Embed Size (px)
Citation preview

Ultrastruktur und Zellwandbildungvon Acanthamoeba sp.
Von
HEINZ BAUERLaboratorium für Elektronenmikroskopie, ETH, Zürich
Inhalt
I. Einleitung 173
H. Material und Methoden 174
1. Objekt 1742. Fixation und Einbettung 1753. Gefrierätzung 1754. Saure Phosphatase 1755. Mazeration und Negativkontrastierung 1756. Mikroveraschung 1757. Hydroxyprolin-Nachweis 1768. Autoradiographie 176
III. Allgemeine Ultrastruktur 176
IV. Zellwandbildung (Encystierung) 178
1. Ablauf der Encystierung 1782. Zellwandproteine 1803. Zellwandeinschlüsse 1814. Diskussion 182
V. Zusammenfassung 183
VI. Literatur 184
VII. Bildlegende 186
I. Einleitung
Acanthamoeba sp. besitzt eine interessante systematische Stellung zwischentierischen und pflanzlichen Mikroorganismen. Ihre heterotrophe Lebensweise sprichtdeutlich zugunsten eines tierischen Organismus, wogegen die Bildung einer zellulose-haltigen Zellwand (TOMLINSON und JOHNES 1962) als typisches pflanzliches Merkmalgedeutet werden darf. In der Natur kennt man zwei, morphologisch und auch

174 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1967
physiologisch deutlich unterscheid bare Formen dieser wärmeliebenden Schlamm-amöbe. Bei genügendem Nährstoffangebot liegt sie als nackter, teilungsfähigerProtoplast vor. Die Fortbewegung erfolgt mit Hilfe von Pseudopodien (amöboideBewegung). Unter ungünstigen Lebensbedingungen (Kohlenstoff- und Stickstoff-mangel; relativ tiefe Temperatur) kapselt sich die Amöbe ein und bildet innerhalbvon 12-18 Stunden eine widerstandsfähige Überdauerungsspore, die Cyste.
Als Versuchsobjekt für die vorliegende Arbeit wurde Acanthamoeba sp. haupt-sächlich aus folgenden Gründen gewählt:
1. Sterile Kulturen lassen sich ohne grossen Aufwand in einem künstlichen,Glucose, Proteose-Pepton, Vitamin B12 und Mineralsalze enthaltenden Nährmedium(KLEIN und NEFF 1960) innerhalb von zwei bis drei Wochen routinemässig im Labora-torium ziehen.
2. Die Zellwandbildung kann durch Übertragen der Protoplasten in eine blossMineralsalze enthaltende Encystierungslösung eingeleitet und jederzeit wieder ab-gestoppt werden. Dadurch erzielt man mehr oder weniger synchrone Kulturen. Nacheiniger Kenntnis des Objektes wurden die Cysten jedoch meist durch eine natürlicheNährstoffverarmung des Kulturmediums erzeugt. Diese Methode birgt den Vorteil,dass für morphologische Studien praktisch sämtliche Entwicklungsstadien neben-einander vorkommen.
3. Durch die Untersuchung von NEFF et al. (1964) ist die chemische Zusammen-setzung der Cystenwand bekannt. In isolierten Wandfragmenten konnten sie 6Lipide, einen erstaunlich grossen Anteil von 33 % Protein sowie 35 % Alkali unlöslichePolysaccharide nachweisen. TOMLINSON und JOHNES (1962) konnten letztere mitHilfe von Infrarot-Absorptionsspektren als Cellulose identifizieren. Bemerkenswertsind zudem 7-8 % anorganisches Material, wovon allein 2 % auf Phosphor entfallen.
4. Eine umfassende Ultrastrukturuntersuchung (einschliesslich einfacher cyto-chemischer Methoden), unter besonderer Berücksichtigung der Cystenwandbildungund der damit verbundenen Biogenese fibrillärer Celluloseverbände, dürfte unsereKenntnisse über den komplizierten Feinbau der Mikroorganismen und derenkomplexe Zellwandstruktur erweitern.
H. Material und Methoden
1. Objekt
Die Züchtung steriler Kulturen von Acanthamoeba sp. erfolgte bei 27 °C in einemvon KLEIN und NEFF (1960) beschriebenen flüssigen Kulturmedium folgender Zu-sammensetzung: 10,0 g Proteose-Pepton, 10,0 g Glucose, 20,0 mg Mg++, 2,0 mg Ca++1,0 μg Vitamin B12, 0,1 ml 0,01 m Eisencitrat und 20,0 ml 0,1 m Phosphatpuffer indest. Wasser ad 1000 ml (pH 5,5). Unter diesen Kulturbedingungen erhält man einemaximale Dichte von ca. 3 . 10 6 Zellen/ccm Nährlösung. Die Encystierung erfolgte ambesten bei pH 7,0 in einem vitaminfreien Medium ohne Stickstoff- und Kohlenstoff-quelle. Die nackten oder encystierten Amöben liessen sich leicht aus dem Kultur-resp. Encystierungsmedium durch Zentrifugation bei 600 g gewinnen.

Jahrgang 112 H. BAUER. Ultrastruktur und Zellwandbildung von Acanthamoeba sp. 175
2. Fixation und Einbettung
Die Fixation des Materials erfolgte entweder mit 2% KMnO 4 in Brunnenwasserwährend 1-18 Stunden oder nach Vorfixation mit Glutaraldehyd (SABATINI et al. 1963)mit 1 % 0s04 nach PALADE (1952). In weiteren Versuchen wurde 8 Min. in einem Ge-misch Von 9,5 % 0S04 und 4,5% K2Cr207 nach BHOWMICK und WOHLFAHRT-BOTTER-
MANN (1965) fixiert. Die fixierten Kulturen wurden während der Entwässerung aufder Stufe von 75 % Aceton mit 1 % UO 2-Acetat über Nacht vorkontrastiert. ZurEinbettung diente Epon. Die Dünnschnitte wurden mittels Glasmessern angefertigtund meistens mit Blei nach KARNOVSKY (1961) oder REYNOLDS (1963) nachkontra-stiert.
3. Gefrierätzung
Für die Gefrierätzung (MooR et al. 1961) verwendete Amöbenkulturen wurdenvorerst 2-6 Std. mit 15 % Glycerin imbibiert; dies erfolgte direkt im Nähr- resp.Encystierungsmedium. Dabei erwies sich, dass dieser Mikroorganismus eine 6stündigeGlycerinimbibition, welche vor allem die Eiskristallbildung im Cytoplasma verhindert,schadlos übeIsteht. Die vorbehandelten Proben wurden in Freon 12 oder Freon 22eingefroren und in flüssigem Stickstoff aufbewahrt. Einzelheiten bezüglich Gefrier-vorgang biologischer Objekte, der Gefrierätztechnik und deren Anwendung sindfolgenden Arbeiten zu entnehmen: MOOR et al. (1961), MOOR und MüHLETHALER
(1963), MooR (1964), MooR et al . (1964), JOST (1965).
4. Saure Phosphatase
Bei 4°C, in 6,25 % Glutaraldehyd (TRIS-Puffer; pH 7,2) vorfixierte Kulturenwurden 20 Min. in kaltem Tris-Puffer (pH 7,2; 0,33 m Saccharose) gewaschen (MILLER
und PALADE 1964) und anschliessend 15-45 Min. in einem Gomori-Medium (GoMORI(1952) mit Na-β-Glycerophosphat inkubiert. Nach der Inkubation musste dasMaterial dreimal, abwechlungsweise in kaltem 0,05 m, 7,5 % Saccharose und 4 % For-mol enthaltenden Na-Acetat-Puffer (pH 5,0) und 2% Essigsäure gewaschen werden.
5. Mazeration und Negativkontrastierung
Zur Gewinnung des Celluloseanteils der Zellwand wurden encystierte Amöben3mal 10 Min. in einem Gemisch von 30 % H2O2 und Eisessig (1:1) mazeriert. Nachkurzer Ultraschallbehandlung und Imprägnierung mit Na-Phosphorwolframsäureliessen sich die einzelnen Cellulosefibrillen elektronenoptisch abbilden.
6. Mikroveraschung
Die Veraschung fixierter Dünnschnitte (THOMAS 1964) erfolgte auf einem zwischenzwei Elektroden eingespannten Molybdänblech während 10-40 Minuten, bei 500 bis600°C in einem Balzers Vakuumbeschatter. Als Objektträger eigneten sich Netz-blenden aus rostfreiem und hitzebeständigem Stahl, die mit einer 100-200 A dickenKohlefolie versehen waren.

176 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1967
7. Hydroxyprolin-Nachweis
Die abzentrifugierten Zellen wurden in 6 n HC1 suspendiert, in eine Glasampulleeingeschmolzen und 18 Stunden bei 108 °C hydrolisiert. Das mit Aktivkohle ge-reinigte Hydrolysat dickte man anschliessend im Rotationsverdampfer zu einem öligenSirup ein. Zur TrennIng der Aminosäuren und der Zucker diente eine Dowex-50-Säule.Der mit 2 n NH4OH erhaltene Auszug der freien Aminosäuren konnte qualitativauf Hydroxyprolin geprüft werden. In Gegenwart von Prolin und Hydroxyprolinerhält man mit 0,2 % Isatin, gelöst in Aceton, nach 2 Minuten bei 105°C eine Blau-färbung, welche sich bei Zugabe von 1 % Ehrlichs Reagens (p-Demethylamino-benzaldehyd) in Aceton-HCI (9:l) klrschrot nachfärbt und spezifisch Hydroxyprolinanzeigt (JEPSON und SMITH 1953).
8. Autoradiographie
20 Tage alte Kulturen wurden während der Encystierung mit H3-Prolin (spezifischeAktivität 720 mc/mM; ca. 20 μc/m1 Nährlösung) gefüttert. In verschiedenen Stadiender Cystenbildung beobachtete man den Einbau der tritiierten Aminosäure in dieZellwandproteine. Das Auftragen einer Einkornschicht (CARO und VAN TUBERGEN
1962) der Ilford -L-4-Emulsion auf die mit Ultradünnschnitten besetzten Netzblendenerfolgte nach der Drahtschlingenmethode von HAASE und JUNG (1964). Die Präparatewurden 4 Wochen exponiert. Die Entwicklung erfolgte entweder in Microdol-X(Eastman Kodak) oder einem von LETTRÉ und PAWELETZ (1966) beschriebenen Ent-wickler folgender Zusammensetzung: 1,5 g Ascorbinsäure, 0,25 g Phenidon (Geigy,Basel), 0,6 g Kaliumbromid, 1,3 g Kaliumcarbonat, 20 g Natriumsulfit (kann weg-gelassen werden) ad 100 ml H2O dest. Gegenüber Microdol-X hat dieser Entwicklerden Vorteil, dass kompakte Körner entstehen. Das Wachstum erfolgt in konzentrischenSchalen um das Primärkorn. Dadurch wird die Zuordnung der Körner zu bestimmtenZellstrukturen erleichtert. Die Fixation erfolgte stets in Kodak-Metafix während 5Minuten.
III. Allgemeine Ultrastruktur
Sowohl nach chemischer als auch nach physikalischer Fixierung kann elektronen-mikroskopisch eine starke Strukturierung des Protoplasten festgestellt werden(Abb. 1, 2). Die meist kugelige Amöbe (bei Züchtung in einem flüssigen Nährmediumunterbleibt oft die Pseudopodienbildung), wird nach aussen durch das dicht mit90-120 A messenden Partikeln besetzte Plasmalemma begrenzt (Abb. 11).
Die Mitochondrien lassen sich klar dem tubulären Typus zuordnen (Abb. 1, 4).Von KLEIN und NEFF (1960) beschriebene anorganische Einschlüsse (Abb. 3) könnenebenfalls nachgewiesen werden. Die meist runden, von einer Membran begrenzten,chemisch noch nicht identifizierten Körper zeigen einen dunklen Kern und einenelektronentransparenten Hof (Abb. 3). Sie sind stets exzentrisch gelagert.
Das Partikelmuster der Vakuolenmembran (Abb. 5) zeigt unregelmässig verteilte,140 A messende Teilchen und dazwischen partikelfreie, glatte Partien. Physiologischmuss den Vakuolen von Acanthamoeba sp. vor allem enzymatische Tätigkeit zuge-

Jahrgang 112 H. BAUER. Ultrastruktur und Zellwandbildung von Acanthamoeba sp. 177
schrieben werden. Bei der permanenten Pinocytose von Amoeba proteus findetWOHLFARTH-BOTTERMANN (1966), dass vom Pinocytosekanal abgeschnürte Primär-vakuolen in mehrere Sekundärvakuolen zerfallen. Letztere verschmelzen mit deneigentlichen Nahrungsvakuolen und geben ihren Inhalt in diese ab. Ein vergleichbarerVorgang scheint bei Acanthamoeba sp. stattzufinden. Aufgenommene Nahrungs-teilchen oder zelleigene, nicht mehr funktionstüchtige, zu eliminierende Strukturenwerden wahrscheinlich zuerst von kleineren Vakuolen aufgenommen. Diese fusio-nieren später mit der grossen zentralen Vakuole (Abb. 6). Da saure Phosphatase bereitsin den kleineren Vakuolen nachgewiesen werden kann, muss der enzymatische Abbauschon in diesen Strukturelementen beginnen. Die vollständige Lyse findet jedoch stetsin der grossen Vakuole statt (Abb. 8). Die membranartigen, «residual bodies» ähn-lichen (HOHL 1965; MILLER und PALADE 1964) Reststoffe (Abb. 7) werden bei nacktenAmöben ins Nährmedium ausgeschieden. Bei encystierenden Amöben dagegen werdendie Abbauprodukte in die entstehende Cystenwand ausgeschleust und dort deponiert(vgl. Abschnitt: IV, 3). Die Lokalisation von saurer Phosphatase in Vakuolen istbereits mehreren Autoren gelungen (MILLER und PALADE 1964; Poux 1963). MATILE
und WIEMKEN (1967) wiesen in isolierten Vakuolen von Hefe mehrere lytische Enzymenach. Die interessante Tatsache, dass den Vakuolen von Acanthamoeba sp. in bezugauf ihre lytische Tätigkeit Doppelfunktion zukommt: Abbau von festen Nahrungs-teilchen, beispielsweise Bakterien, sowie eventuell Lyse zelleigener Strukturen, erinnertwiederum an die systematische Stellung als typischer Übergangsorganismus.
Gelegentlich beobachtet man bei nackten, vor allem aber bei den zur Encystierungangeregten Amöben konzentrische, in sich geschlossene Membranstapel des Endo-plasmatischen Retikulums (ER) (Abb. 9a). Die Aussenseiten der Membranen sinddicht mit Ribosomen besetzt. Allmählich kann an bestimmten Stellen ein lokalesAnschwellen der Membranen unter gleichzeitigem Zerreissen der Stapel festgestelltwerden (Abb. 9 b, c). Zu einem noch späteren Zeitpunkt schnüren sich die nunendständigen im Mittel 600-1000 A messenden und vollkommen ribosomenfreienBläschen ganz vom ER ab (Abb. 9d) und bilden ein dichtes, Golgi-Apparat ähn-liches Vesikelfeld mit einer meist zentral orientierten, grossen Vakuole (Abb. 9 e, f).Die beobachteten Vesikelfelder dürfen kaum als Golgi-Apparate im Sinne der höherenPflanzen bezeichnet werden. Funktionell kommt ihnen jedoch eine vergleichbareBedeutung zu. Nach FREY-WYSSLING et al. (1964) spielen die abgeschnürten Golgi-Vesikel der höheren Pflanzen eine entscheidende Rolle bei der Neubildung der Zell-platte nach der Zellteilung. STAEHELIN (1966) konnte zeigen, dass auch bei Chlorelladen Golgi-Bläschen matrixbildende Funktion zukommt. Inwieweit letztere Eigen-schaft auch auf die vesikelförmigen Zerfallsprodukte des Endoplasmatischen Retiku-lums bei Acanthamoeba sp. zutrifft, wird später beschrieben und diskutiert (vgl. Ab-schnitt: IV, 1 und 4).
Da sich die Amöben bei Züchtung in einem flüssigen Nährsubstrat abrunden undnur die passive Bewegung der Nährlösung mitmachen, ist die Ausbildung von Pseudo-podien und fibrillären Strukturen nur selten zu beobachten. In sehr dichten Popula-tionen sedimentiert jedoch ein grosser Teil der Mikroorganismen und führt aktiveKriechbewegungen aus. Nur unter diesen Kulturbedingungen kommt es zur Aus-bildung von fibrillären Strukturen. Dabei schliessen sich die einzelnen, 40-60 A

178 VierteljahIsschrift der Naturforschenden Gesellschaft in Zürich 1967
messenden Filamente zu Verbänden höherer Ordnung, welche eine Dicke von nahezueinem erreichen können, zusammen (Abb. 10). Diese fädigen Cytoplasmaelementedürfen nicht mit den bis 200 A dicken, röhrenförmigen Mikrotubuli, welchen AM-BROSE (1965) eine grosse Bedeutung für die Zellbewegung zumisst, verwechseltwerden. Letztere sind bei Acanthamoeba sp. nur sehr selten zu beobachten. In Physarum(WOHLFAHRT-BOTTERMANN 1964b) und vielen PrOtOZOen (RANDALL und JACKSON
1958) werden überhaupt keine Mikrotubuli ausgebildet. An ihrer Stelle treten 30-70 Adicke Plasmafilamente auf. Diese bezeichnet WOHLFAHRT-BOTTERMANN (1964b) alsdie eigentlichen kontraktilen Cytoplasmaelemente. Es ist daher naheliegend, dass sieauch bei Acanthamoeba sp. im Dienste der aktiven Fortbewegung stehen. DieseFolgerung wird auch dadurch gestützt, dass bei encystierten Zellen, welche keineaktive Eigenbewegung ausführen, die Plasmafilamente vollständig fehlen.
IV. Zellwandbildung (Encystierung)
NEFF et al. (1964) postulieren auf Grund ihrer vorwiegend chemisch-analytischenUntersuchungen an isolierten Cystenwandfragmenten eine zweischichtige Ze llwand.Die äussere, dünnere und zuerst gebildete Schicht soll vor allem Protein enthalten.Erst spät im Encystierungsprozess wird diese «Primärwand» durch eine bedeutendmächtiger ausgebildete cellulosehaltige « Sekundärwand » verstärkt.
1. Ablauf der Encystierung
Im flüssigen Mangelmedium ist die Encystierungsgeschwindigkeit abhängig von derPopulations-Dichte und vom Alter der Zellkultur sowie von der Temperatur und derBelüftungsintensität. Erwartungsgemäss encystieren ältere Kulturen schneller alsjüngere. 27°C erweist sich als die günstigste Encystierungstemperatur. Meist lassensich bereits nach 3-4 Stunden deutliche cytoplasmatische Veränderungen feststellen.Am auffallendsten ist die starke Ausbildung des ER. Membranstapel lagern sichparallel unterhalb dem Plasmalemma (Abb. 15). Die Aussenseiten der Membranensind dicht mit Ribosomen besetzt (Abb. 15). Endständige Anschwellungen der ER-Kavernen schnüren sich später ganz vom ER ab und bilden ribosomenfreie Bläschenmit einem sehr variablen Durchmesser von 400-2000 A. Gleichzeitig lässt sich eineWanderung dieser Vesikel in die Region unter das Plasmalemma erkennen (Abb. 12,13). Nach längerer Permanganat-Fixation kann der feingranuläre Inhalt der Vesikeldunkel angefärbt werden (Abb. 12).
Das Plasmalemma ist in diesem Entwicklungsstadium immer noch dicht mit90-120 A messenden Partikeln in statistischer Verteilung besetzt (Abb. 11). Ein sichkurz nach Encystierungsbeginn bildendes Lamellensystem ausserhalb des Plasma-lemmas (Abb. 12) zerfällt rasch wieder und ist verantwortlich für die B ildung deräussersten, körnigen «Cuticularschicht» der entstehenden Cystenwand (Abb. 14).Die warzige Oberflächenstruktur dieser Deckschicht (Abb. 23) zeigt eine deutlicheAnalogie zur Ausbildung der Exine bei Pollenkörnern (MüHLETHALER 1953, 1955).
Mikrotubuli (Abb. 14) lassen sich bei zur Encystierung angeregten Amöben selten
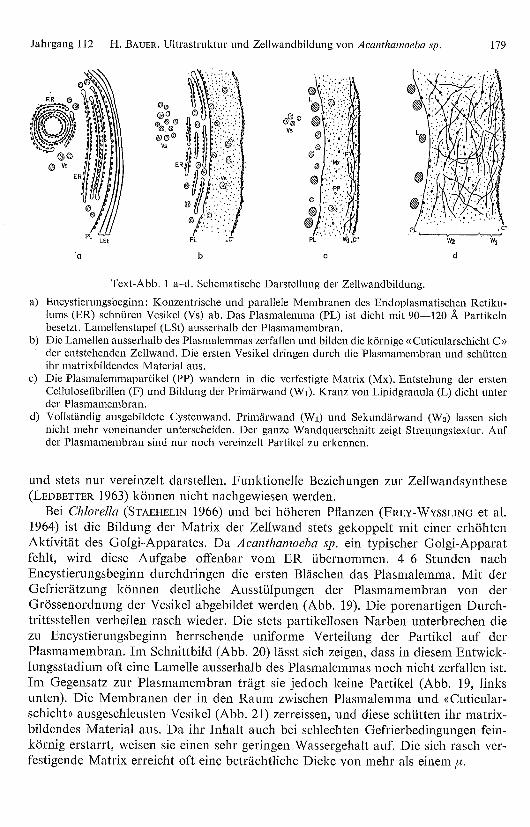
Jahrgang 112 H. BAUER. Ultrastruktur und Zellwandbildung von Acanthamoeba sp. 179
Text-Abb. 1 a–d. Schematische Darstellung der Zellwandbildung.
a) Encystierungsbeginn: Konzentrische und parallele Membranen des Endoplasmatischen Retiku-lums (ER) schnüren Vesikel (Vs) ab. Das Plasmalemma (PL) ist dicht mit 90-120 A Partikelnbesetzt. Lamellenstapel (LSt) ausserhalb der Plasmamembran.
b) Die Lamellen ausserhalb des Plasmalemmas zerfallen und bilden die körnige «Cuticularschicht C»der entstehenden Zellwand. Die ersten Vesikel dringen durch die Plasmamembran und schüttenihr matrixbildendes Material aus.
c) Die Plasmalemmapartikel (PP) wandern in die verfestigte Matrix (Mx). Entstehung der erstenCellulosefibrillen (F) und Bildung der Primärwand (W )). Kranz von Lipidgranula (L) dicht unterder Plasmamembran.
d) Vollständig ausgebildete Cystenwand. Primärwand (W)) und Sekundärwand (W2) lassen sichnicht mehr voneinander unterscheiden. Der ganze Wandquerschnitt zeigt Streuungstextur. Aufder Plasmamembran sind nur noch vereinzelt Partikel zu erkennen.
und stets nur vereinzelt darstellen. Funktionelle Beziehungen zur Zellwandsynthese(LEDBETTER 1963) können nicht nachgewiesen werden.
Bei Chlorella (STAEHELIN 1966) und bei höheren Pflanzen (FREY-WYSSLING et al.1964) ist die Bildung der Matrix der Zellwand stets gekoppelt mit einer erhöhtenAktivität des Golgi-Apparates. Da Acanthamoeba sp. ein typischer Golgi-Apparatfehlt, wird diese Aufgabe offenbar vom ER übernommen. 4-6 Stunden nachEncystierungsbeginn durchdringen die ersten Bläschen das Plasmalemma. Mit derGefrierätzung können deutliche Ausstülpungen der Plasmamembran von derGrössenordnung der Vesikel abgebildet werden (Abb. 19). Die porenartigen Durch-trittsstellen verheilen rasch wieder. Die stets partikellosen Narben unterbrechen diezu Encystierungsbeginn herrschende uniforme Verteilung der Partikel auf derPlasmamembran. Im Schnittbild (Abb. 20) lässt sich zeigen, dass in diesem Entwick-lungsstadium oft eine Lamelle ausserhalb des Plasmalemmas noch nicht zerfallen ist.Im Gegensatz zur Plasmamembran trägt sie jedoch keine Partikel (Abb. 19, linksunten). Die Membranen der in den Raum zwischen Plasmalemma und «Cuticular-schicht» ausgeschleusten Vesikel (Abb. 21) zerreissen, und diese schütten ihr matrix-bildendes Material aus. Da ihr Inhalt auch bei schlechten Gefrierbedingungen fein-körnig erstarrt, weisen sie einen sehr geringen Wassergehalt auf. Die sich rasch ver-festigende Matrix erreicht oft eine beträchtliche Dicke von mehr als einem μ.

180 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1967
12-16 Stunden nach Übertragung der Protoplasten in das Hungermedium lösensich die Plasmalemmapartikel von der Membran ab und wandern in die nun sehrdichte und nach Gefrierätzung homogen verteilte Zellwandmatrix (Abb. 22, 24).Durch ständigen Nachschub der sich von Plasmalemma lösenden Teilchen entstehtschliesslich ein dichtes Partikelfeld (Abb. 25). Erst jetzt treten im Zusammenhangmit der starken Partikelansammlung in der Grundsubstanz, deutlich von der Plasma-membran getrennt, die ersten Cellulosefibrillen auf (Abb. 25, 26). Bei frisch syntheti-sierten Fibrillen kann eine direkte Verbindung mit den Plasmalemmapartikeln(Abb. 26) festgestellt werden. Zwischen den in Streuungstextur angeordneten Fibrillenlassen sich zudem freie Teilchen von der gleichen Grösse (90-120 A) erkennen(Abb. 26). Abb. 27 zeigt erneut überkreuzte, in keiner Weise mit dem Plasmalemmaverbundene, zum Teil immer noch mit Partikeln besetzte Fibrillen, eingebettet inder wasserarmen Matrix. In der Gefrierätzung gelangen sowohl 35-40 A dickeElementarfibrillen (Abb. 26), wie auch 80-240 A messende Mikrofibrillen (Abb. 29)zur Abbildung. Sie sind dicht ineinander verwoben. Mit Ultraschall können die höhe-ren Fibrillenverbände in einzelne Elementarfibrillen aufgespalten und mit Hilfe derNegativkontrastmethode im Elektronenmikroskop sichtbar gemacht werden (Abb. 28).
Auch bei vollständig encystierten Amöben bleibt die anfängliche Streuungs-textur der Celluloserfibrillen erhalten (Abb. 29). Bevorzugte Fibrillenrichtungen sindnicht vorhanden. Das Plasmalemma trägt nur noch ganz vereinzelt globuläre 90-120 APartikel. Im Cytoplasma, dicht unterhalb des Plasmalemmas, ist meist ein Kranz vonLipidtropfen zu finden. Bei der Auskeimung der Cyste wird dieser Reservestoff wiedermobilisiert.
2. Zellwandproteine
Die Streitfrage, ob die Zellwände Protein enthalten oder nicht, ist schon sehr alt.Bereits 1888 postulierte WIESNER, dass die Zellwände, mindestens während demWachstum, eiweisshaltig sein müssen. Seine Resultate wurden aber lange bezweifeltund der Proteingehalt auf plasmatische Verunreinigungen der Wandisolate zurück-geführt. Durch verfeinerte Fraktionierungsmethoden und die Kontrollierbarkeit derIsolate mit Hilfe des Elektronenmikroskopes wurde in letzter Zeit den Zellwand-proteinen immer grössere Beachtung geschenkt. So fanden STEWARD et al. (1958) inProteinhydrolysaten schnell wachsender Gewebe Hydroxyprolin als Leitaminosäure.POLLARD und STEWARD (1959) konnten zeigen, dass mit C14 markiertes Prolin vonsich teilenden Geweben rasch aufgenommen wird. Zudem stellten sie fest, dass dieHydroxylierung von Prolin erst nach dem Einbau dieser Aminosäure in Proteinestattfindet. Im gleichen Jahr fanden NICKERSON und FALCONE (1959) in sprossendenHefezellen eine Protein-Disulfid-Reduktase. KING und BAYLEY (1965) identifiziertenin den Primärwandproteinen höherer Pflanzen 20 % Hydroxyprolin. In einem um-fassenden Bericht macht LAMPORT (1965) die Primärwandproteine der höherenPflanzen für das Flächenwachstum verantwortlich. In diesem Zusammenhang führter für die Hydroxyprolin reichen Primärwandproteine den Begriff «Extensiv» ein.Kürzlich konnten THOMPSON und PRESTON (1967) auch in veIschiedenen Grünalgen-zellwänden bis zu 10 % Protein nachweisen.

Jahrgang 112 H. BAUER. Ultrastruktur und Zellwandbildung von Acanthamoeba sp. 181
Da KLEIN und NEFF (1964) in reifenden Cystenwänden von Acanthamoeba sp.ebenfalls 33 % Protein fanden, war es von grossem Interesse, auch diesen Mikro-organismus auf die für höhere Pflanzen typische Leitaminosäure Hydroxyprolin zuprüfen. Tatsächlich lässt sich nach 18stündiger Hydrolyse mit 6n Salzsäure mit dervon JEPSON und SMITH (1953) entwickelten, spezifischen Farbreaktion bei encystiertenAmöben qualitativ eindeutig Hydroxyprolin nachweisen. Der Hydroxyprolinge-halt der Plasmaproteine nackter Amöben hingegen ist zu gering, um auf dieseTestreaktion positiv anzusprechen. Dies besagt also, dass analog der höheren Pflanzenauch bei Acanthamoeba sp. Wandproteine als «Extensiv» bezeichnet werden können.Die Hydroxylierung der Aminosäure Prolin erfolgt wahrscheinlich ebenfalls erst inder Zellwand.
In einem zweiten Schritt galt es die hydroxyprolinreichen Proteine in der Zellwandzu lokalisieren. Dazu wurden die Amöben während der Encystierung mit H3-Prolingefüttert. Schon nach 4-6 Stunden kann eine aktive Aufnahme der Tritium haltigenAminosäure beobachtet werden. Der Einbau erfolgt vorerst zu ca. 60 % (ermitteltdurch Auszahlung der entwickelbaren Körner) in Proteine der Kernmembran(Abb. 16). Parallel zu der starken Ausbildung des ER bei den zur Encystierung an-geregten Amöben (vgl. Abschnitt: IV, 1) lassen sich immer mehr Körner über Elemen-ten dieses Kavernensystems beobachten (Abb. 17). Diese Tatsache lässt sich mit derKontinIität von Kernmembran und ER, sowie der strukturell an Ribosomen ge-bundenen Synthese von Proteinen, gut erklären. Bei encystierten Amöben liegen dieKörner bis zu 90 % in der Primärwand (Abb. 18). In der viel mächtiger ausgebildetenSekundärwand sind dagegen nur vereinzelt entwickelbare Körner vorhanden. Darausgeht hervor, dass nur die Primärwandproteine hydroxyprolinreich sind und somitauch bei Acanthamoeba sp. als «Extensin» bezeichnet werden dürfen (vgl. auch Ab-schnitt: IV, 4).
3. Zellwandeinschlüsse
Nach chemischer Fixierung lassen sich in bestimmten Bezirken der vollständigausgebildeten Cystenwand membranartige und bläschenförmige Einschlüsse beob-achten (Abb. 30). Morphologisch stimmen diese Strukturen mit den bereits be-schriebenen Überresten der Autolyse-Produkte in den Verdauungsvakuolen überein.Da zudem entlang der Membranfragmente eine deutliche Aktivität der saurenPhosphatase nachgewiesen werden kann, muss es sich um die in die Cystenwand aus-geschleusten Abbauprodukte handeln. Während bei nackten Amöben diese Exkreteohne weiteres ins Kulturmedium ausgeschieden werden können, stellt bei Cysten dieZellwand eine unüberwindbare Barriere dar. Aus diesem Grunde werden die Abbau-produkte in der Zellwand deponiert und, da immer noch saure Phosphatase nach-gewiesen werden kann, weiter abgebaut.
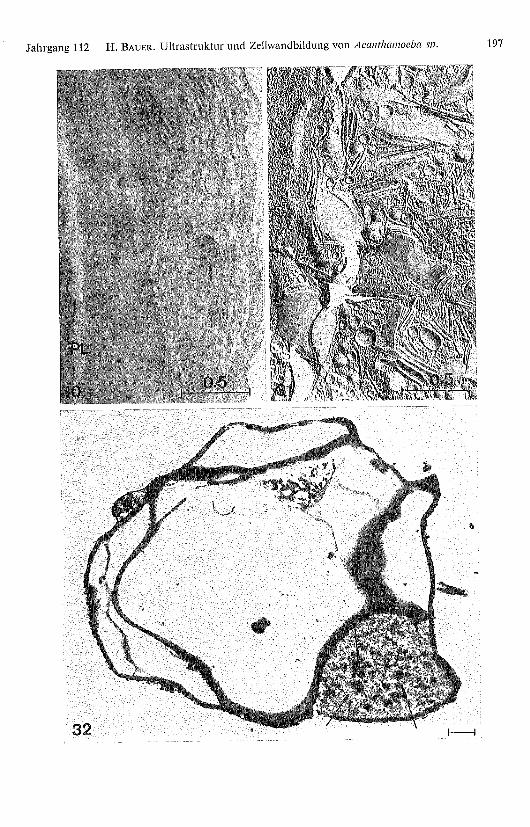
Auch nach Gefrierätzung zeigen die Wandeinschlüsse «unit membrane» Charakter(Abb. 31). Nach der Veraschung von Dünnschnitten encystierten Amöben bleibenin den Wandaussparungen nicht verbrennbare Rückstände übrig (Abb. 32). Diesdeutet darauf hin, dass diese Autolyse-Produkte stark mit anorganischen Substanzeninkrustiert sind. NEFF et al. (1964) gelang es, 2 % der anorganischen Wandsubstanz

182 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1967
als Phosphor zu identifizieren. Es darf daher, in Übereinstimmung mit der Morpholo-gie, angenommen werden, dass ein grosser Teil dieser Wandeinschlüsse Phospholipidedarstellen. Diese Ansicht wird zudem gestützt durch die Arbeit von HENN et al. (1967),welche zeigen konnten, dass synthetische Lipiddoppelschichten ebenfalls eine «unitmembrane» Struktur aufweisen.
4. Diskussion
Die gesteigerte Aktivität des dicht mit Ribosomen besetzten ER der zur Encystie-rung angeregten Amöben muss mit einer stark erhöhten Proteinsynthese verbundensein. Mit Hilfe der elektronenmikroskopischen Autoradiographie lässt sich Proteindirekt über dem ER lokalisieren. Kürzlich konnten MATILE et al. (1967) in denPlasmalemmapartikeln der Bäckerhefe nebst Mannan auch Protein nachweisen. Esist daher durchaus möglich, dass diese proteinhaltigen, globulären Partikel vom ERsynthetisiert werden. Mit dieser Hypothese liesse sich gut die Ausschleusung vonProtein in die Zellwand erklären. Die Genese der Partikel und ihre Einlagerung in diePlasmamembran morphologisch zu erfassen, ist aber ausserordentlich schwierig undbis jetzt noch nicht gelungen. Nach Untersuchungen von KLEIN und NEFF (1964)enthält die reifende Cystenwand von Acanthamoeba sp. 33 % Protein. Es ist daherwahrscheinlich, dass nicht der ganze Proteingehalt der Ze llwand nur von den Plasma-lemmapartikeln stammt. Zumal in der Primärwand bereits vor dem Auswandern derPlasmalemmapartikel hydroxyprolinreiches Protein lokalisiert werden kann, darfangenommen werden, dass dieses von den ausgeschleusten ER-Vesikeln stammt.
SIEVERS (1963) zeigte, dass mit Permanganat stark färbbare Bläschen des Golgi-Apparates bei der Bildung der Zellwand von Wurzelhaaren betei ligt sind. DurchFusion seggregierter Golgi-Vesikel der höheren Pflanzen wird bei der Zellteilung dieneu entstehende Zellplatte gebildet (FREY-WYSSLING et al. 1963). Zudem konnteSTAEHELIN (1966) beobachten, dass bei Chlorella den abgeschnürten Golgi-Bläschenmatrixbildende Funktion zukommt. Da Acanthamoeba sp. keine Golgi-Apparate imSinne der höheren Pflanzen besitzt, muss die Matrixbildung durch ein anderes Zell-organell übernommen werden. Es kann gezeigt werden, dass das ER diese Aufgabezu erfüllen hat. Dabei werden die ganzen Vesikel in den Raum zwischen « Cuticular-schicht » und Plasmalemma ausgeschleust, wo sie platzen und ihr Material ausschütten.Analoge Beobachtungen beschrieben STAEHELIN (1966) bei den Golgi-Bläschen vonChlorella und MATILE (1965) bei den Proteasepartikeln von Neurospora crassa.
Die ersten Hinweise, dass globuläre Teilchen auf der Plasmamembran an derBildung von Fibrillen beteiligt sind, können der Arbeit von MOOR und MüHLETHALER
(1963) entnommen werden. 180 A Partikel liegen zum Teil in hexagonaler An-ordnung vor. In diesen Bereichen lassen sich 50 Å dicke Fibrillen (wahrscheinlichGlucan) abbilden. Ebenso stellte DOST (1965) bei Oscillatoria rubescens von den Plas-malemmapartikeln in die Zellwand hinaus führende Fibrillen fest. Bei Chlorellazeigte STAEHELIN (1966), dass sich die Partikel vom Plasmalemma lösen, in dieMatrix wandern und erst dort die ersten Fibrillen gebildet werden.
Analoge Verhältnisse liegen bei Acanthamoeba sp. vor. Erst spät im Encystierungs-ablauf wandern die durchschnittlich 90-120 A messenden Teilchen in die schon gut

Jahrgang 112 H. BAUER. Ultrastruktur und Zellwandbildung von Acanthamoeba sp. 183
verfestigte Matrix. Zur Bildung der Cellulosefibrillen sind stets mehrere Partikel not-wendig. In einer bestimmten Entwicklungsphase können direkte Verbindungen derTeilchen mit den entstehenden Fibrillen beobachtet werden (Abb. 26). Die Plasma-lemmapartikel finden sich nicht nur in der Primärwand, sondern auch in der Sekundär-wand in Verbindung mit den entstehenden Cellulosefibrillen. Es drängt sich daherdie Vermutung auf, dass diese globulären Teilchen, im Gegensatz zu dem auf diePrimärwand beschränkten «Extensin» und in Analogie zur Bäckerhefe (MATILE et al.1967), Protein-Polysaccharid-Komplexe (nicht hydroxyprolinreich) darstellen. Esmuss also in der Zellwand zumindest zwischen zwei verschiedenen Protein-Komponen-ten unterschieden werden. Zudem bleibt die Frage offen, ob für die Synthese derCellulosefibrillen zusätzlich noch Enzymproteine notwendig sind.
Entgegen den Resultaten von HEYN (1966) an negativ kontrastierten Schnittenverschiedener Pflanzenfasern gelangen mit der Gefrierätzung nicht nur Elementar-fibrillen (MÜHLETHALER 1960, FREY-WYSSLING et al. 1966), sondern auch 80-240 Ådicke Mikrofibrillen zur Abbildung, die allerdings in Elementarfibrillen aufgespaltenwerden können (Abb. 28). Die Fibrillen sind ineinander verwoben und bilden auchnach Abschluss der Encystierung eine Streuungstextur. Ein Zusammenhang mit denvon LEDBETTER (1963), NEWCOMB und BONNET (1965) beschriebenen Mikrotubuliund der Fibrillenbildung kann nicht festgestellt werden. Eine Trennung in Primär-und Sekundärwand ist nach AbschlIss der Encystierung nicht mehr zu beobachten.
Die deutliche Analogie in der Ausbildung einer warzigen Oberflächenstruktur derDeck- oder «Cu ticularschicht », vergleichbar mit der mannigfaltigen Gestaltung vonPollenexinen (MÜHLETHALER 1953, 1955), steht wohl in Zusammenhang mit derähnlichen Funktion von Cysten und Pollenkörnern.
V. Zusammenfassung
Mit verschiedenen Methoden der Elektronenmikroskopie wurde die Ultrastruktur,insbesondere die Zellwandbildung von A canthamoeba sp. untersucht. Zur Feinstrukturder Zellorganelle seien folgende Punkte rekapituliert:
1. Im nackten Zustand liegt die Amöbe als teilngsfähiger, nach aussen lediglichdurch das Plasmalemma begrenzter Protoplast vor.
2. Die Mitochondrien gehören dem tubulären Typus an und zeigen oft Einschlüssevon anorganischem Material.
3. Relativ grosse Vakuolen mit Inhaltsstoffen können sowohl im Dünnschnitt, wieauch mit Hilfe der Gefrierätzung nachgewiesen werden. Ebenfalls gelingt es, in diesenVakuolen saure Phosphatase zu lokalisieren.
4. Ein eigentlicher Golgi-Apparat fehlt der Amöbe; dafür kann ausgehend vonkonzentrischen Lamellen des nur kurz vor der Encystierung stark aktiven ER eineBläschengenese mit resultierendem Vesikelfeld (Art Golgi-Apparat) und meist zentralorientierter Vakuole beobachtet werden.
5. Mit Glutaraldehyd-Fixierung lassen sich im Cytoplasma bis zu einem μ dickeBündel von durchschnittlich 50 A messenden kontraktilen Filamenten abbilden.

184 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1967
Die Bildung der Cystenwand lässt sich in folgende Schritte unterteilen:a) Bei zur Encystierung angeregten Amöben stellt man im Cytoplasma, dicht
unterhalb des Plasmalemmas und parallel zu diesem, Membranstapel des dicht mitRibosomen besetzten Endoplasmatischen Retikulums fest. Die daraus resultierendeerhöhte Proteinsynthese wird durch autoradiographische Befunde gestützt. Zudemist eine Verlagerung der im Mittel 600-1000 Å messenden Vesikel in die Region inner-halb des Plasmalemmas zu erkennen.
b) Die Plasmamembran ist zu Beginn der Encystierung dicht mit 90-120 Åmessenden Partikeln besetzt. Ausserhalb des Plasmalemmas lässt sich eine Lamel-lierung feststellen.
c) Die Lamellen zerfallen wieder und es kommt zur Bildung der körnigen «Cuti-cularschicht » der entstehenden Cystenwand.
d) Die Vesikel durchdringen das Plasmalemma und schütten ihr matrixbildendesMaterial in den Raum zwischen Plasmamembran und «Cuticularschicht».
e) Mit Hilfe der Autoradiographie kann in der Primärwand hydroxyprolin-reiches Protein («Extensiv») nachgewiesen werden.
f) Die globulären Plasmalemmapartikel lösen sich von der Ze llmembran, wandernin die sich verfestigende Matrix und bilden dort ein dichtes Partikelfeld. Erst jetztwerden die ersten Cellulosefibrillen synthetisiert. Es kann eine direkte Verbindung derPartikel mit den entstehenden Fibrillen beobachtet werden. Die Partikel stellen wahr-scheinlich Protein-Polysaccharid-Komplexe dar.
g) Die Streuungstextur der Cellulosefibrillen bleibt auch bei vollständig encystier-ten Amöben erhalten. Eine Trennung in Primär- und Sekundärwand kann nicht mehrbeobachtet werden. Das Plasmalemma ist nach Abschluss der Zellwandsynthese nahe-zu partikelfrei.
Die vorliegende Untersuchung wurde im Laboratorium für Elektronenmikroskopie (Prof. Dr.K. MGHLETHALER) des Institutes für Allgemeine Botanik (Prof. Dr. A. FREY-WYSSLING) ausgeführt.Für die finanzielle Unterstützung danken wir dem Schweizerischen Nationalfonds.
VI. Literatur
AMRROSE, E. J.: «Zellbewegungen». Endeavour, 24, 27-32 (1965).BHOWMICK, D. K. and K. E. WOHLFARTH-BOTTERMANN: An improved method for fixing amoeba
for electron microscopy. Exp. Cell. Res. 40, 252-263 (1965).CARO, L. G. and R. P. VAN TUBERGEN: High resolution autoradiography. I. Methods. J. Cell Biol. 15,
173-188 (1962).FREY-WYSSLING, A., K. MÜHLETHALER und R. MUGGLI: Elementarfibrillen als Grundbausteine der
nativen Cellulose. Holz als Roh- und Werkstoff 24, 443-444 (1966).FREY-WYSSLING, A., J. F. LÓPEZ-SÁEZ and K. MÜHLETHALER: Formation and development of the
cell plate. J. Ultrastr. Res. 10, 422-432 (1964).GOMORI, G.: In microscopic histochemistry principles and practice. University of Chicago Press
(1952).HAASE, G. und G. JUNG: HeIstellungen von Einkornschichten aus photographischen Emulsionen.
Naturw. 51, 404-405 (1964).HEYN, A. N. J.: The microcristalline structure of cellulose in cetl walls of cotton, ramie and jute
fibers as revealed by negativ staining of sections. J. Cell Biol. 29, 181-197 (1966).

Jahrgang 112 H. BAUER. Ultrastruktur und Zellwandbildung von Acanthamoeba sp. 185
HoHL, H. R.: Nature and development of membrane systems in food vacuoles of cellular slimemolds predatory upon bacteria. J. Bact. 90, 755-765 (1965).
JEPSON, J. B. and I. SMITH: «Multiple dipping» procedures in paper chromatography. A specifictest for hydroxyproline. Nature, 172, 1100-1101 (1953).
JosT, M.: Die Ultrastruktur von Oscillatoria rubescens D. C. Arch. Mikrobiol. 50, 211-245 (1965).KARNOVSKY, M. J.: Simple methods for «staining with lead» at high pH in electron microscopy.
J. biophys. biochim. Cytol. 11, 729-732 (1961).KING, N. J. and S. T. BAYLEY: A preliminary analysis of the proteins of the primary walls of some
plant cells. J. Exp. Bot. 16, 294-303 (1965).KLEIN, R. L. and R. J. NEFF: Osmotic properties of mitochondria isolated from Acanthamoeba sp.
Exp. Cell Res. 19, 133-155 (1960).LAMPORT, D. T. A.: The protein component of primaIy cell walls. In: Advances in botanical research
2, 151-218, ed. by R. D. PRESTON. Academic Press London and New York (1965).LEDRETTER, M. C. and K. R. PORTER: A «microtubule» in plant cell fine structure. J. Cell Biol. 19,
239-250 (1963).LETTR, H. and N. PAWELETZ: Probleme der elektronenmikroskopischen Autoradiographie. Naturw.
53, 268-271 (1966).MATILE, P., M. JOST und H. MOOR: Intrazelluläre Lokalisation proteolytischer Enzyme von Neuro-
spora crassa. H. Identifikation von proteasehaltigen Zellstrukturen. Z. Zellforsch. 68, 205-216(1965).
MATILE, P. und A. WIEMKEN: The vacuole as the lysosome of the yeast cell. Arch. Mikrobiol. 56,148-155 (1967).
MATILE, P., H. MooR and K. MÜHLETHALER: Isolation and properties of the plasmalemma in yeast.Arch. Mikrobiol. (1967 im Druck).
MILLER, F. and G. E. PALADE: Lytic activities in renal protein absorption droplets. An electronmicroscopical cytochemical study. J. Cell Biol. 23, 519-552 (1964).
MooR, H.: Die Gefrier-Fixation lebender Zellen und ihre Anwendung in der Elektronenmikroskopie.Z. Zellforsch. 62, 546-580 (1964).
MOOR, H. and K. MÜHLETHALER: Fine structure in frozen-etched yeast cells. J. Cell Biol. 17, 609-628(1963).
MOOR, H., H. WALDNER and A. FREY-WYSSLING: A new freezing ultramicrotome. J. biophys. bio-chim. Cytol. 10, l-13 (1961).
MOOR, H., C. RUSKA und H. RUSKA: Elektronenmikroskopische Darstellung tierischer Zellen mitder Gefrierätztechnik. Z. Zellforsc. 62, 581-601 (1964).
MÜHLETHALER, K.: Die Struktur einiger Pollenmembranen. Planta 46, l-13 (1955).— Untersuchungen über die Struktur der Pollenmembran. Mikroskopie 8, 103-110 (1953).— Die Feinstruktur der Cellulosemikrofibrillen. Z. schweiz. Forstv. 30, Beiheft «Festschrift Prof.
FREY-WYSSLING», 211-222 (1960).NEFF, R. J., W. F. BENTON and R. H. NEFF: The composition of the mature cyst wall of the soil
ameba Acanthamoeba sp. J. Cell Biol. 23, 66A (1964).NEWCOMR, E. H. and H. T. BONNET: Cytoplasmic microtubule and wall microfibril orientation in
root hairs of radish. J. Cell Biol. 27, 575-589 (1965).NICKERSON, W. J. and G. FALCONE: Function of protein disulfide reductase in cellular divison of
yeast. In Sulfur in Proteins 404, edit. by R. A. BENESCH (Academic Press, 1959).PALADE, G. E.: A study of fixation for electron microscopy. J. exp. Med. 95. 285-298 (1952).Poux, N.: Localisation des phosphates et de la phosphatase acide dans les cellules des embryons de
ble (Triticum vulgare Vill.) lors de la germlnation. J. Microscopic 2, 557-568 (1963).POLLARD, J. K. and F. C. STEWARD: The use of C14-proline by growing cells: Its conversion to protein
and hydroxyproline. J. Exp. Bot. 10, 17-32 (1959).RANDALL, J. T. and S. JACKSON: Fine structure and function in Stentor polymorphus. J. biophys.
biochim. Cytol. 4, 807-830 (1958).REYNOLDS, E. S.: The use of lead citrate at high pH as an electron-opaque stain in electron micro-
scopy. J. Cell Biol. 17, 208-212 (1963).

186 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1967
SABATINI, D. D., K. BENSCH and R. J. BARNETT: CytoChemistry and electron microscopy. Thepreservation of cellular ultrastructure and enzymatic activity by aldehyd fixation J. Cell Biol.17, 19-58 (1963).
SIFVERS, A.: Beteiligung des Golgi-Apparates bei der Bildung der Zellwand von Wurzelhaaren.Protoplasma 56, 188-192 (1963).
STAEHELIN, A.: Die Ultrastruktur der Zellwand und des Chloroplasten von Chlorella. Z. Zellforsch.74, 325-350 (1966).
STEWARD, F. C., J. F. THOMPSON and J. K. POLLARD: Contrasts in the nitrogenous composition ofrapidly growing and non-growing plant tissue. J. Exp. Bot. 9, 1-10 (1958).
THOMAS, R. S.: Ultrastructural localisation of mineral matter in bacterial spores by microincine-ration. J. Cell Biol. 23, 113-133 (1964).
THOMPSON, E. W. and R. D. PRESTON: Proteins in the cell walls of some green algae. Nature 213,684-686 (1967).
TOMLINSON, G. and E. A. JOHNES: Isolation of cellulose from the cyst wall of a soil amoeba. Biochim.Biophys. Acta 63, 194-200 (1962).
WIESNER, J.: Über den Nachweis von Eiweisskörper in Pflanzenzellen. Ber. deut. Botan. Ges. 6,187-195 (1888).
WOHLFARTH-BOTTERMANN, K. E.: Differentiations of the ground cytoplasm and their significancefor the generation of the motiv force of ameboid movement. In: Primitive motile system in cellbiology, ed. by R. D. ALLEN and N. KAMIYA, New York Academic Press (1964b).Weitreichende, fibrilläre Protoplasmadifferenzierungen und ihre Bedeutung für die Protoplasma
-strömung. III. Entstehung und experimentell induzierbare Musterbildung. Roux' Arch. Ent-wicklungsmechanik 156, 371-403 (1965).Pinocytose und Bewegung von Amöben. II. Mitteilung: Permanente und induzierte Pinocytosebei Amoeba protects. Z. Zellforsch. 73, 444-474 (1966).
VII. Bildlegende
Tafel Seite 189
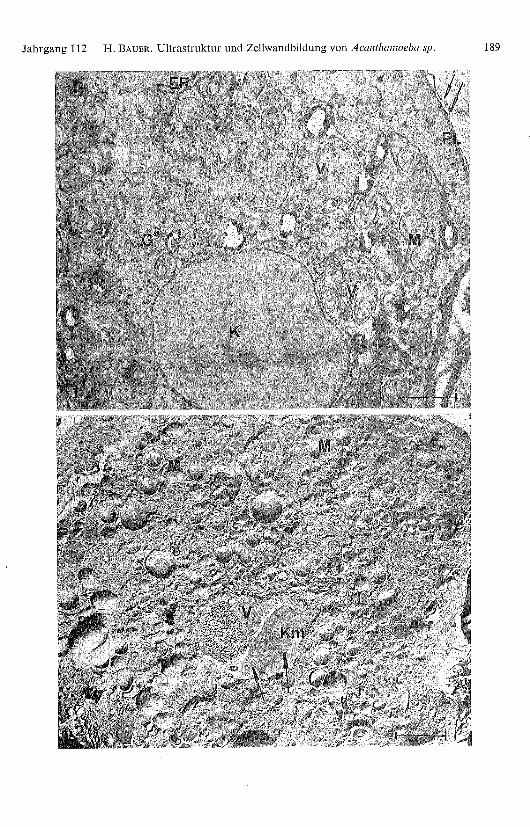
Abb. 1. Teilansicht eines zur Encystierung angeregten Protoplasten nach KMnO 4-Fixierung undUO2-Acetat-Nachkontrastierung. Ausserhalb des Plasmalemmas (PL) sind bereits einige Lamellen(Pfeile) ausgebildet. Durch Abschnürung von Vesikeln bildet das Endoplasmatische Retikulum (ER)Golgi-Apparat ähnliche Strukturen («G»). Weiter erkennt man mehrere Vakuolen (V), Mitochon-drien (M) und den Zellkern (K).
Abb. 2. Protoplast nach Gefrierätzung. Man beobachtet: Mitochondrienaufsichten (M), Lipid-tröpfchen (L), ein Teil der Kernmembran (Km) mit den charakteristischen Kernporen (Pfeile) unddicht daneben eine aufgebrochene Vakuole (V).
Tafel Seite 190
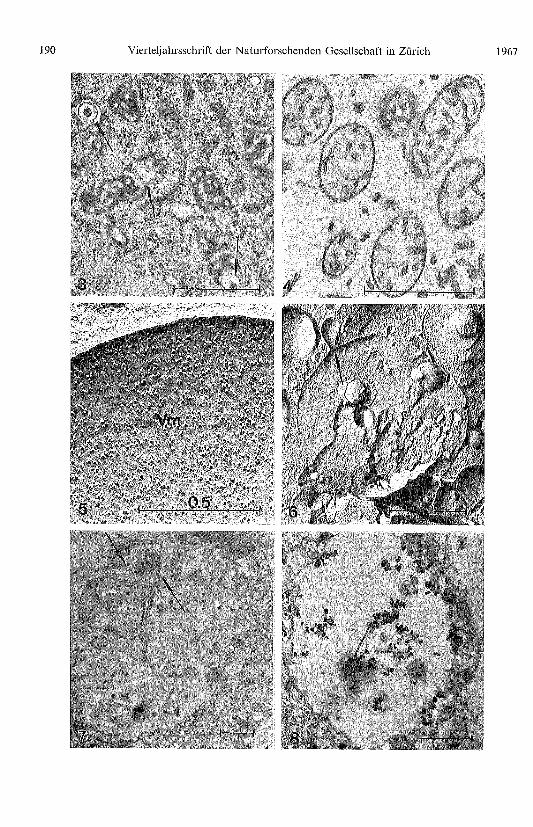
Abb. 3. Mitochondrien nach Glutaraldehyd-Osmium-Fixierung. Man beachte den durch dieUO2-Acetat-Vorkontrastierung bedingten Negativkontrasteffekt und die anorganischen Einschlüsse(Pfeile).
Abb. 4. Der tubuläre Charakter der inneren Mitochondrienstruktur kann besonders gut nachPermanganat-Fixierung festgestellt werden.
Abb. 5. Oberflächenansicht der Vakuolenmembran (Vm). Die 140 Å Partikel sind statistisch ver-teilt.
Abb. 6. Verdauungsvakuole mit Inhaltsstoffen nach Gefrierätzung. Fusion einer kleinerenVakuole mit der grossen zentralen Vakuole (Pfeile).

Jahrgang 112 H. BAUER. Ultrastruktur und Zellwandbildung von Acanthamoeba sp. 187
Abb. 7. Querschnitt durch eine Vakuole mit membranartigen Abbauendprodukten (Pfeile).Glutaraldehyd-Osmium-Fixierung.
Abb. 8. Auf den zum Teil bereits abgebauten Strukturen im Innern der Vakuole lässt sich saurePhosphatase lokalisieren.
Tafel Seite 191
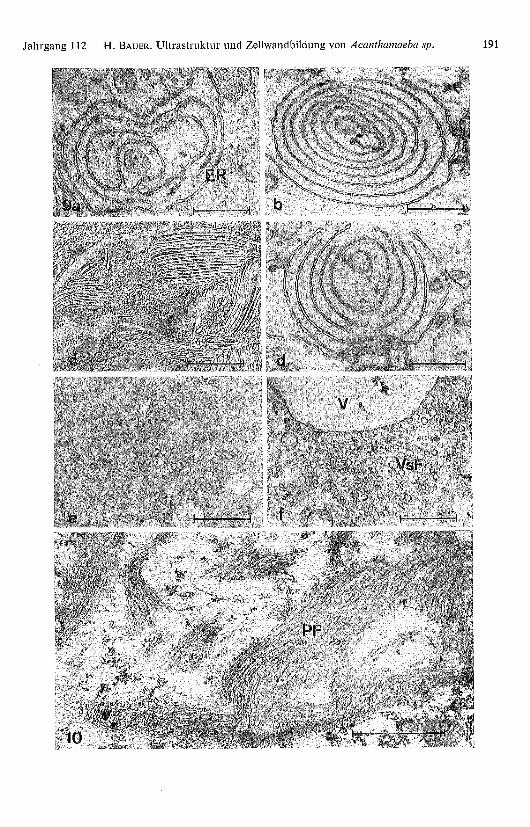
Abb. 9 a-f. Fortschreitender Zerfall konzentrischer Lamellen des Endoplasmatischen Retikulums(ER) und Bildung eines Golgi-Apparat ähnlichen Vesikelfeldes (VsF) mit meist zentral orientierterVakuole (V). Fixation: a, f: K2Cr2O7/0sO4; b, d: KMnO 4 ; c: Gefrierätzung und e: Glutaral-
dehyd/OsO4.
Abb. 10. Die 40-60 Å dicken Plasmafilamente (PF) schliessen sich stets zu Verbänden höhererOrdnung zusammen.
Tafel Seite 192
Abb. 11. Vor der Encystierung ist das Plasmalemma dicht mit 90-120 Å messenden Partikeln instatistischer Verteilung besetzt.
Abb. 12. Die nach 16stündiger Permanganat-Fixierung dunkel gefärbten Vesikel (Vs) wandern indie Region unter das Plasmalemma. Lamellenstapel (LSt) ausserhalb des Plasmalemmas.
Abb. 13. Dle Vesikel (Vs) der Abb. 12 nach Gefrierätzung.
Abb. 14. Durch Zerfall des Lamellensystems entwickelt sich die körnige «Cuticularschicht (C)»der entstehenden Cystenwand. Mikrotubuli (Pfeile) können selten und stets nur vereinzelt abgebildetwerden. Glutaraldehyd-Osmium-Fixierung.
Abb. 15. Dicht mit Ribosomen besetzte Membranstapel des Endoplasmatischen Retikulums (ER)lagern sich parallel unterhalb dem Plasmalemma. Osmium-Kaliumd ichromat-Fixierung.
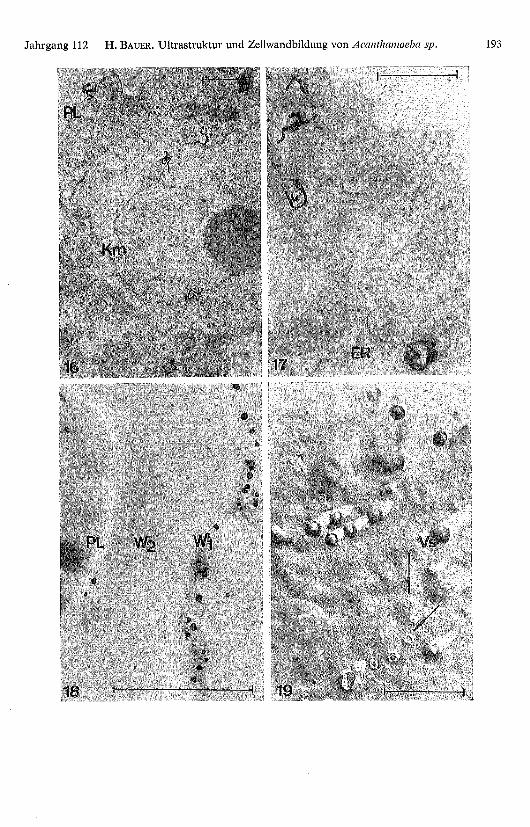
Tafel Seite 193
Abb. 16 Der Einbau von H3 -Prolin erfolgt vorerst zu ca. 60 % in Proteine der Kernmembran(Km). Entwicklung: Microdol-X; Fixation: Osmium-Kaliumdichromat.
Abb. 17. Später können immer mehr Körner über dem sich nun stark entwickelten Endoplasma-tischen Retikulum (ER) lokalisiert werden. Entwicklung: Microdol-X; Fixation: Osmium-Kalium-dichromat.
Abb. 18. Bei encystierten Amöben liegen die Körner in der Primärwand (W i). PL: Plasmalemma,Ws: Sekundärwand. Entwicklung: nach LETTRÉ (1966); Fixation: Osmium-Kaliumdichromat.
Abb. 19. Nach Gefrierätzung können deutliche Ausstülpungen der Plasmamembran von derGrössenordnung der Vesikel (Vs) abgebildet werden. Die porenartigen Durchtrittsstellen (Pfeile)verheilen rasch wieder unter Bildung von partikelfreien Narben.
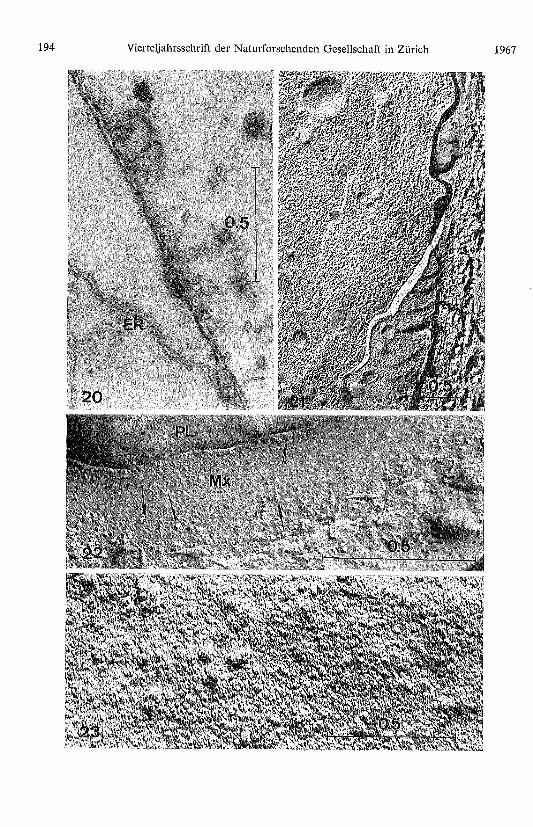
Tafel Seite 194
Abb. 20. Die vom Endoplasmatischen Retikulum (ER) abgeschnürten Vesikel durchdringen diePlasmamembran und schütten ihr matrixbildendes Material in den Raum zwischen Plasmalemmaund «Cuticularschicht». Permanganat-Fixierung.
Abb. 21. Wie Abb. 20 nach Gefrierätzung.

188 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1967
Abb. 22. Die 90-120 Å Partikel (Pfeile) lösen sich vorn Plasmalemma (PL) und wandern in dieMatrix (Mx) aus.
Abb. 23. Nach Platin-Kohle-Beschattung lässt sich die warzige Oberflächenstruktur der körnigen«Cuticularschicht» gut daIstellen.
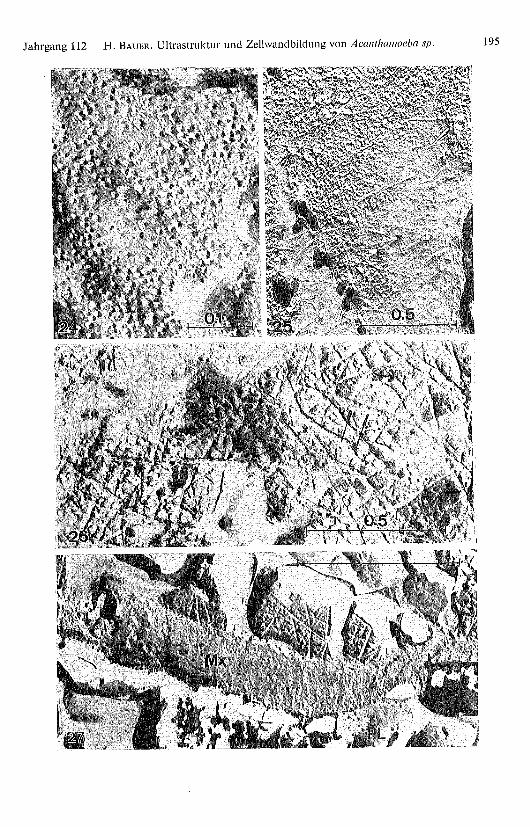
Tafel Seite 195
Abb. 24. Die globulären Plasmalemmapartikel liegen in der Matrix in statistischer Verteilung.
Abb. 25. Durch ständigen Nachschub der sich vom Plasmalemma lösenden Teilchen entstehtschliesslich ein dichtes Partikelfeld. Erst jetzt entstehen die ersten Cellulosefibrillen (F).
Abb. 26. In einer bestimmten, sehr kurzen Entwicklungsphase besteht eine direkte Verbindungder Plasmalemmapartikel mit den entstehenden Fibrillen (Pfeile). Zwischen den Fibrilten finden sichfreie Teilchen.
Abb. 27. Die überkreuzten, in der Matrix (Mx) eingebetteten Fibrillen sind zum Teil immer nochmit Partikeln besetzt. PL: Plasmalemma.
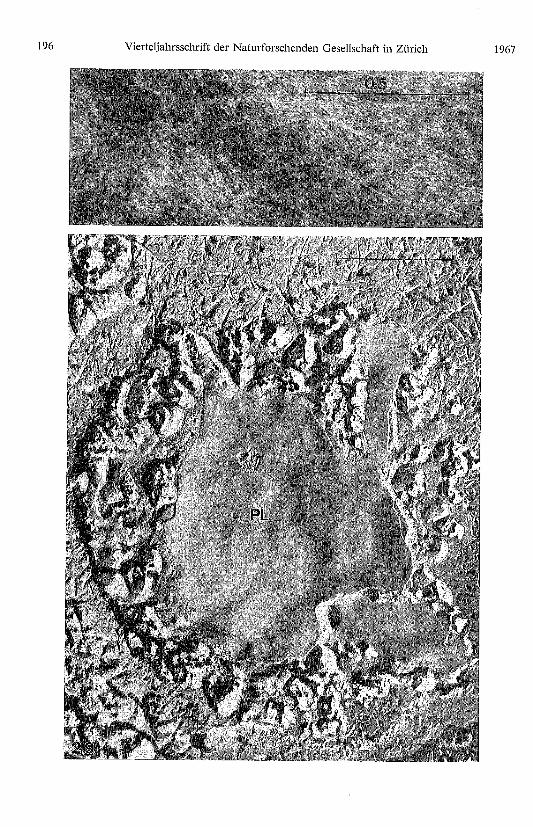
Tafel Seite 196
Abb. 28. Durch Ultraschall können die höheren Fibrillenverbände in einzelne 35-40 Å dickeElementarfibrillen aufgespatten und nach Natriumphosphorwolframsäure-Imprägnierung elektronen-optisch dargestellt werden.
Abb. 29. Vollständig encystierte Amöbe. Die anfängliche Streuungstextur der Fibrillen bleibterhalten. Auf dem Plasmalemma können nach Abschluss der Zellwandbildung keine globulären90-120 Å Partiket mehr beobachtet werden.
Tafel Seite 197
Abb. 30. Membranartige und vesikelförmige Strukturen in Aussparungen der Cystenwand.PL: Plasmalemma. Glutaraldehyd-Osmium-Fixierung.
Abb. 31. Auch mittels Gefrierätzung zeigen die Cystenwandeinschlüsse «unit membrane» Struk-tur.
Abb. 32. Nach Veraschung von Dünnschnitten encystierter Amöben bleiben in Aussparungen derZellwand nicht verbrennbare Rückstände übrig (Pfeile).
Jahrgang 112 H. BAUER. Ultrastruktur und Zellwandbildung von Acanthamoeba sp. 189
190 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1967
Jahrgang 112 H. BAUER. Ultrastruktur und Zellwandbildung von Acanthamoeba sp. 191

192 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1967
Jahrgang 112 H. BAUER. Ultrastruktur und Zellwandbildung von Acantha noeba sp. 193
194 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1967
Jahrgang 112 H. BAUER. Ultrastruktur und Zeltwandbildung von Acanthamoeba sp. 195
196 Vierteljahrsschrift der Naturforschenden Gesellschaft in Zürich 1967
Jahrgang 112 H. BAUER. Ultrastruktur und Zellwandbildung von Acanthmnoeba sp. 197














