Embed Size (px)
Citation preview

446 | Pharm. Unserer Zeit | 34. Jahrgang 2005 | Nr. 6
T R E F F P U N K T FO R SC H U N G |
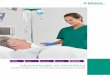
Dies sind die Biosynthese-Enzymevon so genannten Siderophoren. Siderophore sind niedermolekulareSubstanzen (meist Phenol- oder α-Hy-droxycarbonsäuren) und werden vonden Mikroorganismen gebildet, um
dreiwertiges Eisen aufzunehmen undzu speichern. Mycobacterium tuber-culosis und Yersinia pestis synthe-tisieren die Siderophore Mycobactin(MBT) bzw. Yersiniabacin (YBT), bei-de sind durch eine Polypeptid-Poly-
ketid-Hybridstruktur und eine Salicyl-säure als Endgruppe charakterisiert.Diese Siderophore haben eine sosehr hohe Affinität zu Fe3+, dass dasMetall aus dem humanen Transport-protein Transferrin extrahiert und damit für das Bakterium verfügbargemacht werden kann.
Ein wichtiger Schritt in der Bio-synthese der Siderophore ist die ATP-abhängige Übertragung eines Salicyl-säurerestes auf ein Aroyl-Träger-Pro-tein (ArCP).
Der erste Schritt ist eine ATP-ab-hängige Adenylierung des Salicylats,um ein Salicyl-AMP-Intermediat (sie-he Abb.) zu erzeugen, das im aktivenZentrum des Enzyms verbleibt. Dannfolgt eine Esterübertragung desSalicylsäurerestes auf das Thiol derprosthetische Phosphopantetheinyl-Gruppe des ArCP. Die Hemmung die-ser Enzyme MbtA und YbtE, die inden verschiedenen Mikroorganismendiesen Syntheseschritt katalysieren,war das Ziel. Deshalb wurden nichthydrolysierbare Acyl-AMP-Analoga alsMechanismus-basierte Inhibitorensynthetisiert [1]. Dabei wurde einKokristallisat eines verwandtenAdenylat-bildenden Enzyms mit ei-nem 2,3-Dihydroxybenzoyl-Interme-diat [2], DhbE, dessen Enzym einesehr große Homologie zu den obengenannten Enzymen insbesondere imaktiven Zentrum ausweist, genutzt,um ein optimales Intermediat-Analo-gon zu finden. Das nicht geladene Salicyl-AMS, das statt der Phosphat-Gruppe eine Sulfamat-Gruppe auf-weist, ist in der Lage, MtbA und YbtEin nanomolaren Konzentrationen zuhemmen. Damit wird erwartungsge-mäß die Produktion von MTB undYBT so stark inhibiert, dass weder inMycobakterien noch in Yersinien dieSiderophore nachweisbar waren.Außerdem konnte eine Wachstums-hemmung beider Mikroorganismusunter Eisen-limitierten Bedingungenbei mikromolaren Konzentrationendes Inhibitors nachgewiesen werden.
Damit ist eine sehr gute Aus-gangsbasis gefunden, um bessere Enzyminhibitoren und damit Anti-infektiva zu entwickeln, die die Mem-
N
NN
N
NH2
O
OHOH
OS
NH
O O OOH
OH
O
N
R1
NH
OHN
N
O
O O
O
N R2
O
OH
CH3
OH
H
Mycobactine (MBTs)
OH
S
NNH
S
N
S
OH
H3CCH3
O
OHH3C
Yersiniabactin (YBT)
N
NN
N
NH2
O
OHOH
OP
O
O O OOH –
Salicyl-AMP
Salicyl-AMS
Salicylat + ATP + Enzym
PPi
+ Enzym (1)
N
NN
N
NH2
O
OHOH
OP
O
O O OOH –
Salicyl-AMP
+ Enzym Salicyl-ArCP + Enzym (2)
AMP
ArCP
Enzyme: für MBT = MbtA und für YBT = YbtE
A B B . 1 | S I D E RO PH O R- B I OS Y N T H E S E A L S TA RG E T F Ü R A N T I I N F E K T I VA
M E D IZ I N |Neue Wege zu selektiven AntiinfektivaDie Suche nach Antiinfektiva, die nur den Mikroorganismus treffen undnicht den Menschen, ist so alt wie die Antibiotika-Entwicklung begin-nend mit Salvarsan. Jetzt haben amerikanische Forscher ein neues Target gefunden, das es nur bei Bakterien und Mykobakterien gibt, abernicht bei Menschen.
Für die Biosynthese der Siderophore Mycobactin und Yersiniabactin wird ein Salicylsäurerest ATP-abhän-gig auf ein Aroyl-Träger-Protein (ArCP) übertragen. Dieser Schritt wird von den Enzymen MbtA bzw. YbtEkatalysiert. Durch das Analogon Salicyl-AMS lassen sich beide Enzyme sehr effizient inhibieren.

Nr. 6 | 34. Jahrgang 2005 | Pharm. Unserer Zeit | 447
| T R E F F P U N K T FO R SC H U N G
branen besser penetrieren und damitden Mikroorganismus besser treffen.
[1] Ferreras, J. A., Ryu, J.-S., Di Lello, F., Tan,D.S., Quadri, L.E.N.: Small-molecule inhibi-tion of siderophore biosynthesis in Myco-bacterium tuberculosis und Yersinia pestis.Nature Chem. Biol. 1 (2005), 29-32.
[2] May, J.J., Kessler, N., Marahiel, M.A.,Stubbs, M.T.: Crystal structure of DhbE, anarchetype for aryl activating domains ofmodular nonribosomal peptide synthases.Proc. Natl. Acad. Sci. USA 99 (2002),12120-12125.
U. Holzgrabe, Würzburg
M E D IZ I N |Was macht die Telomerase bei Stammzellen?Die Bedeutung von Telomerlänge und Telomerase-Aktivität bei Tumorenund Zellalterung ist inzwischen schon hinreichend aufgeklärt. Wie die Telomerase auf die Proliferation von Stammzellen wirkt, wird derzeitnoch kontrovers diskutiert.
A B B . 1 T E LO M E RV E R K Ü R Z U N G B E I D E R
R E PL I K AT I O N
5'
5' 3' 3' 5'
5' 3'3'
5'
Replikation
5'3'
3'
5'
3'
RNA-Primer
5' 3'3'
5'
5'
3' 3' 5'
Verlust eines Teilsdes Chromosoms
Die linearen DNA-Moleküle der Eu-karyonten-Genome haben im Ver-gleich zu den zirkulären Genomender Prokaryonten den Nachteil, dassunsere Chromosomen bei jeder Ver-dopplung ein kleines Stückchen ver-lieren. Das liegt einfach darin begrün-det, dass DNA-Synthese immer nurgerichtet, vom 5’-Ende zum 3’-Endeerfolgen kann. Während die Replika-tion des einen DNA-Stranges deshalbin „einem Rutsch“ ablaufen kann,muss sich der andere Strang „müh-sam“ aus einzelnen so genanntenOkazaki-Fragmenten zusammenset-zen und dabei bleibt dann immer einkleines Stück DNA von 50 bis 100 Ba-senpaaren „auf der Strecke“ (Abb. 1).
Das ist insofern nicht dramatisch,als diese Chromosomenenden keinewertvolle Information tragen: DieseTelomere bestehen bei unseren Zellen aus der Basenabfolge TTAGGG,die so oft wiederholt wird, dassschließlich Enden von 5.000 bis15.000 Nukleotiden resultieren.Trotzdem führt es dazu, dass sichnormale menschliche Zellen nur ca.50mal teilen können. Danach gehensie in die Replikations-Seneszenzüber. Dass wir uns dennoch aus nureiner befruchteten Eizelle entwickelnkönnen und dabei sicherlich mehrals 50 Zellteilungen stattfinden, liegtdaran, dass pluripotente Stammzellen
und auch Keimzellen ein Enzym ha-ben, das diese Telomere immer wie-der verlängert und so die Alterungder Zellen verzögert. Diese so ge-nannte Telomerase ist ein Komplexaus RNA (TERC) und mehreren Pro-teinen, die unter anderem eine Re-verse-Transkriptase-Aktivität (TERT)aufweisen. Die im Komplex enthal-tene RNA wird von der ReversenTranskriptase als Matrize für dieReplikation der Telomere verwendet,wodurch deren ursprüngliche Längeerhalten bleibt (Abb. 2).
Während die Telomerase in diffe-renzierten Gewebezellen nicht mehraktiv ist, scheinen Tumorzellen inder Lage zu sein, das Enzym wiederzu reaktivieren, weshalb dieses En-zym auch als Zielstruktur für mög-liche Tumortherapeutika gehandeltwird. Eine mutierte Telomerase mitgeringerer Aktivität führt zu Krank-heiten wie z.B. zu kongenitalerDyskeratose oder aplastischer Anä-mie. Und TERC –/–-Knock-out-Mäuse,die keine aktive Telomerase mehr bilden können, altern vorzeitig undhaben adulte Stammzellen, derenProliferationsfähigkeit eingeschränktist.
Diese Knock-out-Mäuse verwen-dete Maria Blasco und ihre beidenMitarbeiter am Spanischen Nationa-len Krebszentrum in Madrid für ihre
Untersuchungen zum Effekt der Telo-merase und der Telomerlänge aufepidermale Stammzellen, die mitteAugust in Science publiziert wurden[1]. Sie konnten zeigen, dass dieTERC –/–-Mäuse in der 3. Generation(G3), die ja wegen der fehlendenTelomerase deutlich kürzere Telome-re als ihre Verwandten in der 1. Ge-neration (G1) aufweisen, deutlichmehr epidermale Stammzellen in denHaarfollikeln aufweisen, als G1-Mäu-se und diese wiederum mehr alsTERC+/+-Wildtyp-Mäuse. Allerdingsließen sich diese Stammzellen beiden G3-Mäusen wesentlich schlech-ter durch den Tumorpromotor 12-O-tetradecanoylphorbol-13-acetat (TPA)mobilisieren als bei G1- bzw. Wild-typ-Mäusen, und sie konnten auchwesentlich weniger Kolonien bilden.So bewirken die kurzen Telomere
Wird die DNA repliziert, so kann einer der beiden Tochter-stränge, der so genannte „Leading“-Strang an einem Stückverdoppelt werden, während der „Lagging“-Strang dadurchentsteht, dass immer wieder kurze RNA-Primer (orange) an-gelagert werden, die dann mit Desoxynukleotiden verlängertund schließlich von der DNA-Polymerase hydrolysiert wer-den. Die entstandenen Okazaki-Fragmente werden zumSchluss zum fertigen Strang ligiert. Wegen der nötigen RNA-Primer verlieren die DNA-Moleküle bei jeder Replikation 50 bis 100 Basenpaare.