Embed Size (px)
Citation preview

Die Meiose ist der Prozeß, in dem aus diploiden haploideZellen entstehen. Dies geschieht durch zwei Zellteilungs-runden ohne dazwischenliegende Replikation. Während diezweite Teilungsrunde weitgehend einer normalen Mitosegleicht, ist die erste deutlich anders, besonders die Prophaseist deutlich länger und wird in Unterphasen unterteilt.
Meiose Ablauf der Meiose
Stufen der erstenProphase der
MeioseLeptotän: Chromatidenwerden sichtbarZygotän:Homologen-paarung, ChiasmataPachytän: Paarung aufganzer LängeDiplotän: Chromatidenmaximal kondensiertDiakinese: Auflösungder Kernmembran,Übergang zurMetaphase
MeioseDabei paaren sich die Homologen jeden Chromosoms, alsodie einander entsprechenden Chromosomen, die ursprünglichvon Vater bzw. Mutter stammen. Die Struktur aus starkkondensierten‚ eng verbundenen Chromosomen wird„synaptonemalerKomplex“ genannt.
Meiose
Die Homologen werden auf die beiden entstehenden Zellenverteilt. Die Verteilung (Vater- oder Mutter-Kopie) ist fürjedes Chromosom unabhängig. Allein dadurch ergeben sichbeim Menschen 223, also ca. 8 Millionen unterschiedlicherVerteilungsmuster.
Meiose
Bei nur zweiChromosomen ergebensich vier haploideGenotypen. Wird dasGenom nur durch dieChromosomenverteilungneu bestimmt, sind diebeiden Zellen jederzweiten meiotischenTeilung genetischidentisch.

Non-DisjunctionBeim Verteilungs-
prozeß kann es zuFehlern kommen,beide Homologebleiben verbundenund wandern indieselbe Zelle:„Non-Disjunction“(Beispiel:Drosophila, dageschieht das ineiner von 2000Meiosen)*
Drosophila und Mensch
*Das Beispiel zeigt auch einen wichtigen Unterschied bei derGeschlechtsbestimmung Mensch und Drosophila. Beidehaben X und Y Chromosomen, wobei XX zu weiblichen, XYzu männlichen Individuen führt. Beim Menschen geschiehtdas über eine dominante Rolle des Y-Chromosoms, XXY-Menschen sind daher männlich, wenn auch mit verzögerterEntwicklung (Klinefelter-Syndrom), X allein weiblich(Ullrich-Turner-Syndrom), XXX praktisch normale Frauen.Bei Drosophila bestimmt dagegen die Gendosis von X dasGeschlecht, Y spielt keine Rolle. XXY ist daher weiblich, Xallein männlich, XXX Weibchen mit stark verminderterLebensfähigkeit.
Meiose
Zusätzlich zu dieser Methode, das Erbmaterial neu zukombinieren, kommt es während der ersten meiotischenTeilung meist zu DNA-Austausch zwischen den gepaartenHomologen durch „crossing-over“ (homologeRekombination). Dadurch entsteht eine noch größere Vielfaltbei der Neukombination der Elterngene. Auch die beidenZellen, die bei der zweiten meiotischen Teilung entstehen,sind durch diese Rekombination unterschiedlich.Ausgangsereignis ist ein Strangbruch in einem Chromatid,der mit DNA eines homologen Stranges verbunden wird(„breakage and reunion“).
MeioseDas Ereignis ist sogar direkt als„Chiasma“ (griechisches Chi: Χ),eine „X-förmige“ Verbindungzwischen den Homologen,sichtbar.
Meiose
Dadurch enthält ein DNA-Bereicheinen Strang vom „Vater-“ und einenvom „Mutter-“Homologen gepaartals Doppelstrang. Man nennt dieseine Heteroduplex oder Hybrid-DNA. Dieser Bereich kann durch„Branch migration“, ein Weiter-wandern des Kreuzungspunktes,vergrößert werden.
MeioseDie crossover-Struktur wird auch„Holliday junction“ genannt. Hollidayprojizierte die Struktur als flaches Kreuz.An diesem Schema wurde deutlich, daßein zweiter Strangbruch, der nötig ist,damit sich die Stränge wieder trennenkönnen, entscheidet, ob durch dascrossing-over nur ein Stück des anderenHomologen übernommen wurde(zweiter Bruch im gleichen Strang), oderob die Teile der Homologen zu einemrekombinanten Chromosom verknüpftwerden (zweiter Bruch im Gegenstrang).

Meiose
Die Auflösung derHolliday-Strukturführt zum Transfereines Stücks („patchrecombinant“) oderzur Bildung vonrekombinantenChromosomen
Meiose
Nach einem neueren Modell startet einDoppelstrangbruch, der zu einer Lückevergrößert wird, die Rekombination. Ansich ist ein Doppelstrangbruch nicht zureparieren, da die beiden Fragmenteauseinander driften. Hier aber werden dieentstehenden Stücke durch densynaptonemalen Komplex nahe demhomologen Partner festgehalten.
Meiose
Bei dieser Rekombination durch Doppelstrangbruch wird dasErbmaterial der beiden Homologen nicht einfach neu verteilt.Die Lücke im gebrochenen Strang (dem „Rezipienten-Strang“) wird durch Replikation des anderen Homologen(dem „Donor“) aufgefüllt, dieser Bereich ist anschließendauf beiden Chromosomen identisch (entspricht nämlich demursprünglichen Donor).
MeioseAnfangs war es unklar, ob das Crossing-over im „Zweistrang-stadium“, also vor der Replikation, oder im Vierstrangstadiumstattfindet.Der Brotschimmel Neurospora crassa produziert 8 Sporen ineinem linearen Ascus, ihre Reihenfolge ergibt sich aus derMeiose (und einer nachfolgenden „postmeiotischen Teilung“).Crossing-over (zwischen dem entsprechenden Gen und demCentromer) führt zu einer 2:2:2:2-alternierenden Verteilungder Allele, muß also nach der Replikation geschehen sein.Diese Verteilung wird „Postreduktion“ genannt, wohl, weildie Gentrennung erst nach der „Reduktionsteilung“, der erstenmeiotischen Teilung, abgeschlossen wird. Eine 4:4-Verteilungist eine „Präreduktion“, da die Alleltrennung in der erstenmeiotischen Teilung geschieht.
Neurosporacrassa
Neurospora crassa
So wäre die Allelverteilung bei Crossing-over im Zweistrangstadium: Das kommtaber nicht vor.

Neurospora crassaAus der Sporenreihen-folge bei Dihybriden läßtsich auch ableiten, welcheder vier Chromatiden(Schwesterchromatiden xHomologes) an denCrossing overs beteiligtwaren.Das Beispiel zeigt auch,daß die Rekombinationnicht auf zwei („innenliegende“) Strängebeschränkt ist!!
Genkonversion
Normalerweise treten die in die Meiose eintretenden Alleleeines Gens in den haploiden Produkten in gleicher Zahl auf(2:2, bei Neurospora wegen postmeiotischer Teilung 4:4).Selten kommt es zu Abweichungen (3:5, 2:6-Verteilungen).Hier ist eines der beiden Allele mit Hilfe der anderen Kopie„repariert“ worden: Genkonversion. Auslöser ist oft einCrossing-over (das Auffüllen der Lücke durch den Donor nachDoppelstrangbruch ist so eine Genkonversion).
Genkonversion beiNeurospora crassa
Genkonversionbei S. cerevisiae
Durch nicht-reparierteHeterodupliceswachsen auseinzelnen SporenMischkolonien, dadie haploide Zellezwei Allele in einemDoppelstrang trägt.
MeioseOb auf einem Chromosom liegende Genezusammen vererbt werden oder getrenntwerden, hängt davon ab, ob im Abschnittzwischen ihnen ein crossing-overstattfindet. Das wirddesto unwahrschein-licher, je kleiner derAbstand ist. WerdenGene meist gemein-sam vererbt (liegenalso auf demChromosom nahebeeinander),bezeichnet man sieals „gekoppelt“.
Meiose
Bevor in den letzten Jahren Genomsequenzierungen möglichwurden, hat man die Reihenfolge der Gene (Genkarte) auf denChromosomen anhand von Genkopplungen erstellt. Jede„Kopplungsgruppe“ (alle Gene, die direkt oder indirektgekoppelt sind) entspricht einem Chromosom im haploidenGenom.Der nach dieser Methode ermittelte Abstand von Genen wirdin „centi-Morgan“ (cM) angegeben. 1 cM entspricht einemAbstand, der zu einer Rekombinationswahrscheinlichkeitvon 1% führt. Die Rekombinationswahrscheinlichkeit istkonstant und reproduzierbar - aber sie ist nicht additiv.
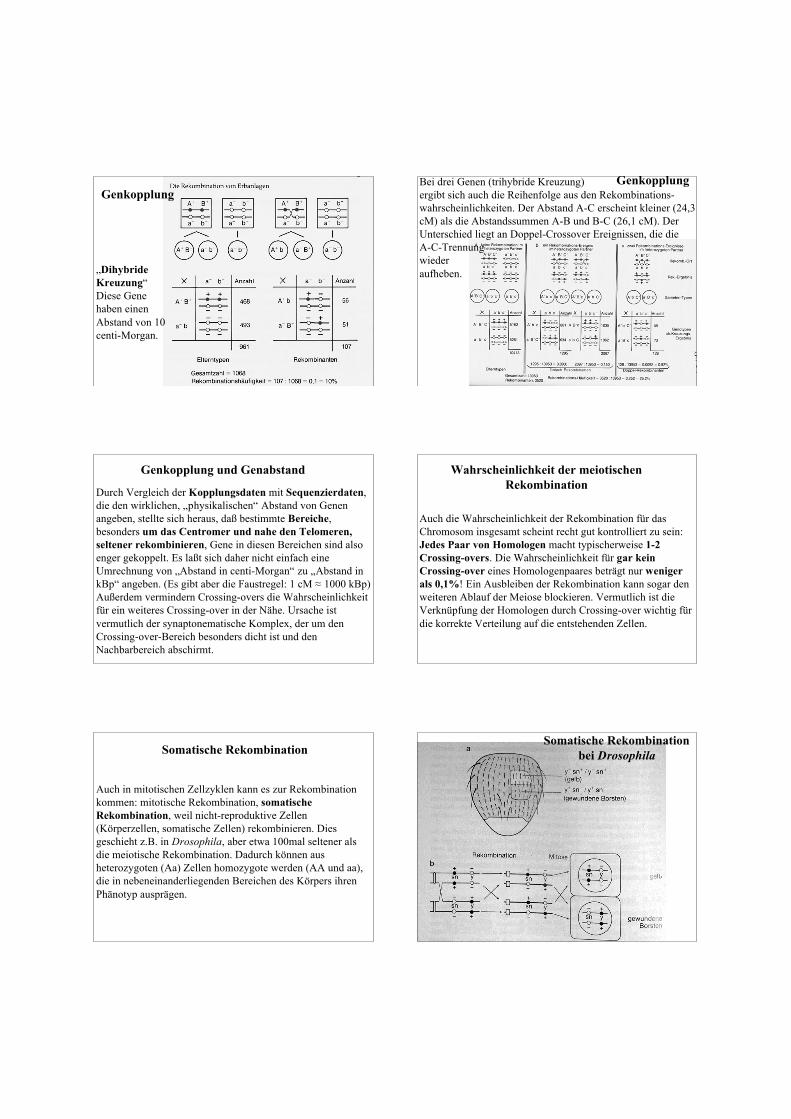
Genkopplung
„DihybrideKreuzung“Diese Genehaben einenAbstand von 10centi-Morgan.
GenkopplungBei drei Genen (trihybride Kreuzung)ergibt sich auch die Reihenfolge aus den Rekombinations-wahrscheinlichkeiten. Der Abstand A-C erscheint kleiner (24,3cM) als die Abstandssummen A-B und B-C (26,1 cM). DerUnterschied liegt an Doppel-Crossover Ereignissen, die dieA-C-Trennungwiederaufheben.
Genkopplung und Genabstand
Durch Vergleich der Kopplungsdaten mit Sequenzierdaten,die den wirklichen, „physikalischen“ Abstand von Genenangeben, stellte sich heraus, daß bestimmte Bereiche,besonders um das Centromer und nahe den Telomeren,seltener rekombinieren, Gene in diesen Bereichen sind alsoenger gekoppelt. Es laßt sich daher nicht einfach eineUmrechnung von „Abstand in centi-Morgan“ zu „Abstand inkBp“ angeben. (Es gibt aber die Faustregel: 1 cM ≈ 1000 kBp)Außerdem vermindern Crossing-overs die Wahrscheinlichkeitfür ein weiteres Crossing-over in der Nähe. Ursache istvermutlich der synaptonematische Komplex, der um denCrossing-over-Bereich besonders dicht ist und denNachbarbereich abschirmt.
Wahrscheinlichkeit der meiotischenRekombination
Auch die Wahrscheinlichkeit der Rekombination für dasChromosom insgesamt scheint recht gut kontrolliert zu sein:Jedes Paar von Homologen macht typischerweise 1-2Crossing-overs. Die Wahrscheinlichkeit für gar keinCrossing-over eines Homologenpaares beträgt nur wenigerals 0,1%! Ein Ausbleiben der Rekombination kann sogar denweiteren Ablauf der Meiose blockieren. Vermutlich ist dieVerknüpfung der Homologen durch Crossing-over wichtig fürdie korrekte Verteilung auf die entstehenden Zellen.
Somatische Rekombination
Auch in mitotischen Zellzyklen kann es zur Rekombinationkommen: mitotische Rekombination, somatischeRekombination, weil nicht-reproduktive Zellen(Körperzellen, somatische Zellen) rekombinieren. Diesgeschieht z.B. in Drosophila, aber etwa 100mal seltener alsdie meiotische Rekombination. Dadurch können ausheterozygoten (Aa) Zellen homozygote werden (AA und aa),die in nebeneinanderliegenden Bereichen des Körpers ihrenPhänotyp ausprägen.
Somatische Rekombinationbei Drosophila

Mechanismen der Rekombination
Obwohl die meiotische Rekombination ein typischeukaryontischer Vorgang ist, sind die molekularenGrundlagen der Rekombination fast ausschließlich anBakterien und Bakteriophagen untersucht worden.Wird eine Bakterie gleichzeitig mit zwei Phagen infiziert,entstehen rekombinante Phagen. Das zeigt auch, daß zurRekombination Chromosomenstrukturen und synapto-nematische Komplexe nicht unbedingt nötig sind.
Rekombination durchPhagen-Doppelinfektion
r- („rapid growth“)führt zu vergrößertenPlaques (Lysehöfeim Bakterienrasen),tu- („turbid“)verursacht trübePlaques.
Mechanismen der RekombinationDurch Mutanten, die nicht zur Rekombination fähig sind,wurden beteiligte Gene identifiziert.Rekombinase RecA (Gen recA) ist eine ATPase, die anDNA-Einzelstränge bindet. Dabei entstehen ganze RecA-Filamente, die dann einen DNA-Doppelstrang nachhomologen Bereichen absuchen. In dem Fall kann RecA denDoppelstrang mit Helicase-Aktivität aufwinden und so dieBindung seines DNA-Einzelstrangs einleiten. Schließlichkann RecA den entstehenden Heteroduplex-Bereichausweiten (>branch migration).RecA ist auch bei der Rekombinations-Reparatur und demSOS-Reparatursystem beteiligt.Jeder Organismus hat ein RecA Homologes, unseres heißtRad51.
Mechanismen der RekombinationDie Aktionen von RecA
Mechanismen der Rekombination
Das RecBCD-System („Exonuclease V“, Gene recB, recC,recD) bindet an DNA-Doppelstrangbrüche und trennt denDNA-Doppelstrang durch Helicase-Aktivität in Einzel-stränge auf. An bestimmten Sequenzmotiven („Chi-Sequenz“) wird die Nucleaseaktivität von recD aktiv undschneidet einen der Einzelstränge ab. Der verbleibendeEinzelstrang kann dann in homologe Doppelsträngeeindringen. Der Komplex leistet sozusagen die Vorarbeit fürRecA, das Ergebnis, Doppelstrangbruch und Lücken-vergrößerung, entspricht dem „Doppelstrangbruch-Modell“bei der Meiose.
Mechanismen derRekombination -
RecBCD

Auflösen der Holliday-Struktur -RuvAB und RuvC, RecG
Für die abschließenden Schritte der Rekombination gibtzwei alternative Systeme. Wenn beide mutiert sind, ist dieRekombination von E. coli vollständig verhindert.RuvA bindet als Tetramer an die vier Stränge der Holliday-Verzweigung, die ATPase RuvB bindet daran als Hexamerund läßt die Verzweigung mit 10-20 bp/sec wandern (branchmigration). Endonuclease RuvC spaltet einen Strang undleitet so die Auslösung des Chiasmas ein.RecG kann ebenfalls die Holliday-Junction auflösen.
Auflösen der Holliday-Struktur -Aktion des RuvAB-Komplexes
Schritte derbakteriellen
Rekombination
Die einleitende Stranglückeentsteht häufig bei derReparatur von DNA-Schäden.
(Anders als gezeigt bindetRecA natürlich an den blauenEinzelstrang und „sucht“ mitihm sein Homologes, trenntden roten Doppelstrang auf.)
Spezialisierte RekombinationDie Integration von Phagen-DNA ins Bakterienchromosomgeschieht nicht über homologe Rekombination, sondern übereine „spezialisierte Rekombination“, bei der bestimmteAnlagerungsstellen (att-Stellen) von Phagen- und Bakterien-DNA attP, attB) reziprok rekombinieren. Fehlt die Lambda-att-Site (attB) in E. coli, kommt es an „sekundärenAnlagerungsstellen“ dennoch zum Einbau des λ-Phagen, abermit über 1000mal schlechterer Effizienz. Zum Wieder-ausschneiden des Prophagen rekombinieren die beiden (durchde Rekombination neu entstandenen) att-Sites (attL, attR)erneut.
SpezialisierteRekombination
Für Ein- und Ausbauwerden die Faktoren Intund IHF benötigt, zurExcision noch zusätzlichFaktor Xis. Durch dieVeränderung der att-Sitesbeim Einbau wird es demPhagen möglich, Ein- undAusbau getrennt zusteuern.
Spezialisierte Rekombination
Die rekombinierenden att-Bereiche binden nicht durchhomologe Bereiche aneinander, sondern beide an einen Core(Kern) aus Int-Protein. Dieser Komplex wird auch Intasomgenannt. Erst bei der Strangaustausch-Reaktion ist eineHomologie der beiden att-Sequenzen erforderlich.Für die Int-Bindung an attR (zur Excision) ist Xis nötig.

Transposition
Noch einen anderen Weg der DNA-Rekombinationverwenden Transposons, im Genom wandernde DNA-Elemente. Hierbei sind keinerlei Homologien der Donor-und Rezipientenbereiche erforderlich.Die Transposone codieren für die notwendigen Enzyme, dieden Bewegungsprozeß katalysieren.
(Entdeckung: Barbara McClintock 1948 bei Mais -Nobelpreis 1983)
Transposition - IS-ElementeDie einfachsten Transposone sind die IS-Elemente(Insertionssequenzen), von denen normale E. coli-Stämmemeist mehrere Exemplare verschiedener Typen enthalten. DieIS-Sequenzen sind von „inverted terminal repeats“ flankiert,also von identischen Sequenzen in entgegengesetzterOrientierung.Viele IS-Elemente integrieren an beliebiger Stelle im Genom,andere haben Vorzugsbereiche („hotspots“). Beim Einbauduplizieren sie eine kurze Sequenz auf dem Chromosom, sodaß sie von einem „direkt repeat“ dieser Sequenz umgebensind.Fast alle IS-Elemente codieren für genau ein Protein, dieTransposase.
Transposition -IS-Elemente
Mechanismen derTransposition
Beim Transposoneinbau wird dieZiel-DNA zuerst in beiden Strängenaufgeschnitten, die beiden Strang-brüche sind um einige Basengegeneinander versetzt. DasTransposon wird an diebeiden überstehenden Endenanligiert, dann die beiden Lückenaufgefüllt und ligiert. Dadurch istder Bereich zwischen denursprünglichen Einzelstrang-brüchen („nicks“) dupliziert.
Mechanismen der Transposition
Das ursprüngliche Transposon kann sich an die neue Stellekopieren (replikative Transposition), oder es kann von derursprünglichen Position an die neue Stelle wandern. Dabeikann es eine gebrochene DNA zurücklassen (nicht-replikative Transposition; hier muß die Zelle denDoppelstrangbruch reparieren oder - wenn mehrere Kopiendes Chromosoms vorhanden sind - den defekten Strangabbauen) oder einen intakten Strang (konservativeTransposition).Einige Transposone benutzen nur einen dieser Mechanismen(TnA nur replikativ), andere können alle durchführen.
Mechanismen derTransposition
nicht-replikative Transposition
replikative Transposition
konservative Transposition

ZusammengesetzteTransposone
Einige Transposone tragenResistenzgene gegen Anti-biotika. Sie werden mit „Tn“und einer Nummer benannt.Viele Transposone dieses Typssind „zusammengesetzt“aufgebaut: Der Genbereich istvon zwei IS-Elementenflankiert. Diese können ingleicher („Tandem“-)Orientierung oder zueinanderinvers orientiert sein.
ZusammengesetzteTransposone
Die flankierenden IS-Elementekönnen sowohl sich selbst (wenn siefunktionell sind) als auch dasgesamte zusammengesetzteTransposon bewegen (dazu muß nureins der IS-Elemente vollfunktionsfähig sein). Dabei ist derBereich zwischen den IS-Elementennur passiver Passagier. Auf einemPlasmid können die IS-Elemente stattihrem Tn-Teil auch den Rest desPlasmids mitnehmen (funktionierenalso „in beiden Richtungen“).
Zusammengesetzte Transposone
Die IS-Elemente bewegen häufiger sich allein als das ganzezusammengesetzte Transposon. Die treibende Kraft für denErhalt der zusammengesetzten Transposone sind ihreResistenzgene, d.h., nur in Gegenwart des Antibiotikums istein echter Vorteil für die Zelle vorhanden.
Mutationen durch Transposone
Beim Einbau eines Transposons in ein Gen wird dies meistinaktiviert. Wird das Transposon durch das Reparatursysteman den duplizierten Enden rekombiniert und ausgeschnitten,bleiben meist kleine Transposonreste zurück („unpräziseExcision“). Dadurch bleibt das Gen mutiert. Nur bei dersehr seltenen präzisen Excision wird der Ursprungszustanddes Gens wiederhergestellt.
Transposition
Durch die Wanderung von Transposonen im Genom kann eszu Rearrangements des Genoms kommen. Einerseits führtdas Transpositionsereignis selbst oft zu Deletionen oderInversionen (Umklappen von DNA-Bereichen). Andererseitsbilden multiple Kopien eines Transposons mobileAngriffsstellen für die homologe Rekombination, da sie jazueinander homolog sind. Das Ergebnis können Deletionen,Inversionen, Insertionen und Translokationen vonGenomabschnitten sein, also eine massive Umsortierung desBakterienchromosoms.10-15% aller spontanen Mutationen in E. coli sind auf IS-Elemente und Transposons zurückzuführen.
Häufigkeit der Transposone
Durch die Genomsequenzierungen der verschiedenenOrganismen stellte sich heraus, das unser Genom fast zurHälfte (45%, Frosch 77%, Mais 60%, Hefe 4%, E. coli0,3%) aus (defekten und stillgelegten) Transposonenbesteht! Auch bei uns dürfte das Rearrangements vonChromosomenteilen und andere Rekombinationsereignisseermöglichen.

Retrotransposone und RetrovirenAndere Transposone bewegen sich über ein RNA-Zwischen-produkt, aus der sie durch Reverse Transkriptase wieder inDNA zurückgeschrieben werden. Dazu gehört das Retro-transposon Ty der Hefe. Nahe verwandt sind Retroviren, zuderen Lebenszyklus diegenomische Integration gehört.Vermutlich sind RetrotransposoneRetroviren, die die Fähigkeit zurZell-Lyse und Infektion verlorenhaben. Durch starke Expressionder Gene auf Ty kann man dieHefezelle mit virusartigenPartikeln füllen >>.
Retrotransposone undRetroviren
Lebenszyklus der Retroviren
Bakterielle Konjugation und DNA-Transfer
Bei Bakterien kommt es auch zu Genom-Neukombinationendurch Mechanismen, die gewisse Ähnlichkeit mit dersexuellen Genomkombination der Eukaryonten hat. Dabeiübertragen Donor-Zellen DNA auf Rezipienten-Zellen. DieDonor-Bakterien besitzen ein F-Teilchen, entweder alsfreies Plasmid, dann sind sie F+, oder integriert insChromosom, dann sind die Zellen Hfr (high frequency ofrecombination). F+-Zellen übertragen das F-Episom in F--Rezipienten. Man bezeichnet das als „infektiöseSexualität“ oder F-Infektion.
Bakterielle Konjugation und DNA-TransferDurch Integration des F-Teilchens entstehen (selten) aus F+
Hfr-Zellen. Auch der umgekehrte Prozeß (Excision) kommtvor, dabei werden oft Teile des Bakterienchromosoms mitdem F-Teilchen ausgeschnitten, es entsteht dann ein F‘-Faktor. Auch F‘ kann in F--Stämme übertragen werden,dabei werden die ehemals chromosomalen Bereichemitübertragen (das wird als F-Duktion bezeichnet) undkönnen in der Akzeptorzelle mit dem Chromosom homologrekombinieren. Durch die langen homologen Bereichebauen sich F‘-Episomen öfter ins Chromosom ein(produzieren also Hfr), werden aber auch öfter wiederexcisiert (da das integrierte F-Teilchen von den homologenBereichen flankiert ist).
Bakterielle Konjugation und DNA-Transfer
Hfr-Zellen übertragen, beginnend an der F-Teilchen-Region,ihr Bakterienchromosom in den Rezipienten(Chromosomentransfer), der durch Rekombination mitseinem eigenen Chromosom die übertragenen Geneübernehmen kann.

Bakterielle Konjugation und DNA-Transfer
Wird der Zell-Zell-Kontakt beim Chromosomentransfervorzeitig (Gesamtdauer 90 Minuten) unterbrochen, werdennur Teile des Chromosoms übertragen. Anhand der Zeit biszu ihrem Transfer (Methode der Transferunterbrechung)ließen sich die Genorte auf dem E. coli-Chromosom sehrgut kartieren. Da die Transfergeschwindigkeit praktischkonstant ist, stimmen die Abstände genau mit denphysikalischen Abständen (kBp) überein.
Genomkartevon E. coli
Bakterielle Konjugation und DNA-Transfer
Wegen der Länge der DNA und der Übertragungszeitgelingt eine komplette Chromosomenübertragung durch Hfrnur sehr selten. Die Rezipienten bleiben daher meist F- (dader Transfer nahe dem Ende des F-Teilchens beginnt, wirdder Großteil des F-Teilchens als letztes übertragen).
Bakterielle Konjugation und DNA-Transfer
33 kBp des F-Teilchens codieren für Transferfunktionen(tra-Region). Die meisten Transfergene liegen auf einemOperon, das von traJ angeschaltet wird. finP codiert eineantisense-RNA, die traJ ausschaltet.
Bakterielle Konjugation und DNA-Transfer
traA codiert für das Pilin, das den Sexualpilus bildet (amZusammenbau sind weitere 12 Gene beteiligt). Damit wirdKontakt zu F--Zellen hergestellt. Die F+-Zellen bildenProteine (Gene traS, traT), die das Andocken des Pilusverhindern, so daß sie nicht untereinander konjugieren.Der Pilus ist auch Andockstelle für einige Phagen, diesekönnen also nur F-positive Zellen infizieren. Der Pilus istnicht ein DNA-Transferkanal, die beiden Zellen müssenzum Transfer direkten Kontakt aufnehmen.
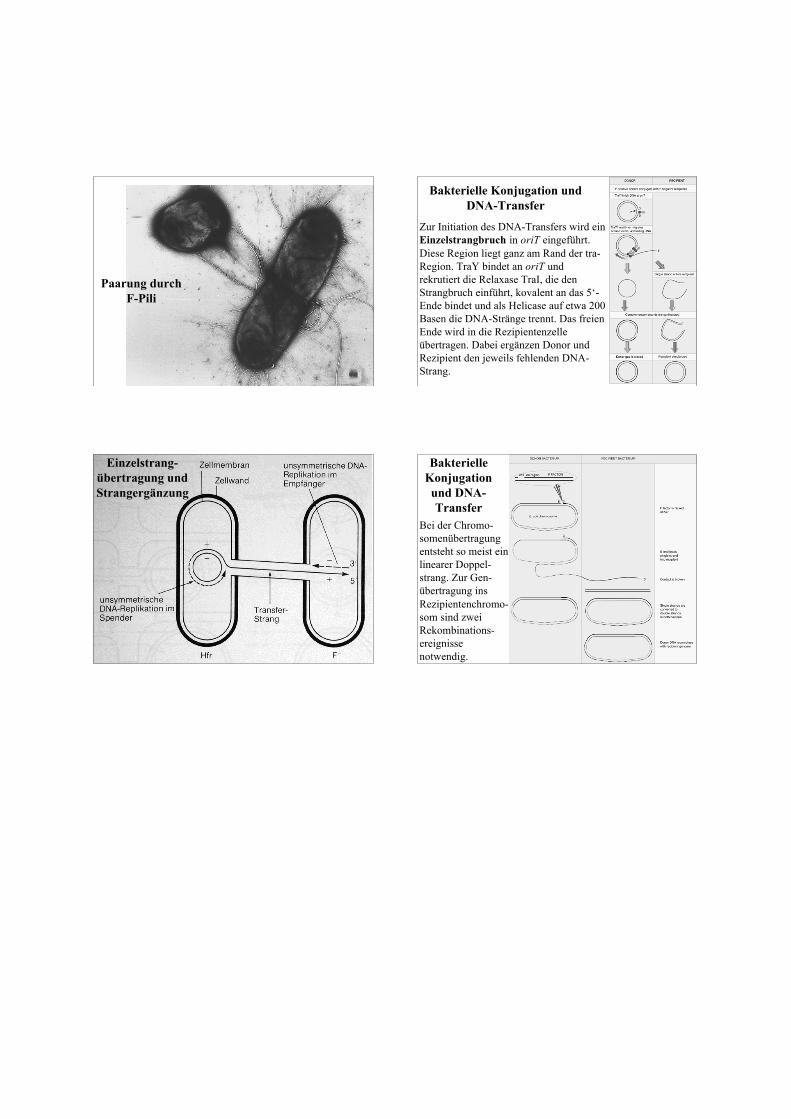
Paarung durchF-Pili
Bakterielle Konjugation undDNA-Transfer
Zur Initiation des DNA-Transfers wird einEinzelstrangbruch in oriT eingeführt.Diese Region liegt ganz am Rand der tra-Region. TraY bindet an oriT undrekrutiert die Relaxase TraI, die denStrangbruch einführt, kovalent an das 5‘-Ende bindet und als Helicase auf etwa 200Basen die DNA-Stränge trennt. Das freienEnde wird in die Rezipientenzelleübertragen. Dabei ergänzen Donor undRezipient den jeweils fehlenden DNA-Strang.
Einzelstrang-übertragung undStrangergänzung
Bei der Chromo-somenübertragungentsteht so meist einlinearer Doppel-strang. Zur Gen-übertragung insRezipientenchromo-som sind zweiRekombinations-ereignissenotwendig.
BakterielleKonjugationund DNA-Transfer





























