zellmembran
beschädigt ist und formt dort einen SYBR-14/PI/DNA-Komplex und quencht dadurch
per Fluoreszenz-Resonanz-Energie-Transfer die Fluoreszenz des SYBR-14 ([76]).
Die Ergebnisse in Abb. 4.7 zeigen, dass eine Laserleistung von 30 mW in Verbin-
dung mit Messzeiten von 160 s pro Zelle bereits zu Schäden in der Zellmembran
führt. Dies ist durch die rote Fluoreszenz, detektiert in Abb. 4.7 (J) - (L) in den Zellen
illustriert, die Laserstrahlung während einer Raman-Messung mit einer Leistung von
30 mW ausgesetzt waren.
CRM-Experimente sind daher mit einer geringeren Laserleistung von 15 mW
wiederholt worden (Abb. 4.8). Die Fluoreszenz von SYBR-14 (s. Abb. 4.8 (C))
zeigt, dass die Raman-Messung keine schädliche Wirkung durch Bestrahlung der
Zellen mit Laserlicht zur Folge hatte. Dies ist weiter durch die Kontrolle des nicht
vorhandenen Fluoreszenz-Signals der PI-Sonde in Abb. 4.8 (D) unterstützt.
Der Diskussion folgend, wie chemische Bilder von einzelnen, lebenden Spermato-
zoen mit CRM unter Verwendung von 15 mW auf eine nicht-invasive Art und Weise
unter physiologieähnlichen Bedingungen gewonnen werden können, behandelt der
abschließende Teil der vorliegenden Arbeit die Optimierung der automatisierten Mes-
sungen. Chemische Bilder von 35 einzelnen, immobilisierten Spermatozoen wurden
über eine automatisierte Messmethode erstellt (Abb. 4.9). Die dafür verwendete
Summenfilteranalyse läuft dabei in Echtzeit während der Messungen. In mehreren
automatisierten Messungen wurden Variationen des S/N Verhältnisses beobachtet
und manuell durch Anpassen der z-Position des Raman-Mikroskops verbessert, da
so eine mögliche Schieflage des CaF2 korrigiert wurde. Während die chemische
55

Diskussion und Ausblick 4
Komposition (verschiedene Kohlenhydrate, DNA, Cytochrom c in Mitochondrien)
generell mit einem guten Signal-zu-Rausch-Verhältnis visualisiert werden konnte,
konnten DNA und Mitochondrien in zwei Fällen nicht aufgelöst werden (Abb. 4.9,
Zelle Nummer 27 und Nummer 32). Dies kann dadurch erklärt werden, dass die
Zellen in diesen Fällen nicht in der gleichen Position in z-Richtung lagen, was zu
einer Verschlechterung des Signal-zu-Rausch Verhältnisses führt, da ein konfokales
Mikroskopiesystem für die automatisierte Messung verwendet wurde. Dies ist ferner
mit dem schwankenden S/N in einer Anzahl von 35 automatisiert gemessenen
Spermatozoen in Abb.4.10 dargestellt. Es wurde zusätzlich nachgewiesen, dass die
z-Fokussierung Ursache der Verschlechterung des S/N ist, indem eine automatische
Messung vor Messungen der letzten drei Spermatozoen unterbrochen wurde, und
manuell in z-Richtung optimiert wurde, bevor die letzten drei Spermatozoen gemes-
sen wurden. Eine deutliche Verbesserung im S/N wurde für die letzten drei Zellen
erreicht, erkennbar im gleichmäßig dunklen Hintergrund und guten Kontrast (s. Abb.
4.11). Dies kann in zukünftigen Studien dadurch vermieden verwenden, dass ein
„automatischer Fokus“ in die Routine aufgenommen wird, der in einer Zeitskala von
ms pro untersuchter Zelle durchgeführt werden kann. Hierbei wird die integrierte
Intensität einer frei wählbaren Bande im Raman-Spektrum in 50 äquidistanten Punk-
ten der Probe in z-Richtung gemessen. Das Mikroskop wird daraufhin automatisch
zu der z-Position gefahren, in der das maximale Signal der integrierten Intensität
gefunden worden ist.
Die vorliegende Arbeit zeigt zum ersten mal, wie die selben lebenden Spermato-
zoen mit automatisierter CRM unter physiologieähnlichen Bedingungen chemisch
untersucht und anschließend mit Fluoreszenz-Mikroskopie die nicht-invasive Natur
der CRM Messungen validiert wurde. Die hier gezeigte neuartige Kombination von
Substrat und Beschichtung ermöglichte die automatisierte Messung einzelner Zellen
in einer Zeitskala von ca. 60 s pro Zelle. Diese Ergebnisse ermöglichen es in zukünf-
tigen Studien eine Vielfalt an unterschiedlichen Parametern bei physiologieähnlichen
Bedingungen zu untersuchen, zum Beispiel den Einfluss oxidativen Schadens, der
Bestrahlung mit UV Photonen oder einer bakteriellen Infektion auf die nukleäre DNA
von lebenden Spermatozoen. Des Weiteren zeigt die hier vorgestellte Methode das
Potential, um nach weiterer stringenter Evaluierung in eine Routine eingearbeitet zu
werden, die nicht-invasiv individuelle, chemisch unbeschädigte Spermatozoen für
die ART auswählt.
56


5Anwendungen konfokaler
Raman-Spektroskopie
Dieses Kapitel beschreibt die Anwendung konfokaler Raman-Spektroskopie im
Rahmen interdisziplinärer Forschung anhand zweier Beispiele aus dem Bereich
anorganischer Chemie. Für Raman-Spektroskopie ist von besonderer Bedeutung
geeignete Substrate auszuwählen, um die Zuordnung von Raman-Banden zu er-
möglichen. Dies ist hier im ersten Beispiel anhand der Substrate Silizium und Glas
gezeigt (vgl. auch Kap. 4, CaF2 als besonders geeignetes Substrat für biologische
Proben). Schichtdicken der untersuchten Dünnschichten von (100±10) nm reichen
bereits für die Detektion von Raman-Banden aus, verglichen zum Beispiel mit den
in Kapitel 4 diskutierten Geisseln menschlicher Spermatozoen mit einem typischen
Durchmesser von 500 nm.
Der Einbau eines Peltier-Thermostats PE 120 (Linkam, Surrey, England) in das
konfokale Raman-Mikrospektroskop erlaubte temperaturabhängige in-situ Mes-
sungen des Hybridmaterials aus Koordinationspolymer und ionischer Flüssigkeit
(C4C1py)[Cu(SCN)2] als Feststoff und Schmelze. Die hier gezeigten Beiträge sind in
[13] und [14] publiziert und zeigen wie die in Tabelle 1.1 genannten Eigenschaften
mit Hilfe konfokaler Raman-Spektroskopie chemisch untersucht und identifiziert
werden können.
58
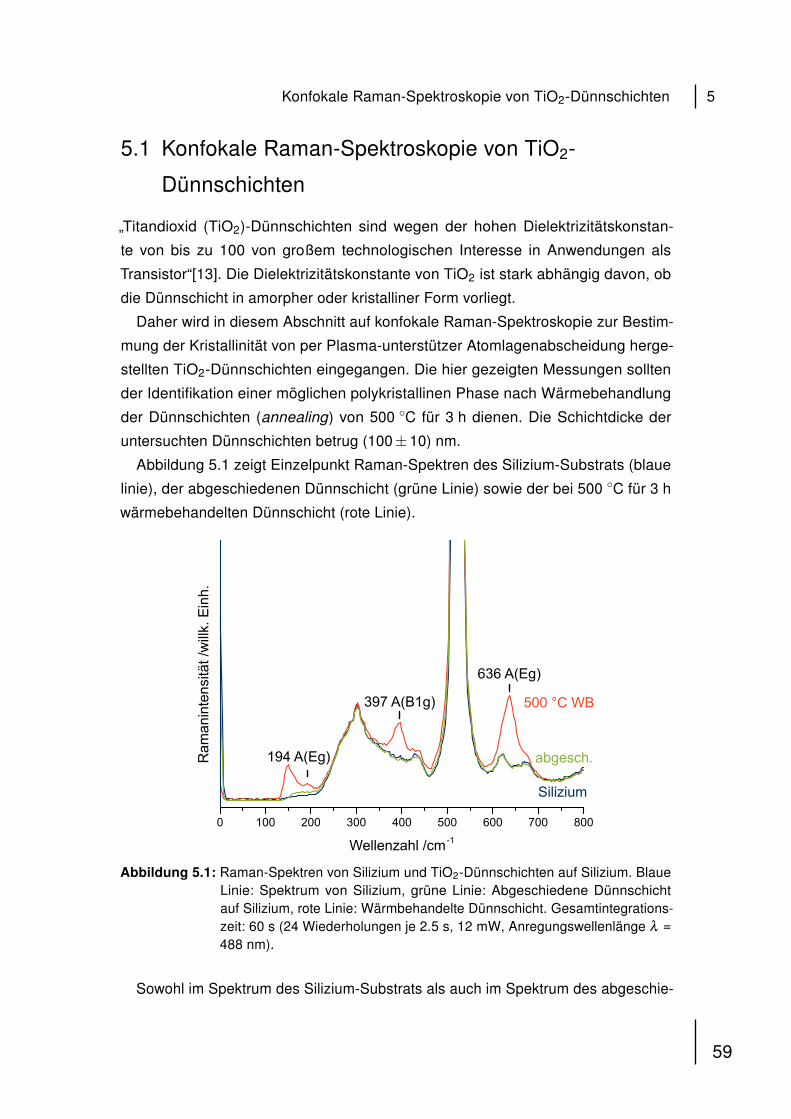
Konfokale Raman-Spektroskopie von TiO2-Dünnschichten 5
5.1 Konfokale Raman-Spektroskopie von TiO2-
Dünnschichten
„Titandioxid (TiO2)-Dünnschichten sind wegen der hohen Dielektrizitätskonstan-
te von bis zu 100 von großem technologischen Interesse in Anwendungen als
Transistor“[13]. Die Dielektrizitätskonstante von TiO2 ist stark abhängig davon, ob
die Dünnschicht in amorpher oder kristalliner Form vorliegt.
Daher wird in diesem Abschnitt auf konfokale Raman-Spektroskopie zur Bestim-
mung der Kristallinität von per Plasma-unterstützer Atomlagenabscheidung herge-
stellten TiO2-Dünnschichten eingegangen. Die hier gezeigten Messungen sollten
der Identifikation einer möglichen polykristallinen Phase nach Wärmebehandlung
der Dünnschichten (annealing) von 500 ◦C für 3 h dienen. Die Schichtdicke der
untersuchten Dünnschichten betrug (100±10) nm.
Abbildung 5.1 zeigt Einzelpunkt Raman-Spektren des Silizium-Substrats (blaue
linie), der abgeschiedenen Dünnschicht (grüne Linie) sowie der bei 500 ◦C für 3 h
wärmebehandelten Dünnschicht (rote Linie).
0 100 200 300 400 500 600 700 800
Ra
ma
nin
ten
sitä
t /w
illk.
Ein
h.
Wellenzahl /cm-1
500 °C WB
abgesch.
Silizium
194 A(Eg)
397 A(B1g)
636 A(Eg)
Abbildung 5.1: Raman-Spektren von Silizium und TiO2-Dünnschichten auf Silizium. BlaueLinie: Spektrum von Silizium, grüne Linie: Abgeschiedene Dünnschichtauf Silizium, rote Linie: Wärmbehandelte Dünnschicht. Gesamtintegrations-zeit: 60 s (24 Wiederholungen je 2.5 s, 12 mW, Anregungswellenlänge λ =488 nm).
Sowohl im Spektrum des Silizium-Substrats als auch im Spektrum des abgeschie-
59
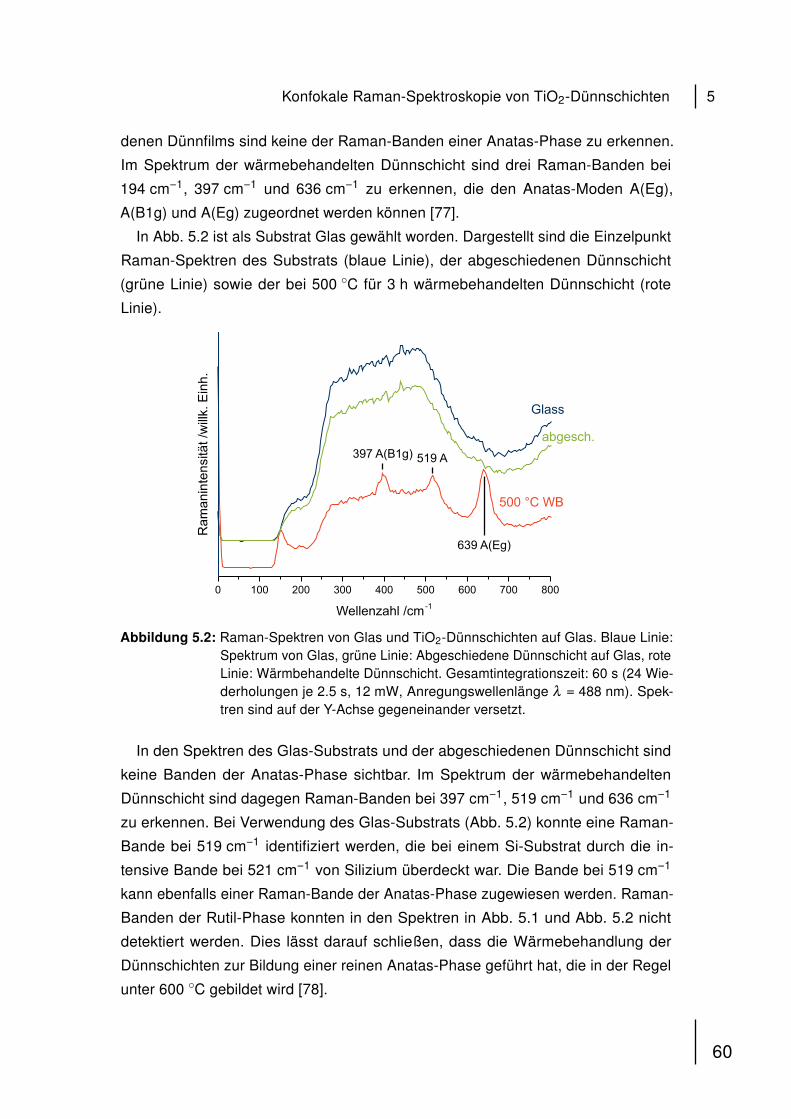
Konfokale Raman-Spektroskopie von TiO2-Dünnschichten 5
denen Dünnfilms sind keine der Raman-Banden einer Anatas-Phase zu erkennen.
Im Spektrum der wärmebehandelten Dünnschicht sind drei Raman-Banden bei
194 cm−1, 397 cm−1 und 636 cm−1 zu erkennen, die den Anatas-Moden A(Eg),
A(B1g) und A(Eg) zugeordnet werden können [77].
In Abb. 5.2 ist als Substrat Glas gewählt worden. Dargestellt sind die Einzelpunkt
Raman-Spektren des Substrats (blaue Linie), der abgeschiedenen Dünnschicht
(grüne Linie) sowie der bei 500 ◦C für 3 h wärmebehandelten Dünnschicht (rote
Linie).
0 100 200 300 400 500 600 700 800
Ra
ma
nin
ten
sitä
t/w
illk.E
inh
.
abgesch.
Glass
Wellenzahl /cm-1
500 °C WB
397 A(B1g) 519 A
639 A(Eg)
Abbildung 5.2: Raman-Spektren von Glas und TiO2-Dünnschichten auf Glas. Blaue Linie:Spektrum von Glas, grüne Linie: Abgeschiedene Dünnschicht auf Glas, roteLinie: Wärmbehandelte Dünnschicht. Gesamtintegrationszeit: 60 s (24 Wie-derholungen je 2.5 s, 12 mW, Anregungswellenlänge λ = 488 nm). Spek-tren sind auf der Y-Achse gegeneinander versetzt.
In den Spektren des Glas-Substrats und der abgeschiedenen Dünnschicht sind
keine Banden der Anatas-Phase sichtbar. Im Spektrum der wärmebehandelten
Dünnschicht sind dagegen Raman-Banden bei 397 cm−1, 519 cm−1 und 636 cm−1
zu erkennen. Bei Verwendung des Glas-Substrats (Abb. 5.2) konnte eine Raman-
Bande bei 519 cm−1 identifiziert werden, die bei einem Si-Substrat durch die in-
tensive Bande bei 521 cm−1 von Silizium überdeckt war. Die Bande bei 519 cm−1
kann ebenfalls einer Raman-Bande der Anatas-Phase zugewiesen werden. Raman-
Banden der Rutil-Phase konnten in den Spektren in Abb. 5.1 und Abb. 5.2 nicht
detektiert werden. Dies lässt darauf schließen, dass die Wärmebehandlung der
Dünnschichten zur Bildung einer reinen Anatas-Phase geführt hat, die in der Regel
unter 600 ◦C gebildet wird [78].
60
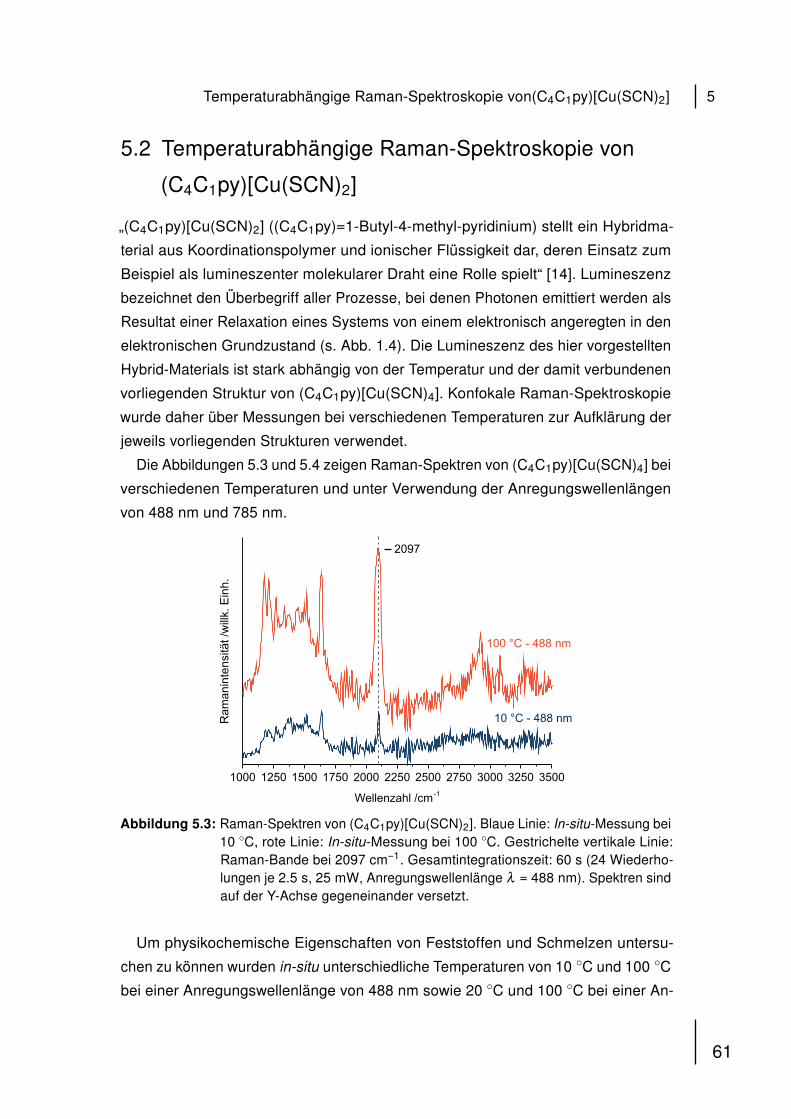
Temperaturabhängige Raman-Spektroskopie von(C4C1py)[Cu(SCN)2] 5
5.2 Temperaturabhängige Raman-Spektroskopie von
(C4C1py)[Cu(SCN)2]
„(C4C1py)[Cu(SCN)2] ((C4C1py)=1-Butyl-4-methyl-pyridinium) stellt ein Hybridma-
terial aus Koordinationspolymer und ionischer Flüssigkeit dar, deren Einsatz zum
Beispiel als lumineszenter molekularer Draht eine Rolle spielt“ [14]. Lumineszenz
bezeichnet den Überbegriff aller Prozesse, bei denen Photonen emittiert werden als
Resultat einer Relaxation eines Systems von einem elektronisch angeregten in den
elektronischen Grundzustand (s. Abb. 1.4). Die Lumineszenz des hier vorgestellten
Hybrid-Materials ist stark abhängig von der Temperatur und der damit verbundenen
vorliegenden Struktur von (C4C1py)[Cu(SCN)4]. Konfokale Raman-Spektroskopie
wurde daher über Messungen bei verschiedenen Temperaturen zur Aufklärung der
jeweils vorliegenden Strukturen verwendet.
Die Abbildungen 5.3 und 5.4 zeigen Raman-Spektren von (C4C1py)[Cu(SCN)4] bei
verschiedenen Temperaturen und unter Verwendung der Anregungswellenlängen
von 488 nm und 785 nm.
1000 1250 1500 1750 2000 2250 2500 2750 3000 3250 3500
100 °C - 488 nm
10 °C - 488 nmRa
ma
nin
ten
sitä
t/w
illk
.E
inh
.
Wellenzahl /cm-1
2097
Abbildung 5.3: Raman-Spektren von (C4C1py)[Cu(SCN)2]. Blaue Linie: In-situ-Messung bei10 ◦C, rote Linie: In-situ-Messung bei 100 ◦C. Gestrichelte vertikale Linie:Raman-Bande bei 2097 cm−1. Gesamtintegrationszeit: 60 s (24 Wiederho-lungen je 2.5 s, 25 mW, Anregungswellenlänge λ = 488 nm). Spektren sindauf der Y-Achse gegeneinander versetzt.
Um physikochemische Eigenschaften von Feststoffen und Schmelzen untersu-
chen zu können wurden in-situ unterschiedliche Temperaturen von 10 ◦C und 100 ◦C
bei einer Anregungswellenlänge von 488 nm sowie 20 ◦C und 100 ◦C bei einer An-
61
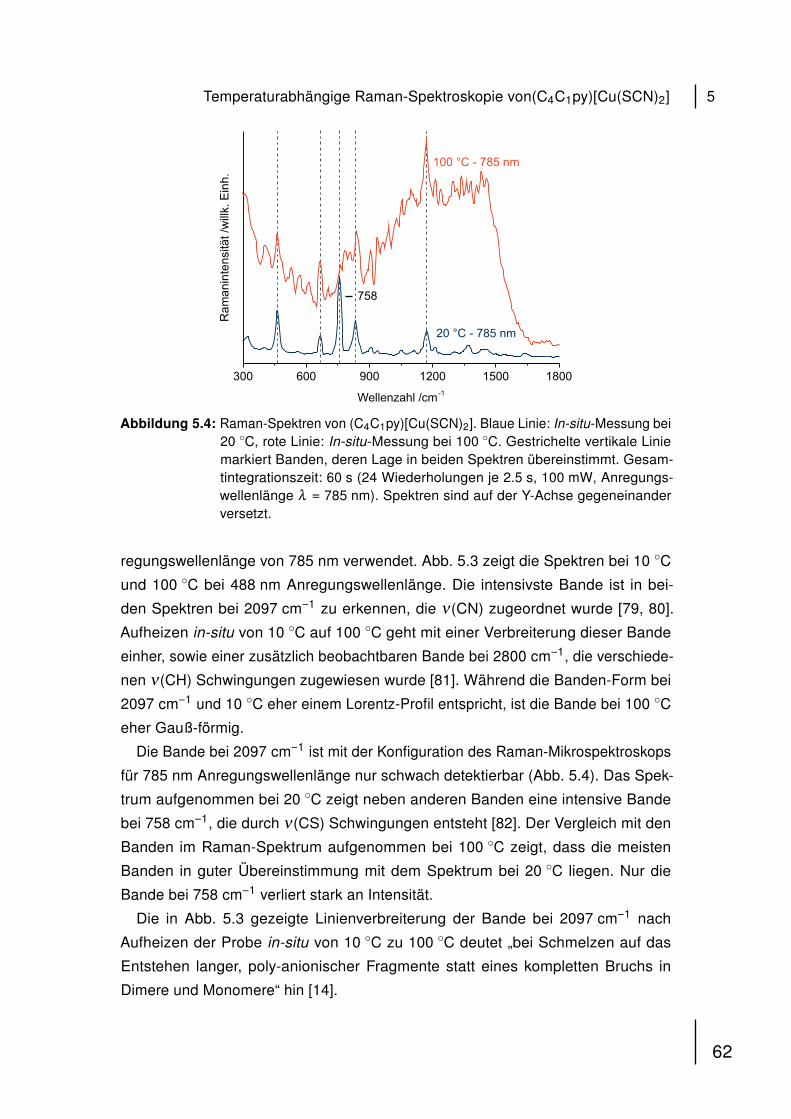
Temperaturabhängige Raman-Spektroskopie von(C4C1py)[Cu(SCN)2] 5
300 600 900 1200 1500 1800
Ra
ma
nin
ten
sitä
t/w
illk
.E
inh
.
Wellenzahl /cm-1
100 °C - 785 nm
20 °C - 785 nm
758
Abbildung 5.4: Raman-Spektren von (C4C1py)[Cu(SCN)2]. Blaue Linie: In-situ-Messung bei20 ◦C, rote Linie: In-situ-Messung bei 100 ◦C. Gestrichelte vertikale Liniemarkiert Banden, deren Lage in beiden Spektren übereinstimmt. Gesam-tintegrationszeit: 60 s (24 Wiederholungen je 2.5 s, 100 mW, Anregungs-wellenlänge λ = 785 nm). Spektren sind auf der Y-Achse gegeneinanderversetzt.
regungswellenlänge von 785 nm verwendet. Abb. 5.3 zeigt die Spektren bei 10 ◦C
und 100 ◦C bei 488 nm Anregungswellenlänge. Die intensivste Bande ist in bei-
den Spektren bei 2097 cm−1 zu erkennen, die ν(CN) zugeordnet wurde [79, 80].
Aufheizen in-situ von 10 ◦C auf 100 ◦C geht mit einer Verbreiterung dieser Bande
einher, sowie einer zusätzlich beobachtbaren Bande bei 2800 cm−1, die verschiede-
nen ν(CH) Schwingungen zugewiesen wurde [81]. Während die Banden-Form bei
2097 cm−1 und 10 ◦C eher einem Lorentz-Profil entspricht, ist die Bande bei 100 ◦C
eher Gauß-förmig.
Die Bande bei 2097 cm−1 ist mit der Konfiguration des Raman-Mikrospektroskops
für 785 nm Anregungswellenlänge nur schwach detektierbar (Abb. 5.4). Das Spek-
trum aufgenommen bei 20 ◦C zeigt neben anderen Banden eine intensive Bande
bei 758 cm−1, die durch ν(CS) Schwingungen entsteht [82]. Der Vergleich mit den
Banden im Raman-Spektrum aufgenommen bei 100 ◦C zeigt, dass die meisten
Banden in guter Übereinstimmung mit dem Spektrum bei 20 ◦C liegen. Nur die
Bande bei 758 cm−1 verliert stark an Intensität.
Die in Abb. 5.3 gezeigte Linienverbreiterung der Bande bei 2097 cm−1 nach
Aufheizen der Probe in-situ von 10 ◦C zu 100 ◦C deutet „bei Schmelzen auf das
Entstehen langer, poly-anionischer Fragmente statt eines kompletten Bruchs in
Dimere und Monomere“ hin [14].
62


Anhang
Abbildung 5.5: Optisches Bild aufgenommen mit dem konfokalen Raman-Mikroskop zeigteinzelne, immobilisierte, lebende Spermatozoen (Vergleich mit Abb. 4.7).Blaue, rote und grüne Vierecke geben Bereiche an, in denen automatisierteCRM Messungen durchgeführt worden sind mit einer Laserleistung von30 mW.
64


6Abbildungs- & Tabellenverzeichnis
66

Abbildungsverzeichnis
1.1 Erläuterung des Raman Effekts (- - - steht für ein virtuelles Energieni-
veau). Adaptiert nach [3]. . . . . . . . . . . . . . . . . . . . . . . . . 2
1.2 Schematisches Raman-Spektrum. . . . . . . . . . . . . . . . . . . . 6
1.3 Schematischer Aufbau und Strahlengang eines Konfokalmikroskops.
Adaptiert nach [6]. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
1.4 Prinzip der Anregung und Emission der Fluoreszenz. Adaptiert aus [27]. 11
1.5 Schematischer Aufbau von DNA, dargestellt im Solvens Wasser (ro-
te Punkte): Die beiden aüßeren Bänder symbolisieren die beiden
Zucker-Phosphat Rückgrate, während die davon horizontal abgehen-
den Stufen Basenpaare darstellen, die die Stränge über Wasserstoff-
brückenbindungen zusammenhalten. Struktur aus PDB-Eintrag 1BNA
[32]. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
2.1 Schematischer Aufbau des Raman-Mikroskops alpha300R (WITec,
Ulm). Der Strahlengang ist in grün angedeutet. Adaptiert nach [6]. . . 17
3.1 Schema der drei µAPPJs, die in dieser Arbeit verwendet wurden, um
das Bio-Makromolekül DNA zu behandeln. Entnommen aus [38]. . . 24
3.2 Raman-Spektren behandelter und unbehandelter dA18 DNA Oligo-
mere. cNB: gekoppelte Nukleobasen, r-v(PO2): Referenzbande aus
dem Phosphat-Rückgrat, bb: Desoxyribose-Rückgrat, A1 - A4: Adenin
spezifisch. Jedes gezeigte Spektrum wurde durch 24 Wiederholungen
für je 2.5 s generiert (Gesamtintegrationszeit: 60 s, Anregungswel-
lenlänge λ = 785 nm). . . . . . . . . . . . . . . . . . . . . . . . . . . 27
67

3.3 Raman-Spektren behandelter und unbehandelter dT18 DNA Homo-
Oligomere. cNB: gekoppelte Nukleobasen, r-v(PO2): Referenzban-
de aus dem Phosphat-Rückgrat, bb: Desoxyribose-Rückgrat, T1 -
T4: Thymin spezifisch. Jedes gezeigte Spektrum wurde durch 24
Wiederholungen für je 2.5 s generiert (Gesamtintegrationszeit: 60 s,
Anregungswellenlänge λ = 785 nm). . . . . . . . . . . . . . . . . . . 27
3.4 Ramanspektren behandelter und unbehandelter dC18 DNA Homo-
Oligomere. cNB: gekoppelte Nukleobasen, r-v(PO2): Referenzban-
de aus dem Phosphat-Rückgrat, bb: Desoxyribose-Rückgrat, C1 -
C3: Cytosin spezifisch. Jedes gezeigte Spektrum wurde durch 24
Wiederholungen für je 2.5 s generiert (Gesamtintegrationszeit: 60 s,
Anregungswellenlänge λ = 785 nm). . . . . . . . . . . . . . . . . . . 29
3.5 Ramanspektren behandelter und unbehandelter dG18 DNA Homo-
Oligomere. cNB: gekoppelte Nukleobasen, r-v(PO2): Referenzban-
de aus dem Phosphat-Rückgrat, bb: Desoxyribose-Rückgrat, G1 -
G3: Guanin spezifisch. Jedes gezeigte Spektrum wurde durch 24
Wiederholungen für je 2.5 s generiert (Gesamtintegrationszeit: 60 s,
Anregungswellenlänge λ = 785 nm). . . . . . . . . . . . . . . . . . . 29
3.6 Ein Sterndiagramm bildet das Integral (algebraische Summe der Tra-
peze, Glg. 3.1) von 300 cm−1 bis 1800 cm−1 ab. Auf jeder Achse sind
die Werte der Integrale der Spektren nach Behandlung mit Helium-
Gas (Kontrolle) oder einem der drei Jets aufgetragen. Die Symbole
repräsentieren dabei die vier DNA Einzelstrang Homo-Oligomere
(dA18, dT18, dG18, dC18). Die Werte der Integrale der Kontrollspektren
wurden auf 1 normiert. . . . . . . . . . . . . . . . . . . . . . . . . . 30
3.7 Ein Sterndiagramm bildet I(cNB)/I(v(PO2)) ab. Auf jeder Achse ist
I(cNB)/I(v(PO2)) nach Behandlung entweder mit Helium-Gas (Kontrol-
le) oder einem der drei Jets aufgetragen. Die Symbole repräsentieren
dabei die vier DNA Einzelstrang Homo-Oligomere (dA18, dT18, dG18,
dC18). Um einen besseren Vergleich der Änderungen nach Behand-
lung mit einem der drei Jets zu gewährleisten wurde I(cNB)/I(v(PO2))
der Kontrollspektren ebenfalls auf 1 normiert. . . . . . . . . . . . . . 31
68

3.8 Normalisierte Raman-Spektren behandelter und unbehandelter dop-
pelsträngiger dA18:dT18 DNA. A1, A14, T1 bis T3: Nukleobasen spe-
zifisch (Annotation entnommen aus Abb.3.9), cNB: gekoppelte Nu-
kleobasen, r-v(PO2): Referenz-Bande aus dem Phosphat-Rückgrat,
bb: Desoxyribose-Rückgrat. Das Kontroll- und Komplett-Jet-Spektrum
wurde generiert durch 24 Wiederholungen bei je 2.5 s (Gesamtinte-
grationszeit: 60 s, Anregungswellenlänge λ = 785 nm). Das Differenz-
Spektrum (unten) der doppelsträngigen dA18:dT18 DNA behandelt
mit dem Komplett-Jet und der unbehandelten dA18:dT18 DNA zeigt
hauptsächlich Rauschbeiträge. . . . . . . . . . . . . . . . . . . . . . 32
3.9 Raman-Spektren von behandelten und unbehandelten Einzelstrang
DNA Homo-Oligomeren. cNB: gekoppelte Nukleobasen, r-v(PO2):
Referenz-Bande aus dem Phosphat-Rückgrat, bb: Desoxyribose-
Rückgrat, (A) dA18 1 - A4: Adenin (B) dT18: T1 - T4: Thymin (C) dC18:
Cytosin (D) dG18 G1 - G3: Guanin, Daten zum Vergleich aus [38].
Jedes gezeigte Spektrum wurde durch 24 Wiederholungen für je 2.5 s
generiert (Gesamtintegrationszeit: 60 s, Anregungswellenlänge λ =
785 nm). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34
4.1 Schematischer Aufbau eines humanen Spermatozoons. Der hoch-
kondensierte haploide Zellkern befindet sich im Kopf. An den Mittelteil,
der Mitochondrien beinhaltet, schließt sich die Geissel an, die der
Fortbewegung der Zelle dient. . . . . . . . . . . . . . . . . . . . . . 39
4.2 Prinzip der Summenfilteranalyse. (A) zeigt Links ein Einzelpunkt-
Spektrum aus einem Raman-Spektrendatensatz. In blau ist der Ar-
beitsbereich des Summenfilters hervorgehoben, also der Bereich in
dem in jedem Spektrum des Datensatzes die Intensitätswerte auf-
summiert wurden. Daneben ist das daraus entstandene farbkodierte
Raman-Bild eines kompletten Spermiums gezeigt mit der korrespon-
dierenden Farbskala, der die summierten Intensitäten in jedem Pi-
xel des Bildes zugewiesen worden sind. (B) zeigt das Ergebnis der
Summenfilteranalyse mit einem anderen Arbeitsbereich, der in der
Visualisierung einer Struktur im Kopf des Spermiums resultiert. (C)
zeigt ein weiteres Beispiel der Aufsummierung der Intensitäten einer
Bande mit der sich das Mittelstück der Zelle darstellen lässt. . . . . . 43
69

4.3 Optische Aufnahme und biochemische Komposition immobilisier-
ter, lebender individueller menschlicher Spermatozoen bei physio-
logieähnlichen Bedingungen. (A) Optische Aufnahme zeigt meh-
rere immobilisierte Spermien. Vierecke in Weiß geben die Mess-
bereiche an, in denen automatisierte CRM Messungen einzelner
Spermien durchgeführt worden sind. Messparameter sind für jede
Zelle: 10 µm x 10 µm bei 333 nm Auflösung pro Pixel, 15 mW La-
serleistung sowie 500 ms Integrationszeit pro Spektrum. (B) - (S)
Chemische Bilder rekonstruiert aus hyperspektralen Datensätzen
durch Summenfilteranalyse. (B) - (G) entspricht verschiedenen CH-
Schwingungen (2800 cm−1 bis 3000 cm−1), (H) - (M) der jeweiligen
Kopfregion 1090 cm−1 bis 1100 cm−1 und (N) - (S) dem Mittelstück
(745 cm−1 bis 755 cm−1). (T) - (Y) Überlagerung der Teilbilder (B) -
(S). blaue Farbskala entspricht der gesamten Zelle, grün dem Mittel-
stück und rot der Kopfregion. Maßstabsbalken entsprechen 1 µm in
(B) - (S) und 2 µm in (T) - (Y). . . . . . . . . . . . . . . . . . . . . . 44
4.4 Durchschnittsspektren der chemischen Bilder aus Abb. 4.3 (B) ver-
schiedene CH-Schwingungen (2800 cm−1 bis 3000 cm−1, Kopfregion
1090 cm−1 bis 1100 cm−1 und Mittelstück (745 cm−1 bis 755 cm−1)
sowie ein Spektrum des CaF2-Substrates (schwarz). Schattierungen
geben die Arbeitsbereiche der verwendeten Summenfilteranalyse an.
Die Spektren sind zum besseren Vergleich auf der Y-Achse gegen-
einander versetzt. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
4.5 Normalisierte Intensitäten der Banden bei 750 cm−1 und 1095 cm−1
aus den Kopfregionen (rot) in Abb. 4.3 (H) – (M) und aus den Mittel-
stückregionen (schwarz) in Abb. 4.3 (N) – (S). Jeweils drei normalisier-
te Mittelwerte mit der höchsten Intensität aus der Summenfilteranalyse
wurden in jedem Teilbild bestimmt, um die Daten als I(750 cm−1) als
Y- und I(1095 cm−1) als X-Werte nach Normalisierung darzustellen.
Fehlerbalken entsprechen dem Standardfehler des jeweiligen Mittel-
wertes. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46
70

4.6 Box Plot der absoluten Werte der Differenz I(750 cm-1) – I(1095 cm−1)
aus den Kopfregionen in Abb. 4.3 (B) – (G) (linke Box) und aus den
Mittelstückbereichen Abb. 4.3 (N) – (S) (rechte Box). Horizontale
Linien geben den Medianwert an. Fehlerbalken sind repräsentativ für
das 95 % Konfidenzintervall des jeweiligen Median. Grau hinterlegt
ist der jeweilige Interquartil-Bereich. Die Sternlinie *** steht für P <
0.0001 aus dem „Unabhängige Proben Mann-Whitney“ Test. . . . . . 47
4.7 Detektierte Fluoreszenzsignale der selben Spermien an der selben
Position nach Laserbestrahlung mit 30 mW. (A) Optisches Bild, das
immobilisierte Spermien zeigt (Sichtfeld zum Vergleich zu Abb. 5.5).
Vierecke in rot, grün und blau geben die Messbereiche an, in denen
automatisierte CRM von einzelnen Spermien mit einer Laserleistung
von 30 mW durchgeführt worden sind. (B) Spermien mit intakter Mem-
bran fluoreszieren hellgrün. (C) Zellen mit beschädigter Zellmembran
fluoreszieren rot. (D) - (F) Digitale Zooms der drei Spermatozoen
von Interesse aus (A). (G) - (I) Digitale Zooms der Spermatozoen
von Interesse aus (B). Das detektierte Fluoreszenzsignal der drei
Spermien, die Laserstrahlung aus CRM ausgesetzt worden sind, ist
deutlich geringer verglichen mit den Zellen in der Umgebung, die
keinem Laserlicht ausgesetzt wurden. (J) - (L) Digitale Zooms der
Spermatozoen von Interesse aus (C). Rote Fluoreszenz konnte für
die Spermien detektiert werden, die vorher mit CRM gemessen und
damit Laserstrahlung ausgesetzt worden sind. . . . . . . . . . . . . 48
71

4.8 Raman-chemische Bilder und detektierte Fluoreszenz aus SYBR
14 und PI des selben Spermiums an der selben Position. (A) Opti-
sches Bild, das immobilisierte Zellen darstellt. Viereck in Grün gibt
den Messbereich an, in dem eine automatisierte CRM Messung mit
einer Laserleistung von 15 mW durchgeführt worden ist. Teilbilder
sind die Raman chemischen Bilder rekonstruiert aus Summenfilter-
analyse der Zelle markiert in (A) mit (1). Von links nach Rechts: Ver-
schiedene CH-Schwingungen (2800 cm−1 bis 3000 cm−1), Kopfregion
(1090 cm−1 bis 1100 cm−1) und dem Mittelstück (745 cm−1 bis 755 cm−1).
Maßstabsbalken sind 2 µm. (C) Detektierte Fluoreszenz von SYBR-
14. Das Spermatozoon (1), welches vorher mit CRM gemessen wor-
den ist erscheint als heller Punkt neben anderen Zellen in der Um-
gebung (2),(3) die nicht vorher Laserstrahlung aus CRM Messungen
ausgesetzt worden sind. (D) Kein Fluoreszenzsignal der PI-Sonde
wurde detektiert. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
4.9 Farbkodierte Raman-Bilder von 35 individuellen, lebenden Sperma-
tozoen bei physiologieähnlichen Bedingungen, die vollständig au-
tomatisiert gemessen und per Summenfilteranalyse rekonstruiert
worden sind. Blaue Farbskala entspricht den verschiedenen Koh-
lenwasserstoffschwingungen (2800 cm−1 bis 3000 cm−1), grün den
Mittelstückbereichen (745 cm−1 bis 755 cm−1) sowie rot den Kopfre-
gionen (1090 cm−1 bis 1100 cm−1). Die gezeigten Maßstabsbalken
entsprechen 2 µm. Geometrie: 13 µm x 13 µm, 20 Punkte pro Linie,
20 Linien pro Bild, 0.15 s Messzeit pro Spektrum, 15 mW Laserleis-
tung. Gesamtmesszeit pro Zelle mit Hintergrund: 60 s. . . . . . . . . 50
4.10 S/N für jedes Teilbild in Abb. 4.9 aufgetragen gegen die laufende
Nummer der Zelle. . . . . . . . . . . . . . . . . . . . . . . . . . . . 51
4.11 Die letzten sechs farbkodierten Teilbilder einer automatisierten Mes-
sung immobilisierter Spermatozoen. Nach Zelle 17 wurde das Mikro-
skop manuell in z-Richtung in die Probe refokussiert. . . . . . . . . . 51
4.12 Detektierte SYBR-14 und PI Fluoreszenzsignale von Spermatozoen
nach Percoll-Dichtegradienten-Zentrifugation, Immobilisierung und
Lagerung für 4 h bei 35 ◦C. (A) Fluoreszenzsignal des SYBR-14/DNA
Komplexes zeigt mehrere helle Punkte. (B) Ein Fluoreszenz-Signal
des PI/SYYBR-14/DNA Komplexes wurde nicht detektiert. . . . . . . 52
72

4.13 Optische Aufnahmen eines einzelnen Spermatozoons direkt nach
Transfer auf das beschichtete CaF2 Substrat. Dargestellt ist die Motili-
tät der Geissel einer Zelle durch die verschiedene Position der Geissel
zu verschiedenen Zeitpunkten t nach Aufbringen auf das Substrat. . 53
5.1 Raman-Spektren von Silizium und TiO2-Dünnschichten auf Silizi-
um. Blaue Linie: Spektrum von Silizium, grüne Linie: Abgeschiedene
Dünnschicht auf Silizium, rote Linie: Wärmbehandelte Dünnschicht.
Gesamtintegrationszeit: 60 s (24 Wiederholungen je 2.5 s, 12 mW,
Anregungswellenlänge λ = 488 nm). . . . . . . . . . . . . . . . . . . 59
5.2 Raman-Spektren von Glas und TiO2-Dünnschichten auf Glas. Blaue
Linie: Spektrum von Glas, grüne Linie: Abgeschiedene Dünnschicht
auf Glas, rote Linie: Wärmbehandelte Dünnschicht. Gesamtintegra-
tionszeit: 60 s (24 Wiederholungen je 2.5 s, 12 mW, Anregungswel-
lenlänge λ = 488 nm). Spektren sind auf der Y-Achse gegeneinander
versetzt. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 60
5.3 Raman-Spektren von (C4C1py)[Cu(SCN)2]. Blaue Linie: In-situ-Messung
bei 10 ◦C, rote Linie: In-situ-Messung bei 100 ◦C. Gestrichelte ver-
tikale Linie: Raman-Bande bei 2097 cm−1. Gesamtintegrationszeit:
60 s (24 Wiederholungen je 2.5 s, 25 mW, Anregungswellenlänge λ
= 488 nm). Spektren sind auf der Y-Achse gegeneinander versetzt. . 61
5.4 Raman-Spektren von (C4C1py)[Cu(SCN)2]. Blaue Linie: In-situ-Messung
bei 20 ◦C, rote Linie: In-situ-Messung bei 100 ◦C. Gestrichelte ver-
tikale Linie markiert Banden, deren Lage in beiden Spektren über-
einstimmt. Gesamtintegrationszeit: 60 s (24 Wiederholungen je 2.5 s,
100 mW, Anregungswellenlänge λ = 785 nm). Spektren sind auf der
Y-Achse gegeneinander versetzt. . . . . . . . . . . . . . . . . . . . . 62
5.5 Optisches Bild aufgenommen mit dem konfokalen Raman-Mikroskop
zeigt einzelne, immobilisierte, lebende Spermatozoen (Vergleich mit
Abb. 4.7). Blaue, rote und grüne Vierecke geben Bereiche an, in
denen automatisierte CRM Messungen durchgeführt worden sind mit
einer Laserleistung von 30 mW. . . . . . . . . . . . . . . . . . . . . 64
73

Tabellenverzeichnis
1.1 Auswahl von mit Raman-Spektroskopie messbaren Eigenschaften. . 7
3.1 Ätzraten des X-Jets in (nmmin−1) von DNA Homo-Oligonukleotide. . 28
4.1 Vergleich und Zuordnung charakteristischer Banden im Raman-Spektrum
zellulärer Komponenten in bisherigen Studien (Ref.) und dieser Arbeit
[EE]. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
74


7Literaturverzeichnis
[1] H. Kuzmany. Solid-State Spectroscopy. Springer, Heidelberg, 2009.
[2] J.M. Hollas. Modern Spectroscopy. Wiley-VCH, Weinheim, 2004.
[3] R. Winter and F. Noll. Methoden der Biophysikalischen Chemie. Teubner
Studienbuecher, Wiesbaden, 1998.
[4] A.H. Compton. A quantum theory of the scattering of x-rays by light elements.
Physical Review, 21:483–502, May 1923.
[5] C.V. Raman and K.S. Krishnan. A new type of secondary radiation. Nature,
121:501–502, 1928.
[6] WITec. WITec Project. WITec, Ulm, 2007.
[7] K. Meister, J. Niesel, U. Schatzschneider, N. Metzler-Nolte, D.A. Schmidt, and
M. Havenith. Label-free imaging of metal-carbonyl complexes in live cells
by raman microspectroscopy. Angewandte Chemie. International Edition in
English, 49(19):3310–3312, Apr 2010.
[8] C. Matthaeus, T. Chernenko, L. Quintero, L. Milan, A. Kale, M. Amiji, V. Torchilin,
and M. Diem. Raman microscopic imaging of cells and applications monitoring
the uptake of drug delivery systems. Biophotonics: Photonic Solutions For
Better Health Care, 6991:99106–99106, 2008.
76

[9] C. Matthaeus, A. Kale, T. Chernenko, V. Torchilin, and M. Diem. New ways
of imaging uptake and intracellular fate of liposomal drug carrier systems
inside individual cells, based on raman microscopy. Molecular Pharmaceutics,
5(2):287–293, 2008.
[10] G. Di Florio, E. Bruendermann, N. S. Yadavalli, S. Santer, and M. Havenith.
Polarized 3d raman and nanoscale near-field optical microscopy of optically
inscribed surface relief gratings: chromophore orientation in azo-doped polymer
films. Soft Matter, 10(10):1544–1554, 2014.
[11] P. J. Pauzauskie, D. Talaga, K. Seo, P. D. Yang, and F. Lagugne-Labarthet. Po-
larized raman confocal microscopy of single gallium nitride nanowires. Journal
of the American Chemical Society, 127(49):17146–17147, December 2005.
[12] N. J. Everall. Measurement of orientation and crystallinity in uniaxially drawn po-
ly(ethylene terephthalate) using polarized confocal raman microscopy. Applied
Spectroscopy, 52(12):1498–1504, December 1998.
[13] V.-S. Dang, H. Parala, J.H. Kim, K. Xu, N.B. Srinivasan, E. Edengeiser, M. Ha-
venith, A.D. Wieck, T. de los Arcos, R.A. Fischer, and A. Devi. Electrical and
optical properties of tio2 thin films prepared by plasma-enhanced atomic layer
deposition. physica status solidi (a), 211:416–424, 2013.
[14] E.T. Spielberg, E. Edengeiser, B. Mallick, M. Havenith, and A.V. Mudring. (1-
butyl-4-methyl-pyridinium)[cu(scn)2 ]: A coordination polymer and ionic liquid.
Chemistry, 20(18):5338–5345, April 2014.
[15] F. Rull, A. C. Prieto, J. M. Casado, F. Sobron, and H. G. M. Edwards. Estimati-
on of crystallinity in polyethylene by raman-spectroscopy. Journal of Raman
Spectroscopy, 24(8):545–550, August 1993.
[16] B. P. Gaber and W. L. Peticolas. Quantitative interpretation of biomembrane
structure by raman-spectroscopy. Biochimica Et Biophysica Acta, 465(2):260–
274, 1977.
[17] M. J. Pelletier. Quantitative analysis using raman spectrometry. Applied Spec-
troscopy, 57(1):20A–42A, January 2003.
[18] N. C. Deno and M. J. Wisotsky. Quantitative raman spectroscopy for deter-
mination of base strengths of weak organic bases. Journal of the American
Chemical Society, 85(12):1735–&, 1963.
77

[19] R. Hooke. Micrographia. Royal Society, London, 1665.
[20] E. Robbins and N. K. Gonatas. Histochemical + ultrastructural studies on hela
cell cultures exposed to spindle inhibitors with special reference to interphase
cell. Journal of Histochemistry & Cytochemistry, 12(9):704–&, 1964.
[21] T. Dieing, O. Hollricher, and J. Toporski. Confocal Raman Microspectroscopy.
Springer, Heidelberg, 2010.
[22] T. H. Maiman. Stimulated optical radiation in ruby. Nature, 187(4736):493–494,
1960.
[23] W. S. Boyle and G. E. Smith. Charge coupled semiconductor devices. Bell
System Technical Journal, 49(4):587, 1970.
[24] J. Zhang, R. E. Campbell, A. Y. Ting, and R. Y. Tsien. Creating new fluorescent
probes for cell biology. Nature Reviews Molecular Cell Biology, 3(12):906–918,
December 2002.
[25] L. Brand and J. R. Gohlke. Fluorescence probes for structure. Annual Review
of Biochemistry, 41:843, 1972.
[26] Hazel H. Szeto, Peter W. Schiller, Kesheng Zhao, and Guoxiong Luo. Fluo-
rescent dyes alter intracellular targeting and function of cell-penetrating tetra-
peptides. FASEB journal : official publication of the Federation of American
Societies for Experimental Biology, 19(1):118–20, January 2005.
[27] J. W. Lichtman and J. A. Conchello. Fluorescence microscopy. Nature Methods,
2(12):910–919, December 2005.
[28] S. W. Hell and J. Wichman. Breaking the diffraction resolution limit by stimulated-
emission - stimulated-emission-depletion fluorescence microscopy. Optics
Letters, 19(11):780–782, June 1994.
[29] S. W. Hell. Toward fluorescence nanoscopy. Nature Biotechnology,
21(11):1347–1355, November 2003.
[30] S. W. Hell. Microscopy and its focal switch. Nature Methods, 6(1):24–32,
January 2009.
[31] J. D. Watson and F. H. C. Crick. Molecular structure of nucleic acids - a structure
for deoxyribose nucleic acid. Nature, 171(4356):737–738, 1953.
78

[32] H.R. Drew, R.M. Wing, T. Takano, C. Broka, S. Tanaka, K. Itakura, and R. E.
Dickerson. Structure of a b-dna dodecamer - conformation and dynamics
.1. Proceedings of the National Academy of Sciences of the United States of
America-biological Sciences, 78(4):2179–2183, 1981.
[33] D. Ellerweg, J. Benedikt, A. von Keudell, N. Knake, and V. Schulz-von der
Gathen. Characterization of the effluent of a he/o-2 microscale atmospheric
pressure plasma jet by quantitative molecular beam mass spectrometry. New
Journal of Physics, 12:013021, January 2010.
[34] M. Laroussi. Nonthermal decontamination of biological media by atmospheric-
pressure plasmas: Review, analysis, and prospects. IEEE Transactions on
Plasma Science, 30(4):1409–1415, August 2002.
[35] D. O’Connell, L. J. Cox, W. B. Hyland, S. J. McMahon, S. Reuter, W. G. Graham,
T. Gans, and F. J. Currell. Cold atmospheric pressure plasma jet interactions
with plasmid dna. Applied Physics Letters, 98(4):043701, January 2011.
[36] X. T. Deng, J. J. Shi, and M. G. Kong. Protein destruction by a helium atmos-
pheric pressure glow discharge: Capability and mechanisms. Journal of Applied
Physics, 101(7):074701, April 2007.
[37] S. Schneider, J. W. Lackmann, D. Ellerweg, B. Denis, F. Narberhaus, J. E.
Bandow, and J. Benedikt. The role of vuv radiation in the inactivation of bacteria
with an atmospheric pressure plasma jet. Plasma Processes and Polymers,
9(6):561–568, June 2012.
[38] J.W. Lackmann, S. Schneider, E. Edengeiser, F. Jarzina, S. Brinckmann,
E. Steinborn, M. Havenith, J. Benedikt, and J.E. Bandow. Photons and par-
ticles emitted from cold atmospheric-pressure plasma inactivate bacteria and
biomolecules independently and synergistically. Journal of the Royal Society,
Interface, 10(89):20130591, Dec 2013.
[39] J.W. Lackmann, S. Schneider, F. Narberhaus, J. Benedikt, and J. E. Ban-
dow. Plasma for Bio-Decontamination, Medicine and Food Security. Springer,
Heidelberg, 2012.
[40] S. Schneider, J. W. Lackmann, F. Narberhaus, J. E. Bandow, B. Denis, and
J. Benedikt. Separation of vuv/uv photons and reactive particles in the effluent
of a he/o-2 atmospheric pressure plasma jet. Journal of Physics D: Applied
Physics, 44(29):295201, July 2011.
79

[41] B. Prescott, W. Steinmetz, and G. J. Thomas. Raman spectral studies of nucleic-
acids 23. characterization of dna structures by laser raman-spectroscopy. Biop-
olymers, 23(2):235–256, 1984.
[42] E. W. Small and W.L. Peticolas. Conformational dependence of raman scat-
tering intensities from polynucleotides 3. order-disorder changes in helical
structures. Biopolymers, 10(8):1377–1416, 1971.
[43] G. J. Thomas, J. M. Benevides, S. A. Overman, T. Ueda, K. Ushizawa, M. Saitoh,
and M. Tsuboi. Polarized raman-spectra of oriented fibers of a-dna and b-dna -
anisotropic and isotropic local raman tensors of base and backbone vibrations.
Biophysical Journal, 68(3):1073–1088, March 1995.
[44] H. Deng, V. A. Bloomfield, J. M. Benevides, and GJ Thomas, Jr. Dependence
of the raman signature of genomic b-dna on nucleotide base sequence. Biopo-
lymers, 50(6):656–666, Nov 1999.
[45] G. J. Lee, Y. W. Kwon, Y. H. Kim, and E. H. Choi. Raman spectroscopic study of
plasma-treated salmon dna. Applied Physics Letters, 102(2):021911, January
2013.
[46] J. G. Duguid, V. A. Bloomfield, J. M. Benevides, and G. J. Thomas. Dna mel-
ting investigated by differential scanning calorimetry and raman spectroscopy.
Biophysical Journal, 71(6):3350–3360, December 1996.
[47] J. Mueller, W. Ibach, K. Weishaupt, and O. Hollricher. Confocal raman micros-
copy. Microscopy and Microanalysis, 9:1084–1085, 8 2003.
[48] K. Fricke, I. Koban, H. Tresp, L. Jablonowski, K. Schroder, A. Kramer, K. D. Welt-
mann, T. von Woedtke, and T. Kocher. Atmospheric pressure plasma: A high-
performance tool for the efficient removal of biofilms. Plos One, 7(8):e42539,
August 2012.
[49] J. Boivin, L. Bunting, J. A. Collins, and K. G. Nygren. International estimates
of infertility prevalence and treatment-seeking: potential need and demand for
infertility medical care. Human Reproduction, 22(6):1506–1512, June 2007.
[50] V. M. Brugh and L. I. Lipshultz. Male factor infertility evaluation and manage-
ment. Medical Clinics of North America, 88(2):367–385, March 2004.
80

[51] D. Mukhopadhyay, A. C. Varghese, M. Pal, S. K. Banerjee, A. K. Bhattacharyya,
R. K. Sharma, and A. Agarwal. Semen quality and age-specific changes: a study
between two decades on 3,729 male partners of couples with normal sperm
count and attending an andrology laboratory for infertility-related problems in
an indian city. Fertility and Sterility, 93(7):2247–2254, May 2010.
[52] M. Rolland, J. Le Moal, V. Wagner, D. Royere, and J. De Mouzon. Decline
in semen concentration and morphology in a sample of 26 609 men close to
general population between 1989 and 2005 in france. Human Reproduction,
28(2):462–470, February 2013.
[53] World Health Organization. WHO laboratory manual for the Examination and
processing of human semen. World Health Organization, Genf (Schweiz), 2010.
[54] W. Schoenfeld. Um die entdeckung der menschlichen samenfaeden. Archiv
fuer Dermatologie und Syphilis, 178(3):358–372, 1938.
[55] L. Stryer. Biochemistry. Freeman, New York, 1995.
[56] C. Avendano, A. Mata, A. M. J. Villanueva, N. Battello, C. A. S. Sarmiento, and
S. Oehninger. Dna fragmentation of sperm subpopulations with different head
morphology. implications for icsi treatment. Fertility and Sterility, 96(3):S160–
S160, September 2011.
[57] X. D. Zhang, R. Balhorn, J. Mazrimas, and J. Kirz. Measuring dna to protein
ratios in mammalian sperm head by xanes imaging. Journal of Structural
Biology, 116(3):335–344, May 1996.
[58] G. S. Bench, A. M. Friz, M. H. Corzett, D. H. Morse, and R. Balhorn. Dna and
total protamine masses in individual sperm from fertile mammalian subjects.
Cytometry, 23(4):263–271, April 1996.
[59] V. Sanchez, J. Wistuba, and C. Mallidis. Semen analysis: update on clinical
value, current needs and future perspectives. Reproduction, 146(6):R249–R258,
December 2013.
[60] J. Jasensky and J. E. Swain. Peering beneath the surface: Novel imaging
techniques to noninvasively select gametes and embryos for art. Biology of
Reproduction, 89(4):105, October 2013.
81

[61] C. Mallidis, V. Sanchez, and J. Wistuba. Raman microspectroscopy: a method
with great promise but which also needs circumspection. Human Reproduction,
28(9):2595–2596, September 2013.
[62] D.A. Schmidt, I. Kopf, and E. Bruendermann, E.ndermann. A matter of scale:
from far-field microscopy to near-field nanoscopy. Laser & Photonics Reviews,
6(3):296–332, 2012.
[63] J. Chan, S. Fore, S. Wachsman-Hogiu, and T. Huser. Raman spectroscopy and
microscopy of individual cells and cellular components. Laser Photonics Rev.,
2(5):325–349, October 2008.
[64] M. Diem, M. Romeo, S. Boydston-White, M. Miljkovic, and C. Matthaus. A
decade of vibrational micro-spectroscopy of human cells and tissue (1994-
2004). Analyst, 129(10):880–885, 2004.
[65] W. L. Kubasek, Y. Wang, G. A. Thomas, T. W. Patapoff, K. H. Schoenwaelder,
J. H. Vandersande, and W. L. Peticolas. Raman-spectra of the model b-dna
oligomer d(cgcgaattcgcg)2 and of the dna in living salmon sperm show that
both have very similar b-type conformations. Biochemistry, 25(23):7440–7445,
November 1986.
[66] T. Huser, C.A. Orme, C.W. Hollars, M.H. Corzett, and R. Balhorn. Raman
spectroscopy of dna packaging in individual human sperm cells distinguishes
normal from abnormal cells. Journal of Biophotonics, 2(5):322–332, May 2009.
[67] K. Meister, D.A. Schmidt, E. Bruendermann, and M. Havenith. Confocal raman
microspectroscopy as an analytical tool to assess the mitochondrial status in
human spermatozoa. Analyst, 135(6):1370–1374, Jun 2010.
[68] C. Mallidis, J. Wistuba, B. Bleisteiner, O. S. Damm, P. Gross, F. Wuebbeling,
C. Fallnich, M. Burger, and S. Schlatt. In situ visualization of damaged dna in
human sperm by raman microspectroscopy. Hum. Reprod., 26(7):1641–1649,
Jul 2011.
[69] Z. Huang, X. Chen, Y. Chen, J. Chen, M. Dou, S. Feng, H. Zeng, and R. Chen.
Raman spectroscopic characterization and differentiation of seminal plasma.
Journal of Biomedical Optics, 16(11):110501, Nov 2011.
[70] V. Sanchez, K. Redmann, J. Wistuba, F. Wubbeling, M. Burger, H. Oldenhof,
W. F. Wolkers, S. Kliesch, S. Schlatt, and C. Mallidis. Oxidative dna damage
82

in human sperm can be detected by raman microspectroscopy. Fertility and
Sterility, 98(5):1124, November 2012.
[71] Z. F. Huang, X. W. Chen, G. N. Chen, J. H. Chen, J. Wang, J. Q. Lin, H. S. Zeng,
and R. Chen. Characterization and differentiation of normal and abnormal sper-
matozoa via micro-raman spectroscopy. Laser Physics Letters, 10(3):035601,
March 2013.
[72] F. Liu, Y. Zhu, Y. F. Liu, X. B. Wang, P. Ping, X. Y. Zhu, H. L. Hu, Z. Li, and
L. He. Real-time raman microspectroscopy scanning of the single live sperm
bound to human zona pellucida. Fertility and Sterility, 99(3):684, March 2013.
[73] C. Krafft, T. Knetschke, R. H. W. Funk, and R. Salzer. Identification of organelles
and vesicles in single cells by raman microspectroscopic mapping. Vibrational
Spectroscopy, 38(1-2):85–93, July 2005.
[74] N. A. Brazhe, M. Treiman, A. R. Brazhe, N. L. Find, G. V. Maksimov, and
O. V. Sosnovtseva. Mapping of redox state of mitochondrial cytochromes in
live cardiomyocytes using raman microspectroscopy. Plos One, 7(9):e41990,
September 2012.
[75] M. Okada, N.I. Smith, A.F. Palonpon, H. Endo, S. Kawata, M. Sodeoka, and
K. Fujita. Label-free raman observation of cytochrome c dynamics during
apoptosis. Proceedings of the National Academy of Sciences of the United
States of America, 109(1):28–32, Jan 2012.
[76] W. Grundler, P. Dirscherl, W. Beisker, F. Weber, R. Stolla, and H. Bollwein.
Quantification of temporary and permanent subpopulations of bull sperm by an
optimized sybr-14/propidium iodide assay. Cytometry Part A, 60A(1):63–72,
July 2004.
[77] T. Ohsaka, F. Izumi, and Y. Fujiki. Raman-spectrum of anatase, tio2. Journal of
Raman Spectroscopy, 7(6):321–324, 1978.
[78] F. Dachille, P. Y. Simons, and R. Roy. Pressure-temperature studies of anatase
brookite rutile and tio2-2. American Mineralogist, 53(11-1):1929, 1968.
[79] G. A. Bowmaker and D. A. Rogers. Infrared and raman-spectra of some
bis(thiocyanato)cuprate(i) and bis(thiocyanato)aurate(i) complexes. Journal of
the Chemical Society - Dalton Transactions, (9):1873–1876, 1982.
83

[80] R.A. Bailey and T.W. Michelse. Infrared-spectra of mixed metal thiocyanate
complexes. Journal of Inorganic & Nuclear Chemistry, 34(8):2671–2673, 1972.
[81] J.J. Cael, J.L. Koenig, and J. Blackwel. Infrared and raman-spectroscopy of
carbohydrates .4. identification of configuration-sensitive and conformation-
sensitive modes for d-glucose by normal coordinate analysis. Carbohydrate
Research, 32(1):79–91, 1974.
[82] H. Krautscheid, N. Emig, N. Klaassen, and P. Seringer. Thiocyanato complexes
of the coinage metals: synthesis and crystal structures of the polymeric pyridine
complexes [agxcuy(scn)(x+y)(py)(z)]. Journal of the Chemical Society-dalton
Transactions, (18):3071–3077, September 1998.
84





























