Embed Size (px)
Citation preview

Aus der Außenstelle für Epidemiologie
der Tierärztlichen Hochschule Hannover in Bakum
Vergleichende Untersuchung zur Effektivität eines Kombinationsimpfstoffes gegen Haemophilus parasuis und
Mycoplasma hyopneumoniae und eines monovalenten Impfstoffes gegen Mycoplasma hyopneumoniae bei Schweinen
INAUGURAL-DISSERTATION
zur Erlangung des Grades einer DOKTORIN DER VETERINÄRMEDIZIN
(Dr. med. vet.)
durch die Tierärztliche Hochschule Hannover
Vorgelegt von Claudia Meistermann
aus Bakum
Hannover 2006

Wissenschaftliche Betreuung: Univ.-Prof. Dr. Th. Blaha 1. Gutachter: Univ.-Prof. Dr. Th. Blaha 2. Gutachter: Univ.-Prof. Dr. K.-H. Waldmann Tag der mündlichen Prüfung: 11. Juli 2006 Diese Arbeit wurde durch Mittel der Firma Fort Dodge, Würselen gefördert.
Für die Unterstützung dieses Versuchsvorhabens möchte ich mich bedanken.

für Johanna und Henrike


Inhaltsverzeichnis
1 Einleitung........................................................................................... 11
2 Literaturübersicht ............................................................................. 13
2.1 Haemophilus parasuis ...................................................................... 13
2.1.1 Allgemeine Eigenschaften................................................................... 13
2.1.2 Ätiologie und Pathogenese ................................................................. 14
2.1.2.1 Allgemeine Einordnung ....................................................................... 14
2.1.2.2 Morphologie ........................................................................................ 15
2.1.2.3 Wachstumsbedingungen..................................................................... 16
2.1.2.4 Biochemische Eigenschaften .............................................................. 18
2.1.2.5 Virulenzfaktoren .................................................................................. 18
2.1.3 Epidemiologie...................................................................................... 22
2.1.4 Klinische Symptomatik ........................................................................ 25
2.1.5 Pathologie ........................................................................................... 27
2.1.6 Diagnostik ........................................................................................... 28
2.1.7 Differentialdiagnosen .......................................................................... 34
2.1.8 Immunität ............................................................................................ 35
2.1.8.1 Immunantwort ..................................................................................... 35
2.1.8.2 Prophylaxe durch Impfung .................................................................. 36
2.1.9 Bekämpfung........................................................................................ 39
2.1.9.1 Erregerkontrolle / Erregereradikation .................................................. 39
2.1.9.2 Medikamentelle Therapie.................................................................... 40
2.1.10 Ko-Infektionen mit anderen Erregern des Schweines ......................... 41
2.2 Mycoplasma hyopneumoniae .......................................................... 43
2.2.1 Ätiologie und Pathogenese ................................................................. 43
2.2.2 Epidemiologie...................................................................................... 45
2.2.3 Klinische Symptomatik ........................................................................ 47
2.2.4 Pathologie ........................................................................................... 48
2.2.5 Diagnostik ........................................................................................... 49
2.2.6 Immunität ............................................................................................ 51

2.2.6.1 Immunantwort ..................................................................................... 51
2.2.6.2 Prophylaxe durch Impfung .................................................................. 53
2.2.7 Bekämpfung........................................................................................ 54
2.2.7.1 Erregerkontrolle................................................................................... 54
2.2.7.2 Medikamentelle Therapie.................................................................... 55
2.2.7.3 Erregereradikation............................................................................... 55
3 Material und Methoden ..................................................................... 57
3.1 Beschreibung des Versuchbetriebes................................................... 57
3.2 Infektionsstatus des Versuchsbetriebes.............................................. 60
3.3 Vakzinen ............................................................................................. 60
3.3.1 Versuchsvakzine ................................................................................. 60
3.3.2 Kontrollvakzine.................................................................................... 61
3.4 Versuchsgruppen ................................................................................ 61
3.5 Datenerfassung und –auswertung ...................................................... 63
3.5.1 Serologie ............................................................................................. 63
3.5.1.1 Untersuchung auf Antikörper gegen H. parasuis Serovar 4 und 5 ...... 64
3.5.1.2 Untersuchung auf Antikörper gegen M. hyopneumoniae .................... 64
3.5.2 Durchschnittliche tägliche Gewichtszunahme ..................................... 65
3.5.3 Futterverwertung ................................................................................. 65
3.5.4 Durchschnittliche Mastdauer ............................................................... 66
3.5.5 Mortalität ............................................................................................. 66
3.5.6 Therapeutische Maßnahmen / Tierbehandlungsindex (TBI) ............... 66
3.5.7 Beurteilung von Lungen nach der Schlachtung / Lungenscore ........... 67
3.5.8 AutoFOM-Daten .................................................................................. 68
3.6 Statistische Methoden......................................................................... 69
4 Ergebnisse......................................................................................... 70
4.1 Allgemeines......................................................................................... 70
4.2 Serologie ............................................................................................. 71
4.3 Durchschnittliche tägliche Gewichtszunahme ..................................... 74
4.4 Futterverwertung ................................................................................. 76
4.5 Durchschnittliche Mastdauer ............................................................... 77
4.6 Mortalität ............................................................................................. 77

4.7 Therapeutische Maßnahmen / Tierbehandlungsindex (TBI) ............... 80
4.8 Lungenscore ....................................................................................... 86
4.9 AutoFOM-Werte .................................................................................. 87
5 Diskussion ......................................................................................... 89
5.1 Allgemeines......................................................................................... 89
5.2 Serologie ............................................................................................. 89
5.3 Mastleistung ........................................................................................ 92
5.4 Mortalität ............................................................................................. 95
5.5 Therapeutische Maßnahmen / Tierbehandlungsindex (TBI) ............... 97
5.6 Lungenscore / AutoFOM-Werte .........................................................100
5.7 Schlussfolgerung................................................................................102
6 Zusammenfassung ..........................................................................104
7 Summary...........................................................................................106
8 Literaturverzeichnis .........................................................................108
9 Anhang..............................................................................................129

Verzeichnis der Abkürzungen A. Actinobacillus Abb. Abbildung AGPT Agargelpräzipitationstest AK Antikörper B. Bordetella BALF Bronchoalveoläre Lavageflüssigkeit ca. circa DCE DNA Cell Equivalence DNA Desoxyribonukleotid Acid E. Escherichia ELISA Enzym Linked Immunosorbent Assay EP Enzootische Pneumonie H. Haemophilus IFT Immunfluoreszenztest IHA Indirekte Haemagglutination ISTME Infektiöse septikämische thrombotisierende Meningo-Enzephalitis Kda Kilodalton kg Kilogramm km Kilometer KMZ Körpermassezuwachs LPS Lipopolysaccharide LT Lebenstag LW Lebenswoche M. Mycoplasma MHDCE Mycoplasma Hyopneumoniae DNA Cell Equivalence mg Milligramm mm Millimeter
m Mikrometer NAD Nikotinamid-Adenin-Dinukleotid NADP Nikotinamid-Adenin-Dinukleotid-Phosphat nm Nanometer OD Optische Dichte OMP Oberflächenmembranproteine P. Pasteurella PCR Polymerase Chain Reaction p.i. post infectionem PHV-1 Porcines Herpesvirus 1 PRCV Porcines Respiratorische Coronavirus PRRS Porcine Reproduction and Respiratory Syndrome Virus S. Streptococcus s.o. siehe oben

SEW segregated early weaning SPF spezifiziert-pathogen-frei St. Staphylococcus Tab. Tabelle TBI Tierbehandlungsindex u. und u. a. unter anderem z.B. zum Beispiel


Einleitung 11
1 Einleitung
In der modernen Schweineproduktion haben Erkrankungen des Respirationstraktes
neben den Erkrankungen des Magen-Darm-Traktes immer mehr an Bedeutung
gewonnen. Atemwegserkrankungen führen aufgrund von vermindertem Zuwachs,
schlechter Futterverwertung, erhöhter Mortalität und hohen Behandlungskosten zu
großen wirtschaftlichen Verlusten. Infektionen mit Haemophilus (H.) parasuis sind in
der Schweineproduktion weit verbreitet, wobei nicht nur die typische Polyserositis in
Form der Glässerschen Krankheit auftritt, sondern der Erreger zunehmend auch an
respiratorischen Erkrankungen beteiligt ist. Es lässt sich zudem seit einigen Jahren
eine weltweite Zunahme der Glässerschen Krankheit beobachten (LOPEZ et al.
2004; MÜLLER et al. 2004; OLIVEIRA u. PIJOAN 2004). Die Gründe hierfür
scheinen die intensiven Haltungsbedingungen, das Zusammenbringen von
Aufzuchtferkeln und Mastschweinen aus verschiedenen Herkünften und ein frühes
Absetzen der Ferkel zu sein.
Die Glässersche Krankheit kann in allen Betriebsarten auftreten, wobei Bestände mit
hohem Gesundheitsstatus (High-health- oder SPF-Betriebe) oftmals stärker betroffen
sind (RAPP-GABRIELSON 1999; VOS 2004).
Faktorenerkrankungen bzw. Krankheitsgeschehen eines gesamten Bestandes
stehen immer mehr im Vordergrund. Beim diagnostischen Vorgehen werden
demzufolge auch häufig mehrere Erreger gefunden, und es ist bedeutsam,
Ergebnisse richtig einzuschätzen und pathogene Erreger herauszufinden. Vor allem
bei Erkrankungen des Respirationstraktes scheinen Ko-Infektionen mit anderen
Erregern häufig zu Problemen zu führen.
Im Rahmen der Bekämpfung von Infektionskrankheiten bei Schweinen hat die
Immunprophylaxe innerhalb der vergangenen Jahre einen erheblichen Stellenwert
eingenommen. Die Anwendung von Impfstoffen ist ein erfolgreicher Weg zur
Reduzierung der Antibiotikaanwendungen. Eine gesicherte Diagnostik bezüglich
beteiligter Erreger und das Wissen um die Epidemiologie des Erregers ist jedoch
unabdingbare Voraussetzung für erfolgreiche immunprophylaktische Maßnahmen.
Aus ökonomischen und tierschützerischen Gründen kann es sinnvoll sein, zwei
Impfungen zu kombinieren, um die Belastung der Ferkel durch eine Vakzination zu
minimieren und die Arbeitsbelastung innerhalb eines Betriebes zu reduzieren.

12 Einleitung
Die vorliegende Untersuchung wurde mit dem Ziel durchgeführt, die Effektivität des
Kombinationsimpfstoffes Suvaxyn® M.hyo-Parasuis unter Feldbedingungen zu
prüfen. Durchgeführt wurde der Versuch im Raum Weser-Ems in einem
Sauenbestand mit eigener Ferkelaufzucht und anschließender Mast, in dem der
Erreger H. parasuis endemisch vorkommt. Die an der Studie beteiligten Tiere wurden
auf zwei Impfgruppen (Kombinationsimpfung gegen H. parasuis und M.
hyopneumoniae und Monoimpfung gegen M. hyopneumoniae) aufgeteilt. Die
Effektivität der Impfung wurde aus den Produktionsparametern während der Ferkel-
aufzucht und der Mast und dem Vorkommen von Lungenveränderungen zum
Zeitpunkt der Schlachtung abgeleitet. Die humorale Reaktion auf die Impfung wurde
anhand einer serologischen Verlaufsuntersuchung verfolgt.

Literatur 13
2 Literaturübersicht
2.1 Haemophilus parasuis
2.1.1 Allgemeine Eigenschaften
Haemophilus (H.) parasuis ist der Erreger der Glässerschen Krankheit.
Unter der Glässerschen Krankheit, auch Glässer`s disease, Transportkrankheit,
Glässersyndrom oder porcine polyserositis and arthritis bezeichnet, versteht man
eine fieberhafte, durch H. parasuis hervorgerufene Allgemeinerkrankung der
Schweine, die klinisch durch Pleuritis, Perikarditis, Peritonitis, Arthritis und Meningitis
gekennzeichnet ist.
Die Affinität zu den Schleimhäuten und Meningen ist stark ausgeprägt, so dass die
Bezeichnung als fibrinöse Serosen- und Gelenksentzündung das typische
Krankheitsbild und zugleich die pathologischen Veränderungen charakterisiert.
Die Glässersche Krankheit ist eine infektiöse Faktorenkrankheit, deren Auftreten und
Verlaufsform entscheidend im Zusammenhang mit der Bestandsgröße, dem
Infektionsdruck, den Temperaturschwankungen, dem Umstallungsstress und den
spezifischen immunologischen Abwehrreaktionen sowie bestimmten biologischen
Eigenschaften des Erregers stehen. Sie tritt in Abhängigkeit vom Wirken dieser
Einflussfaktoren sporadisch oder insbesondere in größeren Beständen enzootisch
mit einer hohen Morbidität auf. Der Erreger ist nur an das Schwein adaptiert.
Die Krankheit ist weltweit verbreitet. Es wird über Fälle in den USA und Kanada
(RADOSTITIS et al. 1963; RILEY et al. 1977; SMART 1987), Asien (ABE et al. 1982;
MORIKOSHI et al. 1990), Australien (PEET et al. 1983; EAVES et al. 1989;
BLACKALL u. PAHOFF 1995) und Europa (HJÄRRE 1957; LITTLE 1970; NIELSEN
u. DANIELSEN 1975; KIELSTEIN 1985) berichtet.
Die Erkrankung wurde erstmals von GLÄSSER (1910) als fibrinöse Pleuro-
Perikardio-Peritonitis beschrieben und von der sogenannten Schweineseuche
abgegrenzt. Wahrscheinlich wurde der Erreger das erste Mal von SCHERMER und
EHRLICHER 1922 isoliert. HJÄRRE und WRAMBY (1943) identifizierten den Erreger

14 Literatur
als Haemophilus suis, 1960 wurde er durch LECCE als Haemophilus influenza suis
beschrieben. Die Bezeichnung wurde dann in Haemophilus parasuis geändert, da
der Erreger im Gegensatz zu H. suis nicht den X-Faktor zum Wachstum benötigt,
sondern nur V-Faktor-abhängig ist (BIBERSTEIN u. WHITE 1969; KILIAN 1976).
Viele der zuerst als H. suis beschriebenen Isolate erwiesen sich später aufgrund
verbesserter Methoden als H.-parasuis-Stämme.
2.1.2 Ätiologie und Pathogenese
2.1.2.1 Allgemeine Einordnung
Die Gattung Haemophilus wird nach BERGEY`s Manual of Determinative
Bacteriology (1994) der Familie der Pasteurellaceae, den fakultativ anaeroben,
sporenlosen, unbeweglichen, gramnegativen Stäbchen zugeordnet.
Die Haemophilen bilden mit den Gattungen Actinobacillus und Pasteurella die
sogenannte HAP-Gruppe. Sie sind Schleimhautbesiedler und hinsichtlich ihrer
Kultivierungsbedingungen recht anspruchsvolle chemoorganotrophe Erreger.
Für die Angehörigen der Gattung Haemophilus ist eine enge Adaptation an die
Wirtsspezies charakteristisch. Bakterien dieser Gattung sind durch folgende
gemeinsame Kriterien charakterisiert (KILIAN u. BIBERSTEIN 1984):
- gramnegatives Färbeverhalten
- kokkoid- bis stäbchenförmig mit Neigung zu Pleomorphismus und
Filamentbildung, Größe 0,2-0,5 x 0,5-2,5 m
- Unbeweglichkeit
- aerobes oder fakultativ anaerobes Wachstum
- Abhängigkeit von bestimmten Wachstumsfaktoren
- Nitratreduktion
- Oxidase- und Katalasereaktionen variieren zwischen den Stämmen
- Fermentativer Abbau von Kohlenhydraten
- Vorkommen als Parasiten auf mukösen Membranen von Mensch und Tier
Literatur 15
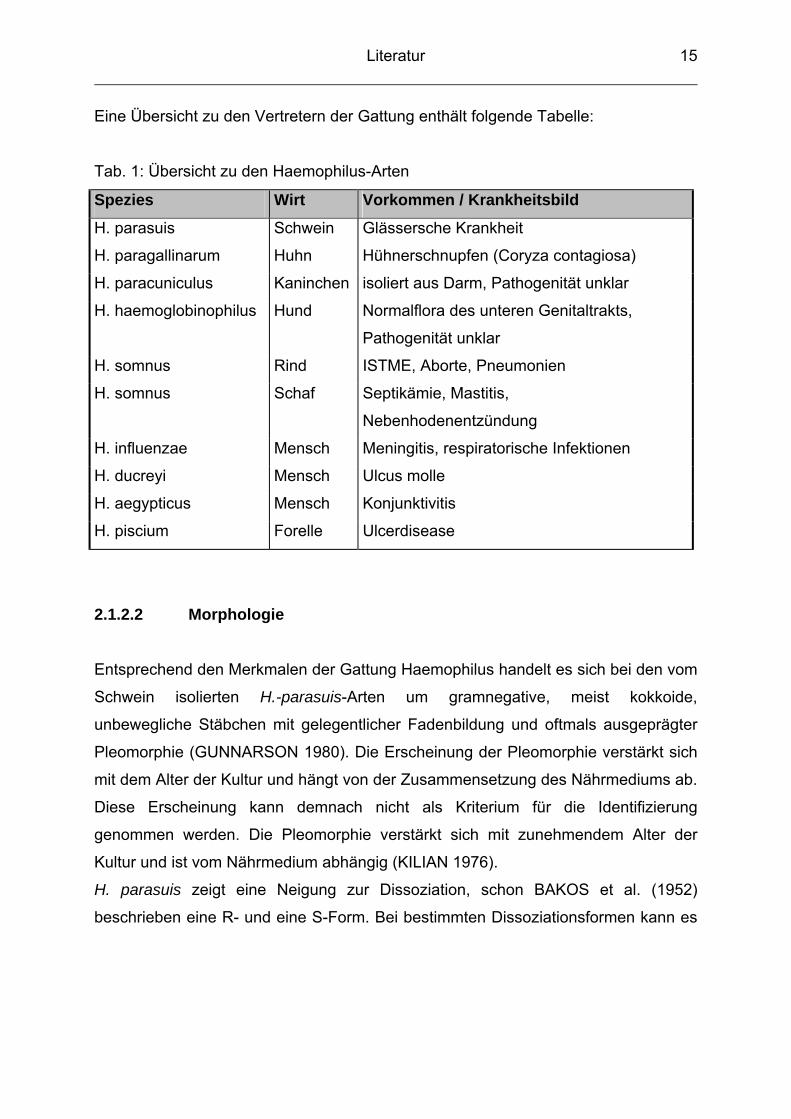
Eine Übersicht zu den Vertretern der Gattung enthält folgende Tabelle:
Tab. 1: Übersicht zu den Haemophilus-Arten
Spezies Wirt Vorkommen / Krankheitsbild
H. parasuis Schwein Glässersche Krankheit
H. paragallinarum Huhn Hühnerschnupfen (Coryza contagiosa)
H. paracuniculus Kaninchen isoliert aus Darm, Pathogenität unklar
H. haemoglobinophilus Hund Normalflora des unteren Genitaltrakts,
Pathogenität unklar
H. somnus Rind ISTME, Aborte, Pneumonien
H. somnus Schaf Septikämie, Mastitis,
Nebenhodenentzündung
H. influenzae Mensch Meningitis, respiratorische Infektionen
H. ducreyi Mensch Ulcus molle
H. aegypticus Mensch Konjunktivitis
H. piscium Forelle Ulcerdisease
2.1.2.2 Morphologie
Entsprechend den Merkmalen der Gattung Haemophilus handelt es sich bei den vom
Schwein isolierten H.-parasuis-Arten um gramnegative, meist kokkoide,
unbewegliche Stäbchen mit gelegentlicher Fadenbildung und oftmals ausgeprägter
Pleomorphie (GUNNARSON 1980). Die Erscheinung der Pleomorphie verstärkt sich
mit dem Alter der Kultur und hängt von der Zusammensetzung des Nährmediums ab.
Diese Erscheinung kann demnach nicht als Kriterium für die Identifizierung
genommen werden. Die Pleomorphie verstärkt sich mit zunehmendem Alter der
Kultur und ist vom Nährmedium abhängig (KILIAN 1976).
H. parasuis zeigt eine Neigung zur Dissoziation, schon BAKOS et al. (1952)
beschrieben eine R- und eine S-Form. Bei bestimmten Dissoziationsformen kann es

16 Literatur
zu einer Kapselbildung kommen, die z.B. durch eine Kapselfärbung sichtbar gemacht
werden kann.
Die Kapselbildung führt zu verschiedenen Kolonieformen (BAKOS 1955):
- Bei der M-Form (mukoid) sind die Kolonien groß und schleimig, trüb und
konfluierend, und weisen eine glänzende Oberfläche auf. Bakterien in der M-
Form besitzen eine vollständige Kapsel.
- Die S-Form (smooth) zeichnet sich durch kleine, feine, nicht konfluierende
Kolonien aus. Die S1-Form besitzt noch teilweise eine Kapsel, die S2-Form
besitzt keine Kapsel mehr.
- Kolonien der R-Form (rough) sind groß, mit teilweise gezackten Rändern. Sie
besitzen keine Kapsel.
Ein Wechsel von der M- in die S-Form, die relativ stabil ist, ist im Gegensatz zum
Wechsel von der S- zur R-Form irreversibel.
BAKOS (1952) stellte keinen wesentlichen Zusammenhang zwischen Herkunft der
Stämme und ihrer Morphologie fest. Im Gegensatz dazu kam MÜNCH (1993) zu dem
Schluss, dass Isolate von an Glässerscher Krankheit erkrankten Tieren auf
Kochblutagar hauptsächlich kleinere Kolonien ausbilden, hingegen bei Stämmen von
Tieren ohne klinische Erscheinungen gehäuft mittelgroße grau-weiße Kolonien
entstehen. Es wurde dabei kein Zusammenhang zwischen der Temperatur bei
Anzüchtung und der Koloniemorphologie festgestellt.
2.1.2.3 Wachstumsbedingungen
Hinsichtlich seiner Wachstumsbedingungen gehört H. parasuis zu den
anspruchsvollen Mikroorganismen.
Charakteristische Merkmale der Gattung Haemophilus sind die spezifischen
Wachstumsfaktoren (X und V), die bei einer aeroben Anzüchtung notwendig sind.
Diese Eigenschaft dient zugleich der Gattungsbestimmung und der Differenzierung
der einzelnen Hämophilusarten. Der X-Faktor ist das Protoporphyrin IX, in einigen

Literatur 17
Fällen auch das Eisen enthaltende Protohäm. Die Quellen des X-Faktors, der
thermostabil ist, sind Blut und Blutderivate einschließlich Hämin. H. parasuis ist in der
Lage Porphyrin aus δ-Aminolävulinsäure zu bilden, somit ist es von der exogenen
Häminzuführung (X-Faktor) unabhängig (BIBERSTEIN u. WHITE 1969). Einige
Haemophilusarten wie z.B. H. influenzae und H. haemoglobinophilus sind nicht zur
enzymatischen Umwandlung von δ-Aminolävulinsäure befähigt und benötigen eine
exogene Zufuhr von Hämin. Der V-Faktor wird als Nikotinamid-Adenin-Dinukleotid
(NAD) oder als NAD-Phosphat (NADP) beschrieben. Er ist thermolabil, im
Hefeextrakt und gering auch im Blut enthalten und für die Oxidationsvorgänge der
Zellen notwendig. H. parasuis ist zur Durchführung seiner Oxidoreduktionsprozesse
auf exogenes NAD, den V-Faktor, angewiesen. Dieser wird z.B. durch
Hitzebehandlung aus den Blutzellen freigesetzt, was in der Routinediagnostik in
Form des sogenannten Schokoladenagar zur Anwendung kommt (BAKOS 1955). Auf
Grund seines hohen NAD-Gehaltes hat sich hier vor allem Pferdeblut bewährt.
Weiterhin nutzt man die NAD-Überproduktion einiger Bakterienarten im sogenannten
Ammen- oder Satellitenphänomen.
Als Ammenkeime werden benutzt:
- Staphylokokken (NICOLET 1981)
- E. coli und Mikrokokken (NIELSEN u. DANIELSEN 1975)
- Pseudomonaden (SHIFRINE u. BIBERSTEIN 1960)
Nach ca. 24 Stunden sind neben der Amme winzige glasige, später gräulich
glänzende Kolonien erkennbar. Ihre Ausmaße verkleinern sich mit dem Abstand zur
Amme. Selektivmedien zur Unterdrückung von Begleitkeimen können verwendet
werden (CRAWFORD et al. 1969). LITTLE (1970) empfiehlt den Zusatz von
Bacitracin und Kristallviolett, und NICOLET (1981) verwandte Bacitracin und
Cloxacillin. NAD kann aber auch durch Zusetzen von Hefeextrakten zur Verfügung
gestellt werden (JORGENSEN et al. 1987). Der Abbau von NAD zu Nicotinamid
erfolgt bei H. parasuis über konventionelle Enzyme.
Als günstigste Bebrütungstemperatur werden 37°C angesehen. H. parasuis ist
mikroaerophil, sein Wachstum wird durch eine CO2-Spannung von 5-10 %
verbessert.

18 Literatur
2.1.2.4 Biochemische Eigenschaften
Neben den Wachstumsfaktoren ist eine weitere Differenzierung von H. parasuis
durch Untersuchung des Hämolyseverfahrens und des biochemischen Verhaltens
möglich. Nach KIELSTEIN (1990) ist H. parasuis eindeutig durch folgende
Eigenschaften charakterisiert: V-Faktorenabhängigkeit, fehlende Urease-,
Hämolysin- und Indolbildung sowie positive Katalase- und Maltosereaktion; die
Kohlenhydrate Arabinose, Lactose und Xylose werden nicht gespalten. In diesen
Eigenschaften bestehen keine Unterschiede zwischen Stämmen, die aus
verschiedenen Organen oder Tieren mit unterschiedlichem Krankheitsbild isoliert
wurden. Es wurde somit kein Zusammenhang zwischen Virulenz und Biochemie
festgestellt. Die Hämophilus-Art „Taxon C“ ist im Gegensatz zu allen anderen
Hämophilus-Arten schwer zu charakterisieren, da hierfür lediglich der Ausfall der
Maltose- und Arabinose-Reaktion herangezogen werden kann.
Die Stämme der sogenannten „minor group“ unterscheiden sich von H. parasuis in
der Ureasebildung (KILIAN 1976). Diesen Stämmen konnte bisher kein klinisches
Bild zugeordnet werden (BRANDRETH u. SMITH 1986; ROSENDAL et al. 1984).
2.1.2.5 Virulenzfaktoren
Über Virulenzfaktoren von H. parasuis ist noch wenig bekannt. In der Literatur wird
u. a. von der Kapsel, von Fimbrien, den Proteinmustern, wie Oberflächenmembran-
proteine (OMP), Lipopolysaccharide (LPS) und dem Polypeptidmuster, sowie der
Enzymausstattung als Virulenzfaktoren berichtet.
Kapsel Viele Bakterien sind von einem extrazellulären Material umgeben, das als Kapsel
bezeichnet wird. Diesen Strukturen kommt in erster Linie die Funktion der Adhäsion
an inerten und auch lebenden Oberflächen, der Adhäsion von Nährsubstraten, aber
auch einer Barrierefunktion zu. Weiterhin dienen Kapseln dem Schutz vor der

Literatur 19
Infektion mit Bakteriophagen, vor Phagozytose und anderen Clearance-
mechanismen.
Erste Untersuchungen über das Vorhandensein einer Kapsel bei H. parasuis wurden
schon früh von CHANDLER (1939) beschrieben. BAKOS (1955) brachte dies mit den
verschiedenen Kolonieformen in Verbindung. Der Kapselnachweis erfolgt neben
verschiedenen Färbemethoden (KIELSTEIN et al. 1992), der Präzipitation mit
Cetavlon und dem Hitzetest sowie der Iridiszenzmethode (MOROZUMI u. NICOLET
1986) vor allem mit Hilfe des Akriflavintests. Die Vielzahl der genutzten Methoden
und deren technische Grenzen bedingen vermutlich die widersprüchlichen Aussagen
über den Zusammenhang von Virulenz und Bekapselung. In verschiedenen Studien
wird über zwei Arten einer Hämophilusinfektion, die von der Kapselbildung abhängen
soll, berichtet. KILIAN (1984) unterscheidet bei H. influenzae zum einen eine akute,
pyogene und meist invasive Infektion mit H. influenzae als primär pathogenem
Erreger, wobei stets eine Kapsel vorhanden ist; zum anderen eine vorwiegend
chronische Infektion, in welcher H. influenzae scheinbar eine sekundäre Rolle spielt.
Diese Stämme sind unbekapselt und werden häufig auch bei gesunden Menschen
isoliert. LITTLE (1971) beschreibt ebenfalls eine solche Einteilung bei der H.-
parasuis-Infektion beim Schwein. KIELSTEIN und ROSNER (1992) konnten
unbekapselte Stämme aus Schweinen mit Glässerscher Krankheit statistisch
gesichert häufiger nachweisen als aus gesunden Schweinen. Sie sehen somit die
fehlende Bekapselung als einen Hinweis auf Virulenz von Feldisolaten.
Anheftungsfaktoren (Fimbrien) Fimbrien sind fädige Gebilde an der Bakterienoberfläche, die aus Proteinen
aufgebaut sind. Fimbrien, häufig auch als Pili oder F-Adhäsine bezeichnet, werden
von allen gramnegativen und einigen grampositiven Bakterien gebildet. Eine
Voraussetzung für das Entstehen von Infektionen ist die Haftung der Erreger an der
Schleimhaut. Diese fimbrienbedingte Adhäsionsfähigkeit wird durch die Testung des
Hämagglutinationsvermögens untersucht (GRUND et al. 1990). Die
Agglutinationsfähigkeit gegenüber einzelnen Tierspezies ist zwischen den Stämmen
sehr unterschiedlich (BAKOS 1955). MÜNCH (1993) beobachtete bei Pneumonie-

20 Literatur
Isolaten keine Agglutination von Hammelerythrozyten und stellte bei Isolaten aus
Fällen von Glässerscher Krankheit selten eine Agglutination von Pferde-, Kälber- und
Meerschweinchenerythrozyten fest. Unterschiedliche Hämagglutinationsmuster
beschreibt auch SCHROER (1992). Er fand im Gegensatz zu MÜNCH (1993) bei
Pulmonal- und Glässer-Stämmen eine hohe Agglutinationsrate gegenüber
Meerschweinchenerythrozyten. An frisch isolierten, auf Kochblutagar angezüchteten
H.-parasuis-Stämmen konnte SCHROER (1992) durch elektronenmikroskopische
Untersuchungen einen dichten Fimbrienrasen feststellen. Dieser ging nach
mehrmaliger Subkultivierung verloren.
Oberflächenmembranproteine (OMP) Die Zellwand gramnegativer Bakterien ist aus drei Schichten aufgebaut. Sie besteht
zum einen aus einer zytoplasmatischen Membran, die die äußere Begrenzung des
Zytoplasmas bildet und aktive Transportsysteme, Enzyme der Atmungskette sowie
des Zitratzyklus enthält (SCHNAITMAN 1970); zum zweiten aus einer dünnen
Peptidoglykanschicht, die zur Aufrechterhaltung der Form und der Stabilität der Zelle
dient; und drittens aus der für gramnegative Bakterien typischen äußeren Membran
(SELTMANN 1982). Die äußere Membran verleiht der Bakterienoberfläche eine
starke Hydrophilie und bietet Schutz vor Phagozytose und Komplementsystem
(DONALDSON et al. 1974). Außerdem stellt sie eine Permeabilitätsbarriere für viele
Antibiotika dar und ist für Prozesse des Stoffaustausches verantwortlich. Wichtige
Bestandteile der äußeren Membran sind die Außenmembranproteine (outer
membrane proteins OMP). Auf Grund der Menge ihres Vorkommens unterscheidet
man Haupt (major)- und Neben (minor)-Proteine. Sie sind für die Ausbildung von
Pathogenität und Virulenz von großer Bedeutung.
Lipopolysaccharide (LPS) Auch Lipopolysaccharide sind Bestandteil der äußeren Membran der Zellwand gram-
negativer Bakterien. Die LPS stellen eine Permeabilitätsbarriere gegenüber
bestimmten Arzneimitteln, Antibiotika oder Detergenzien dar, schützen die Zelle vor
den Abwehrmechanismen des Wirtes, insbesondere der Phagozytose, und können

Literatur 21
als Rezeptoren für Phagen dienen. Sie bestehen aus drei Regionen: 1. aus der O-
spezifischen Seitenkette, 2. der Kernzone und 3. dem Lipid A. Das Lipid A ist das
endotoxische Zentrum der LPS. Es wird bei Tod oder Replikation aus der
Bakterienoberfläche freigesetzt und kann dann im Wirt zirkulieren. Die LPS können
nach verschiedenen Methoden isoliert werden (WESTPHAL u. JANN 1965).
Polypeptidmuster Erste Angaben zum Polypeptidmuster von Hämophilusspezies sind von NEUMANN
und HINZ (1977) bekannt. Sie beobachteten sowohl zwischen als auch innerhalb
einer Bakterienspezies starke Differenzen im Proteinmuster und hielten es deshalb
für eine Speziesidentifizierung für ungeeignet. NICOLET et al. (1980) konnten jedoch
in der SDS-PAGE unter standardisierten Bedingungen reproduzierbare,
speziesspezifische Polypeptidmustertypen für A. pleuropneumoniae und H. parasuis
erstellen. Das Ergebnis wurde durch unterschiedliche Wachstumsbedingungen nicht
beeinflusst. Auch ZUCKER (1993) konnte keinen Einfluss auf die Ausprägung des
Polypeptidmusters durch Kultivierungszeit, -temperatur und Sauerstoffspannung
feststellen. ROSNER und KIELSTEIN (1991) konnten keine eindeutige Beziehung
zwischen den Proteinmustern von Ganzzell-Lysaten, dem Serotyp, der Virulenz
sowie der Kapselausbildung nachweisen.
Enzymausstattung
Neuraminidase ist ein potentieller Virulenzfaktor bei anderen Arten der
Pasteurellaceae. LICHTENSTEIGER und VIMR (1997) beobachteten eine
Produktion von Neuraminidase in über 90 % der Feldisolate. Dieses Enzym kann
gemeinsam mit den Enzymen Permease und Aldolase zur Virulenz beitragen, da es
Kohlenhydrate der Wirtszellen benötigt. Durch die neuraminidase-bedingte Abnahme
von Neuraminsäure kann es zu einer Demaskierung von Rezeptoren, die für die
Kolonisation und Invasion der Wirtszelle benötigt werden, kommen. Ebenso kann es
mit dem Abwehrsystem des Wirtes interferieren, indem die Viskosität des Schleimes
abnimmt (CORFIELD 1990; LICHTENSTEIGER u. VIMR 1997).

22 Literatur
2.1.3 Epidemiologie Seit einigen Jahren lässt sich weltweit eine deutliche Zunahme der Glässerschen
Krankheit beobachten (LOPEZ et al. 2004; MÜLLER et al. 2004; OLIVEIRA u.
PIJOAN 2004). Die Gründe hierfür scheinen die intensiven Haltungsbedingungen,
das Zusammenbringen von Aufzuchtferkeln und Mastschweinen aus verschiedenen
Herkünften sowie ein frühes Absetzen der Ferkel zu sein. Darüber hinaus kommen
weitere Erreger, wie das Virus des Porzinen Respiratorischen und Reproduktiven
Syndroms (PRRSV) oder das Porzine Circovirus Typ 2 (PCV-2), als Kofaktoren in
Betracht (OLIVEIRA et al. 2004; OLIVEIRA u. PIJOAN 2002, 2004). Die Glässersche
Krankheit kann in allen Betriebsarten auftreten, wobei Bestände mit hohem
Gesundheitsstatus (High-health- oder SPF-Betriebe) oftmals stärker betroffen sind
(RAPP-GABRIELSON 1999; VOS 2004). Bei Untersuchungen der verschiedenen
Erreger bei Lungenentzündungen in Norddeutschland wurde H. parasuis in einem
Drittel (31,3 %) der Fälle aus Lungen isoliert, im Bronchialtupfer konnte H. parasuis in
67,5 % der untersuchten Tiere nachgewiesen werden; in Nasentupfern (54 %) und in
bronchioalveolärer Lavage (56,1 %) war der Erreger bei über der Hälfte der
untersuchten Tiere zu finden (NIENHOFF 2005). Allerdings kann davon
ausgegangen werden, dass nicht alle Tiere auch eine H.-parasuis-Infektion
durchgemacht haben, da bei dieser Studie sämtliche zur Sektion anstehenden Tiere
eines Untersuchungsinstitutes beprobt wurden. Andere Studien berichten über die
Isolation von H. parasuis aus gesunden Schweinen (Kielstein et al. 1994; MØLLER u.
KILIAN 1990).
Unter experimentellen Bedingungen liegt die Morbidität bei 50 bis 75 % (teilweise
über 90 %) und die Mortalität bei etwa 10 % (VOS 2004; WIEGAND et al. 1992).
Die Übertragung der Infektion erfolgt aerogen. Nach NICOLET (1981) ist aber auch
der indirekte Infektionsweg, z. B. über Schleim, nicht auszuschließen.
Krankheitsfördernd sind Transport (daher auch die Bezeichnung „Transport-
krankheit“), Umstallen, Zusammenbringen von Tieren unterschiedlicher Herkunft,
Einstallung von Tieren aus H.-parasuis-negativen Beständen, Fütterungswechsel,
schlechtes Stallklima oder allgemeine Stresssituationen. Die Wahrscheinlichkeit,

Literatur 23
dass sich Ferkel von Jungsauen sehr früh infizieren, ist größer als bei pluriparen
Tieren, da im Kolostrum von Jungsauen häufig geringere Mengen an Antikörpern
gebildet werden und sie somit eine schlechtere Immunitätsausbildung haben (DONE
1999).
Infektionen mit anderen bakteriellen Erregern, wie Bordetella bronchiseptica,
begünstigen die Ansiedlung von H. parasuis in der Lunge (BROCKMEIER 2004).
Nach Ansiedlung im Nasen-Rachen-Raum und einer septikämischen Phase
verbreitet sich der Erreger im Organismus. Eine besondere Affinität besteht zu den
serösen Häuten.
H. parasuis kann im oberen Respirationstrakt bei neugeborenen Ferkeln schon
wenige Stunden nach der Geburt nachgewiesen werden (PIJOAN u. OLIVEIRA
2003).
Bei einer experimentellen Infektion (VAHLE et al. 1995) lässt sich der Erreger zwölf
Stunden p.i. in der Nase und 36 Stunden p.i. im Blut nachweisen. Vermehrte
Flüssigkeitsbildung im Peritoneum, in der Pleura und im Perikard zeigt sich zwölf
Stunden nach der Infektion. Nach 36 Stunden bilden sich in den Körperhöhlen
Fibrinfäden, und auch in den Gelenken lässt sich fibrinöses Exsudat finden.
Nach epizootiologischen Erhebungen in zahlreichen Schweinebeständen
unterschiedlicher Größenordnung beschreiben KIELSTEIN et al. (1986)
verschiedene Krankheitsverläufe der Glässerschen Krankheit: Zum einen treten
sporadische Erkrankungen einzelner Tiere oder ganzer Würfe auf, die häufig im
Zusammenhang mit dem Absetzen, dem Transport oder mit starken Stalltemperatur-
schwankungen stehen. Zum anderen treten akute Enzootien mit Befall mehrerer
Würfe oder mehrerer Tiere in Buchten oder ganzen Stalleinheiten auf, die meist nicht
mit erkennbaren Umweltbelastungen in Zusammenhang stehen und nach kurzen
Zeitperioden wieder erlöschen. Der Erreger ist im Bestand jedoch weiterhin
nachweisbar. Außerdem kommen immer wiederkehrende Enzootien mit einer hohen
Morbidität vor, die vor allem in der Saugferkel- und Läuferphase auftreten. Trotz
Einhaltung von Hygienestandards dauern diese Enzootien über Monate hinweg an
und erfordern einen hohen Behandlungsaufwand. Diese Verlaufsform ist nicht nur mit

24 Literatur
dem Auftreten von Stresssituationen erklärbar, es müssen vielmehr auch mögliche
Erregerpassagen mit Virulenzveränderungen, ein hoher Infektionsdruck sowie eine
unzureichende Immunitätsausbildung gerade in größeren Anlagen berücksichtigt
werden.
Zusätzlich zu diesen drei Verlaufsformen hat H. parasuis auch bei respiratorischen
Erkrankungen eine ätiologische Bedeutung, vor allem bei Pneumonien mit Pleuritis,
möglicherweise auch bei Rhinitiden.
Potentiell pathogene Stämme von H. parasuis können häufig auch von gesunden
Schweinen bzw. in gesunden Schweinebeständen von den Schleimhäuten des
Nasen-Rachenraumes und der Trachea isoliert werden (healthy carrier). Naive Tiere
können sich in der Aufzucht anstecken (PIJOAN u. OLIVEIRA 2003). Die
Glässersche Krankheit gehört somit in die Gruppe der infektiösen
Faktorenkrankheiten (KIELSTEIN et al. 1994).
Eine H.-parasuis-Infektion kann in relativ naiven Herden, wie z. B. SPF-Herden oder
in Herden mit SEW-Verfahren, wesentlich problematischer verlaufen als in
konventionellen Betrieben. PIJOAN und OLIVEIRA (2002) haben eine Hypothese
aufgestellt, um diesen Sachverhalt zu klären. In naiven Sauenherden verläuft
demnach die Kolonisation der Saugferkel mit dem Erreger sehr langsam. Wenn diese
Ferkel abgesetzt werden, ist nur eine geringe Anzahl der Tiere infiziert. In
konventionellen Herden verläuft die Infektion bei den Saugferkeln wesentlich
schneller, da es zu Kreuzinfektionen durch die Sauen und durch ältere Ferkel im
Bestand kommt. Ferkel, die isoliert aufgezogen werden, können sich nur durch
andere Ferkel der gleichen Gruppe infizieren. Dieses wird sehr langsam geschehen,
so dass bei den meisten Ferkeln die Infektion so spät stattfindet, dass kein
maternaler Schutz mehr vorhanden ist und die Krankheit so drastischer verläuft.
Nicht-pathogene Stämme, die häufig im oberen Respirationstrakt der Sauen
gefunden werden, besiedeln sehr schnell die Tonsillen, den Nasen-Rachenraum und
die Trachea der Saugferkel. Bei potentiell pathogenen Stämmen verläuft die
Besiedlung wesentlich langsamer, da diese Erreger wahrscheinlich bei säugenden
Sauen nur in geringer Menge vorhanden sind. Infolge der Ausbildung maternaler
Antikörper sind diese Ferkel weitgehend vor einer systemischen Infektion geschützt.

Literatur 25
Maternale Antikörper konnten von PIJOAN und OLIVEIRA (2003) bis zur sechsten
bis achten Lebenswoche nachgewiesen werden. Die Abnahme der maternalen
Antikörpertiter steht im Zusammenhang mit einer Zunahme der Mortalität, die
meistens in der vierten bis sechsten Woche nach dem Absetzen beginnt. In Herden
mit niedrigen maternalen Antikörpertitern beginnt das Krankheitsgeschehen schon
spät in der Saugferkelphase oder früh in der Aufzucht.
KIELSTEIN et al. (1994) haben jedoch in Infektionsversuchen herausgefunden, dass
bei Verwendung virulenter Stämme weniger die Infektionsdosis für die Pathogenese
der Infektion von Bedeutung ist, als vielmehr die individuellen Unterschiede in der
Infektionsabwehr und die für das einzelne Individuum unterschiedlich belastenden
Umweltfaktoren. So fanden sie heraus, dass die aerogene Übertragung grundsätzlich
möglich ist. Jedoch wurden die pathologisch-anatomischen Organveränderungen
nach intratrachealer Applikation nicht beeinflusst, die Ferkel waren also in der Lage,
auch sehr hohe Infektionsdosen abzuwehren, was auf eine hohe Abwehrkapazität
der Lunge eines gesunden Ferkels hinweist.
2.1.4 Klinische Symptomatik
Erkrankungen verursacht durch H. parasuis sind in der Schweineproduktion weit
verbreitet, wobei nicht nur die typische Polyserositis auftritt, sondern der Erreger
zunehmend auch an respiratorischen Erkrankungen beteiligt ist. Am stärksten
betroffen sind Ferkel vier bis sechs Wochen nach dem Absetzen oder Läufer kurz
nach Einstallung in die Mast (OLIVEIRA u. PIJOAN 2002; ZIMMERMANN et al.
2004).
Die Inkubationszeit beträgt fünf bis sieben Tage (BAEHLER et al. 1974), bei
experimenteller Infektion nur 24 Stunden (NEIL et al. 1969; JANETSCHKE et al.
1977). Die Infektion kann verschiedene Verlaufsformen nehmen.
Bei akutem Verlauf sind Apathie, Anorexie und Temperaturerhöhung bis 42,5°C zu
beobachten. Plötzliche Todesfälle können in Verbindung mit einer Septikämie
vorkommen.

26 Literatur
Die typische Verlaufsform der fibrinösen Polyserositis, Polyarthritis und fibrinös-
purulenten Meningitis geht mit Inappetenz einher. Auffällige klinische Symptome
hängen von der Lokalisation der entzündlichen Veränderungen ab. Es kann zu
Lahmheiten, umfangsvermehrten Gelenken, kyphotischer Rückenlinie,
Schmerzäußerungen sowie zentralnervösen Symptomen, wie Inkoordination, Zittern,
Festliegen und Ruderbewegungen teilweise in Seitenlage, kommen (RAPP-
GABRIELSON 1999; VOS 2004; ZIMMERMANN et al. 2004). Bei Befall der
Meningen kommt es außerdem zu Krampfanfällen und Nystagmus (LITTLE u.
HARDING 1971; KIELSTEIN 1985).
Palpatorisch sind Füllung und Fluktuation der Gelenke, insbesondere der
Tarsalgelenke, diagnostizierbar (RITZMANN u. HEINRITZI 2005).
In schweren Fällen können Zyanosen an Ohren, Rüsselscheibe und Gliedmaßen
sowie Konjunktivitiden auftreten (ZIMMERMANN et al. 2004).
Pneumonische Erscheinungen, wie Husten, Niesen, Dyspnoe und verstärkte
abdominale Atmung, treten gleichzeitig oder als separates Krankheitsbild auf. Die
Auskultation der Lunge ergibt häufig Reibegeräusche.
Als seltene klinische Symptomatik wurden eine Panniculitis an den Ohren sowie eine
Myositis und Fasciitis im Bereich der Nackenmuskulatur beschrieben (HOEFLING
1991), außerdem können Aborte bei Sauen und chronische Lahmheiten bei
Zuchtebern vorkommen (RAPP-GABRIELSON 1999).
Der milde Verlauf mit Husten, Dyspnoe oder Lahmheiten geht oft in ein chronisches
Stadium, gekennzeichnet durch Gewichtsverlust, rauem Haarkleid und Kümmern
über (RITZMANN u. HEINRITZI 2005).
Bei älteren Schweinen und in endemisch infizierten Betrieben kann die klinische
Symptomatik auf den Respirationstrakt beschränkt bleiben (OLIVEIRA u. PIJOAN
2002).

Literatur 27
2.1.5 Pathologie
Bei der Sektion der meist gut genährten Tiere finden sich gummiartige Ausgüsse der
Gelenke und gelblich-trübe Flüssigkeitsansammlungen im Pleural- und
Peritonealraum, die serösen Häute sind mit einer bis zu 1,5 mm dicken Fibrinschicht
bedeckt. Außerdem lassen sich katarrhalisch-eitrige Bronchopneumonien finden.
TARASENOK (1982) stellte während einer H.-parasuis-Infektion in einem
Großbestand umfassende pathologisch-anatomische Untersuchungen an. Bei der
Sektion von 26 bis 106 Tage alten Läufern wurden an diesen eine serofibrinöse
Pleuritis, Perikarditis und Peritonitis festgestellt. Arthritiden traten relativ selten auf,
bei der akuten Form wurde vermehrt synoviale Flüssigkeit festgestellt, gleichzeitig
fand man große Mengen serösen Exsudates im Perikard, in der Pleura und im
Peritoneum. Im Gegensatz dazu wurde bei einem subakuten und chronischen
Krankheitsverlauf weniger Exsudat, aber mehr Fibrin festgestellt. Auch die Arthritiden
kamen hierbei häufiger vor.
Bei der akuten Verlaufsform findet man bei der pathologischen Untersuchung
makroskopisch eine fibrinöse Pleuritis, Perikarditis, Peritonitis und Polyarthritis sowie
eine purulente Meningoencephalitis. Neben vorwiegend fibrinösen und serofibrinösen
Ergüssen kommen auch eitrige Verlaufsformen vor (NICOLET 1981).
In selteneren Fällen kann eine Infektion zu einem akuten septikämischen Geschehen
führen, wobei es zu einer Zyanose, subkutanem und pulmonalem Ödem und
plötzlichem Tod ohne die typischen serösen Entzündungen kommen kann (RILEY et
al. 1977; PEET et al. 1983).
Beschrieben wurden auch seltene Fälle von Fasciitis und Myositis (HOEFLING
1991), sowie eine purulente Rhinitis (VAHLE et al. 1995).
JANETSCHKE et al. (1977) führten Untersuchungen zur Pathologie und Histologie
einer experimentellen H.-parasuis-Infektion bei SPF-Ferkeln durch. Die verendeten
bzw. am 7. bis 10. Tag p.i. gemerzten Tiere zeigten eine katarrhalisch-eitrige
Bronchopneumonie, wobei häufig nur lobuläre Bezirke betroffen waren. Gleichzeitig
traten oft charakteristische Pleuraveränderungen in Form von fibrinöser bis fibrinös-
eitriger Pleuritis in großen Teilen der Brusthöhle auf. Zusammenfassend stellten die

28 Literatur
Autoren fest, dass die Lungenveränderungen bei experimenteller Infektion mit H.
parasuis sehr variabel und nicht charakteristisch sind, was vermutlich darauf
zurückzuführen ist, dass nicht immer die gleichen Isolate verwendet wurden.
SOLANO-AGUILAR et al. (1999) beschreiben eitrig-katarrhalische Broncho-
pneumonien, die vor allem in den kranialen und medialen Lungenlappen lokalisiert
sind.
Mikroskopisch sieht man eine fibrinöse Exsudation mit geringgradiger Beteiligung
neutrophiler Granulozyten und einer geringen Anzahl Makrophagen (VAHLE et al.
1995). Im Bereich der Hirn- und Rückenmarkhaut lassen sich dagegen hochgradig
neutrophile Granulozyten finden. Die Fibrinbeteiligung ist im Zentralen Nervensystem
gering- bis mittelgradig.
Sind Husten, Atembeschwerden und hochgradige, periphere Kreislaufstörungen am
Krankheitsgeschehen beteiligt, findet man eine hochgradige Zyanose, sowie Lid- und
Ohrödem. Hirn- und Rückenmarkhäute lassen schon makroskopisch eine sulzige
Imbibition neben fibrinös-eitrigen Auflagerungen erkennen.
Bei der chronischen Verlaufsform kommt es zur adhäsiven Pleuritis, Perikarditis und
Peritonitis. Mikroskopisch findet man korrespondierende fibröse Synechien (SCHULZ
1991).
Wird H. parasuis aus der Schleimhaut gesunder Tiere isoliert, kommt es trotzdem zu
einer Reduzierung der Anzahl der Zilien, man findet neutrophile Granulozyten und
Plasmazellinfiltrationen (VAHLE et al. 1997).
2.1.6 Diagnostik
Die Diagnosestellung der Glässerschen Krankheit erfolgt auf der Grundlage der
Anamnese, klinischer Symptome und pathologischer Untersuchungen. Erste
Hinweise ergeben sich aufgrund des klinischen Bildes sowie anhand der
makroskopisch sichtbaren pathologisch-anatomischen Veränderungen. Diese
umfassen eine serofibrinöse bis fibrino-purulente Entzündung der serösen Häute in

Literatur 29
Form von Polyarthritis, Peritonitis, Pleuritis mit katarrhalisch-eitriger
Bronchopneumonie sowie Perikarditis und Meningoenzephalitis.
Der Nachweis erfolgt durch eine mikrobiologische Untersuchung. Hierfür eignen sich
Tupfer von fibrinösen Auflagerungen, Bauchhöhlenflüssigkeit, Synovia oder Liquor.
Trotz typischer klinischer Symptomatik und entsprechender pathologisch-
anatomischer Veränderungen lässt sich H. parasuis aufgrund seiner Empfindlichkeit
und der häufigen Überwucherung durch andere Keime nicht immer nachweisen
(RAPP-GABRIELSON 1999; VOS 2004). Bei antibiotisch vorbehandelten Tieren
gelingt der Nachweis nicht (OLIVEIRA u. PIJOAN 2002). Auch andere Faktoren, wie
z. B. Chronizität der Erkrankung, nehmen Einfluss auf die Nachweisrate. Sie
verbessert sich, wenn Material von frisch euthanasierten, unbehandelten Schweinen
statt von verendeten Tieren gewonnen wird. Idealerweise werden Proben von
Perikard, Pleura, Peritoneum, Gelenken und aus Meningen genommen (OLIVEIRA
2004). Eine Isolierung des Erregers aus dem Respirationstrakt lässt nicht unbedingt
auf eine systemische Infektion rückschließen, da H. parasuis im oberen
Respirationstrakt auch als kommensaler Besiedler vorkommen kann (OLIVEIRA
2004). Die Isolierungshäufigkeit nimmt parallel zum zeitlichen Abstand vom Eintritt
des Todes des Tieres bis zur Untersuchung ab (KIELSTEIN et al. 1984).

30 Literatur
Abb. 1: Diagnostisches Vorgehen bei Verdacht auf Glässersche Krankheit (nach
RITZMANN und HEINRITZI 2005)
Eine Charakterisierung von H. parasuis kann durch die Unterscheidung in Serotypen
und Genotypen erfolgen. Dazu sind verschiedene Verfahren beschrieben worden.
Im Allgemeinen werden die serologischen Tests als widersprüchlich und ungenau
eingeschätzt (DONE 1999).
In der Literatur sind verschiedene Methoden zur serologischen Typisierung von H.
parasuis beschrieben, die in Abhängigkeit vom verwendeten Testsystem und Antigen
zu unterschiedlichen Einteilungen führen.
Von BAKOS et al. (1952) wurde eine serologische Typisierung von H. parasuis in die
Serovare A, B, C und D auf der Grundlage der Langsamagglutination (LA) entwickelt.
Als Antigen für diesen Test werden Ganzzellen aus Bakteriensuspension sowie
Antiseren gegen diese Ganzzellen verwendet. Die vier Bakos-Typen wurden durch
SCHIMMEL et al. (1985) um weitere drei Serovare ergänzt, die Jena 1, 2 und 3
genannt werden.

Literatur 31
Das Klassifikationssystem von MOROZUMI und NICOLET (1986a) beruht auf der
Verwendung hitzestabiler löslicher Bakterienextrakte sowie serotypspezifischer
Kaninchenhyperimmunseren im Agargelpräzipitationstest (AGPT). Sie konnten so
weitere sieben Serovare beschreiben, die von KIELSTEIN (1991) um weitere sieben
Gruppen sowie von RAPP-GABRIELSON und GABRIELSON (1990) um weitere vier
nicht näher bezeichnete Serovare ergänzt wurden. Mit beiden genannten
Serotypisierungsverfahren werden unterschiedliche Antigenstrukturen erfasst, so
dass nur teilweise Zusammenhänge zwischen den Bakos- und den Morozumi-
Nicolet-Typen besteht. Nach vergleichenden Untersuchungen von KIELSTEIN und
RAPP-GABRIELSON (1992) können somit 15 Serovare unterschieden werden.
Dieses Kielstein-Rapp-Gabrielson-Schema, welches auf hitzestabilen Antigenen
getestet mit dem Agargelpräzipitationstest (AGPT) basiert, ist der international
anerkannte Test zur Serotypisierung von H. parasuis. Die Serovare variieren in ihrer
Pathogenität und treten regional unterschiedlich auf. In den meisten Ländern
dominiert Serovar 5, gefolgt von Serovar 4 und 13. Ein hoher Anteil an
nachweisbaren H.-parasuis-Isolaten lässt sich jedoch nicht typisieren. Das gleiche
Vorkommen von mehreren Serovaren in einem Bestand und sogar in einem Tier ist
möglich (KIRKWOOD et al. 2001; RAPP-GABRIELSON 1993 u. 1999).
Allerdings wurde in allen Studien ein sehr hoher Anteil nicht typisierbarer Serotypen
festgestellt. In den USA und Kanada lag dieser Anteil bei 15 % (RAPP-
GABRIELSON u. GABRIELSON 1992), in Deutschland bei 26 % (KIELSTEIN u.
RAPP-GABRIELSON 1992), in Spanien bei 29 % (RUBIES et al. 1999) und in
Australien bei 41 % (RAFIEE u. BLACKALL 2000). Dieser hohe Anteil nicht
typisierbarer Serotypen zeigt, dass die Möglichkeit einer noch höheren Anzahl von
Serotypen gegeben ist (KIELSTEIN u. RAPP-GABRIELSON 1992; RAFIEE u.
BLACKALL 2000).
H.-parasuis-Stämme aus dem oberen Respirationstrakt sind meist von geringer
Pathogenität und können auch bei klinisch gesunden Schweinen isoliert werden. Als
apathogen werden die Stämme 3, 6, 7, 8, 9 und 11 eingeschätzt (KIELSTEIN et al.
1992; OLIVEIRA u. PIJOAN 2002).

32 Literatur
Eine weitere Methode zur Serotypisierung von H.-parasuis-Isolaten ist die indirekte
Haemagglutination (IHA), bei der erhitzte Bakterienzellen als Antigen für
Schaferythrozyten verwendet werden (MINIATS et al. 1991a). Häufig wurden
negative und sehr variable Ergebnisse erzielt, und die Autoren MINIATS et al. (1991)
halten die IHA für unzuverlässig und für die Serotypisierung ungeeignet. KHYALI und
MITTAL (2002) konnten dagegen im Vergleich zur Immundiffusion, bei der etwa 30 %
der Isolate nicht typisierbar war, mit der indirekten Haemagglutination 90 % der
Isolate typisieren, ohne dass Kreuzreaktionen aufgetreten sind. DEL RÍO et al.
(2003) konnten mit der indirekten Haemagglutination 91 % der Isolate serotypisieren,
mit der Immundiffusion 63 %. Sie empfehlen die IHA als nützliche Methode zur
sensitiven und spezifischen Serotypisierung von H. parasuis.
Die Serotypisierung von H. parasuis kann außerdem mit dem Koagglutinationstest
durchgeführt werden. DEL RÍO et al. (2003) beschreiben den Koagglutinationstest
als einfach, spezifisch und sensitiv, allerdings werden häufig Kreuzreaktionen
beobachtet, dass er nicht zur Serotypisierung für H. parasuis empfohlen wird.
Für den Nachweis von Antikörpern gegen H. parasuis in Serum oder
Kolostrumproben können ELISA-Verfahren (Enzyme Linked Immunnosorbent Assay)
angewendet werden (MINIATS et al. 1991a; SOLANO-AGUILAR et al. 1999).
MÜLLER (2004) hat für seine Untersuchungen einen indirekten spezifischen ELISA
zum Antikörpernachweis gegen H. parasuis Serotyp 5 verwendet. Extrakte dieses
Agens wurden als Antigen genutzt. SOLANO-AGUILAR et al. (1999) benutzten einen
ELISA, der als Antigen formalininaktivierte Bakterienzellen nutzte, um maternale
Antikörpertiter und die humorale Antwort der Ferkel nach einer Impfung zu testen.
Sie fanden bei den Ferkeln schon im Alter von fünf Tagen eine Antikörperbildung als
Reaktion auf die Impfung.
Eine weitere Methode zum Antikörpernachweis ist der Komplementfixationstest.
NIELSEN (1993) konnte mit diesem Verfahren innerhalb einer Woche zirkulierende
Antikörper nach einer Infektion mit H. parasuis nachweisen, aber auch bei dem
Komplementfixationstest kommt es zu Kreuzreaktionen zwischen den einzelnen
Serotypen. TAKAHASHI et al. (2001) benutzten diesen Test, um Antikörpertiter bei

Literatur 33
einem Infektionsversuch zu bewerten. Sie fanden Antikörpertiter 19 Tage nach der
zweiten Vakzination.
Neben den verschiedenen Serotypisierungsverfahren kann zur Identifizierung von H.
parasuis auch eine Genotypisierung durchgeführt werden.
Von OLIVEIRA et al. (2001) wurde ein PCR-Verfahren (Polymerase Chain Reaction)
angewendet, um die Genauigkeit und Schnelligkeit in der H.-parasuis-Diagnostik zu
verbessern. Mit Hilfe der Polymerasekettenreaktion ist es möglich,
Nukleotidsequenzen enzymatisch zu amplifizieren. Das Prinzip der PCR beruht auf
der Fähigkeit von bestimmten zellulären Enzymen, sogenannten Polymerasen, DNA
zu verdoppeln. Die hierfür verwendeten Primer waren hoch spezifisch für den
Nachweis von H. parasuis. Die PCR kann auch im Fall eines negativen
bakteriologischen Ergebnisses herangezogen werden, da Antigene sowohl aus
Bakterienkulturen als auch direkt aus Probenmaterial nachgewiesen werden können
(OLIVEIRA u. PIJOAN 2004). Als Material für die PCR dienen veränderte Organe,
Synovia, Liquor oder Bronchoalveoläre Lavageflüssigkeit (BALF). Die PCR gilt als
gute Screeningmethode zur Beurteilung der einzelnen Betriebe.
Als Modifikationen der konventionellen PCR wurden die Enterobacterial Repetitive
Intergenic Consensus (ERIC)-PCR, die nested-PCR, die Real-time PCR und die
Amplifikationsfragmentlängenpolymorphismus (AFLP)-PCR entwickelt.
Die ERIC-PCR ist eine molekularbiologische Methode, mit deren Hilfe man
stammspezifische Fingerabdrücke produzieren kann, die den Vergleich und die
Differenzierung der verschiedenen Stämme ermöglichen (OLIVEIRA u. PIJOAN
2001). Sie erlaubt Untersuchungen zur Epidemiologie von H. parasuis innerhalb und
zwischen den Herden und verbessert die Charakterisierung der verschiedenen H.-
parasuis-Isolate. In Studien konnte nachgewiesen werden, dass erhebliche
genetische Unterschiede innerhalb der verschiedenen Serovare vorliegen (OLIVEIRA
et al. 2004). OLIVEIRA et al. (2004) haben die Reproduzierbarkeit der Methoden der
AFLP- und der ERIC-PCR miteinander verglichen und sind zu dem Ergebnis
gekommen, dass beide Verfahren gute Methoden zur Genotypisierung sind. Der
Nachteil der ERIC-PCR ist allerdings die schlechte Reproduzierbarkeit der

34 Literatur
Ergebnisse. Die Ergebnisse der AFLP sind abhängig von den benutzten
Restriktionsenzymen und den Parametern der Amplifikation.
Bei der nested-PCR wird DNA aus formalinfixiertem, in Paraffin eingebettetem
Gewebe nachgewiesen. Dieser Nachweis ist für die Routinediagnostik wichtig, da auf
diese Weise viele Proben aufbereitet werden können.
Eine weitere Alternative der H.-parasuis-Diagnostik ist die Immunhistochemie. H.
parasuis konnte aus Proben von experimentell infizierten Schweinen nachgewiesen
werden. Ebenso wird der Nachweis von inaktivierten Erregern aus dem Zytoplasma
phagozytierender Zellen ermöglicht (AMANO et al. 1994; SEGALES et al. 1997). Es
können allerdings Kreuzreaktionen mit A. pleuropneumoniae auftreten, wenn
polyklonale Antikörper für den Nachweis verwendet werden.
2.1.7 Differentialdiagnosen
Differentialdiagnostisch von einer H.-parasuis-Infektion abzugrenzen sind alle
bakteriellen Erreger, die eine septikämische Bakteriämie verursachen, wie z.B.
Streptococcus suis, Erysipelothrix rhusiopathiae, Actinobacillus suis, Salmonella
Choleraesuis und Escherichia coli.
Häufig sind plötzliche Todesfälle der einzige Hinweis auf ein Erkrankungsgeschehen.
In diesem Fall müssen die Ödemkrankheit, verursacht durch E. coli, und
Meningitiden durch S. suis ausgeschlossen werden.
Die Symptome einer Polyserositis müssen von einer Mycoplasma-hyorhinis-Infektion
und die einer Polyarthritis von einer Mycoplasma-hyosynoviae-Infektion abgegrenzt
werden. Purulente Arthritiden kommen aufgrund von Streptokokken oder
Arcanobacter pyogenes in Betracht. Klinisch sind diese Krankheitsbilder kaum zu
unterscheiden. Mycoplasmen sind ähnlich schwer anzuzüchten wie H. parasuis, so
dass bei negativem bakteriologischem Befund eine Untersuchung mittels PCR
angezeigt ist.
Bei Verlauf mit zentralnervösen Erscheinungen müssen insbesondere
Streptokokken-Meningitis und Enterotoxämie berücksichtigt werden. S. suis kann aus

Literatur 35
Liquor oder bei euthanasierten Tieren aus dem Gehirn isoliert werden. Die
Enterotoxämie verläuft ohne Gelenkbeteiligung, klinisch fallen Lidödeme auf.
Ist am Krankheitsgeschehen eine eitrige Bronchopneumonie beteiligt, müssen alle
bakteriellen und viralen Erreger, die an respiratorischen Erkrankungen beteiligt sein
können, ausgeschlossen werden (RAPP-GABRIELSON 1999).
2.1.8 Immunität
2.1.8.1 Immunantwort
Auf Grund der epitheliochorialen Plazentation der Sau kommen die Ferkel praktisch
gammaglobulinfrei zur Welt. Sie sind auf den passiven Immunglobulintransport via
Kolostrum in den ersten Stunden nach der Geburt angewiesen. Hinsichtlich des
Gehaltes und der Zusammensetzung der Gammaglobuline in der Sauenmilch sind
Schwankungen zwischen den Individuen, der Laktation und dem Sauenalter
beobachtet worden (KLOBASA u. BUTLER 1987). PIJOAN und OLIVEIRA (2003)
haben maternale Antikörper bei Ferkeln noch zwischen der sechsten und achten
Lebenswoche nachgewiesen. Die Immunität, die über die kolostralen Immunglobuline
vakzinierter Sauen an die Ferkel übertragen wird, ist serovarspezifisch (RAPP-
GABRIELSON et al. 1997).
Die maternale und die natürliche Immunität sind kritische Faktoren bei der Kontrolle
einer H.-parasuis-Infektion (NIELSEN u. DANIELSEN 1975, SCHIMMEL et al. 1992).
Die Antikörperbildung, also die humorale Immunität ist wegen des septikämischen
Verlaufs der Erkrankung wahrscheinlich der Hauptfaktor im Immunmechanismus
(RAPP-GABRIELSEN 1999).
Die antigenen Eigenschaften von H. parasuis sind in verschiedenen Untersuchungen
der Immunantwort gegen phänotypische Marker, wie OMP, LPS und Kapselprotein,
getestet worden. MINIATS et al. (1991b) benutzten einen Immunoblotassay, um die
Beziehung zwischen der humoralen Antwort geimpfter Ferkel und dem Schutz durch
die Impfung zu beurteilen. Sie fanden heraus, dass nur die Anwesenheit von

36 Literatur
Antikörpern gegen OMP in Zusammenhang mit einem Schutz gegen den Challenge
steht. Geimpfte Tiere, die einen vollen Schutz gegen den Challenge entwickelten,
hatten keine Antikörper gegen LPS oder Kapselproteine. RAPP-GABRIELSON et al.
(1997) demonstrierten, dass unterschiedliche Stämme desselben Serovars einen
unterschiedlichen Schutz gegen einen homologen Challenge ausbilden können,
obwohl sie die gleichen OMP- und LPS-Profile besitzen. Die Serovare 1 bis 7 haben
ähnliche antigene Eigenschaften (MINIATS et al. 1991b). Insgesamt stehen diese
antigenen Eigenschaften nicht im Zusammenhang mit dem Schutz gegen einen
heterologen Challenge.
2.1.8.2 Prophylaxe durch Impfung In der Literatur wird über sehr unterschiedliche Ergebnisse in Infektions- und
Vakzinationsversuchen mit H. parasuis berichtet. Ein Grund hierfür könnte in den
verschiedenen Ausgangsbedingungen in Bezug auf Patientenmaterial,
Infektionsstamm und Infektionsweg liegen.
Es können kommerzielle oder autogene Impfstoffe zur Kontrolle der H.-parasuis-
Infektion eingesetzt werden.
SMART und MINIATS (1989) haben einen Totimpfstoff mit drei Feldstämmen
getestet und erzielten bei geimpften Ferkeln einen Impfschutz, während nicht
vakzinierte Tiere nach Belastungstest mit H.-parasuis-Aerosol erkrankten. Eine
ähnliche Studie (MINIATS 1991a) zeigt ebenso den homologen Schutz bei SPF-
Ferkeln mit demselben Impfstoff.
Bei immunnaiven gnotobiotischen Ferkeln von geimpften Sauen konnte ein Schutz
gegen die homologen Serovare nachgewiesen werden (SOLANO-AGUILAR et al.
1999).
Ein heterologer Schutz gegen Stämme und Serovare wird als sporadisch und
unbeständig beschrieben (RAPP-GABRIELSON et al. 1997, BAK u. RIISING 2002).
RAPP-GABRIELSON et al. (1997) haben in einer Studie die Ausbildung von
Kreuzimmunitäten der Serovare 2, 4, 5, 12, 13 und 14, die in den USA im Jahr 1992

Literatur 37
am häufigsten nachgewiesen wurden, getestet. Die Ergebnisse zeigen eine
Variabilität der Kreuzimmunität zwischen den einzelnen Serovaren. Serovar 4
erzeugte demnach einen homologen und heterologen Schutz, Serovar 5 dagegen
nur einen homologen Schutz. Ein bivalenter Impfstoff mit den Serovaren 4 und 5
reduzierte signifikant das Auftreten von Läsionen und verringerte die Mortalität bei
Tieren, die mit den Serovaren 4 und 5 , aber ebenso mit 13 und 14 getestet wurden.
Kein Schutz wurde dagegen aufgebaut bei Tieren, die mit den Serovaren 2 und 12
oder mit nicht typisierbaren Stämmen belastet wurden. Gegen die Serovare 2 und 12
konnte allerdings auch kein Schutz mit homologen Vakzinen aufgebaut werden.
TAKAHASHI et al. (2001) bestätigen die Ergebnisse der fehlenden Kreuzimmunität
zwischen den Serovaren 2 und 5. BAK und RIISING (2002) konnten einen
homologen Schutz gegen Serovar 5 bei Ferkeln, die im Alter von fünf bis sieben
Wochen geimpft wurden, nachweisen. Der heterologe Schutz gegen die Serovare 1,
12, 13 und 14 konnte nur teilweise gezeigt werden.
RAPP-GABRIELSON et al. (1997) haben in einer Studie gezeigt, dass es möglich ist,
Ferkel im Alter von einer Woche zu impfen, allerdings kann der optimale
Impfzeitpunkt je nach Betrieb variieren. Er ist abhängig von dem Vorhandensein von
maternalen Antikörpern, klinischen Erscheinungen und dem betriebsspezifischen
Management.
Bei Untersuchungen zur Wirkung der maternalen Antikörper gegen eine H.-parasuis-
Infektion wurde festgestellt, dass die Vakzination der Muttertiere (gegen Serotyp 5)
bei den Ferkeln eine anschließende Kolonisation der Nasenschleimhaut zwar nicht
verhindert, diese aber mehrheitlich gegen die Manifestation einer Infektion schützt
(HOFFMANN u. BILKEI 2002).
Maternale Antikörper sind etwa in der sechsten Lebenswoche nicht mehr vorhanden,
die Ausbildung einer aktiven Immunität beginnt mit acht Wochen und ist mit zwölf
Wochen nahezu voll ausgebildet (DONE 1999). Der günstigste Zeitpunkt für eine
Vakzination der Ferkel ist gegeben, wenn die kolostral übertragenen Antikörper
weitgehend abgebaut sind. Bis durch die Impfung ein belastbarerer Schutz aufgebaut
ist, besteht also ein Zeitfenster für eine erhöhte Infektionsanfälligkeit der Tiere.

38 Literatur
SOLANO et al. (1999) fanden in ihren Studien heraus, dass die maternalen
Antikörper keinen negativen Einfluss auf die Impfung der Ferkel haben, wenn diese
im Alter von ein bis drei Wochen geimpft werden.
Der beste Zeitpunkt für eine Impfung ist nach SOLANO-AGUILAR et al. (1999)
abhängig vom Immunstatus des Betriebes. Wenn Infektionen mit H. parasuis
vorwiegend in der frühen Aufzuchtphase stattfinden, ist es am besten, die Sauen vor
der Abferkelung zu vakzinieren, so dass die Ferkel einen ausreichenden Schutz in
den ersten drei Lebenswochen haben. Findet die Hauptinfektion später statt, kann
der beste Schutz erreicht werden, wenn die Ferkel vor dem Absetzen noch einmal
geimpft werden.
Ein ähnliches Impfschema empfiehlt WENDT (2004). Erkranken die Saugferkel sehr
früh, sollten die Sauen in der späten Trächtigkeit geimpft werden. Erkranken sie
dagegen erst beim Absetzen, ist eine Impfung der Saugferkel möglich. Tritt die
Glässersche Krankheit erst später in der Aufzucht auf, reicht eine Immunisierung
zum Zeitpunkt des Absetzens.
Ähnliches beschreiben PIJOAN und OLIVEIRA (2003); um den korrekten
Impfzeitpunkt zu bestimmen, müssen das Vorhandensein der maternalen Antikörper
und der Peak der Ferkelmortalität beachtet werden. Tritt das
Hauptkrankheitsgeschehen um die zweite bis dritte Lebenswoche der Ferkel auf,
sollten die Sauen geimpft werden. Liegt die höchste Ferkelmortalität etwa zwischen
der vierten und sechsten Woche nach dem Absetzen, sollten die Ferkel im
Saugferkelalter und zwei Wochen später geimpft werden.
OLIVEIRA et al. (2002) empfehlen keine Kombination von Sauen- und Ferkel-
impfung, da die maternale Immunität mit der Ferkelimpfung interferieren kann.
Eine alternative Methode zur antibiotischen Behandlung und dem Einsatz
konventioneller oder autogener Vakzine ist die kontrollierte Exposition. Bei der
kontrollierten Exposition, die von OLIVEIRA und PIJOAN (2004) getestet wurde,
werden Saugferkel mit einer geringen Dosis eines lebenden, virulenten H.-parasuis-
Stammes inokuliert, um die Aufzuchtmortalität zu kontrollieren. Diese Methode
basiert auf der Grundlage, dass nur wenige Ferkel vor dem Absetzen mit virulenten

Literatur 39
Stämmen infiziert sind. Diese mit dem Erreger früh besiedelten Ferkel haben einen
guten Schutz durch die maternalen Antikörper gegen eine systemische Erkrankung.
Wenn die maternale Immunität abnimmt, haben diese Ferkel eine aktive Immunität
gegen die virulenten Stämme aufgebaut. Nach dem Absetzen sind diese Tiere
Überträger von Infektionen an naive Tiere, die sich nicht mit den virulenten Stämmen
des Betriebes auseinander gesetzt haben. Naive Tiere sind vor allem in der sechsten
bis achten Lebenswoche, wenn der Titer der maternalen Antikörper absinkt, voll
empfänglich für eine systemische Infektion mit H. parasuis. Aufgrund dieser
Hypothese sollte eine frühe Exposition der Ferkel mit den bestandsprevalenten
Stämmen die Aufzuchtmortalität dadurch senken, dass die Anzahl der naiven, voll
empfänglichen Tiere nach dem Absetzen sinkt und es so nicht mehr zur
systemischen Infektion mit H. parasuis kommt (PIJOAN et al. 1997). OLIVEIRA et al.
(2004) haben diese Methode im Vergleich zu kommerziellen und autogenen
Vakzinen getestet. Sie haben fünf Tage alte Ferkel mit einer geringen Dosis
lebender, virulenter Bakteriensuspension inokuliert. Dabei konnten sie im Vergleich
zu den Vakzinen die Mortalität in der Aufzucht durch die kontrollierte Exposition
signifikant (um 50 %) senken. Außerdem ist diese Methode unabhängig von der
Präsenz von maternalen Antikörpern, die bei der Vakzinierung interferieren können.
OLIVEIRA et al. (2004) weisen allerdings darauf hin, dass Ferkel nicht mit lebenden,
virulenten Stämmen von H. parasuis inokuliert werden sollten, wenn eine akute
PRRS-Infektion im Bestand vorhanden ist.
2.1.9 Bekämpfung
2.1.9.1 Erregerkontrolle / Erregereradikation
H.-parasuis-Infektionen kommen vermehrt in Betrieben mit einem hohen
Gesundheitsstatus vor. Besonders gefährdet sind Ferkel aus diesen Betrieben, wenn
sie mit anderen Tieren zusammenkommen. Die Nasenschleimhaut von Saugferkeln
kann schon sehr früh mit dem Erreger besiedelt sein, ein frühes Absetzen ist für die

40 Literatur
Elimination von H. parasuis alleine nicht ausreichend. CLARK et al. (1994) haben
verschiedene Absetzverfahren getestet. Sie sind zu dem Schluss gekommen, dass
ein Versuch, den Erreger zu eradikieren und das Krankheitsgeschehen unter
Kontrolle zu halten, nur erfolgreich sein kann, wenn ein hoher Hygienestatus
gehalten wird, das SEW-Verfahren (segregated early weaning) angewendet wird und
in Kombination dazu Antibiotika in hohen Dosen eingesetzt werden. Allerdings
scheint eine vollständige Eradikation des Erregers sehr schwierig zu sein, da eine
Besiedlung des oberen Respirationstraktes schon wenige Stunden nach der Geburt
stattfinden kann (PIJOAN u. OLIVEIRA 2003).
2.1.9.2 Medikamentelle Therapie
Aufgrund des schwierigen Nachweises von H. parasuis kann nicht immer ein
Antibiogramm erstellt werden.
H. parasuis weist in der Regel eine gute Empfindlichkeit gegenüber zahlreichen
Antibiotika auf (SELBITZ 1992). Als therapeutisch wirksame Präparate gelten
Amoxicillin, Cephalosporine, Enrofloxacin, Florfenicol, Penicillin, Tiamulin, Tilmicosin
sowie Trimethoprim/Sulfonamid, die bei schweren Fällen und/oder negativem
bakteriologischem Befund auch in Form einer diagnostischen Therapie zum Einsatz
kommen (AARESTRUP et al.2004; RAPP-GABRIELSON 1999; ZIMMERMANN et al.
2004). Als Mittel der Wahl wird Penicillin angesehen, allerdings weisen KIELSTEIN
und LEIRER (1990) auf eine steigende Resistenzrate von H. parasuis gegen
Penicillin hin.
In einer deutschen Studie zeigten sich weniger als 10 % aller H.-parasuis-Keime
resistent gegen Tetracyclin bzw. Oxytetracyclin, Enrofloxacin und Kanamycin, jedoch
zu über 55 % als resistent gegen Sulfonamide (VON ALTROCK 1998).
DANIELS et al. (1998) haben bei Untersuchungen von 1340 Isolaten in den USA
lediglich bei Sulphamethoxin (8 % Empfindlichkeit), Clindamycin (15 %),
Erythromycin (26 %) und Tylosintartrat (39 %) eine höhere Resistenzrate festgestellt.
Alle anderen getesteten Präparate zeigten eine gute Empfindlichkeit gegen H.

Literatur 41
parasuis (Ampicillin 93 %, Apramycin 65 %, Ceftiofur 98 %, Gentamycin 99 %,
Trimethoprim/Sulfonamid 92 %, Neomycin 93 %, Penicillin 59 %, Spectinomycin 86
%, Tetracyclin 91 % und Tiamulin 87 %).
Einer dänischen Studie zufolge sind alle untersuchten Stämme empfindlich
gegenüber allen getesteten Antibiotika. Lediglich zwei von insgesamt 132 Isolaten
zeigten eine Resistenz gegen eine Kombination von Trimethoprim und
Sulfamethoxalin (AARESTRUP et al. 2003). Die Autoren dieser Studie begründen die
hohe Resistenzrate in anderen Studien damit, dass häufig Isolate von behandelten
Tieren verwendet werden.
DESROSIERS et al. (1986) empfehlen bei einem klinischen Ausbruch der
Glässerschen Krankheit so schnell wie möglich eine parenterale Behandlung mit
Antibiotika durchzuführen. Wichtig ist, dass alle Tiere behandelt werden und nicht nur
diejenigen, die klinische Anzeichen zeigen.
2.1.10 Ko-Infektionen mit anderen Erregern des Schweines In der Literatur sind verschiedene Untersuchungen zu Ko-Infektionen und
Interaktionen zwischen H. parasuis und anderen schweinepathogenen Erregern
beschrieben. Vor allem Atemwegserkrankungen sind in der Regel nur sehr selten
Folge von Monoinfektionen sondern werden durch multiple Infektionen verursacht,
die zudem von diversen Umgebungsfaktoren und Eigenschaften des Wirtes
beeinflusst werden.
Die Isolation von H. parasuis von Schweinen mit Pneumonien hat in den letzten
Jahren erheblich zugenommen, und es wird vermutet, dass dieses im
Zusammenhang mit einer steigenden Prävalenz von Mykoplasmen und viralen
Erregern, wie dem Virus des Porzinen Respiratorischen und Reproduktiven
Syndroms (PRRSV), dem Influenzavirus und dem Porzinen Respiratorischen
Coronavirus (PRCV) (RAPP-GABRIELSON 1999), steht.
Die Rolle von H. parasuis bei respiratorischen Erkrankungen beim Schwein sind von
komplexer Natur. Die Tatsache, dass H. parasuis bei purulenten Rhinitiden beteiligt

42 Literatur
ist, unterstützt die Vermutung, dass der Erreger einen prädisponierenden Faktor für
Infektionen mit anderen bakteriellen und viralen Erregern darstellt (GOIS et al. 1983;
VAHLE et al. 1995, 1997).
Bei Pneumonien wird angenommen, dass H. parasuis ein sekundärer Besiedler ist
und Erkrankungen nur im Zusammenhang mit anderen bakteriellen oder viralen
Erregern entstehen. NARITA et al. (1994) haben diesen Zusammenhang bei
experimentellen Infektionen mit H. parasuis Serovar 4 und dem Virus der
Aujetzkyschen Krankheit (Porcines Herpesvirus, PHV-1) dargestellt. Sie zeigten,
dass durch die PHV-1-Infektion die Epithelzellen des Respirationstraktes der
Schweine zerstört wurden und H. parasuis sich in den Lungen vermehren konnte.
Viele neuere Berichte zeigen allerdings H. parasuis eher als Primärerreger bei
fibrinös-eitrigen Bronchopneumonien (PÖHLE et al. 1992; BARIGAZZI et al. 1994;
SOLANO et al. 1998).
SOLANO et al. (1997) haben die Interaktion von PRRS und H. parasuis untersucht.
Sie konnten keine Zunahme der klinischen Symptome, wie Polyserositis bei Ferkeln,
feststellen, die mit beiden Erregern infiziert waren. Allerdings steht dieses im Kontrast
zu Feldbeobachtungen, nach denen endemische PRRS-Infektionen eine Zunahme
der Polyserositiden durch H.-parasuis-Infektion hervorrufen (VAHLE et al. 1994;
DONE u. PATON 1995).
Auch SEGALES et al. (1999) konnten keine potenzierende Wirkung von PRRS auf
die Replikation von H. parasuis und keine Beziehung zwischen der Präsenz von
PRRSV und H.-parasuis-Antigenen bei doppelt infizierten Tieren feststellen. Eine
PRRS-Infektion induziert demnach die Bildung von Interleukinen und Interferon-α in
Lungenzellen, die eine Rolle in der antiviralen Abwehr und der Stimulation nicht-
spezifischer pulmonaler Entzündungsreaktion spielen (VAN REETH 1997).
H. parasuis in Kombination mit Mycoplasma hyorhinis ist in 51,2 % der Isolate
festgestellt worden, die von PRRS-infizierten Schweinen stammten (KOBAYASHI et
al. 1996).
Bei der Untersuchung zum gemeinsamen Vorkommen von Mikroorganismen und
Läsionen beim postweaning multisystemic wasting syndrome stellten KIM et al.

Literatur 43
(2002) in 85 % der Fälle eine duale Infektion fest. Die häufigste Kombination (32 %)
bestand aus PCV-2 und H. parasuis.
Bei Untersuchungen verschiedener Krankheitserreger in der Bronchoalveolar-Lavage
erzielte PABST (2004) Hinweise auf die Bedeutung von H. parasuis als Wegbereiter
für M. hyopneumoniae. Dieser Sachverhalt muss aber noch weiter untersucht
werden.
2.2 Mycoplasma hyopneumoniae
2.2.1 Ätiologie und Pathogenese
Mycoplasma (M.) hyopneumoniae ist der Primärerreger der weltweit verbreiteten
Enzootischen Pneumonie (EP). M. hyopneumoniae wurde erstmalig 1965
beschrieben (GOODWIN et al. 1965; MARE u. SWITZER 1965) und wenig später als
primäre Ursache der Enzootischen Pneumonie erkannt (HODGES et al. 1969). Der
Erreger ist nur an das Schwein adaptiert. Taxonomisch werden Mykoplasmen in die
Klasse der Mollicutes, der kleinsten, selbständig vermehrungsfähigen Bakterien,
eingeordnet (TULLY et al. 1993) . Die Gestalt wird als pleomorph beschrieben, und
der Durchmesser beträgt etwa 0,1 - 0,3 μm, sie vermehren sich durch Querteilung.
Anstelle einer Zellwand verfügen Mykoplasmen über eine einfache, dreischichtige
Plasmamembran (ROSS 1999).
Phylogenetisch lässt sich der Ursprung der Mykoplasmen auf grampositive Bakterien
zurückführen (WOESE 1987). Aufgrund ihrer parasitären Lebensweise haben
Mykoplasmen im Laufe einer reduktiven Evolution genetische Informationen verloren
und besitzen daher nur eine reduzierte Enzymausstattung und eingeschränkte
Stoffwechsel- und Biosynthesewege (RAZIN 1992). Daher ist auch eine Kultivierung
vieler Mykoplasmen bis heute sehr schwierig und bedarf spezieller Nährmedien
(Zusatz von Serum, CO2 und Antibiotika).

44 Literatur
Nach aerogener Aufnahme von M. hyopneumoniae kommt es zur bronchiogenen
Ausbreitung und Bindung des Erregers an das zilientragende Epithel von Trachea,
Bronchien und Bronchiolen (ZIELINSKY u. ROSS 1993). Die Mykoplasmen
replizieren auf der Oberfläche der Zilien und führen innerhalb weniger Tage zu einer
Einschränkung der Ziliaraktivität (JACQUES et al.1992; KOBISCH et al. 1993;
ZIELINSKY u. ROSS 1993). Die Adhäsion geht zudem mit der Bildung oxigener
Radikale und einem Anstieg der Kalziumkonzentration in der Epithelzelle einher, die
zu zytotoxischen Effekten mit Nekrosen, lokaler Desquamation und Deziliation führt
(PARK et al. 2002). Der Verlust der zilientragenden Zellen und eine vermehrte
Schleimproduktion beeinträchtigen die mukoziliäre Clearance und fördern so die
weitere Besiedlung des Atmungstraktes mit pathogenen Keimen (ROSS 1996).
Zilienverluste sind frühestens ab zwei Wochen nach der Infektion
elektronenmikroskopisch festzustellen (KOBISCH et al. 1993).
Die zelluläre Immunantwort hat für den Krankheitsverlauf eine erhebliche Bedeutung.
Die Membranen von M. hyopneumoniae haben einen mitogenen Effekt auf
Schweinelymphozyten und stimulieren das Immunsystem (MESSIER et al. 1990;
KWON et al. 2002). Die Einwanderung von Lymphozyten und Makrophagen in den
peribronchialen, peribronchiolären und perivaskulären Raum beeinflusst infolge
immunpathologischer Veränderungen die Ausprägung der pneumonischen
Veränderungen (YAGIHASHI et al. 1984). Gleichzeitig wird die Phagozytoseaktivität
der Alveolarmakrophagen herabgesetzt, was die Abwehrfunktion einschränkt
(CARUSO u. ROSS 1990; KWON u. CHAE 1999). Neben der Einschränkung der
mechanischen Abwehr (mukoziliäre Clearance) bewirkt die Infektion mit M.
hyopneumoniae somit auch eine partielle Beeinträchtigung der Funktion des
Immunsystems. Infizierte Tiere sind anfälliger für Infektionen mit weiteren viralen
und/oder bakteriellen Erregern von Atemwegserkrankungen (YAGIHASHI et al. 1984;
ROSS 1999; THACKER 2001).

Literatur 45
2.2.2 Epidemiologie
M. hyopneumoniae kommt in Ländern mit intensiver Schweineproduktion bei Zucht-
und Mastschweinen mit hohen Prävalenzen vor, wie anhand serologischer
Untersuchungen mehrfach festgestellt wurde.
Der Anteil seropositiver Bestände (d.h. Bestände mit wenigstens einem positiv
reagierenden Tier) beträgt in Deutschland nach Untersuchungen von HORST et al.
(1997) 81,2 % bei Mastschweinen (ab 60 kg Körpergewicht), 63,02 % bei Jungsauen
und 47,2 % bei Altsauen. In anderen Ländern wurden vergleichbare Zahlen
festgestellt (FALK et al. 1991; KOBISCH et al. 1994; MAES 1997; ERLANDSON
2002).
Für die Einschleppung des Erregers in einen Bestand wird dem Zukauf von
infizierten, aber klinisch unauffälligen Schweinen ein hohes Risikopotential
zugeschrieben (MAES et al. 2000). Durch den Transport, Stall- und Futterwechsel
kann das Immunsystem geschwächt und damit Voraussetzungen für eine erhöhte
Ausscheidung von M. hyopneumoniae geschaffen werden. Eine Infektion bzw.
Übertragung des Erregers zwischen Herden ist aber auch durch die Luft möglich.
Zwischen Herden konnte eine aerogene Übertragung über Distanzen von bis zu 3,2
km nachgewiesen werden (GOODWIN 1985; WHITTLESTONE 1990). Die
Entfernung zur nächsten mit M. hyopneumoniae infizierten Herde ist als einer der
bedeutendsten Risikofaktoren für die Reinfektion von M.-hyopneumoniae-freien
Herden anzusehen. Weitere Risikofaktoren sind das Mischen von Tieren aus
mehreren Herkünften und das Nicht-desinfizieren von Transportfahrzeugen (HEGE et
al. 2002).
Außerhalb des Wirtes kann der Erreger in Abhängigkeit von den Umweltbedingungen
längere Zeit überleben (FRIIS 1973; GOODWIN 1985). Die Überlebenszeit in Wasser
beträgt bei Temperaturen zwischen 2 und 7° C bis zu 31 Tagen (GOODWIN 1985).
Da es sich bei der Enzootischen Pneumonie um eine faktorenabhängige Erkrankung
handelt, treten Probleme vor allem bei ungünstigeren Klimabedingungen im Herbst
und Winter auf (MAES 1998), so dass ein Zusammenhang mit Veränderungen in der

46 Literatur
Luftfeuchtigkeit und Lufttemperatur sowie durch reduzierte Raumbelüftung
anzunehmen ist (ROSS 1999).
Die Übertragung von M. hyopneumoniae innerhalb infizierter Herden erfolgt
hauptsächlich durch direkten Tierkontakt. Das Risiko einer Serokonversion ist für
Schweine mit direktem Kontakt zu infizierten Tieren um den Faktor sieben höher als
für Schweine mit Kontakt zu kontaminierten Vektoren (MORRIS et al. 1995). Belebte
oder unbelebte Vektoren, die mit Sekreten infizierter Tiere kontaminiert sind,
scheinen für die Erregerübertragung insgesamt keine besondere Bedeutung zu
haben (BATISTA et al. 2002). Eine indirekte Erregerübertragung kann aber auch
mittels kontaminierter Aerosole erfolgen (STÄRK 1998).
Eine Erregerübertragung findet von infizierten Sauen auf die Ferkel (CLARK et al.
1992) oder zwischen infizierten und nicht infizierten Schweinen verschiedener
Altersgruppen statt (GOODWIN 1972; WALLGREN u. SCHWAN 1994).
Ein besonderes Risiko einer vertikalen Übertragung wird für Würfe von Jungsauen
angenommen, da jüngere Sauen häufig noch nicht über ausreichend hohe
Antikörperkonzentrationen im Kolostrum verfügen (BERNER 1995). Das Risiko einer
Infektion der Ferkel durch die Sau wird von RAUTIAINEN et al. (1998) als eher
gering eingeschätzt, wenn ausreichend maternale Antikörper auf die Ferkel
übertragen werden.
Eine intrauterine Übertragung von M. hyopneumoniae auf die Feten konnte durch
experimentelle Infektion tragender Sauen nicht erreicht werden (HEITMANN u.
KIRCHHOFF 1981; BÜRGI et al. 1990).
Die horizontale Übertragung kann zwischen gleichaltrigen Schweinen oder von
älteren Schweinen auf jüngere Tiere erfolgen, wenn Abteile kontinuierlich belegt oder
Tiere zurückgestallt werden. Vor allem das Zusammenstallen von Ferkeln aus
verschiedenen Beständen ist mit einem großen Risiko verbunden, da die Gefahr
einer Infektion mit M. hyopneumoniae mit der Anzahl der Bestände steigt, aus denen
Ferkel zugekauft werden (CLARK et al. 1991; MAES et al. 2000). Die Übertragung
von M. hyopneumoniae innerhalb von Herden erfolgt im Vergleich zu viralen Erregern
respiratorischer Erkrankungen langsam, und der Anteil seropositiver Tiere steigt bis
zum Mastende an (MAES et al. 1998; WALLGREN et al. 1998; MATEUSEN et al.

Literatur 47
2002). Infizierte Tiere können den Erreger über Wochen ausscheiden (RUIZ u.
PIJOAN 2002).
2.2.3 Klinische Symptomatik
Charakteristisches Merkmal einer Infektion mit M. hyopneumoniae ist das Auftreten
eines trockenen, chronischen Hustens (PLONAIT 1988; KOBISCH et al. 1993; ROSS
1996). Der Krankheitsverlauf ist in Herden von einer teilweise hohen Morbidität, aber
üblicherweise nur geringen Mortalität geprägt. Erste klinische Anzeichen fallen ab
etwa zwei Wochen p.i. als trockener Husten mit geringer Erhöhung der
Körpertemperatur und Inappetenz auf (DONE 1996; ROSS 1999; KOBISCH et
al.2000).
Akute klinische Erkrankungen sind infolge einer Monoinfektion mit M.
hyopneumoniae unter Feldbedingungen nur bei Schweinen aus SPF-Beständen zu
erwarten. Der Verlauf der Erkrankung unter konventionellen Haltungsbedingungen
wird wesentlich durch Ko-Infektionen mit weiteren bakteriellen Erregern, wie z.B. P.
multocida, B. bronchiseptica, S. suis, H. parasuis und A. pleuropneumoniae,
bestimmt. Das Vorkommen von Ko-Infektionen erschwert den Krankheitsverlauf, so
dass es auch zu einem Anstieg der Mortalität kommen kann (ROSS 1999; KOBISCH
et al. 2000). In Betrieben mit guten Umweltbedingungen verläuft die M.-
hyopneumoniae-Infektion oft subklinisch, oder Infektion und klinische Erkrankung
finden erst spät in der Mast statt (MAES 2002). Bei einer klinisch manifesten M.-
hyopneumoniae-Infektion entstehen durch höhere Behandlungskosten und
Leistungsdepressionen, wie verminderter Zuwachs, reduzierte Futterverwertung und
längere Mastdauer, ökonomische Verluste (KOBISCH et al. 1994; SCHEIDT et al.
1994; ROSS 1999).

48 Literatur
2.2.4 Pathologie Pathologisch-anatomische Lungenläsionen werden innerhalb von ein bis zwei
Wochen nach experimenteller M.-hyopneumoniae-Infektion sichtbar. Die Läsionen
betreffen vorrangig die ventralen Abschnitte der Lobi craniales und mediales in Form
einer katarrhalischen Bronchopneumonie. Das veränderte Gewebe ist atelektatisch,
verfestigt und verfärbt, zuerst blassrot bis blassgelb und im weiteren Verlauf grau bis
dunkelrot. In den Bronchien befindet sich trüber Schleim, und die
Bronchiallymphknoten sind geringgradig vergrößert (BLANCHARD et al. 1992;
KOBISCH et al. 1993). Die als Spitzenlappenpneumonie bezeichneten
Veränderungen werden zwar allgemein als typisch für die Infektion mit M.
hyopneumoniae angesehen, es muss aber beachtet werden, dass auch andere
Pneumonien bakterieller Genese vergleichbare Läsionen erzeugen können (ROSS
1999). 28 Tage nach der Infektion sind die Läsionen am stärksten ausgeprägt und
heilen, sofern es nicht zu bakteriellen Ko-Infektionen kommt, innerhalb von 60 bis
100 Tagen weitgehend aus. Später sind nur noch band- oder sternförmige narbige
Einziehungen auf der Lungenoberfläche zu erkennen (SØRENSEN et a. 1997). Ob
es zu Veränderungen der serösen Häute durch M. hyopneumoniae kommt, ist
umstritten. BERTSCHINGER et al. (1972) vertreten die Meinung, dass
Veränderungen der serösen Häute nicht infolge einer Infektion mit M.
hyopneumoniae auftreten, während ANDREASEN et al. (2001) feststellten, dass im
Zusammenhang mit frühzeitigen Serokonversionen der Anteil der Pleuritiden bei
Schlachtschweinen steigt. Laut BUTTENSCHØN et al. (1997) ist M. hyopneumoniae
der häufigste Erreger von fibrinösen Perikarditiden bei Schlachtschweinen.
Histopathologisch sind Hyperplasien des Bronchialepithels sowie Infiltrationen von
Lymphozyten in das peribronchiale, peribronchioläre und perivaskuläre Gewebe zu
beobachten (ROSS 1999). Bei Vorliegen sekundärer Infektionen bekommen die
Veränderungen einen purulenten Charakter (MAES 2002).

Literatur 49
2.2.5 Diagnostik Unspezifische Anzeichen einer Infektion mit M. hyopneumoniae sind klinische
Symptome sowie makroskopische Veränderungen des Lungengewebes.
Als klinisches Symptom ist das Auftreten eines trockenen, chronischen Hustens
charakteristisch (PLONAIT 1988; KOBISCH et al. 1993; ROSS 1996).
Die betroffenen Lungenareale sind verfestigt und verfärbt, zuerst blassrot bis
blassgelb und im weiteren Verlauf grau bis dunkelrot (KOBISCH et al. 1993). Die als
Spitzenlappenpneumonie bezeichneten Veränderungen lassen aber keine
ätiologische Diagnose zu.
Nur durch den direkten oder indirekten Erregernachweis ist eine spezifische
Diagnose einer Infektion mit M. hyopneumoniae möglich.
Direkter Erregernachweis Der direkte Erregernachweis erfolgt durch die kulturelle Anzüchtung von M.
hyopneumoniae oder die Darstellung von M.-hyopneumoniae-Antigen.
Der kulturelle Nachweis gestaltet sich aufgrund der hohen Ansprüche des Erregers
an den Nährboden schwierig, teuer und sehr zeitaufwändig. Die Isolation von M.
hyopneumoniae wird zudem häufig durch eine Überwucherung mit M. hyorhinis und
M. flocculare erschwert. Auf festen Medien wachsen ca. 0,5 mm große Kolonien, bei
denen das Fehlen eines dunklen Zentrums eine Differenzierung von anderen
Mykoplasmenspezies erlaubt. In flüssigen Medien wächst M. hyopneumoniae
langsam und produziert nach vier bis 15 Tagen einen sauren Farbwechsel mit
schwacher Trübung (KOBISCH et al. 2000).
Um Antigen oder Genomfragmente von M. hyopneumoniae nachzuweisen, sind
verschiedene Methoden möglich.
Die verschiedenen Methoden der Polymerase Chain Reaction (PCR) basieren auf
einer Amplifikation von DNA (MAES 2002). Dabei ist der Nachweis von
Genomfragmenten von M. hyopneumoniae aus Nasentupfern oder Flüssigkeiten
einer bronchoalveolären Lavage möglich (ABIVEN et al. 1992; SØRENSEN et al.
1997). Vorteile ergeben sich aus der im Vergleich zur Kultur schnelleren

50 Literatur
Verfügbarkeit der Ergebnisse, vor allem bei Erregern, deren In-vitro-Kultivierung
schwierig, zeitaufwändig oder unverhältnismäßig teuer ist (KURTH et al. 2002). Mit
Hilfe der PCR können prinzipiell sehr geringe Mengen pathogener Organismen
nachgewiesen werden.
Der Immunfluoreszenztest (IFT) kann zum Nachweis von M. hyopneumoniae in
dünnen Gefrierschnitten der Lunge angewendet werden (MEYLING 1971). M.
hyopneumoniae wird bei diesem Verfahren mit Isothionat markierten Antikörpern
dargestellt. Im Fall einer Infektion wird eine grün-gelblich fluoreszierende Linie auf
der Oberfläche des Bronchial- und Bronchiolarepithels sichtbar (KOBISCH et al.
2000). Die Praktikabilität der Methode ist fraglich, da die Lungen sofort nach der
Tötung der Tiere bearbeitet werden müssen (THACKER 2001).
Als weitere Methoden zum Nachweis von M. hyopneumoniae ist ein Antigen-ELISA
entwickelt worden (PEDERSEN et al. 1990). Ebenfalls ist die Identifizierung mittels
Gensonden beschrieben worden (ABIVEN et al. 1992).
Indirekter Erregernachweis Für den Nachweis von Antikörpern gegen M. hyopneumoniae in Serum, Kolostrum
oder der Bronchoalveolarlavage-Flüssigkeit (BALF) werden ELISA-Verfahren
angewendet.
Serokonversionen sind mit dem ELISA frühestens neun Tage nach einer Infektion
feststellbar; im Mittel vergehen etwa 14 bis 21 Tage zwischen der Infektion und dem
Nachweis einer Serokonversion (KOBISCH et al. 1993; SØRENSEN et al. 1997).
Serumantikörper sind mittels ELISA über einen Zeitraum von ca. 52 Wochen
nachweisbar (DONE 1996). SØRENSEN et al. (1994) konnten maximale Antikörper-
spiegel um den 46. Tag p.i. feststellen, während KOBISCH et al. (1993) die höchsten
Antikörperspiegel 11 bis 12 Wochen p.i. nachweisen konnten. Im Vergleich mit der
Diagnostik pathologisch-anatomischer Veränderungen hat der sogenannte TWEEN-
20-ELISA eine Spezifität von 96 % und eine Sensitivität von 99 % (SHELDRAKE u.
ROMALIS 1992). Ein in Skandinavien entwickelter blocking-ELISA erreichte eine
Spezifität von 96 % und eine Sensitivität von 93 %; Referenzmethode war hierbei der
indirekte Hämagglutinationstest (BARFOD et al. 1994). Der Vorteil des TWEEN-20-

Literatur 51
ELISA liegt in der Möglichkeit, eine Infektion früher nachzuweisen als mit dem
blocking-ELISA (WALLGREN et al. 1996).
Andere Methoden, wie z. B. die Komplementbindungsreaktion (SLAVIK u. SWITZER
1972) oder der indirekte Hämagglutinationshemmungstest (LAM u. SWITZER 1971),
haben aufgrund der geringen Sensitivität und Spezifität kaum Verwendung in der
Routinediagnostik von M.-hyopneumoniae-Infektionen gefunden (ARMSTRONG et
al. 1983).
2.2.6 Immunität 2.2.6.1 Immunantwort M. hyopneumoniae induziert eine systemische und eine lokale Reaktion des
humoralen Immunsystems. Bei der systemischen Antikörperantwort, die nicht direkt
mit der Protektion korreliert, werden Immunglobuline der Klasse M etwa sieben Tage
p.i., die der Klasse G1 zehn bis 14 Tage p.i. und der Klassen G2 und A 21 Tage p.i.
nachgewiesen (PIFFER u. ROSS 1984; SHELDRAKE u. ROMALIS 1992; THACKER
et al. 2000; BOETTCHER et al. 2002). Serumantikörper können mittels ELISA über
einen Zeitraum von etwa einem Jahr nachgewiesen werden (GOODWIN et al. 1969;
HOWARD u. TAYLOR 1985; KOBISCH et al. 1993).
Bei der lokalen Antikörperantwort werden Immunglobuline der Klassen G und A
gebildet (DJORDJEVIC et al. 1997; THACKER et al. 2000). Die protektive Wirkung
von lokalen Antikörpern wird kontrovers diskutiert. DJORDJEVIC et al. (1997)
konnten einen Zusammenhang zwischen lokalen Antikörpern und einer damit
einhergehenden Protektion nicht nachweisen, während THACKER et al. (2000) einen
entsprechenden Zusammenhang feststellten.
Die Bedeutung der zellulären Immunität für den Schutz vor der Infektion mit M.
hyopneumoniae bzw. vor der Entstehung der Enzootischen Pneumonie ist ebenfalls
noch nicht geklärt (BOETTCHER et al. 2002). Es wird von einer Lymphozytenantwort
gegen M. hyopneumoniae berichtet, die aus der Stimulation von Lymphozyten nach

52 Literatur
Impfung bzw. experimenteller Infektion abgeleitet wird (KRISTENSEN et al. 1982;
THACKER et al. 1998). Weitere Untersuchungen zeigen, dass die Impfung gegen M.
hyopneumoniae die Produktion von spezifischen Interferon-Gamma (IFN- )-
produzierenden Zellen induziert (THACKER et al. 1998). IFN- gehört zur Gruppe
der Zytokine und hat Bedeutung für die Makrophagenaktivierung. Die zelluläre
Immunantwort steuert die Abwehrreaktion gegen M. hyopneumoniae demnach durch
eine Aktivierung von Makrophagen (BOETTCHER et al. 2002).
Insgesamt ist davon auszugehen, dass die Infektion mit M. hyopneumoniae eine
andauernde und belastbare Immunität induziert (GOODWIN et al. 1969; KOBISCH et
al. 1993).
Neben der aktiven Immunisierung durch Infektion oder Impfung sind Ferkel durch
maternale Antikörper passiv gegen M. hyopneumoniae geschützt (KOBISCH et al.
1987; RAUTIAINEN u. WALLGREN 2001). Bei tragenden Sauen kommt es vor der
Geburt zu einer Konzentration von Immunglobulinen im Kolostrum und zu einem
Absinken der Antikörperwerte im Serum (KLOBASA et al. 1985; WALLGREN et al.
1998; SCHREIBER 2002). Im Kolostrum können zu diesem Zeitpunkt um 70 %
höhere IgG-Werte als im Blutserum beobachtet werden (NICOLET et al. 1990).
Da Ferkel aufgrund der Plazentationsverhältnisse beim Schwein (Placenta
epitheliochoriales) zum Zeitpunkt der Geburt keine Antikörper besitzen, sind sie auf
die Aufnahme von maternalen Antikörpern mit dem Kolostrum angewiesen. Der
Darm der Ferkel ist innerhalb der ersten 24 bis 36 Stunden post natum vermehrt
durchlässig, maternale Antikörper können während dieser Zeit aus dem Kolostrum
unverändert aufgenommen werden. Während der ersten sechs Lebensstunden stellt
IgG den Hauptanteil der Proteine im Kolostrum (KLOBASA u. BUTLER 1987).
Innerhalb von drei Tagen post partum ändert sich die Zusammensetzung der
Fraktion der Immunglobuline im Kolostrum, so dass anschließend hauptsächlich IgA
enthalten ist, welches direkt im Gesäuge gebildet wird (MESSIER et al. 1990). Die
Konzentration der maternalen Antikörper im Ferkel korreliert eng mit den
Antikörperkonzentrationen des Muttertieres (WALLGREN et al. 1998), wobei ältere
Sauen in der Regel eine höhere Antikörperkonzentration aufweisen als Jungsauen

Literatur 53
(RAUTIAINEN u. WALLGREN 2001). Für den Abbau maternal übertragener
Antikörper wird eine mittlere Halbwertszeit von 15,8 Tagen festgestellt (MORRIS et
al. 1994), wobei die Persistenz maternaler Antikörper im Ferkel von der initialen
Konzentration abhängig ist. Bei initial hohen Antikörperkonzentrationen sind die
Ferkel nach 63 Tagen, bei mittleren Konzentrationen nach 45 Tagen und bei
niedrigen Konzentrationen nach 30 Tagen serologisch negativ.
Verschiedene Veröffentlichungen zeigen, dass maternale Antikörper zumindest mit
der humoralen Immunreaktion auf eine Impfung interferieren können (KLOBASA et
al. 1985; MORRIS et al. 1994; MAES et al. 1998; THACKER et al. 2002). WEGNER
et al. (2002) empfehlen, eine Impfung der Ferkel solange abzuwarten, bis wenigstens
80 % der Ferkel serologisch negativ sind.
2.2.6.2 Prophylaxe durch Impfung
Die Impfung gegen M. hyopneumoniae kann die Ausbildung einer humoralen und
zellulären Immunreaktion induzieren und einen Schutz vor Erkrankung erreichen
(KUHN et al. 2000; THACKER et al. 2000). Die Effektivität der Impfung von Ferkeln
konnte in verschiedenen Studien anhand der Verminderung von Lungenläsionen und
klinischen Symptomen sowie der Verbesserung von Zuwachs und Futterverwertung
nachgewiesen werden (VRAA-ANDERSEN et al. 1994; DOHOO u. MONTGOMERY
1996; RADELOFF 1997; MAES et al. 1999; KUHN 2002). Mit der Impfung ist
allerdings keine sterile Immunität, also eine Verhinderung der Kolonisation von M.
hyopneumoniae zu erreichen (WHITTLESTONE 1990; KOBISCH et al. 2000;
MATEUSEN et al. 2002); sie induziert aber eine Immunreaktion, die bei einer
späteren Erregerexposition geboostert wird, so dass die körpereigene Abwehr
schneller und vermutlich auch effektiver reagieren kann (MAES et al. 1998).
Außerdem kann der Effekt der Impfung unter ungünstigen Haltungs- und
Managementbedingungen sinken und dadurch nur einen partiellen Schutz vermitteln
und klinische Symptome nur unzureichend verhindern (THACKER et al. 1998).

54 Literatur
Durch eine Impfung kann aber grundsätzlich eine Reduzierung der
Erregerausscheidung erreicht werden (DONE 1996).
Inaktivierte Impfstoffe gegen M. hyopneumoniae sind in Europa seit 1992
zugelassen. Diese Impfstoffe waren ursprünglich für die zweimalige intramuskuläre
Applikation bei Saugferkeln konzipiert. Das Angebot einiger Hersteller ist seit 2002
um sogenannte one-shot-Vakzinen erweitert worden, die nur noch einmal appliziert
werden.
Der Impfzeitpunkt wird in den verschiedenen Ländern unterschiedlich gehandhabt. In
Europa ist für two-shot-Vakzinen eine Anwendung in der ersten und dritten bzw.
vierten Lebenswoche weit verbreitet. In Nordamerika ist es dagegen üblich, diese
Vakzine in der dritten und siebten Lebenswoche zu impfen. Die one-shot-Vakzine
wird in Europa hauptsächlich in der dritten Lebenswoche eingesetzt, in Nordamerika
meist in der neunten Lebenswoche (KOBISCH et al. 2000; ERLANDSON et al.
2002). Der Impfzeitpunkt sollte in Abhängigkeit von der Präsenz maternaler
Antikörper und dem Infektionszeitpunkt festgelegt werden.
2.2.7 Bekämpfung 2.2.7.1 Erregerkontrolle
Die Infektionen mit M. hyopneumoniae können grundsätzlich durch medikamentelle
Therapien, Impfungen sowie Hygiene- und Managementmaßnahmen soweit
kontrolliert werden, dass der Infektionsdruck sinkt und die klinischen Folgen der
Infektion kalkulierbar bleiben.

Literatur 55
2.2.7.2 Medikamentelle Therapie Die Behandlung einer M.-hyopneumoniae-Infektion mit antibiotisch wirksamen
Arzneimitteln ist grundsätzlich möglich und kann zu einer klinischen Heilung und
Eingrenzung der pathologischen Lungenveränderungen führen. Prinzipiell kann eine
Behandlung mit Antibiotika aus den Wirkstoffgruppen der Tetracycline (Tetracyclin,
Oxytetracyclin, Chlortetracyclin, Doxicyclin), Makrolide (Tylosin, Spiramycin,
Tilmicosin), Lincosamide (Lincomycin, Clindamycin), Pleuromutiline (Tiamulin,
Valnemulin), Florfenicol oder Chinolone (Enrofloxacin, Marbofloxacin, Danofloxacin,
Norfloxacin, Ofloxacin) erfolgen (HANNAN et al. 1989; INAMOTO et al. 1994;
GRANDE; ANGE et al. 2000).
2.2.7.3 Erregereradikation Neben einer Kontrolle der M.-hyopneumoniae-Infektion ist eine vollständige
Erregereradikation möglich. Der Aufbau von M-hyopneumoiae-freien Herden kann
durch verschiedene Sanierungsprogramme erfolgen.
Beim SPF-Verfahren werden Ferkel per Hysterektomie geboren und anschließend
mutterlos aufgezogen. Diese Methode ist sehr aufwendig und für konventionelle
Betriebe unpraktikabel. Bei großen Zuchtorganisationen wird dieses Verfahren
jedoch zum Aufbau sogenannter Nukleusherden eingesetzt (TAYLOR 1999).
Beim Medicated-Early-Weaning-Verfahren wird die vertikale Übertragung von der
Sau auf die Ferkel durch eine intensive Verabreichung von Antibiotika an die Sau
und deren neugeborene Ferkel unterbrochen. Zusätzlich werden die Ferkel sehr früh
abgesetzt und isoliert aufgezogen (ALEXANDER et al. 1980).
ZIMMERMANN et al. (1989) entwickelten für den schweizerischen Tiergesundheits-
dienst ein als Teilsanierung bezeichnetes Programm, bei dem auf eine komplette
Räumung des Bestandes verzichtet wird. Grundlage des Modells ist die Entfernung
aller Tiere, die jünger als zehn Monate sind und als Hauptinfektionsquelle für M.
hyopneumoniae angesehen werden müssen. Das Risiko einer Übertragung von M.

56 Literatur
hyopneumoniae durch immune Altsauen wird als außerordentlich gering
eingeschätzt. Voraussetzung für den Erfolg der Sanierung ist aber eine Stabilität der
Herde, die anhand des Fehlens klinischer Symptome von Atemwegserkrankungen
festgestellt wird. Der verbleibende Bestand an Alttieren wird für einen Zeitraum von
14 Tagen intensiv mit Antibiotika behandelt. Während dieser Zeit dürfen keine
Abferkelungen stattfinden.
Versuche einer Erregereradikation durch die gezielte Ausmerzung der seropositiven
Tiere führten nicht zum Erfolg (LEVONEN et al. 1992).

Material und Methode 57
3 Material und Methoden Die nachfolgend im Detail beschriebene Untersuchung wurde mit dem Ziel
durchgeführt, die Effektivität des Kombinationsimpfstoffes Suvaxyn® M.hyo-Parasuis
gegen H. parasuis und M. hyopneumoniae im Vergleich zu dem Monoimpfstoff
Suvaxyn® M.hyo gegen M. hyopneumoniae zu prüfen. Berücksichtigt wurden dabei
Produktionsparameter, wie tägliche Gewichtszunahmen, Futterverwertung, Mortalität,
Tierbehandlungsindex, durchschnittliche Mastdauer, AutoFOM-Werte und
Lungenbefunde sowie serologische Reaktionen auf die Impfungen.
Die Untersuchungen wurden in der Zeit zwischen August 2004 und Juni 2005
durchgeführt.
3.1 Beschreibung des Versuchbetriebes Der Versuch wurde in einem Zuchtbestand mit 1700 Sauen in Norddeutschland
durchgeführt, der endemisch mit H. parasuis infiziert ist. Alle Ferkel werden in
eigenen Ferkelaufzuchtställen aufgezogen und anschließend in angeschlossenen
Mastställen gemästet.
Der Versuchsbestand wird im sogenannten Wochenrhythmus geführt, so dass jede
Woche etwa 70 bis 80 Sauen abferkeln. Die Einstallung dieser Sauen erfolgt jeweils
sieben Tage vor dem erwarteten Geburtstermin. Hauptabferkeltag ist der
Donnerstag. In jedem der insgesamt acht Abferkelabteile befinden sich Plätze für
vierzig Sauen. Die einzelnen Buchten sind durch 40 cm hohe Wände voneinander
getrennt. Die Säugezeit der Ferkel beträgt drei Wochen.
Als medizinische Maßnahmen bekommen die Ferkel am 1. Lebenstag Amoxicillin
verabreicht, am 2. Lebenstag ein Präparat zur Prophylaxe der Eisenmangelanämie
und am 5. Lebenstag Langzeitpenicillin.
Am Ende der Säugezeit werden sie in den Ferkelaufzuchtstall gebracht.
Der Aufzuchtstall befindet sich in einer Entfernung von etwa 600 Metern zum
Sauenstall. Nach dem Absetzen werden alle Ferkel einer Wochengruppe zusammen
in einem Abteil eingestallt. Der Aufzuchtstall ist ein Kammstall mit neun Abteilen für je
720 Ferkel. Jedes Abteil ist gleich aufgebaut und besteht aus 24 Buchten für je 30
Ferkel, in der Mitte befindet sich der Futtergang. Vier Abteile sind für Ferkel der

58 Material und Methode
Gewichtsklasse 6 bis 15 kg bestimmt. Die anderen fünf Abteile für die darauffolgende
Gewichtsklasse ab 15 kg sind flächenmäßig größer, um dem erhöhten Flächenbedarf
der älteren Ferkel Rechnung zu tragen. Nach vier Wochen Aufzuchtzeit wird das
gesamte Abteil mit Ferkeln einer Wochengruppe in eines der größeren Abteile
umgestallt.
In jeder Bucht befindet sich ein „AP-Swing-Automat“ der Firma AgroProducts. In den
Automaten befinden sich Sensoren, die die Füllmenge des Futters im Automaten
messen und an den Fütterungscomputer weiterleiten. Somit ist eine Auswertung des
Futterverbrauchs je Bucht möglich.
Die Abluft aus den einzelnen Abteilen wird „unterflur“ abgesaugt, die Zuluft wird
deckenseitig durch Trapezlochbleche zugeführt. Das gesamte Stallklima wird über
den Lüftungscomputer (Fancom) aufgezeichnet.
An den Wandseiten der Buchten befinden sich verstellbare Zwischendecken, um für
die frisch abgesetzten Ferkel ein ihnen angepasstes Mikroklima schaffen zu können.
Geheizt wird der Stall mit warmem Wasser, das über Deltarohre in die jeweiligen
Abteile geführt wird.
In den Abteilen befinden sich Vernebelungsrohre, über die Wasserdampf zugeführt
werden kann, um die Luft anzufeuchten und somit das Stallklima zu verbessern.
Als metaphylaktische Maßnahme bekommen die Ferkel bei Einstallung in den
Aufzuchtstall über sieben Tage Amoxicillin und Colistin über das Trinkwasser
verabreicht. Die Ferkel werden in der siebten Lebenswoche mit 2 ml des Impfstoffes
Porcilis PRRS® (Intervet, 85701 Unterschleißheim) gegen PRRSV geimpft. In der
achten Lebenswoche wird ein Präparat zur Behandlung von Endo- und Ektoparasiten
über das Futter gegeben, und in der zehnten Lebenswoche erfolgt die Boosterung
der PRRS-Impfung.
Die Ferkel werden bei Einstallung in den Aufzuchtstall, beim Umstallen nach vier
Wochen und am Ende der Aufzuchtphase über eine im Stall installierte Waage
gruppen- bzw. buchtenweise gewogen.
Die weitere Mast der Tiere findet in verschiedenen Ställen statt. Alle Ferkel eines
Abteils, also eine Abferkelungsgruppe, werden zusammen in den jeweiligen Maststall
eingestallt.
Die Buchtengrößen sind je Stall unterschiedlich. In den Mastställen befinden sich
jeweils computergesteuerte Flüssigfütterungen. Es ist auch hier eine genaue
Messung und Auswertung des Futterverbrauchs möglich.
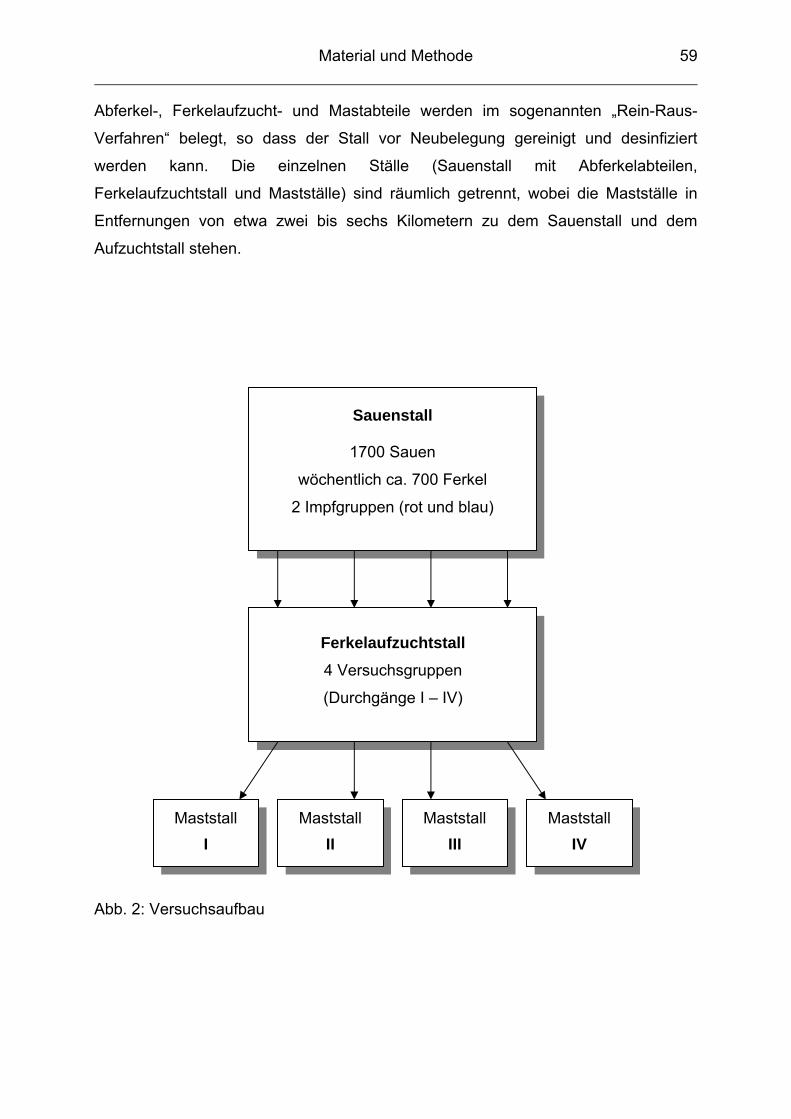
Material und Methode 59
Abferkel-, Ferkelaufzucht- und Mastabteile werden im sogenannten „Rein-Raus-
Verfahren“ belegt, so dass der Stall vor Neubelegung gereinigt und desinfiziert
werden kann. Die einzelnen Ställe (Sauenstall mit Abferkelabteilen,
Ferkelaufzuchtstall und Mastställe) sind räumlich getrennt, wobei die Mastställe in
Entfernungen von etwa zwei bis sechs Kilometern zu dem Sauenstall und dem
Aufzuchtstall stehen.
Abb. 2: Versuchsaufbau
Sauenstall
1700 Sauen
wöchentlich ca. 700 Ferkel
2 Impfgruppen (rot und blau)
Ferkelaufzuchtstall 4 Versuchsgruppen
(Durchgänge I – IV)
Maststall I
Maststall II
Maststall III
Maststall IV

60 Material und Methode
3.2 Infektionsstatus des Versuchsbetriebes
Die endemische Infektion mit H. parasuis im Untersuchungsbetrieb ist schon seit
längerer Zeit bekannt. In den Jahren 2001 bis 2004 sind mehrfach
Bronchoalveolarlavage-Flüssigkeiten (BALF) von Tieren aus dem Ferkelaufzuchtstall
genommen worden. Aus der BALF konnte H. parasuis isoliert werden. Zusätzlich
wurde der Erreger bei der pathologisch-anatomischen Untersuchung aus Tupfern
von Lungengewebe, Perikardflüssigkeit, Synovia und Brust- und Bauchhöhlen-
flüssigkeit isoliert. Klinische Erscheinungen traten vorwiegend im Ferkelaufzuchtstall
in Form von respiratorischen Erkrankungen, wie vermehrtem Husten, Niesen und
Dyspnoe, auf, außerdem wurden zeitweise vermehrt Arthritiden festgestellt.
Vor Untersuchungsbeginn wurde im Juli 2004 bei zehn Ferkeln im Ferkelaufzuchtstall
eine bronchoalveoläre Lavage durchgeführt. Vier dieser Proben enthielten H.-
parasuis-Erreger.
3.3 Vakzinen Im Rahmen der Feldstudie wurde die Effektivität einer Impfung gegen H. parasuis
und M. hyopneumoniae mit einer Impfung gegen M. hyopneumoniae verglichen. Die
Impfungen wurden mit Suvaxyn® M.hyo-Parasuis (Fort Dodge Animal Health Limited,
Southhampton) bzw. mit Suvaxyn® M.hyo (Fort Dodge Veterinär GmbH, 52146
Würselen) durchgeführt. Die intramuskuläre Applikation der Impfstoffe wurde am
Ohrgrund im Bereich des Überganges der behaarten zur unbehaarten Haut
vorgenommen.
3.3.1 Versuchsvakzine Die Erprobung des Impfstoffes Suvaxyn® M.hyo-Parasuis erfolgte auf der Grundlage
einer Ausnahmegenehmigung gemäß § 17c, Absatz 4, Nr. 2 des
Tierseuchengesetzes durch das Niedersächsische Ministerium für den ländlichen
Raum, Ernährung, Landwirtschaft und Verbraucherschutz mit Bescheid vom
04.08.2004.

Material und Methode 61
Suvaxyn® M.hyo-Parasuis ist ein inaktivierter Impfstoff in wässriger Lösung. Eine
Impfdosis (2 ml) enthält mindestens 2 x 109 MHDCE (Mycoplasma hyopneumoniae
DNA Cell Equivalence), mindestens 108 DCE (DNA Cell Equivalence) Haemophilus
parasuis Serotyp 4 und mindestens 108 DCE (DNA Cell Equivalence) Haemophilus
parasuis Serotyp 5. Als Adjuvans dient 4,0 mg Carbopol 941. Zusätzlich sind je
Impfdosis maximal 0,2 mg Thiomersal als Konservierungsmittel zugesetzt. Nach
Herstellerangaben können Ferkel ab dem Alter von einer Woche geimpft werden, die
zweite Impfung sollte im Abstand von zwei bis drei Wochen erfolgen. In dieser Studie
sind die Ferkel am zehnten Lebenstag und in der dritten Lebenswoche geimpft
worden.
3.3.2 Kontrollvakzine Suvaxyn® M.hyo ist ein inaktivierter Impfstoff in wässriger Lösung. Eine Impfdosis (2
ml) enthält mindestens 2 x 109 MHDCE (Mycoplasma hyopneumoniae DNA Cell
Equivalence). Als Adjuvans dient 4,0 mg Carbopol 941. Zusätzlich sind je Impfdosis
maximal 0,2 mg Thiomersal als Konservierungsmittel zugesetzt. Nach Hersteller-
angaben können Ferkel ab dem Alter von drei Tagen geimpft werden, die zweite
Impfung sollte im Abstand von zwei bis drei Wochen erfolgen. In dieser Studie sind
die Ferkel am zehnten Lebenstag und in der dritten Lebenswoche geimpft worden.
3.4 Versuchsgruppen
Grundlage für den Versuch waren alle Ferkel einer Wochenabferkelung.
Die Ferkel einer Abferkelgruppe wurden in den ersten Lebenstagen nach dem
Zufallsprinzip den zwei Gruppen (rot und blau) zugeteilt. Unterschieden wurden sie
durch verschiedenfarbige Ohrmarken. Die Hälfte der Ferkel eines Wurfes wurden der
Impfgruppe rot (Suvaxyn® M.hyo-Parasuis; rote Ohrmarken) zugeordnet, die andere
Hälfte der Impfgruppe blau (Suvaxyn® M.hyo; blaue Ohrmarken). Die Ferkel der
beiden Versuchsgruppen wurden jeweils am zehnten Lebenstag und in der dritten
Lebenswoche mit je zwei Milliliter Impfstoff (Suvaxyn® M.hyo-Parasuis bzw.
Suvaxyn® M.hyo) geimpft.

62 Material und Methode
Bei je fünf Ferkeln pro Gruppe wurden zusätzlich durchnummerierte Ohrmarken
eingezogen, von diesen Ferkeln wurden regelmäßig Blutproben gezogen.
Die Ferkel einer Absetzgruppe wurden in einem Abteil mit je 24 Buchten aufgestallt.
Buchtenweise gehören sie einer Versuchsgruppe (gleiche Ohrmarkenfarbe) an, das
heißt, alle Tiere einer Bucht besitzen den gleichen Impfstatus. Diese
Gruppenzusammensetzung wurde bis zum Ende der Mast beibehalten.
Die Ferkel wurden bei Einstallung in den Aufzuchtstall, beim Umstallen nach vier
Wochen und am Ende der Aufzuchtphase auf einer im Stall installierten Waage
gruppen- bzw. buchtenweise gewogen. Die Anzahl der Ferkel einer Bucht betrug im
Ferkelaufzuchtstall 30 Tiere. Die Anzahl der Tiere pro Bucht in den Mastställen
variierte stallbedingt. Die Tiere wurden demnach vor der Einstallung in die Mast
erneut gewogen, um auch hier eine Feststellung und Auswertung der
Tageszunahmen zu ermöglichen.
Um die Tiere am Schlachthof der jeweiligen Impf- bzw. Versuchsgruppe zuordnen zu
können, sind sie schon im Betrieb per Schlagstempel mit der Betriebsnummer und
einer zusätzlichen fortlaufenden Nummer je Gruppe und Schlachttag gekennzeichnet
worden.
Um die Reproduzierbarkeit der Ergebnisse zu prüfen, wurde der Versuch in vier
Durchgängen (I, II, III und IV) durchgeführt.
Versuchsgruppe rot: Suvaxyn® M.hyo-Parasuis
Kontrollgruppe blau: Suvaxyn® M.hyo
I, II, III, IV: Versuch in vier Durchgängen durchgeführt
Tab. 2: Versuchsgruppen und eingesetzte Impfstoffe
Gruppe / Durchgang Anzahl Impfstoff
rot (I, II, III, IV) ca. 1400 Suvaxyn® M.hyo-Parasuis
blau (I, II, III, IV) ca. 1400 Suvaxyn® M.hyo
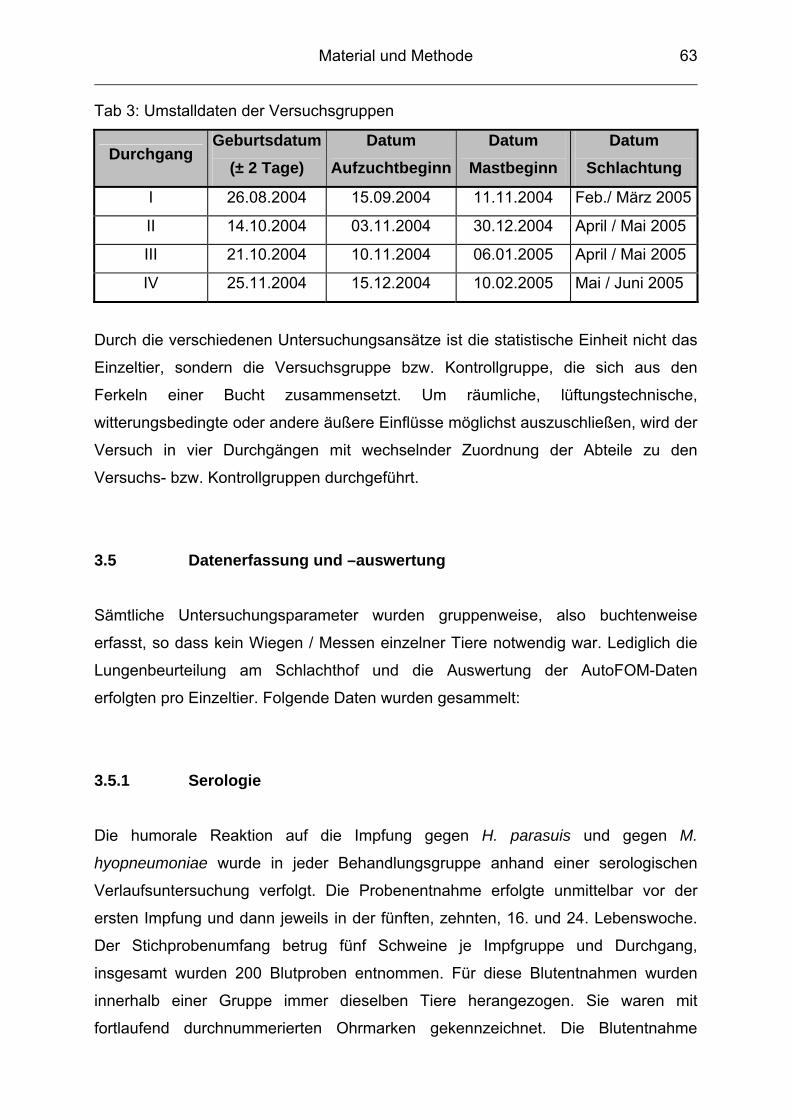
Material und Methode 63
Tab 3: Umstalldaten der Versuchsgruppen
Durchgang Geburtsdatum
(± 2 Tage) Datum
AufzuchtbeginnDatum
Mastbeginn Datum
Schlachtung
I 26.08.2004 15.09.2004 11.11.2004 Feb./ März 2005
II 14.10.2004 03.11.2004 30.12.2004 April / Mai 2005
III 21.10.2004 10.11.2004 06.01.2005 April / Mai 2005
IV 25.11.2004 15.12.2004 10.02.2005 Mai / Juni 2005
Durch die verschiedenen Untersuchungsansätze ist die statistische Einheit nicht das
Einzeltier, sondern die Versuchsgruppe bzw. Kontrollgruppe, die sich aus den
Ferkeln einer Bucht zusammensetzt. Um räumliche, lüftungstechnische,
witterungsbedingte oder andere äußere Einflüsse möglichst auszuschließen, wird der
Versuch in vier Durchgängen mit wechselnder Zuordnung der Abteile zu den
Versuchs- bzw. Kontrollgruppen durchgeführt.
3.5 Datenerfassung und –auswertung
Sämtliche Untersuchungsparameter wurden gruppenweise, also buchtenweise
erfasst, so dass kein Wiegen / Messen einzelner Tiere notwendig war. Lediglich die
Lungenbeurteilung am Schlachthof und die Auswertung der AutoFOM-Daten
erfolgten pro Einzeltier. Folgende Daten wurden gesammelt:
3.5.1 Serologie
Die humorale Reaktion auf die Impfung gegen H. parasuis und gegen M.
hyopneumoniae wurde in jeder Behandlungsgruppe anhand einer serologischen
Verlaufsuntersuchung verfolgt. Die Probenentnahme erfolgte unmittelbar vor der
ersten Impfung und dann jeweils in der fünften, zehnten, 16. und 24. Lebenswoche.
Der Stichprobenumfang betrug fünf Schweine je Impfgruppe und Durchgang,
insgesamt wurden 200 Blutproben entnommen. Für diese Blutentnahmen wurden
innerhalb einer Gruppe immer dieselben Tiere herangezogen. Sie waren mit
fortlaufend durchnummerierten Ohrmarken gekennzeichnet. Die Blutentnahme

64 Material und Methode
erfolgte bei ein bis fünf Wochen alten Ferkeln aus der Vena cava cranialis und bei
älteren Tieren aus der Vena jugularis externa. Das Blut wurde in 10 ml
Serummonovetten (Sarstedt) aufgefangen. Die Monovetten wurden mit der
Ohrmarkennummer des Schweines beschriftet. Im Labor der Praxis WEK wurde das
Serum abzentrifugiert, auf zwei 2 ml Reaktionsgefäße aufgeteilt und bis zur
Untersuchung bei –20° C eingelagert. Die serologischen Untersuchungen auf
Antikörper gegen H. parasuis Serovar 4 und 5 und M. hyopneumoniae wurden zu
einem Zeitpunkt durchgeführt, an dem alle Proben vorlagen. Die Untersuchungen
sind im Labor Fort Dodge Veterinaria (17813 Vall de Bianya, Spanien) durchgeführt
worden.
3.5.1.1 Untersuchung auf Antikörper gegen H. parasuis Serovar 4 und 5 Die Blutproben wurden mittels eines Enzyme Linked Immunosorbent Assay (ELISA)
auf Antikörper gegen H. parasuis Serovar 4 und Serovar 5 untersucht. Bei dem Test
handelte es sich um einen ELISA der Firma Fort Dodge. Als Konjugat wurden H.
parasuis Serovar 4 -Antigen bzw. abgetötete Ganzzellen von H. parasuis Serovar 5
verwendet. Die Extinktion des Serovars 4 wurde bei einer Wellenlänge von 405 nm
und 490 nm gemessen, die des Serovars 5 bei einer Wellenlänge von 405 nm.
3.5.1.2 Untersuchung auf Antikörper gegen M. hyopneumoniae Die Blutproben wurden mittels eines Enzyme Linked Immunosorbent Assay (ELISA)
auf Antikörper gegen M. hyopneumoniae untersucht. Bei dem Test handelte es sich
um einen blocking-ELISA (Mycoplasma hyopneumoniae ELISA, K0043, Firma
DakoCytomation, Cambridgeshire, United Kingdom). Als Konjugat wurden hoch
spezifizierte monoklonale Antikörper gegen das Epitop des 74Kda Protein von M.
hyopneumoniae verwendet. Die Extinktion wurde bei einer Wellenlänge von 450 nm
gemessen. Die Sensitivität dieses ELISA wird anhand von Proben von experimentell
infizierten Schweinen mit 98 bis 100 % und die Spezifität mit 93 bis 100 % (ermittelt
mit Hilfe von Proben von SPF-Kontrolltieren) angegeben (SØRENSEN et al. 1997).

Material und Methode 65
3.5.2 Durchschnittliche tägliche Gewichtszunahme
Mit dem Ziel, den Impferfolg einzuschätzen, wurden die täglichen Zunahmen
gemessen. Der Zuwachs der Tiere wurde buchtenweise anhand von regelmäßigen
Wiegungen zu verschiedenen Zeitpunkten während der Aufzucht und der Mast
festgestellt.
Da das Geburtsgewicht im Vergleich zum Mastendgewicht nur einer geringen
Variationsbreite unterliegt, wurde für alle Ferkel ein Geburtsgewicht von 1,4 kg
angenommen. Untergewichtige Ferkel wurden vom Versuch ausgeschlossen. Die
Gewichtsentwicklung wurde durch Wiegungen jeweils buchtenweise zum Zeitpunkt
des Absetzens, im Alter von sieben Wochen, am Ende der Aufzucht und bei der
Aufstallung zur Mast ermittelt. Im Aufzuchtstall befand sich eine Durchtreibewaage,
bei der buchtenweises Wiegen möglich war. Je nach Art des Maststalles wurden die
Ferkel beim Verladen buchtenweise mit Sprühfarbe gekennzeichnet und mit der
gleichen Zusammensetzung wieder eingestallt. Das Lebendgewicht bei Mastende
wurde aus dem Schlachtgewicht unter Annahme einer 80 % igen Ausschlachtung
des Lebendgewichtes berechnet.
Die Identifikation und Zuordnung der Schweine zu den einzelnen Gruppen erfolgte
anhand einer fortlaufenden Tätowiernummer, die mittels Schlageisen bereits einige
Wochen vor der Schlachtung angebracht worden war.
3.5.3 Futterverwertung
Die Daten der Futterverwertung geben Auskunft darüber, wie viel kg Futter für 1 kg
Körpermassezuwachs (KMZ) benötigt wird. Die Futterverwertung bezieht sich auf
den Gesamtzuwachs inklusive verendeter Tiere.
Die Futterverwertung in der Säugezeit wird nicht berücksichtigt.
In der Aufzucht wurde die Futtermenge durch die sensorengesteuerte
Computerfütterung an den AP-Swing-Automaten buchtenweise gemessen. So
konnte eine genaue Berechnung und Auswertung der Futterverwertung
vorgenommen werden.
In den Mastställen befinden sich computergesteuerte Flüssigfütterungen, die ebenso
eine Auswertung je Bucht ermöglichten.

66 Material und Methode
3.5.4 Durchschnittliche Mastdauer
Die durchschnittliche Mastdauer wird als wirtschaftliches Kennzeichen in der
Schweinemast genutzt. Die Mastdauer wird berechnet vom durchschnittlichen Wert
vom Einstalltag in die Mast bis zum Schlachttag, verendete Tiere und vorzeitig
umgestallte Tiere werden herausgerechnet.
3.5.5 Mortalität
Die Mortalität der einzelnen Behandlungsgruppen wurde pro Versuchsgruppe
erfasst. Alle verendeten Schweine wurden der tierärztlichen pathologisch-
anatomischen Untersuchung zugeführt und insbesondere auf Lungenveränderungen
und Veränderungen der serösen Häute und der Gelenke (Glässersche Krankheit)
untersucht.
3.5.6 Therapeutische Maßnahmen / Tierbehandlungsindex (TBI)
Klinisch manifeste Erkrankungen wurden aus Tierschutzgründen je nach gestellter
Diagnose und Indikation behandelt. Die jeweilige Diagnosestellung erfolgte durch die
Versuchsleiterin, die entsprechende Arzneimittelanwendung wurde abteil- bzw.
buchtenweise im Stallbuch dokumentiert. So konnte eine Auswertung der
Einzeltierbehandlungen und der eventuellen Medikation über das Futter oder das
Trinkwasser je Versuchsgruppe während des Versuchszeitraumes vorgenommen
werden. Zur Quantifizierung der Behandlungen wurde der „Tierbehandlungsindex“
(TBI) nach BLAHA (2005) verwendet. Der TBI gibt die durchschnittliche Anzahl von
Tagen an, die jedes Tier einer Impfgruppe mit einer antimikrobiellen Substanz (oral
oder parenteral) versorgt wurde.
Der Tierbehandlungsindex berechnet sich wie folgt:
Anzahl der Tiere, die behandelt wurden x Anzahl der Behandlungstage TBI =
Anzahl der Tiere in der Gruppe

Material und Methode 67
Um einen besseren Vergleich der beiden Behandlungsgruppen zu bekommen ist der
Tierbehandlungsindex mit zwei verschiedenen Grundlagen berechnet worden.
Der „Tierbehandlungsindex gesamt“ (TBIges.) beinhaltet sämtliche antibiotischen
Behandlungen, wie Bestandsbehandlungen, die allen Tieren desselben Durchgangs
verabreicht wurden, und alle Einzeltierbehandlungen. Der „Tierbehandlungsindex
einzel“ (TBIeinzel) beinhaltet nur Behandlungen, die individuell an einzelnen Gruppen
oder an einzelnen Tieren durchgeführt worden sind. In diesen Index gehen keine
Behandlungen ein, die bei allen Tieren durchgeführt wurden. Vakzinierungen und
Behandlungen gegen Endo- und Ektoparasitenbefall werden in keinem Index
berücksichtigt, da diese Maßnahmen bei allen Tieren durchgeführt wurden und keine
Auswirkungen auf die Effekte der Impfungen erwartet werden.
3.5.7 Beurteilung von Lungen nach der Schlachtung / Lungenscore
Das Vorkommen von pneumonischen Veränderungen ist anhand von
Organuntersuchungen bei der Schlachtung beurteilt worden. Die Lungen wurden
adspektorisch und palpatorisch untersucht und die Qualität der Veränderungen nach
dem Score von MADEC und KOBISCH (1985) klassifiziert. Der Score sieht vor, jeden
einzelnen Lungenlappen entsprechend dem Ausmaß der Veränderungen mit einem
Wert zwischen 0 bis 4 zu beschreiben (Tab. 4). Die Werte der sieben Lungenlappen
werden summiert und ergeben so einen Gesamtwert zwischen 0 und 28. Das
Vorkommen von Abszessen in der Lunge wird mit Werten von 1 oder 2 beurteilt.
Pleuritiden, die mit Verklebungen einhergehen, werden mit 1, solche, die zu
Verwachsungen geführt haben mit 2 bewertet.
Um die Tiere am Schlachthof individuell bzw. den Gruppen zugehörig unterscheiden
zu können, wurden sie vor dem Abliefern zusätzlich zur Betriebskennzeichnung
fortlaufend (je Bucht eine Kennzeichnung) mit dem Schlagstempel durchnummeriert
und so gekennzeichnet.
Die Organbefundung wurde durch die Versuchsleiterin selbst durchgeführt, um
subjektive Beurteilungsunterschiede auszuschließen. Die Lungen wurden im
Rahmen des normalen Schlachtprozesses am „Organband“ beurteilt. Ein weiterer
Untersucher notierte am „Tierkörperband“ die individuelle Tätowierung und die
fortlaufende Schlachtnummer. Beide Bänder laufen in der gleichen Reihenfolge
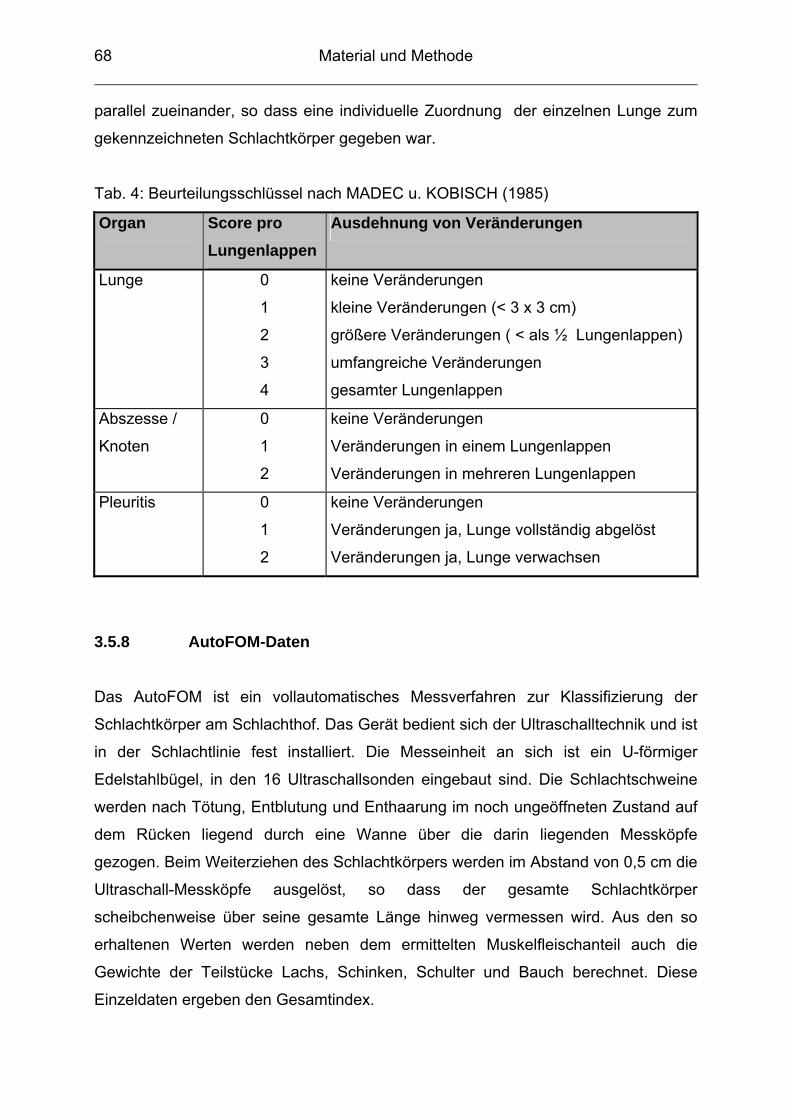
68 Material und Methode
parallel zueinander, so dass eine individuelle Zuordnung der einzelnen Lunge zum
gekennzeichneten Schlachtkörper gegeben war.
Tab. 4: Beurteilungsschlüssel nach MADEC u. KOBISCH (1985)
Organ Score pro Lungenlappen
Ausdehnung von Veränderungen
Lunge 0
1
2
3
4
keine Veränderungen
kleine Veränderungen (< 3 x 3 cm)
größere Veränderungen ( < als ½ Lungenlappen)
umfangreiche Veränderungen
gesamter Lungenlappen
Abszesse /
Knoten
0
1
2
keine Veränderungen
Veränderungen in einem Lungenlappen
Veränderungen in mehreren Lungenlappen
Pleuritis 0
1
2
keine Veränderungen
Veränderungen ja, Lunge vollständig abgelöst
Veränderungen ja, Lunge verwachsen
3.5.8 AutoFOM-Daten
Das AutoFOM ist ein vollautomatisches Messverfahren zur Klassifizierung der
Schlachtkörper am Schlachthof. Das Gerät bedient sich der Ultraschalltechnik und ist
in der Schlachtlinie fest installiert. Die Messeinheit an sich ist ein U-förmiger
Edelstahlbügel, in den 16 Ultraschallsonden eingebaut sind. Die Schlachtschweine
werden nach Tötung, Entblutung und Enthaarung im noch ungeöffneten Zustand auf
dem Rücken liegend durch eine Wanne über die darin liegenden Messköpfe
gezogen. Beim Weiterziehen des Schlachtkörpers werden im Abstand von 0,5 cm die
Ultraschall-Messköpfe ausgelöst, so dass der gesamte Schlachtkörper
scheibchenweise über seine gesamte Länge hinweg vermessen wird. Aus den so
erhaltenen Werten werden neben dem ermittelten Muskelfleischanteil auch die
Gewichte der Teilstücke Lachs, Schinken, Schulter und Bauch berechnet. Diese
Einzeldaten ergeben den Gesamtindex.

Material und Methode 69
3.6 Statistische Methoden
Die Datenerfassung erfolgte mit Hilfe eines Programms zur Tabellenkalkulation
(Microsoft® Excel 2000). Für die statistische Auswertung wurden die Rohdaten
anschließend in das Programm SPSS for Windows (Version 11.5; SPSS Inc.,
Chicago, Illinois, USA) übertragen.
Die Überprüfung der Normalverteilung der einzelnen Parameter wurde mit einem
NPar Test (non parametic test), wie dem One-sample Kolmogorov-Smirnov Test,
durchgeführt.
War eine Normalverteilung der Werte der einzelnen Parameter gegeben, wurde eine
Varianzanalyse durchgeführt. Mit dem Levene`s Test wurde dann überprüft, ob der
T-Test angewendet werden kann.
Eine normale Verteilung wurde bei den Parametern durchschnittliche Mastdauer,
Alter bei Schlachtung, Pneumonie, AutoFOM, Tageszunahmen und Futterverwertung
gefunden. Für diese Werte wurde als Signifikanztest der T-Test angewendet. Das
Signifikanzniveau wurde bei p>0,05 festgelegt.
Lediglich für die Blutprobenauswertung wurde als Signifikanztest der Anova-Test
durchgeführt.
Die Parameter Schlachtgewicht, TBI, Mortalität, Pleuritis und Abszesse zeigten keine
Normalverteilung auf. Hier wurde der Mann-Whitney-Test angewendet.

70 Ergebnisse
4 Ergebnisse
4.1 Allgemeines Im Rahmen der vorliegenden Dissertation wurde in einem Sauenbetrieb mit
angeschlossener Aufzucht und anschließender Mast in vier Durchgängen ein
Kombinationsimpfstoff (Suvaxyn® M.hyo-Parasuis) gegen H. parasuis und M.
hyopneumoniae gegenüber einem monovalenten Impfstoff gegen M.hyopneumoniae
(Suvaxyn® M.hyo) getestet. Auswahlkriterium für diesen Betrieb war zum einen das
endemische Vorkommen von H. parasuis in dem betroffenen Bestand; zum anderen
sind Verlaufsuntersuchungen, in denen Saugferkel bis zum Mastende verfolgt
werden sollen, nur in Beständen mit eigener Aufzucht und Mast oder Beständen mit
konstanten Lieferbeziehungen durchführbar.
Insgesamt sind 3132 Ferkel in vier verschiedenen Wochenabferkelungen geimpft
worden; 1566 Ferkel haben den Kombinationsimpfstoff Suvaxyn® M.hyo-Parasuis
(Impfgruppe rot) erhalten und 1566 Ferkel den Monoimpfstoff Suvaxyn® M.hyo
(Impfgruppe blau). Im Ferkelaufzuchtstall sind davon 2691 Ferkel ausgewertet
worden; in der Mastperiode konnten davon 2107 Tiere bis zum Schlachthof verfolgt
werden. Die Differenzen zwischen den einzelnen Zahlen sind auf
betriebsorganisatorische Bedingungen zurückzuführen, da zum einen gleichgroße
Gruppen in den Versuchsreihen ausgewertet werden sollten und zum anderen die
Maststallgröße nicht immer konform zu der Anzahl ausgestallter Tiere aus dem
Aufzuchtstall war. Überzählige Tiere aus den größeren Gruppen mussten aus dem
Versuch ausscheiden. Zusätzlich konnte am Schlachthof ein Teil der Tiere anhand
der Tätowierung nicht eindeutig identifiziert und demnach auch nicht bewertet
werden.
Um die beiden Versuchsgruppen zu unterscheiden, sind die Ferkel, die mit Suvaxyn®
M.hyo-Parasuis geimpft wurden, mit roten Ohrmarken gekennzeichnet worden, die
mit Suvaxyn® M.hyo geimpften Ferkel erhielten blaue Ohrmarken.
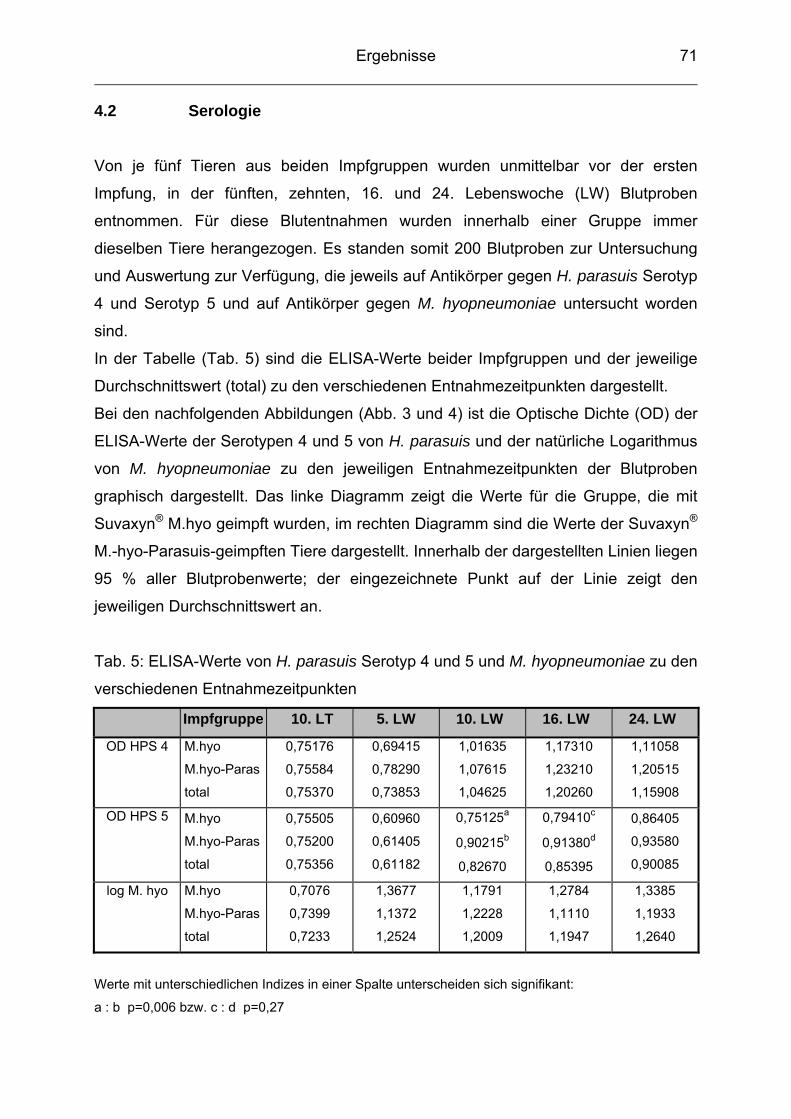
Ergebnisse 71
4.2 Serologie
Von je fünf Tieren aus beiden Impfgruppen wurden unmittelbar vor der ersten
Impfung, in der fünften, zehnten, 16. und 24. Lebenswoche (LW) Blutproben
entnommen. Für diese Blutentnahmen wurden innerhalb einer Gruppe immer
dieselben Tiere herangezogen. Es standen somit 200 Blutproben zur Untersuchung
und Auswertung zur Verfügung, die jeweils auf Antikörper gegen H. parasuis Serotyp
4 und Serotyp 5 und auf Antikörper gegen M. hyopneumoniae untersucht worden
sind.
In der Tabelle (Tab. 5) sind die ELISA-Werte beider Impfgruppen und der jeweilige
Durchschnittswert (total) zu den verschiedenen Entnahmezeitpunkten dargestellt.
Bei den nachfolgenden Abbildungen (Abb. 3 und 4) ist die Optische Dichte (OD) der
ELISA-Werte der Serotypen 4 und 5 von H. parasuis und der natürliche Logarithmus
von M. hyopneumoniae zu den jeweiligen Entnahmezeitpunkten der Blutproben
graphisch dargestellt. Das linke Diagramm zeigt die Werte für die Gruppe, die mit
Suvaxyn® M.hyo geimpft wurden, im rechten Diagramm sind die Werte der Suvaxyn®
M.-hyo-Parasuis-geimpften Tiere dargestellt. Innerhalb der dargestellten Linien liegen
95 % aller Blutprobenwerte; der eingezeichnete Punkt auf der Linie zeigt den
jeweiligen Durchschnittswert an.
Tab. 5: ELISA-Werte von H. parasuis Serotyp 4 und 5 und M. hyopneumoniae zu den
verschiedenen Entnahmezeitpunkten
Impfgruppe 10. LT 5. LW 10. LW 16. LW 24. LW
OD HPS 4 M.hyo
M.hyo-Paras
total
0,75176
0,75584
0,75370
0,69415
0,78290
0,73853
1,01635
1,07615
1,04625
1,17310
1,23210
1,20260
1,11058
1,20515
1,15908
OD HPS 5 M.hyo
M.hyo-Paras
total
0,75505
0,75200
0,75356
0,60960
0,61405
0,61182
0,75125a
0,90215b
0,82670
0,79410c
0,91380d
0,85395
0,86405
0,93580
0,90085
log M. hyo M.hyo
M.hyo-Paras
total
0,7076
0,7399
0,7233
1,3677
1,1372
1,2524
1,1791
1,2228
1,2009
1,2784
1,1110
1,1947
1,3385
1,1933
1,2640
Werte mit unterschiedlichen Indizes in einer Spalte unterscheiden sich signifikant:
a : b p=0,006 bzw. c : d p=0,27
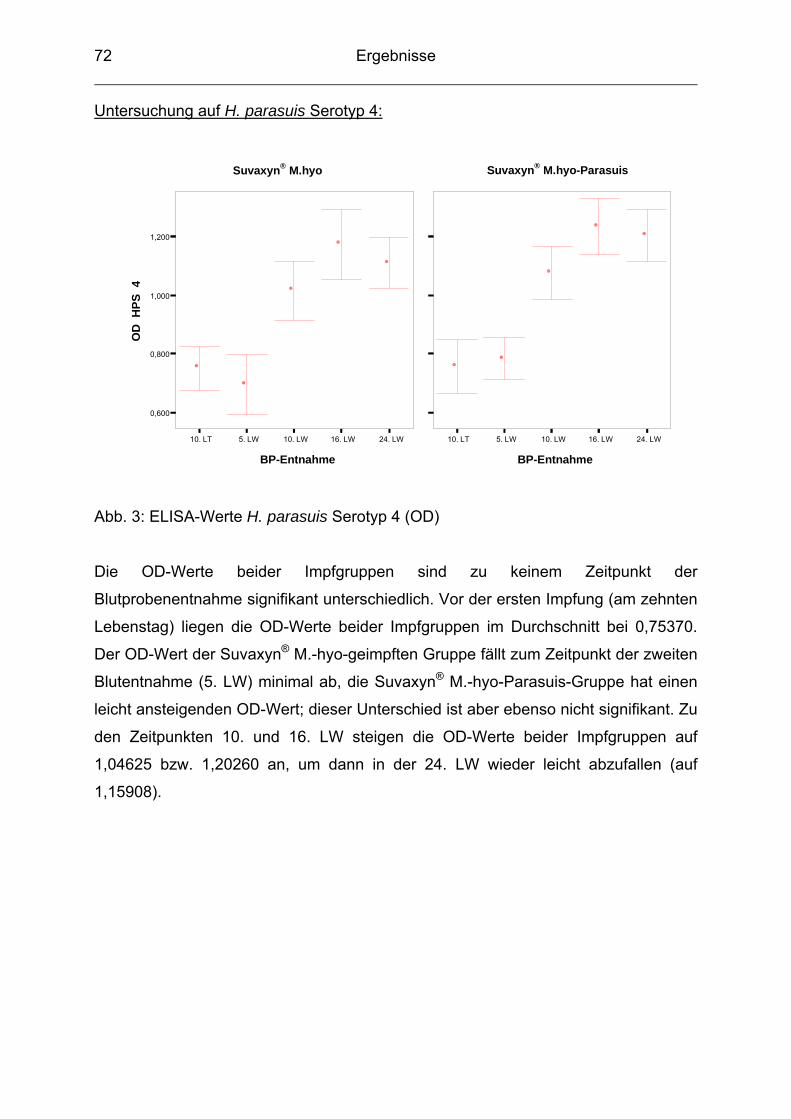
72 Ergebnisse
Untersuchung auf H. parasuis Serotyp 4:
10. LT 5. LW 10. LW 16. LW 24. LW
BP-Entnahme
0,600
0,800
1,000
1,200
OD
HPS
4
Suvaxyn® M.hyo Suvaxyn® M.hyo-Parasuis
10. LT 5. LW 10. LW 16. LW 24. LW
BP-Entnahme
Abb. 3: ELISA-Werte H. parasuis Serotyp 4 (OD)
Die OD-Werte beider Impfgruppen sind zu keinem Zeitpunkt der
Blutprobenentnahme signifikant unterschiedlich. Vor der ersten Impfung (am zehnten
Lebenstag) liegen die OD-Werte beider Impfgruppen im Durchschnitt bei 0,75370.
Der OD-Wert der Suvaxyn® M.-hyo-geimpften Gruppe fällt zum Zeitpunkt der zweiten
Blutentnahme (5. LW) minimal ab, die Suvaxyn® M.-hyo-Parasuis-Gruppe hat einen
leicht ansteigenden OD-Wert; dieser Unterschied ist aber ebenso nicht signifikant. Zu
den Zeitpunkten 10. und 16. LW steigen die OD-Werte beider Impfgruppen auf
1,04625 bzw. 1,20260 an, um dann in der 24. LW wieder leicht abzufallen (auf
1,15908).
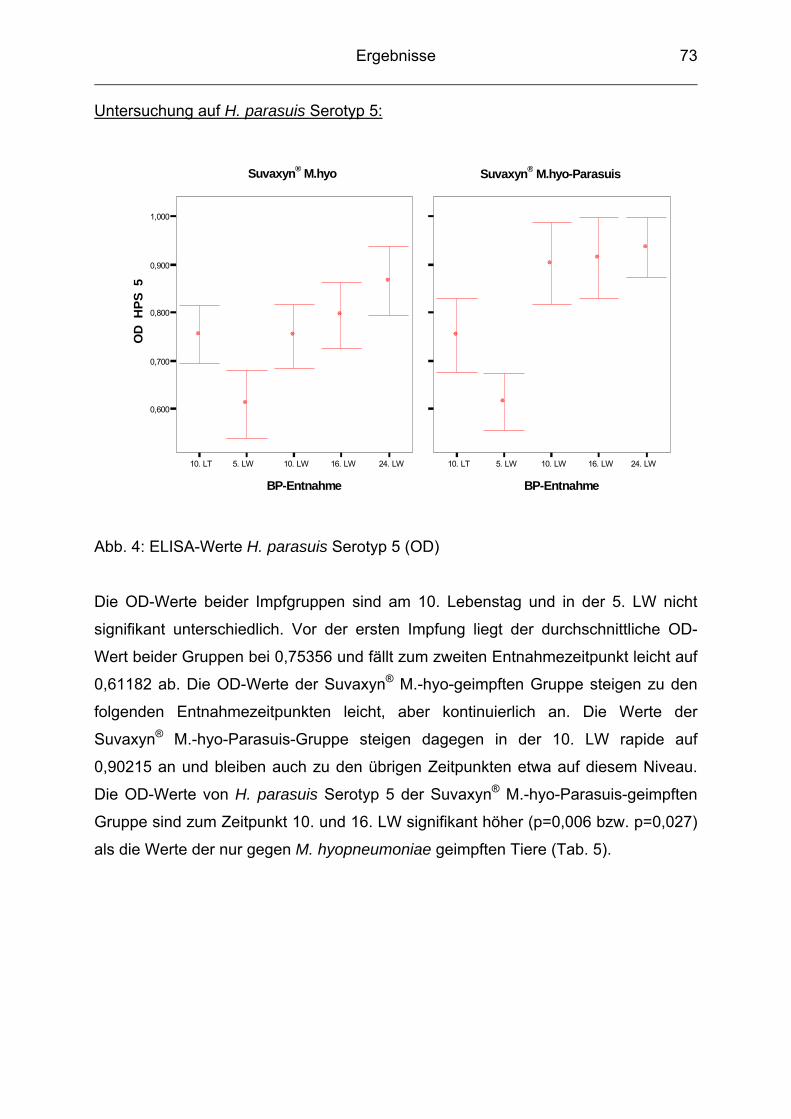
Ergebnisse 73
Untersuchung auf H. parasuis Serotyp 5:
10. LT 5. LW 10. LW 16. LW 24. LW
BP-Entnahme
0,600
0,700
0,800
0,900
1,000
OD
HPS
5
Suvaxyn® M.hyo Suvaxyn® M.hyo-Parasuis
10. LT 5. LW 10. LW 16. LW 24. LW
BP-Entnahme
Abb. 4: ELISA-Werte H. parasuis Serotyp 5 (OD)
Die OD-Werte beider Impfgruppen sind am 10. Lebenstag und in der 5. LW nicht
signifikant unterschiedlich. Vor der ersten Impfung liegt der durchschnittliche OD-
Wert beider Gruppen bei 0,75356 und fällt zum zweiten Entnahmezeitpunkt leicht auf
0,61182 ab. Die OD-Werte der Suvaxyn® M.-hyo-geimpften Gruppe steigen zu den
folgenden Entnahmezeitpunkten leicht, aber kontinuierlich an. Die Werte der
Suvaxyn® M.-hyo-Parasuis-Gruppe steigen dagegen in der 10. LW rapide auf
0,90215 an und bleiben auch zu den übrigen Zeitpunkten etwa auf diesem Niveau.
Die OD-Werte von H. parasuis Serotyp 5 der Suvaxyn® M.-hyo-Parasuis-geimpften
Gruppe sind zum Zeitpunkt 10. und 16. LW signifikant höher (p=0,006 bzw. p=0,027)
als die Werte der nur gegen M. hyopneumoniae geimpften Tiere (Tab. 5).
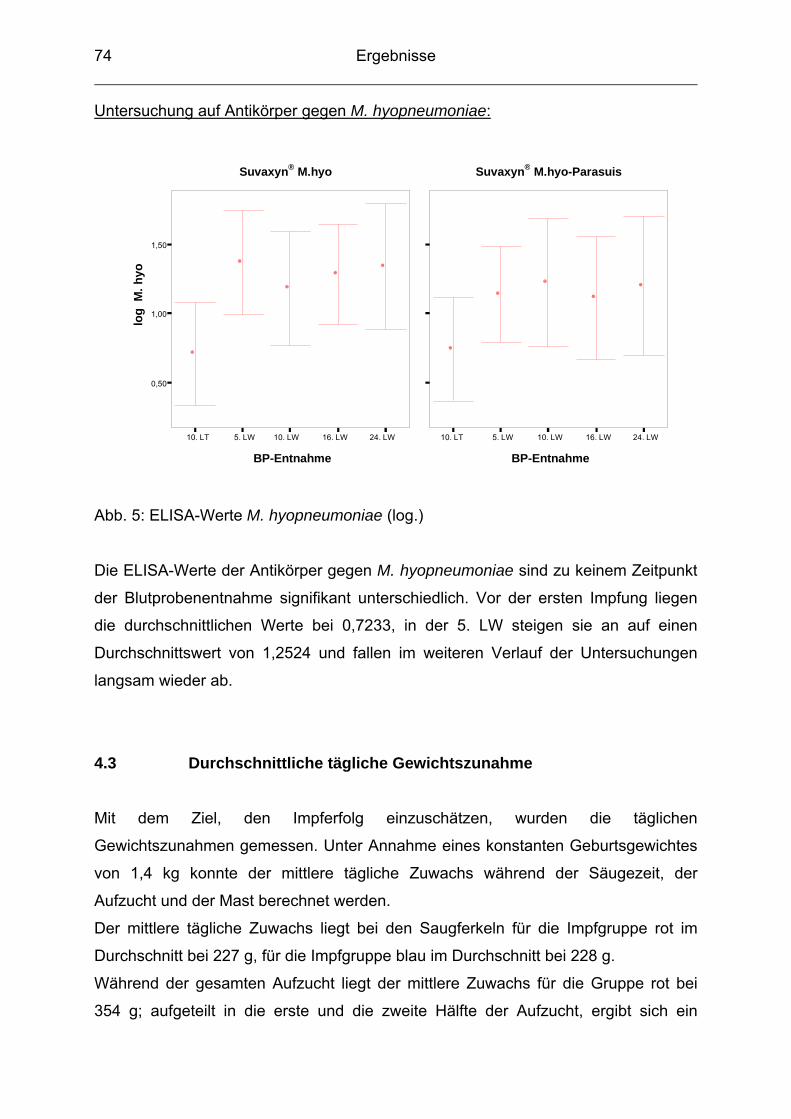
74 Ergebnisse
Untersuchung auf Antikörper gegen M. hyopneumoniae:
10. LT 5. LW 10. LW 16. LW 24. LW
BP-Entnahme
0,50
1,00
1,50
log
M. h
yo
Suvaxyn® M.hyo Suvaxyn® M.hyo-Parasuis
10. LT 5. LW 10. LW 16. LW 24. LW
BP-Entnahme
Abb. 5: ELISA-Werte M. hyopneumoniae (log.)
Die ELISA-Werte der Antikörper gegen M. hyopneumoniae sind zu keinem Zeitpunkt
der Blutprobenentnahme signifikant unterschiedlich. Vor der ersten Impfung liegen
die durchschnittlichen Werte bei 0,7233, in der 5. LW steigen sie an auf einen
Durchschnittswert von 1,2524 und fallen im weiteren Verlauf der Untersuchungen
langsam wieder ab.
4.3 Durchschnittliche tägliche Gewichtszunahme
Mit dem Ziel, den Impferfolg einzuschätzen, wurden die täglichen
Gewichtszunahmen gemessen. Unter Annahme eines konstanten Geburtsgewichtes
von 1,4 kg konnte der mittlere tägliche Zuwachs während der Säugezeit, der
Aufzucht und der Mast berechnet werden.
Der mittlere tägliche Zuwachs liegt bei den Saugferkeln für die Impfgruppe rot im
Durchschnitt bei 227 g, für die Impfgruppe blau im Durchschnitt bei 228 g.
Während der gesamten Aufzucht liegt der mittlere Zuwachs für die Gruppe rot bei
354 g; aufgeteilt in die erste und die zweite Hälfte der Aufzucht, ergibt sich ein
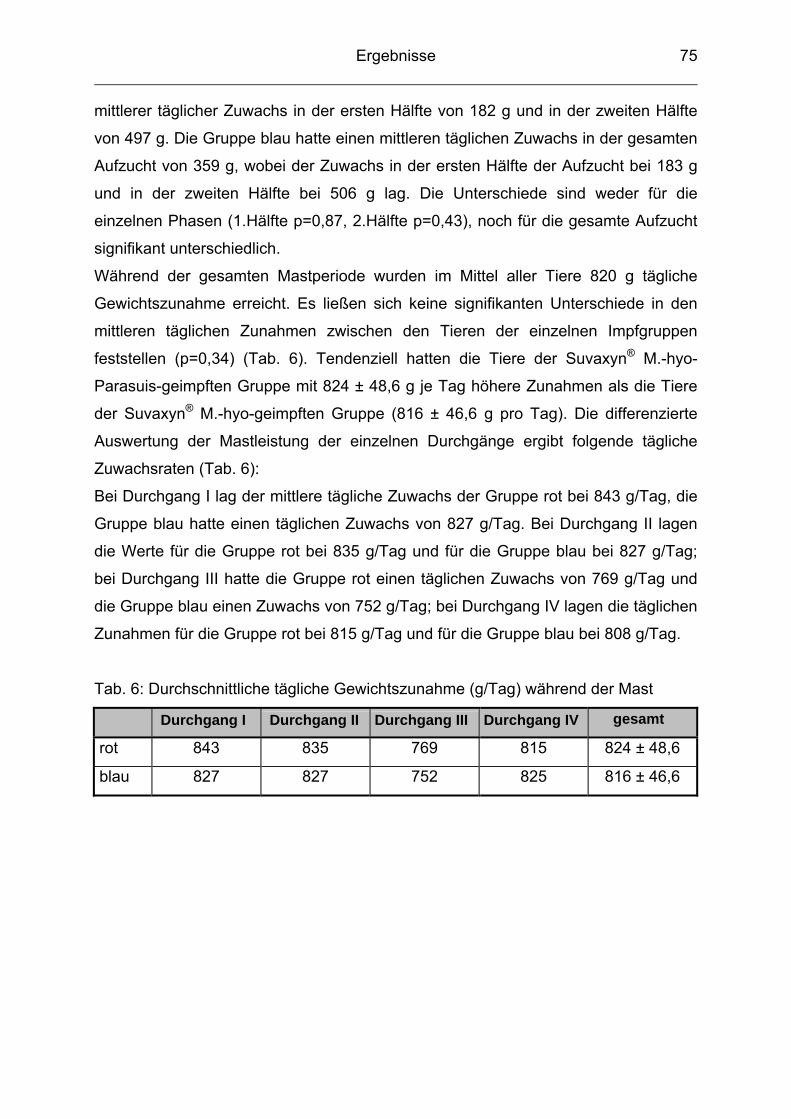
Ergebnisse 75
mittlerer täglicher Zuwachs in der ersten Hälfte von 182 g und in der zweiten Hälfte
von 497 g. Die Gruppe blau hatte einen mittleren täglichen Zuwachs in der gesamten
Aufzucht von 359 g, wobei der Zuwachs in der ersten Hälfte der Aufzucht bei 183 g
und in der zweiten Hälfte bei 506 g lag. Die Unterschiede sind weder für die
einzelnen Phasen (1.Hälfte p=0,87, 2.Hälfte p=0,43), noch für die gesamte Aufzucht
signifikant unterschiedlich.
Während der gesamten Mastperiode wurden im Mittel aller Tiere 820 g tägliche
Gewichtszunahme erreicht. Es ließen sich keine signifikanten Unterschiede in den
mittleren täglichen Zunahmen zwischen den Tieren der einzelnen Impfgruppen
feststellen (p=0,34) (Tab. 6). Tendenziell hatten die Tiere der Suvaxyn® M.-hyo-
Parasuis-geimpften Gruppe mit 824 ± 48,6 g je Tag höhere Zunahmen als die Tiere
der Suvaxyn® M.-hyo-geimpften Gruppe (816 ± 46,6 g pro Tag). Die differenzierte
Auswertung der Mastleistung der einzelnen Durchgänge ergibt folgende tägliche
Zuwachsraten (Tab. 6):
Bei Durchgang I lag der mittlere tägliche Zuwachs der Gruppe rot bei 843 g/Tag, die
Gruppe blau hatte einen täglichen Zuwachs von 827 g/Tag. Bei Durchgang II lagen
die Werte für die Gruppe rot bei 835 g/Tag und für die Gruppe blau bei 827 g/Tag;
bei Durchgang III hatte die Gruppe rot einen täglichen Zuwachs von 769 g/Tag und
die Gruppe blau einen Zuwachs von 752 g/Tag; bei Durchgang IV lagen die täglichen
Zunahmen für die Gruppe rot bei 815 g/Tag und für die Gruppe blau bei 808 g/Tag.
Tab. 6: Durchschnittliche tägliche Gewichtszunahme (g/Tag) während der Mast
Durchgang I Durchgang II Durchgang III Durchgang IV gesamt
rot 843 835 769 815 824 ± 48,6
blau 827 827 752 825 816 ± 46,6
76 Ergebnisse
700
750
800
850
I II III IV gesamt
g / T
ag
Suvaxyn® M.hyo-Parasuis Suvaxyn® M.hyo
Abb. 6: Durchschnittliche tägliche Gewichtszunahme (g / Tag)
4.4 Futterverwertung
Die Futterverwertung in der Saugferkelphase wird nicht berücksichtigt.
Der Vergleich der Futterverwertung während der Aufzucht beider Impfgruppen lässt
keinen signifikanten Unterschied erkennen (p=0,57), beide Impfgruppen haben eine
Futterverwertung von 1,57.
Auch der Vergleich während der Mast zeigt keinen signifikanten Unterschied
zwischen den Gruppen rot und blau (p= 0,82). In der Mastperiode liegt die
Futterverwertung beider Gruppen bei 2,82 (rot 2,82 ± 0,08, blau 2,82 ± 0,10). In der
nachfolgenden Tabelle ist die Futterverwertung der Impfgruppen in den einzelnen
Durchgängen dargestellt. Auch der Vergleich der einzelnen Impfgruppen zeigt keinen
signifikanten Unterschied.
Tab. 7: Futterverwertung während der Mast
Durchgang I Durchgang II Durchgang III Durchgang IV gesamt
rot 2,79 2,83 2,94 2,8 2,82 ± 0,08
blau 2,81 2,84 2,89 2,7 2,82 ± 0,10
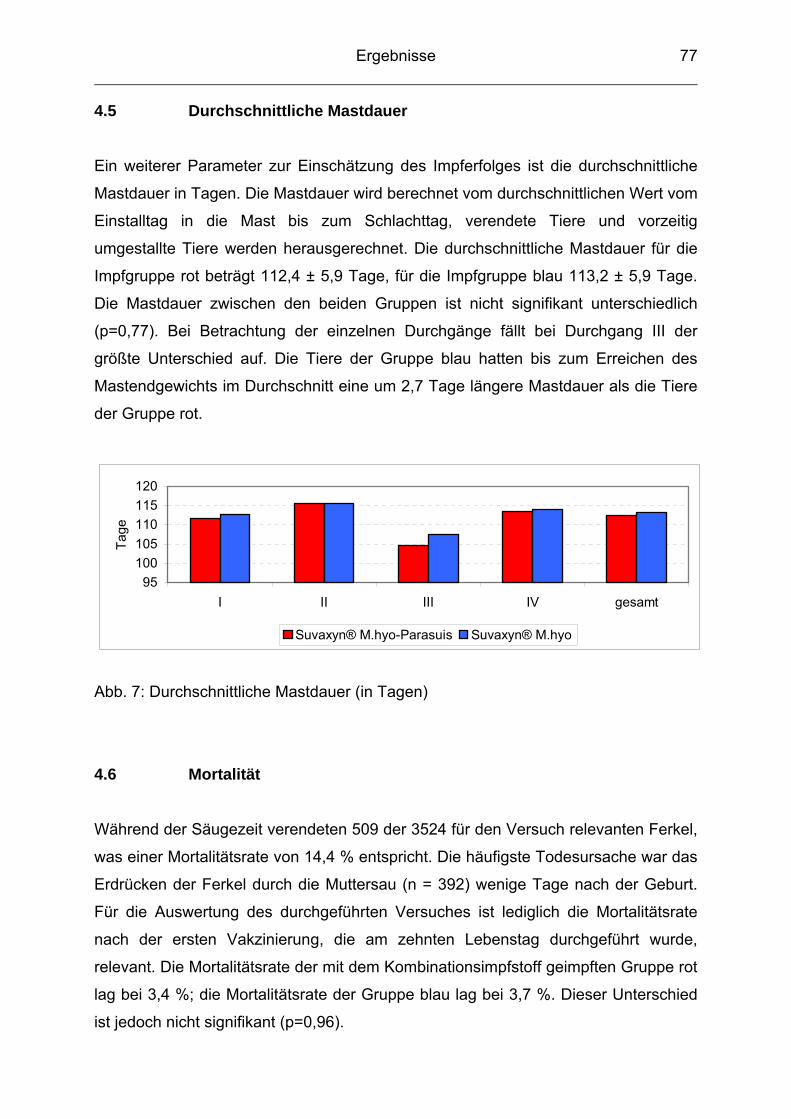
Ergebnisse 77
4.5 Durchschnittliche Mastdauer
Ein weiterer Parameter zur Einschätzung des Impferfolges ist die durchschnittliche
Mastdauer in Tagen. Die Mastdauer wird berechnet vom durchschnittlichen Wert vom
Einstalltag in die Mast bis zum Schlachttag, verendete Tiere und vorzeitig
umgestallte Tiere werden herausgerechnet. Die durchschnittliche Mastdauer für die
Impfgruppe rot beträgt 112,4 ± 5,9 Tage, für die Impfgruppe blau 113,2 ± 5,9 Tage.
Die Mastdauer zwischen den beiden Gruppen ist nicht signifikant unterschiedlich
(p=0,77). Bei Betrachtung der einzelnen Durchgänge fällt bei Durchgang III der
größte Unterschied auf. Die Tiere der Gruppe blau hatten bis zum Erreichen des
Mastendgewichts im Durchschnitt eine um 2,7 Tage längere Mastdauer als die Tiere
der Gruppe rot.
95100105110115120
I II III IV gesamt
Tage
Suvaxyn® M.hyo-Parasuis Suvaxyn® M.hyo
Abb. 7: Durchschnittliche Mastdauer (in Tagen)
4.6 Mortalität
Während der Säugezeit verendeten 509 der 3524 für den Versuch relevanten Ferkel,
was einer Mortalitätsrate von 14,4 % entspricht. Die häufigste Todesursache war das
Erdrücken der Ferkel durch die Muttersau (n = 392) wenige Tage nach der Geburt.
Für die Auswertung des durchgeführten Versuches ist lediglich die Mortalitätsrate
nach der ersten Vakzinierung, die am zehnten Lebenstag durchgeführt wurde,
relevant. Die Mortalitätsrate der mit dem Kombinationsimpfstoff geimpften Gruppe rot
lag bei 3,4 %; die Mortalitätsrate der Gruppe blau lag bei 3,7 %. Dieser Unterschied
ist jedoch nicht signifikant (p=0,96).

78 Ergebnisse
Die Mortalität in der Ferkelaufzucht lag für alle Tiere bei 1,1 %. Die Mortalitätsrate der
Impfgruppe rot (1,0 %) unterscheidet sich nicht signifikant von der Impfgruppe blau
(1,2 %) (p=0,55).
In der Gruppe rot verendeten insgesamt 14 Ferkel, in der Gruppe blau verendeten 16
Ferkel. Alle Tiere sind der tierärztlichen pathologisch-anatomischen Untersuchung
zugeführt worden. Die Anzahl der verendeten Ferkel in den einzelnen
Aufzuchtgruppen ist in Abbildung 9 dargestellt. Auffallend viele Tiere sind im II.
Durchgang verendet, dies waren acht Ferkel aus der Gruppe blau und drei Ferkel der
Gruppe rot. Der häufigste Befund aller untersuchten Ferkel war eine katarrhalische
Enteritis. Dieser Befund konnte bei fünf Ferkeln der Gruppe rot und bei drei Ferkeln
der Gruppe blau festgestellt werden. Fast ebenso häufig konnte der Befund Arthritis
gestellt werden. Je drei Ferkel der Gruppen rot und blau wiesen diesen Befund auf.
Bei jeweils einem dieser Ferkel konnte neben der Arthritis zusätzlich eine
Bronchopneumonie festgestellt werden, und jeweils ein Ferkel beider Gruppen zeigte
neben der Arthritis eine Perikarditis. Zusätzlich zeigte jeweils ein Ferkel beider
Gruppen eine Perikarditis, bei dem Ferkel der Gruppe rot konnte zusätzlich in der
Bakteriologie eine Salmonelleninfektion nachgewiesen werden; das Ferkel der
Gruppe blau zeigte zusätzlich zum Befund Perikarditis das Symptom des
Maulbeerherzens und eine Milzhyperplasie. Bei vier Ferkeln der Gruppe blau war in
der Sektion das Bild einer Polyserositis aufgefallen; bei einem dieser Ferkel wurde
zusätzlich eine Arthritis festgestellt. Bei den verendeten Ferkeln der Gruppe rot
wurde der Befund Polyserositis nicht gefunden. Bei drei der verendeten Ferkel der
Gruppe blau waren im Vorfeld Anzeichen von Kümmern aufgefallen, dies war bei
keinem Ferkel der Gruppe rot der Fall. Alle übrigen Sektionen ergaben
Einzelbefunde. In der Gruppe rot wurde eine Salmonelleninfektion, eine Infektion mit
St. hyicus, eine Infektion mit S. suis und ein Mastdarmvorfall diagnostiziert.
Einzelbefunde der Gruppe blau waren eine Torsio coli und eine Hernia abdominalis.
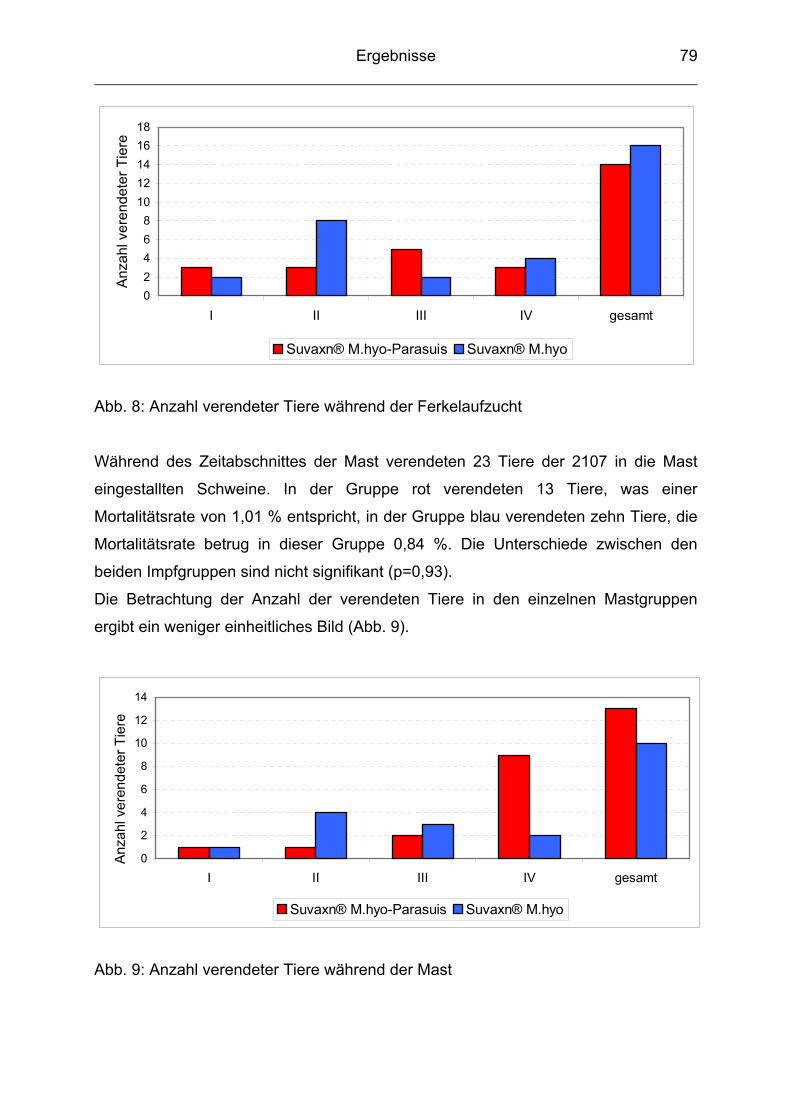
Ergebnisse 79
02468
1012141618
I II III IV gesamt
Anz
ahl v
eren
dete
r Tie
re
Suvaxn® M.hyo-Parasuis Suvaxn® M.hyo
Abb. 8: Anzahl verendeter Tiere während der Ferkelaufzucht
Während des Zeitabschnittes der Mast verendeten 23 Tiere der 2107 in die Mast
eingestallten Schweine. In der Gruppe rot verendeten 13 Tiere, was einer
Mortalitätsrate von 1,01 % entspricht, in der Gruppe blau verendeten zehn Tiere, die
Mortalitätsrate betrug in dieser Gruppe 0,84 %. Die Unterschiede zwischen den
beiden Impfgruppen sind nicht signifikant (p=0,93).
Die Betrachtung der Anzahl der verendeten Tiere in den einzelnen Mastgruppen
ergibt ein weniger einheitliches Bild (Abb. 9).
0
2
4
6
8
10
12
14
I II III IV gesamt
Anz
ahl v
eren
dete
r Tie
re
Suvaxn® M.hyo-Parasuis Suvaxn® M.hyo
Abb. 9: Anzahl verendeter Tiere während der Mast

80 Ergebnisse
Die Sektionen der Masttiere ergaben vorwiegend Einzeltierbefunde, wie Magenulcus,
Enteritis, Darminvagination und Infektion mit PCV-2. Bei Durchgang IV kam es
gehäuft zu Verendungen aufgrund von Kreislaufversagen gegen Ende der Mast,
dieses war bei fünf Tieren der Gruppe rot und bei zwei Tieren der Gruppe blau der
Fall. Im Durchgang II konnte bei drei Tieren der Gruppe blau eine
Bronchopneumonie festgestellt werden, dieser Befund trat bei der Gruppe rot nicht
auf. Ebenfalls eine Bronchopneumonie wurde bei einem Tier der Gruppe blau des
Durchgangs I diagnostiziert. Im III. Durchgang wurde bei drei Schweinen der
Sektionsbefund einer Perikarditis festgestellt; eines dieser Tiere gehörte der
Impfgruppe rot, die anderen beiden der Impfgruppe blau an.
4.7 Therapeutische Maßnahmen / Tierbehandlungsindex (TBI)
In allen Gruppen waren während der Versuchsdurchführung sowohl Einzeltier- als
auch Gruppen- bzw. Bestandsbehandlungen erforderlich. Üblicherweise wurden für
diese Behandlungen, je nach Lage des Falles, antibiotisch wirksame Arzneimittel
über das Futter oder das Trinkwasser verabreicht. Einzeltierbehandlungen sind per
Injektion verabreicht worden. Zur Quantifizierung der Behandlungen wurde der
Tierbehandlungsindex (TBI) mit zwei verschiedenen Berechnungsgrundlagen
verwendet. Der TBIges. beinhaltet alle antibiotischen Behandlungen, in den TBIeinzel
gehen nur Behandlungen einzelner Tiere oder einzelner Gruppen ein.
Die Saugferkel bekamen zur Prophylaxe gegen Streptokokkeninfektionen am ersten
Lebenstag Amoxicillin und am fünften Lebenstag Langzeitpenicillin intramuskulär
verabreicht. Zusätzlich zu diesen Maßnahmen, die generell an allen Ferkeln
durchgeführt wurden, ist der TBIeinzel erfasst worden. Der TBI für die Ferkel der
Gruppe rot liegt bei 0,17 und für die Gruppe blau bei 0,19. Dieser Unterschied lässt
sich statistisch nicht signifikant absichern (p=0,74).
Den Absetzferkeln aller vier Aufzuchtgruppen wurde zur vorbeugenden Behandlung
möglicher Coliinfektionen über sieben Tage Colistin mit dem Trinkwasser verabreicht.
Zur Vorbeugung von Streptokokkeninfektionen wurde zusätzlich über den gleichen
Zeitraum Amoxicillin verabreicht. Die Behandlungen im Ferkelaufzuchtstall sind
nachfolgend für die einzelnen Aufzuchtgruppen dargestellt. Für die einzelnen
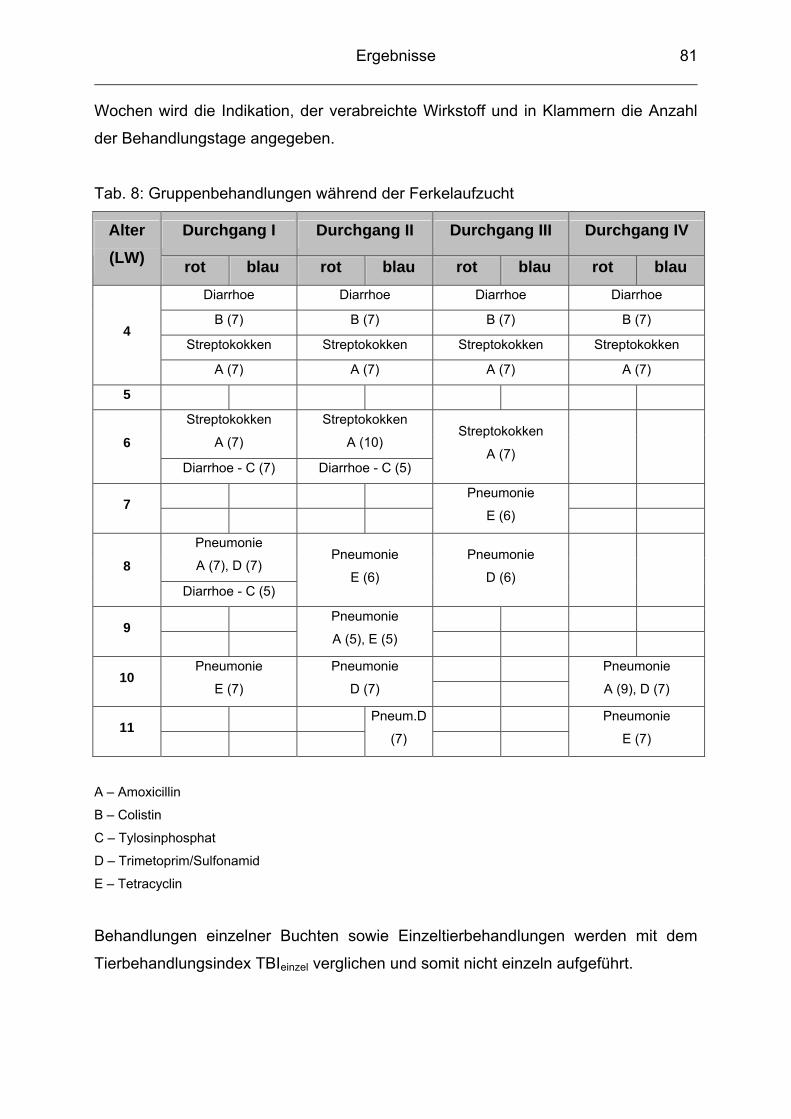
Ergebnisse 81
Wochen wird die Indikation, der verabreichte Wirkstoff und in Klammern die Anzahl
der Behandlungstage angegeben.
Tab. 8: Gruppenbehandlungen während der Ferkelaufzucht
Durchgang I Durchgang II Durchgang III Durchgang IV Alter (LW) rot blau rot blau rot blau rot blau
Diarrhoe Diarrhoe Diarrhoe Diarrhoe
B (7) B (7) B (7) B (7)
Streptokokken Streptokokken Streptokokken Streptokokken 4
A (7) A (7) A (7) A (7)
5
Streptokokken
A (7)
Streptokokken
A (10) 6
Diarrhoe - C (7) Diarrhoe - C (5)
Streptokokken
A (7)
7
Pneumonie
E (6)
Pneumonie
A (7), D (7) 8
Diarrhoe - C (5)
Pneumonie
E (6)
Pneumonie
D (6)
9
Pneumonie
A (5), E (5)
10
Pneumonie
E (7)
Pneumonie
D (7)
Pneumonie
A (9), D (7)
11
Pneum.D
(7)
Pneumonie
E (7)
A – Amoxicillin
B – Colistin
C – Tylosinphosphat
D – Trimetoprim/Sulfonamid
E – Tetracyclin
Behandlungen einzelner Buchten sowie Einzeltierbehandlungen werden mit dem
Tierbehandlungsindex TBIeinzel verglichen und somit nicht einzeln aufgeführt.
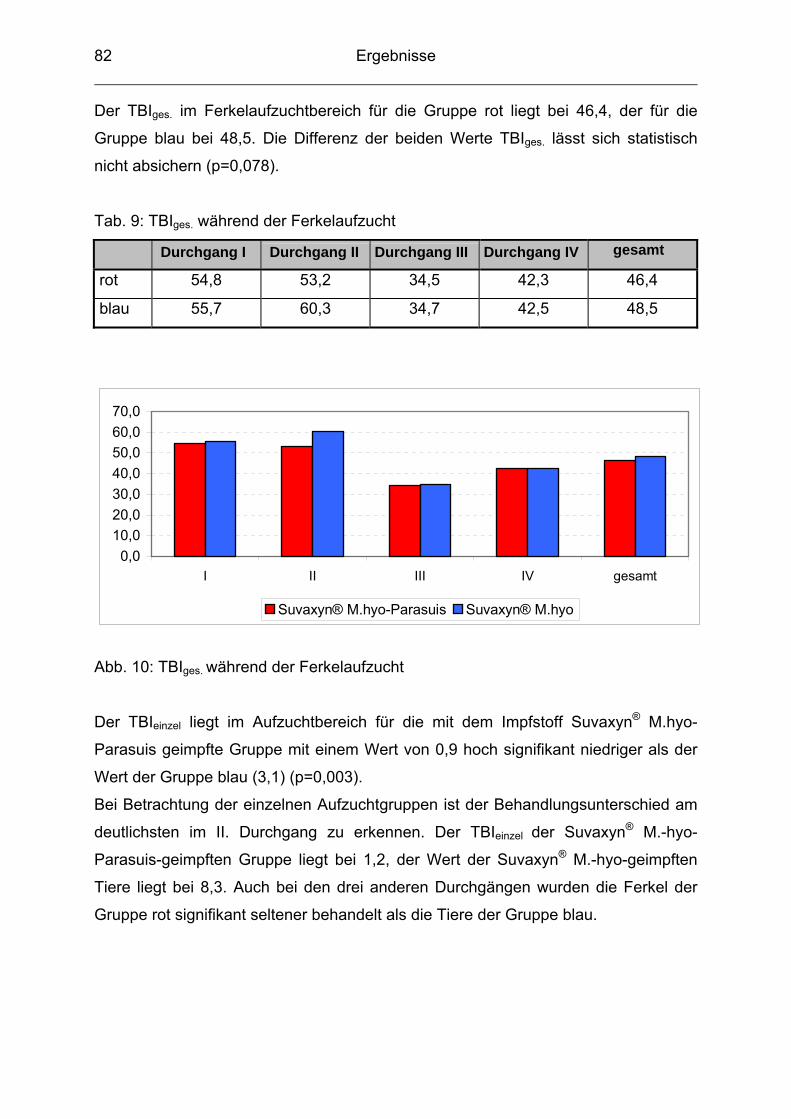
82 Ergebnisse
Der TBIges. im Ferkelaufzuchtbereich für die Gruppe rot liegt bei 46,4, der für die
Gruppe blau bei 48,5. Die Differenz der beiden Werte TBIges. lässt sich statistisch
nicht absichern (p=0,078).
Tab. 9: TBIges. während der Ferkelaufzucht
Durchgang I Durchgang II Durchgang III Durchgang IV gesamt
rot 54,8 53,2 34,5 42,3 46,4
blau 55,7 60,3 34,7 42,5 48,5
0,010,020,030,040,050,060,070,0
I II III IV gesamt
Suvaxyn® M.hyo-Parasuis Suvaxyn® M.hyo
Abb. 10: TBIges. während der Ferkelaufzucht
Der TBIeinzel liegt im Aufzuchtbereich für die mit dem Impfstoff Suvaxyn® M.hyo-
Parasuis geimpfte Gruppe mit einem Wert von 0,9 hoch signifikant niedriger als der
Wert der Gruppe blau (3,1) (p=0,003).
Bei Betrachtung der einzelnen Aufzuchtgruppen ist der Behandlungsunterschied am
deutlichsten im II. Durchgang zu erkennen. Der TBIeinzel der Suvaxyn® M.-hyo-
Parasuis-geimpften Gruppe liegt bei 1,2, der Wert der Suvaxyn® M.-hyo-geimpften
Tiere liegt bei 8,3. Auch bei den drei anderen Durchgängen wurden die Ferkel der
Gruppe rot signifikant seltener behandelt als die Tiere der Gruppe blau.
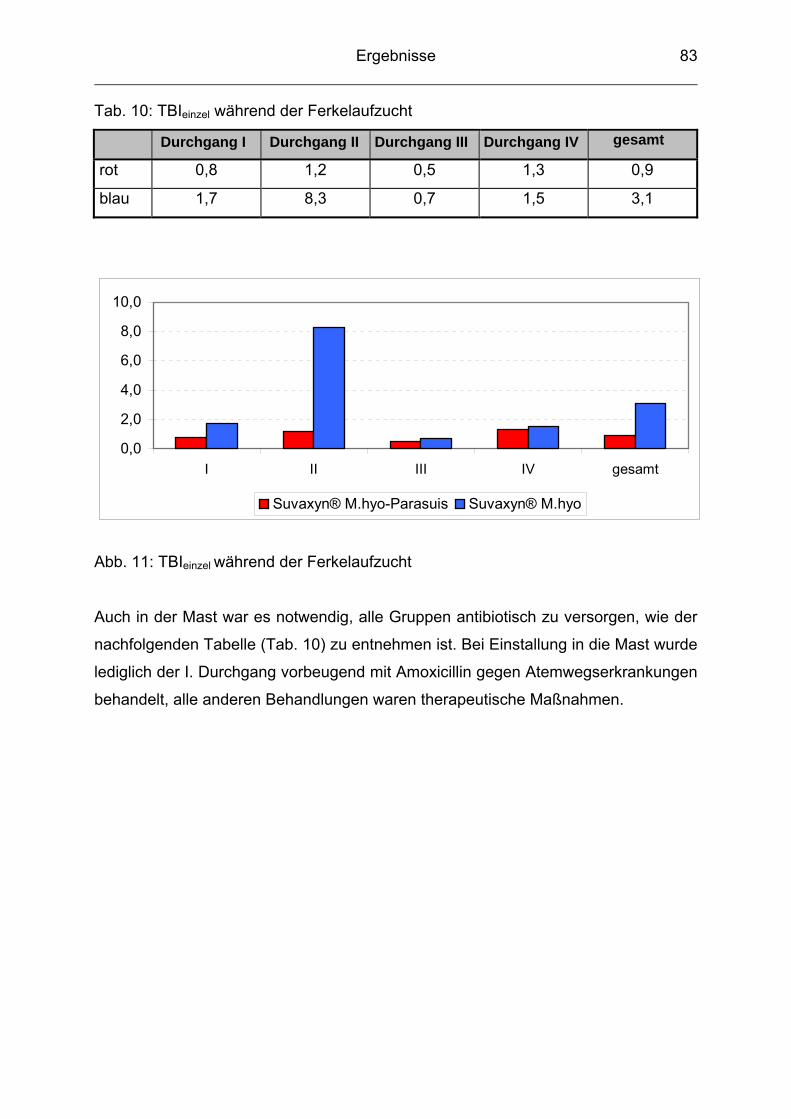
Ergebnisse 83
Tab. 10: TBIeinzel während der Ferkelaufzucht
Durchgang I Durchgang II Durchgang III Durchgang IV gesamt
rot 0,8 1,2 0,5 1,3 0,9
blau 1,7 8,3 0,7 1,5 3,1
0,0
2,0
4,0
6,0
8,0
10,0
I II III IV gesamt
Suvaxyn® M.hyo-Parasuis Suvaxyn® M.hyo
Abb. 11: TBIeinzel während der Ferkelaufzucht
Auch in der Mast war es notwendig, alle Gruppen antibiotisch zu versorgen, wie der
nachfolgenden Tabelle (Tab. 10) zu entnehmen ist. Bei Einstallung in die Mast wurde
lediglich der I. Durchgang vorbeugend mit Amoxicillin gegen Atemwegserkrankungen
behandelt, alle anderen Behandlungen waren therapeutische Maßnahmen.
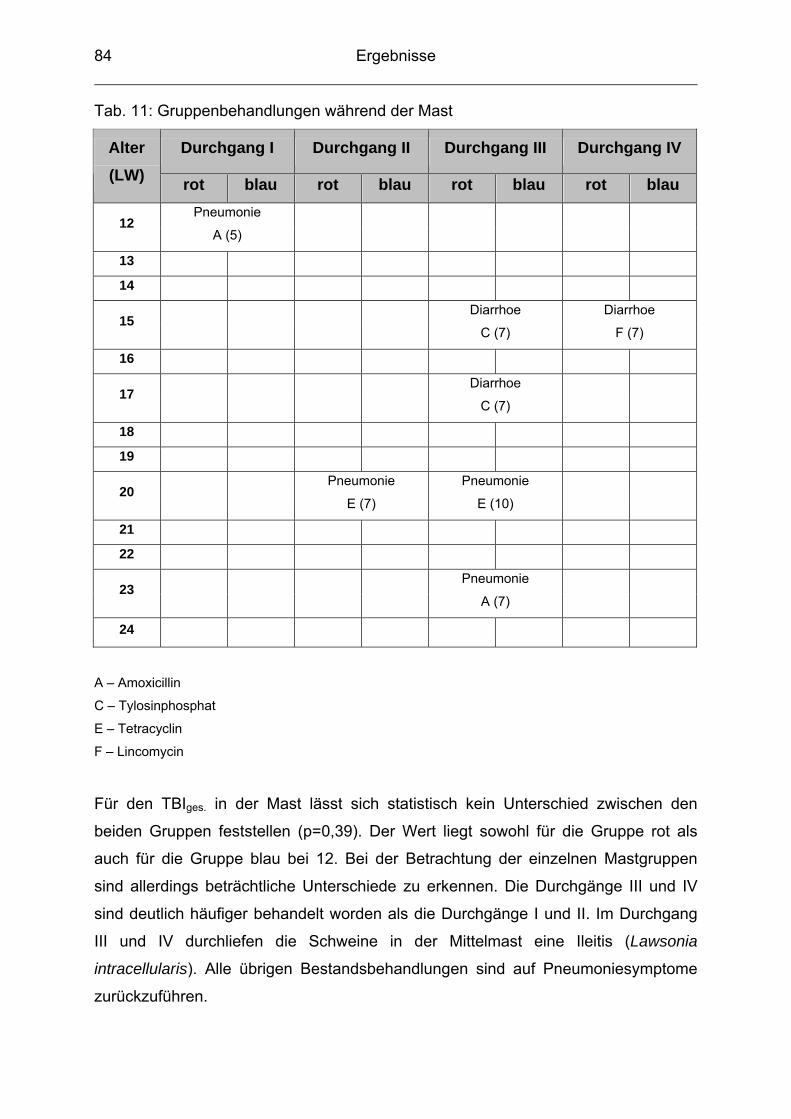
84 Ergebnisse
Tab. 11: Gruppenbehandlungen während der Mast
Durchgang I Durchgang II Durchgang III Durchgang IV Alter (LW) rot blau rot blau rot blau rot blau
12
Pneumonie
A (5)
13
14
15
Diarrhoe
C (7)
Diarrhoe
F (7)
16
17
Diarrhoe
C (7)
18
19
20
Pneumonie
E (7)
Pneumonie
E (10)
21
22
23
Pneumonie
A (7)
24
A – Amoxicillin
C – Tylosinphosphat
E – Tetracyclin
F – Lincomycin
Für den TBIges. in der Mast lässt sich statistisch kein Unterschied zwischen den
beiden Gruppen feststellen (p=0,39). Der Wert liegt sowohl für die Gruppe rot als
auch für die Gruppe blau bei 12. Bei der Betrachtung der einzelnen Mastgruppen
sind allerdings beträchtliche Unterschiede zu erkennen. Die Durchgänge III und IV
sind deutlich häufiger behandelt worden als die Durchgänge I und II. Im Durchgang
III und IV durchliefen die Schweine in der Mittelmast eine Ileitis (Lawsonia
intracellularis). Alle übrigen Bestandsbehandlungen sind auf Pneumoniesymptome
zurückzuführen.
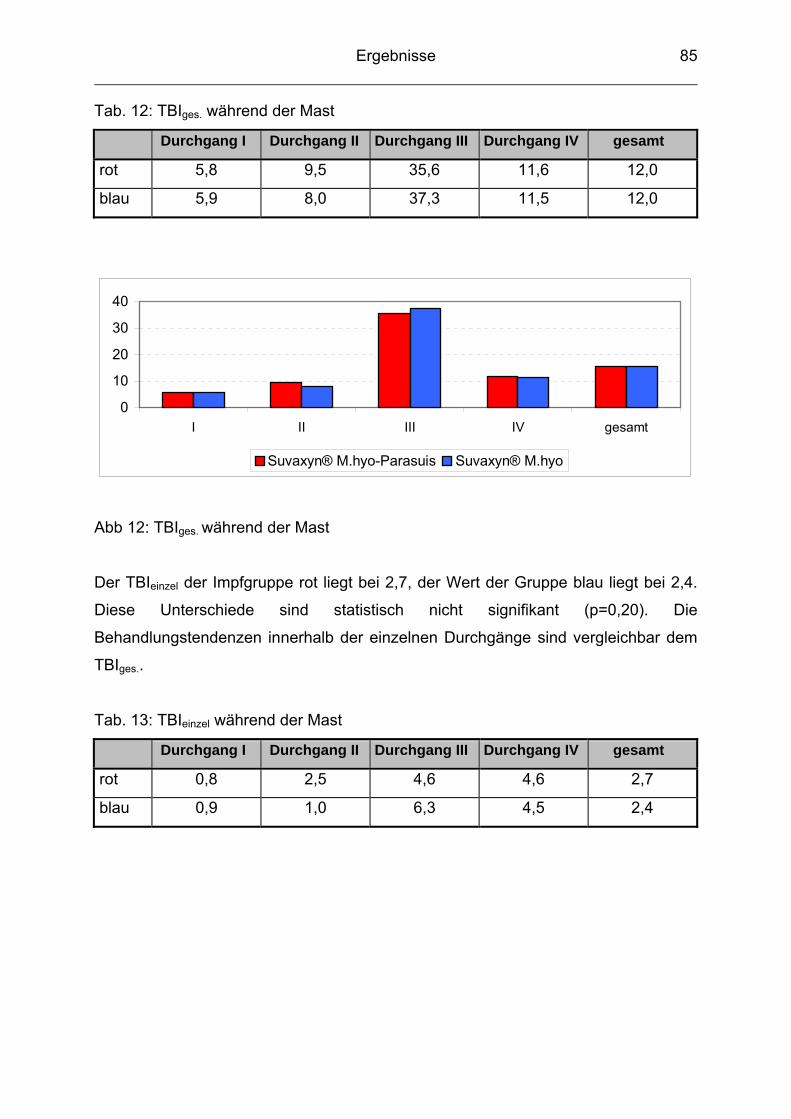
Ergebnisse 85
Tab. 12: TBIges. während der Mast
Durchgang I Durchgang II Durchgang III Durchgang IV gesamt
rot 5,8 9,5 35,6 11,6 12,0
blau 5,9 8,0 37,3 11,5 12,0
0
10
20
30
40
I II III IV gesamt
Suvaxyn® M.hyo-Parasuis Suvaxyn® M.hyo
Abb 12: TBIges. während der Mast
Der TBIeinzel der Impfgruppe rot liegt bei 2,7, der Wert der Gruppe blau liegt bei 2,4.
Diese Unterschiede sind statistisch nicht signifikant (p=0,20). Die
Behandlungstendenzen innerhalb der einzelnen Durchgänge sind vergleichbar dem
TBIges..
Tab. 13: TBIeinzel während der Mast
Durchgang I Durchgang II Durchgang III Durchgang IV gesamt
rot 0,8 2,5 4,6 4,6 2,7
blau 0,9 1,0 6,3 4,5 2,4
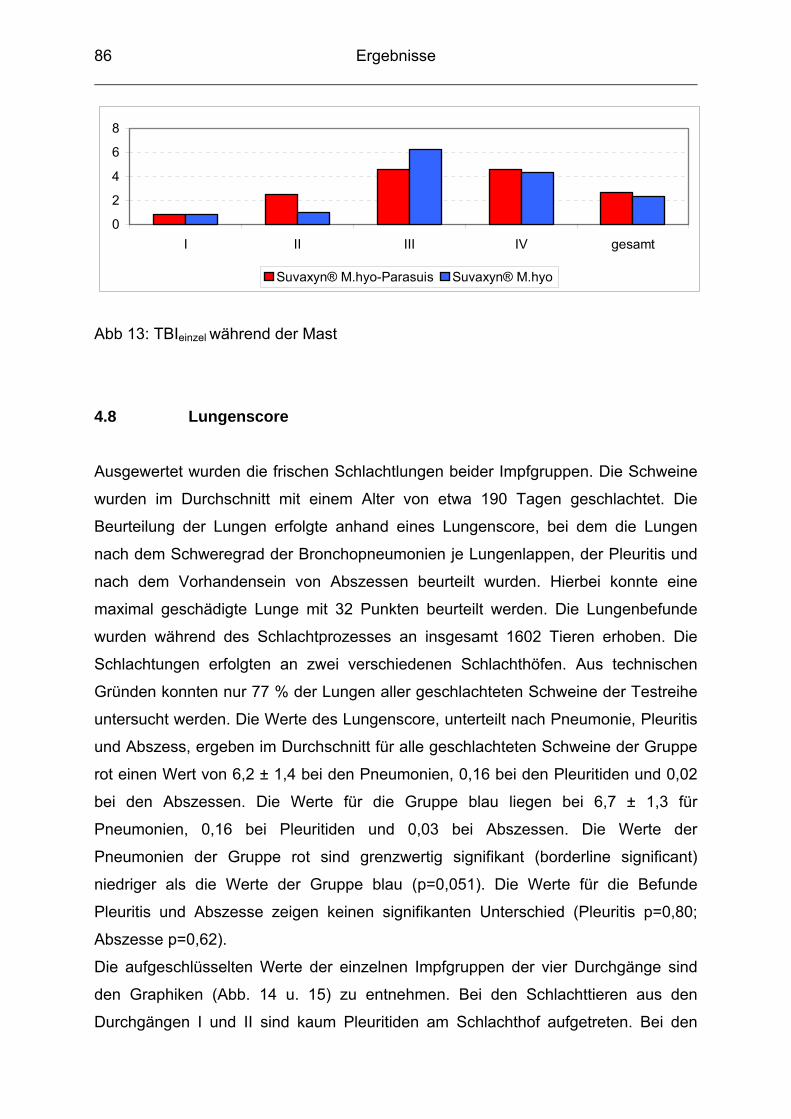
86 Ergebnisse
0
2
4
6
8
I II III IV gesamt
Suvaxyn® M.hyo-Parasuis Suvaxyn® M.hyo
Abb 13: TBIeinzel während der Mast
4.8 Lungenscore
Ausgewertet wurden die frischen Schlachtlungen beider Impfgruppen. Die Schweine
wurden im Durchschnitt mit einem Alter von etwa 190 Tagen geschlachtet. Die
Beurteilung der Lungen erfolgte anhand eines Lungenscore, bei dem die Lungen
nach dem Schweregrad der Bronchopneumonien je Lungenlappen, der Pleuritis und
nach dem Vorhandensein von Abszessen beurteilt wurden. Hierbei konnte eine
maximal geschädigte Lunge mit 32 Punkten beurteilt werden. Die Lungenbefunde
wurden während des Schlachtprozesses an insgesamt 1602 Tieren erhoben. Die
Schlachtungen erfolgten an zwei verschiedenen Schlachthöfen. Aus technischen
Gründen konnten nur 77 % der Lungen aller geschlachteten Schweine der Testreihe
untersucht werden. Die Werte des Lungenscore, unterteilt nach Pneumonie, Pleuritis
und Abszess, ergeben im Durchschnitt für alle geschlachteten Schweine der Gruppe
rot einen Wert von 6,2 ± 1,4 bei den Pneumonien, 0,16 bei den Pleuritiden und 0,02
bei den Abszessen. Die Werte für die Gruppe blau liegen bei 6,7 ± 1,3 für
Pneumonien, 0,16 bei Pleuritiden und 0,03 bei Abszessen. Die Werte der
Pneumonien der Gruppe rot sind grenzwertig signifikant (borderline significant)
niedriger als die Werte der Gruppe blau (p=0,051). Die Werte für die Befunde
Pleuritis und Abszesse zeigen keinen signifikanten Unterschied (Pleuritis p=0,80;
Abszesse p=0,62).
Die aufgeschlüsselten Werte der einzelnen Impfgruppen der vier Durchgänge sind
den Graphiken (Abb. 14 u. 15) zu entnehmen. Bei den Schlachttieren aus den
Durchgängen I und II sind kaum Pleuritiden am Schlachthof aufgetreten. Bei den
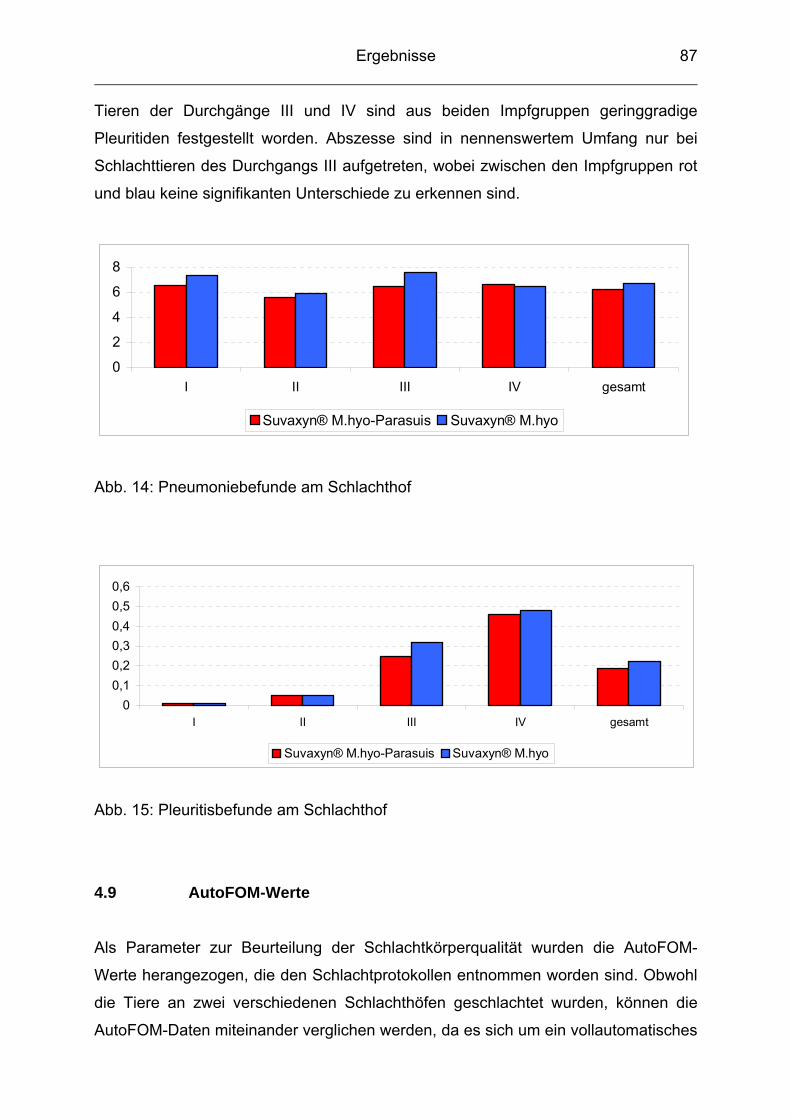
Ergebnisse 87
Tieren der Durchgänge III und IV sind aus beiden Impfgruppen geringgradige
Pleuritiden festgestellt worden. Abszesse sind in nennenswertem Umfang nur bei
Schlachttieren des Durchgangs III aufgetreten, wobei zwischen den Impfgruppen rot
und blau keine signifikanten Unterschiede zu erkennen sind.
0
2
4
6
8
I II III IV gesamt
Suvaxyn® M.hyo-Parasuis Suvaxyn® M.hyo
Abb. 14: Pneumoniebefunde am Schlachthof
00,10,20,30,40,50,6
I II III IV gesamt
Suvaxyn® M.hyo-Parasuis Suvaxyn® M.hyo
Abb. 15: Pleuritisbefunde am Schlachthof
4.9 AutoFOM-Werte
Als Parameter zur Beurteilung der Schlachtkörperqualität wurden die AutoFOM-
Werte herangezogen, die den Schlachtprotokollen entnommen worden sind. Obwohl
die Tiere an zwei verschiedenen Schlachthöfen geschlachtet wurden, können die
AutoFOM-Daten miteinander verglichen werden, da es sich um ein vollautomatisches
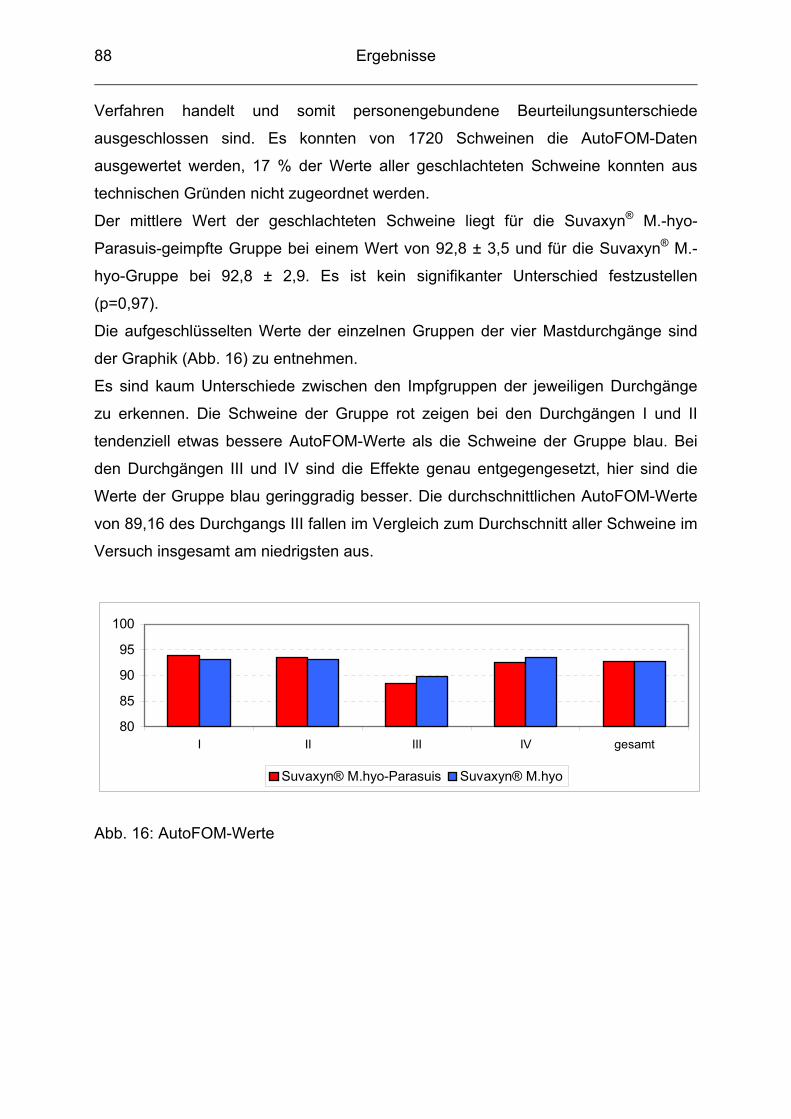
88 Ergebnisse
Verfahren handelt und somit personengebundene Beurteilungsunterschiede
ausgeschlossen sind. Es konnten von 1720 Schweinen die AutoFOM-Daten
ausgewertet werden, 17 % der Werte aller geschlachteten Schweine konnten aus
technischen Gründen nicht zugeordnet werden.
Der mittlere Wert der geschlachteten Schweine liegt für die Suvaxyn® M.-hyo-
Parasuis-geimpfte Gruppe bei einem Wert von 92,8 ± 3,5 und für die Suvaxyn® M.-
hyo-Gruppe bei 92,8 ± 2,9. Es ist kein signifikanter Unterschied festzustellen
(p=0,97).
Die aufgeschlüsselten Werte der einzelnen Gruppen der vier Mastdurchgänge sind
der Graphik (Abb. 16) zu entnehmen.
Es sind kaum Unterschiede zwischen den Impfgruppen der jeweiligen Durchgänge
zu erkennen. Die Schweine der Gruppe rot zeigen bei den Durchgängen I und II
tendenziell etwas bessere AutoFOM-Werte als die Schweine der Gruppe blau. Bei
den Durchgängen III und IV sind die Effekte genau entgegengesetzt, hier sind die
Werte der Gruppe blau geringgradig besser. Die durchschnittlichen AutoFOM-Werte
von 89,16 des Durchgangs III fallen im Vergleich zum Durchschnitt aller Schweine im
Versuch insgesamt am niedrigsten aus.
80
85
90
95
100
I II III IV gesamt
Suvaxyn® M.hyo-Parasuis Suvaxyn® M.hyo
Abb. 16: AutoFOM-Werte

Diskussion 89
5 Diskussion
5.1 Allgemeines
Ziel der vorliegenden Arbeit war, die Effektivität eines Kombinationsimpfstoffes gegen
H. parasuis und M. hyopneumoniae (Suvaxyn® M.hyo-Parasuis) im Vergleich zu
einem Monoimpfstoff gegen M. hyopneumoniae (Suvaxyn® M.hyo) zu beurteilen.
Beurteilungskriterien des Versuches waren verschiedene Daten zur Mastleistung,
serologische Verlaufsuntersuchungen sowie Lungenveränderungen zum Zeitpunkt
der Schlachtung. Durchgeführt wurden die Untersuchungen als Feldstudie in einem
Sauenbetrieb mit angeschlossener Aufzucht und anschließender Mast, so dass die
Versuchstiere von der Geburt bis zur Schlachtung verfolgt werden konnten.
5.2 Serologie
ELISA-Werte H. parasuis Die OD-Werte beider Impfgruppen lagen vor der ersten Vakzinierung, also am 10.
Lebenstag, für die Serotypen 4 und 5 von H. parasuis bei Werten zwischen 0,752
und 0,756. Bei Vergleichsuntersuchungen in etwa zehn europäischen Betrieben mit
demselben ELISA-Testverfahren wurden Normalwerte für Ferkel in einem Alter von
einer Woche von 0,6 bis 0,8 festgestellt (Angaben des Testherstellers Fort Dodge).
Man kann demnach davon ausgehen, dass bei dem nachgewiesenen Niveau
Antikörper aus dem Kolostrum auf die Ferkel übertragen worden sind. Die Höhe der
OD-Werte lässt aber eher auf eine niedrige maternale Immunität schließen.
Demnach können Interferenzen zwischen maternalen Antikörpern und Antikörpern
durch die Impfung nahezu ausgeschlossen werden. Für diese Tatsache spricht auch,
dass bei den am Versuch beteiligten Sauen in der Vergangenheit keine H.-parasuis-
Mutterschutzvakzinierung durchgeführt worden ist. Ebenso ist das Vorkommen
klinischer Erscheinungen einer H.-parasuis-Infektion im Sauenbestand nicht bekannt,
und in der Vergangenheit ist der Erreger bei pathologisch-anatomischen und
bakteriologischen Untersuchungen von Sauen nicht gefunden worden.
Auffallend ist das Absinken der OD-Werte zum Zeitpunkt der zweiten Blutentnahme
(5. LW). Die Werte für H. parasuis Serotyp 5 fallen bei beiden Impfgruppen zwei

90 Diskussion
Wochen nach der letzten Vakzinierung leicht ab. Die H.-parasuis-Serotyp-4-Werte
der Suvaxyn® M.-hyo-Gruppe fallen leicht unter den Wert vom 10. Lebenstag, die
Werte der Suvaxyn® M.-hyo-Parasuis-Gruppe steigen minimal an. Beim Vergleich
beider Werte lassen sich keine signifikanten Unterschiede erkennen. Die Impfungen
sind am 10. Lebenstag und in der dritten Lebenswoche durchgeführt worden. Bei der
Suvaxyn® M.-hyo-Parasuis-Gruppe wäre zum Zeitpunkt der zweiten Blutentnahme,
die zwei Wochen nach der zweiten Impfung durchgeführt wurde, ein Anstieg der OD-
Werte der Serotypen 4 und 5 des ELISAs als Impfreaktion zu erwarten gewesen.
Im weiteren Verlauf der Blutprobenentnahmen steigen die OD-Werte von H. parasuis
Serotyp 4 beider Impfgruppen bis zur 16. LW kontinuierlich an. Dieser Anstieg der
Antikörperkonzentration spricht für eine Feldinfektion mit H. parasuis, da auch die
Ferkel ohne die H.-parasuis-Komponente im Impfstoff erhöhte Antikörperwerte
aufzeigen. Ebenso kontinuierlich steigen die Antikörperwerte gemessen als optische
Dichte des Serotyp 5 der Suvaxyn® M.-hyo-geimpften Tiere an. Die Antikörper-
konzentrationen der Suvaxyn® M.-hyo-Parasuis-geimpften Gruppe steigen dagegen
in der 10. LW steil an und bleiben bis zur 24. LW auf diesem hohen Niveau. Aus
diesen Tatsachen ist wiederum der Schluss zu ziehen, dass eine Feldinfektion
stattgefunden hat, da auch bei Serotyp 5 die Werte der H.-parasuis-ungeimpften
Tiere ansteigen. Der steile Anstieg der Antikörperkonzentrationen der H.-parasuis-
geimpften Tiere spricht für einen Boostereffekt durch die Impfung. Man kann
demnach davon ausgehen, dass gegen H. parasuis geimpfte Tiere mit einer höheren
Antikörperbildung auf eine Feldinfektion reagieren und so eine Schutzwirkung des
Impfstoffes Suvaxyn® M.hyo-Parasuis als gegeben angesehen werden kann.
Eine exakte Eingrenzung des Infektionszeitpunktes durch die
Untersuchungsergebnisse ist nicht möglich. Anhand der Mortalitätsdaten, der
eingesetzten Medikamente und der Antikörperwerte des Durchgangs II im Vergleich
zu den anderen drei Durchgängen kann jedoch vermutet werden, dass die
Feldinfektion etwa zwischen der sechsten und achten Lebenswoche der Ferkel
stattgefunden hat.
Auch in anderen Studien werden Saugferkel und Aufzuchttiere als die Altersgruppe
beschrieben, die am anfälligsten für Keime wie H. parasuis, B. bronchiseptica und S.
suis sind (DELBECK 1995; NIEMANN 1999). Bei den Untersuchungen zum
Vorkommen bakterieller Erreger in pathologisch-anatomisch veränderten Lungen von

Diskussion 91
VON ALTROCK (1998) ging ebenso die Nachweishäufigkeit von H. parasuis nach
einem Anstieg bei den Absetzferkeln mit zunehmendem Alter zurück.
Ausgehend vom Anstieg der OD-Werte beider Serotypen 4 und 5 kann nicht
rückgeschlossen werden, um welchen Serotypen es sich bei der Feldinfektion
handelt, da durch die Vakzine Suvaxyn® M.hyo-Parasuis Kreuzimmunitäten zwischen
den Serotypen 4 und 5 aufgebaut werden, also auch die Werte beider Serotypen
anstiegen. Ebenso wird eine Kreuzreaktion zu anderen Isolaten von H. parasuis
durch den Impfstoff vermutet.
RAPP-GABRIELSON et al. (1996) haben mit dem gleichen Impfstoff wie im
vorliegenden Versuch im Vergleich mit zwei anderen kommerziellen Vakzinen
gezeigt, dass Kreuzimmunitäten zwischen den Serotypen 4, 5, 12 und 13 ausgebildet
werden. Lediglich gegen die Serotypen 2 und 14 wurde kein Schutz aufgebaut. Beim
Vergleich der drei verwendeten Vakzinen konnten die Autoren im Belastungsversuch
zeigen, dass nur der auch in dieser Studie verwendete Impfstoff eine Kreuzprotektion
gegen Serotyp 5 ausbildet. Ebenso konnten sie bei einem Belastungsversuch nach
sechs Monaten zeigen, dass durch den Impfstoff auch bei sehr früher Impfung ein
lebenslanger Schutz für Mastschweine gegen die Serovare 4 und 5 aufgebaut wird.
ELISA-Werte M.hyopneumoniae
Der steile Anstieg der Antikörper in der fünften LW kann als Immunantwort auf die
Vakzination gewertet werden. Aus dem darauf folgenden Absinken der
Antikörperwerte beider Impfgruppen kann gefolgert werden, dass entweder keine
oder nur in geringem Maße eine Feldinfektion mit M. hyopneumoniae stattgefunden
hat. Die Immunität aufgrund der Vakzinierung gegen M. hyopneumoniae scheint im
Laufe der Mast bzw. bis zum Mastende abzufallen. Laut Herstellerangaben gewährt
der Impfstoff Suvaxyn® M.hyo-Parasuis einen Schutz gegen M. hyopneumoniae von
fünf Monaten.
Über die Immunogenität einzelner Komponenten eines Kombinationsimpfstoffes wird
in der Literatur kontrovers diskutiert. OLBERTZ (1990) untersuchte die
immunisierende Wirkung von mono- und polyvalenten Impfstoffen bei kombinierter
Anwendung und kam zu dem Ergebnis, dass die Mehrfachimpfung, d.h. die
simultane Anwendung von bis zu fünf Impfstoffkomponenten einen positiven Einfluss
auf die Antikörperentwicklung hat. Allerdings sollten die einzelnen Komponenten an
verschiedenen Körperstellen injiziert werden. In einer anderen Studie wurde die

92 Diskussion
Antikörperentwicklung polyvalenter Impfstoffe bei simultaner, aber lokal getrennter
Kombinationsimpfung mit einer Kombinationsimpfung mittels Mischinjektion
verglichen (KOHL 1995). Dabei wurden bei Mischinjektionen niedrigere Antikörper-
titer als bei den Einzelinjektionen gemessen. Im Gegensatz dazu konnten
RITZMANN et al. (1999) in einer vergleichenden Untersuchung über den Einsatz
eines Parvovirose-Rotlauf-Kombinationsimpfstoffes und den entsprechenden
Monovakzinen keinen signifikanten Unterschied in der Antikörperentwicklung
nachweisen.
In der vorliegenden Untersuchung weisen die Antikörperbildungen der Suvaxyn® M.-
hyo- und der Suvaxyn® M.-hyo-Parasuis-Impfgruppe in der 5. LW keinen
signifikanten Unterschiede auf. Dies deutet darauf hin, dass das Vorhandensein von
zwei Komponenten in dem Kombinationsimpfstoff Suvaxyn® M.hyo-Parasuis nicht
von Nachteil für die Ausbildung von Antikörpern gegen M. hyopneumoniae ist. Die
Immunogenität der M.-hyopneumoniae-Komponente des Kombinationsimpfstoffes
kann als genauso hoch wie bei dem monovalenten Impfstoff angesehen werden.
5.3 Mastleistung
Im Rahmen des Feldversuches wurde die Effektivität der beiden Impfstoffe Suvaxyn®
M.hyo-Parasuis und Suvaxyn® M.hyo untersucht und verglichen. Als Maß der
Effektivität wurden die Mastleistungen der Tiere durch die Parameter tägliche
Gewichtszunahmen, Futterverwertung und Mastdauer berücksichtigt.
Durchschnittliche tägliche Gewichtszunahme
Unter Annahme eines einheitlichen Geburtsgewichtes von 1,4 kg wurde die mittlere
tägliche Gewichtszunahme separat für die Säugezeit, die Aufzucht und die Mast
kalkuliert.
Bei den Saugferkeln konnte kein signifikanter Unterschied zwischen den beiden
Behandlungsgruppen rot und blau festgestellt werden; die täglichen Zunahmen lagen
bei 227 g/Tag bzw. bei 228 g/Tag. Da die erste Impfung im Alter von zehn Tagen und
die zweite Impfung in der dritten Lebenswoche stattgefunden hat, ist in der
Saugferkelphase mit keinem Effekt der Impfung zu rechnen.

Diskussion 93
Die Ferkelgruppen wiesen bei Einstallung in den Ferkelaufzuchtstall keine
signifikanten Gewichtsunterschiede auf; die Gruppe rot hatte ein durchschnittliches
Gewicht von 5,93 kg, die Gruppe blau von 5,95 kg. Während der Aufzucht sind
zwischen den beiden Behandlungsgruppen (rot 353 g/Tag und blau 359 g/Tag) keine
signifikanten Unterschiede der täglichen Zunahmen zu erkennen.
Während der Mastperiode sind im Mittel 820 g tägliche Zunahmen erreicht worden. In
Niedersachsen werden mittlere tägliche Gewichtszunahmen von 710 g ermittelt,
wobei die besten 25 % der Betriebe 783 g je Tag erreichen und die schlechtesten 25
% der Betriebe 637 g je Tag verzeichnen können (ANONYM 2006). Im vorliegenden
Versuch wurden in beiden Impfgruppen deutlich höhere tägliche Zunahmen
gemessen. Tiere der Gruppe rot erreichten tägliche Zunahmen von 824 g, Tiere der
Gruppe blau erreichten Zunahmen von 816 g.
Futterverwertung
Hinsichtlich der Futterverwertung sind während der Aufzucht- und der Mastperiode
keine signifikanten Unterschiede zwischen den beiden Gruppen zu erkennen. Die
Futterverwertung in der Aufzuchtphase lag für beide Impfgruppen bei 1,57.
In der Praxis werden in der Mastperiode im Mittel 2,98 kg Futter für 1 kg
Körpermassezuwachs (KMZ) benötigt (die 25 % besten Betriebe erreichen dabei
einen Futteraufwand von 2,90 kg/kg, die schlechtesten 25 % der Mastbetriebe 3,11
kg/kg KMZ) (ANONYM 2006).
Der Futteraufwand bzw. die Futterverwertung in diesem Versuch betrug im Mittel bei
beiden Impfgruppen 2,82 und lag somit über den aus der Praxis mitgeteilten Daten.
Im Maststall III zeichnet sich der größte Unterschied zwischen den beiden Gruppen
ab, die Gruppe rot hat eine um 0,05 kg bessere Verwertung des Futters als Gruppe
blau.
Durchschnittliche Mastdauer Die durchschnittliche Mastdauer war in der Suvaxyn® M.-hyo-Parasuis-geimpften
Gruppe um 0,8 Tage kürzer als in der Vergleichsgruppe; die durchschnittliche
Mastdauer der Gesamtgruppe rot lag bei 112,4 Tagen, die der Gruppe blau bei 113,2
Tagen. Im Mittel erreichen Betriebe in Niedersachsen eine durchschnittliche
Mastdauer von 119 Tagen, wobei die besten 25 % der Betriebe eine Mastdauer von
114 Tagen und die schlechtesten 25 % eine durchschnittliche Mastdauer von 129

94 Diskussion
Tagen erreichen (ANONYM 2006). Im vorliegenden Versuch liegt auch die
durchschnittliche Mastdauer unter den Vergleichswerten.
Die größten Unterschiede sind im Maststall III zu finden, dort war die Mastdauer der
Gruppe rot um 2,7 Tage kürzer als in der Gruppe blau.
Die Praxis ist an guten Mastleistungen, gemessen an hohen Tageszunahmen,
interessiert, um die Mastdauer so kurz wie möglich zu halten und damit eine höhere
Wirtschaftlichkeit, eine effektivere Futterverwertung und insgesamt eine bessere
Rentabilität zu erwirtschaften.
Verglichen mit sämtlichen Betrieben in Niedersachsen, die an den
Betriebszweigauswertungen der jeweiligen Beratungsringe teilnehmen, sind die
Mastleistungen der Schweine in diesem Versuch bezüglich der täglichen
Gewichtszunahme, der Futterverwertung und der durchschnittlichen Mastdauer
überdurchschnittlich. Unterschiede zwischen den beiden Impfgruppen rot und blau
sind für diese drei Parameter nicht signifikant. Tendenziell wiesen die Tiere der
Gruppe rot eine höhere tägliche Gewichtszunahme und eine kürzere Mastdauer auf.
Bei einer akuten Infektion mit H. parasuis während der Mastperiode wäre ein
größerer Unterschied zwischen den beiden Gruppen hinsichtlich der Effektivität des
Kombinationsimpfstoffes zu erwarten gewesen. Betrachtet man die einzelnen
Mastgruppen I bis IV, fällt auf, dass die Tiere aus dem Durchgang III insgesamt die
schlechtesten Mastleistungen erzielten. Schaut man sich dann die einzelnen
Impfgruppen an, kann man erkennen, dass die Unterschiede zwischen den beiden
Impfgruppen rot und blau hier am deutlichsten werden. Tiere, die mit Suvaxyn®
M.hyo-Parasuis geimpft wurden, haben eine um 17 g bessere tägliche Zunahme und
eine um 2,7 Tage kürzere Mastdauer. Daraus lässt sich folgern, dass sich bei
Mastdurchgängen mit vergleichsweise niedrigen Leistungen eher Effekte der Impfung
mit dem Impfstoff Suvaxyn® M.-hyo-Parasuis zeigen als bei Durchgängen, bei denen
die Leistungen insgesamt schon relativ gut sind.

Diskussion 95
5.4 Mortalität
Im Saugferkelbereich bestehen zwischen den beiden Gruppen keine nennenswerten
Unterschiede in dem Prozentsatz verendeter Tiere. Die Mortalitätsrate der Gruppe rot
betrug 3,4 %, die der Gruppe blau 3,7 %, berechnet nach der ersten Impfung am
zehnten Lebenstag, also ab Beginn des Versuchs. Es ist in diesem Zeitabschnitt
auch noch mit keinem Effekt der Vakzinierung zu rechnen, da die erste Impfung am
zehnten Lebenstag und die zweite in der dritten Lebenswoche durchgeführt worden
ist.
Im Ferkelaufzuchtstall unterscheiden sich die Gesamtzahlen der verendeten Tiere
kaum. In der Gruppe rot verendeten 14 Ferkel, in der Gruppe blau waren es 16
Ferkel. Bei Betrachtung der einzelnen Sektionsbefunde fallen jedoch Unterschiede
zwischen den beiden Gruppen auf. Der häufigste Befund war eine katarrhalische
Enteritis, die in der Gruppe rot bei fünf Ferkeln und in der Gruppe blau bei drei
Ferkeln festgestellt wurde.
Bei der Beurteilung der Sektionsbefunde, die für eine H.-parasuis-Infektion sprechen
oder zumindest einen Hinweis auf eine solche Infektion geben, sind Unterschiede
zwischen den beiden Gruppen zu erkennen. In der Gruppe rot zeigten drei Ferkel H.-
parasuis-assoziierte Befunde (alle drei Tiere zeigten eine Arthritis, zum Teil in
Kombination mit einer Bronchopneumonie oder einer Perikarditis). In der Gruppe
blau konnte bei vier Ferkeln der Befund Polyserositis, bei drei Ferkeln Arthritis (bei
einem Tier Arthritis in Kombination mit einer Perikarditis und einer
Bronchopneumonie) und bei einem Ferkel eine Bronchopneumonie festgestellt
werden. Die häufigsten H.-parasuis-assoziierten Befunde wurden bei den
Aufzuchtferkeln des II. Durchgangs gefunden. Auch zahlenmäßig war bei diesen
Tieren die Verlustrate am höchsten (Impfgruppe blau acht Tiere, Impfgruppe rot drei
Tiere), demnach kann vermutet werden, dass hier eine Feldinfektion mit H. parasuis
stattgefunden hat.
Bei all diesen Sektionsbefunden sind auch bakteriologische Untersuchungen
durchgeführt worden. Nur bei einem Tier der Gruppe rot konnte H. parasuis aus
Synovia isoliert werden. Bei allen anderen bakteriologischen Untersuchungen wurde
lediglich E. coli und S. suis (häufig Typ 4) aus Lungengewebe, Perikardflüssigkeit
und Synovia nachgewiesen.

96 Diskussion
VON ALTROCK (1998) bezeichnet Keime wie E. coli und -haemolysierende
Streptokokken im Zusammenhang mit Pneumonien als Begleitkeime, die häufig bei
bakteriologischen Untersuchungen in Lungengewebe nachgewiesen werden (E. coli
in 36 %, -haemolysierende Streptokokken in 27 % der untersuchten Lungen). Trotz
der hohen Nachweisrate werden diese Erreger als ätiologisch bedeutungslos für die
Entstehung von Pneumonien erachtet, da sie häufig zusammen mit bekannten
Krankheitserregern isoliert wurden. Nach WEISS und HEIDT (1982) beruhen hohe
Gehalte an Kontaminationen u. a. auf intra- und postmortalen Vorgängen bei
Sektionstieren.
Es ist jedoch möglich, dass H. parasuis seine Vermehrungsfähigkeit schon verloren
hatte und damit die Möglichkeit eines Nachweises nicht mehr gegeben war, da
zwischen Verenden und Sektion vermutlich eine zu lange Zeitspanne lag. Außerdem
gelingt der Nachweis von H. parasuis nicht, wenn die Tiere antibiotisch vorbehandelt
sind (OLIVEIRA u. PIJOAN 2002). In der vorliegenden Studie war dies bei den
meisten Tieren der Fall.
Bei drei der verendeten Ferkel der Gruppe blau waren im Vorfeld Anzeichen von
Kümmern aufgefallen (bei keinem Ferkel der Gruppe rot). Kümmern kann als
typisches Symptom einer H.-parasuis-Infektion angesehen werden, es tritt allerdings
erst bei chronischen Verläufen auf (WENDT 2004).
Laut serologischer Befunde hat während der Ferkelaufzucht (etwa zwischen der
sechsten und achten Lebenswoche) eine Feldinfektion stattgefunden. Trotz
mangelnder Signifikanzen der Mortalitätsdaten und fehlender Nachweise des
Erregers spricht das häufigere Vorkommen der H.-parasuis-typischen
Sektionsbefunde in der Gruppe blau und die durch die serologische Untersuchung
nachgewiesene Feldinfektion während der Aufzucht für eine Schutzwirkung von
Suvaxyn® M.hyo-Parasuis.
Die Serotypisierung des H.-parasuis-Isolats ergab die Serovare 2 und 9. Der
Impfstoff Suvaxyn® M.hyo-Parasuis enthält jedoch die Serovare 4 und 5. Die
Infektion könnte auf eine fehlende Kreuzprotektion zwischen dem Impfstamm und
dem Feldstamm zurückzuführen sein. RAPP-GABRIELSON et al. (1996)
untersuchten mit demselben Impfstoff (Suvaxyn® M.hyo-Parasuis) die Schutzwirkung
und Kreuzimmunitäten gegenüber den Serovaren 2, 4, 5, 12, 13 und 14. Auch sie
konnten keine Kreuzimmunität gegen die Serovare 2 und 14 feststellen, gegen die
übrigen untersuchten Serovare wurde ein wirksamer Schutz aufgebaut. Serovar 9

Diskussion 97
gehört zu den apathogenen Stämmen von H. parasuis (KIELSTEIN et al. 1992;
OLIVEIRA u. PIJOAN 2002).
Auch in der Mast unterscheiden sich die absoluten Zahlen der verendeten Tiere
zwischen den Impfgruppen kaum. In der Gruppe rot sind 13 Tiere verendet, in der
Gruppe blau zehn Tiere. Die Mortalitätsrate aller Schweine der Mastperiode beträgt 1
% und liegt somit weit unter den in der Praxis erzielten Daten. Im Mittel aller Betriebe
in Niedersachsen werden Verluste von 4,1 % verzeichnet (die 25 % der besseren
Betriebe haben Verlustraten von 3,4 %, die 25 % der schlechteren Betriebe Verluste
von 5,0 %) (ANONYM 2006).
Aber auch in diesem Zeitabschnitt scheint es angebracht, die Einzelbefunde der
Sektionen zu betrachten. Im IV. Durchgang verendeten die meisten Schweine durch
Kreislaufversagen gegen Ende der Mast (rot fünf Tiere, blau zwei Tiere). Diese
Verendungen sind wahrscheinlich auf Witterungsbedingungen zurückzuführen, da es
im Frühsommer eine kurze sehr heiße Periode gab. Von der Gesamtzahl der
verendeten Tiere der Durchgänge I bis IV zeigten neun Schweine in der Sektion
Einzelbefunde, die nicht in einem Zusammenhang mit einer Infektion mit H. parasuis
stehen.
H.-parasuis-assoziierte Symptome zeigte nur ein Tier der Impfgruppe rot
(Perikarditis), aus der Impfgruppe blau waren dies sechs Tiere. Bei drei Schweinen
der Gruppe blau wurde in der Sektion eine Bronchopneumonie und bei drei weiteren
Schweinen eine Perikarditis festgestellt. Auch bei diesen Befunden sind
bakteriologische Untersuchungen durchgeführt worden, jedoch konnte auch hier
lediglich E. coli und S. suis nachgewiesen werden. Es stellt sich die Frage, ob H.
parasuis, trotz fehlendem bakteriologischem Nachweis, in ursächlichem
Zusammenhang mit den Sektionsbefunden der Masttiere steht.
5.5 Therapeutische Maßnahmen / Tierbehandlungsindex (TBI)
Die Anzahl der Einzeltierbehandlungen der Saugferkel unterscheidet sich zwischen
den beiden Impfgruppen kaum und kann auch insgesamt als sehr gering
eingeschätzt werden. Eine Infektion mit H. parasuis kann schon in den ersten
Lebenstagen auftreten, da die Besiedlung des oberen Respirationstraktes schon
wenige Stunden nach der Geburt möglich ist (PIJOAN u. OLIVEIRA 2003). Bei den

98 Diskussion
Ferkeln dieses Versuches kann eine so frühzeitige Infektion nahezu ausgeschlossen
werden, da es kaum zu Behandlungen der Tiere gekommen ist und auch die
Mortalitätsrate im normalen Rahmen lag. Im Falle einer frühen Infektion wäre auch
mit Unterschieden zwischen den beiden Impfgruppen zu rechnen gewesen, aber
weder beim Tierbehandlungsindex noch bei der Mortalität sind signifikante
Unterschiede zu erkennen. Allerdings ist auch zu bedenken, dass während der
Saugferkelphase noch keine belastbare Immunität durch die Impfung aufgebaut
werden konnte, da die zweite Impfung erst in der dritten Lebenswoche durchgeführt
wurde.
In der Ferkelaufzucht wurden hoch signifikant weniger Einzelbehandlungen in der
Suvaxyn® M.-hyo-Parasuis-geimpften Gruppe durchgeführt als in der Suvaxyn® M.-
hyo-geimpften Gruppe. Die Gruppenbehandlungen zeigen kaum Unterschiede
zwischen den beiden Impfgruppen rot und blau. Dies ist damit zu begründen, dass im
Krankheitsfall parenterale Bestandbehandlungen durchgeführt wurden. Da jeweils
beide Impfgruppen eines Durchgangs in einem Abteil eingestallt waren, wurden auch
beide Impfgruppen gleich behandelt.
Zwischen den einzelnen Durchgängen (I bis IV) sind jedoch erhebliche Unterschiede
zu erkennen (Tab. 8). Bei dem Durchgang III sind die wenigsten Behandlungen
durchgeführt worden. Neben der prophylaktischen Behandlung gegen eine Coli- und
Streptokokkeninfektion bei Einstallung in den Aufzuchtstall, die bei allen vier
Durchgängen durchgeführt wurden, ist lediglich in der zehnten und elften Woche
wegen einer Bronchopneumonie behandelt worden. Einzeltierbehandlungen sind in
vernachlässigbarem Maß vorgenommen worden. Bei insgesamt geringem
antibiotischem Einsatz sind auch kaum Unterschiede zwischen den beiden
Impfgruppen zu erwarten. Anders sieht dieses beim II. Durchgang aus, der den
höchsten Einsatz von Antibiotika zu verzeichnen hat. Von Mitte bis kurz vor Ende der
Aufzucht waren klinische Anzeichen von Bronchopneumonie zu erkennen, und
dementsprechend sind auch Gruppenbehandlungen durchgeführt worden. Der letzte
Antibiotikaeinsatz in der Aufzucht betraf nur die Ferkel der Gruppe blau (Tab. 8).
Auch bei den Behandlungen einzelner Tiere bzw. Buchten ist der Index der Gruppe
blau hochsignifikant größer als der TBIeinzel der Gruppe rot. Bei der Sektion von
Tieren aus diesem Durchgang konnte bei drei Tieren eine Polyserositis (blau), bei
einem Tier eine Bronchopneumonie (blau), bei einem Tier eine Bronchopneumonie in
Kombination mit Arthritis (blau) und bei einem Tier eine Perikarditis in Kombination

Diskussion 99
mit einer Arthritis (rot) festgestellt werden. Bei letztgenanntem Ferkel konnte H.
parasuis aus Synovia isoliert werden.
Der signifikant höhere TBIeinzel, die Befunde der pathologisch-anatomischen
Untersuchungen, die Indikation Pneumonie für die Behandlungen sowie die hohe
Mortalitätsrate in diesem Durchgang sprechen für eine H.-parasuis-Infektion. Dieses
wird auch durch die Auswertung der Blutprobenergebnisse bestätigt.
Differentialdiagnostisch sind auch andere Infektionen des Respirationstraktes in
Betracht zu ziehen, allerdings fehlt für diese Erreger der bakteriologische Nachweis
und es wären vermutlich keine Unterschiede hinsichtlich der Behandlungen zwischen
den beiden Impfgruppen aufgetreten. Es kann davon ausgegangen werden, dass die
Suvaxyn® M.-hyo-Parasuis-geimpften Ferkel einen besseren Schutz besaßen.
Die Behandlungen während der Mast, quantitativ gemessen als TBIges. und TBIeinzel,
unterscheiden sich zwischen den beiden Impfgruppen nicht signifikant. Zwischen den
einzelnen Durchgängen gibt es Differenzen in der Behandlungshäufigkeit.
Durchgang I wurde lediglich vorbeugend gegen Atemwegserkrankungen bei der
Einstallung in die Mast behandelt, auch Einzeltierbehandlungen sind kaum
durchgeführt worden. Auch im II. und IV. Durchgang ist verhältnismäßig wenig
behandelt worden. Durchgang III jedoch weist die höchste Behandlungshäufigkeit
auf. In der Mittelmast war eine zweifache Behandlung wegen einer
Durchfallerkrankung (Ileitis) nötig, und gegen Ende der Mast musste noch zweimal
wegen einer Atemwegserkrankung behandelt werden. Entsprechend höher fallen
auch die Einzeltierbehandlungen aus. Diese Behandlungsunterschiede der einzelnen
Durchgänge können ätiologisch nicht auf eine H.-parasuis-Infektion zurückgeführt
werden, da die Unterschiede zwischen den Impfgruppen nicht signifikant sind. Die
Unterschiede liegen lediglich zwischen den einzelnen Durchgängen, was damit
begründet werden kann, dass die einzelnen Mastdurchgänge in verschiedenen
Mastställen stattgefunden haben und die Tiere jeweils von unterschiedlichen
Tierhaltern betreut wurden.

100 Diskussion
5.6 Lungenscore / AutoFOM-Werte
Die Bewertung der Lungenbefunde erfolgte nach dem Klassifikationssystem von
MADEC und KOBISCH (1985), wobei die Lungen am Schlachtband anhand der
Oberflächenveränderungen und der palpatorischen Befunde beurteilt wurden. In der
vorliegenden Untersuchung wurde so verfahren, dass ein Untersucher am
„Tierkörperband“ die individuelle Tätowierung der Tiere in der Reihenfolge notierte, in
der die Schlachtung erfolgte. Ein zweiter Untersucher, die Versuchsleiterin, beurteilte
die Lungen, die in der gleichen Reihenfolge wie die Tierkörper am Schlachtband
hingen, aber über das „Organband“ liefen.
Neben den Mastleistungen wurden Vorkommen und Ausmaß von
Lungenveränderungen als Maß für die Effektivität der Impfungen herangezogen.
Lungenveränderungen sind relativ eng mit dem Zuwachs korreliert (PAISLEY 1993).
Die Erfassung der AutoFOM-Daten haben auf vielen Schlachthöfen die Beurteilung
nach Magerfleischanteilen abgelöst. AutoFOM-Werte und der Magerfleischanteil
werden zur Beurteilung der Schlachtkörperqualität herangezogen und sind
Grundlage für die Bezahlung jedes einzelnen Schlachttieres.
Ein direkter Einfluss der Impfung auf die AutoFOM-Werte ist nicht zu erwarten.
Indirekt ist jedoch eine positive Beeinflussung über ein besseres Wachstum und
damit eine verkürzte Mastdauer möglich.
In einer Studie an einem Schlachthof in Weser-Ems wurde der Zusammenhang
zwischen Lungenveränderungen und dem Schlachtkörpergewicht untersucht
(BLAHA u. BLAHA 1994). Aus diesen Untersuchungsergebnissen geht hervor, dass
Schlachtschweine mit mittel- bis hochgradig krankheitsbedingt veränderten Lungen
hochsignifikant niedrigere Schlachtkörpergewichte (bis zu 8,8 kg) aufweisen als
Schweine mit unveränderten Lungen. Die tägliche Gewichtszunahme ist demnach
umso geringer, je stärker krankheitsbedingte Lungenveränderungen auftreten. Dies
ist wirtschaftlich umso bedeutsamer, als in aller Regel die Tiere mit den geringsten
Gewichten zuletzt ausgestallt werden und mehr Zeit benötigen, um höhere
Schlachtgewichte zu erzielen und es so zu einer verlängerten Mastdauer kommt. Zu
gleichen Ergebnissen kommt auch HOY (1994), der Auswirkungen von
Atemwegserkrankungen auf Mast- und Fruchtbarkeitsleistungen untersucht hat.

Diskussion 101
Demnach führen Atemwegserkrankungen bei Mastschweinen zu einer Verminderung
der Schlachtmasse um zwei bis sechs Kilogramm.
Dieser Zusammenhang wird auch in der vorliegenden Studie bestätigt. Die
Schlachtschweine mit den grenzwertig signifikant geringeren Lungenläsionen der mit
Suvaxyn® M.-hyo-Parasuis-geimpften Gruppe wiesen auch tendenziell, allerdings
nicht signifikant, eine kürzere Mastdauer auf.
In der oben erwähnten Studie von BLAHA und BLAHA (1994) wurde auch der
Zusammenhang zwischen Lungenveränderungen und der Handelsklasseneinstufung
untersucht. Die Einstufung der Schlachtkörper in Handelsklassen resultiert aus dem
Magerfleischanteil bzw. dem AutoFOM-Wert. Einen Magerfleischanteil von über 55 %
wiesen signifikant mehr Tiere ohne Lungenbefunde auf als Tiere mit
Lungenveränderungen, während Tiere mit Lungenveränderungen vornehmlich einen
Magerfleischanteil von 50 bis 55 % hatten. Dieser Zusammenhang konnte in der
vorliegenden Studie nicht bestätigt werden. Die AutoFOM-Werte, die synonym dem
Magerfleischanteil in die Handelsklasseneinstufung eingehen, der Schlachtschweine
dieses Versuches weisen keinen signifikanten Unterschied auf.
Eine Infektion mit M. hyopneumoniae führt zu typischen Lungenveränderungen in
den ventralen Abschnitten der Lobi craniales und mediales. Das veränderte Gewebe
ist verfestigt, verfärbt und kann am Schlachtband visuell und palpatorisch erfasst
werden (BERTSCHINGER et al. 1972; MADEC u. KOBISCH 1985).
Lungenveränderungen durch Infektionen mit H. parasuis sind durch katarrhalisch-
eitrige Bronchopneumonien gekennzeichnet (RITZMANN u. HEINRITZI 2005), die
jedoch sehr variabel sein können und nicht charakteristisch sind (JANETSCHKE et
al. 1977).
Lungenveränderungen traten sowohl in der mit dem Kombinationsimpfstoff als auch
in der mit dem Monoimpfstoff vakzinierten Gruppe auf. Die mittleren Werte bezüglich
der Einzelbefunde für Pleuritiden und Abszesse zeigen keine signifikanten
Unterschiede. Der Lungenbefund Bronchopneumonie trat in der Suvaxyn® M.-hyo-
Parasuis-geimpften Gruppe grenzwertig signifikant seltener auf als in der Suvaxyn®
M.-hyo-geimpften Gruppe.
In der Literatur wird ebenso von geringeren Lungenläsionen bei Tieren, die gegen H.
parasuis geimpft wurden, berichtet. BAUMANN und BILKEI (2002) haben in
Versuchen bei Ferkeln von geimpften Sauen signifikant weniger

102 Diskussion
Lungenveränderungen als bei den Ferkeln von ungeimpften Sauen gefunden. Zu den
gleichen Ergebnissen kommen auch SOLANO-AGUILAR et al. (1999); Tiere ohne
Impfschutz hatten einen deutlich höheren Anteil an Pneumonien als geimpfte Tiere.
Die Differenzen der Lungenveränderungen im vorliegenden Versuch sind nicht sehr
gravierend, was darauf zurückzuführen sein kann, dass die H.-parasuis-Feldinfektion
in der Aufzucht stattgefunden hat. Das Krankheitsgeschehen liegt vermutlich so
lange zurück, dass die Lungenaffekte bis zum Mastende teilweise wieder ausgeheilt
sind. Zu einem analogen Ergebnis kommt JØRGENSEN (1992), der geringere
Häufigkeiten pathologisch-anatomischer Lungenveränderungen bei Tieren mit länger
zurückliegenden klinischen Erkrankungen fand als bei Tieren, die in späteren
Mastabschnitten erkrankt waren.
NOYSE et al. (1990) geben ebenso zu bedenken, dass der Anteil an pneumonisch
verändertem Lungengewebe bei der Schlachtung eine irreführende Information bei
der Ermittlung von Auswirkungen, die Lungenerkrankungen auf die Mastleistung
haben, sein kann. Nach deren Untersuchungsergebnissen können
Lungenerkrankungen zu Beginn der Mast die Mastleistung stark beeinträchtigen und
nur zu geringen Veränderungen am Schlachtband führen, da die pathologisch-
anatomischen Veränderungen weitgehend abgeheilt sind. Dagegen können
Erkrankungen in der Endphase der Mast zu erheblichen Läsionen führen, die jedoch
die Mastleistungen nicht mehr entsprechend beeinflussen zu können.
5.7 Schlussfolgerung
Die Ergebnisse der vorliegenden Untersuchung lassen sich zu folgenden
Schlussfolgerungen zusammenfassen:
Durch die Impfung mit dem Kombinationsimpfstoff Suvaxyn® M.hyo-Parasuis im
Vergleich mit dem monovalenten Impfstoff Suvaxyn® M.hyo konnten bei fast allen
untersuchten Produktionsparametern tendenziell bessere Ergebnisse erzielt werden.
Allerdings lässt sich das nicht für jeden einzelnen Untersuchungspunkt statistisch
absichern.
In der Ferkelaufzucht hat laut serologischer Untersuchungen eine Feldinfektion mit H.
parasuis stattgefunden. Hier konnte gezeigt werden, dass die mit Suvaxyn® M.hyo-
Parasuis geimpften Ferkel hoch signifikant weniger behandelt worden sind.

Diskussion 103
Bei der Lungenbeurteilung am Schlachthof traten grenzwertig signifikant weniger
Pneumonien auf.
Zusammenfassend kann gesagt werden, dass bei beiden Impfgruppen während des
Untersuchungszeitraumes insgesamt sehr gute Aufzucht- und Mastleistungen erzielt
wurden. Bei den einzelnen Parametern wären größere Unterschiede zwischen den
beiden Impfgruppen zu erwarten gewesen. Eine Begründung hierfür kann darin
liegen, dass trotz einer Feldinfektion während der Ferkelaufzucht der Infektionsdruck
durch H. parasuis nicht sehr hoch war, was wiederum nur zu geringen klinischen
Ausprägungen geführt hat.
Insbesondere anhand der serologischen Untersuchungen konnte gezeigt werden,
dass durch die M.-hyopneumoniae-Komponente des Kombinationsimpfstoffes eine
dem Monoimpfstoff vergleichbare Immunitätsausbildung gewährleistet ist.
Auch die H.-parasuis-Komponente des Impfstoffes kann als immunogen angesehen
werden, da die Feldinfektion bei den geimpften Tieren zu einer höheren
Antikörperreaktion geführt hat und so eine Boosterwirkung durch die Impfung
entstanden ist. Daraus lässt sich auf einen Schutz der geimpften Ferkel gegen H.
parasuis schließen.
Die Ergebnisse der Serologie lassen durchaus den Schluss zu, dass trotz
zeitgleicher und lokal gleicher Applikation der beiden Komponenten des Impfstoffes
eine gute Antikörperentwicklung gegeben ist. Aus der Kombinationsimpfung ergeben
sich somit Vorteile, da die Belastung der Ferkel durch nur eine Vakzination minimiert
und die Arbeitsbelastung innerhalb eines Betriebes reduziert werden kann.
Die Anwendung des untersuchten Kombinationsimpfstoffes kann aus
immunologischen, tierschützerischen und ökonomischen Gründen empfohlen
werden.

104 Zusammenfassung
6 Zusammenfassung Claudia Meistermann
Vergleichende Untersuchung zur Effektivität eines Kombinationsimpfstoffes gegen Haemophilus parasuis und Mycoplasma hyopneumoniae und eines monovalenten Impfstoffes gegen Mycoplasma hyopneumoniae bei Schweinen
Infektionen mit H. parasuis sind in der Schweineproduktion weit verbreitet, wobei
nicht nur die typische Polyserositis als Glässersche Krankheit auftritt, sondern der
Erreger zunehmend auch an respiratorischen Erkrankungen beteiligt ist. Weltweit
lässt sich seit einigen Jahren eine Zunahme der Erkrankung beobachten. Die Gründe
hierfür scheinen die intensiven Haltungsbedingungen, das Zusammenbringen von
Aufzuchtferkeln und Mastschweinen aus verschiedenen Herkünften und frühes
Absetzen der Ferkel zu sein.
Die vorliegende Untersuchung wurde mit dem Ziel durchgeführt, die Effektivität eines
Kombinationsimpfstoffes (Suvaxyn® M.hyo-Parasuis) gegen H. parasuis und M.
hyopneumoniae und eines monovalenten Impfstoffes (Suvaxyn® M.hyo) gegen M.
hyopneumoniae unter Feldbedingungen zu überprüfen. Durchgeführt wurde der
Versuch im Raum Weser-Ems in einem Sauenbestand mit eigener Ferkelaufzucht
und anschließender Mast, in dem der Erreger H. parasuis endemisch vorkommt.
Die Studienpopulation bestand aus etwa 2800 Schweinen, die in vier Durchgängen
jeweils gleichmäßig auf die beiden Impfgruppen verteilt waren. Die Impfungen
wurden bei beiden Gruppen am zehnten Lebenstag und in der dritten Lebenswoche
durchgeführt. Die Effektivität wurde dabei anhand von Produktionsparametern, wie
tägliche Gewichtszunahmen, Futterverwertung, Mortalität, Tierbehandlungsindex,
durchschnittliche Mastdauer sowie Lungenveränderungen zum Zeitpunkt der
Schlachtung, abgeleitet. Außerdem wurde die humorale Reaktion auf die Impfung in
beiden Impfgruppen anhand einer serologischen Verlaufsuntersuchung verfolgt.
Die Ergebnisse können folgendermaßen zusammengefasst und interpretiert werden:
In der vorliegenden Untersuchung konnte anhand der serologischen
Verlaufsuntersuchungen eine Infektion mit H. parasuis während der Ferkelaufzucht
festgestellt werden.

Zusammenfassung 105
Die mit dem Impfstoff Suvaxyn® M.hyo-Parasuis geimpften Tiere wiesen bezüglich
der untersuchten Produktionsparameter tendenziell bessere Leistungen auf als die
Tiere, die nur gegen M. hyopneumoniae geimpft wurden.
Bei den gegen H. parasuis geimpften Tieren wurde während der Ferkelaufzucht
hochsignifikant weniger Antibiotika eingesetzt.
Ebenso zeigten die gegen H. parasuis geimpften Tiere grenzwertig signifikant
weniger und weniger schwere Lungenveränderungen zum Zeitpunkt der
Schlachtung.
Die M.-hyopneumoniae-Komponente des Kombinationsimpfstoffes gewährleistet eine
dem Monoimpfstoff vergleichbare Immunitätsausbildung.
Die H.-parasuis-Komponente induziert nach einer Feldinfektion eine höhere
Antikörperreaktion und damit eine Boosterwirkung.
Bei Einsatz des Kombinationsimpfstoffes ist, trotz zeitgleicher und lokal gleicher
Applikation der beiden Komponenten, eine gute Antikörperentwicklung und somit ein
guter Schutz sowohl gegen eine H.-parasuis- als auch gegen eine M.-
hyopneumoniae-Infektion gegeben.

106 Summary
7 Summary Claudia Meistermann
Comparative efficacy of a combination vaccine against Haemophilus parasuis and Mycoplasma hyopneumoniae and of a monovalent vaccine against Mycoplasma hyopneumoniae in pigs
Infections with H. parasuis are widely spread in swine production - and it is not only
the typical polyserositis occurring as Glässer disease, but the pathogen causes more
and more respiratory diseases. Since a few years, an increasing number of diseases
have been seen all over the world. The reasons for this seem to be the intensive
conditions of production, the allocation of nursery pigs and fattening pigs of different
herds and the early weaning of the piglets.
The aim of this study was to compare the efficiency of a combination vaccine
(Suvaxyn® M.hyo-Parasuis) against H. parasuis and M. hyopneumoniae and a
monovalent vaccine (Suvaxyn® M.hyo) against M. hyopneumoniae under field
conditions. The study has been carried out in a closed herd in the Northwest of
Germany, endemically infected with H. parasuis.
The study population consisted of about 2,800 pigs which, in four turns, were
allocated randomly and equally to the two treatment groups. The vaccinations were
performed for the two groups on the tenth day of life and in their third week of life.
The efficiency was evaluated by production parameters as the average daily weight
gain, feed conversion ratio, mortality, treatment index, average duration of fattening
as well as changes of the lung at slaughter. Additionally, the humoral reaction after
vaccination was compared using a longitudinal serological study.
Results and conclusion:
By means of serology an infection with H. parasuis during nursery phase could be
demonstrated.
The pigs vaccinated with the Suvaxyn® M.hyo-Parasuis vaccine showed a trend to
better performance than the pigs which only were vaccinated against M.
hyopneumoniae.

Summary 107
The pigs vaccinated with H. parasuis obtained highly significantly less antibiotics
during nursery phase.
Moreover, they showed borderline sigificantly less and less severe lung changes at
slaughter.
The M. hyopneumoniae component of the combination vaccine induces a humoral
immunity level comparable to that of the monovalent vaccine.
The H. parasuis component induces higher antibody reactivity and a booster effect
after field infection, compared to the control.
Despite of the same application time and location of the two components, the use of
the combination vaccine assures a good antibody development and therefore a good
protection against H. parasuis as well as against M. hyopneumoniae infections.

108 Literaturverzeichnis
8 Literaturverzeichnis AARESTRUP, F. M., A. M. SEYFARTH u. A. ØYSTEIN (2004): Antimicrobial susceptibility of Haemophilus parasuis and Histophilus somni from pigs and cattle in Denmark. Vet. Microbiol. 101, 143-146 ABE, T., M. IWASAKI, Y. YOSHIKAWA, T. MOROZUMI u. T. HIRAMUNE (1982): An outbreak of Glässer`s disease due to Haemophilus parasuis in piglets. Natl. Inst. Anim. Health Q 22, 182 ABIVEN, P., B. BLANCHARD, C. SAILLARD, M. KOBISCH u. J.M. BOVE (1992): A specific DNA probe for detecting Mycoplasma hyopneumoniae in experimentally infected piglets. Mol. Cell. Probs. 6, 423-429 ANDREASEN, M., J. MOUSING u. L. K. THOMSEN (2001): No simple association between time elapsed from seroconversion until slaughter and the extend of lung lesions in Danish swine. Prev. Vet. Med. 52, 147-161 ANGEN Ø., B. SVENSMARK u. K. R. MITTAL (2004): Serological characterization of Danish Haemophilus parasuis isolates. Vet. Microbiol. 103, 143-146 ANONYM (2006): Arbeitskreis Betriebszweigauswertung Schwein Niedersachsen (ABSN); Vereinigte Informationssysteme Tierhaltung w.V. (VIT) (2006): Berichte aus Verden, Ferkelerzeugung, Schweinemast 2005 ARMSTRONG, C.H., M.J. FREEMAN, L. SANDS-FREEMAN, M. LOPEZ-OSUNA, T.YOUNG u. L.J. RUNNELS (1983): Comparison of the enzyme-linked immunosorbent assay and the indirect hemagglutination and complement fixation test for detecting antibodies to Mycoplasma hyopneumoniae. Can. J. Comp. Med. 47, 464-470 BAK, H., u. H.-J. RIISING (2002): Protection of vaccinated pigs against experimental infections with homologous and heterologous Haemophilus parasuis. Vet. Rec. 151, 502-505 BAKOS, K., A. NILSSON u. E. THAL (1952): Untersuchungen über Haemophilus suis. Nord. Vet. Med. 4, 241-255 BAKOS, K. (1955): Studien über Haemophilus suis mit besonderer Berücksichtigung der serologischen Differenzierung. Vet. Med. Diss., Stockholm

Literaturverzeichnis 109
BARFOD, K., V. SØRENSEN u. N.C. FELD (1994): Evaluation of a monoclonal blocking ELISA detecting antibodies to Mycoplasma hyopneumoniae on a single-pig level. in: 13th Congr. Int. Pig Vet. Soc., Bangkok, Thailand1994, Proc. 189 BARRIGAZZI, G., F. VALENZA, E. BOLLO, F. GUARDA, P. CANDOTTI, A. RAFFO u. E. FONI (1994): Anatomohistopathological features related to Haemophilus parasuis infection in pigs. in: 13th Congr. Int. Pig Vet. Soc., Bangkok, Thailand 1994, Proc.13, 235 BATISTA, L., A. RUIZ, V. UTRERA u. C. PIJOAN (2002): Assessment of mechanical transmission of Mycoplasma hyopneumoniae. in: 17th Congr. Int. Pig Vet. Soc., Ames, USA 2002, Proc., 1, 285 BERGEY´S MANUAL OF DETERMINATIV BACTERIOLOGY (1994): Williams and Wilkins, Baltimore, USA BERNER, H. (1995): Impfung – eine neue Methode der Bekämpfung der Enzootischen Pneumonie des Schweines. Prakt. Tierarzt 14, 668-682 BERTSCHINGER, H. U., H. KELLER, A. LÖHRER u. W. WEGMANN (1972): Der zeitliche Verlauf der experimentellen Enzootischen Pneumonie beim SPF-Schwein. Schweiz. Arch. Tierheilk. 114, 107-116 BIBERSTEIN, E. L., u. D. C. WHITE (1969): A proposal for the astablishment of two new Haemophilus species. J. Med. Microbiol. 2, 75-78 BLAHA, T. (2005): Epidemiologische Ansätze zur Umsetzung der “risiko-orientierten“ Fleischuntersuchung. Vortrag auf der Fortbildungsveranstaltung „Das Ende der staatlichen Kontrolle?“ an der TiHo Hannover, 24.06.2005 BLAHA, T., u. M.-L. BLAHA (1994): Qualitätssicherung in der Schweinefleischerzeugung. Gustav Fischer Verlag, Jena, Stuttgart 1995, S.55-61 BLACKALL, P. J., u. J. L. PAHOFF (1995): Characterisation of porcine haemophili isolated from Australian pigs between 1988 and 1992. Austr. Vet J. 72, 18-21

110 Literaturverzeichnis
BLANCHARD, B., M. M. VENA, A. CAVALIER, J. Le LANNIC, J. GOURANTON u. M. KOBISCH (1992): Electronic microscopic observation of the respiratory tract of SPF piglets inoculated with Mycoplasma hyopneumoniae. Vet. Microbiol. 30, 329-341 BOETTCHER, T. B., B. J. THACKER, P. G. HALBUR, W. R. WATERS, R. NUTSCH u. E. T. THACKER (2002): Vaccine efficacy and immune response to Mycoplasma hyopneumoniae challenge in pigs vaccinated against Porcine Reproductive and Respiratory Syndrome Virus and Mycoplasma hyopneumoniae. Swine Health Prod. 10, 259-264 BRANDRETH S. R., u. I. M. SMITH (1986): Lack of pathogenity of Haemophili of the “minor group” taxon for the gnotobiotic piglet. Res. Vet Sci. 40, 273-275 BROCKMEIER, S. (2004): Prior infection with Bordetella bronchiseptica increases nasal colonization by Haemophilus parasuis in swine. Vet. Rec. 151, 502-505 BÜRGI, E., O. SEIZ u. H.U. BERTSCHINGER (1990): Lack of transmission of Mycoplasma hyopneumoniae from dam to fetus in experimentally infected pregnant gilts. in: 11th Congr. Int. Pig Vet. Soc., Lausanne, Schweiz 1990, Proc. 89 BUTTENSCHØN, J., N. F. FRIIS, B. AALBAEK, T. K. JENSEN, T. IBURG u. J. MOUSING (1997): Microbiology and pathology of fibrinous pericarditis in Danish slaughter pigs. J. Vet. Med. B 44, 271-280 CARUSO, J. P., u. R.F. ROSS (1990): Effects of Mycoplasma hyopneumoniae and Actinobacillus (Haemophilus) pleuropneumoniae infections on alveolar macrophage functions in swine. Am. J. Vet. Res. 51, S. 227-231 CHANDLER, C. A., L. D. FOTHERGILL u. J. H. DIGLE (1939): The pattern of dissociation in Haemophilus influenzae. J. Bacteriol. 37, 415-425 CLARK, L. K., C.H. ARMSTRONG, A.B. SCHEIDT, L. SANDS-FREEMAN u. K. KNOX (1992): Investigating the transmission of Mycoplasma hyopneumoniae in a swine herd with enzootic pneumonia. Vet. Med. 86, 543-550 CLARK, K., T. S. KNIFFEN et al. (1994): An evaluation of the components of medicated early weaning. Swine Health Prod. 2, 5-11

Literaturverzeichnis 111
CRAWFORD J. J., L. BARDEN u. J. B. KIRKMAN (1969): Selective culture medium to survey the incidence of Haemophilus species. Appl. Microbiol. 18, 646-649 DELBECK, F. (1995): Erfolgskontrolle einer oralen Chlortetracyclin-Behandlung bei lungenkranken Schweinen mittels der bronchoalveolären Lavage. Hannover, Tierärztliche Hochschule, Diss. DEL RÍO, M. L., C. B. GUTIĖRREZ u. E. F. RODRÍGUEZ FERRI (2003): Value of Indirect Hemagglutination and Coagglutination Tests for Serotyping Haemophilus parasuis. J. Clin. Microbiol. 41, 880-882 DESROSIERS, R., J. B. PHANEUF u. A. BROERS (1986): An outbreak of a typical Glasser`s disease in Quebec. in: 9th Congr. Int. Pig Vet. Soc., Barcelona, Spain 1986, Proc. 9, 277 DJORDJEVIC, S. P., G. J. EAMENS, L. F. ROMALIS, P. J. NICHOLLS, V. TAYLOR u. J. CHIN (1997): Serum and mucosal antibody responses and protection in pigs vaccinated against Mycoplasma hyopneumoniae with vaccines containingdenatured membrane antigen pool and adjuvant. Aust. Aet. J. 75, 504-511 DOHOO, I. R., u. M.E. MONTGOMERY (1996): A field trial to evaluate a Mycoplasma hyopneumoniae vaccine: Effects on lung lesions and growth rates in swine. Can. Vet. J. 36, 299-302 DONALDSON, D. M., R. R. ROBERTS, H. S. LARSEN u. J. C. TEW (1974): Interrelationship between serum beta-lysin, lysozyme and antibody-complement system in killing Escherichia coli. Infect. Immun. 10, 657-666 DONE, H. S. (1996): Enzootic pneumonia (mycoplasmosis) revisted. Pig Vet. J. 38, 40-61 DONE, H. S. (1999): Haemophilus parasuis: a synopsis. Pig Journ. 44, 207-221 DONE, H. S., u. D. J. PATON (1995): Porcine reproductive and respiratory syndrome: Clinical disease, pathology and immunosuppression. Vet. Rec. 136, 32-35 EAVES, L. E., P. J. BLACKALL u. M. FEGAN (1989): Characterisation and antimicrobial sensitivity of Haemophili isolated from pigs. Austr. Vet. J. 66, 1-4

112 Literaturverzeichnis
FALK, K., S. HØIE u. B.M. LIUM (1991): An abattoir survey of pneumonia and pleuritis in slaughter weight swine from 9 selected herds. II. Enzootic pneumonia of pigs: Microbiological findings and their relationship to pathomorphology. Acta Vet. Scand. 32, 67-77 FRIIS, N.F. (1973): Resistance of porcine mycoplasmas to drying. Acta Vet. Scand. 14, 489-491 GLÄSSER, K. (1910): Untersuchungen über die Schweineseuche mit besonderer Berücksichtigung ihrer Ätiologie und Pathologie. Dtsch. Tierärztl. Wochenschrift 18, 729-733 GOIS, M., H. J. BARNES u. R. F. ROSS (1983) : Potentation of turbinate atrophy in pigs by long-term nasal colonization with Pasteurella multocida. Am. J. Vet. Res. 44, 372-278 GOODWIN, R.F.W. (1972): Isolation of Mycoplasma suipneumoniae from the nasal cavaties and lungs of pigs affected with enzootic pneumonia or exposed to this infection. Res. Vet. Sci. 13, 262-267 GOODWIN, R.F.W. (1985): Apparent re-infection of enzootic-pneumonia-free pig herds: Search for possible causes. Vet. Rec. 116, 690-694 GOODWIN, R.F.W., R. HODGSON, P. WHITTELSTONE u. R. WOODHAMS (1969): Immunity in experimentally induced enzootic pneumonia of pigs. J. Hyg. 67, 193-208 GOODWIN, R.F.W., A. POMEROY u. P. WHITTELSTONE (1965): Production of enzootic pneumonia in pigs with a mycoplasma. Vet. Rec. 77, 1247-1249 GRUND S., P. SCHÖSS u. B. THIESSEN (1990): Pasteurella multocida (Typ D und A) und Rhinitis atrophicans des Schweines Hämagglutination, Fimbrien, sowie Adhäsion in vitro an der Nasenschleimhaut von Schweinefeten. Berl. Münch. Tierärztl. Wschr. 103, 365-371 GUNNARSON A. (1979): Serologic studies on porcine strains of Haemophilus parahaemolyticus (pleuropneumoniae): Extraction of type-specific antigens. Am. J. Vet. Res. 40, 469-472

Literaturverzeichnis 113
HEITMANN, J., u. H. KIRCHHOFF (1981): Untersuchungen zur Übertragbarkeit von Mykoplasmen auf den Fetus beim Schwein. Zbl. Vet. Med. B 28, S. 378-385 HEGE, R., W. ZIMMERMANN, R. SCHEIDEGGER u. K. D. C. STÄRK (2002): Incidence of reinfection with Mycoplasma hyopneumoniae and Actinobacillus pleuropneumoniae in pig farms located in respiratory-disease-free regions of Switzerland Acta Vet. Scand. 40, 145-156 HJÄRRE, A. (1957): Glässer`s Krankheit. Arch. Exp. Vet. Med. 11, 3-8 HJÄRRE, A., u. G. WRAMBY (1949): Experimental investigation into swine influenza in Sweden with spezial regard to the etiological importance of haemoglobinophilic bacteria. Rept. XIVth Intern. Vet. Cong. London 2, 465-471 HODGES, R. T., A. O. BETTS u. A. R. JENNINGS (1969): Production of pneumonia in gnotobiotic pigs with pure cultures of Mycoplasma hyopneumoniae. Vet. Rec. 84, 268-272 HOEFLING, D. C. (1991): Acute myositis associated with Haemophilus parasuis in primary SPF sows. J. Vet. Diagn. Invest: 3, 354-355 HOFFMANN, C. R., u. G. BILKEI (2002): The effect of a homologous bacterin given to sows prefarrowing on the development of Glässer`s disease in postweaning pigs after i.v. challenge with Haemophilus parasuis serotype 5. Dtsch. Tierärztl. Wschr. 109, 271-276 HORST, I., A. LINDNER, M. KRÜGER, H. R. GINDELE u. R. STING (1997): Verbreitung der Mycoplasma-hyopneumoniae-Infektion in Deutschland - Schlussfolgerungen für die Bekämpfung der Enzootischen Pneumonie der Schweine. Tierärztl. Umsch. 52, 508-514 HOWARD, C. J., u. G. TAYLOR (1985): Immune responses to Mycoplasma infections of the respiratory tract. Vet. Immunol. Immunopathol. 10, 3-32 HOY, St. (1994): Zu den Auswirkungen von Atemwegserkrankungen auf die Mast- und Fruchtbarkeitsleistungen der Schweine. Prakt. Tierarzt 2, 121-127

114 Literaturverzeichnis
JANETSCHKE, P., P. KIELSTEIN, W. SCHÖNHERR, J. MARTIN u. H. GÜNTHER (1977): Beitrag zur experimentellen Haemophilus-infektion (H. parahaemolyticus, H. parasuis) bei SPF-Ferkeln. Arch. Exp. Vet.-med. 31, 129-137, 347-357 JASQUES, M., BLANCHARD, B., FOIRY, B., GIRARD, G. u. M. KOBISCH (1992) : In vitro colonisation of porcine trachea by Mycoplasma hyopneumoniae. Ann. Rec. Vet. 23, 239-247 JØRGENSEN, B. (1992): Interrelations between diseases and effects of treatment procedures on some production parameters at Danish pig-breeding test stations. Prev. Vet. Med. 14, 267-279 JORGENSEN, J. H., J. S. REDDING, L. A. MAHER u. A. HOWELL (1987): Improved medium for antibicrobiol suscebility testing of Haemophilus influenzae. J. Clin. Microbiol. 25, 2105-2113 KIELSTEIN, P. (1985): Zur Glässerschen Krankheit und Chemotherapeutikaempfindlichkeit ihres Erregers. Mh. Vet.-med. 40, 806-809 KIELSTEIN, P. (1991): Zur Glässerschen Krankheit des Schweines. Untersuchungen über Zusammenhänge zwischen serologischen Eigenschaften, Kapselausbildung und Virulenz von H.-parasuis-Stämmen. Mh. Vet.-med. 46, 137 KIELSTEIN, P., u. R. LEIRER (1990): Zur Glässerschen Krankheit des Schweines – Ätiologisch-epizootiologische Untersuchungen zum Erregerspektrum. Mh. Vet.-med. 45, 577-582 KIELSTEIN, P., W. METHLING, D. SEYFARTH u. A. KELLER (1986): Zur Bedeutung der Glässerschen Krankheit in Schweinebeständen. Arch. exper. Vet. Med., Leipzig 40, 170-182 KIELSTEIN, P., V. J. RAPP-GABRIELSEN, et al. (1992): Designation of 15 serovars of Haemophilus parasuis on the basis of immunodiffusion using heatstable antigen extracts. J. Clin. Microbiol. 30, 862-865 KIELSTEIN, P., A. RAßBACH, D. PÖHLE, U. JOHANNSEN, M. WIEGAND u. M. SCHÄFER (1994): Zur Pathogenese der Haemophilus-parasuis-Infektin des Schweines (Glässersche Krankheit). Mh. Vet.-med. 46, 71-75

Literaturverzeichnis 115
KILIAN, M. (1976): A taxonomic study of the genus Haemophilus with the proposal of a new species. J. Gen. Microbiol. 93, 9-63 KIM J., H. CHUNG, T. JUNG, W. CHO, C. CHOI u. C. CHAE (2002): Postweaning multisystemic wasting syndrome of pigs in Korea: prevalence, microscopic lesions an coexisting microorganisms. J. Vet. Med. Sci. 64, 57-62 KLOBASA, F., u. J. E. BUTLER (1987): Absolute and relative concentrations of immunoglobulins G, M and A, and albumin in the lacteal secretion of sows of different lactation numbers. Am. J. Vet. Res. 48, 176-182 KLOBASA, F., F. HABE, E. WERHAHN u. J. E. BUTLER (1985): Changes in the concentration of serum IgG, IgA and IgM of sows throughout the reproductive cycle. Vet Immunol. Immunopathol. 10, 341-353 KOBAYASHI, H., T. MOROZUMI, C. MIYAMOTO, M. SHIMIZU, S. YAMADA, S. OHASHI, M. KUBO, K. KIMURA, K. MITANI, N. ITO u. K. YAMAMOTA (1996): Mycoplasma hyorhinis infection levels in lungs of piglets with porcine reproductive and respiratory syndrome (PRRS). J. Vet. Med. Sci. 58, 109-113 KOBISCH, M., B. BLANCHARD u. M. F. LE POTIER (1993): Mycoplasma hyopneumoniae infection in pigs: duration of the disease and resistance to reinfection. Vet. Res. 24, 67-77 KOBISCH, M., B. BLANCHARD u. M. F. LE POTIER (2000): Mycoplasma hyopneumoniae in pigs - old disease still causing trouble. in: 16th Congr. Int. Pig Vet. Soc., Melbourne, Australia 2000, Proc. 434-438 KOBISCH, M., A. LABBÉ, P. MORVAN u. R. CARIOLET (1994): Evaluation of Mycoplasma hyopneumoniae vaccine in pigs experimantally infected with Mycoplasma hyopneumoniae and Pasteurella multocida. in: 13th Congr. Int. Pig Vet. Soc., Bangkok, Thailand 1994, Proc. 194 KOBISCH, M., u. J. NICOLET (1987): Comparison of enzyme-linked immunosorbent assay (ELISA) and indirect haemagglutination (IHA) in experimental Mycoplasma hyopneumoniae infection of pigs. Isr. J. Med. Sci. 23, 644-646 KOHL, A. (1995): Untersuchungen beim Schwein über die Impftiterentwicklung nach einfacher oder kombinierter Vakzination mittels Mischinjektion mit mono- und polyvalenten Impfstoffen. Hannover, Tierärztliche Hochschule Hannover, Diss.

116 Literaturverzeichnis
KRISTENSEN, B., P. PAROZ, J. NICOLET, M. WANNER u. A. L. DE WECK (1982): Cell-mediated and humoral immune responses in swine after vaccination and natural infection with Mycoplasma hyopneumoniae. Am. J. Vet. Res. 42, 784-788 KUHN, M. J. (2002): Vaccine efficiacy and production performance of pigsvaccinated with RespiSure-ONETM.
in: 33rd Ann. Meeting Americ. Assoc. Swine Pract., Kansas City, Proc. 139-141 KUHN, M. J., B.J. THACKER, P.A. HAWKINS, W.R. WATERS u. E. L. THACKER (2000): Evaluation of the mode of action of RespiSure® Mycoplasma hyopneumoniae Bacterin. in: 31st Ann Meeting Americ. Assoc. Swine Pract., Indianapolis, Proc. 123 KURTH, K.T., T. HSU, E.R. SNOOK, E.L. THACKER, B.J. THACKER u. F.C. MINION (2002): Use of a Mycoplasma hyopneumoniae nested polymerase chain reaction test to determine the optimal sampling sites in swine. J. Vet. Diagn. Invest. 14, 463-469 KWON, D., u. C. CHAN (1999): Detection and localization of Mycoplasma hyopneumoniae DNA in lungs from naturally infected pigs by in situ hybridization using a digoxigeninlabeled probe. Vet. Pathol. 36, 308-313 LAM, K.M., u. W.P. SWITZER (1971) : Mycoplasmal pneumonia of swine: Development of an indirect hemagglutination test. Am. J. Vet. Res. 32, 1731-1736 LICHTENSTEIGER, C. A., u. E. R. VIMR (1997): Neuraminidase (sialidase) activity of Haemophilus parasuis. FEMS Microbiol. Lett. 152, 269-274 LITTLE, T. W. A. (1970): Haemophilus infection in pigs. Vet. Rec. 87, 399-402 LITTLE, T. W. A., u. J. D. J. HARDING (1971): The comparative pathogenicity of two porcine haemophilus species. Vet. Rec. 88, 540-545 LÓPEZ, R., S. MENDOZA, E. ESQUIVEL, A. ROMERO et al. (2004): Production of an immunofluorescent conjugate for the diagnosis of Glässer`s disease using Haemophilus parasuis as an antigen. in: 18th Congr. Int. Pig Vet. Soc., Hamburg 2004, Proc. 1, 210

Literaturverzeichnis 117
MADEC, F., u. KOBISCH, M. (1985): Disease registration on pigs at slaughter as a method of preventive and therapeutic veterinary medicine in swine production. Can. Vet. J. 26, 381-384 MAES, D. (1997): Risk indicators for the seroprevalence of Mycoplasma hyopneumoniae in slaughter pigs from farrow-to-finish pig herds. Epidemiol. Sante Anim. 5, 31-32 MAES, D. (2002): Herd factors associated with the seroprevalences of four major respiratory pathogens in slaughter pigs from farrow-to-finish pig herds. Vet. Res. 31, 313-327 MAES,D., H. DELUYKER, M. VERNDONCK, F. CASTRYCK, C. MIRY, A. LEIN, B. VRIJENS u. A. DE KRUIF (1998): The effect of vaccination against Mycoplasma hyopneumoniae in pig herds with a continuous production system. J. Vet. Med. B. 45, 495-505 MAES, D., H. DELUYKER, M. VERDONCK, F. CASTRYCK, C. MIRY, B. VRIJENS u. A. DE KRUIF (2000): Herd factors associated with the seroprevalences of four major respiratory pathogens in slaughter pigs from farrow-to-finish pig herds. Vet. Res. 31, 313-327 MAES,D., H. DELUYKER, M. VERNDONCK, F. CASTRYCK, C. MIRY, B. VRIJENS, W. VERBEKE, J. VIAENE u. A. DE KRUIF (1999): Effect of vaccination against M. hyopneumoniae in pig herds with an all-in/all-out production system. Vaccine 17, 1024-1034 MARE, C.J., u. W.P. SWITZER (1965): New species: Mycoplasma hyopneumoniae, a causative agent of virus pig pneumonia. Vet. Med. 60, 841-846 MATEUSEN, B., D. MAES, M. VAN GOUBERGEN, M. VERNDONCK u. A. DE KRUIF (2002): Effectiveness of treatment with lincomycin hydrochloride and/or vaccination against Mycoplasma hyopneumoniae for controlling chronic diseaese in herd of pigs. Vet. Rec. 151, 135-140 MESSIER, S., R. F. ROSS u. S. PAUL (1990): Humoral and cellular immune responses of pigs inoculated with Mycoplasma hyopneumoniae. Am. J. Vet. Res. 51, 52-58

118 Literaturverzeichnis
MEYLING, A. (1971): Mycoplasma suipneumoniae and Mycoplasma hyorhinis demonstrated in pneumonic pig lungs by the fluorescent antibody technique. Acta Vet. Scand. 12, 137-141 MINIATS, O. P., N. L. SMART u. E. EWERT (1991a): Vaccination of gnotobiotic primary specific pathogen-free pigs against Haemophilus parasuis. Can. J. Vet. Res. 55, 33-36 MINIATS, O. P., N. L. SMART u. S. ROSENDAL (1991b): Cross-protection among Haemophilus parasuis strains in immunized gnotobiotic pigs. Can. J. Vet. Res. 55, 37-41 MØLER, K., u. M. KILIAN (1990): V-factor dependent members of the family Pasteurellaceae in the porcine upper respiratory tract. J. clin. Microbiol. 28, 2711-2716 MORIKOSHI, T., K. KOBAYASHI, T. KAMINO, S. OWAKI, S. HAYASHI u. S. HIRANO (1990) Characterisation of Haemophilus parasuis isolated in Japan. Jpn. J. Vet. Sci. 52, 1990, 667-669 MOROZUMI, T., u. J. NICOLET (1986a): Morphological variations of Haemophilus parasuis strains. J. Clin. Microbiol. 23, 138-142 MOROZUMI, T., u. J. NICOLET (1986b): Some antigenic properties of Haemophilus parasuis and a proposal for new serological classification. J. Clin. Microbiol. 23, 1022-1025 MORRIS, C.R., I. A. GARDNER, S. K. HIETALA, T. E. CARPENTER, R. J. ANDERSON u. K. M. PARKER (1994): Persistence of passively acquired antibodies to Mycoplasma hyopneumoniae in a swine herd. Prev. Vet. Med. 21, 29-41 MORRIS, C.R., I. A. GARDNER, S. K. HIETALA, T. E. CARPENTER, R. J. ANDERSON u. K. M. PARKER (1995): Seroepidemiologic study of natural transmission of Mycoplasma hyopneumoniae in a swine herd. Prev. Vet. Med. 21, 323-337 MÜLLER, C. A. (2004): Haemophilus parasuis-Infektion: eine sero-epidemiologische Studie in geimpften und ungeimpften sowie infizierten und nicht infizierten Schweinebeständen. Bern, Vetsuisse-Fakultät der Universität Bern, Diss.

Literaturverzeichnis 119
MÜLLER, C., M. DOHERR, C. EGLI, D. SICHER et al. (2004): Haemophilus parasuis infection: vaccination and serological follow-up. in: 18th Congr. Int. Pig Vet. Soc., Hamburg 2004, Proc. 1, 817 MÜNCH, S. (1993): Untersuchungen zum Nachweis von Virulenzfaktoren bei Haemophilus parasuis. HU-Berlin, Diss. NARITA, M., K. KAWASHIMA, S. MATSUURA, A. UCHIMURA u. Y. MIURA (1994): Pneumonia in pigs infected with pseudorabies virus and Haemophilus parasuis serovar 4. J. Comp. Path. 110, 329-339 NEIL D., K. MC KAY, C. L. ECUYER u. A. H. CORNER (1969): Glässer´s disease of swine produced by the intratracheal inoculation of Haemophilus suis. Can. J. Comp. Med. 33, 187-193 NEUMANN, U., u. K.-H. HINZ (1977): Elektrophoretische Auftrennung von Haemophilus-Proteinen im Polyacrylamid-Gel. Zbl. Bakt. Hyg., I. Abt. Orig. A 238, 244-250 NICOLET, J., P. PAROZ u. M. KRAWINKLER (1980): Polyacrylamide gel electrophoresisof whole-cell proteins of porcine strainsof Haemophilus. Int. J. Syst. Bacteriol. 30, 69-76 NICOLET, J., P. PAROZ u. S. BRUGGMANN (1990): Tween 20 soluble proteins of Mycoplasma hyopneumoniae as antigen for an ELISA. Res. Vet. Sci. 29, 305-309 NIELSEN, R. (1993): Pathogenicity and immunity studies of Haemophilus parasuis serovars. Acta. Vet. Scand. 34, 193-198 NIELSEN, R., u. V. DANIELSEN (1975): An outbreak of Glässer`s Disease. Nord. Vet. Med. 27, 20-25 NIEMANN, O. (1999): Kontrolle der Wirksamkeit eines inaktivierten Impfstoffes gegen Mycoplasma hyopneumoniae bei Vakzination von Saugferkeln mittels klinischer, serologischer, bakteriologischer und zytologischer Untersuchung. Hannover, Tierärztliche Hochschule Hannover, Diss. NIENHOFF, H. (2005): Vor Streptokokken ist keiner sicher. Top agrar, 10, 10-13

120 Literaturverzeichnis
NOYSE, E. P., D. A. FEENEY u. C. PIJOAN (1990): Comparison of the effect of pneumonia detected during lifetime with pneumonia detected at slaughter on growth in swine. J. Am. Vet. Med. Assoc. 197, 1025-1029 OLBERTZ, B. (1990): Untersuchungen der Impftiter-Entwicklung beim Schwein nach Verabreichung mono- und polyvalenter Impfstoffe als Einfach- oder Kombinationsimpfung. Hannover, Tierärztliche Hochschule Hannover, Diss. OLIVEIRA, S., L. GALINA u. C. PIJOAN (2001): Development of a PCR test to diagnose Haemophilus parasuis infections. J. Vet. Diagn. Invest. 13, 495-501 OLIVEIRA, S., J. MAHLBERG u. R. SIMONSON (2004): Safety of controlled exposure to Haemophilus parasuis: the role of sow vaccination and PRRSV infection. in: 18th Congr. Int. Pig Vet. Soc., Hamburg 2004, Proc. 1, 189 OLIVEIRA, S., T. OLESON, M. D. TITUS u. R. SIMONSON (2004): Comparison of Haemophilus parasuis genotyping using amplified fragment lenght polymorphism (AFLP) and ERIC-PCR. Am. Assoc. Sw. Vet., 2004, 273-276 OLIVEIRA, S., u. C. PIJOAN (2002): Diagnosis of Haemophilus parasuis in affected herds and use of epidemiological data to control disease. J. Swine Health and Prod., 10, 221-225 OLIVEIRA, S., u. C. PIJOAN (2004): Haemophilus parasuis: new trends on diagnosis, epidemiology and control. Vet. Microbiol. 99, 1-12 OLIVEIRA, S., C. PIJOAN u. R. MORRISON (2002): Role of Haemophilus parasuis in nursery mortality. Proc. of the Allen D. Leman Swine Conference, Minneapolis USA, S. 111 OLIVEIRA, S., C. PIJOAN u. R. MORRISON (2004): Evaluation of Haemophilus parasuis control in the nursery using vaccination and controlled exposure. J. Swine Health Prod., 12(3), 123-128 PABST, T. (2004): Ausbreitung von M. hyopneumoniae in endemisch infizierten Schweineherden und Bekämpfung der Enzootischen Pneumonie mittels one-shot Impfung (Ingelvac®M.hyo). Hannover, Tierärztliche Hochschule Hannover, Diss.

Literaturverzeichnis 121
PAISLEY, L. G., L. VRAA-ANDERSEN, L. DYBKJAER, K. MØLLER, G. CHRISTENSEN, J. MOUSING u. J. F. AGGER (1993): An epidemiologic and economic study of respiratory diseases in two conventional danish swine herds: II: Associations between lesions present at slaughter and mean daily gains during specific intervals of the growth period. Acta. Vet. Scand. 34, 331-334 PARK, S. C., S. YIBCHOK-ANUN, H. CHENG, T. F. YOUNG, E. L. THACKER, F. C. MINION, R. F. ROSS u. W. H. HSU (2002): Mycoplasma hyopneumoniae increase intracellular calcium release in porcine ciliated tracheal cells. Infect. Immun. 6, 2502-2506 PEDERSEN, M. W., N. F. FRIIS u. P. AHRENS (1990): Detection of Mycoplasma hyopneumoniae in pneumonic pig lungs by an ELISA using monoclonal antibodies. in: 8th Congr. of I.O.M., Istanbul, Abstr., 399-400 PEET, R. L., J. FRY, J. LLOYD, J. HENDERSON, J. CURRAN u. D. MOIR (1983): Haemophilus parasuis septicaemia in pigs. Austr. Vet. J. 60, 187 PIFFER, I. A., u. R. F. ROSS (1984): Effect of age on susceptibility of pigs to Mycoplasma hyopneumoniae pneumonia. Am. J. Vet. Res. 45, 478-481 PIJOAN, C., M. TORREMOLL u. G. SOLANO (1997): Colonization patterns by the bacterial flora of young pigs. Am. Assoc. Swine Pract., 463-464 PIJOAN, C., u. S. OLIVEIRA (2002): Haemophilus parasuis: challenges and changes. Am. Assoc. Sw. Vet. 2002, 251-253 PIJOAN, C., u. S. OLIVEIRA (2003): Haemophilus parasuis: trends and new knowledge. Am. Assoc. Sw. Vet. 2003, 401-403 PLONAIT, H. (1988): Geburt, Puerperium und perinatale Verluste. in: Lehrbuch der Schweinekrankheiten, Verlag P. Parey, Berlin u. Hamburg, S. 309-348 PÖHLE, D., U. JOHANNSEN, P. KIELSTEIN, A. RAßBACH u. M. WIEGAND (1992): Investigations on pathology and pathogenesis of Haemophilus parasuis infection of swine. in: 12th Congr. Int. Pig Vet. Soc., The Hague, Netherlands 1992, Proc. 12., 335

122 Literaturverzeichnis
RADELOFF, I. (1997): Untersuchungen zur Wirksamkeit eines inaktivierten Mycoplasma hyopneumoniae-Impfstoffes (Stellamune® Mycoplasma) bei unterschiedlichen Vakzinationszeitpunkten. München, Ludwig-Maximilians-Universität, Tierärztl. Fak., Diss. RADOSTITIS, O. M., H. L. RUHNKE u. G. J. LOSOS (1963): An outbreak of meningitis in swine caused by Haemophilus species of bacterium. Can. Vet. Jour. 4, 265-270 RAFIEE, M., u. P. J. BLACKALL (2000): Establishment, validation and use of the Kielstein-Rapp-Gabrielson serotyping sheme for Haemophilus parasuis. Aust. Vet. J. 78, 172-174 RAPP-GABRIELSON, V. J. (1999): Haemophilus parasuis. in: B.E. STRAW, W.L. MENGELING, S. D’ALLAIRE u. D.J. TAYLOR (Hrsg.): Diseases of swine, 8th edition, Wolfe Publishing Ltd., S. 475-479 RAPP-GABRIELSON, V. J., u. D. A. GABRIELSON (1990): Serotyping of Haemophilus parasuis by Agar-Gel-Precipitation Test (AGPT). in: 11th Congr. Int. Pig Vet. Soc., Lausanne, Schweiz 1990, Proc. 523 RAPP-GABRIELSON, V. J., u. D. A. GABRIELSON (1992): Prevalence of Haemophilus parasuis serovars among isolates from swine. Am. J. Vet. Res. 53, 659-664 RAPP-GABRIELSON, V. J., G. J. KOCUR, J. T. CLARK, S. MUIR u. R. WHITE (1996): Efficiacy and duration of immunity of the Haemophilus parasuis fractions of Suvaxyn® Respifend® MH/HPS. in: 14th Congr. Int. Pig Vet. Soc., Bologna, Italy 1996, Proc. 300 RAPP-GABRIELSON, V. J., G. J. KOCUR, J. T. CLARK u. K. M. STEPHEN (1997): Haemophilus parasuis: immunity in swine after vaccination. Vet. Med. 92, 83-90 RAUTIAINEN, E., u. P. WALLGREN (2001): Aspects of the transmission of protection against Mycoplasma hyopneumoniae from sow to offspring. J. Vet. Med. B 48, 55-65 RAUTIAINEN, E., P. WALLGREN, P. VEIJALAINEN, K. E. JOHANSSON u. I. LAAMANEN (1998): The transmission of Mycoplasma hyopneumoniae between sow and piglets during the first 14 days of piglets life. in: 15th Congr. Int. Pig Vet. Soc., Birmingham, Great Britain 1998, Proc. S. 250

Literaturverzeichnis 123
RAZIN, S. (1992): Peculiar properties of mycoplasmas: The smallest self-replicating prokaryotes. FEMS Microbiol. Lett. 100, 423-432 RILEY, M. G. I., E. G. RUSSEL u. R. B. CALLINAN (1977): Haemophilus parasuis infection in swine. J. Am. Vet. Med. Ass. 171, 649-651 RITZMANN, M., H. GERBERMANN, H. GYRA, H. M. EICHINGER u. K. HEINRITZI (1999): Vergleichende Untersuchungen über den Einsatz eines Parvovirose-Rotlauf-Kombinationsimpfstoffes sowie entsprechender Monovakzinen bei unterschiedlichen Revakzinationszeitpunkten. Tierärztl. Praxis 27 (G), 168-174 RITZMANN, M., u. K. HEINRITZI (2005): Klinisches Bild, Diagnostik und Differenzialdiagnostik der Glässer’schen Krankheit. Tierärztl. Praxis 33 (G), 61-64 ROSENDAL S., D. A. BOYD u. K. A. GILBRIDE (1985): Comparative virulence of porcine Haemophilus bacteria. Can. J. Comp. Med. 49, 68-74 ROSNER, H., P. KIELSTEIN, W. MÜLLER u. B. ROHMANN (1991): Beziehung zwischen Serotyp, Virulenz und SDS-PAGE-Proteinmuster von Haemophilus parasuis. Dtsch. Tierärztl. Wschr. 98, 327-330 ROSS, R.F. (1996): Pathogenetic factors in, and pathogenesis of, mycoplasmal pneumonia. in: A. D. Leman Swine Conf., St. Paul 1996, Proc., S. 124-179 ROSS, R. F. (1999): Mycoplasmal diseases. in: B.E. STRAW, W.L. MENGELING, S. D’ALLAIRE u. D.J. TAYLOR (Hrsg.): Diseases of swine, 8th edition, Wolfe Publishing Ltd., S. 495-509 RUBIES, X., P. KIELSTEIN, L.I. COSTA, P. RIERA, C. ARTIGAS u. E. ESPUNA (1999): Prevalence of Haemophilus parasuis serovars isolated in Spain from 1993 to 1997. Vet. Microbiol. 66, 245-248 RUIZ, A. u. C. PIJOAN (2002): Use and interpretation of nPCR from nasal swabs for Mycoplasma hyopneumoniae diagnosis. in: 17th Congr. Int. Pig Vet. Soc., Ames,USA 2002, Proc. 1, 79

124 Literaturverzeichnis
SCHEIDT, A., V. MAYROSE, W. VAN ALSTINE, L. CLARK, T. CLINE u. M. EINSTEIN (1994): The effects of vaccinating pigs for mycoplasmal pneumonia in a swine herd affected by enzootic pneumonia. Swine Health Prod. 2, No. 1, S. 7-11 SCHIMMEL, D., A. HOGG u. J. UHLEMANN (1992): Prevention of Haemophilus parasuis infection with an inactivated vaccine. in: 12th Congr. Int. Pig Vet. Soc., The Hague, Netherlands 1992, Proc. 12, 336 SCHIMMEL, D., P. KIELSTEIN u. R. HASS (1985): Zur serologischen Typisierung von Haemophilus parasuis. Arch. Exper. Vet.-med. 39, 944 SCHNAITMANN, C. A. (1970): Protein composition of the cell wall and cytoplasmatic membrane of Escherichia coli. J. Bacteriol. 104, 890-901 SCHREIBER, A. (2002): Untersuchungen zum Einfluß maternaler Antikörper auf die humorale Immunantwort bei Ferkeln, die in der ersten und vierten bzw. vierten und achten Lebenswoche gegen M. hyopneumoniae (Hyoresp®, Firma Merial) geimpft wurden. Hannover, Tierärztl. Hochschule, Diss. SCHROER W. (1992): Untersuchung und Charakterisierung der Oberflächeneigenschaften von Haemophilus parasuis. Justus-Liebig-Universität Gießen, Diss. SCHULZ, L. C. (1991): Glässersche Krankheit. in: Pathologie der Haustiere, Gustav Fischer Verlag, Jena, S. 118-119 SEGALES, J., M. DOMINGO, G. I. SOLANO u. C. PIJOAN (1999): Porcine reproductive and respiratory syndrome virus and Haemophilus parasuis antigen distribution in dually infected pigs. Vet. Microbiol. 64, 287-297 SELTMANN, G. (1982): Die bakterielle Zellwand. Gustav Fischer Verlag, Jena SHELDRAKE, R.F., I. A. GARDNER, M. M. SAUNDERS u. L.F. ROMALIS (1990): Serum antibody response to Mycoplasma hyopneumoniae measured by enzym-linkedimmunosorbent assay after experimental and natural infection of pigs. Aust. Vet. J. 67, 39-42 SHELDRAKE, R.F., u. L.F. ROMALIS (1992): Evaluation of an enzyme-linked immunosorbent assay for the detection of Mycoplasma hyopneumoniae antibodies in porcine serum. Aust. Vet. J. 67, 39-42

Literaturverzeichnis 125
SHIFRINE M., u. E. L. BIBERSTEIN (1960): A growth factor for Haemophilus species secreted by a pseudomonad. Natur 187, 623 SLAVIK, M. F., u. W. P. SWITZER (1972): Development of a microtitration complement-fixation test for diagnosis of mycoplasmal swine pneumonia. Iowa State J. Res. 47, 117-128 SMART, N. L. (1987): Haemophilus parasuis: Subclinical infection and disease in southern Ontario swine; restriction endonuclease analysis of field isolates and prevention of disease through vaccination. M Sc thesis, University of Guelph, Guelph, Ontario, 1987 SMART, N.L., u. O. P. MINIATS (1989): Preliminary assessment of a Haemophilus parasuis bacterin for use in specific pathogen free swine. Can. J. Vet. Res. 53, 390-393 SOLANO, G., J. SEGALES, J. COLLINS, T. W. MOLITOR u. C. PIJOAN (1997): Porcine reproductive and respiratory syndrome virus (PRRSv) interaction with Haemophilus parasuis. Vet. Microbiol. 55, 247-257 SOLANO-AGUILAR, G. I., C. PIJOAN, V. RAPP-GABRIELSEN, J. COLLINS, L. F. CARVALHO u. N. WINKELMAN (1999): Protective role of maternal antibodies against Haemophilus parasuis infection. Am. J. Vet. Res. 60, 81-87 SØRENSEN, V., P. AHRENS, K. BARFOD, A. A. FEENSTRA, N. C. FELD, N. F. FRIIS, V. BILLE-HANSEN, N. E. JENSEN u. M. W. PEDERSEN (1997): Mycoplasma hyopneumoniae infection in pigs: Duration of the disease and evaluation of four diagnostic assays. Vet. Microbiol. 54, 247-257 TAKAHASHI, K., S. NAGAI, T. YAGIHASHI, T. IKEHATA, Y. NAKANO, K. SENNA, T. MARUYAMA u. J. MUROFUSHI (2001): A cross-protection experiment in pigs vaccinated with Haemophilus parasuis serovars 2 and 5 bacterin, and evaluation of a bivalent vaccine under laboratory and field conditions. J. Vet. Med. Sci. 63, 487-491 TARASENOK, N. J. (1982): Carriage of Haemophilus parasuis the causal agent of polyserositis by healthy swine. Veterinarija Moskva, 6, 67-68

126 Literaturverzeichnis
THACKER, E.L (2001): Mycoplasma diagnosis and immunity. in: 32th Ann. Meeting Americ. Assoc. Swine Pract., Nashville, 467-469 THACKER, E.L., B. J. THACKER, T.B. BOETTCHER u. H. JAYAPPA (1998): Comparison of antibody production, lymphocyte stimulation and protection induced by four commercial Mycoplasma hyopneumoniae bacterins. Swine Health Prod. 6, 107-112 THACKER, E.L., B. J. THACKER, M. KUHN, P. A. HAWKINS u. W. R. WATERS (2000): Mucosal and systemic characteristics of protective activity of a Mycoplasma hyopneumoniae bacterin. Am. J. Vet. Res. 61, 1384-1389 TRIGO, E., A. V. MENDEZ-TRIGO u. R. SIMONSON (1996): Antimicrobiol susceptibility profiles of Haemophilus parasuis: A retrospective study from clinical cases submitted during 1994 and 1995 to a veterinary diagnostic laboratory. in: 14th Congr. Int. Pig Vet. Soc., Bologna, Italy 1996, Proc. 14, 313 TULLY, J. G., J. M. BOVÉ, F. LAIGRET u. R. F. HWITCOMB (1993): Revised taxonomy of the class Mollcutes: proposed evaluation of a monophyletic cluster of arthropod-associated mollicutes to orinal rank (Entomoplamatales ord. Nov.), with provisionfor tamilial rank to separate species (Spiroplasmataceae), and emended descriptions the Order Mycoplasmatales, family Mycoplasmataceae. Int. J. Syst. Bacteriol. 43, 378-385 VAHLE, J. L., J. S. HARNESS u. J. J. ANDREWS (1995): Experimental reproduction of Haemophilus parasuis infection in swine: Clinical, bacteriological and morphologic findings. J. Vet. Diagn. Invest 7, 476-480 VALHE, J. L., J. S. HAYNES u. J. J. ANDREWS (1997): Interaction of Haemophilus parasuis with nasal and tracheal mucosa following intranasal inoculation of cesarean derived colostrum deprived (CDCD) swine. Can. J. Vet. Res. 61, 200-206 VAN REETH, K. (1997): Pathogenesis and clinical aspects of a respiratory porcine and reproductive and respiratory syndrome virus infection. Vet. Microbiol. 55, 223-230 VON ALTROCK, A. (1998): Untersuchungen zum Vorkommen bakterieller Infektionserreger in pathologisch-anatomisch veränderten Lungen von Schweinen und Zusammenstellung der Resistenzspektren. Berl. Münch. Tierärztl. Wschr. 111, 164-172

Literaturverzeichnis 127
VOS, J. (2004): Glässer`s Disease. in: 18th Congr. Int. Pig Vet. Soc., Hamburg, Proc. VRAA-ANDERSEN, L., G. CHRISTENSEN u. R. KUIPER (1994): Vaccine efficacy trial with Suvaxyn® M.hyo in Denmark. in: 13th Congr. Int. Pig Vet. Soc., Bangkok, Thailand 1994, Proc. 192 WALLGREN, P., G. BÖLSKE, S. GUSTAFSSON, S. MATTSON u. C. FOSSUM (1998): Humoral immune responses to Mycoplasma hyopneumoniae in sows and offspring following an outbreak of mycoplasmosis. Vet. Microbiol. 60,193-205 WALLGREN, P., O. SCHWAN, S. MATTSON u. G. BÖLSKE (1996): Comparison of the sensitivity of two ELISA systems for detection of antibodies to Mycoplasma hyopneumoniae in naturally infected pigs. in: 14th Congr. Int. Pig Vet. Soc., Bologna, Italy 1996, Proc. 217 WEGNER, M. D., B. J. THACKER u. E. L. THACKER (2002): Evaluation of the decay in colostrum derived maternal antibodies against Mycoplasma hyopneumoniae in swine. in: 33rd Ann. Meeting Americ. Assoc. Swine Pract., Kansas City, 71-74 WENDT, M. (2004): Die Glässersche Krankheit ist nicht tot. Top agrar 2, S10-13 WEISS, R., u. M. HEIDT (1982): Zum Vorkommen bakterieller Infektionserreger in pathologisch-anatomisch veränderten Lungen von Schweinen unter besonderer Berücksichtigung der Gattung Haemophilus. 1. Isolierungsergebnisse aus Lungen von Schlachtschweinen und Sektionsfällen. Berl. Münch. Tierärztl. Wochenschr. 95, 442-446 WHITTELSTONE, P. (1990): Control of Enzootic Pneumonia infection in pigs. in: G. STANEK, G.H. CASSEL, J.G. TULLY u. R.F. WHITCOMB (Hrsg.): Recent advances in mycoplasmology. Proc. 7th Congress of I.O.M., Baden 1990, Zbl. Suppl. 20, Gustav Fischer Verlag, Stuttgart, New York, S. 254-259 WIEGAND, M., M. SCHÄFER, P. KIELSTEIN, A. RAßBACH et al. (1992): Clinical, haematological and cerebrospinal fluid findings of primary specific-pathogen-free piglets after experimental infection with Haemophilus parasuis. in: 12th Congr. Int. Pig Vet. Soc.,The Hague, Netherlands 1992 , Proc. 332 WOESE, C. R. (1987): Bacterial evolution. Microbiol. Rev. 51, 221-271

128 Literaturverzeichnis
YAGIHASHI T., T. NUNOYA, T. MITIU u. M. TAJIMA (1984): Effect of Mycoplasma hyopneumoniae infection on the development of Haemophilus pleuropneumoniae pneumonia in pigs. Jap. J. Vet. Sci. 46, 705-713 ZIELINSKI, G.C., u. R.F. ROSS (1993): Adherence of Mycoplasma hyopneumoniae to porcine ciliated respiratory tract cells. Am. J. Vet. Res. 54, 1262-1269 ZIMMERMANN, W., u. C. MÜLLER (2004): Bedeutung und Bekämpfung der „Glässerschen Krankheit“ – Transportkrankheit – Haemophilus parasuis-Infektion in der Schweiz. in: 18th Congr. Int. Pig Vet. Soc.,Hamburg 2004, Proc. 332 ZUCKER, B.-A. (1993): Die Charakterisierung von Bordetella bronchiseptica, Haemophilus parasuis und Yersinia enterocolitica mittels Lipopolysaccharid- und Polypeptidmusternalyse in der SDS-PAGE. HU-Berlin, Diss.

Anhang 129
9 Anhang
In den Tabellen verwendete Abkürzungen: Absz. Abszess Anz. Anzahl Ausst. Ausstallung DG Durchgang Einst. Einstallung Futterverw. Futterverwertung g Gramm Gew. Gewicht H. Hälfte kg Kilogramm Mort. Mortalität PL Pleuritis PN Pneumonie Schlacht. Schlachtzeitpunkt tägl. Zun. tägliche Zunahmen Umst. Umstallung
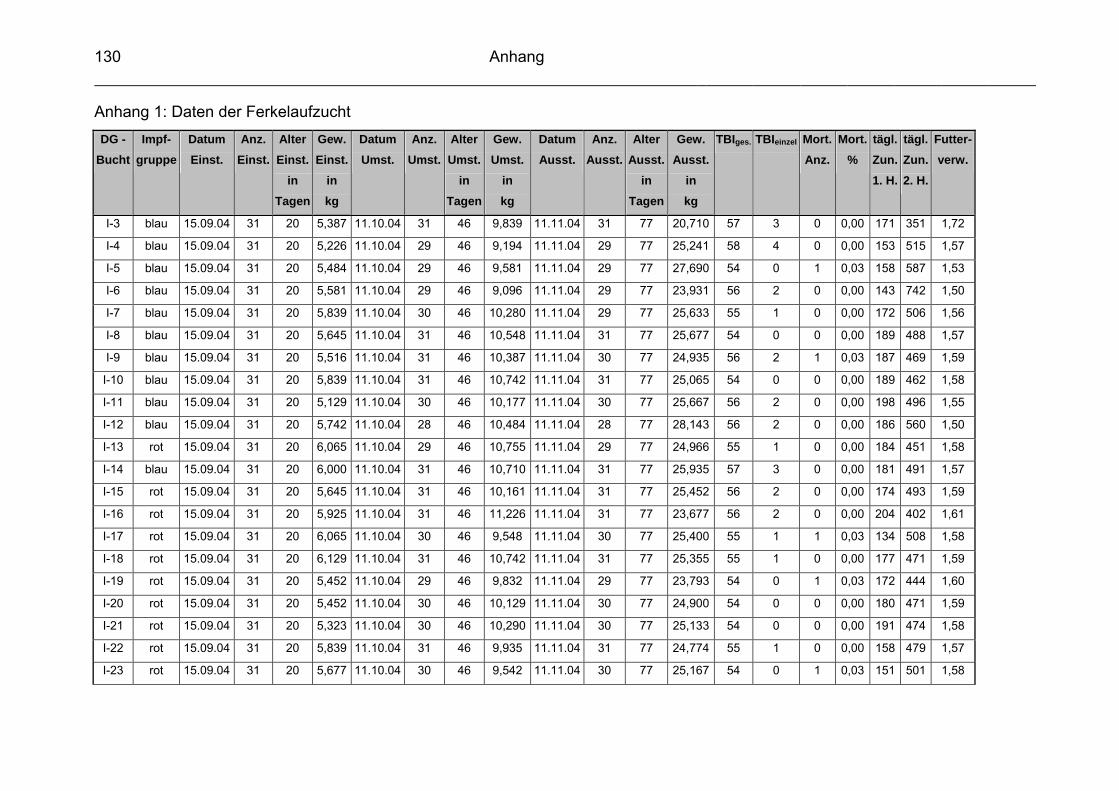
130 Anhang
Anhang 1: Daten der Ferkelaufzucht DG -
Bucht Impf-
gruppeDatumEinst.
Anz. Einst.
Alter Einst.
in Tagen
Gew. Einst.
in kg
DatumUmst.
Anz. Umst.
Alter Umst.
in Tagen
Gew. Umst.
in kg
DatumAusst.
Anz. Ausst.
Alter Ausst.
in Tagen
Gew.Ausst.
in kg
TBIges. TBIeinzel Mort.Anz.
Mort.%
tägl.Zun.1. H.
tägl.Zun.2. H.
Futter-verw.
I-3 blau 15.09.04 31 20 5,387 11.10.04 31 46 9,839 11.11.04 31 77 20,710 57 3 0 0,00 171 351 1,72
I-4 blau 15.09.04 31 20 5,226 11.10.04 29 46 9,194 11.11.04 29 77 25,241 58 4 0 0,00 153 515 1,57
I-5 blau 15.09.04 31 20 5,484 11.10.04 29 46 9,581 11.11.04 29 77 27,690 54 0 1 0,03 158 587 1,53
I-6 blau 15.09.04 31 20 5,581 11.10.04 29 46 9,096 11.11.04 29 77 23,931 56 2 0 0,00 143 742 1,50
I-7 blau 15.09.04 31 20 5,839 11.10.04 30 46 10,280 11.11.04 29 77 25,633 55 1 0 0,00 172 506 1,56
I-8 blau 15.09.04 31 20 5,645 11.10.04 31 46 10,548 11.11.04 31 77 25,677 54 0 0 0,00 189 488 1,57
I-9 blau 15.09.04 31 20 5,516 11.10.04 31 46 10,387 11.11.04 30 77 24,935 56 2 1 0,03 187 469 1,59
I-10 blau 15.09.04 31 20 5,839 11.10.04 31 46 10,742 11.11.04 31 77 25,065 54 0 0 0,00 189 462 1,58
I-11 blau 15.09.04 31 20 5,129 11.10.04 30 46 10,177 11.11.04 30 77 25,667 56 2 0 0,00 198 496 1,55
I-12 blau 15.09.04 31 20 5,742 11.10.04 28 46 10,484 11.11.04 28 77 28,143 56 2 0 0,00 186 560 1,50
I-13 rot 15.09.04 31 20 6,065 11.10.04 29 46 10,755 11.11.04 29 77 24,966 55 1 0 0,00 184 451 1,58
I-14 blau 15.09.04 31 20 6,000 11.10.04 31 46 10,710 11.11.04 31 77 25,935 57 3 0 0,00 181 491 1,57
I-15 rot 15.09.04 31 20 5,645 11.10.04 31 46 10,161 11.11.04 31 77 25,452 56 2 0 0,00 174 493 1,59
I-16 rot 15.09.04 31 20 5,925 11.10.04 31 46 11,226 11.11.04 31 77 23,677 56 2 0 0,00 204 402 1,61
I-17 rot 15.09.04 31 20 6,065 11.10.04 30 46 9,548 11.11.04 30 77 25,400 55 1 1 0,03 134 508 1,58
I-18 rot 15.09.04 31 20 6,129 11.10.04 31 46 10,742 11.11.04 31 77 25,355 55 1 0 0,00 177 471 1,59
I-19 rot 15.09.04 31 20 5,452 11.10.04 29 46 9,832 11.11.04 29 77 23,793 54 0 1 0,03 172 444 1,60
I-20 rot 15.09.04 31 20 5,452 11.10.04 30 46 10,129 11.11.04 30 77 24,900 54 0 0 0,00 180 471 1,59
I-21 rot 15.09.04 31 20 5,323 11.10.04 30 46 10,290 11.11.04 30 77 25,133 54 0 0 0,00 191 474 1,58
I-22 rot 15.09.04 31 20 5,839 11.10.04 31 46 9,935 11.11.04 31 77 24,774 55 1 0 0,00 158 479 1,57
I-23 rot 15.09.04 31 20 5,677 11.10.04 30 46 9,542 11.11.04 30 77 25,167 54 0 1 0,03 151 501 1,58
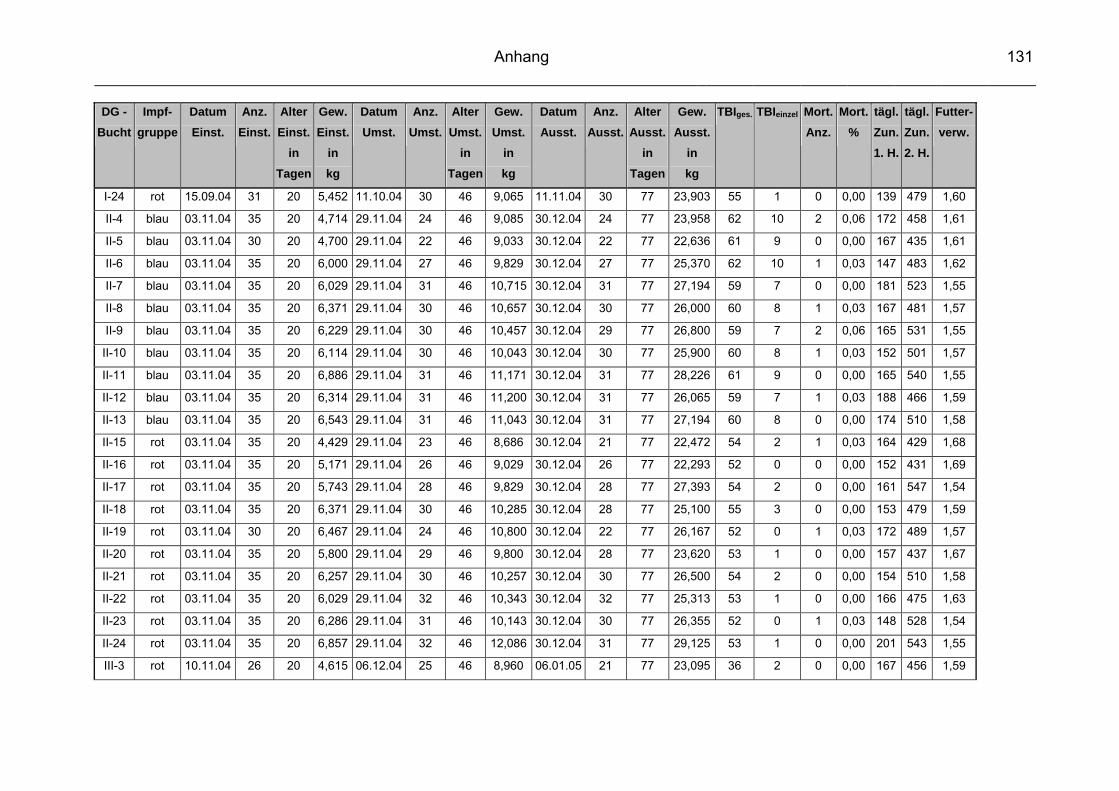
Anhang 131
DG -
Bucht Impf-
gruppeDatumEinst.
Anz. Einst.
Alter Einst.
in Tagen
Gew. Einst.
in kg
DatumUmst.
Anz. Umst.
Alter Umst.
in Tagen
Gew. Umst.
in kg
DatumAusst.
Anz. Ausst.
Alter Ausst.
in Tagen
Gew.Ausst.
in kg
TBIges. TBIeinzel Mort.Anz.
Mort.%
tägl.Zun.1. H.
tägl.Zun.2. H.
Futter-verw.
I-24 rot 15.09.04 31 20 5,452 11.10.04 30 46 9,065 11.11.04 30 77 23,903 55 1 0 0,00 139 479 1,60
II-4 blau 03.11.04 35 20 4,714 29.11.04 24 46 9,085 30.12.04 24 77 23,958 62 10 2 0,06 172 458 1,61
II-5 blau 03.11.04 30 20 4,700 29.11.04 22 46 9,033 30.12.04 22 77 22,636 61 9 0 0,00 167 435 1,61
II-6 blau 03.11.04 35 20 6,000 29.11.04 27 46 9,829 30.12.04 27 77 25,370 62 10 1 0,03 147 483 1,62
II-7 blau 03.11.04 35 20 6,029 29.11.04 31 46 10,715 30.12.04 31 77 27,194 59 7 0 0,00 181 523 1,55
II-8 blau 03.11.04 35 20 6,371 29.11.04 30 46 10,657 30.12.04 30 77 26,000 60 8 1 0,03 167 481 1,57
II-9 blau 03.11.04 35 20 6,229 29.11.04 30 46 10,457 30.12.04 29 77 26,800 59 7 2 0,06 165 531 1,55
II-10 blau 03.11.04 35 20 6,114 29.11.04 30 46 10,043 30.12.04 30 77 25,900 60 8 1 0,03 152 501 1,57
II-11 blau 03.11.04 35 20 6,886 29.11.04 31 46 11,171 30.12.04 31 77 28,226 61 9 0 0,00 165 540 1,55
II-12 blau 03.11.04 35 20 6,314 29.11.04 31 46 11,200 30.12.04 31 77 26,065 59 7 1 0,03 188 466 1,59
II-13 blau 03.11.04 35 20 6,543 29.11.04 31 46 11,043 30.12.04 31 77 27,194 60 8 0 0,00 174 510 1,58
II-15 rot 03.11.04 35 20 4,429 29.11.04 23 46 8,686 30.12.04 21 77 22,472 54 2 1 0,03 164 429 1,68
II-16 rot 03.11.04 35 20 5,171 29.11.04 26 46 9,029 30.12.04 26 77 22,293 52 0 0 0,00 152 431 1,69
II-17 rot 03.11.04 35 20 5,743 29.11.04 28 46 9,829 30.12.04 28 77 27,393 54 2 0 0,00 161 547 1,54
II-18 rot 03.11.04 35 20 6,371 29.11.04 30 46 10,285 30.12.04 28 77 25,100 55 3 0 0,00 153 479 1,59
II-19 rot 03.11.04 30 20 6,467 29.11.04 24 46 10,800 30.12.04 22 77 26,167 52 0 1 0,03 172 489 1,57
II-20 rot 03.11.04 35 20 5,800 29.11.04 29 46 9,800 30.12.04 28 77 23,620 53 1 0 0,00 157 437 1,67
II-21 rot 03.11.04 35 20 6,257 29.11.04 30 46 10,257 30.12.04 30 77 26,500 54 2 0 0,00 154 510 1,58
II-22 rot 03.11.04 35 20 6,029 29.11.04 32 46 10,343 30.12.04 32 77 25,313 53 1 0 0,00 166 475 1,63
II-23 rot 03.11.04 35 20 6,286 29.11.04 31 46 10,143 30.12.04 30 77 26,355 52 0 1 0,03 148 528 1,54
II-24 rot 03.11.04 35 20 6,857 29.11.04 32 46 12,086 30.12.04 31 77 29,125 53 1 0 0,00 201 543 1,55
III-3 rot 10.11.04 26 20 4,615 06.12.04 25 46 8,960 06.01.05 21 77 23,095 36 2 0 0,00 167 456 1,59
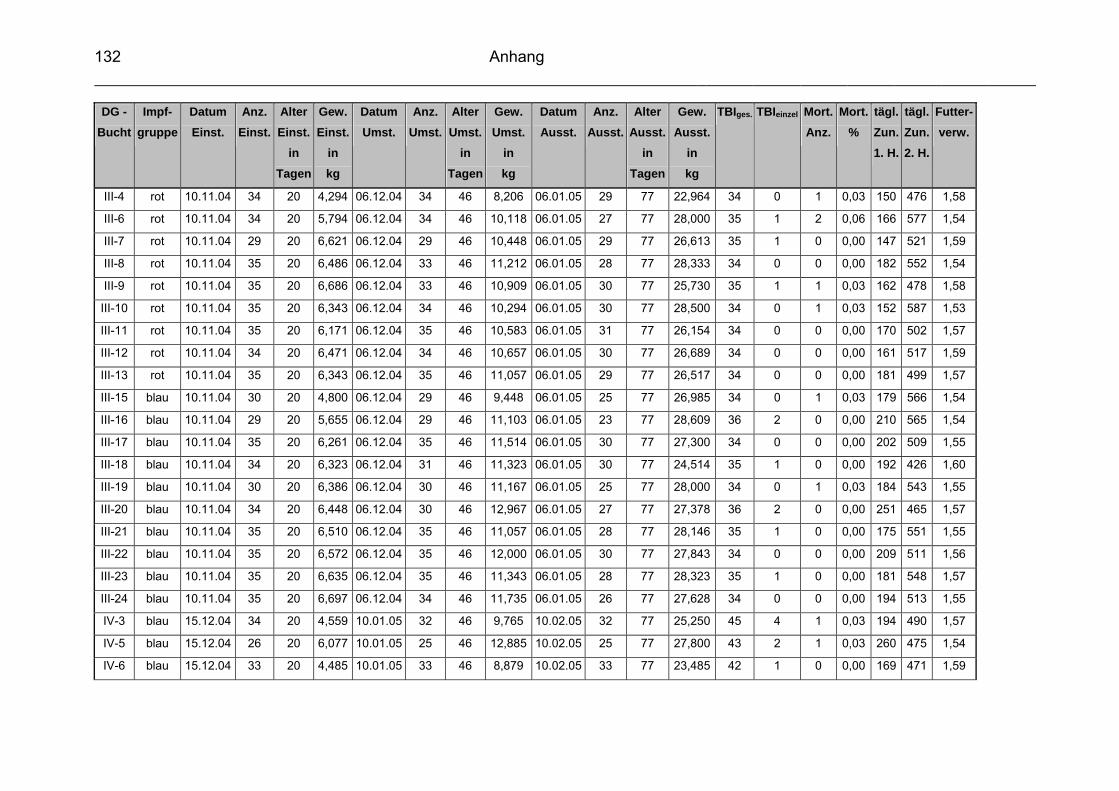
132 Anhang
DG -
Bucht Impf-
gruppeDatumEinst.
Anz. Einst.
Alter Einst.
in Tagen
Gew. Einst.
in kg
DatumUmst.
Anz. Umst.
Alter Umst.
in Tagen
Gew. Umst.
in kg
DatumAusst.
Anz. Ausst.
Alter Ausst.
in Tagen
Gew.Ausst.
in kg
TBIges. TBIeinzel Mort.Anz.
Mort.%
tägl.Zun.1. H.
tägl.Zun.2. H.
Futter-verw.
III-4 rot 10.11.04 34 20 4,294 06.12.04 34 46 8,206 06.01.05 29 77 22,964 34 0 1 0,03 150 476 1,58
III-6 rot 10.11.04 34 20 5,794 06.12.04 34 46 10,118 06.01.05 27 77 28,000 35 1 2 0,06 166 577 1,54
III-7 rot 10.11.04 29 20 6,621 06.12.04 29 46 10,448 06.01.05 29 77 26,613 35 1 0 0,00 147 521 1,59
III-8 rot 10.11.04 35 20 6,486 06.12.04 33 46 11,212 06.01.05 28 77 28,333 34 0 0 0,00 182 552 1,54
III-9 rot 10.11.04 35 20 6,686 06.12.04 33 46 10,909 06.01.05 30 77 25,730 35 1 1 0,03 162 478 1,58
III-10 rot 10.11.04 35 20 6,343 06.12.04 34 46 10,294 06.01.05 30 77 28,500 34 0 1 0,03 152 587 1,53
III-11 rot 10.11.04 35 20 6,171 06.12.04 35 46 10,583 06.01.05 31 77 26,154 34 0 0 0,00 170 502 1,57
III-12 rot 10.11.04 34 20 6,471 06.12.04 34 46 10,657 06.01.05 30 77 26,689 34 0 0 0,00 161 517 1,59
III-13 rot 10.11.04 35 20 6,343 06.12.04 35 46 11,057 06.01.05 29 77 26,517 34 0 0 0,00 181 499 1,57
III-15 blau 10.11.04 30 20 4,800 06.12.04 29 46 9,448 06.01.05 25 77 26,985 34 0 1 0,03 179 566 1,54
III-16 blau 10.11.04 29 20 5,655 06.12.04 29 46 11,103 06.01.05 23 77 28,609 36 2 0 0,00 210 565 1,54
III-17 blau 10.11.04 35 20 6,261 06.12.04 35 46 11,514 06.01.05 30 77 27,300 34 0 0 0,00 202 509 1,55
III-18 blau 10.11.04 34 20 6,323 06.12.04 31 46 11,323 06.01.05 30 77 24,514 35 1 0 0,00 192 426 1,60
III-19 blau 10.11.04 30 20 6,386 06.12.04 30 46 11,167 06.01.05 25 77 28,000 34 0 1 0,03 184 543 1,55
III-20 blau 10.11.04 34 20 6,448 06.12.04 30 46 12,967 06.01.05 27 77 27,378 36 2 0 0,00 251 465 1,57
III-21 blau 10.11.04 35 20 6,510 06.12.04 35 46 11,057 06.01.05 28 77 28,146 35 1 0 0,00 175 551 1,55
III-22 blau 10.11.04 35 20 6,572 06.12.04 35 46 12,000 06.01.05 30 77 27,843 34 0 0 0,00 209 511 1,56
III-23 blau 10.11.04 35 20 6,635 06.12.04 35 46 11,343 06.01.05 28 77 28,323 35 1 0 0,00 181 548 1,57
III-24 blau 10.11.04 35 20 6,697 06.12.04 34 46 11,735 06.01.05 26 77 27,628 34 0 0 0,00 194 513 1,55
IV-3 blau 15.12.04 34 20 4,559 10.01.05 32 46 9,765 10.02.05 32 77 25,250 45 4 1 0,03 194 490 1,57
IV-5 blau 15.12.04 26 20 6,077 10.01.05 25 46 12,885 10.02.05 25 77 27,800 43 2 1 0,03 260 475 1,54
IV-6 blau 15.12.04 33 20 4,485 10.01.05 33 46 8,879 10.02.05 33 77 23,485 42 1 0 0,00 169 471 1,59
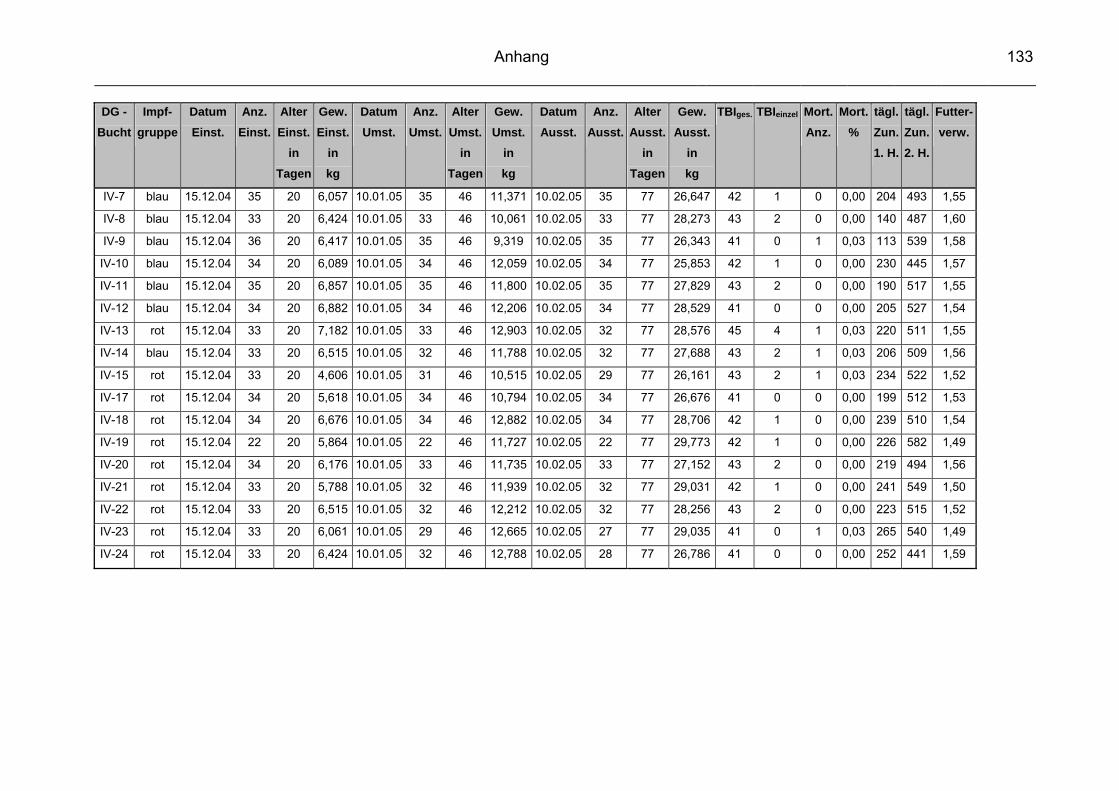
Anhang 133
DG -
Bucht Impf-
gruppeDatumEinst.
Anz. Einst.
Alter Einst.
in Tagen
Gew. Einst.
in kg
DatumUmst.
Anz. Umst.
Alter Umst.
in Tagen
Gew. Umst.
in kg
DatumAusst.
Anz. Ausst.
Alter Ausst.
in Tagen
Gew.Ausst.
in kg
TBIges. TBIeinzel Mort.Anz.
Mort.%
tägl.Zun.1. H.
tägl.Zun.2. H.
Futter-verw.
IV-7 blau 15.12.04 35 20 6,057 10.01.05 35 46 11,371 10.02.05 35 77 26,647 42 1 0 0,00 204 493 1,55
IV-8 blau 15.12.04 33 20 6,424 10.01.05 33 46 10,061 10.02.05 33 77 28,273 43 2 0 0,00 140 487 1,60
IV-9 blau 15.12.04 36 20 6,417 10.01.05 35 46 9,319 10.02.05 35 77 26,343 41 0 1 0,03 113 539 1,58
IV-10 blau 15.12.04 34 20 6,089 10.01.05 34 46 12,059 10.02.05 34 77 25,853 42 1 0 0,00 230 445 1,57
IV-11 blau 15.12.04 35 20 6,857 10.01.05 35 46 11,800 10.02.05 35 77 27,829 43 2 0 0,00 190 517 1,55
IV-12 blau 15.12.04 34 20 6,882 10.01.05 34 46 12,206 10.02.05 34 77 28,529 41 0 0 0,00 205 527 1,54
IV-13 rot 15.12.04 33 20 7,182 10.01.05 33 46 12,903 10.02.05 32 77 28,576 45 4 1 0,03 220 511 1,55
IV-14 blau 15.12.04 33 20 6,515 10.01.05 32 46 11,788 10.02.05 32 77 27,688 43 2 1 0,03 206 509 1,56
IV-15 rot 15.12.04 33 20 4,606 10.01.05 31 46 10,515 10.02.05 29 77 26,161 43 2 1 0,03 234 522 1,52
IV-17 rot 15.12.04 34 20 5,618 10.01.05 34 46 10,794 10.02.05 34 77 26,676 41 0 0 0,00 199 512 1,53
IV-18 rot 15.12.04 34 20 6,676 10.01.05 34 46 12,882 10.02.05 34 77 28,706 42 1 0 0,00 239 510 1,54
IV-19 rot 15.12.04 22 20 5,864 10.01.05 22 46 11,727 10.02.05 22 77 29,773 42 1 0 0,00 226 582 1,49
IV-20 rot 15.12.04 34 20 6,176 10.01.05 33 46 11,735 10.02.05 33 77 27,152 43 2 0 0,00 219 494 1,56
IV-21 rot 15.12.04 33 20 5,788 10.01.05 32 46 11,939 10.02.05 32 77 29,031 42 1 0 0,00 241 549 1,50
IV-22 rot 15.12.04 33 20 6,515 10.01.05 32 46 12,212 10.02.05 32 77 28,256 43 2 0 0,00 223 515 1,52
IV-23 rot 15.12.04 33 20 6,061 10.01.05 29 46 12,665 10.02.05 27 77 29,035 41 0 1 0,03 265 540 1,49
IV-24 rot 15.12.04 33 20 6,424 10.01.05 32 46 12,788 10.02.05 28 77 26,786 41 0 0 0,00 252 441 1,59
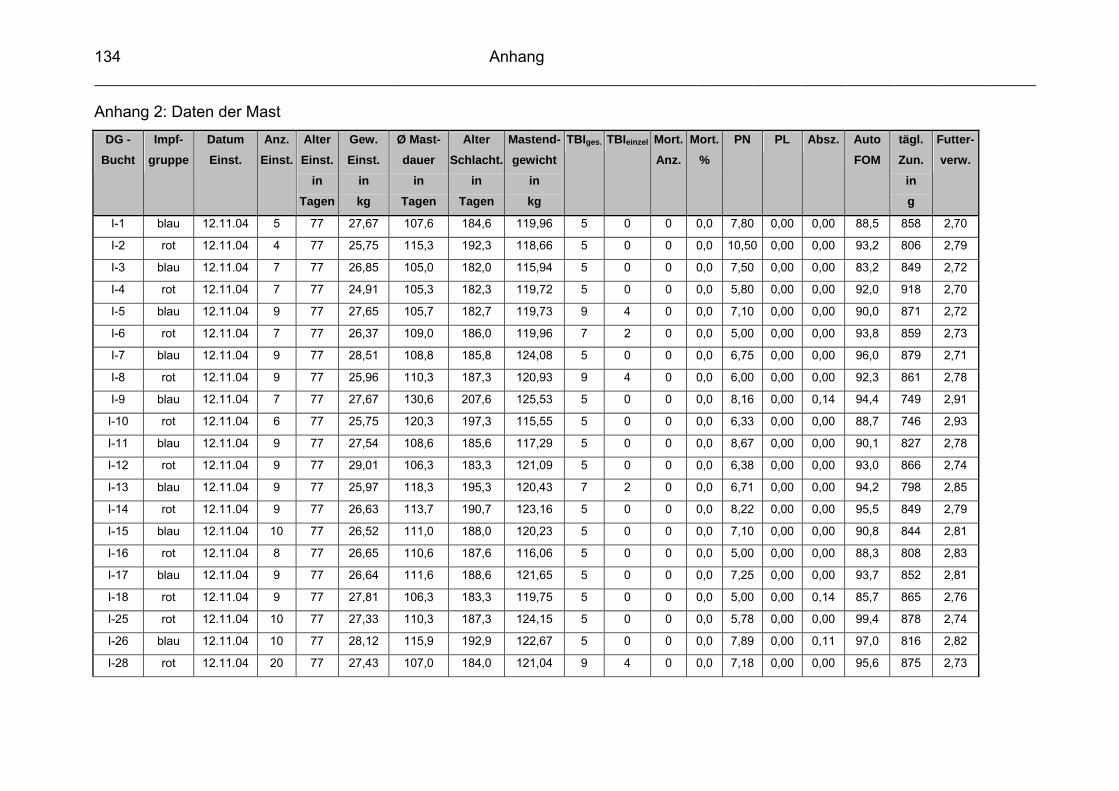
134 Anhang
Anhang 2: Daten der Mast DG -
Bucht Impf-
gruppeDatum Einst.
Anz. Einst.
AlterEinst.
in Tagen
Gew.Einst.
in kg
Ø Mast-dauer
in Tagen
Alter Schlacht.
in Tagen
Mastend-gewicht
in kg
TBIges. TBIeinzel Mort.Anz.
Mort.%
PN PL Absz. AutoFOM
tägl.Zun.
in g
Futter-verw.
I-1 blau 12.11.04 5 77 27,67 107,6 184,6 119,96 5 0 0 0,0 7,80 0,00 0,00 88,5 858 2,70
I-2 rot 12.11.04 4 77 25,75 115,3 192,3 118,66 5 0 0 0,0 10,50 0,00 0,00 93,2 806 2,79
I-3 blau 12.11.04 7 77 26,85 105,0 182,0 115,94 5 0 0 0,0 7,50 0,00 0,00 83,2 849 2,72
I-4 rot 12.11.04 7 77 24,91 105,3 182,3 119,72 5 0 0 0,0 5,80 0,00 0,00 92,0 918 2,70
I-5 blau 12.11.04 9 77 27,65 105,7 182,7 119,73 9 4 0 0,0 7,10 0,00 0,00 90,0 871 2,72
I-6 rot 12.11.04 7 77 26,37 109,0 186,0 119,96 7 2 0 0,0 5,00 0,00 0,00 93,8 859 2,73
I-7 blau 12.11.04 9 77 28,51 108,8 185,8 124,08 5 0 0 0,0 6,75 0,00 0,00 96,0 879 2,71
I-8 rot 12.11.04 9 77 25,96 110,3 187,3 120,93 9 4 0 0,0 6,00 0,00 0,00 92,3 861 2,78
I-9 blau 12.11.04 7 77 27,67 130,6 207,6 125,53 5 0 0 0,0 8,16 0,00 0,14 94,4 749 2,91
I-10 rot 12.11.04 6 77 25,75 120,3 197,3 115,55 5 0 0 0,0 6,33 0,00 0,00 88,7 746 2,93
I-11 blau 12.11.04 9 77 27,54 108,6 185,6 117,29 5 0 0 0,0 8,67 0,00 0,00 90,1 827 2,78
I-12 rot 12.11.04 9 77 29,01 106,3 183,3 121,09 5 0 0 0,0 6,38 0,00 0,00 93,0 866 2,74
I-13 blau 12.11.04 9 77 25,97 118,3 195,3 120,43 7 2 0 0,0 6,71 0,00 0,00 94,2 798 2,85
I-14 rot 12.11.04 9 77 26,63 113,7 190,7 123,16 5 0 0 0,0 8,22 0,00 0,00 95,5 849 2,79
I-15 blau 12.11.04 10 77 26,52 111,0 188,0 120,23 5 0 0 0,0 7,10 0,00 0,00 90,8 844 2,81
I-16 rot 12.11.04 8 77 26,65 110,6 187,6 116,06 5 0 0 0,0 5,00 0,00 0,00 88,3 808 2,83
I-17 blau 12.11.04 9 77 26,64 111,6 188,6 121,65 5 0 0 0,0 7,25 0,00 0,00 93,7 852 2,81
I-18 rot 12.11.04 9 77 27,81 106,3 183,3 119,75 5 0 0 0,0 5,00 0,00 0,14 85,7 865 2,76
I-25 rot 12.11.04 10 77 27,33 110,3 187,3 124,15 5 0 0 0,0 5,78 0,00 0,00 99,4 878 2,74
I-26 blau 12.11.04 10 77 28,12 115,9 192,9 122,67 5 0 0 0,0 7,89 0,00 0,11 97,0 816 2,82
I-28 rot 12.11.04 20 77 27,43 107,0 184,0 121,04 9 4 0 0,0 7,18 0,00 0,00 95,6 875 2,73
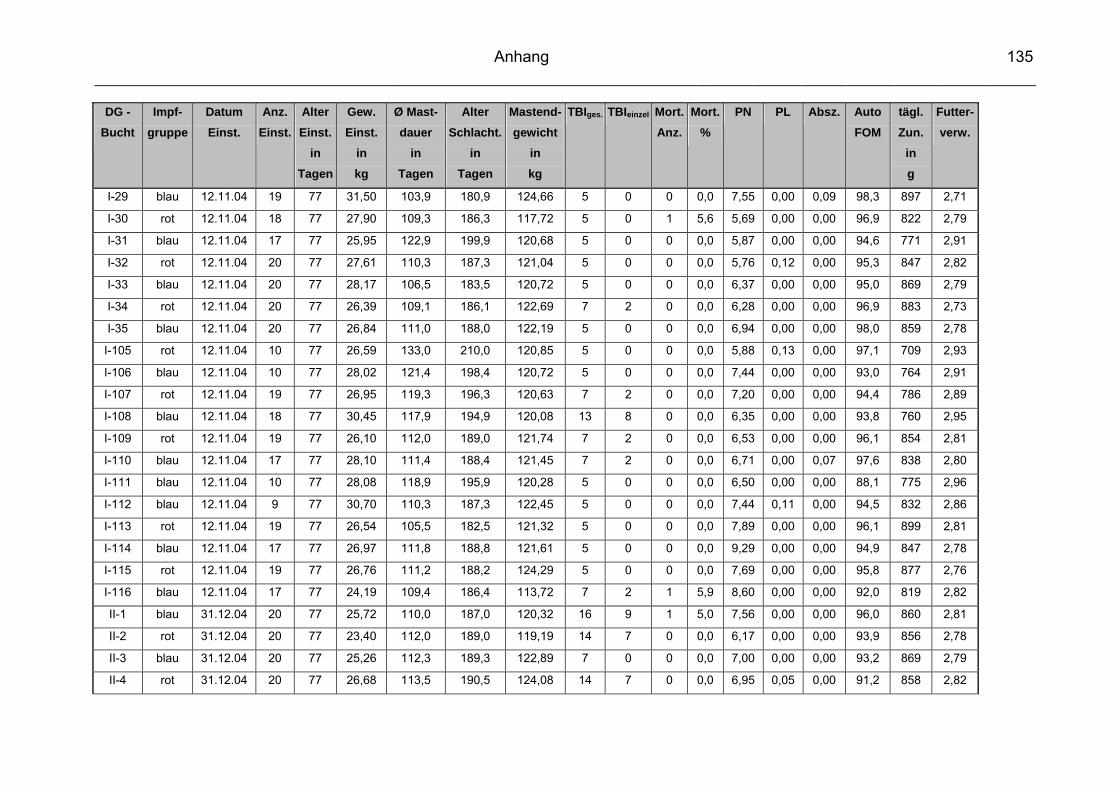
Anhang 135
DG -
Bucht Impf-
gruppeDatum Einst.
Anz. Einst.
AlterEinst.
in Tagen
Gew.Einst.
in kg
Ø Mast-dauer
in Tagen
Alter Schlacht.
in Tagen
Mastend-gewicht
in kg
TBIges. TBIeinzel Mort.Anz.
Mort.%
PN PL Absz. AutoFOM
tägl.Zun.
in g
Futter-verw.
I-29 blau 12.11.04 19 77 31,50 103,9 180,9 124,66 5 0 0 0,0 7,55 0,00 0,09 98,3 897 2,71
I-30 rot 12.11.04 18 77 27,90 109,3 186,3 117,72 5 0 1 5,6 5,69 0,00 0,00 96,9 822 2,79
I-31 blau 12.11.04 17 77 25,95 122,9 199,9 120,68 5 0 0 0,0 5,87 0,00 0,00 94,6 771 2,91
I-32 rot 12.11.04 20 77 27,61 110,3 187,3 121,04 5 0 0 0,0 5,76 0,12 0,00 95,3 847 2,82
I-33 blau 12.11.04 20 77 28,17 106,5 183,5 120,72 5 0 0 0,0 6,37 0,00 0,00 95,0 869 2,79
I-34 rot 12.11.04 20 77 26,39 109,1 186,1 122,69 7 2 0 0,0 6,28 0,00 0,00 96,9 883 2,73
I-35 blau 12.11.04 20 77 26,84 111,0 188,0 122,19 5 0 0 0,0 6,94 0,00 0,00 98,0 859 2,78
I-105 rot 12.11.04 10 77 26,59 133,0 210,0 120,85 5 0 0 0,0 5,88 0,13 0,00 97,1 709 2,93
I-106 blau 12.11.04 10 77 28,02 121,4 198,4 120,72 5 0 0 0,0 7,44 0,00 0,00 93,0 764 2,91
I-107 rot 12.11.04 19 77 26,95 119,3 196,3 120,63 7 2 0 0,0 7,20 0,00 0,00 94,4 786 2,89
I-108 blau 12.11.04 18 77 30,45 117,9 194,9 120,08 13 8 0 0,0 6,35 0,00 0,00 93,8 760 2,95
I-109 rot 12.11.04 19 77 26,10 112,0 189,0 121,74 7 2 0 0,0 6,53 0,00 0,00 96,1 854 2,81
I-110 blau 12.11.04 17 77 28,10 111,4 188,4 121,45 7 2 0 0,0 6,71 0,00 0,07 97,6 838 2,80
I-111 blau 12.11.04 10 77 28,08 118,9 195,9 120,28 5 0 0 0,0 6,50 0,00 0,00 88,1 775 2,96
I-112 blau 12.11.04 9 77 30,70 110,3 187,3 122,45 5 0 0 0,0 7,44 0,11 0,00 94,5 832 2,86
I-113 rot 12.11.04 19 77 26,54 105,5 182,5 121,32 5 0 0 0,0 7,89 0,00 0,00 96,1 899 2,81
I-114 blau 12.11.04 17 77 26,97 111,8 188,8 121,61 5 0 0 0,0 9,29 0,00 0,00 94,9 847 2,78
I-115 rot 12.11.04 19 77 26,76 111,2 188,2 124,29 5 0 0 0,0 7,69 0,00 0,00 95,8 877 2,76
I-116 blau 12.11.04 17 77 24,19 109,4 186,4 113,72 7 2 1 5,9 8,60 0,00 0,00 92,0 819 2,82
II-1 blau 31.12.04 20 77 25,72 110,0 187,0 120,32 16 9 1 5,0 7,56 0,00 0,00 96,0 860 2,81
II-2 rot 31.12.04 20 77 23,40 112,0 189,0 119,19 14 7 0 0,0 6,17 0,00 0,00 93,9 856 2,78
II-3 blau 31.12.04 20 77 25,26 112,3 189,3 122,89 7 0 0 0,0 7,00 0,00 0,00 93,2 869 2,79
II-4 rot 31.12.04 20 77 26,68 113,5 190,5 124,08 14 7 0 0,0 6,95 0,05 0,00 91,2 858 2,82
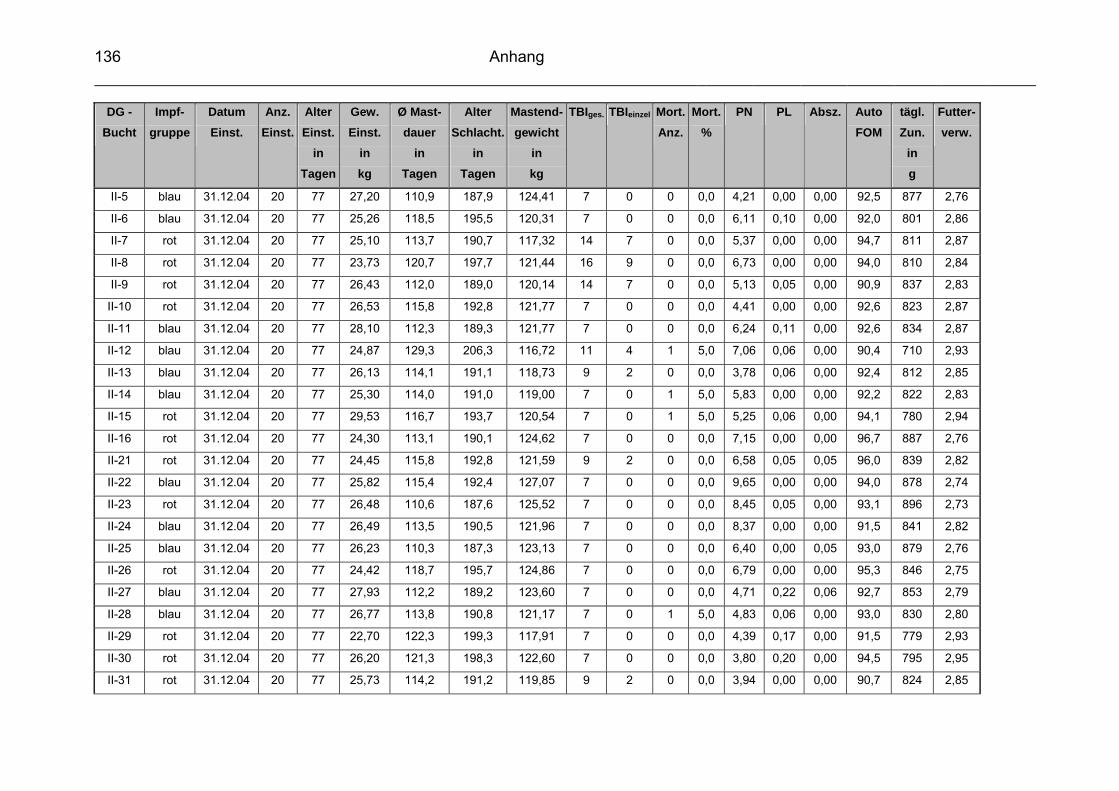
136 Anhang
DG -
Bucht Impf-
gruppeDatum Einst.
Anz. Einst.
AlterEinst.
in Tagen
Gew.Einst.
in kg
Ø Mast-dauer
in Tagen
Alter Schlacht.
in Tagen
Mastend-gewicht
in kg
TBIges. TBIeinzel Mort.Anz.
Mort.%
PN PL Absz. AutoFOM
tägl.Zun.
in g
Futter-verw.
II-5 blau 31.12.04 20 77 27,20 110,9 187,9 124,41 7 0 0 0,0 4,21 0,00 0,00 92,5 877 2,76
II-6 blau 31.12.04 20 77 25,26 118,5 195,5 120,31 7 0 0 0,0 6,11 0,10 0,00 92,0 801 2,86
II-7 rot 31.12.04 20 77 25,10 113,7 190,7 117,32 14 7 0 0,0 5,37 0,00 0,00 94,7 811 2,87
II-8 rot 31.12.04 20 77 23,73 120,7 197,7 121,44 16 9 0 0,0 6,73 0,00 0,00 94,0 810 2,84
II-9 rot 31.12.04 20 77 26,43 112,0 189,0 120,14 14 7 0 0,0 5,13 0,05 0,00 90,9 837 2,83
II-10 rot 31.12.04 20 77 26,53 115,8 192,8 121,77 7 0 0 0,0 4,41 0,00 0,00 92,6 823 2,87
II-11 blau 31.12.04 20 77 28,10 112,3 189,3 121,77 7 0 0 0,0 6,24 0,11 0,00 92,6 834 2,87
II-12 blau 31.12.04 20 77 24,87 129,3 206,3 116,72 11 4 1 5,0 7,06 0,06 0,00 90,4 710 2,93
II-13 blau 31.12.04 20 77 26,13 114,1 191,1 118,73 9 2 0 0,0 3,78 0,06 0,00 92,4 812 2,85
II-14 blau 31.12.04 20 77 25,30 114,0 191,0 119,00 7 0 1 5,0 5,83 0,00 0,00 92,2 822 2,83
II-15 rot 31.12.04 20 77 29,53 116,7 193,7 120,54 7 0 1 5,0 5,25 0,06 0,00 94,1 780 2,94
II-16 rot 31.12.04 20 77 24,30 113,1 190,1 124,62 7 0 0 0,0 7,15 0,00 0,00 96,7 887 2,76
II-21 rot 31.12.04 20 77 24,45 115,8 192,8 121,59 9 2 0 0,0 6,58 0,05 0,05 96,0 839 2,82
II-22 blau 31.12.04 20 77 25,82 115,4 192,4 127,07 7 0 0 0,0 9,65 0,00 0,00 94,0 878 2,74
II-23 rot 31.12.04 20 77 26,48 110,6 187,6 125,52 7 0 0 0,0 8,45 0,05 0,00 93,1 896 2,73
II-24 blau 31.12.04 20 77 26,49 113,5 190,5 121,96 7 0 0 0,0 8,37 0,00 0,00 91,5 841 2,82
II-25 blau 31.12.04 20 77 26,23 110,3 187,3 123,13 7 0 0 0,0 6,40 0,00 0,05 93,0 879 2,76
II-26 rot 31.12.04 20 77 24,42 118,7 195,7 124,86 7 0 0 0,0 6,79 0,00 0,00 95,3 846 2,75
II-27 blau 31.12.04 20 77 27,93 112,2 189,2 123,60 7 0 0 0,0 4,71 0,22 0,06 92,7 853 2,79
II-28 blau 31.12.04 20 77 26,77 113,8 190,8 121,17 7 0 1 5,0 4,83 0,06 0,00 93,0 830 2,80
II-29 rot 31.12.04 20 77 22,70 122,3 199,3 117,91 7 0 0 0,0 4,39 0,17 0,00 91,5 779 2,93
II-30 rot 31.12.04 20 77 26,20 121,3 198,3 122,60 7 0 0 0,0 3,80 0,20 0,00 94,5 795 2,95
II-31 rot 31.12.04 20 77 25,73 114,2 191,2 119,85 9 2 0 0,0 3,94 0,00 0,00 90,7 824 2,85
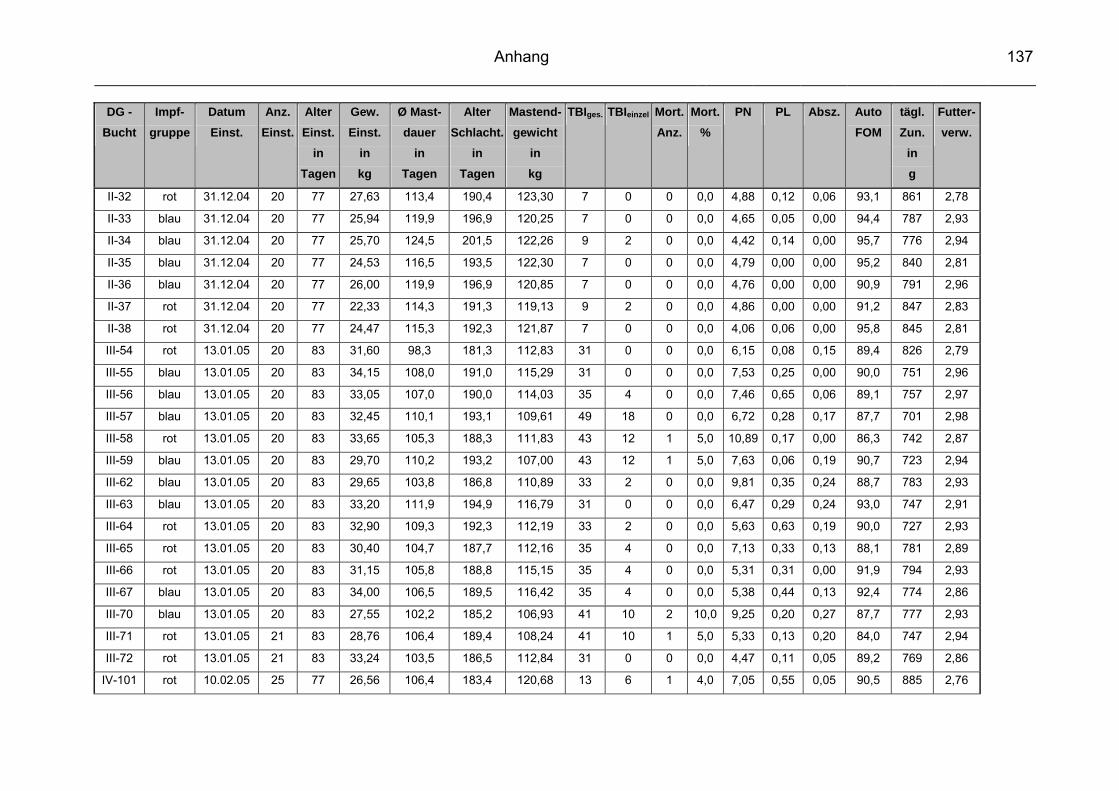
Anhang 137
DG -
Bucht Impf-
gruppeDatum Einst.
Anz. Einst.
AlterEinst.
in Tagen
Gew.Einst.
in kg
Ø Mast-dauer
in Tagen
Alter Schlacht.
in Tagen
Mastend-gewicht
in kg
TBIges. TBIeinzel Mort.Anz.
Mort.%
PN PL Absz. AutoFOM
tägl.Zun.
in g
Futter-verw.
II-32 rot 31.12.04 20 77 27,63 113,4 190,4 123,30 7 0 0 0,0 4,88 0,12 0,06 93,1 861 2,78
II-33 blau 31.12.04 20 77 25,94 119,9 196,9 120,25 7 0 0 0,0 4,65 0,05 0,00 94,4 787 2,93
II-34 blau 31.12.04 20 77 25,70 124,5 201,5 122,26 9 2 0 0,0 4,42 0,14 0,00 95,7 776 2,94
II-35 blau 31.12.04 20 77 24,53 116,5 193,5 122,30 7 0 0 0,0 4,79 0,00 0,00 95,2 840 2,81
II-36 blau 31.12.04 20 77 26,00 119,9 196,9 120,85 7 0 0 0,0 4,76 0,00 0,00 90,9 791 2,96
II-37 rot 31.12.04 20 77 22,33 114,3 191,3 119,13 9 2 0 0,0 4,86 0,00 0,00 91,2 847 2,83
II-38 rot 31.12.04 20 77 24,47 115,3 192,3 121,87 7 0 0 0,0 4,06 0,06 0,00 95,8 845 2,81
III-54 rot 13.01.05 20 83 31,60 98,3 181,3 112,83 31 0 0 0,0 6,15 0,08 0,15 89,4 826 2,79
III-55 blau 13.01.05 20 83 34,15 108,0 191,0 115,29 31 0 0 0,0 7,53 0,25 0,00 90,0 751 2,96
III-56 blau 13.01.05 20 83 33,05 107,0 190,0 114,03 35 4 0 0,0 7,46 0,65 0,06 89,1 757 2,97
III-57 blau 13.01.05 20 83 32,45 110,1 193,1 109,61 49 18 0 0,0 6,72 0,28 0,17 87,7 701 2,98
III-58 rot 13.01.05 20 83 33,65 105,3 188,3 111,83 43 12 1 5,0 10,89 0,17 0,00 86,3 742 2,87
III-59 blau 13.01.05 20 83 29,70 110,2 193,2 107,00 43 12 1 5,0 7,63 0,06 0,19 90,7 723 2,94
III-62 blau 13.01.05 20 83 29,65 103,8 186,8 110,89 33 2 0 0,0 9,81 0,35 0,24 88,7 783 2,93
III-63 blau 13.01.05 20 83 33,20 111,9 194,9 116,79 31 0 0 0,0 6,47 0,29 0,24 93,0 747 2,91
III-64 rot 13.01.05 20 83 32,90 109,3 192,3 112,19 33 2 0 0,0 5,63 0,63 0,19 90,0 727 2,93
III-65 rot 13.01.05 20 83 30,40 104,7 187,7 112,16 35 4 0 0,0 7,13 0,33 0,13 88,1 781 2,89
III-66 rot 13.01.05 20 83 31,15 105,8 188,8 115,15 35 4 0 0,0 5,31 0,31 0,00 91,9 794 2,93
III-67 blau 13.01.05 20 83 34,00 106,5 189,5 116,42 35 4 0 0,0 5,38 0,44 0,13 92,4 774 2,86
III-70 blau 13.01.05 20 83 27,55 102,2 185,2 106,93 41 10 2 10,0 9,25 0,20 0,27 87,7 777 2,93
III-71 rot 13.01.05 21 83 28,76 106,4 189,4 108,24 41 10 1 5,0 5,33 0,13 0,20 84,0 747 2,94
III-72 rot 13.01.05 21 83 33,24 103,5 186,5 112,84 31 0 0 0,0 4,47 0,11 0,05 89,2 769 2,86
IV-101 rot 10.02.05 25 77 26,56 106,4 183,4 120,68 13 6 1 4,0 7,05 0,55 0,05 90,5 885 2,76
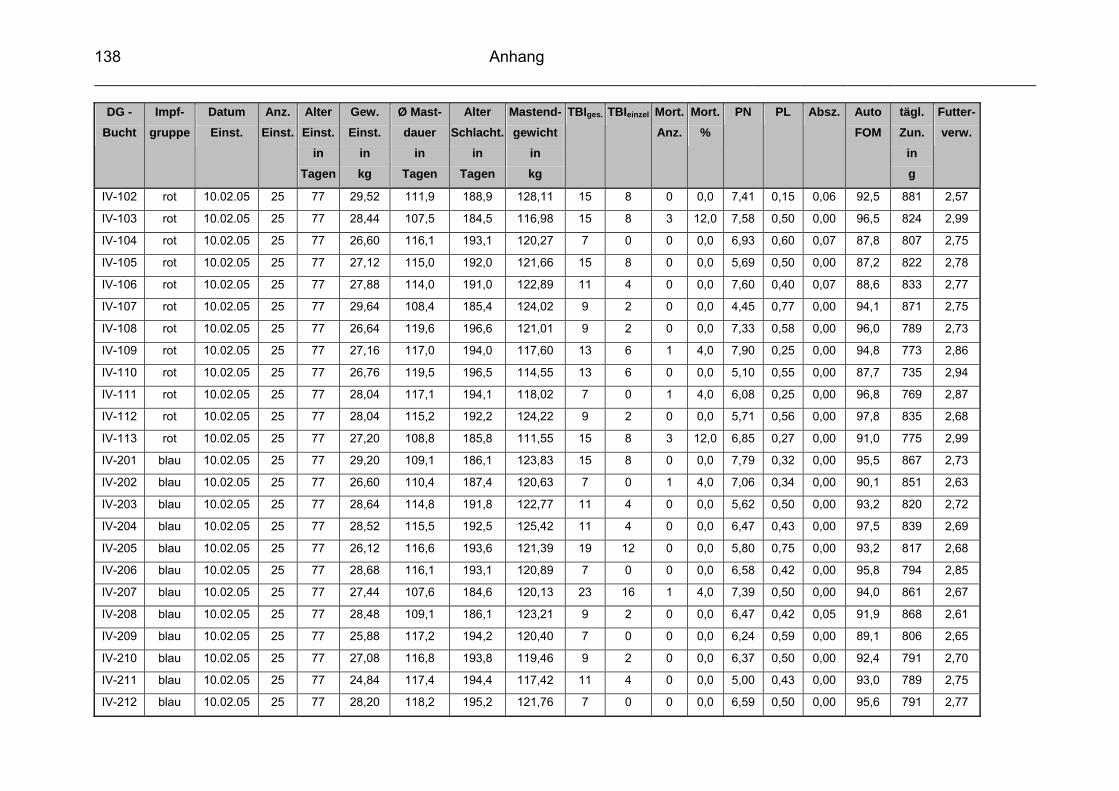
138 Anhang
DG -
Bucht Impf-
gruppeDatum Einst.
Anz. Einst.
AlterEinst.
in Tagen
Gew.Einst.
in kg
Ø Mast-dauer
in Tagen
Alter Schlacht.
in Tagen
Mastend-gewicht
in kg
TBIges. TBIeinzel Mort.Anz.
Mort.%
PN PL Absz. AutoFOM
tägl.Zun.
in g
Futter-verw.
IV-102 rot 10.02.05 25 77 29,52 111,9 188,9 128,11 15 8 0 0,0 7,41 0,15 0,06 92,5 881 2,57
IV-103 rot 10.02.05 25 77 28,44 107,5 184,5 116,98 15 8 3 12,0 7,58 0,50 0,00 96,5 824 2,99
IV-104 rot 10.02.05 25 77 26,60 116,1 193,1 120,27 7 0 0 0,0 6,93 0,60 0,07 87,8 807 2,75
IV-105 rot 10.02.05 25 77 27,12 115,0 192,0 121,66 15 8 0 0,0 5,69 0,50 0,00 87,2 822 2,78
IV-106 rot 10.02.05 25 77 27,88 114,0 191,0 122,89 11 4 0 0,0 7,60 0,40 0,07 88,6 833 2,77
IV-107 rot 10.02.05 25 77 29,64 108,4 185,4 124,02 9 2 0 0,0 4,45 0,77 0,00 94,1 871 2,75
IV-108 rot 10.02.05 25 77 26,64 119,6 196,6 121,01 9 2 0 0,0 7,33 0,58 0,00 96,0 789 2,73
IV-109 rot 10.02.05 25 77 27,16 117,0 194,0 117,60 13 6 1 4,0 7,90 0,25 0,00 94,8 773 2,86
IV-110 rot 10.02.05 25 77 26,76 119,5 196,5 114,55 13 6 0 0,0 5,10 0,55 0,00 87,7 735 2,94
IV-111 rot 10.02.05 25 77 28,04 117,1 194,1 118,02 7 0 1 4,0 6,08 0,25 0,00 96,8 769 2,87
IV-112 rot 10.02.05 25 77 28,04 115,2 192,2 124,22 9 2 0 0,0 5,71 0,56 0,00 97,8 835 2,68
IV-113 rot 10.02.05 25 77 27,20 108,8 185,8 111,55 15 8 3 12,0 6,85 0,27 0,00 91,0 775 2,99
IV-201 blau 10.02.05 25 77 29,20 109,1 186,1 123,83 15 8 0 0,0 7,79 0,32 0,00 95,5 867 2,73
IV-202 blau 10.02.05 25 77 26,60 110,4 187,4 120,63 7 0 1 4,0 7,06 0,34 0,00 90,1 851 2,63
IV-203 blau 10.02.05 25 77 28,64 114,8 191,8 122,77 11 4 0 0,0 5,62 0,50 0,00 93,2 820 2,72
IV-204 blau 10.02.05 25 77 28,52 115,5 192,5 125,42 11 4 0 0,0 6,47 0,43 0,00 97,5 839 2,69
IV-205 blau 10.02.05 25 77 26,12 116,6 193,6 121,39 19 12 0 0,0 5,80 0,75 0,00 93,2 817 2,68
IV-206 blau 10.02.05 25 77 28,68 116,1 193,1 120,89 7 0 0 0,0 6,58 0,42 0,00 95,8 794 2,85
IV-207 blau 10.02.05 25 77 27,44 107,6 184,6 120,13 23 16 1 4,0 7,39 0,50 0,00 94,0 861 2,67
IV-208 blau 10.02.05 25 77 28,48 109,1 186,1 123,21 9 2 0 0,0 6,47 0,42 0,05 91,9 868 2,61
IV-209 blau 10.02.05 25 77 25,88 117,2 194,2 120,40 7 0 0 0,0 6,24 0,59 0,00 89,1 806 2,65
IV-210 blau 10.02.05 25 77 27,08 116,8 193,8 119,46 9 2 0 0,0 6,37 0,50 0,00 92,4 791 2,70
IV-211 blau 10.02.05 25 77 24,84 117,4 194,4 117,42 11 4 0 0,0 5,00 0,43 0,00 93,0 789 2,75
IV-212 blau 10.02.05 25 77 28,20 118,2 195,2 121,76 7 0 0 0,0 6,59 0,50 0,00 95,6 791 2,77

Danksagung
Mein herzlicher Dank gilt Herrn Prof. Dr. Th. Blaha für die Überlassung des Themas
und die jederzeit gewährte unkomplizierte Unterstützung bei der Anfertigung der
Dissertation.
Bei den Mitarbeitern der Praxis WEK, insbesondere bei A. Wilms-Schulze Kump und
dem Laborteam bedanke ich mich für die Unterstützung während des
Untersuchungszeitraumes.
Weiterhin danke ich ganz herzlich den Landwirten, die ihre Bestände für die
Untersuchung zur Verfügung gestellt haben.
Stefan, Anne und meinem Vater danke ich für die Hilfe bei der Entnahme der
Blutproben, beim Ferkel wiegen und impfen sowie für die vielen Stunden auf dem
Schlachthof.
Ganz besonders möchte ich mich bei Monika für die praktische Hilfe während des
Versuchs und vor allem für den gegenseitigen Motivationsaufbau während der
gesamten Zeit Bedanken.








![MYCOPLASMA DUO 62739 62740 - Bio-Rad Laboratories · † Mycoplasma hominis(MH) wurde als eine Ursache für die Entzündung der Gebärmutterschleimhaut und der Eileiter [12] und in](https://img.pdfslide.org/doc/110x75/5f0ccaf67e708231d4372828/mycoplasma-duo-62739-62740-bio-rad-a-mycoplasma-hominismh-wurde-als-eine-ursache.jpg)












