Embed Size (px)
Citation preview

Planta (Berl.) 121, 225--235 (1974) �9 Springer-Verlag 1974
[Jber den EinfluB yon NaC1- oder Abscisins~iurebehandlung auf den Proteinmetabolismus und einige weitere Enzyme
des Aminos~iurestoft~veehsels in Keimlingen yon Pennisetum typhoides*
W. t Iuber
Institut ffir Botanik der Technischen Universit//t 1V[iinchen, Arcisstr. 2I, D-8 ~iinehen 2, Federal Republic of Germany
Eingegangen am 24. Oktober / 5. November 1974
Influence of NaC1- and Abscisic Acid Treatment on Protein Metabolism and Some Further Enzymes of Amino Acid Metabolism in Seedlings of Pennisetum typhoides
Summary. The effects of sodium chloride (NaC1) and abscisic acid (ABA) on protein synthesis, protein hydrolysis, glutamine synthetase, A-pyrroline-5-carboxylate reduetase and amino acid composition have been studied in aerial parts of Pennisetum typhoides (Stapf et Hubbard) seedlings. Whereas protein synthesis is inhibited by NaC1 and ABA, the activity or the synthesis of leucine-arylamidase, glutamine synthetase and zi-pyrroline-5-carboxylate reductase is promoted by both the substances. The results of the amino acid analysis showed that in the presence of NaCI and ABA the amounts of alanine, aspartafe and ammonia decreased, while the concentrations of glutamate, and especially that of proline increased greatly. The possible significance of the results in explaining the proline accumulation under stress conditions and ABA-treatment is discussed.
Einleitung
Unter StreBbedingungen steigt der endogene Abscisins/iurespiegel in den Pflanzen stark an (Literaturangaben u. a. bei Hsiao, 1973 ; Mizrahi et al., 1970, 1971, 1972). Mit dem ABA-Anstieg unter Stregbedingungen ist eine Ver/~nderung des Aminos/~uregehaltes, vor allem ein Anstieg an freiem Prolin verbunden (Barnett und Naylor, 1966; Singh et al., 1972; u.a.). Den gleiehen Prolinanstieg kann man durch exogene Behandlung der Pflanzen mit ABA erzielen (Aspinall et al., 1973). Wir haben daher sowohl unter NaC1-Streg als aueh nach exogener ABA-Behandlung den enzymatisehen Mechanismus der Prolinanh/~ufung unter- sucht. Es sollte geprfift warden, ob ABA aueh bei NaC1-Behandlung eine Kompo- nente ist, die den Synthesemechanismus des Prolins beeinflussen karm. Da Prolin offensichtlich dureh de novo Syathese aus anderen Aminos~uren syntheti- siert wird (Palfi, 1968; Morris etal., 1969), die durch Proteinabbau entstehen, wurde auch die Proteinsynthese und die Proteinhydrolyse unter ABA-Behandlung oder NaC1-StreB untersueht. Es liegen zwar sehon Befunde vor fiber eine I tem- mung der Proteinsynthese oder eine gesteigerte Proteinhydrolyse unter NaCI-Streg
* Abkiirzung: ABA = Abseisins~ure.

226 W. Huber
(Kahane und Pol jakoff -Mayber , 1968 ; Prisco und O 'Leary , 1970 ; cf. Waisel , 1970) oder A B A - B e h a n d l u n g (Addicot t , 1972; A d d i c o t t und Lyon, 1969; Mizrahi et al., 1972), doch soll ten auch un te r unseren Bedingungen und mi t unserer Pfl~nze diese Vorg~nge un te r such t werden, um eJnen Gesamtf iberbl ick fiber den St icks tof fmeta- bol ismus in Pennisetum typhoides Keiml ingen zu erhal ten.
W~hrend Unte r suchungen fiber die Transaminasen und die Glutaminsi~ure- Dehydrogenase schon anderwei t ig publ iz ier t wurden (Huber und Sankhla , 1973; Sankhla und Huber , 1974) und zu sehr i~hnlichen Ergebnissen nach NaC1-Strel~ wie naeh A B A - B e h a n d l u n g f i ihr ten, soll hier fiber den Pro te inmetabo l i smus und die Pro l insynthese ber ich te t werden.
Material und Methoden
Pflanzenmaterial. Ffir die Untersuchungen wurden 4 Tage alte Keimlinge yon Pennisetum typhoide8 (Burro. f.) Stapf und Hubbard verwendet. Die Keimlinge wurden bei 28 • 1 7 6 und ca. 3000 lx (Osram-L Fluora Lampen) in 9 em Petrischalen auf Filterpapier angezogen, das mi$ 5 ml TestlSsung angefeuehtet worden war. Es wurden 3 Konzentrationen yon w~$riger ABA (Fluka), viz. 3,8 • 10 -~, 7,4 • 10 -~ und 1,5 • 10 -5 M und 3 Konzentrationen yon NaC1, viz. 1,7 • 10 -a, 8,5 • 10 -a und 1,7 • 10-~M als TesblSsung eingesetzt. Die Kontrolle enthielt nur dest. Wasser. Zu den Enzymbestimmungen wurden die BlOtter und Koleoptilen der Keimlinge herangezogen.
Enzymextraktionen. Die Extraktion der A-Pyrrolin-5-Carboxylat (P-5-C)-Reduktase und der Leuein-Arylamidase erfolgte naeh Sankhla und Huber (1974). Zur Isolierung der Glutamin- Synthetase wurde eine besondere Aufarbeitung vorgenommen. Es wurden ca. 1 g BlOtter und Koleoptilen yon Pennisetum typhoides in 2,5 ml Isolierungsmedium (0,1 1~ Phosphatpuffer pH 7,4; 0,375 M Saecharose und 0,001 M EDTA) in einem Potter-Elvehjem ttomogenisator (Potter-S, Braun l~Ielsungen, 1200 Umin) etwa 1 min unter Eiskfihlung homogenisiert. Darauf wurde der Rohexbrakt sofort in einer Sorvall SS 1 Zentrifuge bei 4 ~ C 15 min mit 20000 • g abzentrifugiert und der ~berstand als Enzym verwendet (Loomis, 1959). Die Extrakte Ifir die Enzymbestimmungen wurden jeweils ffiseh bereite$.
Bestimmung der Enzymaktivitiiten und Zusammensetzung der Inkubationsgemische. Die 3-Pyrrolin-5-Carboxylat-Reduktase (EC 1.5.1.2), die Glutaminsynthetase (EC 6.3.1.2) und die Leuein-Arylamidase (EC 3.4.1.--) wurden mit folgenden l~ethoden und Enzymans~tzen gemessen:
1. A- Pyrrolin-5.Carboxylat- Redu~tase Dieses Enzym katalysiert den letzten Sehritt der Prolinbildung, die Reduktion von
A-Pyrrolin-5-Carboxylat zu Prolin mit Hilfe yon NADH oder I~ADPI-I (l~azelis und Fowden, 1969; Splittstoesser und Splittstoesser, 1973). Daher kann das Enzym fiber die Oxidation yon NADH oder NADPH gemessen werden. Vor der Bestimmung des Enzyms muBte zun~chst das d-Pyrrolin-5-Carboxylat hergestellt werden. Dies gesehah durch Synthese aus DL-~- Amino-A-hydroxyvalerians~mre (Splittstoesser und Splittstoesser, 1973). Das Inkubations- gemisch war wie folgt zusammengesetzt: 25 mM A-Pyrrolin-5-Carboxylat; 100 tzmol NADH oder NADPH; 200 n ~ Tris-ttC1-Puffer pH 7,4 und 500 ~zl Enzym in einem Gesamtvolumen yon 2,5 ml. Die Absorptionsi~nderungen bei 340 nm wurden mit einem Zeiss DMR 21 Photo- meter mit Schreiber gemessen. Die Konzentration yon P-5-C nach der Synthese wurde nach Streeker (1971) mit einem Extinktionskoeffizienten yon 2,71 bestimmt.
2. Glutamin-Synthetase Die Glutamin-Synthetase wurde fiber ihre :Nebenaktivi$iit, die ~bertragung des y-Glut-
amylrestes auf Hydroxylamin, gemessen:
Glutamins~iure -{- ATP + NHpOH--* Glutamylhydroxamat + ADP.

EinfluB yon NaC1 und ABA auf Enzyme 227
Das Glutamylhydroxamat bildet mit Eisen(III)-ionen einen Chelatkomplex, der zur kolorimetrischen Bestimmung bei 540 nm herangezogen werden kann (Loomis, 1959). Die Inkubationsmischung enthielt: 120~tmol Tris-HC1-Puffer pH 7,4; 200~mol K-Glutamat; 6 izmol ATP; 60 ~zmol MgSO~; 20 ~xmol Na-Thioglykolat; 200 fxmol Hydroxylamin (neutrali- siert mit KOH) und 500 ~l Enzym. Das Gesamtvolumen war 2 ml. Inkubiert wurde 60 min bei 37 ~ C. Wegen eines eventuellen Abbaus yon ATP dureh ATPasen wurden naeh 20 und naeh 40 min Inkubationszeit jeweils noch einmal 6 ~zmol ATP zugegeben. Einen Vergleichs- wert ergab eine Probe, in die das K-Glutamat erst nach Beendigung der Inkubation hin- zugefiigt wurde.
3. Leucin-Arylamidase
Das Enzym hydrolysier~ eine groBe Anzahl yon Peptiden und Aminosi~ureamiden der L-Konfigurar (Bergmeyer, 1970). Die Ak~ivit~tsbestimmung erfolgte fiber das bei der Hydrolyse yon L-Leucin-p-nitranilid ~reiwerdende p-Nitranilin:
L-Leuein-p-nitranilid-t- H20--> p-Nitranilin ~ L-Leucin.
Das Nitranilin kann bei 405 nm kolorimetrisch bestimmt werden. Die Inkubations- misehung enthielt: 300 9tool Phosphatpu~fer pH 7,2; 2,4 ~tmol Leuein-p-nitranilid und 200 ~1 Enzym in einem Gesamtvolumen yon 3,2 ml. Die Inkubationszeit betrug 30 rain bei 25 ~ C. Eine Blindprobe ohne Enzym wurde mitinkubiert.
Messung der Proteinsynthese. Zur Messung der Proteinsynthese warden ca. 50 mg Blatt- stfieke und Koleoptilen yon e~wa 5 mm L~nge 4 h bei 37~ in einem Medium tolgender Zusammensetzung inkubier~: 4 ~mol ATP; 50 ~mol Tris-HC1/EDTA Puffer pH 7,5 und 1 ~zCi Aminos~uremisehung (i4C-markiert; Radiochemical Center Amersham, CFB 104, spez. Aktivi- t~t 52 mCi/milliAtom Kohlenstoff) in einem Gesamtvolumen yon 0,54 ml. Nach Beendigung der Inkubation wurden die Blattstiicke aus dem Inkubationsmedium enbfernt, 3 real mit H~O gewasehen und ansehliel3end in einem Potter-Elvejhem Homogenisator mit 0,5 ml H20 homogenisiert. Dann wurden die Proben 10 min bei 3000 • g abzentrifugiert und 0,1 ml des t~berstandes, zusammen mit 0,05 ml (200 ~g) Rinderserumalbumin, auf Whatman GF/A Glasfiberfilter (2,1 cm 2~ ) pipettiert. Auf dem Filterpapier wurden die Proteine mit 10% Tri- chloressigs/iure (TCA) ausgef~llt. AnschlieBend wurde 3real mit kalter TCA (5%), einmal 15 rain mit 5% TCA bei 90~ und erneut einmal mit kalter TCA gewaschen. Die Lipide wurden durch zweimaliges Wasehen mit einer Athanol/Diithyl~thermisehung (1:1) und dreimaliges Wasehen mit Aceton entfemt (Jones und Stoddard, 1970). Nach sorgf~ltiger Troeknung der Filterseheibchen warden diese in vorgez~hlte Szintillationsgl~ser in 5 ml Cocktail (5 g PPO, 100 g Naphthalin ad 1000 ml mit Dioxan) gegeben und in einem Berthold- ~'rieseke (BF-5000) Szintillationsziihler gemessen. Zu jedem Experiment wurden Blindproben durehgefiihrt, in die die markierte Aminos~uremischung erst naeh Beendigung der Inkubation hinzugegeben wurde. Um Bakterienkontaminationen auszusehlieBen, wurde bei einigen An- s~tzen Chloramphenieol (10 -3 M) in die Ans~tze gegeben. Dieses Chloramphenieol hatte jedoeh keinerlei Wirkung, so dal3 bakterielle Kontaminationen ausgesehlossen werden kSnnen.
Bestimmung der Aminosiiurekonzentrationen. Etwa 50 mg gefriergetroeknete Bl~ter mad Koleoptilen yon Pennisetum typhoides warden in 2 ml Methanol-Chloroform-Wasser-Gemiseh (12:5:1, v/v) bei Raumtemperatur in einem Potter-Elvejhem Homogenisa~r homogenisiert. Das Homogenat wurde darauf abzentrifugiert (10 min bei 3 000 • g) und der klare ~berstand aufbewahrt. Der Rfickstand wurde noch 2mal mit je 2 ml MCW-Gemiseh bei --20 ~ C je 8 h eluiert. Alle ~0berst~nde wurden danaeh vereinigt und zur Troekenheit eingedampft. Die ge- trockneten Extrakte wurden in 0,021K Citratpuffer pi t 2,2 gelSst mad anschliel~end mit einem Beckmarm LS 120 Aminosiiureanalysator im Einsiulenveffahren aufgetrennt oder das Prolin wurde direkt naeh Singh et al. (1973) im Extrakt bestimm~.
Berechnung der Ergebniase. Die Experimente wurden mindestens 4real wiederholt. Wo immer es m5glich war, wurde die Standardabweichung oder das Signifikanzniveau ange- geben. Die Enzymaktivit~ten der Kontrollen wurden in ~zmol Substratumsatz/g Frisehgewicht rain angegeben, wiihrend die Aktivit~iten der mit NaC1 oder mit ABA behandelten Proben in % des Kontrollwertes dargestell~ wurden.

228 W. Huber
Ergebnisse Wie in anderen yon uns untersuchten Systemen zeigen sich sehr /~hnliche Wirkungen yon ABA- und NaC1-Behandlungen (Huber und Sankhla, 1973; Sankhla und Huber, 1974a, b, c; Huber et al., 1974; Huber und Sankhla, 1974). Auch hier werden cinige Enzyme durch Anzucht der Pflanzen in NaC1- oder ABA-L6sungen aktiviert.
Die Ergebnisse fiber die Wirkungen yon NaC1 oder ABA auf die Synthese oder die Aktivit~t yon A-Pyrrolin-5-Carboxylat-Reduktase zeigen eine deutliche Aktivits des fiir die Prolinsynthese wichtigen Enzyms (Abb. 1). Dutch NaC1 ist die Steigerung wesentlich deutlieher als durch ABA. Bei der h6chsten Salzkonzentration wird eine Aktivit/it yon mehr als 225 % der Kontrolle erreicht. Dies steht im Einldang mit den Prolinbestimmungen, die zeigen, dab durch NaC1- Behandlung die Prolinkonzentration his auf das 10fache ansteigen kann (Tabelle 1), durch ABA jedoch nur auf das 2,5fache. Die P-5-C-Reduktase ist mit NADH ca. 4real aktiver als mit NADPH (Abb. 1), jedoch bewegt sich die Steigerung der Enzymaktiviti~t mit beiden Pyridinnukleotiden in der gleiehen Gr6Benordnung.
Neben der Prolinsynthese wird aber auch die Glutaminsynthese beeinfluBt. Auch hier ist eine Aktivierung durch NaC1 und durch ABA festzustellen, wobei die It6chstwerte bei NaC1 wieder h6her liegen als bei ABA. Wi~hrend durch die h6chste NaC1-Konzentration eine Steigerung der Enzymwirkung um 188% er- zielt wird, gelingt dies mit ABA nur um etwa 150% (Abb. 2).
Sowohl durch NaC1 als auch durch ABA wird die Proteinhydrolyse ver- st&rkt. Die Leucin-Arylamidase wird durch NaC1 urn 60% und durch ABA um 150% bei der h6chsten Konzentrat ion aktiviert (Abb. 3).
Mit dem vergr6Berten Proteinabbau ist eine verringerte Proteinsynthese korreliert. Wie Abb. 4 sichtbar macht, finden wir sowohl durch ABA als auch durch NaC1 eine Verringerung der Proteinsyntheseaktivit~t. Der EinfluB yon NaC1 ist deutlich ausgepriigter als der yon ABA.
Die Ergebnisse der Aminos/~ureanalyse sind in Tabelle 2 dargestellt. Aus den Befunden wird klar, dab unter unseren Bedingungen die Konzentrationen yon Alanin, Asparagins/iure und Ammonium abnehmen. W/~hrend die Glutamin- s/iurekonzentration nur leicht ansteigt, ist eine deutliche Zunahme der Prolin- konzentration zu erkennen. Auch hier wird sichtbar, dab der EinfluB des Salz- stresses viel gr6Ber ist als der der exogenen ABA-Gabe.
Tabelle 1. Prolinkonzentra~ionen in den mit ABA- oder NaC1 behandelten Pflanzen
Behandlung der Keimlinge ~g Prolin/mg Trockengewicht % Werte
Kontrolle 1,58 100 3,8 • 10 -6 M ABA 2,1 a 132,9 7,4 • 10 -6 M ABA 6,0 c 380,4 1,5 • 10 -~ 1YI ABA 4,1 b 258,2 1,7 • 10 -3 M NaC1 2,6 a 165,2 8,5 • 10 -81YI NaC1 8,5 c 538,0 1,7 • 10 -2 M NaC1 16,3 c 1031,0
a-c Signifikant verschieden von dem Kontrollwert mit P = 0,05, 0,01 und 0,001.
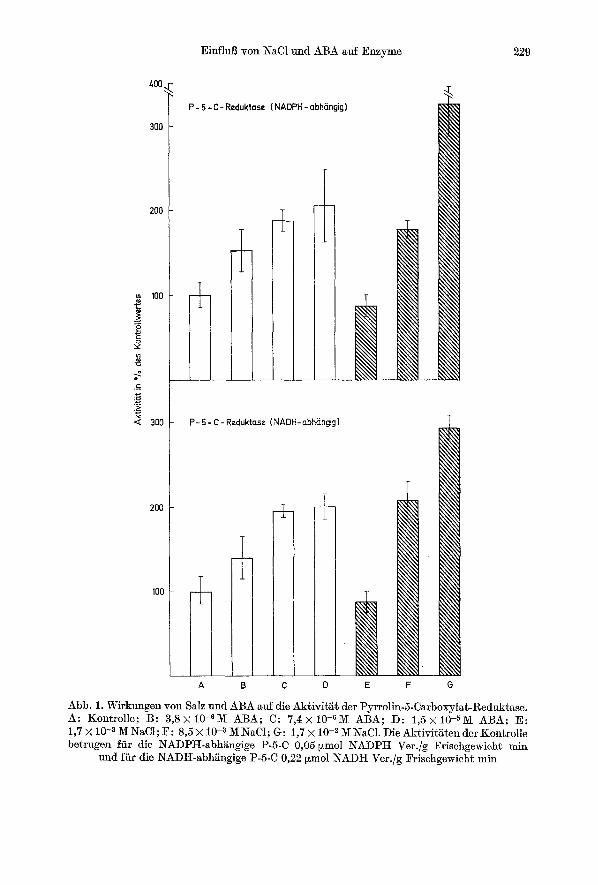
EinfluB yon NaC1 und ABA auf Enzyme 229
/.00
300
200
100 2
-o
.c
.>
< 300
200
100
P - 5 - C - Reduktosr ( NADPH- abhangig)
+
+ t
P - 5 - C - Rcduktose ( NADH - abh;Hngig )
i i
A B C D
11
E F G
Abb. 1. Wirkungen yon Salz und ABA auf die AktivitE~ der Pyrrolin-5-Carboxylat-Reduktase. A: Kontrolle; B: 3,8• 10-617[ ABA; C: 7,4x 10-6M ABA; D: 1,5• 10-sM ABA; E: 1,7 X 10 -a M NaCI; F: 8,5 x 10 -a M NaC1; G: 1,7 • 10 -2 M NaC1. Die AktivitEten der Kontrolle betrugen ffir die NAI)PH-abhfingige P-5-C 0,05 Fmol NADPH Ver./g Frischgewicht rain
und fiir die NADIt-abhi~ngige P-5-C 0,22 Fmol NADH Ver./g Frischgewicht min
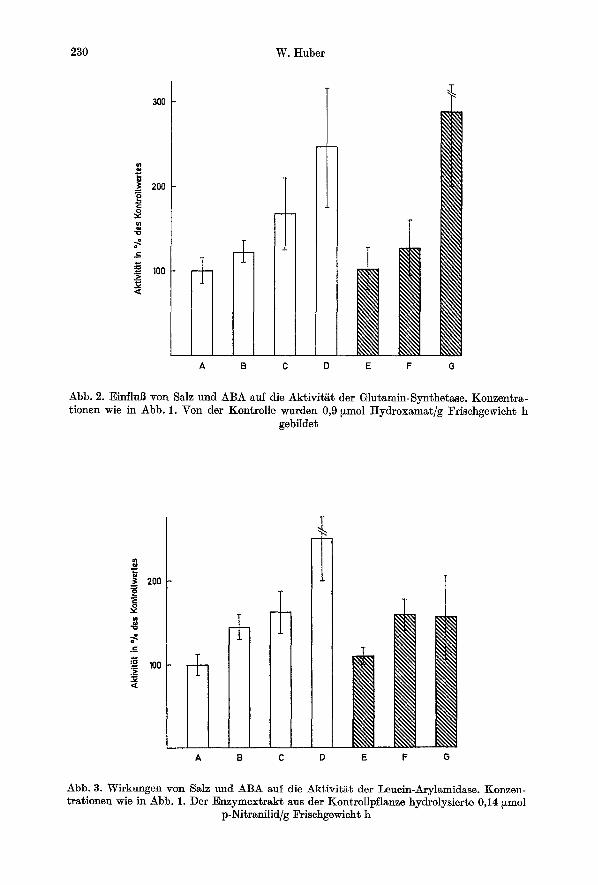
230 W. Huber
==
200
.~_
100 e:
+ §
A B C D E F G
Abb. 2. Einflul3 yon Salz und ABA auf die Aktivit~t der Glutamin-Synthetase. Konzentra- tionen wie in Abb. 1. Von der Kon~rolle wurden 0,9 ~mol Hydroxamat/g Frischgewieh~ h
gebilde~
~= 2oo
. .e
. E
1oo
A
t t
E B C O F G
Abb. 3. Wirkungen yon Salz und ABA auf die Aktivit~t der Leuein-Arylamidase. Konzen- trationen wie in Abb. 1. Der Enzymextrakt aus der Kontrollpflanze hydrolysierte 0,14 ~mol
p-Nitranilid/g Frisehgewicht h
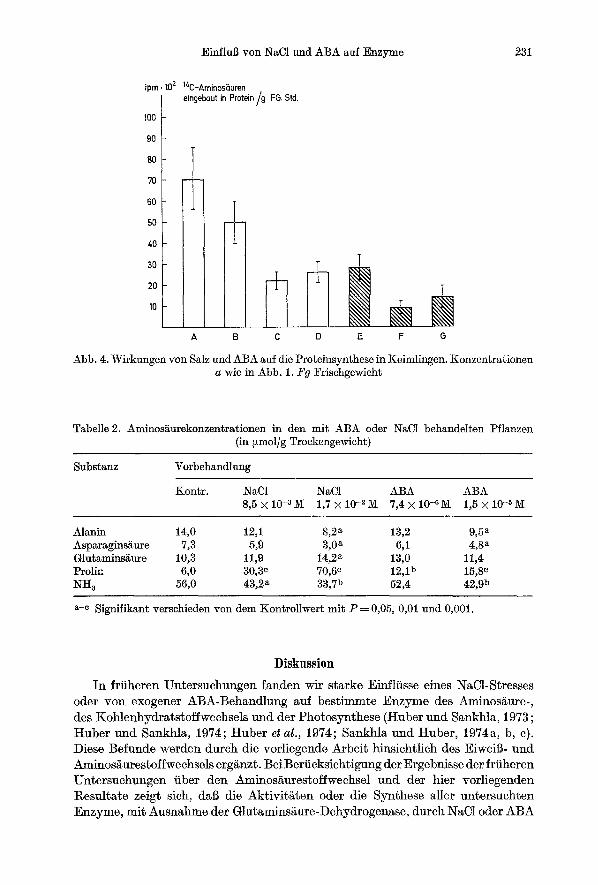
Einfluil yon NaC1 und ABA auf Enzyme 231
ipm- 102 14C-Aminosiiuren eingebaut in Protein/g FG. Std.
100
90
80
?(3
60
50
Z,0
30
20
10
A B C D E F G
Abb. 4. Wirkungen yon Salz und ABA auf die Proteinsynthese in Keimlingen. Konzentrationen a wie in Abb. 1. Fg Frischgewicht
Tabelle 2. Aminos~urekonzentrationen in den mit ABA oder NaC1 behandelten Pflanzen (in ~mol/g Trockengewicht)
Substanz Vorbehandlung
Kontr. NaC1 NaC1 ABA ABA 8,5 • 10 -3 IV[ 1,7 X 10 -2 !V~ 7,4 X 10 -8 M 1,5 • 10 -5
Alanin 14,0 12,1 8,2 a 13,2 9,5 a Asparagins~ure 7,3 5,9 3,0 a 6,1 4,8 a Glutamins~ure 10,3 11,9 14,2 a 13,0 11,4 Prolin 6,0 30,3c 70,6 e 12,1 b 15,8 c Ni t 3 56,0 43,2 a 33,7 b 52,4 42,9 b
a-c Signifikant verschieden yon dem Kontrollwert mit P = 0,05, 0,01 und 0,001.
Diskussion
I n frf iheren Unte r suchungen f anden ~ r s t a rke Einflfisse eines NaC1-S~resses oder yon exogener ABA-Behand lung auf bes t immte E n z y m e des Aminos/~ure-, des Kohlenhydra t s to f fwechse l s und der Pho tosyn these (Huber und Sankhla , 1973 ; H u b e r und Sankhla , 1974; H u b e r et al., 1974; Sankh la und Huber , 1974a, b, c). Diese Befunde werden durch die vor l iegeade Arbe i t hinsicht l ich des Eiwei6- und Aminos~urestoffwechsels erg/~nzt. BeiBerf ieks ieht igung der Ergebnisse tier frf iheren Ua te r suchungen fiber den Aminos~urestoffwechsel und tier b ier vor l i egeaden Resu l t a t e zeigt sich, da$ die Aktivi t /~ten oder die Synthese aller un te r sueh ten Enzyme , mi t Ausnahme der Glu tamins~ure-Dehydrogenase , durch NaC1 oder A B A

232 W. Huber
im Anzuchtsmedium stimuliert werden. Wahrscheinlich wird die Synthese ver- st/~rkt, da wir keinen direkten EinfluS beider Substanzen auf die Enzyme in vitro messen konnten.
Die vermehrten Syntheseraten unter diesen Bedingungen erschienen uns zunachst unsinnig, da ja die Proteinsynthese durch NaC1 und ABA gehemmt wird. Auch die verst&rkte Proteinhydrolyse past scheinbar nicht in dieses Bild. Eine vergrSSerte Synthese der Transaminasen z.B. wiirde bei Wirkung der Enzyme in l~ichtung Alanin- bzw. Asparagins/~uresynthese nur zu einem unn6tigen Anstau dieser Aminosauren ffihren, da diese bei der Proteinsyathese nieht mehr ver- wertet werden k6rmen. Wit haben daher die Itypothese aufgestellt, daS die ver- mehrte Synthese der Transaminasen nur sinnvoll ist, wenn angenommen wird, daS die Transaminasen in Riehtung yon Alanin- oder Asparaginsaureabbau wirken, daS also vermehrt Glutamins/~ure gebildet wird, die eine zentrale Position im Aminos/~urestoffwechsel einnimmt, auch in Bezug auf die Prolinsynthese (Huber und Sankhla, 1973; Sankhla und Ituber, 1974a). Diese Itypothese wird dutch die Befunde der vorliegenden Arbeit weitgehend gestfitzt.
Die Aminos/~urebestimmungen zeigten n/~mlich eine deutliche Abnahme der Alanin- und Asparagins/~urekonzentrationen. Der Befund fiir die Asparagins/~ure ist allerdings nieht ganz eindeutig, da auch Asparagin gebildet worden sein k6nnte. Zwar kormten wit weder eine Zunahme der Asparagins/iurekonzentration noch eine Aktivit~t der Asparagin-Synthetase messen (Ergebnisse nieht dargestellt), aber nach Untersuehungen yon Barnett und Naylor (1966) und Singh et al. (1973) steigt das Asparagin nnter SalzstreS stark an. Die nut leichte Zunahme der Glutamins/~ure ist damit zu erkl/~ren, alas es sieh bei dieser Substanz nicht um ein Endprodukt handelt, sondern daS aus Glutamins/~ure noch andere Substanzen gebildet werden. Durch die Zunahme der Syatheseraten tier Glutamin-Synthetase und der Pyrrolin-5-Carboxylat-Reduktase karm die GlutamJns/~ure zu Glutamin und vor allem zu Prolin umgebildet werden. Diese Verschiebung des Amino- s/~urestoffweehsels in Riehtung Prolinsynthese wird auch dadurch begfinstigt, daS gleichzeitig die Syatheserate der Glutamins/iure-Dehydrogenase erniedrigt wird, daS also eine Desaminierung der Glutamins~ure mit gleichzeitigem Freiwerden yon toxisehem Ammoniak verhindert wird (Huber und Sankhla, 1973; Sankhla und Huber, 1974).
Es 1/~uft somit sowohl unter SalzstreS als auch naeh exogener ABA-Behand- lung in den Pflanzen ein sehr sirmvoller, gleichartiger MechanJsmus ab, der es diesen ermSglieht, die Mangelsubstanz Stickstoff zun/~chst festzuhalten (Abb. 5). Dieser Mechanismus ist sicher nur bis zu einer bestimmten StreSst/~rke wirksam. ErhSht man n/~mlich die NaC1-Konzentration fiber einen bestimmten Bereich, so wird tier Aminos/~urestoffwechsel pathologiseh und die Prohnkonzentration sinkt wieder ab (Kreutmeier, Ituber und Sankhla, unverSffentlicht). Warum in den Pflanzen haupts/~chlich das Prolin unter StreSbedingungen angestaut wJrd, bleibt noch unklar. Doeh gibt es versehiedene Erkl/~rungsmSgliehkeiten. Einmal soll Prolin weniger toxisch als andere Aminos&uren sein (Levitt, 1972), es so!l leiehter transportiert werden k6rmen (Splittstoesser und Splittstoesser, 1973) und es kann dem Stoffweehsel nach Beendigung der StreSeinwirkung wieder sehr sehnell dureh Uberffihrung in Glutamins/~ure zug/~nglieh gemaeht werden. Sowohl
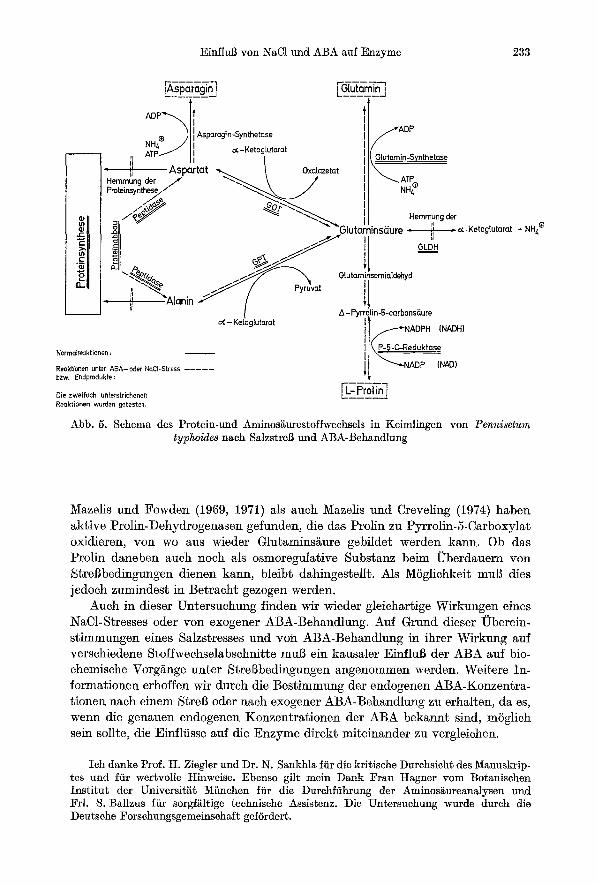
EinfluB yon NaCI und ABA auf Enzyme 233
u____r __o____J
A D P " ~ NH~ / Aspctmgin-Synthetase ~//ADP A T P / r
'l "~.~ ~ Oxo l Glutamin-Synthetose - w j Aspctrtot Oxal~.etat ~ . ~ Hemmung der ...,.~ ~ ~ ..,,~ AW ProteinsyntEese/" NH40
~-~1 ~1 "~']'~ ~-~ "~ ~"~"~ PY r u~vO-t �9 Hemmung der GlotomlflS~iure r, . NHz| ...----.~--~j or Ket og[utarat
~ . - . ~ I ..-J~ iI GLDH
-%\ \ .,o,=,.~ II A - Pyrrqlin-5-carbonsaure
c~- KetogIutarat ] =NADPH (NADH)
i-;, ------..-:--I Die zweifoch unterstrich . . . . LL-P._r2Un~ Reaktionen wurden getesteL
Abb. 5. Schema des Protein-und Aminos~urestoffwechse]s in Kcim]ingen yon Pennisetum typhoides nach SalzstreB and ABA-Behandlung
Mazelis und Fowden (1969, 1971) als auch Mazelis und Creveling (1974) haben aktive Prolin-Dehydrogenasen gefunden, die das Prolin zu Pyrrolin-5-Carboxylut oxidieren, yon wo aus wieder Glutamins/ture gebfldet werden kann. Ob das Prolin daneben such noeh Ms osmoregulative Substanz beim Uberdauem yon Stregbedingungen dienen kann, bleibt dahingestellt. Als M6glichkeit mug dies jedoch zumindest in Betracht gezogen werden.
Auch in dieser Untersuehung finden wir wieder gleiehartige Wirkungen eines NaC1-Stresses oder yon exogener ABA-Behandlung. Auf Grund dieser l~berein- st immungen eines Salzstresses und voJa ABA-Behandlung in ihrer Wirkung auf versehiedene Stoffweehselabschnitte mug ein kausaler Einflug der ABA auf bio- ehemische Vorg~nge unter Strel3bedingungen angenommen werden. Weitere In- tormationen erhoffen wit dutch die Bestimmung der endogenen ABA-Konzentra- tionen naeh einem Streg oder nach exogener ABA-Behandlung zu erhalten, da es, wenn die genauen endogenen Konzentrationen der ABA bekarm~ sind, m6glich sein sollte, die Einfliisse auf die Enzyme direkt miteinander zu vergleiehen.
Ich danke Prof. H. Ziegler und Dr. N. Sankhla ffir die kritische Durchsicht des Manuskrip- tes und fiir wertvolle Hinweise. Ebenso gilt mein Dank Frau Hagner yore Botanischcn Institut der Universitgt Mfinchen flit die Durctffiihrung der Aminosaureanalysen und Frl. S. Ballzus ffir sorgfaltige technische Assistenz. Die Untersuchung wurde dutch die Deutsche Forschungsgemeinschaft gefSrdert.

234 W. Huber
Literatur
Addicott, F. T.: Biochemical aspects of abscisic acid. In: Plant growth substances 1970, D.J. Carr, ed., p. 272--280. Berlin-Heidelberg-New York: Springer 1972
Addicott, F. T., Lyon, J. L. : Physiology of abseisic acid and related substances. Ann. Rev. Plant Physiol. 29, 139--164 (1969)
Aspinall, D., Singh, N. T., Paleg, L. G.: Stress metabolism. V. Abscisic acid and nitrogen metabolism in barley and Lollum temulentum L. Aust. J. biol. Sci. 26, 319--327 (1973)
Barnett, N. M., Naylor, A.W. : Amino acid and protein metabolism in Bermuda grass during water stress. Plant Physiol. 41, 1222---1230 (1966)
Bergmeyer, H. U. (Ed.): Methoden der enzymatischen Analyse. Weinheim: Verlag Chemic 1970
Hsiao, T. C. : Plant responses to water stress. Ann. Rev. Plant Physiol. 24, 519-570 (1973) Huber, W., Sankhla, N.: Eco-physiologieal studies on Indian arid zone plants. II . Effects
of salinity and gibberellin on the activity of the enzymes of amino-acid metabolism in leaves of Pennisetum typhoides. Oecologia 13, 271--277 (1973)
Huber, W., Rustagi, P. N., Sankhla, N. : Eeo-physiological studies on Indian arid zone plants. I II . Effect of sodium chloride and Gibberellin on the activity of the enzymes of carbo- hydrate metabolism in leaves of Pennlsetum typhoides. Oecologia 15, 77-86 (1974)
Huber, W., Sankhla, N. : Untersuehungen fiber den Einflu~ yon Abscisin- und Gibberellin- s~urebehandlung auf die Aktivit~ten einiger Enzyme des Kohlenhydratstoffwechsels in Bl~ttern yon Pennisetum typhoides Keimlingen. Planta (Berl.) 116, 55--64 (1974)
Jones, T. W. A., Steddart, J. L. : Gibberellin induced changes in protein synthesis and enzyme activity in shoot apices of Tri/olium pratense. J. exp. Bot. 21, 452---461 (1970)
Kahane, J., Poljakoff-Mayber, A. : Effects of substrate salinity on the ability for protein synthesis in pea roots. Plant Physiol. 43, 1115--1119 (1968)
Levitt, J . : Responses of plants to environmental stresses. New York-London: Academic Press 1972
Loomis, W. D. : Amide metabolism in higher plants. III.Distribution of glutamyl-transferase and glutamine synthetase activity. Plant Physiol. 34, 541--546 (1959)
Mazelis, M., Creveling, R. K.: L-Proline dehydrogenase of Triticum vulgare germ: Purification, properties and eofactor interactions. Phytochemistry 13, 559--565 (1974)
Mazelis, M., Fowden, L.: Conversion of ornithine into proline by enzymes from germinating peanut cotyledons. Phyteehemistry 8, 801--809 (1969)
Mazelis, M., Fowden, L. : The metabolism of proline in higher plants. II . L-proline dehydro- genase from cotyledons of germinating peanut (Arachis hypogea L.) seedlings. J. exp. Bot. 22, 137--145 (1971)
Mizrahi, Y., Blumenfeld, A., Bittner, S., Richmond, A. E.: Abscisic acid and cytokinin content of leaves in relation to salinity and relative humidity. Plant Physiol. 48, 752--755 (1971)
Mizrahi, Y., Blumenfeld, A., Richmond, A. E.: Abseisic acid and transpiration in leaves in relation to osmotic root stress. Plant Physiol. 46, 169--171 (1970)
Mizrahi, Y., Blumenfeld, A., Richmond, A. E.: The role of abseisic acid and salination in the adaptive response of plants to reduced root aeration. Plant & Cell Physiol. 18, 15--21 (1972)
Morris, C.J., Thompson, J. F., Johnson, C. M. : Metabolism of glutamic acid and N-acetyl- glutamic acid in leaf discs and cell free extracts of higher plants. Plant Physiol. 44, 1023--1026 (1969)
Palfi, G. : Changes in the amino acid content of detached wilting leaves of Solanum la~iniatum Air. in the light and in the dark. Aeta Agron. Acad. Sei. Hung. 17, 381--388 (1968)
Prisco, J .T. , O'Leary, J . W . : Effect of salt and water stresses on protein synthesizing capacity of embryo-axis of germinating Phaseolus vulgaris L. seeds. Rev. Brasil. Biol. 86, 317--321 (1970)
Sankhla, N., Huber, ~u Enzyme activities in Pennisetum seedlings germinated in the presence of abscisic and gibberelic acids. Phytochemistry 18, 543--546 (1974a)
Sankhla, N., Huber, W. : Effect of abscisie acid on the activities of photosynthetic enzymes and l~CO~-fixation products in leaves of Pennisetum typhoide8 seedlings. Physiol. Plant. 30, 291--294 (1974b)

Einflu~ yon NaC1 und ABA auf Enzyme 235
Sankhla, N., Huber, W.: Eco-physiological studies on Indian arid zone plants. IV. Effect of salinity and gibberellin on the activities of photosynthetic enzymes and 14C02-fixation products in leaves of Pennisetum typhoides seedlings. Bioch. Physiol. Pflanzen (ira Druek) (1974c)
Singh, T. N., Paleg, L. G., Aspinall, D. : Stress metabolism I. Nitrogen metabolism and growth in the barley plant during water stress. Aust. J. biol. Sei. 26, 45--56 (1973)
Splittstoesser, S. A., Splittstoesser, W. E. : Pyrroline-5-carboxylate reductase from Cucurbita cotyledons. Phytochemistry 12, 1565--1568 (1973)
Strecker, H.J . : The preparation of animal proline oxidase (Rat liver), and its use for the preparation of A-Pyrroline-5-carboxylate. In: Methods of enzymology, Tabor, H., Tabor, C. W., eds., vol. 17 B, p. 251--252. New York: Academic Press 1971
Waisel, Y. : Biology of halophytes. London: Academic Press 1972





























