Embed Size (px)
Citation preview

Aus dem Universitätsklinikum Münster
Institut für Medizinische Mikrobiologie
- Direktor: Univ.-Prof. Dr. med. Georg Peters -
Transkriptionsprofil wichtiger Regulatoren und Virulenzgene klinischer
Staphylococcus aureus-Isolate aus dem Respirationstrakt von
Mukoviszidose-Patienten
INAUGURAL – DISSERTATION
zur
Erlangung des doctor medicinae
der Medizinischen Fakultät
der
Westfälischen Wilhelms-Universität Münster
vorgelegt von
Ulrich, Katharina Maria Elisabeth
aus Münster
2013

Gedruckt mit Genehmigung der Medizinischen Fakultät der
Westfälischen Wilhelms-Universität Münster

Dekan: Univ.-Prof. Dr. med. Dr. h.c. Wilhelm Schmitz
1. Berichterstatterin: Prof. Dr. med. Barbara Kahl
2. Berichterstatter: Priv.-Doz. Dr. med. Alexander Mellmann
Tag der mündlichen Prüfung: 06.11.2013

Aus dem Universitätsklinikum Münster
Institut für Medizinische Mikrobiologie
Direktor: Univ.-Prof. Dr. med. Georg Peters
Referentin: Prof. Dr. med. Barbara Kahl
Koreferent: Priv.-Doz. Dr. med. Alexander Mellmann
Zusammenfassung
Transkriptionsprofil wichtiger Regulatoren und Virulenzgene klinischer
Staphylococcus aureus-Isolate aus dem Respirationstrakt von Mukoviszidose-Patienten
Ulrich, Katharina
In der vorliegenden Arbeit wurde die Genexpression von 16 klinischen S. aureus-Isolaten aus dem
Respirationstrakt von Mukovizidose-Patienten untersucht. Die verwendeten Isolate stammten aus einer
multizentrischen prospektiven Observationsstudie von Mukoviszidose-Patienten. Fokus der Untersuchung
war das Transkriptionsprofil der Regulatoren agr, in Form von RNA III, asp23 als sigB-abhängiges
Transkript und sae sowie das Transkriptionsprofil der Virulenzgene hla und spa. Zum Vergleich wurden
neben den Isolaten Laborstämme (SH 1000, RN 6390, 8325-4) und Mutanten (agr-, sar-, agr-/sar-, sigB-)
mitgeführt. Die phänotypische Analyse der Stämme erfolgte nach Anzucht auf Columbia-Blut- und
Schaedler-Agar, wobei letztgenannter Unterschiede im Phänotyp, wie den Hämolysegrad und die
Pigmentierung, noch besser erfasste. Die Isolate wurden je nach Ausmaß der Hämolyse in drei Gruppen
zusammengefasst. Da die RNA-Isolierung von Agarplatten nicht üblich ist, wurde die RNA der Stämme
zusätzlich noch aus BHI-Flüssigkulturen isoliert. Ziel war es, die relative Genexpression der Regulatoren
und Virulenzgene mittels der RT-PCR zu bestimmen, um mögliche Rückschlüsse auf den Phänotyp
ziehen zu können. Arbeiten, die eine Korrelation zwischen Phänotyp und erwarteter Genexpression
herstellten, gab es bisher nicht.
Ergebnis der Untersuchung ist die große Variation der Genexpression zwischen den klinischen Isolaten
untereinander aber auch im Vergleich zu den Mutanten und Laborstämmen. Der Phänotyp von 8325-4
entwickelte eine maximale Hämolyse und exprimierte wie erwartet hla in hohem Maße. Diese Hypothese
ließ sich bei den klinischen Isolaten nicht belegen. Stark hämolysierende Phänotypen (S 568 I) wiesen
teilweise auch eine geringere Hämolyse auf Schaedler-Agar auf. Allerdings folgte einer erfolgreichen
Transkription von sae die ebenfalls erfolgreiche Expression von hla bei Isolaten der Gruppe III.
Interessanterweise stellten sich zwei klinische Isolate (S 348 I, S 179 I) offenbar als spa-Mutanten heraus.
Bei zwei weiteren klinischen Stämmen (S 1368 II und III) könnte es sich um natürliche agr-Mutanten
handeln, da die RNA III-Expression auf den Agarplatten sehr gering war. Diese Erkenntnisse dienen als
Anstoß für weitere Untersuchungen, die sich mit den Mutationen in klinischen S. aureus-Isolaten aus
Mukoviszidose-Patienten befassen könnten.
Tag der mündlichen Prüfung: 06.11.2013

Eidesstattliche Erklärung
Ich gebe die eidesstattliche Erklärung ab, dass ich meine Dissertation über das
Transkriptionsprofil wichtiger Regulatoren und Virulenzgene klinischer
Staphylococcus aureus-Isolate aus dem Respirationstrakt von
Mukoviszidose-Patienten
Im Institut für Medizinische Mikrobiologie des
Universitätsklinikums Münster
Unter Anleitung von
Frau Prof. Dr. med. Barbara Kahl
1. selbstständig angefertigt,
2. weder in der gegenwärtigen, noch in der anderen Fassung als Dissertation,
Semesterarbeit, Prüfungsarbeit oder zur Erlangung eines akademischen Grades
vorgelegt,
3. nur unter der Benutzung der im Literaturverzeichnis angegebenen Arbeiten angefertigt
und sonst kein anderes gedrucktes oder ungedrucktes Material verwendet,
4. keine unerlaubte fremde Hilfe in Anspruch genommen habe.
Hamburg, den 05.05.2013
Katharina Ulrich

1 Inhalt
6
1 Inhalt
1 Inhalt ..........................................................................................................................6
2 Einleitung und Fragestellung .....................................................................................8
2.1 Staphylococcus aureus ........................................................................................8
2.2 Zystische Fibrose ................................................................................................9
2.3 Genexpression bei Bakterien ............................................................................11
2.4 Expression der Virulenzgene und Virulenzfaktoren von S. aureus ..................11
2.4.1 Der agr-Regulator......................................................................................13
2.4.2 Der sar-Regulator ......................................................................................14
2.4.3 Der sae-Regulator ......................................................................................15
2.4.4 Der sigB-Regulator ....................................................................................15
2.4.5 Die Hämolysine .........................................................................................16
2.4.6 Das Protein A.............................................................................................16
2.5 Bestimmung der Genexpression mittels Real-Time-PCR ................................17
2.6 Fragestellung und Ziel dieser Arbeit.................................................................19
3 Material und Methoden ...........................................................................................20
3.1 Materialien ........................................................................................................20
3.1.1 Bakterielle Stämme....................................................................................20
3.1.2 Chemikalien ...............................................................................................21
3.1.3 Geräte.........................................................................................................23
3.1.4 Software .....................................................................................................23
3.2 Methoden ..........................................................................................................23
3.2.1 Bakterienanzucht auf Columbia-Blut- und Schaedler-Agar ......................23
3.2.2 Bakterienanzucht in Flüssigkulturen zur Ermittlung des
Wachstumsverlaufes ................................................................................................24

1 Inhalt
7
3.2.3 RNA–Isolierung.........................................................................................25
3.2.4 Bestimmung von Konzentration und Reinheit der RNA ...........................26
3.2.5 cDNA-Transkription..................................................................................26
3.2.6 Real Time-Polymerasekettenreaktion........................................................27
4 Ergebnisse................................................................................................................30
4.1 Wachstumsverhalten auf Columbia-Blut- und Schaedler-Agar........................30
4.1.1 Häufigkeitsverteilung der Merkmale der klinischen Isolate ......................31
4.1.2 Das phänotypische Erscheinungsbild der Kolonien auf Columbia-Blut-
und Schaedler-Agar .................................................................................................33
4.2 Der Wachstumsverlauf in den Flüssigkultur .....................................................35
4.3 Die Ergebnisse der Real Time-PCR..................................................................37
4.3.1 Das Transkriptionsprofil der Laborstämme und Mutanten .......................37
4.3.2 Das Transkriptionsprofil der klinischen Isolate .........................................52
5 Diskussion ...............................................................................................................78
6 Literaturverzeichnis .................................................................................................85
7 Abbildungsverzeichnis ............................................................................................91
8 Tabellenverzeichnis .................................................................................................93
9 Abkürzungen ...........................................................................................................94
10 Anhang.....................................................................................................................95
11 Lebenslauf ...............................................................................................................96

2 Einleitung und Fragestellung
8
2 Einleitung und Fragestellung
2.1 Staphylococcus aureus
Staphylococcus aureus (S. aureus) gehört zur Familie der Staphylokokken (griech.
staphylos = Traube). Staphylokokken sind grampositive, fakultativ anaerobe,
Katalase-positive, typischerweise in der Gramfärbung in Haufen gelagerte, ca. 0,5-
1,7µm große Bakterien, die unbeweglich und nicht sporenbildend sind. Sie sind relativ
hitzestabil und können noch nach längerer Zeit aus getrocknetem Material kultiviert
werden.
Die Abgrenzung zu anderen Koagulase-negativen Staphylokokken (CoNS) geschieht
bei S. aureus durch die Produktion des Enzyms Koagulase, das Fibrinogen in Fibrin
umwandelt. Beim Wachstum auf Blut-Agar neigt S. aureus zur Bildung von weißlich
bis goldgelben, ß-hämolysierenden Kolonien. Daher stammt auch der Name S. aureus
(lat. aureus =golden).
S. aureus, ein opportunistischer Erreger, kolonisiert 20-50% der gesunden
Normalbevölkerung dauerhaft oder vorübergehend. Am häufigsten sind die vorderen
Nasenvorhöfe und der Perinealbereich besiedelt. Höher ist die Kolonisationshäufigkeit
bei chronisch Kranken, wie insulinpflichtigen Diabetikern und Hämodialyse-Patienten.
Zu einer hohen Morbidität und Mortalität führen vor allem die nosokomialen
Infektionen (38), aber auch bei Gesunden kann eine S. aureus-Infektion schwere
Verläufe aufweisen.
S. aureus kann oberflächliche Hautinfektionen wie Abszesse oder Impetigo
verursachen. Eine Infektion mit S. aureus kann aber auch zu schweren invasiven
Infektionen wie Arthritis, Osteomyelitis, Endokarditis oder Sepsis führen (38). Ein
erhöhtes Risiko haben vor allem hospitalisierte Patienten nach invasiven Eingriffen, wie
nach Anlage intravenöser Katheter oder chirurgischen Operationen, bei denen die
nosokomialen Infektionen mit S. aureus eine wichtige Rolle spielen. Ein großes
Problem bei der Behandlung von S. aureus-Infektionen ist das Auftreten von
Methicillin-resistenten S. aureus Stämmen (MRSA), bei denen inzwischen schon

2 Einleitung und Fragestellung
9
Resistenzen gegen das Reserveantibiotikum Vancomycin aufgetreten sind (25).
Zunehmend treten so genannte „community-acquired“ MRSA-Infektionen auf, an denen
vor allem gesunde Menschen erkranken, die nicht in einer medizinischen Einrichtung
behandelt worden sind (59).
S. aureus exprimiert eine Vielzahl an Virulenzfaktoren, die z.T. dazu führen, dass der
Erreger der Immunabwehr des Menschen entgeht und die zur Zerstörung des Gewebes
führen. Die Virulenzfaktoren werden eingeteilt in die Gruppe der Oberflächenproteine
und in die der extrazellulären Proteine. Wichtige Virulenzfaktoren, die in der Zellwand
verankert sind, sind das Protein A, das Fibronektin-bindende Protein und der Clumping-
Faktor A. Viele S. aureus-Stämme produzieren zudem eine Polysaccharidkapsel (48).
Ein Virulenzfaktor, der zunehmend bei CA-MRSA-Infektionen an Bedeutung gewinnt,
ist das Panton-Valentine-Leukozidin (PVL), das mit schweren Hautinfektionen und
nekrotisierenden Pneumonien assoziiert ist. So wurden auch schon PVL-positive
MRSA-Stämme aus der Lunge von CF-Patienten isoliert (16). Zu den extrazellulären
Proteinen gehört eine große Gruppe von Exotoxinen, wie die zu den Superantigenen
(SAgs) gehörenden Enterotoxine, SEs, SEIs und Toxic-Shock-Syndrome-Toxin-1
(TSST-1), die Exfoliativtoxine A und B und die Zytolysine, wie das PVL und die
Hämolysine.
Typischerweise werden die verschiedenen Virulenzfaktoren von S. aureus in
bestimmten Phasen des Wachstumszyklus exprimiert. So werden die
Oberflächenproteine in der exponentiellen Phase und die sezernierten Proteine vor allem
in der postexponentiellen und stationären Phase produziert (55).
2.2 Zystische Fibrose
Die zystische Fibrose (cystic fibrosis, CF, Synonym: Mukoviszidose) ist eine autosomal
rezessiv vererbbare Stoffwechselerkrankung. Es liegt eine Mutation eines Gens auf
Chromosom 7 vor, das für den „cystic fibrosis transmembrane conductance regulator“
(CFTR) kodiert. Die häufigste Mutation (ca. 70%) beruht auf einer Deletion von drei
Basenpaaren, die zum Fehlen von Phenylalanin (F) an Position 508 (∆ F508) im
CF-Genprodukt führt (13). Die Prävalenz liegt in den westlichen Ländern bei 1 von

2 Einleitung und Fragestellung
10
3000 Lebendgeburten. Über 50% der Patienten mit CF sind homozygot für die Mutation
∆ F508 (13), wobei die Schwere der Erkrankung stark variieren kann.
Typischerweise kommt es durch eine epitheliale Dysfunktion mit erhöhter
Natriumabsorption und verminderter Chloridsekretion zur Funktionsstörung der
exokrinen Drüsen. Ursächlich dafür ist das Fehlen der cAMP-aktivierten Chloridionen-
Transportfunktion des CFTR-Proteins (15). Außerdem scheint CFTR als Inhibitor der
epithelialen Natriumkanäle zu wirken. Das CFTR-Protein kommt in der apikalen
Epithelmembran verschiedener Organe vor und führt dort bei gestörter Funktion zu
einer erhöhten Viskosität der Sekrete.
Charakterisiert wird die zystische Fibrose klinisch durch verzögertes Wachstum,
Steatorrhoe, Infektionen und Pankreasinsuffizienz.
Die Infektionen in der Lunge werden durch die hohe Viskosität des Bronchialsekrets,
welches durch die erhöhte Natriumchloridkonzentration bedingt ist, begünstigt. Die
gestörte mukoziliäre Clearance verhindert, dass der Schleim abgehustet werden kann
und somit ein idealer Nährboden für bakterielle Keime entsteht. Folgen sind
rezidivierende Bronchitiden aufgrund der chronischen Inflammation und
Bronchiektasen, die zu Atelektasen führen. Typischerweise werden im Sputum von
CF-Patienten S. aureus, Pseudomonas aeruginosa und Burkholderia cepacia sowie
gelegentlich Escherichia coli und Klebsiellen nachgewiesen. S. aureus und
Haemophilus influenzae sind die Keime, die meist schon im frühen Kindesalter im
Sputum nachgewiesen werden, wenn in der Lunge noch keine ausgeprägte strukturelle
Schädigung vorliegt (51). Bei Jugendlichen und Erwachsenen ist P. aeruginosa der
führende Keim im bronchopulmonalen System. S. aureus ist in der Lage für Monate bis
Jahre in der Lunge der CF-Patienten trotz adäquater antibiotischer Therapie zu
persistieren (31). Ein Mechanismus, um sich an die Langzeittherapie mit Antibiotika zu
adaptieren, ist das Auftreten von „small colony variants“ (SCV), die noch resistenter
gegen die gängigen bei CF eingesetzten Antibiotika sind als der normale Phänotyp von
S. aureus (33).
Die chronische Beteiligung der Lunge führt zu weiteren Komplikationen, wie
Pneumothorax, Hämoptysen und im fortgeschrittenen Stadium zur respiratorischen
Insuffizienz und chronischem Cor pulmonale (15). Die Patienten fallen meist im frühen
Kindesalter mit chronischer Rhinosinusitis und Nasenpolypen auf.

2 Einleitung und Fragestellung
11
Die wesentlichen Säulen der Therapie sind die Förderung der Sekret-Clearance durch
gezielte Physiotherapie, die Gabe von rekombinanter humaner DNAase und die
rechtzeitige prolongierte antibiotische Therapie bei akut entzündlichen Schüben. Zur
Aufrechterhaltung einer regelrechten Verdauung werden Pankreasenzyme substituiert.
Wichtige Punkte in der antibiotischen Therapie im Zusammenhang von CF und
S. aureus sind die primäre Prophylaxe, die Eradikation und die Behandlung von
pulmonalen Exazerbationen.
Durch eine Verbesserung der therapeutischen Möglichkeiten in den letzten Jahren
beträgt die Lebenserwartung heute bei diesen Patienten mehr als 30 Jahre (13).
2.3 Genexpression bei Bakterien
Gene liegen nicht verstreut auf der DNA, sondern sind meistens zu mehreren als
regulierte Einheit zusammengefasst. Diese Einheiten werden als Operon bezeichnet.
Das zentrale Element der Signaltransduktion bei Bakterien bezieht sich auf ein
Zwei-Komponenten-System. Dieses besteht gewöhnlich aus zwei Proteinen, die in
einem einzelnen Operon kodiert werden. Bei einem der beiden Proteine handelt es sich
um den transmembrane sensor und bei dem anderen um den Transkriptionsfaktor oder
auch response regulator (52).
Alle Bakterien exprimieren ihre genetische Information in einem bestimmten Muster
abhängig von der Wachstumsphase, relativ wenige sogar durchgehend während des
gesamten Zellwachstums (konstitutiv).
2.4 Expression der Virulenzgene und Virulenzfaktoren von
S. aureus
Eine Reihe verschiedener regulatorischer Gene sind für die Expression der
Virulenzfaktoren von S. aureus verantwortlich.
Die Produktion der Virulenzfaktoren wird eng in Bezug auf die Zelldichte,
Umgebungseinflüsse und zur Verfügung stehender Energie kontrolliert und findet nur
bei entsprechendem Bedarf statt (45). Die Expression der Virulenzfaktoren bei
S. aureus ist abhängig von der jeweiligen Wachstumsphase. Bei der Kultivierung eines
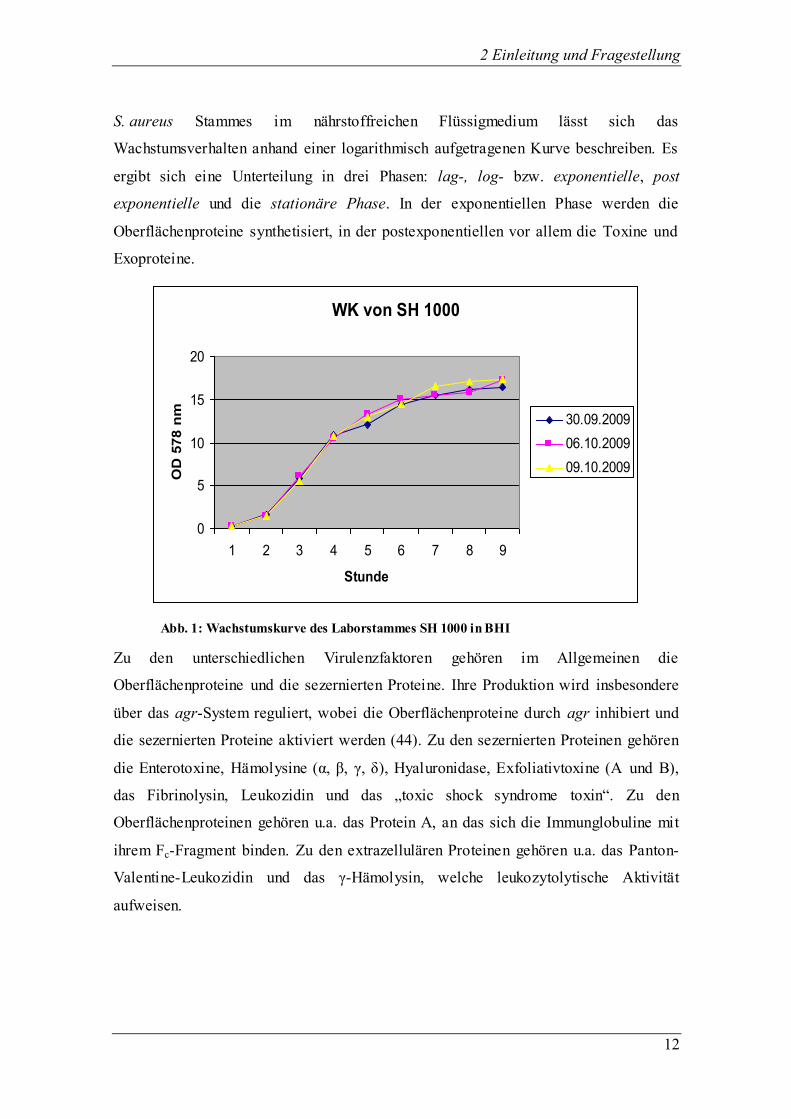
2 Einleitung und Fragestellung
12
S. aureus Stammes im nährstoffreichen Flüssigmedium lässt sich das
Wachstumsverhalten anhand einer logarithmisch aufgetragenen Kurve beschreiben. Es
ergibt sich eine Unterteilung in drei Phasen: lag-, log- bzw. exponentielle, post
exponentielle und die stationäre Phase. In der exponentiellen Phase werden die
Oberflächenproteine synthetisiert, in der postexponentiellen vor allem die Toxine und
Exoproteine.
Abb. 1: Wachstumskurve des Laborstammes SH 1000 in BHI
Zu den unterschiedlichen Virulenzfaktoren gehören im Allgemeinen die
Oberflächenproteine und die sezernierten Proteine. Ihre Produktion wird insbesondere
über das agr-System reguliert, wobei die Oberflächenproteine durch agr inhibiert und
die sezernierten Proteine aktiviert werden (44). Zu den sezernierten Proteinen gehören
die Enterotoxine, Hämolysine (α, β, γ, δ), Hyaluronidase, Exfoliativtoxine (A und B),
das Fibrinolysin, Leukozidin und das „toxic shock syndrome toxin“. Zu den
Oberflächenproteinen gehören u.a. das Protein A, an das sich die Immunglobuline mit
ihrem Fc-Fragment binden. Zu den extrazellulären Proteinen gehören u.a. das Panton-
Valentine-Leukozidin und das γ-Hämolysin, welche leukozytolytische Aktivität
aufweisen.
WK von SH 1000
0
5
10
15
20
1 2 3 4 5 6 7 8 9
Stunde
OD
578 n
m
30.09.2009
06.10.2009
09.10.2009

2 Einleitung und Fragestellung
13
2.4.1 Der agr-Regulator
agr (accessory gene regulator) ist ~3kb groß und besteht aus verschiedenen
Transkriptionseinheiten, die von den Promotoren P2 und P3 gesteuert werden (45). Es
handelt sich um einen zentralen Transkriptionsregulator von S. aureus, der die
Expression von Genen, die für Virulenz-assoziierte Proteine kodieren, kontrolliert. agr
wurde erstmals von Recsei et al. 1986 als globaler Regulator von S. aureus beschrieben
und nimmt eine Schlüsselfunktion in der Kontrolle der Expression von Exoproteinen ein
(54).
Das Regulator-Gen agr repräsentiert ein „quorum-sensing-System“, welches über die
bakterielle Dichte von S. aureus reguliert wird. Dabei werden sogenannte
autoinduzierende Peptide (AIP) sezerniert, die in Abhängigkeit von der bakteriellen
Populationsdichte zur Aktivierung des Regulators führen. Es werden vier verschiedene
agr-Gruppen I-IV synthetisiert. Die AIPs induzieren die Aktivierung des
Zwei-Komponenten-System von S. aureus derselben agr-Gruppe, während die AIPs
einer anderen agr-Gruppe den Regulator inhibieren. Das agr-Operon besteht aus den
beiden Promotoren P2 und P3, die für die Produktion der Transskriptionseinheiten RNA
II und RNA III verantwortlich sind (45). Das P3-Transkript RNA II besteht aus dem
Operon agrA, -B, -C und -D, die alle für die Aktivierung des Promotors P2 und die
folgende RNA III Transkription benötigt werden. RNA III ist das agr-Effektormolekül,
das im Wesentlichen für die Kontrolle der Synthese von Oberflächenproteinen und
extrazellulären Proteinen verantwortlich ist (44).
Die einzelnen vier agr-Gruppen werden mit verschiedenen S. aureus-bedingten
Krankheiten assoziiert. So wird die agr-Gruppe III mit dem „toxic-shock-Syndrom“ in
Verbindung gebracht (41) und die Gruppe IV korreliert mit dem „staphylococcal-
scaled-Skin-Syndrom“ (29). Bei den aus CF-Patienten isolierten S. aureus-Isolaten
konnte keine Korrelation zu einer bestimmten agr-Gruppe gefunden werden (32).
Bei RNA III handelt es sich um den agr-Effektor, der für das δ-Hämolysin Gen kodiert
und für die agr-vermittelte Regulation verantwortlich ist (44). Es agiert wechselseitig,
indem es die Transkription der meisten extrazellulären Proteine aktiviert und im
Gegenzug viele Oberflächenproteine inhibiert. So werden hla und spa von RNA III
reguliert, indem RNA III mit dem hla Transkript interagiert und dessen Translation
erleichtert (40). Die Unterdrückung von spa geschieht nicht nur auf

2 Einleitung und Fragestellung
14
Transkriptionsebene, sondern auch auf der RNA III-vermittelten Ebene der Translation
und ebenso durch den Abbau der spa-mRNA mit Hilfe der RNase III (27).
Der agr-Lokus spielt eine wichtige Rolle bei der Virulenz von S. aureus im Tiermodell
und daher wird schlussfolgernd vermutet, dass er ebenso eine Rolle bei humanen
Infektionen spielt. Obwohl die meisten klinischen S. aureus-Isolate, die bei humanen
Infektionen isoliert werden, agr+ sind, gibt es mehrere Berichte über die Isolation von
agr-defekten Mutanten aus Materialen infizierter Patienten. Es ist bekannt, dass der
agr-Lokus in vitro sehr labil ist.
Traber et al. beschäftigten sich mit Mutationen im agr-Lokus von klinischen S. aureus-
Isolaten und kamen zu der Erkenntnis, dass die Mutationen im Verlauf der Infektion
zustande gekommen waren (57).
Eine Studie von Goerke et al. kam zu dem Schluss, dass die Aktivität von agr bei einer
Infektion mit S. aureus bei der zystischen Fibrose keine wesentliche Relevanz hat. Sie
konnten zeigten, dass RNA III in den klinischen S. aureus-Isolaten nur gering
exprimiert wurde, was für ein inaktives agr in vivo sprach (21).
2.4.2 Der sar-Regulator
Bei sar (staphylococcal accessory regulator) handelt es sich um einen
Regulationslokus, der das agr-Operon in dessen Funktion beeinflusst. Es ist aus drei
verschiedenen sich überlappenden Transkripten sarA, -B und -C aufgebaut, die ihren
Ursprung in den Promotoren P1, P2 und P3 nehmen. Die Arbeitsgruppe um Manna
analysierte 1998 die Expression dieser Promotoren, um die Aktivierung des sar-Lokus
zu untersuchen. Die maximale Expression wurde bei einer Verbindung der P2-P3-P1-
Promoteren beobachtet. Desweiteren wurde mit der Untersuchung der Expression bei
einer sigB-Mutante gezeigt, dass P3 sigB abhängig ist (39). So kann sigB die Expression
von sarA steigern während es gleichzeitig die Expression von agr reduziert (3). Die
Expression der drei Transkripte variiert während des Wachstumszyklus, mit einem
Maximum der Expression von sarA und -B in der frühen exponentiellen Phase und von
sarC in der späten stationären Phase (2). sarA selbst ist in der Lage seine Produktion zu
unterdrücken, indem es die Transkription von P1 und P3 limitiert (12).

2 Einleitung und Fragestellung
15
Darüber hinaus ist sar notwendig für die agr-abhängige Regulation von RNA III (7).
Ebenso hat sar einen Einfluss auf die Transkription der Exoproteine α- und β-
Hämolysin (8).
2.4.3 Der sae-Regulator
Der Lokus staphylococcal accessory gene (sae) besteht aus einem Zwei-Komponenten-
Signaltransduktionssystem, das für die Gene saeR (response regulator), saeS (Histidin
Proteinkinase), saeQ (Membranprotein) und saeP (Lipoprotein) kodiert und das eine
Schlüsselfunktion in der Genregulation der Exoproteine aufweist. Es beeinflusst deren
Produktion während der Wachstumsphase und agiert auf der Transkriptionsebene.
Gesteigert wird die Expression von α- und β-Hämolysinen (hla, hlb), DNase und
Koagulase (coa). In einigen Fällen scheint es sich synergistisch zu agr zu verhalten. Es
findet aber keine Beeinflussung der Regulationsgene agr, sarA und σB statt, wobei diese
Gene allerdings im Gegenzug die Transkription von sae beeinflussen (46). sae wird vor
allem in der postexponentiellen Phase transkribiert (19).
2.4.4 Der sigB-Regulator
Das Operon für den alternativen Stress-Sigma-Faktor B (sigB) besteht aus vier Genen,
rsbU, rsbV, rsbW und sigB, dem ein σA-ähnlicher Promotor vorausgeht und das eine
innere σB-ähnliche Promotorsequenz enthält (63). Der alternative Sigma Faktor σB spielt
eine wichtige Rolle in der Regulation der Exoproteine und verhält sich generell
antagonistisch zu agr und kontrolliert zumindest teilweise die Transkription des
globalen Regulatorgens sar (14). Eine Inaktivierung des σB-Gens scheint im Vergleich
zum Wildtyp (RsbU-) zu einer gesteigerten Expression von sae zu führen (46).
Die Arbeitsgruppe um Gertz identifizierte 1999 das Alkalische-Schock-Protein asp23,
dessen Gen vom alternativen Stress-Sigma Faktor σB transkribiert wird und damit einzig
und allein von sigB abhängig ist. Dadurch wird asp23 zu einem idealen Marker, um die
Aktivität von sigB zu messen (18).
Phänotypisch zeigt eine ∆σB-Mutante eine verminderte Pigmentierung, eine
beschleunigte Sedimentierung und eine höhere Sensitivität gegenüber Katalase in der
stationären Phase (36).

2 Einleitung und Fragestellung
16
2.4.5 Die Hämolysine
Bekannt sind vier Membran schädigende Hämolysine, die von jedem Stamm exprimiert
werden können. Dazu gehören das α-, β-, γ- und das δ-Hämolysin bzw. -Toxin. Sie sind
zytotoxisch und begünstigen die Invasion des Erregers ins Gewebe.
Das hla-Gen kodiert für das α-Hämolysin und ist eines der am besten untersuchten
Virulenzfaktoren von S. aureus. Es wird in seiner Expression von den drei Loki agr,
sarA und sae reguliert. Eine Mutation im sae Gen führt zu einer erheblich verminderten
Expression von hla in vivo und in vitro während Mutationen in agr und sarA dagegen
keine Konsequenz auf die hla-Expression in vivo aufwiesen (20).
Das α-Toxin bildet Poren in der Zellmembran der Zielzellen, vor allem der
Erythrozyten, und führt dort zur osmotischen Lyse. Weiterhin stimuliert das α-Toxin die
Apoptose von Lymphozyten (30). Viele Studien zeigen, dass α-Toxin die S. aureus-
Infektion begünstigt und besonders bei durch S. aureus-induzierten Pneumonien ein
wichtiger Virulenzfaktor ist (53). Denn eine hla-Mutante war im Mausmodell weniger
virulent als der α-Toxin produzierende Elternstamm (62).
Das β-Hämolysin ist ebenfalls ein zytolytisches Toxin, welches in seiner Funktion wie
eine Sphingomyelinase agiert (60). Aarestrup et al. fanden 1999 heraus, dass nur 11%
von humanen Nasenabstrichen Gesunder und 13% von septischen Isolaten β-Hämolysin
positiv waren; allerdings waren Isolate von Rindern mit Mastitis zu 72% positiv. Bei
Versuchen in vitro wurde deutlich, dass das β-Hämolysin antagonistisch zum α-Toxin
reagiert, indem das β-Hämolysin an der Membran der Erythrozyten bindet und damit
die Fähigkeit des α-Toxins an dieser zu binden und zu multimerisieren inhibiert. Die
Wirkung von β-Hämolysin auf Blut-Agar wird als „hot-cold-lysis“ bezeichnet, da sich
die Hämolyse der Erythrozyten nach Inkubation unter Kälte deutlich klarer darstellt als
unter normalen Bedingungen bei 37°C (28).
2.4.6 Das Protein A
Das „staphylococcal“ Protein A (spa) ist ein wichtiger Virulenzfaktor, der ebenfalls
über agr reguliert wird. Er soll besonders bei S. aureus-induzierten Pneumonien als
inflammatorischer Faktor eine Rolle spielen (22). Bei SpA handelt es sich um ein in der
Zellwand von S. aureus verankertes Oberflächenprotein, das in über 90% der S. aureus-

2 Einleitung und Fragestellung
17
Stämme vorkommt (61). Es gehört zur Gruppe der MSCRAMM (microbial surface
components recognizing adhesive matrix molecules) (56). SpA bindet an die FC-Region
des IgG und verhindert dadurch die Antikörper-vermittelte Opsonierung und
beeinträchtigt so die Phagozytose (17). Ebenso bindet es den von-Willebrand-Faktor,
ein Serum-Glykoprotein, das die Plättchenaggregation induziert, und an TNFR1, einen
Rezeptor des Tumor-Nekrose-Faktors α (22). In früheren Studien konnte gezeigt
werden, dass SpA als Virulenzfaktor in Mausmodellen mit septischer Arthritis oder
Pneumonien eine wichtige Rolle spielt (49). Außerdem wurden in PVL-positiven CA-
MRSA-Isolaten eine verstärkte Produktion von SpA beobachtet (37).
Im Gegensatz zu den Exoproteinen wird spa kontinuierlich bis zur stationären Phase im
Wachstumszyklus transkribiert. Man geht davon aus, dass agr und sar die spa-
Produktion auf Transkriptionsebene regulieren, indem sie dessen Synthese unterdrücken
(10). Somit führt eine Inaktivierung von agr zu einem Anstieg des spa-Transkripts.
Sobald es aber zur Induktion des Effektorgens RNA III kommt, wird die Synthese von
spa eingestellt (58).
2.5 Bestimmung der Genexpression mittels Real-Time-PCR
Das Verfahren der quantitativen Real-Time PCR wurde erstmals 1992 von Higuchi et
al. beschrieben. Die Zugabe von Ethidiumbromid in den Reaktionsansatz ermöglichte
die simultane Amplifikation der doppelsträngigen DNA-Sequenzen und die Detektion
des Produkts durch den Anstieg der Fluoreszenz (24).
Zur Normalisierung der experimentellen Fehler ist es notwendig, die Transkription der
untersuchten Gene zur Transkription interner Referenzgene ins Verhältnis zu setzen.
Diese Gene werden als Haushalts-Gene bezeichnet, da sie im Metabolismus der
Bakterien konstitutiv und nicht-reguliert exprimiert werden und nicht durch äußere
Einflüsse, Zelltyp oder Zellstadium beeinflusst werden. So werden zur Normalisierung
sowohl ein als auch mehrere Haushalts-Gene verwendet, wobei die Verwendung
mehrerer Haushalts-Gene zu exakteren Ergebnissen in der RT-PCR führt, besonders
dann, wenn kleinere Änderungen in der Expression festgestellt werden sollen (47).
Die RT-PCR beruht auf dem Prinzip der konventionellen PCR. Der Unterschied
zwischen beiden Methoden besteht darin, dass bei der qRT-PCR die Messung des

2 Einleitung und Fragestellung
18
Amplifikats quantitativ in Echtzeit erfolgt. Die Quantifizierung erfolgt mit Hilfe von
Fluoreszenz-Messungen am Ende eines jeden PCR-Zyklus. Eine Möglichkeit, die
Fluoreszenz zu messen, ist der Gebrauch von interkalierenden Farbstoffen, wie das
SYBR Green I, das sich in die doppelsträngige DNA einlagert. Die Menge der
gemessenen Fluoreszenz erhöht sich exponentiell mit jedem weiteren PCR-Zyklus und
wird als relatives Maß für die amplifizierte Menge des Genfragmentes verwendet. Der
Vorteil des SYBR Green I besteht darin, dass es hitzestabil, selektiv für dsDNA und
hoch sensitiv ist (64).
Mit der RT-PCR gelingt es, die Transkription der interessierenden Gene zu messen.
Dazu muss zur Bestimmung der Genexpression in einem ersten Schritt die RNA der
Bakterien extrahiert werden. Als nächstes werden für die Amplifikation der cDNA zwei
Oligonukelotid-Primer benötigt, die die cDNA an beiden Enden flankieren. Dann wird
die cDNA durch reverse Transkription aus der ursprünglichen RNA synthetisiert. Die
eigentliche Reaktion der RT-PCR besteht aus Denaturation, Annealing und Elongation
(Abb. 2) und wird durch die Veränderung der Temperatur eingeleitet.
Abb. 2: RT-PCR Zyklus aus (34)
Zu Beginn des Zyklus wird eine hohe Temperatur von üblicherweise 95°C benötigt, um
die beiden Stränge der cDNA auf zu trennen. Die Höhe der Schmelztemperatur und die
Dauer des Schmelzvorganges sind abhängig von der Länge und der Sequenz des zu

2 Einleitung und Fragestellung
19
amplifizierenden DNA-Abschnittes. Damit sich die Primer an die DNA anlagern
können, wird die Temperatur gesenkt. Die Höhe dieser Temperatur hängt von den
Primern ab. Am Ende des Zyklus wird die Temperatur auf 72°C gesetzt. Das ist das
Optimum für die Amplifikation des DNA-Abschnittes (34).
Um die PCR-Produkte in Echtzeit messen zu können, lagert sich der
Fluoreszenzfarbstoff während der Amplifikation in die amplifizierte doppelsträngige
DNA ein. Zu Beginn der Reaktion ist das Fluoreszenzsignal schwach und reflektiert
dementsprechend nur eine kleine Menge an PCR-Produkt. Mit der Akkumulation des
Produktes kommt es im Verlauf zu einem exponentiellen Anstieg des Signals. Zum
Ende hin pendelt sich dies durch Sättigung auf dem Höhepunkt ein. Im Verlauf
überschreitet das Signal eine Schwelle, deren Schnittpunkt als Ct-Wert bezeichnet wird.
Der Ct-Wert gibt an, wie viele Zyklen benötigt werden, um diese Schwelle zu
überschreiten.
2.6 Fragestellung und Ziel dieser Arbeit
Die vorliegende Arbeit baut auf eine über zwei Jahre laufende multizentrische Studie
auf, in der aus 17 Mukoviszidose-Zentren Untersuchungsmaterial von 195 Patienten
mikrobiologisch untersucht wird und die S. aureus-Isolate besonders charakterisiert
werden, um durch Untersuchung verschiedener patienten- und pathogen-spezifischer
Parameter eine Unterscheidung zwischen S. aureus-Kolonisation und -Infektion treffen
zu können. Bei dem Untersuchungsmaterial handelt es sich um Nasen-, Rachenabstriche
und Sputum. Vierteljährlich werden die Materialien von den Mukoviszidose-Patienten
eingesandt und aus ihnen die S. aureus-Stämme isoliert.
In dieser Arbeit wurden S. aureus-Isolate, die auf Columbia-Blut- und Schaedler-Agar
ein bestimmtes Wachstumsmuster zeigten, hinsichtlich ihrer Genexpression mittels
Real-Time PCR untersucht. Dabei wurde der Frage nachgegangen, ob sich ein
Zusammenhang zwischen dem Wachstum der S. aureus-Isolate in Bezug auf
Hämolyseverhalten und Pigmentierung auf Columbia-Blut-Agar, bebrütet bei 37ºC, und
Schädler-Agar, bebrütet bei 37ºC unter 5% CO2, und dem Transkriptionsprofil dieser
Isolate ein Bezug auf die Aktivität der Regulatoren agr, sigB und sae und der
abhängigen Virulenzgene hla und spa herstellen lässt.

3 Material und Methoden
20
3 Material und Methoden
3.1 Materialien
3.1.1 Bakterielle Stämme
Die klinischen S. aureus-Isolate, die in der vorliegenden Arbeit untersucht wurden,
stammen aus einer multizentrischen prospektiven Observationsstudie von
Mukoviszidose-Patienten. Die Studie ist auf www.clinicaltrials.gov unter der
Kennzeichnung NCT00669760 abrufbar.
Es wurden 23 S. aureus-Stämme hinsichtlich ihres phänotypischen Erscheinungsbildes
auf Columbia-Blut- und Schaedler-Agar und ihrer Genexpression hin untersucht. Bei
den 23 Stämmen handelt es sich dabei um drei Wildtyp-Laborstämme (Tabelle 1), vier
isogene Deletions-Mutanten (Tabelle 2) und 16 klinische Isolate (Tabelle 3).
Tabelle 1: Laborstämme
Stamm Beschreibung Referenz
SH 1000 8325-4 rsbU+ (26)
RN 6390 genetischer Hintergrund dem von 8325-4 sehr
ähnlich; natürliche 11bp Deletion im rsbU Gen (42,50)
8325-4 Wildtyp, Standard Laborstamm (43,26)
Tabelle 2: Mutanten
Stamm Beschreibung Referenz
RN 6911
(RN 6390 ∆agr)
von RN 6390 abstammend, in dem der agr Lokus durch das tetM-Gen ersetzt wurde
(44)
ALC 136
(RN 6390 ∆sar)
isogenetische Mutante von RN6390 mit einer
sarA::ermC Mutation
(10)
ALC 135
(RN 6390 ∆agr/sar)
RN6390 mit agr::tetM und sar::Tn917LTV1 Insertion
(9)
ALC 1001
(RN 6390 ∆sigB)
sigB-Mutante von RN6390 abstammend (11)

3 Material und Methoden
21
Tabelle 3: klinische Isolate
Stamm Referenz
S 179 I aktuelle Studie
S 198 II „
S 348 I „
S 385 IV „
S 376 I „
S 501 II „
S 516 I „
S 568 I „
S 592 II „
S 605 I „
S 66 I „
S 1344 „
S 1365 I „
S 1365 II „
S 1368 II „
S 1368 III „
3.1.2 Chemikalien
- BBL™ Columbia-Blut-Agar BD, USA
Pro Liter: pankreatisch abgebautes Casein 10g
peptisch abgebautes Fleisch 5g
Hefeextrakt 5g
pankreatisch abgebautes Herzmuskelgewebe 3g
Maisstärke 1g
Natriumchlorid 5g
Agar 13,5g
5% steriles, defibriniertes Blut
- Schaedler-Agar BD, USA
Pro Liter: pankreatisch abgebautes Casein 8,2g
peptisch abgebautes Tiergewebe 2,5g
papainisch abgebautes Sojamehl 1g
Glucose 5,8g
Hefeextrakt 5g
Natriumchlorid 1,7g

3 Material und Methoden
22
Dikaliumphosphat 0,8g
L-Cystin 0,4g
Hämin 0,01g
Tris-(hydroxymethyl)-aminomethan 3g
Agar 13,5g
- Bacto™ Tryptische-Soja-Bouillon BD, USA
Pro Liter: pankreatisch abgebautes Casein 17g
papainisch abgebautes Soja 3g
Dextrose 2,5g
Natriumchlorid 5g
Dikaliumphosphat 2,5g
- Brain-Heart-Infusion (Hirn-Herz-Bouillon) Merck, Darmstadt
Pro Liter: Nährsubstrat (Hirn-, Herzextrakt u. Peptone) 27,5g
D(+)-Glucose 2,0g
Natriumchlorid 5,0g
di-Natriumhydrogenphosphat 2,5g
- LiChrosolv® Wasser für die Chromatographie Merck, Darmstadt
- RNA Protect ® Qiagen, Hilden
- FastRNA® Pro Blue Kit MP Biomedicals, Eschwege
RNApro™ Solution
DEPC-H2O
Lysing B Matrix
- Quantitect® Reverse Transcription Kit Qiagen, Hilden
gDNA Wipeout Buffer, 7x
Quantiscript® Reverse Transcriptase
Quantiscript RT Buffer, 5x
RT Primer Mix
RNase-Free Water
- iCycler iQ™ PCR Plates, 96 well Bio-Rad, USA
- iQTM SYBR green Supermix Bio-Rad, USA
- qPCR EvaGreen® Mastermix (5x concentrated) Segenetic, Borken
+ Fluorescein 100x concentrated Segenetic, Borken

3 Material und Methoden
23
3.1.3 Geräte
- Fast Prep FP120 Cell Disrupter Qbiogene, Frankreich
- Megafuge 1.OR Heraeus Instruments, Osterode
- Eppendorf Centrifuge 5417R Eppendorf, Hamburg
- Photometer Ultrospec 1100pro AmershamBiosciences, USA
- Thermomixer comfort Eppendorf, Hamburg
- Biophotometer 6131 Eppendorf, Hamburg
- iQ™5 Multicolor Real-Time PCR Detection System Bio-Rad, USA
- iCycler Bio-Rad, USA
3.1.4 Software
Bio-Rad iQ™5 Optical System Software, Version 2.1 Bio-Rad, USA
3.2 Methoden
3.2.1 Bakterienanzucht auf Columbia-Blut- und Schaedler-Agar
Der Columbia-Blut- und Schaedler-Agar wurden im 3-Ösen-Ausstrich mit S. aureus
beimpft und über Nacht im Brutschrank bei 37°C bebrütet, wobei der Columbia-Blut-
Agar unter aeroben Bedingungen und der Schaedler-Agar unter 5%igem CO2 bebrütet
wurde.
Am darauffolgenden Tag wurde die Beschreibung des Phänotyps der S. aureus-
Kolonien auf Blut- und Schaedler-Agar mit Hilfe eines standardisierten
Beurteilungsbogens (Anhang) durchgeführt, der für die CF-Studie entwickelt wurde.
Beurteilt wurden Hämolysegrad, β-Toxin, Pigmentierung und Größe der Kolonien,
Wachstum und Konsistenz. Der Hämolysegrad wurde eingeteilt in die Stufen 0 bis 4,
wobei „Null“ keine Hämolyse und „vier“ maximale Hämolyse bedeutete. Bei der
Pigmentierung wurde zwischen weiß, grau und gelb unterschieden. Die Größe der
Kolonien wurde von winzig über klein bis hin zu groß beschrieben. Beim Wachstum
gab es die Unterteilung in normal, klein und winzig. Die Konsistenz der Kolonien war
trocken, feucht oder schleimig.
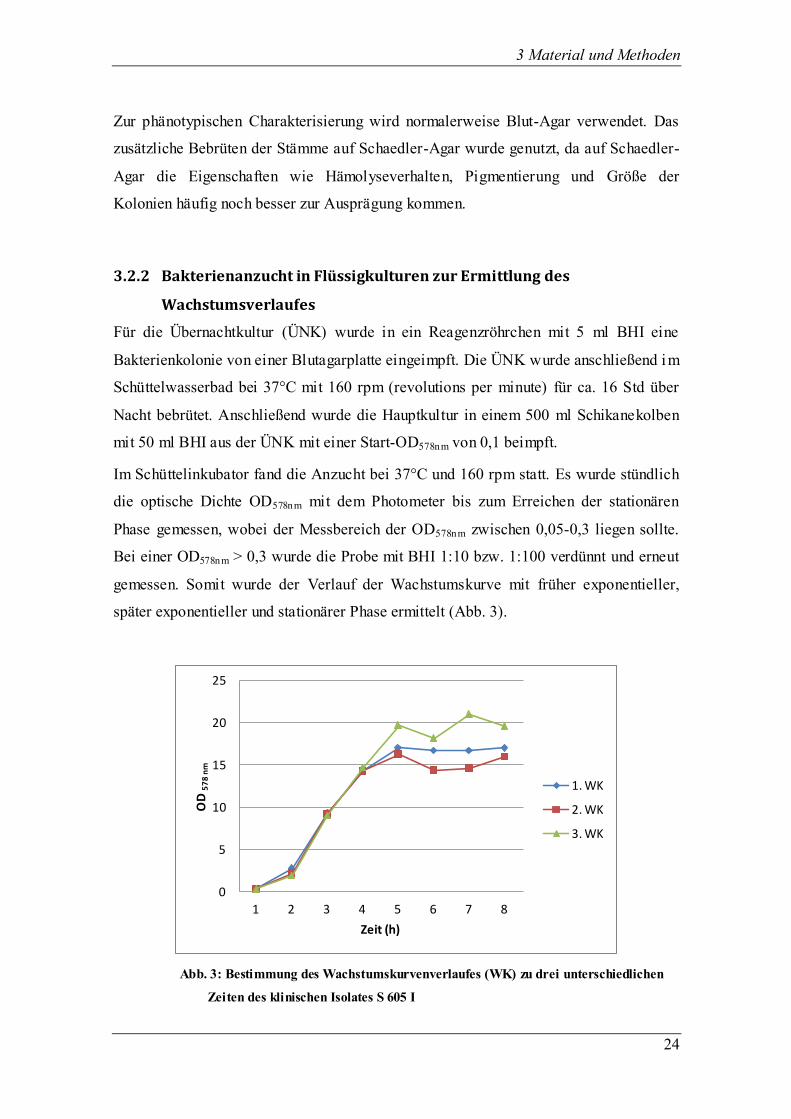
3 Material und Methoden
24
Zur phänotypischen Charakterisierung wird normalerweise Blut-Agar verwendet. Das
zusätzliche Bebrüten der Stämme auf Schaedler-Agar wurde genutzt, da auf Schaedler-
Agar die Eigenschaften wie Hämolyseverhalten, Pigmentierung und Größe der
Kolonien häufig noch besser zur Ausprägung kommen.
3.2.2 Bakterienanzucht in Flüssigkulturen zur Ermittlung des
Wachstumsverlaufes
Für die Übernachtkultur (ÜNK) wurde in ein Reagenzröhrchen mit 5 ml BHI eine
Bakterienkolonie von einer Blutagarplatte eingeimpft. Die ÜNK wurde anschließend im
Schüttelwasserbad bei 37°C mit 160 rpm (revolutions per minute) für ca. 16 Std über
Nacht bebrütet. Anschließend wurde die Hauptkultur in einem 500 ml Schikanekolben
mit 50 ml BHI aus der ÜNK mit einer Start-OD578nm von 0,1 beimpft.
Im Schüttelinkubator fand die Anzucht bei 37°C und 160 rpm statt. Es wurde stündlich
die optische Dichte OD578nm mit dem Photometer bis zum Erreichen der stationären
Phase gemessen, wobei der Messbereich der OD578nm zwischen 0,05-0,3 liegen sollte.
Bei einer OD578nm > 0,3 wurde die Probe mit BHI 1:10 bzw. 1:100 verdünnt und erneut
gemessen. Somit wurde der Verlauf der Wachstumskurve mit früher exponentieller,
später exponentieller und stationärer Phase ermittelt (Abb. 3).
Abb. 3: Bestimmung des Wachstumskurvenverlaufes (WK) zu drei unterschiedlichen
Zeiten des klinischen Isolates S 605 I
0
5
10
15
20
25
1 2 3 4 5 6 7 8
OD
57
8 n
m
Zeit (h)
1. WK
2. WK
3. WK

3 Material und Methoden
25
3.2.3 RNA–Isolierung
Die RNA-Isolierung der S. aureus-Stämme erfolgte nach Anzucht dieser auf Blut- bzw.
Schaedler-Agar und in dem Flüssigmedium Brain-Heart-Infusion (BHI) mit Hilfe des
FastRNA® Pro Blue Kit von qBiogene.
3.2.3.1 RNA-Isolierung von Columbia-Blut- und Schaedler-Agar
Es wurde 500 µl RNA Protect in einem 2 ml biopur Eppendorf Zentrifugenröhrchen
vorgelegt, anschließend der gesamte erste Ausstrich von der Agarplatte zugegeben und
beides gründlich homogenisiert. Dann wurde die Probe für fünf Minuten bei 5000 g bei
Raumtemperatur zentrifugiert. Der Überstand wurde verworfen. Nach diesem
Arbeitsschritt konnte das Pellet auch bei -20°C für einige Zeit eingefroren werden.
Die nächsten Arbeitsschritte durften nur mit speziellen Pipetten, gestopften
Pipettenspitzen, Handschuhen und unter dem Abzug durchgeführt werden, um jegliche
Verunreinigung durch Kontamination oder Partikel aus der Luft zu vermeiden.
Im nächsten Schritt wurde das Pellet in 1 ml RNApro™ Solution durch gründliches
Durchmischen im Vortex resuspendiert und in die Lysing B Matrix überführt. Diese
wurde in das Homogenisationssystem FastPrep® gesetzt und für 40 s bei einer
Aufschlussintensität von 6 gerüttelt, um die bakteriellen Zellen zu lysieren.
Anschließend wurde die Probe bei 12000 g für 5 min bei 4°C zentrifugiert. Der
Überstand wurde in ein neues 1,5 ml Zentrifugenröhrchen überführt, für 5 min bei
Raumtemperatur inkubiert und anschließend 300 µl Chloroform hinzugefügt und 10 s
im Vortex durchmischt. Nach weiteren 5 min Inkubationszeit bei Raumtemperatur
wurde wieder bei 12000 g für 5 min bei 4°C zentrifugiert. Die oberste RNA-enthaltende
Phase wurde, ohne die Interphase zu zerstören, abpipettiert und in ein neues
Zentrifugenröhrchen überführt. Nach Hinzufügen von 500 µl absolutem Ethanol und
fünfmaligem Invertieren folgte eine mindestens 30 minütige Lagerung bei -20°C.
Anschließend wurde die Probe erneut bei 12000 g für 5 min bei 4°C zentrifugiert und
der Überstand verworfen. Das weiße Pellet am Rand des Zentrifugenröhrchens, welches
die aufgereinigte RNA enthält, wurde mit 500 µl kaltem 75%igen Ethanol, hergestellt
mit DEPC-H2O, gewaschen und der Ethanol verworfen. Nach fünfminütigen Trocknen

3 Material und Methoden
26
bei Raumtemperatur wurde das Pellet in 100 µl DEPC-H2O resuspendiert. Eine
Lagerung der Probe war bei -80°C für längere Zeit möglich.
3.2.3.2 RNA-Isolierung aus der BHI-Flüssigkultur
Für die Isolierung der RNA aus der Flüssigkultur wurde der gleiche Ansatz des BHI-
Mediums verwendet wie zur Ermittlung des Wachstumsverlaufes.
An den zuvor ermittelten Zeitpunkten für die frühe exponentielle, späte exponentielle
und stationäre Phase wurden Proben entnommen. Zum Zeitpunkt der frühen
exponentiellen Phase wurden 2 ml, zu den beiden anderen Zeitpunkten je 500 µl Kultur
abpipettiert. Die Proben wurden im Zentrifugenröhrchen zu gleichen Teilen mit RNA
Protect versetzt und anschließend im Vortex gründlich durchmischt. Sie wurden zum
Inkubieren 5 min bei Raumtemperatur stehen gelassen und anschließend bei 4000 rpm
für 10 min bei 20°C zentrifugiert. Der Überstand wurde verworfen und das Pellet bei -
20°C zur Lagerung eingefroren oder es wurde die RNA-Isolierung (s.o.) direkt
angeschlossen.
3.2.4 Bestimmung von Konzentration und Reinheit der RNA
Zur Bestimmung der RNA-Konzentration wurde die RNA im Verhältnis 1:100 mit
HPLC-H2O und für die Bestimmung der Reinheit ebenfalls im Verhältnis 1:100 mit 10
mM Tris HCL versetzt. Gemessen wurde nach Eichen der Leerprobe mit dem
Eppendorf Photometer. Der Quotient aus A260nm/A280nm gab einen Hinweis auf die
Reinheit der RNA. Er lag idealerweise zwischen einer OD von 1,8 und 2,0, da ein Wert
unter 1,8 für eine Verunreinigung der Probe sprach und ein Wert von 2 eine optimale
Aufreinigung widerspiegelte.
3.2.5 cDNA-Transkription
Zur Transkription der RNA in cDNA wurde das QuantiTect® Reverse Transcription Kit
von der Firma Qiagen verwendet.
Sämtliche Arbeitsschritte wurden auf Eis durchgeführt. Das RNA-Template wurde auf
Eis aufgetaut, ebenso die Reagenzien gDNA Wipeout Buffer, Quantiscript Reverse

3 Material und Methoden
27
Transcriptase, Quantiscript RT Buffer und der RT Primer Mix. Das RNAse-freie
Wasser wurde bei Raumtemperatur aufgetaut. Folgender Reaktionsansatz wurde zur
Elimination der genomischen DNA verwendet:
2 µl gDNA Wipeout Buffer 7x sowie eine nach Konzentrationsbestimmung berechnete
Menge RNA wurden in ein 1,5 ml biopur Zentrifugenröhrchen gegeben und zuletzt
wurde RNAse freies Wasser auf einen Gesamtansatz von 24 µl ergänzt. Die Probe
wurde 2 min bei 42°C inkubiert. Danach wurde der Reverse-Transkriptions-Mastermix
zur Probe hinzugefügt, bestehend aus 2 µl Quantiscript Reverse Transkriptase, 8 µl
Quantiscript RT Buffer 5x und 2 µl RT Primer Mix. Es folgte ein weiterer
Inkubationsschritt für 15 min bei 42°C und direkt anschließend für 3 min bei 95°C, um
die reverse Transkriptase zu inaktivieren. Die erhaltene cDNA wurde bei -20°C
gelagert.
3.2.6 Real Time-Polymerasekettenreaktion
Die quantitative Real-Time-PCR (qRT-PCR) wurde in der vorliegenden Arbeit
verwendet, um das Transkriptionsprofil mehrerer wichtiger regulatorischer Gene von
S. aureus zu quantifizieren und mit dem phänotypischen Erscheinungsbild auf
Columbia-Blut- und Schädler-Agar in Verbindung zu bringen. Dazu wurden aus Agar-
und Flüssigkulturen RNA-Isolierungen vorgenommen, die anschließend in
komplementäre DNA umgeschrieben wurden. Mit der cDNA wurde dann die RT-PCR
durchgeführt.
Die Real-Time-Polymerasekettenreaktion (RT-PCR) leitet sich von der ursprünglichen
PCR ab, bei der DNA-Abschnitte mit Hilfe von Primern amplifiziert werden. Der
Unterschied in der RT-PCR liegt darin, dass bei der Amplifikation der DNA-Abschnitte
ein Fluoreszenzfarbstoff in die DNA interkaliert, der proportional zur PCR
Produktmenge ist(34). Damit lässt sich in Echtzeit das PCR-Produkt ermitteln. Die
Zyklen, die benötigt werden, um das Produkt zu erhalten, gestatten einen Rückschluss
auf die in der Probe vorhandene Menge der DNA.
Zur Normalisierung der RNA-Expression der Regulator-Gene dienen sogenannte
Haushalts-Gene oder auch Referenz-Gene genannt. Bei ihnen handelt es sich um Gene,
die konstitutiv und nicht reguliert exprimiert werden, deren Transkription somit

3 Material und Methoden
28
unabhängig vom Zellstadium, Zelltyp und äußeren Einflüssen ist. Als Referenzgen
wurde gyrB eingesetzt (21).
Tabelle 4: Sequenzen der Referenzgene
Oligonukleotid
Forward
Reverse
aro E 5‘- CTA TCC ACT TGC CAT CTT TTA T - 3‘
5’- ATG GCT TTA ATA TCA CAA TCC C - 3’
gmk 5’- AAG GTG CAA AGC AAG TTA GAA - 3’
5’ - CTT TAC GCG CTT CGT TAA TAC - 3’
gyr B 5’ - TGG TTC AAT GGA TGG AGA TGG - 3’
5’ - CAT ACG AGC CTT ATT CAC TTG G - 3’
Tabelle 5: Sequenzen der in dieser Arbeit untersuchten Gene
Oligonukleotid
Forward
Reverse
Referenz
hla 5‘ - TTT CAC CAG ACT TCG CTA CAG - 3‘
5’ - CCA ATT TGT TGA AGT CCA ATG - 3’
(5)
sae 5‘ - CAA TTT ACG CCT TAA CTT TA - 3‘
5’ - TAG TCA TAT CCC CAA ACT T - 3’
diese Arbeit
rna III 5‘ - TGA ATT TGT TCA CTG TGT CG - 3‘
5‘ - AGG AAG GAG TGA TTT CAA TG - 3‘
(5)
asp23 5‘ - CAA GAA CAA AAT CAA GAG CCT CAA T - 3‘
5‘ - CTT CAC GTG CAG TAC CA - 3‘
(6)
spa 5‘ - GCA ACG TAA CGG CTT CAT TC - 3‘
5’ - CTT TCG GTG CTT GAG ATT CG - 3’
diese Arbeit
Der Reaktionsansatz setzte sich aus folgenden Reagenzien zusammen:
11 µl HPLC-H2O
12,5 µl SYBR Green Supermix
1 µl Primer Mix (enthielt forward und reverse)
0,5 µl cDNA
entsprachen insgesamt 25 µl Gesamtreaktionsansatz
Es wurde ein Mastermix mit H2O, SYBR Green und Primer-Mix vorgelegt und
anschließend die cDNA hinzugefügt.

3 Material und Methoden
29
Folgendes Temperaturprofil wurde für die Amplifikation der cDNA verwendet:
Initiale Denaturierung: 1 min bei 95°C
Annealing: 35x 0:10 min bei 95°C
0:30 min bei 56°C
0:30 min bei 72°C
Elongation: 81x Anstieg der Temperatur von initial 55°C in 0,5°C
Schritten alle 0:10 min bis auf 95°C
Die relative Expression der Transkripte wurde als Anstieg (n-Fold) in Bezug zu den
Werten der Haushalts-Gene ausgedrückt(6).

4 Ergebnisse
30
4 Ergebnisse
4.1 Wachstumsverhalten auf Columbia-Blut- und Schaedler-Agar
Zunächst wurden die S. aureus-Kolonien phänotypisch charakterisiert. Bestimmt
wurden die Pigmentierung, die Größe, das Hämolyseverhalten, der Nachweis von β-
Toxin sowie die Konsistenz der Kolonien. Bei der Pigmentierung wurden weiße, gelbe
und graue Kolonien unterschieden. Die Einteilung der Hämolyse reichte von 0 (keine
Hämolyse) bis 4 (maximale Hämolyse). Die Konsistenz wurde definiert von trocken
über feucht bis hin zu schleimig. Bei der Größe galt es winzige, kleine oder große
Kolonien zu beschreiben.
Wie in Kapitel 3.2.1 beschrieben, wurden die Isolate auf Columbia-Blut- und
Schaedler-Agar überimpft und über Nacht bei 37°C bebrütet, wobei der Schaedler-Agar
unter 5%igem CO2 inkubiert wurde. Ausschlaggebend für die Bebrütung auf Schaedler-
Agar war, dass einige Merkmale der Kolonien, wie die Pigmentierung oder auch die
Hämolyse durch α-Toxin und β-Toxin, deutlich stärker ausgeprägt waren als auf Blut-
Agar. Einen Einfluss auf das veränderte Wachstum der Kolonien auf Schaedler-Agar
wird auch die Bebrütung unter 5%igem CO2 gehabt haben; im Gegensatz zur Bebrütung
des Blut-Agars unter Raumluft.
Typischerweise wird S. aureus zur Analyse der Transkriptionsprofile in Flüssigkulturen
angezüchtet. Über die Untersuchung von S. aureus von Kulturplatten ist bisher wenig
bekannt. Daher wurde in der vorliegenden Arbeit zum einen das Transkriptionsprofil
sämtlicher Stämme zusätzlich mit dem Flüssigmedium BHI ermittelt und zum anderen
zur Korrelation wichtige Laborstämme (SH 1000; RN 6390; 8325-4) sowie deren
Regulator-Mutanten (agr-; sar-; agr-/sar-; sigB-) untersucht.

4 Ergebnisse
31
4.1.1 Häufigkeitsverteilung der Merkmale der klinischen Isolate
Die isolierten S. aureus Stämme aus dem Respirationstrakt der CF-Patienten zeigten auf
Blut-Agar in Hinblick auf die Farbe der Kolonien eine andere Häufigkeitsverteilung als
auf Schaedler-Agar. Zum Zeitpunkt der Berechnung der Häufigkeit befanden sich 740
klinische Isolate in der Studie.
Abb. 4 Häufigkeitsverteilung der Pigmentierung der Kolonien auf Blut- und Schaedleragar
Auf Blut-Agar traten weiße Kolonien mit 52% etwas häufiger auf als gelbe Kolonien
(46%). Auf Schaedler-Agar war das Verhältnis zugunsten der gelben Kolonien deutlich
verschoben. Diese traten in 72% der Fälle auf, im Gegensatz zu den weißen Kolonien
mit einem Anteil von nur 24 %. Graue Kolonien traten auf beiden Nährböden nur zu
einem sehr geringen Prozentsatz auf.
Abb. 5 Häufigkeitsverteilung der Hämolysestufe der Kolonien auf Blut- und Schaedler-Agar
gelbe Kolonien
42% weiße
Kolonien 52%
graue Kolonien
6%
Columbia-Blut-Agar
gelbe Kolonien
72%
weiße Kolonien
24%
graue Kolonien
4%
Schaedler-Agar
keine Hämolyse
31%
Hämolyse 1
25%
Hämolyse 2
25%
Hämolyse 3
14%
Hämolyse 4
5%
Columbia-Blut-Agar keine
Hämolyse 5% Hämolyse
1 14%
Hämolyse 2
20%
Hämolyse 3
23%
Hämolyse 4
38%
Schaedler-Agar

4 Ergebnisse
32
Die Verteilung der Hämolyse zeigte auf Blut- und Schaedler-Agar große Unterschiede
(Abb. 5). Auf Blut-Agar hatten die Kolonien ohne Hämolyse den größten Anteil mit
31%. Dagegen waren diese auf Schaedler-Agar nur mit 5% vertreten. Den größten
Anteil hatten auf Schaedler-Agar diejenigen mit der Hämolysestufe 4 mit 38%. Den
zweitgrößten Anteil mit 23% hatten hier die Kolonien mit Hämolysestufe 3. Auf
Blutagar trat Hämolysestufe 1 und 2 mit jeweils 25% auf.
Abb. 6 Häufigkeitsverteilung der β-Toxin-Expression der klinischen Isolate
Auf beiden Medien, Blut- und Schaedler-Agar, lag der Anteil der Kolonien, die kein β-
Toxin produzierten bei knapp über 90%.
kein β-Toxin 92%
β-Toxin exprimiert
8%
Columbia-Blut-Agar
kein β-Toxin 91%
β-Toxin exprimiert
9%
Schaedler-Agar
4 Ergebnisse
33
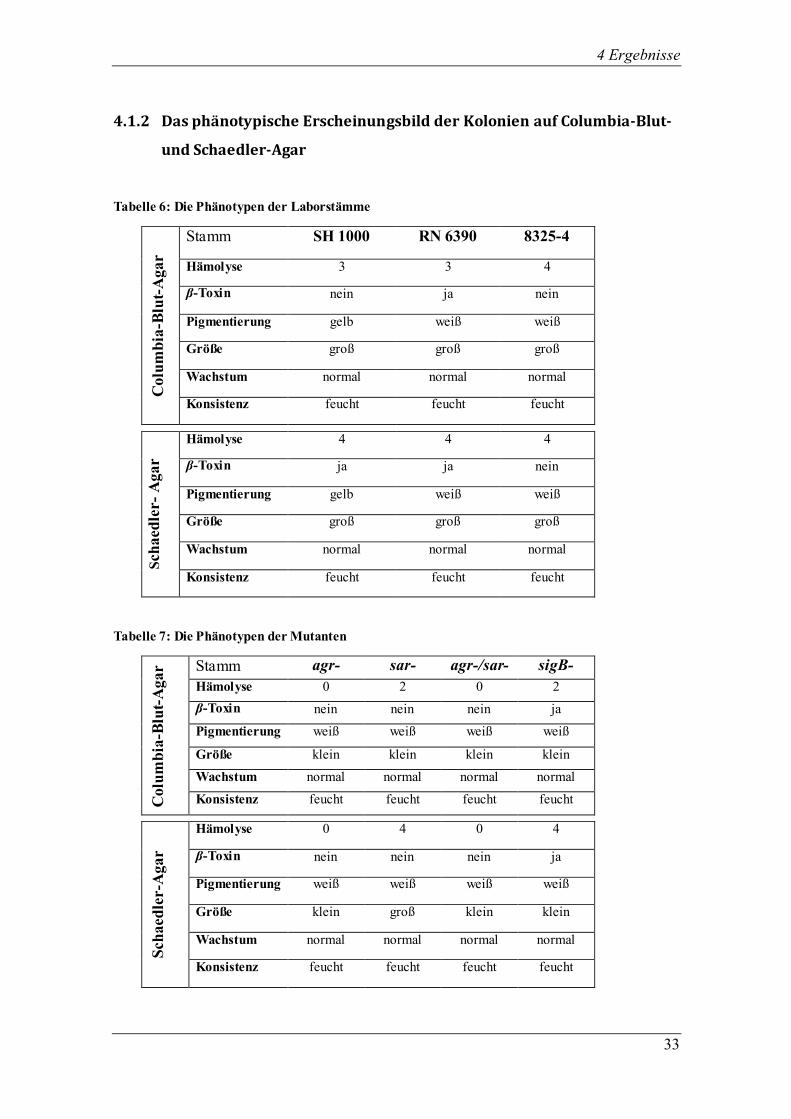
4.1.2 Das phänotypische Erscheinungsbild der Kolonien auf Columbia-Blut-
und Schaedler-Agar
Tabelle 6: Die Phänotypen der Laborstämme
Colu
mb
ia-B
lut-
Agar
Stamm SH 1000 RN 6390 8325-4
Hämolyse 3 3 4
β-Toxin nein ja nein
Pigmentierung gelb weiß weiß
Größe groß groß groß
Wachstum normal normal normal
Konsistenz feucht feucht feucht
Sch
aed
ler-
Agar
Hämolyse 4 4 4
β-Toxin ja ja nein
Pigmentierung gelb weiß weiß
Größe groß groß groß
Wachstum normal normal normal
Konsistenz feucht feucht feucht
Tabelle 7: Die Phänotypen der Mutanten
Colu
mb
ia-B
lut-
Agar Stamm agr- sar- agr-/sar- sigB-
Hämolyse 0 2 0 2
β-Toxin nein nein nein ja
Pigmentierung weiß weiß weiß weiß
Größe klein klein klein klein
Wachstum normal normal normal normal
Konsistenz feucht feucht feucht feucht
Sch
aed
ler-A
gar
Hämolyse 0 4 0 4
β-Toxin nein nein nein ja
Pigmentierung weiß weiß weiß weiß
Größe klein groß klein klein
Wachstum normal normal normal normal
Konsistenz feucht feucht feucht feucht
4 Ergebnisse
34
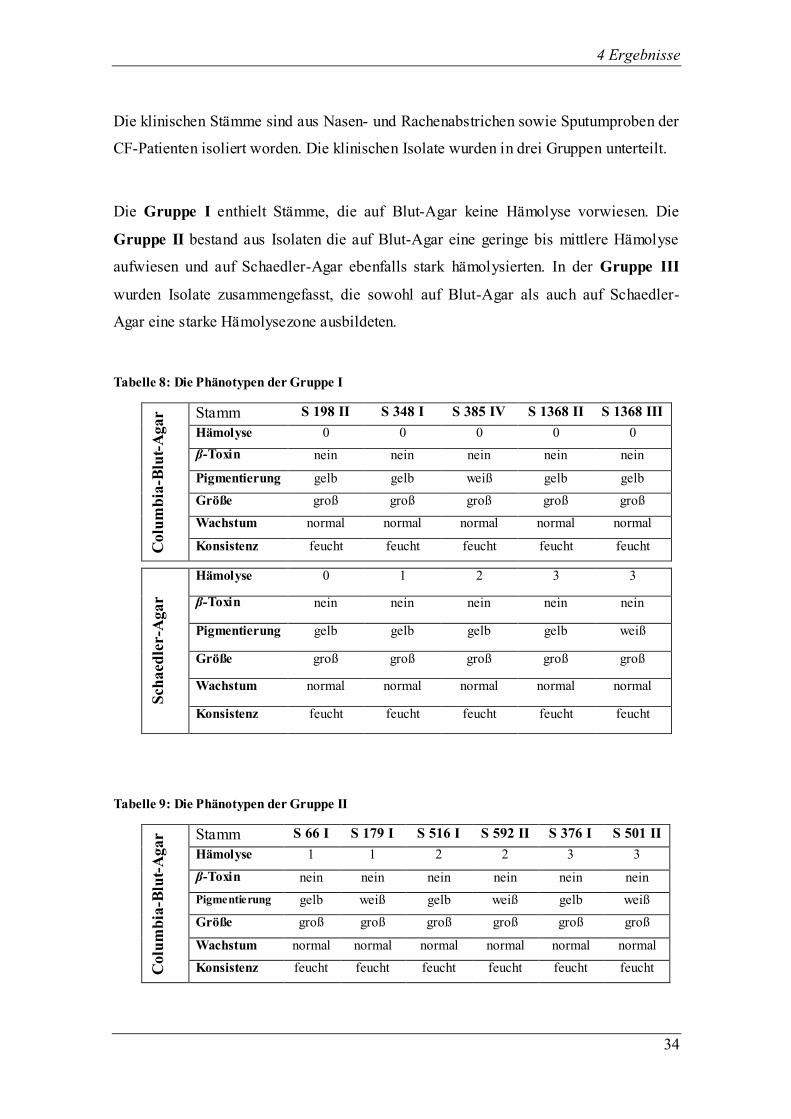
Die klinischen Stämme sind aus Nasen- und Rachenabstrichen sowie Sputumproben der
CF-Patienten isoliert worden. Die klinischen Isolate wurden in drei Gruppen unterteilt.
Die Gruppe I enthielt Stämme, die auf Blut-Agar keine Hämolyse vorwiesen. Die
Gruppe II bestand aus Isolaten die auf Blut-Agar eine geringe bis mittlere Hämolyse
aufwiesen und auf Schaedler-Agar ebenfalls stark hämolysierten. In der Gruppe III
wurden Isolate zusammengefasst, die sowohl auf Blut-Agar als auch auf Schaedler-
Agar eine starke Hämolysezone ausbildeten.
Tabelle 8: Die Phänotypen der Gruppe I
Colu
mb
ia-B
lut-
Agar Stamm S 198 II S 348 I S 385 IV S 1368 II S 1368 III
Hämolyse 0 0 0 0 0
β-Toxin nein nein nein nein nein
Pigmentierung gelb gelb weiß gelb gelb
Größe groß groß groß groß groß
Wachstum normal normal normal normal normal
Konsistenz feucht feucht feucht feucht feucht
Sch
aed
ler-A
gar
Hämolyse 0 1 2 3 3
β-Toxin nein nein nein nein nein
Pigmentierung gelb gelb gelb gelb weiß
Größe groß groß groß groß groß
Wachstum normal normal normal normal normal
Konsistenz feucht feucht feucht feucht feucht
Tabelle 9: Die Phänotypen der Gruppe II
Colu
mb
ia-B
lut-
Agar Stamm S 66 I S 179 I S 516 I S 592 II S 376 I S 501 II
Hämolyse 1 1 2 2 3 3
β-Toxin nein nein nein nein nein nein
Pigmentierung gelb weiß gelb weiß gelb weiß
Größe groß groß groß groß groß groß
Wachstum normal normal normal normal normal normal
Konsistenz feucht feucht feucht feucht feucht feucht
4 Ergebnisse
35
Sch
aed
ler-A
gar
Hämolyse 2 4 3 4 4 4
β-Toxin ja nein nein nein nein ja
Pigmentierung gelb gelb gelb gelb gelb gelb
Größe groß groß groß groß groß groß
Wachstum normal normal normal normal normal normal
Konsistenz feucht feucht feucht feucht feucht feucht
Tabelle 10: Die Phänotypen der Gruppe III
Colu
mb
ia-B
lut-
Agar Stamm S 1365 I S 1365 II S 605 I S 1344 S 568 I
Hämolyse 4 4 4 4 4
β-Toxin nein nein nein nein nein
Pigmentierung weiß weiß weiß weiß gelb
Größe groß groß groß groß groß
Wachstum normal normal normal normal normal
Konsistenz feucht feucht feucht feucht feucht
Sch
aed
ler-A
gar
Hämolyse 4 4 4 4 4
β-Toxin nein nein ja nein nein
Pigmentierung weiß gelb gelb weiß gelb
Größe groß groß groß groß groß
Wachstum normal normal normal normal normal
Konsistenz feucht feucht feucht feucht feucht
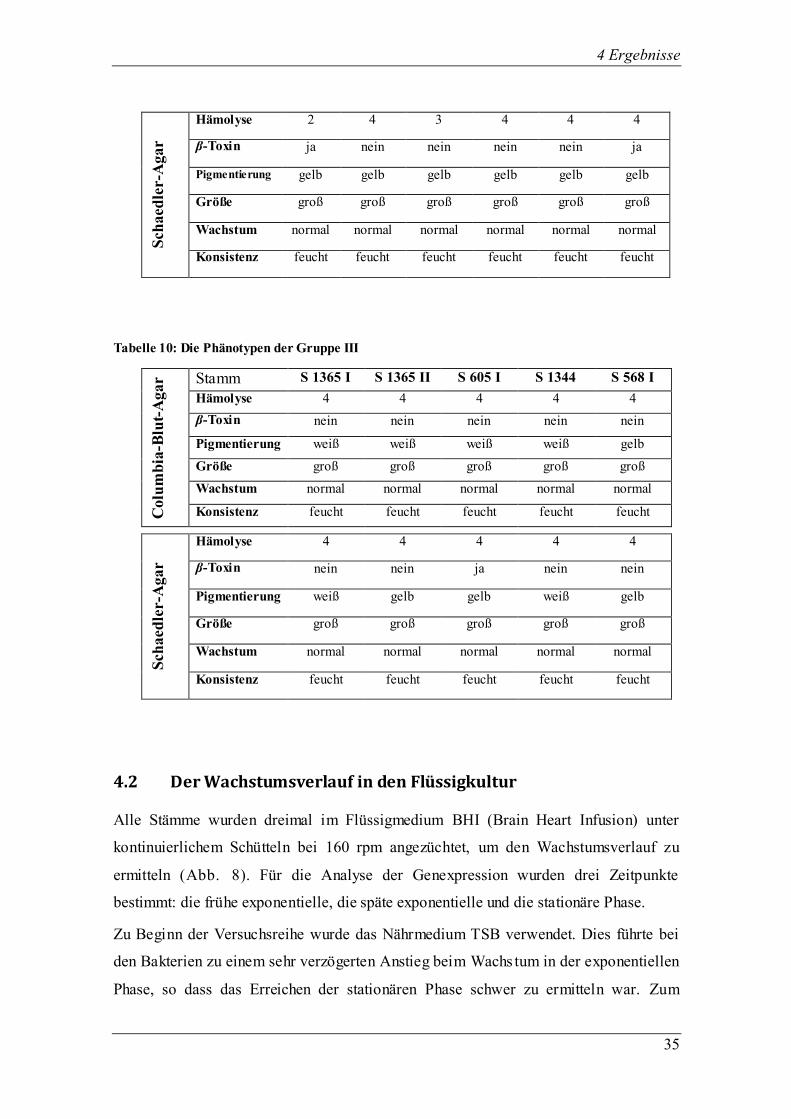
4.2 Der Wachstumsverlauf in den Flüssigkultur
Alle Stämme wurden dreimal im Flüssigmedium BHI (Brain Heart Infusion) unter
kontinuierlichem Schütteln bei 160 rpm angezüchtet, um den Wachstumsverlauf zu
ermitteln (Abb. 8). Für die Analyse der Genexpression wurden drei Zeitpunkte
bestimmt: die frühe exponentielle, die späte exponentielle und die stationäre Phase.
Zu Beginn der Versuchsreihe wurde das Nährmedium TSB verwendet. Dies führte bei
den Bakterien zu einem sehr verzögerten Anstieg beim Wachs tum in der exponentiellen
Phase, so dass das Erreichen der stationären Phase schwer zu ermitteln war. Zum
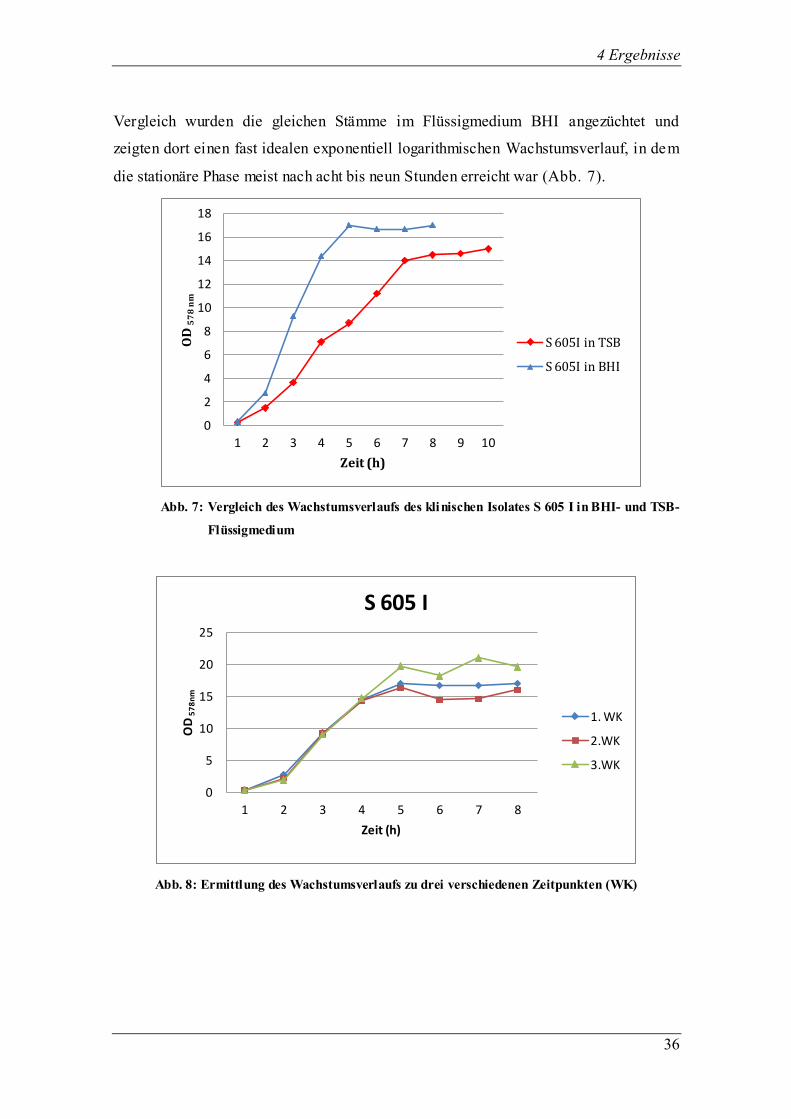
4 Ergebnisse
36
Vergleich wurden die gleichen Stämme im Flüssigmedium BHI angezüchtet und
zeigten dort einen fast idealen exponentiell logarithmischen Wachstumsverlauf, in dem
die stationäre Phase meist nach acht bis neun Stunden erreicht war (Abb. 7).
Abb. 7: Vergleich des Wachstumsverlaufs des klinischen Isolates S 605 I in BHI- und TSB-
Flüssigmedium
Abb. 8: Ermittlung des Wachstumsverlaufs zu drei verschiedenen Zeitpunkten (WK)
0
2
4
6
8
10
12
14
16
18
1 2 3 4 5 6 7 8 9 10
OD
57
8 n
m
Zeit (h)
S 605I in TSB
S 605I in BHI
0
5
10
15
20
25
1 2 3 4 5 6 7 8
OD
57
8n
m
Zeit (h)
S 605 I
1. WK
2.WK
3.WK

4 Ergebnisse
37
4.3 Die Ergebnisse der Real Time-PCR
Die RNA von den Kolonien der Blut- und Schaedler-Agarplatten und aus den
Flüssigkulturen mit folgender cDNA-Transkription wurde in drei unabhängigen Läufen
extrahiert. Die RT-PCR wurde mit jeder Probe doppelt durchgeführt. Zur Analyse der
Ergebnisse konnten daher von jeder Probe sechs Werte herangezogen werden, um den
Mittelwert und die Standardabweichung bestimmen zu können.
Es wurden die Transkription der Regulatoren agr, sae und sigB untersucht, sowie die
der Virulenzgene hla und spa. Für die Bestimmung der agr-Transkription wurde RNA
III und für die von sigB das sigB-abhängige Transkript asp23 untersucht. Als
Referenzgene wurden aroE, gmk und gyrB verwendet.
4.3.1 Das Transkriptionsprofil der Laborstämme und Mutanten
Um das Transkriptionsprofil der klinischen S. aureus-Isolate besser beurteilen und in
Bezug zur Regulation bekannter Laborstämme setzen zu können, wurden die drei
Laborstämme 8325-4, RN 6390 und SH 1000 sowie definierte Regulator-Mutanten
mitgeführt und hinsichtlich ihres Transkriptionsprofils beim Wachstum sowohl auf
Blut- und Schaedler-Agar als auch in der Flüssigkultur mit BHI untersucht.
4 Ergebnisse
38
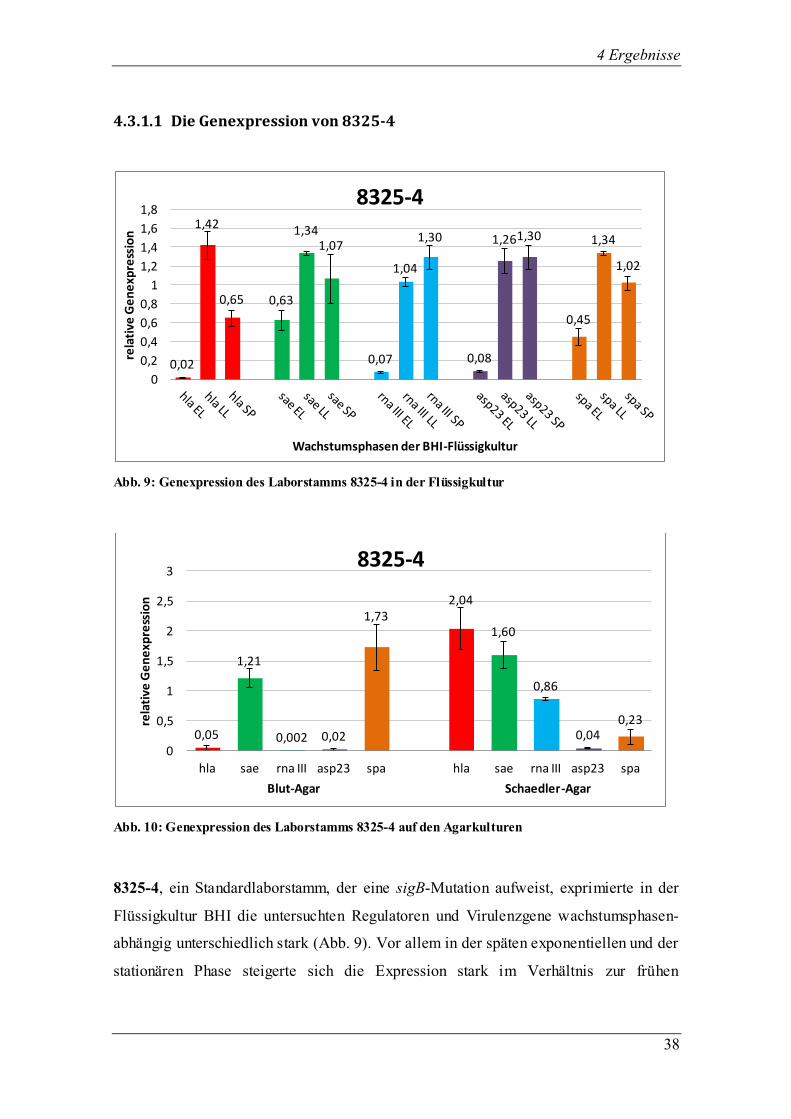
4.3.1.1 Die Genexpression von 8325-4
Abb. 9: Genexpression des Laborstamms 8325-4 in der Flüssigkultur
Abb. 10: Genexpression des Laborstamms 8325-4 auf den Agarkulturen
8325-4, ein Standardlaborstamm, der eine sigB-Mutation aufweist, exprimierte in der
Flüssigkultur BHI die untersuchten Regulatoren und Virulenzgene wachstumsphasen-
abhängig unterschiedlich stark (Abb. 9). Vor allem in der späten exponentiellen und der
stationären Phase steigerte sich die Expression stark im Verhältnis zur frühen
0,02
1,42
0,65 0,63
1,34 1,07
0,07
1,04
1,30
0,08
1,26 1,30
0,45
1,34
1,02
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkultur
8325-4
0,05
1,21
0,002 0,02
1,73 2,04
1,60
0,86
0,04 0,23
0
0,5
1
1,5
2
2,5
3
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
8325-4

4 Ergebnisse
39
exponentiellen Phase. Der Regulator sae wurde zu allen drei Zeitpunkten erfolgreich
exprimiert, wobei die stärkste Transkription wie erwartet in der späten exponentiellen
Phase zu verzeichnen war. Das von sigB kontrollierte asp23 wurde trotz der sigB-
Defizienz insbesondere in der exponentiellen und stationären Phase erfolgreich
transkribiert. RNA III steigerte seine Expression von der frühen exponentiellen bis zur
stationären Phase stetig. Das Virulenzgen hla machte wie erwartet von der frühen zur
späten exponentiellen Phase einen großen Sprung hinsichtlich der relativen Menge der
Genexpression. In dieser Phase wurde das Maximum der Transkription erreicht.
Unerwartet und anders als in der beschriebenen Literatur war der Abfall des hla-
Transkripts in der stationären Phase. Das könnte möglicherweise auf das von uns
benutzte BHI-Medium zurück zu führen sein. Ähnlich sah es bei spa aus, wo auch die
maximale Expression des Gens in der späten exponentiellen Phase stattfand.
Auf Blut- und Schaedler-Agar entwickelte 8325-4 ein Hämolyseverhalten der Stufe 4,
bildete weiß pigmentierte große Kolonien, zeigte ein normales Wachstum und eine
feuchte Konsistenz (Abb. 10). Es wurde kein β-Toxin produziert. Im Vergleich zur
Flüssigkultur war die relative Menge der Genexpression beim Regulator sae sowohl auf
Blut- als auch auf Schaedler-Agar annähernd gleich hoch. Interessant war die fast nicht
vorhandene Transkription von RNA III auf Blut-Agar, wobei es auf Schaedler-Agar
auch wie oben erwähnt in der Flüssigkultur erfolgreich exprimiert wurde. Die
Transkription von asp23 fiel im Gegensatz zur Flüssigkultur auf Blut- und Schaedler-
Agar deutlich geringer aus. Das Virulenzgen hla zeigte eine stark erhöhte Expression
des Gens auf Schaedler-Agar im Vergleich zur Transkription auf Blut-Agar. Wie auch
in der Flüssigkultur wurde spa auf beiden Agarplatten erfolgreich exprimiert. Allerdings
war die Transkription auf Blut-Agar höher als auf Schaedler-Agar.
4 Ergebnisse
40
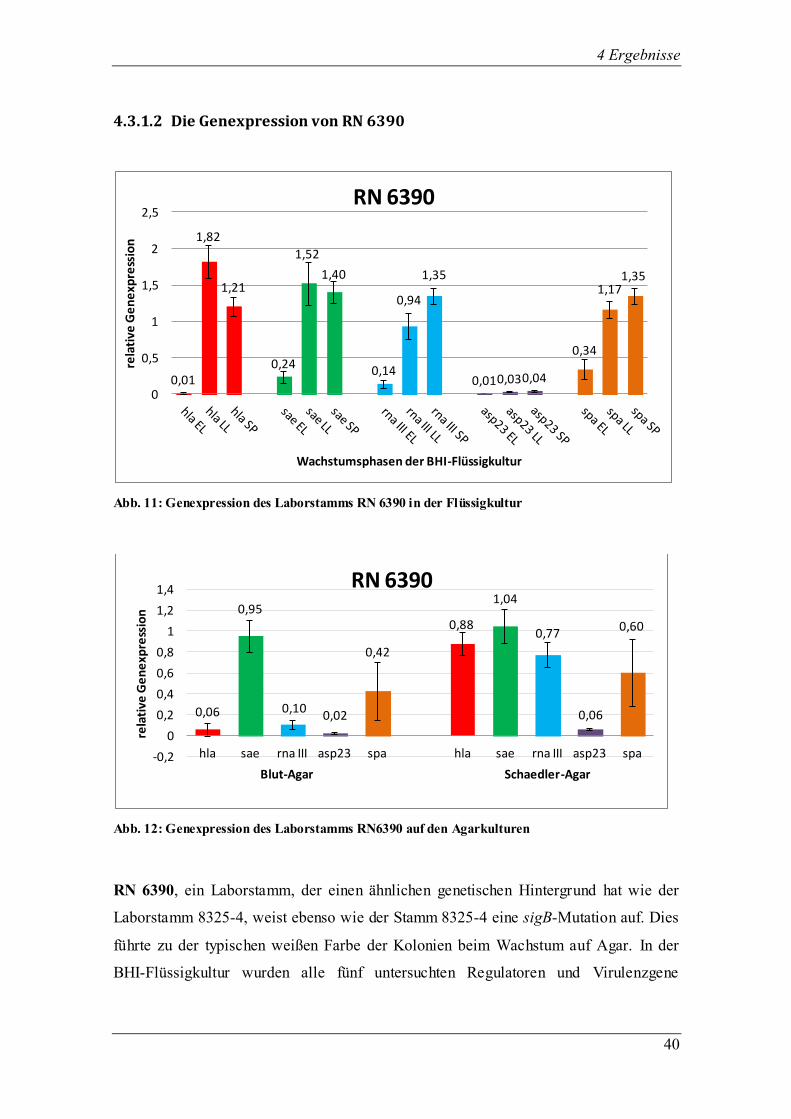
4.3.1.2 Die Genexpression von RN 6390
Abb. 11: Genexpression des Laborstamms RN 6390 in der Flüssigkultur
Abb. 12: Genexpression des Laborstamms RN6390 auf den Agarkulturen
RN 6390, ein Laborstamm, der einen ähnlichen genetischen Hintergrund hat wie der
Laborstamm 8325-4, weist ebenso wie der Stamm 8325-4 eine sigB-Mutation auf. Dies
führte zu der typischen weißen Farbe der Kolonien beim Wachstum auf Agar. In der
BHI-Flüssigkultur wurden alle fünf untersuchten Regulatoren und Virulenzgene
0,01
1,82
1,21
0,24
1,52
1,40
0,14
0,94
1,35
0,01 0,03 0,04
0,34
1,17 1,35
0
0,5
1
1,5
2
2,5
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkultur
RN 6390
0,06
0,95
0,10 0,02
0,42
0,88
1,04
0,77
0,06
0,60
-0,2
0
0,2
0,4
0,6
0,8
1
1,2
1,4
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
RN 6390

4 Ergebnisse
41
erfolgreich exprimiert (Abb. 11). Der Regulator sae transkribierte die größte Menge an
RNA in der späten exponentiellen Phase. Da eine Mutation im sigB-Gen vorliegt,
konnte die Transkription von asp23 in allen drei Phasen kaum nachgewiesen werden.
Das Effektorgen RNA III steigerte seine Expression von der frühen exponentiellen bis
zur stationären Phase, in der es sein Maximum erreichte. Wie auch schon vom
Laborstamm 8325-4 wurde hla in der späten exponentiellen Phase am stärksten
exprimiert. Das Virulenzgen spa wies die höchste Transkription in der stationären Phase
auf.
Auf Blut- und Schaedler-Agar zeigte RN 6390 ein starkes Hämolyseverhalten (Stufe 3),
die Kolonien waren weiß pigmentiert, groß, hatten ein normales Wachstum und eine
feuchte Konsistenz. Auf Blut- und Schädler-Agar wurde das β-Toxin exprimiert. Alle
fünf Gene wurden auf beiden Agarmedien transkribiert (Abb. 12). Der Regulator sae
zeigte sowohl auf Blut- als auch auf Schaedler-Agar eine ungefähr gleich hohe
Genexpression wie zum Zeitpunkt der stationären Phase in der Flüssigkultur.
Interessanterweise wurde asp23 auf beiden Agarmedien genauso gering exprimiert wie
in der Flüssigkultur. RNA III wurde auf Schaedler-Agar stärker exprimiert als auf Blut-
Agar. Ebenso sah es bei den Virulenzgenen hla und spa aus, bei denen die
Transkription auf Schaedler-Agar stärker ausfiel.
4 Ergebnisse
42
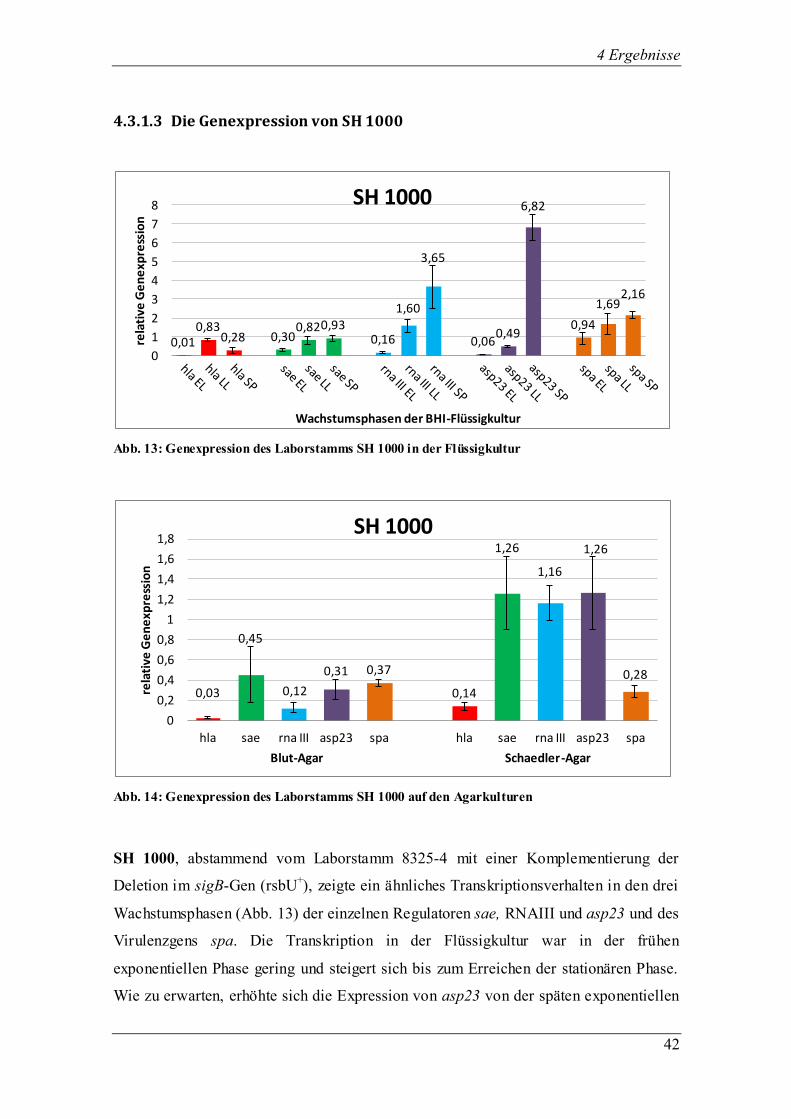
4.3.1.3 Die Genexpression von SH 1000
Abb. 13: Genexpression des Laborstamms SH 1000 in der Flüssigkultur
Abb. 14: Genexpression des Laborstamms SH 1000 auf den Agarkulturen
SH 1000, abstammend vom Laborstamm 8325-4 mit einer Komplementierung der
Deletion im sigB-Gen (rsbU+), zeigte ein ähnliches Transkriptionsverhalten in den drei
Wachstumsphasen (Abb. 13) der einzelnen Regulatoren sae, RNAIII und asp23 und des
Virulenzgens spa. Die Transkription in der Flüssigkultur war in der frühen
exponentiellen Phase gering und steigert sich bis zum Erreichen der stationären Phase.
Wie zu erwarten, erhöhte sich die Expression von asp23 von der späten exponentiellen
0,01 0,83
0,28 0,30 0,82 0,93
0,16
1,60
3,65
0,06 0,49
6,82
0,94
1,69 2,16
0
1
2
3
4
5
6
7
8
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkultur
SH 1000
0,03
0,45
0,12
0,31 0,37
0,14
1,26
1,16
1,26
0,28
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
SH 1000

4 Ergebnisse
43
zur stationären Phase sehr stark, da es sich bei SH1000 um eine komplementierte sigB-
Mutation handelt und folglich war die Transkription von asp23 entsprechend hoch. Der
sae-Regulator zeigte keine großen Schwankungen zwischen den drei
Wachstumsphasen. Zwar war die Expression in der frühen exponentiellen Phase am
geringsten, bis zur stationären Phase kam es jedoch nur zu einem geringen Anstieg in
der Transkriptionsmenge. Bei RNA III, dem Effektorgen von agr, war die Expression in
der frühen exponentiellen Phase am niedrigsten und erreichte ihr Maximum in der
stationären Phase. Die erfolgreiche Expression von RNA III unterstrich auch die
positive Transkription des Virulenzgens hla, da dessen Expression durch RNA III
aktiviert wurde. hla erreichte sein Maximum in der späten exponentiellen Phase und
wurde als einziges Transkript in der stationären Phase in geringerer Menge produziert.
Obwohl spa, das für das Oberflächenprotein Protein A kodiert, normalerweise durch
RNA III supprimiert wird, zeigte sich die maximale Transkription der spa-RNA in der
stationären Phase.
Die Hämolyse von SH 1000 erreichte auf Schaedler-Agar Stufe 4 und war somit stärker
als auf Blut-Agar. Die Pigmentierung war auf beiden Agarplatten goldgelb, die
Kolonien wuchsen groß und hatten eine feuchte Konsistenz. Auf Schaedler-Agar konnte
jedoch im Gegensatz zum Blut-Agar die Hämolyse durch das β-Toxin beobachtet
werden. Abb. 14 zeigt das Transkriptionsprofil von SH 1000 nach Isolierung der RNA
von Blut- und Schaedler-Agar. Auf den ersten Blick fiel auf, dass alle untersuchten
Gene auf Schaedler-Agar stärker exprimiert wurden. Der Genregulator sae wurde auf
Schaedler-Agar stärker exprimiert als auf Blut-Agar und in der Flüssigkultur. RNA III
wies auf Schaedler-Agar eine höhere Expression als auf Blut-Agar auf. Die erfolgreiche
sigB-Transkription spiegelte sich ebenso auf den beiden Agarplatten wie in der
Flüssigkultur wieder, da asp23 auf Blut- und Schaedler-Agar erfolgreich exprimiert
wurde. Das Virulenzgen hla zeigte auf Blut- und Schaedler-Agar eine verminderte
Transkription als in der Flüssigkultur. Das spa-Transkript fand sich auf Blut- und
Schaedler-Agar in annähernd gleicher Menge.
4 Ergebnisse
44
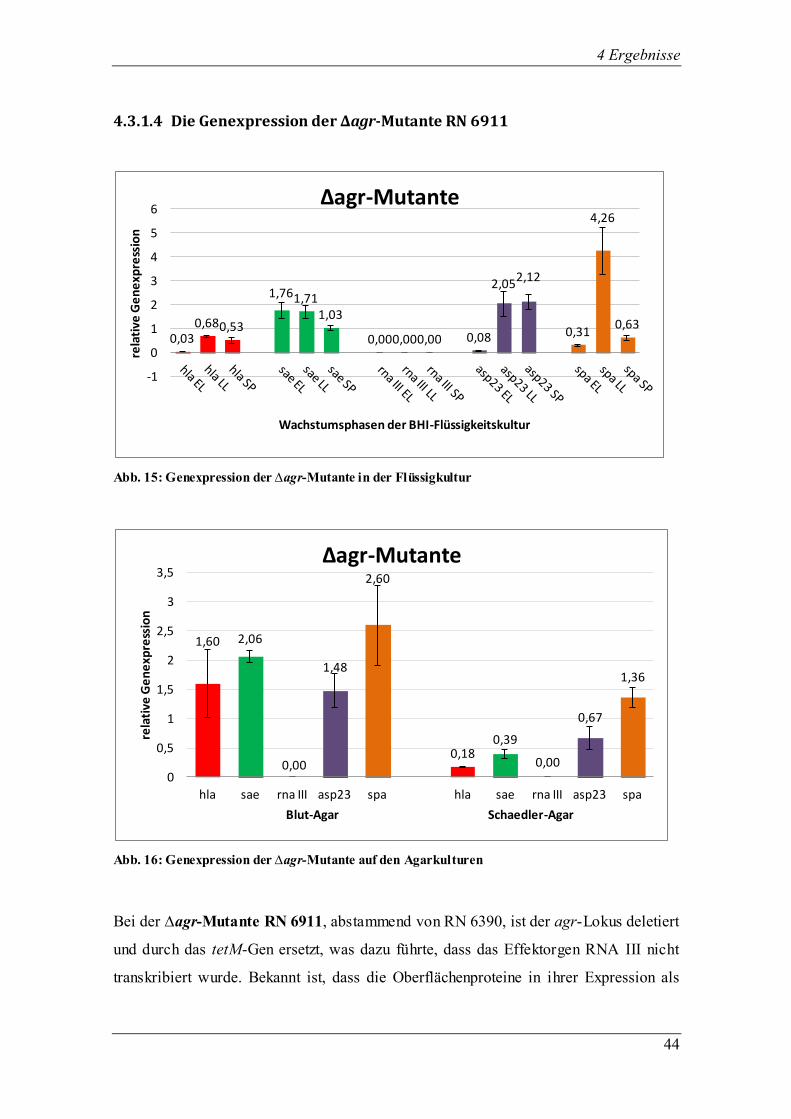
4.3.1.4 Die Genexpression der ∆agr-Mutante RN 6911
Abb. 15: Genexpression der ∆agr-Mutante in der Flüssigkultur
Abb. 16: Genexpression der ∆agr-Mutante auf den Agarkulturen
Bei der ∆agr-Mutante RN 6911, abstammend von RN 6390, ist der agr-Lokus deletiert
und durch das tetM-Gen ersetzt, was dazu führte, dass das Effektorgen RNA III nicht
transkribiert wurde. Bekannt ist, dass die Oberflächenproteine in ihrer Expression als
0,03 0,68 0,53
1,76 1,71 1,03
0,00 0,00 0,00 0,08
2,05 2,12
0,31
4,26
0,63
-1
0
1
2
3
4
5
6
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkeitskultur
∆agr-Mutante
1,60 2,06
0,00
1,48
2,60
0,18 0,39
0,00
0,67
1,36
0
0,5
1
1,5
2
2,5
3
3,5
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
∆agr-Mutante

4 Ergebnisse
45
Folge der agr-Mutation gesteigert und die der Exoproteine reduziert werden.
Desweiteren besteht eine reduzierte Virulenz der Mutante. Alle übrigen vier
untersuchten Gene wurden in allen Wachstumsphasen der Flüssigkultur erfolgreich
exprimiert (Abb. 15). Bei sae wurde erwartet, dass dessen Transkription in einer ∆agr-
Mutante erniedrigt ist. Hier wurde es in der frühen exponentiellen Phase am stärksten
transkribiert mit sinkender Tendenz bis zur stationären Phase. asp23, das die Aktivität
des sigB-Gens widerspiegelt, steigerte seine Transkriptionsmenge kontinuierlich bis
zum Erreichen der stationären Phase. Dafür zeigten hla und spa als agr-abhängige Gene
die stärkste Transkription in der späten exponentiellen Phase und wurden nur gering in
der frühen exponentiellen und stationären Phase exprimiert.
Die ∆agr-Mutante RN 6911 entwickelte sowohl auf Blut- als auch auf Schädler-Agar
keine Hämolyse, kein β-Toxin, hatte kleine weiß pigmentierte Kolonien, ein normales
Wachstum und eine feuchte Konsistenz. Auch beim Wachstum auf Agar wurde RNA III
aufgrund der Mutation im agr-Lokus wie erwartet nicht exprimiert (Abb. 16). Auffällig
war hier, dass die Transkription der vier erfolgreich exprimierten Gene auf Blut-Agar
stärker war als auf Schaedler-Agar. sae erreichte auf Blut-Agar eine ähnlich hohe
Expression wie in der frühen exponentiellen Phase in der Flüssigkultur, wohingegen die
Transkription auf Schaedler-Agar im Vergleich erheblich niedriger war. Das in der
Flüssigkultur ebenfalls erfolgreich exprimierte asp23 zeigte eine erfolgreiche
Expression auf den Agarplatten, wobei die Transkription auf Blut-Agar ungefähr
doppelt so stark war wie auf Schaedler-Agar. Auch die agr-abhängigen Virulenzgene
hla und spa wurden auf Blut-Agar stärker exprimiert als auf Schaedler-Agar.
4 Ergebnisse
46
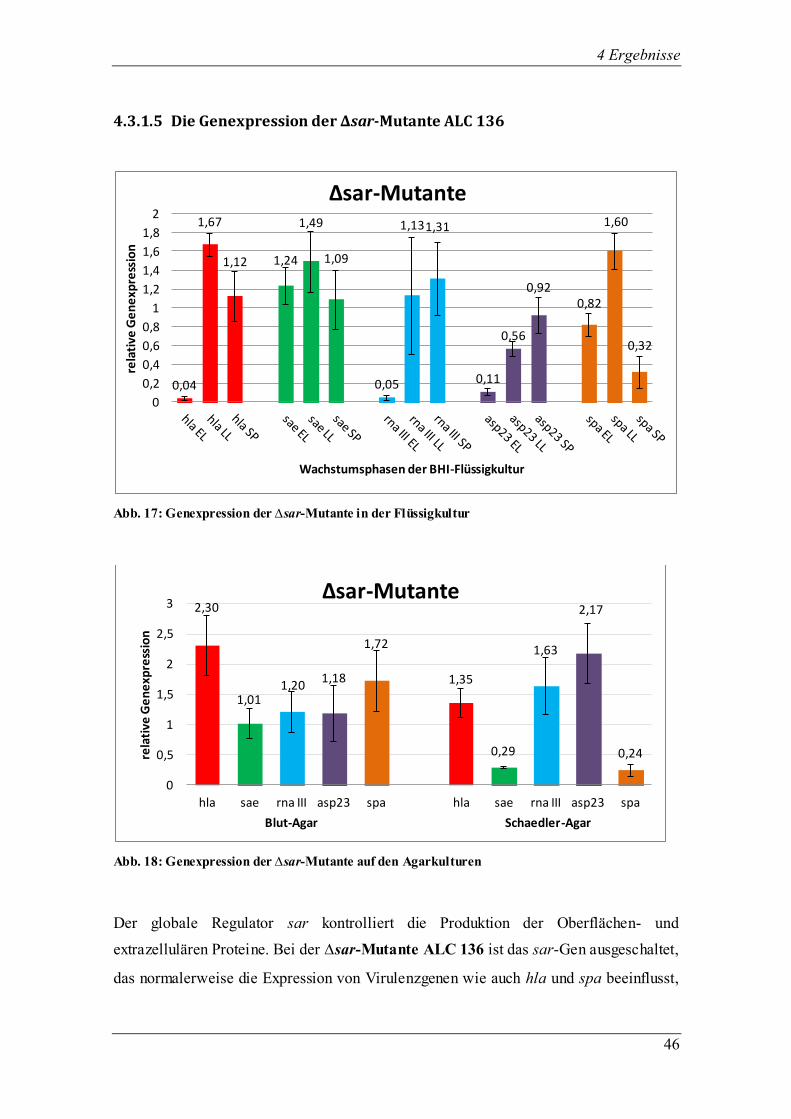
4.3.1.5 Die Genexpression der ∆sar-Mutante ALC 136
Abb. 17: Genexpression der ∆sar-Mutante in der Flüssigkultur
Abb. 18: Genexpression der ∆sar-Mutante auf den Agarkulturen
Der globale Regulator sar kontrolliert die Produktion der Oberflächen- und
extrazellulären Proteine. Bei der ∆sar-Mutante ALC 136 ist das sar-Gen ausgeschaltet,
das normalerweise die Expression von Virulenzgenen wie auch hla und spa beeinflusst,
0,04
1,67
1,12 1,24
1,49
1,09
0,05
1,13 1,31
0,11
0,56
0,92 0,82
1,60
0,32
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkultur
∆sar-Mutante
2,30
1,01 1,20
1,18
1,72
1,35
0,29
1,63
2,17
0,24
0
0,5
1
1,5
2
2,5
3
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
∆sar-Mutante

4 Ergebnisse
47
allerdings nur indirekt. Es handelt sich um eine isogenetische Mutante von RN 6390 mit
einer sarA::ermC Mutation. Daher wurden im folgenden Transkriptionsprofil alle fünf
untersuchten Gene in den einzelnen Wachstumsphasen erfolgreich exprimiert (Abb. 17).
asp23 steigerte die Menge der relativen Genexpression stetig bis zum Erreichen der
stationären Phase, in der es sein Maximum erreichte. Dagegen wurde der Regulator sae
in der späten exponentiellen Phase am stärksten transkribiert, ebenso auch spa und hla.
Der Regulator agr, dessen Transkriptionsmenge durch die Expression des Effektorgens
RNA III gemessen wurde, erreichte das Maximum der relativen Genexpression erst mit
dem Eintreten in die stationäre Phase. Der Sprung bezüglich der Transkriptionsmenge
zwischen der frühen exponentiellen und der späten exponentiellen Phase war sehr hoch.
Die ∆sar-Mutante erreichte auf Blut-Agar Hämolysestufe 2 und auf Schaedler-Agar
Hämolysestufe 4. Sie bildete kein β-Toxin, die Kolonien waren klein und weiß
pigmentiert. Das Wachstum war normal und die Kolonien hatten eine feuchte
Konsistenz. Die untersuchten fünf Gene wurden alle wie in der Flüssigkultur erfolgreich
transkribiert (Abb. 18). Bei sae fiel auf, dass der Regulator auf Blut-Agar deutlich
stärker exprimiert wurde als auf Schaedler-Agar. RNA III zeigte wie auch schon in der
Flüssigkultur eine positive Transkription, die auf Schaedler-Agar etwa gleich stark
ausgeprägt war wie auf Blut-Agar. Dagegen war asp23 das einzige Virulenzgen,
welches auf Schaedler-Agar eine höhere Expression als auf Blut-Agar aufwies.
Interessanterweise wurden die beiden agr-abhängigen Virulenzgene hla und spa auf
Blut-Agar stärker transkribiert.
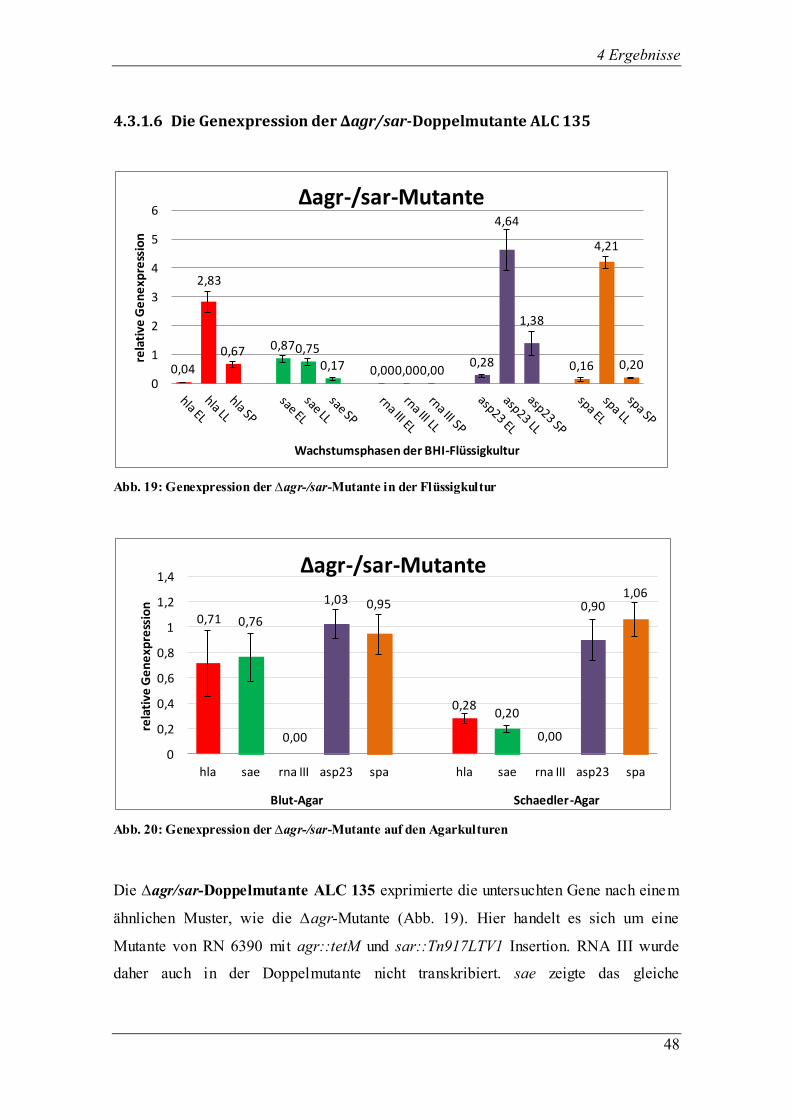
4 Ergebnisse
48
4.3.1.6 Die Genexpression der ∆agr/sar-Doppelmutante ALC 135
Abb. 19: Genexpression der ∆agr-/sar-Mutante in der Flüssigkultur
Abb. 20: Genexpression der ∆agr-/sar-Mutante auf den Agarkulturen
Die ∆agr/sar-Doppelmutante ALC 135 exprimierte die untersuchten Gene nach einem
ähnlichen Muster, wie die ∆agr-Mutante (Abb. 19). Hier handelt es sich um eine
Mutante von RN 6390 mit agr::tetM und sar::Tn917LTV1 Insertion. RNA III wurde
daher auch in der Doppelmutante nicht transkribiert. sae zeigte das gleiche
0,04
2,83
0,67 0,87 0,75 0,17 0,00 0,00 0,00
0,28
4,64
1,38
0,16
4,21
0,20
0
1
2
3
4
5
6
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkultur
∆agr-/sar-Mutante
0,71 0,76
0,00
1,03 0,95
0,28 0,20
0,00
0,90 1,06
0
0,2
0,4
0,6
0,8
1
1,2
1,4
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
∆agr-/sar-Mutante

4 Ergebnisse
49
Transkriptionsmuster wie in der agr-Mutante. Bei asp23 sah das Transkriptionsprofil in
den drei Wachstumsphasen im Gegensatz zur agr-Mutante anders aus. Hier erreichte
asp23 sein Maximum schon in der späten exponentiellen Phase. Da sar die Expression
von anderen Virulenzgenen nur indirekt beeinflusst, wurden diese auch in der
Doppelmutante erfolgreich exprimiert. Die Expression von hla und spa war in der
späten exponentiellen Phase am stärksten.
Auch der Phänotyp der Kolonien auf den Blut- und Schaedler-Agar ähnelte dem der
agr-Mutante sehr. Sie wies ebenfalls auf beiden Agarplatten keine Hämolyse auf, kein
β-Toxin und entwickelte kleine weiße Kolonien mit normalem Wachstum und feuchter
Konsistenz. Ebenso ließ sich ein ähnliches Transkriptionsmuster zur agr-Mutante
feststellen (Abb. 20). RNA III wurde nicht exprimiert, die Expression des Regulators
sae war auf Blut-Agar erheblich stärker als auf Schaedler-Agar und asp23 zeigte hier
ein ebenso erfolgreiches Transkriptionsmuster, was schon in der Flüssigkultur zu
erkennen war. Die Höhe der Expression war auf den beiden Agar-Platten ungefähr
gleich stark. Auch die Transkription von hla ähnelte dem der agr-Mutante, da deren
Expression von hla auf Blut-Agar wesentlich höher ausfiel. Dagegen steigerte spa in der
Doppelmutante auf Schaedler-Agar seine Expression nicht mehr.
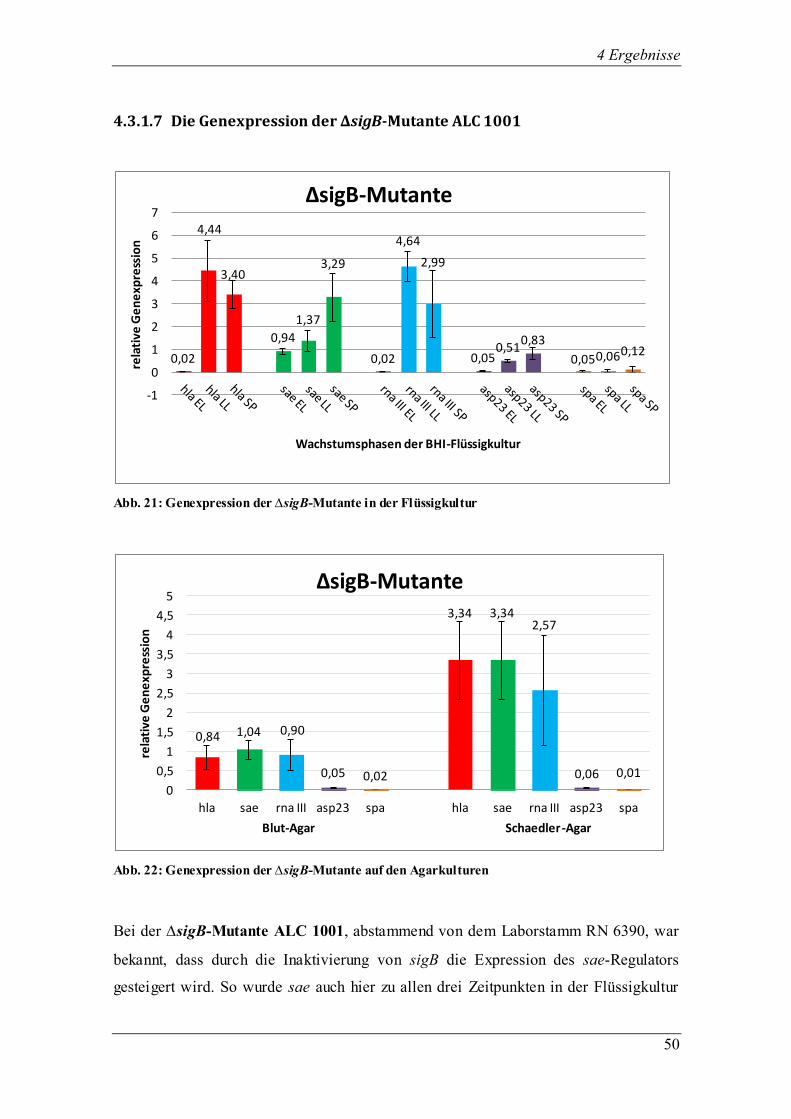
4 Ergebnisse
50
4.3.1.7 Die Genexpression der ∆sigB-Mutante ALC 1001
Abb. 21: Genexpression der ∆sigB-Mutante in der Flüssigkultur
Abb. 22: Genexpression der ∆sigB-Mutante auf den Agarkulturen
Bei der ∆sigB-Mutante ALC 1001, abstammend von dem Laborstamm RN 6390, war
bekannt, dass durch die Inaktivierung von sigB die Expression des sae-Regulators
gesteigert wird. So wurde sae auch hier zu allen drei Zeitpunkten in der Flüssigkultur
0,02
4,44
3,40
0,94
1,37
3,29
0,02
4,64
2,99
0,05 0,51
0,83
0,05 0,06 0,12
-1
0
1
2
3
4
5
6
7
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkultur
∆sigB-Mutante
0,84 1,04 0,90
0,05 0,02
3,34 3,34 2,57
0,06 0,01
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
5
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
∆sigB-Mutante

4 Ergebnisse
51
erfolgreich exprimiert (Abb. 21). Es fand eine kontinuierliche Steigerung von der frühen
exponentiellen bis zur stationären Phase statt. Interessanterweise wurde asp23 trotz der
sigB-Defizienz wenn auch nur gering transkribiert. RNA III hatte das Maximum schon
in der späten exponentiellen Phase erreicht. spa wurde im Gegensatz zu den anderen
Mutanten in der sigB-Mutante nur ganz gering transkribiert und es war ein kaum
merklicher Unterschied zwischen den einzelnen Wachstumsphasen erkennbar. Es
bestand somit ein Unterschied zu RN 6390, bei dem spa erfolgreich in allen drei
Wachstumsphasen mit Maximum in der stationären Phase exprimiert wurde. Wie
erwartet wurde das Virulenzgen hla erfolgreich exprimiert und hatte sein Maximum in
der Transkription während der späten exponentiellen Phase. Es bestätigte sich außerdem
die höhere Expression von hla in der sigB-Mutante im Vergleich zu RN 6390.
Weiterhin hat die sigB-Mutation einen Einfluss auf die Transkripte von spa, die in den
Flüssigkeiten und auf Blut- und Schaedler-Agar zu erkennen ist.
Auf Blut- und Schaedler-Agar ähnelte der Phänotyp der sigB-Mutante sehr dem von
RN 6390. Die Mutante entwickelte auf Blut-Agar eine geringere Hämolyse als auf
Schaedler-Agar, bildete aber auf beiden ein β-Toxin. Die Kolonien waren klein, weiß
pigmentiert und wuchsen normal mit feuchter Konsistenz. Der Virulenzregulator sae
wies auf Schaedler-Agar ein stärkeres Transkriptionsprofil als auf Blut-Agar auf (Abb.
22). Ebenso verhielt es sich mit dem Effektorgen RNA III. asp23 wurde auf beiden
Agarplatten gering exprimiert. spa wurde wie schon in der Flüssigkultur sowohl auf
Blut- als auch auf Schaedler-Agar wenig exprimiert. hla wurde in dieser Mutante auf
beiden Agar-Medien erfolgreich exprimiert, jedoch wesentlich stärker auf Schaedler-
Agar.

4 Ergebnisse
52
4.3.2 Das Transkriptionsprofil der klinischen Isolate
In der vorliegenden Arbeit wurden 16 klinische S. aureus-Isolate aus dem
Respirationstrakt von Mukoviszidose-Patienten ausgewählt, um zu untersuchen, ob
abhängig vom phänotypischen Erscheinungsbild auf Blut- und Schaedler-Agar,
Rückschlüsse auf das Transkriptionsverhalten der Isolate zu ziehen sind.
Die RNA-Isolierung von den Agarplatten erfolgte nach Inkubation der Stämme über 16-
24h bei 37°C. Der Blut-Agar wurde aerob und der Schaedler-Agar unter 5%igem CO2
bebrütet. Durch die Verwendung von Columbia-Blut- und Schaedler-Agar ist eine
bessere Charakterisierung und Vergleich der einzelnen Phänotypen von klinischen
S. aureus-Isolaten untereinander möglich. Zumeist ist das Hämolyseverhalten auf
Schaedler-Agar noch stärker ausgeprägt als auf Blut-Agar. Auch das Pigment der
Kolonien ist häufig unterschiedlich. So haben die Kolonien auf Schaedler-Agar häufig
eine gelblichere Pigmentierung als diejenigen auf Blut-Agar.
Desweiteren wurde ein Teil der klinischen S. aureus-Isolate in dem Flüssigmedium BHI
bebrütet und deren Wachstumsverlauf ermittelt. So konnten die Zeitpunkte des
Erreichens der frühen exponentiellen, der späten exponentiellen und der stationären
Phase ermittelt werden. Anschließend wurde nach erfolgter RNA-Isolierung das
Transkriptionsprofil der Gene mittels RT-PCR bestimmt. Es wurden die S. aureus-
Regulatoren agr, sigB und sae sowie die Virulenzgene hla und spa untersucht.
Bekannt ist, dass die verschiedenen Virulenzfaktoren von S. aureus in bestimmten
Phasen des Wachstumszyklus mehr oder weniger stark exprimiert werden. So werden
die Oberflächenproteine vor allem in der exponentiellen Phase und die Exoproteine und
-toxine eher in der postexponentiellen und stationären Phase transkribiert. Das in dieser
Arbeit untersuchte Virulenzgen spa gehört zur Gruppe der Oberflächenproteine, somit
wurde eine stärkere Expression in der exponentiellen Phase erwartet, während hla zur
Gruppe der sezernierten Proteine gehört, sodass eine stärkere Expression in der
stationären Phase erwartet wurde.
Die klinischen Isolate wurden zu drei Gruppen zusammengefasst, wobei die Einordnung
entsprechend der Ausprägung der Hämolyse erfolgte.

4 Ergebnisse
53
Die Gruppe I der klinischen Isolate, die auf Blut-Agar keine Hämolyse entwickelten,
zeigten in der Flüssigkultur ein sehr unterschiedliches Verhalten in der Transkription
der einzelnen Gene in den verschiedenen Phasen des Wachstumsverlaufes.
Die fünf S. aureus-Isolate S 198 II, S 348 I, S 385 IV, S 1368 II und S 1368 III
entwickelten auf Blut-Agar keine Hämolyse, auf Schaedler-Agar zeigten sie eine
Hämolysezone von Stufe 0 bis 3. Abb. 23 zeigt S 1368 II ohne Hämolyse auf Blut-Agar
und Hämolysestufe 3 auf Schaedler-Agar.
Abb. 23: Isolat S 1368 II mit Hämolyse 0 auf Blut-Agar (li) und mit Hämolyse 3 auf Schaedler-Agar
(re)
Die Pigmentierung der Kolonien war bei drei Isolaten auf Blut- und Schaedler-Agar
gelb, bei einem Isolat auf Blut-Agar weiß und auf Schaedler-Agar gelb und das fünfte
Isolat entwickelte ein gelbes Pigment auf Blut-Agar und ein weißes auf Schaedler-Agar
(Tabelle 8, s.S. 34).
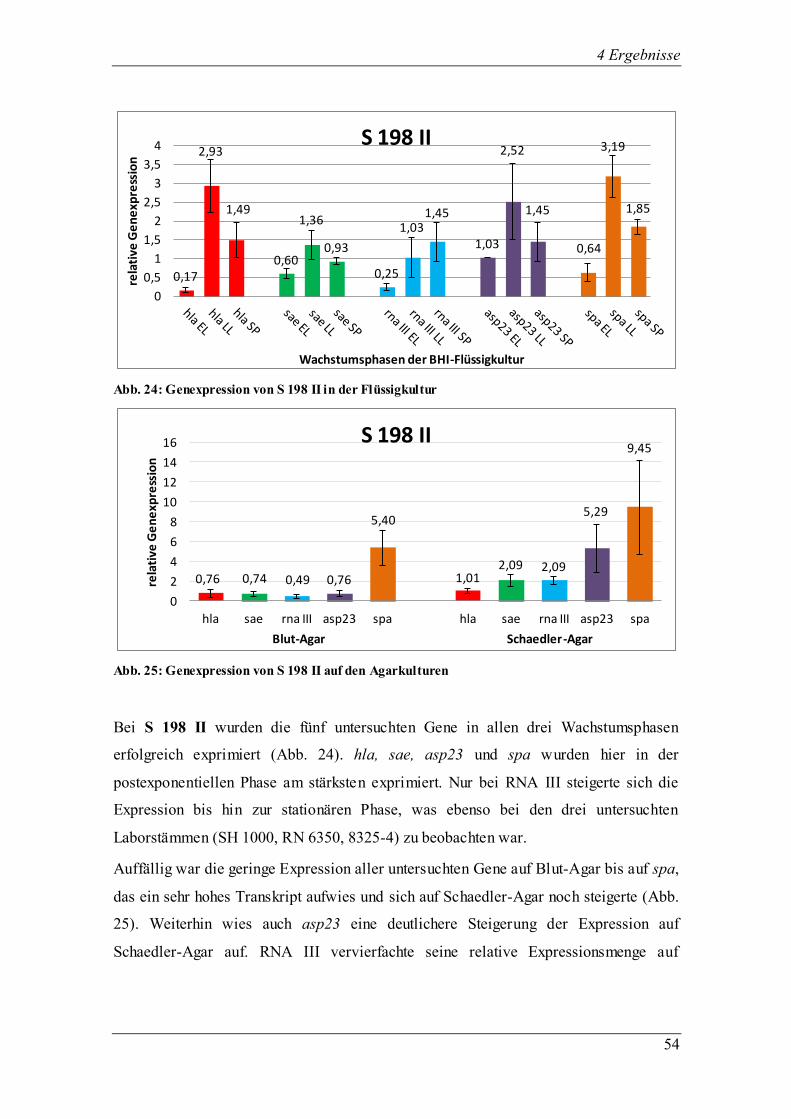
4 Ergebnisse
54
Abb. 24: Genexpression von S 198 II in der Flüssigkultur
Abb. 25: Genexpression von S 198 II auf den Agarkulturen
Bei S 198 II wurden die fünf untersuchten Gene in allen drei Wachstumsphasen
erfolgreich exprimiert (Abb. 24). hla, sae, asp23 und spa wurden hier in der
postexponentiellen Phase am stärksten exprimiert. Nur bei RNA III steigerte sich die
Expression bis hin zur stationären Phase, was ebenso bei den drei untersuchten
Laborstämmen (SH 1000, RN 6350, 8325-4) zu beobachten war.
Auffällig war die geringe Expression aller untersuchten Gene auf Blut-Agar bis auf spa,
das ein sehr hohes Transkript aufwies und sich auf Schaedler-Agar noch steigerte (Abb.
25). Weiterhin wies auch asp23 eine deutlichere Steigerung der Expression auf
Schaedler-Agar auf. RNA III vervierfachte seine relative Expressionsmenge auf
0,17
2,93
1,49
0,60
1,36
0,93
0,25
1,03 1,45
1,03
2,52
1,45
0,64
3,19
1,85
0
0,5
1
1,5
2
2,5
3
3,5
4 re
lati
ve G
en
exp
ress
ion
Wachstumsphasen der BHI-Flüssigkultur
S 198 II
0,76 0,74 0,49 0,76
5,40
1,01 2,09 2,09
5,29
9,45
0
2
4
6
8
10
12
14
16
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
S 198 II
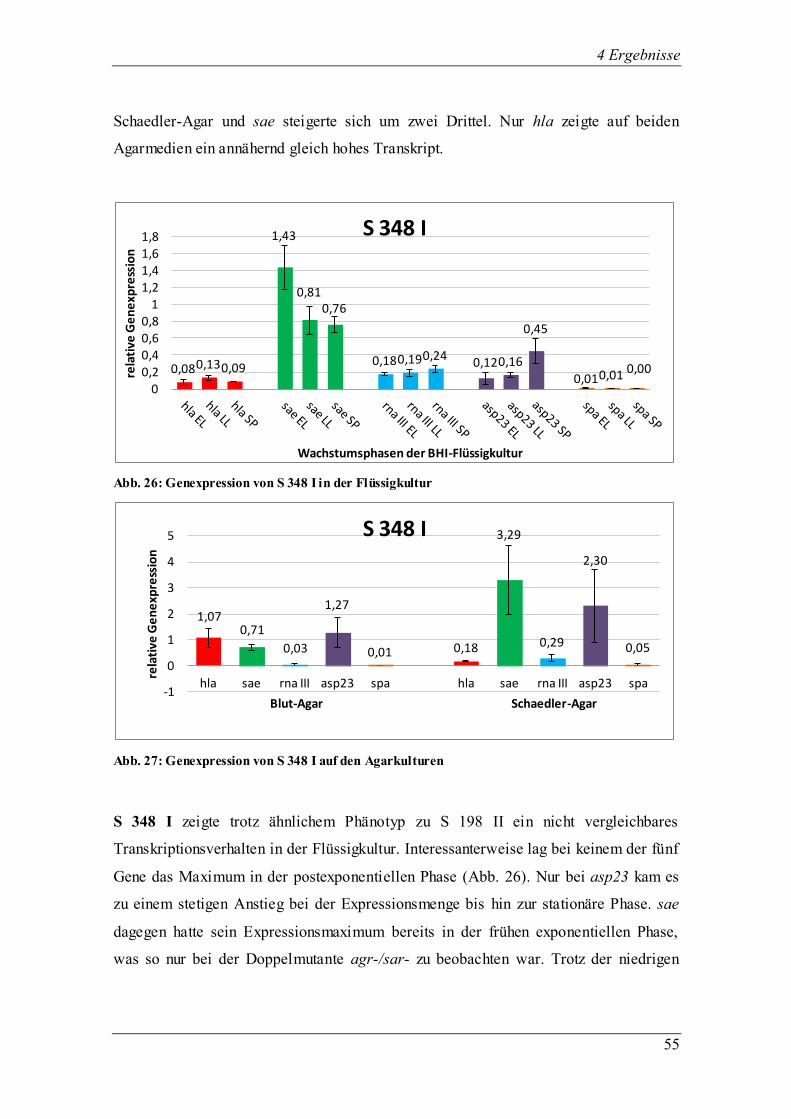
4 Ergebnisse
55
Schaedler-Agar und sae steigerte sich um zwei Drittel. Nur hla zeigte auf beiden
Agarmedien ein annähernd gleich hohes Transkript.
Abb. 26: Genexpression von S 348 I in der Flüssigkultur
Abb. 27: Genexpression von S 348 I auf den Agarkulturen
S 348 I zeigte trotz ähnlichem Phänotyp zu S 198 II ein nicht vergleichbares
Transkriptionsverhalten in der Flüssigkultur. Interessanterweise lag bei keinem der fünf
Gene das Maximum in der postexponentiellen Phase (Abb. 26). Nur bei asp23 kam es
zu einem stetigen Anstieg bei der Expressionsmenge bis hin zur stationäre Phase. sae
dagegen hatte sein Expressionsmaximum bereits in der frühen exponentiellen Phase,
was so nur bei der Doppelmutante agr-/sar- zu beobachten war. Trotz der niedrigen
0,08 0,13 0,09
1,43
0,81 0,76
0,18 0,19 0,24 0,12 0,16
0,45
0,01 0,01 0,00
0 0,2 0,4 0,6 0,8
1 1,2 1,4 1,6 1,8
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkultur
S 348 I
1,07 0,71
0,03
1,27
0,01 0,18
3,29
0,29
2,30
0,05
-1
0
1
2
3
4
5
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
S 348 I
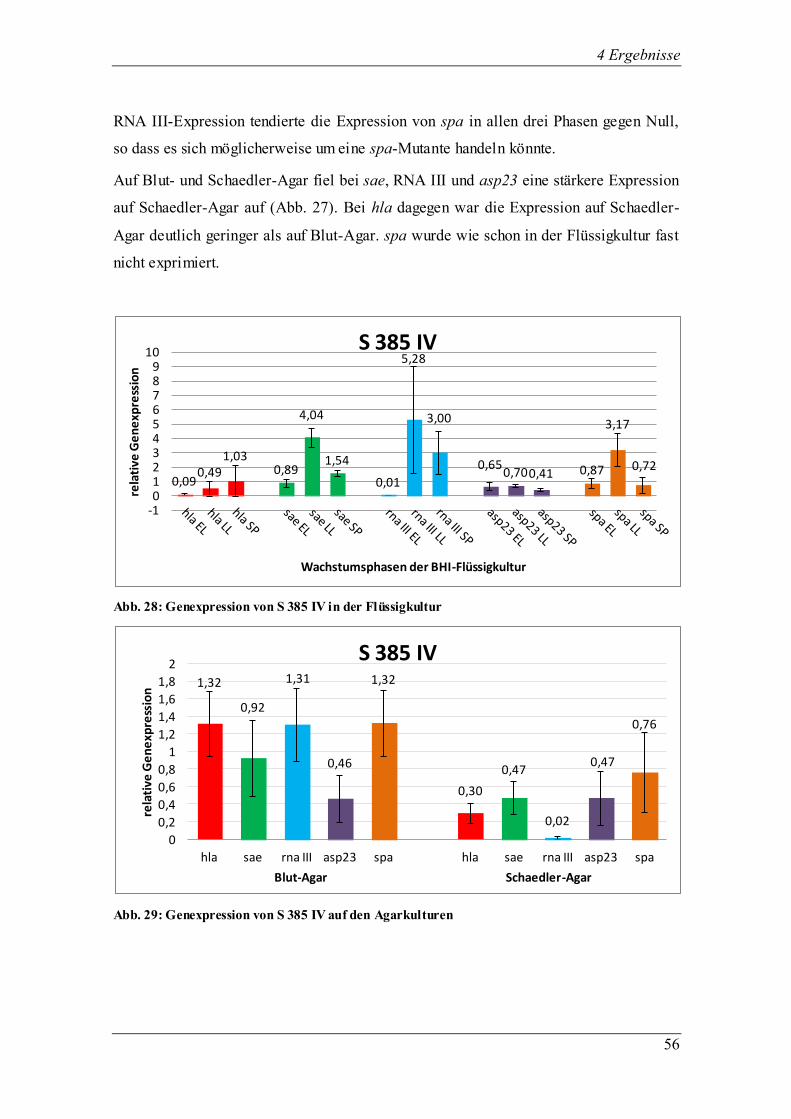
4 Ergebnisse
56
RNA III-Expression tendierte die Expression von spa in allen drei Phasen gegen Null,
so dass es sich möglicherweise um eine spa-Mutante handeln könnte.
Auf Blut- und Schaedler-Agar fiel bei sae, RNA III und asp23 eine stärkere Expression
auf Schaedler-Agar auf (Abb. 27). Bei hla dagegen war die Expression auf Schaedler-
Agar deutlich geringer als auf Blut-Agar. spa wurde wie schon in der Flüssigkultur fast
nicht exprimiert.
Abb. 28: Genexpression von S 385 IV in der Flüssigkultur
Abb. 29: Genexpression von S 385 IV auf den Agarkulturen
0,09 0,49
1,03 0,89
4,04
1,54
0,01
5,28
3,00
0,65 0,70 0,41 0,87
3,17
0,72
-1 0 1 2 3 4 5 6 7 8 9
10
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkultur
S 385 IV
1,32
0,92
1,31
0,46
1,32
0,30
0,47
0,02
0,47
0,76
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
S 385 IV
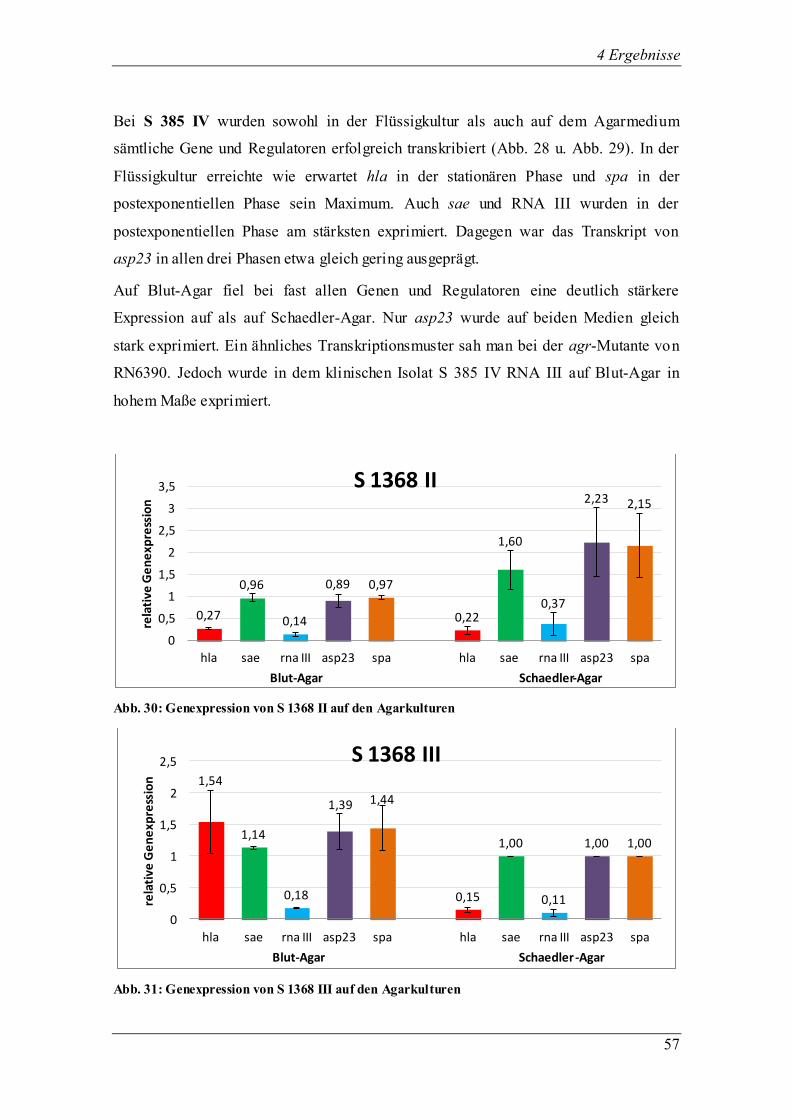
4 Ergebnisse
57
Bei S 385 IV wurden sowohl in der Flüssigkultur als auch auf dem Agarmedium
sämtliche Gene und Regulatoren erfolgreich transkribiert (Abb. 28 u. Abb. 29). In der
Flüssigkultur erreichte wie erwartet hla in der stationären Phase und spa in der
postexponentiellen Phase sein Maximum. Auch sae und RNA III wurden in der
postexponentiellen Phase am stärksten exprimiert. Dagegen war das Transkript von
asp23 in allen drei Phasen etwa gleich gering ausgeprägt.
Auf Blut-Agar fiel bei fast allen Genen und Regulatoren eine deutlich stärkere
Expression auf als auf Schaedler-Agar. Nur asp23 wurde auf beiden Medien gleich
stark exprimiert. Ein ähnliches Transkriptionsmuster sah man bei der agr-Mutante von
RN6390. Jedoch wurde in dem klinischen Isolat S 385 IV RNA III auf Blut-Agar in
hohem Maße exprimiert.
Abb. 30: Genexpression von S 1368 II auf den Agarkulturen
Abb. 31: Genexpression von S 1368 III auf den Agarkulturen
0,27
0,96
0,14
0,89 0,97
0,22
1,60
0,37
2,23 2,15
0
0,5
1
1,5
2
2,5
3
3,5
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
S 1368 II
1,54
1,14
0,18
1,39 1,44
0,15
1,00
0,11
1,00 1,00
0
0,5
1
1,5
2
2,5
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
S 1368 III

4 Ergebnisse
58
Ergänzt wurde diese Gruppe um die Stämme S 1368 II und S 1368 III, die aus
derselben klinischen Probe eines Patienten isoliert wurden. Auffallend war hier, dass
beide Isolate auf Blut-Agar gelbe Kolonien entwickelten, S 1368 III im Gegensatz zu S
1368 II allerdings auf Schaedler-Agar weiße Kolonien produzierte. Es wurden alle Gene
und Regulatoren erfolgreich exprimiert. Es fiel aber auf, dass sich auf Schaedler-Agar
bei S 1368 III eine schwächere Expression aller Gene im Vergleich zur Transkription
auf Blut-Agar zeigte.
Zusammenfassend lässt sich für die Gruppe I sagen, dass sich die Transkriptionsprofile
der untersuchten Regulatoren und Virulenzgene im Vergleich der Stämme miteinander
sehr unterschiedlich darstellten. Der globale Regulator sae wurde bei allen fünf Isolaten
erfolgreich exprimiert. Es bestand ein Unterschied in der Höhe der Expression zwischen
Blut- und Schaedler-Agar, wobei die Transkription auf Schaedler-Agar bei drei Isolaten
(S 198 II, S 348 I, S 1368 II) stärker war. Auch das Effektorgen RNAIII transkribierte
auf Schaedler-Agar eine größere Menge RNA bei diesen drei Isolaten. Der sigB-
abhängigen Virulenzfaktor asp23 wurde bei den Isolaten, die auf beiden Agarplatten
gelbe Kolonien produzierten, auf Schaedler-Agar in höherem Maße exprimiert.
Interessant war, dass die Expression von asp23 bei S 1368 III, das auf Blut-Agar gelbe
und auf Schaedler-Agar weiße Kolonien entwickelte, entsprechend auch auf Blut-Agar
eine stärkere Transkription aufwies. Der Virulenzfaktor hla wurde bei allen 3 Isolaten
auf Blut-Agar im Gegensatz zu Schaedler-Agar deutlich stärker exprimiert. Auch die
Transkription des agr-abhängigen Faktors spa zeigte gewisse Unterschiede auf. So war
die spa-Transkription von S 1368 II wie schon bei asp23 auf Blut-Agar höher. Bei S
198 II und S 1368 II war die Expression auf Schaedler-Agar stärker. Die anderen beiden
Isolate exprimierten spa auf Blut-Agar in stärkerem Maße, wobei die im Vergleich zu
den anderen Isolaten sehr geringe Expression von spa bei S 348 I auffiel.
Es lässt sich bei allen Isolaten kein Zusammenhang zwischen den Transkripten von hla
und dem Grad der Hämolyse auf Blut- und Schaedler-Agar darstellen. Obwohl alle
Isolate auf Blut-Agar ohne Hämolyse wuchsen, konnte bei allen Isolaten eine
Expression für hla auf Blut-Agar aufgezeigt werden.
Beim Isolat S 198 II fiel eine sehr starke Expression von spa, sowohl auf Blut- als auch
auf Schaedler-Agar auf. Ebenfalls wies asp23 eine sehr hohe Transkription auf. Bei S

4 Ergebnisse
59
348 I handelte es sich möglicherweise um eine RNA-Mutation mit kombinierter spa-
Mutation. Bei S 385 IV fiel eine ungewöhnliche Regulation auf, da die Transkripte auf
Blut- höher als auf Schaedler-Agar waren. Weiterhin zeigte sich ein starkes sae-
Transkript in BHI. Bei den Isolaten S 1368 II und III könnte es sich um natürliche agr-
Mutanten handeln, da die Expression von RNA III sowohl auf Blut- als auch auf
Schaedler-Agar sehr gering war.
Zu der Gruppe II gehörten die klinischen Isolate S 66 I, S 179 I, S 516 I, S 592 II, S
376 I und S 501 II, die auf Blut- und Schaedler-Agar eine mäßige bis starke
Hämolysezone entwickelten.
Abb. 32: Isolat S 516 I mit Hämolysestufe 2 auf Blut-Agar (li) und Hämolysestufe 3 auf Schaedler-
Agar (re)
Die Pigmentierung von 3 Isolaten war auf Blut- und Schaedler-Agar gelb (S 66 I, S 516
I, S 376 I) und bei den übrigen drei jeweils auf Blut-Agar weiß und auf Schaedler-Agar
gelb (S 179 I, S 592 II, S 501 II) (Tabelle 9, s.S. 34).
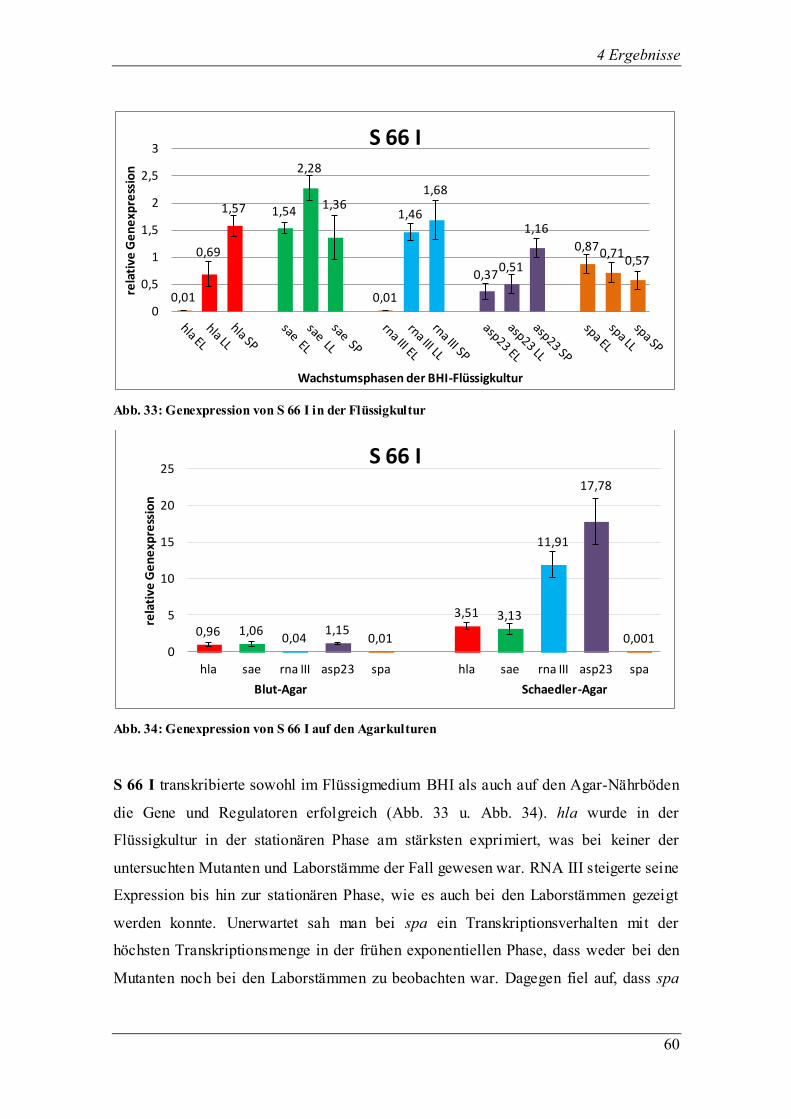
4 Ergebnisse
60
Abb. 33: Genexpression von S 66 I in der Flüssigkultur
Abb. 34: Genexpression von S 66 I auf den Agarkulturen
S 66 I transkribierte sowohl im Flüssigmedium BHI als auch auf den Agar-Nährböden
die Gene und Regulatoren erfolgreich (Abb. 33 u. Abb. 34). hla wurde in der
Flüssigkultur in der stationären Phase am stärksten exprimiert, was bei keiner der
untersuchten Mutanten und Laborstämme der Fall gewesen war. RNA III steigerte seine
Expression bis hin zur stationären Phase, wie es auch bei den Laborstämmen gezeigt
werden konnte. Unerwartet sah man bei spa ein Transkriptionsverhalten mit der
höchsten Transkriptionsmenge in der frühen exponentiellen Phase, dass weder bei den
Mutanten noch bei den Laborstämmen zu beobachten war. Dagegen fiel auf, dass spa
0,01
0,69
1,57 1,54
2,28
1,36
0,01
1,46
1,68
0,37 0,51
1,16
0,87 0,71 0,57
0
0,5
1
1,5
2
2,5
3
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkultur
S 66 I
0,96 1,06 0,04
1,15 0,01
3,51 3,13
11,91
17,78
0,001 0
5
10
15
20
25
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
S 66 I
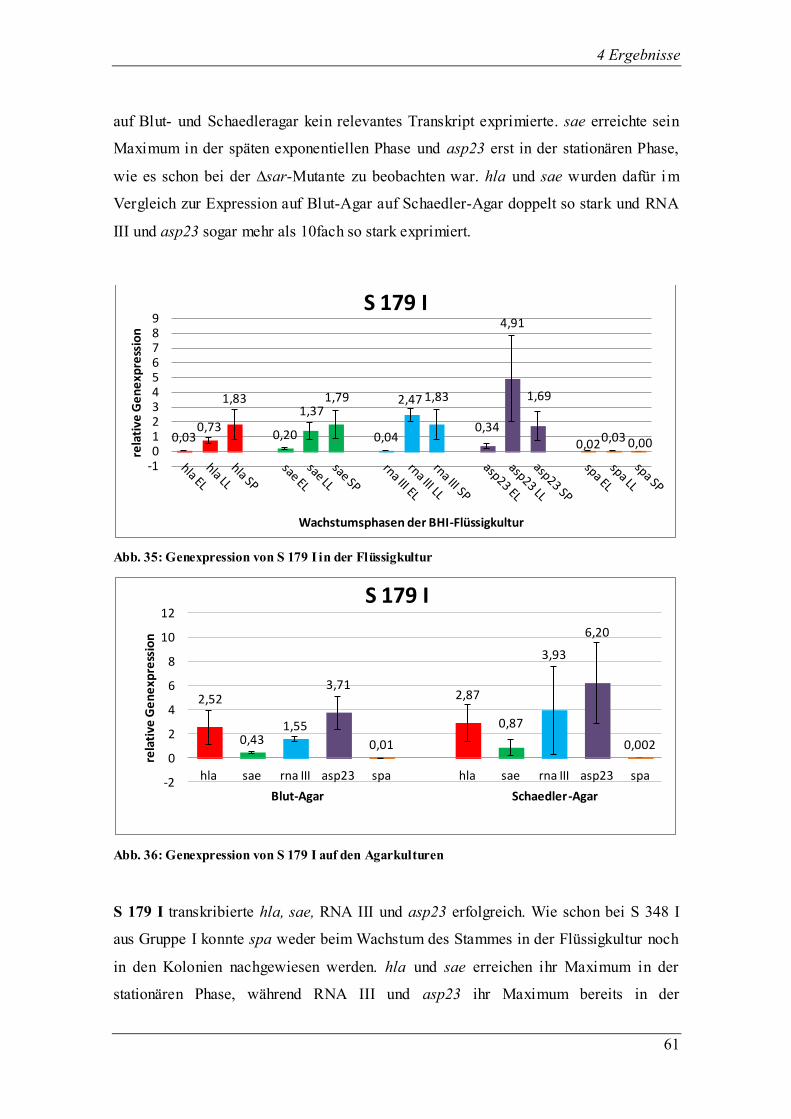
4 Ergebnisse
61
auf Blut- und Schaedleragar kein relevantes Transkript exprimierte. sae erreichte sein
Maximum in der späten exponentiellen Phase und asp23 erst in der stationären Phase,
wie es schon bei der ∆sar-Mutante zu beobachten war. hla und sae wurden dafür im
Vergleich zur Expression auf Blut-Agar auf Schaedler-Agar doppelt so stark und RNA
III und asp23 sogar mehr als 10fach so stark exprimiert.
Abb. 35: Genexpression von S 179 I in der Flüssigkultur
Abb. 36: Genexpression von S 179 I auf den Agarkulturen
S 179 I transkribierte hla, sae, RNA III und asp23 erfolgreich. Wie schon bei S 348 I
aus Gruppe I konnte spa weder beim Wachstum des Stammes in der Flüssigkultur noch
in den Kolonien nachgewiesen werden. hla und sae erreichen ihr Maximum in der
stationären Phase, während RNA III und asp23 ihr Maximum bereits in der
0,03 0,73
1,83
0,20
1,37 1,79
0,04
2,47 1,83
0,34
4,91
1,69
0,02 0,03 0,00
-1 0 1 2 3 4 5 6 7 8 9
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkultur
S 179 I
2,52
0,43 1,55
3,71
0,01
2,87
0,87
3,93
6,20
0,002
-2
0
2
4
6
8
10
12
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
S 179 I
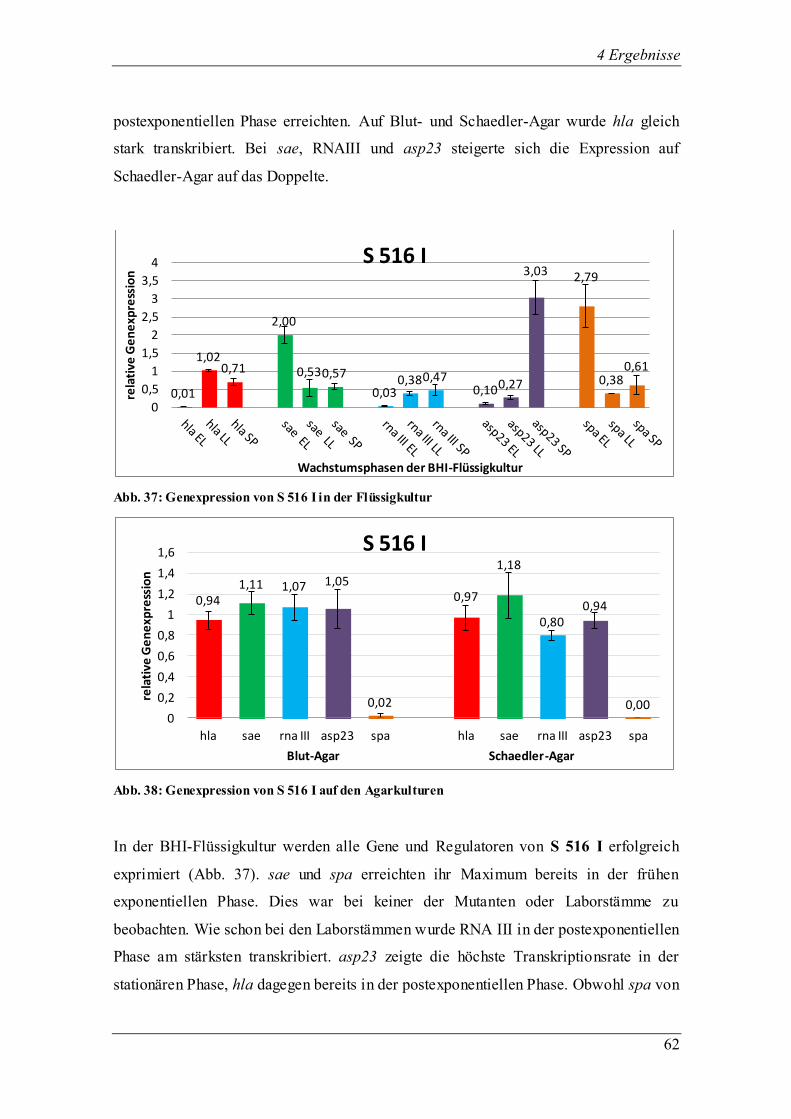
4 Ergebnisse
62
postexponentiellen Phase erreichten. Auf Blut- und Schaedler-Agar wurde hla gleich
stark transkribiert. Bei sae, RNAIII und asp23 steigerte sich die Expression auf
Schaedler-Agar auf das Doppelte.
Abb. 37: Genexpression von S 516 I in der Flüssigkultur
Abb. 38: Genexpression von S 516 I auf den Agarkulturen
In der BHI-Flüssigkultur werden alle Gene und Regulatoren von S 516 I erfolgreich
exprimiert (Abb. 37). sae und spa erreichten ihr Maximum bereits in der frühen
exponentiellen Phase. Dies war bei keiner der Mutanten oder Laborstämme zu
beobachten. Wie schon bei den Laborstämmen wurde RNA III in der postexponentiellen
Phase am stärksten transkribiert. asp23 zeigte die höchste Transkriptionsrate in der
stationären Phase, hla dagegen bereits in der postexponentiellen Phase. Obwohl spa von
0,01
1,02 0,71
2,00
0,53 0,57
0,03 0,38 0,47
0,10 0,27
3,03 2,79
0,38 0,61
0
0,5
1
1,5
2
2,5
3
3,5
4
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkultur
S 516 I
0,94 1,11 1,07 1,05
0,02
0,97
1,18
0,80 0,94
0,00 0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
S 516 I
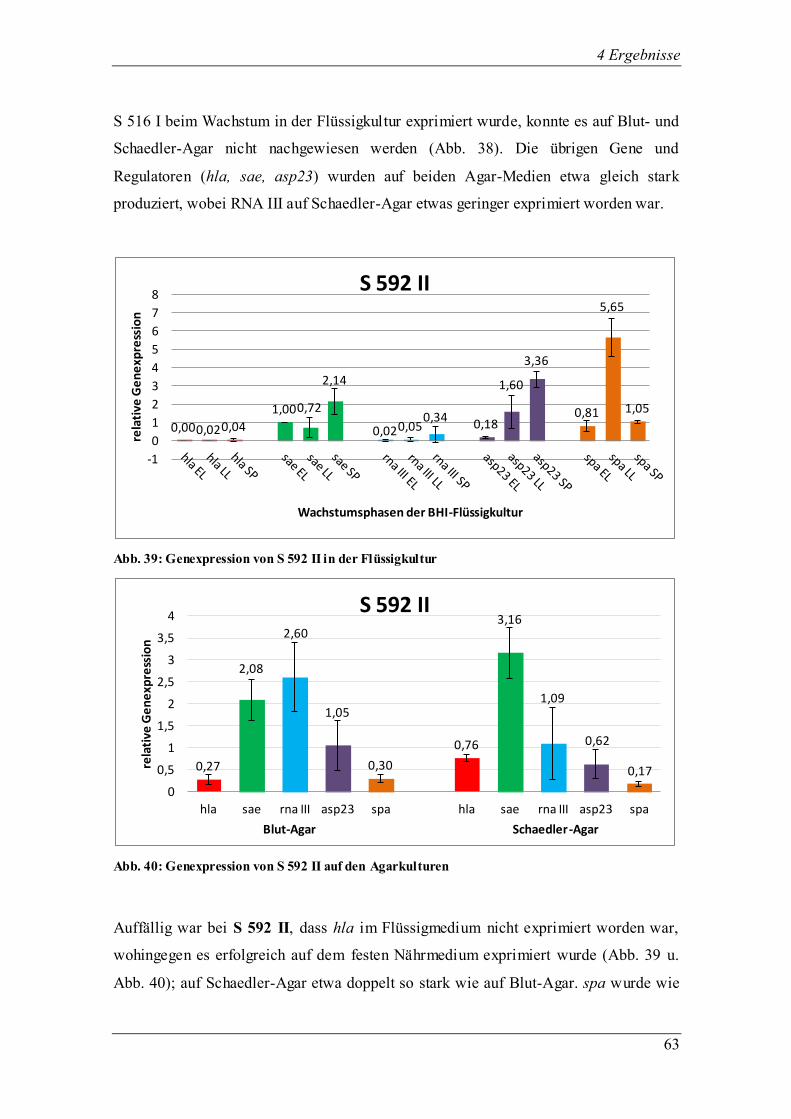
4 Ergebnisse
63
S 516 I beim Wachstum in der Flüssigkultur exprimiert wurde, konnte es auf Blut- und
Schaedler-Agar nicht nachgewiesen werden (Abb. 38). Die übrigen Gene und
Regulatoren (hla, sae, asp23) wurden auf beiden Agar-Medien etwa gleich stark
produziert, wobei RNA III auf Schaedler-Agar etwas geringer exprimiert worden war.
Abb. 39: Genexpression von S 592 II in der Flüssigkultur
Abb. 40: Genexpression von S 592 II auf den Agarkulturen
Auffällig war bei S 592 II, dass hla im Flüssigmedium nicht exprimiert worden war,
wohingegen es erfolgreich auf dem festen Nährmedium exprimiert wurde (Abb. 39 u.
Abb. 40); auf Schaedler-Agar etwa doppelt so stark wie auf Blut-Agar. spa wurde wie
0,00 0,02 0,04
1,00 0,72
2,14
0,02 0,05 0,34
0,18
1,60
3,36
0,81
5,65
1,05
-1
0
1
2
3
4
5
6
7
8
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkultur
S 592 II
0,27
2,08
2,60
1,05
0,30
0,76
3,16
1,09
0,62
0,17
0
0,5
1
1,5
2
2,5
3
3,5
4
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
S 592 II
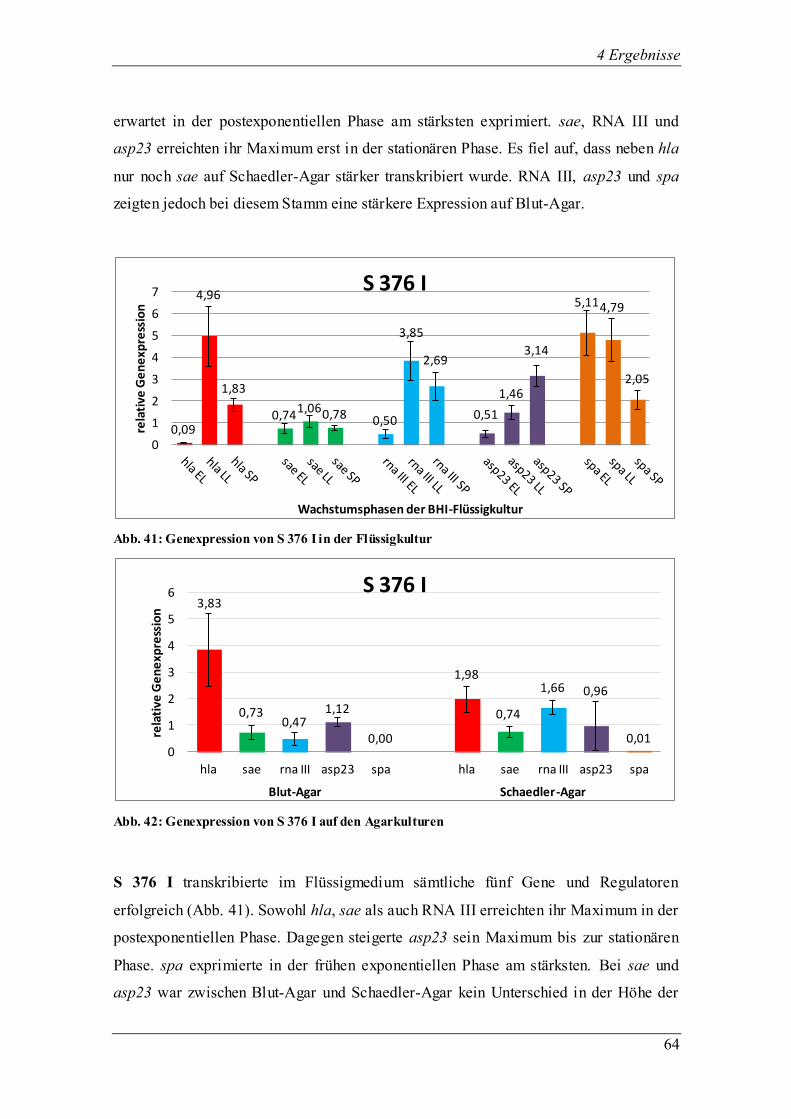
4 Ergebnisse
64
erwartet in der postexponentiellen Phase am stärksten exprimiert. sae, RNA III und
asp23 erreichten ihr Maximum erst in der stationären Phase. Es fiel auf, dass neben hla
nur noch sae auf Schaedler-Agar stärker transkribiert wurde. RNA III, asp23 und spa
zeigten jedoch bei diesem Stamm eine stärkere Expression auf Blut-Agar.
Abb. 41: Genexpression von S 376 I in der Flüssigkultur
Abb. 42: Genexpression von S 376 I auf den Agarkulturen
S 376 I transkribierte im Flüssigmedium sämtliche fünf Gene und Regulatoren
erfolgreich (Abb. 41). Sowohl hla, sae als auch RNA III erreichten ihr Maximum in der
postexponentiellen Phase. Dagegen steigerte asp23 sein Maximum bis zur stationären
Phase. spa exprimierte in der frühen exponentiellen Phase am stärksten. Bei sae und
asp23 war zwischen Blut-Agar und Schaedler-Agar kein Unterschied in der Höhe der
0,09
4,96
1,83
0,74 1,06 0,78 0,50
3,85
2,69
0,51
1,46
3,14
5,11 4,79
2,05
0
1
2
3
4
5
6
7
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkultur
S 376 I
3,83
0,73 0,47
1,12
0,00
1,98
0,74
1,66 0,96
0,01 0
1
2
3
4
5
6
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
S 376 I
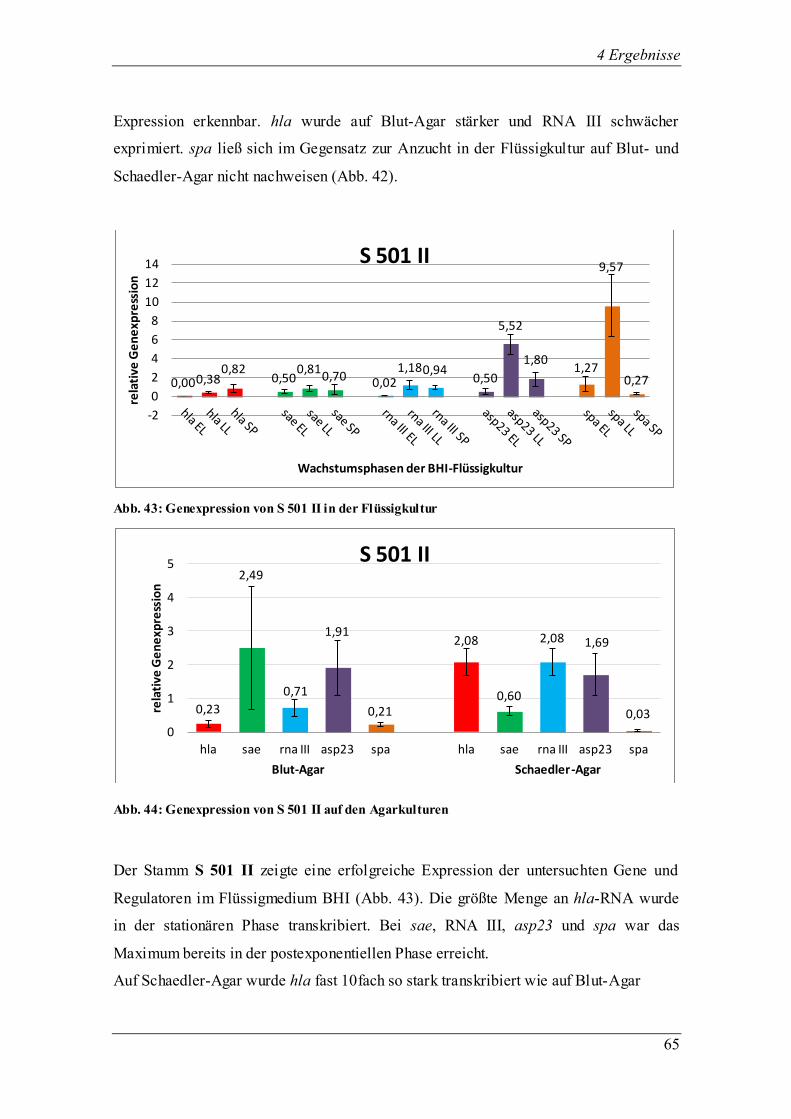
4 Ergebnisse
65
Expression erkennbar. hla wurde auf Blut-Agar stärker und RNA III schwächer
exprimiert. spa ließ sich im Gegensatz zur Anzucht in der Flüssigkultur auf Blut- und
Schaedler-Agar nicht nachweisen (Abb. 42).
Abb. 43: Genexpression von S 501 II in der Flüssigkultur
Abb. 44: Genexpression von S 501 II auf den Agarkulturen
Der Stamm S 501 II zeigte eine erfolgreiche Expression der untersuchten Gene und
Regulatoren im Flüssigmedium BHI (Abb. 43). Die größte Menge an hla-RNA wurde
in der stationären Phase transkribiert. Bei sae, RNA III, asp23 und spa war das
Maximum bereits in der postexponentiellen Phase erreicht.
Auf Schaedler-Agar wurde hla fast 10fach so stark transkribiert wie auf Blut-Agar
0,00 0,38 0,82
0,50 0,81
0,70 0,02 1,18 0,94
0,50
5,52
1,80 1,27
9,57
0,27
-2
0
2
4
6
8
10
12
14
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkultur
S 501 II
0,23
2,49
0,71
1,91
0,21
2,08
0,60
2,08 1,69
0,03
0
1
2
3
4
5
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
S 501 II

4 Ergebnisse
66
Abb. 44). Eine höhere Transkription auf Blut-Agar ließ sich bei sae, asp23 und spa
nachweisen. Dagegen war die Expression von RNA III auf Schaedler-Agar am
stärksten.
In der Gruppe II wurden in der Flüssigkultur von fast allen Stämmen die untersuchten
Regulatoren und Virulenzgene exprimiert. Es fiel auf, dass hla des Stamms S 592 II in
BHI kein relevantes Transkript bildete, jedoch war hla auf Blut- und Schaedler-Agar
nachweisbar. Bei dem Isolat S 179 I scheint es sich möglicherweise um eine spa-
Mutante zu handeln, da in keinem der Medien ein spa-Transkript nachgewiesen wurde.
Auch in den Kolonien, die auf Blut- und Schaedler-Agar wuchsen, konnte bei drei
Stämmen (S 66 I, S 516 I, S 376 I) kein relevantes spa-Transkript nachgewiesen
werden. Allerdings gelang eine erfolgreiche Expression in den Flüssigkulturen dieser
Isolate, so dass bei der Untersuchung der Kolonien möglicherweise die spa-RNA
bereits schon abgebaut worden war. Die übrigen vier Gene bzw. Regulatoren dieser drei
Isolate wurden dafür auf beiden Agarmedien erfolgreich exprimiert. S 66 I zeigte im
Vergleich zu den anderen fünf Isolaten das schwächste Hämolyseverhalten (Blut-Agar
Stufe 1, Schaedler-Agar Stufe 2). Dafür waren hier die Unterschiede in der relativen
Expressionsmenge von Blut- zu Schaedler-Agar am größten (Abb. 34). Bei den
Stämmen, die auf beiden Kulturplatten einen hohen Hämolysegrad erreichten, wie S
376 I und S 501 II, zeigte sich eine Steigerung des Transkripts auf Schaedler-Agar nur
beim Regulator RNA III. hla wurde sogar auf Schaedler-Agar weniger stark exprimiert
und bei sae und asp23 waren keine relevanten Veränderungen in der Expressionsmenge
nachweisbar (Abb. 42).

4 Ergebnisse
67
Der Gruppe III wurden aufgrund ihrer phänotypischen Gemeinsamkeit die Isolate S
605 I, S 568 I, S 1344, S 1365 I und S 1365 II zugeordnet, da sie sowohl auf Blut- als
auch auf Schaedler-Agar die stärkste Hämolyse entwickelten.
Abb. 45: Isolat S 1365 I mit Hämolysestufe 4 auf Blut -(li) und Schaedler-Agar (re)
Zwei Isolate (S 1365 I, S 1344) wiesen sowohl auf Blut- als auch auf Schaedler-Agar
eine weiße Pigmentierung auf. S 568 I war auf beiden Agarplatten gelb pigmentiert und
die restlichen zwei Isolate (S 1365 II, S 605 I) zeigten eher eine weiße Pigmentierung
auf Blut-Agar und ein gelbes Pigment auf Schaedler-Agar (Tabelle 10, s.S. 35).
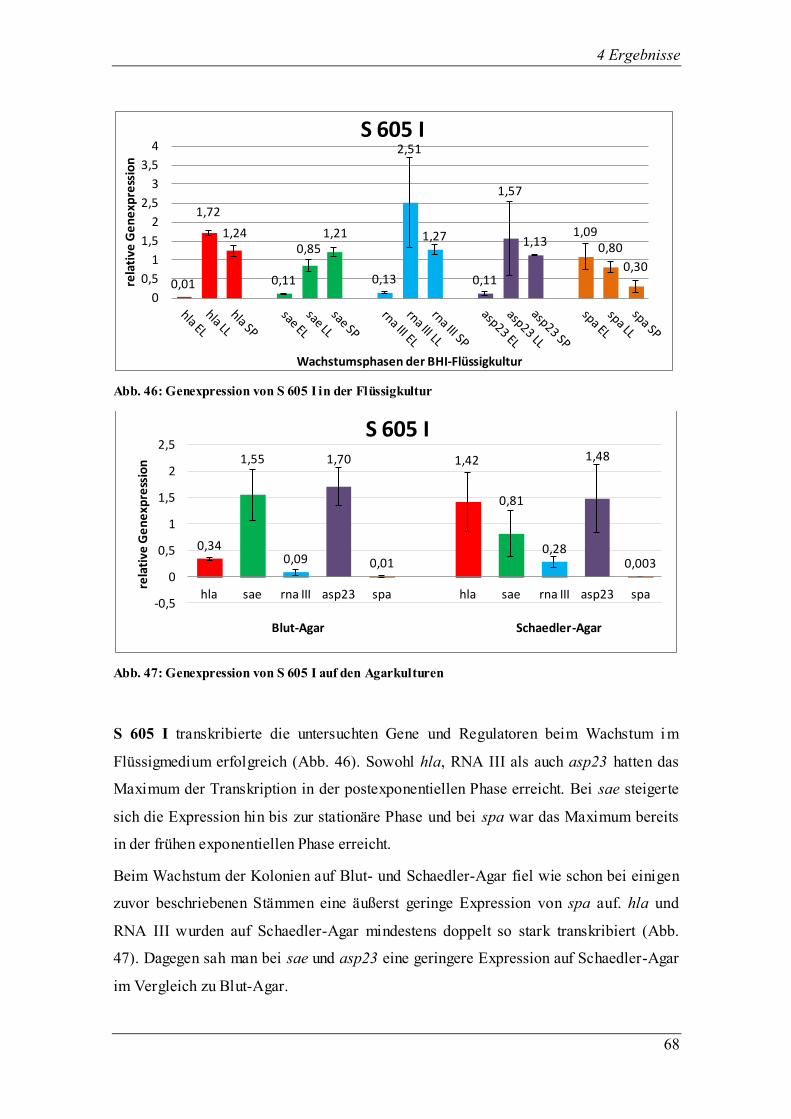
4 Ergebnisse
68
Abb. 46: Genexpression von S 605 I in der Flüssigkultur
Abb. 47: Genexpression von S 605 I auf den Agarkulturen
S 605 I transkribierte die untersuchten Gene und Regulatoren beim Wachstum im
Flüssigmedium erfolgreich (Abb. 46). Sowohl hla, RNA III als auch asp23 hatten das
Maximum der Transkription in der postexponentiellen Phase erreicht. Bei sae steigerte
sich die Expression hin bis zur stationäre Phase und bei spa war das Maximum bereits
in der frühen exponentiellen Phase erreicht.
Beim Wachstum der Kolonien auf Blut- und Schaedler-Agar fiel wie schon bei einigen
zuvor beschriebenen Stämmen eine äußerst geringe Expression von spa auf. hla und
RNA III wurden auf Schaedler-Agar mindestens doppelt so stark transkribiert (Abb.
47). Dagegen sah man bei sae und asp23 eine geringere Expression auf Schaedler-Agar
im Vergleich zu Blut-Agar.
0,01
1,72
1,24
0,11
0,85 1,21
0,13
2,51
1,27
0,11
1,57
1,13 1,09
0,80
0,30
0
0,5
1
1,5
2
2,5
3
3,5
4
rela
tive
Ge
ne
xpre
ssio
n
Wachstumsphasen der BHI-Flüssigkultur
S 605 I
0,34
1,55
0,09
1,70
0,01
1,42
0,81
0,28
1,48
0,003
-0,5
0
0,5
1
1,5
2
2,5
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
S 605 I
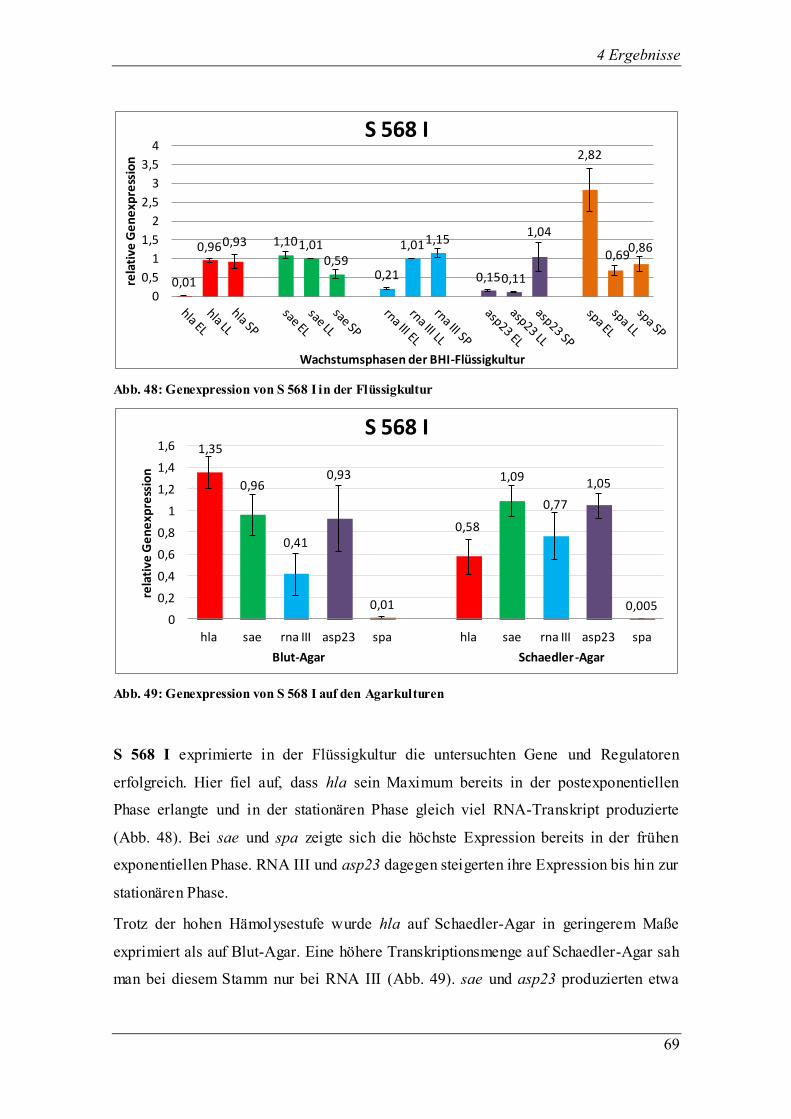
4 Ergebnisse
69
Abb. 48: Genexpression von S 568 I in der Flüssigkultur
Abb. 49: Genexpression von S 568 I auf den Agarkulturen
S 568 I exprimierte in der Flüssigkultur die untersuchten Gene und Regulatoren
erfolgreich. Hier fiel auf, dass hla sein Maximum bereits in der postexponentiellen
Phase erlangte und in der stationären Phase gleich viel RNA-Transkript produzierte
(Abb. 48). Bei sae und spa zeigte sich die höchste Expression bereits in der frühen
exponentiellen Phase. RNA III und asp23 dagegen steigerten ihre Expression bis hin zur
stationären Phase.
Trotz der hohen Hämolysestufe wurde hla auf Schaedler-Agar in geringerem Maße
exprimiert als auf Blut-Agar. Eine höhere Transkriptionsmenge auf Schaedler-Agar sah
man bei diesem Stamm nur bei RNA III (Abb. 49). sae und asp23 produzierten etwa
0,01
0,96 0,93 1,10 1,01 0,59
0,21
1,01 1,15
0,15 0,11
1,04
2,82
0,69 0,86
0
0,5
1
1,5
2
2,5
3
3,5
4 re
lati
ve G
en
exp
ress
ion
Wachstumsphasen der BHI-Flüssigkultur
S 568 I
1,35
0,96
0,41
0,93
0,01
0,58
1,09
0,77
1,05
0,005 0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
S 568 I
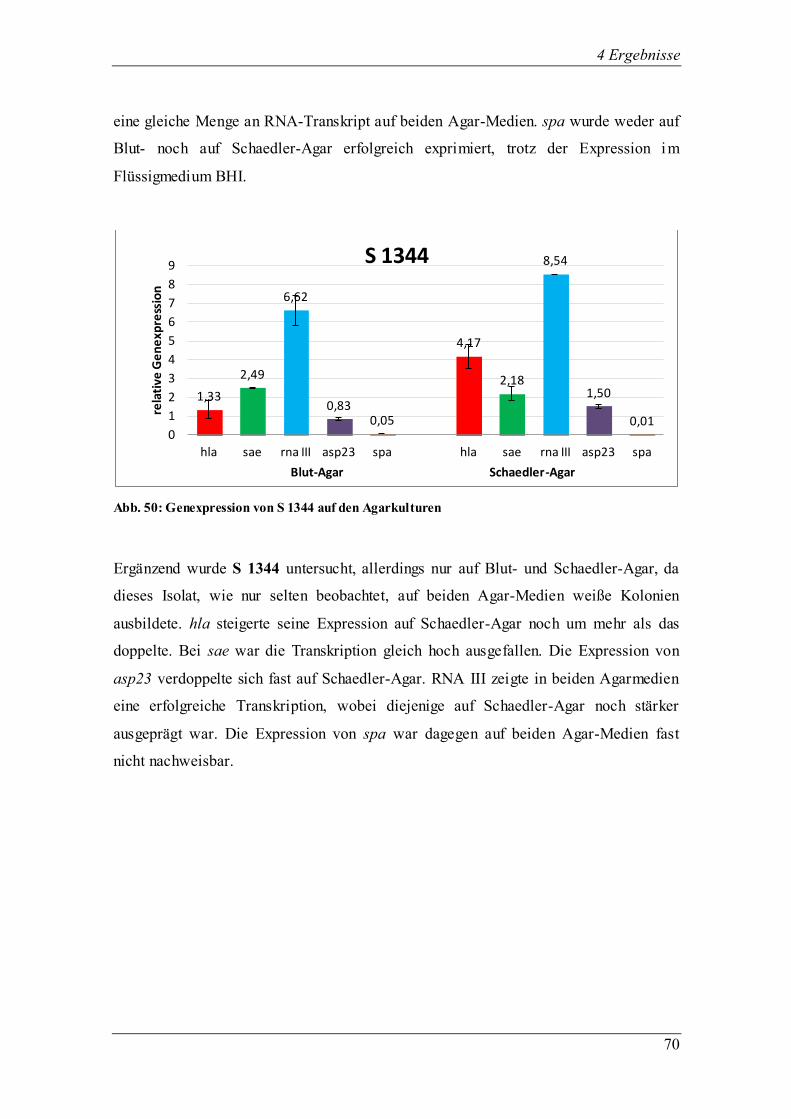
4 Ergebnisse
70
eine gleiche Menge an RNA-Transkript auf beiden Agar-Medien. spa wurde weder auf
Blut- noch auf Schaedler-Agar erfolgreich exprimiert, trotz der Expression im
Flüssigmedium BHI.
Abb. 50: Genexpression von S 1344 auf den Agarkulturen
Ergänzend wurde S 1344 untersucht, allerdings nur auf Blut- und Schaedler-Agar, da
dieses Isolat, wie nur selten beobachtet, auf beiden Agar-Medien weiße Kolonien
ausbildete. hla steigerte seine Expression auf Schaedler-Agar noch um mehr als das
doppelte. Bei sae war die Transkription gleich hoch ausgefallen. Die Expression von
asp23 verdoppelte sich fast auf Schaedler-Agar. RNA III zeigte in beiden Agarmedien
eine erfolgreiche Transkription, wobei diejenige auf Schaedler-Agar noch stärker
ausgeprägt war. Die Expression von spa war dagegen auf beiden Agar-Medien fast
nicht nachweisbar.
1,33
2,49
6,62
0,83 0,05
4,17
2,18
8,54
1,50
0,01 0
1
2
3
4
5
6
7
8
9
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
S 1344
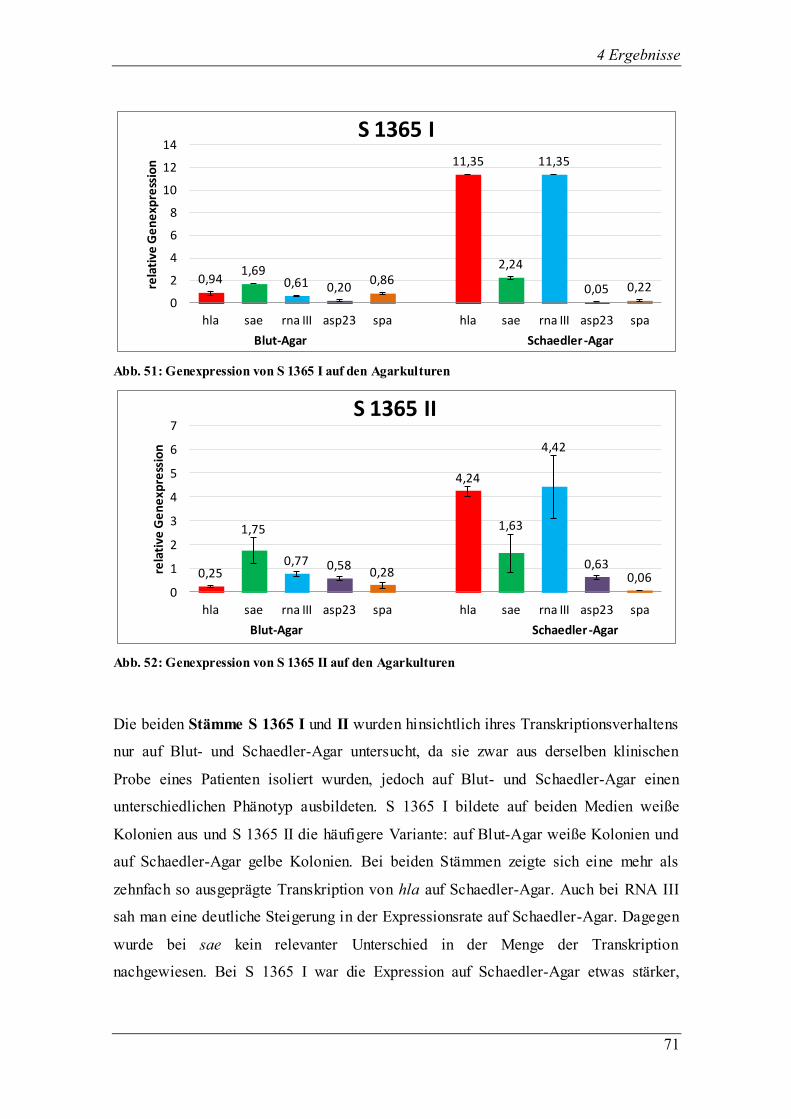
4 Ergebnisse
71
Abb. 51: Genexpression von S 1365 I auf den Agarkulturen
Abb. 52: Genexpression von S 1365 II auf den Agarkulturen
Die beiden Stämme S 1365 I und II wurden hinsichtlich ihres Transkriptionsverhaltens
nur auf Blut- und Schaedler-Agar untersucht, da sie zwar aus derselben klinischen
Probe eines Patienten isoliert wurden, jedoch auf Blut- und Schaedler-Agar einen
unterschiedlichen Phänotyp ausbildeten. S 1365 I bildete auf beiden Medien weiße
Kolonien aus und S 1365 II die häufigere Variante: auf Blut-Agar weiße Kolonien und
auf Schaedler-Agar gelbe Kolonien. Bei beiden Stämmen zeigte sich eine mehr als
zehnfach so ausgeprägte Transkription von hla auf Schaedler-Agar. Auch bei RNA III
sah man eine deutliche Steigerung in der Expressionsrate auf Schaedler-Agar. Dagegen
wurde bei sae kein relevanter Unterschied in der Menge der Transkription
nachgewiesen. Bei S 1365 I war die Expression auf Schaedler-Agar etwas stärker,
0,94 1,69
0,61 0,20 0,86
11,35
2,24
11,35
0,05 0,22 0
2
4
6
8
10
12
14
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler-Agar
S 1365 I
0,25
1,75
0,77 0,58 0,28
4,24
1,63
4,42
0,63 0,06
0
1
2
3
4
5
6
7
hla sae rna III asp23 spa hla sae rna III asp23 spa
rela
tive
Ge
ne
xpre
ssio
n
Blut-Agar Schaedler -Agar
S 1365 II

4 Ergebnisse
72
jedoch sah man bei S 1365 II keinen großen Unterschied. spa zeigte bei beiden
Stämmen eine relevante Reduktion in der Transkriptionsmenge.
Die Gruppe III fasst diejenigen Isolate zusammen, die auf Blut- und Schaedler-Agar
die größte Hämolysezone (Stufe 4) entwickelten. Wie schon in der Gruppe II, fiel bei
zwei Isolaten (S 605 I und S 568 I) auf, dass ein spa-Transkript in den Kolonien auf den
Agarmedien nicht nachgewiesen werden konnte, trotz einer erfolgreichen Expression
von spa in der Flüssigkultur. Der Regulator RNA III wurde in allen fünf Stämmen in
den Kolonien auf Schaedler-Agar stärker exprimiert als auf Blut-Agar. Bei S 1365 I war
die Transkription um mehr als das 10fache gesteigert. asp23 zeigte nur bei S 1344 einen
relevanten Unterschied, in dem es seine Expression auf Schaedler-Agar fast
verdoppelte. Die übrigen Isolate wiesen keinen relevanten Unterschied in ihrer
Transkriptionsmenge von asp23 im Agarmedium auf. S 568 I entwickelte als einziges
Isolat der Gruppe auf beiden Agarmedien gelbe Kolonien. Die Expression von hla war
auch nur bei diesem Isolat auf Blut-Agar stärker als auf Schaedler-Agar. Alle anderen
Isolate wiesen auf Blut-Agar eine geringere Expression von hla auf und produzierten
auf Blut-Agar weiße Kolonien. Außerdem ließ sich die Verbindung zwischen sae und
hla darstellen. Es ist bekannt, dass sae die Expression der α- und β-Hämolysine steigert.
Dies spiegelt sich auch in den Ergebnissen dieser Gruppe wieder. Einer erfolgreichen
Transkription von sae folgte die ebenfalls erfolgreiche Expression von hla im
entsprechenden Medium.

4 Ergebnisse
73
4.3.2.1 Die relative Expression der klinischen Isolate bei dem Wachstum auf
Blut- und Schaedler-Agar im Vergleich
Es wurde die relative Expression der mRNA beim Wachstum der Isolate auf Blut- und
Schädler-Agar untersucht.
Die Expression des Virulenzgens hla, welches vom agr Lokus reguliert wird und für
das α-Toxin kodiert, zeigte bei den 16 klinischen S. aureus-Isolaten z. T. Unterschiede
in der relativen Transkriptionsmenge. 7 Isolate hatten auf Schaedler-Agar eine stärkere
Transkription als auf Blut-Agar, 5 Isolate eine geringere Expression auf Schaedler-
Agar. Bei den restlichen 4 Isolaten konnte keine Änderung in der Expression dargestellt
werden.
Das Virulenzgen spa, dass ebenfalls vom agr-Lokus reguliert wird, kodiert für das
Protein A. 3 Isolate zeigten auf Blut-Agar eine geringere Transkription als auf
Schaedler-Agar und bei 12 Isolaten verhielt es sich umgekehrt. Bei einem Isolat konnte
keine veränderte Transkription festgestellt werden.
Auffällig war, dass RNA III als Effektorgen von agr bei 12 Isolaten auf Schaedler-Agar
stärker exprimiert wurde als auf Blut-Agar. Bei 3 Isolaten verhielt es sich umgekehrt.
Ein Stamm zeigte eine gleich starke Expression auf beiden Agarmedien.
asp23 kodiert für das Alkalische-Schock-Protein und steht unter der Kontrolle des
alternativen Stress-Sigma-Faktors σB. 6 Isolate exprimierten asp23 auf Schaedler-Agar
stärker als auf Blut-Agar. Eine höhere Transkription auf Blut-Agar zeigte sich dagegen
bei 5 Isolaten. In weiteren 5 Isolaten konnte keine vermehrte Expression nachgewiesen
werden.
Der Genlokus sae, bestehend aus den zwei Komponenten saeR und saeS, kodiert für
viele Exoproteine. Mit 7 Isolaten war bei fast der Hälfte der Isolate die Transkription
auf Schaedler-Agar höher als auf Blut-Agar. 4 Isolate wiesen eine höhere Expression
auf Blut-Agar auf. Die übrigen 5 Isolate zeigten keinen relevanten Unterschied in der
Expression im Vergleich von Blut- auf Schaedler-Agar.
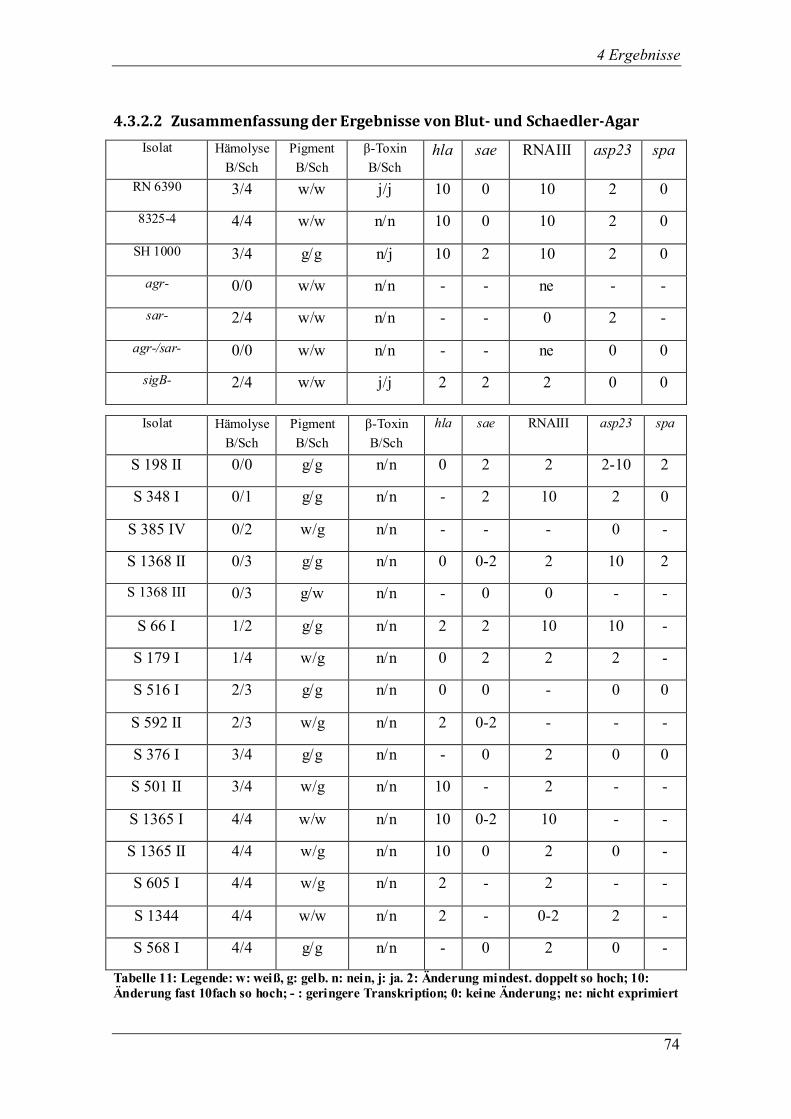
4 Ergebnisse
74
4.3.2.2 Zusammenfassung der Ergebnisse von Blut- und Schaedler-Agar
Isolat Hämolyse
B/Sch
Pigment
B/Sch
β-Toxin
B/Sch
hla sae RNAIII asp23 spa
RN 6390 3/4 w/w j/j 10 0 10 2 0
8325-4 4/4 w/w n/n 10 0 10 2 0
SH 1000 3/4 g/g n/j 10 2 10 2 0
agr- 0/0 w/w n/n - - ne - -
sar- 2/4 w/w n/n - - 0 2 -
agr-/sar- 0/0 w/w n/n - - ne 0 0
sigB- 2/4 w/w j/j 2 2 2 0 0
Isolat Hämolyse
B/Sch
Pigment
B/Sch
β-Toxin
B/Sch
hla sae RNAIII asp23 spa
S 198 II 0/0 g/g n/n 0 2 2 2-10 2
S 348 I 0/1 g/g n/n - 2 10 2 0
S 385 IV 0/2 w/g n/n - - - 0 -
S 1368 II 0/3 g/g n/n 0 0-2 2 10 2
S 1368 III 0/3 g/w n/n - 0 0 - -
S 66 I 1/2 g/g n/n 2 2 10 10 -
S 179 I 1/4 w/g n/n 0 2 2 2 -
S 516 I 2/3 g/g n/n 0 0 - 0 0
S 592 II 2/3 w/g n/n 2 0-2 - - -
S 376 I 3/4 g/g n/n - 0 2 0 0
S 501 II 3/4 w/g n/n 10 - 2 - -
S 1365 I 4/4 w/w n/n 10 0-2 10 - -
S 1365 II 4/4 w/g n/n 10 0 2 0 -
S 605 I 4/4 w/g n/n 2 - 2 - -
S 1344 4/4 w/w n/n 2 - 0-2 2 -
S 568 I 4/4 g/g n/n - 0 2 0 -
Tabelle 11: Legende: w: weiß, g: gelb. n: nein, j: ja. 2: Änderung mindest. doppelt so hoch; 10:
Änderung fast 10fach so hoch; - : geringere Transkription; 0: keine Änderung; ne: nicht exprimiert

4 Ergebnisse
75
Tabelle 11 gibt eine vergleichende Übersicht über die Expression der einzelnen
Virulenzgene und Regulatoren auf Blut- und Schaedler-Agar. „2“ bedeutet, dass die
relative Menge des jeweiligen Transkripts auf Schaedler-Agar mindestens doppelt so
hoch war als auf Blut-Agar. Die „10“ steht für ein fast 10fach so hohes Transkript auf
Schaedler-Agar. Das Minus besagt, dass die Expression auf Schaedler-Agar geringer
war als auf Blut-Agar. Ergänzend dazu sind das Ausmaß der Hämolyse auf Blut- und
Schaedler-Agar, die Ausbildung von β-Toxin sowie die Pigmentierung aller Stämme
mit aufgelistet.
4.3.2.3 Die Veränderung des Hämolyse-Verhaltens verglichen mit der
Veränderung der Transkription von RNAIII auf Blut- und Schaedler-
Agar
Da RNA III hla positiv reguliert und hla für die Hämolyse verantwortlich ist, wird
davon ausgegangen, dass bei einer gesteigerten Expression von RNAIII auch das
Hämolyseverhalten auf Blutagar stärker ausgeprägt sein sollte.
Die Einteilung der Hämolyse fand nach einem vorgefertigten Beschreibungsbogen
(Anhang) statt und wurde mit 0 (keine Hämolyse) bis 4 (maximale Hämolyse)
beschrieben. Dabei kam es vor, dass der Unterschied in der Hämolyse im Vergleich der
Kolonien von Blut- und Schaedler-Agar nur eine oder auch mehrere Stufen betrug.
Möglich war auch eine sehr starke Hämolysezone auf Schaedler-Agar und gar keine
oder nur geringste Hämolyse auf Blut-Agar.
Abb. 53: S 179 I mit Hämolysegrad 1 auf Blut-Agar (li) und Hämolysegrad 4 auf Schaedler-Agar
(re)

4 Ergebnisse
76
Bei 12 Isolaten zeigte sich eine Steigerung der RNA III-Transkription auf Schaedler-
Agar. Dies trat bei 3 von 5 Isolaten ohne Hämolyse auf Blut-Agar, bei 4 von 6 Isolaten
mit geringer bis mäßiger Hämolyse auf Blut-Agar und bei 5 von 5 Isolaten mit starker
Hämolyse auf Blut-Agar auf.
4.3.2.4 Die Unterschiede in der Pigmentierung der Kolonien auf Blut- und
Schaedler-Agar verglichen mit der jeweiligen asp23-Expression
In früheren Arbeiten konnte nachgewiesen werden, dass ein Zusammenhang zwischen
der phänotypischen Pigmentierung und der Expression des Stress-Sigma-Faktors σB
besteht. Ein Produkt des σB-Faktors, das eine Aussage über die Aktivität von σB erlaubt,
ist das asp23. Wenn das σB-Gen in einem S. aureus Stamm nicht transkribiert wird, sind
die Kolonien des Phänotyps weiß. Bei vorhandener Transkription des σB-Gens können
die Kolonien eine gelb-goldene Färbung annehmen. Es stellt sich somit die Frage, ob
sich bei einer Veränderung der Pigmentierung der Kolonien von Blut- nach Schaedler-
Agar auch die Expression des σB, gemessen durch asp23, entsprechend verändert.
Zum Vergleich wurde von einer ∆sigB-Mutante das Transkriptionsprofil des
Wachstums auf Blut- und Schaedler-Agar ermittelt. Die Kolonien hatten eine weiße
Farbe, was auf eine verminderte sigB-Transkription schließen lässt. In der RT-PCR war
das Transkript asp23 sowohl von Blut- als auch von Schaedler-Agar nahezu Null.
Bei den klinischen Isolaten zeigte sich häufig eine Pigmentierung von weißen Kolonien
auf Blut- und gelben Kolonien auf Schaedler-Agar. Dies Verhalten trat bei 6 der 16
untersuchten klinischen Isolate auf. Von diesen wies lediglich 1 Isolat (S179 I) auf
Schaedler-Agar die vermutete höhere Expression von asp23 auf. 4 Isolate zeigten ein
annähernd gleich hohes Transkript auf beiden Agarmedien. Ein Transkript (S 592 II)
zeigte sogar auf Blutagar die höhere Expression von asp23.
Ebenfalls häufig zu beobachten war das Auftreten von Isolaten mit Kolonien einer gelb-
goldenen Farbe. So wuchsen 7 von 16 Isolaten mit gelbem Pigment auf beiden
Agarmedien. Auch hier war eine Differenz zwischen der relativen Menge von asp23 auf
Blut- und Schaedler-Agar zu beobachten. 4 Isolate zeigten eine doppelt bis zehnfach
erhöhte Expression von asp23 auf Schaedler-Agar. 3 Isolate exprimierten asp23 auf
beiden Agarmedien gleich stark.

4 Ergebnisse
77
Seltener ist das Auftreten von weißen Kolonien sowohl auf Blut- als auch auf
Schaedler-Agar, was nur bei 2 der untersuchten klinischen Isolate der Fall war. Beide
Isolate transkribierten auf beiden Agarmedien eine geringe Menge an asp23.

5 Diskussion
78
5 Diskussion
Die zystische Fibrose ist in der westlichen Bevölkerung die häufigste autosomal-
rezessiv vererbbare Stoffwechselerkrankung, die durch eine Mutation im CFTR-Gen
zustande kommt. Einer der wichtigsten therapeutischen Ansätze ist die Behandlung
rezidivierender bronchopulmonaler Infekte, die leider auch heute noch einen
lebenslimitierenden Faktor bei CF-Patienten darstellen. Einer der bedeutsamsten
Erreger dieser Infektionen ist S. aureus. Die Identifizierung der Erreger findet in erster
Linie in der mikrobiologischen Diagnostik durch die Kultivierung des Erregermaterials
aus Nasen-, Rachenabstrichen oder Sputum statt. Üblicherweise wird Selektivagar
verwendet, der andere Erreger im Wachstum inhibiert, und das Wachstum von S. aureus
fördert. Für die Identifizierung von S. aureus wird gewöhnlich Columbia-Blut-Agar und
Mannitol-Salz-Agar verwendet.
Zur phänotypischen Analyse der S. aureus Isolate aus dem Respirationstrakt von CF-
Patienten wurden die klinischen Proben auf verschiedene Agarplatten ausgestrichen und
das Wachstum der Kolonien nach Bebrütung über Nacht charakterisiert. Üblich ist dabei
die Verwendung von Columbia-Blut-Agar. In der CF-Studie, die die Grundlage für die
vorliegende Arbeit bildete, wurden die S. aureus-Isolate zusätzlich auf Schaedler-Agar
ausgestrichen und unter 5% CO2 bebrütet. Denn es wurde beobachtet, dass Unterschiede
im Phänotyp der einzelnen Isolate meist auf Schaedler-Agar noch besser zu erfassen
sind, da sich die Pigmentierung der Kolonien und das Hämolyseverhalten noch besser
ausprägen. Bei Schaedler-Agar handelt es sich um einen nicht-selektiven Nährboden,
der üblicherweise zur Kultivierung von anaeroben Bakterien eingesetzt wird.
In der vorliegenden Arbeit wurden ausgewählte klinische Isolate aus der CF-Studie und
ausgewählte Laborstämme und Mutanten auf Blut- und Schaedler-Agar bebrütet, deren
Phänotyp beschrieben und anschließend eine RNA-Isolierung für die Untersuchung der
Transkription vorgenommen. Ziel war es, das Transkriptionsprofil verschiedener
Virulenzgene und -regulatoren dieser Stämme auf Blut- und Schaedler-Agar mit dem
Verfahren der RT-PCR vergleichen zu können. Es sollte die Frage beantwortet werden,
ob es möglich sei, vom phänotypischen Erscheinungsbild der Kolonien auf
Agarmedien, Columbia-Blut- und Schaedler-Agar, auf das Transkriptionsprofil

5 Diskussion
79
wichtiger Virulenzregulatoren und -gene von S. aureus schließen zu können. Es wurden
die globalen Regulatoren sae, agr durch die Ermittlung von RNAIII und sigB durch die
Bestimmung von asp23, sowie die Virulenzgene hla und spa untersucht. Da es nicht
üblich ist, RNA-Isolierung von Agarplatten durchzuführen, wurden die untersuchten
Stämme zusätzlich noch in einer BHI-Flüssigkultur angezüchtet und aus diesen zu
verschiedenen Zeitpunkten des Wachstumsverlaufes RNA-Isolierungen durchgeführt.
Zu drei voneinander unabhängigen Zeitpunkten wurden die Wachstumsphasen eines
jeden Stammes ermittelt: frühe exponentielle, späte exponentielle und stationäre Phase.
BHI wurde als Flüssigmedium gewählt, da sich hier nach ca. 8 Stunden der typische
exponentielle Wachstumsverlauf entwickelte. Zum Vergleich wurde zu Beginn auch das
Flüssigmedium TSB verwendet, in dem sich allerdings kein klarer exponentieller
Wachstumsverlauf abzeichnete, so dass die einzelnen Wachstumsphasen nicht eindeutig
abzugrenzen waren. Daher wurde TSB als Flüssigmedium wieder verworfen.
Jeder S. aureus-Stamm wurde dreimal auf Blut- und Schaedler-Agar bzw. in der
Flüssigkultur angezüchtet, um zu drei unabhängigen Zeitpunkten die RNA isolieren zu
können. Von jedem RNA-Isolat wurde eine Doppelbestimmung in der RT-PCR
durchgeführt.
Unterschiede im genetischen Hintergrund der einzelnen S. aureus-Stämme können zu
einem völlig unterschiedlichen Transkriptionsprofil bei der Genexpression führen. Die
klinischen Isolate befanden sich längere Zeit (Einschlusskriterium war eine Persistenz
von mindestens sechs Monaten) im Respirationstrakt der CF-Patienten. Bei
rezidivierenden Infektionen mit entsprechender klinischer Symptomatik aufgrund der
Abwehrschwäche ist es notwendig, diese Patienten wiederholt mit Antibiotika zu
behandeln. Die Erreger entwickeln zunehmend Resistenzen und Veränderungen im
Erbgut, um der Immunantwort des Wirts und der Wirkung der Antibiotika zu entgehen.
So kommt es zu einer veränderten Transkription der Regulatoren und Virulenzgene von
S. aureus. Longitudinalstudien haben bereits gezeigt, dass Stämme, die denselben
genetischen Hintergrund haben, wiederholt aus dem Respirationstrakt von CF-Patienten
isoliert wurden und somit chronische Infektionen durch einen einzel nen Stamm
verursacht werden können (4). Vor allem bei chronischen Infektionen ist S. aureus
einem starken Selektionsdruck unterworfen, der durch die Therapie mit Antibiotika,
durch die Immunantwort des Wirts aber auch durch Co-Infektionen mit anderen

5 Diskussion
80
Mikroorganismen verursacht wird (23). So lässt sich möglicherweise die durchaus sehr
unterschiedliche Expression der in dieser Arbeit untersuchten Gene erklären, auch wenn
diese Isolate ein ähnliches phänotypisches Erscheinungsbild aufwiesen. In dieser Arbeit
wurden Isolate in Gruppen zusammengefasst, die aus dem Respirationstrakt von
verschiedenen Patienten isoliert worden waren. Hier haben sich vermutlich sehr
unterschiedliche adaptive Vorgänge abgespielt, die somit auch zu einer unterschiedlich
starken Expression der Virulenzgene und Regulatoren führen konnten. Ein interessanter
Aspekt für weitere Untersuchungen wäre es, die Expression der Virulenzgene von
Isolaten eines einzelnen Patienten zu vergleichen, um die Hypothese zu prüfen, ob die
Isolate möglicherweise einen ähnlichen genetischen Hintergrund vorzuweisen haben.
Zu unterschiedlichen Ergebnissen führen auch minimale Unterschiede in den
Wachstumsbedingungen. Die Ergebnisse dieser Arbeit zeigten, dass die Expression der
Virulenzgene und -regulatoren beim Wachstum auf festem Medium wie Blut- und
Schaedler-Agar und in dem Flüssigmedium BHI sehr unterschiedlich sind. Bei allen
klinischen Isolaten unterschied sich das Transkriptionsprofil zwischen Flüssigmedium,
Blut-Agar und Schaedler-Agar. So wurde auch asp23 des Laborstamms 8325-4 zwar in
der Flüssigkultur in allen drei Wachstumsphasen erfolgreich exprimiert, dagegen konnte
dafür asp23 auf den Agarplatten nicht suffizient nachgewiesen werden. Allerdings
zeigte sich bei RN6390, da dieser Stamm eine sigB-Mutation aufweist, wie erwartet,
eine nicht erfolgreiche Expression von asp23 in allen drei Medien. Zum Vergleich war
jedoch bei den vier Mutanten ∆agr, ∆sar, ∆agr/sar und ∆sigB eine ähnlich erfolgreiche
Expression zwischen dem festem Agar-Medium und dem Flüssigmedium zu erkennen.
Bei den klinischen Isolaten fiel vor allem bei den Isolaten aus der Gruppe II mit S 66 I,
S 516 I und S 376 I und aus der Gruppe III mit S 605 I und S 568 I auf, dass diese zwar
spa in der Flüssigkultur erfolgreich exprimierten, dass ein spa-Transkript allerdings auf
den Agarmedien nicht erfolgreich nachzuweisen war. Möglicherweise spielte hier die
Dauer der Inkubationszeit eine Rolle, so dass die längere Inkubation von 24 h bei den
Agarplatten zu einer Instabilität der RNA von spa geführt haben könnte.
Wichtig war es, die Transkription der Regulatoren und Gene zusätzlich in der
Flüssigkultur zu untersuchen, da hier die einzelnen Wachstumsphasen (prä-,
postexponentielle und stationäre Phase) genau ermittelt werden konnten. Auf den
Agarmedien wurden dagegen nur die Produkte der Gene und Regulatoren

5 Diskussion
81
nachgewiesen, die nach 24 Stunden Inkubation noch stabil waren. Mögliche instabile
Produkte, die zu diesem Zeitpunkt schon abgebaut wurden, konnten so nicht mehr
erfasst werden. Dass die Regulatoren und Virulenzgene jedoch erfolgreich exprimiert
wurden, ließ sich somit durch die zusätzliche Anzucht der Isolate in den Flüssigkulturen
bestätigen.
Viele Untersuchungen zur Genregulation und Expression von Virulenzfaktoren basieren
auf Studien mit bekannten S. aureus-Laborstämmen, wie dem Stamm 8325-4.
Durch die Wahl von BHI als Flüssigmedium zeigten die Laborstämme teilweise schon
ein unterschiedliches Verhalten als bisher in der Literatur beschrieben. Zum Beispiel lag
das Maximum der Expression von hla bei 8325-4 in der späten exponentiellen Phase.
Auch zeigte dieser Stamm eine Expression von sigB, die eigentlich nicht vorhanden sein
sollte, da es sich um eine Deletion in RsbU handelt. RN 6390 zeigte ebenfalls ein frühes
Maximum von hla in der späten exponentiellen Phase. Hingegen weißt dieser Stamm
tatsächlich die sigB-Deletion auf, was auch durch die vorliegenden Ergebnisse bestätigt
werden konnte.
Ein wichtiges Ergebnis dieser Untersuchung ist, dass eine große Variation in der
Genexpression der klinischen S. aureus-Stämme untereinander, aber auch im Vergleich
zu den Laborstämmen und Mutanten herrscht.
Desweiteren ist die initial phänotypische Charakterisierung der Kolonien auf den Agar-
Platten in gewisser Weise der subjektiven Betrachtungsweise des einzelnen
Untersuchers unterworfen. So können Kolonien, die nicht eindeutig in ihrer Farbe zu
beschreiben sind, von einem Untersucher anders interpretiert werden als von einem
anderen. Dadurch ist es z. B. schwer, definitiv zwischen weißen und gelben Kolonien zu
trennen. Die in dieser Arbeit erfolgte Zusammenfassung der klinischen Isolate in
Gruppen nach dem Hämolyseverhalten konnte auf diesem Hintergrund auch nicht rein
objektiv erfolgen. Es ergab sich daher in der Gruppe II mit Isolaten mit gering bis
mäßiger Hämolyse auf Blut-Agar die ungenaueste Einteilung. Hierdurch lassen sich
möglicherweise die wenigen Gemeinsamkeiten im Transkriptionsverhalten erklären.
Nur bei spa zeigte sich bei vier von sechs Isolaten eine geringere Expression des
Transkripts auf Schaedler-Agar. Alle anderen vier Virulenzgene und -regulatoren waren
bei den einzelnen Isolaten dieser Gruppe nicht miteinander zu korrelieren. Ein weiterer
Ansatz wäre somit, die Isolate zum einen von mehreren unabhängigen Untersuchern

5 Diskussion
82
hinsichtlich der Pigmentierung der Kolonien beurteilen zu lassen. Zum anderen wäre für
weitere Folgearbeiten eine strikte Trennung zwischen Isolaten mit und ohne
Hämolysezone sinnvoller. Dadurch wäre eine ungenaue Beurteilung der phänotypischen
Beschreibung der Kolonien vermeidbar.
Eine weitere wichtige Hypothese der Untersuchung war es, festzustellen, ob eine
Korrelation zwischen dem Hämolyseverhalten und der Expression von hla darzustellen
ist. Hierzu können vor allem die Ergebnisse der Gruppe III herangezogen werden, da in
dieser Gruppe das Hämolyseverhalten am stärksten ausgeprägt war. Sowohl auf Blut-
als auch auf Schaedler-Agar wurde die Stufe 4 des Hämolysegrads erreicht. Unsere
Vermutung ließ sich jedoch nicht belegen. Ein Isolat (S 568 I) wies sogar eine
reduzierte Expression von hla auf Schaedler-Agar auf, was nicht vermutet worden war.
Interessanterweise konnte bei dem Laborstamm 8325-4, der auf beiden Agarmedien die
maximale Hämolyse erreichte, eine sehr hohe Expression von hla dokumentiert werden.
Dieses Ergebnis lässt sich eventuell damit begründen, dass die klinischen Isolate durch
adaptive Vorgänge im Wirt Veränderungen auf RNA-Ebene durchgemacht haben, die
dadurch zu nicht vergleichbaren Ergebnissen mit den Laborstämmen führen.
Interessanterweise stellten sich bei der Untersuchung 2 klinische Isolate (S 348 I, S 179
I) offenbar als spa-Mutanten heraus. Keines der beiden Isolate exprimierte spa in den
einzelnen Wachstumsphasen der Flüssigkultur suffizient. Auch auf Blut- und Schaedler-
Agar wurde spa nicht transkribiert. Im Rahmen der Studie konnte in einer anderen
Untersuchung der spa-Typ bei beiden Isolaten ermittelt werden. Für S 179 I war dies
t015, für S348 I t2845. Damit kann die Mutation zwar nicht im dem Bereich der Primer-
Bindungsstelle für die spa-Typisierung, aber möglicherweise an anderer Stelle im Gen,
erfolgt sein. In einer früheren Arbeit (1) konnte bereits 2009 gezeigt werden, dass
klinische S. aureus-Stämme, bei denen eine spa-Typisierung nicht gelingt, häufig spa-
Mutanten sind. Bei der Sequenzierung zeigten sich vor allem Deletionen in den IgG-
bindenden Domainen. Allerdings konnte auch nachgewiesen werden, dass Stämme mit
Deletionen in mehreren Abschnitten in der RT-PCR sehr hohe Transkriptionen von spa
aufwiesen. Dies ist in der Weise interessant, da spa als wichtiger Virulenzfaktor gilt,
und somit spa-Mutanten in S.aureus-Isolaten nachgewiesen werden, die zu Infektionen
bei CF-Patienten geführt haben. Andererseits muss gerade bei diesen im Allgemeinen
abwehrgeschwächten Patienten, der Stamm gar nicht so virulent sein, um klinisch

5 Diskussion
83
relevante Infektionen zu verursachen. Es wäre somit ein weiterer interessanter Ansatz,
die klinischen Isolate aus dem Respirationstrakt von CF-Patienten hinsichtlich
möglicher Mutationen in den Virulenzgenen zu untersuchen. Bei den Isolaten der
Gruppe I, ausgenommen die spa-Mutante S 348 I, fielen relativ hohe spa-Transkripte
im Vergleich zu den anderen Gruppen auf.
Bei zwei klinischen Isolaten (S 1368 II und III), die zum gleichen Zeitpunkt aus dem
Respirationstrakt desselben Patienten isoliert worden waren, fiel eine ganz geringe
Expression von RNA III auf. Hierbei könnte es sich um natürliche agr-Mutanten
handeln. Ein interessantes Ergebnis, so dass man in weiteren Untersuchungen der Frage
nachgehen könnte, was ursächlich für die Mutation gewesen ist.
Die Transkription von asp23 wird durch den Regulator sigB beeinflusst (35). Da bei
einer hohen Aktivität von sigB auf eine verstärkte Pigmentierung der Kolonien
geschlossen werden kann, kann dies indirekt über asp23 hergeleitet werden. Somit
bildete die sigB-Mutante in dieser Untersuchung auch nur weiße Kolonien. Eine
Transkription von asp23 fand erwartungsgemäß nicht statt. Bei den klinischen Isolaten
fand sich bei allen Isolaten, die auf beiden Agarmedien gelbe Kolonien entwickelten,
eine erfolgreiche Expression von asp23. Allerdings wies auch der Stamm S 1344, der
nur weiße Kolonien bildete, eine erfolgreiche Expression von asp23 auf.
Interessanterweise zeigte sich jedoch bei den klinischen Isolaten S 1365 I und II ein sehr
ähnliches Bild in der relativen Menge der Genexpression, obwohl diese in ihrer
Pigmentierung unterschiedlich waren. Denn S 1365 I zeigte auf Schaedler-Agar die
seltene weiße Pigmentierung, die nur zu 24 % in der Studie beobachtet worden war.
Passend dazu wurde asp23, als sigB-abhängiges Transkript, nicht erfolgreich
exprimiert. Dafür zeigte S 1365 II die mit 72 % häufigeren gelben Kolonien auf
Schaedler-Agar und wie erwartet, eine Expression von asp23.
Aus den Ergebnissen dieser Arbeit lässt sich ableiten, dass es nicht oder nur bedingt
möglich ist, von der phänotypischen Erscheinung der klinischen Isolate auf das
Transkriptionsprofil wichtiger Regulatoren und Virulenzgene von S. aureus zu
schließen. Da bisher keine Untersuchungen zum Transskriptionsprofil der Erreger auf
Agarplatten vorlagen, war ein wichtiges Ergebnis dieser Arbeit, dass es möglich ist,
RNA der Bakterien von Agarplatten nachzuweisen, dass es Unterschiede zum
Transskriptionsverhalten der Erreger auf den verschiedenen Agarplatten (Blut- und

5 Diskussion
84
Schaedler-Agar) unter den verschiedenen Kultivierungsbedingungen (normale
Bedingungen und CO2) gibt und dass sich das Transskriptionsverhalten auf Agarplatten
zu dem in Flüssigkulturen unterscheidet. Interessant waren auch die Ergebnisse des
Transskriptionsverhaltens wichtiger Laborstämme und deren Mutanten, mit denen die
klinischen Stämme verglichen wurden. Weiterhin wurden im Rahmen dieser Arbeit
durch Auswahl der klinischen Isolate einige interessante klinische Mutanten gefunden
(spa-Mutanten, agr-Mutanten).
Die Arbeit dient somit als Anstoß für weitere Experimente und Fragestellungen: Wie
unterscheidet sich das Transkriptionsverhalten von Isolaten eines einzelnen Patienten?
Treten hier möglicherweise ähnliche Mutationen auf? Inwieweit und wie häufig treten
Mutationen in den Genen von diesen klinischen Stämmen auf, die die adaptiven
Vorgänge der S. aureus-Stämme im Wirt erklären könnten? Treten diese Mutationen
erst bei chronischer Infektion mit langer Persistenz im Wirt oder möglicherweise auch
schon nach kurzer Zeit bei akuten Infektexazerbationen auf?

6 Literaturverzeichnis
85
6 Literaturverzeichnis
(1) Baum C, Haslinger-Loffler B, Westh H, Boye K, Peters G, Neumann C, et al. (2009) Non-spa-typeable clinical Staphylococcus aureus strains are naturally occurring protein
A mutants. J.Clin.Microbiol. 47 (11): 3624-3629.
(2) Bayer M, Heinrichs J, Cheung A (1996) The molecular architecture of the sar locus
in Staphylococcus aureus. J.Bacteriol. 178 (15): 4563-4570.
(3) Bischoff M, Entenza JM, Giachino P (2001) Influence of a Functional sigB Operon
on the Global Regulators sar and agr in Staphylococcus aureus. J.Bacteriol. 183 (17):
5171-5179
(4) Branger C, Gardye C, Lambert-Zechovsky N (1996) Persistence of Staphylococcus
aureus strains among cystic fibrosis patients over extended periods of time. Journal of
Medical Microbiology 45 (4): 294-301.
(5) Chatterjee I, Kriegeskorte A, Fischer A, Deiwick S, Theimann N, Proctor RA, et al.
(2008) In vivo mutations of thymidylate synthase (encoded by thyA) are responsible for
thymidine dependency in clinical small-colony variants of Staphylococcus aureus.
J.Bacteriol. 190 (3): 834-842.
(6) Chatterjee I, Becker P, Grundmeier M, Bischoff M, Somerville GA, Peters G, et al.
(2005) Staphylococcus aureus ClpC Is Required for Stress Resistance, Aconitase
Activity, Growth Recovery, and Death. J.Bacteriol. 187 (13): 4488-4496.
(7) Cheung AL, Projan SJ (1994) Cloning and sequencing of sarA of Staphylococcus
aureus, a gene required for the expression of agr. J.Bacteriol. 176 (13): 4168-4172.
(8) Cheung AL, Ying P (1994) Regulation of alpha- and beta-hemolysins by the sar
locus of Staphylococcus aureus. J.Bacteriol. 176 (3): 580-585.
(9) Cheung AL, Eberhardt KJ, Chung E, Yeaman MR, Sullam PM, Ramos M, et al.
(1994) Diminished virulence of a sar-/agr- mutant of Staphylococcus aureus in the
rabbit model of endocarditis. J.Clin.Invest. 94 (5): 1815-1822.
(10) Cheung A, Eberhardt K, Heinrichs J (1997) Regulation of protein A synthesis by
the sar and agr loci of Staphylococcus aureus. Infect.Immun. 65 (6): 2243-2249.
(11) Cheung AL, Chien Y, Bayer AS (1999) Hyperproduction of Alpha-Hemolysin in a
sigB Mutant Is Associated with Elevated SarA Expression in Staphylococcus aureus.
Infect.Immun. 67 (3): 1331-1337.
(12) Cheung AL, Nishina K, Manna AC (2008) SarA of Staphylococcus aureus Binds to
the sarA Promoter To Regulate Gene Expression. J.Bacteriol. 190 (6): 2239-2243.

6 Literaturverzeichnis
86
(13) Davis PB (2006) Cystic Fibrosis Since 1938. Am.J.Respir.Crit.Care Med. 173 (5):
475-482.
(14) Deora R, Tseng T, Misra TK (1997) Alternative transcription factor sigmaSB of
Staphylococcus aureus: characterization and role in transcription of the global
regulatory locus sar. J.Bacteriol. 179 (20): 6355-6359.
(15) Köhnlein, T, Welte T (2008) Mukoviszidose. In: Harrisons Innere Medizin. Dietel
M, Suttorp N, Zeitz M [Hrsg] ABW Wissenschaftsverlag GmbH, Berlin, Bd. 2, 17.
Aufl.: 2009-2013.
(16) Elizur A, Orscheln RC, Ferkol TW, Atkinson JJ, Dunne WM,Jr, Buller RS, et al.
(2007) Panton-Valentine Leukocidin-positive methicillin-resistant Staphylococcus
aureus lung infection in patients with cystic fibrosis. Chest 131 (6): 1718-1725.
(17) Foster TJ (2005) Immune evasion by staphylococci. Nat.Rev.Microbiol. 3 (12):
948-958.
(18) Gertz S, Engelmann S, Schmid R, Ohlsen K, Hacker J, Hecker M (1999)
Regulation of sigmaB-dependent transcription of sigB and asp23 in two different
Staphylococcus aureus strains. Mol.Gen.Genet. 261 (3): 558-566.
(19) Giraudo AT, Mansilla C, Chan A, Raspanti C, Nagel R (2003) Studies on the
expression of regulatory locus sae in Staphylococcus aureus. Curr.Microbiol. 46 (4):
246-250.
(20) Goerke C, Fluckiger U, Steinhuber A, Zimmerli W, Wolz C (2001) Impact of the
regulatory loci agr, sarA and sae of Staphylococcus aureus on the induction of alpha-
toxin during device-related infection resolved by direct quantitative transcript analysis.
Mol.Microbiol. 40 (6): 1439-1447.
(21) Goerke C, Campana S, Bayer MG, Doring G, Botzenhart K, Wolz C (2000) Direct
Quantitative Transcript Analysis of the agr Regulon of Staphylococcus aureus during
Human Infection in Comparison to the Expression Profile In Vitro. Infect.Immun. 68
(3): 1304-1311.
(22) Gomez MI, Lee A, Reddy B, Muir A, Soong G, Pitt A, et al. (2004)
Staphylococcus aureus protein A induces airway epithelial inflammatory responses by
activating TNFR1. Nat.Med. 10 (8): 842-848.
(23) Harrison F (2007) Microbial ecology of the cystic fibrosis lung. Microbiology 153
(4): 917-923.
(24) Higuchi R, Dollinger G, Walsh PS, Griffith R (1992) Simultaneous amplification
and detection of specific DNA sequences. Biotechnology (N.Y) 10 (4): 413-417.
(25) Hiramatsu K (2001) Vancomycin-resistant Staphylococcus aureus: a new model of
antibiotic resistance. Lancet Infect.Dis. 1 (3): 147-155.

6 Literaturverzeichnis
87
(26) Horsburgh MJ, Aish JL, White IJ, Shaw L, Lithgow JK, Foster SJ (2002)
{sigma}B Modulates Virulence Determinant Expression and Stress Resistance:
Characterization of a Functional rsbU Strain Derived from Staphylococcus aureus 8325-
4. J.Bacteriol. 184 (19): 5457-5467.
(27) Huntzinger E, Boisset S, Saveanu C, Benito Y, Geissmann T, Namane A, et al.
(2005) Staphylococcus aureus RNAIII and the endoribonuclease III coordinately
regulate spa gene expression. EMBO J. 24 (4): 824-835.
(28) Huseby M, Shi K, Brown CK, Digre J, Mengistu F, Seo KS, et al. (2007) Structure
and Biological Activities of Beta Toxin from Staphylococcus aureus. J.Bacteriol. 189
(23): 8719-8726.
(29) Jarraud S, Lyon GJ, Figueiredo AM, Gerard L, Vandenesch F, Etienne J, et al.
(2000) Exfoliatin-producing strains define a fourth agr specificity group in
Staphylococcus aureus. J.Bacteriol. 182 (22): 6517-6522.
(30) Jonas D, Walev I, Berger T, Liebetrau M, Palmer M, Bhakdi S (1994) Novel path
to apoptosis: small transmembrane pores created by staphylococcal alpha-toxin in T
lymphocytes evoke internucleosomal DNA degradation. Infect.Immun. 62 (4): 1304-
1312.
(31) Kahl B, Herrmann M, Everding AS, Koch HG, Becker K, Harms E, et al. (1998)
Persistent infection with small colony variant strains of Staphylococcus aureus in
patients with cystic fibrosis. J.Infect.Dis. 177 (4): 1023-1029.
(32) Kahl BC, Becker K, Friedrich AW, Clasen J, Sinha B, Von Eiff C, et al. (2003)
agr-dependent bacterial interference has no impact on long-term colonization of
Staphylococcus aureus during persistent airway infection of cystic fibrosis patients.
J.Clin.Microbiol. 41 (11): 5199-5201.
(33) Kahl BC, Duebbers A, Lubritz G, Haeberle J, Koch HG, Ritzerfeld B, et al. (2003)
Population Dynamics of Persistent Staphylococcus aureus Isolated from the Airways of
Cystic Fibrosis Patients during a 6-Year Prospective Study. J.Clin.Microbiol. 41 (9):
4424-4427.
(34) Kubista M, Andrade JM, Bengtsson M, Forootan A, Jonák J, Lind K, et al. (2006)
The real-time polymerase chain reaction. Mol.Aspects Med. 27 (2-3): 95-125.
(35) Kullik II, Giachino P (1997) The alternative sigma factor sigmaB in
Staphylococcus aureus: regulation of the sigB operon in response to growth phase and
heat shock. Arch.Microbiol. 167 (2/3): 151-159.
(36) Kullik I, Giachino P, Fuchs T (1998) Deletion of the Alternative Sigma Factor
sigma B in Staphylococcus aureus Reveals Its Function as a Global Regulator of
Virulence Genes. J.Bacteriol. 180 (18): 4814-4820.

6 Literaturverzeichnis
88
(37) Labandeira-Rey M, Couzon F, Boisset S, Brown EL, Bes M, Benito Y, et al.
(2007) Staphylococcus aureus Panton-Valentine Leukocidin Causes Necrotizing
Pneumonia. Science 315 (5815): 1130-1133.
(38) Lowy FD (1998) Staphylococcus aureus infections. N.Engl.J.Med. 339 (8): 520-
532.
(39) Manna AC, Bayer MG, Cheung AL (1998) Transcriptional Analysis of Different
Promoters in the sar Locus in Staphylococcus aureus. J.Bacteriol. 180 (15): 3828-3836.
(40) Morfeldt E, Taylor D, von Gabain A, Arvidson S (1995) Activation of alpha-toxin
translation in Staphylococcus aureus by the trans-encoded antisense RNA, RNAIII.
EMBO J. 14 (18): 4569-4577.
(41) Musser JM, Schlievert PM, Chow AW, Ewan P, Kreiswirth BN, Rosdahl VT, et al.
(1990) A single clone of Staphylococcus aureus causes the majority of cases of toxic
shock syndrome. Proc.Natl.Acad.Sci.U.S.A. 87 (1): 225-229.
(42) Novick RP (1990) The Staphylococcus as a molecular genetic system. In: R.P.
Novick (Hrsg) Molecular biology of the staphylocci. VCH Publishers, New York, New
York, S. 1-40.
(43) Novick R (1967) Properties of a cryptic high-frequency transducing phage in
Staphylococcus aureus. Virology 33 (1): 155-166.
(44) Novick RP, Ross HF, Projan SJ, Kornblum J, Kreiswirth B, Moghazeh S (1993)
Synthesis of staphylococcal virulence factors is controlled by a regulatory RNA
molecule. EMBO J. 12 (10): 3967-3975.
(45) Novick RP (2003) Autoinduction and signal transduction in the regulation of
staphylococcal virulence. Mol.Microbiol. 48 (6): 1429-1449.
(46) Novick RP, Jiang D (2003) The staphylococcal saeRS system coordinates
environmental signals with agr quorum sensing. Microbiology 149 (Pt 10): 2709-2717.
(47) Ohl F, Jung M, Xu C, Stephan C, Rabien A, Burkhardt M, et al. (2005) Gene
expression studies in prostate cancer tissue: which reference gene should be selected for
normalization? J.Mol.Med. 83 (12): 1014-1024.
(48) O'Riordan K, Lee JC (2004) Staphylococcus aureus capsular polysaccharides.
Clin.Microbiol.Rev. 17 (1): 218-234.
(49) Palmqvist N, Foster T, Tarkowski A, Josefsson E (2002) Protein A is a virulence
factor in Staphylococcus aureus arthritis and septic death. Microb.Pathog. 33 (5): 239-
249.

6 Literaturverzeichnis
89
(50) Peng HL, Novick RP, Kreiswirth B, Kornblum J, Schlievert P (1988) Cloning,
characterization, and sequencing of an accessory gene regulator (agr) in Staphylococcus
aureus. J.Bacteriol. 170 (9): 4365-4372.
(51) Pletz MW, Sauer-Heilborn A, Kohnlein T, Seidler U, Lamprecht G (2010) Cystic
fibrosis in adults. Internist (Berl) 51 Suppl 1: 277-288.
(52) Projan SJ, Novick RP The molecular Basis of Pathogenicity. In: The molecular
Basis of Pathogenicity. The Staphylococci in Human Disease. Wiley-Blackwell, S. 55-
81.
(53) Ragle BE, Bubeck Wardenburg J (2009) Anti-Alpha-Hemolysin Monoclonal
Antibodies Mediate Protection against Staphylococcus aureus Pneumonia.
Infect.Immun. 77 (7): 2712-2718.
(54) Recsei P, Kreiswirth B, O'Reilly M, Schlievert P, Gruss A, Novick RP (1986)
Regulation of exoprotein gene expression in Staphylococcus aureus by agar.
Mol.Gen.Genet. 202 (1): 58-61.
(55) Schlievert PM, Strandberg KL, Lin YC, Peterson ML, Leung DY (2010) Secreted
virulence factor comparison between methicillin-resistant and methicillin-sensitive
Staphylococcus aureus, and its relevance to atopic dermatitis. J.Allergy Clin.Immunol.
125 (1): 39-49.
(56) Smeltzer MS, Lee CY, Harik N, Hart ME (2009) Molecular Basis of Pathogenicity.
In: K. B. Crossley, K. K. Jefferson, G. L. Archer (Hrsg) Staphylococci in Human
Disease. Wiley-Blackwell, 2. Aufl., S. 65-109.
(57) Traber KE, Lee E, Benson S, Corrigan R, Cantera M, Shopsin B, et al. (2008) agr
function in clinical Staphylococcus aureus isolates. Microbiology 154 (8): 2265-2274.
(58) Vandenesch F, Kornblum J, Novick RP (1991) A temporal signal, independent of
agr, is required for hla but not spa transcription in Staphylococcus aureus. J.Bacteriol.
173 (20): 6313-6320.
(59) Vandenesch F, Naimi T, Enright MC, Lina G, Nimmo GR, Heffernan H, et al.
(2003) Community-acquired methicillin-resistant Staphylococcus aureus carrying
Panton-Valentine leukocidin genes: worldwide emergence. Emerg.Infect.Dis. 9 (8):
978-984.
(60) Wadstrom T, Mollby R (1971) Studies on extracellular proteins from
Staphylococcus aureus. VII. Studies on -haemolysin. Biochim.Biophys.Acta 242 (1):
308-320.
(61) Waldvogel FA (1985) Staphylococcus aureus. In: Staphylococcus aureus.
Principles and practice of infectious diseases. John Wiley & Sons, New York, 5. Aufl.,
S. 1097-1116.

6 Literaturverzeichnis
90
(62) Wardenburg JB, Patel RJ, Schneewind O (2007) Surface Proteins and Exotoxins
Are Required for the Pathogenesis of Staphylococcus aureus Pneumonia. Infect.Immun.
75 (2): 1040-1044.
(63) Wu S, de Lencastre H, Tomasz A (1996) Sigma-B, a putative operon encoding
alternate sigma factor of Staphylococcus aureus RNA polymerase: molecular cloning
and DNA sequencing. J.Bacteriol. 178 (20): 6036-6042.
(64) Zipper H, Brunner H, Bernhagen J, Vitzthum F (2004) Investigations on DNA
intercalation and surface binding by SYBR Green I, its structure deter mination and
methodological implications. Nucl.Acids Res. 32 (12): e103.

7 Abbildungsverzeichnis
91
7 Abbildungsverzeichnis
Abb. 1: Wachstumskurve des Laborstammes SH 1000 in BHI ......................................12
Abb. 2: RT-PCR Zyklus aus (34) ....................................................................................18
Abb. 3: Bestimmung des Wachstumskurvenverlaufes (WK) zu drei unterschiedlichen
Zeiten des klinischen Isolates S 605 I .............................................................................24
Abb. 4 Häufigkeitsverteilung der Pigmentierung der Kolonien auf Blut- und
Schaedleragar...................................................................................................................31
Abb. 5 Häufigkeitsverteilung der Hämolysestufe der Kolonien auf Blut- und Schaedler-
Agar .................................................................................................................................31
Abb. 6 Häufigkeitsverteilung der β-Toxin-Expression der klinischen Isolate ................32
Abb. 7: Vergleich des Wachstumsverlaufs des klinischen Isolates S 605 I in BHI- und
TSB- Flüssigmedium .......................................................................................................36
Abb. 8: Ermittlung des Wachstumsverlaufs zu drei verschiedenen Zeitpunkten (WK)..36
Abb. 9: Genexpression des Laborstamms 8325-4 in der Flüssigkultur ...........................38
Abb. 10: Genexpression des Laborstamms 8325-4 auf den Agarkulturen ......................38
Abb. 11: Genexpression des Laborstamms RN 6390 in der Flüssigkultur .....................40
Abb. 12: Genexpression des Laborstamms RN6390 auf den Agarkulturen....................40
Abb. 13: Genexpression des Laborstamms SH 1000 in der Flüssigkultur ......................42
Abb. 14: Genexpression des Laborstamms SH 1000 auf den Agarkulturen ...................42
Abb. 15: Genexpression der ∆agr-Mutante in der Flüssigkultur ....................................44
Abb. 16: Genexpression der ∆agr-Mutante auf den Agarkulturen .................................44
Abb. 17: Genexpression der ∆sar-Mutante in der Flüssigkultur .....................................46
Abb. 18: Genexpression der ∆sar-Mutante auf den Agarkulturen ..................................46
Abb. 19: Genexpression der ∆agr-/sar-Mutante in der Flüssigkultur .............................48
Abb. 20: Genexpression der ∆agr-/sar-Mutante auf den Agarkulturen ..........................48
Abb. 21: Genexpression der ∆sigB-Mutante in der Flüssigkultur ...................................50
Abb. 22: Genexpression der ∆sigB-Mutante auf den Agarkulturen ................................50
Abb. 23: Isolat S 1368 II mit Hämolyse 0 auf Blut-Agar (li) und mit Hämolyse 3 auf
Schaedler-Agar (re) .........................................................................................................53
Abb. 24: Genexpression von S 198 II in der Flüssigkultur .............................................54
Abb. 25: Genexpression von S 198 II auf den Agarkulturen ..........................................54
Abb. 26: Genexpression von S 348 I in der Flüssigkultur ...............................................55

7 Abbildungsverzeichnis
92
Abb. 27: Genexpression von S 348 I auf den Agarkulturen............................................55
Abb. 28: Genexpression von S 385 IV in der Flüssigkultur ............................................56
Abb. 29: Genexpression von S 385 IV auf den Agarkulturen .........................................56
Abb. 30: Genexpression von S 1368 II auf den Agarkulturen ........................................57
Abb. 31: Genexpression von S 1368 III auf den Agarkulturen .......................................57
Abb. 32: Isolat S 516 I mit Hämolysestufe 2 auf Blut-Agar (li) und Hämolysestufe 3 auf
Schaedler-Agar (re) .........................................................................................................59
Abb. 33: Genexpression von S 66 I in der Flüssigkultur .................................................60
Abb. 34: Genexpression von S 66 I auf den Agarkulturen..............................................60
Abb. 35: Genexpression von S 179 I in der Flüssigkultur ...............................................61
Abb. 36: Genexpression von S 179 I auf den Agarkulturen............................................61
Abb. 37: Genexpression von S 516 I in der Flüssigkultur ...............................................62
Abb. 38: Genexpression von S 516 I auf den Agarkulturen............................................62
Abb. 39: Genexpression von S 592 II in der Flüssigkultur .............................................63
Abb. 40: Genexpression von S 592 II auf den Agarkulturen ..........................................63
Abb. 41: Genexpression von S 376 I in der Flüssigkultur ...............................................64
Abb. 42: Genexpression von S 376 I auf den Agarkulturen............................................64
Abb. 43: Genexpression von S 501 II in der Flüssigkultur .............................................65
Abb. 44: Genexpression von S 501 II auf den Agarkulturen ..........................................65
Abb. 45: Isolat S 1365 I mit Hämolysestufe 4 auf Blut -(li) und Schaedler-Agar (re) ...67
Abb. 46: Genexpression von S 605 I in der Flüssigkultur ...............................................68
Abb. 47: Genexpression von S 605 I auf den Agarkulturen............................................68
Abb. 48: Genexpression von S 568 I in der Flüssigkultur ...............................................69
Abb. 49: Genexpression von S 568 I auf den Agarkulturen............................................69
Abb. 50: Genexpression von S 1344 auf den Agarkulturen ............................................70
Abb. 51: Genexpression von S 1365 I auf den Agarkulturen ..........................................71
Abb. 52: Genexpression von S 1365 II auf den Agarkulturen ........................................71
Abb. 53: S 179 I mit Hämolysegrad 1 auf Blut-Agar (li) und Hämolysegrad 4 auf
Schaedler-Agar (re) .........................................................................................................75

8 Tabellenverzeichnis
93
8 Tabellenverzeichnis
Tabelle 1: Laborstämme ..................................................................................................20
Tabelle 2: Mutanten .........................................................................................................20
Tabelle 3: klinische Isolate ..............................................................................................21
Tabelle 4: Sequenzen der Referenzgene ..........................................................................28
Tabelle 5: Sequenzen der in dieser Arbeit untersuchten Gene ........................................28
Tabelle 6: Die Phänotypen der Laborstämme .................................................................33
Tabelle 7: Die Phänotypen der Mutanten ........................................................................33
Tabelle 8: Die Phänotypen der Gruppe I .........................................................................34
Tabelle 9: Die Phänotypen der Gruppe II........................................................................34
Tabelle 10: Die Phänotypen der Gruppe III ....................................................................35
Tabelle 11: Legende: w: weiß, g: gelb. n: nein, j: ja. 2: Änderung mindest. doppelt so
hoch; 10: Änderung fast 10fach so hoch; - : geringere Transkription; 0: keine Änderung;
ne: nicht exprimiert..........................................................................................................74

9 Abkürzungen
94
9 Abkürzungen
BHI Brain heart infusion
cAMP Cyclisches Adenosinmonophosphat
CF Cystische Fibrose
cDNA complementary deoxyribonucleic acid
dsDNA Doppelstrang DNA
EL early log phase
g Vielfaches der Erdbeschleunigung
kb Kilobase
li links
LL late log phase
OD optische Dichte
PCR Polymerase Ketten Reaktion
re rechts
rpm rounds per minute
RT-PCR Real-Time PCR
s Sekunde
SP stationary phase
TSB Tryptic soy broth
WK Wachstumskurve
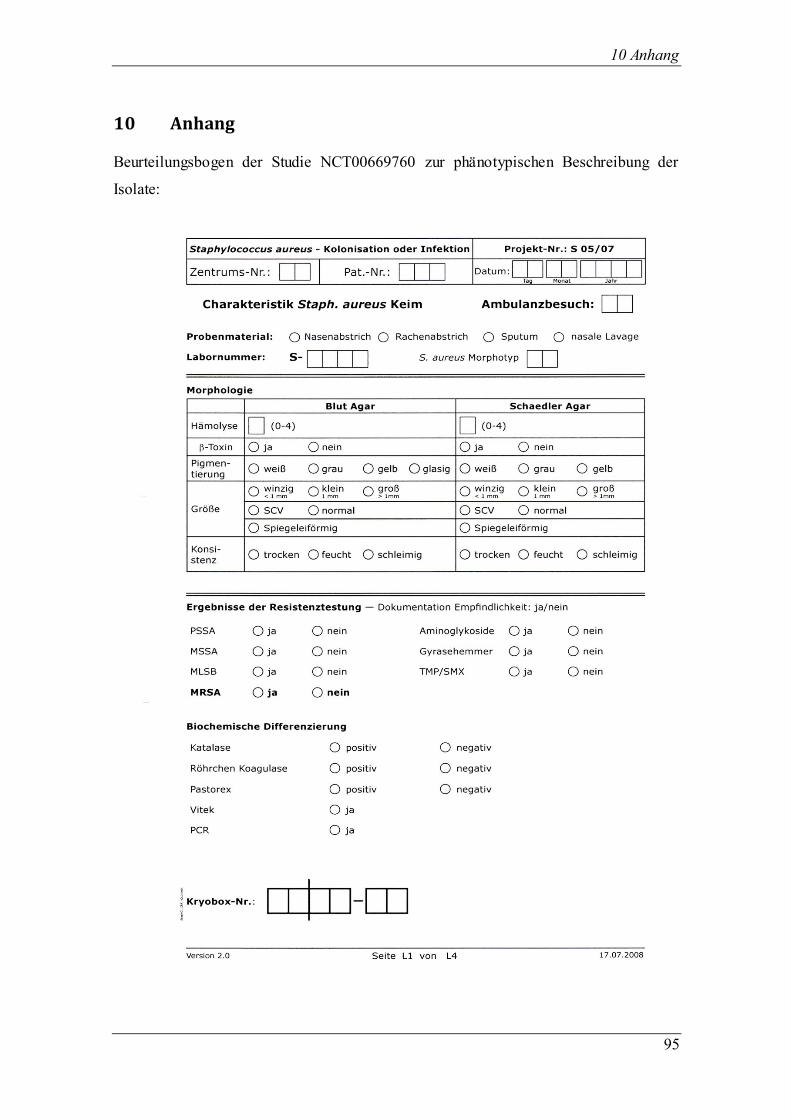
10 Anhang
95
10 Anhang
Beurteilungsbogen der Studie NCT00669760 zur phänotypischen Beschreibung der
Isolate:

11 Lebenslauf
96
11 Lebenslauf

11 Lebenslauf
97





























