Embed Size (px)
Citation preview

Zeitschrift fiir Zellforschung 72, 446 - 461 (1966)
[ J B E R D A S R A U M S Y S T E M D E S E N D O P L A S M A T I S C H E N
R E T I K U L U M S V O N H A U T N E R V E N F A S E R N .
U N T E R S U C H U N G E N A N S E R I E N S C H N I T T E N *
WERNER REITER **
Anatomisches Ins t i tu t der Universi t~t Kiel (Direktor: Prof. Dr. W. BARGMANN)
Eingegangen am 21. Dezember 1965
Summary. Serial sections of skin-nerve fibres of Guinea-pig ears fixed in glutaraldehyde- osmium tetroxide and embedded in Aralditerevealed a number of electron microscopical findings.
1. The agranular axoplasmic reticulum of the axonal section of skin-nerve fibres appears in the three-dimensional reconstruction as a system of complicately shaped cavities. These cavities are arranged along the fibres and interweave with the axoplasm, either diffusely or in labyrinth-like formations.
2. Junct ions exist between the axoplasmic reticulum and mitoehondria on the one hand, and the periaxonal space on the other hand. In addition, direct connections between the periaxonal space and the mitochondria of the axon are observed.
3. In areas where the axoplasmic retieulum is missing, neurotubuli (microtubuli) may take its place. Like the axoplasmic reticulum, the neurotubuli are connected with the plasma- lemma and the mitochondria of the axon.
4. As revealed by the investigation, the granular endoplasmie reticulum of the Schwann cell has a structure similar to tha t of the axoplasmie reticulum. With in the cell, however, the granular endoplasmie retieulum is not orientated in a definite direction. I t is, however, connected with the mitochondria of the Schwann cell and the periaxonal space of unmyelinated O , x o n s .
5. The peripheral vesicular parts of the Golgi apparatus of the Schwann cell seem to origi- nate from proliferations of the granular endoplasmic reticulum which fuse to form the Golgi vesicles in the centre. Other vesicles communicate with the periaxonal space of unmyelinated axons.
6. The probable functional significance of these structures is briefly discussed.
Zusamme~/assung. An Seriensehnitten von Hautnervenfasern des Meerschweinchen- ohres wurden nach Fixierung mit Glutaraldehyd-Osmiumtetroxyd und Araldi te-Einbet tung folgende elektronenmikroskopisehen Befunde erhoben:
1. Das agranulis axoplasmatisehe Retikulum der axonalen Abschnit te der Hautnerven- laser erscheint in der Rekonstrukt ion als kompliziert gestaltetes Hohlraumsystem, das sieh l~ings der Faser formiert und das Axoplasma entweder in diffuser Verteilung oder in mehr randstis gestrecktcr Anordnung labyrinthar t ig durchsetzt.
2. Es bestehen Verbindungen vom axoplasmatischen Ret ikulum zu den Miteehondrien und zum periaxonalen Spaltraum, von wo aus auch direkte (~berg~nge zu den Mitoehondrien des Axons zu beobachten sind.
3. Die Neurotubuli (Mikrotubuli) kSnnen stellvertretend fiir das axoplasmatische Reti- kulum an Stellen erseheinen, wo dieses fehlt. Sie stehen wie dieses in Verbindung mit dem Axolemm und den Mitochondrien des Axons.
4. Das granuliire endoplasmatische Retikulum (Ergastoplasma) der Schwannzelle weist, wie die Rekonstrukt ion erkennen liigt, eine dem axoplasmatischen Ret ikulum 5~hnliche Struktur auf. Es ist jedoeh nicht in einer best immten Riehtung in der Zelle orientiert. Auch hier zeigen sich Verbindungen zu den Mitoehondrien der Schwannzelle und dem periaxonalen Spaltraum markloser Axone.
* Mit dankenswerter Unterst i i tzung durch die Deutsche Forsehungsgemeinsehaft. ** Unter Anleitung yon Priv.-Doz. Dr. K. H. ANDRES.

Endoplasmatisches Retikulum von Hautnervenfasern 447
5. Die peripheren vesikul/~ren Anteile des Golgiapparates der Schwannzelle k6nnen aus dem Ergastoplasma durch Abschnfirung entstehen und dann im Zentrum zu den bekannten Golgis~ickchen konfluieren. Andere Vesikel stehen in verbindendem Kontakt zum periaxonalen Raum markloser Axone.
6. Die m6gliche funktionelle Bedeutung der Strukturen wird kurz diskutiert.
Seitdem die Theorie der Erregungsleitung in den Neurofibrillen aufgegeben werden mul]te, wurde die Membran der Nervenfaser und die sie umgebende Markscheide als morphologisches Substrat f/Jr den Funktionsablauf am Nerven verst/~rkt in Betracht gezogen. Die Struktur des Inneren und der Umgebung dieses Membranrohres hat infolgedessen weiterhin wenig Beachtung gefunden. Auch die Analyse der Nervenfaserstruktur durch die Elektronenmikroskopie 1/il~t dies erkennen.
Schon 1953 konnten HEss und LANSING und 1954 BENNETT und DE ROBERTIS ein axoplasmatisches Retikulum beobachten. Neurotubuli haben bereits GASSER (1955, 1956), THORNBURG und DE ROBERTIS (1956), PALAu (1958) und VAN BREE- MEN et al. (1958) beschrieben. Nach weiteren Untersuchungen der Nervenfaser, der Entwicklung ihrer Markscheiden und der Neurofilamente (Obersiehten bei LEHMANN, 1959; CROSBY et al., 1962; HAGER, 1964) hat sich jedoch erst METU- ZALS (1963) wieder mehr der Struktur des axoplasmatisehen Retikulums gewidmet. Er sprieht auf Grund seiner Beobachtungen an markhaltigen Nervenfasern des ZNS yon der M6gliehkeit eines systematischen Zusammenhangs dieses Reti- kulums, das sich langs des Axons formieren mfisse.
ROBERTSON (1961) und DE ROBERTIS und BLEICHMAR (1962) diskutieren die M6glichkeit, dab Mitoehondrien aus den Formationen des axoplasmatischen Retikulums hervorgehen, weil Verbindungen zwischen diesen Zellbestandteilen zu beobachten seien. J~hnliehe Verbindungen wurden von SANDBORN et al. (1964) gesehen, der die Mikrotubuli als Bestandteil vermutlich jeder Zelle fordert, wo sie in einem feinsten Raumsystem auch Kontakte zur Zellmembran aufnehmen k6nnen.
ANDRES (1965 a) erkennt das axoplasmatisehe Retikulum in einem strukturellen Zusammenhang mit den Neurotubuli einerseits, den synaptisehen Vesikeln andererseits, wobei den Neurotubuli die Funktion einer Art yon Leitungsapparat zwischen Perikaryon und Faserendigung zugesprochen wird. Derselbe Verfasser beschreibt auch eine direkte Verbindung zwisehen axoplasmatischem Retikulum und dem periaxonalen Interzellularspalt (ANDRES, 1965b).
Alle diese Befunde wurden an Einzelschnitten erhoben. Im folgenden Beitrag wird fiber die ri~umliche Struktur des axoplasmatischen Retikulums und des Ergastoplasmas der Schwannzelle auf Grund einer Rekonstruktion aus Serien- schnitten berichtet. Au2erdem werden Verbindungen dieser Systeme zu anderen Zellbestandteilen an Befunden dargestellt, die gleichfalls an Serienschnitten yon Hautnervenfasern des Meerschweinchens erhoben wurden.
Material und Methode Untersucht wurden ca. 1--2 mm grolle Gewebestfickchen aus den Ohrmuscheln von
300--400 g schweren m/innlichen Meerschweinchen. Dieses Hautareal wurde vor anderem bevorzugt, weil es schnell und leicht zug/inglich ist (Fixierung), st6rendes Fett in der Subcutis nur in ganz geringer Menge vorkommt (Prgparation, Fixierung, Orientierung), die elastischen Knorpelanteile eine schienende und stabilisierende Eigenschaft haben (Prgparation, mechani-

448 WERNER REITER :
sche Fixierungsartefakte) und die doppelseitig vorhandene Epidermis und der mittelst~ndige Knorpel eine schnelle und leichte Orientierung im Gewebeanschnitt erm6glichen (s. u.).
Die Tiere wurden mit Nembutal-Natrium (Nembutal-Abbott) in einer Dosis yon 50 mg/kg K6rpergewicht i. p. narkotisiert. 5--7 mm vom Rande der Ohrmuschel entfernt wurden ca. 5 • 5 mm grofle Gewebestficke des ganzen Ohrmuschelquerschnittes entnommen und die durch die Schere gequetschten R~nder auf einem Paraffinpl~ttchen vorsichtig mit einer Rasierklinge abgeschnitten (ca. 2 • 2 mm Endgr6Be).
Die Erstfixierung erfolgte im 5 %igen Glutaraldehydgemisch yon pH 7,4 (SABATINI et al., 1963) bei Zimmertemperatur 60 rain lang. (Die Myelin|amellen waren in ihrer Lagerung zu- einander eindeutig besser erhalten als bei dem gleichen Fixierungsprozefl, der bei 0 ~ ~ C vor- genommen wurde (vgl. Abb. 3 mit Abb. 4). Zu einer giiltigen Aussage bedarf es aber noch systematischer Untersuchungen!). Ohne Zwischenspfilung wurde dann mit 1%iger Osmium- s~ure (d-5% Saccharose, pH 7,4) 2 Std nachfixiert. Als Einbettungsmittel diente Araldite.
Die einzubettenden Stfickchen des untersuchten Gewebes wurden also schon vor der Fixierung auf ihre Endgr613e zurechtgeschnitten, da bei vorsichtiger Pri~paration rein mecha- niseh bedingte Artefakte weitaus seltener waren als die Fixierungsartefakte bei ungeniigender, d .h. nicht rechtzeitig einsetzender Fixierung im Inneren zu grofler Gewebestiicke. Selbst bei der o. a. Gr61~e lagen die gut fixierten Bereiche meist nur in der Aul~enzone.
Zur Anfertigung m6glichst vieler Seriendiinnschnitte war eine Verkleinerung der An- schnittfl~che auf nut einen Hautnervenquerschnitt n6tig. Dazu wurden die Nerven zun~chst in 600--700 m~z dicken, nach RICnARDSO~ et al. (1960) gef~rbten Schnitten mit dem Licht- mikroskop aufgesucht. Dank der leichten Orientierung an den fibrigen Gewebebestandteilen (s. o.) konnte die Stelle mittels eines Binokulars im Anschnitt wiedergefunden und zu einer Fl~che von 0,2--0,4 mm 2 beschnitten werden. Um ein gerades Schnittband zu gew~hrleisten, muBten mindestens zwei Schnittkanten parallel orientiert werden. Mit einem Servall-Ultra- mikrotom und Glasmessern (PORTER und BLUM, 1953) wurden B~nder 700--800 A dicker Serienschnitte (20--30 Stfick) hergestellt, mit Chloroformdiimpfen gestreckt (GELBER, 1957) und auf Formvar-(Dioxan-)befilmten Kupfernetztr~gern mit einem Stegabstand yon 85 ~z aufgefangen, 10 min lang mit Bleicitrat kontrastiert (REYNOLDS, 1963) und mit dem Zeiss E ~ 9 mikroskopiert. Dabei mui~te peinlich auf eine sehr langsame Steigerung der Bildhellig- keit ( = Elektronendichte) geachtet werden, um eine ffir die plastische Rekonstruktion nach- teilige Verzerrung der Schnitte und der in ihnen enthaltenen Strukturen zu vermeiden.
Ffir die Rekonstruktion erschien eine Endvergr6Berung der Aufnahmen yon 60000fach (12000 • 5) giinstig. Die interessierenden Membranstrukturen wurden in Durchsicht entweder auf Glasplatten (Modell des axoplasmatischen Retikulums) oder auf Kunststoffplatten (Er- gastoplasmamodell) fibertragen und bei jedem Schnitt mit Hilfe mehrerer nicht zu rekon- struierenden Membranstrukturen des vorhergehenden Schnittes topographisch orientiert. Kfinstliche Markierungen lichtmikroskopischer Rekonstruktionsverfahren mit senkrechten Rillen am Gewebe etc. sind in der Elektronenmikroskopie nicht zu verwirklichen.
Fiir das ErgastoplasmamodeU (23 Schnitte) wurden 0,5 mm dicke, schneidbare Kunststoff- platten verwendet, wobei unter Berficksichtigung des Klebematerials fiir jeden Schnitt eine Schichth6he yon ca. 0,6 mm resultierte. Bei einer 60000fachen Vergr6flerung und einer Schnittdicke yon 700-800/~ ergab sich daraus eine Stauchung des Modells in der Vertikalen von ca. 1:8,7. Sie wurde sparer durch die photographisch-zeichnerische Streckung in der gleichen Ebene ausgegliehen (Abb. 8).
Bei der Rekonstruktion des axoplasmatischen Retikulums (24 Schnitte) wurden Glasplatten verwendet, da nach einer ersten Einsicht der Serienaufnahmen mit frei im Axoplasma liegen- den Vesikeln oder beginnenden Teilen des Retikulums zu rechnen war, die im Modcll keinen oder nur geringen Halt an den iibrigen Teilen haben konnten. Die interessierenden Strukturen wurden auf die Glasplatten gezeichnet und diese zu B16cken zusammengeffigt. Um ein Strich- gewirr fiir die photographisch-zeichnerische ~bertragung zu vermeiden, wurden zuerst 4 Plattenbl6cke mit nur je 6 Platten ( Schnitte) angefertigt und die danach gezeichneten Teildarstellungen des Modells zu einer Gesamtdarstellung vereinigt (Abb. 6). Eine zeichneri- sche Streekung war hier jedoch nicht n6tig, da die Dicke der Glasplatten der Vergr6Berung entsprechend gew~hlt werden konnte (4,6 mm + 0,2 mm Klebematerial pro Platte).
Die zeichnerische Wiedergabe einer Modellansicht kann natiirlich immer nur einen Tell der komplizierten r~umliehen Verh~ltnisse wiedergeben.

Endoplasmatisches Retikulum yon Hautnervenfasern 449
Befunde
A xoplasmatisches Retikulum
Die rSumliche Struktur des agranul/iren axoplasmatischen Retikulums wurde in erster Linie an Serienqucrschnittcn markloser Fasern untersucht (Abb. 1 a--e). Auch ffir die Rekonstruktion wurde ein solches Axon verwertet. Nach Durchsicht zahlreicher Querschnittserien markhaltiger Fasern und dem Vergleich mit Serienl/ingsschnitten (Abb. 2) muff als sicher angenommen werden, dal~ auch die markhaltige Faser ein axoplasmatisches Retikulum gleichen Typs enth~ilt. Da bei beiden Fasertypen nur Anteile mit einer ausgepr/~gten Schwannzellumschei- dung untersucht wurden, gelten die vorliegenden Befunde im besonderen ffir die axonalen Abschnitte motorischer oder sensibler Hautnervenfasern.
Das Neuroplasma der marklosen Fasern ist relativ reich an Neurotubuli. Die weitaus sp/~rlicher vorhandenen Neurofilamente liegen meist in Gruppen beiein- ander. Zwischen diesen Elementen sind die unterschiediich geformten Anschnitte des axoplasmatischen Retikulums mit Lumina zwischen 200 und 900 A diffus verteilt (Abb. la--e) . Daneben zeigen sich vereinzelt Glykogengranula.
Bei den markhaltigen Axonen k6nnen drei verschiedene Fasertypen unter- schieden werden:
a) Der Bau des Axoplasmas gleicht dem der marklosen Fasern (Abb. 2). b) Daneben sind Fasern zu beobachten, die kaum Neurotubuli, jedoch viele
Neurofilamente enthalten. Das axoplasmatische Retikulum ist auch hier diffus verteilt (Abb. 3).
c) Schliel~lich kommen Axone vor, die nur wenige Neurotubuli haben und reich an Neurofilamenten sind, jedoch ein auffallend randst/~ndiges axoplasmatisches Retikulum aufweisen (Abb. 4). Dies ist in unmittelbarer N~he des Axolemms bemerkenswert regelmKl~ig angeordnet, wobei Verbindungen unter den rand- st/~ndigen und zu nur vereinzelt zentral gelegenen Anschnitten des Retikulums vorhanden sind. Selbst an intraplasmatischen Einfaltungen des Axolemms 1/iBt sich diese regelm/~Bige Anordnung weiterverfolgen. Bemerkenswert ist auBerdem eine Art Stellvertretung durch Neurotubuli an Orten, an denen das randst/~ndige axoplasmatische Retikulum fehlt.
Die Auswertung der Quer- und L/~ngsschnittserien erlaubt nun eine Aussage fiber die rdumliche Struktur des axoplasmatischen Retikulums und seiner Ver- bindungen zum periaxonalen Spaltraum und zu den Mitochondrien der Faser.
1. In der Rekonstruktion erscheint das axoplasmatische Retikulum als kom- pliziertes, in der L/ingsachse der Faser orientiertes Hohlraumsystem. Es besteht aus unterschiedlich weiten R6hren und Vesikeln, die auch Querverbindungen untereinander aufweisen. Mitunter sind sackf6rmige Auftreibungen mit dicht gelagerten Granula (60--80 A) zu beobachten, die in das zusammenh/~ngende Raumsystem mit eingegliedert sind. Isoherte Vesikel kommen relativ selten vor (Abb. 6).
2. Dieses Hohlraumsystem hat Verbindungen zum periaxonalen Interzellular. spalt. Aus Einzelquerschnitten allein ist dies selten zu entnehmen, denn dort zeigt sich meistens nur eine begrenzte Invagination des Axolemms in das Plasma der Faser (Abb. 1a--c, 5a). Die starke SchlKngelung des axoplasmatischen Retikulums im Verh/iltnis zur Schnittdicke macht dies verst/~ndiich. Verfolgt man nun diese Stelle in den anschliel]enden Serienschnitten, so erkennt man eine
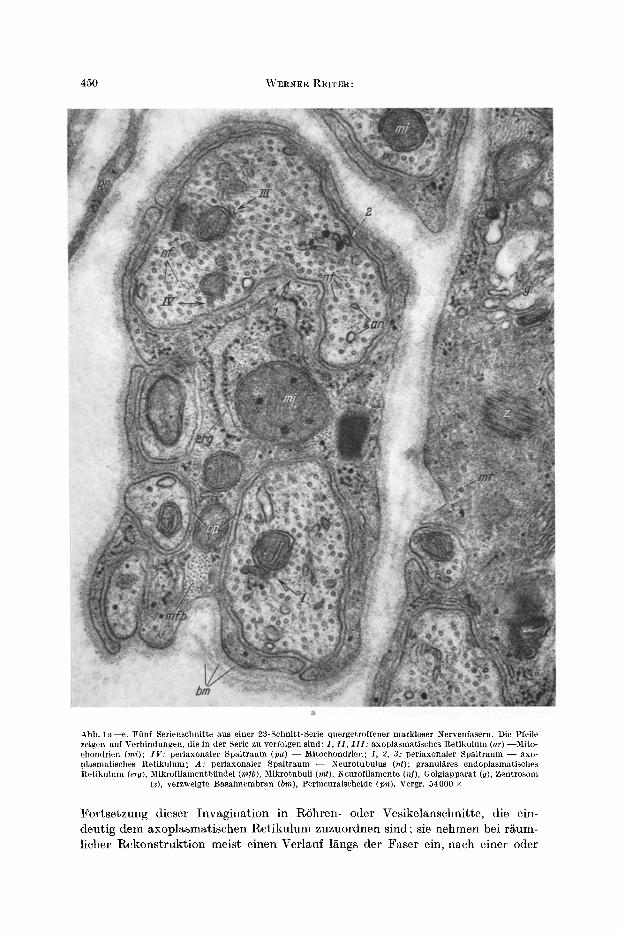
4 5 0 W E R N E R R E I T E R :
Abb. l a - - e . F i inf Serienschnitte aus einer 23-Schnit t-Serie quergetroffener markloser Nervenfasern . Die Pfeile zeigen auf Verbindungen, die in der Serie zu verfolgen s ind: I , I I , I I I : axoplasmat ischcs Re t iku lum (ar) - -Mi to - chondrien (mi); I F : periaxonaler Spa l t raum (pa) - - Mitochondricu; 1, 2, 3: periaxonuler Spa l t raum - - axo- plasmatisches Re t iku lum; A: periaxonaler Spa l t raum - - Neurotubulus (nt); granul~res cndoplasmatisches Re t iku lum (erg), Mikrofi lamcntbfindel (m/b), Mikrotubuli (rot), Neurof i lamente (hi), Golgiappara t (g), Zentrosom
(z), verzweigte Basa lmembran (bin), Perineuralscheide (pn). Vergr. 54000 •
Fortsetzung dieser Invagination in RShren- oder Vesikelanschnitte, die ein- deutig dem axoplasmatischen Retikulum zuzuordnen sind; sie nehmen bei r/~um- licher Rekonstruktion meist einen Verlauf 1Kngs der Faser ein, nach einer oder
Endoplasmatisches Retikulum von Hautnervenfasern 45I
Abb. 1 b - - e
nach zwei Seiten hin. An Serienl~ngsschnitten k6nnen diese Befunde best/~tigt werden (Abb. 2).
Die besehriebenen r6hrenf6rmigen Einstfilpungen der Fasermembran und die sie fortsetzenden Teile des axoplasmatisehen Retikulums miissen yon den/l~ichen- haften Einfaltungen untersehieden werden, die sieh dutch die ganze Sehnittserie verfolgen lassen. Sie k6nnen entweder fingerf6rmige Ausl/~ufer von Sehwannzellen (Abb. 3) oder Anteile des Cytoplasmas von Sehwannzellen am inneren Mesaxon umschliegen (Abb. 4). Doeh aueh yon diesem eingefalteten Axolemm k6nnen die besproehenen Verbindungen zum axoplasmatisehen Retikulum ausgehen (Abb. 3).

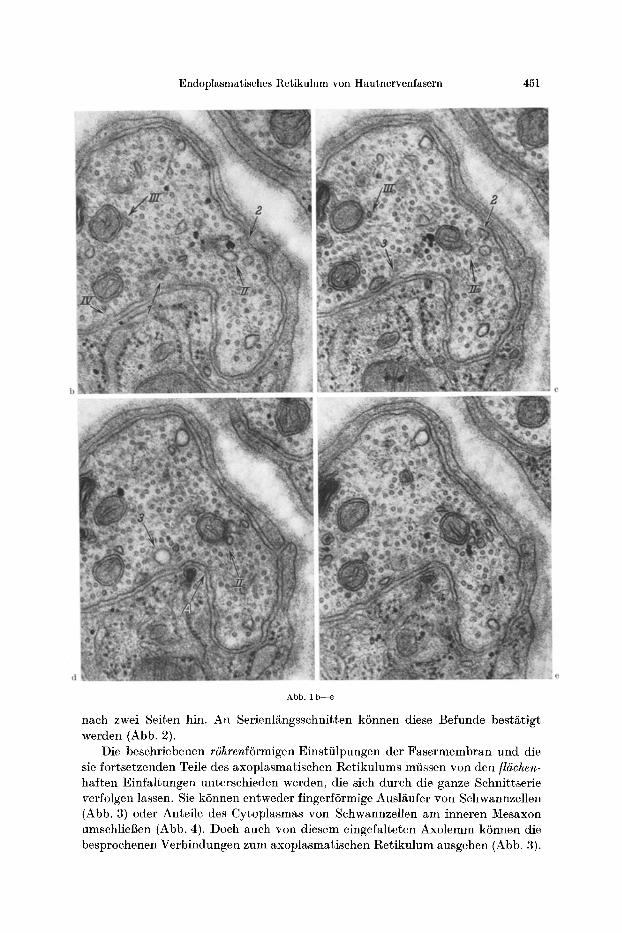
452 WERNER ~EITER :
Abb. 2
Abb. 3
Abb. 2. L~ngsschni t t einer markha l t igen Hau tne rvenfase r (aus einer Schnittserie). Es ist der Zugang v o m peri- axonalen t t a u m (pa) zum axoplasmat i schen Re t iku lum (ar) zu beobachten (Pfeil). Das Axo lemm (axl) knick t spi tzwinklig in die Membran des axoplasmat i schen l~etikulums urn. Viele Neurotubuli (nt). Vergr. 45000 •
Abb. 3. Querschni t t einer markha l t i gen Hau tne rvenfase r (aus einer Schnittserie). Verzweigte, f l~chenhafte Axolemmeinfa l tungen (ale), die f ingerfSrmige Schwannzellausstf i lpungen (sa) umscheiden, ncben rShrenfSrmiger
Verb indung zum axoplasmat i schen Re t iku lum (ar, Pfeil). Viele Neurof i lamente (nf). u 45000 •
3. Bei den Verbindungen des axoplasmatischen Retikulum8 zu den Mitochondrien liegen ganz /~hnliche Verh~ltnisse vor. Es zeigen sich entweder 40--100 m~ lange nabelfSrmige Ausstfilpungen (Abb. 1a- -b) oder aber sackf6rmige Aus- buchtungen der AuBenmembran der Mitochondrien (Abb. l c). In den folgenden Serienschnitten sind auch in diesem Fall isoliert liegende Anschnitte des axo- plasmatischen Retiku]ums neben den Mitochondrien zu sehen. Somit ergibt sich nach Mai~gabe der Rekonstruktion eine r6hrenfSrmige Verbindung zwischen axoplasmatischem Retikulum und den Mitochondrien (Abb. 6).
4. Nach dem Gesagten ist also eine indirekte Verbindung zwischen dem Peri- axonalen Spaltraum und den Mitochondrien des Axons fiber den Weg des axo- plasmatischen Retikulums vorhanden. Jedoch sind daneben Stellen zu beobach- ten, an denen das Axolemm mehr oder minder direkt in die ~ul~ere Membran der Mitochondrien umschl~gt, die nahe am Axolemm liegen (Abb. 1 a - -b , 5b). Auch hier sind oft zwei aufeinanderfolgende Serienschnitte zur Auswertung nStig.
N eurotubuli ( M ikrotubuli )
Selbst die verbesserte elektronenmikroskopische Technik unserer Tage erlaubt es kaum, Aussagen fiber die membran6sen Beziehungen der Neurotubuli zu den anderen Raumsystemen der Zelle zu machen, Das liegt an dem Gr6Benverh~ltnis yon Neurotubulusdurchmesser (150--200 A) und den in der Elektronenmikro-
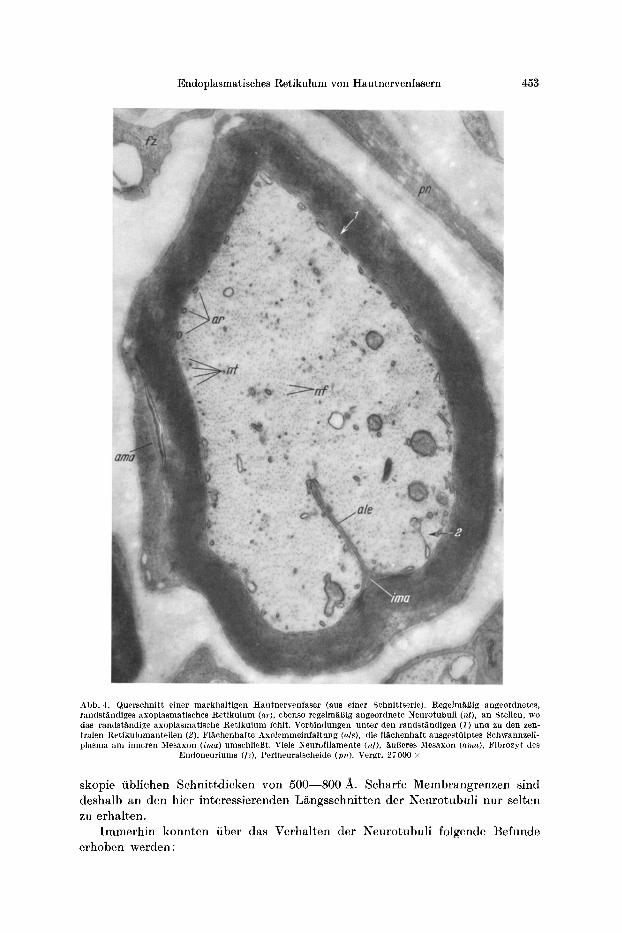
Endoplasmatisches Retikulum yon Hautnervenfasern 453
Abb. 4. Querschnitt einer markhaltigen Hautnervenfaser (aus einer Sehnittserie). Regehnfi, gig angeordnetes, randst~ndiges axoplasmatisches Retikulum (ar), ebenso r ege lm~ig angeordnete Neurotubuli (nt), an Stellen, wo das randstandige axoplasmatisehe Retikulum Iehlt. Verbindungen unter dell randstgndigen (1) und zu den zen- tralen Rctikulumanteilen (2). Fl~chenhafte Axolemmeinfaltung (ale), die fl~chenhaft attsgestOlptes Schwannzell- plasma am inncren Mesaxon (ima) umschliegt. Viele Neurofilamente (M), ~ugeres Mesaxon (area), Fibrozyt des
Endoneuriums ([z), Perineuralscheide (pn). Vergr. 27000 •
skopie fiblichen Schni t td icken von 500--800 A. Scharfe Membrangrenzen sind deshalb an den hier interessierenden L/~ngsschnitten der Neurotubul i nur selten zu erhalten.
Immerh in konn t en fiber d~s Verhal ten der Neuro tubul i folgende Bcfunde erhoben werden :

454 WER~ER REITER :
Abb. 5a, Teil eines Anschni t tes einer Schwannzelle mi t marklosen Nervenfasern (aus einer Schnittserie). Bci 1: Verbindung zwischen Ergas top lasma (erg) und Mitochondr ium (mi) dcr Schwannzelle. Bei 2: Schr~ganschni t t eines Zuganges v o m pcriaxonalen Spa l t raum zum axoplasmat i schen Re t iku lum (ar) eines mark losen Axons; im n~tchsten (nicht gezeigten) Serienschnit t ist dort ein v o m Axolemm get rennt liegender Anschni t t des Re t i ku lums
zu beobachten. Yergr. 54 000 •
Abb. 5b. Teil eines Anschni t tes einer Schwannzelle mi t mark losen Axonen (aus einer Schnittserie). Bei 1: Das Ergas top lasma (erg) der Schwannzelle ha t einen Zugang zum periaxonalen Spa l t r aum eines mark losen Axons. Bei 2: Die A u 6 e n m e m b r a n eines axonalen Mitochondriums schlRgt in das Axo lemm um. Bei 3: NabelfSrmige
Verbindung eines axonalen Mitochondriums zu einem Neurotubulus (nt). Vergr. 54000 x
1. Direkte Verbindungen zwischen Neurotubuli und axoplasmatischem Retikulum sind in den untersuchten Abschnitten der Hautnerven nicht sieher nachzuweisen.
2. In den Querschnitten markloser Nervenfasern kSnnen offene Verbindungen zwischen Neurotubuli und periaxonalem Spaltraum beobachtet werden. Diese Ver- bindungstubuli sind meist 50--60 m~ lang, laufen quer zum allgemeinen Verlauf der Neurotubuli im Axon und liegen in Zonen, in denen sonst nur Querschnitte der Neurotubuli zu sehen sind (Abb. 7 a u. b). Die mit dem Axolemm verbundenen Profile von Neurotubuli(quersehnitten) haben in den beiderseits folgenden Serien- schnitten Fortsetzungen in einer Position, die nach dem allgemeinen Tubulus- verlauf l~ngs der Faser zu erwarten ist und nicht etwa in einer Seitenverschiebung um die Li~nge des Verbindungstubulus. Eine 0ffnung des Tubulusquerschnittes zum Verbindungstubulus hin ist jedoch wie auch bei den unten noch zu schildern- den Befunden in keinem Fall zu erkennen. Auch dies ist Folge der zu gro~en Schnittdicke (s. o.).
3. Wie beim axoplasmatisehen Retikulum sind aueh bei den Neurotubuli membranSse Beziehungen zu den Mitochondrien zu erkennen. Auch hier stellen sie sich als nabelfSrmige Ausstfilpungen der Au6enmembran dar, die meist zu einem nahegelegenen Neurotubulus ffihren (Abb. 7c). Kalibersti~rke und Membran- kontrast entsprechen den in Schri~g- und Li~ngsschnitten gewohnten Darstel- lungen der Neurotubuli. Ffir die Deutung an den beiderseits folgenden Serien- schnitten gilt das unter 2. Gesagte.
Ergastoplasma und Golgiapparat der Schwannzelle
1. Die besonders enge topographische Beziehung des Ergastoplasmas der Schwannzelle zu den Axonen, insbesondere den marklosen Fasern, machte eine genauere Untersuchung dieser Strukturen anhand yon Serienschnitten wiin- schenswert. Es ergab sich das in Abb. 8 dargestellte Raumsystem, das weitaus komplizierter erscheint als der yon PALAY und PALADE (1955) untersuchte

Endoplasmatisches Retikulum yon Hautnervenfasern 455
Abb. 6. R~umliches Verhal ten des axoplasmat i schen Re t iku lums einer mark losen Hau tne rvenfase r (Rekonst ruk- t ion aus 24 Serienschnitten). I - - V I I I : Verbindungen zwischcn Mitochondrien (mi) und axoplasmat i schem Reti- ku tum; I X : direkter Zugang v o m periaxonalen R a u m zu einem Mitochondr ium; 1--4: Invag ina t ion des Axo- lemms (axl) mit Zugang zum axoplasmat i schen Re t iku lum; 5: [Tbergang v o m periaxonalen R a u m zum granularen endoplasnmtischen Re t iku lum (erg) der Schwannzelle; in das axoplasmat ische Re t iku lum eingegliederte Granula-
s~ickchen (grs), isoliert l iegende vesikul~re Anteile des Re t iku lums (vs). MaBstab ca. 1:85000
Ergastoplasmatyp in Nervenzellen. Gemeinsam ist beiden jedoch die Kontinuit/~t ihres Internums, die auch durch einen komplizierten Membranverlauf nicht unter- brochen wird. Das Ergastoplasma scheint das Schwannzellpl~sma auf diese Weise labyrinthartig zu durchflechten.
In Analogie zum axoplasmatischen Retikulum sind auch beim Ergastoplasma Verbindungen zu ~nderen Raumsystemen zu beobachten. Auch hier gibt cs r6hren- f6rmige Verbindungen zu benachb~rten periaxonalen R~umen markloscr Fasern (Abb. 5b, 6, 8), ebenso solche zu Mitochondrien der Schwannzelle (Abb. 5a, 8).

4 5 6 W E R N E R R E I T E R :
Abb. 7a u. b. Teil eines Anschni t tes einer Schwannzelle (sz) m i t marklosen Axonen (ax) (aus einer Schnittserie). Verbindungen zwischen per iaxonalem l~aum und Neurotubuli (1)feile). Vergr. 72000 •
Abb. 7c. Tell eines Anschni t tes einer Schwannzelle (sz) m i t mark losem Axon (ax) (aus einer Schnittserie). ~abel- fSrmige Ausst i i lpung der Aul~enmembran eines axonalen Mitochondr iums m i t Verb indung zu e inem Neurotubulus
(Pfeil). Vergr. 72000 •
Diese Verbindungen waren aber im gesamten Untersuchungsmaterial weitaus seltener als die bei dem axoplasmatischen Retikulum beobachteten.
2. Der Golgiapparat der SchwannzeUe ist in der 57~he der Zentriolen sehr aus- gepriigt (Abb. 1 a) und kann dort besonders gut in seiner Beziehung zu benach- barren Raumsystemen beobachtet werden. (Eine genaue Rekonstruktion ist einer sp~teren Arbeit vorbehalten und liegt in einer Teilrekonstruktion schon bei REALE und LUCIANO, 1964, vor.) Auch hier sind zum periaxonalen Interzellular- spalt markloser Axone Verbindungen zu beobachten, die sich im untersuchten Material jedoch nur auf die vesikul~ren Teile der Golgizone beschr~nken (Abb. 9).
Dies gilt genauso ffir die Beziehung zum Ergastoplasma, doch erinnert die Art dieser Verbindung stark an das flfichtige Zustandsbild eines Abschnfirungsvor- ganges (Abb. 10). Die sich abschniirenden oder bereits abgeschnfirten Golgivesikel gehSren zu den peripher gelegenen Anteilen des Golgiapparates mit einem Inhalt, dessen Zentrum dem fein granulierten Internum des Ergastoplasmas gleicht, unter der Membran jedoch einen dichteren Granulabesatz hat. Diese Bl~schen scheinen nahe dem Golgizentrum zu den abgeplatteten GolgisKckchen zu kon- fluieren, die auch dann noch die geschilderte Internumstruktur aufweisen kSnnen (Abb. 9).
Diskussion
Die Vorstellung unserer Befunde steht mit der erw~hnten Forderung METU- ZALs (1963) nach einem im Axon liingsgerichteten Raumsystem des axoplasma- tischen Retikulums sehr weitgehend in Einklang. Dieses Zellorganell nimmt im Axon einen weitaus grSBeren Raum ein, als es nach den ersten elektronenmikro- skopischen Beobachtungen yon HESS und LANSING (1953) und BENNETT und DE ROBERTIS (1954) ZU erwarten war und wie es auf Einzelquer- und -l~ngs- sehnitten yon Nervenfasern erscheint.
Neben der diffusen Verteilung des axoplasmatischen Retikulums ist die rand- sti~ndige Anordnung in neurotubulusarmen, markhaltigen Axonen besonders interessant (Abb. 4). Diese Erseheinung kann nur hypothetisch gedeutet werden.
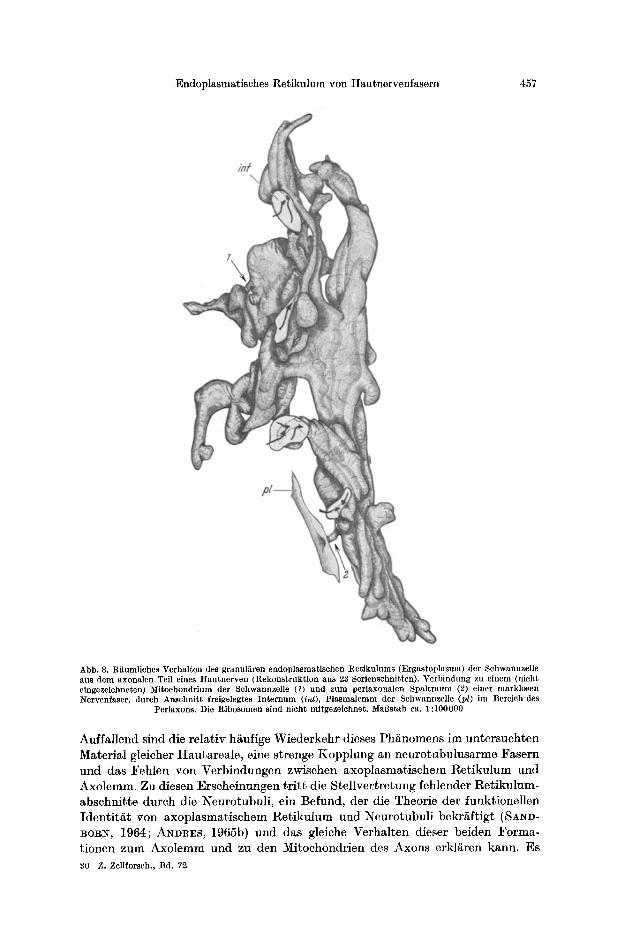
Endoplasmatisches Retikulum von Hautnervenfasern 457
Abb. 8. Riiumliches Verhal ten des granul~ren endoplasmat ischen Re t iku lums (Ergas toplasma) der Schwannzelle aus dem axonalen Tell eines H a u t n e r v e n (Rekons t ruk t ion aus 23 Serienschnitten). Verb indung zu einem (nicht eingezeichneten) Mi tochondr ium der Schwannzelle (1) und zum periaxonalen Spa l t raum (2) einer marklosen Nervenfaser , durch AnschniSt freigelegtes I n t e r n u m (int), P l a s m a l e m m der Schwannzelle (lad im Bereich des
Per iaxons. Die Ribosome, n s ind nicht mitgezeichnet . Ma[3stab ca. 1:100000
Auffallend sind die relativ h/iufige Wiederkehr dieses Ph~nomens im untersuchten Material gleicher Hautareale, eine strenge Kopplung an neurotubulusarme Fasern und das Fehlen von Verbindungen zwischen axoplasmatischem l~etikulum und Axolemm. Zu diesen Erscheinungen tr i t t die Stellvertretung fehlender Retikulum- abschnitte durch die Neurotubuli, ein Befund, der die Theorie der funktionellen Identit/i t yon axoplasmatischem Retikulum und Neurotubuli bekr~ftigt (SAND- BO~N, 1964; ANDRES, 1965b) und das gleiche Verhalten dieser beiden Forma- tionen zum Axolemm und zu den Mitochondrien des Axons erkl/~ren kann. Es 30 Z. Zellforsch., Bd. 72

458 WERNER REITER :
Abb. 9 Abb. 10 Abb. 9. Teil eines Anschnittes einer Schwannzelle mit mark|osen Axonen (ax) (aus einer Schnittserie). Die vesiku- lf~ren Anteile des Golgiapparates (g) stehen im Kontakt zum periaxonalen Raum (1, 2). Im Zentrum der Golgizone zu S~ckchen konfluierende Vesikel des Goigiapparates (3), die in ihrer Innenstruktur den in Abb. 10 gezeigten
~hnlich sind, Vergr. 54000 •
Abb. 10, Teile zweier Anschnitte yon Schwannzellen (sz) (aus einer Schnittserie). Vom granulfiren endoplasmati- schen Retikulum (erg) schnQren sich peripher gelegene, vesikul~re Anteile des Golgiapparates (vg) ab (Pfeil). Die Fortsetzung des Golgiappas in der Serie steht senkrecht zur Bildebene. Die Vesikel haben in ihrer Struktur
~hnlichkeit mit den in Abb. 9 gezeigten, konfiuierenden Anteilen der Golgizone. Vergr. 67000 •
ist also mSglich, dal~ stoffwechselmechanische Griinde zur Affinit/~t des axo- plasmatischen Retikulums zum elektrophysiologisch wichtigen Axolemm ffihren. Worin der Sinn der beobachteten ,,unerkl/irlichen" Axolemmeinfaltung (Dixon, 1963) besteht, ist nicht zu entscheiden. Mit morphologischer Methodik kann nicht gekl/~rt werden, welcher Funktion die beiden Retikulumtypen im Zellstoff- wechsel dienen, doch kSnnte man die Neurotubuli als gestreckte Direktverbin- dungen des axonalen Stofftransportes deuten, denen die komplizierteren Raum- systeme des axoplasmatischen Retikulums mit einer hSheren Stoffwechsel- und Transportkapazit~t zur SeRe stehen. Daffir wfirde auch eine (vorl~ufige ?) Ab- lagerung yon Stoffwechselzwischen- oder -endprodukten in Granulas~ckchen des Retikulums sprechen (Abb. 6). Sind an bestimmten Abschnitten der Fasern nur wenige Neurotubuli vorhanden, wfirde sich das axoplasmatische Retikulum in einem gestreckteren ( ~ direkteren) Verlauf in der AuBcnzone des Axons for- mieren, wo auch ein osmoregulatorischer Stoffwechselprozel~ besser mSglich ist. Warum gerade dann keine offenen Verbindungen zur AxonoberfliLche bestehen, kann nicht erkl~rt werden.
Die Einfaltungen des Axolemms markhaltiger Fasern waren Ifir DE ROBERTXS und BLEICHMAR (1962) Anlal~ zu einer besonderen Theorie der Mitochondrio- genese. Ein Teil ihrer Befunde ist mit denen in Abb. 3 gezeigten identisch. Die von den Veffassern als ,,Promitochondrien" gedeuteten Bildungen am Ende der Axolemmeinfaltungen sind jedoch - - nach Auswertung langer Schnittserien - - fingerfSrmige Aussackungen der Schwannzelle, die vom Axon gleichsam um- scheidet werden. Dies konnte auch A~])RES (1965b) an markhaltigen Fasern im Kleinhirn der Ratte zeigen. Das Axolemm dieser ]Idchenhaften Einfaltungen sollte in seiner morphologischen Wertigkeit in keiner Weise yon den fibrigen Axolemm- anteilen differenziert werden und ist von den rein r6hrenfSrmigen Verbindungen zwischen axoplasmatischem Retikulum und periaxonalem Interzellularspalt streng zu unterscheiden. Beide Ph~nomene sind in Abb. 3 nebeneinander dar- gestellt.
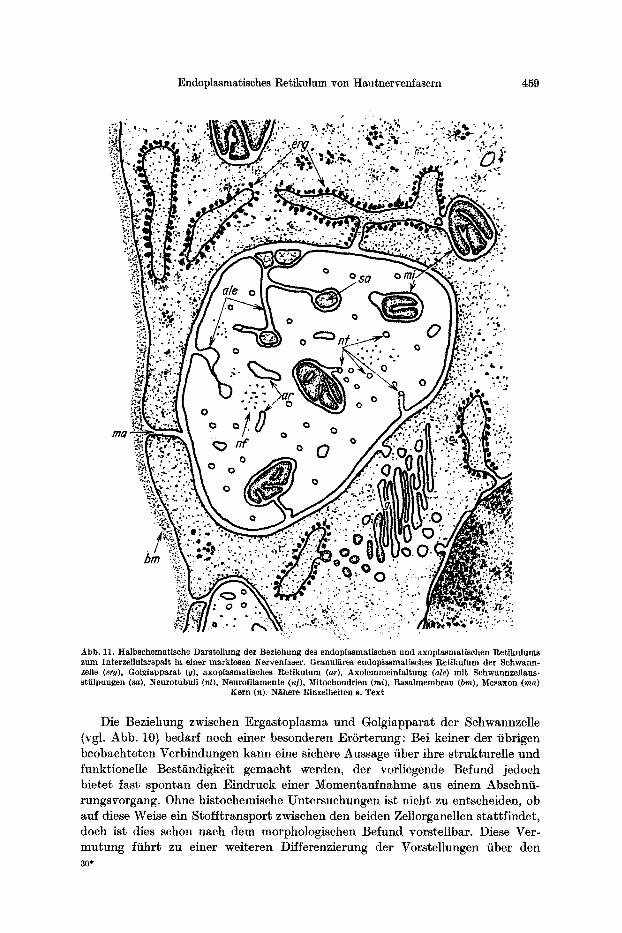
Endoplasmatisches Retikulum yon Hautnervenfasern 459
Abb. 11, Halbschematische Darstellung der Beziehung des endoplasmatisehen und axoplasmatisehen Retikulums zum Interzel|ularspalt, in einer marklosen Nervenfaser. Granulates endoplasmatisehes Ret ikulum der Schwann- zeUe (erg), Golgiapparat (g), axoplasmatisches Retikulum (ar), Axolemmeinfaltung (ale) mit Schwannzellaus- stiilpungen (sa), ~eurotubuli (nt), Neurofilamente (n]), Mitoehondrien (mi), Basalmembran (bin), Mesaxon (ma)
Kern (n). N~there Einzelheiten s. Text
Die Beziehung zwischen Ergastoplasma und Golgiapparat der Schwannzelle (vgl. Abb. 10) bedarf noch einer besonderen ErSrterung: Bei keiner der fibrigen beobachteten Verbindungen kann eine sichere Aussage fiber ihre strukturelle und funktionelle Best~ndigkeit gemaeht werden, der vorliegende Befund jedoch bietet fast spontan den Eindruck einer Momentaufnahme aus einem Abschnfi- rungsvorgang. Ohne histochemische Untersuchungen ist nicht zu entscheiden, ob auf diese Weise ein Stofftransport zwischen den beiden Zellorganellen stattfindet, doch ist dies schon nach dem morphologischen Befund vorste|lbar. Diese Ver- mutung ffihrt zu einer weiteren DiSferenzierung der Vorstellungen fiber den 30*

460 WERNER REITER :
Stoffwechsel des Golg iappara tes a n h a n d submikroskopischer Befunde (ROTtI und PORTER, 1964; A~DRES, 1964).
Es is t anzunehmen, dab die un te r such ten Membransys t eme des Axop la smas und der Schwannzelle zu e inem Tell die Funk t ions t f i ch t igke i t des Axons aufrecht- erhal ten. Die Er regungs le i tung f indet zwar durch die Vermi t t lung der A xonober- /lgche s ta t t , doch sind endoplasmat i sches Re t iku lum, Mikro tubu l i und Mito- chondr ien des Axoplasmas Organellen, die als H i l f sappa ra t e ffir die Regenera t ion der e r regten Fase r dienen und fiberdies in die Erni~hrung und sicher auch in die mechanische Stabi l is ierung des Axons e ingeordnet sind. Bei der mark losen Fase r komra t auBerdem ein besonders enger K o n t a k t ihrer Oberfl~che zu den Raum- sys temen der Schwannzelle hinzu, die, wie beschrieben, ihre eigenen charakter i s t i - schen Merkmale haben, jedoch in keiner Weise gegeneinander isoliert sind. Auch sie dienen der E rha l t ung des Axons und seiner Arbei ts f~higkei t .
Berf icksicht igt man die Verb indung zwischen endop lasmat i schem Re t iku lum und per inukle~rem R a u m (WATSOn, 1955), die wi t h~ufig beobach te ten , so ergibt sich ein morphologisch f iberaus kompl iz ie r te r Zusammenhang verschiedener Sys teme nicht nur einer, sondern mehrerer bena c hba r t e r Zellen, die funkt ionel l ane inander gebunden sind (Abb. 11). Sie s ind durch pri~formierte Wege des Stoff- aus tausches mi t e inander verknfipft , und es b le ib t zu untersuchen, in welcher Weise diese Transpor twege genu tz t werden und ob sie eine s ts E in r i ch tung oder daue rnde r s t ruk ture l l e r W a n d l u n g unterworfen sind. ,,Die Regu la t ion des Chemismus der Zelle kann nicht ohne Beach tung ihrer S t ruk tu re inhe i t en ver- s tanden werden" (NETTER, 1959).
L i t e r a t u r
ANDRES, K. IX. : Mikropinozytose im Zentralnervensystem. Z. Zellforsch. 64, 63--73 (1964). - - Der Feinbau des Bulbus olfactorius der Ratte unter besonderer Beriicksichtigung der
synaptischen Verbindungen. Z. Zellforsch. 65, 530--561 (1965a). - - l~ber die Feinstruktur besonderer Einrichtungen in markhaltigen Nervenfasern des Klein-
hirns der Ratte. Z. Zellforsch. 65, 701--712 (1965b). BENNETT, IX. S., and E. DE P~OBERTIS: Some observations on the fine structure of the giant
nerve fibers. Proc. Internat. Conf. on Electron Microscopy, London 1954. BREEME~, V. L. VAN, E. A~DERSON, and J. F. REOER: An attempt to determine the origin
of synaptic vesicles. Exp. Cell. l~es., Suppl. 6, I53--167 (1958). CROSBY, E. C., T. HUMP~IREY, and E. W. LAUER: Correlative anatomy of the nervous system.
New York: Macmillan & Co. 1962. DIXON, A. D. : The ultrastructure of nerve fibers in the trigeminal ganglion of the rat. J.
Ultrastruct. l~es. 8, 107--121 (1963). GASSER, H. S. : Properties of dorsal root unmedullated fibers on the two sides of the ganglion.
J. gen. Physiol. 38, 709--728 (1955). - - Olfactory nerve fibers. J. gen. Physiol. 38, 473--496 (1956). GELBER, D. : Thin sectioning: Details of techniques. J. biophys, biochem. Cytol. 3, 311--316
(1957). HAGER, IX. : Feinere Cytologie und Cytopathologie des Nervensystems. In: VerSffentl. morph.
Path., H. 67. Stuttgart: Gustav Fischer 1964. HESS, A., and A. I. LANSING: The fine structure of peripheral nerve fibers. Anat. Rec. 117,
175--199 (1953). L E G M A N . , H.-J.: Die Nervenfaser. In: Handbueh der mikroskopisehen Anatomie des Men-
schen, Teil IV, Erg. z. Bd. IV/1. Berlin-GSttingen-ixeidelberg: Springer 1959. MI~TUZALS, J. : Ultrastrueture of myelinated nerve fibers in the central nervous system of the
frog. J. Ultrastruct. Res. 8, 30--47 (1963).

Endoplasmatisches Retikulum von Hautnervenfasern 461
NETTER, I-I.: Theoretische Biochemie. PhysikMisch-chemische Grundlagen der Lebens- vorgange. Berlin-GSttingen-Heidelberg: Springer 1959.
PALAY, S. L. : The morphology of synapses in the central nervous system. Exp. Cell Res., Suppl. 5, 275--293 (1958).
- - , and G. E. PaLADE: The fine structure of neurons. J. biophys, biochem. Cytol. 1, 69--88 (1955).
PORTER, K. R., and J. BLU~: A study in microtomy for electron microscopy. Anat. Rec. 117, 685--712 (1956).
REALE, E., u. L. LVCIANO: Beobachtung an parallel zur Lage der Sacculi des Golgi-Apparates gefiihrten Schnitten. J. Microscopie 8, 545--550 (1964).
REYNOLDS, E. S. : The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 17, 208--212 (1963).
RICHARDSON, K. C., L. JANETT, and E. H. FINKE: Embedding in epoxy resins for ultrathin sectioning in electron microscopy. Stain Technol. 85, 313--323 (1960).
ROBERTIS, E. DE, and It. BLEICI~MAR: Mitochondriogenesis in nerve fibers of the infrared receptor membrane of pit vipers. Z. Zellforsch. ~7, 572--582 (1962).
ROBERTSON, J. D . : Cell membranes and the origin of mitochondria. In: Regional neuro- chemistry (S. KETY and J. ELKES, eds.), p. 497--530. Oxford: Pergamon Press 1961.
ROTIt, T. :F., and K. R. PORTER: Specialized sites on the cell surface for protein uptake, in 5th Internat. Congr. for Electron microscopy, Philadelphia 1962 (S. S. BREESE jr. Edit.) 2. LL-4. New York: Academic Press Inc. 1962.
SABATINI, D. D., K. G. BENSCH, and R. J. BARRNETT: The preservation of cellular ultra- structure and encymatic activity by aldehyde fixation. J. Cell Biol. 17, 19--58 (1963).
SANDBORN, E., P. F. KOEN, J. D. Mc NABB, and G. MOORE: Cytoplasmic microtubules in mammalian cells. J. Ultrastruct. Res. 11, 123--138 (1964).
T~ORNBURG, W., and E. ~)E RO~RTIS: Polarization and electron microscope study of frog nerve axoplasm. J. biophys, biochem. Cytol. 2, 475--482 (1956).
WATSOn, M. L. : The nuclear envelope. Its structure and relation to cytoplasmic membranes. g. Biophys. Biochem. 1, 257--270 (1955).
WERNER REITER Anatomisches Institut der Universit~it 23 Kiel, Neue Universit~t, Eingang F 1





























