Embed Size (px)
Citation preview

E I N F Ü H R U N G I N D I E M E T H O D E N
D E R T I E R H Y S I O L O G I E
Die Tierphysiologie ist eine Teildisziplin der Zoologie. Sie befasst sich mit den
Funktionen tierischer Gewebe, Organe und Organsysteme, und damit, wie diese
Funktionen und deren Zusammenspiel gesteuert und reguliert werden. Da die
Mechanismen, die zu einer spezifischen Organfunktion führen, auf den physi-
kalischen, biochemischen und informationsverarbeitenden Eigenschaften der
beteiligten Zellen und Geweben beruhen, sind die Arbeitsweisen der Tier-
physiologie entsprechend vielfältig. Tierphysiologen wenden daher sowohl Me-
thoden aus der Molekularbiologie an, etwa wenn die Strukturen von Ionen-
kanälen oder Antigenen erforscht werden sollen, häufig kommen aber auch
Messverfahren zum Einsatz, die ganz andere physikalische Messgrößen erfassen,
wenn beispielsweise die räumliche Auflösung eines Sehorgans, der zeitliche
Verlauf eines Aktionspotenzials, oder die aerodynamischen Eigenschaften eines
Vogelflügels experimentell ermittelt werden.
In diesem Kurs werden Sie in vier verschiedenen Kurseinheiten mit Frage-
stellungen und Messmethoden der Tierphysiologie vertraut gemacht. Dabei
werden Sie Experimente durchführen, die Einblicke in die Funktionsweise
tierischer Organismen und ihrer Organe demonstrieren. Die Experimente werden
teilweise an Tierpräparaten vorgenommen, die Sie selbst herstellen, wie im Herz-
und dem Muskelversuch oder beim Elektroretinogramm der Fliege, andererseits
werden Sie auch an Ihrem eigenen Körper arbeiten, wie z.B. in der Psychophysik.
THEORETISCHE VORAUSSETZUNGEN
MESSVERFAHREN UND SENSOREN
Da die Arbeitsweise der tierphysiologischen Forschung so vielfältig ist, ist es
notwendig, dass Sie sich bei jedem Versuch klar machen, welche Messgröße Sie
erfassen und dass Sie das Verfahren kennen, mit dem diese Größe erhoben wird.
Man unterscheidet dabei zwischen direkten und indirekten Messverfahren.
Während bei einem direkten Messverfahren das Messergebnis direkt am Mess-
gerät abgelesen werden kann - beispielsweise bei der Längenmessung mit einem
Lineal - wird bei einer indirekten Messung ein Messwandler (Sensor) eingesetzt,
der eine primäre Messgröße, die nur schwierig zu erfassen ist, in eine gut
messbare (sekundäre) Ausgangsgröße umsetzt. Sehr häufig eingesetzt werden
Sensoren, die abhängig von der zu messenden Größe ihre elektrischen Eigen-
schaften (Widerstand, Ausgangsspannung, Kapazität, usw.) ändern, da
elektrische Signale gut handhabbar sind, insbesondere, wenn noch weitere Signal
verarbeitende Stufen, wie Verstärker oder Filter nachgeschaltet werden müssen.
Jeder Sensor wird durch seine Kennlinie charakterisiert, mit deren Hilfe man die
Umrechnung der Ausgangsgröße zurück in die primäre Messgröße vornehmen
kann. Diese Kennlinie findet man in der Regel im Datenblatt des Herstellers des
Sensors. Hat man dieses Datenblatt nicht zur Hand, oder ändern sich die
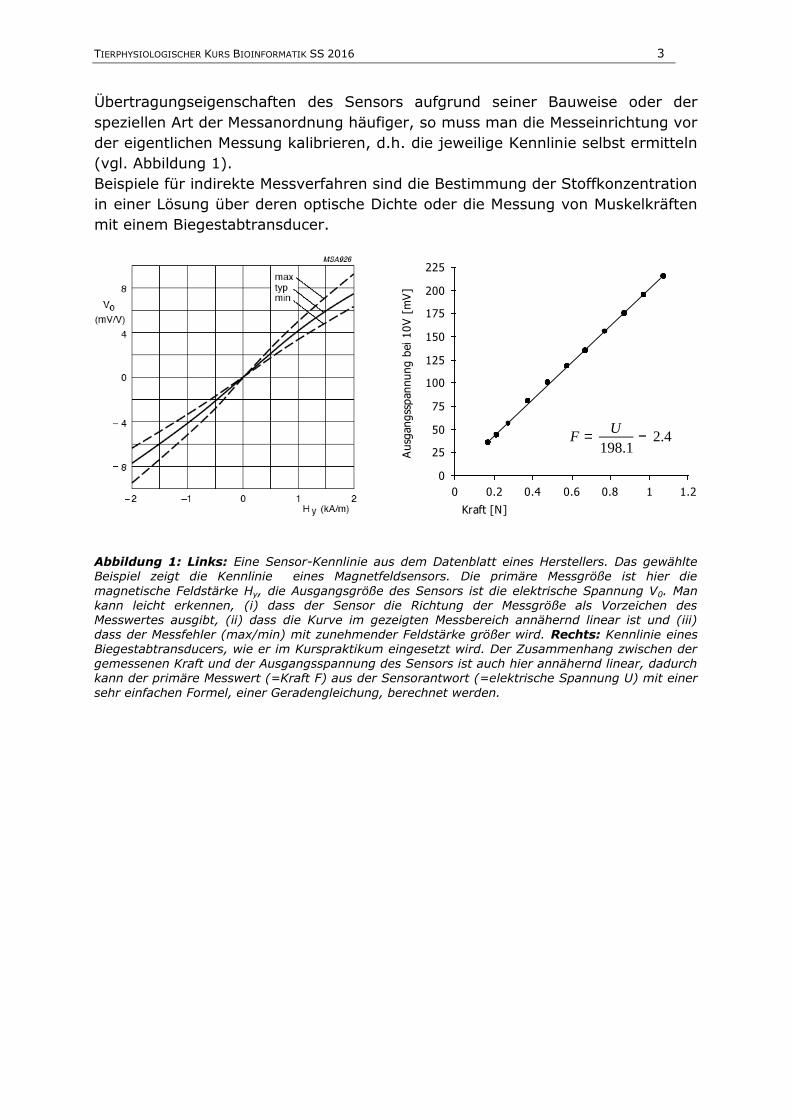
TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 3
Übertragungseigenschaften des Sensors aufgrund seiner Bauweise oder der
speziellen Art der Messanordnung häufiger, so muss man die Messeinrichtung vor
der eigentlichen Messung kalibrieren, d.h. die jeweilige Kennlinie selbst ermitteln
(vgl. Abbildung 1).
Beispiele für indirekte Messverfahren sind die Bestimmung der Stoffkonzentration
in einer Lösung über deren optische Dichte oder die Messung von Muskelkräften
mit einem Biegestabtransducer.
Abbildung 1: Links: Eine Sensor-Kennlinie aus dem Datenblatt eines Herstellers. Das gewählte Beispiel zeigt die Kennlinie eines Magnetfeldsensors. Die primäre Messgröße ist hier die
magnetische Feldstärke Hy, die Ausgangsgröße des Sensors ist die elektrische Spannung V0. Man kann leicht erkennen, (i) dass der Sensor die Richtung der Messgröße als Vorzeichen des Messwertes ausgibt, (ii) dass die Kurve im gezeigten Messbereich annähernd linear ist und (iii) dass der Messfehler (max/min) mit zunehmender Feldstärke größer wird. Rechts: Kennlinie eines Biegestabtransducers, wie er im Kurspraktikum eingesetzt wird. Der Zusammenhang zwischen der gemessenen Kraft und der Ausgangsspannung des Sensors ist auch hier annähernd linear, dadurch kann der primäre Messwert (=Kraft F) aus der Sensorantwort (=elektrische Spannung U) mit einer
sehr einfachen Formel, einer Geradengleichung, berechnet werden.
y = 198.08 x + 2.3653
0
25
50
75
100
125
150
175
200
225
0 0.2 0.4 0.6 0.8 1 1.2
Kraft [N]
Ausg
angss
pannung b
ei 10V
[m
V]
F =U
198.1− 2.4

+
-U
R3
R4
R2
R1
KASTEN 1: FUNKTIONSWEISE EINES BIEGESTABTRANSDUCERS ZUR MESSUNG VON KRÄFTEN
Ein Beispiel für ein indirektes Messverfahren, das Sie in diesem Kurs kennen lernen
werden, ist die Messung von Kräften mit Hilfe eines Biegestabtransducers. Der hier im
Kurs verwendete Typ ist aufgebaut aus einem Metallstab, auf den vier einzelne
Dehnungsmessstreifen (DMS) befestigt sind. Bei den DMS handelt es sich um Bauteile,
deren elektrischer Widerstand zunimmt, wenn sie gedehnt werden. Die vier DMS in
unserem Biegestabtransducer sind zu einer so genannten Wheatstone-Messbrücke
verschaltet, wie sie in der
nebenstehenden Abbildung
dargestellt ist. An zwei
„gegenüber liegenden“
Knotenpunkten der Brücke
wird eine konstante
Spannungsquelle (U) ange-
legt, an die anderen zwei ein
Spannungs-Messgerät.
R1 und R4 bzw. R2 und R3
bilden jeweils einen
Spannungsteiler, wobei die
beiden Widerstände eines
Zweiges jeweils auf gegen-
über liegenden Seiten des
Biegestabes angebracht werden: R4 und R2 auf der einen, R1 und R3 auf der anderen
Seite. Da im Ruhezustand (=Metallstab gerade) alle DMS gleich lang sind, sind auch alle
Widerstände gleich groß, das Spannungsmessgerät zeigt deshalb eine Spannungs-
differenz von 0 V an.
Belastet man den Stab nun mechanisch, wird er in eine Richtung verbogen. In beiden
Spannungsteilern wird daher jeweils ein DMS gedehnt, der andere gestaucht, die
Widerstandswerte in den Spannungsteilern ändern sich entsprechend ihrer Lage auf
dem Stab, so dass sich die Potenziale an beiden Anschlüssen des Messgerätes nun
gegensinnig verändern. Die Spannungsdifferenz ist dabei proportional zur
aufgewendeten Kraft, ihr Vorzeichen gibt die Richtung der Kraftwirkung an.
Die Wheatstone - Messbrücke ist in Sensorschaltungen weit verbreitet, weil sie einfach
aufzubauen ist und sehr präzise arbeitet. Durch Verwendung unterschiedlicher
Materialien können ganz unterschiedliche Messgrößen in Spannungen umgewandelt
werden.
Eine anschauliche, interaktive Animation zum Prinzip der Messbrücke finden Sie im
Internet unter: http://www.bipede.de/Downloads/DMS_Voll.swf
(Anmerkung: In dieser Animation müssen Sie auf den schwarzen Punkt klicken und ihn
bewegen, um den virtuellen Biegestab zu verformen)

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 5
MESSGERÄTE
Die meisten Sensoren wandeln eine primäre Messgröße in eine Spannung um. Da
sich im Verlauf eines Experiments in der Regel der Betrag der Messgröße und
damit die Ausgangsspannung des Sensors ändert, ist es zweckmäßig, die
Messung mit einem Gerät vorzunehmen, das nicht nur das momentane
Messsignal, sondern das auch dessen zeitlichen Verlauf sichtbar macht. Bis vor
wenigen Jahren verwendete man dafür ein Oszilloskop, dessen Funktionsweise
Sie in jedem Physikbuch nachlesen können. Stand der Technik sind heutzutage
volldigitale Datenerfassungssysteme, die alle Funktionen eines Oszilloskops
bieten und sich ganz ähnlich bedienen lassen, die zusätzlich aber auch alle
ankommenden Signale digital speichern, so dass die Messwerte auch nach der
Messung zur Verfügung stehen. Im Kurs verwenden wir als Datenerfassungs-
und -analysesystem PowerLab und die Software Chart. Eine Einführung zur
Bedienung der Software erhalten Sie im Kurs.
Abbildung 2: Prinzip der digitalen Datenerfassung mit Powerlab
ERFASSUNG ANALOGER SIGNALE MIT DIGITALER TECHNIK
Die Messdaten, die bei biologischen Messungen anfallen, sind in der Regel
kontinuierliche („analoge“) Signale. Während diese im herkömmlichen
Oszilloskop lediglich verstärkt werden mussten und dann direkt auf die
Ablenkplatten der Vertikalablenkung geschaltet wurden, ist es bei der Erfassung
analoger Signale mit digitalen Geräten notwendig, den Signalverlauf mit Hilfe
eines Analog-Digital Wandlers (Analog-Digital Converter oder kurz: ADC) in eine
Folge von diskreten Daten umzuformen. Die wichtigsten Parameter, über die ein
ADC verfügt, sind die Abtastrate und die Abtasttiefe. Da sie ganz wesentlich die
Qualität der Messdaten bestimmen, wird hier auf die Bedeutung beider Größen
näher eingegangen.

DIE ABTASTRATE
Die Abtastrate oder Abtastfrequenz (auch sampling rate oder Samplerate
genannt) wird in Hz angegeben und bestimmt die zeitliche Auflösung, mit der die
Messdaten erfasst werden. Eine Abtastrate von 44 kHz bedeutet zum Beispiel,
dass jede Sekunde des gemessenen Signals in 44.000 einzelne Datenpunkte
umgewandelt wird.
Je höher die Abtastrate ist, desto besser ist die Qualität der Aufzeichnung in der
zeitlichen Domäne, d.h. höhere Frequenzen können noch ausreichend in einer
späteren Analyse ausgewertet werden. Da die Anzahl der je Zeiteinheit
aufgenommenen Datenpunkte mit der Abtastrate ansteigt, führt eine zu hohe
Abtastrate allerdings auch zu einem höheren Speicherverbrauch, weswegen die
Abtastrate an die zu erwartende Geschwindigkeit angepasst werden sollte, mit
denen sich das zu messende Signal ändert.
Eine Regel für die richtige Wahl der Abtastrate ergibt sich aus dem Nyquist-
Shannon-Theorem. Demnach ist es möglich, den Wellenverlauf eines analogen
Signals vollständig zu rekonstruieren, wenn die Samplerate dem doppelten der
höchsten Frequenz entspricht, die im analogen Signal vorkommt. Will man also
noch Änderungen in einem Signal aufspüren können, die sich innerhalb einer
Millisekunde abspielen, ist eine Samplerate von mindestens 2 kHz erforderlich,
denn 1 ms entspricht der Periodendauer eines Signals mit einer Frequenz von 1
kHz.
DIE ABTASTTIEFE
Die Abtasttiefe oder Bit-Tiefe des Signals bestimmt, wie viele Bits der Rechner
intern verwendet, um den analogen Wert eines einzelnen Samples darzustellen.
Je größer die Abtasttiefe ist, desto geringer ist der minimale
Amplitudenunterschied, der bei der A/D Konversion erkannt wird: Bei einer
Abtasttiefe von 8 Bit wird der analoge Messwert als Zahl zwischen 0 und 255
dargestellt, bei 16 Bit kann er bereits 65536 unterschiedliche Werte annehmen.
Auch für die Abtasttiefe gilt: Je höher sie ist, desto mehr Speicher wird
verbraucht. Allerdings ist die Abtasttiefe in vielen Fällen durch die verwendete
Hardware festgelegt.
Der von Ihnen im Rahmen dieses Kurses verwendete ADC heißt PowerLab 26T
und verfügt neben 4 unabhängigen Eingangskanälen noch über Ausgänge, über
die Signale zur Stimulation (z.B. für den Nerv- oder den Muskelkurs) erzeugt
werden können. Seine Abtasttiefe beträgt 24 Bit, die maximale Samplerate
beträgt 400 kHz, wenn nur ein Kanal verwendet wird. Beim Mehrkanal-Betrieb
müssen sich die Kanäle diese Abtastrate (bzw. die Rechenleistung des ADCs)
„teilen“ – entsprechend sinkt sie beim Einsatz von vier Messkanälen auf 100 kHz
je Kanal ab.
TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 7
PHYSIKALISCHE GRÖßEN UND IHRE EINHEITEN
Im Praktikum - und ganz allgemein in empirischen Experimenten werden Messergebnisse immer in der zur Messgröße passenden Einheit angegeben werden. Auch wenn Sie wegen der Anwendung eines indirekten Messverfahrens
letztlich eine ganz andere Größe bestimmen, Sie also z.B. Spannungsänderungen anstelle von Kräften messen, müssen Sie die Daten für das Protokoll immer in
die Messgröße umrechnen und die richtige Einheit angeben. Oft werden die gemessenen Signale noch verstärkt, was Sie bei der Angabe der Messgröße berücksichtigen müssen. Durch die Verwendung der oben genannten volldigitalen
Messanordnungen wird Ihnen diese Arbeit extrem erleichtert: Sie können bei der Kalibrierung angeben, wie die am ADC ankommenden elektrischen Signale in die
jeweils richtige Messgröße und deren Einheit umgerechnet werden sollen, den Rest erledigt die Software für Sie und Sie erhalten die Messergebnisse automatisch in den von Ihnen vorgegebenen Einheiten und mit den
entsprechenden Umrechnungsfaktoren. Damit dabei allerdings keine Fehler auftreten, müssen Sie sich umso mehr darüber klar sein, welche Messgröße sie
eigentlich erfassen wollen und wie die Messwerte des Sensors in die jeweilige Messgröße umgerechnet werden. Zu Ihrer Erinnerung enthält eine (unvollständige) Auflistung von häufig in der Tierphysiologie vorkommender
Messgrößen und ihrer Einheiten.
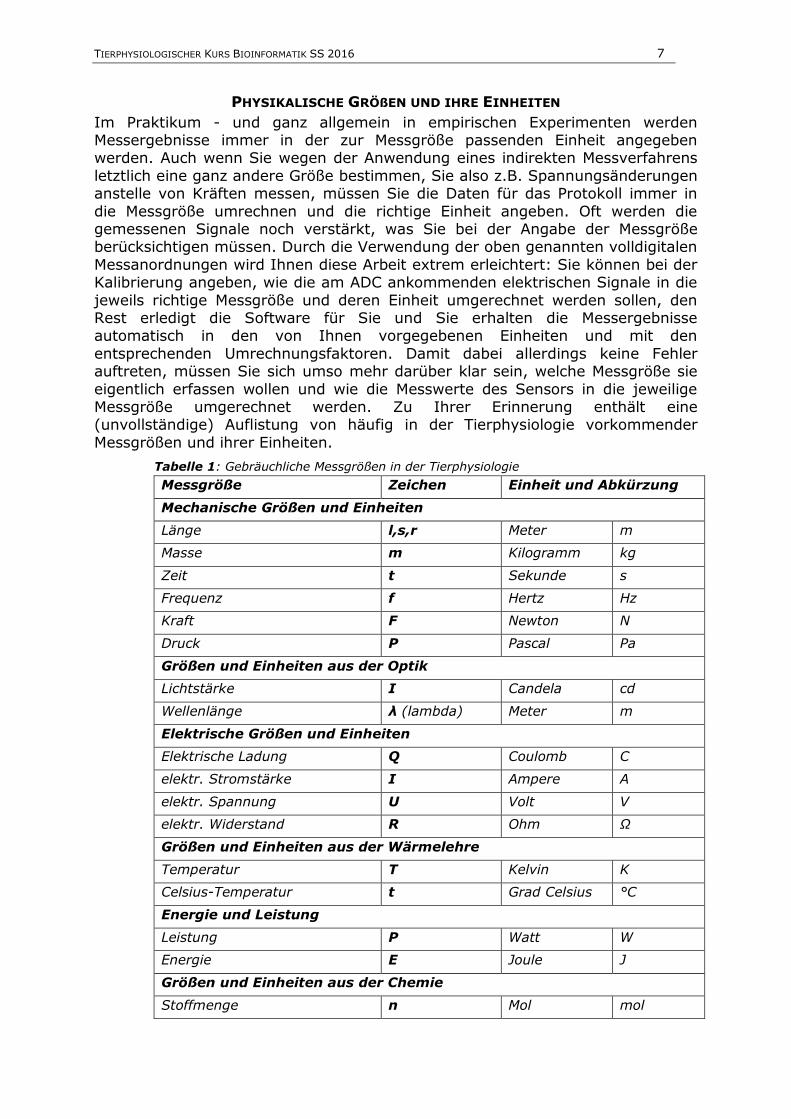
Tabelle 1: Gebräuchliche Messgrößen in der Tierphysiologie
Messgröße Zeichen Einheit und Abkürzung
Mechanische Größen und Einheiten
Länge l,s,r Meter m
Masse m Kilogramm kg
Zeit t Sekunde s
Frequenz f Hertz Hz
Kraft F Newton N
Druck P Pascal Pa
Größen und Einheiten aus der Optik
Lichtstärke I Candela cd
Wellenlänge λ (lambda) Meter m
Elektrische Größen und Einheiten
Elektrische Ladung Q Coulomb C
elektr. Stromstärke I Ampere A
elektr. Spannung U Volt V
elektr. Widerstand R Ohm Ω
Größen und Einheiten aus der Wärmelehre
Temperatur T Kelvin K
Celsius-Temperatur t Grad Celsius °C
Energie und Leistung
Leistung P Watt W
Energie E Joule J
Größen und Einheiten aus der Chemie
Stoffmenge n Mol mol

... EIN PAAR WORTE ZUR HILFSEINHEIT BEL BZW. DEZIBEL
Häufig werden Messwerte in Bel (B) oder Dezibel (dB) angegeben. Das Dezibel
taucht in Tabelle 1 nicht auf, weil es lediglich eine Hilfseinheit ist, die das
Verhältnis zweier Signalpegel beschreibt. Es wird immer dann verwendet, wenn
Messwerte in Relation zu einem Referenzwert gemessen werden – das ist
beispielsweise bei Verstärkungs- oder Dämpfungsfaktoren zweckmäßig. Die
Hörschwelle, die Sie im Kursteil Psychophysik bestimmen werden, ist ein Beispiel
aus dem Kurs, in dem sie in dB messen werden. Ein Bel kennzeichnet den
dekadischen Logarithmus des Verhältnisses zweier gleichartiger Leistungs- bzw.
Energiegrößen P1 und P2, das Dezibel ist einfach der zehnte Teil eines Bels:
Da das dB eine logarithmische Größe ist, sollte man sich wieder ein wenig mit
den Rechenregeln vertraut machen, die beim dekadischen Logarithmus zum
Glück recht einfach sind:
10 dB entsprechen einem Verhältnis des Signals P2 zur Referenz P1 von 10:1.
Und weil
gilt, entspricht ein Signalpegel von -10 dB einem Zehntel des Referenzsignals.
Die Leistung eines Signals mit einem Pegel von 3dB ist deswegen ungefähr
doppelt so groß, wie die Referenz:
Zu beachten ist noch, dass Leistungs- bzw. Energiegrößen proportional zu den
Quadraten der einwirkenden Effektivwerte der betreffenden Feldgrößen sind (in
der Akustik wäre dies der Schalldruck; in der Elektrizität die Stromstärke oder
auch die Spannung).
Das trifft auf die von uns gemessenen Messgrößen (Spannungspegel oder
Schalldruck) zu; daher erweitern wir die Gleichung und formen um:
... aber dazu mehr im Psychophysik-Teil des Praktikums ...

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 9
DARSTELLUNG VON MESSERGEBNISSEN IN EINEM PROTOKOLL
Messwerte werden in einem Protokoll übersichtlich in Diagrammen oder Tabellen
dargestellt und zusätzlich im Textteil des Ergebnisteils verbal beschrieben.
Bei der Verwendung von Diagrammen sind folgende Dinge zu beachten:
Ein Diagramm kann nur dann vollständig sein, wenn an den Achsen die
richtigen Messgrößen angegeben sind (das gilt insbesondere auch in
Klausuren!). Hierbei können die Namen der Messgrößen, ihre gängigen
Formelzeichen (vergl. Tabelle 1) oder beides verwendet werden.
Bei Diagrammen, die gemessene Daten enthalten, sind die Achsen
zusätzlich mit Intervallmarken und Zahlenwerten zu beschriften. Die
Einheiten der Messgrößen müssen dann (in runden oder eckigen
Klammern) hinter die Bezeichnung der Messdaten gesetzt werden (vgl.
Abbildung 1). Diagramme, die dagegen lediglich schematisch einen
Zusammenhang zwischen zwei Größen veranschaulichen sollen, die aber
nicht auf gemessenen Daten beruhen, müssen keine Einheiten und
Zahlenwerte an den Achsen enthalten - außer, dieser Zahlenwert ist für
die vollständige Darstellung des Zusammenhangs der dargestellten Größen
notwendig (dies gilt z.B. oft für den Nullpunkt eines Diagramms).
Im Allgemeinen werden die Messgrößen so auf die Achsen des Diagramms
übertragen, dass „y von x abhängt“ und nicht umgekehrt - bei Dia-
grammen, die eine zeitliche Abfolge von Messwerten enthalten, wird daher
normalerweise die Zeit entlang der x-Achse dargestellt.
Vorsicht bei der Erstellung von Diagrammen in Excel oder ähnlichen
Programmen! Häufig ist die Funktion „Kurve glätten“ in der Grund-
einstellung des Grafik-Editors aktiviert, was zu „runderen“ Kurven-
verläufen führt, die zwar „harmonischer“ aussehen, die aber nicht die
tatsächlich gemessenen Werte darstellen. Die Verwendung derartiger
Funktionen sollte vermieden werden, oder, wenn überhaupt, nur unter
Angabe eines vernünftigen Grundes geschehen (meistens gibt es dafür
keinen ...)
Diagramme und Tabellen müssen Legenden enthalten, ähnlich denen, die Sie
unter den Abbildungen in diesem Kapitel finden. Eine Abbildungslegende soll den
Inhalt der Abbildung kurz beschreiben, ohne auf die Details einzugehen, diese
sollen im Text des Ergebnisteils beschrieben werden. Die in einer Abbildung
verwendeten Symbole und ihre Bedeutung müssen dagegen in der
Abbildungslegende angegeben sein.
... Und noch ein Hinweis: Die meisten Textverarbeitungsprogramme bieten
Funktionen an, mit denen Abbildungen unmittelbar nach dem Einfügen in das
Dokument beschriftet werden können. Bei der Erstellung der Abbildungs-
beschriftung vergeben diese Programme der Abbildung oder Tabelle auch gleich
eine laufende Nummer, auf die Sie aus dem Text quer verweisen können. Wenn
Sie danach noch weitere Abbildungen auf dieselbe Weise einfügen, so

aktualisieren diese Programme automatisch die Nummerierung der Abbildungen
und auch die Nummern in den entsprechenden Querverweisen im Text. Auf jede
Abbildung oder Tabelle muss sich übrigens mindestens ein Querverweis aus dem
Text beziehen.
Machen Sie sich bei der Erstellung Ihrer Protokolle am besten gleich mit den
Möglichkeiten vertraut, die Ihr bevorzugtes Textverarbeitungsprogramm bietet,
sie werden diese Funktionen später, z.B. bei der Abfassung Ihrer Bachelor- oder
Masterarbeit zu schätzen wissen
Möglichkeiten für Bioinformatik-Studierende in den
Neurowissenschaften
Für die Bioinformatik-Studierenden existieren zahlreiche Möglichkeiten, sich
näher mit Neurobiologie oder Neurowissenschaften an der Universität Tübingen
zu beschäftigen. Es sind hier an erster Stelle das Institut für Neurobiologie mit
den Professuren Neuroethologie, Tierphysiologie und Kognitive
Neurowissenschaften, das Hertie-Institut für klinische Hirnforschung (HIH) und
das Werner-Reichardt Zentrum für Integrative Neurowissenschaften (CIN) zu
nennen. Diese Einrichtungen bieten zahlreiche Lehrveranstaltungen (Vorlesungen
und Praktika) an, stellen Themen für das Anfertigen einer BSc oder MSc-Arbeit
zur Verfügung und bieten unter Umständen auch Möglichkeiten für eine
Dissertation.

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 11
1. N E R V E N P H Y S I O L O G I E
EINFÜHRENDE BEMERKUNGEN
Nervensysteme sind aus einzelnen Nervenzellen
aufgebaut. Im Gegensatz zu vielen anderen
Körperzellen sind Nervenzellen erregbar, d. h. sie
antworten bei Depolarisation des Membranpotentials nach Überschreiten der
Feuerschwelle mit Aktionspotentialen, die im Axon (Nervenfaser) fortgeleitet
werden. Solche Depolarisationen werden in der biologischen Situation bei
Nervenzellen durch postsynaptische Potentiale im Bereich der Synapsen, bzw. bei
Sinnesrezeptoren durch die durch physikalische oder chemische Reize erzeugten
Rezeptorpotenziale ausgelöst. Im physiologischen Experiment können sie durch
elektrische Reize ausgelöst werden.
Einige Eigenschaften der Erregungsvorgänge bei Nervenzellen sollen in diesem
Versuchsabschnitt experimentell untersucht werden. Leider können Sie nicht -
was didaktisch besonders günstig wäre - am einzelnen Axon mit intrazellulärer
Ableittechnik arbeiten. Aus methodischen Gründen müssen wir einen Nerv mit
vielen Axonen (N. ischiadicus des Krallenfrosches) verwenden und extrazelluläre
Summenaktionspotenziale ableiten. Das Summenaktionspotenzial ist die
Resultierende aus allen extrazellulär messbaren Aktionspotenzialen der einzelnen
Axone in einem Nerv.
THEORETISCHE VORAUSSETZUNGEN
Folgende Kenntnisse aus dem Modul Tierphysiologie I und aus der Wirbel-
tieranatomie sind für eine erfolgreiche Durchführung des Kurses unerlässlich:
Neuroanatomie: Aufbau eines motorischen, eines sensorischen und eines
gemischten Nervs (afferente, efferente Fasern), Neuron, Soma, Dendrit,
Axon, Kollaterale, Synapse, myelinisierte und unmyelinisierte Axone,
Ranvier'scher Schnürring.
Ruhepotential: Intrazelluläre Potentialmessung, Ionenverteilung an der
Axonmembran, Na+- und K+-Gleichgewichtspotential, Leitfähigkeit
(Permeabilität) für Na+ und K+, Ionenpumpen, elektrisches Ersatzschaltbild
für die Entstehung des Ruhepotentials, Nernst- und Goldmann-Gleichung,
Veränderung des Ruhepotentials bei Änderung der Na+- und K+-
Konzentration.
Passive elektrische Eigenschaften der Nervenzelle: Ruhemembranwider-
stand, Membrankapazität, intrazellulärer Längswiderstand, Elektrotonische
(passive) Ausbreitung, Membranzeitkonstante, Membranlängskonstante,
lokale Antwort.
Na+/K+-Aktionspotential (AP): Zeitverlauf des APs, Erregungsschwelle,
Alles-oder-Nichts Regel, Permeabilität für Na+ und K+ während des APs,

schnelles Na+-System mit Inaktivierung, Hodgkin-Huxley Zyklus, relative
und absolute Refraktärzeit, Veränderung des APs bei Änderung der Na+-
und K+-Konzentration, Unterschied zwischen intrazellulär und extrazellulär
abgeleiteten APs.
Fortleitung des APs: Unterschied zwischen passiver und aktiver Erregungs-
fortleitung, Erregungsleitung an myelinisierten und unmyelinisierten
Axonen, Abhängigkeit der Leitungsgeschwindigkeit vom Axondurchmesser.
Summenaktionspotential (SAP): Unterschied zwischen dem AP einer Einzel-
faser und dem SAP eines Nervs, Form und Ableitung eines diphasischen
SAP, Beziehung zwischen Amplitude des SAP und Reizintensität.
Methodische Kenntnisse, die im Praktikum erworben werden sollen:
Verkabelung einer elektrophysiologischen Messapparatur, extrazelluläre Ab-
leitung eines Summenpotentials, Auswertung und Aufbereitung
elektrophysiologischer Daten.
LITERATUR
KANDEL/SCHWARZ/JESSEL: Neurowissenschaften, Spektrum
Hervorragend verständliche Zusammenfassung der gesamten
Neurowissenschaften mit Betonung der Verhältnisse beim Säuger.
SCHMIDT/THEWS/LANG: Einführung in die Physiologie des Menschen, Springer. -
- Hervorragendes Lehrbuch für die gesamte allgemeine Physiologie und die
Physiologie des Menschen. Leider ohne vergleichende tierphysiologische
Aspekte.
MOYES/SCHULTE: Principles of Animal Physiology, Pearson -Gibt guten Überblick.
Aufgrund der Kürze manchmal etwas schwer verständlich.
ECKERT/RANDALL: Animal Physiology, Freeman. -Gutes Lehrbuch für allgemeine
und vergleichende Physiologie. (Deutsche Übersetzung: Thieme)
MÜLLER: Tier- und Humanphysiologie, Springer -Gutes Lehrbuch für allgemeine
und vergleichende Physiologie
Weiterhin empfehlenswert (für Spezialisten):
ZIGMOND/BLOOM/LANDIS/ROBERTS/SQUIRE: Fundamental Neuroscience.
Modernes Standardwerk für angehende Neurobiologen mit Schwerpunkt
Physiologie und zelluläre Mechanismen.
KANDEL/SCHWARZ/JESSEL: Principles of Neural Sciences, 4th Edition. (2000)
Ebenfalls ein Standardwerk mit Schwerpunkt Säuger/Mensch. Achtung -
ältere Auflagen sind wirklich veraltet!
PRÄPARIERBESTECK

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 13
1 spitze Pinzette
1 feine spitze Schere
1 größere Schere
1 feine Schere
VERSUCHE
1. Passive Eigenschaften der Nervenzellmembran
Modell der passiven Eigenschaften der Nervenmembran
Dieser Versuchsteil dient dem Kennenlernen der Ableitmethode und der Messung
von passiven Membraneigenschaften einer Nervenzelle.
Die Messungen werden an einem Modell der Nervenzellmembran (Kette von RC-
Gliedern) durchgeführt. Jedes Glied dieser Kette repräsentiert einen kleinen
Membranabschnitt mit Membranwiderstand und Membrankapazität. Die einzelnen
Glieder sind durch den Innenwiderstand der "Intrazellulärflüssigkeit" verbunden.
Der Außenwiderstand der Extrazellulärflüssigkeit wird als sehr klein angenom-
men. An diesem Modell sollen Sie die Membranzeitkonstante und die Membran-
längskonstante messen.
2. Versuchsdurchführung
Öffnen Sie in Chart den File „Membranmodell.adiset“. Verbinden Sie den
Stimulusausgang des Powerlabs mit dem Eingang des Membranmodells.
Verbinden Sie außerdem den Stimulusausgang mit dem Kanal 1 des Powerlabs.
Verbinden Sie die Ableitelektroden mit Kanal 2 des Powerlabs. Stellen Sie in
Chart im Stimulator Panel eine Reizamplitude von 5 V und eine Reizdauer von
200 ms ein.
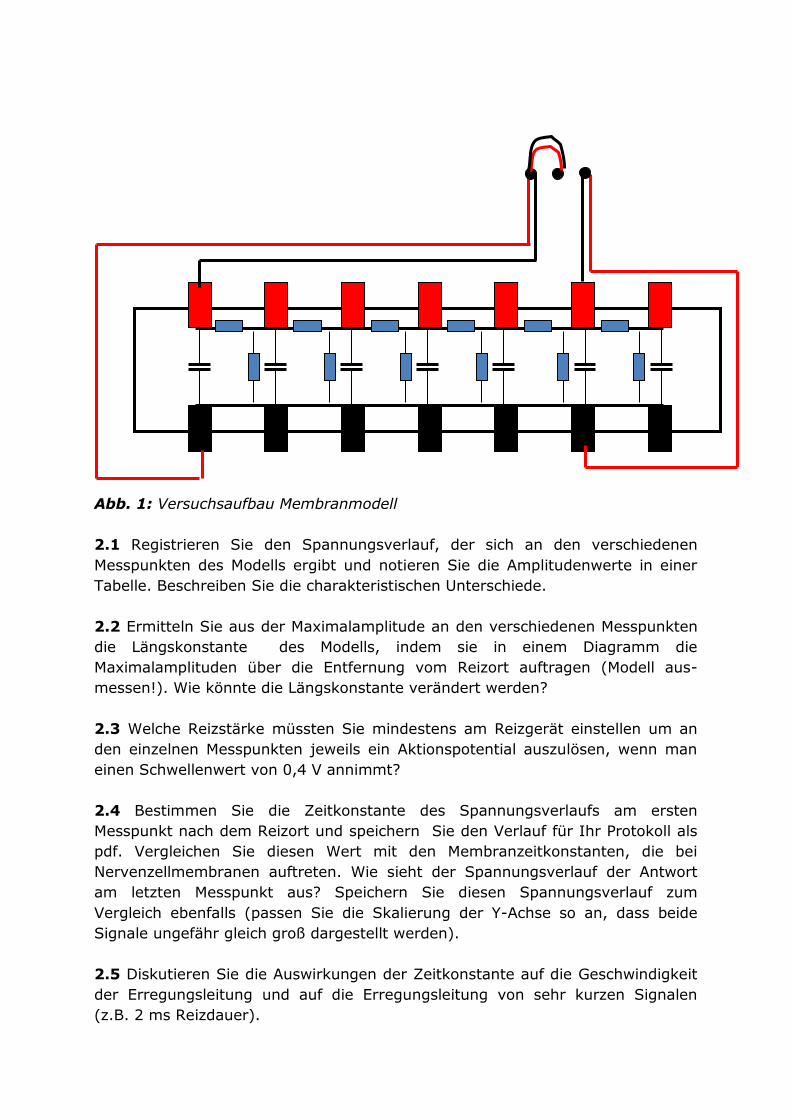
Abb. 1: Versuchsaufbau Membranmodell
2.1 Registrieren Sie den Spannungsverlauf, der sich an den verschiedenen
Messpunkten des Modells ergibt und notieren Sie die Amplitudenwerte in einer
Tabelle. Beschreiben Sie die charakteristischen Unterschiede.
2.2 Ermitteln Sie aus der Maximalamplitude an den verschiedenen Messpunkten
die Längskonstante des Modells, indem sie in einem Diagramm die
Maximalamplituden über die Entfernung vom Reizort auftragen (Modell aus-
messen!). Wie könnte die Längskonstante verändert werden?
2.3 Welche Reizstärke müssten Sie mindestens am Reizgerät einstellen um an
den einzelnen Messpunkten jeweils ein Aktionspotential auszulösen, wenn man
einen Schwellenwert von 0,4 V annimmt?
2.4 Bestimmen Sie die Zeitkonstante des Spannungsverlaufs am ersten
Messpunkt nach dem Reizort und speichern Sie den Verlauf für Ihr Protokoll als
pdf. Vergleichen Sie diesen Wert mit den Membranzeitkonstanten, die bei
Nervenzellmembranen auftreten. Wie sieht der Spannungsverlauf der Antwort
am letzten Messpunkt aus? Speichern Sie diesen Spannungsverlauf zum
Vergleich ebenfalls (passen Sie die Skalierung der Y-Achse so an, dass beide
Signale ungefähr gleich groß dargestellt werden).
2.5 Diskutieren Sie die Auswirkungen der Zeitkonstante auf die Geschwindigkeit
der Erregungsleitung und auf die Erregungsleitung von sehr kurzen Signalen
(z.B. 2 ms Reizdauer).

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 15
ABLEITUNG EINES SUMMENAKTIONSPOTENTIALS (SAP)
Präparation des Nervus ischiadicus
- wird vom Kursbetreuer durchgeführt
Ein Frosch wird mit einer Guillotine dekapitiert und sein Rückenmark durch
Einführen einer Sonde in den Rückenmarkskanal zerstört. Anschließend wird das
Präparat enthäutet und mit Ringerlösung abgespült. Nach diesem Arbeitsgang
sollten auch alle Instrumente und die Hände sorgfältig gereinigt werden, um das
giftige Hautsekret zu entfernen.
Die Bauchhöhle des Frosches wird geöffnet und die Eingeweide entnommen.
Die beiden nun freiliegenden Ischiadicus-Nerven werden mit einem Bindfaden
abgebunden. Dazu wird vorsichtig eine kleine Pinzette unter einen Ischiadicus
kurz hinter seinem Austritt aus dem Wirbelkanal durchgeschoben und ein mit
Ringerlösung angefeuchteter Zwirnfaden unter dem Nerv durchgezogen und fest
um den Nerv geknotet. Das kürzere Ende des Fadens wird dicht am Knoten
abgeschnitten, das andere Ende dient später als Haltegriff. Die Nerven werden
nun proximal vom Knoten mit einer kleinen Schere durchtrennt und bis zum
Eintritt in den Oberschenkel freipräpariert. Nun wird das Becken durch einen
Schnitt in der Medianebene halbiert. Jede Gruppe führt nun an einem der beiden
Froschbeine die Präparation fort.
- weitere Präparation wird von den Praktikanten durchgeführt
Ein isolierter Nerv ist ein lebendes und sehr empfindliches Gewebe; er sollte
daher schonend behandelt werden. Dehnen Sie oder fassen Sie den Nerv niemals
mit den Fingern oder einer Pinzette an.
Die dorsale Oberschenkelmuskulatur wird nun mit den Daumen auseinan-
dergedrückt bis der Nerv sichtbar wird. Präparieren Sie den Nerv über die ganze
Länge des Oberschenkels von Blutgefäßen und Bindegewebe frei. Beim Anheben
des Nervs mit dem Haltefaden (nicht dehnen!) können seitlich abgehende
Nervenäste mit einer kleinen Schere durchtrennt werden. Durchtrennen Sie
Kollaterale direkt am Nerv, indem Sie vom Nerv weg schneiden. Beim
Abschneiden sollte die Schere vom Nerv weg weisen. Entfernen Sie mit einer
Pinzette alle restlichen Bindegewebskontakte.
Wenn Sie am Kniegelenk angelangt sind, durchtrennen Sie den Nerv und legen
Sie ihn in eine Petrischale mit Ringerlösung. Den Rest des Froschbeines legen Sie
bitte in eine Präparierschale, decken es mit ringer-feuchtem Filtrierpapier ab und
stellen es in den Kühlschrank. Eventuell kann es von den Studenten des "Muskel-
Versuches" noch als Ersatzpräparat verwendet werden.
Versuchsaufbau Ableitapparatur
Verkabeln Sie den Versuchsaufbau. Eine nahezu detailgetreue Wiedergabe der
Verschaltung finden Sie in der folgenden Abb.1 "Versuchsaufbau Nerv".
Verbinden Sie dabei die Reizelektroden mit der Ableitkammer so, dass die
Kathode (schwarze Buchse) benachbart zu den Ableitelektroden liegt (warum?).

Abb. 2: Versuchsaufbau Nerv
3. Messung des Reizartefakts
Öffnen Sie in Chart den File „Reizamplitude.adiset“. Wählen Sie im Stimulator
Panel eine Reizamplitude von ca.3 V und eine Reizdauer von 200µs. Legen Sie
einen mit Ringerlösung befeuchteten Faden über die Elektroden der Ableit-
kammer und reizen Sie mit Einzelreizen. Speichern Sie das abgeleitete Signal für
Ihr Protokoll. Wie lässt sich dieses erklären?
4. Ableitung eines fortgeleiteten diphasischen Summenaktionspotentials
bei unterschiedlichen Reizstärken
Ein Summenaktionspotential (SAP) entsteht bei synchroner Erregung mehrerer
oder sämtlicher Axone eines Nervs. Die Ableitung erfolgt extrazellulär. Gemessen
wird die Summe aller extrazellulär abgeleiteten Aktionspotenziale der einzelnen
Axone im Nerv. Der N. ischiaticus setzt sich aus unterschiedlichen Fasertypen
zusammen. Da diese unterschiedliche Leitungsgeschwindigkeiten besitzen, hängt
die Form des SAPs von Abstand zwischen Reiz- und Ableitelektroden, als auch
vom Abstand zwischen den Ableitelektroden ab. Die Amplitude des SAPs hängt
von der Anzahl und Dicke der erregten Axone ab. Je höher die Reizamplitude,
desto mehr Axone werden erregt und desto höher ist die gemessene Amplitude

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 17
des SAPs. Bei der Reizamplitude unterscheidet man zwischen der Schwellen-
reizstärke (kleinste Reizamplitude, die eben noch ein messbares SAP auslöst)
und der Maximalreizstärke (Reizamplitude, ab der eine weitere Vergrößerung
keine Zunahme der SAP-Amplitude bewirkt).
Versuchsdurchführung und Auswertung:
Bevor der Nerv jeweils für eine Messreihe in die Ableitkammer quer über die
Reiz- und Messelektroden gelegt wird, müssen Sie sich über die gesamte
Aufgabe klar geworden sein und alle Einstellungen richtig vorgenommmen
haben. Erst dann wird der Nerv platziert, eine Messreihe zügig durchgemessen,
und dann der Nerv in die Ringerlösung zurückgelegt. Zwischen zwei Messreihen –
jedoch nie innerhalb einer Messreihe – kann der Nerv auch mit Ringerlösung
beträufelt werden. Das Beträufeln mit Ringerlösung, das Bewegen des Nervs auf
den Elektroden, sowie das Antrocknen des Nervs bei langen Messzeiten
verändern die Ableitbedingungen und somit die Messergebnisse innerhalb einer
Messreihe!
4.1 Darstellung eines SAPs:
Wählen Sie im Stimulator Panel eine Reizamplitude von ca.3 V. Platzieren Sie
nun das Präparat und stellen Sie ein typisches SAP dar, das Sie für Ihr Protokoll
abspeichern. Verändern Sie nun den Abstand zwischen den Reiz- und den
Ableitelektroden sowie zwischen den Ableitelektroden. Wie verändern sich die
Form und die Amplitude des SAPs? Erklären Sie Ihre Beobachtungen.
4.2 Abhängigkeit der SAP Amplitude von der Reizstärke:
Stecken Sie nun die Ableitelektroden reizortnah in die Ableitkammer, stellen Sie
die Reizamplitude auf 10 mV und erhöhen schrittweise langsam die
Reizamplitude. Messen Sie die Amplitude des SAPs in Abhängigkeit von der
Reizamplitude und tragen Sie die Werte in eine Tabelle ein. Achtung: der
Differenzverstärker verstärkt die Signale um den Faktor 100. Erstellen Sie ein
Diagramm über die Zunahme der SAP-Amplitude in Abhängigkeit von der
Reizamplitude. Bestimmen sie die Minimal- und Maximalreizstärke.
5. Bestimmung der Geschwindigkeit der Erregungsleitung
In diesem Experiment sollen Sie feststellen, wie groß die Geschwindigkeit ist, mit
der Aktionspotentiale im Froschnerv weitergeleitet werden.
Das Prinzip der Geschwindigkeitsmessung besteht darin, dass das vom Reiz
ausgelöste SAP einmal nahe am Reizort und einmal in einem weiteren Abstand
vom Reizort registriert wird. Aus dem Abstand zwischen den beiden
Ableitelektrodenpaaren (s) und dem ermittelten Zeitunterschied (∆t) zwischen
den abgeleiteten SAPs kann die Leitungsgeschwindigkeit (v) errechnet werden
(v=Δs/Δt). Die Reizung sollte mit der Maximalreizstärke erfolgen.

Durchführung und Auswertung:
5.1 Leiten Sie mit einer Serie von Einzelreizen erst das SAP an reiznahen
Ableitelektroden ab. Stecken Sie die Ableitelektroden in eine reizfernere Position
um und wiederholen Sie die Messung. Die Elektrodenpaare sollten bei diesem
Versuch möglichst weit auseinander stehen, um die Länge des Nervs maximal zu
nutzen.
5.2 Bestimmen Sie die Zeitdifferenz zwischen den beiden SAP-Gipfeln und den
Abstand zwischen den beiden Ableitelektrodenpaaren. Errechnen Sie daraus die
Leitungsgeschwindigkeit in m/s.
5.3 Vergleichen Sie Ihre Werte mit den Literaturwerten für die verschiedenen
Fasertypen des Froschnervs.
6. Bestimmung der Refraktärzeit beim Froschnerv
Die Refraktärzeit eines Nervs ist die Zeitspanne, in der er während und nach
einer Erregung überhaupt nicht (absolute Refraktärzeit) oder aber nur mit
höheren Reizamplituden (relative Refraktärzeit) erneut erregt werden kann bzw.
in der bei gleicher Reizamplitude die Amplitude des zweiten SAPs kleiner ist. Zum
Nachweis dieses Phänomens werden zwei Reize benötigt, deren zeitlicher
Abstand variiert werden kann. Mit dem zweiten Reiz wird das refraktäre
Verhalten des Nervs nach dem ersten Reiz bestimmt.
Durchführung und Auswertung:
6.1 Stecken Sie die Ableitelektroden in eine reizferne Position. Öffnen Sie in
Chart die Datei „Refraktärzeit.adiset“. Wählen Sie eine Reizstärke nahe der
Maximalreizstärke. Beginnen Sie bei einem Reizabstand von 5 ms und reduzieren
Sie diesen kontinuierlich. Registrieren Sie beide SAP-Antworten auf den
Doppelreiz und messen Sie die Amplituden des zweiten SAPs. Tragen Sie die
Werte in Abhängigkeit vom Doppelreizabstand in eine Tabelle ein.
6.2 Speichern sie Sie einige repräsentative Messungen für Ihr Protokoll.
6.3 Zeichnen Sie ein Diagramm der Amplitude des zweiten SAPs in Abhängigkeit
vom Doppelreizabstand. Bestimmen Sie aus dieser Messreihe die absolute und
relative Refraktärzeit.
6.4 Überlegen Sie, welche maximale Reizfrequenz der vorliegende Froschnerv
ohne Ausfall von Aktionspotentialen beantworten kann. Bedenken Sie, wie sich
bei einem SAP der Ausfall von Aktionspotentialen einzelner Axone äußert.

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 19
7. Unterbrechung der Erregungsleitung
Ein diphasisches SAP kommt durch Wandern der Erregungswelle entlang der
Axone über zwei Ableitelektroden hinweg zustande. Zuerst wird die erste
Elektrode und dann die zweite Elektrode negativ gegenüber der jeweils anderen.
Wird die Erregungsleitung zwischen den beiden Ableitelektroden unterbrochen,
sollte das diphasische SAP in ein monophasisches SAP umgewandelt werden.
Durchführung und Auswertung:
7.1 Öffnen Sie wieder die Datei „Reizamplitude. adiset“. Reizen Sie wieder mit
einer Amplitude von 3 V und leiten Sie zuerst ein diphasisches SAP ab.
Quetschen Sie dann den Nerv zwischen den beiden Ableitelektroden ab, ohne
dessen Lage zu verändern. Leiten Sie erneut ein SAP ab. Speichern Sie beide
Ableitungen für Ihr Protokoll
7.2 Welche Unterschiede bestehen zwischen einem diphasischen und einem
monophasischen SAP?
Wie lässt sich die Form des diphasischen SAP aus den monophasischen SAPs
erklären?
Bestimmen Sie die Dauer des monophasichen und des diphasischen SAPs.
Wodurch wird die Dauer dieser Potentiale beeinflusst?
Überprüfen Sie, ob das SAP mehrere Gipfel (Schultern) aufweist, und versuchen
Sie solche Gipfel zu erklären.
8. Leitungsanästhesie am peripheren Nerv
Sprechen Sie sich untereinander so ab, dass eine Teilgruppe mit ihrem Präparat
Versuch 8, die andere Versuch 9 durchführt. Vergleichen Sie danach Ihre
Ergebnisse.
Die meisten von Ihnen haben schon einmal die angenehme Wirkung einer
örtlichen Betäubung (Lokalanästhesie) verspürt. Die dabei verwendeten
Lokalanästhetika sind Medikamente, die eine reversible Blockade der
Nervenleitung bewirken. Sie wirken nicht schlagartig, sondern es wird eine
gewisse Zeit benötigt, bis ihre Wirkung eintritt. Dies liegt daran, dass die
verschiedenen Fasern eines gemischten Nervenstammes nicht alle zur selben
Zeit vom Lokalanästhetikum erfasst werden. Seit den dreißiger Jahren wird als
Lokalanästhetikum das von Ihnen hier im Praktikum benutzte Xylocain
(Wirkstoff: Lidocain) verwendet, ein Abkömmling des Kokains.

Durchführung und Auswertung:
8.1 Öffnen Sie die Datei „Reizamplitude.adiset“. Wählen Sie als Reizamplitude
wieder eine Wert um die Maximalreizamplitude und machen Sie eine
Kontrollmessung.
8.2 Besprühen Sie den Nerv in der Ableitkamammer im Bereich zwischen Reiz -
und Ableitelektroden mit Xylocain und beginnen sie dann sofort mit den
Messungen. Reizen Sie das Präparat mit Einzelreizen von gleicher Reizamplitude
im Abstand von 30 s und messen Sie die SAP-Amplitude aus. Setzen Sie die
Messung fort, bis kein SAP mehr ausgelöst werden kann. Falls sich nach 2 min.
noch kein Amplitudenabfall des SAPs zeigt, sprühen Sie erneut Xylocain auf und
wiederholen die Messung. Speichern Sie die Messungen für Ihr Protokoll.
8.3 Erstellen Sie ein Diagramm der SAP-Amplitude als Funktion der Zeit vor und
während der Xylocaineinwirkung. Worauf beruht die Wirkung von Xylocain? Wie
lange dauert es bis Xylocain den Nerv vollständig betäubt hat?
9. Betäubung eines Nervs mit Äther
Durchführung und Auswertung:
9.1 Öffnen Sie die Datei „Reizamplitude.adiset“ . Wählen Sie als Reizamplitude
wieder eine Wert um die Maximalreizamplitude und machen Sie eine
Kontrollmessung.
9.2 Legen Sie ein Stück Filterpapier zwischen Kammer und Abdeckplatte, das mit
etwas Äther getränkt wurde (feucht, aber nicht tropfend). Beginnen Sie sofort
mit der Messung. Reizen Sie das Präparat mit Einzelreizen von gleicher
Reizamplitude im Abstand von 10 s.. Setzen Sie die Messung fort, bis kein SAP
mehr ausgelöst werden kann. Entfernen Sie anschließend das Filterpapier und
legen Sie es unter den Abzug.
Warten Sie nun einige Minuten. Meistens kann man, im Gegensatz zur Lidocain-
Betäubung, eine Erholung des Nervs beobachten (Warum?). Messen Sie die SAP-
Amplituden aus und speichern Sie die Registrierungen für Ihr Protokoll.
9.2 Zeichnen Sie ein Diagramm der SAP-Amplitude als Funktion der Zeit vor und
während der Äthereinwirkung. Wie lange dauert es, bis Äther den Nerv voll-
ständig betäubt hat?
9.3 Worauf beruht die Wirkung von Äther?

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 21
ERFOLGSKONTROLLE
Nach diesem Praktikumsteil sollten Sie in der Lage sein:
den Aufbau eines peripheren Nervs am Beispiel des Nervus ischiadicus zu
beschreiben,
die nacheinander ablaufenden Vorgänge von der synaptischen Reizung
eines Motoneurons, passiver Fortleitung im Dendriten, Auslösen eines
Aktionspotentials, aktiver Fortleitung im (myelinisierten) Axon, bis hin zur
synaptischen Übertragung an der motorischen Endplatte zu erläutern,
Den Unterschiede zwischen aktiver und passiver Erregungsleitung
erläutern können
die Bedeutung der Längs- und Zeitkonstante bei der elektrotonischen
Erregungsausbreitung zu erklären,
den Zusammenhang zwischen Axondurchmesser und Leitungsge-
schwindigkeit zu beschreiben,
zu erläutern, warum die Form und die Amplitude von den Ableit-
bedingungen abhängt
den Unterschied zwischen einem AP und einem SAP zu erklären,
den Entstehungsmechanismus des diphasischen SAPs zu erklären,
die Größenordnung der Amplitude eines vom Froschnerv abgeleiteten SAPs
anzugeben,
die ungefähre Dauer eines mono- und diphasischen SAPs anzugeben,
eine Methode zur Umwandlung eines diphasischen in ein monophasisches
SAP erklären zu können,
anzugeben, wie man die Nervenleitungsgeschwindigkeit bestimmt,
die Leitungsgeschwindigkeit der schnellen Fasern bei Frosch und Mensch
anzugeben,
zu erklären, weshalb das SAP in der relativen Refraktärphase kleiner wird,
die Dauer der absoluten und relativen Refraktärphase am Froschnerv zu
nennen,
die Ursache der relativen und absoluten Refraktärzeit zu nennen,
den Versuchsaufbau zu skizzieren,
die Versuchsapparatur selbständig zu verschalten
und die in Ihrem Protokoll aufgeführten Kurven zu skizzieren.

2. M U S K E L P H Y S I O L O G I E
EINFÜHRENDE BEMERKUNGEN
In diesem Versuch sollen grundlegende Eigenschaften von zwei
Vertebratenmuskeln, dem quergestreiften Skelettmuskel von Frosch und Mensch
und dem Herzmuskel des Frosches, besprochen und experimentell erarbeitet
werden. Im Mittelpunkt der theoretischen Vorbereitung auf den Versuch steht die
vergleichende Betrachtung der physiologischen Eigenschaften bei der
Erregungsentstehung und Erregungsfortleitung der beiden Muskelarten. Im
praktischen Teil dieses Versuchs sollen bei der Skelettmuskulatur die
elektromechanischen Eigenschaften anhand des Kontraktionsverhaltens von
Krallenfrosch-Präparaten sowie durch die Registrierung des Elektromyogramms
(EMG) eines menschlichen Handmuskels untersucht werden. Am Vertebraten-
Herz sollen Versuche zur Pharmakologie der Regulation des Herzschlages
exemplarisch durch die Applikation verschiedener Pharmaka auf ein
Krallenfrosch-Präparat untersucht werden
THEORETISCHE VORAUSSETZUNGEN
Folgende Kenntnisse aus der Vorlesung "Einführung in die Tierphysiologie" und
aus der Wirbeltieranatomie sind für eine erfolgreiche Durchführung des Kurses
unerlässlich:
Anatomie: Hierarchischer Aufbau von der organischen zur subzellulären Ebene.
Quergestreifte Muskelfasern mit motorischer Endplatte, Sarkolemm,
Myofibrillen, transversale Tubuli (T-System), longitudinale Tubuli
(sarkoplasmatisches Retikulum), Sarkomer, Z-Scheibe, A- und I-Bande, H-
Zone, Actin, Myosin, Tropomyosin, Troponin.
Neuromuskuläre Endplatte: Bau, Endplattenpotenzial, Transmitterwirkung,
Abbau des Acetylcholins, synaptische Latenz, neuromuskuläre Blockade,
Elektroplaques (Elektrozyten), Entstehung und Weiterleitung des
Aktionspotenzials.
Molekulare Mechanismen der Kontraktion: elektromechanische Kopplung,
Muskelaktionspotenzial, Verkürzung der Sarkomere, Querbrücken, Rolle der
Ca++-Ionen, ATP, Calciumpumpe, Erregungsleitung, neurogener Tonus,
myogener Tonus, oxidativer und glykolytischer Energiegeumsatz, muskuläre
Ermüdung.
Wirkung elektrischer Reize: Elektrotonus, anodische und kathodische
Reizung, Depolarisation, Hyperpolarisation.
Elektromyogramm (EMG): Entstehung, Motoneuron, motorische Einheit,
Muskelfasertypen, Rekrutierung, Adduktion, Abduktion, Flexion, Extension.
Mechanik: Regulation der Muskelkraft, Summation und Rekrutierung, un-

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 23
vollständiger und vollständiger Tetanus, isometrische, isotonische,
auxotonische Kontraktion.
Anatomie: Kreislaufsystem bei Amphibien (Frosch) und bei Säugern (Mensch),
Bau von Amphibien- und Säugerherz, sympathische und parasympathische
Innervation des Säugerherzens, Morphologie der Herzmuskelzellen
Erregungsentstehung und –weiterleitung: Autorhythmische Zentren,
Mechanismus und Strukturen der Erregungsentstehung und –weiterleitung:
Sinusknoten, Atrioventrikularknoten. Erregungsleitende Strukturen, Unterschied
zwischen myogenem und neurogenem Herzen, Form der Aktionspotenziale der
Schrittmacherzellen und des Arbeitsmyokards, Refraktärzeit, Sympathikus- und
Parasympathikuseinwirkung, Pharmakologie der Herz-Innervation, elektro-
mechanische Kopplung, Nicht-Tetanisierbarkeit des Herzens, EKG lesen und
interpretieren.
Mechanik der Herzkontraktion: Systole, Diastole, Herzklappentätigkeit,
Druck-Volumen-Diagramm, Abhängigkeit des Herzminutenvolumens von Sym-
pathikus- und Vaguseinwirkung, Extrasystole, kompensatorische Pause.
Methodische Kenntnisse, die im Praktikum erworben werden sollen:
Herstellen eines Nerv-Muskelpräparates des M. gastrocnemius des Krallen-
frosches. Bedienen eines elektrischen Reizgerätes. Registrieren der Skelett-
muskelkontraktionen (Mechanogramme) mit Biegestabtransducern. Eichung der
Messapparatur. Umrechnung der Messwerte des Transducers in Kontraktions-
kräfte anhand der Kennlinie des Transducers. Registrieren des EMGs eines Hand-
muskels. Darstellen der Signale mittels Oszilloskop (Speichern, Triggern,
Bestimmen von Signalamplitude, Signaldauer und Signalfrequenz) sowie die
Dokumentation der Signale über den angeschlossenen Grafik-Drucker.
LITERATUR
SCHMIDT/THEWS/LANG: Einführung in die Physiologie des Menschen, Springer. -
- Hervorragendes Lehrbuch für die gesamte allgemeine Physiologie und die
Physiologie des Menschen. Leider ohne vergleichende tierphysiologische
Aspekte.
ECKERT/RANDALL: Animal Physiology, Freeman. --Gutes Lehrbuch für allgemeine
und vergleichende Physiologie. (Deutsche Übersetzung: Thieme)
MÜLLER: Tier- und Humanphysiologie, Springer --Gutes Lehrbuch für allgemeine
und vergleichende Physiologie
PENZLIN: Lehrbuch der Tierphysiologie, Elsevier, Spektrum, Akad. Verl.
Umfassendes Lehrbuch für allgemeine und vergleichende Physiologie
PRÄPARIERBESTECK
2 feine Pinzetten (wichtig)
1 feine spitze Schere (wichtig)
1 größere Schere
VERSUCHSTEIL I:
PHYSIOLOGISCHE UNTERSUCHUNGEN AN DER SKELETTMUSKULATUR
Versuchsaufbau:
Machen Sie sich zunächst mit der Versuchsapparatur vertraut und verkabeln
Sie diese richtig, damit sofort nach Fertigstellung der Präparation mit den
Messungen begonnen werden kann. Eine nahezu detailgetreue Wiedergabe der
Verschaltung finden Sie in Abbildung 1.
Wenn Sie sich versichert haben, dass alle Geräte richtig angeschlossen und
eingeschaltet sind, starten Sie auf Ihrem Computer das Programm „Chart“. Die
Abbildung1: Schema des Versuchsaufbaus für die Experimente am Froschmuskel

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 25
Grundeinstellungen für das Muskelexperiment können Sie jetzt aus einer
vorbereiteten Konfigurationsdatei laden
(Hauptmenü: File/Open).
Die Konfigurationsdatei für Ihren Kurs befindet sich dann im Ordner:
Ihr_Username\WahrnehmungPsychophysik\
Kalibrierung der Messanordnung:
Zunächst müssen Sie Ihre Messapparatur eichen: Wie bereits in Kapitel 1
beschrieben wurde, müssen Sie dafür sorgen, dass die gemessenen
Spannungswerte des Biegestabtransducers [mV] in die entsprechenden Kräfte
[N] umgerechnet werden, die auf ihn einwirken. Gehen Sie dabei
folgendermaßen vor:
Kalibrierung Channel 1:
Suchen Sie sich aus den Ihnen zur Verfügung stehenden Gewichten zwei
verschiedene im Bereich zwischen 20 und 200 g aus und hängen Sie diese
nacheinander an den Transducer. Starten Sie die Messung in Kanal 1. Wenn das
Gewicht an den Biegestabtransducer hängt, können Sie den Spannungswert des
Transducers (in mV) links neben dem Anzeigefeld ablesen. Gewicht und
Spannung werden notiert. Beachten Sie dabei, dass Sie die Masse der Gewichte
[g] noch in die entsprechende Gewichtskraft [N] umrechnen müssen! Jetzt haben
Sie zwei Wertepaare, die die Eichgerade ihres Transducers genau festlegen.
Um diese Werte in das Chart-Programm einzugeben, klicken Sie nun auf die
Schaltfläche Channel1.
Wählen Sie dann im Drop-Down-Menü den Punkt
Units Conversion
aus und tragen Sie die gerade gemessenen Wertepaare dort ein:
Die Einheit mV setzt das Programm selber in das jeweils erste Editfenster
ein. Anschließend wählen Sie noch die Einheit aus, die an der y-Achse stehen
soll (N) und die Anzahl der anzuzeigenden Nachkommastellen.

Weitere Einstellungen, die Sie überprüfen sollten:
→ Input Amplifier auswählen, als Range 50mV und als Low Pass Frequenz
100 Hz einstellen.
→ Arithmetic auswählen, als unit „N“ eintragen
Damit ist die Eichung für Channel1 abgeschlossen.
Hinweis: Diese Art von Eichung, die mit nur zwei Messpunkten
auskommt, ist nur dann zulässig, wenn Sie davon ausgehen können, dass
der Messwertaufnehmer eine lineare Kennlinie hat, was auf den
Biegestabtransducer zutrifft, aber keineswegs immer der Fall ist ...
Kalibrierung Channel 2:
Am Channel 2, über den der Muskel stimuliert werden soll, sollten Sie
ebenfalls kurz das Input Amplifier Fenster auswählen und dort eventuell
eingeschaltete Low Pass bzw. High Pass Einstellungen löschen, also auf off
stellen. Da Sie mit elektrischen Impulsen reizen werden, können Sie die anderen
Einstellungen für diesen Kanal in der Grundeinstellung belassen.
PRÄPARATION DES NERV-SKELETTMUSKEL-PRÄPARATES
wird vom Kursbetreuer durchgeführt
Ein Krallenfrosch (Xenopus spec.) wird mit einer Guillotine dekapitiert und
das Rückenmark durch Einführen einer Sonde in den Rückenmarkskanal zerstört.
Anschließend wird der Frosch enthäutet und der Körper mit Ringerlösung
abgespült. Nach diesem Arbeitsgang sollten auch alle Instrumente und die Hände
sorgfältig gereinigt werden, um das giftige Hautsekret zu entfernen.
Weitere Präparation des Nerv-Muskel-Präparates
Öffnen Sie die Bauchhöhle und entfernen Sie die Eingeweide. Suchen Sie
zunächst den Ischiadicus-Nerv, und knoten Sie an den beiden Nervenstümpfe
möglichst nah am Rückenmark einen Bindfaden an. Schieben Sie dazu vorsichtig
eine kleine, stumpfe oder gebogene Pinzette unter den Ischiadicus kurz hinter
seinem Austritt aus dem Wirbelkanal. Ziehen Sie einen mit Froschringer
angefeuchteten Zwirnsfaden mit der Pinzette unter dem Nerven durch. Knoten
Sie den Faden fest um den Nerv, und schneiden Sie vorsichtig das kürzere Ende
des Fadens dicht am Knoten ab. Der Faden dient später als Haltegriff. Dehnen
Sie oder fassen Sie den Nerven niemals mit den Fingern oder einer Pinzette an.
Durchtrennen Sie den Nerven proximal vom Knoten mit einer kleinen Schere.
Heben Sie den Ischiadicus mit Hilfe des Fadens an, ohne ihn zu dehnen.
Entfernen Sie mit einer kleinen Schere alle restlichen Bindegewebs-Kontakte,
und durchtrennen Sie Kollaterale direkt am Nerven, indem Sie vom Nerven weg

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 27
schneiden. Legen Sie den Nerven auf den zugehörigen Oberschenkel zurück.
Verfahren Sie nun in gleicher Weise auf der anderen Seite.
Nun können Sie Ober- und Unterkörper des Frosches nach Anweisung des
Kursleiters trennen: Den Oberkörper erhält die Gruppe, die den Herz-Versuch
durchführt, Sie fahren mit der Präparation der Beine fort, indem Sie das Becken
durch einen Schnitt in der Medianebene ebenfalls in Absprache mit dem
Kursbetreuer halbieren. Jede Gruppe fährt nun mit der Präparation eines Beines
fort.
Die dorsale Oberschenkelmuskulatur wird nun mit den Daumen
auseinandergedrückt, bis der Nerv sichtbar wird. Präparieren Sie den Nerven
über die ganze Länge des Oberschenkels von Blutgefäßen und Bindegewebe frei.
Beim Anheben des Nervs mit dem Haltefaden (nicht dehnen!) können seitlich
abgehende Nervenäste mit einer kleinen Schere durchtrennt werden. Beim
Abschneiden sollte die Schere wieder vom Nerven wegweisen. Wenn Sie am
Kniegelenk angelangt sind, wenden Sie sich zunächst dem M. gastrocnemius zu:
Einen Faden unter der Achillessehne durchziehen und diese sehr fest anbinden.
Die Sehne distal des Sesambeines durchschneiden. Den M. gastrocnemius mit
dem Faden sehr vorsichtig anheben und bis zum Ansatz am Femurstumpf vom
Unterschenkel ablösen. Dann den Unterschenkel um etwa die Hälfte kürzen und
die Tibia mit den restlichen Muskeln abschneiden. Weiterhin präparieren Sie alle
Oberschenkelmuskeln bis zum Kniegelenk ab und schneiden den Femur in
Beckennähe mit einer scharfen Schere durch, so dass ein ca. 1-2 cm langer
Femurstumpf übrigbleibt.
Das Nerv-Muskel-Präparat muss während des ganzen Versuchs mit
Ringerlösung feucht gehalten werden!!
1. Kontraktionsverhalten des M. gastrocnemius des Frosches
Das Nerv-Muskel-Präparat wird mit dem Femurstumpf in die Knochenklemme
eingespannt und mit dem an der Achillessehne befestigten Fadenstück an einem
Biegestabtransducer befestigt. Der Nervenstumpf wird vorsichtig über die
Reizelektroden gelegt (Kathode muskelnah). Das Präparat, vor allem der Nerv,
muss fortwährend mit Ringer feucht gehalten werden. Wenn gerade keine
Messungen durchgeführt werden, kann der Nerv an den Muskel angelegt werden,
um ihn vor dem Austrocknen zu schützen.
Richten Sie das Präparat mit dem Feintrieb des Stativs so ein, dass der Faden
gerade eben gespannt ist. Lesen Sie am Kanal 1 einen eventuell vorhandenen
Offset (ca 0.2 bis 0.5N) ab und kompensieren Sie diesen Offset unter
Zuhilfenahme der Funktionen, die Ihnen im Menüpunkt Arithmetic zur
Verfügung stehen
Hinweis: Da das Muskelpräparat sich im Laufe des Kurses verändern kann,
ist es möglich, dass sie diese Offsetkompensation öfters während des Kurses
wiederholen müssen.

Versuch 1.1: Abhängigkeit der Kontraktionskraft von der Reizamplitude.
Rufen Sie die
Stimulatorfunktion von
PowerLab auf
(Setup/Stimulate ...).
Das Fenster ist mehr oder
weniger selbst erklärend.
Stellen Sie zunächst
Einzelreizungen mit 0.2 ms
Reizdauer und einer
Amplitude von 10mV ein.
Achten Sie darauf, dass die
Stimulatorfunktion auf „On“
steht
Hinweis: Während der Messung können Sie sich im sog. Stimulatorpanel
eine Kurzform der Stimulatoreinstellungen anzeigen lassen.
Menü: Setup/Stimulator Panel.
Die Stimulatorfunktion kann nur bei laufender Messung („Start“) genutzt
werden.
Durchführung und Auswertung:
Bestimmen Sie, ausgehend von 10mV die Reizamplitude, bei der Einzelreize
den Muskel gerade zur Kontraktion bringen (=Minimalreizamplitude). Erhöhen
Sie dann stufenweise die Reizamplitude, bis durch weitere Erhöhung der
Reizamplitude keine weitere Steigerung der Kontraktionskraft mehr erzielt wird
(=Maximalreizamplitude).
Wählen Sie diese beiden ermittelten Grenzwerte und zusätzlich 3 Werte in
geeigneten Intervallen zwischen ihnen als Voreinstellungen für die
Reizamplitude, für die sie dann die Kontraktionskraft des Muskels bestimmen:
Registrieren Sie jeweils 5 Einzelzuckungen mit dem Oszilloskop für jede der
5 eingestellten Reizamplituden. Erstellen Sie ein Diagramm aus den Messwerten,
das die Kontraktionskraft in Abhängigkeit von der Reizamplitude darstellt.
Erklären Sie das Ergebnis unter dem Gesichtspunkt der „Alles-oder-Nichts“
Regel.

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 29
Hinweis: Nachträgliches Durchsuchen der Messergebnisse:
Alle Messwerte, die Sie zwischen „Start“ und „Stop“ aufgenommen haben,
können Sie sich nach der Messung noch mal ansehen: Mit dem kleinen
Schieber am unteren Bildrand gehen Sie in der Zeit vor- und zurück.
Den Maßstab der Zeitachse können Sie mit den anderen Schaltflächen rechts
unten verändern.
Versuch 1.2: Abhängigkeit der Kontraktionskraft von der Reizfrequenz.
Reizen Sie den Muskel mit der Maximalreizamplitude, die Sie soeben
bestimmt haben. Wählen sie jetzt im Stimulator-Fenster Mehrfachreizungen aus
und zeichnen Sie die Kontraktionen des Muskels nacheinander bei Reizungen von
jeweils 5 Sekunden Länge mit 1,2,5,10,20 und 50Hz Reizfrequenz auf. Überlegen
Sie sich vorher, welche Einstellungen am Oszilloskop und Stimulator eingestellt
werden müssen, damit die Reize auch aufgenommen werden – häufige
Wiederholungen dieses Experiments sind zwar möglich, der Muskel wird
allerdings allmählich ermüden, da im Präparat kein ATP nachgebildet werden
kann.
Ermitteln Sie die Reizfrequenzen, bei denen zum ersten mal ein
unvollständiger bzw. ein vollständiger Tetanus eintritt. Verwenden Sie im
Protokoll die Oszillogramme der Kontraktionskraft und der zugehörigen
Reizmuster, um die einsetzende Tetanisierung bei steigender Reizfrequenz zu
verdeutlichen. Erklären Sie kurz den Mechansimus, der der Tetanisierung zu
Grunde liegt.
1.3 Abhängigkeit der Kontraktionskraft von der Ausgangslänge
Messen Sie die Ausgangslänge des Muskels mit einem Lineal. Reizen Sie den
Muskel mit der Maximalreizamplitude und registrieren Sie jeweils 5 Einzel-
zuckungen mit dem Oszilloskop. Führen Sie diesen Versuch bei verschiedenen
Ausgangslängen durch. Die Länge des Muskels können Sie durch Drehen an der
Feinjustierung am Biegestab verändern. Was passiert dabei im Muskel? Erstellen
Sie ein Diagramm, in dem die Kontraktionskraft in Abhängigkeit von der
Ausgangslänge dargestellt ist. Mit welcher Ausgangslänge würde der Muskel wohl
am effizientesten im Skelettsystem des Frosches arbeiten?
Machen Sie sich klar (und diskutieren Sie in Ihrem Protokoll), welche
physiologischen Vorgänge zwischen der elektrischen Reizung am Nervenstumpf
und der Kontraktion des Muskels ablaufen.

Überlegen Sie, welche Art von Kontraktion (isometrisch, isotonisch oder
auxotonisch) in diesem (und den anderen) Experimenten vorliegt. Die
Auslenkung des Biegestabs, die bei Belastung zu einer kleinen Verkürzung führt,
kann für diese Überlegung vernachlässigt werden.
Hinweis: Sollte der Zustand des Nerv-Muskelpräparates so schlecht sein, dass
die Reizung am Nervenstumpf unmöglich wird, so können Sie das vorge-
schriebene Programm auch mit direkter Muskelreizung durchführen. In diesem
Fall wird eine Elektrode in den Muskel eingestochen, die zweite Elektrode leitet
über die Knochenklemme zu. Die Reizdauer sollten Sie dann auf 5 ms
verlängern.
2. Messen der Latenzzeit für die Kontraktion bei elektrischer Reizung am
Nervenstumpf bzw. direkt am Muskel
Hierzu wird zuerst über den Nervenstumpf und dann direkt über den Muskel
gereizt. In zweiten Fall wird die Elektrode in den Muskel eingestochen, die zweite
Elektrode hat Kontakt über die Knochenklemme. Die Reizdauer bei Reizung über
den Nerv beträgt wieder 0,2 ms, bei Reizung direkt am Muskel 5 ms.
Durchführung und Auswertung:
Reizen Sie mit 'Einzelreizen' (Reizabstand von 5 s einstellen) von ca. 300 mV am
Nervenstumpf, und bestimmen Sie am Oszilloskop die Latenz zwischen
Reizbeginn und Beginn der vom Transducer angezeigten Kontraktion. Überlegen
Sie bitte zuvor, in welchem Zeitbereich Sie die Latenzen erwarten, und stellen
Sie die Zeitachse am Oszilloskop entsprechend in einen sinnvollen Bereich.
Wiederholen Sie diesen Versuch mit elektrischer Reizung direkt am Muskel
(Amplitude hier ca. 3 V: warum?). Auch hier bestimmen Sie die Latenz zwischen
Reizbeginn und Kontraktion.
Vergleichen Sie die Latenzen, und diskutieren Sie die Ergebnisse in Ihrem
Protokoll.

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 31
VERSUCHSTEIL II:
ELEKTROMYOGRAMM (EMG) EINES MENSCHLICHEN SKELETTMUSKELS
Die Fragestellung dieses Versuchsteils bezieht sich auf die Mechanismen, die
eine feine Regulation der Muskelkraft ermöglichen. Bei den Wirbeltieren wird jede
Faser eines Muskels von genau einem Motoneuron kontaktiert1. Allerdings
können einzelne Motoneurone mehrere Muskelfasern innervieren. Ein
Motoneuron und alle davon innervierten Muskelfasern werden als „motorische
Einheit“ bezeichnet. Jeder Muskel besteht aus 100 bis 1000 solcher motorischen
Einheiten. Ein Aktionspotenzial eines Motoneurons führt zu einer Zuckung aller
kontaktierten Muskelfasern. Die motorische Einheit kann daher als die
elementare Größe der Muskelkraft betrachtet werden.
Grob lassen sich zwei unterschiedliche Typen von Muskelfasern unterscheiden:
1. Typ I / ST-Fasern (= slow twitch): langsame, nicht ermüdende Fasern,
2. Typ II / FT-Fasern (= fast twitch): schnelle, rasch ermüdende Fasern.
Die Typ II Fasern verfügen im Vergleich zu Typ I über nur wenig Myoglobin – der
im Muskel dominierenden Isoform des Hämoglobins. Sie sind gekennzeichnet
durch eine vorwiegend anaerobe Energiegewinnung (Glycolyse). Muskeln, in
denen der Fasertyp II vorherrscht, sind daher auch deutlich heller (= ’weiße’
Muskeln) als die rot gefärbte Typ-I Muskulatur. Unter den Typ II-Fasern wird
noch einmal unterschieden zwischen Typ IIA und IIB Fasern. Die IIB-Fasern sind
extrem schnell in der Kraftentfaltung, ermüden allerdings ebenso schnell. Sie
gewinnen ihr ATP ausschließlich aus Glykogen, während die Typ IIA Fasern
zumindest teilweise ATP oxidativ gewinnen und daher eine Mischform darstellen.
Neben der maximalen Kraftentwicklung und der Ermüdungs-Schwelle nehmen
innerhalb dieser drei Typen von Muskelfasern auch der Faserdurchmesser, die
ATPase-Tätigkeit und der Glykogen-Gehalt in den Fasern zu (I < IIA < IIB), der
Myoglobingehalt und die Kapillardichte (oxidative Energie-Gewinnung) nehmen
dagegen ab. Die Zusammensetzung aus den einzelnen Typen variiert stark von
Muskel zu Muskel. So besteht der bereits im Stand oder im Gehen voll aktivierte
Schollenmuskel (M. soleus) zu 90% aus langsamen Fasern, während der erst bei
schnellkräftigen Bewegungen wie dem Springen voll aktivierte Zwillings-
wadenmuskel (M. gastrocnemius) zu 45% aus schnellen, ermüdenden und zu
25% aus schnellen, nicht-ermüdenden Fasern besteht.
Die Verteilung des Anteils der verschiedenen Fasertypen in der
Skelettmuskulatur ist in hohem Maß genetisch vorbestimmt, Sportler können
allerdings durch Training speziell den Anteil an Typ II – Fasern positiv beein-
flussen. Während Ausdauerathleten einen hohen Anteil an ST-Fasern benötigen,
besitzen Kraftsportathleten dagegen mehr FT-Fasern.
1 Bei Wirbellosen ist dieses Prinzip nicht gültig! Die neuronale Verrechnung, die bei Wirbeltieren im
Rückenmark stattfindet und auf der Ebene des Motoneurons abgeschlossen ist, findet bei den Wirbellosen noch an der Muskelfaser statt. So existieren bei Invertebraten inhibitorische Motoneurone (wie z.B. der common inhibitor).

Notwendige Präparation
Ein EMG lässt sich extrazellulär ableiten mit Hilfe von Elektroden, die über
dem Muskel auf der Haut aufgeklebt werden. Es liegt auf der Hand, dass Sie mit
einer derartigen Methode lediglich das Summenpotenzial vieler motorischer
Endplatten und den dazu führenden Motoneuronen aufnehmen. Trotzdem werden
Sie charakteristische Zusammenhänge zwischen dem Signalverlauf und der vom
jeweiligen Muskel entfalteten Kraft messen können.
KONTRAKTION EINES MUSKELS DER HAND
Eine Kontraktion des M. interosseus dorsalis I führt zu einer Abduktion des
Zeigefingers. Dieser intrinsische, dorsale Handmuskel eignet sich besonders gut
zur Aufzeichnung eines EMGs, da es keinen anderen Muskel (der
korrespondierende ventrale Handmuskel fehlt beim Zeigefinger) gibt, der
ebenfalls eine Abduktion dieses Fingers bewirken würde. Der Muskel kann bei
einer Abduktion des Zeigefingers leicht ertastet werden. Eine Elektrode sollte
direkt auf den Muskel geklebt werden, die Referenzelektrode wird seitlich dazu
aufgeklebt. Die Qualität des EMGs kann drastisch verbessert werden, wenn der
Hautwiderstand unter den Elektroden durch Abreiben mit Alkohol verringert wird.
Eine Erdung (Masse-Armband) sollte nicht vergessen werden.
1 M. interosseus dorsalis I
2 Os metacarpale I
3 Os metacarpale II
4 Dorsalaponeurose
5 Phalanx proximalis
Versuchsaufbau
Isometrische Kontraktionen können durch die Belastung des Zeigefingers mit
unterschiedlichen Gewichten erreicht werden. Die Gewichte werden dabei an eine
Abbildung 2: Schema zur Anbringung der Elektroden auf der rechten Hand.

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 33
Schnur gehängt; ihre Gewichtskraft wird über einen Faden und eine Rolle auf den
Zeigefinger übertragen. Die aktuelle Position des Zeigefingers (Abduktion) wird
über einen Steuerknüppel gemessen, der mit dem Finger bewegt wird. So kann
auch das EMG bei isotonischen Bewegungen aufgezeichnet werden. Auf dem
Oszilloskop werden sowohl das EMG als auch die Position des Fingers dargestellt.
Zur Messung kleben Sie zwei Elektroden auf die Hand der Versuchsperson auf,
wie in Abbildung dargestellt. Schließen Sie die Elektroden an Kanal 1 des
vierpoligen Adapterkabels an, das andere Ende schließen Sie am A/D-Wandler an
der Buchse ‚BioAmp’ an. Vergessen Sie nicht, das Masseband anzuschließen und
um das Handgelenk der Versuchsperson zu binden.
An den zweiten Eingangskanal des A/D-Wandlers schließen Sie den
Steuerknüppel des Positionsmelders an. (Vergessen Sie nicht, den
Positionsmelder an das 10V-Netzgerät anzuschließen). Danach richten Sie die
entsprechenden Kanäle in PowerLab ein.
1. Isometrische Kontraktion
Belasten sie den Zeigefinger ihrer Versuchsperson mit unterschiedlichen
Gewichten bis zur maximalen Belastbarkeit (je nach Versuchsperson bis etwa 3
kg). Achten Sie darauf, dass die Position des Fingers konstant bleibt (das Signal
vom Steuerknüppel darf sich nicht ändern). Messen und dokumentieren Sie das
EMG für jede Belastung einmal über 10 s zur Übersicht sowie einmal über 200
ms zur Identifikation einzelner Summenpotenziale.
Auswertung
Können tatsächlich die Summenaktionspotenziale einzelner motorischer Einheiten
erkannt werden? Was bedeuten unterschiedliche Spannungsverläufe?
Bei welcher Kraft werden die einzelnen Muskelfasern rekrutiert?
Gibt es eine Kodierung der Muskelkraft in der Frequenz der Aktionspotenziale
einer Muskelfaser?
2. Dynamische Kontraktion
Besonders deutlich wird der Zusammenhang zwischen der Abduktion des
Zeigefingers und des EMGs, wenn der Finger tatsächlich bewegt wird. Zeichnen
Sie das EMG bei verschiedenen Frequenzen der Fingerbewegung auf. Bitten Sie
die Versuchsperson, ihren Finger mit konstanter Frequenz hin und her zu
bewegen. Stellen Sie die Zeitbasis des Oszilloskop so ein, dass mindestens eine
volle Periode auf dem Schirm (bzw. Ausdruck) abgebildet wird. Warum nimmt
das EMG mit steigender Frequenz der Fingerbewegung zu?

VERSUCHSTEIL III:
PHYSIOLOGISCHE UNTERSUCHUNGEN AM FROSCHHERZEN
PRÄPARATION DES HERZMUSKELPRÄPARATS
Sie erhalten von dem Nerv- und dem Muskelversuch den Torso eines Frosches.
Der Froschtorso wird mit der Bauchseite nach oben in die Präparierschale gelegt,
an beiden Vorderextremitäten und seitlich am Thorax mit Stecknadeln
festgesteckt. Durch Anheben des Sternums mit einer Pinzette wird das Herz im
Thorakalraum bereits sichtbar. Ziel ist es das Herz in situ frei zu präparieren. In
situ bedeutet, dass das Herz im Körper verbleibt und dort weiterschlägt.
Schneiden Sie vorsichtig links und rechts des Sternums in Richtung des Kopfes.
Trennen Sie Clavicula und Coracoid ab und entfernen Sie das Brustbein mit den
anliegenden Gewebeteilen. Das Präparat gründlich mit Ringer-Lösung spülen.
Eröffnen Sie den silbrig schimmernden Herzbeutel vorsichtig mit einer feinen
Pinzette und einer kleinen Schere und reinigen Sie das Herz sorgfältig von allen
Resten des Perikards. Zuletzt schneiden Sie das unter dem Herzen liegende
Herzbändchen durch.
Wichtig: Das Präparat immer reichlich mit Ringer-Lösung feucht halten und
vorsichtig behandeln!

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 35
VERSUCHSAUFBAU UND APPARATUREN
Machen Sie sich vor der Präparation des Frosches mit der Apparatur vertraut.
Eine lange zeitliche Verzögerung zwischen dem Töten des Frosches und der
Untersuchung des Herzens, kann die Funktion des Herzens stark einschränken.
Achten Sie darauf, dass der Biegestabtransducer fest im Dreifuß eingespannt ist.
Der Transducer wird über ein dreipoliges Kabel sowohl mit dem Netzteil als auch
mit dem Differenzverstärker verbunden. Der Ausgang des Differenzverstärkers
ist mit Eingang 1 des PowerLab 26T analog-digital Konverters von
ADInstruments verbunden, welcher über den USB-Eingang mit dem Computer in
Verbindung steht. Achten Sie darauf, dass alle Geräte eingeschaltet und mit
Strom oder Batterie versorgt sind. Dieser Versuchsaufbau wird für Versuch 2
verwendet. Für Versuch 3 müssen die beiden Elektroden mit dem Ausgang des
Reizgerätes verbunden werden. Außerdem wird der Ausgang des Reizgerätes mit
dem zweiten Eingang des analog-digital Konverters verbunden.
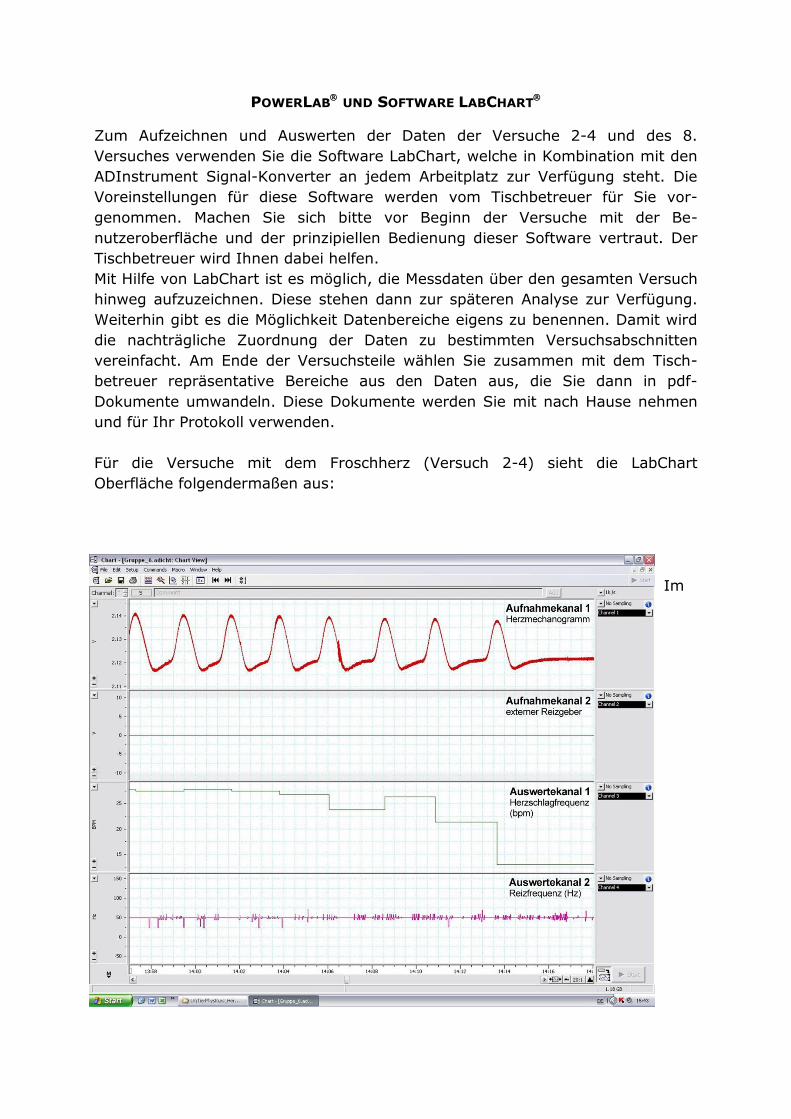
POWERLAB UND SOFTWARE LABCHART
Zum Aufzeichnen und Auswerten der Daten der Versuche 2-4 und des 8.
Versuches verwenden Sie die Software LabChart, welche in Kombination mit den
ADInstrument Signal-Konverter an jedem Arbeitplatz zur Verfügung steht. Die
Voreinstellungen für diese Software werden vom Tischbetreuer für Sie vor-
genommen. Machen Sie sich bitte vor Beginn der Versuche mit der Be-
nutzeroberfläche und der prinzipiellen Bedienung dieser Software vertraut. Der
Tischbetreuer wird Ihnen dabei helfen.
Mit Hilfe von LabChart ist es möglich, die Messdaten über den gesamten Versuch
hinweg aufzuzeichnen. Diese stehen dann zur späteren Analyse zur Verfügung.
Weiterhin gibt es die Möglichkeit Datenbereiche eigens zu benennen. Damit wird
die nachträgliche Zuordnung der Daten zu bestimmten Versuchsabschnitten
vereinfacht. Am Ende der Versuchsteile wählen Sie zusammen mit dem Tisch-
betreuer repräsentative Bereiche aus den Daten aus, die Sie dann in pdf-
Dokumente umwandeln. Diese Dokumente werden Sie mit nach Hause nehmen
und für Ihr Protokoll verwenden.
Für die Versuche mit dem Froschherz (Versuch 2-4) sieht die LabChart
Oberfläche folgendermaßen aus:
Im
TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 37
Aufnahmekanal 1 werden die verstärkten Signale des Transducers als
Herzmechanogramm aufgezeichnet.
Aufnahmekanal 2 stellt die Impulse des externen Reizgebers dar. Zwei weitere
Kanäle (Auswertekanäle) können so genutzt werden, dass sie, basierend auf den
Daten der Aufnahmekanäle, automatisch die Herzschlagfrequenz (bpm - beats
per minute) und die Reizfrequenz (Hz) berechnen.
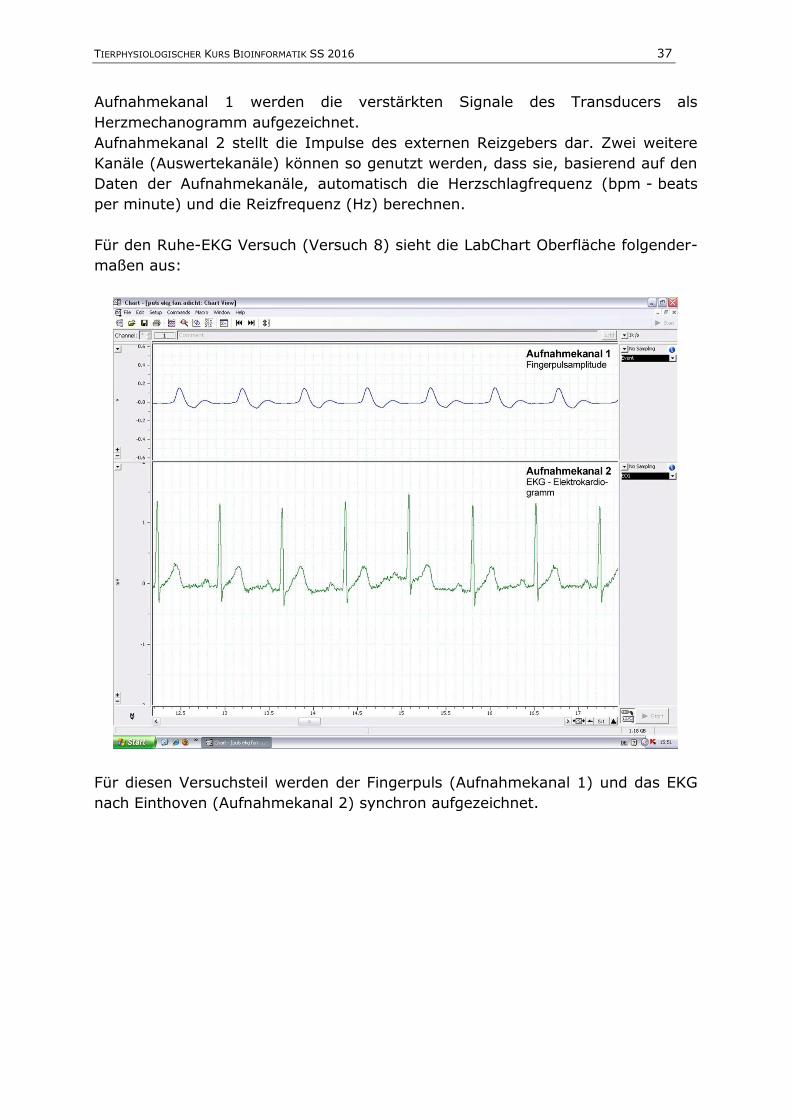
Für den Ruhe-EKG Versuch (Versuch 8) sieht die LabChart Oberfläche folgender-
maßen aus:
Für diesen Versuchsteil werden der Fingerpuls (Aufnahmekanal 1) und das EKG
nach Einthoven (Aufnahmekanal 2) synchron aufgezeichnet.

VERSUCHE
Versuch 1:
Beobachten der Herzbewegung
Identifizieren Sie zunächst wichtige Herzabschnitte, wie den Sinus
venosus, die Atrien, den Ventrikel und den Truncus arteriosus. Beobachten
Sie die Kontraktionsfolgen der einzelnen Herzabschnitte. Wo beginnt die
Kontraktionswelle und wo endet sie wieder?
Versuch 2:
Registrierung eines Mechanogramms
In den folgenden Versuchsteilen (2-4) dient das Mechanogramm der Froschherz-
Kontraktion als Messparameter. Um ein Mechanogramm des Herzens registrieren
zu können, muss das Herz mit der Versuchsapparatur verbunden werden. Heben
Sie dazu die Ventrikelspitze leicht an und befestigen Sie die Herzklammer
vorsichtig an der äußersten Spitze des Ventrikels. Ein dünner Faden verbindet die
Herzklammer mit dem Biegestabtransducer. Justieren Sie die Versuchsapparatur
so, dass der Herzklammerdraht senkrecht über dem Herzen nach oben zum
Biegestab verläuft. Das ist wichtig, um eine gute Übertragung der Herz-
kontraktion auf den Transducer und damit eine saubere Registrierung zu be-
kommen. Stellen Sie das Mechanogramm nun auf dem Computerbildschirm dar.
Wählen Sie dazu sinnvolle Dimensionen für die Zeit- und Spannungsachse in den
LabChart Einstellungen.
Wichtig: Das Herz muss ständig mit reichlich Ringer-Lösung feucht gehalten
werden!
Speichern Sie ein Mechanogramm, auf dem mehrere Herzzyklen registriert
werden. Ordnen Sie den beobachteten Kontraktionsverlauf des Herzens
den Registrierungen des Mechanogramms zu. Ermitteln Sie aus dem
registrierten Mechanogramm die Herzfrequenz.
Speichern Sie am Ende der Herzversuche die Messungen als pdf-
Datei ab und fügen Sie diese später in Ihr Protokoll ein.

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 39
Versuch 3:
Einfluss von Atropin, Noradrenalin und Acetylcholin auf die Herztätigkeit
Bei Wirbeltieren wirken die Transmitter Noradrenalin des postganglionären
Sympathikus-Nervensystems und Acetylcholin der parasympathischen Vagus-
Nerven auf die autorhythmischen Zentren des Herzens und zum Teil auch auf das
Arbeitsmyokard. Im folgenden Versuchsabschnitt soll die Wirkungsweise dieser
Pharmaka untersucht werden.
Versuchsdurchführung
Der Versuchsaufbau entspricht dem des 2. Versuchs.
Im Folgenden wird die Herzfrequenz unter Einfluss verschiedener Substanzen
gemessen. Beobachten und dokumentieren Sie jeweils die Reaktion des Herzens.
Nutzen Sie hier die Möglichkeit der LabChart Software und benennen Sie die
entsprechenden Datenbereiche während der Aufnahme eindeutig, um diese
später wieder zu finden.
Wichtig: Das Präparat muss zwischen den einzelnen Arbeitsschritten immer
gründlich mit Ringer-Lösung gereinigt und feucht gehalten werden.
Registrieren Sie eine Zeit lang die normale Herztätigkeit. Bestimmen Sie
die Herzfrequenz und die Amplitude der Kontraktion. Diese Werte sind Ihre
Kontrollwerte.
Geben Sie Ringer-Lösung, die direkt aus dem Kühlschrank kommt, auf das
Präparat.
Beträufeln Sie das Herz mit zimmerwarmer Ringer-Lösung.
Stellen Sie die Ringer-Lösung an einen warmen Ort (Heizung, Sonne) und
wiederholen Sie den oben genannten Arbeitsschritt.
Geben Sie einige Tropfen der vorbereiteten Adrenalinlösung auf das Herz.
Nach der Registrierung muss gut mit der Ringer-Lösung gespült werden.
Sobald das Herz wieder seinen Normalrhythmus erreicht hat, tropfen Sie
die Acetylcholinlösung auf das Herz auf und beobachten, was geschieht.
Sollte es zu einem Herzstillstand kommen, sofort etwas von der Atropin-
lösung auf das Herz auftropfen, um die Herztätigkeit wieder anzuregen.
Zuletzt tropfen Sie etwas von der Atropinlösung auf das Herz.
Speichern Sie später typische Mechanogramme eines jeden Arbeitsschrittes für
Ihr Protokoll und diskutieren Sie die Wirkungen von Temperatur und
verschiedener Transmitter auf die Herz-tätigkeit.

ERFOLGSKONTROLLE
Nach diesem Praktikumsteil sollten Sie in der Lage sein:
den anatomischen Grobaufbau von einem Skelettmuskel zu beschreiben,
den anatomischen Feinbau von Muskelfasern zu beschreiben,
die nacheinander ablaufenden physiologischen Vorgänge von der elek-
trischen Reizung am Nerv bis hin zur Kontraktion des Muskels zu
erläutern,
die molekularen Mechanismen der Kontraktion zu beschreiben,
ein Nerv-Muskel-Präparat herzustellen,
zu erklären, wie ein Tetanus entsteht,
zu erläutern, wovon die Kontraktionskraft abhängig ist,
die Versuchsapparatur selbständig aufzubauen,
den Versuchsaufbau im Muskelversuch zu skizzieren,
eine EMG-Ableitung vom M. interosseus dorsalis durchzuführen,
den Begriff Rekrutierung inklusive Hennemannsches Größenprinzip zu
erläutern,
die in Ihrem Protokoll aufgeführten Kurven zu skizzieren und zu erläutern,
ein funktionelles Herz skizzieren zu können,
zu beschreiben, wie die Erregungswelle über das Herz läuft,
nachzuvollziehen wie das Blut durch das Herz gepumpt wird und welche
Mechanismen dabei wichtig sind,
die Entstehung des Schrittmacherpotenzials zu erläutern,
ein charakteristisches Aktionspotenzial zu skizzieren und zu erklären
und die Wirkung verschiedener Pharmaka auf die Herztätigkeit zu
erklären.

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 41
3. I N S E K T E N - E R G
EINFÜHRENDE BEMERKUNGEN
Sinnessysteme ermöglichen es einem
Organismus, Reize aus seiner Umwelt
wahrzunehmen. Die Reize wirken in den
Sinnesorganen auf spezifische Rezeptoren und
erzeugen an den Zellmembranen der
Rezeptorzellen Potenzialänderungen, die zur
Erregung afferenter sensorischer Nervenfasern
führen. Diese Potenzialänderungen lassen sich
extrazellulär ableiten und ermöglichen die
quantitative Beschreibung von Rezeptor-
leistungen. Am heutigen Kurstages werden bestimmte Eigenschaften von
Photorezeptoren (z.B. Kennlinie, Farbempfindlichkeit, Latenzzeit, zeitliches
Auflösungsvermögen) experimentell durch elektrophysiologische Ableitung
(objektive Sinnesphysiologie) am Fliegenauge dargestellt. Einige Eigenschaften
des Facettenauges werden mit den Leistungen des visuellen Systems des
Menschen verglichen, die in psychophysischen Experimenten (subjektive
Sinnesphysiologie) ermittelt werden.
THEORETISCHE VORAUSSETZUNGEN
Folgende theoretischen Vorkenntnisse aus der Vorlesung "Grundlagen der
Tierphysiologie" und aus den Lehrbüchern werden für die sinnvolle Durchführung
der Versuche benötigt und deshalb vorausgesetzt:
Allgemeine Sinnesphysiologie: Phasische und tonische Rezeptoren
(Beispiele), Kennlinien, Beziehung zwischen Reiz, Rezeptorpotenzial und
Impulsfolgefrequenz, adäquater Reiz.
Sehphysiologie: Bau und Funktion des Wirbeltierauges am Beispiel des
menschlichen Auges: Aufbau, dioptrischer Apparat, Bildentstehung,
Akkomodation, Fehlsichtigkeit, Aufbau der Netzhaut, Photorezeptoren von
Wirbellosen und Wirbeltiere (Besonderheiten des Ruhe- und
Belichtungspotenzials sowie die ursprünglichen lichtempfindlichen
Organelle), Sehpigmente und Transduktionsprozess, Farbempfindlichkeit,
Adaptation, räumliches und zeit-liches Auflösungsvermögen, magno- und
parvozelluläres System.
Bau und Funktion des Insektenauges: Bau eines Ommatidiums,
Appositionsauge, optisches Superpositionsauge, neurales Superpositions-
auge, Umwandlung der Photopigmente, Transduktionsprozess, räumliches
Auflösungsvermögen, zeitliches Auflösungsvermögen (Flimmerver-

schmelzungs-frequenz), Spektralempfindlichkeit, Elektroretinogramm
(ERG).
Methodische Kenntnisse, die Sie im Praktikum erwerben sollen:
Präparation der Fliege, Durchführen einer extrazellulären Summenableitung.
Datenerfassung mit dem Programm Chart. Eine Einführung in die Bedienung des
Programms und der im Praktikum verwendeten technischen Geräte wird durch
den Kursbetreuer gegeben. Grundlagen eines psychophysischen Experiments.
LITERATUR
PENZLIN: Lehrbuch der Tierphysiologie, Fischer. -- Erläutert ausführlich die
optischen Eigenschaften verschiedener Augentypen. Viele vergleichende
Aspekte.
DUDEL/MENZEL/SCHMIDT: Neurowissenschaft, Springer. -- Anspruchsvolles und
gutes Kapitel über Photorezeption im Facetten- und Linsenauge.
SCHMIDT/THEWS/LANG: Einführung in die Physiologie des Menschen, Springer. -
- Erläutert ausführlich die Prinzipien der Sehphysiologie wie z.B. den
Mechanismus der Phototransduktion und die anatomischen und
physiologischen Verhältnisse beim Säugerauge. Leider ohne vergleichende
tierphysiologische Aspekte.
Weiterhin empfehlenswert:
MÜLLER: Tier- und Humanphysiologie, Springer -- Erläutert die Prinzipien der
Sehphysiologie wie z.B. den Mechanismus der Phototransduktion und die
anatomischen Verhältnisse beim Säugerauge und bei den verschiedenen
Facettenaugen-Typen der Insekten.
ECKERT/RANDALL: Animal Physiology, Freeman. (Deutsche Übersetzung:
Thieme) -- Erläutert die Prinzipien der Sehphysiologie wie z.B. den
Mechanismus der Phototransduktion und die anatomischen Verhältnisse
beim Säugerauge. Bei den Facettenaugen liegt die Betonung nicht auf den
Insekten.
KIRSCHFELD K: (1971) Aufnahmen und Verarbeitung optischer Daten im
Komplexauge der Insekten. Naturwissenschaften 58: 201-209. - neuronales
Superpositionsauge.

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 43
HANDWERKSZEUG
1 kleine Pinzette
1 Federstahlpinzette
Schreibzeug
Geodreieck
Datenspeicher
VERSUCHE
Die Leistungsfähigkeit des optischen Systems eines Organismus wird durch
sein räumliches Auflösungsvermögen (Sehschärfe), sein zeitliches
Auflösungsvermögen, seine spektrale Empfindlichkeit und seine absolute
Lichtempfindlichkeit beschrieben. Diese Eigenschaften sollen am Fliegenauge
geprüft und, soweit möglich, mit denen des menschlichen Auges verglichen
werden.
1. Räumliches Auflösungsvermögen (Sehschärfe)
Wie exakt und feinkörnig das Bild der Umwelt von einem Wirbeltier
wahrgenommen wird, hängt entscheidend von der Dichte der Rezeptoren in der
Retina ab. Das Auflösungsvermögen eines Komplexauges wird im Wesentlichen
von zwei Parametern bestimmt: dem physiologischen Öffnungswinkel, der die
Richtungsempfindlichkeit des Rezeptors wiedergibt (er kann nur durch
komplizierte intrazelluläre Ableittechnik aus einem Photorezeptor oder durch
Verhaltensexperimente bestimmt werden), und dem Divergenzwinkel, der die
optischen Achsen benachbarter Ommatidien einschließt (er kann aus der
Anatomie des Komplexauges ermittelt werden). Im Kurs bestimmen Sie diesen
Divergenzwinkel bei verschiedenen Insekten und berechnen daraus deren
Auflösungsvermögen.
1.1 Messung des Auflösungsvermögens bei Insekten und beim Mensch
Im Kurs werden Ihnen die Horizontalschnitte durch das Facettenauge der
Wachsmotte (hell- und dunkel adaptiert) und des Kaisermantels zur Verfügung
gestellt.
Um welche Augentypen handelt es sich?
Identifizieren Sie die wichtigsten Teile des Insektenauges und der an-
grenzenden Gehirnteile.
Zeichnen Sie den Verlauf der Ommatidien mit dem Lineal nach, und
verlängern Sie die Linien zu einem Schnittpunkt
Bestimmen Sie den Divergenzwinkel der beiden Augenpräparate durch
Auswertung und Mittelwertbildung von 10 benachbarten Ommatidien.
Bestimmen Sie das räumliche Auflösungsvermögen einer Versuchsperson
in der Fovea centralis mit Hilfe des psychophysischen Messprogramms
(AUFLOESE). Eine detaillierte Einführung in das Programm erhalten Sie im
Kurs.

Berechnen Sie das Auflösungsvermögen von Mensch, Wachsmotte und
Kaisermantel. Wie groß muss der Abstand zwischen zwei Punkten in 1 m
Entfernung sein, damit diese gerade noch getrennt wahrgenommen
werden können? Das Auflösevermögen der Facettenaugen berechnen Sie
nach folgender Formel:
tan α = Gegenkathete/Ankathete d/E
tan α/2=d/2E d = 2E tan α/2
2. Exkurs: Das Elektroretinogramm (ERG) der Fliege
Belichtet man das Auge von Wirbeltieren oder Wirbellosen, so treten infolge
der durch den Transduktionsprozess ausgelösten Membranprozesse
Potenzialschwankungen im Auge auf. Die Summenpotenziale der gesamten elek-
trischen Aktivität des Auges kann man mit geeigneten Elektroden als
Potenzialdifferenz zwischen Auge und Körpermilieu ableiten. Ein ERG erhalten
Sie, wenn Sie die Potentialdifferenzen werden über der Zeit auftragen. Dabei
handelt es sich um eine extrazelluläre Summenableitung (vgl. SAP im
Nervversuch). Die Potenziale haben z.T. komplizierte Kurvenverläufe, die bis
heute nicht endgültig geklärt werden können (vgl. Summenableitungen EKG,
EEG), die aber eine einfache Methode darstellen, um quantifizierbare Aussagen
über Erregungsvorgänge in den Photorezeptoren zu machen. Die ERG-Kurve der
Insekten ist z.B. stark von den Ableitbedingungen abhängig (Alter des Präparats,
Lage der Ableitelektrode, Elektrodenmaterial, Eingangswiderstand des Ver-
stärkers etc.).
Während man bei "langsamen" Insekten (Bsp. Heuschrecken, Schaben)
tendenziell eher monophasische Potenzialverläufe (Abb. 1A) feststellen kann,
sind bei schnellfliegenden Formen, zu denen die Schmeißfliege Calliphora zählt,
diphasische Potenzialverläufe (Abb. 1B) charakteristisch.
E
d

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 45
In dieser Form des diphasischen ERG's spiegelt sich sowohl die summierte
Aktivität der Rezeptoren (Depolarisation) als auch die Summenaktivität
nachgeschalteter neuronaler Elemente, hauptsächlich vom 1. optischen Ganglion
(Lamina ganglionaris) wieder. Die Antwort der Laminaneurone ist ähnlich
graduiert von der Reizintensität abhängig wie das Rezeptorpotenzial (keine
Aktionspotenziale!). Die jeweiligen Anteile dieser beiden Strukturen an der Form
des ERG lassen sich durch eine Betäubung der Erregungsübertragen in die
Lamina darstellen.
* Bei allen abgeleiteten Potenzialen handelt es sich um Summenpotenziale, die extrazellulär
abgeleitet werden.
VERSUCHSAUFBAU
Bevor Sie die Fliege narkotisieren, machen Sie sich mit dem Versuchsaufbau
vertraut. Als Lichtquelle (L) dienen LEDs verschiedener Farben, die in die
Ableitkammer integriert sind. Die Farbe kann durch einen Drehschalter im
Bedienfeld eingestellt werden. Die Lichtleistung wird ebenfalls durch einen
Drehschalter gewählt. Die absolute Lichtleistung bei entsprechenden
Schalterpositionen entnehmen Sie der Tabelle auf der Ableitkammer. Die
Belichtungsdauer (Pulsbreite) kann ebenfalls im Bedienfeld eingestellt werden.
Der Reiz kann entweder als Einzelreiz, als kontinuierliches Licht oder als Abfolge
mit einer wählbaren Frequenz präsentiert werden. Zur Abdunkelung des Objekts
und zur Abschirmung gegenüber elektrischen Störfeldern wird ein Kasten als
Faraday-Käfig über das Präparat geklappt. Der Reiz wird über den Ausgang
„Reiz“ ausgegeben. Verbinden sie diesen mit dem Kanal 2 des PowerLabs. Die
differente (DE) und indifferente (IE) Elektrode sind mit den Eingängen eines in
die Elektronik der Ableitkammer integrierten Differenzverstärkers verbunden, der
die Signale 100x verstärkt (Beachten Sie dies bitte bei der Berechnung Ihrer
ERG-Amplituden). Das Antwortsignal wird über den Ausgang „Antwort“
ausgegeben. Verbinden sie diesen mit Kanal 1 des PowerLabs. Öffnen Sie im
Programm Chart die Datei „ERG_RI-Kennlinien.adiset“ und machen Sie sich unter
Anleitung des/der Tischbetreuer/in mit der Software vertraut. Stellen Sie nun
eine Reizdauer von 500 ms ein und wählen Sie Weißlicht bei einer mittleren
Lichtleistung. Wählen Sie im Bedienfeld die Einstellung Einzelreiz.
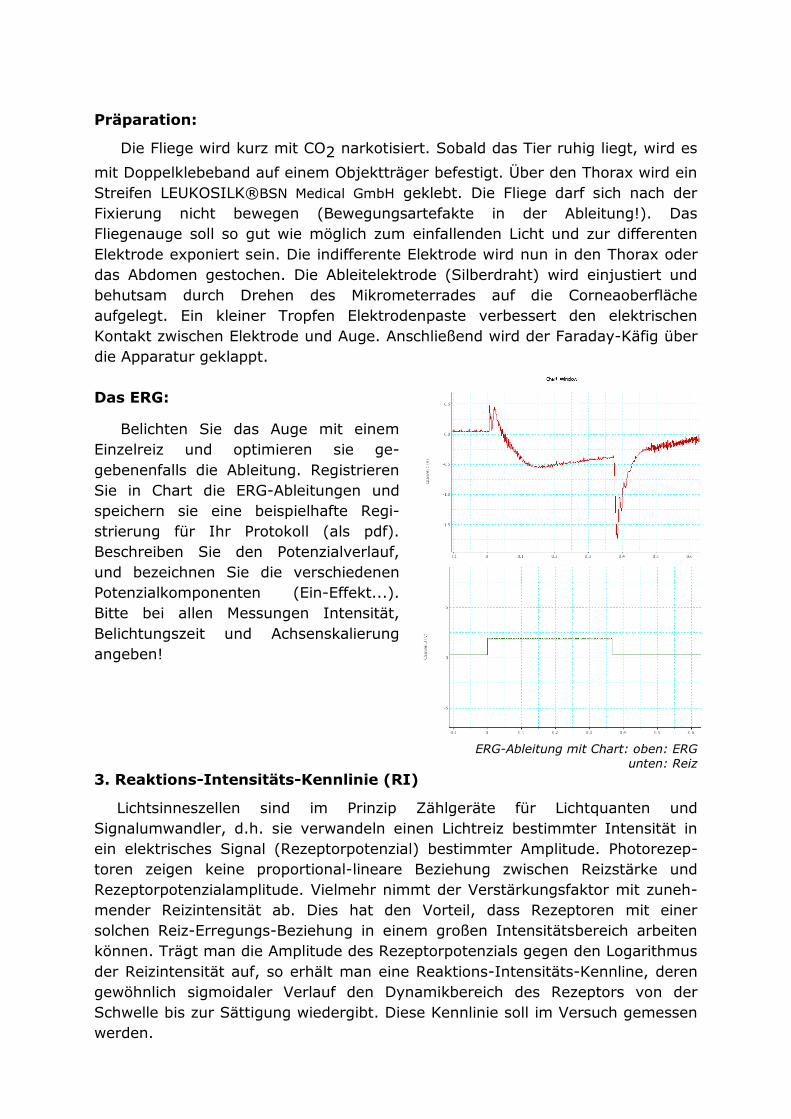
Präparation:
Die Fliege wird kurz mit CO2 narkotisiert. Sobald das Tier ruhig liegt, wird es
mit Doppelklebeband auf einem Objektträger befestigt. Über den Thorax wird ein
Streifen LEUKOSILK®BSN Medical GmbH geklebt. Die Fliege darf sich nach der
Fixierung nicht bewegen (Bewegungsartefakte in der Ableitung!). Das
Fliegenauge soll so gut wie möglich zum einfallenden Licht und zur differenten
Elektrode exponiert sein. Die indifferente Elektrode wird nun in den Thorax oder
das Abdomen gestochen. Die Ableitelektrode (Silberdraht) wird einjustiert und
behutsam durch Drehen des Mikrometerrades auf die Corneaoberfläche
aufgelegt. Ein kleiner Tropfen Elektrodenpaste verbessert den elektrischen
Kontakt zwischen Elektrode und Auge. Anschließend wird der Faraday-Käfig über
die Apparatur geklappt.
Das ERG:
Belichten Sie das Auge mit einem
Einzelreiz und optimieren sie ge-
gebenenfalls die Ableitung. Registrieren
Sie in Chart die ERG-Ableitungen und
speichern sie eine beispielhafte Regi-
strierung für Ihr Protokoll (als pdf).
Beschreiben Sie den Potenzialverlauf,
und bezeichnen Sie die verschiedenen
Potenzialkomponenten (Ein-Effekt...).
Bitte bei allen Messungen Intensität,
Belichtungszeit und Achsenskalierung
angeben!
ERG-Ableitung mit Chart: oben: ERG
unten: Reiz 3. Reaktions-Intensitäts-Kennlinie (RI)
Lichtsinneszellen sind im Prinzip Zählgeräte für Lichtquanten und
Signalumwandler, d.h. sie verwandeln einen Lichtreiz bestimmter Intensität in
ein elektrisches Signal (Rezeptorpotenzial) bestimmter Amplitude. Photorezep-
toren zeigen keine proportional-lineare Beziehung zwischen Reizstärke und
Rezeptorpotenzialamplitude. Vielmehr nimmt der Verstärkungsfaktor mit zuneh-
mender Reizintensität ab. Dies hat den Vorteil, dass Rezeptoren mit einer
solchen Reiz-Erregungs-Beziehung in einem großen Intensitätsbereich arbeiten
können. Trägt man die Amplitude des Rezeptorpotenzials gegen den Logarithmus
der Reizintensität auf, so erhält man eine Reaktions-Intensitäts-Kennline, deren
gewöhnlich sigmoidaler Verlauf den Dynamikbereich des Rezeptors von der
Schwelle bis zur Sättigung wiedergibt. Diese Kennlinie soll im Versuch gemessen
werden.

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 47
Methode:
Das dunkel adaptierte Fliegenauge (ca. 2 min.) wird mit Lichtreizen (500 ms)
verschiedener Intensität gereizt. Das Reizlicht wird, mit der geringsten Intensität
beginnend, stufenweise bis zur maximalen Lichtleistung erhöht. Die absolute
Lichtleistung entnehmen Sie der Tabelle. Messen Sie für jede Lichtintensität 3
Zwischenpotenzialamplituden. Tragen Sie alle Werte in das Datenblatt, das Sie
im Kurs erhalten ein und berechnen Sie den Mittelwert. Zwischen zwei
aufeinander folgenden Reizen sollte mindestens die Zeit von 1 Minute liegen, bei
starken Reizen 2 Minuten, um Adaptationseffekte gering zu halten Speichern sie
für jede Lichtintensität ein exemplarisches ERG für Ihr Protokoll.
Tragen Sie die Amplituden des Zwischenpotenzials in einer halblogarith-
mischen Darstellung gegen die Intensität des Lichtes auf.
Ordinate: ERG-Amplitude in mV (linear)
Abszisse: log. der Reiz-Intensität (Lichtleistung)
Diskutieren Sie den Kurvenverlauf und die allgemeine biologische Bedeutung
von logarithmischen Kennlinien.
4. Spektralempfindlichkeit der Fliegenrezeptoren
Die Absorptionseigenschaften des dioptrischen Apparates und des Photo-
pigments (bzw. mehrerer Sehfarbstoffe) in den Rezeptoren bestimmen die
spektrale Empfindlichkeit des Auges. Der Mensch hat im normal hell adaptierten
Zustand eine Spektralempfindlichkeit von ca. 400 nm bis 750 nm mit einem
Maximum bei 550 nm. Bei Arthropoden ist der sichtbare Teil des Spektrums weit
in den kurzwelligen Bereich verschoben (z.B. Biene: 300 nm bis 650 nm). Sie
können also UV wahrnehmen und sind im Rotbereich blind. Im folgenden Versuch
soll die Spektralempfindlichkeit der Fliegenrezeptoren ermittelt werden. Fliegen
besitzen zwei Rezeptorsysteme, die Grün-Rezeptoren (Maximum bei 515 nm,
Sinneszellen 1-6 eines Ommatidiums) und die Blaurezeptoren (Maximum bei 470
nm, Sinneszellen 7 und 8). Die Grünrezeptoren sind etwas empfindlicher als die
Blaurezeptoren.
Methode:
Die Potenzialamplitude (Zwischenpotenzial) des ERG's wird bei verschiedenen
Wellenlängen und verschiedenen Intensitäten bestimmt. Messen Sie für jede
Farbe die Zwischenpotenzialamplitude des ERG's am Oszilloskop bei 3 ver-
schiedenen Lichtintensitäten. Messen Sie jeden Wert dreimal und berechnen Sie
den Mittelwert. Wählen Sie die Kombination von Farbe und Lichtleistung so, dass
für alle Farben etwa der gleiche Bereich der ERG-Amplitude erreicht wird.
Notieren Sie die Werte in einer Tabelle.

Auswertung:
Tragen Sie für jede Wellenlänge die mittleren Zwischenpotenzialamplituden
(mV) als Funktion der Lichtintensität in ein Diagramm ein (halblogarithmische
Darstellung). Für jede Wellenlänge erhält man also eine Kennlinie.
Aus diesen Diagrammen ermitteln Sie dann für die einzelnen Farben die
Lichtintensitäten, die zur selben Zwischenpotenzialamplitude (z.B. 1 mV, 2 mV, 3
mV) führen. Tragen Sie diese Werte in ein weiteres Diagramm (log Reizintensität
gegen Wellenlänge) ein. Aus den so erhaltenen Isopotenzialkurven ergibt sich die
Farbempfindlichkeit des Tieres.
Vergleichen Sie die Kurve mit der Farbempfindlichkeit des Menschen.
5. Verschmelzungsfrequenz bei verschiedenen Lichtintensitäten
Film und Fernsehen basieren letztendlich auf dem Phänomen, dass der
Mensch oberhalb einer bestimmten Bildfrequenz die Folge stationärer Bilder nicht
mehr getrennt wahrnimmt. Die Einzelbilder verschmelzen miteinander und
vermitteln bei geringfügigen Konfigurationsänderungen von Bild zu Bild den
Eindruck eines Bewegungsablaufes. Diese Fusionsfrequenz, die für einen
zeitlichen Hell- Dunkelwechsel den Eindruck einer kontinuierlichen Beleuchtung
erzeugt wird als Flimmerfusionsfrequenz (kritische Flimmerfrequenz CFF)
bezeichnet. Ihr Wert ist von der Wellenlänge, der Intensität und von der Tierart
abhängig. Bei Wirbeltieren unterscheidet sich die Flimmerfusionsfrequenz
außerdem zwischen Fovea und Peripherie.
5.1 Verschmelzungsfrequenz der Fliege
Öffnen sie in Chart die Datei „ERG_Flimmerfusionsfrequenz.adiset“. Wählen
Sie an der Apparatur eine mittlere Lichtleistung bei Weißlicht und stellen Sie die
Reizeinstellung auf das Symbol für Reizfolge. Registrieren sie nun den fort-
laufenden den Reiz sowie das ERG. Erhöhen Sie mit dem Potentiometerknopf die
Reizfrequenz solange, bis sie keine Korrelation mehr zwischen ERG und Reiz
erkennen können. Wie verändern sich Amplitude und Potenzialverlauf bei
steigender Frequenz? Bestimmen Sie die Verschmelzungsfrequenz ist bei 2 oder
3 Lichtintensitäten. Welche Tendenz ergibt sich?
5.2 Verschmelzungsfrequenz des Menschen
Für diesen Versuch benötigen Sie Apparatur mit einer LED. Verbinden Sie den
Reizausgang der Apparatur mit Kanal 2 des PowerLabs und öffnen Sie die Datei
„Flimmerfusion Mensch.adiset“. Projizieren Sie den Lichtkegel der LED auf ein
weißes Papier und fixieren sie diesen, dass der Lichtkegel auf die Fovea zentralis
fällt, oder schauen Sie direkt in den Lichtstrahl. Wählen Sie zunächst eine
niedrige Intensität und bestimmen Sie die Verschmelzungsfrequenz für Ihren
optischen Sinn. Wiederholen Sie den Versuch bei verschiedenen Lichtintensitäten

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 49
und vergleichen Sie die Werte mit den bei der Fliege gefundenen. Wiederholen
Sie das Experiment im peripheren Gesichtsfeld. Wie unterscheidet sich die
Flimmerfusionsfrequenz zwischen Peripherie und Fovea? Wodurch ist dieser
Unterschied begründet?
6. Blockade der synaptischen Verbindungen zwischen Lamina und
Rezeptor
Um das eigentliche Rezeptorpotenzial sichtbar zu machen, wird der Fliege ein
Tropfen 2%-ige Procainlösung mit Hilfe einer Injektionskanüle in die hintere
Kopfkapselwand appliziert. Von dort diffundiert die Lösung zu den optischen
Ganglien.
Registrieren sie die Veränderungen des ERG's. Speichern Sie die Daten für Ihr
Protokoll. Erklären Sie die Vorgänge (vgl. Absatz 2. Exkurs: Das Elektroretino-
gramm der Fliege).
7. Latenzzeit des Rezeptorsummenpotenzials beim procainbehandelten
Präparat (Wahlaufgabe)
Die Gesamtzeit für alle Sehfarbstoffreaktionen und den nachfolgenden
Erregungsvorgängen an der Membran bis zum Auftreten des Rezeptorpotenzials
nennt man Latenzzeit. Bestimmen Sie bei Weißlicht bei verschiedenen
Intensitäten die Latenzzeit des Rezeptorpotenzials. Spreizen Sie dazu die x-
Achse entsprechend. Wie unterscheiden sich die Ergebnisse bei verschiedenen
Intensitäten? Warum?
Untersuchen Sie die Latenzzeit für violett (420 nm) und rot (660 nm).
Wählen Sie die Lichtintensität so, dass Sie vergleichbare ERG-Amplituden
erhalten.

ERFOLGSKONTROLLE
Nach diesem Praktikumsteil sollten Sie in der Lage sein:
zu beschreiben, welche Eigenschaften das Rezeptorpotenzial kennzeichnet,
wie es sich vom Aktionspotenzial unterscheidet, wie sich eine intrazelluläre
Ableitung von einer extrazellulären Ableitung unterscheidet,
die Beziehung zwischen Reiz, Rezeptorpotenzial und Impulsfolgefrequenz
wiederzugeben,
die im Kurs verwendeten Messmethoden zu beschreiben,
den Entstehungsmechanismus des diphasischen Fliegen-ERG's zu erläutern
und anzugeben, welche Komponenten daran beteiligt sind,
eine Möglichkeit anzugeben, die Komponenten des ERG's zu trennen, um
das Rezeptorsummenpotenzial darzustellen,
zu erläutern, was man unter der Kennlinie eines Rezeptors versteht und
einige verschiedene Kennlinienverläufe anzugeben,
die Unterschiede in der Spektralempfindlichkeit zwischen Insekten und
Menschen aufzuzeigen und ihre Ursachen zu nennen,
die im Kurs gemessenen Kurvenverläufe zu skizzieren,
zu beschreiben, wovon das räumliche Auflösungsvermögen des Menschen
und der Fliege abhängt und die ungefähren Werte angeben zu können,
das ungefähre zeitliche Auflösungsvermögen von menschlichem Auge und
Fliegenauge zu nennen und zu erläutern, wie es sich bei geringerer oder
größerer Beleuchtungsstärke verhält,
die Unterschiede im zeitlichen Auflösungsvermögen beim Menschen
zwischen Fovea und Peripherie erläutern zu können und
den prinzipielle Aufbau vom menschlichem Auge und vom Insektenauge
darzustellen und die Funktion der einzelnen Komponenten zu erläutern.
TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 51
4. P S Y C H O P H Y S I K, A U G E N B E W E G U N G E N
und R E F L E X E
Psychophysik
Die Psychophysik ist die Lehre von der
Quantifizierbarkeit von Wahrnehmungsleistungen und
etwa erst 150 Jahre alt. Als ein Begründer der modernen
Psychophysik gilt G.T. Fechner der als Physiker die
exakten Messmethoden der Physik für die Wahr-
nehmungsforschung nutzbar machte. Bis dahin widmete
man sich der Wahrnehmung v. a. philosophisch bzw.
durch die Introspektion. Die Grundidee der Psychophysik
ist die, einen Probanden nach seiner Empfindung zu
physikalisch exakt definierten Reizen zu befragen. Da
keine direkten Messungen vorgenommen werden können
und immer Proband oder Versuchstier über ihre
Empfindung Auskunft geben müssen, wird diese Mess-
methodik als subjektive Sinnesphysiologie bezeichnet.
Dem gegenüber steht die objektive Sinnesphysiologie, bei der Erregungen von
Zellen oder Rezeptoren gemessen werden (z.b. Insekten ERG). Am heutigen
Kurstag werden Sie Schwellenmessungen für das menschliche Hören erheben.
Reflexe
Bestimmte Reize führen zu einer direkten motorischen
Reaktion. Dieser Zusammenhang zwischen Reiz und
Reaktion wird als Reflex be-schrieben. Unwillkürliche,
mit kurzer Latenz ab-laufende Reflexe spielen
besonders für die Regulation der Körperhaltung und
die Orientierung im Raum eine große Rolle.
Beispielhaft wird im Praktikum der
Patellarsehnenreflex unter-sucht.

THEORETISCHE VORAUSSETZUNGEN
Folgende Kenntnisse aus der Vorlesung "Einführung in die Tierphysiologie" und
aus den Lehrbüchern werden für die sinnvolle Durchführung der Versuche be-
nötigt und daher vorausgesetzt:
Grundlagen der Psychophysik: Sinnesmodalität, Reizqualität, Reizintensität,
adäquater Reiz, absolute Schwelle, Unterschiedsschwelle, Weber-Fechner-
Gesetz, Stevens'sche Potenzfunktion, Forced-Choice Paradigma.
Physikalische Grundlagen: Schallwellen, Schalldruck, Dezibel (dB) als Maß-
einheit, dB SPL, Frequenz, Oktave, spektrale Zusammensetzung von
Schallereignissen (Ton, Klang, Geräusch).
Grundlagen der Hörphysiologie: Aufbau des Ohres beim Menschen,
Frequenzrepräsentation in der Cochlea, Schalldruckpegel, Lautstärke,
Lautheit, Phon, dB(A), Verlauf der menschlichen Hörschwelle,
Wanderwellentheorie (Georg von Békésy) Hauptsprachbereich,
Richtungshören - Bedeutung von Laufzeit- und Schalldruckunterschieden an
den beiden Ohren, horizontale und vertikale Schallokalisation.
Motorische Rückenmarksreflexe: Elemente des Kniesehnenreflexes, Aufbau
des Rückenmarks, afferente und efferente Bahnen sowie spinale
Interneurone (Transmitter), Muskelspindeln und ihre efferente Kontrolle,
monosynaptische und polysynaptische Reflexe, Eigenreflex, Fremdreflex,
Regelung durch negative Rückkopplung.
Augenbewegungen: extraokuläre Muskeln, Hirnnerven, Gleichgewichtssinn,
Innenohr, Retina, visueller Kortex, extrastriärer Kortex, Sakkaden,
elementare Bewegungsdetektoren, Bewegungssehen, bedingter und
unbedingter Reflex
LITERATUR
SCHMIDT/THEWS -- Physiologie des Menschen, Springer. Behandelt in hervor-
ragender Weise alle für diesen Versuch nötigen theoretischen Grundlagen
SILBERNAGL/DESPOPOULOS: Taschenatlas der Physiologie
HANDWERKSZEUG
Protokollheft
Schreibzeug
weite Beinkleidung für Kniesehnenreflex

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 53
VERSUCHSTEIL I : PSYCHOPHYSIK
Experiment Ia: Bestimmung der absoluten Hörschwelle des Menschen
Der Versuchsperson wird bei verschiedenen Frequenzen ein Dauerton mit
variablem Schalldruck (dB) vorgespielt. Bei jeder gemessenen Frequenz wird der
Schalldruckpegel notiert, bei dem ein Ton gerade gehört werden kann. Dadurch
wird die Hörkurve des Probanden ermittelt.
Versuchsdurchführung:
Mit dem Hörschwellenmessgerät wird ein Sinuston variablen Schalldruck-
pegels und variabler Frequenz erzeugt und auf einem Kopfhörer ausgegeben, den
die Versuchsperson trägt.
Bestimmen Sie bei jeder Frequenz den Schwellenwert durch eine adaptive
„staircase-Prozedur“. Dazu verändern Sie bei konstanter Frequenz den Schall-
druckpegel zunächst in groben Schritten so lange, bis Sie den Schwellenbereich
für die jeweilige Frequenz gefunden haben. Dazu erhöhen Sie den SPL, wenn der
Ton nicht gehört wird, und erniedrigen den SPL, wenn der Ton gehört wird.
Engen Sie diesen Bereich weiter ein, indem Sie mit feinerer SPL-Änderung die
Schwelle besser bestimmen. Ermitteln Sie nun den genauen Wert, indem Sie in 1
oder 0.5 dB-Schritten leiser bzw. wieder lauter stellen, und notieren Sie den
Wert, bei dem die Versuchsperson den Ton gerade noch wahrnimmt. Gehen Sie
dabei folgendermaßen vor: Antwortet Ihre Versuchsperson zweimal hin-
tereinander korrekt, so verringern Sie den Schalldruckpegel. Antwortet Ihre
Versuchsperson einmal falsch, erhöhen Sie ihn wieder. Notieren Sie den
niedrigsten SPL für diese Frequenz, bei welcher die Versuchsperson gerade noch
2 mal korrekt geantwortet hat.
Achten Sie auch darauf, dass die Versuchsperson schnell und ohne zu
überlegen antwortet. Stellen Sie im Interesse der Versuchsperson vor jedem
Frequenzwechsel den Schalldruckpegel auf einen niedrigen dB-Wert ein.
Stellen Sie nun die Frequenz auf ca. 20 kHz und höchsten Schalldruck (etwa
80 dB) ein. Erniedrigen Sie nun langsam die Reizfrequenz. Welche höchste
Frequenz kann unter den gegebenen Versuchsbedingungen gerade noch gehört
werden?

Technische Anleitung zur Messung der Hörschwelle
Achtung: nach Versuchsende unbedingt den Kopfhörer ausstecken!!
Menüschalter A (gelb)
Menüschalter B (blau)
Das Gerät wird eingeschaltet, indem der Kopfhörer seitlich eingestöpselt wird.
Durch Drücken des Menüschalters A kann zwischen folgenden Menüs
gewechselt werden.
Menü 1 Pulsetime
Mute Ch
Bestimmt die Dauer des Testtons. Die Einstellung „None“ bewirkt einen Dauerton. Zur Bestimmung der Hörschwelle stellen Sie einen Sinuston mit einer Tonlänge von 500ms ein
(außer bei oberer Hörschwelle, s. Skript). Auf „Right“ stellen: Nur ein Ohr messen
Menü 2 Light
Für das Display kann ein Licht eingeschaltet werden Headphone bleibt auf 84 SPL/V
Menü 3 Frequenz
Lautstärke
Die Lautfrequenz wird verändert
Der Schalldruckpegel wird verändert
In jedem Menü können Werte verändert werden. Dazu betätigt man die
Drehscheibe. Die Position des Pfeils wird mit dem Menüschalter B verändert.
Auf Wunsch kann die Auswertung der Experimente mit Excel erfolgen. Hierzu
steht ein Rechner zur Verfügung. Tabellen, in die Sie Ihre Messdaten eintragen
sind vorbereitet. Die Abbildungen bzw. Tabellen können auf einem USB-Stick
mitgenommen werden.
Achten Sie bitte darauf, dass die Versuchsperson das Display des Hörschwellen-
messgeräts nicht sieht. Die Versuchsperson kann z.B. durch Handzeichen an-
geben, ob der Ton gehört wurde.
[Gebe
n Sie
ein
Zitat
aus
dem
Doku
ment
oder
die
Zusam
menfa
ssung
eines
interes
santen
Punkte
s ein.
Sie
könne
n das
Textfe
ld an
einer
beliebi
gen
Stelle
im
Doku
ment
positio
nieren.
Verwe
nden
Sie die
Regist
erkarte
'Textfe
ldtools
',
wenn
Sie
das
Forma
t des
Textfe
lds
'Textzi
tat'
ändern
möcht
en.]

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 55
Ermitteln Sie die Hörschwellen (in dB) für folgende Frequenzen:
Frequenz Schalldruckpegel
1 kHz
500 Hz
200 Hz
100 Hz
50 Hz
20 Hz
Stellen Sie zu Beginn der nächsten Messung den Schalldruck wieder auf etwa 0dB!!!
Frequenz Schalldruckpegel
2 kHz
5 kHz
10 kHz
15 kHz
Bestimmung der höchsten, wahrnehmbaren Frequenz
Stellen Sie einen Dauerton (Pulsetime „none“) auf den Schalldruck von 80 dB
ein, die Frequenz auf über 20 kHz. Welches ist die höchste, wahrnehmbare
Frequenz für die Versuchsperson? Ermitteln sie diesen Wert durch Veränderung
der Frequenz.
Kopfhörer ausstecken!
Auswertung:
Tragen Sie die auf diese Weise gemessene Schwellenkurve graphisch über
einer logarithmischen Frequenzskala auf. Ergänzen Sie diese Abbildung durch
den Verlauf der durchschnittlichen menschlichen Hörschwellenkurve (z.B. aus
Lehrbuch). Tragen Sie auch Ihre obere Hörgrenze (höchste wahrgenommene
Frequenz) in das Audiogramm ein.

Experiment Ib: Akustische Richtungslokalisation
- Bestimmung der binauralen Zeitdifferenzschwelle
Um die horizontale Richtung einer Schallquelle zu bestimmen, benutzt der
Mensch (bei Frequenzen über ca. 1 kHz) sowohl die Intensitäts- als auch die
Laufzeitunterschiede, welche der Schall beim Auftreffen auf die beiden Ohren
hat. Im folgenden Versuch soll geklärt werden, wie gering diese Unterschiede
sein können, damit sie immer noch als Abweichung von der Vorausrichtung em-
pfunden werden, und wie die relative Gewichtung der beiden Parameter
Schalldruckunterschied und Laufzeitdifferenz für die Richtungsempfindung ist.
Für diese Messungen steht Ihnen eine Messapparatur zur Verfügung, mit der
für einen binauralen Reiz der Zeitversatz (in µs) zwischen rechtem und linkem
Ohr sowie (für 2. Versuchsteil) die Schalldruckpegeldifferenz (dB) eingestellt
werden kann. Der Reiz für das rechte Ohr wird jeweils konstant gehalten, der
Reiz für das linke Ohr variiert. Die Zeitverzögerung des Reizes auf dem rechten
Ohr beträgt konstant 1000 µs und der relative Schalldruckpegel 0 dB. Die Zeit-
verzögerung des linken Reizes kann eingestellt werden; der Schalldruckpegel auf
dem linken Ohr kann in Schritten von 1.5 dB relativ zum rechten Ohr variiert
werden.
Versuchsdurchführung:
Über einen Kopfhörer wird ein Klickreiz auf beiden Ohren gegeben. Der Klick ist
auf beiden Ohren gleich laut, die Schalldruckpegeldifferenz wird dazu konstant
auf 0 dB eingestellt. Variiert wird der relative Zeitversatz des Klickens zwischen
den beiden Ohren, er wird auf dem linken Ohr in Schritten von 20 µs zwischen -
100 µs und +100 µs relativ zum rechten Ohr eingestellt (links eingestellte,
absolute Zeitverzögerung 900 µs – 1100 µs). Die Versuchsperson muss benen-
nen, ob sie den Klicklaut "rechts" oder "links" gehört hat, die Angabe "Mitte" ist
nicht zugelassen ("forced choice"). Versuchen Sie, die Messung schnell
durchzuführen; die Versuchsperson sollte nicht lange überlegen. Es werden in
zufälliger Reihenfolge Klickreize mit den genannten Zeitdifferenzen vorgespielt,
bis für jede angegebene Zeitdifferenz 10 Messwerte ("rechts" oder "links")
vorliegen.
Technische Anleitung zur binauralen Zeitdifferenzschwelle (Abb. s. nächste Seite).
Netzstecker bitte einstöpseln.
Mit diesem Gerät können Sie kontrolliert Klicklaute für das rechte bzw. linke Ohr
erzeugen. Durch Einstellung der Zeitverzögerung verändern Sie immer nur den Reiz für
das rechte Ohr, der Reiz für das linke Ohr bleibt jeweils konstant. Dieser wird mit einer
Zeitverzögerung von 1000µSekunden präsentiert.
Achten Sie bitte darauf, dass die Versuchsperson die Einstellungen am Reizgerät nicht
sehen kann. Die Reize sollten in zufälliger Reihenfolge präsentiert werden.
Achtung: Den Kopfhörer richtig aufsetzen, 900 µs sollte rechts empfunden werden. SPL
auf 0 dB stellen.
Der Kippschalter oben links muss auf „leise“ eingestellt sein. In den Zahlfeldern in denen
der Schalldruckpegel angezeigt wird, können nur ganze Zahlen dargestellt werden.
Betätigt man den „+-Schalter“, so erhöht sich der Schalldruck jeweils um 1.5dB. Die
Nachkommazahl wird durch die kleine rote Lampe angegeben.

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 57
LED: leuchtet beim
Klicken
Zeitverzöge-
rung linker Klicklaut
Antwort der Versuchsperson
Summe
Antwort „rechts“
Summe
Antwort „links“
Zeit in µs 1 2 3 4 5 6 7 8 9 10
900
920
940
960
980
1000
1020
1040
1060
1080
1100
Pulsgeber: Mit diesem Knopf wird Klicklaut ausgelöst
Rotes Lämpchen (zeigt Nachkommastelle 0.5dB )
N
Rädchen, mit denen die Zeitverzögerung des rechten Lautsprechers eingestellt wird (1000 µs = Mitte)
Erhöht bzw. erniedrigt den Schalldruck um jeweils 1.5 dB dB
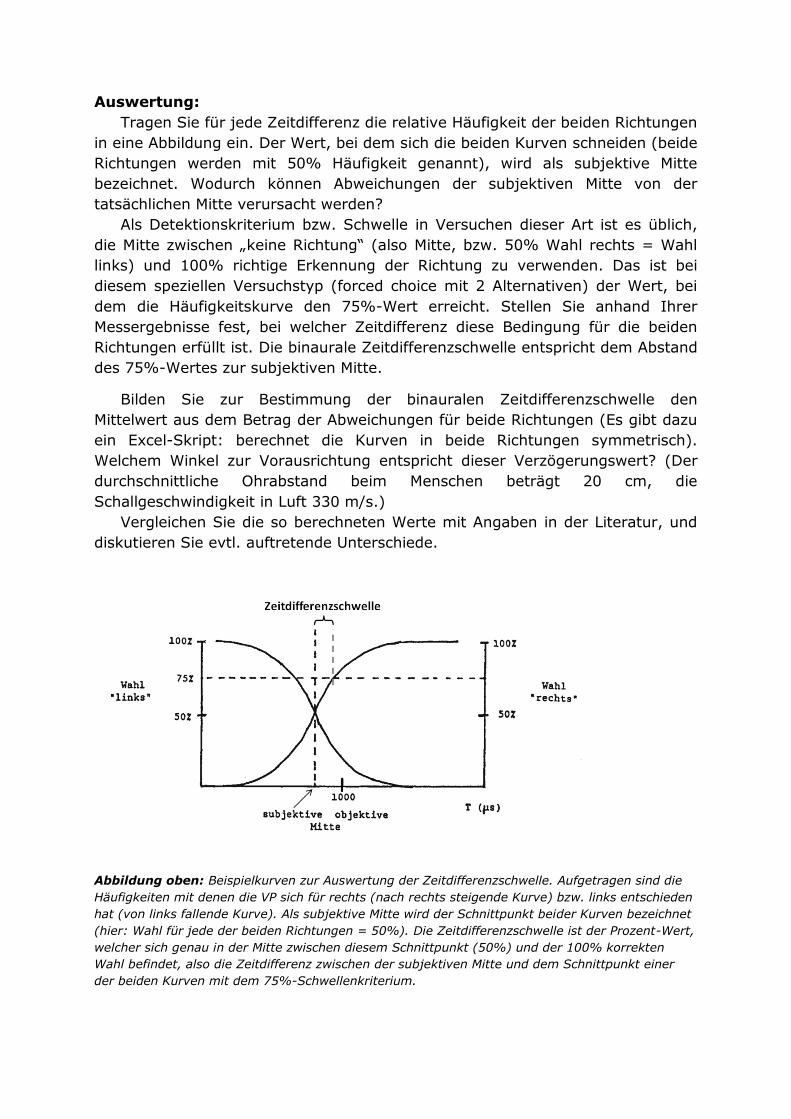
Auswertung:
Tragen Sie für jede Zeitdifferenz die relative Häufigkeit der beiden Richtungen
in eine Abbildung ein. Der Wert, bei dem sich die beiden Kurven schneiden (beide
Richtungen werden mit 50% Häufigkeit genannt), wird als subjektive Mitte
bezeichnet. Wodurch können Abweichungen der subjektiven Mitte von der
tatsächlichen Mitte verursacht werden?
Als Detektionskriterium bzw. Schwelle in Versuchen dieser Art ist es üblich,
die Mitte zwischen „keine Richtung“ (also Mitte, bzw. 50% Wahl rechts = Wahl
links) und 100% richtige Erkennung der Richtung zu verwenden. Das ist bei
diesem speziellen Versuchstyp (forced choice mit 2 Alternativen) der Wert, bei
dem die Häufigkeitskurve den 75%-Wert erreicht. Stellen Sie anhand Ihrer
Messergebnisse fest, bei welcher Zeitdifferenz diese Bedingung für die beiden
Richtungen erfüllt ist. Die binaurale Zeitdifferenzschwelle entspricht dem Abstand
des 75%-Wertes zur subjektiven Mitte.
Bilden Sie zur Bestimmung der binauralen Zeitdifferenzschwelle den
Mittelwert aus dem Betrag der Abweichungen für beide Richtungen (Es gibt dazu
ein Excel-Skript: berechnet die Kurven in beide Richtungen symmetrisch).
Welchem Winkel zur Vorausrichtung entspricht dieser Verzögerungswert? (Der
durchschnittliche Ohrabstand beim Menschen beträgt 20 cm, die
Schallgeschwindigkeit in Luft 330 m/s.)
Vergleichen Sie die so berechneten Werte mit Angaben in der Literatur, und
diskutieren Sie evtl. auftretende Unterschiede.
Abbildung oben: Beispielkurven zur Auswertung der Zeitdifferenzschwelle. Aufgetragen sind die
Häufigkeiten mit denen die VP sich für rechts (nach rechts steigende Kurve) bzw. links entschieden
hat (von links fallende Kurve). Als subjektive Mitte wird der Schnittpunkt beider Kurven bezeichnet
(hier: Wahl für jede der beiden Richtungen = 50%). Die Zeitdifferenzschwelle ist der Prozent-Wert,
welcher sich genau in der Mitte zwischen diesem Schnittpunkt (50%) und der 100% korrekten
Wahl befindet, also die Zeitdifferenz zwischen der subjektiven Mitte und dem Schnittpunkt einer
der beiden Kurven mit dem 75%-Schwellenkriterium.
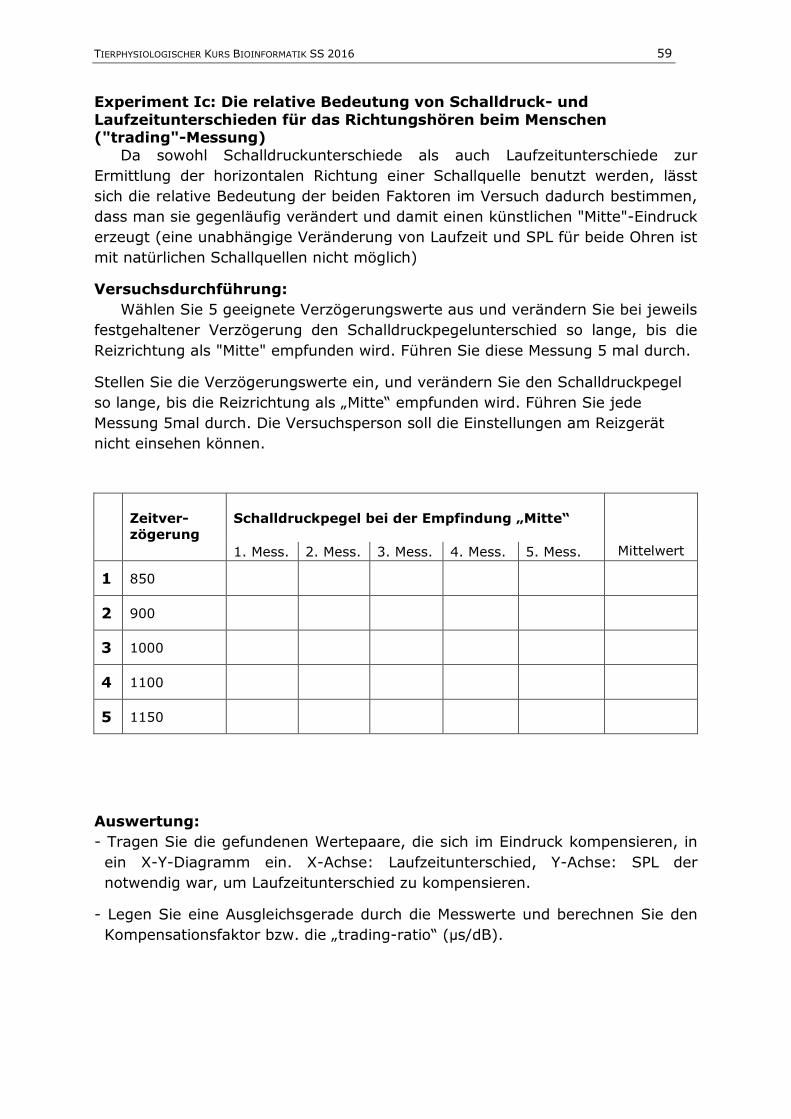
TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 59
Experiment Ic: Die relative Bedeutung von Schalldruck- und
Laufzeitunterschieden für das Richtungshören beim Menschen ("trading"-Messung)
Da sowohl Schalldruckunterschiede als auch Laufzeitunterschiede zur
Ermittlung der horizontalen Richtung einer Schallquelle benutzt werden, lässt
sich die relative Bedeutung der beiden Faktoren im Versuch dadurch bestimmen,
dass man sie gegenläufig verändert und damit einen künstlichen "Mitte"-Eindruck
erzeugt (eine unabhängige Veränderung von Laufzeit und SPL für beide Ohren ist
mit natürlichen Schallquellen nicht möglich)
Versuchsdurchführung:
Wählen Sie 5 geeignete Verzögerungswerte aus und verändern Sie bei jeweils
festgehaltener Verzögerung den Schalldruckpegelunterschied so lange, bis die
Reizrichtung als "Mitte" empfunden wird. Führen Sie diese Messung 5 mal durch.
Stellen Sie die Verzögerungswerte ein, und verändern Sie den Schalldruckpegel
so lange, bis die Reizrichtung als „Mitte“ empfunden wird. Führen Sie jede
Messung 5mal durch. Die Versuchsperson soll die Einstellungen am Reizgerät
nicht einsehen können.
Zeitver-
zögerung
Schalldruckpegel bei der Empfindung „Mitte“
1. Mess. 2. Mess. 3. Mess. 4. Mess. 5. Mess. Mittelwert
1 850
2 900
3 1000
4 1100
5 1150
Auswertung:
- Tragen Sie die gefundenen Wertepaare, die sich im Eindruck kompensieren, in
ein X-Y-Diagramm ein. X-Achse: Laufzeitunterschied, Y-Achse: SPL der
notwendig war, um Laufzeitunterschied zu kompensieren.
- Legen Sie eine Ausgleichsgerade durch die Messwerte und berechnen Sie den
Kompensationsfaktor bzw. die „trading-ratio“ (μs/dB).

VERSUCHSTEIL II: REFLEXE
Dehnungsreflexe und Vestibularreflexe sind ganz wesentlich an der Kontrolle
unserer Körperhaltung unter wechselnden Einflüssen beteiligt. Im Folgenden wird
eine vergleichende Messung des Kniesehnenreflexes durchgeführt.
Experiment IIa: Messung der Bruttoreflexzeit des Patellarsehnenreflexes
Dehnungsreflexe ermöglichen bereits auf spinalem Niveau und ohne die
Notwendigkeit von Willkürbewegungen die Kompensation von Störungen einer
stabilen Position. Dehnungsreflexe lassen sich in der klinischen Praxis leicht und
reproduzierbar auslösen und sind damit eine wichtige neurologische
Untersuchungsmethode. Mit einem Reflexhammer wird durch einen leichten
Schlag auf die entsprechende Sehne der Muskel gedehnt und damit eine
Reflexkontraktion ausgelöst. Z.B. beim Kniesehnenreflex (Patellarsehnenreflex)
führt ein Schlag auf die Sehne unterhalb der Kniescheibe zu einer Kontraktion
des Streckers im Oberschenkel und damit zum Heben des unbelasteten
Unterschenkels. An dieser Reaktion lassen sich die Eigenschaften eines
monosynaptischen, spinalen Reflexes gut beschreiben. Als Maß für die Stabilität
eines solchen Reflexes lässt sich die Varianz der Reflexzeit verwenden.
Modulierende Einflüsse durch andere motorische Aktivitäten können ebenfalls
anhand der Reflexzeit nachgewiesen werden.
Die Bruttoreflexzeit ist der Zeitraum zwischen dem Einfluss eines Reizes und
dem Beginn der hierdurch ausgelösten motorischen Reaktion. Sie umfasst also
die Zeiten, die für die sensorische Verarbeitung, Erregungsleitung, Verschaltung
im ZNS und die Muskelkontraktion nötig sind. Am Beispiel des
Patellarsehnenreflexes (PSR) soll diese Reflexzeit unter verschiedenen
Randbedingungen untersucht werden.
Die Messung der Bruttoreflexzeit des PSR erfolgt durch eine elektronische
Stoppuhr. Die Uhr wird durch den Schlag mit dem Reflexhammer auf die Sehne
gestartet (getriggert) und durch die reflektorische Bewegung des Unterschenkels
wieder gestoppt.
Versuchsdurchführung:
Die Versuchsperson sitzt entspannt auf einem hohen Stuhl oder Tisch; der
Unterschenkel muss frei hängen. Schließen Sie nun den elektrischen Zeitnehmer
an. Hierzu muss die Versuchsperson an der Hand geerdet werden (elektrische
Verbindung mit der schwarzen Buchse des Zeitnehmers). Hinter ihrer
Ferse/Achillessehne wird ein Stativ mit einem Elektrokontakt so aufgestellt, dass
der Kontakt zwischen Ferse und Stativ in Ruhestellung geschlossen ist
(Verbindung mit der blauen Buchse des Zeitnehmers). Der Reflexhammer wird
mit der gelben Buchse des Zeitnehmers verbunden. Das Knie muss für den
elektrischen Kontakt mit dem Hammer frei sein.

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 61
Drücken Sie vor jeder Messung den "Reset"-Knopf. Der Zeitnehmer wird
genullt und die "Ready"-Lampe leuchtet, wenn ein guter Kontakt an Erdung und
Ferse besteht. Mit einem leichten Schlag auf die Patellarsehne lösen Sie den
Reflex aus. Durch den Reflexhammer wird zu diesem Zeitpunkt der Zeitnehmer
gestartet; die Zeit zwischen Hammer“schlag“ und Bewegung des Beines wird
gemessen. Diese Zeit nennt man die Bruttoreflexzeit.
Führen Sie die folgenden Messung jeweils 16 mal durch.
1. Bestimmen Sie die Bruttoreflexzeit in Ruhe. Die Versuchsperson sollte
hierzu entspannt sitzen.
2. Bestimmen Sie die Bruttoreflexzeit bei Belastung. Die Versuchsperson
spannt einen Expander. Während der Spannung wird der Reflex ausgelöst.
3. Messen Sie die Reaktionszeit für eine bewusst ausgelöste Bewegung des
Unterschenkels. Dazu wird das Knie seitlich nur leicht mit dem
Reflexhammer berührt und die Versuchsperson streckt das Bein, sobald sie
diese Berührung spürt (Augen zu!).
Technische Durchführung:
1. Auf die Ferse und die Spitze des Hammers Elektrodenpaste auftragen. Dies muss
im Verlauf des Experiments evtl. wiederholt werden.
2. Die Versuchsperson nimmt das schwarze Kabel in die Hand. Ist der Kontakt gut,
leuchtet nach Drücken des Reset-Knopfes die „Ready“- Lampe auf. Falls das nicht
der Fall ist muss der Kontakt verbessert werden: ggf. mehr Elektrodenpaste
verwenden, ggf. Schleifpapier verwenden. Nochmals Reset drücken.
3. Die folgenden drei Versuchen 16 mal durchführen und die Reaktionszeiten
aufschreiben. Vor jeder Messung muss Ready leuchten. Versuchsbeschreibung s.o.
Versuch 1: Bruttoreflexzeit des Kniesehnenreflexes in Ruhe
Versuch 2: Bruttoreflexzeit bei Belastung
Versuch 3: Reaktionszeit bewusste Bewegung
4. a. Elektrodenpaste bitte wegputzen
b. Kniesehnenreflexkästchen ausschalten und an das Ladegerät hängen
Messung Versuch 1 Versuch 2 Versuch 3
Zeiten Rang
Vgl. Ve. 2
Rang
Vgl. Ve. 3 Zeiten
Rang
Vgl. Ve. 1 Zeiten
Rang
Vgl. Ve. 1
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
Mittelwert
Rangsum. Rangsum. Rangsum. Rangsum.
Standard-
abweichung
Kniesehnenreflexkästchen ausschalten und an Ladegerät hängen!

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 63
Auswertung:
Berechnen Sie für jede der drei Messungen jeweils den Mittelwert und die Standardabweichung. Die Standardabweichung ist (i.A.) ein stabiles Maß für die
Schwankungen eines Messwertes. Eine kleine Standardabweichung besagt also, dass ein Messwert sehr reproduzierbar ist.
Überprüfen Sie mit einem geeigneten statistischen Testverfahren (Wilcoxon-Test,
s.u.), ob sich die gemessenen Reaktionszeiten (1) beim Reflex in Ruhe und unter
Belastung bzw. (2) beim Reflex in Ruhe und bei willkürlicher Bewegung
signifikant voneinander unterscheiden.
Der Wilcoxon-Test (anderer Name: Man-Whitney-U-Test) ist ein
statistischer Test, der unabhängig von der Verteilung der Daten verwendet
werden kann (im Gegensatz zum t-Test, bei welchem oft Normalverteilung
vorausgesetzt ist). Wenn z.B. die Belastung in unserem Experiment keinen
systematischen Einfluss auf die Reflexzeit hat, dann erwartet man, dass die
beiden Mittelwerte ähnlich sind; exakt gleich werden sie nie sein, das wäre ein
großer Zufall. Wenn aber die Belastung einen systematischen Einfluss hat, dann
sollten die beiden gemessenen mittleren Reflexzeiten signifikant verschieden
sein, nach statistischer Testung also mit einer Irrtumswahrscheinlichkeit < 5%
bzw. < 0.05.
Beim Wilcoxon-Test werden die beiden zu vergleichenden Datensätze (à
16 Daten) „in einen Topf geworfen“, dann bekommen sie gemeinsame Ränge (im
Beispiel von Rang 1 bis Rang 32). D.h., die niedrigste Reaktionszeit der beiden
Datensätze bekommt den Rang 1, die nächstniedrigste den Rang 2, usw., die
längste Reaktionszeit den Rang 32.
Wenn zwei Reaktionszeiten gleich sind: z.B. 193 ms hat den Rang 4, dann
kommen die Zeiten 196 ms und 196 ms. In diesem Fall sollen auf die beiden
Werte die Ränge 5 und 6 verteilt werden, dies macht man, indem man die zu
vergebenden Ränge mittelt, und den mittleren Rang (5.5 in diesem Beispiel)
jedem der beiden Reaktionszeiten zuweist.
Wenn 3 oder mehr Reaktionszeiten gleich sind: Man verfährt analog, d.h.
man mittelt die 3 oder mehr Ränge, und weist jedem der Messwerte den
gemittelten Rang zu.
Wenn man die beiden Rangsummen vergleicht, dann sollten diese fast
gleich sein, wenn sich die Mittelwerte nicht systematisch unterscheiden. Im
Beispiel hat man 32 Werte in einen Topf geworfen, Ränge von 1 bis 32 vergeben,
deren Summe ergibt 528. (Probe: die Summe der beiden Rangsummen im
Vergleich sollte 528 sein). Bei exakter Gleichheit erwartet man also eine
Rangsumme von 264 für jede der beiden Gruppen (da in jede Gruppe gleich viele
Werte eingehen). Exakte Gleichheit findet man in der Realität fast nie vor, d.h.
es gibt immer kleine Rangsummenunterschiede. Bis zu einer Grenze von 212-
316 nimmt man an, dass diese Unterschiede nicht signifikant sind
(Nullhypothese). Wenn aber eine der Rangsummen außerhalb dieses kritischen
Bereichs liegt, dann nimmt man einen signifikanten Unterschied an
(Alternativhypothese). Die Irrtumswahrscheinlichkeit ist in diesem Fall 0.05 (oder
5%). Bemerkung: die Grenze 212-316 gilt nur für den Fall von genau je 16
Datenpunkten in jeder Gruppe, sie wurde also für diesen Praktikumsversuch aus
der Tabelle entnommen.
Interpretieren Sie das Ergebnis!

VERSUCHSTEIL III Augenbewegungen
Zu Beginn des 20. Jahrhundert wurde eine erste, mittlerweile klassisch
gewordene Einteilung der Augenbewegungen eines Menschen in 5 verschiedene Klassen vorgenommen (Dodge 1903):
1) Sakkaden: schnelle, ruckartige Augenbewegungen, die den Blick von einem zum anderen Fixationsziel bringen.
2) Glatte Augenfolgebewegungen: Augenbewegungen, die in ihrer Geschwindigkeit exakt an die Geschwindigkeit eines bewegten Objekts angepasst sind. Diese Augenbewegungen führen dazu, dass das retinale Bild eines bewegten Objekts stationär in der Fovea zentralis ruht.
3) Vestibulo-okulärer Reflex (VOR): Die Bewegung des Kopfes führt zu einer Stimulation des Gleichgewichtsorgans im Innenohr, was wiederum zu einer stabilisierenden Augenbewegung führt. Das heißt, die Bewegung der Augachsen gleicht exakt die Bewegung des Kopfes aus, der Blick im Raum bleibt somit stationär.
4) Optokinetischer Reflex (OKR): Bewegen sich große Anteile des Gesichtsfeldes kohärent, d.h. mit identischer Geschwindigkeit und gleicher Richtung, unter natürlichen Umständen geschieht dies nur bei Eigenbewegung, so werden ebenfalls kompensatorische Augenbewegungen ausgelöst, die das Bild der bewegten Umwelt auf der Retina stabilisieren.
5) Vergenzbewegungen: Werden Blickziele in unterschiedlicher Tiefe im Raum fixiert, so müssen Augenbewegungen ausgeführt werden, die den Winkel zwischen den Sehachsen des linken und des rechten Auges ändern.
Diese Einteilung der Augenbewegung war rein phänomenologisch bedingt, wesentlich sinnvoller erscheint es heute, einen funktionalen Aspekt zu berücksichtigen und deshalb die Augenbewegungen in blickstabilisierende und blickführende Bewegungen einzuteilen. Zu den stabilisierenden Augenbewegungen kann der OKR und VOR gezählt werden, die blickführenden Bewegungen setzen sich aus Sakkaden, Glatten Folgebewegungen und Vergenzbewegungen zusammen. Versuchsaufbau
Es steht ein Infrarot-Messystem (IRIS) zur Aufzeichnung der horziontalen und vertikalen Position des Auges einer Versuchsperson zur Verfügung. Das Prinzip der Infrarot-Okulographie ist in der Abbildung 5 gezeigt. Es wird infrarotes Licht auf das Auge einer Versuchsperson gestrahlt und aus der Intensität der Reflexion an zwei verschiedenen Punkten kann die Position des Blickes bestimmt werden. Aus Gründen der Einfachheit wird lediglich die horizontale Position des linken Auges gemessen.

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 65
Abb. 5: Grundprinzip der Infrarot-Okulographie
Um reflektorsiche Augenbewegungen auszuschließen, die aus einer möglichen Kopfbewegung resultieren würden, wird der Kopf der Versuchsperson durch eine Kinnauflage sowie durch ein Stirnbrett immobilisiert. Ein Bildschirm befindet sich in 57 cm (warum gerade 57?) Entfernung vor der Versuchsperson, über den die visuelle Stimulation erfolgt. Der Ausgang des IRIS Systems ist einmal mit einem Oszilloskop verbunden und gleichzeitig mit dem Analog-Digital-Konverter eines Computers verbunden. 1. Linearität der IR-Okulographie
Auf dem Bildschirm wird an definierten Positionen ein weißer Punkt präsentiert (Programm CAL). Der rote Punkt repräsentiert die Position des Blickes. Durch eine geeignete Einstellung des Verstärkungsfaktors sowie
der Nullposition muss die Apparatur zunächst geeicht werden.
Oszillator Spannungsquelle
Tiefpass-Filter
Bandpass-Filter
IR-Sender
IR-Empfänger
Augenposition

Abb. 6: Bedienungselemente des IRSI Systems. 6: Einstellung der Verstärkung, 7: Einstellung der Nulllage, 8: Wählschalter für horizontale oder vertikale Blickposition, 9: grobe Positionsanzeige
Nach erfolgter Kalibrierung wird das horizontale und vertikale
Ausgangssignal für je 5 verschiedene Zielpositionen im Bereich zwischen 0 und 10° links, rechts, oben und unten in Grad Sehwinkel protokolliert. Gibt es ein Übersprechen zwischen vertikaler und horizontaler Position? Wie gut ist die Linearität der Messung? Tragen Sie dazu die Messwerte in ein x-y-Diagramm und berechnen Sie eine lineare Regression. 2. Glatte Augenfolgebewegungen
Das räumliche Auflösevermögen ist sehr unterschiedlich für die verschiedenen Bereiche des Gesichtsfeldes. Nur in der Fovea ist die Auflösung in der Größenordnung von etwa 60 Linien pro Grad. Bewegt sich ein Objekt, so sollten unsere Augen in der Lage sein, dieser Bewegung zu folgen. Das Programm FOLGE bietet eine Reihe von periodischen Zielbewegungen, die als Blickziel einer Versuchsperson dienen können. Dokumentieren Sie die Augenbewegungen in dieser Situation. Wie sieht die Initiierung der Folgebewegung aus? Bitten Sie die Versuchsperson, eine langsame Augenbewegung ohne einen bewegten Stimulus auszuführen. Wie sehen die Augenbewegungen in dieser Situation aus.
Können diese Augenbewegungen auch dann ausgeführt werden, wenn sich das Ziel vor einem strukturierten Hintergrund bewegt? Die Augenbewegung selbst führt zu einer Bewegung des retinalen Bildes des Hintergrund entgegengesetzt zur Blickfolgebewegung.
Bestimmen Sie den Frequenzgang der Augenfolgebewegungen. Messen Sie dazu die Augenbewegungen bei verschiedenen Frequenzen und bestimmen Sie zur Quantifizierung der Güte der Folgebewegung das Fehlerintegral (d.h. die Summe der Abweichung der Augengeschwindigkeit von der Zielgeschwindigkeit).
Frage für das Protokoll: Nehmen Sie an, dass die Augenbewegungen tatsächlich exakt der Zielbewegung entsprechen. Dies hat zur Folge, dass sich das Bild des Zieles auf der Retina nicht mehr bewegt. Wird in dieser Situation die subjektive Wahrnehmung von der retinalen Bildverschiebung bestimmt? 3. Sakkaden
Sakkaden sind sehr schnelle ruckartige Augenbewegungen zwischen zwei Fixationsphasen. Sie werden auch als „visueller Greifreflex“ bezeichnet. Dieser Begriff deutet die reflex-ähnliche Eigenschaft dieser Augenbewegungen an. Messen Sie mit einem geeigneten Messprogramm diese Augenbewegungen und analysieren Sie ihre Latenz. Ist die Latenz einer Sakkade abhängig von der Position des Ziels? Vergleichen Sie die sakkadische Latenz mit der Latenz des Kniesehnenreflexes.

TIERPHYSIOLOGISCHER KURS BIOINFORMATIK SS 2016 67
4. Augenbewegungen beim Lesen Beim Lesen führen wir ein sehr gut zu beschreibendes Augenbewegungsmuster aus. Wie sieht dieses Muster aus, wie lässt es sich erklären? Gibt es Abhängigkeiten des Augenbewegungsmusters von Eigenschaften des Textes?
ERFOLGSKONTROLLE
Nach diesem Praktikumsteil sollten Sie in der Lage sein, - den Unterschied zwischen Lautstärke und Schalldruck zu erklären,
- zu erklären, was ein Schallereignis physikalisch ist,
- den Unterschied zwischen Sinnesmodalität, Reizqualität und Reizintensität zu erklären,
- den Verlauf der menschlichen Hörschwelle zu skizzieren,
- die Maßeinheiten "dB SPL" und "Phon" zu definieren,
- das Verfahren zur Messung der Hörschwelle anzugeben,
- den Frequenzbereich des menschlichen Hörens anzugeben,
- den Hauptsprachbereich anzugeben,
- anzugeben, welche Kurvenform sich beim intermodalen Intensitätsvergleich in einer psychophysischen Messung ergibt,
- anzugeben, welche Parameter der Mensch zur horizontalen Richtungslokalisation verwendet,
- anzugeben, wie groß die Zeitdifferenzschwelle beim Menschen ist,
- ein Messverfahren anzugeben, mit dem die relative Bedeutung von Laufzeit- und Schalldruckunterschieden für das Richtungshören bestimmt werden kann,
- die Elemente eines monosyptischen Reflexkreises am Beispiel des Patellarsehnenreflexes zu nennen,
- den Unterschied zwischen einem monosynaptischen und einem polysynaptischen Reflexkreis zu beschreiben,
- die Wirkung zusätzlicher zentraler Einflüsse auf die Ausprägung eines monosynaptischen Reflexes zu nennen,
- die extraokulären Muskeln und die sie versorgenden Hirnnerven aufzählen zu können
- und Fixationen, Sakkaden und Glatte Augenfolgebewegungen unterscheiden zu können.





























