Embed Size (px)
Citation preview

1MULTIPLICATION

2
Der Replikationszyklus
• Virusreplikation kann in 8 Phasen eingeteilt werden– Anlagerung -Anheftung -Adsorption– Eindringen -Penetration– Freisetzung der viralen Nukleinsäuren -Uncoating– Expression -Transkription– Replikation -Translation– Zusammenbau -Replication– Reifung -Assembly– Ausschleussung -Release

3
Der Replikationszyklus
Es handelt sich um eine mehr oder weniger willkürliche Unterteilung, die zur einfacheren Erklärung des Replikationszyklus eines nicht-existenten “typischen” Virus dienen soll.
• Unabhängig vom jeweiligen Wirt, durchlaufen alle Viren diese Stadien annäherungsweise während ihrer Replikation.
• Nicht alle der hier beschriebenen Schritte sind erkennbar für jeden Virustyp vorhanden, oft gehen sie fließend ineinander über oder laufen fast zeitgleich ab.
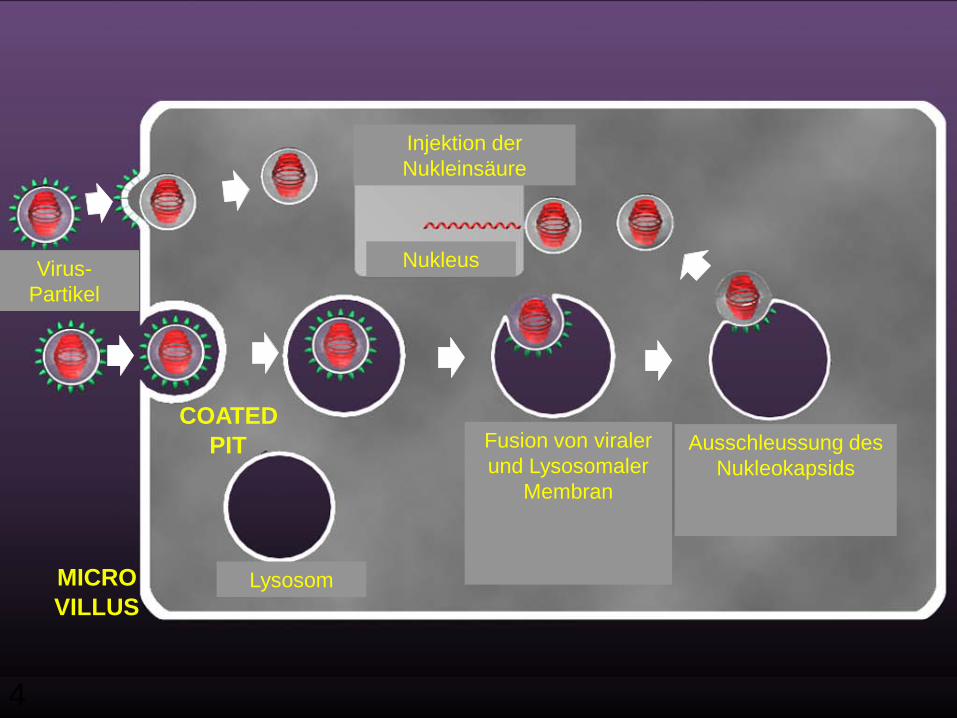
4
Injection ofNucleic Acid
NucleusVIRUSPARTICLES
COATEDPIT
LYSOSOME
FUSIONOF VIRAL
ANDLYSOSOMALMEMBRANE
EJECTIONOF
NUCLEOCAPSID
MICROVILLUS
Fusion von viralerund Lysosomaler
Membran
Ausschleussung desNukleokapsids
Nukleus
Injektion derNukleinsäure
Lysosom
Virus-Partikel
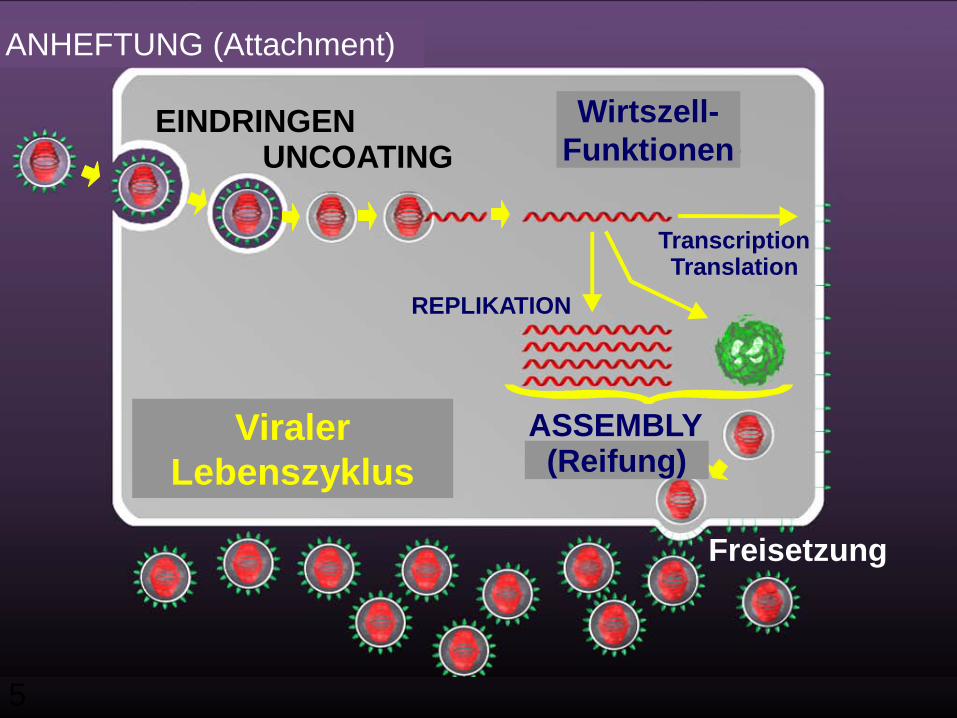
5
VIRAL LIFE CYCLE
EINDRINGENUNCOATING FUNCTIONS
ASSEMBLY
TranscriptionTranslation
(MATURATION)
REPLIKATION
Freisetzung
Viraler Lebenszyklus
ANHEFTUNG (Attachment)
Wirtszell-Funktionen
(Reifung)

6
Genom-Replikation & Gen-Expression
• Die Replikationsstrategie eines beliebigen Virus hängt von der Art seines genetischen Materials ab.
• Viren können in sieben Gruppen eingeteilt werden –ein Schema dazu wurde zuerst von David Baltimore1971 vorgeschlagen.
• Anfänglich enthielt diese Klassifikation nur sechs Gruppen, sie wurde jedoch um die Gruppe der Hepadna- und Caulimoviren erweitert, die eine spezielle Art der Genom-Replikation aufweisen.

7
Baltimore - KlassifikationssystemKlasse I: Doppelsträngige DNA Papovaviridae, Adenoviridae, Herpesviridae,
Poxviridae
Klasse II: Einzelsträngige (+) DNA Parvoviridae
Klasse III: Doppelsträngige RNA Reoviridae
Klasse IV: Einzelsträngige (+) RNA Picornaviridae, Caliciviridae, Flaviviridae, Togaviridae, Coronaviridae
Klasse V: Einzelsträngige (-) RNA (i) Segmentiert: Orthomyxoviridae (i) (ii) Nicht-segmentiert:
Rhabdoviridae
Klasse VI: Einzelsträngige (+) RNA mit DNA als Zwischenstufe Retroviridae
Klasse VII: Doppelsträngige DNA mit RNA als ZwischenstufeHepadnaviridae, Caulimovirus

8
Klasse I: doppelsträngige DNA
• Diese Klasse kann in zwei Untergruppen unterteilt werden:
• A) Die Replikation findet ausschließlich im Nukleus statt. Die Replikation dieser Viren hängt im wesentlichen von zellulären Faktoren ab.
• B) Die Replikation findet im Zytoplasma statt (Poxviridae). Diese Viren haben die notwendigen Faktoren zur Transkription & Replikation ihrer Genome entwickelt (oder sich angeeignet) und sind deswegen relativ unabhängig von zellulären Proteinen.

9
DNA-VIREN
• brauchen mRNAs für die Protein-Herstellung– Der Wirt besitzt eine DNA-abhängige RNA Polymerase, und
alle anderen Proteine, die zur Herstellung von richtig modifizierter mRNA nötig sind
• müssen ihre DNA replizieren– Der Wirt hat eine DNA Polymerase, plus zusätzliche Proteine,
die zur Replikation der DNA nötig sind
• DIESE MASCHINERIE BEFINDET SICH IM NUKLEUS• Pockenviren replizieren im Zytoplasma und müssen
deswegen alle diese Funktionen mitbringen
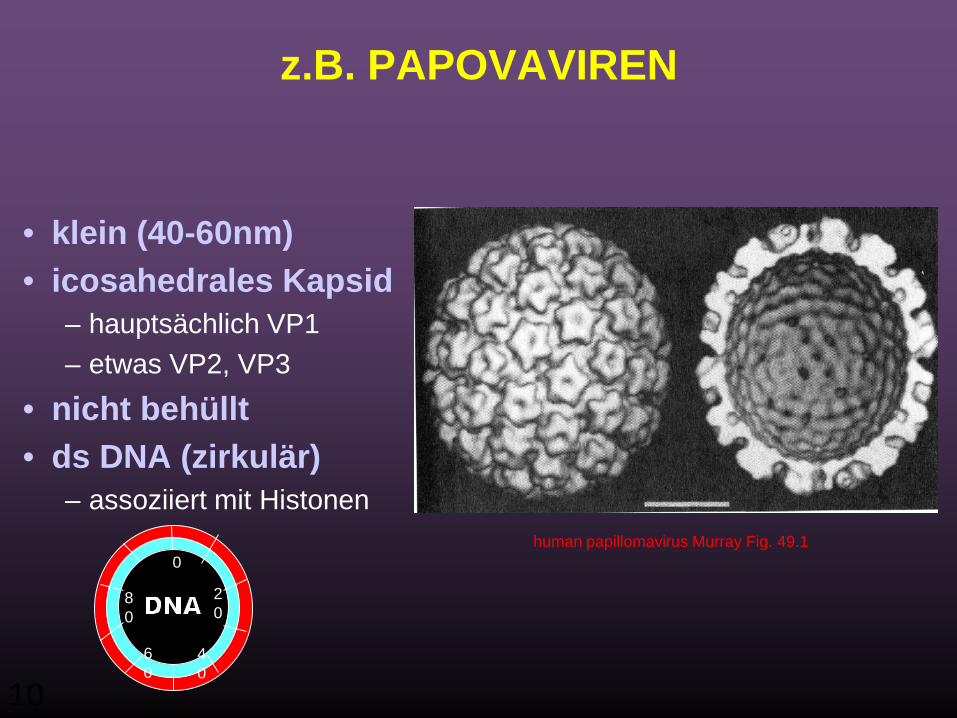
10
z.B. PAPOVAVIREN
• klein (40-60nm) • icosahedrales Kapsid
– hauptsächlich VP1– etwas VP2, VP3
• nicht behüllt• ds DNA (zirkulär)
– assoziiert mit Histonenhuman papillomavirus Murray Fig. 49.1
20
40
60
80
0
11
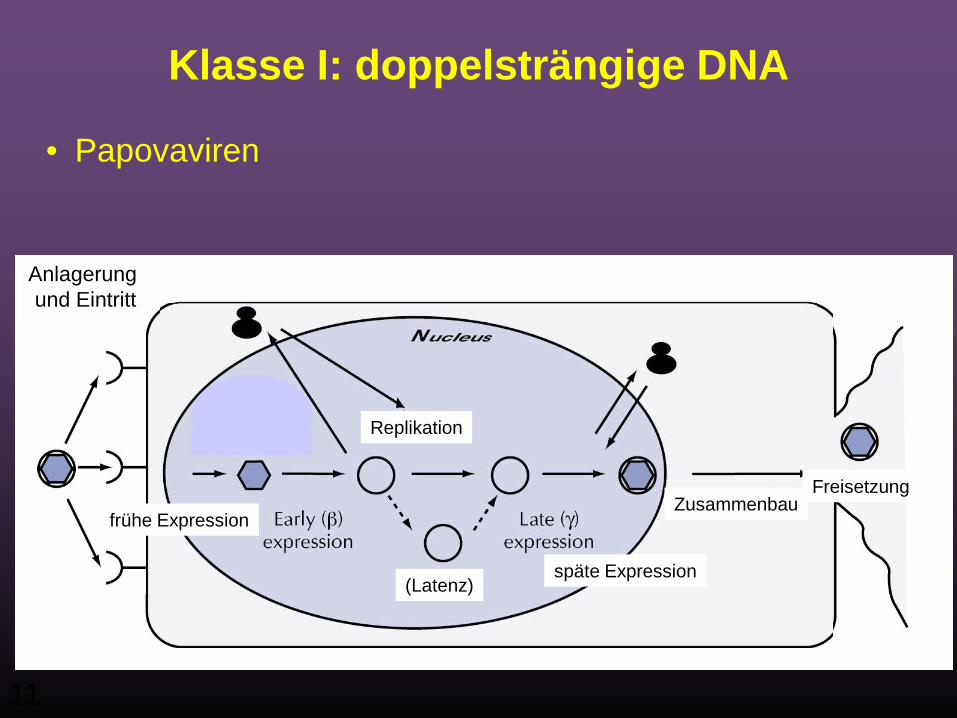
Klasse I: doppelsträngige DNA
• Papovaviren
Anlagerung und Eintritt
Replikation
frühe Expression
späte Expression(Latenz)
FreisetzungZusammenbau

13
LATENTER und LYTISCHER ZYKLUS• FRÜHE PHASE
– PROTEINE, DIE ZUR DNA REPLIKATION BENÖTIGT WERDEN
– REGULATORISCHE PROTEINE– OFT PROTEINE, DIE DIE WIRTSZELLE
VERÄNDERN
• SPÄTE PHASE– DNA REPLIKATION– NEUE STRUKTUR PROTEINE
NUR LYTISCHER ZYKLUS
14
20
4060
80
0
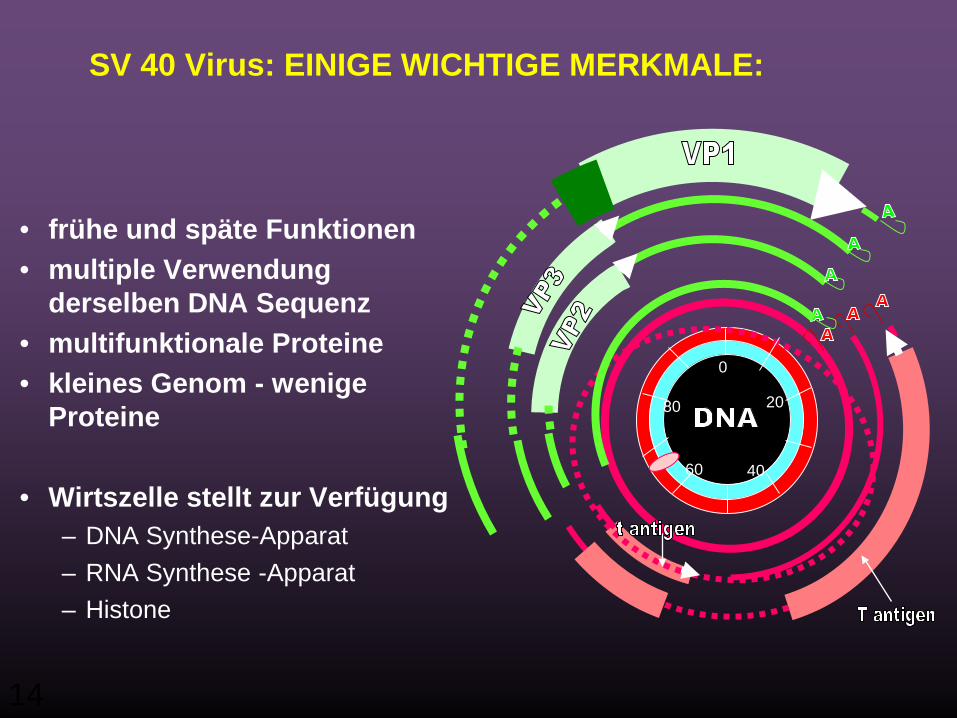
SV 40 Virus: EINIGE WICHTIGE MERKMALE:
• frühe und späte Funktionen• multiple Verwendung
derselben DNA Sequenz • multifunktionale Proteine• kleines Genom - wenige
Proteine
• Wirtszelle stellt zur Verfügung– DNA Synthese-Apparat– RNA Synthese -Apparat– Histone

15
nur LYTISCHER ZYKLUS• SPÄTE PHASE
– DNA REPLIKATION– NEUE STRUKTURPROTEINE
• Replikation ist bidirektional – (Es existieren zwei Replikationsgabeln pro zirkulärem DNA
Genom und die Replikation läuft über einen Führungs- und einen nachfolgenden Strang (“leading/lagging” strands, Okazaki Fragmente, DNA Ligase, etc.) ab.
• Dieser Prozess der DNA Replikation ist sehr ähnlich zu dem, der in der Wirtszelle vor sich geht – das ist nicht erstaunlich, da das Virus hauptsächlich die
Wirtszellmaschinerie benutzt, ausser was das T Antigen betrifft.• Histone der Wirtszelle komplexieren die neu
synthetisierte DNA.

16
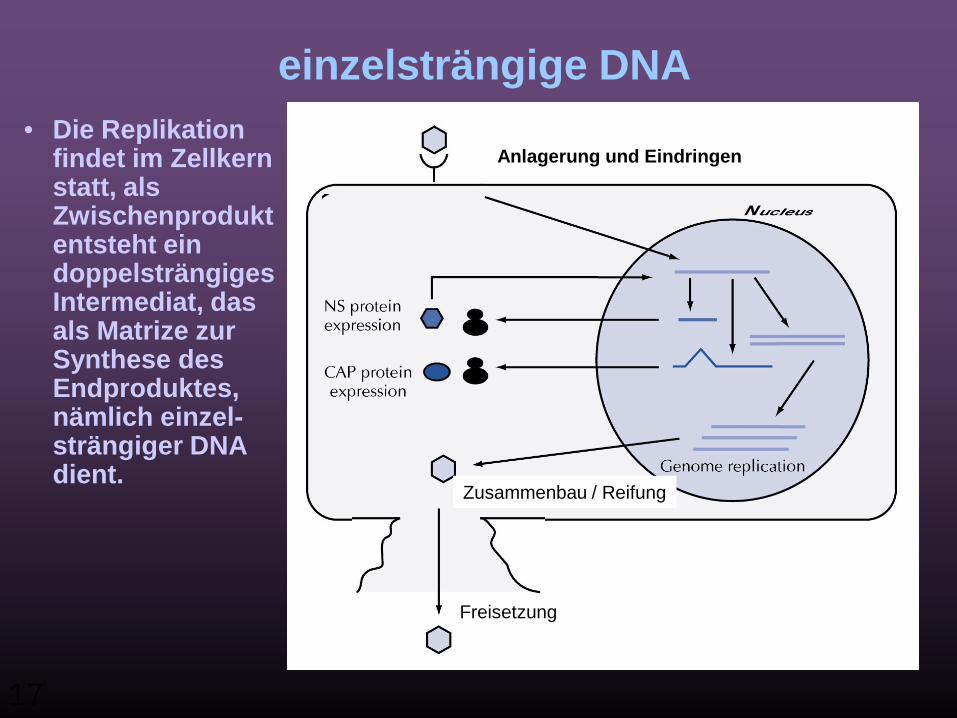
II) ss DNA Viren: Beispiel: Parvovirus
17
einzelsträngige DNA• Die Replikation
findet im Zellkern statt, als Zwischenprodukt entsteht ein doppelsträngiges Intermediat, das als Matrize zur Synthese des Endproduktes, nämlich einzel-strängiger DNA dient.
Anlagerung und Eindringen
Zusammenbau / Reifung
Freisetzung

18
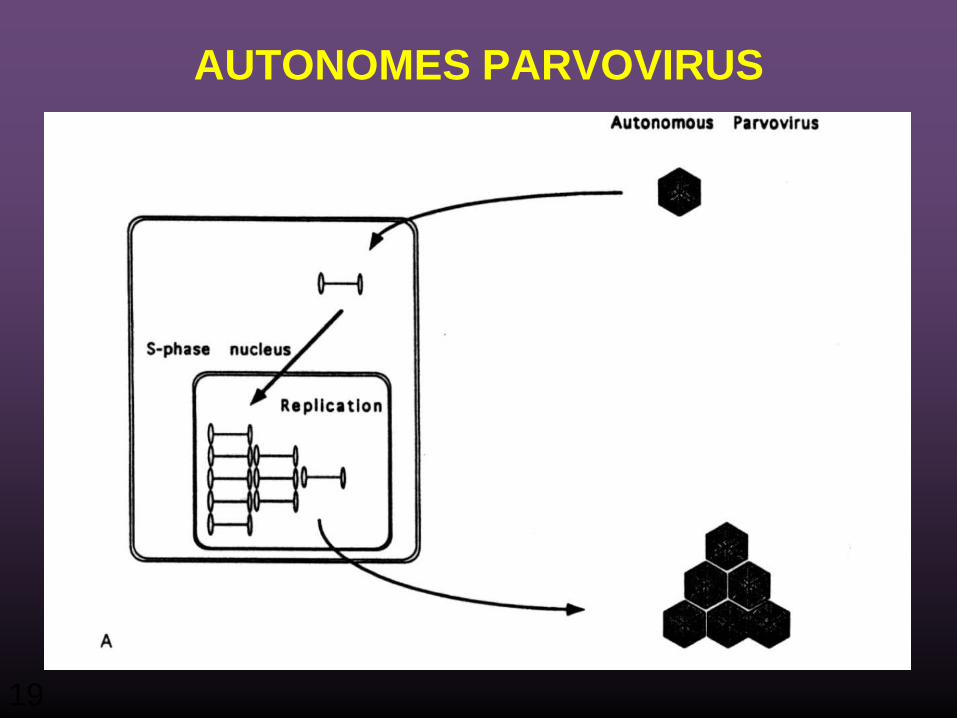
PARVOVIRUS
• Drei Gattungen – Parvovirus – Dependovirus (adeno-assoziiertes Virus) – Erythrovirus
• Nicht behülltes (nacktes) icosahedrales Kapsid, 20 – 25nm• linear, ss DNA, 5 kb • Parvovirus: verpackt ausschließlich negativ-Strang-DNA • Andere Gattungen verpacken beide, positiv und negativ-
strängige DNA • Einzigartiger Mechanismus der DNA Replikation bindet
die Enden der Genome ein.
19
AUTONOMES PARVOVIRUS
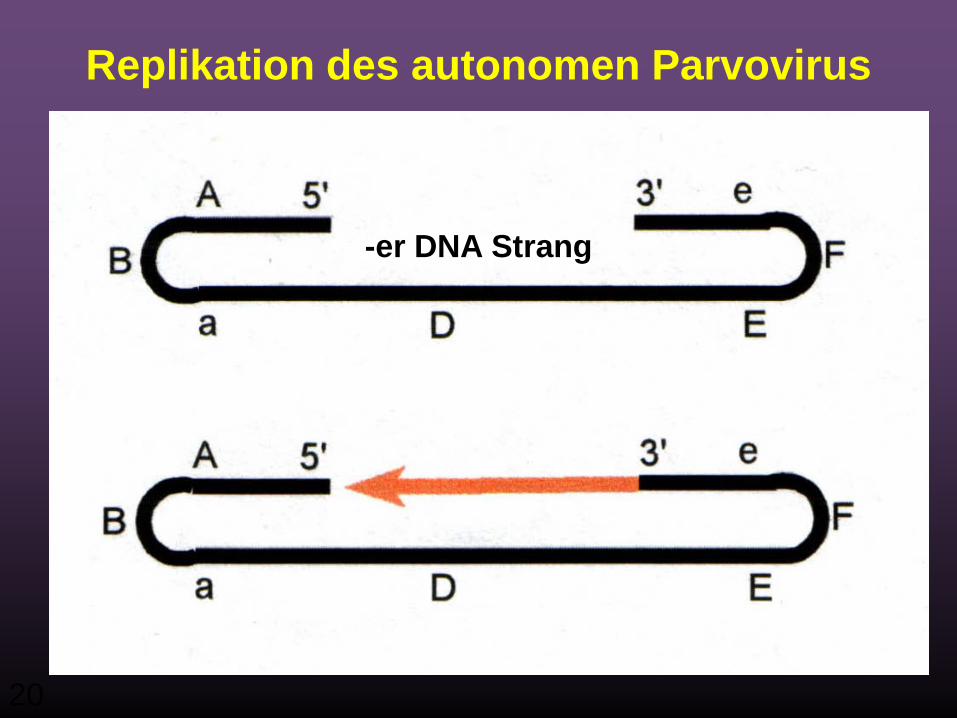
20
Replikation des autonomen Parvovirus
-er DNA Strang
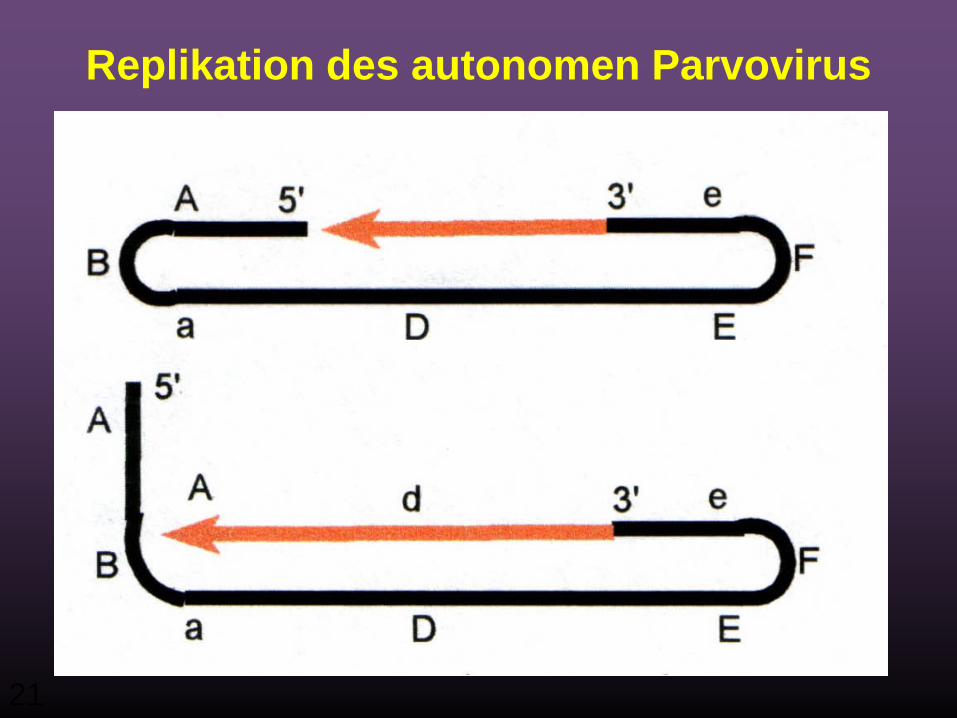
21
Replikation des autonomen Parvovirus
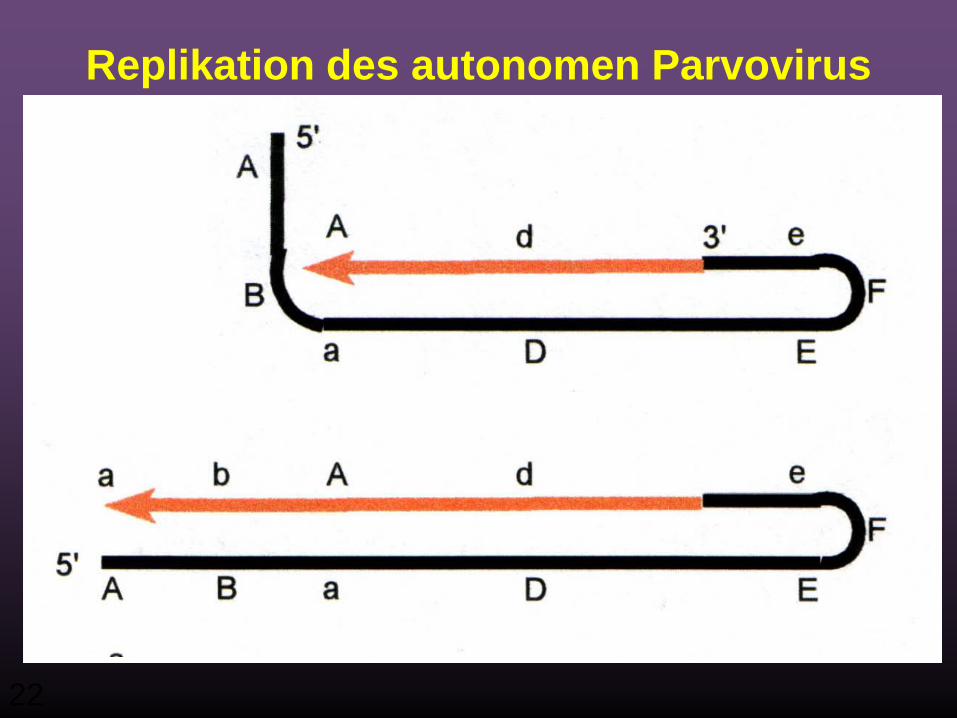
22
Replikation des autonomen Parvovirus
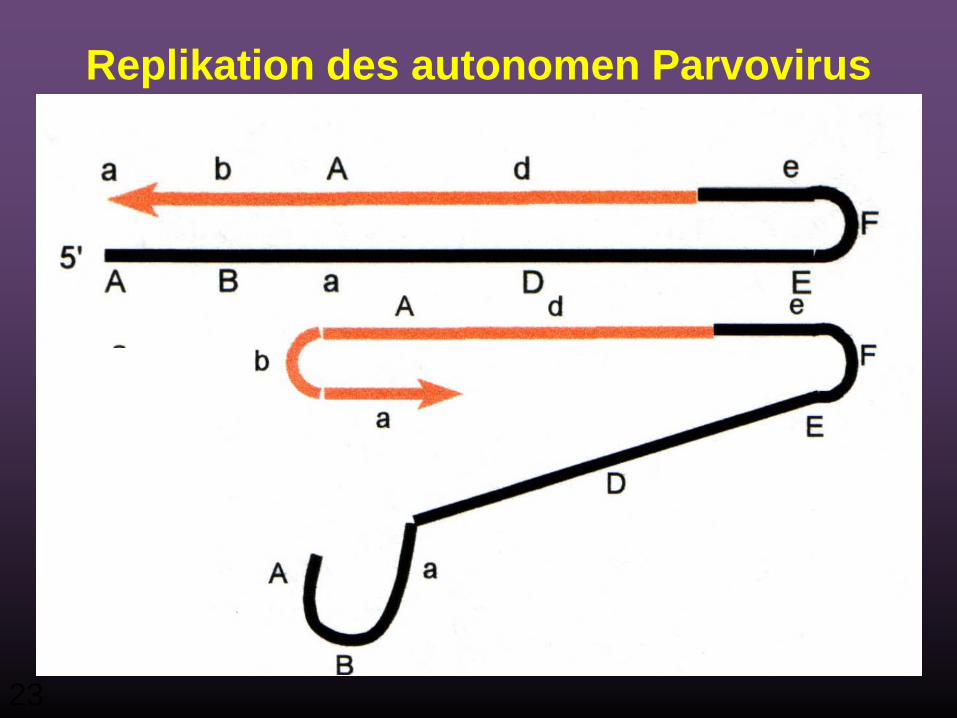
23
Replikation des autonomen Parvovirus
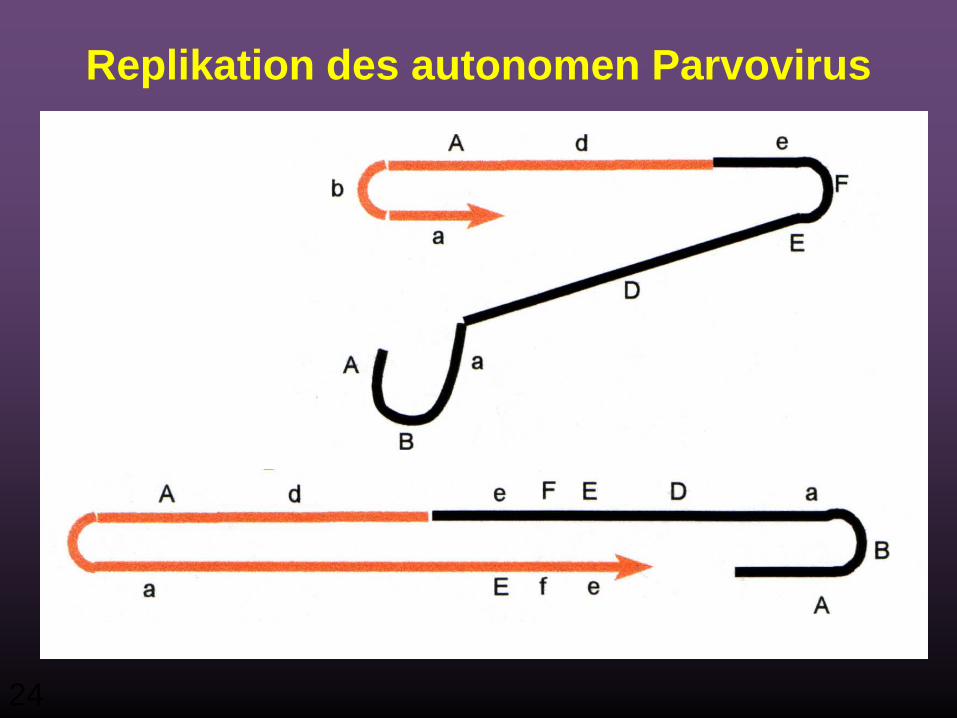
24
Replikation des autonomen Parvovirus
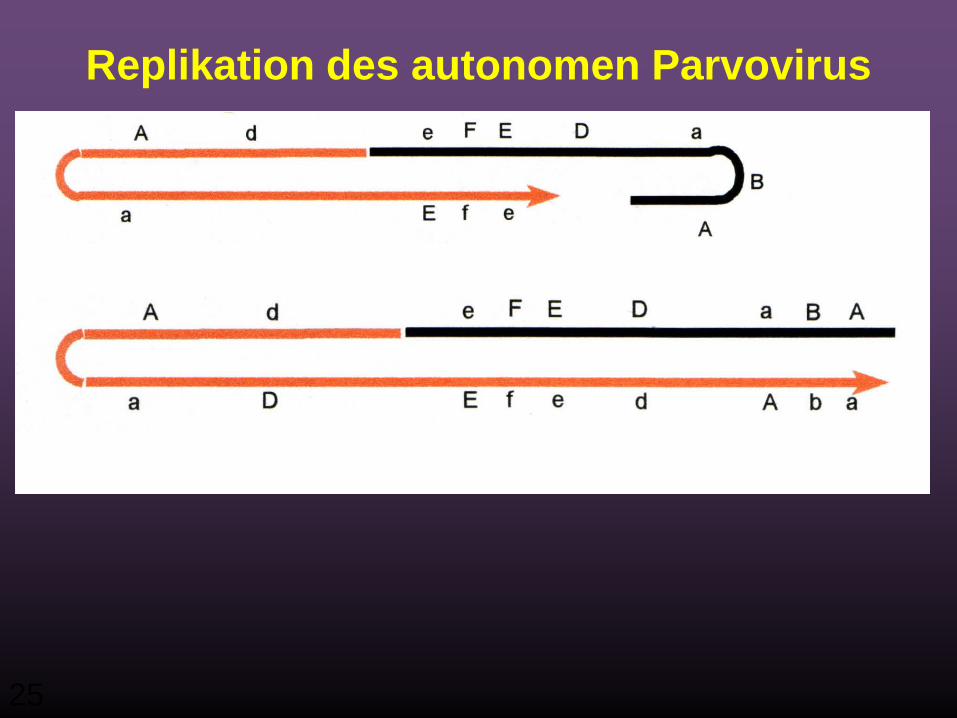
25
Replikation des autonomen Parvovirus
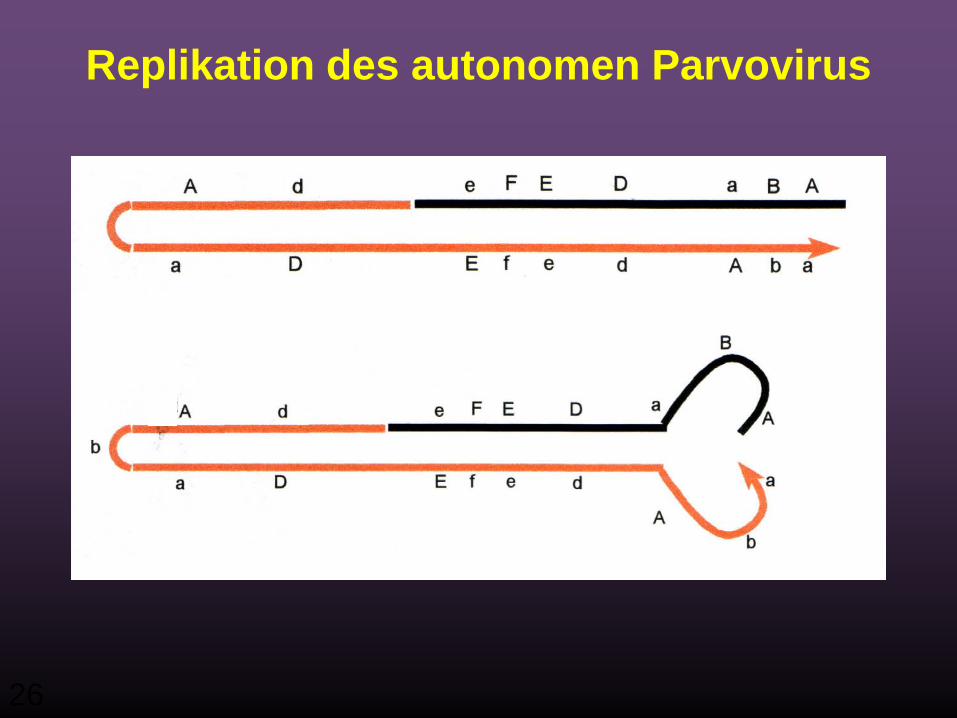
26
Replikation des autonomen Parvovirus
27
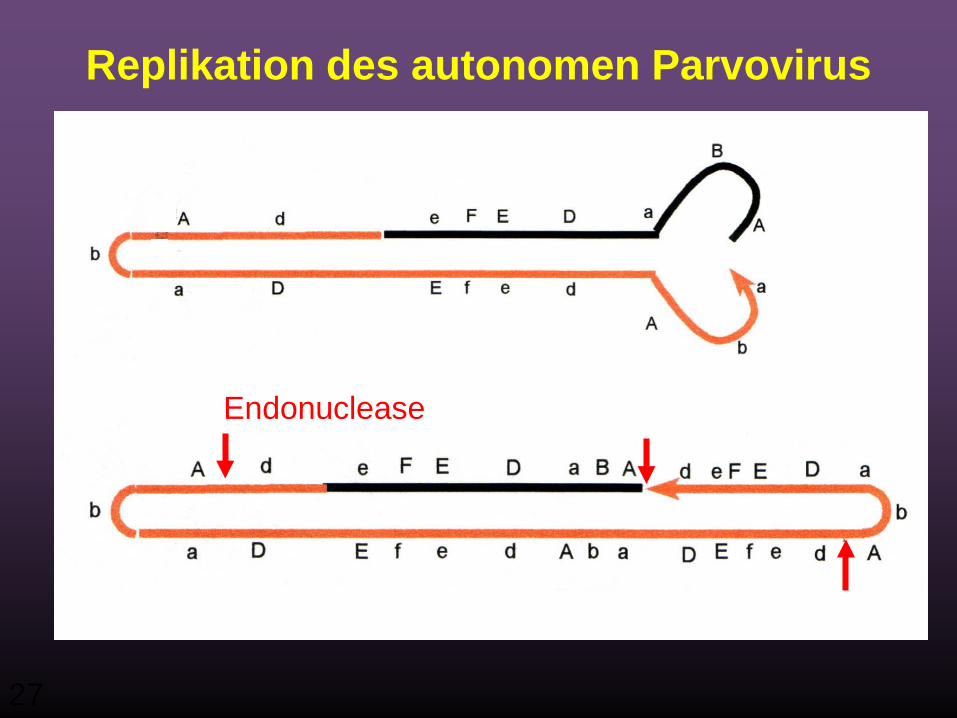
Endonuclease
Replikation des autonomen Parvovirus
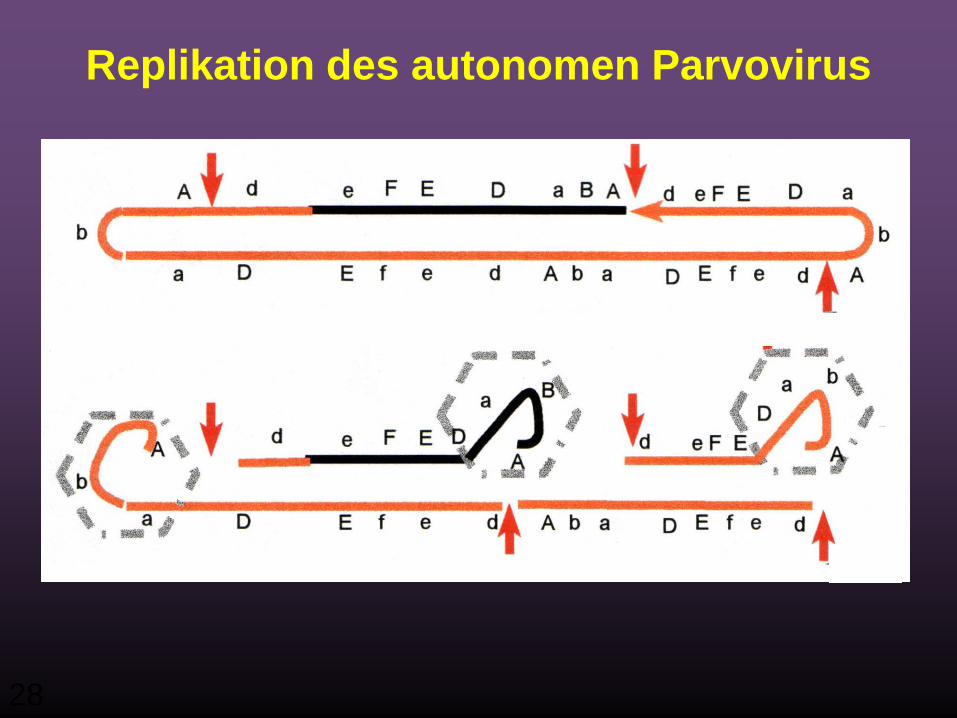
28
Replikation des autonomen Parvovirus
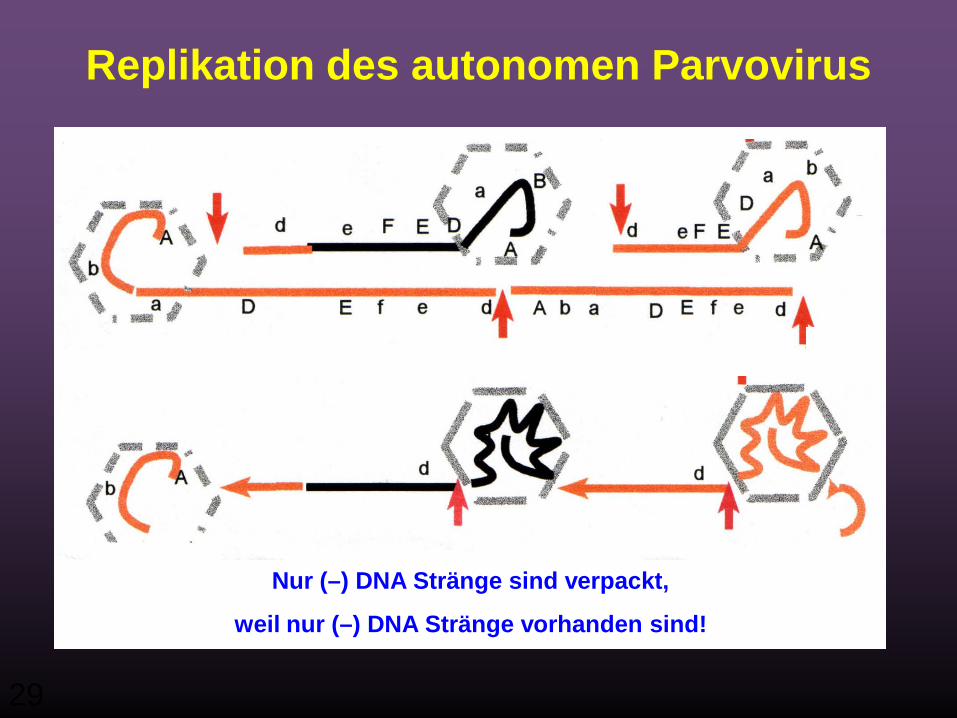
29
Nur (–) DNA Stränge sind verpackt,
weil nur (–) DNA Stränge vorhanden sind!
Replikation des autonomen Parvovirus

30
EXKURS: RNA-VIRUS STRATEGIEN

31
RNA-VIRUS STRATEGIEN
RNA -> RNARNA-abhängige RNA Polymerase
RNA -> DNARNA-abhängige DNA Polymerase- reverse Transkriptase
WIRTSZELL-DNA -> RNADNA-abhängige RNA Polymerase

32
ALLE TIER- RNA-VIREN KODIEREN FÜR EINE
POLYMERASE
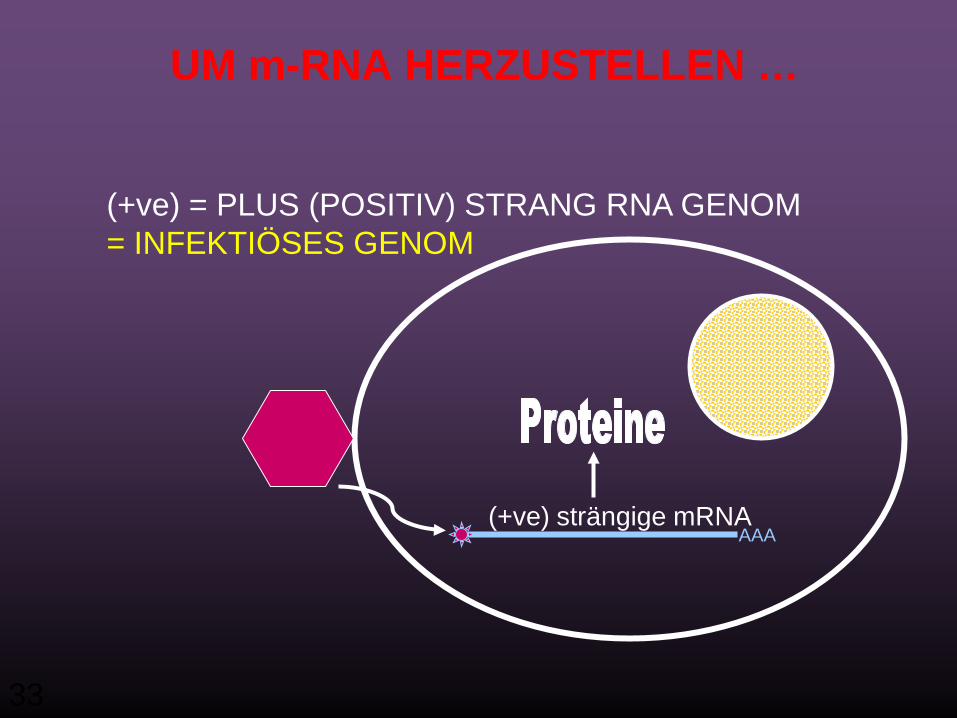
33
UM m-RNA HERZUSTELLEN …
(+ve) = PLUS (POSITIV) STRANG RNA GENOM= INFEKTIÖSES GENOM
AAA(+ve) strängige mRNA
34
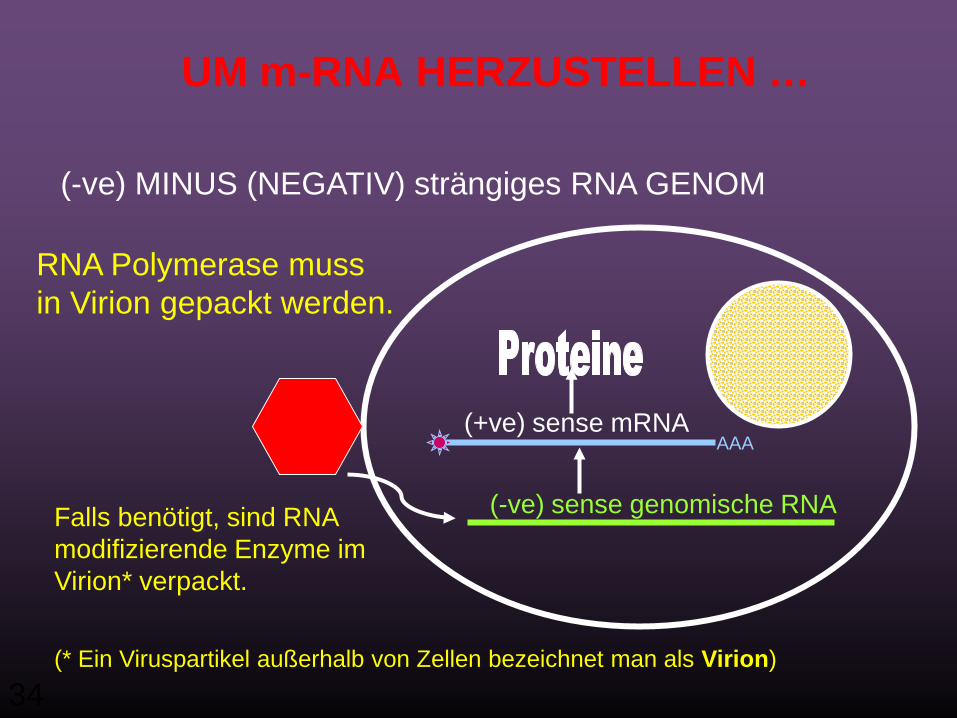
UM m-RNA HERZUSTELLEN …
(-ve) MINUS (NEGATIV) strängiges RNA GENOM
AAA(+ve) sense mRNA
(-ve) sense genomische RNAFalls benötigt, sind RNA modifizierende Enzyme im Virion* verpackt.
RNA Polymerase mussin Virion gepackt werden.
(* Ein Viruspartikel außerhalb von Zellen bezeichnet man als Virion)
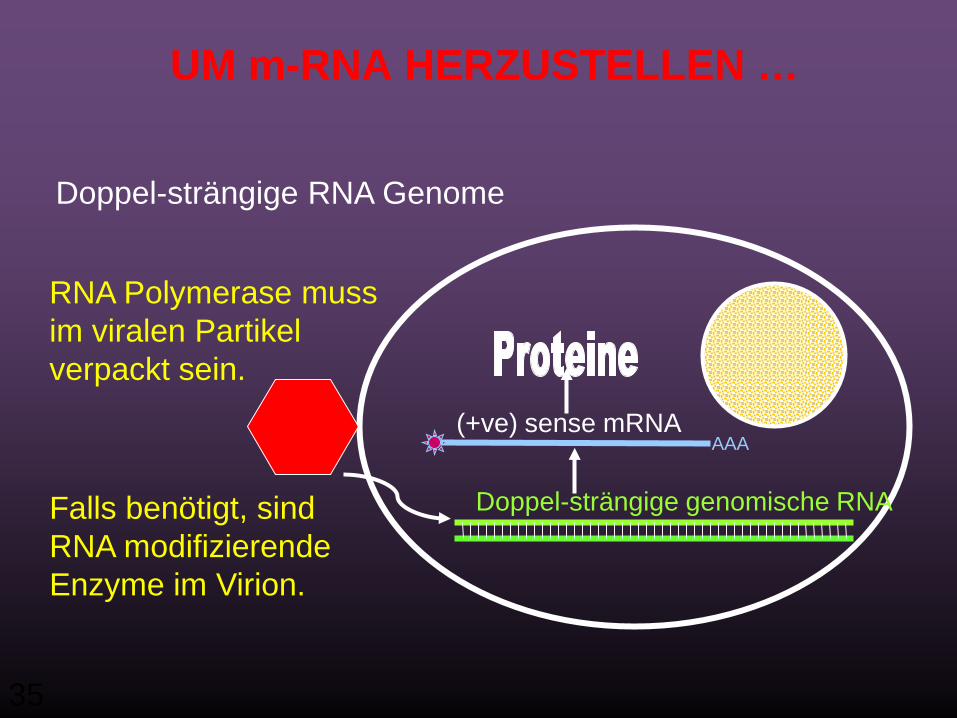
35
UM m-RNA HERZUSTELLEN …
Doppel-strängige RNA Genome
AAA(+ve) sense mRNA
Doppel-strängige genomische RNAFalls benötigt, sind RNA modifizierende Enzyme im Virion.
RNA Polymerase muss im viralen Partikel verpackt sein.
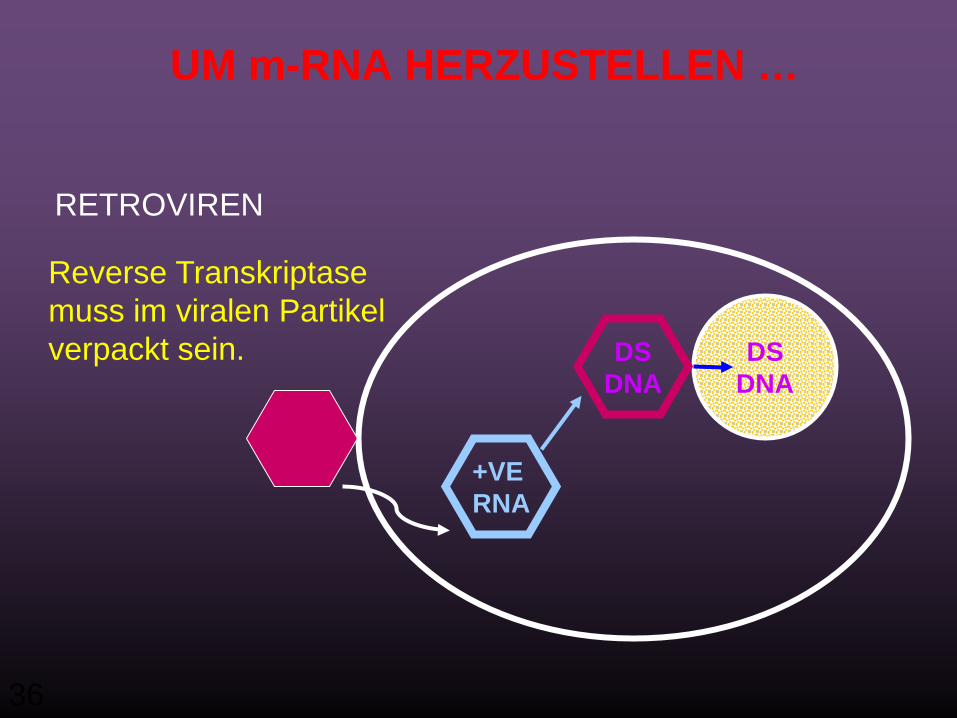
36
UM m-RNA HERZUSTELLEN …
RETROVIREN
DSDNA
+VE RNA
DSDNA
Reverse Transkriptase muss im viralen Partikelverpackt sein.

37
TranskriptionNicht infektiösJaDoppel-strängige RNA
TranskriptionNicht infektiösJaNegativ-strängige RNA
TranslationinfektiösNeinPlus-strängige RNA
Erster Vorgang in der Zelle
Infektiösität der RNA
RNA-abhängige RNA Polymerase (Transkriptase) im Virion
Genom
RNA Viren, die keine DNA-Phase haben
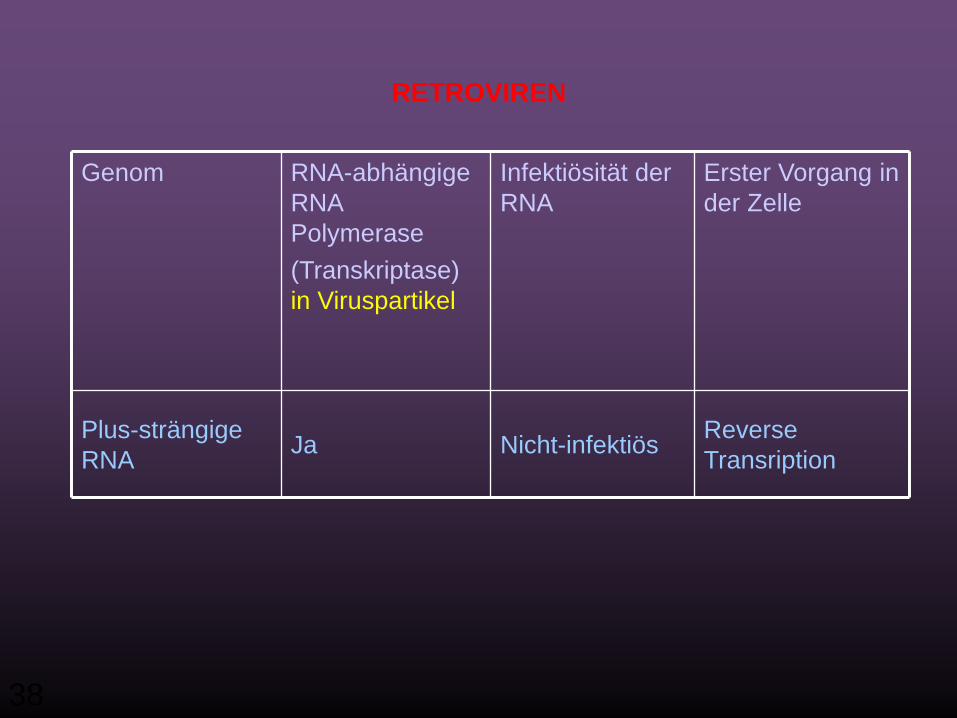
38
Reverse TransriptionNicht-infektiösJaPlus-strängige
RNA
Erster Vorgang in der Zelle
Infektiösität der RNA
RNA-abhängige RNA Polymerase (Transkriptase) in Viruspartikel
Genom
RETROVIREN

39
III) ds RNA Viren:
Beispiel: Reovirus

40
Replikation: ds RNA Virus
• Reovirus• Segmentierte Genome
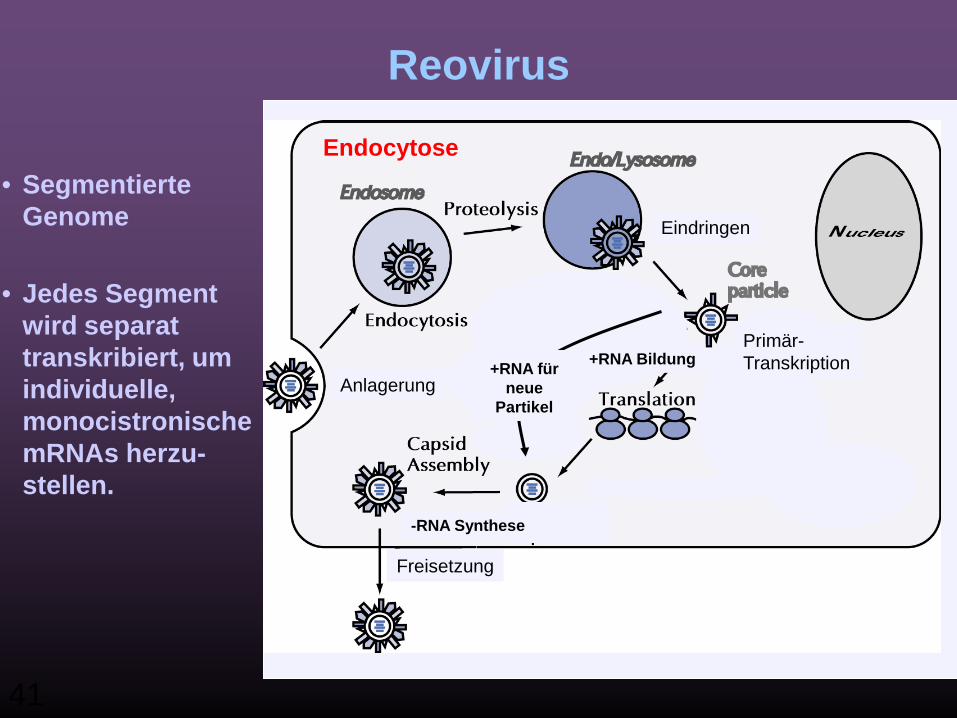
41
Reovirus
• SegmentierteGenome
• Jedes Segment wird separat transkribiert, um individuelle, monocistronische mRNAs herzu-stellen.
+RNA Bildung
-RNA Synthese
+RNA für neue
Partikel
Endocytose
Anlagerung
Eindringen
Freisetzung
Primär-Transkription

42
ISVPs• Reoviren: Zwei Kapside
– Ein äusseres und ein inneres-Kapsid
• Reovirus Virion oder Viruspartikel– infektiöses, intaktes Partikel, (äusseres-Kapsid, inneres-
Kapsid und virales RNA Genom) • Während natürlicher Infektion
– Virus dringt in den Darm ein – Teilweise abgebaut durch intestinale Proteasen– Struktureller Übergang zu einem “infectious subviral particle”
(ISVP).• ISVPs
– Haben äusseren Kapsid-Proteine (sigma-3) verloren– Haben lange Spikes, die aus den Kapsid-Scheitelpunkten
(vertices) herausragen

43
Neue genomische RNA
• die generierten (+) RNA Stränge sind teilweise– NICHT gecapped– verpackt in den neu translatierten viralen Proteinen.
– Es ist nicht bekannt wie das Virus sicherstellt, dass jedes Partikel eine Kopie der 11 verschiedenen (+) strängigen RNAs bekommt.
• verpackte (+) strängige RNA– Wird als Matrize zur Zweitstrang-Synthese der (–) genomischen RNA benutzt
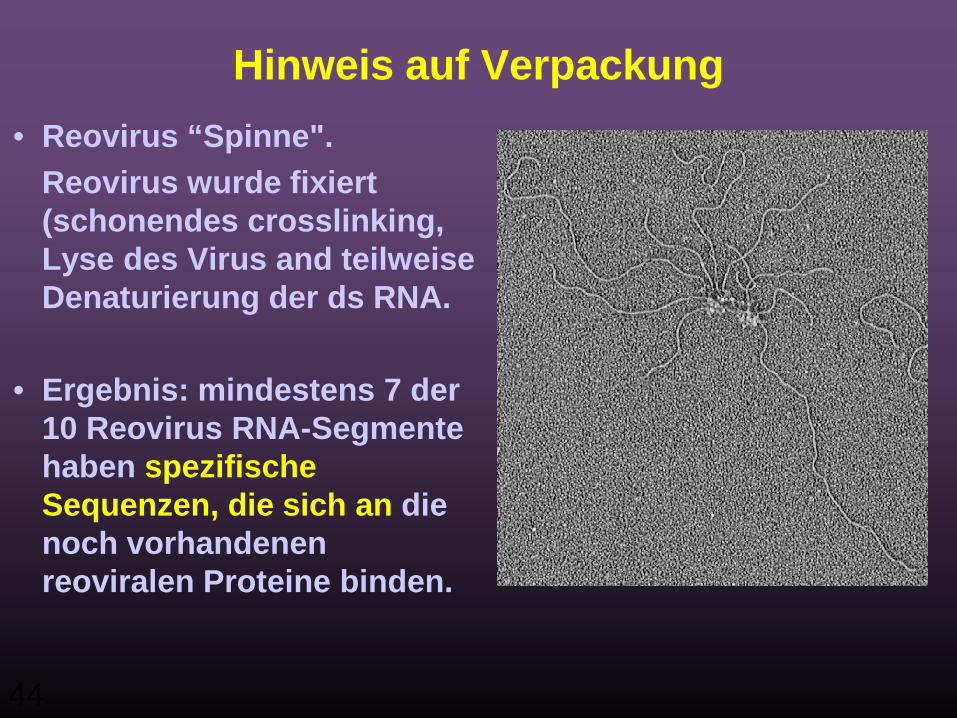
44
Hinweis auf Verpackung• Reovirus “Spinne".
Reovirus wurde fixiert (schonendes crosslinking, Lyse des Virus and teilweise Denaturierung der ds RNA.
• Ergebnis: mindestens 7 der 10 Reovirus RNA-Segmente haben spezifische Sequenzen, die sich an die noch vorhandenen reoviralen Proteine binden.
45
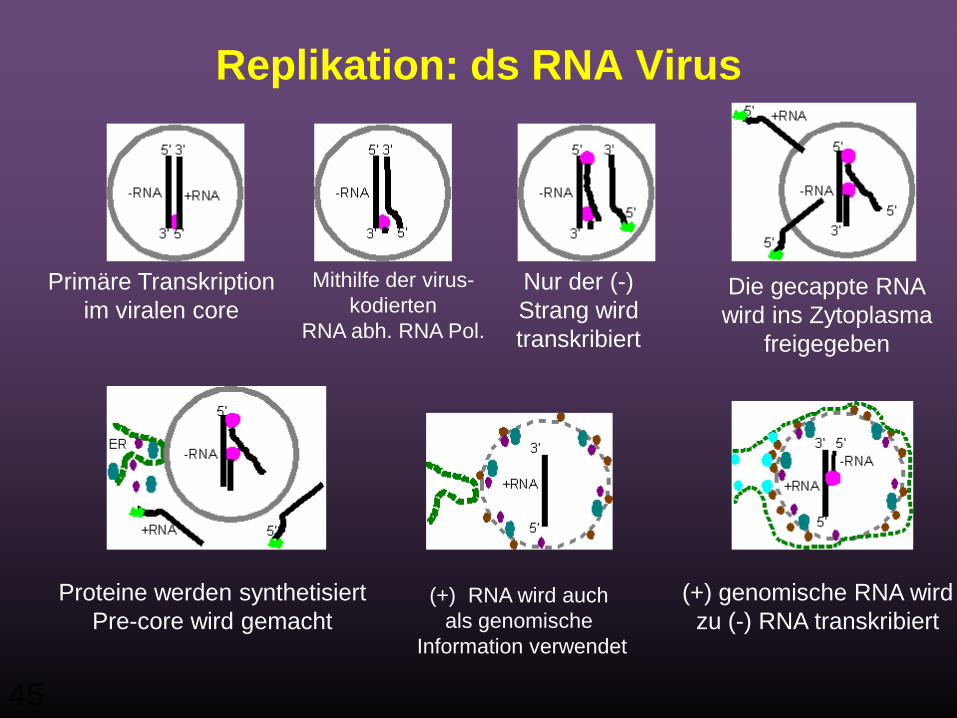
Replikation: ds RNA Virus
Primäre Transkriptionim viralen core
Mithilfe der virus-kodierten
RNA abh. RNA Pol.
Nur der (-) Strang wird transkribiert
Die gecappte RNAwird ins Zytoplasma
freigegeben
Proteine werden synthetisiertPre-core wird gemacht
(+) RNA wird auch als genomische
Information verwendet
(+) genomische RNA wirdzu (-) RNA transkribiert

46
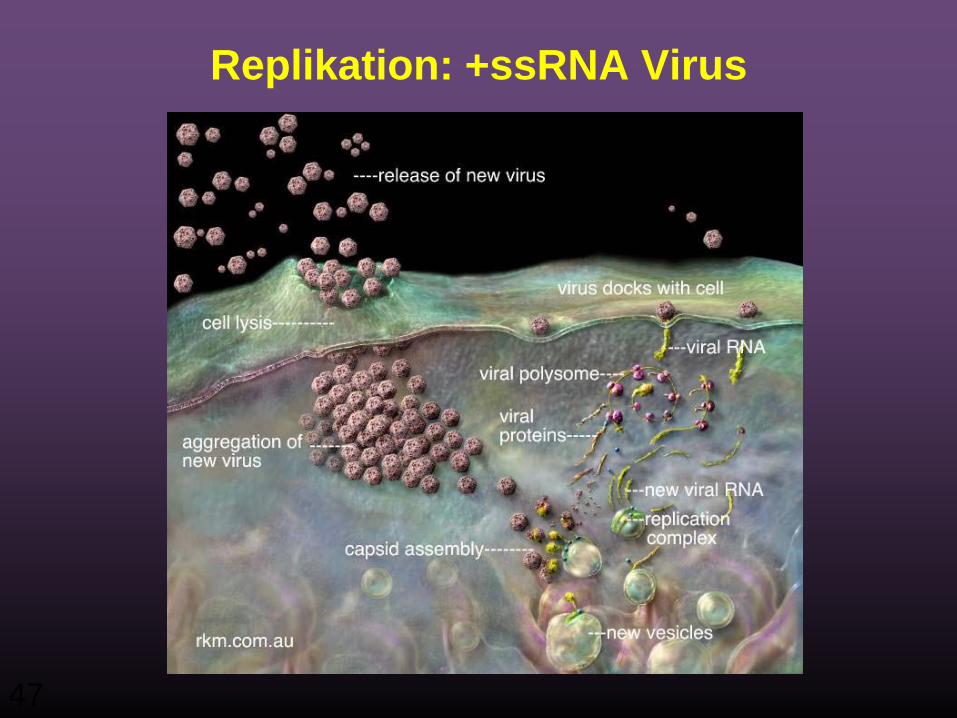
IV) ss+ RNA
Beispiel: Poliovirus
47
Replikation: +ssRNA Virus

48
SYNTHESE VIRALER PROTEINE
• Die im Poliovirus Virion enthaltene RNA funktioniert wie eine RNA, besitzt aber nicht die methylierte Cap-Struktur, wie sie eukaryotische m-RNA hat.
• Stattdessen hat sie ein "ribosome landing pad" (internal ribosome entry site, IRES), die es den Ribosomen erlaubt, trotzdem an sie zu binden.
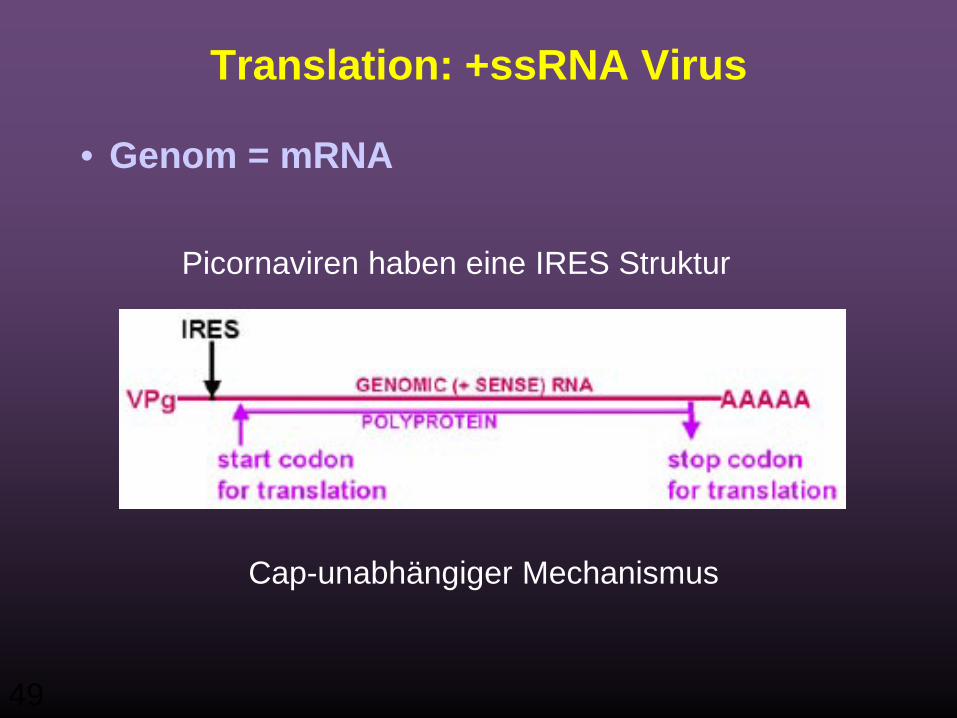
49
Translation: +ssRNA Virus
• Genom = mRNA
Picornaviren haben eine IRES Struktur
Cap-unabhängiger Mechanismus

50
Translation: +ssRNA Virus
• IRES(Internal ribosomal Entry Site)
Picornaviren haben eine IRES Struktur

51
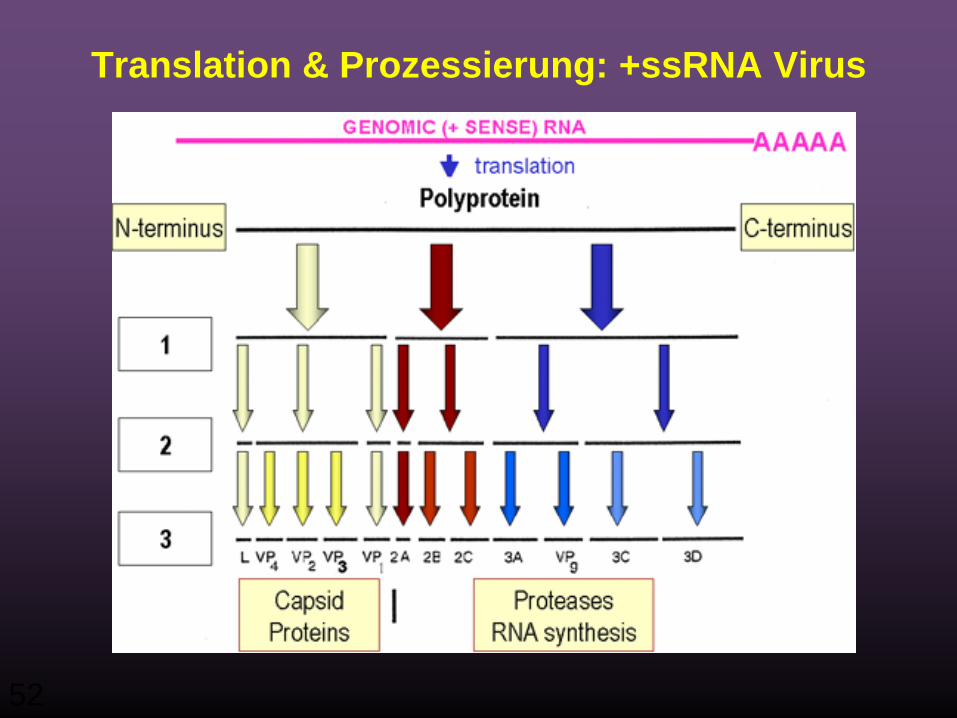
Post-translationale Prozessierung
• Die m-RNA wird zu einem einzigen Polypeptid (Polyprotein) translatiert, das danach geschnitten (prozessiert) wird.
• Dies geschieht co-translationell, das heisst an der entstehenden Polypeptidkette und zwar durch virus-kodierte Proteasen.
Prozessierungs-Produkte sind z.B.:– Eine RNA Polymerase (Replikase)– Strukturkomponenten des Virions– Proteasen
52
Translation & Prozessierung: +ssRNA Virus

53
RNA Replikation
• Jetzt stehen also neu gemachte virale Proteine zur Verfügung, die bei der Replikation helfen .
• Die Virale RNA Polymerase schreibt plus-strängige genomische RNA in komplementäre minus-Strang RNA um:

54
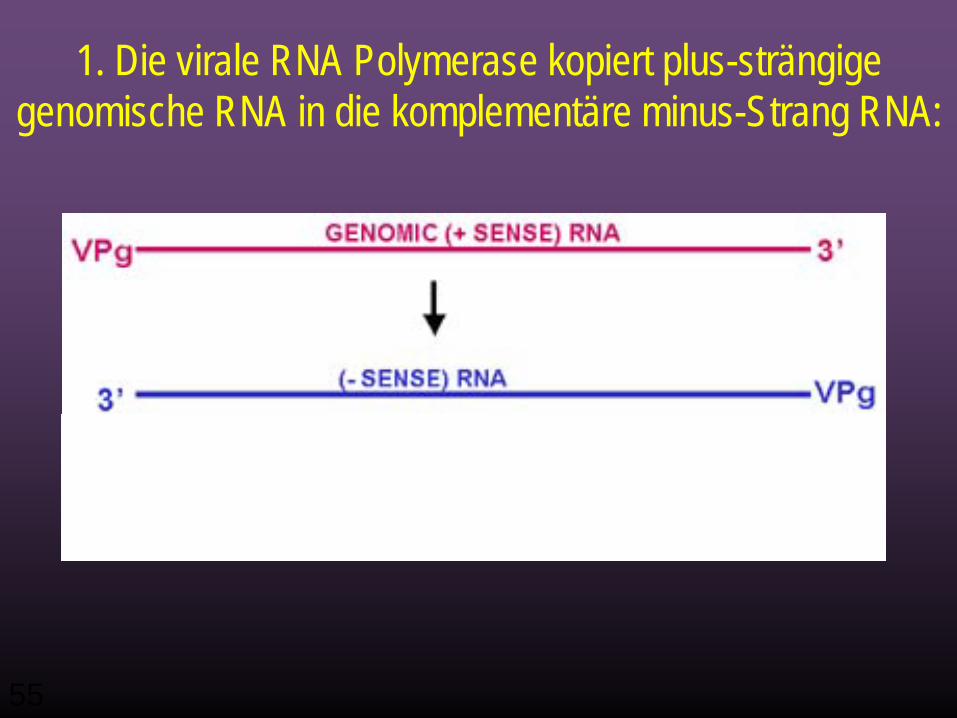
RNA Replikation
• Jetzt stehen also neu gemachte virale Proteine, die bei der Replikation helfen zur Verfügung.
• Die Virale RNA Polymerase schreibt plus-strängige genomische RNA in komplementäre minus-Strang RNA um:
55
1. Die virale RNA Polymerase kopiert plus-strängige genomische RNA in die komplementäre minus-Strang RNA:

56
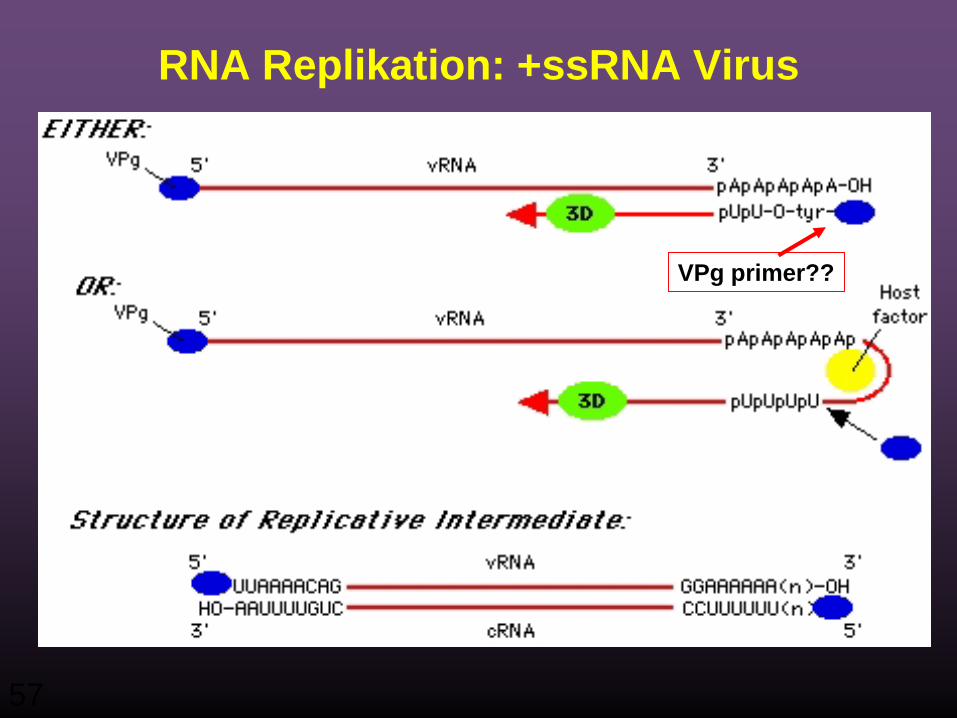
RNA Replikation
• Für diesen Vorgang braucht man:– VPg (oder ein Vorläuferprotein, das VPg enthält)
RNA Polymerase (Replikase)Wirtsproteine
• VPg funktioniert vermutlich als Primer für die RNA Synthese, dies würde erklären, warum es an allen 5' Enden neu synthetisierter RNAs vorhanden ist.
57
RNA Replikation: +ssRNA Virus
VPg primer??

58
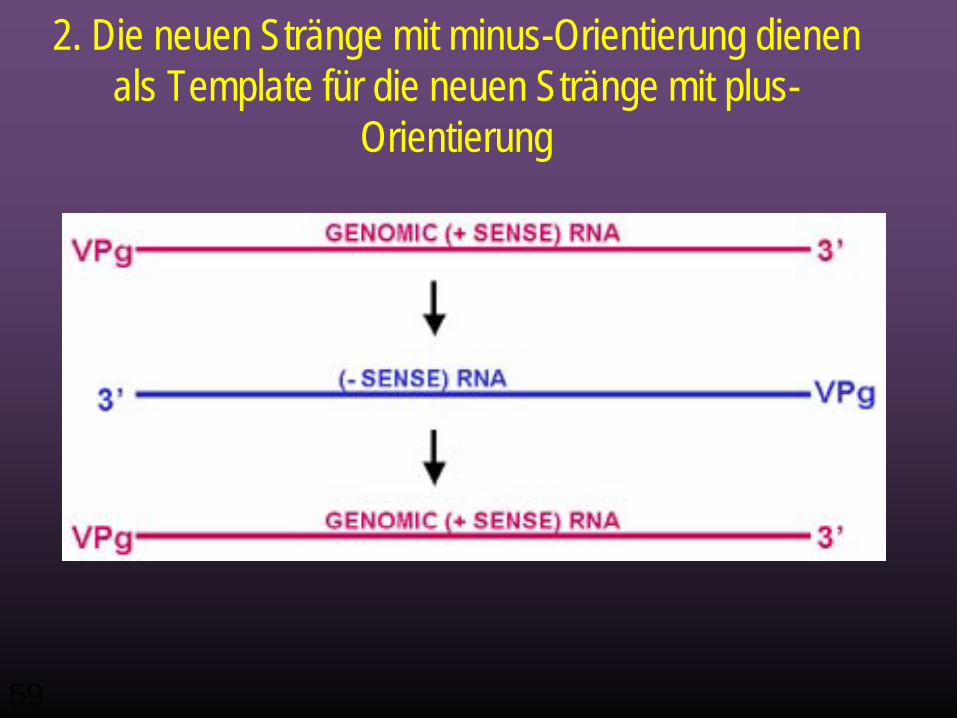
RNA Replikation
• Die neuen Minus-Stränge dienen als Template zur Synthese von neuen Plus-Strängen.
• Wieder werden RNA Replikase und VPg gebraucht. • VPg ist an die 5' Enden der neuen plus-strängigen
RNA gebunden (wie oben dient es vermutlich als Primer).
59
2. Die neuen Stränge mit minus-Orientierung dienen als Template für die neuen Stränge mit plus-
Orientierung

60
REPLIKATION DES GENOMS PLUS-STRÄNGIGER RNA VIREN
Parental (+) Strang
Protein Synthesis
Minus StrandSynthesis (Stage 1)
Plus StrandSynthesis (Stage 2)
Progeny (+) Strands
5
5
5
5
3
3
3
3
(+)
(+)
(+)
(-)(-)(-)
(+)(+)
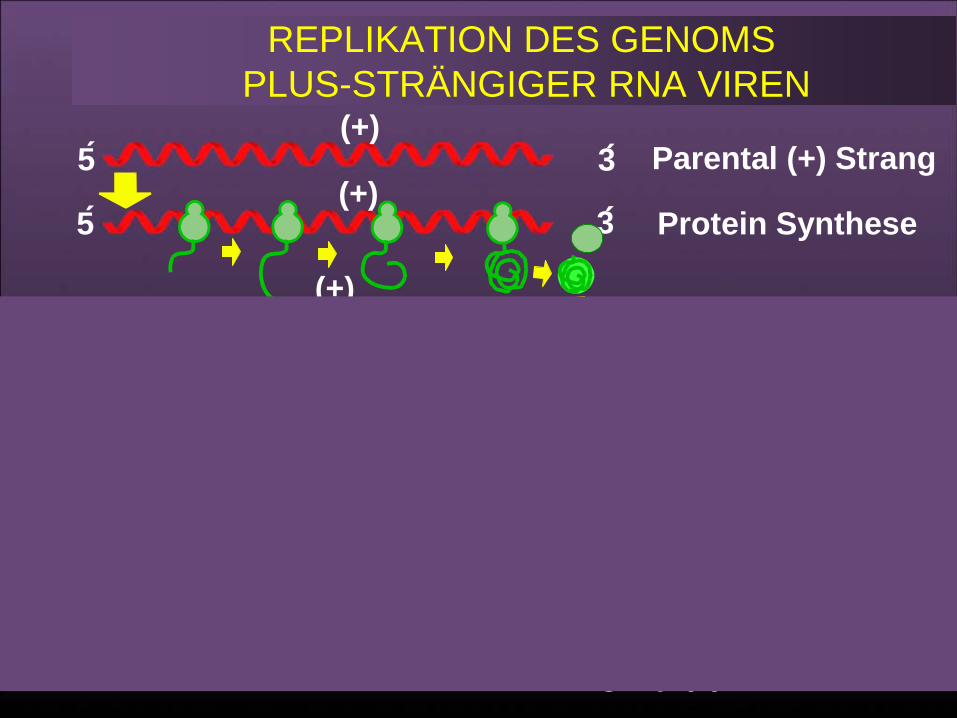
61
REPLIKATION DES GENOMS PLUS-STRÄNGIGER RNA VIREN
Parental (+) Strang
Protein Synthese
Minus StrandSynthesis (Stage 1)
Plus StrandSynthesis (Stage 2)
Progeny (+) Strands
5
5
5
5
3
3
3
3
(+)
(+)
(+)
(-)(-)(-)
(+)(+)
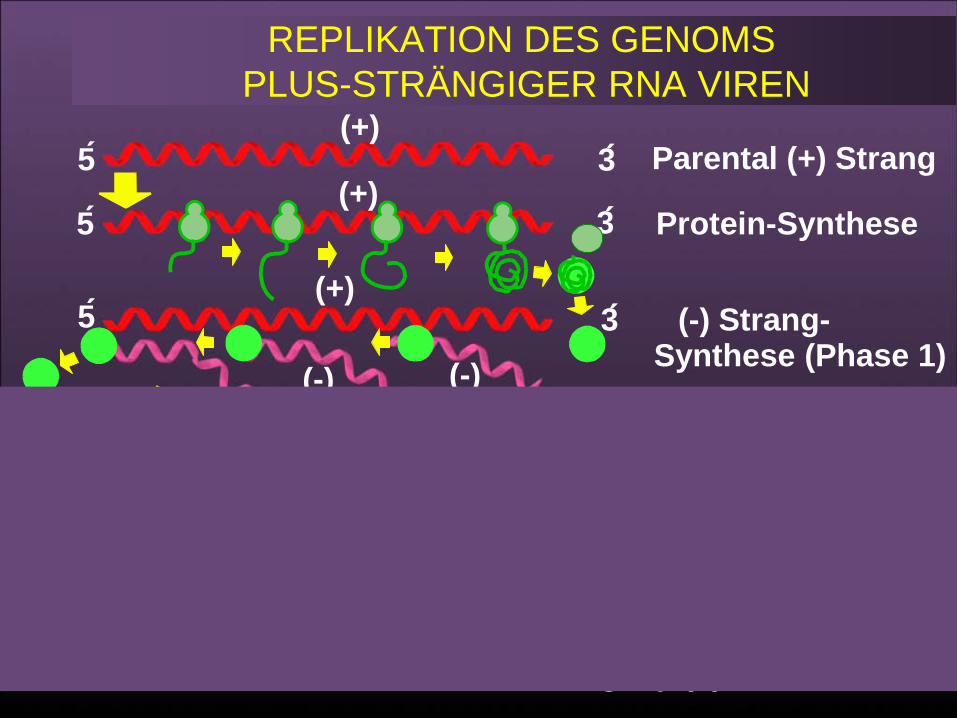
62
REPLIKATION DES GENOMS PLUS-STRÄNGIGER RNA VIREN
Parental (+) Strang
Protein-Synthese
(-) Strang-Synthese (Phase 1)
Plus StrandSynthesis (Stage 2)
Progeny (+) Strands
5
5
5
5
3
3
3
3
(+)
(+)
(+)
(-)(-)(-)
(+)(+)
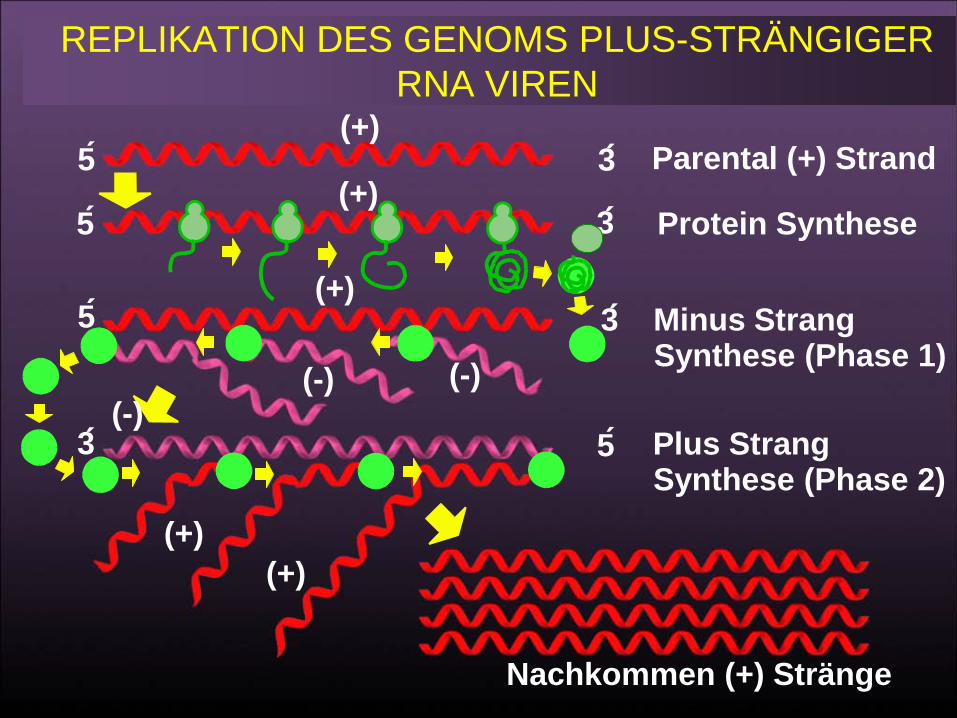
63
REPLIKATION DES GENOMS PLUS-STRÄNGIGER RNA VIREN
Parental (+) Strand
Protein Synthese
Minus StrangSynthese (Phase 1)
Plus StrangSynthese (Phase 2)
Nachkommen (+) Stränge
5
5
5
5
3
3
3
3
(+)
(+)
(+)
(-)(-)(-)
(+)(+)

64
RNA REPLIKATION
• virale RNA Polymerase (Replikase)• Ebenfalls involviert: Wirtszellfaktoren als
akzesorische Proteine • neue (+) Stränge
– verpackt – Templates für weitere Replikation– Templates für weitere Translation (nach
Entfernen von VpG)

65
V) ss-RNA
z.b Rhabdovirus

66
Capping und Polyadenylierung
• Das multifunktionelle Enzyme, das vom sogenannten L Protein kodiert wird, ermöglicht das 5' capping.
• Die Polyadenylierung erfolgt über eine sogenannte "polymerase slippage" an jeder nicht-kodierenden intergenen Sequenz

67
Zusammenfassung
• Die mRNAs sind gecapped, methyliert, und polyadenyliert.
• Da es sich um zytoplasmatische, negative-strängige RNA Viren handelt, befinden sich die Enzyme für die m-RNA Synthese und Modifikation im Virus.

68
(+) Strang RNA Synthese des (–) ssRNA Virus
5’
5’
3’(-)
(-)
(-) (-)
Eltern (-) Strang
PROTEIN SYNTHESIS
PLUS STRANDSYNTHESIS
(Stage 1)
MINUS STRANDSYNTHESIS
(Stage 2)
5’
5’
3’
3’
3’
Progeny (-) Strands
(+)(+)
(+)
(+)
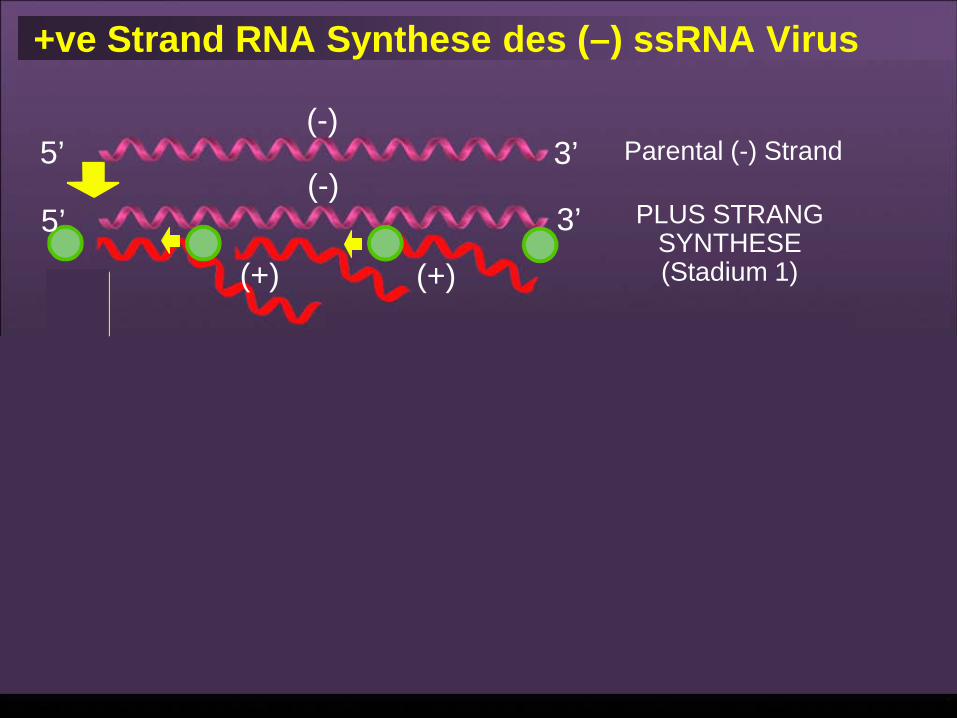
69
+ve Strand RNA Synthese des (–) ssRNA Virus
5’
5’
3’(-)
(-)
(-) (-)
Parental (-) Strand
PROTEIN SYNTHESIS
PLUS STRANGSYNTHESE(Stadium 1)
MINUS STRANDSYNTHESIS
(Stage 2)
5’
5’
3’
3’
3’
Progeny (-) Strands
(+)(+)
(+)
(+)
70
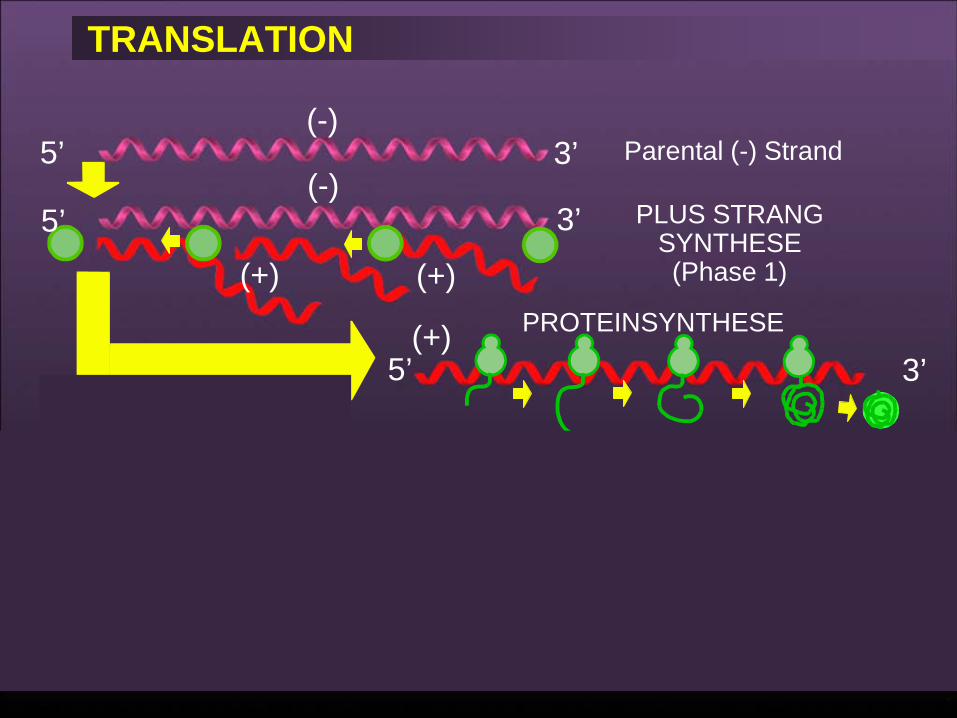
TRANSLATION
5’
5’
3’(-)
(-)
(-) (-)
Parental (-) Strand
PROTEINSYNTHESE
PLUS STRANGSYNTHESE(Phase 1)
MINUS STRANDSYNTHESIS
(Stage 2)
5’
5’
3’
3’
3’
Progeny (-) Strands
(+)(+)
(+)
(+)

71
Translation
• m-RNAs werden auf Wirtszell-Ribosomen translatiert.
• Alle 5 Proteine werden gemacht.
• Es gibt keine Unterscheidung zwischen frühen und späten Funktionen

72
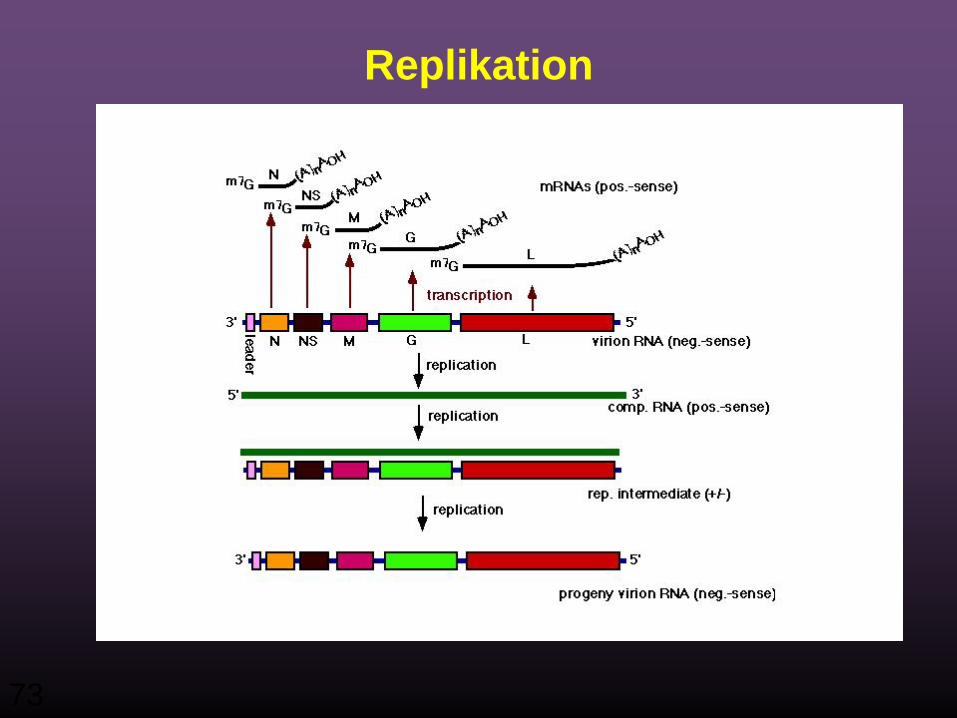
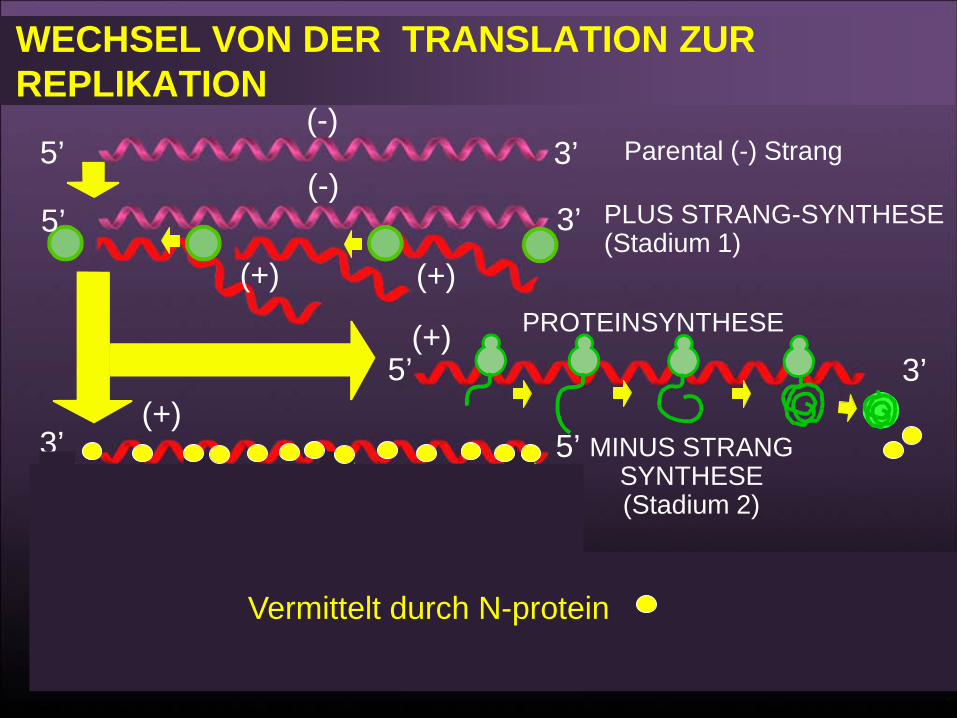
Replikation
• Das Virus beginnt die Replikation nach der Synthese des N Proteins das es ermöglicht, die Polyadenylierungs- und Capping- sowie des intergenischen Transkriptions-Terminations-Signals zu ignorieren,
• Produziert den komplementären positiv-Strang der genomischen RNA.
• Dieser positive-Strang dient als Template, von dem aus die neuen Genome gemacht werden
73
Replikation
74
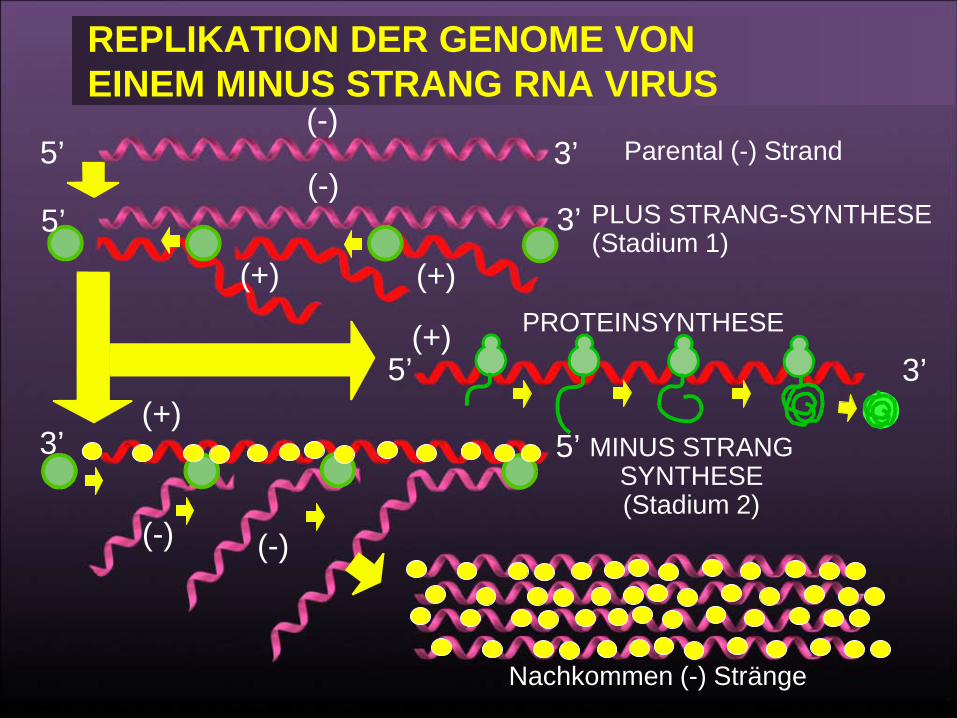
WECHSEL VON DER TRANSLATION ZUR REPLIKATION
5’
5’
3’(-)
(-)
(-) (-)
Parental (-) Strang
PROTEINSYNTHESE
PLUS STRANG-SYNTHESE(Stadium 1)
MINUS STRANGSYNTHESE(Stadium 2)
5’
5’
3’
3’
3’
Progeny (-) Strands
(+)(+)
(+)
(+)
Vermittelt durch N-protein
75
REPLIKATION DER GENOME VONEINEM MINUS STRANG RNA VIRUS
5’
5’
3’(-)
(-)
(-) (-)
Parental (-) Strand
PROTEINSYNTHESE
PLUS STRANG-SYNTHESE(Stadium 1)
MINUS STRANGSYNTHESE(Stadium 2)
5’
5’
3’
3’
3’
Nachkommen (-) Stränge
(+)(+)
(+)
(+)

76
Neu-synthetisierte (–) Strang RNAs
• Neue (-) Strang RNA Genome werden auch mit N-Protein „gecoated“ (= ummantelt, eingehüllt).
• Neue negative Stränge können:– i. Als Vorlage für die Synthese von weiteren volle-Länge (+)
Strängen benutzt werden– ii. Als Vorlage für die Synthese mehrerer mRNAs verwendet
werden. – iii. In Viruspartikel verpackt werden

77
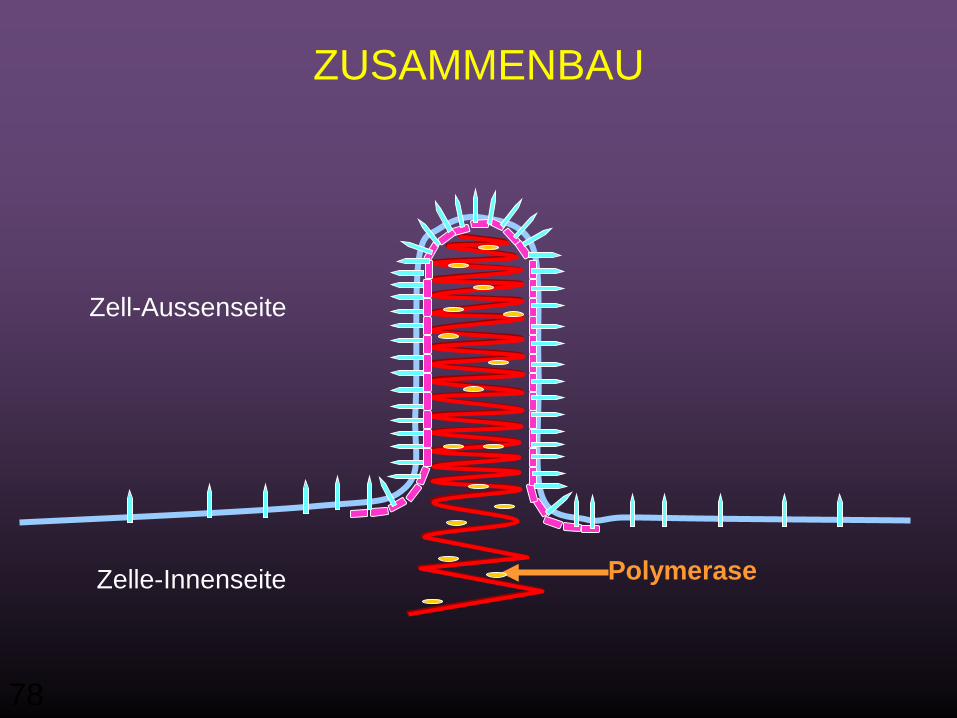
Assembly• Das Virus wird in zwei Phasen wieder zusammengebaut:
• Die erste Phase findet im Zytoplasma statt, wo sich die neuen (+) Strang RNA Genome mit N Proteinen assoziiert haben
• Und nachfolgend mit Polymerase-Proteinen L und NS, das Nukleocapsid zu bilden
• Die Interaktion zwischen dem M Protein und den durch Plasmamembrane-Glycoprotein G-modifizierten Bereichen führt dazu, dass sich das Nucleocapsid zurück in seine kondensierte Form verändert
• In der zweiten Phase, wird das Nukleocapsid an der Plasmamembran behüllt und freigesetzt.
78
PolymeraseZelle-Innenseite
Zell-Aussenseite
ZUSAMMENBAU

79
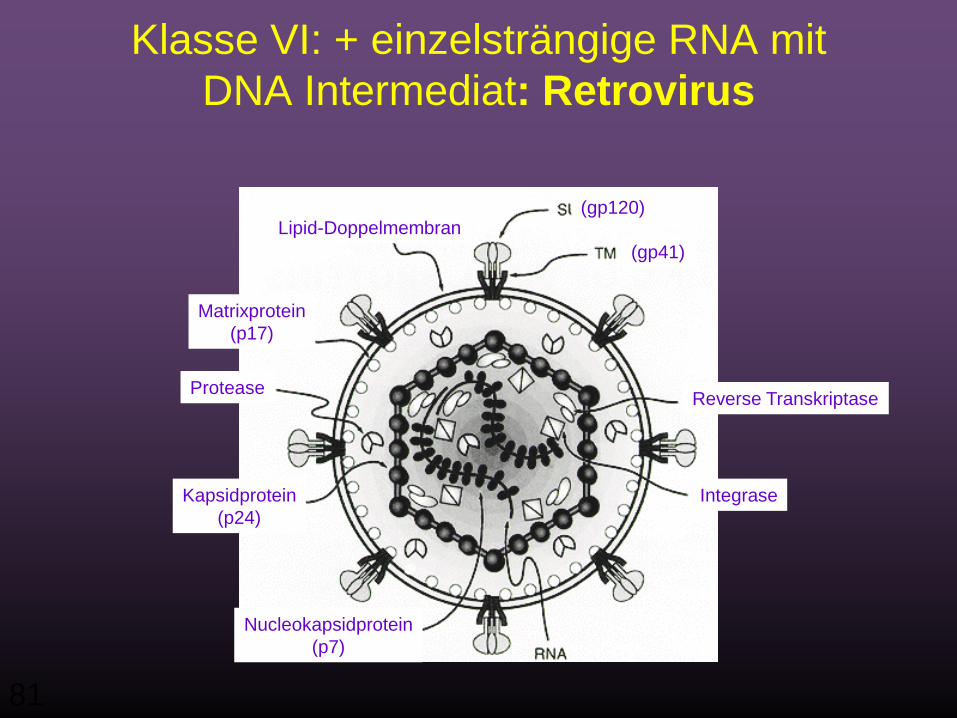
VI) ss+RNA mit DNA als Zwischenstufe
z.B. Retroviren
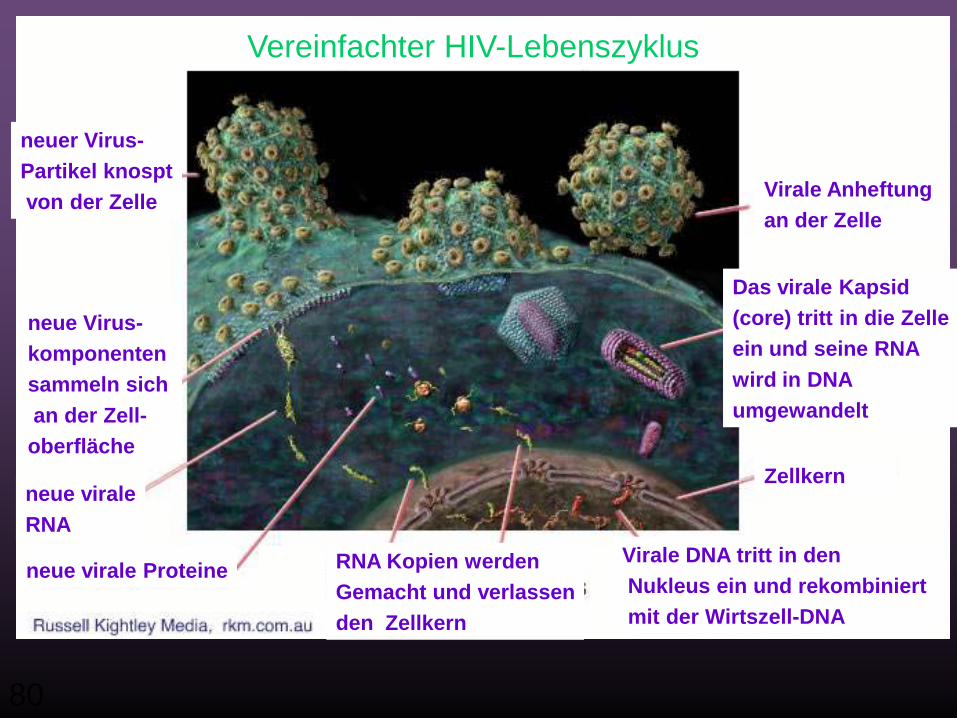
80
Vereinfachter HIV-Lebenszyklus
neuer Virus-Partikel knosptvon der Zelle
neue Virus-komponenten sammeln sichan der Zell-oberfläche
neue viraleRNA
neue virale Proteine
Virale Anheftung an der Zelle
Das virale Kapsid(core) tritt in die Zelleein und seine RNA wird in DNA umgewandelt
Zellkern
Virale DNA tritt in denNukleus ein und rekombiniertmit der Wirtszell-DNA
RNA Kopien werdenGemacht und verlassenden Zellkern
81
Klasse VI: + einzelsträngige RNA mit DNA Intermediat: Retrovirus
Matrixprotein(p17)
Lipid-Doppelmembran
Kapsidprotein(p24)
Nucleokapsidprotein(p7)
Reverse Transkriptase
Integrase
Protease
(gp120)
(gp41)
82
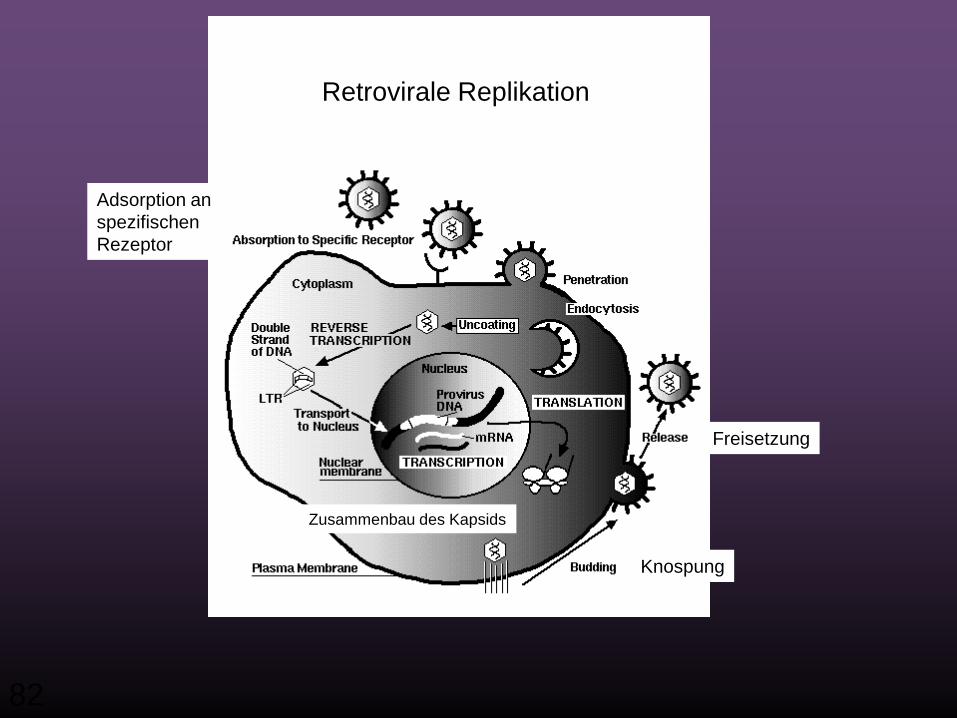
Knospung
Freisetzung
Retrovirale Replikation
Adsorption an spezifischenRezeptor
Zusammenbau des Kapsids

83
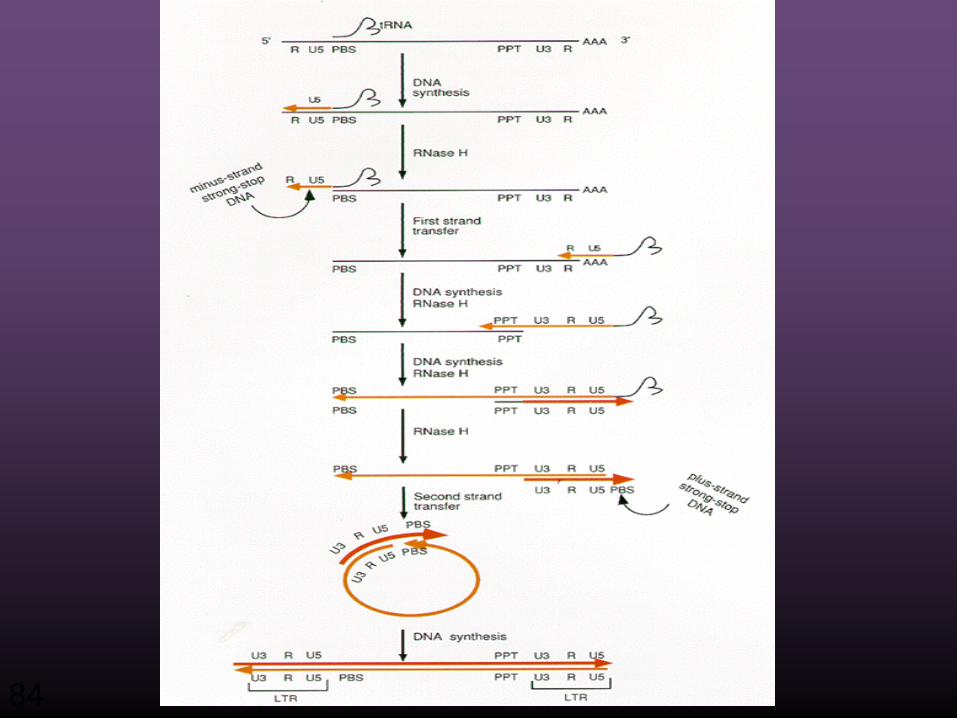
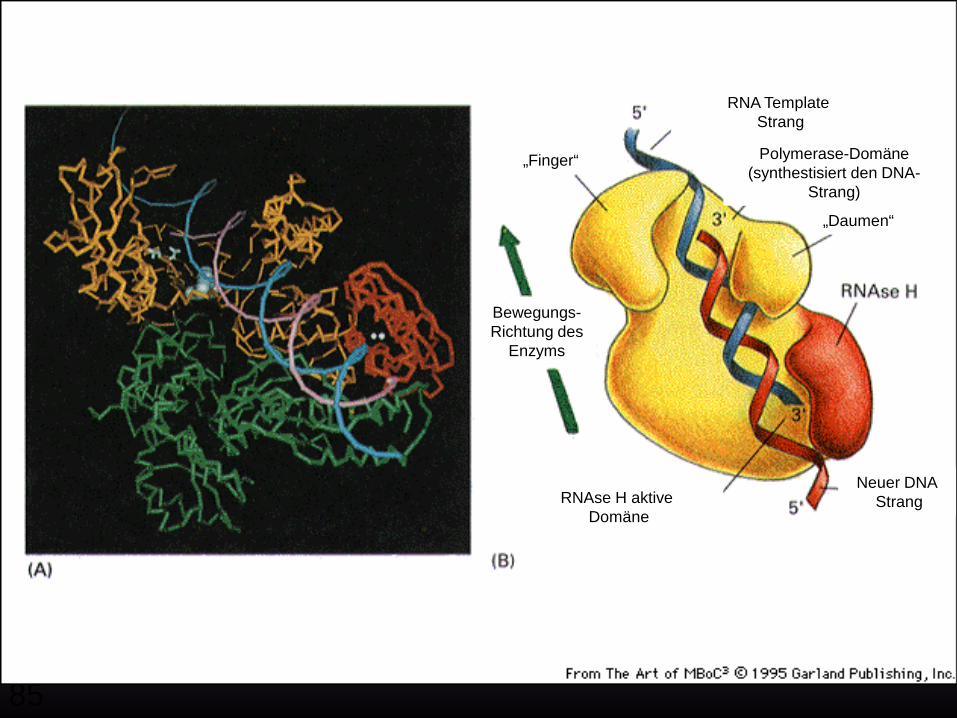
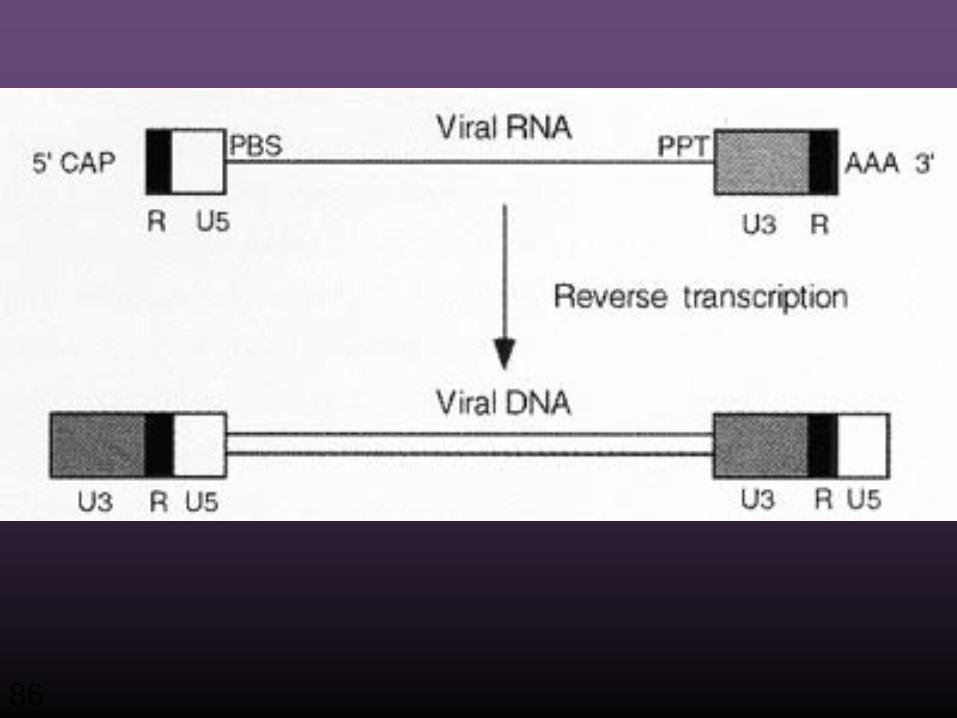
Zentrales Dogma der Biologie
Reverse Transkription
84
85
RNAse H aktiveDomäne
Bewegungs-Richtung des
Enzyms
Polymerase-Domäne (synthestisiert den DNA-
Strang)
„Finger“
„Daumen“
Neuer DNAStrang
RNA TemplateStrang
86
87
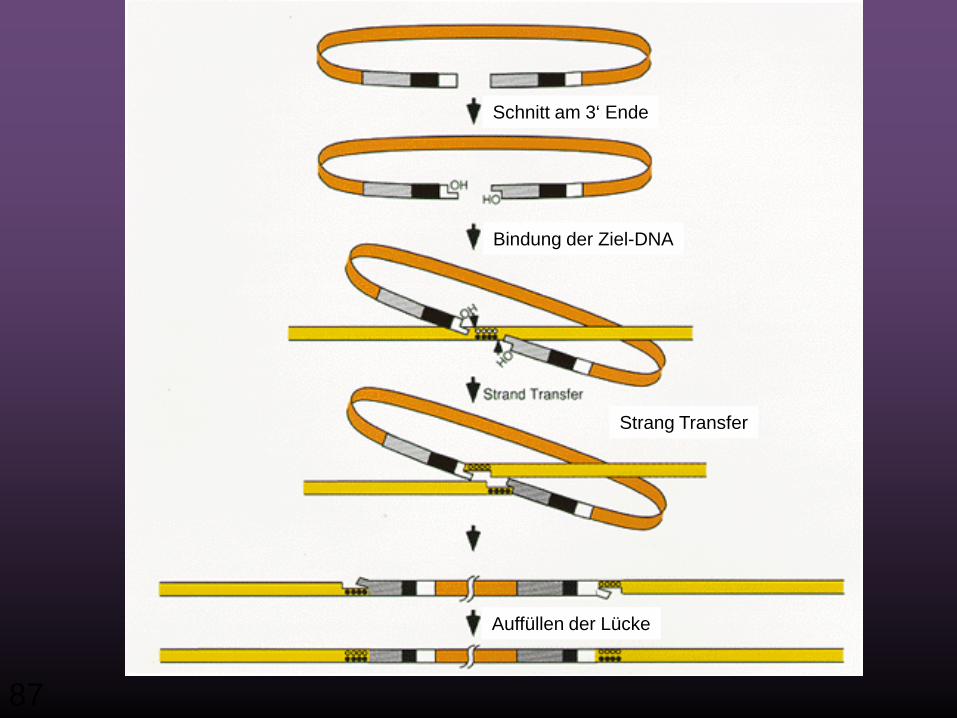
Schnitt am 3‘ Ende
Bindung der Ziel-DNA
Strang Transfer
Auffüllen der Lücke
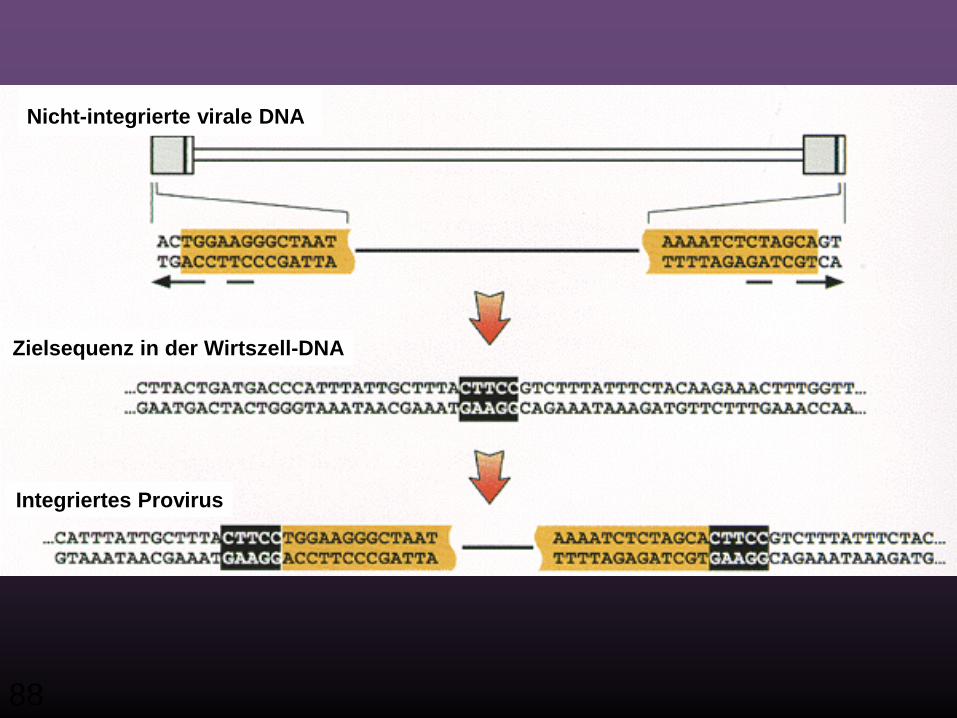
88
Nicht-integrierte virale DNA
Zielsequenz in der Wirtszell-DNA
Integriertes Provirus
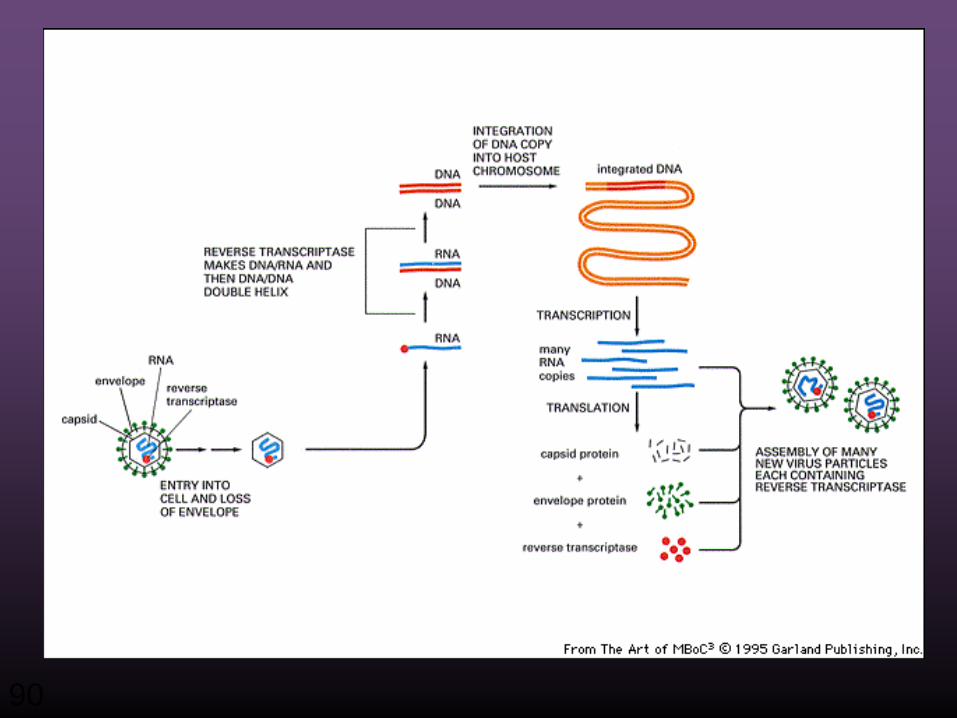
90

91
VII) ds DNA mit RNA als Zwischenstufe
z.B. Hepadnaviren (Hepatitis B Virus),Caulimovirus (Pflanzenvirus)

92
VII) ds DNA mit RNA als Zwischenstufe
1. Twisted Virus-DNADrei Lücken in der DNA an defin. Stellen:eine im (-) strangzwei im (+) strang
2. Supercoiled DNA,„Minichromosom“TranskriptionstemplateExpression durch zweiPromotoren kontrolliert(P) einmal fürgenomische 35S RNA +subgenom. 19S RNA
3. 35S RNATemplate für Translationnund Reverse Transkript.

93
Virus-Rezeptoren
• Virus-Rezeptoren fallen unter verschiedene Klassen, z.B.:
• Immunoglobulin-ähnliche Moleküle• membran-assoziierte Rezeptoren• Transmembran-Transporter & Kanäle
• Allen gemeinsam ist, dass diese Rezeptoren von den Zellen nicht als Rezeptoren für Viren vorgesehen wurden
• Viren haben sich diese Moleküle, die zelluläre Funktionen besitzen als Rezeptoren zu eigen gemacht
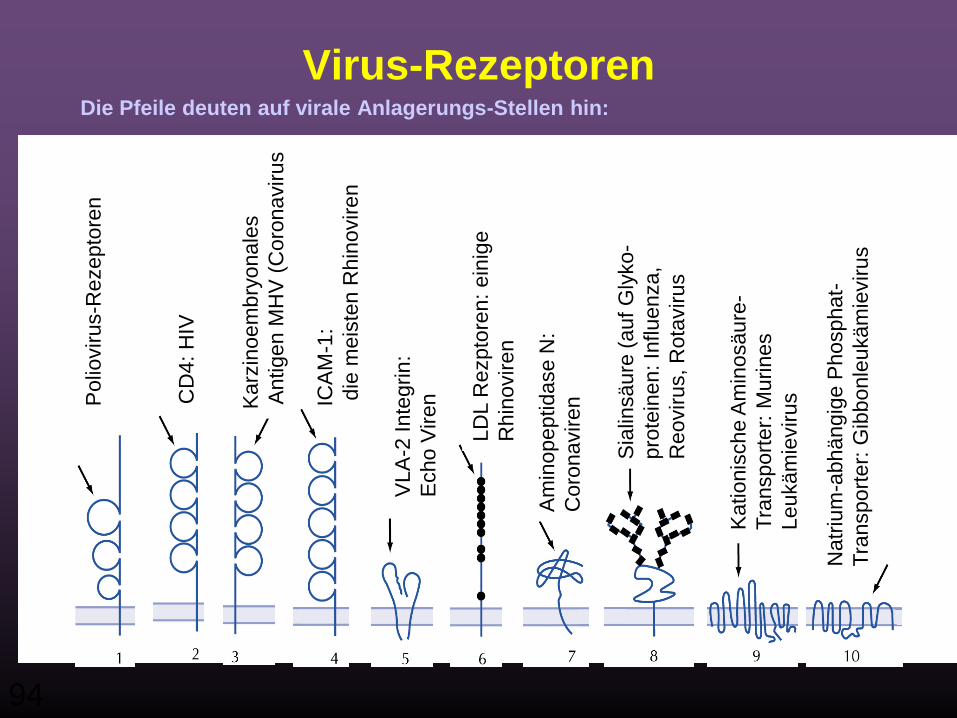
94
Virus-RezeptorenDie Pfeile deuten auf virale Anlagerungs-Stellen hin:
Pol
iovi
rus-
Rez
epto
ren
CD
4: H
IV
ICA
M-1
:di
e m
eist
en R
hino
vire
n
VLA
-2 In
tegr
in:
Ech
o Vi
ren
LDL
Rez
ptor
en: e
inig
e R
hino
vire
n
Kar
zino
embr
yona
les
Ant
igen
MH
V (C
oron
aviru
s
Sia
linsä
ure
(auf
Gly
ko-
prot
eine
n: In
fluen
za,
Reo
viru
s, R
otav
irus
Am
inop
eptid
ase
N:
Cor
onav
iren
Kat
ioni
sche
Am
inos
äure
-Tr
ansp
orte
r: M
urin
es
Leuk
ämie
viru
s
Nat
rium
-abh
ängi
ge P
hosp
hat-
Tran
spor
ter:
Gib
bonl
eukä
mie
viru
s

95
Rezeptoren
• Anwesenheit determiniert das Wirtsspektrum• Anlagerung ist anfänglich oft reversibel• Rezeptoren sind NORMALE zelluläre Proteine• Nur wenige Rezeptoren sind charakterisiert• Sie führen in den meisten Fällen zu
IRREVERSIBLEN ÄNDERUNGEN viraler Proteine
• Co-Rezeptoren können beteiligt sein
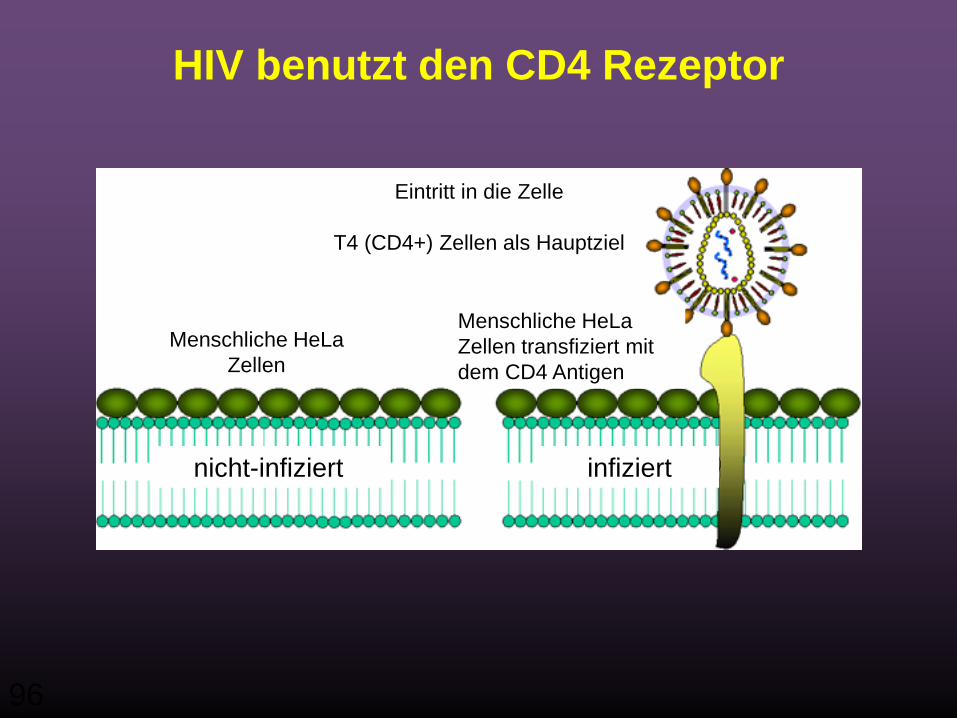
96
HIV benutzt den CD4 Rezeptor
nicht-infiziert infiziert
Menschliche HeLa Zellen
Menschliche HeLa Zellen transfiziert mit dem CD4 Antigen
Eintritt in die Zelle
T4 (CD4+) Zellen als Hauptziel

97
Eindringen
• Das Eindringen in die Zielzelle geschieht normalerweise innerhalb kurzer Zeit nach der Anheftung des Virus an seinen Rezeptor in der Zellmembran
• Anders als bei der Anheftung, ist das Eindringen in die Zielzelle meist energieverbrauchend, d.h. die Zelle muss dafür metabolisch aktiv sein.
• Es gibt hier zwei Hauptmechanismen:
–Direkte Fusion–Endozytose
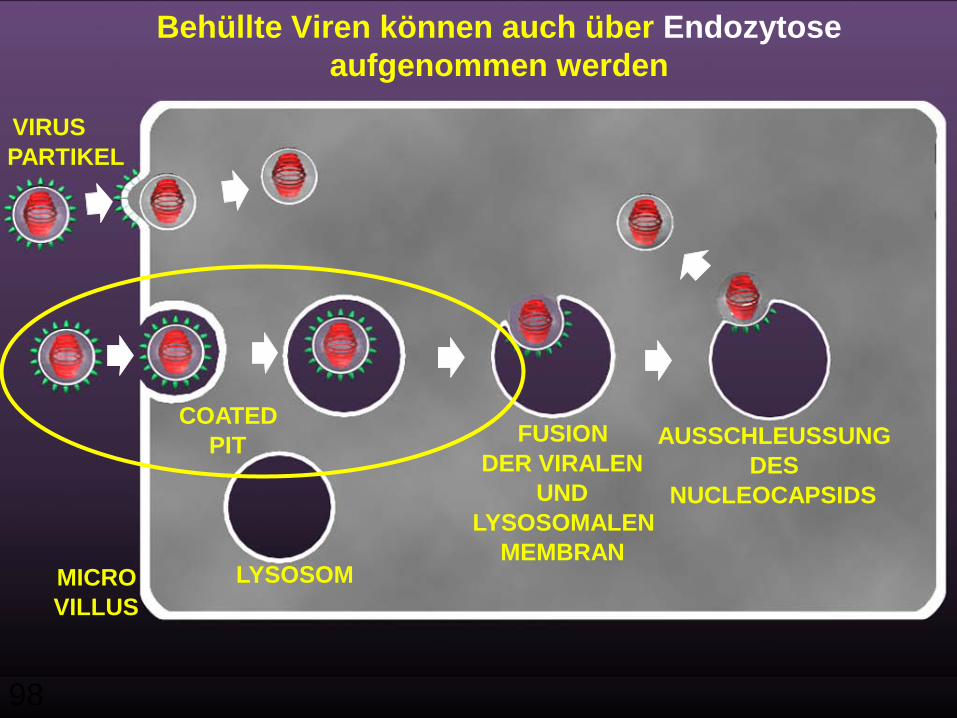
98
VIRUSPARTIKEL
COATEDPIT
LYSOSOM
FUSIONDER VIRALEN
UNDLYSOSOMALEN
MEMBRAN
AUSSCHLEUSSUNGDES
NUCLEOCAPSIDS
MICROVILLUS
Behüllte Viren können auch über Endozytose aufgenommen werden
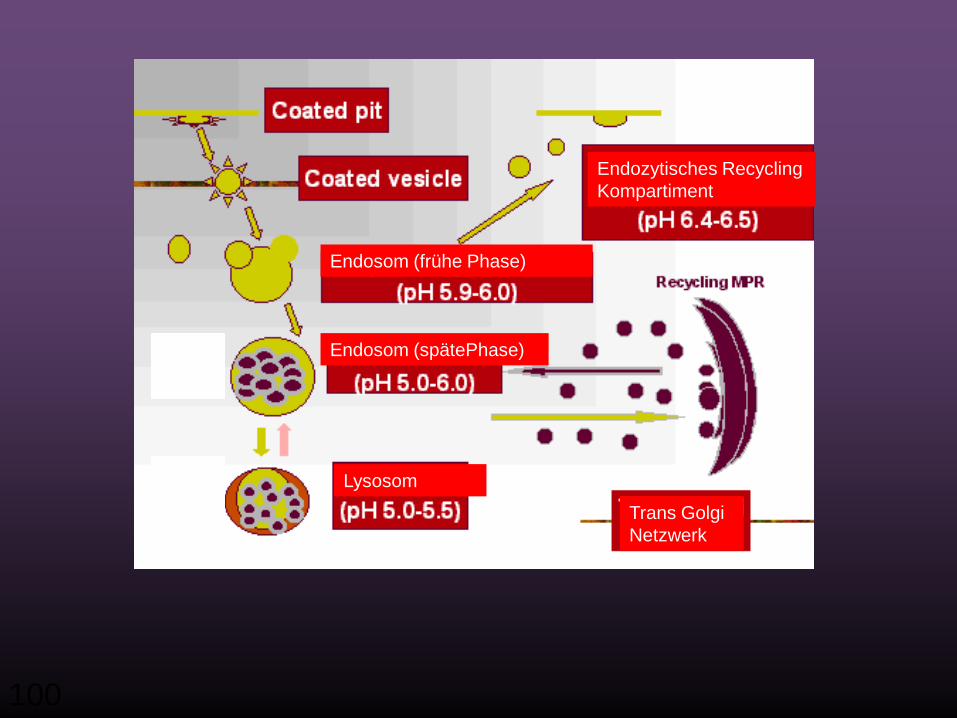
100
Endosom (frühe Phase)
Trans Golgi Netzwerk
Endosom (spätePhase)
Endozytisches RecyclingKompartiment
Lysosom
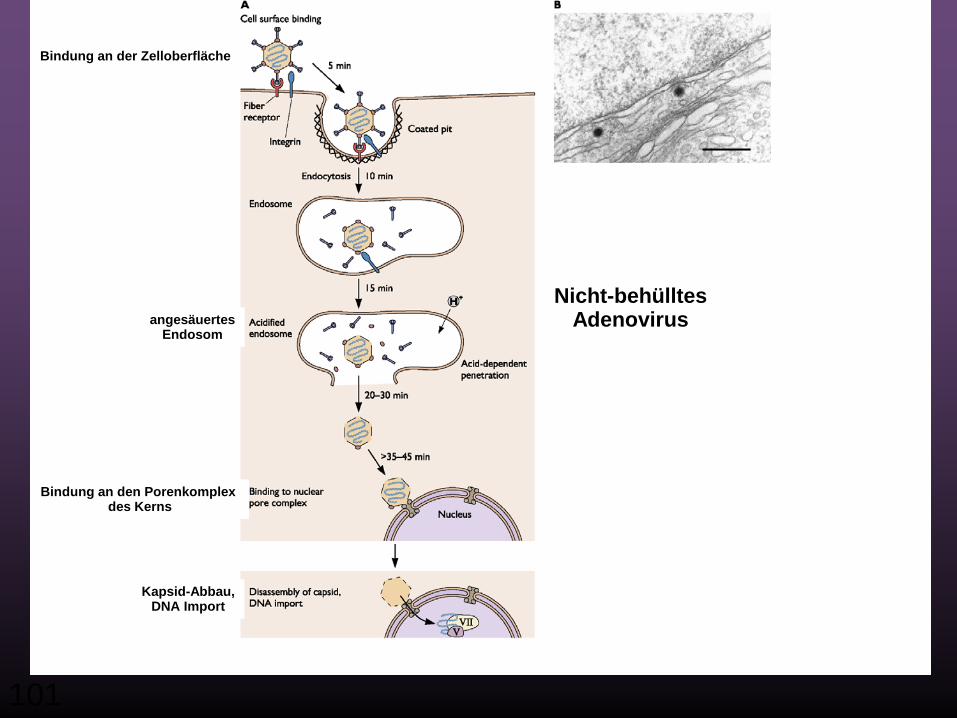
101
Nicht-behülltesAdenovirus
Kapsid-Abbau,DNA Import
angesäuertesEndosom
Bindung an den Porenkomplex des Kerns
Bindung an der Zelloberfläche
102
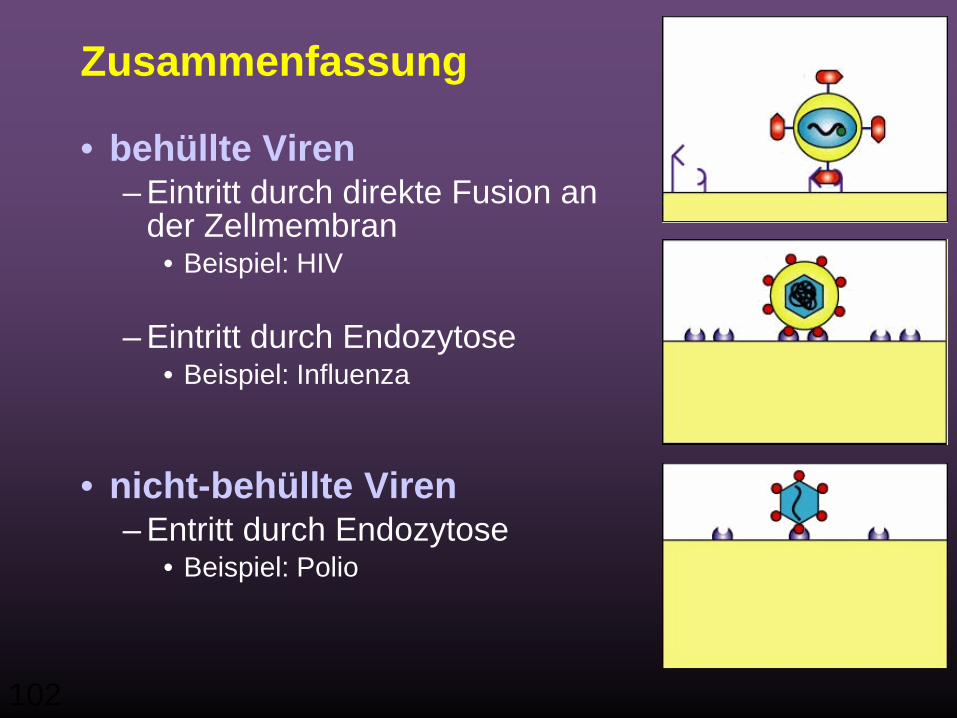
Zusammenfassung
• behüllte Viren– Eintritt durch direkte Fusion an
der Zellmembran• Beispiel: HIV
– Eintritt durch Endozytose• Beispiel: Influenza
• nicht-behüllte Viren– Entritt durch Endozytose
• Beispiel: Polio














![SOFTWARE Open Access ExprEssence - Revealing the essence ... · (DNA methylation, DNA/Histone acetylation) on a large scale has become possible only recently [1,2]. Measuring transcription](https://img.pdfslide.org/doc/110x75/5f8aa6bc776cef3194554896/software-open-access-expressence-revealing-the-essence-dna-methylation-dnahistone.jpg)