Embed Size (px)
Citation preview
- 97 -
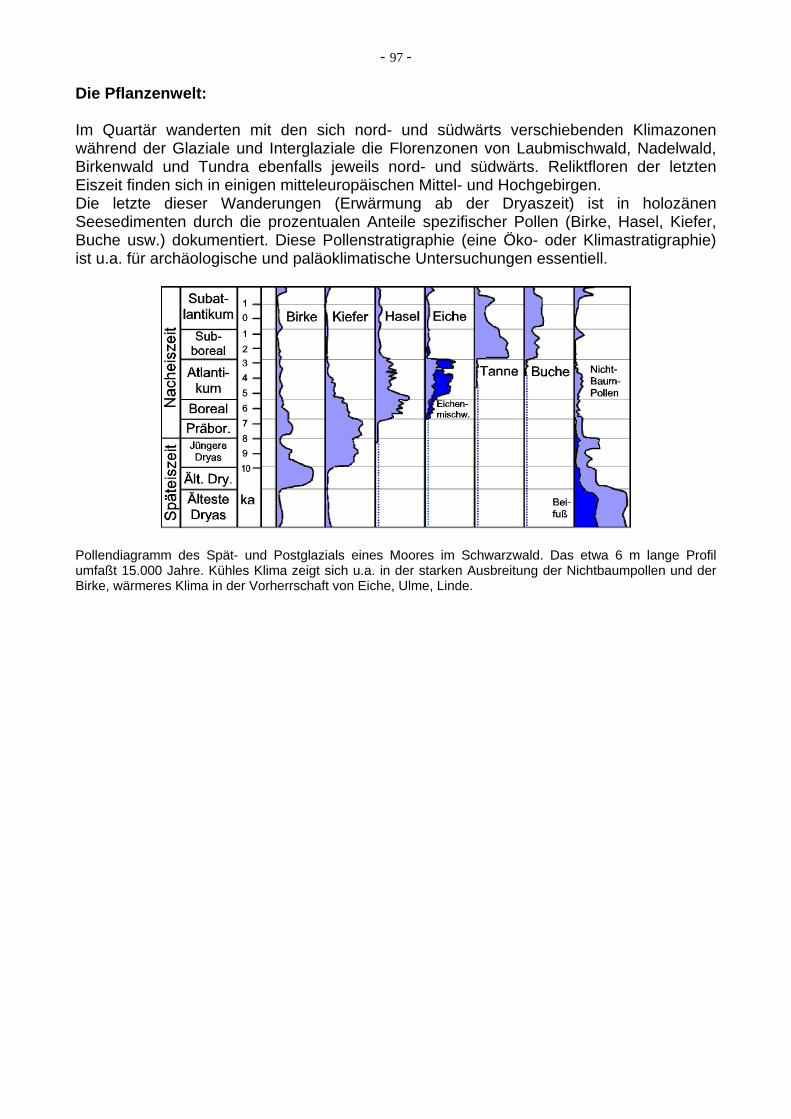
Die Pflanzenwelt: Im Quartär wanderten mit den sich nord- und südwärts verschiebenden Klimazonen während der Glaziale und Interglaziale die Florenzonen von Laubmischwald, Nadelwald, Birkenwald und Tundra ebenfalls jeweils nord- und südwärts. Reliktfloren der letzten Eiszeit finden sich in einigen mitteleuropäischen Mittel- und Hochgebirgen. Die letzte dieser Wanderungen (Erwärmung ab der Dryaszeit) ist in holozänen Seesedimenten durch die prozentualen Anteile spezifischer Pollen (Birke, Hasel, Kiefer, Buche usw.) dokumentiert. Diese Pollenstratigraphie (eine Öko- oder Klimastratigraphie) ist u.a. für archäologische und paläoklimatische Untersuchungen essentiell.
Pollendiagramm des Spät- und Postglazials eines Moores im Schwarzwald. Das etwa 6 m lange Profil umfaßt 15.000 Jahre. Kühles Klima zeigt sich u.a. in der starken Ausbreitung der Nichtbaumpollen und der Birke, wärmeres Klima in der Vorherrschaft von Eiche, Ulme, Linde.

- 98 -
7. Spezialthemen von besonderem erdgeschichtlichem Interesse Im weiten Feld erdgeschichtlicher Fragestellungen und Schwerpunkte sind einige Punkte von besonderem Interesse und werden auch in der Wissenschaftspresse derzeit häufig diskutiert, da hier immer wieder spektakuläre neue Erkenntnisse gewonnen werden. Einige der bekanntesten Beispiele für solche Fragen sind: Themen aus den Bereichen der Geologie, Plattentektonik und Klimaentwicklung: 7.1. Plattentektonik im Archaikum und im Proterozoikum/Phanerozoikum im Vergleich 7.2. Die Entwicklung der Hydrosphäre 7.3. Die Entwicklung der Atmosphäre 7.4. Zusammensetzung der Atmosphäre und Greenhouse/Icehouse-Zyklen im Phanerozoikum 7.5. Temperaturentwicklung im Phanerozoikum 7.6. Eustatische Meeresspiegelschwankungen im Phanerozoikum 7.7. Die Beziehungen zwischen Plattentektonik und Klimaentwicklung 7.8. Eiszeiten in der Erdgeschichte 7.9. Superkontinentzyklen 7.10. Die drei großen Orogenesen in Europa Themen aus dem Bereich der Paläontologie (Lebensgeschichte) 7.11. Die Entstehung des Lebens auf der Erde 7.12. Die Entwicklung der Pflanzenwelt 7.13. Die Entwicklung der Tierwelt 7.14. Massenaussterben im Phanerozoikum 7.1. Plattentektonik im Archaikum und danach im Vergleich Dimensionen und Mechanismen der Plattentektonik waren im Archaikum und im Phanerozoikum (bis heute) sehr verschieden. Mit dem Beginn des Proterozoikums wurden diese den heutigen zumindest deutlich ähnlicher, allerdings meinen manche Wissenschaftler, dass der heutige Typus so ganz erst vor ca. 1 Ga erreicht wurde, also im Übergang bereits zum Neoproterozoikum. Deshalb sei hier im folgenden nur vom archaischen und vom modernen Typus der Plattentektonik gesprochen. Archaische Plattentektonik: In der Frühzeit der Erde dominierten SiO2-arme Magmen sehr niedriger Viskosität. Sie schufen grosse Plateaus und flache Schildvulkane. Evident war die Abwesenheit von großen Kontinentalmassen, stattdessen gab es viele große ozeanische Plateaus und vulkanische Inseln, einen hohen Anteil an submarinen Eruptionen. Pillowlaven, vulkani-sche Brekzien und Abkühlungsrinden waren weitverbreitet. Der Mantel war heißer, die Lithosphäre (mechanisch rigide) dünner. Plumes mit Förderung von Komatiiten waren häufig. Eine Vertikaltektonik war vielleicht wichtiger als horizontale Plattentektonik. Es gab viele kleinere, schnellere Konvektionszellen, mehr Rücken, mehr Subduktionszonen als heute. Die subduzierende Kruste war generell heißer und jünger als heute, schmolz bei geringerer Tiefe, hatte viel Auftrieb, und war wahrscheinlich schwerer zu subduzieren (flache Eintauchwinkel). Es gab nur schlecht begrenzte, nicht-lineare Inselbögen. Aus vielen sehr kleinen Protokontinenten wuchsen im Verlaufe des Archaikums größere Protokontinente zusammen, an ihren Nahtstellen wurden aus Forearc Basin Sedimenten, deformierter ozeanischer Kruste und vulkanischen Inselbögen die sogenannten Greenstone Belts (Grünstein-Gürtel) zusammengeschoben, die heute mit den BIFs (Banded Iron Formations) die wichtigsten Eisenerzlagerstätten der Erde darstellen.
- 99 -
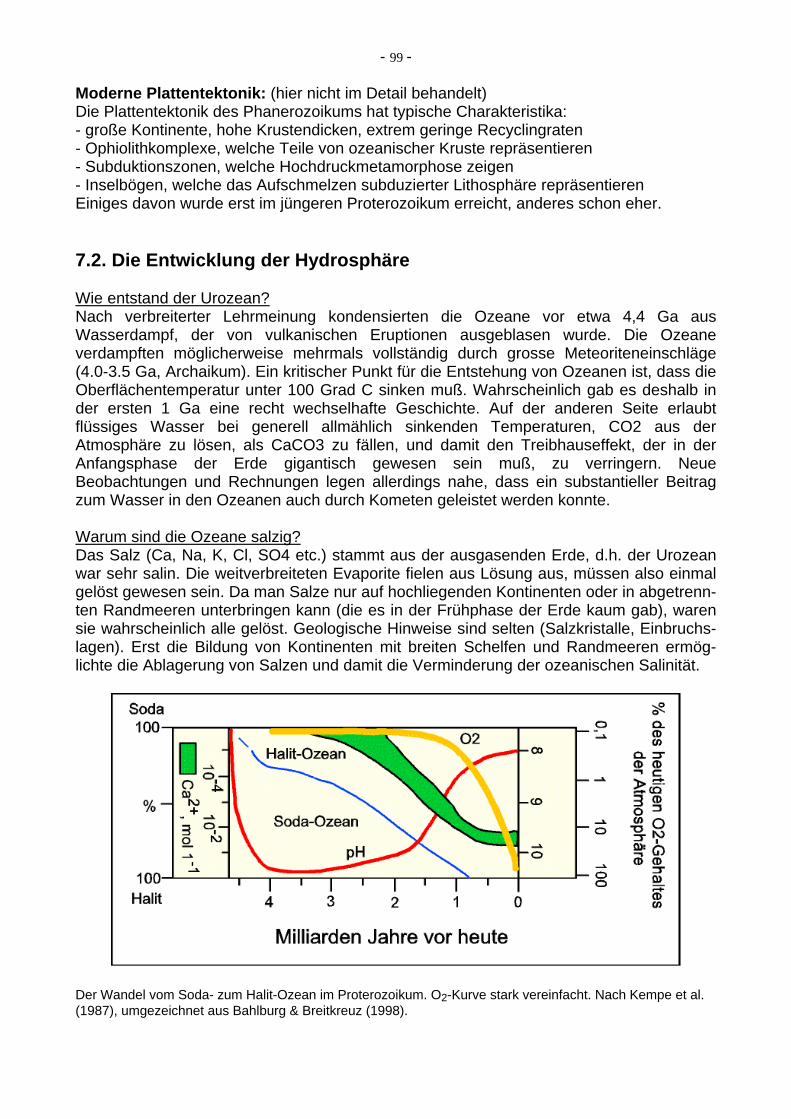
Moderne Plattentektonik: (hier nicht im Detail behandelt) Die Plattentektonik des Phanerozoikums hat typische Charakteristika: - große Kontinente, hohe Krustendicken, extrem geringe Recyclingraten - Ophiolithkomplexe, welche Teile von ozeanischer Kruste repräsentieren - Subduktionszonen, welche Hochdruckmetamorphose zeigen - Inselbögen, welche das Aufschmelzen subduzierter Lithosphäre repräsentieren Einiges davon wurde erst im jüngeren Proterozoikum erreicht, anderes schon eher. 7.2. Die Entwicklung der Hydrosphäre Wie entstand der Urozean? Nach verbreiterter Lehrmeinung kondensierten die Ozeane vor etwa 4,4 Ga aus Wasserdampf, der von vulkanischen Eruptionen ausgeblasen wurde. Die Ozeane verdampften möglicherweise mehrmals vollständig durch grosse Meteoriteneinschläge (4.0-3.5 Ga, Archaikum). Ein kritischer Punkt für die Entstehung von Ozeanen ist, dass die Oberflächentemperatur unter 100 Grad C sinken muß. Wahrscheinlich gab es deshalb in der ersten 1 Ga eine recht wechselhafte Geschichte. Auf der anderen Seite erlaubt flüssiges Wasser bei generell allmählich sinkenden Temperaturen, CO2 aus der Atmosphäre zu lösen, als CaCO3 zu fällen, und damit den Treibhauseffekt, der in der Anfangsphase der Erde gigantisch gewesen sein muß, zu verringern. Neue Beobachtungen und Rechnungen legen allerdings nahe, dass ein substantieller Beitrag zum Wasser in den Ozeanen auch durch Kometen geleistet werden konnte. Warum sind die Ozeane salzig? Das Salz (Ca, Na, K, Cl, SO4 etc.) stammt aus der ausgasenden Erde, d.h. der Urozean war sehr salin. Die weitverbreiteten Evaporite fielen aus Lösung aus, müssen also einmal gelöst gewesen sein. Da man Salze nur auf hochliegenden Kontinenten oder in abgetrenn-ten Randmeeren unterbringen kann (die es in der Frühphase der Erde kaum gab), waren sie wahrscheinlich alle gelöst. Geologische Hinweise sind selten (Salzkristalle, Einbruchs-lagen). Erst die Bildung von Kontinenten mit breiten Schelfen und Randmeeren ermög-lichte die Ablagerung von Salzen und damit die Verminderung der ozeanischen Salinität.
Der Wandel vom Soda- zum Halit-Ozean im Proterozoikum. O2-Kurve stark vereinfacht. Nach Kempe et al. (1987), umgezeichnet aus Bahlburg & Breitkreuz (1998).
- 100 -
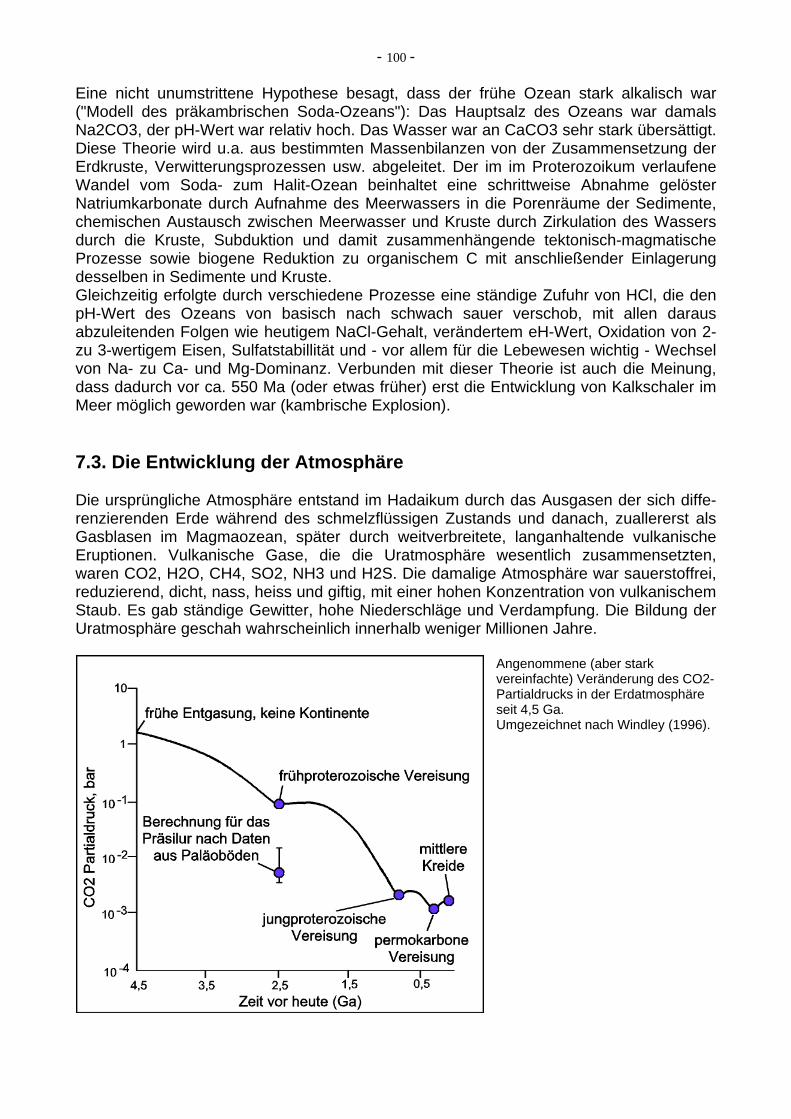
Eine nicht unumstrittene Hypothese besagt, dass der frühe Ozean stark alkalisch war ("Modell des präkambrischen Soda-Ozeans"): Das Hauptsalz des Ozeans war damals Na2CO3, der pH-Wert war relativ hoch. Das Wasser war an CaCO3 sehr stark übersättigt. Diese Theorie wird u.a. aus bestimmten Massenbilanzen von der Zusammensetzung der Erdkruste, Verwitterungsprozessen usw. abgeleitet. Der im im Proterozoikum verlaufene Wandel vom Soda- zum Halit-Ozean beinhaltet eine schrittweise Abnahme gelöster Natriumkarbonate durch Aufnahme des Meerwassers in die Porenräume der Sedimente, chemischen Austausch zwischen Meerwasser und Kruste durch Zirkulation des Wassers durch die Kruste, Subduktion und damit zusammenhängende tektonisch-magmatische Prozesse sowie biogene Reduktion zu organischem C mit anschließender Einlagerung desselben in Sedimente und Kruste. Gleichzeitig erfolgte durch verschiedene Prozesse eine ständige Zufuhr von HCl, die den pH-Wert des Ozeans von basisch nach schwach sauer verschob, mit allen daraus abzuleitenden Folgen wie heutigem NaCl-Gehalt, verändertem eH-Wert, Oxidation von 2- zu 3-wertigem Eisen, Sulfatstabillität und - vor allem für die Lebewesen wichtig - Wechsel von Na- zu Ca- und Mg-Dominanz. Verbunden mit dieser Theorie ist auch die Meinung, dass dadurch vor ca. 550 Ma (oder etwas früher) erst die Entwicklung von Kalkschaler im Meer möglich geworden war (kambrische Explosion). 7.3. Die Entwicklung der Atmosphäre Die ursprüngliche Atmosphäre entstand im Hadaikum durch das Ausgasen der sich diffe-renzierenden Erde während des schmelzflüssigen Zustands und danach, zuallererst als Gasblasen im Magmaozean, später durch weitverbreitete, langanhaltende vulkanische Eruptionen. Vulkanische Gase, die die Uratmosphäre wesentlich zusammensetzten, waren CO2, H2O, CH4, SO2, NH3 und H2S. Die damalige Atmosphäre war sauerstoffrei, reduzierend, dicht, nass, heiss und giftig, mit einer hohen Konzentration von vulkanischem Staub. Es gab ständige Gewitter, hohe Niederschläge und Verdampfung. Die Bildung der Uratmosphäre geschah wahrscheinlich innerhalb weniger Millionen Jahre.
Angenommene (aber stark vereinfachte) Veränderung des CO2-Partialdrucks in der Erdatmosphäre seit 4,5 Ga. Umgezeichnet nach Windley (1996).
- 101 -
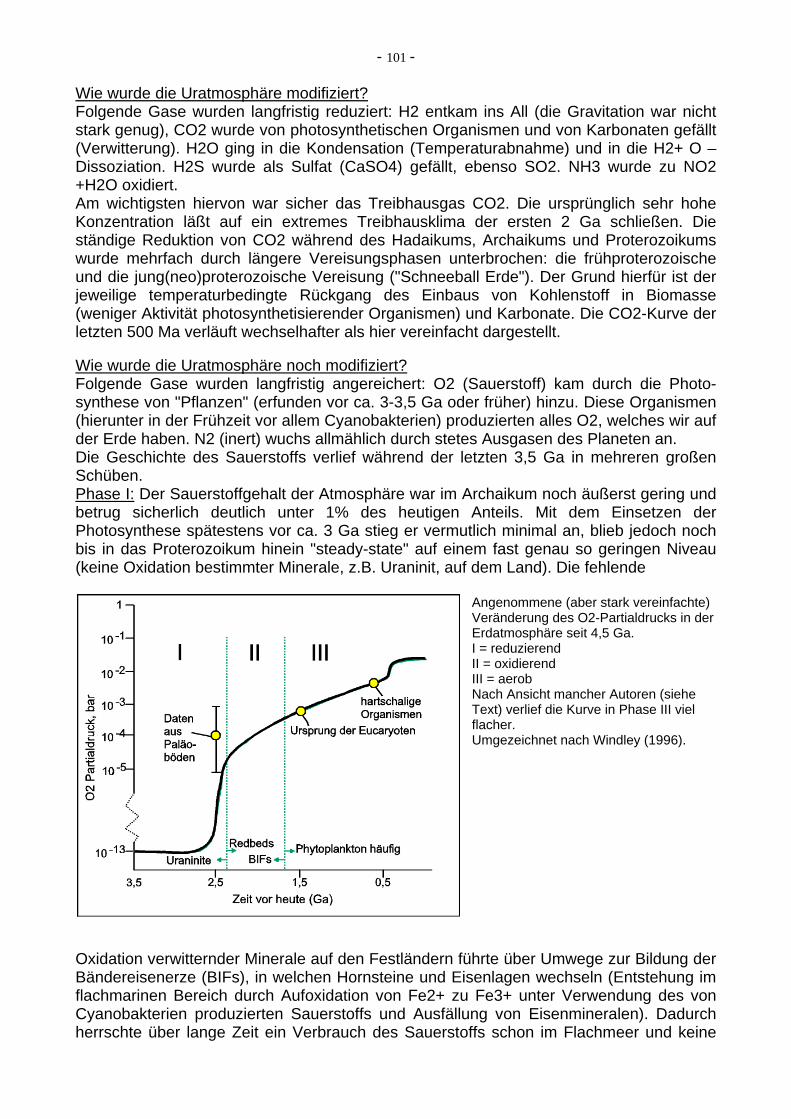
Wie wurde die Uratmosphäre modifiziert? Folgende Gase wurden langfristig reduziert: H2 entkam ins All (die Gravitation war nicht stark genug), CO2 wurde von photosynthetischen Organismen und von Karbonaten gefällt (Verwitterung). H2O ging in die Kondensation (Temperaturabnahme) und in die H2+ O – Dissoziation. H2S wurde als Sulfat (CaSO4) gefällt, ebenso SO2. NH3 wurde zu NO2 +H2O oxidiert. Am wichtigsten hiervon war sicher das Treibhausgas CO2. Die ursprünglich sehr hohe Konzentration läßt auf ein extremes Treibhausklima der ersten 2 Ga schließen. Die ständige Reduktion von CO2 während des Hadaikums, Archaikums und Proterozoikums wurde mehrfach durch längere Vereisungsphasen unterbrochen: die frühproterozoische und die jung(neo)proterozoische Vereisung ("Schneeball Erde"). Der Grund hierfür ist der jeweilige temperaturbedingte Rückgang des Einbaus von Kohlenstoff in Biomasse (weniger Aktivität photosynthetisierender Organismen) und Karbonate. Die CO2-Kurve der letzten 500 Ma verläuft wechselhafter als hier vereinfacht dargestellt. Wie wurde die Uratmosphäre noch modifiziert? Folgende Gase wurden langfristig angereichert: O2 (Sauerstoff) kam durch die Photo-synthese von "Pflanzen" (erfunden vor ca. 3-3,5 Ga oder früher) hinzu. Diese Organismen (hierunter in der Frühzeit vor allem Cyanobakterien) produzierten alles O2, welches wir auf der Erde haben. N2 (inert) wuchs allmählich durch stetes Ausgasen des Planeten an. Die Geschichte des Sauerstoffs verlief während der letzten 3,5 Ga in mehreren großen Schüben. Phase I: Der Sauerstoffgehalt der Atmosphäre war im Archaikum noch äußerst gering und betrug sicherlich deutlich unter 1% des heutigen Anteils. Mit dem Einsetzen der Photosynthese spätestens vor ca. 3 Ga stieg er vermutlich minimal an, blieb jedoch noch bis in das Proterozoikum hinein "steady-state" auf einem fast genau so geringen Niveau (keine Oxidation bestimmter Minerale, z.B. Uraninit, auf dem Land). Die fehlende
Angenommene (aber stark vereinfachte) Veränderung des O2-Partialdrucks in der Erdatmosphäre seit 4,5 Ga. I = reduzierend II = oxidierend III = aerob Nach Ansicht mancher Autoren (siehe Text) verlief die Kurve in Phase III viel flacher. Umgezeichnet nach Windley (1996).
Oxidation verwitternder Minerale auf den Festländern führte über Umwege zur Bildung der Bändereisenerze (BIFs), in welchen Hornsteine und Eisenlagen wechseln (Entstehung im flachmarinen Bereich durch Aufoxidation von Fe2+ zu Fe3+ unter Verwendung des von Cyanobakterien produzierten Sauerstoffs und Ausfällung von Eisenmineralen). Dadurch herrschte über lange Zeit ein Verbrauch des Sauerstoffs schon im Flachmeer und keine
- 102 -
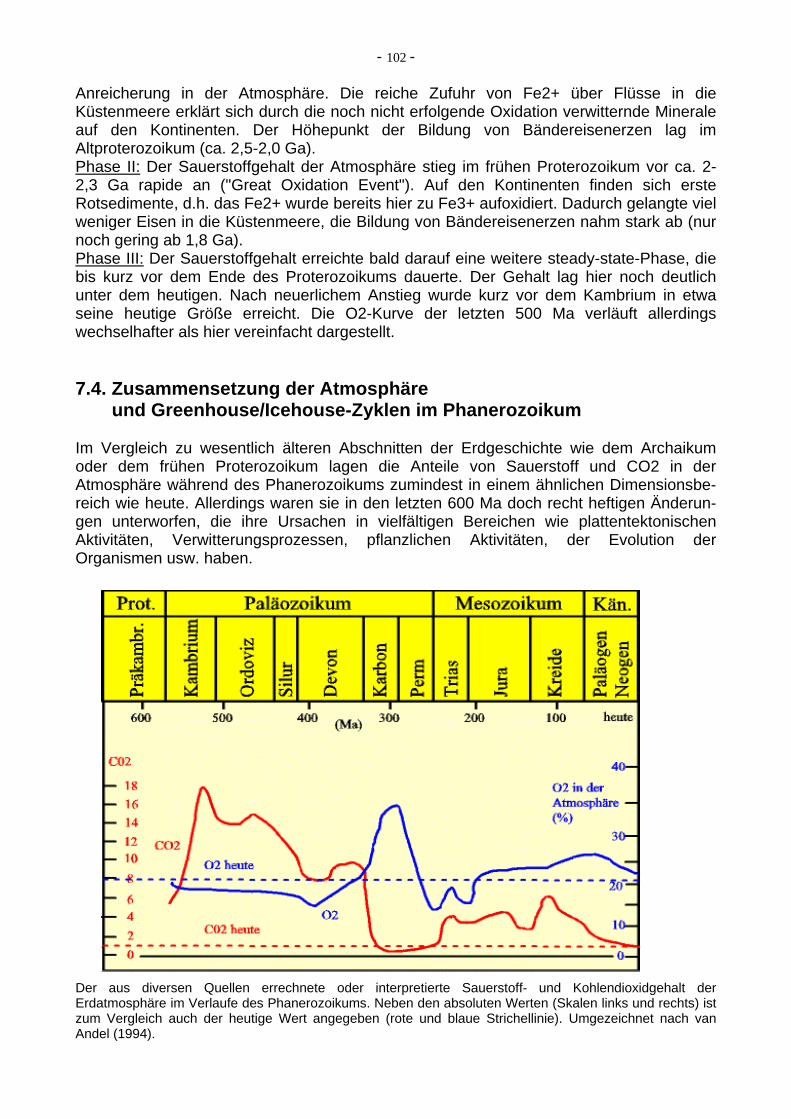
Anreicherung in der Atmosphäre. Die reiche Zufuhr von Fe2+ über Flüsse in die Küstenmeere erklärt sich durch die noch nicht erfolgende Oxidation verwitternde Minerale auf den Kontinenten. Der Höhepunkt der Bildung von Bändereisenerzen lag im Altproterozoikum (ca. 2,5-2,0 Ga). Phase II: Der Sauerstoffgehalt der Atmosphäre stieg im frühen Proterozoikum vor ca. 2-2,3 Ga rapide an ("Great Oxidation Event"). Auf den Kontinenten finden sich erste Rotsedimente, d.h. das Fe2+ wurde bereits hier zu Fe3+ aufoxidiert. Dadurch gelangte viel weniger Eisen in die Küstenmeere, die Bildung von Bändereisenerzen nahm stark ab (nur noch gering ab 1,8 Ga). Phase III: Der Sauerstoffgehalt erreichte bald darauf eine weitere steady-state-Phase, die bis kurz vor dem Ende des Proterozoikums dauerte. Der Gehalt lag hier noch deutlich unter dem heutigen. Nach neuerlichem Anstieg wurde kurz vor dem Kambrium in etwa seine heutige Größe erreicht. Die O2-Kurve der letzten 500 Ma verläuft allerdings wechselhafter als hier vereinfacht dargestellt. 7.4. Zusammensetzung der Atmosphäre und Greenhouse/Icehouse-Zyklen im Phanerozoikum Im Vergleich zu wesentlich älteren Abschnitten der Erdgeschichte wie dem Archaikum oder dem frühen Proterozoikum lagen die Anteile von Sauerstoff und CO2 in der Atmosphäre während des Phanerozoikums zumindest in einem ähnlichen Dimensionsbe-reich wie heute. Allerdings waren sie in den letzten 600 Ma doch recht heftigen Änderun-gen unterworfen, die ihre Ursachen in vielfältigen Bereichen wie plattentektonischen Aktivitäten, Verwitterungsprozessen, pflanzlichen Aktivitäten, der Evolution der Organismen usw. haben. Der aus diversen Quellen errechnete oder interpretierte Sauerstoff- und Kohlendioxidgehalt der Erdatmosphäre im Verlaufe des Phanerozoikums. Neben den absoluten Werten (Skalen links und rechts) ist zum Vergleich auch der heutige Wert angegeben (rote und blaue Strichellinie). Umgezeichnet nach van Andel (1994).

- 103 -
Der Sauerstoffanteil hatte, anders als noch bis vor wenigen Jahren vermutet, wohl schon vor dem Kambrium in etwa das heutige Level erreicht. Er sank dann im Verlaufe des Altpaläozoikums unwesentlich ab, um mit dem Devon und vor allem im Karbon extrem (auf einen vorher und nachher nie erreichten Wert) anzusteigen. Der Grund hierfür liegt im Entstehen der ersten ausgedehnten Wälder auf der Erde und der damit verbundenen Steigerung der photosynthetischen Sauerstoffproduktion. Danach sank der Sauerstoffgehalt der Atmosphäre schnell wieder ab, im Zuge allgemeiner Klimaabkühlungen, zusätzlich aber wohl auch infolge der Entwicklung der sauerstoffzehrenden Tierwelt und der allgemeinen Anpassung der Regelkreisläufe an die "neuen" Quellen. Das folgende Mesozoikum verzeichnet wieder leicht steigende Sauerstoffgehalte (durch Wiederansteigen der Bioproduktion durch vermehrtes Gedeien der Pflanzenwelt in der mesozoischen "Greenhouse"-Ära, in der Kreide wohl auch durch die Neuevolution der effizienteren Angiospermen). Im Verlaufe des Känozoikums sanken die Sauerstoffgehalte infolge der sukzessiven Klimaabkühlungen (känozoische "Icehouse"-Ära) wieder ab. Der Kohlendioxidanteil der Atmosphäre ist besonders stark mit der vulkanischen Entgasung an mittelozeanischen Rücken, also mit der plattentektonischen Aktivität, korreliert. Deshalb stieg er im Zuge des Zerfalls des Rodinia-Superkontinentes zu Beginn des Phanerozoikums zuerst stark an, um anschließend mit dem zunehmenden Aufbau des Pangäa-Superkontinentes in mehreren Stufen wieder abzusinken. Ein besonders starker Abfall ist mit erdgeschichtlich relativ schnellen Aufkommen erster weitausgedehnter Wälder im Karbon und dementsprechend hohem CO2-Entzug aus der Atmosphäre verbunden. Beide Faktoren kamen hier zusammen (wenngleich langfristig natürlich die zur Kernzeit Pangäas geringe plattentektonischen Aktivität die größere Rolle spielte), so dass die Erde anschließend bis in die Trias hinein in eine ausgeprägte (und damit CO2-arme) "Icehouse"-Ära geriet. Im Jura und besonders in der Kreide stieg der CO2-Gehalt wieder an, um, parallel zum O2-Gehalt, im Känozoikum auf das heutige Niveau abzusinken. Eine kurzfristiger Rückgang während des Überganges vom Jura zur Kreide zeigt sich auch in der Temperaturentwicklung.
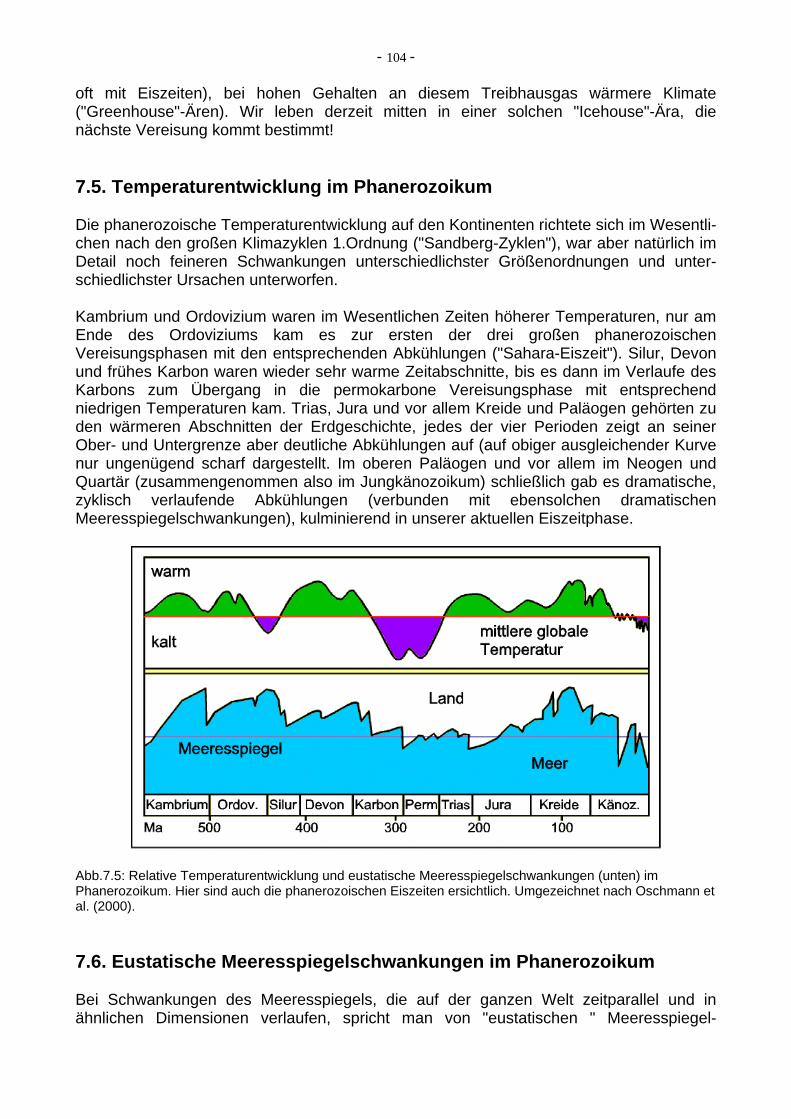
Abb.7.4: Klimayzklen 1. Ordnung ("Sandberg-Zyklen") der letzten 600 Millionen Jahre. Mehrfach wechselten langfristige Kaltphasen (Icehouse) und Warmphasen (Greenhouse). Der CO2-Gehalt der Atmosphäre korreliert sowohl mit den Temperaturen als auch mit der Skelett-Mineralogie einiger Gruppen mariner Kalkschaler. Umgezeichnet nach Sandberg (1983), aus Crowley & North (1991). Die intensiven Beziehungen zwischen Plattentektonik, CO2-Gehalt der Atmosphäre und Temperaturen zeigen sich auch in langfristigen Klimazyklen (nach ihrem Erstautoren auch "Sandberg-Zyklen genannt), die die letzten 600 Ma der Erdgeschichte beherrschten. Bei geringen CO2-Gehalten dominieren kältere Klimate ("Icehouse"-Ären, nicht immer, aber
- 104 -
oft mit Eiszeiten), bei hohen Gehalten an diesem Treibhausgas wärmere Klimate ("Greenhouse"-Ären). Wir leben derzeit mitten in einer solchen "Icehouse"-Ära, die nächste Vereisung kommt bestimmt! 7.5. Temperaturentwicklung im Phanerozoikum Die phanerozoische Temperaturentwicklung auf den Kontinenten richtete sich im Wesentli-chen nach den großen Klimazyklen 1.Ordnung ("Sandberg-Zyklen"), war aber natürlich im Detail noch feineren Schwankungen unterschiedlichster Größenordnungen und unter-schiedlichster Ursachen unterworfen. Kambrium und Ordovizium waren im Wesentlichen Zeiten höherer Temperaturen, nur am Ende des Ordoviziums kam es zur ersten der drei großen phanerozoischen Vereisungsphasen mit den entsprechenden Abkühlungen ("Sahara-Eiszeit"). Silur, Devon und frühes Karbon waren wieder sehr warme Zeitabschnitte, bis es dann im Verlaufe des Karbons zum Übergang in die permokarbone Vereisungsphase mit entsprechend niedrigen Temperaturen kam. Trias, Jura und vor allem Kreide und Paläogen gehörten zu den wärmeren Abschnitten der Erdgeschichte, jedes der vier Perioden zeigt an seiner Ober- und Untergrenze aber deutliche Abkühlungen auf (auf obiger ausgleichender Kurve nur ungenügend scharf dargestellt. Im oberen Paläogen und vor allem im Neogen und Quartär (zusammengenommen also im Jungkänozoikum) schließlich gab es dramatische, zyklisch verlaufende Abkühlungen (verbunden mit ebensolchen dramatischen Meeresspiegelschwankungen), kulminierend in unserer aktuellen Eiszeitphase.
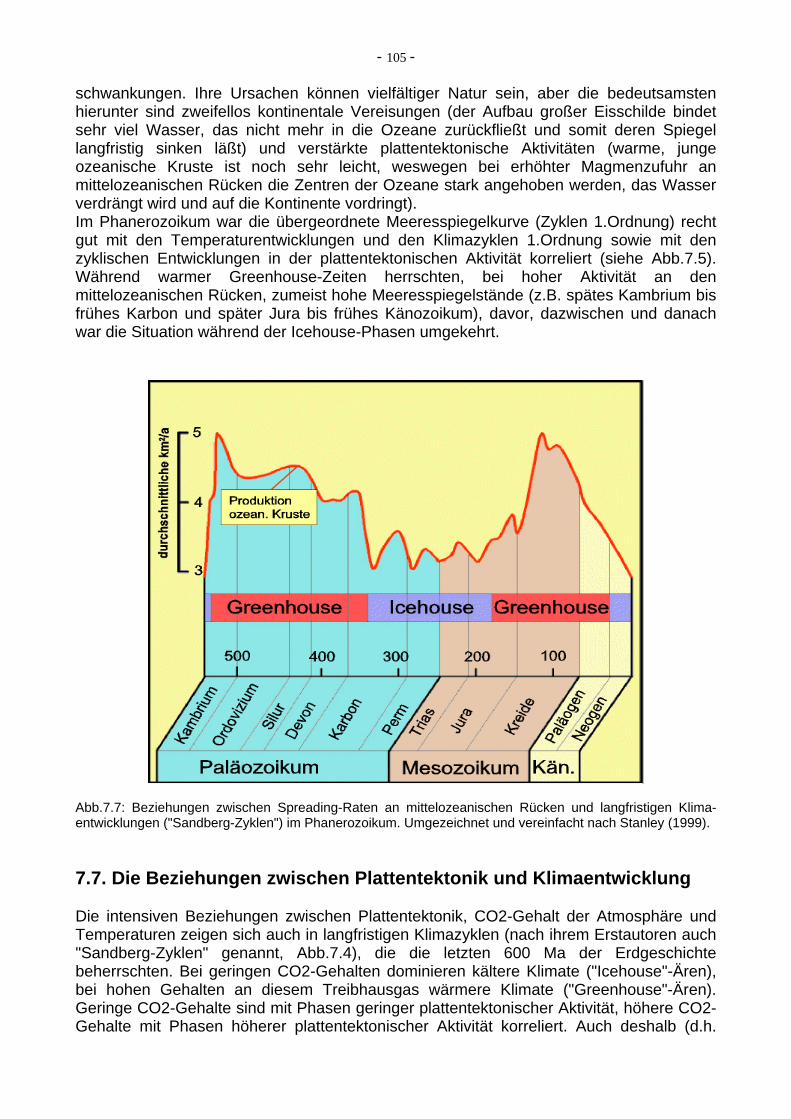
Abb.7.5: Relative Temperaturentwicklung und eustatische Meeresspiegelschwankungen (unten) im Phanerozoikum. Hier sind auch die phanerozoischen Eiszeiten ersichtlich. Umgezeichnet nach Oschmann et al. (2000). 7.6. Eustatische Meeresspiegelschwankungen im Phanerozoikum Bei Schwankungen des Meeresspiegels, die auf der ganzen Welt zeitparallel und in ähnlichen Dimensionen verlaufen, spricht man von "eustatischen " Meeresspiegel-
- 105 -
schwankungen. Ihre Ursachen können vielfältiger Natur sein, aber die bedeutsamsten hierunter sind zweifellos kontinentale Vereisungen (der Aufbau großer Eisschilde bindet sehr viel Wasser, das nicht mehr in die Ozeane zurückfließt und somit deren Spiegel langfristig sinken läßt) und verstärkte plattentektonische Aktivitäten (warme, junge ozeanische Kruste ist noch sehr leicht, weswegen bei erhöhter Magmenzufuhr an mittelozeanischen Rücken die Zentren der Ozeane stark angehoben werden, das Wasser verdrängt wird und auf die Kontinente vordringt). Im Phanerozoikum war die übergeordnete Meeresspiegelkurve (Zyklen 1.Ordnung) recht gut mit den Temperaturentwicklungen und den Klimazyklen 1.Ordnung sowie mit den zyklischen Entwicklungen in der plattentektonischen Aktivität korreliert (siehe Abb.7.5). Während warmer Greenhouse-Zeiten herrschten, bei hoher Aktivität an den mittelozeanischen Rücken, zumeist hohe Meeresspiegelstände (z.B. spätes Kambrium bis frühes Karbon und später Jura bis frühes Känozoikum), davor, dazwischen und danach war die Situation während der Icehouse-Phasen umgekehrt.
Abb.7.7: Beziehungen zwischen Spreading-Raten an mittelozeanischen Rücken und langfristigen Klima-entwicklungen ("Sandberg-Zyklen") im Phanerozoikum. Umgezeichnet und vereinfacht nach Stanley (1999). 7.7. Die Beziehungen zwischen Plattentektonik und Klimaentwicklung Die intensiven Beziehungen zwischen Plattentektonik, CO2-Gehalt der Atmosphäre und Temperaturen zeigen sich auch in langfristigen Klimazyklen (nach ihrem Erstautoren auch "Sandberg-Zyklen" genannt, Abb.7.4), die die letzten 600 Ma der Erdgeschichte beherrschten. Bei geringen CO2-Gehalten dominieren kältere Klimate ("Icehouse"-Ären), bei hohen Gehalten an diesem Treibhausgas wärmere Klimate ("Greenhouse"-Ären). Geringe CO2-Gehalte sind mit Phasen geringer plattentektonischer Aktivität, höhere CO2-Gehalte mit Phasen höherer plattentektonischer Aktivität korreliert. Auch deshalb (d.h.

- 106 -
zusätzlich zu anderen Gründen) korreliert die Kernphase des Pangäa-Superkontinentes, in der die Aktivität an den kaum noch vorhandenen mittelozeanischen Rücken äußerst eingeschränkt war, mit einer langfristigen Icehouse-Ära. Analoges gilt für die weiteren hier dargestellen Greenhouse- und Icehouse-Ären. 7.8. Eiszeiten in der Erdgeschichte Im Verlaufe der Erdgeschichte hat es wiederholt längere Zeitabschnitte gegeben, in denen die klimatischen Bedingungen der Erde oder von Teilen der Erde sich derart veränderten, dass im Winter gefallener Schnee auch im Flachland im Sommer nicht wieder auftaute. Hierdurch bildeten sich weitverbreitet kontinentale Gletscher, diese Phasen werden als Eiszeiten bezeichnet. Bei den bedeutendsten dieser Eiszeiten konnten die Eisschilde mehrere 10er Millionen Quadratkilometer bedecken und mehrere 1000 Meter dick sein, und das (allerdings mit zyklisch wiederholtem Abtauen und Wiederaufbau) über Zeiträume von bis zu mehreren 10er Millionen Jahren. Die wichtigsten 5 seien hier dargestellt: 7.8.1. Die Vereisung im frühen Proterozoikum Erste ausgedehnte Vereisungen gab es im ältesten Proterozoikum vor 2,45-2,2 Ga, offenbar im Zusammenhang mit einer rapiden Abnahme des CO2-Gehaltes der Atmo-sphäre und der damit verbundenen Klimaabkühlung. Diese Eiszeitphase wird nach den am eindrucksvollsten erschlossenen Gletscherablagerungen (jahrezeitliche Bändertone, Moränenablagerungen, "Dropstones", auch Gletscherschrammen) nördlich des Huron-Sees in Südkanada auch als "Huronische Vereisung" bezeichnet. Gleich alte Gletscherablagerungen findet man auch anderswo in Kanada sowie in Wyoming, Finnland, Südafrika und Indien. Die huronische Vereisung scheint also eine weltweite Klimaabkühlung zu repräsentieren. 7.8.2. Die Vereisung im ausgehenden Proterozoikum ("Schneeball Erde") Eine sehr ausgeprägte Icehouse-Ära mit mehreren Vereisungen herrschte am Ende des Proterozoikums, darunter die vermutlich stärkste Eiszeit der Erdgeschichte ("Schneeball Erde") vor ca. 600 Ma. Indizien hierfür sind Tillite (Moränenablagerungen) und Gletscherschrammen, "Dropstones", Bändereisenerze im Meer (kein Sauerstoff unter der Eisdecke des Meeres) und die Tatsache, dass Kohlenstoffisotope in Karbonaten direkt über Tilliten extrem reich an dem leichten Kohlenstoffisotop C12 sind (ein Hinweis auf Zusammenbruch der Bioproduktion). Während der Hauptvereisungen soll ein 1 km dicker Eisschild die Ozeane bedeckt haben (möglicherweise auch die tropischen Meere), minus 50 Grad Celsius, Eis und Wüsten herrschten auf den Kontinenten. Leben konnte sich nur noch in Eisseen oder submarin an heißen Quellen halten. Während der Warmphasen (Auftauen nach vulkanischer CO2-Anrechung) sollen die Temperaturen auf den Kontinenten sich über 50 Grad Celsius erhitzt haben.

- 107 -
Die Gründe für diese extreme Vereisung gerade zu diesem Zeitpunkt sehen die Protagonisten dieser (umstrittenen) Theorie in folgenden Fakten: Der Superkontinent Rodinia zerbrach äquatornah zu vielen kleinen Kontinenten. Dies brachte wegen der größeren Küstennähe der meisen Regionen mehr Regen, der CO2 aus der Atmosphäre auswusch (das Treibhausgas nahm ab, das Klima kühlte sich ab). Polare Meereisschilde wuchsen daraufhin schnell äquatorwärts und erhöhten die Albedo der Erde, was, da ja damals die Sonnenenergie sechs Prozent geringer war als heute, zur Überschreitung eines Schwellwertes und rapiden Vergletscherung der gesamten Erde führte. Die ständig weiter ablaufende vulkanische Entgasung reicherte schließlich nach einiger Zeit dieses Treibhausgas wieder so sehr an, dass Wiedererwärmung und Wiederaufschmelzen der Gletscher einsetzten. Der ganze Ablauf soll sich zyklisch wiederholt haben. Andere Autoren sehen in der Existenz von Vereisungsspuren nahe des damaligen Äquators die Folgen einer extremen Kippung der Erdachse (Pole lagen zur Sonne gerichtet). Auch dies ist aber heftigst umstritten. 7.8.3. Die Vereisung am Ende des Ordoviziums ("Sahara-Eiszeit") Mehrere Millionen Jahre vor dem Ende des Ordoviziums breiteten sich im Bereich des Südpols (damals im Gebiet der heutigen Nordwest-Sahara) auf dem Gondwana-Kontinent Gletscher aus, die im Übergang zum Silur ihr Maximum erreichten. In einem weiten Gebiet von Nordafrika finden sich Tillite (Moränenablagerungen), Gletscherschrammen und eis-verfrachtete Geschiebe ("Dropstones") aus jener Zeit. Weniger häufig sind Spuren auf anderen Kontinenten, die im Falle einiger noch umstrittener Funde aus Nord- und Südamerika etwas jünger (frühes Silur) zu sein scheinen. Diese nach ihrem Haupt-Dokumentationsgebiet auch "Sahara-Eiszeit" genannten Vereisungen waren verbunden mit einem starken welten Meeresspiegelrückgang und einem der fünf großen Massenaussterben, bzw. haben die Vereisungen diese beiden Vorgänge letztlich bedingt. Auch im Falle des oberordovizischen Massenaussterbens scheint dieser Zusammenhang relativ klar zu sein, deswegen ist dieses auch - im Gegensatz zu einigen der anderen großen Massenaussterben - ein relativ langandauernder Vorgang gewesen zu sein. Nicht ganz klar ist allerdings die Ursache für die "Sahara-Eiszeit", denn sie fällt eigentlich in eine ausgeprägte Greenhouse-Ära, die sich über lange Zeiträume (Kambrium bis

- 108 -
Devon) eher durch höhere Temperaturen auszeichnet. Möglicherweise hat die längerandauernde Position des Südpols über einer großen Kontinentalmasse hierzu beigetragen (auf Kontinenten können sich Eisschilde schneller aufbauen), aber ausrei-chend scheint diese Erklärung noch nicht. 7.8.4. Die Vereisung in Karbon und Perm ("Gondwana-Vereisung") Die stärkste und am längsten andauernde Eiszeit des Phanerozoikums war die Vereisung Gondwanas in Karbon und Perm ("permokarbone Vereisung"). In den kältesten Abschnitten war ein großer Teil des Gondwana-Kontinentes (d.h. der südhemisphärischen Hälfte des Superkontinentes Pangäa) vergletschert (über 15 Millionen Quadratkilometer). Bezogen auf die langfristige Klimaentwicklung des Phanerozoikums fällt diese Vereisung, wie es sich "gehört", in eine langandauernde "Icehouse"-Phase. Diese war korreliert mit (und also offenbar bedingt durch) einen/m geringen Gehalt des Treibhausgases CO2 in der damaligen Atmosphäre. Vermutlich war diese Entwicklung sowohl durch die geringe vulkanische Entgasung in dieser Zeit geringer plattentektonischer Aktivität als auch durch das Aufkommen ausgedehnter Wälder und damit verbundenen Kohlenstoffeinbau in pflanzliche Substanz verursacht. Hinzu kam die Lage des damaligen Südpols, der auf der Antarktis, somit auf einem Kontinent (und nicht einem Ozean) lag und somit den Aufbau eines kontinentalen Eisschildes erleicherte. Erste Vergletscherungen hatten sich bereits im Mississippium (Unterkarbon) gebildet, der große Eisschild entstand jedoch erst im Pennsylvanium (Oberkarbon). Der Höhepunkt der Vereisungen war etwa an der Wende Karbon/Perm erreicht, Spuren der Gletscher (Morä-nenablagerungen, gekritzte Geschiebe, Gletscherschrammen usw.) finden sich auf allen heutigen Südkontinenten.
Ausdehnung der Inlandseismassen auf dem Südteil von Gondwana etwa an der Karbon / Perm - Wende. Die Lage der heutigen Kontinente ist skizziert. Die Pfeile geben eine repräsentative Auswahl geologisch dokumentierter Bewegungsrichtungen an. Umgezeichnet und verändert nach Seyfert & Sirkin (1979) aus Faupl (2000). Höheres Karbon und tieferes Perm sind als Hauptzeiten der permokarbonen Vereisung weltweit durch Sedimentationsrhythmen geprägt, die über Meeresspiegeltrans- und -

- 109 -
regressionen (bedingt durch Aufbau und Abschmelzen der riesigen Gletschermassen) regelmäßige Klimazyklen widerspiegeln. Die bekanntesten dieser Zyklen sind die soge-nannten "Kohlezyklen" des Pennsylvanium in Nordamerika und Europa. Im Verlaufe des frühen Perm gingen die Vereisungen der Südkontinente allmählich wieder zu Ende. Das Oberperm war weitestgehend eisfrei. 7.8.5. Die Vereisung im Jungkänozoikum (heutige Eiszeit)
Ozeanographische und klimatische Auswirkungen der Schließung des Isthmus von Panama zwischen Nord- und Südamerika. Die Zahlen geben den Salzgehalt des Meerwassers in Promille an. Umgezeichnet und verändert nach Stanley (1999).
Das ozeanische Strömungssystem, wie es mit dem thermohalinen Zirkulationsband heute vorherrscht (Abb.7.8.5: rechts) unterscheidet sich wesentlich von jenem, welches noch im frühen Pliozän (Abb.7.8.5: links) herrschte, bevor die Vereisung der nördlichen Hemi-sphäre begann. Heute bringen die westlichen Passatwinde Feuchtigkeit in den Pazifik, welche sie vorher in dem erhitzten Westatlantik (Karibik) evaporiert hatten. Dadurch wird das Atlantikwasser sehr salzreich und schwer, sodass es nach Transport in Nordostrichtung (Nordatlantik- oder Golfstrom) bei noch weiterer Abkühlung bereits im Raum zwischen Grönland und Norwegen so schwer wird, dass es absinkt und sauerstoffreiches Tiefenwasser bildet. Dadurch gelangen warme Strömungen nicht mehr so weit nach Norden wie noch im frühen Pliozän, als warme und nicht so salzreich (deshalb nicht so schwere) Ozeanwässer in das arktische Meer gelangen und den arktischen Raum warm halten konnten. Die Schließung des Isthmus von Panama (ein plattentektonischer Vorgang) hat also den Startschuß für die Vereisung der Nordhemisphäre gegeben. Der Schwellwert hierfür war vor 4,6 Millionen Jahren erreicht. Das Schwanken der Sonneneinstrahlung aufgrund überlagerter zyklisch schwankender orbitaler Parameter (Lage der Erdachse 23 ka, Schiefe der Erdachse 40 ka, Exztrenzitität der Erdumlaufbahn 100 ka) bewirkte besonders im Quartär (letzte 1,75 Ma) Aufbau und Abschmelzen großer nordhemisphärischer Eisschilde. Dies ist der Grund, dass erst lange nach dem Erreichen des Schwellwertes bei 4,6 Ma (Schließung des Panamaseeweges, s.o.) die Bedingungen zum Aufbau der ersten großen Eisschilde auch im Norden im Zusammenhang mit den orbitalen Parametern vor ca. 3 Ma erreicht waren.
- 110 -
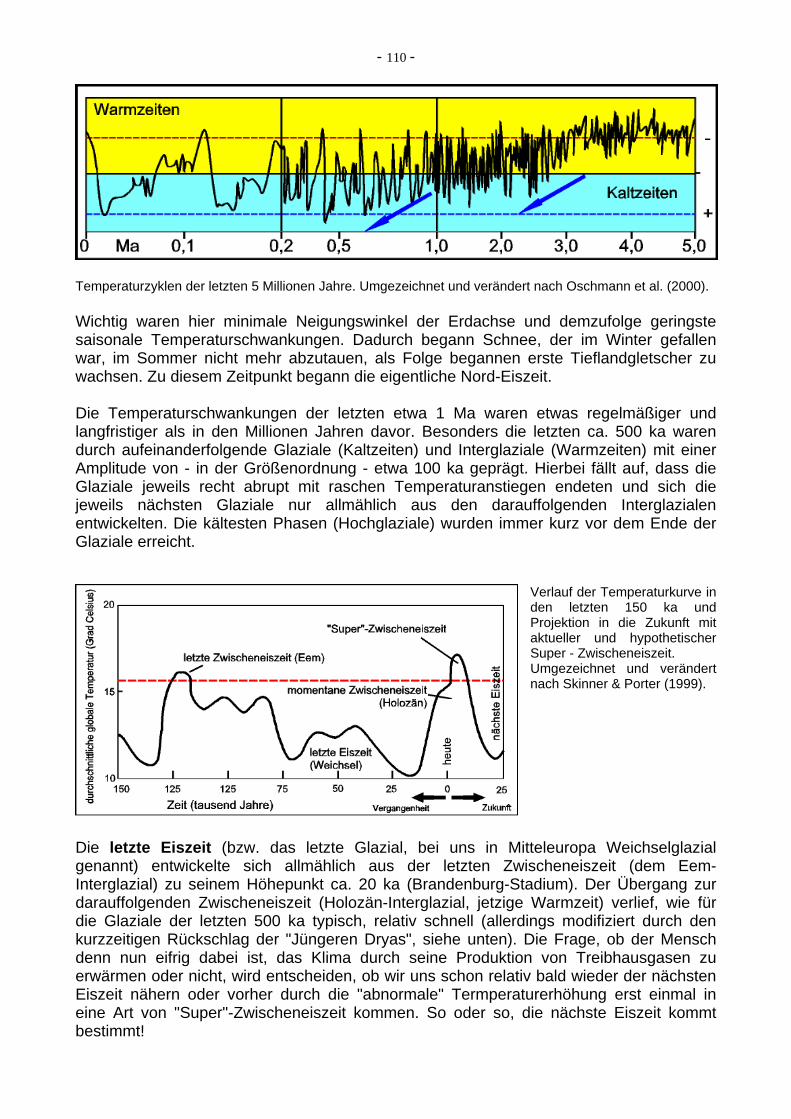
Temperaturzyklen der letzten 5 Millionen Jahre. Umgezeichnet und verändert nach Oschmann et al. (2000). Wichtig waren hier minimale Neigungswinkel der Erdachse und demzufolge geringste saisonale Temperaturschwankungen. Dadurch begann Schnee, der im Winter gefallen war, im Sommer nicht mehr abzutauen, als Folge begannen erste Tieflandgletscher zu wachsen. Zu diesem Zeitpunkt begann die eigentliche Nord-Eiszeit. Die Temperaturschwankungen der letzten etwa 1 Ma waren etwas regelmäßiger und langfristiger als in den Millionen Jahren davor. Besonders die letzten ca. 500 ka waren durch aufeinanderfolgende Glaziale (Kaltzeiten) und Interglaziale (Warmzeiten) mit einer Amplitude von - in der Größenordnung - etwa 100 ka geprägt. Hierbei fällt auf, dass die Glaziale jeweils recht abrupt mit raschen Temperaturanstiegen endeten und sich die jeweils nächsten Glaziale nur allmählich aus den darauffolgenden Interglazialen entwickelten. Die kältesten Phasen (Hochglaziale) wurden immer kurz vor dem Ende der Glaziale erreicht.
Verlauf der Temperaturkurve in den letzten 150 ka und Projektion in die Zukunft mit aktueller und hypothetischer Super - Zwischeneiszeit. Umgezeichnet und verändert nach Skinner & Porter (1999).
Die letzte Eiszeit (bzw. das letzte Glazial, bei uns in Mitteleuropa Weichselglazial genannt) entwickelte sich allmählich aus der letzten Zwischeneiszeit (dem Eem-Interglazial) zu seinem Höhepunkt ca. 20 ka (Brandenburg-Stadium). Der Übergang zur darauffolgenden Zwischeneiszeit (Holozän-Interglazial, jetzige Warmzeit) verlief, wie für die Glaziale der letzten 500 ka typisch, relativ schnell (allerdings modifiziert durch den kurzzeitigen Rückschlag der "Jüngeren Dryas", siehe unten). Die Frage, ob der Mensch denn nun eifrig dabei ist, das Klima durch seine Produktion von Treibhausgasen zu erwärmen oder nicht, wird entscheiden, ob wir uns schon relativ bald wieder der nächsten Eiszeit nähern oder vorher durch die "abnormale" Termperaturerhöhung erst einmal in eine Art von "Super"-Zwischeneiszeit kommen. So oder so, die nächste Eiszeit kommt bestimmt!

- 111 -
Das Ende der letzten Eiszeit verlief nicht ohne Komplikationen. Nachdem sich das Klima vor ca. 13 ka schon relativ stark erwärmt hatte ("Dryas-Zeit", benannt nach der sich nun ausbreitenden Silberwurz), zeigen die an vielen Orten der Nordhemisphäre unabhängig voneinander noch einmal die Rückkehr zu kühleren Temperaturen an ("Jüngere Dryas vor ca. 11-12 ka), bis dann mit einer abrupten Erwärmung der endgültige Übergang zum Holozän erfolgte. Für diesen raschen Klimawechsel werden Extremwerte von 7 Grad Celsius in nur 40 Jahren (oder gar noch schneller) genannt, was alle für die derzeitig von uns Menschen gemachte Erwärmung gemessene oder vorhergesagte Geschwindigkeiten bei weitem überschreitet. Der Grund hierfür kann nur eine plötzliche Änderung des nordatlantischen Strömungssystems sein. Offenbar sprang das System nach Überschreiten eines Schwellwertes in einen anderen stabilen Zustand um, der uns seitdem mehr Wärme in das Nordmeer liefert.
Die "Jüngere Dryas" vor ca. 11-12 ka. Relativer Temperaturverlauf nach Analysen der stabilen Sauerstoffisotope in Sedimenten eines schweizer Sees und eines grönländischen Eiskerns. Umgezeichnet und verändert nach Skinner & Porter (1999).
Den Grund hierfür sieht man mittlerweile recht einvernehmlich in dem (wiederholten?) katastrophalen Ausfließen großer Eisstauseen ("Lake Agassiz", heutige Große Seen, allerdings mit vielfacher Ausdehnung) am Südrand des nach Norden zurückweichenden kanadischen Eisschildes. Hierdurch wurde das aus der Karibik stammende salzreiche Wasser des Nordatlantikstroms (fälschlich auch Golfstrom genannt) so stark verdünnt, dass das thermohaline Strömungssystem zusammenbrach. Dadurch kühlte der Nordatlantik ab, und hier entstehende kühle Winde ließen die Temperaturen in Europa, aber auch auf Grönland wieder sinken. Nach weiterem Abtauen der kanadischen Gletscher ließ dieser Effekt nach, und es kam zu der beschriebenen abrupten Erwärmung im Übergang zum Holozän. 7.9. Superkontinentzyklen Das Phanerozoikum (die letzten 540 Ma der Erdgeschichte) war beherrscht von einem plattentektonischen Großzyklus, in dem der Superkontinent Pangäa sich im Verlaufe des Paläozoikums aufbaute und im Verlaufe des Mesozoikums und Känozoikums allmählich wieder zerfiel. Beginn und Ende eines solchen "Superkontinent-Zyklus" genannten Vor-ganges sind nicht genau zu definieren, aber man muß wohl eine zeitliche Größenordnung von 400-500 Ma für derartige Phänomene annehmen. Heute glaubt man, dass sich in der Erdgeschichte mindestens fünf solcher Zyklen abspielten und wir momentan eigentlich schon in der Frühphase eines sechsten leben. Vier davon (das ist allerdings umstritten)

- 112 -
sollen im Proterozoikum abgelaufen sein. Deren jüngster (der Rodinia (Vendium) – Super-kontinentzyklus) reichte bis in das Kambrium hinein und ist unstrittig. Am bekanntesten ist der der Pangäa - Superkontinentzyklus des Phanerozoikums. Der zukünftige Superkontinentzyklus wird in ca. 250-300 Ma wieder zu einem neuen Superkontinent führen, der von manchen als "Pangaea ultima" bezeichnet wird. Der Motor für diese Zyklizität liegt im Erdinneren und in einigen logischen Zwängen aus der Plattentektonik: Wenn kontinentale Platten nur lange genug über die Erde wandern, muß es irgendwann zur weitestgehenden Vereinigung dieser Platten zu einem Superkontinent kommen, denn kontinentale Kruste wird normalerweise nicht subduziert und aufgeschmolzen - wie die ozeanische Kruste - sondern dem Kontinent, mit dem die Kollision erfolgt, angegliedert. So müssen Kontinente einfach langfristig wachsen, und ein großer Superkontinent muß entstehen. Ist dies einmal geschehen, kommt die Tatsache zum Tragen, dass die Erde ja ständig Wärme nach außen abgibt und, da kontinentale Lithosphäre ein schlechterer Wärmeleiter ist als ozeanische Lithosphäre, sich unter einer großen kontinentalen Platte ein Wärmestau bilden muß. Hierauf muß der Mantel mit der Ausbildung eines Konvek-tionssystems reagieren, welches unter dem Superkontinent aufsteigendes Mantelmaterial seitlich nach außen führt – der Kontinent bricht auseinander. 7.9.1. Der Pangäa - Superkontinentzyklus Das Phanerozoikum war beherrscht von einem plattentektonischen Großzyklus, in dem der Superkontinent Pangäa sich im Verlaufe des Paläozoikums aufbaute und im Verlaufe des Mesozoikums und Känozoikums allmählich wieder zerfiel.

- 113 -
Dieser Zyklus steuerte langfristig die klimatische Entwicklung und die Meeresspiegelkurve, somit auch die Evolution der Organismen bis hin zu den Wirbeltieren. Im Verlaufe des Paläozoikums wurden durch aufeinanderfolgende Orogenesen (die kaledonische und die variszische im europäischen Raum) die Kontinente Gondwana, Sibiria, Baltica und Laurentia sowie weitere kleinere Krustenstücke zu dem Superkontinent zusammengeschweißt, der im Perm und in der Trias seine Kernphase hatte. Schon in der Obertrias begann erstes Rifting zwischen Nordwestafrika und Nordamerika, und im Jura setzte der Zerfall Pangäas mit der Entstehung des Zentralatlantiks endgültig ein. Heute leben wir in einer Übergangsphase, eigentlich hat der nächste Superkontinentzyklus bereits begonnen. 7.9.2. Der zukünftige Superkontinentzyklus Aus den derzeitigen Bewegunsrichtung und –geschwindigkeiten der Lithosphärenplatten läßt sich die plattentektonische Zukunft der Erde für einige hundert Millionen Jahre voraus-berechnen. (alle Abb. umgezeichnet und vereinfacht nach http://www.scotese.com)
Die Erde in 50 Millionen Jahren. Der Atlantik wird sich zuerst noch aus-weiten, allerdings wird an seinem Westrand bereits eine Subduktions-zone entstehen. Die fortschreitende Kollision von Afrika und Europa (alpi-dische Gebirgsbildung) wird das Mittelmeer vollständig schließen und das bis Asien hineinreichende "mediterrane Gebirge" (entsprichten den Alpiden) vergrößern. Die Erde in 150 Millionen Jahren. Das Mittelmeer wird endgültig verschwunden, Afrika und Eurasien zu einem Kontinent vereinigt sein. Südatlantik und Indik werden zusammen einen neuen Ozean bilden. Australien wird sich wieder mit der Antarktis vereinigen. Der mittelatlantische Rücken wird fast ganz verschwunden sein, der Atlantik wieder schmaler werden. An seinem Westrand wird er unter Amerika subduziert. Die Erde in 250 Millionen Jahren. Der Atlantik ist völlig verschwunden, Amerika, Afrika und Eurasien haben sich wieder zu einem Superkontinent vereinigt, der ein großes Binnenmeer umschließt. Scotese (siehe Internet-Quelle) bezeichnet diesen Kontinent als "Pangea ultima".

- 114 -
7.10. Die drei großen Orogenesen in Europa 7.10.1. Die kaledonische Orogenese (Kambrium bis Devon) Die Kaledoniden stellen ein Gebirgssystem des Altpaläozoikums dar, das aus der ersten der drei großen phanerozoischen Orogenesen Europas entstand. Heute besteht es aus Teilen von Spitzbergen, Grönland, Norwegen, Westschweden, Schottland, Wales, Irland, den Appalachen und - allerdings verborgen unter sehr viel jüngeren Sedimenten - Gebieten unter der Nordsee, Nordeutschland und Polen. "Caledonia" ist der keltische Name für Nordschottland, eines der wichtigsten Gebiete der Kaledoniden. Geschaffen wurde das kaledonische Gebirge durch die Kollision der Kontinente Laurentia und Baltica sowie von kleineren, vom Nordrand des großen Südkontinentes Gondwana abgelösten Krustensplittern ("perigondwanische Terranes"). Zwischen Laurentia und Baltica lag ursprünglich der Iapetus-Ozean, der durch die kaledonische Orogenese geschlossen wurde. Auch nördliche Teile des Rhea-Ozeans (auch Proto-Tethys genannt) wurden mit einbezogen, aber die Hauptrolle spielte dieser Ozean erst später bei der variszischen Orogenese. Während der Hauptphasen der Kaledoniden im Silur wurden Teile der Gebirge nach Westen gegen Laurentia (Westvergenzen in Abb.unten) und Teile nach Osten gegen Baltica (Ostvergenzen in Abb.unten) bewegt.
Das Gebirgssystem der Kaledoniden und die vorherrschenden tektonischen Transportrichtungen (Vergenzen, Pfeile). Umgezeichnet nach Faupl (2000).
Die ältesten Sedimente, die später in den Kaledoniden verfaltet wurden, waren bereits im Proterozoikum abgelagert worden, und zwar im Zuge des Zerfalls des Rodinia-Superkon-tinentes. Im Kambrium begann dann die vermehrte Bildung ozeanischer Kruste des Iapetus-Ozeans, der sich bis in das Ordovizium hinein verbreiterte. Gleichzeitig kam es aber auch schon zur Subduktion ozeanischer Kruste im Westen, später (im Ordovizium) auf beiden Seiten. Im späteren Ordovizium (Caradoc) wurde der Iapetus-Ozean bereits wieder schmaler, im Silur schloß er sich unter Vereinigung von Baltica und Laurentia zum Old-Red-Kontinent (auch Eurameria oder Laurussia genannt) schließlich ganz.
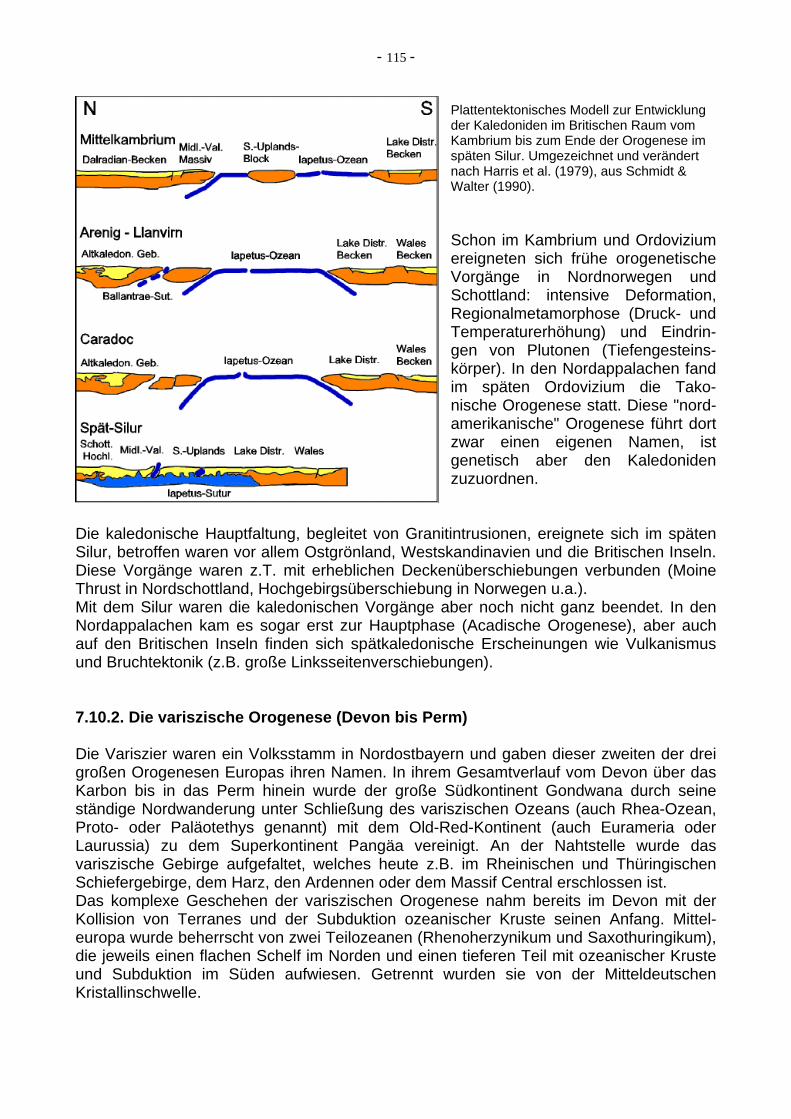
- 115 -
Plattentektonisches Modell zur Entwicklung der Kaledoniden im Britischen Raum vom Kambrium bis zum Ende der Orogenese im späten Silur. Umgezeichnet und verändert nach Harris et al. (1979), aus Schmidt & Walter (1990). Schon im Kambrium und Ordovizium ereigneten sich frühe orogenetische Vorgänge in Nordnorwegen und Schottland: intensive Deformation, Regionalmetamorphose (Druck- und Temperaturerhöhung) und Eindrin-gen von Plutonen (Tiefengesteins-körper). In den Nordappalachen fand im späten Ordovizium die Tako-nische Orogenese statt. Diese "nord-amerikanische" Orogenese führt dort zwar einen eigenen Namen, ist genetisch aber den Kaledoniden zuzuordnen.
Die kaledonische Hauptfaltung, begleitet von Granitintrusionen, ereignete sich im späten Silur, betroffen waren vor allem Ostgrönland, Westskandinavien und die Britischen Inseln. Diese Vorgänge waren z.T. mit erheblichen Deckenüberschiebungen verbunden (Moine Thrust in Nordschottland, Hochgebirgsüberschiebung in Norwegen u.a.). Mit dem Silur waren die kaledonischen Vorgänge aber noch nicht ganz beendet. In den Nordappalachen kam es sogar erst zur Hauptphase (Acadische Orogenese), aber auch auf den Britischen Inseln finden sich spätkaledonische Erscheinungen wie Vulkanismus und Bruchtektonik (z.B. große Linksseitenverschiebungen). 7.10.2. Die variszische Orogenese (Devon bis Perm) Die Variszier waren ein Volksstamm in Nordostbayern und gaben dieser zweiten der drei großen Orogenesen Europas ihren Namen. In ihrem Gesamtverlauf vom Devon über das Karbon bis in das Perm hinein wurde der große Südkontinent Gondwana durch seine ständige Nordwanderung unter Schließung des variszischen Ozeans (auch Rhea-Ozean, Proto- oder Paläotethys genannt) mit dem Old-Red-Kontinent (auch Eurameria oder Laurussia) zu dem Superkontinent Pangäa vereinigt. An der Nahtstelle wurde das variszische Gebirge aufgefaltet, welches heute z.B. im Rheinischen und Thüringischen Schiefergebirge, dem Harz, den Ardennen oder dem Massif Central erschlossen ist. Das komplexe Geschehen der variszischen Orogenese nahm bereits im Devon mit der Kollision von Terranes und der Subduktion ozeanischer Kruste seinen Anfang. Mittel-europa wurde beherrscht von zwei Teilozeanen (Rhenoherzynikum und Saxothuringikum), die jeweils einen flachen Schelf im Norden und einen tieferen Teil mit ozeanischer Kruste und Subduktion im Süden aufwiesen. Getrennt wurden sie von der Mitteldeutschen Kristallinschwelle.

- 116 -
Modell zur Entwicklung der mitteleuropäischen Varisziden im Devon. Umgezeichnet nach Franke (1989) aus Faupl (2000).
Die mit diesen Subduktionen und Kollisionen kleinerer Platten verbundenen frühvariszischen Phasen werden als reussisch und bretonisch bezeichnet. Typisch war auch ein frühorogener submariner basischer Vulkanismus, dessen Ergebnisse heute vielerorts als Diabase für den Straßenbau abgebaut werden.
Prinzipieller Ablauf der tektonischen und magmatischen Ereignisse während der variszischen Gebirgsbildung in Mitteleuropa. Zacken = tektonische "Phasen". Schwarz = basischer Vulkanismus. Weiß mit Signatur = saurer Vulkanismus und Plutonismus. Die variszische Hauptfaltung (sudetische Phase) ereignete sich in Europa etwa an der Wende Mississippium / Pennsylvanium (bzw. Unter/Oberkarbon). Sie war mit intensiven Verfaltungen (nördlich der Mitteldeutschen Kristallinschwelle meist NW-vergent, südlich davon meist SE-vergent) und der Intrusion großer Tiefengesteinskörper (saure Plutonite, z.B. Brocken im Harz) verbunden und ging in die erzgebirgische Phase über. Wie in den Kaledoniden (beachten Sie überhaupt die z.T. ähnlichen Abfolgen, fast mag man von einer Regelhaftigkeit in der Entwicklungsgeschichte von Orogenen sprechen) folgten noch über längere Zeit einige Spätphasen (asturisch, saalisch), die von (meist saurem) Vulkanismus und intensiver Bruchtektonik begleitet wurden.

- 117 -
Ebenfalls sehr regelhaft entwickelt ist in den Varisziden die Abfolge typischer Sedimente vom Flysch (Wechsellagerung feinkörniger Beckenablagerungen mit von den Schelfkanten ins tiefere Becken abgerutschten Trübeströmen, sogenannten "Turbiditen") zur Molasse (Abtragunsschutt der bereits aufgefalteten Gebirge). Der variszische Flysch wird z.B. durch die Tonschiefer/Grauwacken-Wechsellagerungen des Mississippium in Kulm-Fazies, die Molasse durch die klastischen Sedimente des Perm in Rotliegendfazies repräsentiert.
Geologischer Schnitt von NW (links) nach SE (rechts) durch das variszische Gebirge in Mitteleuropa mit den Kossmat' schen Zonen. KTB = Kontinentale Tiefbohrung. Umgezeichnet nach Bahlburg & Breitkreuz (1998). Diese Abbildung zeigt das Endergebnis der variszischen Orogenese in Mitteleuropa. Das komplizierte Bild mit zahlreichen Unterschiebungen und Deckenbahnen (enorme Krustenverkürzung!) ist offensichtlich. Sie zeigt auch eine seit der ersten Hälfte des letzten Jahrhunderts (nach Kossmat) übliche Gliederung des Variszikums von NW (links) nach SE (rechts) in breite, SW-NE-verlaufende Zonen jeweils ähnlicher geologischer Geschichte, tektonischen und sedimentologischen Inventars, Metamorphosegrade und Durchsetzung mit Plutoniten: - Das Subvariszikum besteht fast ausschließlich aus nach Norden vorgeschütteter Molasse des von Süden heranrückenden variszischen Gebirges, hierin auch die wichtigsten großen Steinkohlenvorkommen in Frankreich, Belgien, Deutschland und Polen. - Das Rhenoherzynikum mit einer Vielfalt an Sedimenten meist geringeren Metamorphosegrades, überwiegender NW-Vergenz der Faltung und geringem Anteil an sauren Plutonen (z.B. Brocken) ist heute z.B. in den Ardennen, der Eifel, dem Sauerland und dem Harz erschlossen. - Im Saxothuringikum (heute z.B. Frankenwald, Thüringer Schiefergebirge, Spessart, Odenwald) waren die Faltungen (meist SE-vergent) bereits vor dem Pennsylvanium abgeschlossen, es gibt kein marines höheres Karbon. Die Metamorphosegrade sind oft etwas größer, die Plutonite häufiger, der Deckenbau ist intensiver. - Das Moldanubikum (Böhmische Masse, Schwarzwald, Vogesen u.a.) ist sehr stark metamorph und von zahlreichen Tiefengesteinskörpern durchsetzt.
- 118 -
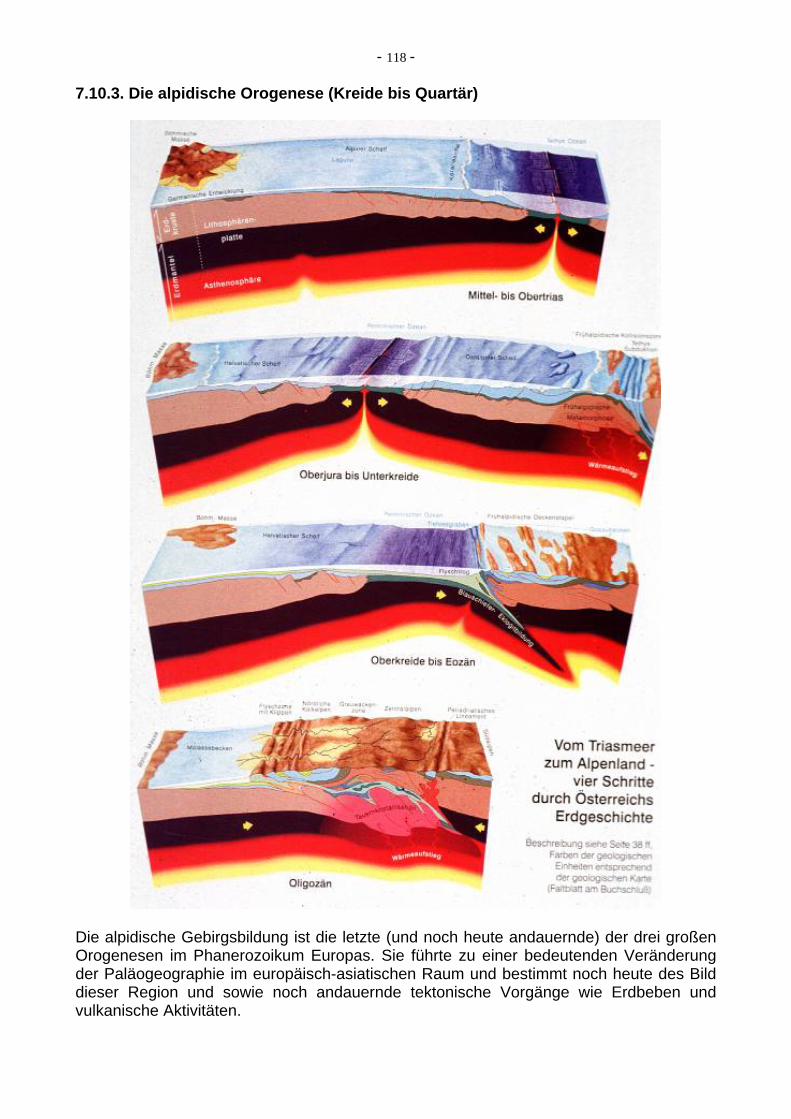
7.10.3. Die alpidische Orogenese (Kreide bis Quartär)
Die alpidische Gebirgsbildung ist die letzte (und noch heute andauernde) der drei großen Orogenesen im Phanerozoikum Europas. Sie führte zu einer bedeutenden Veränderung der Paläogeographie im europäisch-asiatischen Raum und bestimmt noch heute des Bild dieser Region und sowie noch andauernde tektonische Vorgänge wie Erdbeben und vulkanische Aktivitäten.

- 119 -
Die Vorgeschichte begann in
der Obertrias mit erstem Rifting auf Pangäa und an-chließender Bildung des penninischen Ozeans wähend des Jura. Die eigentlichen gebirgsbildenden Vorgänge begannen in der Unterkreide und hielten in einigen Regioen bis in das Quartär an. In der Oberkreide wurden die Aktivitäten enorm verstärkt.Die alpidische Orogenese erlebte ihre Höhepunkte mit der illyrischen Phase im Obereozän und der pyrenäischen Phase an der Wende Eozän / Oligozän. Die Vergenzen sind hauptsächlich nach Norden gerichtet, die Subduktion nach Süden. Im Miozän wurden die Alpen weiter eingeengt, Bruchtektonik bekam eine höhere Bedeutung. Erst im Zuge dieser Vorgänge kam es Heraushebung als Hochgebirge. Im außeralpinen Mitteleuropa waren vor allem bruchtektonische Vorgänge und vulkanische Aktivitäten mit der alpidischen Orogenese verbunden. Im Himalaya lag der Bewegungshöhepunkt an der Wende Oligozän/Miozän. Der Hintergrund für die orogenetischen Vorgänge ist die seit der Kreide wirkende Subduktion am Südrand der eurasiatischen Platte und deren Kollision mit den von Süden heranrückenden Teilstücken des Gondwana-Blockes (Afrika, Arabien, Indien). 7.11. Die Entstehung und die frühe Geschichte des Lebens auf der Erde Das Leben auf der Erde muss sich auf natürliche Weise aus lebloser Materie in weit zurückliegender Zeit aus einfachsten Stoffen entwickelt haben. Es gibt zwar auch Theo-rien, dass das Leben (oder Vorstufen davon) mit kosmischem Staub aus em Weltall zu uns gekommen ist, aber wahrscheinlicher ist doch, dass die frühe Erde als chemisches Labor zur Erzeugung ersten Lebens diente. Agenzien und chemische Grundstoffe hierzu waren die Urgase CO2, NH3, CH4, H2O und H2, elektrische Entladungen, Meteori-teneinschläge, Lava, heisse Quellen im Meer, vulkanische, radioktive Hitze und/oder Evaporation. Über Zwischenprodukte wie Aminosäuren, Alkane, Säuren, Lipide u.a wur-den erste Zellen gebildet. Bereits vor ca. 4,4 Ga (also schon im Hadaikum) hat vermutlich die abiotische Bildung von Bausteinmolekülen begonnen, anschließend folgte die abiotische Polymerisierung zu Makromolekülen, schließlich der Zusammenbau zu grösseren organell- und zellähnlichen Strukturen (Präzellen) und - als Abschluß der bis hierher so genannten "chemischen Evolution" - die Entstehung von Protobionten.

- 120 -
Links: Versuch einer Rekonstruktion der "universalen Minimalzelle" mit den wichtigsten Bestandteilen. Mitte: Einige der Energiequellen, die zur Verfügung gestanden haben, um die chemischen Reaktionen in den Ozeanen der frühen Erde anzutreiben. Rechts: Stanley Millers berühmtes Experiment simulierte die Bedingungen der frühen Erde. In einer Atmosphäre aus Methan, Stickstoff und Wasserstoff wurden elektrische Entladungen (Blitze) erzeugt, die Reaktionsprodukte abgekühlt, kondensiert und in ein Gefäß (Ozean) abgeregnet. Unter den Reaktionsprodukten fanden sich Aminosäuren ! (umgezeichnet nach Cowen 2000) Mit deren Vermehrung, Erbvariation und Selektion setzte die biologische Evolution und somit das Leben ein. Über den Zeitpunkt besteht noch Unklarheit, aber noch durch geochemische Methoden (stabile Kohlenstoffisotope) ist dies auf fast 4 Ga datiert worden. Der Nachweis von Bioproduktion auf geochemischem Wege beruht auf der Tatsache, dass hierbei das leichte Kohlenstoffenisotop C12 bevorzugt eingebaut und somit dem Meerwasser entzogen wird. Für die Bildung von Karbonaten steht somit nur noch ein am schweren Kohlenstoffenisotop C13 angereichertes Verhältnis zur Verfügung. Bei laufender Bioproduktion ergibt sich somit immer ein deutlich unterschiedliches C12/C13-Verhältnis in organischem Kohlenstoff und dem Kohlenstoff aus Karbonaten. Körperliche Reste von Lebenswesen gibt es erst aus dem Archaikum: In der 3,8 Ga Isua Supergroup (Grönland) sind Chemobakterien (Präzellen?) gefunden worden, in 3,5 Ga alten Gesteinen des Pilbara Kraton (Australien) und des Barberton Greenstone Belts (Südafrika) Biomatten (Stromatolithe) sowie kugelige und fädige Bakterien. Es läßt sich aus verschiedenen geochemischen Quellen ableiten, dass etwa zu diesem Zeitpunkt auch die Photoynthese erfunden wurde. Ab ca. 3,0 Ga waren Stromatolithe (Blaugrün"algen"-Matten) dann bereits weit verbreitet. Für die Entstehung des Lebens (vermutlich schon im Hadaikum) selbst gibt es verschie-dene Theorien. Ursprüngliche Theorie war die Entstehung in einer "Ursuppe" aus einfa-cheren organischen Verbindungen unter Beteiligung von Energiequellen wie UV-Strahlung und elektrischen Entladungen. Es wird heute auch diskutiert, ob die ersten einfachen organischen Verbindungen mit Meteoriten aus dem Weltall auf die Erde gelangt sind. Diese scheinen hier im interstellaren Staub wirklich zu existieren. Andere Theorien beschäftigen sich mit der Entstehung komplizierterer organischer Moleküle an Matrizen aus Tonmineralen, unter Beteiligung von framboidalem Pyrit (Schwefelkies) oder der Enstehung des Lebens an submarinen heißen Quellen (black smokers, hot seams).
- 121 -
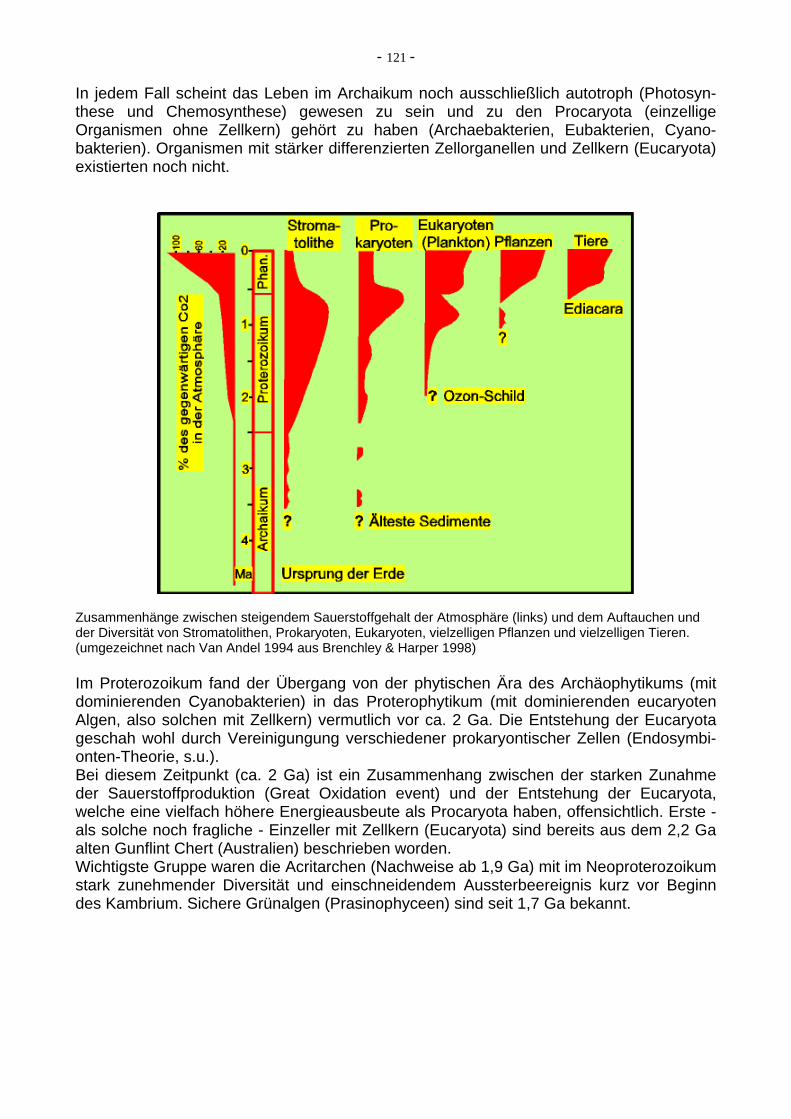
In jedem Fall scheint das Leben im Archaikum noch ausschließlich autotroph (Photosyn-these und Chemosynthese) gewesen zu sein und zu den Procaryota (einzellige Organismen ohne Zellkern) gehört zu haben (Archaebakterien, Eubakterien, Cyano-bakterien). Organismen mit stärker differenzierten Zellorganellen und Zellkern (Eucaryota) existierten noch nicht.
Zusammenhänge zwischen steigendem Sauerstoffgehalt der Atmosphäre (links) und dem Auftauchen und der Diversität von Stromatolithen, Prokaryoten, Eukaryoten, vielzelligen Pflanzen und vielzelligen Tieren. (umgezeichnet nach Van Andel 1994 aus Brenchley & Harper 1998)
Im Proterozoikum fand der Übergang von der phytischen Ära des Archäophytikums (mit dominierenden Cyanobakterien) in das Proterophytikum (mit dominierenden eucaryoten Algen, also solchen mit Zellkern) vermutlich vor ca. 2 Ga. Die Entstehung der Eucaryota geschah wohl durch Vereinigungung verschiedener prokaryontischer Zellen (Endosymbi-onten-Theorie, s.u.). Bei diesem Zeitpunkt (ca. 2 Ga) ist ein Zusammenhang zwischen der starken Zunahme der Sauerstoffproduktion (Great Oxidation event) und der Entstehung der Eucaryota, welche eine vielfach höhere Energieausbeute als Procaryota haben, offensichtlich. Erste - als solche noch fragliche - Einzeller mit Zellkern (Eucaryota) sind bereits aus dem 2,2 Ga alten Gunflint Chert (Australien) beschrieben worden. Wichtigste Gruppe waren die Acritarchen (Nachweise ab 1,9 Ga) mit im Neoproterozoikum stark zunehmender Diversität und einschneidendem Aussterbeereignis kurz vor Beginn des Kambrium. Sichere Grünalgen (Prasinophyceen) sind seit 1,7 Ga bekannt.

- 122 -
Die Endosymbiontentheorie: Die wahrscheinliche Abfolge der Entstehung vielzelligen Lebens (Pflanzen und Tiere) aus zellkernlosen Einzellern über die Zwischen-stufen der Protista (Einzeller mit Zellkern und Zellorganellen). Umgezeichnet nach Stanley (1999).
Im Neoproterozoikum fand auch schon eine rapide Entwicklung von tangähnlichen Makroalgen in den Flachmeeren statt, berühmte Funde kommen v.a. aus China. Die Entwicklung erster (noch hartteilloser) vielzelliger Tiere ist durch Spurenfossilien bei etwa 1 Ga (also etwa an der Grenze vom Meso- zum Neoproterozoikum) nachgewiesen. Dies dürften wurmartige Tiere gewesen sein. Seitdem dokumentiert eine ständige Diversifizierung der Spurenfauna eine allmähliche Entwicklung der Metazoen (vielzellige Tiere). Etwa vor 650-600 Ma (Neoproterozoikum) waren erste Makrofaunen mit erhaltenen Körpermerkmalen (nicht nur Spuren) weltweit verbreitet: Die Ediacara-Fauna, ausschließlich mit Abdrücken der Weichteile und noch ohne Hartteile. Ihre systematische Zugehörigkeit ist umstritten, einige Formen mögen zu den Coelenteraten, Anneliden oder Arthropoden gehört haben. Manche haben sich unlängst sogar als Makroalgen herausgestellt. Im allerjüngsten Proterozoikum gelangen jüngst auch schon Erstnachweise einiger heute noch existierender Tierstämme, besonders noch hartteiloser Anneliden, auch von Schwämmen. Hinzu kommen wenige sehr kleine hartschalige Fossilien von röhrenförmiger Ausbildung, allerdings ebenfalls unsicherer systematischer Stellung. Bei vielen Gruppen fand im Zuge der neoproterozoischen Vereisungen ("Schneeball Erde") ein Massenaussterben statt. Nach einigen Autoren war die hiermit verbundene Klimainstabilität einer der Gründe für die darauf folgende "Kambrische Explosion" der Tierwelt.

- 123 -
7.12. Die Entwicklung der Pflanzenwelt Die letzten 4 Ga (4 Milliarden Jahre) der Erdgeschichte lassen sich nach Entwicklungs-stand und dominierenden Pflanzengruppen in fünf sogenannte "phytische Ären" oder "Phytoären" zusammenfassen. Im Unterschied zu den Ären in der Entwicklung der Tierwelt (Paläozoikum, Mesozoikum, Känozoikum) sind die Grenzen zwischen diesen Phytoären nicht durch kurze Zeiten des Massenaussterbens gekennzeichnet, denn es handelt sich nicht um relativ scharfe Grenzen, sondern eher um länger andauernde Zeiten des Florenwandels. Die Untergrenze jeder Phytoära ist sehr unscharf, nicht zeitgleich auf verschiedenen Teilen der Erde (anisochron), und sie ist nicht durch Aussterben von Gruppen, sondern vielmehr durch Hinzukommen („Dominierendwerden“) einer neuen, fortschrittlicheren Großgruppe definiert.
Die Einteilung der Entwicklungsgeschichte der Pflanzenwelt in phytische Ären, analog zu den Ären in der Entwicklung der marinen Tierwelt. Das Archaikum ist hier des besseren Vergleichs wegen als "Archäozoi-kum" bezeichnet. Umgezeichnet nach Traverse (1988). 7.12.1. Das Archäophytikum (etwa 4,0-2,0 Ga): Dominanz der Blaugrünalgen Wenn man als "Pflanzen" im weiteren Sinne alle Photosynthese betreibenden Lebewesen zusammenfaßt (also auch zellkernlose Cyanobakterien oder Blaugrün"algen"), beginnt die Geschichte der Pflanzenwelt bereits im frühen Archaikum. Aus dieser Zeit (vielleicht schon vor etwa 4 Ga) sind aus geochemischen Untersuchungen bereits Hinweise auf photosynthetische Aktivitäten bekannt, wenn auch die eigentlichen Fossilbelege noch fehlen. Diese Frühphase der Pflanzenentwicklung, in der es noch keine Algen mit Zellkern gab, wird als die phytische Ära des "Archäophytikum" bezeichnet.

- 124 -
Die Cyanobakterien des Archäophytikums lebten im flachen Meer und trugen mit ihrer Photosynthese zu der damals noch geringen Sauerstoffproduktion bei. Wirtschaftlich bedeutend ist ihre Rolle bei der Bildung der Bändereisenerze (BIFs = Banded Iron Formations, oder auch Itabirite), da der von ihnen im Flachmeer gebildete Sauerstoff die von den damaligen Festländern eingeschwemmten Eisenoxide aufoxidierte und zum Ausfällen brachte. Dokumentiert sind die Cyanobakterien nur selten als Körperfossilien (meist fädige Auf-reihungen von Einzelzellen), sondern viel häufiger in den durch sie induzierten Bildungen lagig-kuppelförmiger Stromatolithe (durch Algenfäden eingefangene Sedimentlagen und ausgefällte Karbonatlagen). Vor etwa 2 Milliarden Jahren (2 Ga) entwickelten sich aus den verschiedenen Procaryota (zellkernlose Einzeller) die ersten Eucaryota (Einzeller mit Zellkern), also echte Algen. Dies und die damit verbundene enorme Steigerung der Sauerstoffproduktion markiert den Übergang zum Proterophytikum, der nächsten phytischen Ära. 7.12.2. Das Proterophytikum (etwa 2 Ga - 430 Ma): Dominanz der Algen mit Zellkern Als Proterophytikum bezeichnen wir die Phase, in der zusätzlich zu den Cyanobakterien höher entwickelte Algen mit Zellkern hinzukommen. Dies dürfte vor ca. 2 Ga der Fall gewesen sein und einen besonderen Schub in der Sauerstoffanreichung der Atmosphäre ausgelöst haben. Gefäßpflanzen gab es im Proterophytikum noch nicht, stattdessen dominierten immer höher entwickelte Algen (bis hin zu Makroalgen mit Zelldifferenzierung) die Meere. Noch im jüngeren Proterozoikum entwickelten sich die verschiedensten Gruppen, darunter auch Grünalgen. Besonders im Kambrium und Ordovizium waren die Böden der flachen Meere von diesen Formen bedeckt, während die Kontinente (bis auf winzige Algen, Moose und Flechten) noch reltiv kahl waren. Eine vor allem im Proterozoikum wichtige Algengruppe waren die Acritarchen. Hierunter werden äußerst vielgestaltige, organischwandige (Sporopollenin), einzellige Algen unsicherer genauerer systematischer Zugehörigkeit zusammengefaßt. Acritarchen sind nicht selten wichtige Leitfossilien, z.B. im Bereich der Proterozoikum/Kambrium-Grenze. Nach einer langen Zeit ständig steigender Diversität fast durch das gesamte Proterozoikum hindurch erlitten sie im Verlaufe der Eiszeiten im Neoproterozoikum ("Schneeball Erde") einen immensen Diversitätseinbruch (das erste Massenausterben der Erdgeschichte), von dem sie sich nie wieder richtig erholten. Das Proterophytikum endete mit der Entwicklung der Gefäßpflanzen und der Eroberung des Landes durch diese (Paläophytikum). Der Zeitpunkt ist noch umstritten und liegt entweder im Oberordovizium (nach mancher Ansicht sogar schon im Kambrium), spätestens jedoch im Silur (430-435 Ma). Im Oberordovizium der Sahara fanden sich erste Reste möglicher Landpflanzen, allerdings ausschließlich Sporentetraden und Cuticulen unsicherer systematischer Zugehörigkeit. Von den meisten Paläobotanikern werden sie nicht als gesicherte Reste von Gefäßpflanzen anerkannt, sondern z.B. zu den Lebermoosen gestellt.

- 125 -
Stammesgeschichte und Diversität der wichtigsten Gruppen der Gefäßpflanzen. Die Urfarne (Psilophyta) sind eine hier künstlich zusammengefaßte inhomogene Gruppe (ein Paraphylum). Sphenopsiden = Schach-telhalmgewächse. Lycopoden = Bärlappgewächse. Cordaiten = eine ausgestorbene Gruppe baumförmiger Nacktsamer. Cycadeen = Palmfarne. Cycadeoidea = eine ausgestorbene Gruppe von Palmfarnen. Angiospermen = Becktsamer ("Blütenpflanzen"). Umgezeichnet nach Stanley (1999). 7.12.3. Das Paläophytikum (etwa 430-270 Ma): Dominanz der Sporenpflanzen Das Paläophytikum begann mit der Entwicklung der Gefäßpflanzen und der Eroberung des Landes durch diese. Der Zeitpunkt ist noch umstritten und liegt entweder im Oberordovizium (nach mancher Ansicht sogar schon im Kambrium), spätestens jedoch im Silur (430-435 Ma). Für etwa 150-200 Ma waren nun Sporenpflanzen (Urfarne, Bärlappe, Farne und Schachtelhalme) die dominierenden Gruppen. Schon im Oberdevon entwickelten sich jedoch die ersten Samenpflanzen, und Nacktsamer wie die Cordaiten und die Samenfarne waren im Karbon bedeutend Gruppen (Stammbaum der Gefäßpflanzen). Der Übergang vom Proterophytikum in das Paläophytikum vollzog sich spätestens im Verlaufe des Silurs. Erste Funde möglicher Gefäßpflanzen bereits im Oberordovizium werden mittlerweile von den meisten Spezialisten als Reste von Lebermoosen angesehen. Die Evolution der Gefäßpflanzen nahm im Devon einen raschen Verlauf. Schnell entwickelten sich die Urfarne von einfachen und kleinen, blatt- und (fast) wurzellosen Formen zu komplizierteren und größeren Vertretern. Sukzessive kamen echte Farne, Bärlapp- und Schachtelhalmgewächse hinzu. Im Oberdevon gab es schon die ersten großen Wälder auf der Erde (meist baumförmige Progymnospermen), vereinzelt auch schon Nacktsamer (Vertreter der heute ausgestorbenen Samenfarne).

- 126 -
Das Karbon war dann die große Zeit der Sporenpflanzen, die ausgedehnte Wälder bildeten ("Steinkohlenwälder", siehe obige Abbildung). Die Pflanzengemeinschaften wurden immer diverser und differenzierter: An besonders feuchten Standorten dominierten die Schachtelhalmgewächse, in den Sumpfwäldern vor allem Bärlappbäume ("Rinden-bäume", z.B. Lepidodendron und Sigillaria) sowie Farne, Samenfarne und weitere Schachtelhalmgewächse (auch diese z.T. baumförmig). Nacktsamer wurden immer diverser und begannen, mit den Cordaiten und den Koniferen (diese im Karbon noch selten) auch die etwas trockeneren Anhöhen und Mittelgebirge zu erobern. Im Perm (siehe auch die nächste Abbildung) verarmte die typische Karbonflora, und es kam mit zunehmenden Anteilen von Nacktsamern (v.a. Koniferen) zum Florenübergang zum Mesophytikum. 7.12.4. Das Mesophytikum (etwa 270-110 Ma): Dominanz der Nacktsamer Das Mesophytikum entwickelte sich aus dem Paläophytikum mit dem allmählichen Überhandnehmen der Nacktsamer, insbesondere der Koniferen, während des Perm (lokal sogar erst in der Trias), also in den meisten Gebieten der Erde vor ca. 270-250 Ma. In vielen Sedimenten ist dies am besten an der Häufigkeit der sogenannten "taeniaten Pollen" zu erkennen. In Trias, Jura und Unterkreide dominierten dann die Gymnospermen (Nacktsamer)-Gruppen der Koniferen, Palmfarne und Ginkgogewächse die Kontinente. Der Übergang vom Paläo- zum Mesophytikum (dokumentiert durch den Anteil der Trockenheit anzeigenden taeniaten Pollen) geschah auf verschiedenen Teilen von Pangäa zu unterschiedlichen Zeiten, zuerst äquatornah in Nordamerika und zuletzt weiter östlich und polwärts. Mit der Trias begann die Hauptphase des Mesophytikums. Wichtigste Gruppen waren die Koniferen, Ginkgogewächse und Palmfarne (Cycadeen und Cycadoideen), daneben die Samenfarne. An feuchten Standorten dominierten weiterhin die Sporenpflanzen, v.a.
- 127 -
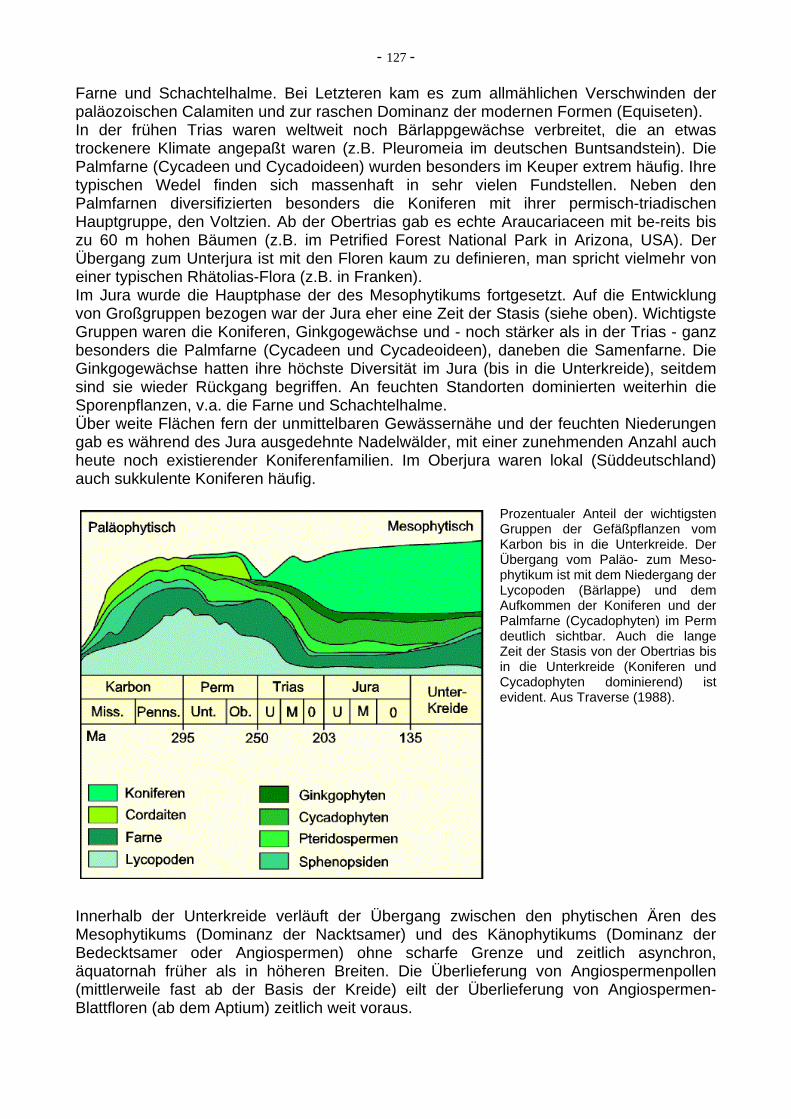
Farne und Schachtelhalme. Bei Letzteren kam es zum allmählichen Verschwinden der paläozoischen Calamiten und zur raschen Dominanz der modernen Formen (Equiseten). In der frühen Trias waren weltweit noch Bärlappgewächse verbreitet, die an etwas trockenere Klimate angepaßt waren (z.B. Pleuromeia im deutschen Buntsandstein). Die Palmfarne (Cycadeen und Cycadoideen) wurden besonders im Keuper extrem häufig. Ihre typischen Wedel finden sich massenhaft in sehr vielen Fundstellen. Neben den Palmfarnen diversifizierten besonders die Koniferen mit ihrer permisch-triadischen Hauptgruppe, den Voltzien. Ab der Obertrias gab es echte Araucariaceen mit be-reits bis zu 60 m hohen Bäumen (z.B. im Petrified Forest National Park in Arizona, USA). Der Übergang zum Unterjura ist mit den Floren kaum zu definieren, man spricht vielmehr von einer typischen Rhätolias-Flora (z.B. in Franken). Im Jura wurde die Hauptphase der des Mesophytikums fortgesetzt. Auf die Entwicklung von Großgruppen bezogen war der Jura eher eine Zeit der Stasis (siehe oben). Wichtigste Gruppen waren die Koniferen, Ginkgogewächse und - noch stärker als in der Trias - ganz besonders die Palmfarne (Cycadeen und Cycadeoideen), daneben die Samenfarne. Die Ginkgogewächse hatten ihre höchste Diversität im Jura (bis in die Unterkreide), seitdem sind sie wieder Rückgang begriffen. An feuchten Standorten dominierten weiterhin die Sporenpflanzen, v.a. die Farne und Schachtelhalme. Über weite Flächen fern der unmittelbaren Gewässernähe und der feuchten Niederungen gab es während des Jura ausgedehnte Nadelwälder, mit einer zunehmenden Anzahl auch heute noch existierender Koniferenfamilien. Im Oberjura waren lokal (Süddeutschland) auch sukkulente Koniferen häufig.
Prozentualer Anteil der wichtigsten
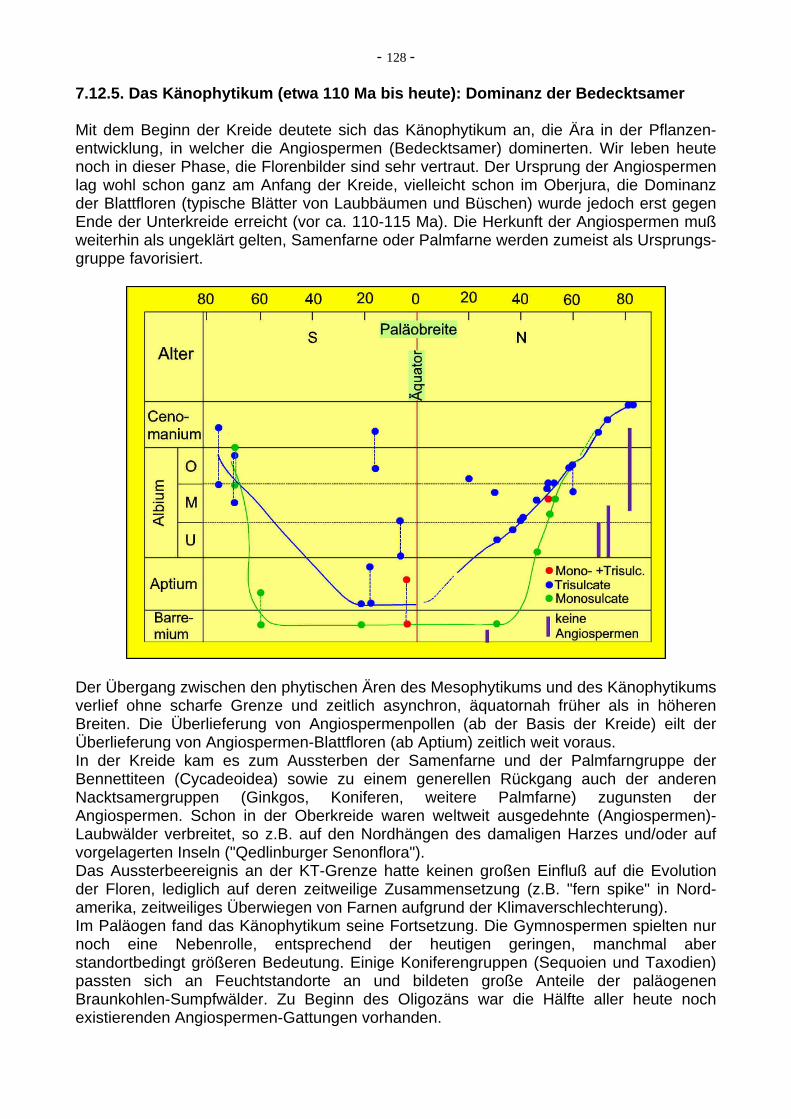
Gruppen der Gefäßpflanzen vom Karbon bis in die Unterkreide. Der Übergang vom Paläo- zum Meso-phytikum ist mit dem Niedergang der Lycopoden (Bärlappe) und dem Aufkommen der Koniferen und der Palmfarne (Cycadophyten) im Perm deutlich sichtbar. Auch die lange Zeit der Stasis von der Obertrias bis in die Unterkreide (Koniferen und Cycadophyten dominierend) ist evident. Aus Traverse (1988).Innerhalb der Unterkreide verläuft der Übergang zwischen den phytischen Ären des Mesophytikums (Dominanz der Nacktsamer) und des Känophytikums (Dominanz der Bedecktsamer oder Angiospermen) ohne scharfe Grenze und zeitlich asynchron, äquatornah früher als in höheren Breiten. Die Überlieferung von Angiospermenpollen (mittlerweile fast ab der Basis der Kreide) eilt der Überlieferung von Angiospermen-Blattfloren (ab dem Aptium) zeitlich weit voraus.
- 128 -
7.12.5. Das Känophytikum (etwa 110 Ma bis heute): Dominanz der Bedecktsamer Mit dem Beginn der Kreide deutete sich das Känophytikum an, die Ära in der Pflanzen-entwicklung, in welcher die Angiospermen (Bedecktsamer) dominerten. Wir leben heute noch in dieser Phase, die Florenbilder sind sehr vertraut. Der Ursprung der Angiospermen lag wohl schon ganz am Anfang der Kreide, vielleicht schon im Oberjura, die Dominanz der Blattfloren (typische Blätter von Laubbäumen und Büschen) wurde jedoch erst gegen Ende der Unterkreide erreicht (vor ca. 110-115 Ma). Die Herkunft der Angiospermen muß weiterhin als ungeklärt gelten, Samenfarne oder Palmfarne werden zumeist als Ursprungs-gruppe favorisiert.
Der Übergang zwischen den phytischen Ären des Mesophytikums und des Känophytikums verlief ohne scharfe Grenze und zeitlich asynchron, äquatornah früher als in höheren Breiten. Die Überlieferung von Angiospermenpollen (ab der Basis der Kreide) eilt der Überlieferung von Angiospermen-Blattfloren (ab Aptium) zeitlich weit voraus. In der Kreide kam es zum Aussterben der Samenfarne und der Palmfarngruppe der Bennettiteen (Cycadeoidea) sowie zu einem generellen Rückgang auch der anderen Nacktsamergruppen (Ginkgos, Koniferen, weitere Palmfarne) zugunsten der Angiospermen. Schon in der Oberkreide waren weltweit ausgedehnte (Angiospermen)-Laubwälder verbreitet, so z.B. auf den Nordhängen des damaligen Harzes und/oder auf vorgelagerten Inseln ("Qedlinburger Senonflora"). Das Aussterbeereignis an der KT-Grenze hatte keinen großen Einfluß auf die Evolution der Floren, lediglich auf deren zeitweilige Zusammensetzung (z.B. "fern spike" in Nord-amerika, zeitweiliges Überwiegen von Farnen aufgrund der Klimaverschlechterung). Im Paläogen fand das Känophytikum seine Fortsetzung. Die Gymnospermen spielten nur noch eine Nebenrolle, entsprechend der heutigen geringen, manchmal aber standortbedingt größeren Bedeutung. Einige Koniferengruppen (Sequoien und Taxodien) passten sich an Feuchtstandorte an und bildeten große Anteile der paläogenen Braunkohlen-Sumpfwälder. Zu Beginn des Oligozäns war die Hälfte aller heute noch existierenden Angiospermen-Gattungen vorhanden.

- 129 -
Besonders wichtig war die Entwicklung der Gräser, die im Eozän und tieferen Oligozän noch auf bewaldete und sumpfige Gebiete beschränkt waren. Mit der Entwicklung des kontinuierlichen Wachstumsprozesses und der Windbestäubung wurden ab dem Oberoligozän die offenen Länder erobert, Steppen und Grasländer breiteten sich aus, größeren Massen von Pflanzenfressern konnten durch Nachwachsen der Halme Nahrung finden. Diese Entwicklung korreliert mit dem allmählichen Trockenerwerden des Klimas (Versteppung) und der Evolution einiger Säugetiergruppen (z.B. Pferde). 7.13. Die Entwicklung der Tierwelt Die Entwicklung erster (noch hartteilloser) vielzelliger Tiere ist durch Spurenfossilien bei etwa 1 Ga (also etwa an der Grenze vom Meso- zum Neoproterozoikum) nachgewiesen. Dies dürften wurmartige Tiere gewesen sein. Seitdem dokumentiert eine ständige Diver-sifizierung der Spurenfauna eine allmähliche Entwicklung der Metazoen (vielzellige Tiere). Etwa vor 650-600 Ma (Neoproterozoikum) waren erste Makrofaunen mit erhaltenen Körpermerkmalen weltweit verbreitet: Die Ediacara-Fauna, ausschließlich mit Abdrücken der Weichteile und noch ohne Hartteile. Ihre systematische Zugehörigkeit ist umstritten, einige Formen mögen zu den Coelenteraten, Anneliden oder Arthropoden gehört haben. Manche haben sich unlängst sogar als Makroalgen herausgestellt. Im allerjüngsten Proterozoikum gelangen jüngst auch schon Erstnachweise einiger heute noch existierender Tierstämme, besonders noch hartteiloser Anneliden, auch von Schwämmen. Hinzu kommen wenige sehr kleine hartschalige Fossilien von röhrenförmi-ger Ausbildung, allerdings ebenfalls unsicherer systematischer Stellung. Bei vielen Gruppen fand im Zuge der neoproterozoischen Vereisungen ("Schneeball Erde") ein Massenaussterben statt. Nach einigen Autoren war die hiermit verbundene Klimainstabilität einer der Gründe für die darauf folgende "kambrische Explosion" der Tierwelt. Diese gab den Startschuß für die Evolution der hartteiltragenden Tierwelt und die darauf basierend gute Fossilüberlieferung mit allen Konsequenzen für die Biostratigraphie. Im Anschluß an die Proterozoikum/Kambrium-Grenze fand während der letzten 540 Ma der Erdgeschichte die Entwicklung der marinen Invertebratenfaunen im Phanerozoikum (die drei großen "evolutionären Faunen" des Phanerozoikums). Diese wurde wiederholt von bedeutenden Phasen des Massenaussterbens unterbrochen, in denen bis zu 95 % (geschätzt) aller Gattungen mariner Wirbelloser ausstarben. Besonders heftig litten unter diesen Zeiten immer wieder die besonders anfälligen Riff-Ökosysteme. Die Stammesgeschichte der Wirbeltiere setzte zwar bereits im Kambrium ein, erste Landtiere gab es aber erst gegen Ende des Devons. Die Entwicklungsgeschichte des Menschen ist in diesem großen Zeitrahmen nicht mehr als ein Hauch, setzte sie doch erst im späten Miozän vor ca. 6 Ma ein. 7.13.1. Die "Ediacara-Fauna" des späten Proterozoikums Seit etwa 1950 wurden in vielen Teilen der Erde (zuerst entdeckt in den Ediacara Hills in Südaustralien, daher der Name für diesen gesamten Faunentyp) Abdrücke verschiedenster, bis 50 cm großer Tiere mit Weichkörpern, aber ohne Hartteile gefunden. Ihr Alter liegt bei ca. 600-560 Ma. In Rekonstruktionen sind diese Tiere teils wie Quallen, Seefedern oder bodenliegende Seeanemonen dargestellt. Ihre systematische Zugehörigkeit ist aber umstritten, einige Formen mögen zu den Coelenteraten, Anneliden oder Arthropoden gehört haben. Manche haben sich unlängst sogar als Makroalgen herausgestellt.

- 130 -
Andere Autoren wiederum haben für den Großteil dieser z.T. offenbar ähnlich wie gesteppte Luftmatratzen gebauten Organismen einen eigenen Tierstamm (Vendobionta, nach Vendium, dem alten Namen für das Neoproterozoikum) eingeführt. Nach dieser, allerdings sehr stark angefochtenen, Anschauung des derzeit wohl berühmtesten deutschen Paläontologen Adolf Seilacher (emer. Prof. in Tübingen) waren die meisten Ediacara-Tiere riesengroße Einzeller, die nur durch die Unterteilung ihrer Zelle diese Größen erreichen konnten und in Symbiose mit Algen friedlich auf den Meereböden lebten (Stichwort "Garten Ediacara").
Vertreter dieser völlig hartteil- und schutzlosen Faunen sind wohl nur deshalb erhalten geblieben, weil es in dieser Frühzeit der Tierevolution nur wenige (oder keine) Räuber oder Aasfresser gegeben hat, die die abgestorbenenm Organismen verzehren bzw. die am Meeresboden liegenden Leichen durch ihre Aktivitäten zerstören konnten. 7.13.2. Die "kambrische Explosion" der marinen Tierwelt Etwa an der Wende vom Proterozoikum zum Kambrium (vor 540 Ma) änderte sich die Überlieferung von Organismenresten fast schlagartig: Neben den vorher dominierenden Weichkörperfossilien (z.B. Ediacara-Fauna und diverse Spurenfossilien) kommen zunächst sehr kleine ("small shelly fossils"), dann recht bald aber auch größere skelett-tragende Organismen hinzu. Dieses Phänomen wird allgemein als "kambrische Explosion" oder "Revolution" bezeichnet, gemeint ist hiermit die - in geologischen Zeitbegriffen - rapide Beschleunigung in der Evolution mariner Invertebratenfaunen. Die Entwicklungen an der Wende vom Proterozoikum zum Kambrium führten zum abrupten Auftreten vieler Metazoen-Gruppen, die so hochgradig verschieden sind, dass diese Plötzlichkeit sehr rätselhaft erscheint. Schon im Unterkambrium (zumindest im weiteren Verlauf des Kambriums) tauchten fast alle heutigen Tierstämme auf! Daneben

- 131 -
entwickelten sich viele Metazoen, die von den heute bekannten Formen und Bauplänen so verschieden sind, dass sie als eigene, heute ausgestorbene Stämme aufgefaßt werden müssen. Viele Vertreter dieser Faunen sind nur aus Schichten mit außergewöhnlich guter Fossilerhaltung (auch von Weichteilen) bekannt geworden (z.B. Burgess-Schiefer in Kanada oder Chenjiang-Schichten in China). Zusammen mit jenen Vertretern aus auch heute noch vertretenen Tierstämmen, die im schon Kambrium ihren Höhepunkt erlebten und später bis zum Ende des Paläozoikums sukzessive unbedeutender wurden (z.B. Trilobiten und inarticulate Brachiopoden), werden diese Gruppen als "kambrische evolutionäre Fauna" zusammengefasst (siehe unten).
Geologische Verbreitung der wichtigsten Gruppen mariner wirbelloser Tiere. Die Höhe der jeweiligen Kurven soll in etwa die relative Anzahl von Gattung (Diversität) in den jeweiligen Gruppen und Zeiten repräsentieren. (umgezeichnet nach Lehmann & Hillmer 1997).
Im Detail ist die Fossilüberlieferung (das gestaffelte Erstauftreten ver-schiedenster Gruppen) an der Proterozoikum / Kambrium - Grenze bzw. im Unterkambrium sehr kompli-ziert verlaufen. Mittlerweile ist schon eine Reihe von Skelettresten auch aus dem spätesten Proterozoikum bekannt geworden (z.B. Schwamm-skleren und kalkige Röhrchen von Cloudina). Generell traten aber zunächst fast ausschließlich phos-phatische Reste auf, kalkige Nadeln, Sklerite und Röhrchen ("small shelly fossils") folgten etwas später, deutlich schon im frühen Kambrium. Einige dieser Reste mögen von den ersten Mollusken stammen. Kurz danach traten die ersten Archaeo-cathiden (schwammähnliche sessile Strudler) auf, erst dann die ersten Trilobiten und weitere Metazoen-gruppen mit Harteilen.
- 132 -
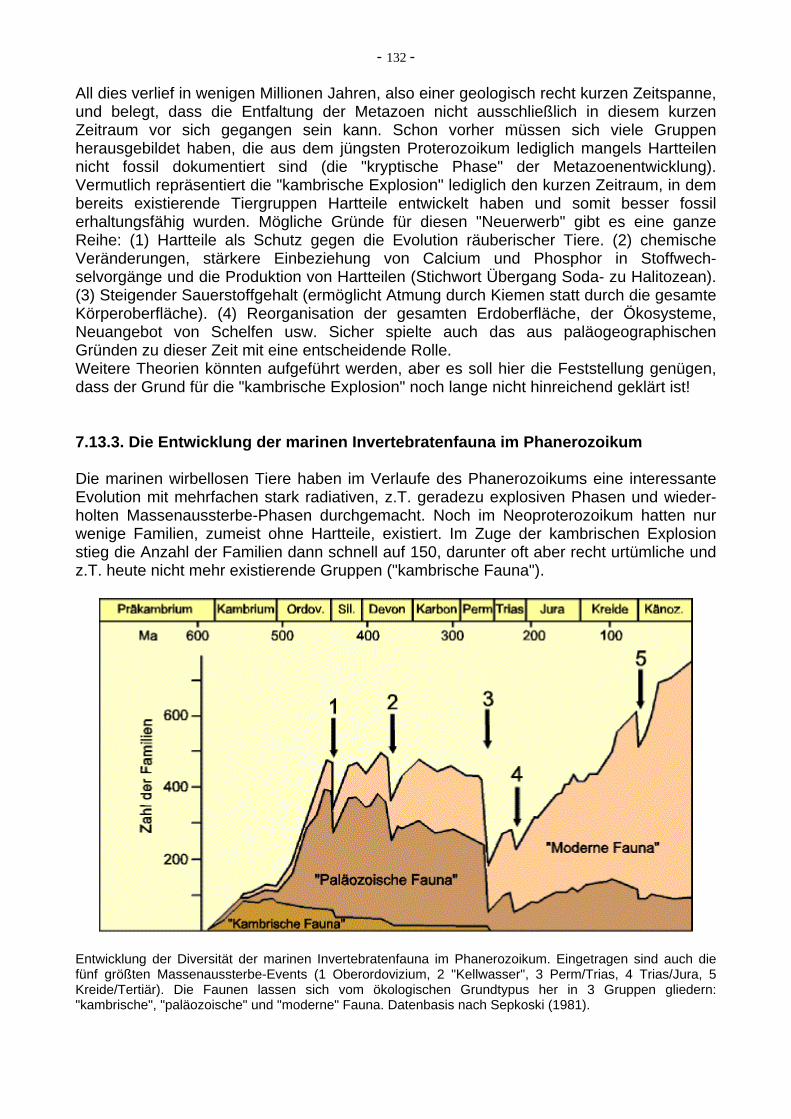
All dies verlief in wenigen Millionen Jahren, also einer geologisch recht kurzen Zeitspanne, und belegt, dass die Entfaltung der Metazoen nicht ausschließlich in diesem kurzen Zeitraum vor sich gegangen sein kann. Schon vorher müssen sich viele Gruppen herausgebildet haben, die aus dem jüngsten Proterozoikum lediglich mangels Hartteilen nicht fossil dokumentiert sind (die "kryptische Phase" der Metazoenentwicklung). Vermutlich repräsentiert die "kambrische Explosion" lediglich den kurzen Zeitraum, in dem bereits existierende Tiergruppen Hartteile entwickelt haben und somit besser fossil erhaltungsfähig wurden. Mögliche Gründe für diesen "Neuerwerb" gibt es eine ganze Reihe: (1) Hartteile als Schutz gegen die Evolution räuberischer Tiere. (2) chemische Veränderungen, stärkere Einbeziehung von Calcium und Phosphor in Stoffwech-selvorgänge und die Produktion von Hartteilen (Stichwort Übergang Soda- zu Halitozean). (3) Steigender Sauerstoffgehalt (ermöglicht Atmung durch Kiemen statt durch die gesamte Körperoberfläche). (4) Reorganisation der gesamten Erdoberfläche, der Ökosysteme, Neuangebot von Schelfen usw. Sicher spielte auch das aus paläogeographischen Gründen zu dieser Zeit mit eine entscheidende Rolle. Weitere Theorien könnten aufgeführt werden, aber es soll hier die Feststellung genügen, dass der Grund für die "kambrische Explosion" noch lange nicht hinreichend geklärt ist! 7.13.3. Die Entwicklung der marinen Invertebratenfauna im Phanerozoikum Die marinen wirbellosen Tiere haben im Verlaufe des Phanerozoikums eine interessante Evolution mit mehrfachen stark radiativen, z.T. geradezu explosiven Phasen und wieder-holten Massenaussterbe-Phasen durchgemacht. Noch im Neoproterozoikum hatten nur wenige Familien, zumeist ohne Hartteile, existiert. Im Zuge der kambrischen Explosion stieg die Anzahl der Familien dann schnell auf 150, darunter oft aber recht urtümliche und z.T. heute nicht mehr existierende Gruppen ("kambrische Fauna").
Entwicklung der Diversität der marinen Invertebratenfauna im Phanerozoikum. Eingetragen sind auch die fünf größten Massenaussterbe-Events (1 Oberordovizium, 2 "Kellwasser", 3 Perm/Trias, 4 Trias/Jura, 5 Kreide/Tertiär). Die Faunen lassen sich vom ökologischen Grundtypus her in 3 Gruppen gliedern: "kambrische", "paläozoische" und "moderne" Fauna. Datenbasis nach Sepkoski (1981).

- 133 -
Ein neuer rasanter Anstieg auf etwas über 400 Familien fand im Ordovizium statt ("ordovizische Radiation"). Dies sind überwiegend Vertreter ökologisch etwas fortge-schrittenerer Grundtypen ("paläozoische Fauna"). Trotz mehrfacher Massenaussterbe-Ereignisse (Oberordovizium und Oberdevon) hielt sich das Level dann im Paläozoikum recht konstant bei etwas über 400 Familien. An der Perm/Trias-Grenze kam es dann zu einem dramatischen Rückgang der marinen Invertebraten auf ca. 200 Familien. Seitdem ist ein kontinuierlicher Anstieg auf über 600 Familien (zumeist Vertreter der sogenannten "modernen Fauna") zu verzeichnen, mit Rückschlägen an der Trias/Jura- und an der Kreide/Paläogen-Grenze (K/T -Event). Die drei großen "evolutionären" Faunen des Phanerozoikums unterscheiden sich grund-säztlich hinsichtlich der Lebensweisen ihrer Hauptvertreter (unterschiedliche "Gilden" der Lebensgemeinschaften und Nahrungsketten, Details siehe unten). Neue Faunentypen addieren immer mehr ökologische Komponenten zu den Meeresboden-Assoziationen (mehr "Gilden"). 7.13.3.1. Die "kambrische Fauna" (oder „kambrische evolutionäre Fauna“) Die "kambrische Fauna" (oder "kambrische evolutionäre Fauna") bestand v.a. aus Trilobiten, Würmern und inarticulaten Brachiopoden, zusätzlich einige urtümliche Mollusken- und Echinodermengruppen. Meist waren das Sedimentfresser, oft sehr flache Tiere, grabende Tiere waren nur sehr oberflächlich tätig. Die Diversität war generell gering, Räuber spielten noch keine allzu große Rolle.
Die "kambrische Fauna" und ihre Diversitätsentwicklung im Phanerozoikum sowie einige typische Vertreter. Dargestellt ist die Anzahl der Familien mariner Invertebraten. Datenbasis nach Sepkoski (1981). Der Name mag anfangs etwas verwirren, da Vertreter der "kambrischen Fauna" bis zu Ende des Perms (Perm/Trias-Krise) existierten (und wenige noch heute). Der Faunentyp definiert sich aber ausschließlich durch die in ihm vertretenen ökologische Gilden, nicht durch seine stratigraphische Reichweite. Vertreter der "kambrischen Fauna" waren die Gewinner der sogenannten "kambrischen Explosion" der marinen Tierwelt. Hierzu gehört neben bekannteren und durch Hartteilfossilien häufiger erhaltenen Gruppen wie den Trilobiten und den inarticulaten Brachiopoden auch eine größere Anzahl von Tierstämmen mit heute völlig unbekannten Grundbauplänen, oft nur überliefert in Fundstellen mit außergewöhnlich guter Erhaltung wie z.B. den Burgess Shales in Kanada. Nach ihrer Blütezeit im Kambrium gingen die Vertreter der "kambrischen Fauna" in ihrer Diversität während des ganzen Paläozoikums allmählich zurück. Grund hierfür war der Konkurrenzdruck der höher entwickelten Vertreter der "paläozoischen Fauna". Am Ende

- 134 -
des Perms starb die "kambrische Fauna" nahezu aus, heute existieren nur noch wenige Vertreter wie die inarticulaten Brachiopoden (z.B. Lingula). 7.13.3.2. Die "paläozoische Fauna" (oder „paläozoische evolutionäre Fauna“) Die "paläozoische Fauna" dominerte, nach erster Entstehung schon im Kambrium, seit dem Ordovizium während des gesamten restlichen Paläozoikums. Es waren v.a. articulate Brachiopoden, Crinoiden, Ostracoden, Cephalopoden und Korallen. Die Lebensgemein-schaften waren dominiert von sessilen Suspensionsfressern des Epibenthos. Räuber wurden wichtiger, waren aber eher langsam beweglich ("Wegelagerer"-Typus). Die trophi-schen Strukturen wurden deutlich komplexer, die Filtrierer reichten höher in das Wasser hinauf. Der Name mag anfangs etwas verwirren, da Vertreter der "paläozoischen Fauna" vom Kambrium bis heute existierten. Der Faunentyp definiert sich aber ausschließlich durch die in ihm vertretenen ökologische Gilden, nicht durch seine stratigraphische Reichweite. Nach dem großen Aussterbeereignis an der Perm/Trias-Grenze haben sich die Vertreter der im Paläozoikum so dominierenden "paläozoischen Fauna" nie wieder auf das alte Niveau von etwa 300 Familien zurückentwickeln können. Stattdessen verblieben sie bis heute bei nur noch ca. 100 Familien. Gewinner der Perm/Trias-Krise war die "moderne Fauna". Obwohl die Unterschiede zwischen typisch "paläozoischer" und typisch "moderner" Fauna nicht ganz so scharf sind, wie der bisherige Fossilbericht dies weismachen möchte, ist er trotzdem sehr evident.
Die "paläozoische Fauna" und ihre Diversitätsentwicklung im Phanerozoikum sowie einige typische Vertreter. Dargestellt ist die Anzahl der Familien mariner Invertebraten. Datenbasis nach Sepkoski (1981). Warum hat sich nach der Perm/Trias-Grenze die "paläozoische" Fauna nicht wieder durchgesetzt? Sicher gibt es mehrere zusammenspielende Faktoren: Die Entwicklung leistungsfähigerer Räuber scheint einer der Gründe hierfür zu sein. Bei den Schnecken z.B. ist das ganz besonders evident, ebenso bei Krebsen und bei modernen Fischen. Zum Teil geschah dies aber nach bisheriger Fossilüberlieferung etwas später (bei Schnecken z.B. ab der mittleren Kreide). Die sessilen ("paläozoischen") Nahrungsfilterer, fixiert am Meeresboden, scheinen zu angreifbar gewesen zu sein. Die

- 135 -
neu entwickelten vergraben lebenden Filter-Feeder (Muscheln etc.) waren besser geschützt, ergo erfolgreicher. Ein weiterer Grund: Die Entwicklung leistungsfähigerer Kiemen bei Muscheln bewirkte die Möglichkeit, sich tiefer einzugraben, ein effektiver Schutz gegen Räuber, auch gegen Austrockenen und Erosion. Die Bulldozer-Hypothese: Bei der "modernen Fauna" gibt es viele intensiv grabende Tiere (Muscheln, Krebse, Würmer, Echiniden usw.), so dass die Larven "paläozoischer" Strudler schwer Fuß fassen konnten, Adulte "mitverarbeitet" werden und überhaupt aufgewühlte Schlammmassen ihnen die Kiemen verkleben. Alles zusammengenommen sind auch deshalb mobile Nahrungsfilterer (eine Gilde der "modernen Fauna") konkurrenzfähiger als sessile Nahrungsfilterer (eine Gilde der "paläozoischen Fauna"). 7.13.3.3. Die "moderne Fauna" (oder „moderne evolutionäre Fauna“) Die Vertreter der "modernen Fauna" waren die Gewinner der Perm/Trias-Krise. Auch sie waren mit ihren ersten Vertretern bereits im Kambrium entstanden, jedoch während des gesamten Paläozoikums nur von untergeordneter (aber ständig steigender) Bedetung. Es sind v.a. Muscheln und Schnecken, Knochenfische, Krebse und Seeigel sowie bestimmte moderne Bryozoen, Schwämme und Foraminiferen. Dominiert wurden und werden sie von schwimmenden Räubern (also im Gegensatz zu den langsamen Räubern der "paläozoischen Fauna" schnelle Räuber) und grabenden, also vagilen (ebenfalls im Gegensatz zur "paläozoischen Fauna") Suspensions- und Sedimentfressern (Mollusken, Seeigel, Krebse) sowie von vielseitigen Opportunisten wie Crustaceen und Gastropoden. Die ökologische Komplexität stieg enorm an, bis zu ihrem heutigen Level.
Die "moderne Fauna" und ihre Diversitätsentwicklung im Phanerozoikum sowie die wichtigsten Vertreter. Dargestellt ist die Anzahl der Familien mariner Invertebraten. Datenbasis nach Sepkoski (1981). Zusätzlich sind hier als „schnelle Räuber“ schwimmende Wirbeltiere dargestellt.
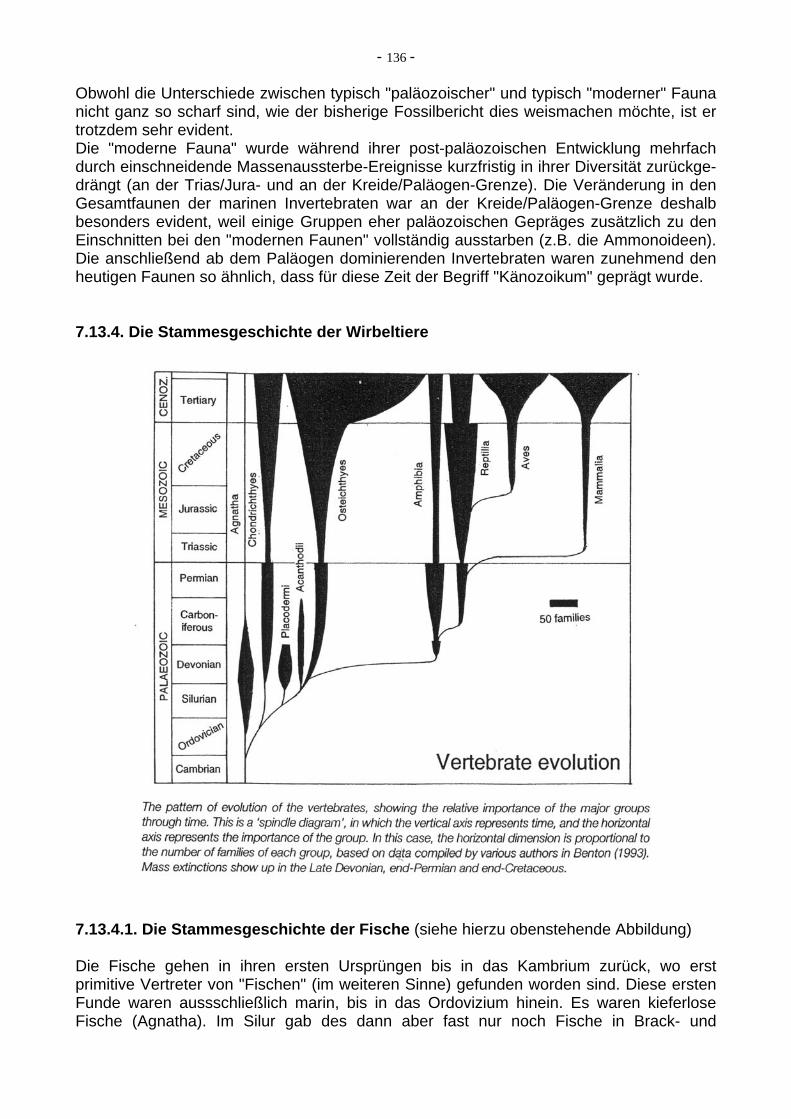
- 136 -
Obwohl die Unterschiede zwischen typisch "paläozoischer" und typisch "moderner" Fauna nicht ganz so scharf sind, wie der bisherige Fossilbericht dies weismachen möchte, ist er trotzdem sehr evident. Die "moderne Fauna" wurde während ihrer post-paläozoischen Entwicklung mehrfach durch einschneidende Massenaussterbe-Ereignisse kurzfristig in ihrer Diversität zurückge-drängt (an der Trias/Jura- und an der Kreide/Paläogen-Grenze). Die Veränderung in den Gesamtfaunen der marinen Invertebraten war an der Kreide/Paläogen-Grenze deshalb besonders evident, weil einige Gruppen eher paläozoischen Gepräges zusätzlich zu den Einschnitten bei den "modernen Faunen" vollständig ausstarben (z.B. die Ammonoideen). Die anschließend ab dem Paläogen dominierenden Invertebraten waren zunehmend den heutigen Faunen so ähnlich, dass für diese Zeit der Begriff "Känozoikum" geprägt wurde. 7.13.4. Die Stammesgeschichte der Wirbeltiere
7.13.4.1. Die Stammesgeschichte der Fische (siehe hierzu obenstehende Abbildung) Die Fische gehen in ihren ersten Ursprüngen bis in das Kambrium zurück, wo erst primitive Vertreter von "Fischen" (im weiteren Sinne) gefunden worden sind. Diese ersten Funde waren aussschließlich marin, bis in das Ordovizium hinein. Es waren kieferlose Fische (Agnatha). Im Silur gab des dann aber fast nur noch Fische in Brack- und

- 137 -
Süßwasserablagerungen. Hier kamen neben den Agnathen auch schon erste kiefertra-gende Fische (Acanthodii) vor. Das Devon war die wichtigste Zeit in der Entwicklung der Fische, sie eroberten alle aquatischen Bereiche, und verschiedenste, z.T. heute ausgestorbene oder weniger bedeutende Gruppen kiefertragender Fische wurden dominant (Placodermii). Am Ende des Devons starben die meisten Gruppen kieferloser Fische aus, bis heute haben nur noch die Schleimaale und Neunaugen überlebt. Die Stammesgeschichte der Fische verlief im Weiteren sukzessive hin zu immer größeren Ähnlichkeiten mit den heutigen Fischfaunen und ihren Hauptgruppen der Chondrichthyes (Knorpelfische, also Haie und Rochen) und vor allem der Osteichthyes (Knochenfische, Strahlenflosser). Typische paläozoische und typische post-paläozoische Fischfaunen unterscheiden sich wesentlich, einen starken Einschnitt gab es im Verlaufe des Massenaussterbens an der Perm/Trias-Grenze. 7.13.4.2. Die Stammesgeschichte der Reptilien Die Stammesgeschichte der Reptilien setzte mit ihren ersten Vertretern im Verlaufe des Karbons ein. Die Entwicklung des Amnion-Eies ermöglichte ein gegenüber den in ihrem Fortpflanzungszyklus auf das Wasser angewiesenen Amphibien eine Lebensweise auch in gewässerferneren und trockeneren Gebieten.
Das amniotische Ei ist in vielerlei Hinsicht analog zu
einem Raumschiff. So wie dieser neue Ei-Typus geeignet war, unter bisher ungeeigneten Bedingungen die Fortpflanzung zu sichern, ermöglicht das Raumschiff das Überleben des Menschen in eigentlich für diesen völlig ungeeigneter Umgebung. (umgezeichnet nach Cowen 2000)Es ist sicherlich kein Zufall, dass sich die Reptilien im Verlaufe des Perm parallel zu weltweit immer arider werdenden Klimaten gegenüber den Amphibien durchsetzten und zur dominierenden Wirbeltiergruppe wurden. Im frühen Perm dominierten zuerst die Pelycosaurier mit ihren bekannten "Rückenflügeln" (dienten zur Thermorgulation), während sich im höheren Perm die säugetierähnlichen Reptilien (Therapsida) mit ihren für eine räuberische Lebensweise besser geeigneten Eigenschaften (Gebißdifferenzierung, Stellung der Beine mehr unter als neben dem Körper?, Haarkleid und Warmblütigkeit?) durchsetzen konnten.
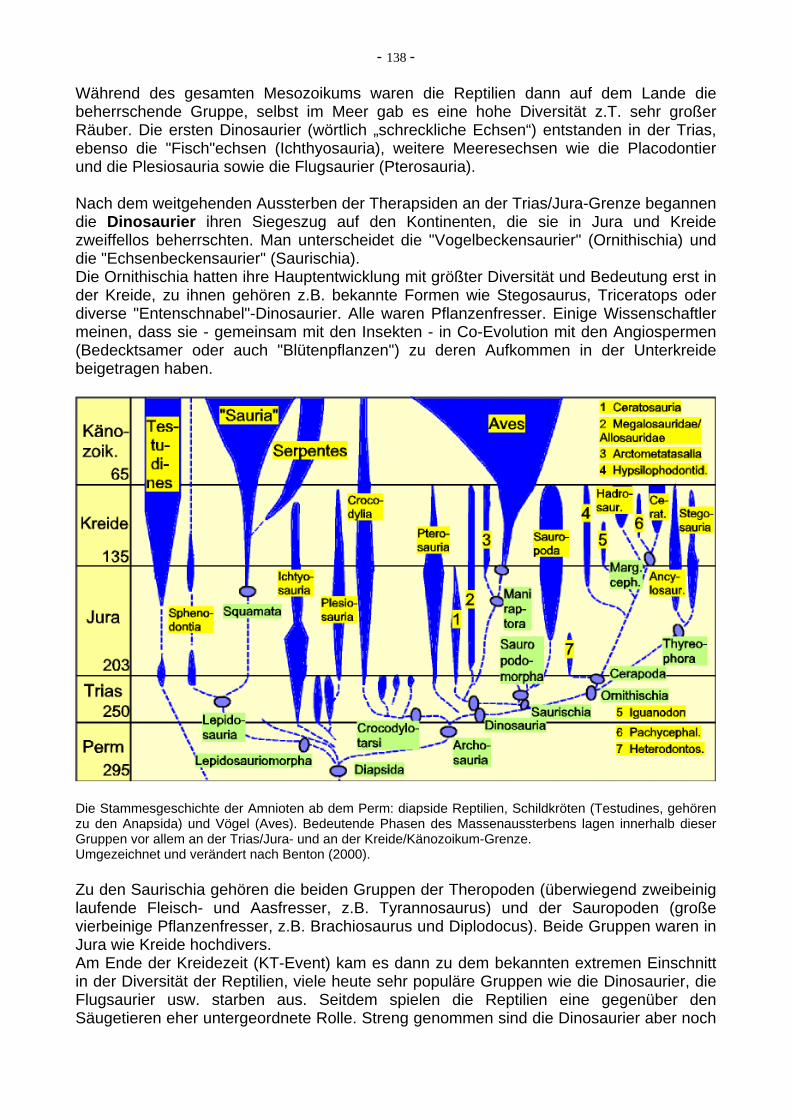
- 138 -
Während des gesamten Mesozoikums waren die Reptilien dann auf dem Lande die beherrschende Gruppe, selbst im Meer gab es eine hohe Diversität z.T. sehr großer Räuber. Die ersten Dinosaurier (wörtlich „schreckliche Echsen“) entstanden in der Trias, ebenso die "Fisch"echsen (Ichthyosauria), weitere Meeresechsen wie die Placodontier und die Plesiosauria sowie die Flugsaurier (Pterosauria). Nach dem weitgehenden Aussterben der Therapsiden an der Trias/Jura-Grenze begannen die Dinosaurier ihren Siegeszug auf den Kontinenten, die sie in Jura und Kreide zweiffellos beherrschten. Man unterscheidet die "Vogelbeckensaurier" (Ornithischia) und die "Echsenbeckensaurier" (Saurischia). Die Ornithischia hatten ihre Hauptentwicklung mit größter Diversität und Bedeutung erst in der Kreide, zu ihnen gehören z.B. bekannte Formen wie Stegosaurus, Triceratops oder diverse "Entenschnabel"-Dinosaurier. Alle waren Pflanzenfresser. Einige Wissenschaftler meinen, dass sie - gemeinsam mit den Insekten - in Co-Evolution mit den Angiospermen (Bedecktsamer oder auch "Blütenpflanzen") zu deren Aufkommen in der Unterkreide beigetragen haben.
Die Stammesgeschichte der Amnioten ab dem Perm: diapside Reptilien, Schildkröten (Testudines, gehören zu den Anapsida) und Vögel (Aves). Bedeutende Phasen des Massenaussterbens lagen innerhalb dieser Gruppen vor allem an der Trias/Jura- und an der Kreide/Känozoikum-Grenze. Umgezeichnet und verändert nach Benton (2000). Zu den Saurischia gehören die beiden Gruppen der Theropoden (überwiegend zweibeinig laufende Fleisch- und Aasfresser, z.B. Tyrannosaurus) und der Sauropoden (große vierbeinige Pflanzenfresser, z.B. Brachiosaurus und Diplodocus). Beide Gruppen waren in Jura wie Kreide hochdivers. Am Ende der Kreidezeit (KT-Event) kam es dann zu dem bekannten extremen Einschnitt in der Diversität der Reptilien, viele heute sehr populäre Gruppen wie die Dinosaurier, die Flugsaurier usw. starben aus. Seitdem spielen die Reptilien eine gegenüber den Säugetieren eher untergeordnete Rolle. Streng genommen sind die Dinosaurier aber noch
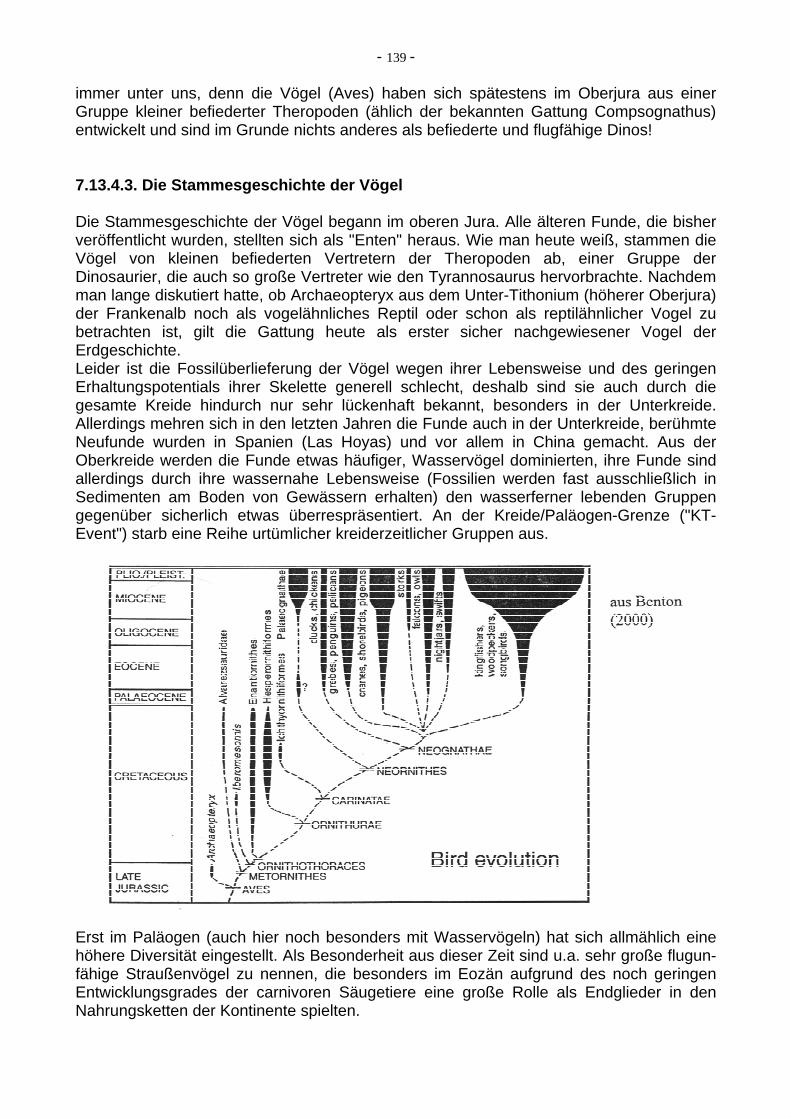
- 139 -
immer unter uns, denn die Vögel (Aves) haben sich spätestens im Oberjura aus einer Gruppe kleiner befiederter Theropoden (ählich der bekannten Gattung Compsognathus) entwickelt und sind im Grunde nichts anderes als befiederte und flugfähige Dinos! 7.13.4.3. Die Stammesgeschichte der Vögel Die Stammesgeschichte der Vögel begann im oberen Jura. Alle älteren Funde, die bisher veröffentlicht wurden, stellten sich als "Enten" heraus. Wie man heute weiß, stammen die Vögel von kleinen befiederten Vertretern der Theropoden ab, einer Gruppe der Dinosaurier, die auch so große Vertreter wie den Tyrannosaurus hervorbrachte. Nachdem man lange diskutiert hatte, ob Archaeopteryx aus dem Unter-Tithonium (höherer Oberjura) der Frankenalb noch als vogelähnliches Reptil oder schon als reptilähnlicher Vogel zu betrachten ist, gilt die Gattung heute als erster sicher nachgewiesener Vogel der Erdgeschichte. Leider ist die Fossilüberlieferung der Vögel wegen ihrer Lebensweise und des geringen Erhaltungspotentials ihrer Skelette generell schlecht, deshalb sind sie auch durch die gesamte Kreide hindurch nur sehr lückenhaft bekannt, besonders in der Unterkreide. Allerdings mehren sich in den letzten Jahren die Funde auch in der Unterkreide, berühmte Neufunde wurden in Spanien (Las Hoyas) und vor allem in China gemacht. Aus der Oberkreide werden die Funde etwas häufiger, Wasservögel dominierten, ihre Funde sind allerdings durch ihre wassernahe Lebensweise (Fossilien werden fast ausschließlich in Sedimenten am Boden von Gewässern erhalten) den wasserferner lebenden Gruppen gegenüber sicherlich etwas überrespräsentiert. An der Kreide/Paläogen-Grenze ("KT-Event") starb eine Reihe urtümlicher kreiderzeitlicher Gruppen aus.
Erst im Paläogen (auch hier noch besonders mit Wasservögeln) hat sich allmählich eine höhere Diversität eingestellt. Als Besonderheit aus dieser Zeit sind u.a. sehr große flugun-fähige Straußenvögel zu nennen, die besonders im Eozän aufgrund des noch geringen Entwicklungsgrades der carnivoren Säugetiere eine große Rolle als Endglieder in den Nahrungsketten der Kontinente spielten.
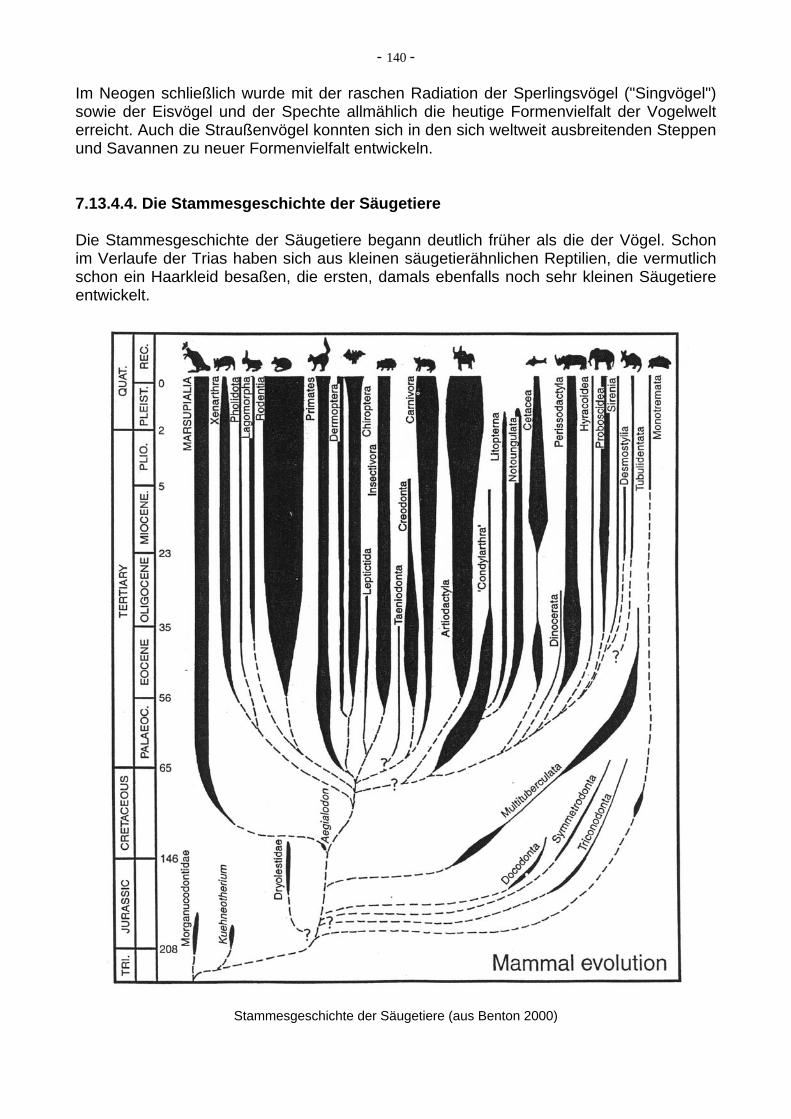
- 140 -
Im Neogen schließlich wurde mit der raschen Radiation der Sperlingsvögel ("Singvögel") sowie der Eisvögel und der Spechte allmählich die heutige Formenvielfalt der Vogelwelt erreicht. Auch die Straußenvögel konnten sich in den sich weltweit ausbreitenden Steppen und Savannen zu neuer Formenvielfalt entwickeln. 7.13.4.4. Die Stammesgeschichte der Säugetiere Die Stammesgeschichte der Säugetiere begann deutlich früher als die der Vögel. Schon im Verlaufe der Trias haben sich aus kleinen säugetierähnlichen Reptilien, die vermutlich schon ein Haarkleid besaßen, die ersten, damals ebenfalls noch sehr kleinen Säugetiere entwickelt.
Stammesgeschichte der Säugetiere (aus Benton 2000)

- 141 -
Im weiteren Verlauf des Mesozoikums blieben die Säugetiere dann, trotz allmählich steigender Formenvielfalt, klein und - relativ - unbedeutend, die Reptilien waren eindeutig die vorherrschende Gruppe. Die Säugetiere waren im Jura schon etwas formenreicher als in der Trias (vor allem im Oberjura), blieben aber noch immer sehr klein (bis rattengroß). Auch in der Kreide gab es weiterhin nur kleine Formen. Es entwickelten sich aber schon die ersten Beuteltiere (Marsupialia) und Plazentatiere. Die dominierende Gruppe waren die im Paläogen ausgestorbenen Multituberculaten. Das änderte sich aber - in geologischen Zeitbegriffen - schlagartig nach dem Aussterben der meisten Reptiliengruppen mit dem Ende der Kreidezeit (KT-Event). Die wichtigsten Konkurrenten in den terrestrischen Ökosystemen waren verschwunden, eine große Zahl von Nischen neu zu besetzen. Die daraufhin einsetztende Radiation der Säugetiere im Paläogen führte relativ schnell zu einer hohen Diversität und - vor allem im Oligozän - zu immensen Körpergrößen. Zu Beginn des Paleozäns gab es noch auschließlich relativ kleine Tiere (max. Hundegröße), später jedoch erfolgte ein rasches Größenwachstum in vielen Gruppen. Im Unter-Eozän existierten bereits die meisten modernen Ordnungen der Mammalia. Während des weiteren Eozäns ist eine ständig zunehmende Formenvielfalt und Verdoppelung der Familienzahl zu verzeichnen, hierunter modernere Pflanzenfresser der Unpaarhufer und der Paarhufer. Die bekannte Pferdereihe nahm ihren Anfang, ebenfalls die der Rüsseltiere. Die Nagetiere verdrängten die Multituberculaten. Als Carnivoren kamen erste Hunde, Katzen und Wiesel vor. Ab dem Oligozän gab es Affen. Im Oligozän fand bei einigen Gruppen von Pflanzenfressern ein enormes Größenwachs-tum statt, es gab Riesenformen (Schulterhöhe = Gesamthöhe der heutigen Giraffen) unter den Nashörnern. Dieses Phänomen wurde sicherlich durch die klimatisch bedingte zunehmende Versteppung der Waldlandschaften mit verursacht. 7.13.4.5. Die Entwicklungsgeschichte des Menschen Erste wirklich echte Affen gab es bereits im Oligozän. Noch ältere Funde sind den Affen immer unähnlicher, aber doch zu den Primaten zu rechnen. Ab dem Miozän entwickelten sich die Hominoiden (menschenähnliche Primaten). Frühe Menschenaffen gab es schon am Beginn des Miozäns mit den Dryopithecinen, die ab ca. 20 Ma. in Afrika entstanden und nach der Kollision von Afrika mit Asien vor ca. 15 Ma Eauch nach Eurasien einwanderten. In diese Zeit fällt auch die Entstehung einer anderen frühen Menschenaffengruppe, den Ramapithecinen. Beide Gruppen waren im Miozän weit verbreitet und relativ divers, vor allem in der alten Welt. Ihre Vertreter waren z.T. bodenlebend und sehr groß (Giganthopithecus, etwa wie ein Gorilla), z.T. lebten sie auf Bäumen (bekannteste Gattung Proconsul).
Die Familien der höher entwickelten Primaten. Moderne Affen (Pongiden) sind fossil kaum bekannt. Die Hominiden reichen ca. 6 Ma zurück (siehe Abbildung unten). Verändert nach Stanley (1999).
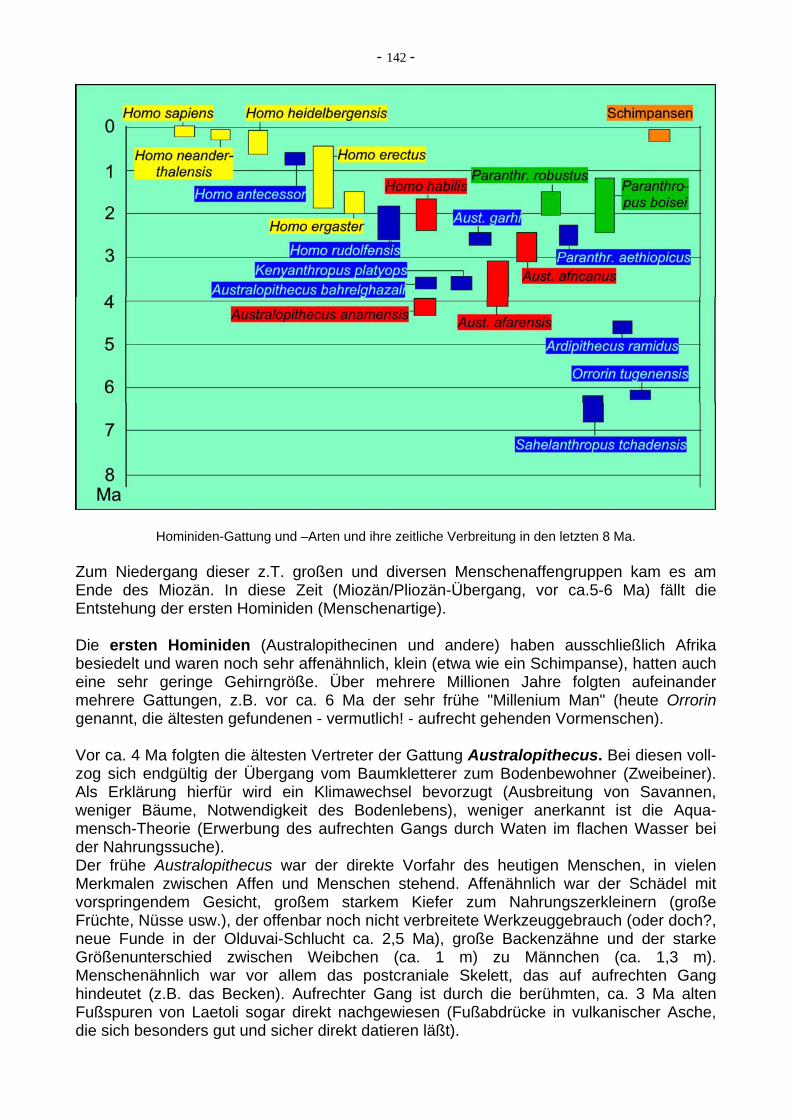
- 142 -
Hominiden-Gattung und –Arten und ihre zeitliche Verbreitung in den letzten 8 Ma. Zum Niedergang dieser z.T. großen und diversen Menschenaffengruppen kam es am Ende des Miozän. In diese Zeit (Miozän/Pliozän-Übergang, vor ca.5-6 Ma) fällt die Entstehung der ersten Hominiden (Menschenartige). Die ersten Hominiden (Australopithecinen und andere) haben ausschließlich Afrika besiedelt und waren noch sehr affenähnlich, klein (etwa wie ein Schimpanse), hatten auch eine sehr geringe Gehirngröße. Über mehrere Millionen Jahre folgten aufeinander mehrere Gattungen, z.B. vor ca. 6 Ma der sehr frühe "Millenium Man" (heute Orrorin genannt, die ältesten gefundenen - vermutlich! - aufrecht gehenden Vormenschen). Vor ca. 4 Ma folgten die ältesten Vertreter der Gattung Australopithecus. Bei diesen voll-zog sich endgültig der Übergang vom Baumkletterer zum Bodenbewohner (Zweibeiner). Als Erklärung hierfür wird ein Klimawechsel bevorzugt (Ausbreitung von Savannen, weniger Bäume, Notwendigkeit des Bodenlebens), weniger anerkannt ist die Aqua-mensch-Theorie (Erwerbung des aufrechten Gangs durch Waten im flachen Wasser bei der Nahrungssuche). Der frühe Australopithecus war der direkte Vorfahr des heutigen Menschen, in vielen Merkmalen zwischen Affen und Menschen stehend. Affenähnlich war der Schädel mit vorspringendem Gesicht, großem starkem Kiefer zum Nahrungszerkleinern (große Früchte, Nüsse usw.), der offenbar noch nicht verbreitete Werkzeuggebrauch (oder doch?, neue Funde in der Olduvai-Schlucht ca. 2,5 Ma), große Backenzähne und der starke Größenunterschied zwischen Weibchen (ca. 1 m) zu Männchen (ca. 1,3 m). Menschenähnlich war vor allem das postcraniale Skelett, das auf aufrechten Gang hindeutet (z.B. das Becken). Aufrechter Gang ist durch die berühmten, ca. 3 Ma alten Fußspuren von Laetoli sogar direkt nachgewiesen (Fußabdrücke in vulkanischer Asche, die sich besonders gut und sicher direkt datieren läßt).

- 143 -
Eines der ältesten Exmplare von Australopithecus ist Lucy, sicher das berühmteste Einzelfossil der Welt. Gefunden wurde Lucy 1973 in Äthiopien, sie war ein erwachsenes Weibchen (sehr klein!) von A. afarensis. Die Männchen dazu waren wesentlich größer. Trotz des aufrechten Ganges verbrachten diese frühen Arten auch viel Zeit in Bäumen, denn sie hatten noch lange Arme, lange gebogene Finger und Zehen und andere darauf hindeutende Skelettmerkmale. Die "robusten" Typen von Australopithecus und nahe verwandten Gattungen (boisei, robustus und andere Neufunde) waren noch etwas größer (1,5 Meter im Durchschnitt), robuster, hatten kräftigere Zähne und Kiefer, einen Scheitelkamm auf dem Schädel (Muskelansatz) und extreme Überaugenwülste. Diese kräftigen Typen waren sicher nicht die direkten Vorfahren des heutigen Menschen, sondern stellen ausgestorbene Spezialan-passungen dar (siehe Abbildung oben).
Die wesentlich fortgeschrittenere Gattung Homo entstand vor ca. 2-2,5 Ma ebenfalls in Afrika. Erste gesicherte Funde sind H. rudolfensis und H. habilis). Sie zeigen ein größeres Hirnvolumen als Australopithecus (ca. 650-800 ccm), erste sichere Steinwerkzeuge, aber noch einen primitiven, schräg abfallenden Schädel. Homo erectus (1,6-0,3 Ma) war noch größer und höher entwickelt. Seine Vertreter wurden früher oft mit anderen Art- und Gattungsnamen belegt (Pithecantropus, Peking-Mensch, Java-Mensch, Heidelberg-Mensch etc.). Es war der erste Mensch, der Afrika verließ und nach Eurasien auswanderte. Auch in Europa war er verbreitet (Heidelberg-Mensch, Speere von Schöningen usw.). H. erectus war schon ähnlich groß wie der moderne Mensch und nach seinem Knochenbau (v.a Becken) sogar ein besserer Läufer. Das Becken war enger, also besser fürs Laufen geeignet, aber schlechter für Intelligenz, da große Babyköpfe bei der Geburt nicht hindurchpaßten. Beim modernen Menschen ist dies umgekehrt - ergo wurde das Laufen der Intelligenz geopfert! Das Hirnvolumen von H. erectus betrug 800 bis (selten)1300 ccm und nahm im Laufe der Evolution etwas zu, sein Schädel war noch etwas länger und niedriger als bei H. sapiens (ein primitives Merkmal).
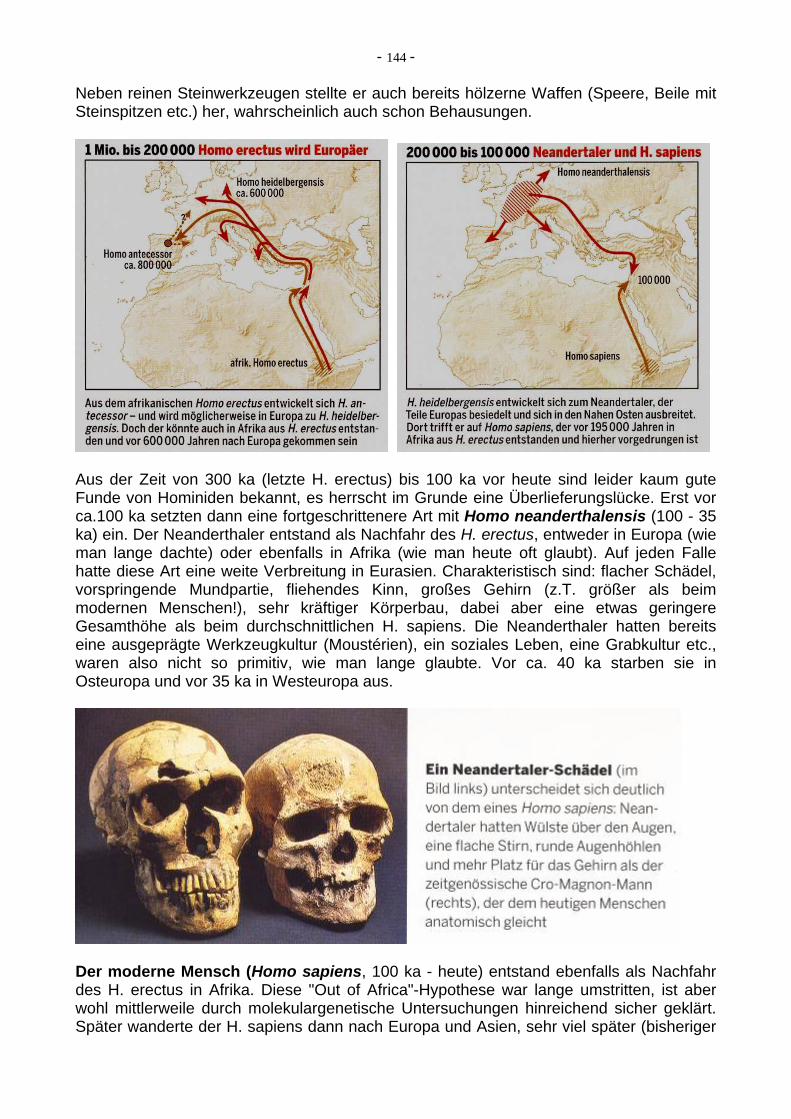
- 144 -
Neben reinen Steinwerkzeugen stellte er auch bereits hölzerne Waffen (Speere, Beile mit Steinspitzen etc.) her, wahrscheinlich auch schon Behausungen.
Aus der Zeit von 300 ka (letzte H. erectus) bis 100 ka vor heute sind leider kaum gute Funde von Hominiden bekannt, es herrscht im Grunde eine Überlieferungslücke. Erst vor ca.100 ka setzten dann eine fortgeschrittenere Art mit Homo neanderthalensis (100 - 35 ka) ein. Der Neanderthaler entstand als Nachfahr des H. erectus, entweder in Europa (wie man lange dachte) oder ebenfalls in Afrika (wie man heute oft glaubt). Auf jeden Falle hatte diese Art eine weite Verbreitung in Eurasien. Charakteristisch sind: flacher Schädel, vorspringende Mundpartie, fliehendes Kinn, großes Gehirn (z.T. größer als beim modernen Menschen!), sehr kräftiger Körperbau, dabei aber eine etwas geringere Gesamthöhe als beim durchschnittlichen H. sapiens. Die Neanderthaler hatten bereits eine ausgeprägte Werkzeugkultur (Moustérien), ein soziales Leben, eine Grabkultur etc., waren also nicht so primitiv, wie man lange glaubte. Vor ca. 40 ka starben sie in Osteuropa und vor 35 ka in Westeuropa aus.
Der moderne Mensch (Homo sapiens, 100 ka - heute) entstand ebenfalls als Nachfahr des H. erectus in Afrika. Diese "Out of Africa"-Hypothese war lange umstritten, ist aber wohl mittlerweile durch molekulargenetische Untersuchungen hinreichend sicher geklärt. Später wanderte der H. sapiens dann nach Europa und Asien, sehr viel später (bisheriger

- 145 -
Stand, von einigen Forschern aber bestritten) nach Amerika aus, unter Verdrängung und/oder Ausrottung der Neanderthaler (möglicherweise auch Vermischung?). Aus der Frühphase dieser Wanderungen in Europa am bekanntesten ist der "Cro-Magnon-Mensch" mit seinen Höhlenmalereien im Mittelmeerraum. Offenbar trug der Mensch auch zur Ausrottung großer Säugetiere am Ende der letzten Eiszeit bei.
Eine der interessantesten und kontrovers diskutierten Fragen der letzten Jahre in diesem Umfeld ist: Waren Neanderthaler und moderner Mensch zwei getrennte Arten oder nur Rassen einer einzigen Biospezies? Nach dem postcranialem Skelett zu urteilen waren sie vielleicht eine Arte, nach dem Schädel aber eher nicht (trotz großen Hirns des Neanderthaler). Nach DNA-Untersuchungen waren sie eher zwei getrennte Arten, nach der "molekularen Uhr" erfolgte die Trennung sogar schon vor ca. 500-700 ka.

- 146 -
7.14. Massenaussterbe-Events im Phanerozoikum Seit es auf der Erde Organismen in einer gewissen Artenzahl (Diversität) gibt, sind diese immer wieder von Phasen erhöhter Aussterberaten dezimitert worden. Alle Tier- und Pflanzenarten sterben nach einer mehr oder weniger langen (im geologischen Zeit-maßstab eher kurzen) Zeit aus, aber in diesen Phasen des Massenaussterbens (auch Massenaussterbe-"Events" genannt) verstärkte sich das normale Aussterbe-"Hintergrund-rauschen" um eine Vielfaches. Im Proterozoikum gab es noch keine genügend große Diversität der Organismen, um Zeiten des Massenaussterbens erkennen zu können. Allenfalls ist ein mit der neoproterozoischen Eiszeit ("Schneeball Erde") zeitgleicher (und wohl dadurch bedingter) starker Artenrückgang bei den Acritarchen (einzellige Algen mit Sporopolleninhülle) in den jungpräkambrischen Meeren so zu bezeichnen. Die wirklich großen Massenaussterbe-Phasen, z.T. erhebliche Einschnitte in die Faunen-Diversität (nicht so in die Floren-Diversität!) fanden jedoch im Phanerozoikum statt. Neben einer ganzen Reihe von weniger bedeutenden Phasen verstärkten Massenaussterbens sind es vor allem fünf Zeitscheiben, in denen innerhalb weniger Millionen Jahre (oder sogar nur einer halben Millionen Jahre) bis zu 95 % aller marinen Tierarten ausstarben. Diese als "Die großen Fünf" bekannten Events sind: 1. das Massenaussterben im Oberordovizium 2. das Massenaussterben im Oberordevon ("Kellwasser-Event") 3. das Massenaussterben an der Perm/Trias-Grenze 4. das Massenaussterben an der Trias/Jura-Grenze 5. das Massenaussterben an der Kreide/Paläogen-Grenze ("KT-Event") Ein Problem bei der quantitativen Analyse der Aussterberaten ist natürlich immer die Tatsache, dass ja in der fossilen Überlieferung diejenigen Gruppen stark überrepräsentiert sind, die mineralisierte Hartteile haben und somit wesentlich besser erhaltungsfähig sind als z.B. "Würmer" ohne jegliche Skelette, Schalen etc. Trotzdem gibt es genügend Fossil-dokumentation (als Abdrücke oder Spurenfossilien z.B.), um auch bei solchen Gruppen die Überlebensraten im Zuge bedeutenderer Massenaussterbe-Events abschätzen zu können. Aus numerischen Berechnungen ist jedoch deutlich ersichtlich, dass das Ereignis an der Perm/Trias-Grenze, welches gleichzeitig die Grenze zwischen Paläozoikum und Mesozoikum definiert, das mit Abstand bedeutendste war. Das KT-Event hingegen (Kreide / Paläogen-Grenze) ist zwar aufgrund des damit verbundenen spektakulären Aussterbens der Dinosaurier und des Aufpralls eines großen Asteroiden auf der Erde das bekannteste, aber auf die marine Invertebratenfauna bezogen (und damit für das System Erde in all seinen komplizierten Stoffkreisläufen) das am wenigsten bedeutende Massenaussterbe-Event der "großen Fünf".
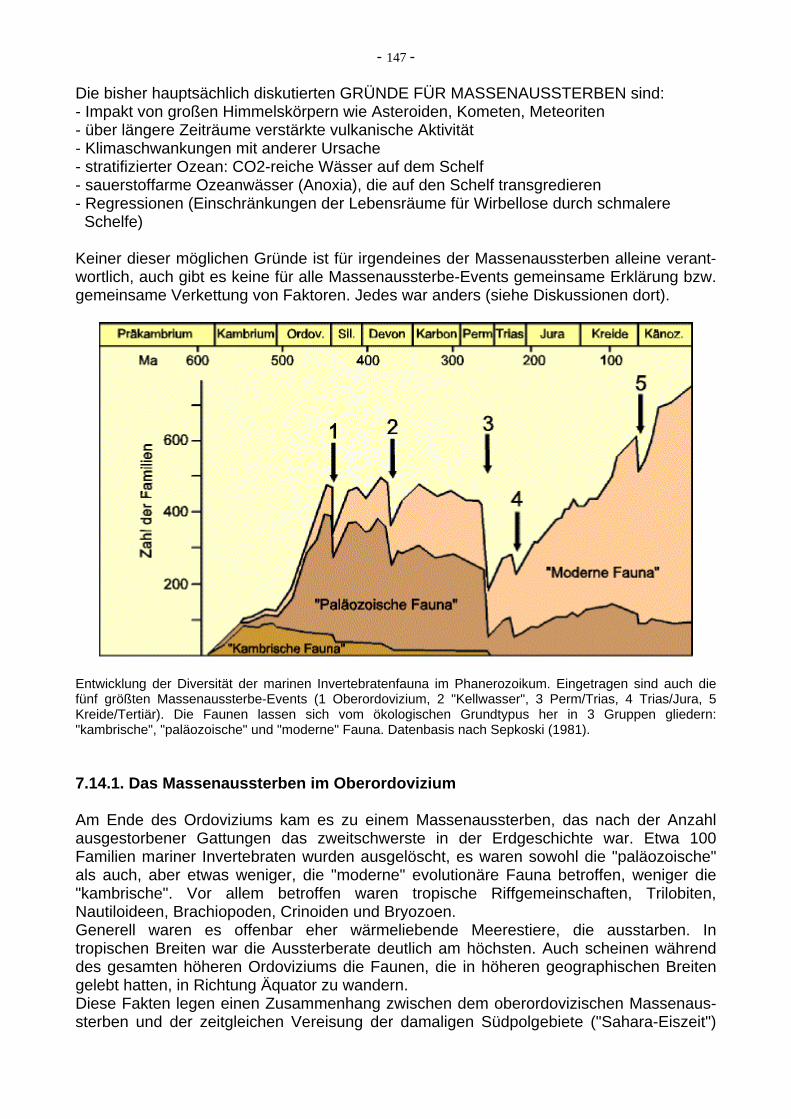
- 147 -
Die bisher hauptsächlich diskutierten GRÜNDE FÜR MASSENAUSSTERBEN sind: - Impakt von großen Himmelskörpern wie Asteroiden, Kometen, Meteoriten - über längere Zeiträume verstärkte vulkanische Aktivität - Klimaschwankungen mit anderer Ursache - stratifizierter Ozean: CO2-reiche Wässer auf dem Schelf - sauerstoffarme Ozeanwässer (Anoxia), die auf den Schelf transgredieren - Regressionen (Einschränkungen der Lebensräume für Wirbellose durch schmalere Schelfe) Keiner dieser möglichen Gründe ist für irgendeines der Massenaussterben alleine verant-wortlich, auch gibt es keine für alle Massenaussterbe-Events gemeinsame Erklärung bzw. gemeinsame Verkettung von Faktoren. Jedes war anders (siehe Diskussionen dort).
Entwicklung der Diversität der marinen Invertebratenfauna im Phanerozoikum. Eingetragen sind auch die fünf größten Massenaussterbe-Events (1 Oberordovizium, 2 "Kellwasser", 3 Perm/Trias, 4 Trias/Jura, 5 Kreide/Tertiär). Die Faunen lassen sich vom ökologischen Grundtypus her in 3 Gruppen gliedern: "kambrische", "paläozoische" und "moderne" Fauna. Datenbasis nach Sepkoski (1981). 7.14.1. Das Massenaussterben im Oberordovizium Am Ende des Ordoviziums kam es zu einem Massenaussterben, das nach der Anzahl ausgestorbener Gattungen das zweitschwerste in der Erdgeschichte war. Etwa 100 Familien mariner Invertebraten wurden ausgelöscht, es waren sowohl die "paläozoische" als auch, aber etwas weniger, die "moderne" evolutionäre Fauna betroffen, weniger die "kambrische". Vor allem betroffen waren tropische Riffgemeinschaften, Trilobiten, Nautiloideen, Brachiopoden, Crinoiden und Bryozoen. Generell waren es offenbar eher wärmeliebende Meerestiere, die ausstarben. In tropischen Breiten war die Aussterberate deutlich am höchsten. Auch scheinen während des gesamten höheren Ordoviziums die Faunen, die in höheren geographischen Breiten gelebt hatten, in Richtung Äquator zu wandern. Diese Fakten legen einen Zusammenhang zwischen dem oberordovizischen Massenaus-sterben und der zeitgleichen Vereisung der damaligen Südpolgebiete ("Sahara-Eiszeit")

- 148 -
nahe. Die zeitliche Auflösung der Ereignisse legt nahe, dass dieses Massenaussterben sich über einen wesentlich längeren Zeitraum erstreckte als die anderen "vier Großen" des Phanerozoikums. Auch geologische Ereignisse, die zu einem Massenaussterben in kürzerem Zeitrahmen hätten beitragen können, wie Asteroiden-Impacts, extrem langanhaltende Vulkanausbrüche oder Zusammenbrüche der ozeanischen Zirkulation konnten für das Oberordovizium bisher nicht nachgewiesen werden. Dies paßt auch zu der offenbar langen Zeitdauer des oberordovizischen Massenaussterbens. 7.14.2. Das Massenaussterben im Oberdevon ("Kellwasser-Event") Das Oberdevon war eine Zeit zyklische wiederholter Anoxia (sauerstoffarme bis -freie Wassermassen) auf den Schelfen der Kontinente. Diese Anoxia waren auch gleichzeitig verbunden mit (bzw. bewirkten) Zeiten verstärkten Aussterbens der wirbellosen Flachwas-serfaunen. Das stärkste dieser Events liegt recht genau an der Grenze zwischen unterem (Frasnium) und oberem (Famennium) Oberdevon und wird nach einem kleinen Flüßchen im Harz (dort ist ein bekanntes Profil, das diese Schichten gut erschließt) auch als "Kellwasser-Event" oder "Kellwasser-Horizont" bezeichnet. Unter den "großen Fünf" Massenaussterben des Phanerozoikums war das Kellwasser-Event nach der Anzahl ausgestorbener Gattungen das drittschwerste. Es waren sowohl die "paläozoische" als auch, etwas geringer, die "moderne" evolutionäre Fauna betroffen, weniger die "kambrische". Vor allem betroffen waren Riffgemeinschaften, Suspensionsfresser und große Räuber, weniger die Detritusfresser und kleinere bis mittelgroße Räuber. Eine Interpretation des oberdevonischen Massenaussterbens muß die in dieser Zeit besonders auffälligen zyklischen Anoxia berücksichtigen. Trotz gelegentlicher "Sensationsmeldungen" ist ein Nachweis populärerer Ereignisse wie Asteroiden-Impakts und Vulkan-Katastrophen bisher nicht gelungen.
Schematisches Modell zur Erklärung der Vorgänge um die Kellwasser-Horizonte und andere Anoxia im Oberdevon. Nach Joachimski & Buggisch (1996).

- 149 -
Über die Untersuchung der stabilen Isotopen des Kohlenstoffs (C12 und C13), die gute Aussagen über Bioaktivitäten, Sauerstoffgehalte in Wassermassen usw. (und generell über den Kohlenstoffkreislauf) zulassen, konnte ein kompliziertes Szenario wahrscheinlich gemacht werden (siehe Abbildung). Entscheidender Punkt für das wiederholte (und an der Frasnium/Famennium-Grenze besonders starke) Aussterben ist hierbei das Entstehen warmer salzhaltiger Tiefenwässer (WSDW = warm saline deep water), das im Zuge von Meeresspiegelanstiegen ozeanische Anoxia (sauerstoffarme bis -freie Wassermassen) auf die Schelfe drückte ("anoxic overturns") und so zum Absterben besonders der Riffsyste-me, der festgewachsenen Supensionsfresser sowie großer langsamer Räuber führte. Die Frage, warum diese Ereignisse gerade im Oberdevon solche tiefgreifenden Folgen hatten, ist nicht neu. Als Antwort darauf wurde wiederholt die Tatschache ins Spiel gebracht, dass das Oberdevon ja die Zeit der Erdgeschichte war, in der erstmalig ausgedehnte Wälder zumindest die Flachländer der Kontinente bedeckten. Vielleicht hat der verstärkte Eintrag organischen Materials aus dieser Quelle zu einem im Vergleich zu späteren Zeitabschnitten besonders starken Einfluß auf den Kohlenstoffkreislauf beigetragen, da die marinen Ökosysteme sich hierauf noch nicht haben einstellen können. Hinzu kommt, dass die Ablagerungssysteme auf dem Festland in der ersten Hälfte des Paläophytikums noch wenige "Fallen" für organischen Detritus bereitstellten, mäandrierende Flußsysteme mit stabilisierenden Uferwällen, Detritus aufnehmenden Altarmen und Überflutungssenken usw. dazwischen gab es noch kaum, totes Pflanzenmaterial konnte relativ ungehindert in größeren Mengen mit Flüssen in die Küstenregionen der Meere befördert werden. 7.14.3. Das Massenaussterben an der Perm/Trias-Grenze Der seit dem 19. Jahrhundert bekannte auffallende Unterschied zwischen den Fossilge-meinschaften vor und jenen nach der Perm-Trias Grenze hat schon früh zur Abgrenzung des Paläozoikums vom darauffolgenden Mesozoikum Anlaß gegeben. Es ist das mit Abstand größte Massenaussterbe-Ereignis der Erdgeschichte und damit auch das größte der "großen Fünf" des Phanerozoikums. Nach fundierten Einschätzungen starben 95% der Gattungen und Familien aus. Ganz eklatant betroffen waren unter den marinen Wirbellosen die Vertreter der "paläozoi-schen" evolutionären Fauna, etwas weniger die der "modernen" Fauna, während die "kambrische" evolutionäre Fauna, die aber schon vorher fast völlig verschwunden war, bis auf wenige "lebende Fossilien" (z.B. die inarticulate Brachiopode Lingula) endgültig von der Erde verschwand. Auch das Perm/Trias-Massenaussterben ist nicht auf eine einzige Erklärung zurückzu-führen, zumal es auch nicht einfach, sondern vielmehr aus mindestens zwei im Abstand von mehreren Millionen Jahren aufeinanderfolgenden Aussterbe-Höhepunkten zusam-mengesetzt ist. Diskutiert werden und wurden verschiedenste Faktoren wie Impakts, Vulkanismus, ozeanische Anoxia, Klimaänderungen, Regressionen usw. (siehe Auflistung unter Massenaussterben). Obwohl auch für die Perm/Trias-Grenze immer wieder "Sensationsmeldung" in der Presse auftauchen, ist der weithin (nicht von allen!) akzeptierte Stand bisher folgender: Mit der Vereinigung vieler Kontinente zum Superkontinent Pangäa kam es zur Zeit der Perm/Trias-Wende zu Regressionen und einer Verschmälerung der Schelfe in der ganzen Welt. Dies allein war schon negativ für eine hohe Biodiversität von Flachwasserfaunen, hinzu kam eine sehr eingeschränkte Anzahl von möglichen West-Ost-Wanderungswegen in niederen Breiten (auch dies bewirkte geringe Biodiversität). Somit war den Organismen bei Klimaveränderungen das Ausweichen sehr erschwert, die Gefahr des Aussterbens hoch.
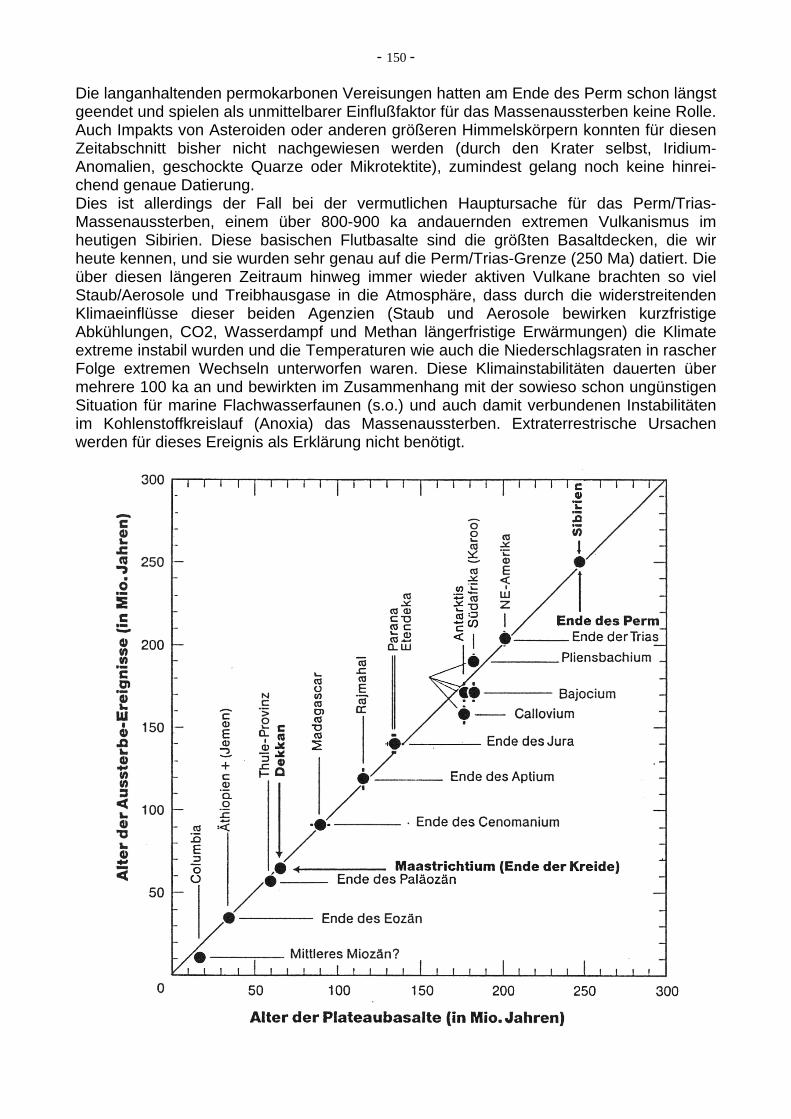
- 150 -
Die langanhaltenden permokarbonen Vereisungen hatten am Ende des Perm schon längst geendet und spielen als unmittelbarer Einflußfaktor für das Massenaussterben keine Rolle. Auch Impakts von Asteroiden oder anderen größeren Himmelskörpern konnten für diesen Zeitabschnitt bisher nicht nachgewiesen werden (durch den Krater selbst, Iridium-Anomalien, geschockte Quarze oder Mikrotektite), zumindest gelang noch keine hinrei-chend genaue Datierung. Dies ist allerdings der Fall bei der vermutlichen Hauptursache für das Perm/Trias-Massenaussterben, einem über 800-900 ka andauernden extremen Vulkanismus im heutigen Sibirien. Diese basischen Flutbasalte sind die größten Basaltdecken, die wir heute kennen, und sie wurden sehr genau auf die Perm/Trias-Grenze (250 Ma) datiert. Die über diesen längeren Zeitraum hinweg immer wieder aktiven Vulkane brachten so viel Staub/Aerosole und Treibhausgase in die Atmosphäre, dass durch die widerstreitenden Klimaeinflüsse dieser beiden Agenzien (Staub und Aerosole bewirken kurzfristige Abkühlungen, CO2, Wasserdampf und Methan längerfristige Erwärmungen) die Klimate extreme instabil wurden und die Temperaturen wie auch die Niederschlagsraten in rascher Folge extremen Wechseln unterworfen waren. Diese Klimainstabilitäten dauerten über mehrere 100 ka an und bewirkten im Zusammenhang mit der sowieso schon ungünstigen Situation für marine Flachwasserfaunen (s.o.) und auch damit verbundenen Instabilitäten im Kohlenstoffkreislauf (Anoxia) das Massenaussterben. Extraterrestrische Ursachen werden für dieses Ereignis als Erklärung nicht benötigt.
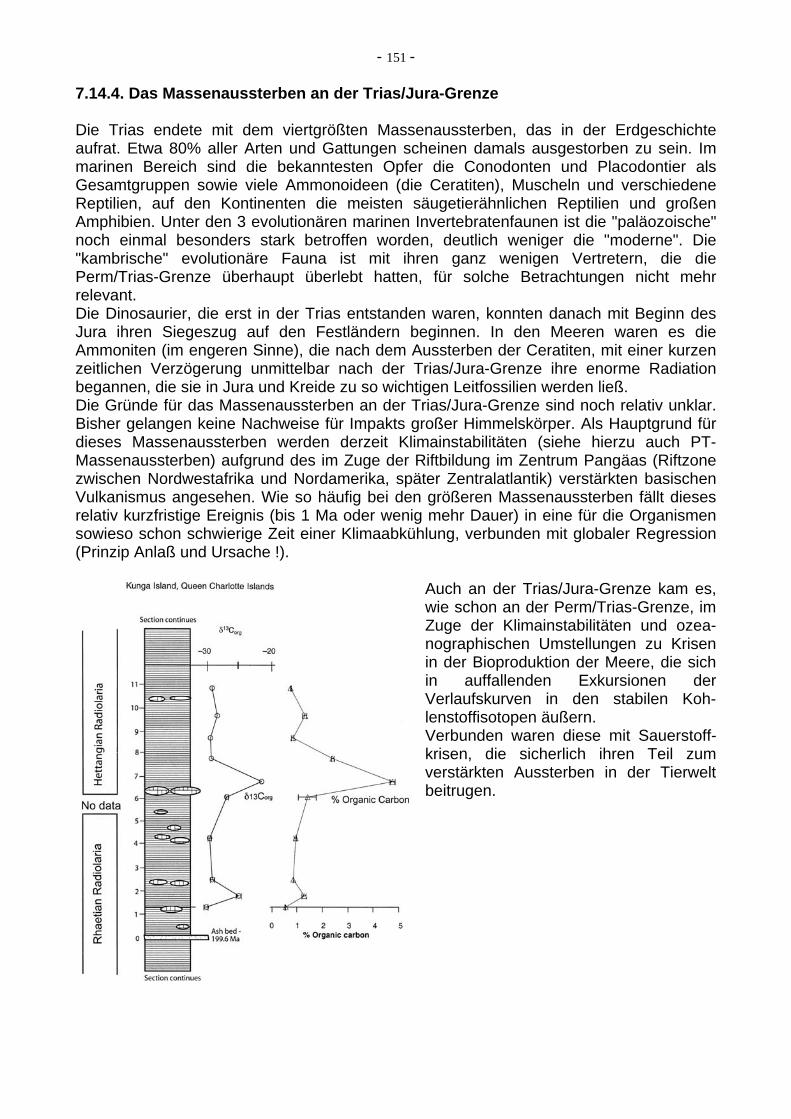
- 151 -
7.14.4. Das Massenaussterben an der Trias/Jura-Grenze Die Trias endete mit dem viertgrößten Massenaussterben, das in der Erdgeschichte aufrat. Etwa 80% aller Arten und Gattungen scheinen damals ausgestorben zu sein. Im marinen Bereich sind die bekanntesten Opfer die Conodonten und Placodontier als Gesamtgruppen sowie viele Ammonoideen (die Ceratiten), Muscheln und verschiedene Reptilien, auf den Kontinenten die meisten säugetierähnlichen Reptilien und großen Amphibien. Unter den 3 evolutionären marinen Invertebratenfaunen ist die "paläozoische" noch einmal besonders stark betroffen worden, deutlich weniger die "moderne". Die "kambrische" evolutionäre Fauna ist mit ihren ganz wenigen Vertretern, die die Perm/Trias-Grenze überhaupt überlebt hatten, für solche Betrachtungen nicht mehr relevant. Die Dinosaurier, die erst in der Trias entstanden waren, konnten danach mit Beginn des Jura ihren Siegeszug auf den Festländern beginnen. In den Meeren waren es die Ammoniten (im engeren Sinne), die nach dem Aussterben der Ceratiten, mit einer kurzen zeitlichen Verzögerung unmittelbar nach der Trias/Jura-Grenze ihre enorme Radiation begannen, die sie in Jura und Kreide zu so wichtigen Leitfossilien werden ließ. Die Gründe für das Massenaussterben an der Trias/Jura-Grenze sind noch relativ unklar. Bisher gelangen keine Nachweise für Impakts großer Himmelskörper. Als Hauptgrund für dieses Massenaussterben werden derzeit Klimainstabilitäten (siehe hierzu auch PT-Massenaussterben) aufgrund des im Zuge der Riftbildung im Zentrum Pangäas (Riftzone zwischen Nordwestafrika und Nordamerika, später Zentralatlantik) verstärkten basischen Vulkanismus angesehen. Wie so häufig bei den größeren Massenaussterben fällt dieses relativ kurzfristige Ereignis (bis 1 Ma oder wenig mehr Dauer) in eine für die Organismen sowieso schon schwierige Zeit einer Klimaabkühlung, verbunden mit globaler Regression (Prinzip Anlaß und Ursache !).
Auch an der Trias/Jura-Grenze kam es, wie schon an der Perm/Trias-Grenze, im Zuge der Klimainstabilitäten und ozea-nographischen Umstellungen zu Krisen in der Bioproduktion der Meere, die sich in auffallenden Exkursionen der Verlaufskurven in den stabilen Koh-lenstoffisotopen äußern. Verbunden waren diese mit Sauerstoff-krisen, die sicherlich ihren Teil zum verstärkten Aussterben in der Tierwelt beitrugen.
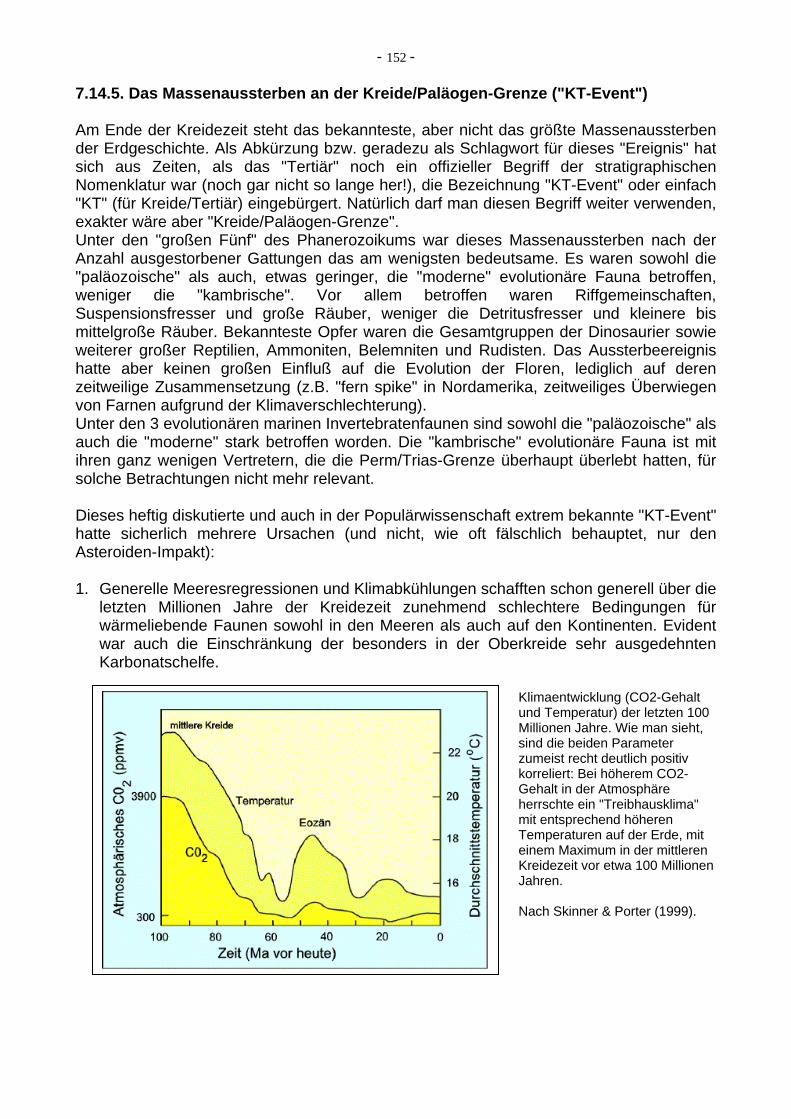
- 152 -
7.14.5. Das Massenaussterben an der Kreide/Paläogen-Grenze ("KT-Event") Am Ende der Kreidezeit steht das bekannteste, aber nicht das größte Massenaussterben der Erdgeschichte. Als Abkürzung bzw. geradezu als Schlagwort für dieses "Ereignis" hat sich aus Zeiten, als das "Tertiär" noch ein offizieller Begriff der stratigraphischen Nomenklatur war (noch gar nicht so lange her!), die Bezeichnung "KT-Event" oder einfach "KT" (für Kreide/Tertiär) eingebürgert. Natürlich darf man diesen Begriff weiter verwenden, exakter wäre aber "Kreide/Paläogen-Grenze". Unter den "großen Fünf" des Phanerozoikums war dieses Massenaussterben nach der Anzahl ausgestorbener Gattungen das am wenigsten bedeutsame. Es waren sowohl die "paläozoische" als auch, etwas geringer, die "moderne" evolutionäre Fauna betroffen, weniger die "kambrische". Vor allem betroffen waren Riffgemeinschaften, Suspensionsfresser und große Räuber, weniger die Detritusfresser und kleinere bis mittelgroße Räuber. Bekannteste Opfer waren die Gesamtgruppen der Dinosaurier sowie weiterer großer Reptilien, Ammoniten, Belemniten und Rudisten. Das Aussterbeereignis hatte aber keinen großen Einfluß auf die Evolution der Floren, lediglich auf deren zeitweilige Zusammensetzung (z.B. "fern spike" in Nordamerika, zeitweiliges Überwiegen von Farnen aufgrund der Klimaverschlechterung). Unter den 3 evolutionären marinen Invertebratenfaunen sind sowohl die "paläozoische" als auch die "moderne" stark betroffen worden. Die "kambrische" evolutionäre Fauna ist mit ihren ganz wenigen Vertretern, die die Perm/Trias-Grenze überhaupt überlebt hatten, für solche Betrachtungen nicht mehr relevant. Dieses heftig diskutierte und auch in der Populärwissenschaft extrem bekannte "KT-Event" hatte sicherlich mehrere Ursachen (und nicht, wie oft fälschlich behauptet, nur den Asteroiden-Impakt): 1. Generelle Meeresregressionen und Klimabkühlungen schafften schon generell über die
letzten Millionen Jahre der Kreidezeit zunehmend schlechtere Bedingungen für wärmeliebende Faunen sowohl in den Meeren als auch auf den Kontinenten. Evident war auch die Einschränkung der besonders in der Oberkreide sehr ausgedehnten Karbonatschelfe.
Klimaentwicklung (CO2-Gehalt
und Temperatur) der letzten 100 Millionen Jahre. Wie man sieht, sind die beiden Parameter zumeist recht deutlich positiv korreliert: Bei höherem CO2-Gehalt in der Atmosphäre herrschte ein "Treibhausklima" mit entsprechend höheren Temperaturen auf der Erde, mit einem Maximum in der mittleren Kreidezeit vor etwa 100 Millionen Jahren. Nach Skinner & Porter (1999).

- 153 -
2. In diese schon länger anhaltenden schlechten und bereits zu einem leichten Ansteigen der Aussterberaten führenden Bedingungen hinein kam es nun zusätzlich für ca. 500 ka zu enorm verstärktem basischem Vulkanismus (Dekkantrapp in Indien) mit Schwefelaerosol- und Staubbildung (kurzfristige Abkühlungen) und Ausstoß von Treibhausgasen (langfristige Erwärmungen). Folge waren starke Klimainstabilitäten in diesem Zeitraum und darüber hinaus, die ähnlich wie an der Perm/Trias-Grenze diverse Folgen nach sich zogen, darunter auch Störungen im normalen Kohlenstoffkreislauf und in den ozeanischen Systemen, was zu nochmals beschleunigten Aussterberaten führte. Der zeitliche Zusammenhang ist mittlerweile so gut datiert (siehe Abbildung auf der nächsten Seite), dass es hierüber keinen Zweifel geben kann. Ein Zufall kann dieses Zusammentreffen kaum sein!
Synthese der paläomagneti-
3. D
YNKGEgBRbsgKob
schen, paläontologischen und geochronologischen Daten der Dekkan-Basalte. Die Laven flossen in mehreren Phasen während ca. 500 ka aus, mitten hierin befindet sich die K/T-Grenze.
Nach Courtillot (1999).
er vielzitierte Einschlag eines großen Himmelskörpers im Gebiet des heutigen ucatan (Chicxulub-Krater) mit seinen klimatischen Folgen (Zusammenbruch der ahrungsketten) ist ebenfalls mittlerweile sicher nachgewiesen, und zwar durch den rater selbst sowie Iridium-Anomalien, geschockte Quarze und Tektite in der KT-renzschicht weltweit. Aktuelle Daten könnten allerdings darauf hindeuten, dass der inschlag 200-300 ka zu früh erfolgte. Die Folgen des Impakts waren über einen eologisch kurzen Zeitraum ohne Zweifel äußerst dramatisch. Es sind gut fundierte erechnungen und Szenarien publiziert worden, die riesige Flutwellen, extrem saure egen, "atomare Winter", Zusammenbrüche der Nahrungsketten, weltweite Wald-rände, extrem hohe Temperaturen durch den Rücksturz in den Weltraum ge-chleuderten Fallouts ("microwave model") und ähnlich schlimme Dinge wahrscheinlich emacht haben. Auf keinen Fall stellt dieser Impalt aber die alleinige Ursache für das T-Massenaussterben dar, sondern er kam noch zusäztlich zu den beiden bengenannten und längerandauernden Faktoren 1 + 2 hinzu, was schließlich zu den ekannten Folgen führte.

- 154 -
Allgemeine Literatur zur Erdgeschichte Brinkmann, Roland (1991) (14.Aufl., durchgesehen von Strauch, F., neubearbeitet von
Krömmelbein, K.): Historische Geologie. Erd- und Lebensgeschichte. - 404 S. (+ 8 Falttafeln mit 21 strat.Übersichten); Enke-Verlag (später Spektrum-Verlag). (wird nicht mehr gedruckt, steht aber in allen Bibliotheken, wichtiges Nachschlagewerk).
Courtillot, Vincent (1999): Das Sterben der Saurier. – 136 S.; Enke-Verlag. (wird nicht mehr gedruckt, gebraucht um die 10 €).
Cowen, Richard (2000) (2. Aufl.): History of Life. – 432 S.; Blackwell Science. (34,99 GBP bei Blackwell – April 2007)
Faupl, Peter (2004): Historische Geologie. Eine Einführung. - 271 S; UTB Uni-Taschenbücher. (18,90 € bei Amazon – April 2007).
Gradstein,F., Ogg,J. & Smith,A. (eds.) (2004): A geologic time scale 2004. – 589 S.; Cambridge University Press. 61,90 €.
Lunine, Jonathan I. (1999): Earth – Evolution of a habitable world. – 344 S.; Cambridge University Press. (46,62 € bei Amazon – April 2007).
Murphy, M.A. & Salvador, A. (1999): International stratigraphic guide, an abridged version. – Episodes, 22: 255-271.
Rey, Jaques (1991): Geologische Altersbestimmung. – 195 S.; Enke-Verlag. (nicht mehr im Druck – leider !)
Rothe, Peter (2000): Erdgeschichte. Spurensuche im Gestein. – 240 S.; Wissenschaftliche Buchgesellschaft. (39.90 € bei Amazon – April 2007)
Stanley, Steven M. (2001) (Hrsg. v. Volker Schweizer, 2. Auflage): Historische Geologie. Eine Einführung in die Geschichte der Erde und des Lebens. - 710 S.; Spektrum Akademischer Verlag. (43,00 € bei Amazon - April 2007).
Stanley, Steven M. (2005): Earth System History. – 608 S.; Palgrave MacMillan. (65,60 € bei Amazon – April 2007)
Steininger, F.F. & Piller, W.E. (1999): Empfehlungen (Richtlinien) zur Handhabung der stratigraphischen Nomenklatur. – Courier Forsch.-Inst. Senckenberg, 209: 1-19; Frankfurt am Main.
Walter, Roland (2003): Erdgeschichte. Die Entstehung der Kontinente und Ozeane. 5. Auflage. – 325 S.; de Gruyter. (24,95 € bei Amazon – April 2007)
Windley, Brian F. (1996): The Evolving Continents. – 526 S.; Wiley. (wird nicht mehr gedruckt, gebraucht um die 40-50 €).
Meine derzeitigen Empfehlungen: Stanley (2001 oder 2005) für die Entwicklung der Organismen und allgemeine Zusammenhänge Faupl (2004) oder Walter (2003) für die Geologie. .... und natürlich dieses Skriptum !! Kaufempfehlung (wenn es denn ein Buch sein soll) Stanley 2005 Wichtige Websites: http://jan.ucc.nau.edu/~rcb7/globaltext.htmlhttp://jan.ucc.nau.edu/~rcb7/globaltext2.htmlhttp://www.scotese.com/
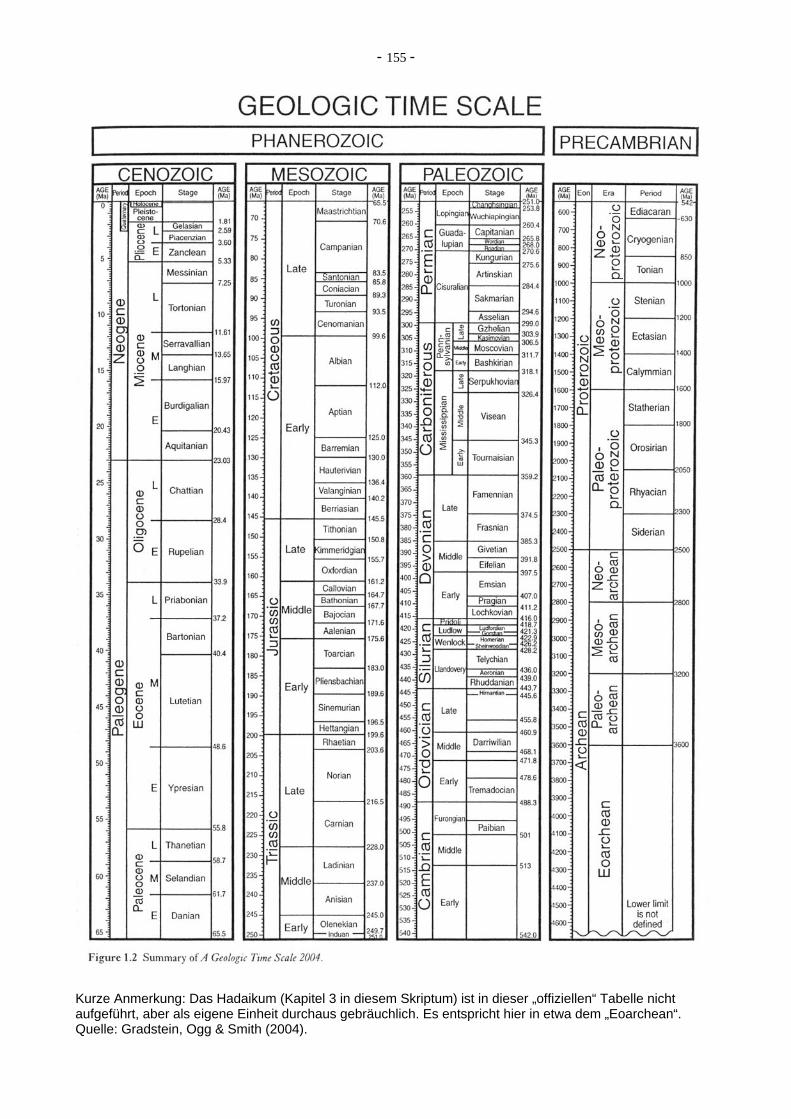
- 155 -
Kurze Anmerkung: Das Hadaikum (Kapitel 3 in diesem Skriptum) ist in dieser „offiziellen“ Tabelle nicht aufgeführt, aber als eigene Einheit durchaus gebräuchlich. Es entspricht hier in etwa dem „Eoarchean“. Quelle: Gradstein, Ogg & Smith (2004).
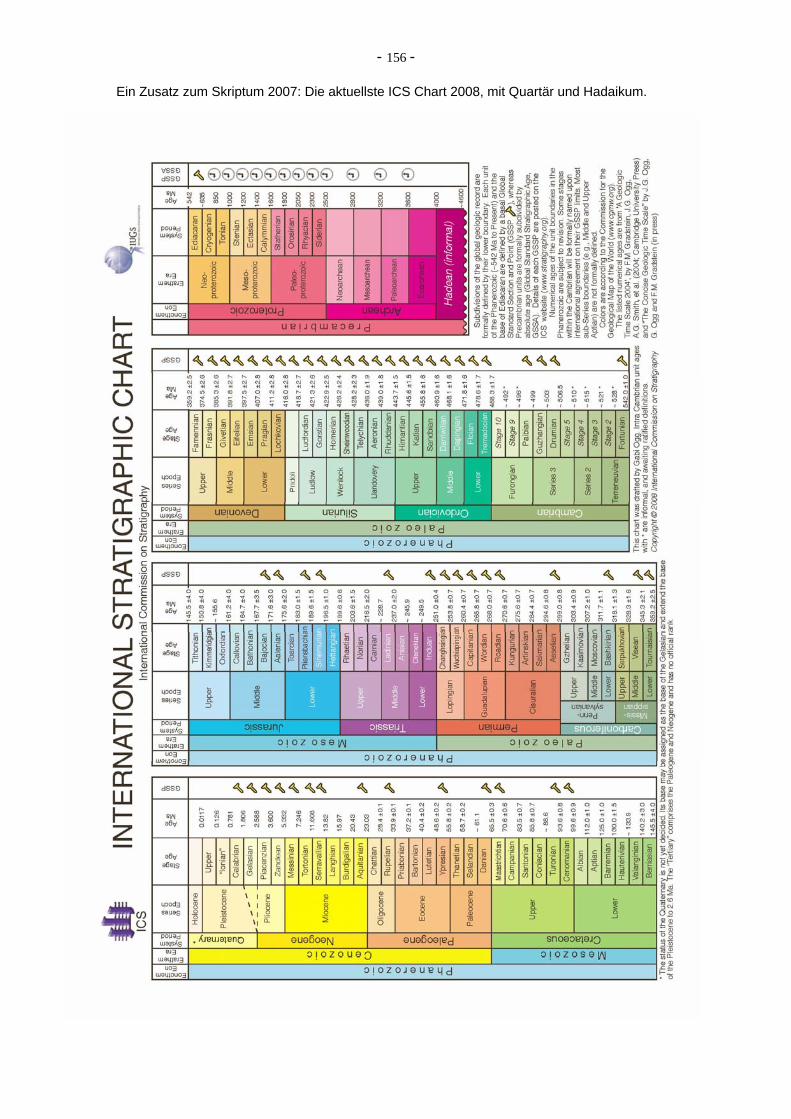
- 156 -
Ein Zusatz zum Skriptum 2007: Die aktuellste ICS Chart 2008, mit Quartär und Hadaikum.



















![GRAMMATIKÜBUNGEN Hauptstufe 1/2nemcinacezskype.weebly.com/uploads/4/1/2/2/... · Ich befürchte, dass ich zu spät komme. Ich befürchte[,] zu spät zu kommen. Ich befürchtete,](https://img.pdfslide.org/doc/110x75/5b9f90e809d3f2e02c8d5b28/grammatikuebungen-hauptstufe-1-ich-befuerchte-dass-ich-zu-spaet-komme-ich.jpg)









