Embed Size (px)
Citation preview

Ruprecht-Karls-Universität Heidelberg
Südasien-Institut
im Sommersemester 1999
Konsequenzen der Plattentektonik für die
Biogeographie
Hauptseminar
Biogeographie
Dozent: PD Dr. Wolfgang Werner
von
Sebastian Klüsener

1
Inhaltsverzeichnis
1 Einleitung-----------------------------------------------------------------------------------------------------3
2 Einführung in die wissenschaftliche Diskussion ------------------------------------------------------4
2.1 Die Theorie der Plattentektonik --------------------------------------------------------------------42.1.1 Entstehungsgeschichte der Theorie ----------------------------------------------------------------- 42.1.2 Die Theorie in ihrer heutigen Form ----------------------------------------------------------------- 9
2.1.2.1 Plattengrenzen ---------------------------------------------------------------------------------------102.1.2.1.1 Divergierende Plattengrenzen------------------------------------------------------------------------- 102.1.2.1.2 Konvergierende Plattengrenzen ---------------------------------------------------------------------- 112.1.2.1.3 Transformströmungen---------------------------------------------------------------------------------- 12
2.1.2.2 Hotspots-----------------------------------------------------------------------------------------------122.1.2.3 Mikroplatten -----------------------------------------------------------------------------------------13
2.2 Wirkung plattentektonischer Prozesse auf die Lithosphäre und das Ökosystem ------- 142.2.1 Tektonische Veränderungen -------------------------------------------------------------------------142.2.2 Klimatische und ozeanographische Veränderungen --------------------------------------------15
2.2.2.1 Global -------------------------------------------------------------------------------------------------152.2.2.1.1 Lage der Platten ----------------------------------------------------------------------------------------- 172.2.2.1.2 Orogenese ------------------------------------------------------------------------------------------------ 202.2.2.1.3 Vulkanismus --------------------------------------------------------------------------------------------- 212.2.2.1.4 Synthese anhand der zeitlichen Wirkung ----------------------------------------------------------- 21
2.2.2.2 Regional: Geographischer Formenwandel -----------------------------------------------------212.2.3 Wirkung auf eustatische Meeresspiegelschwankungen ----------------------------------------232.2.4 Synthese -------------------------------------------------------------------------------------------------24
2.3 Einführung in die Biogeographie mit besonderer Berücksichtigung der historischenBiogeographie ------------------------------------------------------------------------------------------------ 24
3 Konsequenzen der Plattentektonik für die Biogeographie---------------------------------------- 26
3.1 Konzepte der Aufrechterhaltung der Art ------------------------------------------------------- 283.1.1 Geoökologie---------------------------------------------------------------------------------------------283.1.2 Barrieren-------------------------------------------------------------------------------------------------303.1.3 Provinzen ------------------------------------------------------------------------------------------------32
3.2 Konzepte der Änderung des Verbreitungsmusters-------------------------------------------- 343.2.1 Sprungverbreitung über Barrieren hinweg--------------------------------------------------------353.2.2 Gleichgewichtstheorie und Inselbiogeographie--------------------------------------------------363.2.3 Erdgeschichte (TEKO-Ereignisse und Palaeogeographie) ------------------------------------363.2.4 Verbreitungsspuren als Resultat durch erdgeschichtliche Ereignisse bedingten
ökologischen Wandel ----------------------------------------------------------------------------------38
3.3 Konzepte der Entstehung neuer Arten ---------------------------------------------------------- 393.3.1 Empirisch phylogenetische (die Abstammung untersuchende) Biogeographie -----------413.3.2 Vikarianz als Resultat erdgeschichtlicher Veränderungen ------------------------------------413.3.3 Vikarianz als Resultat durch erdgeschichtliche Ereignisse bedingten ökologischen
Wandels oder ökologischen Wandel an sich ------------------------------------------------------423.3.4 Randisolierung als Resultat durch erdgeschichtliche Ereignisse bedingten ökologischen
Wandels--------------------------------------------------------------------------------------------------433.3.5 Sprungverbreitung über Barrieren hinweg--------------------------------------------------------433.3.6 Entstehungsgebiete neuer Arten (Centres of Origin) -------------------------------------------443.3.7 Geoökologie über Zeit --------------------------------------------------------------------------------44
4 Schlußbemerkung----------------------------------------------------------------------------------------- 46

2
5 Literaturverzeichnis -------------------------------------------------------------------------------------- 47
Appendix I: Die Geologische Zeittafel -----------------------------------------------------------------------------53
Appendix II: Die Biosphäre und darauf wirkende Prozesse ----------------------------------------------------54
Appendix III: Biogeographische Karten nach Ergebnissen des Paleomap Projects -----------------------55

3
1 Einleitung„But if rivers come into being and perish and if the same parts of the earth are not al-
ways moist the sea also must necessarily change correspondingly. And if in places the
sea recedes while in others it encroaches, then evidently the same parts of the earth as a
whole are not always sea, nor always mainland, but in process of time all change.“
Aristoteles, Meteorologica
Die Biogeographie beschäftigt sich mit der Ökologie und Entwicklung von lebender Materie
in Zeit und Raum. Man könnte dieses auf die scheinbar einfache Frage zurückführen: Warum
leben bestimmte Organismen zu einem bestimmten Zeitpunkt an einem oder mehreren be-
stimmten Orten? (vgl. Huggett, 1998).
Damit Materie entstehen, geformt und lebendig werden kann, benötigt es Energie: E =
m*c² (E = Energie, m = Materie, c = Lichtgeschwindigkeit) (Einstein, 1922). Für die Erdober-
fläche und die oberflächennahen Schichten, in denen hauptsächlich das terrestrische Leben
stattfindet (die Biosphäre), ist insbesondere die Sonne als exogene (von außen wirkende) E-
nergiequelle von Bedeutung. Daneben spielt aber auch die endogene (ohne äußeren Antrieb
von innen kommende) Energiezufuhr aus dem Planeten eine Rolle, deren Quellen der ca.
2.600 °C warme Erdkern und radioaktive Mineralien sind (Briggs, u.a. 1997) (siehe auch Ap-
pendix III für systematische Darstellung der Einflüsse auf die Biosphäre). Zwar macht die
endogene Energiezufuhr an der Erdoberfläche heute nur im Durchschnitt ca. 0,0001 Promille
der überwiegend solaren Gesamteinstrahlung aus (Briggs u.a., 1997), jedoch werden dadurch
die Prozesse der Plattentektonik bewirkt.
Theoretische Grundlage der Arbeit
Ursache und Wirkung zwischen Plattentektonik und dem Untersuchungsgebiet der Bioge-
ographie scheinen relativ einfach einordbar zu sein: Erst entstand das Sonnensystems aus dem
solaren Urnebel, es bildeten sich die Sonne und die Planeten. Die Erde kühlte ab, jedoch ist
ihr heißer Kern weiterhin die Energiequelle für plattentektonische Prozesse. Diese wirken in
vielfacher Art und Weise auf das komplexe ökologische Wirkungsgefüge, in dem das Leben
als Wasser-Kohlenstoff-Derivat aus geologischer Fraktionierung entstanden ist (Briggs,
1997), sich evolutionär angepaßt und in Raum und Zeit ausgebreitet hat.
Die Wissenschaft ist jedoch nicht in der Lage, diese Kausalkette von den Anfängen bis
heute aufzuarbeiten. Das liegt zum einen daran, daß die Erforschung von Ursache und Wir-

4
kung im Ökosystem mit positiven und negativen Rückkopplungen noch nicht weit genug fort-
geschritten bzw. das Ökosystem einfach zu komplex ist, als daß die Wissenschaft auf alle
Fragen Antworten geben könnte. Zum anderen wird das Datenmaterial auch umso rarer, je
weiter man sich in der Erdgeschichte zurückbegibt. Dieses liegt zum einen daran, daß eine
große Anzahl von Daten noch nicht erhoben sind, darüber hinaus sind aber auch viele Be-
weisstücke bereits durch erdgeschichtliche Prozesse wieder zerstört worden. So versucht die
Biogeographie, mit dem vorhandenen Daten und Erkenntnissen die Erdgeschichte hypothe-
tisch zu rekonstruieren.
Wenn also die vorliegende Arbeit die Konsequenzen der Plattentektonik für die Bioge-
ographie betrachtet, so zeigt sie auf, inwieweit Konzepte und Methoden der Biogeographie
diese berücksichtigen, da sie erdhistorische Komponenten beinhalten. Sie wird aber auch dar-
legen, daß im wissenschaftsevolutionären Sinne fast schon von „Konsequenzen der
Biogeographie für die Plattentektonik“ gesprochen werden kann. Brenchley und Harper
(1998) stellen etwa fest, das Theorien zur Plattentektonik und palaeogeographische Rekon-
struktionen auch heute zum größten Teil anhand von biologischen Daten vermutet und
getestet werden. Dieses birgt für die Biogeographie Fehlerquellen, wie unter 3.1.2 aufgezeigt
wird.
2 Einführung in die wissenschaftliche Diskussion
2.1 Die Theorie der Plattentektonik
2.1.1 Entstehungsgeschichte der Theorie
Francis Bacon wies schon 1620 auf den paßgenauen Verlauf der Westküste Afrikas und der
Ostküste Südamerikas hin (Bacon, 1620), ebenso machte der Botaniker und Geograph Ale-
xander von Humboldt auf auffällige Parallelitäten aufmerksam:
„Man kann diese (amerikanischen) Gebirgsketten jenseits des Ozeans im alten Konti-
nent nach Osten hin verfolgen, und man erkennt, daß unter der gleichen Breite die alten
Gebirge in den Gebieten von Pernambuco, von Minas, Bahia und Rio de Janeiro denen
am Kongo entsprechen; (...) Dieser Gedanke scheint weniger gewagt, wenn man die alte
und die neue Welt als gewaltsam durch das Wasser getrennt ansieht. (...) Der so ge-
schaffene Kanal hat bei Brasilien und bei Grönland die geringste Breite, aber die Tier-
und Pflanzengeographie deuten darauf hin, daß er sich schon zu einer Zeit bildete, wo
es auf der Erde keine oder nur wenige Keime organischen Lebens gab.“ (zitiert von
Reinke-Kunze, 1994)

5
Jedoch ging die geologische Lehrmeinung bis in die Mitte dieses Jahrhunderts von einem fi-
xistischen Weltbild aus, in dem die Kontinente sich nicht bewegten. Gegen dieses Weltbild
sprachen aber u.a. Befunde aus der Biologie und der Paläontologie. So hatte man etwa Fossi-
lien identischer Arten von Reptilien sowohl in Afrika als auch in Südamerika gefunden,
obwohl diese nicht in der Lage gewesen waren, den Ozean zu überqueren.
Fossilien des Reptils Mesosaurus
Quelle: Press und Siever (1995)
Die Fixisten erklärten diese Befunde mit der Kontraktionshypothese, die davon ausging,
daß der Erdkörper in seiner Geschichte abgekühlt sei, was einen vertikalen Schrumpfungspro-
zeß zur Folge gehabt hatte. Dadurch wurden nicht nur Kontinente auseinandergerissen,
sondern auch die Gebirge der Erde geformt, gleich den Runzeln in der Schale eines austrock-
nenden Apfels. Ein anderer Erklärungsansatz nahm an, daß ehemals Landbrücken zwischen
den Kontinenten bestanden hätten, die jedoch später im Meer versunken sein sollten. Einige
vermuteten, daß die Sintflut die Ursache für jene „Katastrophe“ war (vgl. Reinke-Kunze,
1994).
Diesem Weltbild stellte sich der Geophysiker Alfred Wegener entgegen, der als Wegbe-
reiter der modernen Theorie der Plattentektonik anzusehen ist. Zwar hatten schon vor und
unabhängig von ihm Wissenschaftler ähnliche Hypothesen vorgelegt (vgl. u.a. Snider Pel-

6
legrini (1858)), doch hatte keiner vor ihm die wissenschaftliche Diskussion nachhaltig in
Richtung eines Paradigmenwechsels lenken können (zu Paradigmenwechsel vgl. Kuhn, 1967).
Grundlage seiner Hypothese war zum einen die Entdeckung der natürlichen Reaktivität
durch Becquerel (1896), die Wegener zu der Vermutung geleitet hat, „daß schon mäßige Vor-
räte an solchen radioaktiven Stoffen im Erdinnern genügen, um den Wärmehaushalt zu
balancieren“ (zitiert von Closs u.a., 1985). Das widersprach der Grundannahme der Kontrak-
tionstheorie, dem Abkühlen des Erdkörpers. Darüber hinaus war ein Absinken der
Landbrücken zwischen den Kontinenten nicht mit dem Anfang des 20. Jahrhunderts entwi-
ckelten und heute allgemein akzeptierten Konzept der Isostasie vereinbar, das davon ausging,
daß die Kontinente aus leichterem Material (Sial) bestehen als die Ozeanböden und der unter
dem Kontinenten liegende Bereich (Sima). Da die Schwerkraft überall auf der Erdoberfläche
annähernd gleich war, kam man zu der Annahme, daß die Kontinente gleich Schwimmkörpern
im Wasser im isostatischen Gleichgewicht auf dem flüssigen Erdmantel schwimmen. Nach
dieser Theorie wäre ein Absinken der leichten Landbrücken in den dichteren Meeresboden
nicht möglich.
Isostasie
Quelle: Press und Siever (1995)
Es mußte also eine andere Erklärung für die bemerkenswerte Gleichartigkeit der Gesteine,
der geologischen Strukturen und der Fossilien auf den gegenüberliegenden Seiten des Atlan-
tiks in Südamerika und Afrika geben. So kam Wegener zur Hypothese der
Kontinentalverschiebung, die er zum ersten Mal 1912 auf einem Geologentag in Frankfurt
vorstellte. In der Folge lieferte er weitere Argumente für die Kontinentaldrift, wobei seine

7
Theorie davon ausging, daß alle heutigen Kontinente einmal in einem Kontinent, Pangaea
(„alles Land“), vereint waren.
Besonders in den zwanziger Jahren wurde seine Theorie heftig diskutiert und anschließend
von der Mehrheit der Wissenschaftler verworfen. Dieses lag zum einen daran, daß das ganze
geologische Weltbild aufgrund der Hypothese eines „Fachfremden“ hätte revidiert werden
müssen. So sagte Schuchart zu der Theorie:
„Die Schlacht um die Theorie von der Beständigkeit aller Großformen der Erde ist von
den Amerikanern längst ausgefochten und gewonnen. In Europa freilich geht der Kampf
noch weiter, weil dort führende Geologen immer noch Lyell folgen und an die Unbe-
ständigkeit der Kontinente und Ozeane glauben, während andere sich nicht entblöden,
die Pole hierhin und dorthin zu verlegen, nur um ein paar Sonderbarkeiten der Flora
und Fauna zu erklären.“(zitiert von Reinke-Kunze, 1994)
Weitere zitierte Reaktionen, die offenbaren, wie sehr sich das Wissenschaftsverständnis am
Anfangs des 20. Jahrhunderts von dem heutigen unterscheidet, finden sich in Reinke-Kunzes
Wegener-Biographie (Reinke-Kunze, 1994).
Neben diesen eher prinzipiellen als wissenschaftlichen Einwänden konnte sich die Theorie
auch deshalb nicht durchsetzen, da es den Anhängern der Theorie nicht gelang, eine schlüssi-
ge Erklärung für die Antriebsenergie zu finden, die Pangaea auseinanderbrechen und die
Kontinente voneinander fortbewegen ließ. Der Astronom und Geophysiker Jeffreys griff die
Theorie etwa an, indem er aufzeigte, daß die Erdkruste viel zu massiv und stark sei, um von
Kräften innerhalb der Erde angetrieben zu werden. So berechnete er etwa, daß eine Anzie-
hungskraft, die ausreichend wäre, um Kontinente zu bewegen, die Rotation der Erde in
weniger als einem Jahr zum stehen bringen würde (Jeffreys, 1924).
Die Befürworter der Theorie konnten sich dadurch in ihrer Beweisführung anfangs ledig-
lich auf die Überlieferung von Fossilien, Klimadaten und die Hypothese der Gebirgsbildung
an kollidierenden „Schollen“ stützen. Hinsichtlich der Fossilien zeigten die Vertebraten (Wir-
beltiere) und Landpflanzen bis zum vermuteten Auseinanderbrechen in ihren
Entwicklungslinien auffallende Ähnlichkeiten, während sie in der Folge wohl aufgrund der
Isolation und sich verändernder Klimabedingungen auseinanderliefen. Auch die Verteilung
der permokarbonischen Gletscherablagerungen in Südamerika, Afrika, Indien und Australien
war in Form getrennter Gletscher, von denen einige in Äquatornähe lagen, schwerlich zu er-
klären. Ging man jedoch davon aus, daß sich in der Erdgeschichte diese

8
Gletscherablagerungen im Südpolargebiet über einen damals zusammenhängenden Kontinent
(Gondwana) erstreckten, machten die Funde Sinn (Du Toit, 1937).
Hinsichtlich der Antriebsenergie veröffentlichte Holmes 1928 eine erste schlüssige Hypothe-
se. Er nahm an, daß der Mechanismus der thermischen Konvektion, wie er etwa in einem von
unten erhitzen Wassertopf zu finden ist, im Erdmantel die Kontinentaldrift antreiben würde.
Konvektionsbewegungen
Quelle: Press und Siever (1995)
Holmes vermutete, daß subkrustale Strömungen „die beiden Hälften des ursprünglichen
Kontinents auseinanderzogen, mit anschließender Gebirgsbildung an den Rändern der Konti-
nente, dort, wo die Strömungen nach unten abtauchen, und der Bildung von Ozeanboden in
der Lücke, wo die Strömungen aufsteigen“ (Holmes, 1928). Er konnte aber seine schon sehr
nahe an der heutigen Lehrmeinung befindliche Hypothese nicht beweisen, da es ihm an empi-
rischen Befunden fehlte. So räumte er selber ein, daß rein spekultativen Vorstellungen dieser
Art, die ausschließlich entwickelt werden, um den Erfordernissen zu genügen, kein wissen-
schaftlicher Wert zukommt, bis sie durch unabhängige Beweise gestützt werden.
Diese lieferte schließlich eine systematische Erforschung des Meeresbodens. Die Entde-
ckung des spaltenartigen Rifts auf dem mittelatlantischen Rücken löste zahlreiche
Spekulationen aus, die Hess und Dietz (Hess, 1962) zur Theorie des Sea-Floor-Spreadings
(siehe 2.1.2.1.1) ausbauten. Unterstützt wurde ihre These durch Probeentnahmen, die anhand
von paläomagnetischen Untersuchungen bewiesen, daß das Alter der Gesteine im Atlantik
zum mittelatlantischen Rücken hin abnimmt. Ende der 60er Jahre waren die Beweise so über-
zeugend, daß sich die meisten Geowissenschaftler der Theorie anschlossen.

9
Für Press und Siever (1995) ist die Entdeckung der Plattentektonik für die Geowissen-
schaften von ähnlich fundamentaler Bedeutung, wie etwa die Entdeckung der
Relativitätstheorie für die Physik, die der DNA für die Biologie oder der Urknalltheorie für
die Astronomie. Die Stärke der Theorie liegt ihrer Meinung nach in der Einfachheit und All-
gemeingültigkeit. Sie liefert für viele endogene Vorgänge Erklärungen und hat zahlreiche
Falsifizierungsversuche überstanden, so daß viele Wissenschaftler sie heute als Tatsache an-
sehen. Sie sollte jedoch ihrer Ansicht nach keinesfalls als Dogma verstanden werden (Press
und Siever, 1995).
2.1.2 Die Theorie in ihrer heutigen Form
Nach der heutigen Lehrmeinung finden die Vorgänge der Plattentektonik auf der äußersten
Schale unseres Planeten statt, der Lithosphäre. Diese ist eine starre und dadurch brechbare 50
-100 km dicke Schicht, welche die Kruste und die äußeren Schichten des Erdmantels umfaßt.
Die Lithosphäre ist in etwa ein Dutzend starre und passiv driftende Platten gebrochen, die im
isostatischen Gleichgewicht auf der sogenannten Astehnosphäre schwimmen, einer unter ho-
hem Druck stehenden teilweise flüssigen und verformbaren Schicht. Man geht davon aus, daß
in ihr die von Holmes angenommenen Konvektionsbewegungen auftreten (Press und Siever,
1995). Diese sollen die Antriebskraft für die meßbaren Lithosphärenplattenbewegungen sein,
die wenige Zentimeter pro Jahr betragen (entspricht etwa der Wachstumsgeschwindigkeit
menschlicher Fingernägel).
Die relative Geschwindigkeit (cm/ Jahr) und Richtung der Plattenbewegungen

10
Press und Siever (1995)
Neben horizontalen Plattenbewegungen sind aber auch vertikale auszumachen, die sich durch
das Prinzip der Isostasie erklären. So sinken Platten tiefer in den Mantel ab, wenn sie schwe-
rer werden (Subsidenz). Dieses kann etwa mechanisch durch die Bildung von Eisschilden, die
Ablagerung von Sedimenten oder einen höheren Meeresspiegel bewirkt werden. Der umge-
kehrte Prozeß wird als Elevation bezeichnet. Vertikale Plattenbewegungen können daneben
nach Briggs u.a. (1997) auch thermisch durch isolierte Hotspots (siehe 2.1.2.3) oder Magmen-
kammern bedingt sein.
2.1.2.1 Plattengrenzen
Hinsichtlich der Plattengrenzen, an denen großräumige geologische und geophysikalische
Vorgänge auftreten, werden drei Typen unterschieden. Divergierende Plattengrenzen, an de-
nen sich die Platten voneinander fortbewegen, konvergierende, an denen Platten kollidieren,
und Transformströmungen, an denen sie aneinander vorbeigleiten.
2.1.2.1.1 Divergierende Plattengrenzen
An divergierenden Grenzen wird der entstehende Freiraum zwischen den auseinanderdriften-
den Platten mit Magma gefüllt, welches aus dem oberen Mantel aufsteigt und in den Spalten
zu Gestein erstarrt, wobei es zu Erdbebentätigkeit und Vulkanismus kommt. Durch die Ak-
kumulation frischen basaltreichen Gesteins mit einem Gewicht von 2.8 - 3,4 g/ cm³ in 7 -10
km dichten Schichten (Briggs u.a., 1997) werden die sich trennenden Lithosphärenplatten
ständig vergrößert. Dieses tritt in der Regel auf dem Ozeangrund (Seafloor-Spreading) auf, da
die neugebildete „ozeanische“ Kruste dünner und spezifisch schwerer ist als ältere leichtere
und dickere „kontinentale“ Krusten, wodurch sie im isostatischen Gleichgewicht tiefer liegt.
Divergierende Grenzen zeichnen sich durch eine zentrale Grabenstruktur aus, ein soge-
nanntes Rift, das typischerweise auf dem Kamm von Gebirgsketten im Meer verläuft, den
sogenannten mittelozeanischen Rücken. Es gibt aber auch Stellen, an denen das Rift über dem
Meeresspiegel liegt (z.B. auf Island).
Frühstadien der Plattentrennung sind auch auf den Kontinenten zu finden. Diese Gebiete
zeichnen sich durch langgestreckte Grabensenken, sogenannte Rift-Valleys, aus. Beispiele
hierfür sind die Ostafrikanischen Gräben oder der Oberrheingraben, deren schwache tektoni-
sche Aktivität jedoch zur Vermutung veranlaßt, daß bei diesen Grabensystemen der Prozeß

11
zum Stillstand gekommen ist. Eine sehr aktive junge Riftzone stellt dagegen das Becken des
Roten Meeres dar, welches die Arabischen Halbinsel vom Afrikanischen Kontinent trennt.
2.1.2.1.2 Konvergierende Plattengrenzen
Wenn Platten sich vergrößern und auseinanderdriften, müssen an anderer Stelle auch Platten
kollidieren. Hierbei sind aufgrund der Tatsache, daß kontinentale Platten leichter als ozeani-
sche sind, drei Formen zu unterscheiden: die Kollision zwischen zwei kontinentalen Platten
und die sehr ähnlichen Kollision einer ozeanischen mit einer kontinentalen Platte bzw. Kolli-
sion zweier ozeanischer Platten.
Driften zwei kontinentale Platten aufeinander zu, wie es etwa momentan an der Grenze der
Indischen zur Eurasischen Platte der Fall ist, können die Platten wegen ihrer geringen Dichte
nicht nach unten abtauchen. Im konkreten Fall führt das dazu, daß die Eurasische die Indische
Platte überfährt, wodurch die Krustenmächtigkeit in diesem Raim verdoppelt wird, so daß das
momentan höchste Gebirge der Welt, der Himalaya, entstehen konnte.
Driftet eine ozeanische auf eine kontinentale Platte, wird erstgenannte von der leichteren
kontinentalen Platte überfahren und nach unten in den darunterliegenden Erdmantel gezogen
(Subduktion), in dem sie schließlich aufgeschmolzen wird. Bei der Kollision zweier ozeani-
scher Platten wird ebenfalls die schwerere Platte subduziert. Es kommt zur Bildung von
Tiefseerinnen, von Gebirgen mit Falten und Bruchtektonik und von magmatischen Gürteln.
Bei diesen kann es sich um einen Gebirgszug auf dem Festland oder um eine Kette von Vul-
kaninseln auf dem Meeresboden, einen sogenannten Inselboden handeln. Die in den
Erdmantel subduzierte Lithosphäre wird erwärmt und teilweise aufgeschmolzen, wobei sie
sich mit heißen Mantelmaterial vermischt. Das dadurch entstehende Magma steigt teilweise
auf, speist und bildet Vulkane und vulkanische Plateaus. Die durch die Kollision entstehenden
gewaltigen Kräfte äußern sich in Bruchtektonik, die Flach- und Tiefherdbeben auslöst. Ein
gutes Beispiel für die beschriebene Kollision ist die der Nazca-Platte mit der Südamerikani-
schen Platte.
Nach Burchfield (1983) sind die beiden zuletzt beschriebenen konvergierende Plattengren-
zen die Entstehungsräume neuer kontinentaler Kruste. Der Anteil der kontinentalen Platten
scheint dabei gegenüber der ozeanischen in der letzten Milliarde Jahre jährlich um etwa 1.3
km³ gewachsen zu sein (Briggs u.a. 1997). Kontinentale Kruste ist dicker (25 - 75 km), leich-
ter (2,7 g cm-3) und dadurch beständiger als die schwere eher subduzierte ozeanische Kruste.
So kann kontinentale Kruste bis zu 4 Milliarden Jahre alt sein, während die älteste datierte
ozeanische etwa 160 - 190 Millionen Jahre alt ist. Die kontinentale Kruste umfaßt dabei nicht

12
nur die ca. 29% Oberfläche, die momentan nicht von Ozeanen und Meeren bedeckt sind, son-
dern 39%, da sie teilweise von epikontinentalen Meeren wie etwa der Nordsee bedeckt
werden. Diese Kontinentalschelfe sind ein Zentrum der marinen Biodiversität (Briggs u.a.,
1997).
2.1.2.1.3 Transformströmungen
An Transformströmungen gleiten Platten aneinander vorbei, wobei praktisch weder neue Li-
thosphäre gebildet noch bestehende zerstört wird. Sie treten dort auf, wo eine divergierende
Plattengrenze in ihrem Verlauf unterbrochen und seitlich versetzt wird. Ein klassisches Bei-
spiel für eine Transformstörung ist die San-Andreas-Störung in Kalifornien, bei der die
Pazifische an der Nordamerikanischen Platte entlanggleitet. Transformströmungen sind von
Flachbeben mit horinzontalem Versatz geprägt.
Vereinfachte Darstellung der drei Typen von Plattengrenzen
Quelle: Press und Siever (1995)
2.1.2.2 Hotspots
Vulkanismus tritt nicht nur an Plattenrändern sondern auch inmitten von Platten auf. Dieser
Intraplattenvulkanismus war lange schwer in die Theorie der Plattentektonik einzuordnen, bis
die Hypothese der Hot Spots aufkam. Nach ihr sind Hot Spots vulkanische Erscheinungsfor-
men von sogenannten Manteldiapiren. Ein Manteldiapir ist ein begrenzter Strahl von heißem
Material, der aus dem Inneren des Mantels aufsteigt, die Lithosphäre durchdringt und an der
Oberfläche ausfließt. Diese Strömungen sind vermutlich relativ ortsfest und bewegen sich
nicht mit den Lithosphärenplatten. Wenn sich eine Platte über einen Hot-Spot hinwegbewegt,
entsteht so eine Reihe erloschener Vulkane, die zunehmend älter werden, wie etwa die Ha-

13
waii- und Emperor-Seamount-Kette. Deren jüngste Insel ist Hawaii, der immer ältere nach
Nordwesten folgen, die überwiegend bereits wieder unter dem Meeresspiegel liegen (Press
und Siever, 1995).
2.1.2.3 Mikroplatten
Die kontinentalen Lithosphärenplatten stellen im Gegensatz zu den neugebildeten Ozeanplat-
ten keine homogenen Körper dar. So finden sich etwa in Krustenblöcke mit Ausdehnungen
bis zu 100 Kilometern, die sich in der Paragenese (Vorkommen von Mineralien in Gesteinen),
der Art der Faltung und Bruchtektonik, dem Paläomagnetismus und der Metamorphosege-
schichte sehr von angrenzenden Bereichen unterscheiden. Auch Fossilfunde, falls vorhanden,
sprechen dafür, daß diese Blöcke aus anderen Bildungsräumen und geologischen Zeitaltern
stammen.
Solche Mikroplatten können Bruchstücke anderer Kontinente, unterseeischer Tafelberge,
vulkanischer Inselbögen oder Späne ozeanischer Kruste sein, die bei Kollisionsvorgängen von
ihrem Unterlager abgeschürft und an andere Kontinente „angeschweißt“ werden. Mikroplatten
finden sich etwa in den Appalachen und in den Kodilleren Nordamerikas, in denen etwa 100
nachgewiesen wurden. Aber auch in Japan, Südostasien, China und Sibirien sind sie belegt.
Ein interessanter Fall ist Florida, das wohl ehemals zum afrikanischen Kontinent gehörte. Es
verblieb jedoch bei Nordamerika, als sich beide Kontinente vor 200 Millionen Jahren nach
einem vorübergehenden Zusammenschluß wieder trennten (Press und Siever, 1995).
Die Entstehung von Mikroplatten

14
Quelle: Press und Siever (1995)
2.2 Wirkung plattentektonischer Prozesse auf die Lithosphäre und das
Ökosystem
Die Prozesse der Plattentektonik sind als endogene (unbeeinflußte) Prozeßresponssysteme von
großer Bedeutung für die Entwicklung der Lithosphäre und des Reliefs der Erde. Sie beeinflu-
ßen darüber hinaus auch auf vielfältige Weise das komplexe globale Ökosystem. Dieses soll
anhand von Hypothesen bezüglich der räumlichen und zeitlichen Wirkung der Prozesse auf
tektonische, eustatische (hier nur auf Meeresspiegelschwankungen bezogen), klimatische und
ozeanographische Prozesse (TEKO-Veränderungen) aufgezeigt werden, die für die Bioge-
ographie von großer Bedeutung sind.
2.2.1 Tektonische Veränderungen
Plattentektonische Prozesse bewirken global über Zeiträume von 106-8 Jahren (Briggs u.a.,
1997) den immerwährenden Wechsel der Konfiguration von Ozeanbecken, Kontinenten, Ge-
birgen und Gräben. Einige Wissenschaftler vermuten, daß dieses zyklischen Charakter hat,
was etwa alle 500 Millionen Jahre zur Bildung von Superkontinenten wie Pangaea vor 200
Millionen Jahren oder Rhondinia vor 750 Millionen Jahren führt (vgl. Wilson, 1963, Kearey

15
und Vine, 1996). Eine hypothetische Rekonstruktion der Kontinentbewegungen der letzten
200 Millionen Jahre durch das Palaeokartenprojekt des Internationalen Lithosphä-
renprogramms (Scotese, 1992) Jahre findet sich in Appendix IV.
Durch Plattenbewegungen entstehen im Zeitraum von 106-7 Jahren (Briggs u.a., 1997) glo-
bale seimische Gürtel, in welchen der größte Teil der Erdbeben- und Vulkanaktivität zu
finden ist, die über die Erdgeschichte starken Schwankungen unterworfen war. Innerhalb von
101-9 Jahren (Briggs u.a., 1997) werden Gesteine gebildet, zerstört und neuformiert. Darüber
hinaus werden durch die horizontale und vertikale Verschiebung der kontinentalen Kruste
bzw. durch die eustatische (2.2.3) Veränderung des Meeresspiegels geomorphologische Pro-
zesse angetrieben (Briggs u.a., 1997).
Die Wirkung der Plattentektonik auf regionale bis kleinräumige geomorphologische, pedolo-
gische und hydrologische Prozesse wird hier vernachlässigt, da diese überwiegend durch
solare Energie bewirkt werden und nur selten explizit auf Erdenergie zurückzuführen sind.
Auch Schwankungen des Meeresspiegels durch Änderung der Ozeangeometrie oder durch
vertikale Plattenbewegungen bedingte isostatische Meeresspiegelschwankungen werden nicht
weiter berücksichtigt. Diese treten zwar auf, jedoch wird ihre Wirkung etwa auf die Verände-
rung von Meeresströmungen durch diemit einer höheren Amplitide auftretenden eustatischen
Meeresspiegeländerungen überdeckt.
2.2.2 Klimatische und ozeanographische Veränderungen
2.2.2.1 Global
Schönwiese (1994) unterscheidet bei potentiellen Ursachen für Klimaänderungen extraterrest-
rische von terrestrischen. Daneben unterteilt Schönwiese (1994) die Einflüsse in externe,
unbeeinflußte und in interne, die durch andere bewirkt werden und im allgemeinen vernetzt
sind. Von den terrestrischen Ursachen für Klimaänderungen sieht Schönwiese (1994) mit
Vulkanismus, Kontinentaldrift und Orogenese alleine Prozesse der Plattentektonik als extern
an., die er als eine primäre Ursache für den festzustellenden erdgeschichtlichen Wechsel zwi-
schen Eis- und Warmzeiten einordnet, wobei dafür aber auch extraterrestrische Ursachen als
Faktoren diskutiert werden (vgl. Fischer, 1984; Williams, 1981).
Übersicht der wichtigsten natürlichen Ursachen von Klimaänderungen
Extraterrestrisch Terrestrisch

16
Solarkonstante, langfristiger (geolo-
gischer Trend)
Solarkonstante, Variation (durch Son-
nenaktivität bzw. Pulsationen)
Rotation der Milchstraße und kosmi-
sche Materie
Meteore und Meteoriten
Mond
Gezeitenkräfte allgemein (Wirkung auf
Sonne und Erde
Kontinentaldrift
Orogenese
Vulkanismus
Waldbrände
Zusammensetzung der Atmosphäre allge-
mein²
Zirkulation der Atmosphäre
Zirkulation und Salzgehalt des Oze-
ans1
Eis- und Schneebedeckung1
Bewölkung1
Vegetation1
Autovariationen, Chaos
Orbitalparameter
Rückkopplungen
1 Interne Mechanismen des Klimasystems (in Wechselwirkungen eingebunden)
² Teils intern, teils extern beeinflußt
Quelle: Schönwiese (1994)

17
2.2.2.1.1 Lage der Platten
“If no great continents existed, but a great number of islands without any remarkable
preponderance of land towards the tropical or the polar regions, the mean temperature
of the earth would be increased, and the distribution of heat over its surface rendered
far more uniform.” Hennessey (1860)
Schnee kann auf Land liegenbleiben, auf Wasser ist dieses nicht möglich. Sobald dieses ge-
schieht, kommt es zu einer positiven Eis-Albedo-Rückkopplung (Crowley und North, 1991).
Da nämlich Schnee eine sehr hohe Albedo hat, verringert sich die Strahlungsbilanz der Ober-
fläche, was eine weitere Abkühlung zur Folge hat. So geht die von Fairbridge (1972)
aufgestellte Koinzidenzhypothese davon aus, daß ein Übergang von einem warmzeitlichen
Gleichgewicht in ein eiszeitliches umso wahrscheinlicher ist, je mehr Landoberfläche sich um
den Pol befindet bzw. der Übergang zu einer Warmzeit wahrscheinlicher wird, je weniger
Land am Pol liegt (vgl. Beaty, 1978; Schwarzbach, 1993). Für die Entstehung ausgeprägter
Wärmeperioden wie zur Zeit der Kreide reichen diese Prozesse nicht aus, sondern wohl ihr
gleichzeitiges Eintreten mit extraterrestrischen Veränderungen und/oder einem Ansteigen des
CO2-Gehalts durch erhöhte vulkanische Aktivität (2.2.2.3.1) und dem Ansteigen des Meeres-
spiegels (2.2.3), der die Verwitterung von Silikaten auf den Kontinenten einschränkt (Barron
und Washington, 1985).
Hinsichtlich der Eiszeiten ist belegt, daß die Eisbildungen des Silur-Ordovizischen Zeital-
ters auftraten, als der sich durch plattentektonische Prozesse relativ bewegende Südpol vom
Meer kommend Gondwana „erreichte“ (Schönwiese, 1994). Auch die Abkühlung zu Beginn
des Tertiärs, der vermutlich 75% der Faunen-Arten zum Opfer fielen (Schönwiese, 1994),
kann die Kontinentalverschiebung zur Ursache haben. Jedoch wird hier auch ein Meteoriten-
einschlag auf Yucatan als primäre Ursache in Betracht gezogen. Auch wenn dieser die
Hauptursache war, hat die Lage Antarktis am geographischen Südpol und die Gruppierung
großer Landgebiete um den geographischen Nordpol (Nordamerika, Grönland, Asien) den
Übergang ins heutige Eiszeitalter gefördert.
Auswirkungen der Stellung der Platten auf die Ozeanzirkulation
Die Änderung der Ausdehnung und Tiefengestaltung der Ozeane über die Erdgeschichte wird
von Schwarzbach (1993) als paläoklimatischer Faktor ersten Ranges bezeichnet, da durch sie
die gesamte atmosphärische Zirkulation und der Verlauf der für das Klima so wichtigen Mee-

18
resströmungen beeinflußt wird. Experimente mit Ozeanzirkulationsmodellen konnten etwa
belegen, daß Veränderungen der Position der Kontinente einen signifikanten Einfluß auf den
für das Globalklima bedeutsamen polwärts gerichteten Wärmetransport haben können (Sei-
dov, 1986). Dieses belegen etwa die Untersuchungen von Maier-Reimer u.a. (1990). Anhand
eines Ozean-GCMs stellten sie für den Fall eines offenen Isthmuses zwischen Nord- und Süd-
amerika, wie er etwa bis vor 3 Millionen bestanden hat, zwei signifikante Effekte fest. Zum
einen würde der Wegfall der westlichen Barriere des Atlantiks den Golfstrom schwächen.
Darüber hinaus würde sich auch die 1,5 ‰-Differenz des Salzgehalts zwischen Nordpazifik
und Nordatlantik annähernd ausgleichen. Der Salzgehalt des Nordatlantik würde zurückgehen,
wodurch das Wasser weniger anfällig gegen vertikale Konvektion wäre, die momentan als
positive Rückkopplung auf den Wärmetransport wirkt (vgl. Broecker u.a. (1985)). Diese Ef-
fekte führten im Modell zu einem Rückgang des polwärtigen Wärmetransport und schließlich
zu Eisbildungen in der Norwegischen See.
Vergleich des polwärtigen Wärmetransports im Atlantik für eine Ozean-GCM-
Simulation mit gegenwärtigen Begrenzungen bzw. mit einem offenen zentral-
amerikanischen Isthmus
Erstellt nach Ergebnissen von Maier-Reimer u.a., 1990
Quelle: Crowley und North (1991)
Auch die Entwicklung der Vereisung auf dem antarktischen Kontinent scheint nach Hart-
mann (1994) durch die Drift der Kontinente und die dadurch bedingte Veränderung der
Ozeanzirkulation beeinflußt worden zu sein. Obwohl der antarktische Kontinent schon mehre-
re Millionen Jahre über dem Südpol lag, war er bis vor etwa 20 Millionen Jahren von einem

19
kühlgemäßigten Klima geprägt, daß den Wuchs von Wäldern zuließ. Durch die Nord-
wärtsdrift von Südamerika und Australien wurde jedoch mit der Öffnung der Drake-Passage
auf der Südhalbkugel eine zirkumpolare Ozeanzirkulation möglich. Diese führte nach Hart-
mann (1994) dazu, daß sich Oberflächen- und Tiefengewässer um mehr als 10°C abkühlten.
Genau zu dieser Zeit entstanden Gletscher, die schließlich vor 14 Millionen Jahren über der
Ostantarktis einen Eisschild formten, der heutige Vereisungsstand wurde etwa vor 5 Millionen
Jahren erreicht. Dem widersprechend hat Oglesby (1989) jedoch ermittelt, daß eine 10° C
wärmere antarktische zirkumpolare Ozeanzirkulation nur in den Küstenregionen einen signi-
fikanten Effekt hätte, im Inneren würde die durchschnittliche Temperatur weiter unter Null
liegen. Das wärmere Klima würde zu vermehrten Schneefällen und zu einer Erhöhung der
Albedo führen, welche negativ auf die Erwärmung im Sommer wirken.
Zonal gemittelte mit GCM berechnete Temperaturen für die hohen Breiten der
Südatmosphäre basierend auf vier verschiedene Experimente
(1) mit einer um 10° C wärmeren zirkumpolaren Ozeanströmung und der heuti-
gen Topographie, (2) mit den heutigen Strömungsverhältnissen, aber ohne
topographisches Relief, (3) mit einer um 10° C wärmeren zirkumpolaren Oze-
anströmung, jedoch ohne topographisches Relief, (4) heutige
Strömungsverhältnisse und Topographie (erstellt nach Oglesby, 1989).
Quelle: Crowley und North (1991)
Es ist also noch schwierig, allgemeingültige Hypothesen bezüglich des Einflusses des
Wärmetransports durch Ozeane auf Klimaänderungen aufzustellen (vgl. auch Matthews und
Poore, 1980). Die Modelle und die ihnen zugrundeliegenden Daten beinhalten noch zu viele
Unsicherheiten, so daß weitere Untersuchungen notwendig sind (Crowley und North, 1991).

20
2.2.2.1.2 Orogenese
Die Orogense hat wahrscheinlich auch Einfluß auf die Langzeitentwicklung des Klimas
(Crowley und North (1991); vgl. auch Ruddiman u.a. 1986a; Ruddiman und Raymo, 1988).
Anhand von General Circulation Models (GCM) können etwa Aussagen gemacht werden,
inwieweit sich die Gebirgsbildung auf das Klima auswirken kann. Betrachtet man etwa die
Hebung des westlichen Nordamerikas, der Anden und des Himalaya seit dem Pliozän, so hat
diese signifikant die atmosphärische Zirkulation beeinflußt (Manabe und Terpstra, 1974;
Kutzbach u.a., 1989; Manabe und Broccoli., 1990).
Effekte der Veränderung der Topographie auf den saisonalen Bodendruck in
der nördlichen Hemisphäre.
Die Resultate sind von GCM-Simulationen für Szenarien jeweils mit (M) oder
ohne Gebirgsbildung (NM) (Gebiete mit Gebirgsbildung sind gepunktet). Die
dicken Pfeile zeigen den ermittelten Verlauf der Jet-Streams auf (nach Rud-
diman und Kutzbach, 1989).
Quelle: Crowley und North (1991)

21
Die Orogenese bedingt auch durch die Bildung der jungen Faltengebirge hohe gletscherfähige
Terrains, auf denen Schnee ganzjährig liegen bleibt. Dieses führt zu obenbeschriebenen Eis-
Albedo-Rückkopplungen, weswegen die jüngste alpidische Gebirgsbildung von Schönwiese
(1994) als eine Ursache für den Übergang in das Eiszeitalter des Quartärs angesehen wird.
2.2.2.1.3 Vulkanismus
Beim Vulkanismus ist besonders der explosive von Bedeutung, der Partikel und Gase in die
Stratosphäre, in extremen Fällen sogar in die Mesosphäre schleudert. Klimawirksam sind hier
besonders Sulfatpartikel, die in diesen Schichten aus schwefelhaltigen Gasen entstehen und
Aerosolschichten bilden, die sich rasch west-ostwärts und langsam nord-südwärts ausbreiten.
So können insbesondere tropische Vulkanausbrüche global wirksam werden. Durch die Aero-
solschichten kommt es zu verstärkter Absorption und Streuung der solaren Einstrahlung, was
zu einer Erwärmung der Stratosphäre führt. Dagegen werden die bodennahen Luftschichten
durch die verringerte atmosphärische Transmission auf die Erdoberfläche abgekühlt (Schön-
wiese, 1994).
2.2.2.1.4 Synthese anhand der zeitlichen Wirkung
Als Synthese ziehen Crowley und North (1991) hinsichtlich der zeitlichen Wirkungen plat-
tentektonischer Prozesse auf klimatische Änderungen folgende Schlüsse:
Die Drift der Kontinente und die durch sie bewirkte Änderung der ozeanischen Zirkulation
sind ein wichtiger Klimafaktor für Fluktuationen im Zeitrahmen von 107-8 Jahren. Intensive
tektonische Fluktuationen werden von Crowley und North (1991) als eine mögliche Ursache
für die mit hoher Signifikanz auftretenden Klimaschwankungen innerhalb von 106 Jahren
vermutet. Vulkanismus kann etwa über einen Zeitraum von 100-2 Jahren auf das Klima wir-
ken. Daneben gibt es noch diverse indirekt nicht nur durch die Plattentektonik bewirkte
Prozesse wie interne Interaktionen im Land-See-Luft-Eis-System, die sich gegenseitig auf-
schaukeln können und in einer Zeitskala von 103-5 Jahren auf das Klima wirken (Crowley und
North, 1991).
2.2.2.2 Regional: Geographischer Formenwandel
Die Relativlage der Kontinente, die Verteilung von Land und die Lage von Gebirgen ist von
großer Bedeutung für die auszumachenden geographischen Formenwandel, die regelhafte

22
Abwandlung der Geoökofaktoren über die Erde in Raum und Zeit. Dieses soll hier ansatzwei-
se skizziert werden.
So läßt sich vom Äquator zu den Polen ein durch die Höhe der Sonneneinstrahlung be-
dingter Wandel von tropischen über gemäßigte bis hin zu polaren Klimaten ausmachen. Die
latitudinale Verschiebung von Kontinenten, Kontinentalschelfen oder Ozeanbecken kann die-
se daher in eine andere Klimazone bringen, was auf ihnen zu einer Klimaänderung führt, wie
exemplarisch anhand der Klimageschichte Australiens aufgezeigt wird (Schwarzbach, 1993).
Klimageschichte Australiens
Oben: aus geologischen Klimazeugen rekonstruiert. Dargestellt ist die Lage
des Kontinents zu den verschiedenen Klimagürteln. Unten: Änderung der Brei-
tenlage nach paläomagnetischen Messungen; nach Irving (1964) & Green (1961)
(absolute Datierung geändert). Beide Diagramme stimmen wenigstens seit dem
Devon auffällig überein, obgleich sie auf völlig verschiedene Weise kon-
struiert wurden. (...) (nach Schwarzbach, 1968)
Quelle: Schwarzbach, 1993
Gebirge bewirken einen hypsometrischen Formenwandel, der durch einen vertikal sinken-
den Temperaturgradienten infolge eines durch Abnahme der Dichte der Atmosphäre
veränderten Strahlungshaushaltes geprägt ist. Dieser kann sogar in den Tropen von tropischem
Klima über gemäßigte Gebiete bis zu Polarklimaten in Hochgebirgen wie etwa den Anden
reichen.

23
Ein zentral-peripherer Formenwandel ist insbesondere in den Mittelbreiten und in der borea-
len Zone auszumachen. Da Meere sich nicht so schnell erwärmen wie Kontinente, umgekehrt
die Wärme aber auch länger speichern, sind Küstengebiete in der Regel von geringeren Tages-
und Jahresschwankungen der Temperatur geprägt als Gebiete in den Kontinenten. Darüber
hinaus kann es im Regenschatten von Gebirgen oder in Lagen, die zu weit von der Küste ent-
fernt liegen, um von wasserführenden Luftmassen erreicht zu werden, zur Genese von Wüsten
kommen. Über großen Landmassen wie Eurasien können sich darüber hinaus im Sommer
thermische Tiefdruckgebiete bilden, die Passate über die innertropische Konvergenzzone hin-
aus anziehen, die durch den Flächendivergenzeffekt zu instabilen Monsunwinden werden. So
waren Großkontinente wie Pangaae oder Gondwana von Megamonsunen und im Zentrum von
großen Temperaturschwankungen geprägt (Huggett, 1991).
Durch die Corioliskraft bewirkt sich ein öst-westlicher Formenwandel. Sie hat zur Konse-
quenz, daß in den mittleren Breiten vorherrschend Westwinde wehen. So sind Westküsten von
zyklonaler Tätigkeit dieser Winde geprägt, die oft warme Luftmassen aus Südwesten heran-
führen. Ostküsten sind dagegen oft von polaren Kälteeinbrüchen und insgesamt von höheren
saisonalen Schwankungen betroffen. In niederen Breiten entstehen durch die Corioliskraft an
den Westküsten von Ozeanen Meeresströmungen, die kalte Wassermassen aus polaren Ge-
bieten bis in die Tropen bringt. Das widerum führt dort zur Abkühlung und
Küstennebelbildung, wodurch Küstengebiete keine wasserführenden Luftmassen erhalten, was
die Entstehung von Küstenwüsten zur Folge hat.
2.2.3 Wirkung auf eustatische Meeresspiegelschwankungen
Eustatische Meeresspiegelschwankungen werden in einem Zeitraum von 101-5 Jahren (Briggs
u.a., 1997) in erster Linie durch klimabedingte Änderungen des globalen Wasserhaushalts
bewirkt, wobei hauptsächlich die Verschiebung des Anteils zwischen dem festen und dem
flüssigen Aggregatzustand von Bedeutung ist.
Daher ist insbesondere der erdgeschichtliche Wechsel zwischen Warm- und Kaltzeiten von
Interesse, für die plattentektonische Prozesse eine der primären Ursachen sind (siehe 2.2.2.1).
Dieses kann jedoch nur als Faktor für langfristige Meeresspiegelschwankungen, wie etwa dem
Rückgang des Meeresspiegels zwischen der Kreidezeit und dem Quartär, herangezogen wer-
den.
Bedeutender sind aber die zwischen Eiszeiten und Interglazialen auftretenden Schwankun-
gen, die nicht direkt auf Plattentektonik zurückgeführt werden können. So war im letzten

24
Zyklus von Eiszeit und Zwischeneiszeit über 125.000 Jahre der Meeresspiegel durch den
Aufbau von Gletschern einer Schwankung von 130 - 165 m unterworfen (Briggs u.a., 1997).
Die Veränderung kann dabei in einigen Zeiten sehr schnell vorangehen. So stieg der Meeres-
spiegel zwischen 15.000 und 9000 BP von - 130 auf - 60 m (Briggs u.a., 1997).
2.2.4 Synthese
Zusammenfassend läßt sich sagen, daß plattentektonische Prozesse in der Regel erdgeschicht-
lich langfristig wirken. In diesem Zeitraum haben sie einen prägnanten Einfluß auf die Genese
und Lage der Großformen der Erde, auf die Verteilung von Land und See und auf das Klima.
Vulkanausbrüche können darüber hinaus aber auch kurz- und mittelfristig regional bis global
wirksam sein.
Ausgehend von der These, daß „Life fits the world“ vergleicht Whitfield (1993) die durch
plattentektonische Prozesse bewirkten Änderungen daher mit einem Schachspiel, in dem sich
die schwarzen und weißen Feldern andauernd ändern. Bewegungen, die einen bestimmten
Satz an Regeln benötigen, können dadurch unmöglich werden bzw. ermöglicht werden. So hat
die Plattentektonik in der Erdgeschichte einige Interaktionen erlaubt, andere verhindert und
dadurch die makroevolutionäre Geschichte prägend mitbestimmt.
2.3 Einführung in die Biogeographie mit besonderer Berücksichtigung
der historischen Biogeographie
Nach Myers und Giller (1988) läßt sich die Bandbreite biogeographischer Forschung zwi-
schen zwei Eckpunkten festmachen, der ökologischen und der historischen Biogeographie
(vgl. Candolle (1820)). Die ökologische Biogeographie beschäftigt sich mit ökologischen Pro-
zessen, die (in der Regel heute beobachtbar) in kurzen Zeiträumen in kleinräumlichen
Maßstäben auftreten, während die historische Biogeographie auf der anderen Seite des Spekt-
rums evolutionäre Prozesse erforscht, die innerhalb von mehreren Millionen Jahren
stattgefunden haben. (Myers und Giller, 1988). Eine detailliertere Einteilung nimmt Rosen
(1988a) vor, der die Konzepte der Biogeographie in Aufrechterhaltung der Art (Maintenance);
Änderung des Verbreitungsmusters (Distributional Change) und Enstehung neuer Arten (Ori-
ginations) einreiht, wobei ersteres eher der ökologischen und letzteres der historischen
Biogeographie zuzuordnen sind, während sich bei der Änderung der Verbreitungsmuster die
Interessen überschneiden.

25
Innerhalb der beschriebenen Bandbreite gibt es eine Vielzahl von Theorien, Hypothesen und
Modellen, die versuchen, heutige oder ehemalige Verbreitungsmuster zu erklären. Myers und
Giller (1988) bemängeln hierbei jedoch eine oft mangelnde Interaktivität zwischen For-
schungsergebnissen. Als Ursache hierfür vermuten sie, daß es keine Studiengänge für
Biogeographie gibt, so daß Biogeographen heute aus unterschiedlichen Disziplinen kommen
(etwa Botaniker, Genetiker, Geologen, Geographen, Paläobiologen, Paläogeographen, Palä-
ontologen, Paläoökologen, Taxonomisten, Zoologen). Sie verfolgen bei ihren Forschungen oft
sehr unterschiedliche Fragestellungen und Ziele, wodurch sich ihre Forschungsergebnisse
vielfach nur peripher oder gar nicht überschneiden. Ihnen fehlt oft das Verständnis für das
ganze Spektrum ökologischer und historischer Prozesse, was Myers und Giller (1988) jedoch
als fundamentale Voraussetzung für die Untersuchung von Verbreitungsmustern von Orga-
nismen ansehen.
Um einer weiteren Fragmentierung dieses interdisziplinären Feldes entgegenzuwirken,
sollte nach Myers und Giller (1988) die Biogeographie versuchen, die divergenten Interessen
zu integrieren. Sie sollte determinieren, inwieweit Artenbildung, Adaption, Extinktion und
ökologische Prozesse sich gegenseitig beeinflussen, bzw. durch geologische oder klimatische
Prozesse bedingt sind (Myers und Giller, 1988).
Entwicklung der Historischen Biogeographie
Der Wissenschaftszweig der historischen Biogeographie hat seinen Ursprung in der Debatte
über Hypothesen, wie die Erde nach der Sintflut von den Tieren aus Noahs Arche wieder be-
siedelt wurde, nachdem diese am Berge Ararat gestrandet war. Auch wenn die grundlegende
Annahme der Debatte bald verworfen wurde, regte die Diskussion Darwin und Wallace zur
Entwicklung der Hypothese an, daß alle Arten von Taxa ein Ursprungszentrum haben, von
dem aus sie sich ausbreiten (centre-of-origin-dispersal-model) (Darwin, 1872; Wallace, 1876).
Darwin argumentierte dabei, daß während des Ausbreitungsprozesses Grenzen die Bewe-
gung der Spezies aufhalten, so daß dieser Ablauf langsam genug stattfindet, um natürliche
Selektion zu verursachen und die Art zu verändern. Dieses Modell wurde insbesondere durch
Mayr, Simpson und Darlington zu einer grundlegenden Theorie der Biogeographie weiterent-
wickelt (Huggett, 1998).
Jedoch richteten sich schon bald Kritiken gegen eine Hegemonie der „Ausbreitungsbioge-
ographie“, die schließlich von Croizat in seiner Vikarianzhypothese manifestiert wurden.
Croizat testete das darwinistische Centre-of-origin-dispersal Modell anhand der Verteilung

26
hunderter Pflanzen- und Tierarten (Croizat 1958, 1964). Hierbei kam er zu dem Ergebnis, daß
Arten mit sehr unterschiedlichen Ausbreitungsneigungen und Kolonisierungsfähigkeiten eine
sehr ähnliche geographische Verbreitung hatten. Er nannte diese gemeinsamen Verbreitungs-
muster generalisierte bzw. standardisierte Routen, die jedoch nicht Migrationslinien
entsprächen. Vielmehr deutet er die heutigen Verbreitungsmuster als Teilmenge bzw. als indi-
viduelle Überbleibsel ehemals zusammenhängender Muster. Diese von ihm Vikarianz
genannte Fragmentierung konnte Vorgänge der Plattentektonik, Änderungen des Meeresspie-
gelniveaus oder Klimaänderungen als Ursache haben.
Auch Anhänger dieser Hypothese gehen davon aus, daß sich Spezies ausbreiten müssen,
um eine weiträumige Verbreitung zu erhalten, daß diese aber zum überwiegenden Teil dann
passiert, wenn keine Barrieren (Wüsten, Ozeane, Kontinente, Berge, Täler o.a.) vorhanden
sind. Eine Verbreitung auch über Barrieren hinaus ist möglich, wird jedoch eher als unsignifi-
kante Ausnahme angesehen (Huggett, 1998).
Synthetiker, die beide Theorien verbinden, halten die Vikarianzhypothese im historischen
Rahmen für wichtig, in der Gegenwart ist für sie jedoch die Verbreitung von größerer Bedeu-
tung. Letztere ist eine alltägliche Erscheinung, die von fast allen Spezies vorgenommen wird,
während Vikarianz viel seltener auftritt, da in ihr eine Barriere entstehen muß, die zusammen-
hängende Ausbreitungsmuster trennt. Wichtig ist hierbei, daß vikariante Vorgänge
gleichzeitig für andere Lebewesen neue Ausbreitungsmöglichkeiten bieten. So ermöglichte
etwa die Entstehung des Isthmus von Panama als Landbrücke zwischen Nord- und Südameri-
ka im Pliozän einen einfachen Austausch von Land- und Frischwasserorganismen.
Gleichzeitig wurden jedoch die Populationen der tropischen Meere in der neuen Welt in zwei
Teile geteilt: Die des Pazifiks westlich und die des Atlantiks östlich des Isthmus. So versteht
Briggs Ausbreitung als einen andauernder Prozeß, während Vikarianz für ein Habitat norma-
lerweise Ausbreitung in einem anderen stimuliert (Briggs, 1995).
3 Konsequenzen der Plattentektonik für die BiogeographieWie unter 2.3 aufgezeigt wurde, ist die Biogeographie kein homogenes Forschungsfeld, in ihr
überschneiden sich die Interessen verschiedenster Fächer. Wenn es also nicht „die Bioge-
ographie“ gibt, ist es schwierig, die Konsequenzen der Plattentektonik für sie zu erörtern,
ohne sich in den differierenden Auffassungen zu verlieren. Daher orientiert sich meine Arbeit
an der von Rosen (1988a, 1988b) aus den unterschiedlichen Fächern zusammengetragenen
Aufstellung biogeographischer Konzepte.

27
Waren bis zu Croizat in der Biogeographie Verbreitungskonzepte bestimmend, die auf der
heutigen Lage der Kontinete basierten, so hat sich das in den letzten 40 Jahren auch durch die
Einführung der Theorie der Plattentektonik in die allgemeine Lehrmeinung stark verändert. So
beinhalten heute selbst viele ökologische Konzepte erdhistorische Elemente, wie die folgende
Tabelle zeigt. In welcher Form die Konzepte die Erdgeschichte berücksichtigen, wird in die-
sem Abschnitt erörtert.
Konzepte und Prozesse
Konzepte der Biogeographie Einzuordnen in
Aufrechterhaltung der Art a) b) c) d)
Ausbreitung X
Geoökologie X X
Gleichgewichtstheorie und Inselbiogeographie X
Barrieren X X
Geoökologische Zusammensetzung X
Provinzen und Provenzialität X X X
Änderung des Verbreitungsmusters a) b) c) d)
Vergrößerung des Verbreitungsgebietes X X
Sprungverbreitung über Barrieren hinweg X X X
Gleichgewichtstheorie und Inselbiogeographie X X X
Erdgeschichte (TEKO1-Ereignisse und Palaeogeographie) X X X X
Verbreitungsspuren als Resultat ökologischer Veränderungen X X
Verbreitungsspuren als Resultat durch erdgeschichtliche Er-
eignisse bedingter ökologischer VeränderungenX X X X
Anpassung einer Taxa an veränderte Umweltbedingungen X X
Entstehung neuer Arten a) b) c) d)
Empirische phylogenetische Biogeographie X X X
Vikarianz als Resultat erdgeschichtlicher Veränderungen X X X
Vikarianz als Resultat durch erdgeschichtliche Ereignisse
bedingter ökologischer VeränderungenX X X
Randisolierung als Resultat ökolgischer Veränderungen X
Randisolierung als Resultat durch erdgeschlichte Ereignisse
bedingter ökologischer VeränderungenX X
Sprungverbreitung über Barrieren hinweg X X
Entstehungsgebiete neuer Arten X X
Geoökologie über die Zeit X X X
a) Ökologische Biogeographie, b) Historische Biogeographie, c) Von der Erdge-
schichte beeinflußte Konzepte, d) Konzepte mit praktischen Wert für die
Rekonstruktion der Erdgeschichte1 TEKO = Tektonische, Eustatische und Klimatische oder Ozeanographische
Quelle: Zusammengefügt aus Rosen (1988a) und Rosen (1988b)

28
3.1 Konzepte der Aufrechterhaltung der Art
Die Konzepte der Aufrechterhaltung der Art sind überwiegend ökologischer Natur. Rosen
(1988a) definiert sie als Prozesse, mit denen eine Art ihre Präsenz in einem bestimmten
Verbreitungsgebiet aufrechterhält. Das schließt auch alle Faktoren ein, die das Überleben in
diesem Raum effektiv determinieren. Diese Gegebenheiten treten innerhalb der Lebensspanne
eines einzelnen Organismus auf, sind also erdhistorisch als sehr kurzfristig einzuordnen.
Man kann anhand dieser Konzepte auch Hypothesen entwickeln, ob die Abwesenheit einer
Art an einem bestimmten Ort durch einen vorübergehenden Wandel der Umweltbedingungen
verursacht ist, welcher auch durch plattentektonische Prozesse bedingt sein kann. Dabei kann
sich zum einen das Ökosystem seit dem störenden Ereignis bis zum beobachteten Zeitpunkt
noch nicht wieder erholt haben, zu anderen kann es sein, daß sich die Population trotz einer
Wiederherstellung der Umweltbedingungen noch nicht wieder erholt hat (Rosen, 1988a).
3.1.1 Geoökologie
Die Geoökologie umfaßt sowohl das Verhalten und die ökophysiologischen Prozesse, durch
die eine Taxa die Aufrechterhaltung seiner Art innerhalb eines bestehenden Ausbreitungsge-
biet sichert, als auch die Umweltfaktoren, die seine Präsenz begünstigen (Rosen, 1988a).
Umgekehrt ist aber auch jedes Verbreitungsgebiet durch limitierende Faktoren begrenzt (Hug-
gett, 1998). Diese sind zwar selten direkt auf plattentektonische Prozesse zurückzuführen,
jedoch sind sie für das Verständnis vieler Konzepte wichtig, weshalb sie hier dargestellt wer-
den.
Limitierende Faktoren
Der Begriff des limitierender Faktor geht auf Liebig (1840) zurück. Er hatte anhand des
Wachstums einer Feldfrucht festgestellt, daß sie für ihr Wachstum diverse Nährstoffe wie
etwa Phosphor oder Stickstoff braucht. Wenn aber auch nur einer dieser Nährstoffe nicht in
ausreichendem Maße vorhanden ist, würde das Wachstum verlangsamt.
Aus dieser Erkenntnis entwickelte er das Gesetz des Minimums: Produktivität, Wachstum
und Reproduktion von einer Population werden eingeschränkt, wenn einer oder mehrere Um-
weltfaktoren unter dem optimalen Niveau liegen. Diesem haben später Ökologen ein Gesetz
des Maximums entgegengesetzt, da etwa zu viel Phosphor auf einem Feld genauso das
Wachstum einschränkt, wie zu wenig (Huggett, 1998).

29
Ein limitierender Faktor muß nicht gleich tödlich für eine Art wirken, er macht einfach die
Arbeit der Physiologie oder das Verhalten weniger effizient. Dieses kann neben einer niedri-
geren Reproduktionsrate auch zu Benachteiligungen im Kampf mit anderen Lebensformen um
Essen oder Lebensraum führen (Cox u.a., 1976). Dabei ist auch die Toleranzrate einer Pflanze
gegenüber einem bestimmten Umweltfaktor von Bedeutung. So hat ein Kamel eine deutlich
höhere Toleranzrate gegenüber Wassermangel, als etwa eine Nacktschnecke. Innerhalb der
Toleranzrate für einen Umweltfaktor gibt es nach Shelford (1911) in der Regel eine obere und
eine untere Grenze, über bzw. unter der ein Leben nicht möglich ist, einen Optimalbereich
und eine zwischen letzteren und den Grenzen liegende Zone mit physiologischem Streß.
Toleranzbereich und seine Grenzen
Entwickelt nach Shelford (1911)
Quelle: Huggett (1998)
Ein Beispiel für ein Verhalten unter psychologischem Streß ist etwa das von Libellen an der
Nordgrenze ihres Verbreitungsgebietes in Europa, das primär durch niedrige Nachttemperatu-
ren begrenzt ist. In der Grenzregion sind die Temperaturen in der Nacht nicht so tief, daß sie
die Libellen töten würden. Jedoch werden sie infolgedessen dazu gezwungen, ihre Metamor-
phose während des Tages durchzuführen, wodurch sie eher von Vögeln gefressen werden
(Cox u.a., 1976).
Generelle limitierende Faktoren
Die limitierenden Faktoren sind unzählig und stehen in komplexen Wirkungsgefügen zuein-
ander. Sie sind nicht nur räumlich, sondern auch zeitlich großen Schwankungen unterworfen.

30
Ein wichtiger Faktor ist die Sonneneinstrahlung, wobei Intensität, Qualität und Dauer der Ein-
strahlung (u.a. bestimmt durch die saisonale Schwankung der Tageslänge) von Bedeutung
sind. Auch die Temperatur hat eine stark limitierende Wirkung. Ebenso spielen biotische
Faktoren wie Nährstoffe, Plagen, Anwesenheit von Feinden oder Mitstreitern um Ressourcen
und Störungen etwa durch Winde, Feuer, Fluten, Vulkanausbrüche oder menschliche Eingrif-
fe eine Rolle (Cox u.a., 1976 / Huggett, 1998).
An Land
Auf dem Land sind daneben insbesondere Wasser mitsamt zyklischer Schwankungen, die
Zusammensetzung des Bodens und topographische Faktoren zu beachten (Huggett, 1998).
Im Meer
Für Meeresorganismen sehen Brenchley und Harper (1998) in küstennahen Gewässern Salz-
gehalt, Substratmobilität und Turbulenz als wichtige Faktoren an, während in größeren Tiefen
in erster Linie Sonneneinstrahlung, Sauerstoffgehalt, Druck und Nahrung wirken und die
Verteilung vertikal begrenzen. Von genereller Bedeutung wird der Faktor Temperatur, der
auch gerade latitudinal die marine Biota eingrenzt (Brenchley und Harper, 1998). Hinsichtlich
der Verteilung spielen auch die Topographie und Ozeanströmungen eine Rolle.
3.1.2 Barrieren
Selten kann eine Art alle Räume besiedeln, die für ihre ökologische Toleranz geeignet sind.
Häufiger Grund dafür ist, daß dazwischen liegende Barrieren sie daran hindern. Diese können
nicht oder nur schwer überwunden werden, da einer oder mehrere limitierende Faktoren au-
ßerhalb der Toleranzgrenze sind. Eine der größten Barrieren ist die zwischen Land und Meer,
da sich in diesen Lebensräumen Bewegung, Sauerstoffaufnahme und Nahrung stark unter-
scheiden. Großräumig wirken etwa Gebirge, ozeanisch Rücken, Tiefseegräben, Wüsten oder
Regenwälder. Je nach Lebensform kann aber auch schon der Abstand von ein paar Metern
zwischen zwei Tümpeln unüberbrückbar sein (Cox u.a., 1976).
Die Verbindung des ökologischen Barrierenkonzeptes mit der Erdgeschichte führt ein
Langzeitelement in die Prozesse der Artaufrechterhaltung ein. Darüber hinaus ist es auch ein
wichtiger Teil der Konzepte der Veränderung der Verbreitung. Die Entstehung einer Barriere
nennt man ein Divergenzereignis, während das Vergehen ein Konvergenzereignis ist (eine
detailliertere Darstellung ist unter 3.1.2.3 und 3.1.2.4 zu finden).
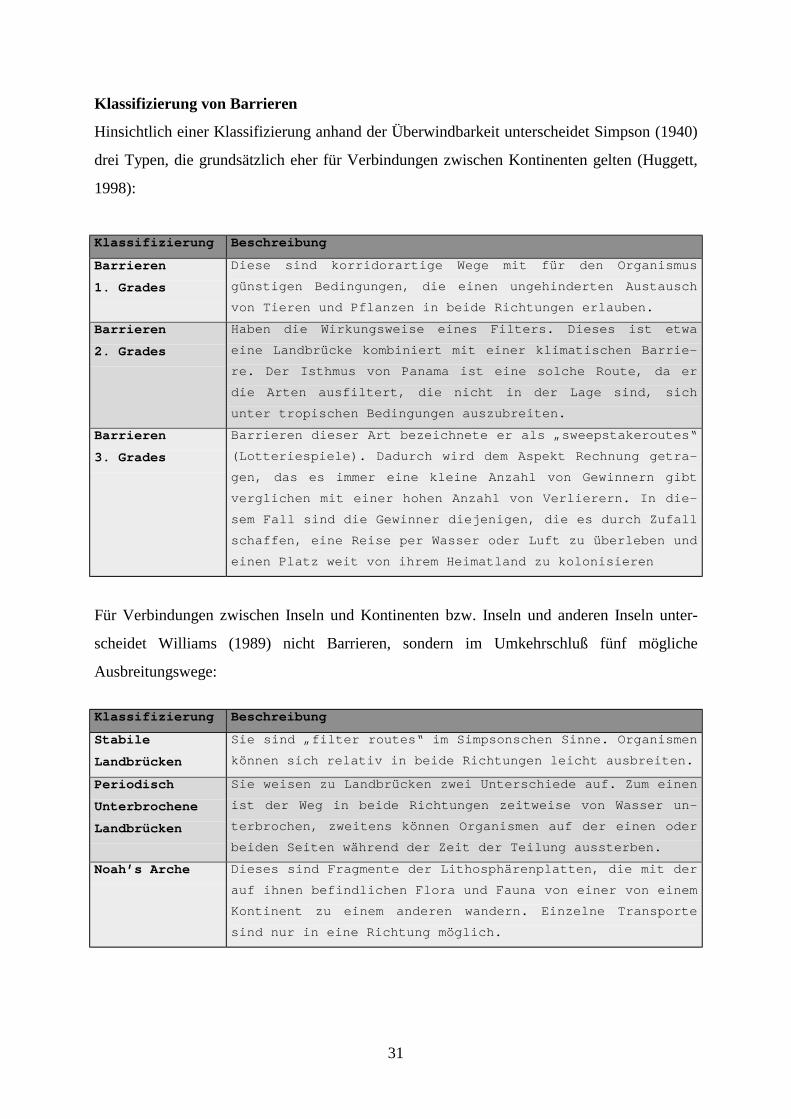
31
Klassifizierung von Barrieren
Hinsichtlich einer Klassifizierung anhand der Überwindbarkeit unterscheidet Simpson (1940)
drei Typen, die grundsätzlich eher für Verbindungen zwischen Kontinenten gelten (Huggett,
1998):
Klassifizierung Beschreibung
Barrieren
1. Grades
Diese sind korridorartige Wege mit für den Organismus
günstigen Bedingungen, die einen ungehinderten Austausch
von Tieren und Pflanzen in beide Richtungen erlauben.
Barrieren
2. Grades
Haben die Wirkungsweise eines Filters. Dieses ist etwa
eine Landbrücke kombiniert mit einer klimatischen Barrie-
re. Der Isthmus von Panama ist eine solche Route, da er
die Arten ausfiltert, die nicht in der Lage sind, sich
unter tropischen Bedingungen auszubreiten.
Barrieren
3. Grades
Barrieren dieser Art bezeichnete er als „sweepstakeroutes“
(Lotteriespiele). Dadurch wird dem Aspekt Rechnung getra-
gen, das es immer eine kleine Anzahl von Gewinnern gibt
verglichen mit einer hohen Anzahl von Verlierern. In die-
sem Fall sind die Gewinner diejenigen, die es durch Zufall
schaffen, eine Reise per Wasser oder Luft zu überleben und
einen Platz weit von ihrem Heimatland zu kolonisieren
Für Verbindungen zwischen Inseln und Kontinenten bzw. Inseln und anderen Inseln unter-
scheidet Williams (1989) nicht Barrieren, sondern im Umkehrschluß fünf mögliche
Ausbreitungswege:
Klassifizierung Beschreibung
Stabile
Landbrücken
Sie sind „filter routes“ im Simpsonschen Sinne. Organismen
können sich relativ in beide Richtungen leicht ausbreiten.
Periodisch
Unterbrochene
Landbrücken
Sie weisen zu Landbrücken zwei Unterschiede auf. Zum einen
ist der Weg in beide Richtungen zeitweise von Wasser un-
terbrochen, zweitens können Organismen auf der einen oder
beiden Seiten während der Zeit der Teilung aussterben.
Noah’s Arche Dieses sind Fragmente der Lithosphärenplatten, die mit der
auf ihnen befindlichen Flora und Fauna von einer von einem
Kontinent zu einem anderen wandern. Einzelne Transporte
sind nur in eine Richtung möglich.

32
Klassifizierung Beschreibung
Trittstein-
Inseln
Sie sind eine Serie relativ permanenter bzw. temporärer
Inseln, unterbrochen von schmalen bis zu mittleren Gewäs-
sern. Der Austausch könnte in beide Richtungen erfolgen,
meistens erfolgt er jedoch von Inseln mit höherer Biodi-
versität zu Inseln mit niedrigerer.
Ozeaninseln Diese befinden sich weit entfernt von anderen Landflächen.
Sie sind den Trittsteininseln ähnlich, auf ihnen finden
jedoch viel seltener Ankünfte von neuen Organismen statt.
Außerdem ist die Wahrscheinlichkeit auch stark abhängig
von den Kolonisationsfähigkeiten der jeweiligen Arten.
Letztendlich ist jedoch für Cox u.a. (1976) die ultimative Barriere in der Regel nicht von
feindlichen Umweltfaktoren bedingt, sondern die jeder Art eigenen Physiologie, welche sie in
die Lage versetzt, eine mehr oder weniger limitierte Schwankungsbreite der Werte einzelner
Umweltfaktoren zu tolerieren. So ist eine Art hinsichtlich ihrer Ausbreitung immer ein Gefan-
gener ihrer eigenen evolutionären Entwicklung (Cox u.a., 1976). Umgekehrt sind sie aber
auch durch die Evolution in der Lage, sich an Amplitude, Frequenz und Voraussagbarkeit von
Umweltfluktuationen anzupassen (Futuyma, 1990).
3.1.3 Provinzen
„Why doth Africa breed so many venomous beasts, Ireland none? Athens owls, Crete
none? Why hath Daulis and Thebes no swallows (so Pausanias informeth us) as well as
the rest of Greece, Ithaca no hares, Pontus (no) asses, Scythia (no) wine? Whence co-
mes this variety of complexions, colours, plants, birds, beasts, metals, peculiar to almost
every place.“ Robert Burton (1628)
Die Idee von biogeographischen Provinzen ist eine der ältesten der Biogeographie (Nelson
und Platnick, 1981). Die Abgrenzung einzelner Provinzen kann auf unterschiedliche Art und
Weise vorgenommen werden. Zum einen ist es möglich, sie anhand der An- oder Abwesen-
heit von Daten bestimmen. Dabei kann man sowohl Regionen mit einer hohen Anzahl
endemischer (nur in einem bestimmten Areal zu findender) Arten festlegen oder auch statisti-
sche geographische Zusammensetzungen suchen. Ein anderer Ansatz betrachtet Provinzen als
endemische Regionen, deren charakteristische Taxa sich dadurch auszeichnet, daß sie von
gleichen biogeo-graphischen Barrieren begrenzt ist. Diese Definition erklärt Provinzen eher
durch historische und evolutionäre Prozesse wie Änderung des Verbreitungsgebietes, Geolo-

33
gie und Ökologie von Barrieren oder allopatrische Artenbildung (siehe 4.1.3), als durch die
pure Ökologie.
Provinzen wurden aber nicht nur über die An- oder Abwesenheit von Daten bestimmt, es
besteht auch eine lange Tradition, Verbreitungsdaten mit geographischen oder geologischen
Charakteristiken zu verbinden, um Demarkationslinien festzulegen. Dieses liegt wohl daran,
daß die Verbreitungsgebiete von Organismen oft ein Kontinuum von überlappenden Mosaiken
darstellen, wodurch es oft zu kontroversen Debatten kommt, wo denn nun die Grenze zu zie-
hen sei (Rosen, 1988a).
Die heutigen biogeographischen Provinzen
Wenn man die von Pielou (1979) durch Synthese aus phyto- und zoogeographischer Abgren-
zungen abgeleiteten globalen terrestrischen Provinzen mit plattentektonischen Ereignissen
oder deren Rückkopplungen in Relation setzt, sind auffällige Übereinstimmungen festzustel-
len.
Biogeographische Regionen Welt, ermittelt durch die Kombination traditio-
neller zoogeographischer und phytogeographischer Reiche
Nach Pielou (1979)

34
Quelle: Archibold, 1995
So liegt zwar die Konvergenz Indiens mit Asien erdgeschichtlich zu weit zurück, um Indien
als eigenständige biogeographische Provinz einzuordnen. Dafür scheint der durch die Kollisi-
on entstehende Himalaya die Nordgrenze der Region Oriental zu bilden. Auch der hohe Anteil
von endemen Arten in der australasiatischen Provinz erklärt sich daraus, daß die Landmassen
über 30 Millionen Jahre isoliert waren. Erst vor 15 Millionen Jahren begann die Kollision mit
Asien, wobei aber bis heute zu keiner Zeit eine durchgehende Landbrücke bestanden hat (vgl.
Whitmore, 1981).
Vermutete Zeiten (Million Jahre vor heute), in denen Konvergenz- oder Di-
vergenzereignisse durch Kontinentaldrift stattgefunden haben
Nach Pielou (1979)
Quelle: Archibold (1995)
3.2 Konzepte der Änderung des Verbreitungsmusters
Die Änderung des Verbreitungsmusters umfaßt alle Prozesse, bei der sich die Verbreitung
eines Organismus über die Zeit räumlich verändert (Rosen, 1988a). Dieses kann das Resultat
von ökologischen, TEKO-Prozessen oder Kombinationen der beiden Formen sein.

35
Das Konzept birgt jedoch ein durch plattentektonische Prozesse bedingtes methodisches
Problem: Wenn sich die geographische Verbreitung einer Art auf einem Kontinent nicht ver-
ändert hat, sich jedoch der Kontinent im Verhältnis zu den anderen Kontinenten verschoben
hat, kann dann von einer Veränderung des Verbreitungsgebietes gesprochen werden? Hier
stellt sich die Frage nach der geographischen Bezugsgröße, wenn diese über die Zeit Verände-
rungen unterworfen ist (Rosen, 1988a).
Auch besteht die Gefahr der Zirkularität, wenn man etwa das Verbreitungsgebiet einer Ta-
xa anhand einer bestimmten Drift der Kontinente erklärt, deren Bewegung gerade
hypothetisch mit Hilfe der Ausbreitung der Art rekonstruiert wurde (Rosen, 1988a).
3.2.1 Sprungverbreitung über Barrieren hinweg
Das Konzept Sprungverbreitung basiert explizit darauf, daß eine Art von einer Ausbreitung
durch eine Barriere behindert wird. Jedoch gelingt es mit der Zeit durch eine sehr unwahr-
scheinliche Sprungverbreitung einem oder mehreren Organismen dieser Art, einen geeigneten
Lebensraum hinter der Barriere zu erreichen. Wenn zwischen solchen Ereignissen eine längere
Zeitspanne liegt, kann das zur Artenbildung führen (siehe 3.1.3.5). So kann dieser Prozeß auf
allen Zeitebenen von kurz- bis langfristig auftreten, wobei es am kurzfristigen Ende sich mit
Verbreitung überschneidet. Die Barriere sollte jedoch zumindest für eine signifikante Zeit-
spanne bestehen, um von einer Sprungverbreitung zu sprechen. Dabei ist auch scheinbar
Unmögliches möglich, wie dieses Beispiel belegt:
„One morning (in Glacier Park), dark streaks were observed extending downward at
various angles from saddles or gaps in the mountains to the east of us. Later in the day,
these streaks appeared to be much longer and at the lower end of each there could be
siscerned a dark speck. Through binoculars these spots were seen to be animals floun-
deringdownward in the deep, soft snow. As they reached lower levels not so far distant,
they proved to be porcupines. From every little gap there poured forth a dozen or
twenty, or in one case actually fifty five, of these animals, wallowing down to the tim-
berline on the west side. Hundreds of porcupines were crossing the main range of the
Rockies.“ Cox (1936)
Bei Konzepten der Sprungverbreitung muß heute aber auch die Rolle des Menschens be-
achtet werden, der mit Schiffen oder Flugzeugen absichtlich oder unabsichtlich Organismen
helfen kann, Barrieren zu überwinden, wie etwa die Verbreitung des europäischen Kaninchen
zeigt (Flux und Fullagar, 1992).

36
3.2.2 Gleichgewichtstheorie und Inselbiogeographie
Die Gleichgewichtstheorie und Inselbiogeographie (vgl. MacArthur und Wilson, 1967) bietet
ein Modell für die Kolonisation von neuem Terrain, insbesondere von Ozeaninseln, die durch
plattentektonischen Vulkanismus entstehen. Diese sind wie etwa die Osterinsel oft isoliert und
müssen durch Sprungverbreitung besiedelt werden. Man kann diese Konzepte sowohl als Al-
ternative, als integralen oder optionalen Teil von Verbreitungsmodellen nutzen (Rosen,
1988a).
3.2.3 Erdgeschichte (TEKO-Ereignisse und Palaeogeographie)
Konzepte der Erdgeschichte umfassen die Wirkung geologischer Faktoren, TEKO-Events und
Kombinationen derselben (vergl. Rosen 1984) und die u.a. daraus resultierende Palaeoge-
ographie (physische Geographie der Erdgeschichte).
Die einfachste Vorstellung von Erdgeschichte als Faktor des Wandels der Verbreitung ist
empirisch, basierend auf dokumentierten und über die Zeit sich verändernden Ausbreitungen
von Taxa oder Zusammensetzungen von Taxa (bis hin zu Provinzen). Dabei ist natürlich das
Problem des geographischen Bezugsrahmen zu beachten, weshalb Rosen und Smith (1988)
die Daten einfach als unverbundene, aus derselben Zeit stammende Musterpunkte betrachten,
um Vorurteile zu vermeiden. Diese würden einfließen, wenn man die gefundenen Daten
gleich in bereits ermittelten palaeogeographischen Hypothesen zur Verteilung der Kontinente
einzupassen versuchte. Aufbauend auf den Musterpunkten kann man schließlich Hypothesen
über die historische Lage der Kontinente ziehen.
Für dieses Konzept gibt es zwei wichtige Modelle, die jedoch auch für die Wirkung ande-
rer Faktoren verwendet werden können (vergleiche etwa 3.1.2.4):
Divergenzereignisse (Entstehung von Barrieren)
Sich neubildende Barrieren trennen die Populationen von Organismen in Teilpopulationen,
die sich mehr oder minder untereinander austauschen können. Die Evolution geht von da an in
den voneinander getrennten Gruppen relativ unabhängig vor sich und kann zu Lebensformen
führen, die sich untereinander stark unterscheiden können. Oft sterben Arten auch auf einer
oder beiden Seiten der Barriere aus.
Für die Forschung ist heute besonders das Auseinanderbrechen des alle heutigen großen
Landmassen umfassenden Großkontinents Pangaea vor etwa 200 Millionen Jahren von Be-

37
deutung, der über mehrere 107 Jahre bestanden hatte und der damaligen Flora und Fauna weite
Verbreitungsmöglichkeiten geboten hat (Huggett, 1998).
Konvergenzereignisse (Vergehen von Barrieren)
Generell können zwei Konvergenzereignisse unterschieden werden: In der wahren Konver-
genz werden zu dem Zeitpunkt lebende Arten zusammengebracht, die in Interaktion treten
können, in der anderen werden fossile Relikte von zu diesem Zeitpunkt bereits ausgestorbenen
Arten zusammengebracht. Auf terrestrische Lebewesen bezogen titulierte McKenna (1973)
die beiden Konvergenzformen als Noahs Arche bzw. wikingisches Leichenschiff.
Bei einer wahren Konvergenz sind nach Marshall (1981) bei terrestrischen Lebensformen
vier Prozesse möglich:
1. Es kann zu einem aktiven Wettbewerb zwischen den Spezies bzw. Arten kommen, welche
auf den beiden Kontinenten relativ ähnliche Nischen besiedelt haben. Wenn die heimische
Form effektiver bei der Nutzung der Ressourcen ist, kann der Eindringling gestoppt wer-
den. Erfolglose Invasionsversuche sind relativ selten, sie konnten bisher nur durch fossilen
Aufzeichnungen belegt werden. Der alternative Ausgang ist die erfolgreiche Invasion mit
Vertreibung der heimischen Spezies.
2. Zu einer passiven Ersetzung kann es kommen, wenn eine einheimische Spezie, die eine
Nische mit einer eindringenden teilt, ausstirbt, ohne daß es zu einem Wettbewerb zwischen
den beiden kommt. In diesem Fall hatte die überlebende Form lediglich Glück, zur richti-
gen Zeit am richtigen Ort zu sein.
3. Eine weitere Form ist die der Einschmeichelung, die auftritt, wenn eine Spezies beim ein-
dringen in ein Biom bisher unbesetzte ökologische Nischen besiedelt, ohne das dieses
Auswirkungen auf die existierende Gesellschaft hat. Solche Einschmeichler stehen ökolo-
gisch allein ohne wahrnehmbare Interaktion mit den anderen Organismen. Sie bereichern
die biotische Vielfalt eines Biosystems.
4. Wenn die eindringende und die einheimische Form bisher die gleiche Nische besetzen,
kann es auch zu einem Merkmalsverschiebung genannten evolutionären Anpassungsprozeß
kommen, in dem sich eine oder beide Arten so verändert, daß sie nebeneinander existieren
können.
Die von Marshall klassifizierten Prozesse lassen sich auch auf Meeresbiome übertragen.
Ebenso kann man sie bei der Betrachtung einer Barrierenüberwindung etwa durch Sprung-
verbreitung heranziehen. So hängt etwa bei Galápagosfinken die Dicke des Schnabels davon
ab, wieviel der drei Arten die jeweilige Insel besiedeln konnten (Futuyma, 1990).

38
Eines der Beispiele für Merkmalsverschiebung bei den Galápagos-Finken
Kommen Geospiza fortis und G. fuliginosa allopatrisch vor (auf den Inseln
Daphne und Crossman), ist die Dicke des Schnabels ähnlich, sie unterschei-
det sich jedoch auf den Inseln, auf denen beide Arten sympatrisch
vorkommen. (Verändert übernommen von Lack (1947))
Quelle: Futuyma (1990)
3.2.4 Verbreitungsspuren als Resultat durch erdgeschichtliche Ereignisse bedingten
ökologischen Wandel
Die in diesem Konzept enthaltenen Prozesse sind eine Kombination der Effekte der Erdge-
schichte auf den ökologischen Wandels, der das Verändern der Verbreitungsgebiete bewirkt.
Es können vier Hauptprozesses stattfinden, die schon unter 2.2 angedeutet wurden. Erstens
kann ein Gebiet in eine andere geoökologische Zone verschoben werden, zweitens können
etwa klimatische Veränderungen die regionale Verbreitung von ökologischen Parametern wie
der Temperatur oder der Niederschlagsrate bewirken. Drittens können ökologisch ähnliche
Zonen zusammengebracht werden, wodurch Organismen schwimmend, treibend, fliegend

39
oder gehend Gebiete erreichen, die sie vorher nicht bewohnen konnten (Konvergenz). Vier-
tens können sie durch die divergierende Entstehung von Wüsten, Gebirgen, Landbrücken u.ä.
eine homogene Region fragmentieren (Divergenz).
Der Vorteil dieses Modells gegenüber des reinen geologischen ist, daß es plausible kausale
Beziehungen zwischen der Erdgeschichte und der Verbreitung von Organismen bietet. Die
Nachteile sind eher konzeptioneller Natur, da der Vorteil der Plausibilität mit dem Verlust der
empirischen Einfachheit einhergeht, was besonders kladistische Biogeographen betonen (Ro-
sen, 1988a).
Ein Beispiel ist die Erklärung der Verbreitung des alpinen Murmeltiers (Marmota marmo-
ta), daß während der Eiszeiten auf den Ebenen Zentraleuropas heimisch war, nun aber bedingt
durch die holozäne Erwärmung nur noch in den Hochgebirgen Rückzugsgebiete findet:
Das alpine Murmeltier - ein klimatisches Relikt
Nach Bjärvall und Ullström (1986)
Quelle: Huggett (1998)
3.3 Konzepte der Entstehung neuer Arten
Im biogeographischen Kontext bezieht sich Artentstehung auf die Prozesse, in denen eine
geographische Dimension zu finden ist, die neue Taxa bedingt. Umgekehrt gehört auch die
geographische Dimension des Aussterbens von Arten dazu.

40
Rosen (1988a) unterteilt die biogeographischen Konzepte der Artentstehung in zwei Formen.
Die erste umfaßt empirische Modelle (4.1.3.1), mit denen Biogeographen ihre Basisdaten
nach Korrelationen untersuchen, die zweite besteht aus Prozeßmodellen (4.1.3.2 - 4.1.3.7), die
sich überwiegend mit der allopatrischen Artentstehung beschäftigen, während parapatrische
und sympatrische Konzepte kaum berücksichtigt werden, obwohl auch sie vikarianten Cha-
rakter haben können (Barton, 1988).
Artentstehungsformen nach Mayr (1963)
Klassifizierung Beschreibung
allopatrisch Evolution von Fortpflanzungsbarrieren in Populationen,
die durch geographische Barrieren am Genaustausch behin-
dert werden.
parapatrisch (C) Evolution von Fortpflanzungsbarrieren zwischen benachbar-
ten Populationen, zwischen denen ein moderater
Genaustausch besteht.
sympatrisch (D) Evolution von Fortpflanzungsbarrieren zwischen zufälligen
Subgruppen einer einzigen Population.
Quelle: Futuyma, 1998
Die allopatrische Artentstehung läßt sich in die perapatrische (B) und die vikariante (A)
Artbildung teilen. In der erstgenannten findet die Evolution in einer isolierten Kolonie statt, in
der anderen werden zwei weitverbreitete Populationen getrennt (Futuyma,1998).
Diagramm von sukzessiven Stadien in vier Modellen von Artentstehung
Quelle: Futuyma, 1998

41
3.3.1 Empirisch phylogenetische (die Abstammung untersuchende) Biogeographie
Die empirische phylogenetische Biogeographie geht davon aus, daß phylogenetische Bezie-
hungen mit den heutigen oder vergangenen geographischen Gegebenheiten verbunden sein
könnten. In der reinsten empirischen Form basiert der Ansatz auf kladistischen Methoden,
ohne das Annahmen gemacht werden, warum und wie Verbindungen mit geographischen
Faktoren bestehen könnten. Die Betonung liegt hier also einfach auf der Suche nach überein-
stimmenden Mustern, jedoch sind auch in der empirischen Kladistik verdeckte Hinweise auf
Prozesse zu sehen (Hull, 1980).
3.3.2 Vikarianz als Resultat erdgeschichtlicher Veränderungen
Dieses Konzept nimmt angelehnt an die Hypothese der allopatrischen vikarianten Artentste-
hung an, daß die Entstehung neuer Arten ein Resultat der Teilung ehemals
zusammenhängender Verbreitungsgebiete einer Art durch TEKO-Ereignisse sein kann, die
neue Barrieren bzw. Divergenzen bewirken. Diese Gegebenheiten werden als zeitlich mitein-
ander verflochten angesehen. Wenn jedoch eine zeitliche Verzögerung festzustellen ist, wird
angenommen, das die durch TEKO-Ereignisse bedingte Divergenz vorausgehen muß, bevor
Artenentstehung daraus folgen kann (Rosen, 1988a).
Verbreitung der Familie der Iguanidae
Erstellt nach Brown und Gibson, 1983
Quelle: Futuyma, 1998

42
Ein Exempel hierfür ist die Verbreitung der Familie der Eidechsenfamilie der Iguanidae (siehe
S. 40). Die meisten Igannidae leben in Amerika, jedoch bewohnen einige Arten Madagaskar,
Fiji und Tonga. Die Entwicklung ihrer Verbreitung und Artentstehung läßt sich auf einen Ah-
nen zurückführen, der ehemals auf Gondwanaland gelebt hat und dessen Verbreitungsgebiet
durch die Divergenz des Großkontinents unterteilt wurde, wobei die Familie bis zum heutigen
Zeitpunkt in den anderen Gebieten ausgestorben ist.
3.3.3 Vikarianz als Resultat durch erdgeschichtliche Ereignisse bedingten ökologi-
schen Wandels oder ökologischen Wandel an sich
Diese unterscheidet sich von dem unter 4.1.3.2 vorgestellten Konzept dadurch, daß explizit
ökologische Faktoren eingebunden werden. Dadurch werden die Ideen der Verbreitungsspuren
mit jenen der Vikarianz verbunden.
Bei dem Divergenzereignis, das in zumindest einer der geteilte Teilpopulationen Artenbil-
dung bewirkt, kann es sich um ein TEKO-Ereignis handeln, es werden aber auch ökologische
wie Verlust des Habitats in der Mitte eines Verbreitungsgebietes als Möglichkeit in Betracht
gezogen, wodurch die Population in zwei Hälften geteilt wird. Ökologische Fälle lassen sich
aber auch oft auf TEKO-Ereignisse zurückführen (Rosen, 1988a).
Die westliche und östliche Diamantklapperschlange und ihre heutigen
Verbreitungsgebiete
Photographien: Klauber, 1972; Karte nach Conant (1958)
Quelle: Futuyma, (1998)
Als Beispiel ist etwa die Entwicklung der Diamantklapperschlange aufzuführen, welche die
letzte Eiszeit in Nordamerika in zwei Refugien im Südosten und Südwesten Nordamerikas
überleben konnte. In diesen hat sich die Art in die westliche (Crotalus atrox) und die östliche
Diamantklapperschlange (C. adamanteus) differenziert. Ob die Eiszeit nun als Konsequenz
eines TEKO-Ereignisses ist, sei dahingestellt, jedoch determinierte die latitudinale Lage

43
Nordamerikas verbunden mit ihrer ökologischen Toleranz die Rückzugsmöglichkeiten der
Schlange.
3.3.4 Randisolierung als Resultat durch erdgeschichtliche Ereignisse bedingten ökolo-
gischen Wandels
Diese peripatrische Hypothese basiert auf der Annahme, daß ein kleiner Teil der Population
einer Art durch TEKO-Ereignisse am Rand des Verbreitungsgebietes isoliert wird. Durch die
geringe Populationgröße ist nach Mayr (1942) hier eine Artenentstehung wahrscheinlicher.
Rosen (1988a) vermutet auch, daß die isolierte Teilpopulation in diesen Randgebieten unter
höherem ökologischen Streß steht, da einer oder mehrere limitierende Faktoren nahe an der
Toleranzgrenze liegen, wodurch sie angetrieben wird, sich den Bedingungen evolutionär an-
zupassen.
Mayrs These wird durch Untersuchungen von Selander (1970) hinsichtlich der Varianz
(Spannweite) der Allelenfrequenz (unterschiedliche Zustände einer Erbeinheit wie etwa eine
unterschiedliche Blütenfarben) unterstützt. So war die mittlere Allelenfrequenz bei beiden
Populationsgrößen sehr ähnlich, die Varianz (Streuweite) der Allelen bei den kleineren Popu-
lationen jedoch viel größer.
Frequenz der Allelen von Habitaten von Hausmäusen mit unterschiedlicher Po-
pulationsgröße
Mittlere
Allelenfrequenz
Varianz der Allelenfre-
quenz
Vermutete
Populations-
größe
Anzahl der
untersuchten
Populationen Es-3b Hbb Es-3b Hbb
ca. 10 29 0.418 0.849 0.0506 0.1883
ca. 200 13 0.372 0.843 0.0125 0.0083
Nach Selander (1970)
Quelle: Futuyma (1998)
3.3.5 Sprungverbreitung über Barrieren hinweg
Wenn eine Taxa eine Barriere per Sprungverbreitung überwunden hat und beidseits der Bar-
riere heimisch geworden ist, eine weitere Überwindung der Barriere aber sehr
unwahrscheinlich ist, so daß sie nur sehr selten stattfindet, so können sich beide Teilpopulati-
onen isoliert voneinander durch evolutionäre Prozesse auseinander entwickeln, was
schließlich zu verschiedenen Arten führen kann (Rosen, 1988a). Dieses ist etwa bei xenodon-
tiner Schlangen auf den Antillen der Fall. Dort sind 9 Genera nachgewiesen, die den

44
südamerikanischen sehr ähnlich sind. Daraus läßt sich ableiten, daß sie die Inseln erst vor kur-
zer Zeit schwimmend von Südamerika aus besiedelt haben (Williams, 1989; Futuyma, 1998)
und nicht zur Kreidezeit, als eine Landverbindung zwischen den Antillen und Zentralamerika
bestand.
3.3.6 Entstehungsgebiete neuer Arten (Centres of Origin)
Unter dem Titel „Centers of Origin“ findet sich nicht ein, sondern mehrere Konzepte der Bio-
geographie (Rosen,1988a). Einige lassen sich schon aus den bereits vorgestellten Konzepten
der Artenentstehung ableiten, ein interessanten Ansatz hinsichtlich der globalen Biodiversität
findet sich unter 3.1.3.7. Letztendlich sollten Centres of Origin nach Rosen (1988a) primär als
räumliches Muster verstehen, in dem mehrere auftretende Prozesse die Entstehung neuer Ar-
ten begünstigen.
3.3.7 Geoökologie über Zeit
Die Geoökologie über Zeit (auch Paläoökologie) geht davon aus, daß ökologische Faktoren
die Verbreitung bedingen. Es werden aber auch geologische Faktoren (TEKO-Events) berück-
sichtigt, die über die Zeit globale und regionale ökologische Bedingungen verändert haben.
Diese Kombination von Geologie und Ökologie wird als Stimulus für die Bildung und das
Aussterben von Arten, und dadurch als Artenvielfaltregulator betrachtet (Rosen, 1988a).
Die evolutionäre Komponente ist hierbei nicht auf einzelne Arten ausgelegt, wie etwa bei
der Vikarianz oder Randisolierung, sondern betrachtet das Aussterben, die Vielfalt und die
Entstehung für Hauptgruppen oder größere ökologische Wirkungsgefüge. In der Regel ge-
schieht dieses statistisch.
In erster Linie werden empirisch Korrelationen zwischen zeitlichen und räumlichen taxo-
nomischen Daten mit geologischen Bedingungen und Ereignissen untersucht. In der Praxis
findet die gesamte Bandbreite der Prozeßannahmen aus der Geoökologie Verwendung (etwa
Gleichgewicht, Wettbewerb, Nischentheorie, Ressourcenteilung). TEKO-Ereignisse werden
als über die Ökologie wirkender Antrieb der evolutionärer Entwicklung, weniger im vikari-
anten Sinne sondern als Ursache für Anpassungen von Organismen oder Gruppen von
Organismen (Rosen, 1988a), angesehen.
Zeitlich betrachtet beeinflußt so etwa die Drift der Platten die Anzahl der Bioreiche. Theo-
retisch wäre die Anzahl und Unterschiedlichkeit von Bioreichen am höchsten bei einer hohen

45
Fragmentation in schmale Kontinente und tiefe Ozeanbecken, die latitudinal voneinander ge-
trennt liegen (Valentine u.a., 1978).
Die niedrigste Anzahl von Bioreichen ist zu erwarten, wenn alle Kontinente zusammen
einen Superkontinent bilden (Brenchley und Harper, 1998). Diese Theorie wird durch Unter-
suchungen von Smith hinsichtlich der sich ändernden Vielfalt der skeletal shelf benthos
(Smith, 1990) unterstützt.
Changing family diversity of skeletal benthos through time in relation to
plate configurations
Schwankung der Familiendiversität der skeletal benthos über die Zeit in Be-
ziehung zu Lage der Lithosphärenplatten: hohe Diversität tritt kohärent zu
hoher kontinentaler Fragmentation auf, so etwa zur Zeit des Ordoviziums,
des Devons und zu Kreidezeit/ Tertiär. Abkürzungen: I - Iapetus Ozean; A -
Prä-appalachisch-variskischer Ozean; U - Präuralischer Ozean; H - Spani-
scher Korridor (Grafik erstellt nach Smith (1990)).
Quelle: Brenchley und Harper (1998)

46
Das Auseinanderbrechen Rhondinia schuf zum Paläozoikum eine größere Anzahl von Konti-
nenten: Avalonia, Baltica, Laurentia, Siberia und die große Landmasse von Gondwana. Diese
verteilten sich im frühen Ordovizium über das gesamte latitudinale Spektrum von polaren bis
äquatorialen Breiten. Genau zu dieser Zeit läßt sich ein Ansteigen der Artenvielfalt ausma-
chen. Dieses verstärkte sich zur Mitte des Ordoviziums. Zu dieser Zeit ließen intensive
magmatische und tektonische Aktivität Gondwana weiter auseinanderbrechen, Inselbögen und
Archipels entstanden (Harper u.a., 1996), welche Brenchley und Harper (1998) für Zentren
der Artenbildung halten. Sie vermuten, daß dort möglicherweise auch die makroevolutionären
Prozesse abliefen, welche die Entwicklungsgeschichte der Meeresorganismen im Palaeozoi-
kum maßgeblich prägten (Brenchley und Harper, 1998).
4 SchlußbemerkungHinsichtlich dem Stand der biogeographischen Forschung stimme ich Myers und Giller
(1988) zu, daß eine verstärkte interdisziplinäre Zusammenarbeit wünschenswert wäre. Jedoch
macht auch gerade die Vielfalt der Forschungsansätze die Biogeographie zu einer lebendigen
Wissenschaft.
Letztendlich steckt die historische Biogeographie, wie allgemein die Paläowissenschaften,
immer noch in den Kinderschuhen. Viele Makrohypothesen stützen sich lediglich auf wenige
Forschungsergebnisse, es wird also auch in Zukunft noch viel zu entdecken geben.
Ein Ausbau der Arbeit um die Konsequenzen der Plattentektonik auf die Methodik der Bi-
ogeographie wäre möglich, wurde von mir aber unterlassen, da er den Rahmen der Arbeit
gesprengt hätte.

47
LiteraturverzeichnisAchibold, O.W. (1995): Ecology of World Vegetation. London.
Bacon, F. (1620): im Novum Organum
Barron, E.J. / Washington, W.M. (1985): Warm Cretaceous climates : High atmospheric CO2-
Levels as a pausible mechanism. In: Sundquist, E.T. und Broecker, W.S. (Hg.) (1985): The
Carbon Cycle and Atmospheric CO2 : Natural Variations Archean to Present. Geophysical
Monographies 32. American Geophysical Union. Washington DC. S. 546 -553.
Barton, N.H. (1988). Spciation. In: Myers, A.A. und Giller, P.S. (Hrsg.) (1988): Analytical
Biogeography : An integrated approach to the study of animal and plant distributions. Lon-
don. S. 185 - 218.
Beaty, C. (1978): Ice ages and continental drift. New Science 80. S. 776-777.
Becquerel, H. (1896): Sur les radiations émises par phosphorescence. In: Comptus Rendus
122. 420.
Bjärvall, A. / Ullström, S. (1986): The Mammals of Britain and Europe. London.
Brenchley, P.J. / Harper, D.A.T. (1998): Palaeoecology : Ecosystems, environments and evo-
lution. London.
Briggs, D. / Smithson, P. / Addison, K. / Atkinson, K. (1997): Fundamentals of the Physical
Environment. 2. Aufl. London.
Briggs, J.C. (1995): Global Biogeography. Amsterdam. Developments in Palaeontology and
Stratigraphy 14.
Broecker, W.S. / Peteet, D.M. und Rind, D. (1985): Does the ocean-atmosphere system have
more than one stable mode of operation? In: Natue 315. S. 21 - 26.
Brown, J.H. / Gibson, A.C. (1983): Biogeography. St. Louis MO.
Brundin, L. (1965): On the real nature of transantarctic relationships. In: Evolution 19, S. 496
- 505.
Burchfield, B.C. (1983): The continental crust. In. Scientific American (September 1983). S.
130
Burton, R. (1628): The Anatomy of Melancholy. Neuaufgelegt durch: Shilleto (Hg.) (1896):
The Anatomy of Melancholy. London.
Candolle, A.-P. (1820): Geographie botanique. In: Dictionnaire des Sciences Naturelles 18, S.
359 - 422.

48
Closs, H. / Giese, P. / Jacobshagen, V. (1985): Alfred Wegeners Kontinentalverschiebung aus
heutiger Sicht. In: Spektrum der Wissenschaft: Ozeane und Kontinente. Heidelberg. S. 40 -
53.
Conant, R. (1958): A Field Guide to Reptiles and Amphibians. Boston MA.
Cox, W.T. (1936): Snowshoe rabbit migration, tick infestation and weather cycles. Journal of
Mammalogy 17. S. 216 - 221.
Cox, C.B. / Healey, I.N. / Moore, P.D. (1976): Biogeography : An ecological and evolutionary
approach. 2. Aufl. Oxford.
Croizat, L. (1958): Pangeography. 2 Bände. Caracas.
Croizat, L. (1964): Space, Time, Form : The Biological Synthesis. Caracas.
Crowley, T.J. und North, G.R. (1991): Paleoclimatology. Oxford Monographs on Geology
and Geophysics 16.
Darwin, C.R. (1872): The Origin of Species. 6. Aufl. London.
Du Toit, A. (1937): Our Wandering Continents. Edinburgh.
Einstein, A. (1922): Grundzüge der Relativtätstheorie. Braunschweig.
Fairbridge, R.W. (1972): Glaciation and plate migration. In: Tarling, D.H. / Runcorn, S.K.
(Hg.) (1973): Implication of continental drift to the earth sciences. I. London.
Fischer, A.G. (1984): The two Phanerozoic supercycles. In Berggren W.A. / Van Couvering,
J.A. (Hg.): Catastrophes and Earth history : The new uniformitarianism. Princeton. S. 129-
150.
Flux, J.E.C. / Fullagar, P.J. (1992): World distribution of the rabbit Oryctolagus cuniculus on
islands. In: Mammal Review 22. S. 151 -205.
Futuyma, D.G. (1990): Evolutionsbiologie. Basel.
Futuyma, D.G. (1998): Evolutionary Biology. 3. Aufl. Sunderland.
Grenn, R. (1961): Palaeoclimatic significance of evaporites. In: Nairn, A.E.M. (Hg.) (1961):
Descriptive Paleoclimatology. New York.
Harper, D.A.T. / Mac Niocaill, C. / Williams, S.H. (1996): The palaeogeography of early Or-
dovician Iapetus terranes : An integration of faunal and palaeomagnetic constraints. In:
Palaeogeography, Palaeoclimatology and Palaeoecology 121, S. 297 - 312.
Hartmann, D.L. (1994): Global Physical Climatology. International Geophysics Series 56. San
Diego.
Hennessey, H. (1860): Change of Climate. The Atheanaeum 1717. S. 384 - 386.

49
Hess, H.H. (1962): History of ocean bassins. In: Engel, A.E.J. / James, H.L & Leonard, B.F.
(Hg.) (1962): Petrological Studies. Denver. S. 599 - 620.
Holmes, A. (1928): Rede vor der Geological Society of Glasgow. Publiziert in: Transactions
of the Geological Society of Glasgow 18. S. 559.
Huggett, R.J. (1991): Climate, Earth Processes and Earth History. Berlin.
Huggett, R.J. (1998): Fundamentals of Biogeography. London.
Hull, D.L. (1980): Cladism gets sorted out. (Review of: Cracraft, J. / Eldredge, N. (1979):
Phylogenetic analysis and paleontology. New York.) In: Paleobiology 6. S. 131 - 136.
Irving, E. (1964): Paleomagnetism. New York
Jablonski, D. / Flessa, K.W. / Valentine, J.W. (1985): Biogeography and paleobiology. In:
Paleobiology 11. S. 75 - 90.
Jablonski, D. / Erwin, D.H. / Lipps, J.H. (Hg.) (1996): Evolutionary Paleobiology : Essays in
Honor of James W. Valentine. Chicago.
Jeffreys, H. (1924): The Earth : Ist Origins, History and Physical Constitution. Cambridge.
Kearey, P. / Vine, F. (1996): Global Tectonics. 2. Aufl. Oxford.
Klauber, L.M. (1972): Rattlesnakes : Their Habits, Life Histories, and Influence on Mankind.
berkeley.
Kuhn, T. (1967): Die Struktur wissenschaftlicher Revolutionen. Frankfurt a.M.
Kutzbach, J.E. / Guetter, P.J. / Ruddiman, W.F. / Prell, W.L. (1989): The sensitivityof climate
to late Cenozoic uplift in southeast Asia and the American southwest : Numerical experi-
ments.
Lack, D. (1947): Darwin’s finches. Cambridge.
Liebig, J. (1840): Organic Chemistry and ist Application to Agriculture and Physiology. Lon-
don.
MacArthur, R.H. / Wilson, E.O. (1967): The Theory of Island Biogeography. Princeton.
Maier-Reimer, E. / Mikolajewicz, U. und Crowley, T.J. (1990): Ocean GCM sensitivity expe-
riment with an open central American isthmus. Paleoceanography 5. S. 349 - 366.
Manabe, S. / Broccoli, A.J. (1990): Mountains and arid climates of middle latitudes. In:
Science 247. S. 192 - 195
Manabe, S. / Terpstra, T.B. (1974): The effects of mountains on the general circulation of the
atmosphere as identified by numerical experiments. In: Journal of Atmospheric Scienes 31.
S. 3 - 42.

50
Marshall, L.G. (1981): The Great American Interchange - an invasion induced crisis for South
American mammals. S. 133 - 229. In: Nitecki, M.H. (Hrsg.) (1981): Biotic Crisis in Ecolo-
gical and Evolutionary Time. New York.
Matthews, R.K. / Poore, R.Z. (1980): Tertiary 18O record and glacioeustatic sea-level fluctua-
tion. In: Geology 8. 501 - 504.
Mayr, E. (1942): Systematics and the Origin of Species. New York.
Mayr, E. (1963): Animal Species and Evolution. Cambridge MA.
McKenna, M.C. (1973): Sweepstakes, filters, corridors, Noah´s Arks and beached Viking fu-
neral ships in palaeogeography. In: Tarling, D.H. / Runcorn, S.K. (Hg.) (1973): Implication
of continental drift to the earth sciences. I. London.
Myers, A.A. und Giller, P.S. (1988): Process, pattern and scale in biogeography. In: Myers,
A.A. und Giller, P.S. (Hrsg.) (1988): Analytical Biogeography : An integrated approach to
the study of animal and plant distributions. London. S. 3 - 12.
Nelson, G. / Platnick, N. (1981): Systematics and Biogeography : Cladistics and vicariance.
New York.
Oglesby, R.J. (1989): A GCM study of Antarctic glaciation. In: Climate Dynamics 3. S. 135 -
156.
Pielou, E.C. (1979): Biogeography. New York.
Press F. & Siever, R. (1995): Geologie : Eine Einführung. Heidelberg.
Reinke-Kunze, C. (1994): Alfred Wegener : Polarforscher und Entdecker der Kontinentaldrift.
Basel.
Rosen, B.R. (1984): Reef coral biogeography and climate through the late Cainozoic : Just
islands in the sun or a critical pattern of islands. In: Brenchley, P.J. (1984): Fossils and
Cllmate. Geological Journal Special Issue 11. S. 201 - 262.
Rosen, B.R. (1988a): Biogeographic patterns : a perceptual overview. In: Myers, A.A. und
Giller, P.S. (Hrsg.) (1988): Analytical Biogeography : An integrated approach to the study
of animal and plant distributions. London. S. 23 - 55.
Rosen, R.B. (1988b): From fossils to earth history : Applied historical biogeography. In: My-
ers, A.A. und Giller, P.S. (Hrsg.) (1988): Analytical Biogeography : An integrated
approach to the study of animal and plant distributions. London. S. 437 - 481.
Rosen, B.R. / Smith, A.B. (1988): Tectonics from fossils? Analysis of reef coral and sea ur-
chin distributions from late Cretaceous to Recent, using a new method. In: Audley-Charles,

51
M.G. / Hallam, A. (1988): Gondwana and Tethys. Geological Society of Lonson Special
Publication.
Ruddiman, W.F: / Kutzbach, J.E. (1989): Forcing of late Cenozoic northern hemisphere cli-
mate by plateau uplift in southeast Asia and the American southwest. In: Journal of
Geophysical Research 94. S. 18409 - 18427.
Ruddiman, W.F. / Raymo, M.E. (1988): Northern hemisphere climate regimes during the past
3 MA : Possible tectonic connections. In : Philosophical Transactions of the Royal Society
of London B 318. S. 411 - 430.
Ruddiman, W.F. / Raymo, M.E. und McIntyre, A. (1986): Matuyama 41.000-year cycles :
North Atlantic Ocean and northern hemisphere ice sheets. In: Earth and Planetary Science
Letters 80. S. 117 - 129.
Schönwiese, C.D. (1994): Klimatologie. Stuttgart.
Schwarzbach, M. (1968): Neuere Eiszeithypothesen. In: Jahrbuch der Universität Köln
(1968). Köln.
Schwarzbach, M. (1993): Das Klima der Vorzeit : Eine Einführung in die Paläoklimatologie.
5. Aufl. Stuttgart.
Scotese, C.R. (1992): Paleomap project. Progress Report Nomber 36. Arlington.
Seidov, D.G. (1986): Numerical modelling of the ocean circulation and paleocirculation. In:
Hsü, K. (Hg.) (1986): Mesozoic and Cenozoic Oceans. Geodynamic Series15. American
Geophysical Union. Washington DC. S. 11 - 26.
Selander, R.K. (1970): Behavior and genetic variation in natural populations. In: American
Zoologist 10. S. 53 -66.
Shelford, V.E. (1911): Physiological animal geography. In: Journal of Morphology 22. S. 551
- 618.
Simpson, G.G. (1940): Mammals and land bridges. In: Journal of the Washington Academy of
Science 30. S. 137 - 163.
Smith, P.L. (1990): Paleobiogeography and plate tectonics. Geoscience Canada 15, S. 261 -
279.
Snider-Pellegrini, A. (1858): La Création et ses Mystères Dévoilees. Paris.
Valentine, J.W.; Foin, T.C. und Peart, D. (1978): A provincial model of Phanerozoic diversi-
ty. In: Paleobiology 4, S. 55 - 66.

52
Wallace, A.R. (1876): The Geographical Distribution of Animals; with a Study of the Relati-
ons of Living and Extinct Faunas as Elucidating the Past Changes of the Earth's Surface. 2
Bände. London.
Wegener, A. (1921): Die Theorie der Kontinentalverschiebung. In: Zeitschrift der Gesellschaft
für Erdkunde zu Berlin. S. 95.
Whitfield, P. (1993): From so Simple a Beginning : The Book of Evolution. New York.
Whitmore, T.C. (1981) (Hg.): Wallace’s Line and Plate Tectonics. Oxford.
Williams, G.E. (Hg.) (1981): Megacycles : long-term eepisodicity in Earth and planetary
history. Benchmark Papers in Geography 57. Stroudsberg.
Williams, E.E. (1989): Old problems and new opportunities in West Indian Biogeography. In:
Woods, C.A. (Hg.) (1989): Biogeography of the West Indies : Past Present and Future.
Gainesville. S. 1 - 46.
Wilson, J.T. (1963): Continental Drift. Science America. S. 1 - 16.
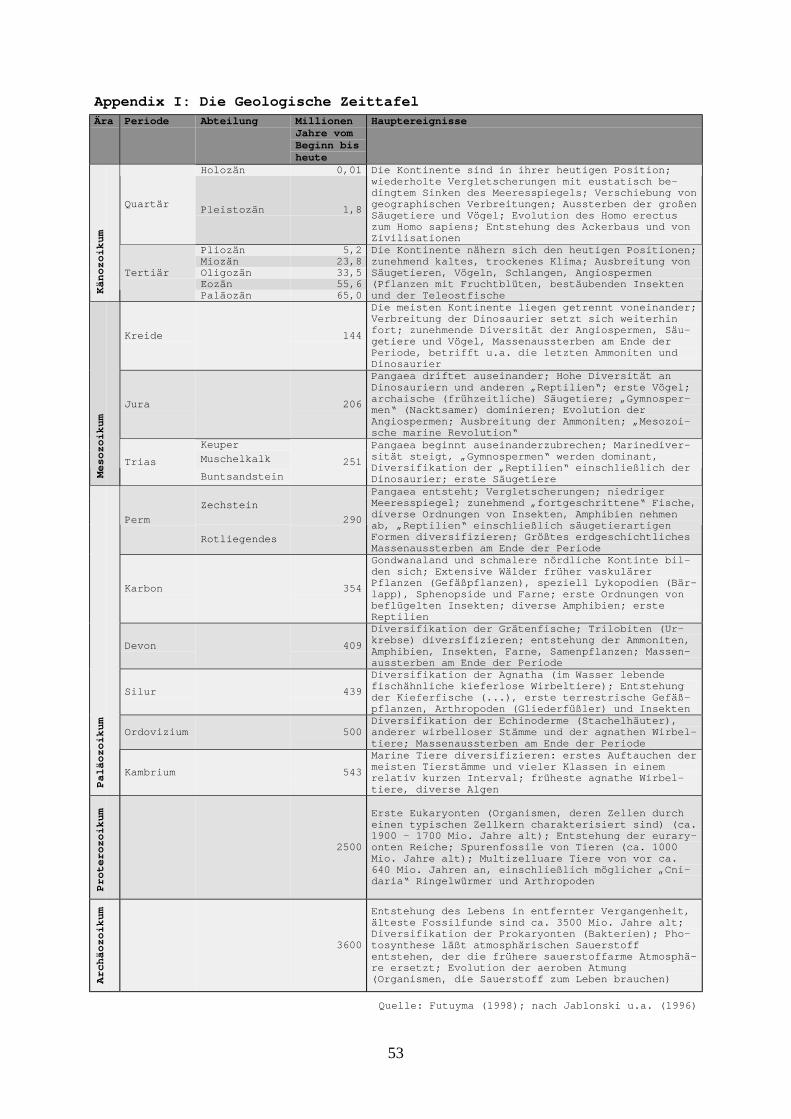
53
Appendix I: Die Geologische ZeittafelÄra Periode Abteilung Millionen
Jahre vomBeginn bisheute
Hauptereignisse
Holozän 0,01
Quartär Pleistozän 1,8
Die Kontinente sind in ihrer heutigen Position;wiederholte Vergletscherungen mit eustatisch be-dingtem Sinken des Meeresspiegels; Verschiebung vongeographischen Verbreitungen; Aussterben der großenSäugetiere und Vögel; Evolution des Homo erectuszum Homo sapiens; Entstehung des Ackerbaus und vonZivilisationen
Pliozän 5,2Miozän 23,8Oligozän 33,5Eozän 55,6
Känozoikum
Tertiär
Paläozän 65,0
Die Kontinente nähern sich den heutigen Positionen;zunehmend kaltes, trockenes Klima; Ausbreitung vonSäugetieren, Vögeln, Schlangen, Angiospermen(Pflanzen mit Fruchtblüten, bestäubenden Insektenund der Teleostfische
Kreide 144
Die meisten Kontinente liegen getrennt voneinander;Verbreitung der Dinosaurier setzt sich weiterhinfort; zunehmende Diversität der Angiospermen, Säu-getiere und Vögel, Massenaussterben am Ende derPeriode, betrifft u.a. die letzten Ammoniten undDinosaurier
Jura 206
Pangaea driftet auseinander; Hohe Diversität anDinosauriern und anderen „Reptilien“; erste Vögel;archaische (frühzeitliche) Säugetiere; „Gymnosper-men“ (Nacktsamer) dominieren; Evolution derAngiospermen; Ausbreitung der Ammoniten; „Mesozoi-sche marine Revolution“
Keuper
Muschelkalk
Mesozoikum
Trias
Buntsandstein
251
Pangaea beginnt auseinanderzubrechen; Marinediver-sität steigt, „Gymnospermen“ werden dominant,Diversifikation der „Reptilien“ einschließlich derDinosaurier; erste Säugetiere
Zechstein
Perm
Rotliegendes
290
Pangaea entsteht; Vergletscherungen; niedrigerMeeresspiegel; zunehmend „fortgeschrittene“ Fische,diverse Ordnungen von Insekten, Amphibien nehmenab, „Reptilien“ einschließlich säugetierartigenFormen diversifizieren; Größtes erdgeschichtlichesMassenaussterben am Ende der Periode
Karbon 354
Gondwanaland und schmalere nördliche Kontinte bil-den sich; Extensive Wälder früher vaskulärerPflanzen (Gefäßpflanzen), speziell Lykopodien (Bär-lapp), Sphenopside und Farne; erste Ordnungen vonbeflügelten Insekten; diverse Amphibien; ersteReptilien
Devon 409
Diversifikation der Grätenfische; Trilobiten (Ur-krebse) diversifizieren; entstehung der Ammoniten,Amphibien, Insekten, Farne, Samenpflanzen; Massen-aussterben am Ende der Periode
Silur 439
Diversifikation der Agnatha (im Wasser lebendefischähnliche kieferlose Wirbeltiere); Entstehungder Kieferfische (...), erste terrestrische Gefäß-pflanzen, Arthropoden (Gliederfüßler) und Insekten
Ordovizium 500Diversifikation der Echinoderme (Stachelhäuter),anderer wirbelloser Stämme und der agnathen Wirbel-tiere; Massenaussterben am Ende der Periode
Paläozoikum
Kambrium 543
Marine Tiere diversifizieren: erstes Auftauchen dermeisten Tierstämme und vieler Klassen in einemrelativ kurzen Interval; früheste agnathe Wirbel-tiere, diverse Algen
Proterozoikum
2500
Erste Eukaryonten (Organismen, deren Zellen durcheinen typischen Zellkern charakterisiert sind) (ca.1900 - 1700 Mio. Jahre alt); Entstehung der eurary-onten Reiche; Spurenfossile von Tieren (ca. 1000Mio. Jahre alt); Multizelluare Tiere von vor ca.640 Mio. Jahren an, einschließlich möglicher „Cni-daria“ Ringelwürmer und Arthropoden
Archäozoikum
3600
Entstehung des Lebens in entfernter Vergangenheit,älteste Fossilfunde sind ca. 3500 Mio. Jahre alt;Diversifikation der Prokaryonten (Bakterien); Pho-tosynthese läßt atmosphärischen Sauerstoffentstehen, der die frühere sauerstoffarme Atmosphä-re ersetzt; Evolution der aeroben Atmung(Organismen, die Sauerstoff zum Leben brauchen)
Quelle: Futuyma (1998); nach Jablonski u.a. (1996)

54
Appendix II: Die Biosphäre und darauf wirkende Prozesse
Quelle: Huggett (1991)

55
Appendix III: Biogeographische Karten, modifiziert von Briggs (1995)nach Ergebnissen des Paleomap Projects des Internationalen Li-thosphärenprogramms (Karten 9 - 16) (Scotese, 1992).
Karte 9: Mittleres Trias (Muschelkalk): Pangaea ist komplett zusammen. fastalle Tetrapodenfamilien (Vierfüßler) waren cosmo-politisch, ohne deutlichsichtbare latitudinale Variationen.

56
Karte 10: Spätes Jura. Pangaea beginnt, auseinanderzubrechen. Die Turgaiseehat Asien von Euramerika, Südamerika und Afrika gehen auseinander. Madagas-kar hat sich von Afrika getrennt, Euramerika und Afrika sind nur noch übereine kliene Landbrücke verbunden.
Karte 11: Frühe Kreide: Die nördlichen Laurasischen Kontinente haben sichkomplett von Gondwana getrennt, wodurch das zirkum-tropische Thetysmeerentstanden ist. Afrika und Südamerika könnten schon auseinandergedriftetsein, liegen aber noch nah beieinander.

57
Karte 12: Späte Kreide: Ein hoher Meeresspiegelstand hat zu extensiven epi-kontinentalen Schelfmeeren geführt. Asien und Westamerika formen eineneigenen Kontinent, getrennt von Euramerika. Indien ist eventuell schon mitAsien verbunden.
Karte 13: Paläozän: Die epikontinentalen Meere sind zurückgegangen. Es gibtnur einen zusammenhängenden Nordkontinent. Der zentralamerikanische Isthmuskann schon existiert haben, Indien ist sehr wahrscheinlich mit Asien ver-bunden. Die nahe Lage von Südamerika, der Abtarktis und Australien stelltewohl eine Filterroute dar.

58
Karte 14: Spätes Eozän: Nordeuropa hat sich von Grönland getrennt, Zentral-amerika existiert als ein Inselarchipel. Die Antillen haben sich gebildet.Der atlantische Ozean geht weiter auseinander.
Karte 15: Frühes Miozän: Durch den Zusammenschluß Afrikas mit Asien überdie Arabische Halbinsel wurde die Thetyssee überwiegend eliminiert, ledig-lich das Mittelmeer und das Meer in der zentralasiatischen Senkeverbleiben. Zentralamerika ist wiederhergestllt mit Ausnahme einer kleinenLücke zwischen Panama und Kolumbien. Australien-Neuguinea sind nordwärtsgewandert und haben fast ihre heutige Position erreicht.

59
Karte 16: Heute: Die Beringlandbrücke ist unterbrochen, der Zentralamerika-nische Isthmus geschloßen. Das Schwarze und das Kaspische Meer haben sichgeformt.





























