Embed Size (px)
Citation preview

Arch. exper. Path. u. Pharmakol., Bd. 229, S. 207--219 (1956)
Aus der Universit~ts-Kinderklinik Freiburg im Breisgau (Direktor: Prof. Dr. W. KELLER)
0xydation menschlicher und tierischer 0xyh~imoglobine dutch Kaliumferrieyanid ~
Yon
K. BETKE~ I . GREINACHER und F. HECKER
Mit 6 Textabbfldungen
(Eingegangen am 9. April 1956)
Kal iumfe r r i cyan id (KaFe(CN)e) reagier t q u a n t i t a t i v mi t Hi~moglobin- eisen, indem es das Eisen aus der zwei- in die dre iwer t ige Stufe fiber- ff ihr t 1, s, 9, ,15. Es b i lde t sich Hi~miglobin (Methi~moglobin, HbI I I ) . Lag der Blu t fa rbs to f f als Oxyhi~moglobin (02Hb) vor, d a n n wird der moleku- lar gebundene Sauerstoff bei diesem Vorgang ffei ; eine Tatsache , die ffir die vo lumet r i sche Messung des gebundenen Sauerstoffs ausgenu tz t wirdl0, 22. Die Oxyda t ion yon 02Hb n i m m t bei geeigneten Versuchs- bed ingungen einige Minuten in Anspruch, is t also leicht messend zu ver- folgen. Bei vergle ichenden Messungen fanden wir Speciesunterschiede der Oxydat ionsgeschwindigke i ten . ~ b e r diese Befunde soll im folgenden be- r ich ter werden.
Material und Methodik OxyMimoglobinl6sungen. Die fiir eine Priifung erfordcrliche Mindestblutmenge
betrug 0,1--0,2 cm 3. Die Erythrocyten wurden auf der Zentrifuge 3mal mit 1,5~oiger KochsalzlSsung gewaschen und mit Wasser h~molysiert. Zu dem H~mo- lysat wurde kolloidales Aluminiumhydroxyd in einer Menge zugesetzt, die 4--5 real das Volumen des benutzten Erythrocytenbreies ausmachte. Nach Schiitteln wurde filtriert. Die resultierende vollkommen klare BlutfarbstofflSsung konnte als weit- gehend gereinigt und doch ungesch~digt angesehen werden a. - - Die Blutfarbstoff- konzentration wurde nach BETKE U. SAVELSBERG an einer Probe fiber Cyanh~mi- globin mit dem Zeiss-Spektralphotometer bei 540 m# gemessen. Durch Verdfinnung mit Wasser wurde eine Konzentration yon 105 mg-~o eingestellt. AnschlieBend wurde 1/20 Vol. eines 2,8 (teilweise auch 2,0) mol. Phosphatpuffers vom PH 6,8 oder 7,0 zugesetzt, so dab endgfiltig eine Hb-Konzentration yon 100 mg-~o resultierte.
Kalium/erricyanid. Ffir die Versuche wurde meist eine 0,01 tool. L5sung ver- wendet. Diese wurde jeweils frisch aus einer 0,5 mol. StammlSsung hergestellt, die 0,25~o Natriumcarbona~ enthielt. Die StammlSsung wurde im Dunklen gehalten und alle drei Wochen erneuert. Verwendet wurde Kaliumferricyanid p.A. (Merck).
Versuchsvorgang. Die angegebenen Daten beziehen sich auf das zuletzt als Standardmethode angenommene Vorgehen. Prinzip: Es wird das Anwachsen der HbIILKonzentration am Anwachsen der Extinktion bci 630 m/~ im Ablauf der Zeit
* Mit Unterstfitzung der Deutschen Forschungsgemeinschaft. Arch. exper. Path. u. Pharmakol. Bd. 229 15

208 K. BETKE, I. GREINACHER und F. HECKER:
verfolgt. - - 2,5 cm a der 02Hb-LSsung wurde in eine Cuvette von 1 em Schichtdicke des Zeiss-Spektralphotometers pipettiert. Bei 630 m~ wurde die Extinktion (Eo) gegen Wasser als Vergleichswert gemessen. Mit einer schlanken Blutzuekerpipette wurden 0,07 cm 3 der 0,01 n KaFe(CN)6-LSsung eingeblasen; es wurde rasch zwehnal aufgesaugt und wieder ausgeblasen und mit der Pipette durchmischt. Im Augen- blick des Einblasens wurde eine Stoppuhr in Gang gesetzt. 15 sec sparer konnte die erste Ablesung des Galvanometerausschlags gemacht werden. Es wurde au•erdem naeh 20, 30, 40, 50, 60, 80, 100 und 120 sec abgelesen. Nach 3, 4 oder 8 rain, d. h. zu einem Zeitpunkt, an dem sich der Galvanometerausschlag nicht mehr veri~nderte, wurde der Endwert (Ee) abgelesen. Er repr~sentierte den Wert nach v6lliger Um- wandlung des Blutfarbstoffs in HbIII, Ee---Eo den gesamten Zuwachs an HbIII. Der Prozentsatz an umgewandeltem Blutfarbstoff zu der Zeit t lieI3 sich aus der zu-
Et - - Eo gehSrigen Extinktion E t als E ¢ - - - E o " 100 berechnen.
Die dutch den Zusatz yon 0,07 cm a K3Fe(CN)6-LSsung bedingte Volumen- vermehrung und damit Verkleinerung yon Eo wurde vernachl~ssigt, da sie in der Berechnung nicht ins Gewicht fiel. Der Farbwert yon KaFe(CN) ~ brauchte ebenfalls nicht beriieksichtigt zu werden, da er in der gew~hlten Konzentration bei 630 m]~ unmei~bar klein ist.
Im Reaktionsansatz befindet sich bei der gew~hlten Anordnung eine Kon- zentration yon 2,72 • 10 -4 Mo]/1 KaFe(N)6 und 5,72 ~ 10-SAqu./1 02Hb (1 Aquivalent
1/4 Molekiil, Aquivalentgewicht 17000). Vom Oxydans ist also das 4,75faehe der ftir eine quantitative Reaktion erforderlichen Menge vorhanden.
Alle Untersuchungen wurden als Doppelversuche, teilweise auch als Dreifach- versuche durchgefiihrt, und zwar so, daI] in einer Versuchsreihe immer erst alle Erstversuche nacheinander und dann anschliel~end die Zweitversuche absolviert wurden. Ffir 20 ohne Auswahl herausgegriffene, im Zusammenhang durehgeftihrte Doppelversuche h~ben wir eine mittlere Streuung gegenfiber dem Mittelwert yon -4- 1,85% HbIII mit einem mittleren Fehler yon ± 0,41% errechnet (fiir die Oxy- dation nach 20 sec) ; die Sicherheit der Einzelbestimmung war also recht gut.
Da die FerricyanidlSsung zwar in der Hauptmenge schlagartig eingebraeht wird, bis zur vSlligen S~uberung der Pipette und Durchmischung des Cuvetten- inhalts jedoeh etwa 2 sec verstreichen, wurden auch noch andere Wege ausprobiert, z. B. ein Einspritzen mit einer Recordspritze nnter starkem Druek durch eine sehr feine Kaniile. Dadurch wurde keine Verbesserung der Methode erzielt. Auch die ver- suchte photographische Registrierung des jeweiligen Standes des wandernden Galvanometerzeigers anstelle der einfachen Ablesung brachte keine Vortefle, die die erheblichen Umst~nde dieses Verfahrens aufgewogen h~tten.
~eben 02Hb wurde auch in einigen F~llenCOHb untersucht, t t ierzuverwendeten wir speziell angefertigt e Cuvetten, die einen verschliel~baren Einffillstutzen be- sal~en. Die BlutfarbstofflSsung wurde jewefls vor dem Versueh in der Cuvette mit CO durchperlt. Auf diese Weise konnte der Oxydationsablauf in einem abgeschlos- senen, mit CO geffillten Raum studiert werden. Da bei COHb die Oxydation aul]er- ordentlich ]angsam abl~uft, muflte wesentlich mehr KaFe(CN)~ in Reaktion ge- braeht werden. Die LSsung (0,6 cm a einer 0,75 oder 1 tool. LSsung auf 4 cm a COHb- LSsung) wurde mit einer Tuberkulinspritze und feiner Kaniile eingespritzt und die Cuvette rasch mit der Hand durchgeschfittelt. Die Auswertung geschah analog den Versuchen mit O~H, nur wurde Eo wegen der hSheren Volumenverdfinnung und der st~rkeren Eigenabsorption yon COHb korrigiert.
Untersuchungsgut. Die Ergebnlsse der Arbeit grtinden sieh auf die Untersuehung der Blutproben yon fiber 50 Erwachsenen und Kindern, 28 Neugeborenen (~abel- schnurblut), 7 erwachsenen Rindern, 1 Kalb, 4 Rinderfeten (bei Schlachtung der
Oxydation menschl, u. tierischer Oxyh~moglobine durch Kaliumferricyanid 209
Muttertiere gewonnen), 2 Hirschen, 5 Schweinen, 2 Hammeln, 8 Kaninchen, 9 Meer- schweinchen, 16 M~usen und 1 Huhn. Aul3erdem konnten noch 3 Blutproben von Sichelzellan~mie-Patienten und 1 Blutprobe eines heterozygoten Tr~gers der H~mo- globinanomalie C (Hb A und Hb C im Blut) untersucht werden. Herrn Prof. SIN~ER, Chicago, der uns diese pathologischen menschlichen Blutproben uuf dem Luftweg zuschickte, mSchten wir auch an dieser Stelle herzlich danken.
E r g e b n i s s e
1. A u s w e r t u n g der Versuchsergebn i s se
Nach Zusatz des Oxydat ionsmi t te l s setzte die Oxyda~ion sofort mi t
raschem Anst ieg ein und war, al lm~hlich langsamer werdend, in wenigen
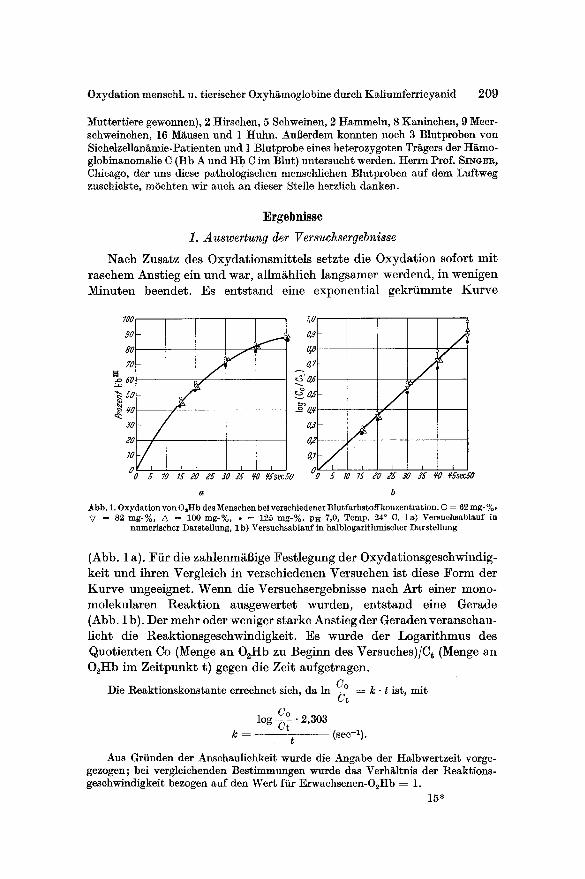
Minuten beendet . Es en t s tand eine exponent ia l gekrf immte K u r v e
zoo z,o
3o ~ o,9
~6o
" , j ° gO ~ O,q
2o o~
70 ~I
t 0 / l I r I / r I b" lg 7b" 20 25- 3g 3£ q0 ~'secS0
b Abb. 1. Oxydation yon OsHb des :V[enschen bei verschiedener Blur farbst offkonzent ration. © = 62 rag- %, ~7 = 82 rag-%, • = 100 rag-%, • ~ 125 mg-%. PH 7,0, Temp. 24 ° C. la) Versuchsablauf in
numeriseher Darstellung, I b) Versuchsablauf in halblogarithmischer Darstellung
(Abb. 1 a). Fi i r die zahlenm~Bige Fes t legung der Oxydat ionsgeschwindig-
kei t und ihren Vergleich in verschiedenen Versuchen ist diese F o r m der
K u r v e ungeeignet . W e n n die Versuchsergebnisse nach Ar t einer mono-
molekularen Reak t ion ausgewerte t wurden, en t s tand eine Gerade
(Abb. 1 b). Der mehr oder weniger s tarke Anst ieg der Geraden veranschau-
l icht die Reakt ionsgeschwindigkei t . Es wurde der Logar i thmus des
Quot ien ten Co (Menge an O2Hb zu Beginn des Versuches)/Ct (Menge an
O~Hb im Ze i tpunk t t) gegen die Zeit aufgetragen.
Die Reaktionskonstante errechnet sich, da In C_~ ~ / c • t ist, mit
Co log ~ - - 2,303
k -- t (see-Z)"
Aus Griinden der Anschaulichkeit wurde die Angabe der Halbwertzeit vorge- gezogen; bei vergleichenden Bestimmungen wurde das Verh~ltnis der Reaktions- geschwindigkeit bezogen auf den Wert fiir Erwachsenen-O2Hb : 1.
15"

210 K. BETKE, I. GREINACltER und F. HECKER'-
Auch der Ablauf der Oxydation yon COHb liel~ sich am ehesten nach der Art einer monomolekularen Reaktion graphisch darstellen. Es fiel aber auf, dab die ersten Werte (15--30 sec) stets ein wenig oberhalb der erhaltenen Geraden lagen (s. Abb. 6).
2. Ein/lu[3 der i~u/3eren Versuchsbedingungen
a) Vorbereitung der 02Hb-Ldsungen. Es wurden verglichen: Einfaches H/~molysat aus gewaschenen Erythrocyten, durch Adsorption an Alumi- niumhydroxyd gereinigtes H/£molysat, durch Adsorption und Dialyse gereinigtes H~molysat. Die Oxydation lief in allen drei Pr~parationen gleich schnell ab. Auf eine Dialyse konnte daher ftir die Versuche ver-
zichtet werden. Die Reini- Tabelle 1. Halbwertzeit der Oxydation von Er- wachsenen-O2Hb durch KaFe(CN)s bei verschie-
denem PH (Phosphat, 0,1 mo O
PH 6,1 6,58 6,88
Halbwertzeit sec • • . 8 17 31 Reaktionsgeschwindig- keit relativ, abgerundet 4 2 1
gung durch Adsorption an Aluminiumhydroxyd wur- de jedoch wegen des Vor- teils der vollkommenen Klarheit der 02Hb-LSsun- gen beibehalten.
Wurde den 02Hb-L6- sungen vor Versuchsbeginn
eine geringe Menge Ascorbins~ure zugesetzt (rund 1/1O der vorhandenen O2Hb-Xquivalente ), dann stSrte das die Oxydation nicht (ira Gegensatz zu der Oxyd~tion mit N~triumnitrit, die durch einen solchen Zusatz enorm verlgngert wirdS). Ebenso wenig war eine Vorbehandlung mit Jod (~enge dem O2Hb ~quivalent) yon Einflul~.
b) Pu//erung. In Best~tigung ~iterer eigener Befunde wurde gefunden, dab die Reaktionsgeschwindigkeit mit fallendem pH steigt (Tab. 1). Die pH-Werte wurden in den fertigen 02Hb-LSsungen mit der Glaselektrode ermittelt.
Auch die Pufferkonzentration hatte einen gewissen Einflul~. Je hSher die Menge an zugesetztem Phosphat, desto rascher der Oxydationsvor- gang.
c) Temperatur. Exakte Messungen der Temperatur~bh~ngigkeit konnten mangels geeigneter Vorrichtungen nicht gemacht werden. Durch Vorkiihlen oder Vorw~rmen der Re~ktionsl6sungen konnte jedoch orientierend festgestellt werden, dab die Reaktionsgeschwindigkeit mit steigender Temperatur betr/~chtlich zu- nimmt. Alle in dieser Arbeit referierten Versuche wurden bei Zimmertemperatur vorgenommen. Sie konnte nicht konstant gehalten werden und schwankte von Versuchsreihe zu Versuchsreihe im Extrem zwischen 20 ° und 26 ° C.
d) Konzentration an KaFe(CN)s. In der Umgebung des gew~hlten Konzentrationsbereichs stieg mit steigender Menge an K3Fe(CN)s die l~eaktion annghernd im gleichen AusmaB (Tab. 2).
Bei COHb konnte ein grSl~erer Konzentrationsbereich an KsFe(CN)~ geprtift werden. Bei den Konzentrationen (in Mol/1) 1,63.10 -e, 3,26.10 -2,

Oxydation mensehl, u. tierischer Oxyh/~moglobine dureh Kaliumferricyanid 211
6,52- 10 -2 und 11,74.10 -2 (d. h. also einem Verh/~ltnis von 1:2:4:7,2) verhielten sich die Reaktionsgeschwindigkeiten wie 1 : 1,8 : 3,3 : 5,1.
e) Konzentration an Blut/arbsto]]. Abb. 1 zeigt den Oxydationsablauf bei folgenden Konzentrationen yon O2Hb : 62, 82, 100 und 125 mg-~o. Die Reaktionsgesehwindigkeit wurde kaum beinfluBt; immerhin lagen die Werte fiir die Konzentraton 125 nag-~o konstant unter denen fiir die Konzentration 62 rag-~o. In Versuehen mit COHb, bei denen ein sehr hoher KzFe(CN)6-~berschuB bestand, war die Variation der Blutfarbstoff- konzentration bis zum 4fachen ohne EinfluB auI die Reaktionsge- sehwindigkeit.
TabeUe 2. Halbwertzeit der Oxydation yon Erwachsenen-OzHb bei verschiedenen Konzentrationen yon KsFe(CN)6
Menschl. Erwachs. O2Hb
M/~use O2Hb
KsFe(CN)6 tool/1
KaFe(CN)6-Konzentr. relativ . .
Halbwertzeit sec Reaktionsgeschwindigkeit relativ
Halbwertzeit sec . . . . . . . Reaktionsgesehwindigkeit relativ
2,18 • 10 -412 ,72 .10 -4
0,8
37,5 0,83
13 0,85
1,0
31 1,0
11 1,0
3,26 "]0 -4
1,2
26 1,19
9 1,22
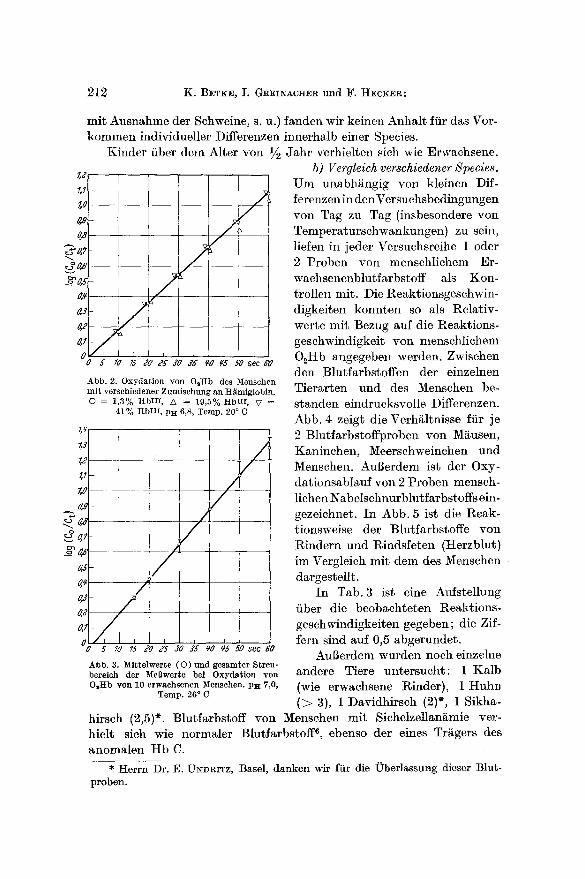
/) Gehalt an Hdmiglobin vor Versuchsbeginn. Aus Erythrocyten, die mit NaN% behandelt worden waren, wurde eine gereinigte, dialysierte HbHi-LSsung hergestellt. Von ihr wurden verschieden grofle Anteile zu einer O~Hb-LSsung gesetzt. Die Oxydationsgeschwindigkeit des An- teils an O2Hb in jeder dieser Mischungen war gleich der einer reinen O2Hb-LSsung (Abb. 2). Die Beimischung yon Hi~miglobin war also ohne Einfiul~.
Zusammen/assung. Der Reaktionsablauf wird mal]gebend vom pH- Wert, yon der Temperatur und yon der Menge des Oxydans bee~nfluBt. Auch die Menge an zugesetztem Puffer ist nicht gleichgiiltig. Ffir ver- gleichende Untersuchungen ist daher erforderlich, diese GrSl]en exakt konstant zu halten. Ohne EinfluB sind dagegen Differenzen in bezug auf AusmaB der Reinigung der H/~molysate, in bezug auf den Gehalt an vor Versuchsbeginn vorhandenem H/~miglobin und auch - - allerdings in gewissen Grenzen - - in bezug auf die Blutfarbstoffkonzentration.
3. Vergleichende Untersuchungen
a) Vergleich verschiedener lndividuen einer Species. In Abb. 3 ist der gesamte Streubereich der Werte von 10 erwachsenen Menschen dar- gestellt. Ffir den Stand der Oxydation nach 20 sec errechnete sich eine mittlere Streuung yon ~ 1,16~o Hb HI mit einem mittleren Fehler yon
0,37~o. Danach sind individuelle Differenzen der Oxydationsge- schwindigkeit der Blutfarbstoffe verschiedener Menschen unwahrschein- lich. Auch bei den bisher yon uns untersuehten Tieren (mSglieherweise
2 1 2 K . BETKE, I . GREINACHER u n d F . H E C K E R :
mit Ansnahme der Schweine, s. u.) fanden wir keinen Anhalt ffir das Vor- kommen individueller Differenzen innerhalb einer Species.
Kinder fiber dem Alter yon 1/2 Jahr verhielten sich wie Erwachsene.
7,21
q,o o,81 o,8
~o,e ~ 0 ~ / ' -
g¢
O,q
0 g I0 75 20 Z£30 3S ~0 g5 50secGO
Abb. 2. Oxydation yon DgHb des Me~ehen mit verschiedener Zumischung an H~miglobin. © = 1,3% ]tbtII, ~ = 19,5% ttbIII, V =
41% t tbHL p~ 6,8, Temp. 20 ° C
1,1
o,9 .~.o,Q
o~
~2
Abb. 3. ~i t te lwer te ( 0 ) und gesamter Streu- bereieh der ~[eBwerte bei Oxydat ion yon O,Hb yon 10 erwachsenen Mensehen. Plt 7,0,
Temp. 26 ° C
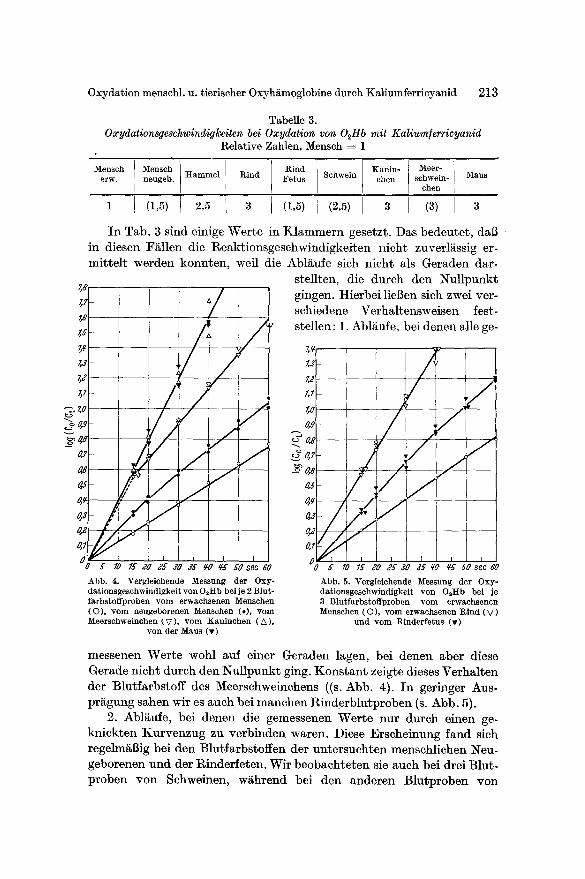
b) Vergleich verschiedener Species. Um unabh~ngig von kleinen Dif- ferenzenin denVersuchsbedingungen yon Tag zu Tag (insbesondere yon Temperaturschwankungen) zu sein, liefen in jeder Versuchsreihe 1 oder 2 Proben yon menschlichem Er- wachsenenblutf~rbstoff als Kon- trollen mit. Die Reaktionsgeschwin- digkeiten konnten so als Relativ- werte mit Bezug auf die Reaktions- geschwindigkeit von menschlichem O2Hb angegeben werden. Zwischen den Blutfarbstoffen der einzelnen Tierarten und des Menschen be- standcn eindrucksvolle Differenzen. Abb. 4 zeigt die Verhi~ltnisse fiir je 2 Blutfarbstoffproben von M~usen, Kaninchen, Meerschweinchen und Menschen. Au~erdem ist der Oxy- dationsablauf yon 2 Proben mensch- lichen Nabelschnurblutfarbstoffs ein- gezeiehnet. In Abb. 5 ist die Reak- tionsweise der Blutfarbstoffe yon Rindern und Rindsfeten (Herzblut) im Vergleich mit dem des Menschen dargestellt.
In Tab. 3 ist eine Aufstellung fiber die beobachteten Reaktions- geschwindigkeiten gegeben; die Zif- fern sind auf 0,5 abgerundet.
Aul]erdem wurden noch einzelne andere Tiere untersucht : 1 Kalb (wie erwachsene Rinder), 1 Huhn ( ~ 3), 1 Davidhirsch (2) ~, 1 Sikha-
hirsch (2,5) ~. Blutfarbstoff yon )/[enschen mit Siehelzellan~mie ver- hielt sich wie normaler Blutfarbstoff 6, ebenso der eines Tr~gers des anomalen Hb C.
* Herrn Dr. E. U~DRITZ, Basel, danken wir fiir die Uberlassung dleser Blut- proben.
Oxydation menschl, u. tierischer Oxyh~moglobine dureh Kaliumferricyanid 213
Tabelle 3. Oxydationsgeschwlndigkeiten bei Oxydation yon O~Hb mit Kalium/erricyanid
Relative Zahlen, Mensch = 1
Mensch Mensch I Rind :Kanin- Meer- erw. neugeb. I I tammel Rind Fetus Schwein chen schwein- Maus
chen
1 (l,S) I 2,s i 3 (1,5) (2,s) 3 (3) 3
In Tab. 3 sind einige Werte in Klammern gesetzt. Das bedeutet, dab in diesen lV/~llen die Reaktionsgesehwindigkeiten nicht zuverl~ssig er- mittel t werden konnten, weil die Abl/iufe sich nicht als Geraden dar-
:,5
7,3- 7/ 7,Z
o,~ ~6
F
(
/ /
/ /
, . , o ~ 70 IE 20 2£ 30 3E ~0 ~£ 50secGO
Abb. 4. Vergleichende Messung der Oxy- dationsgeschwindigkeit von O2Hb bei je 2 Blut- farbstoffproben yore erwachsenen Menschen (O), vom neugeborenen Menschen (.), vom Meerschweinchen (V), vom Kaninchen (A),
yon der Maus (v)
stellten, die durch den Nullpunkt gingen. Hierbei lieBen sich zwei ver- schiedene Verhaltensweisen fest- stellen: 1. Abl/£ufe, bei denen alle ge-
;7
0 , ~
~ 0,~
o,~
o,~
6 0 35 ¢0 ~5 50sec6"O
Abb. 5. Vergleichende ]Vfessung der Oxy- dationsgeschwindigkeit von 02ttb bei je 8 Blutfarbstoffproben vom erwachsenen Menschen (O), vom erwachsenen Rind (V)
und vom Rinderfetus (v)
messenen Werte wohl auf einer Geraden lagen, bei denen aber diese Gerade nicht durch den Nullpunkt ging. Konstant zeigte dieses Verhalten der Blutfarbstoff des Meerschweinchens ((s. Abb. 4). In geringer Aus- prBgung sahen wit es auch bei manchen Rinderblutproben (s. Abb. 5).
2, AblBufe, bei denen die gemessenen Werte nur durch einen ge. knickten Kurvenzug zu verbinden waren. Diese Erscheinung land sich regelm/~Big bei den Blutfarbstoffen der untersuchten menschlichen Neu- geborenen und der Rinderfeten. Wir beobachteten sie auch bei drei Blur. proben yon Schweinen, w~hrend bei den anderen Blutproben yon
214 K . ~ E T K E , I . GREINACttER u n d F . HECKER:
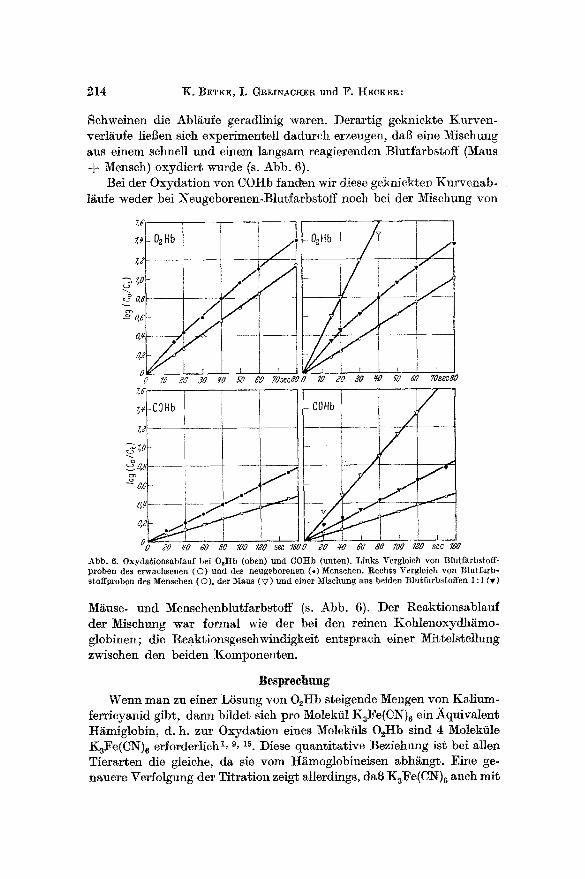
Schweinen die Abl~ufe geradlinig waren. Derartig geknickte Kurven- verl~ufe liel]en sioh experimenteH dadurch erzeugen, dab eine Mischung aus einem schnell und ehlem langsam reagierenden Blutl~rbstoff (Maus
Mensch) oxydiert wurde (s. Abb. 6). Bei der Oxydation yon COHb fand~n wir diese geknickten Kurvenab-
1/~u~e weder bei Neugeborenen-Blutfarbstoff noch bei der Mischung yon
I,~ - 02Hb Hb / v
o,z
0 i I , , 0 !# 20 JO q# ~YO gO 7#sec8gO 70 Z# 30 g-# YO ~ 70secSO
7,6 l / ~,~ .COHb -COHb
], I
~0 Zo #0 #0 30 :00 120 ssc 7600 20 gO $0 80 10# 120 sea 1~
Abb. 6. Oxyda~ionsab/auf bei O2Itb (oben) nnd COHD (unten). Links Yergleich yon Blu~farbstoff- proben des erwachsenen (O) und des neugeborenen ( , )Menschen. Rechts Vergleich yon ]~lutfarb- stoffproben des ]~enschen (©), tier ~faus ( V ) und einer .M[schung aus beiden Blutfarbs~offen 1 : 1 Or)
M~use- und Menschenblutfarbstoff (s. Abb. 6). Der Reaktionsablauf der Mischung war formal wie der bei den reinen Kohlenoxydh/imo- globinen; die Reaktionsgeschwindigkeit entsprach einer Mi~telstellung zwischen den beiden Xomponenten.
B e s p r e c h u n g
Wenn man zu einer LSsung von O2Hb steigende Mengen yon Kalium- ferricyanid gibt, dann bildet sich pro Molekiil KsFe(CN)s ein Aquivalent Hgmiglobin, d. h. zur Oxydation eines Molekfils O oHb sind 4 Molektile KsFe(CN) s erforderlich 1, 9, is. Diese quantitative Beziehung ist bei allen Tierarten die gleiche, da sie vom ttiimoglobineisen abhgngt. Eine ge- nauere Veffolgung der Titration zeigt allerdings, daJ~ KaFe(CN)s auch mi~

Oxydation menschl, u. tierischer Oxyhamoglobine durch Kaliumferricyanid 215
Anteilen der EiweiBkomponente des Blutfarbstoffs reagieren kann TM, 21. Es handelt sich hierbei um eine relativ langsame Reaktion, yon der bei den kurzen yon uns angewandten Beobachtungszeiten - - jedenfalls in den Versuchen mit O2Hb - - kaum StSrungen zu erwarten sind.
Obwohl die quanti tat iven Beziehungen zwischen KaFe(CN)s und H~moglobineisen bei allen Species die gleichen sind, konnten wir zeigen, dab die Geschwindigkeit, mit der sich O2Hb auf diese Weise oxydieren l~Bt, yon Species zu Species merklich differiert. Es l~Bt sich vorerst nicht erkennen, ob diese Differenzen irgendwelche Beziehungen zu bio- logischen Eigenarten der betreffenden Tiere haben. Interessant ist, dab von allen bisher yon uns gepriiften Blutfarbstoffen der des Menschen die geringste Oxydationsgeschwindigkeit aufwies. Wichtiger aber als die Aufz~hlung der einzelnen Reaktionsgeschwindigkeiten scheint uns die Tatsache zu sein, dab sie bei jeder Tierart eine recht scharf definierte und reproduzierbare GrSfle darstellt.
Da es sich gezeigt hat, dab die Oxydationsgeschwindigkeit recht un- empfindlich gegeniiber dem Reinigungsgrad der H~moglobinlSsungen ist, daft geschlossen werden, dal~ die beobachteten Differenzen tats~Lchlich speciesspezifische Eigenschaften der verschiedenen Blutfarbstoffe repr~L- sentieren und nicht etwa durch irgendwelche andere den LSsungen bei- gemischte Substanzen bedingt sind.
Der formale Ablauf der Oxydation war am besten nach der Gesetz- m~Bigkeit einer monomolekularen Reaktion darstellbar. Damit steht in Einklang, dab sich die Reaktionsgeschwindigkeit bei Variation der Hb- Konzentrat ion kaum ~nderte. Wurde der K3Fe(CN)6-t3berschuB hoch genug gew~hlt, wie im l%lle der Oxydation yon COHb, dann war tat- s~chlich die Oxydationsgesehwindigkeit v o n d e r Blutfarbstoffkonzentra- tion unabhgngig. Es handelt sich offensichtlich um eine der in biologischen Systemen so hgufig beobachteten pseudomonomolekularen Reaktionen. I m Reaktionsansatz laufen 4 verschiedene Vorggnge ab:
H b l I <
(Oxydation) - - e
t t b I I I
(Assoziation) +02 _02 * 02 Hb II
(Dissoziation)
Fe(CN)6,,, + e -~ Fe(CN) 6 ....
1. und 2. : Assoziation und Dissoziation yon 0 2 und Hb. Das Gleich- gewicht dieses Vorganges ist bei Anwesenheit yon Luft von normalem

216 K. BETKE, I. GI%EINACHER und F. HEOKER:
Atmosph/~rendruck sower nach rechts verschoben, dab der Blutfarb- stoff im wesentlichen als O2Hb vorhegt.
3. Die l%eaktion Hb rI --* I-[b In , also die Oxydation. Sie ist in vitro spontan irreversibel. Sie ist nut mSglich, wenn das Eisen keinen moleku- laren Sauerstoff tr~gt.
4. Die Reduktion yon Ferricyanid zu Ferrocyanid. Wenn man sauerstofffreies Hb I I m i t K~Fe(CN)~ reagieren li~Bt, ge-
schieht die Oxydation schlagartig; jedenfalls 1/iuft sie so schnell ab, dab sie in einer Versuchsanordnung, wie sie bier benutzt wurde, nieht meBbar ist 2. Die Assoziationsgesehwindigkeit yon Hb II und O 2 zu 02Hb ist auBerordentlieh hoeh 12. Wesentlich langsamer li~uft die Dissoziation yon O2Hb zu Hb und 02 ab 11. Sie ist abet Vorbedingung daffir, dab tiber- haupt eine Oxydation zu Hb IH stattfinden kann, da nut sauerstoff- freies Hb n oxydierbar ist s, la. Es leuchtet daher ein, dab dig Dissoziation als offensichtlieh langsamster Vorgang den formalen Ablauf tier Reaktion bestimmt ; die tatsaehliche Geschwindigkeit h/~ngt dann welter yon dem Verh/~ltnis der Gesehwindigkeiten der Oxydation einerseits und der Rekombination des durch Dissoziation entstandenen Hb II mit 02 zu O2Hb andererseits ab.
Die Sauerstoffdissoziation ist - - mit gewissen Korrekturen 2° - - ein mono- molekularer Vorgang 11. Ihre Abhangigkeit yon Temperatur u, pH 11, Salzkonzen- tration TM erkl/~rt wenigstens zu einem Tefl die gefundenen Abh&ngkeiten der Oxy- dationsgeschwindigkeit yon diesen GrSBen. Hinzu kommt, dab die Abh/~ngigkeit des Redoxpotentials Hbn ~ HbnI yon Temperatur und PH in die gleiche Richtung wirkt xa, 18. Die Assoziationsgeschwindigkeit des 02 mit Hb n ist dagegen unabh~ngig yon der Temperatur, pr~ und Salzkonzentration TM.
Die Speeiesdifferenzen der Oxydationsgeschwindigkeiten yon Oxy- h/imoglobinen kSnnten ihre Wurzel sowohl in Differenzen der Sauerstoff- assoziation als auch in solchen der Dissoziation oder auch in solchen der eigentlichen Oxydation des Hb-Eisens haben. Da direkte Messungen dieser Vorg~nge schwierig sind, existieren nut wenige Unterlagen, die man zu einer Entscheidung der Frage heranziehen k6nnte. Nach Unter- suchungen yon LASZT ist es unwahrseheinlieh, dab wesentliche Species- differenzen in bezug auf die Geschwindigkeit der Sauerstoffassoziation bestehen. Auch ffir den l~bergang von Hb II zu Hb u I liegen bisher nur Angaben vet, die keine Speciesdifferenzen anzeigen. HAVE~A~Z¢ land bei Pferd und Rind das Redoxpotential Hb n ~-- Hb HI identisch. In eigenen Untersuchungen wurde die Geschwindigkeit der Reduktion yon Hb H~ zu t tb H bei Nabelschnur- und Erwachsenenblutfarbstoff des Menschen nieht different gefunden a, was die SchluBfolgerung erlaubt, dab wahr- scheinlich auch der umgekehrte Vorgang, die Oxydation, nicht differieren dtirfte. Dagegen wurden bei eigenen Messungen der Gesehwindigkeit tier Dissoziation yon O2Hb zu 02 und Hb eindeutige Differenzen zwischen Nabelschnur- und Erwaehsenenblutfarbstoff des Mensehen gefunden,

Oxydation mensehl, u. tierischer Oxyh~moglobine durch Kaliumferricyanid 217
indem der Vorgang bei dem ersteren rascher ablief 3. Man kann also fiir den Fall des fetalen und bleibenden Blutfarbstoffs des Mensehen mit einiger Begrfindung behaupten, dad die Differenz in der Oxydierbarkeit der Oxyh~moglobine auf einer unterschiedliehen Dissoziationsgeschwin- digkeit des O~Hb beruht. Ffir die Differenzen zu den Tierblutfarbstoffen ist eine Analogie naheliegend, abet vorerst nicht bewiesen.
Die Oxydation einer Misehung aus einem schnell und einem langsam oxydierbaren Blutfarbstoff (M~use- und Menschen-O2Hb) erfolgte mit einer intermedi~ren Geschwindigkeit und formal insofern besonders, als der Kurvenzug in der halblogarithmischen Darstellung geknickt war (Abb. 6). Diese Kurvenform l~l~t sich ohne weiteres als Summation zweier mit verschiedener Geschwindigkeit unabh~ngig voneinander ablaufender pseudomonomolekularer Reaktionen erkl~ren. Wir haben an anderer Stelle gezeigt, daD man den erhaltenen Kurvenzug rechneriseh in seine beiden Komponenten zerlegen kann, wobei zwei Gerade mit Steigungs- maBen erhalten werden, die denen der beiden Ausgangsblutfarbstoffe entspreehen 5. Der Versuch mit dem Blutfarbstoffgemisch bietet eine Handhabe zur Erkl~rung der geknickten Kurvenform bei menschlichem Nabelschnurblutfarbstoff und dem Blutfarbstoff yon Rinderfeten. Wir wissen, dad diese Blutfarbstoffe nicht einheitlich sind, sondern aus 2 Komponenten bestehen, aus fetalem Blutfarbstoff und Blutfarbstoff, wie ihn der erwachsene Organismus besitzt. Dutch rechnerische Auf- 15sung der erhaltenen Kurven in beide Komponenten lied sieh ermitteln, dab reines fetales 02Hb des Menschen 2 ~ m a l so raseh oxydierbar ist wie bleibendes 02I-Ib. Umgekehrt war zu schliel~en, dal~ reiner fetaler l~inder- blutfarbstoff sieh nur halb so raseh wie der erwachsener l~inder oxydiert und etwa 1,5real so sehnell wie mensehliehes bleibendes O~Hb.
Das erw~hnte abweichende Verhalten einiger Blutfarbstoffproben yon Sehweinen kSnnte ebenso auf eine Uneinheitliehkeit dieser Blutfarbstoffe hinweisen. Wir haben diese Frage nicht weiter verfolgt. Ebenso kSnnen wit nicht die Besonderheit des Meerschweinchenblutfarbstoffs erkl~ren. Mit Hilfe yon LSslichkeitskurven und der Alkalidenaturierung haben wir keinen Anhalt ffir eine Uneinheitlichkeit des Blutfarbstoffs dieser Tiere gefunden.
Wir hat ten gehofft, da[~ uns die Untersuchung yon COHb noch ein- gehendere Aufsehlfisse fiber die Kinetik des Oxydationsablaufs geben wfirde, weft hierbei die Gesehwindigkeit niedriger ist, also der Vorgang besser studiert werden kann. Diese Hoffnung hat insofern getrogen, als bei der Oxydation yon COHb neue Ph~nomene auftraten, die die Beurtei- lung stSrten. Einmal wich der Kurvenablauf von der monomolekularen Geraden ab : die Werte bis etwa zu 40 sec lagen stets darfiber (Abb. 6), sie sehwenkten dann in den Verlauf der Geraden ein und entfernten sich bei l~ngerer Beobachtung h~ufig wieder etwas davon naeh oben. Zum anderen

218 K. BETKE, I. GREII~ACHER und F. HECKER:
war iiberraschend, da2 eine Mischung aus einem schnell und einem lang- sam reagierenden Blutfarbstoff (Mause- und Menschen-COHb) keinen zusammengesetzten Kurvenverlauf ergab (Abb. 6). Ebensowenig war ein zusammengesetzter Kurvenablauf bei COHb aus I~abelsehnurblutfarb- stoff festzustellen. Die Blutfarbstoffmischungen reagierten sozusagen als ,,Kollektiv" mit einheitlichem Ablauf mitt]erer Geschwindigkeit. Aus der Untersuehung yon Blutfarbstoffproben mit weehselndem Gehalt an fetalem Hb lie2 sich durch Extrapolation schlie2en, dab reines fetales COHb des Menschen etwa 2½real so rasch oxydierbar ist wie Er- wachsenen-COHb 17, was mit den Verhaltnissen bei 02Hb gut iiberein- stimmt.
Woran die Besonderheiten des Oxydationsablaufs bei COttb liegen, ist aus unseren Versuchsergebnissen nicht zu entnehmen. Man muB in Betracht ziehen, dab bei dieser trotz hohen KaFe(CN)6-Ubcrschusses doch ziemlich langsam ablaufenden Reaktion schon eine Oxydationswirkung am EiweiBantefl des Blutfarbstoffs ins Spiel kommt is, 21. Es ware mSglich, dal~ ein derartiger Vorgang fiir die Oxydation des Hb-Eisens neue Verhaltnisse schafft.
Zusammenfassung
Der zeitliche Ablauf der Oxydation yon Oxyhamoglobinen ver- sehiedener Herkunft durch Kaliumferricyanid wurde untersucht. Die Oxydationsgesehwindigkeit hatte fiir jede Tierart eine eharakteristische und reproduzierbare Gr52e. Zwischen den Blutfarbstoffen mehrerer In- dividuen einer Species wurden keine Differenzen gefunden. Von allen bis- her untersuchten Blutfarbstoffen hatte der des erwachsenen Menschen die geringste Oxydationsgeschwindigkeit.
Die Oxydation lief formal wie eine (pseudo-)monomolekulare Reaktion ab. Sie beschleunigte sich mit fallendem pH, steigender Temperatur und steigender K~Fe(CN)6-Konzentration. Die Blutfarbstoffkonzentration und der Anteil an Hamiglobin (Methamoglobin) vor Versuchsbeginn waren ohne Einflul~ auf die Geschwindigkeit.
Der fiir den formalen Ablauf der Oxydation verantwortliche limi- tierende Vorgang diirfte die Dissoziation yon 02Hb zu 02 und Hb se in ; nur sauerstofffreies Hb kann oxydiert werden.
Kiinstliche wie natiirliche Mischungen aus einem schnel] und einem langsam oxydierbaren Blutfarbstoff lieferten bei der Oxydation abge- knickte Kurven, die sich als Summation zweier mit verschiedener Ge- schwindigkeit ablaufender pseudomonomolekularer l%eaktionen inter- pretieren lie~en.
Die Oxydation yon COHb war im gro~en und ganzen der von 02Hb analog. Es waren erheblich hShere K~Fe(CN)6-Mengen erforderlich, um eine fiir die Messungen ausreichend rasche Reaktion zu erhalten. Mischun- gen yon einem schnell und einem langsam oxydierbaren COHb ergaben im Gegensatz zu dem Verhalten yon O2Hb keine Summationskurven.

O x y d a t i o n mensch l , u. t ie r i scher O x y h ~ m o g l o b i n e d u r c h K a l i u m f e r r i c y a n i d 219
L i t e r a t u r
i AI~STIN, J . H. , a n d D. L. DRABKI~: J . o f Biol. Chem. 112, 67 (1935--36) . - - 3 BETXE, K . : N a t u r w i s s e n s c h a f t e n 39, 481 (1952). - - 3 BETKE, K . : K l i n . W s c h r . 1953, 573. - - 4 BETKE, K. : Der mensch l i che ro te Blu t fa rbs tof f , Ber l in -G5t t ingen- Heide lberg : Spr inger 1954. - - ~ BETKE, K . : S y m p o s i o n fiber S t r u k t u r u n d F u n k t i o n der E r y t h r o e y t e n . Ber l in 1955. - - 6 BETKE, K . , u. I. GREINACHER: Kl in . W s c h r . 1955, 611. - - 7 BETKE, K. , u. W. SAVELSBERG: Biochem. Z. 320, 431 (1950). - - s CO,ANT, J . B. : J . o f Biol. Chem. 57, 401 (1923). - - 9 DRI~CKREY, H. , P . DAI~NE- BERG, K. KAISER, J . FROMME U . H . SCHNEIDER: Biochem. Z. 322, 535 (1952). - - 10 I-IALDANE, M.: J . o f Phys io l . 22, 298 (1897--1898) . - - 1 1 HARTRIDGE, H., a n d F. J . W. ROI~GI~TON: Proc. Roy . Soc. Lond . Ser. A. 104, 395 ( 1 9 2 3 ) . - 12 HART- RIDGE, H., a n d F. J . W. ROIIGHTON: Proe. Roy . Soc. Lond . Ser. A, 107, 654 (1925). _ _ 13 HAVEMANN, R. : B iochem. Z. 314, 118 (1943). - - 14HAVEMANN, R. , u. W. HEUB- ~ER: Bioehem. Z. 299, 222 (1938). - - 16 HAVEMA~N, R. , F . JUNG u. B. y o n ISSE- KUTZ: Biochem. Z. 301, 116 (1939). - - 1 ~ HAVEMA2~N, R. , U. K . WOLFF: Biochem. Z. 293, 399 (1937). - - 17 HECKER, F . : I n a u g u r a l - D i s s e r t a t i o n , F re ibu rg 1955. - - is KoBoWlrZ, F. : PersSnl . Mi t te i lung 1955. - - 19 LASZT, L. : He lve t . physiol . A c t a 3, 291 (1945). - - s0 ROCOHTO~, F. J . W. , J . W. LEGGE a n d P. I~ICOLSOI~: H a e m o - g l o b i n - S y m p o s i n m . L o n d o n : B u t t e r w o r t h s Sei. Pub l . 1 9 4 9 . - el SCALER, H . : B iochem. Z. 255, 474 (1932). - - 22 ZEYNEKVO~, R . : Arch . f. Phys io l . 1899, 460.
Doz. Dr . K . BETKE, Fre iburg /Br . , U n i v . - K i n d e r k l i n i k





























