Embed Size (px)
Citation preview

uber die spezifische Tatigkeit der Lungen bei der respiratorischen Gasaufnahme und ihr Verhalten zu der
durch die Alveolarwand stattfindenden Gasdiffusion. Von
Christian Bohr.'
Uurch die seit 1888 von dem hiesigen Laboratorium veriiffentlichten Untersuchungen ist festgestellt. worden, dal3 die Lunge bei der Gas- aufnahme eine aktive Rolle spielt. Dieser Umstand ist selbstrerstand- lich von weittragender Bedeutung fur das Verstindnis der respirato- rischen Vorgiinge uberhaupt, und zwar besonders fiir das Verstindnis der Mittel, die der Organismus zu deren Regulierung benutzt.
Eine trotz verschiedener darauf gerichteten Untersuchungen bisher nicht hinllnglich aufgeklarte Frage hierbei ist es indes, wie hoch die nnzmeifelhaft durch die Alveolarwand stattfindende Gasdiffusion zu schiitzen ist. Nur hierdurch wird man Aussicht erhalten, den quant i - ta t iven Umfang der spezifischen Tatigkeit des Lungengewebes wiihrend der wechselnden Lagen des Organismus zu ermitteln.
Die folgenden Untersuchungen haben hauptsichlich diese Frage im Auge, indem sie die Bedingungen der physikalischen Gasdiffusion und die effektive Bedeutung dieses Faktors unter variierenden Ver- hiiltnissen zu bestimmen suchten. Als Einleitung hierzu wird es jedoch am Platze sein, eine kurze Darstellung dessen zu geben, was bisher in den Hauphugen iiber die spezifische Tiitigkeit der Lunge klar- gestellt worden ist.
A bsohnit t I. Spezilasche Ttitigkeit der Lunge.
Die aktive Rolle der Lunge wihrend der respiratorischen Vor- gange spielt sich dem bisher Ermittelten zufolge in zwei Richtungen
Der Redaktion am 30. April 1909 zugegangen. Vgl. Nagel , Hmdbuch der Physiologie. I. I. S. 156-157.

222 CHRISTIAN BOHR :
ab; teils hat sie Bedeutuug fur die Wanderung der Gase durch die Alveolarwand, teils vermittelt sie in wechselndem Umfanp einen be- sonderen respiratorischen ProzeB, mittels dessen intermediare Stoff- wechselprodukte aus den Geweben wahrend eines in der Lunge selbst, stattfindenden Verbrauchs von Sauerstoff, bzw. einer Bildung yon Kohlensaure, umgesetzt werden. Die letztere Form der Tiitigkeit werden wir vorerst betrachten.
8 1. Anteil der Lunge an der inneren Atmnng.
Jndem wir mit Bezug auf die Einzelheitcn der Technik und die Ergebnisse der mehr speziellen Versuche auf die originalen A bhand- lungen 1 und die in N agels Handbuch gegebene zusammenfassende Obersicht verweiseq2 werden wir hier wesentlich nur die Grundlinien der Untersuchungen uber den in der Lunge selbst stattfindenden Respirationsproze6 aufzeichnen.
\Venn man an einem Tiere gleichzeitig einerseits die wahrend der Zeiteinheit durch die Lunge aufgenommene Menge (0) Sauerstoff und anderseits den prozentigen Sauerstoffgehalt sowohl der Venen- (v) als des Arterienblutes (a) und au6erdem die Menge (M) des wihrend der Zeiteinheit die Lunge passierenden Blutes bestimmt, so hat man die erforderlichen Daten, um zu entscheiden, ob der gesamte, wahrend einer gegebenen Zeit von der Lunge aufgenommene Sauerstoff in dem derselben entstromenden Arterienblute wirklich wiedergefunden wird; man mii6te dann natiirlich haben:
a i v 100
0 = M . -. 1st dies nicht der Fall, und findet man im Versuche, daS
so ist ein Teil des Sauerstoffes zu Vorgangen verbraucht worden, die in der Lunge selbst erfolgen. Ganz analoge Bestimmungen lassen sich selbstverstiindlich gleichzeitig mit Bezug auf die Kohlensaure ausfuhren. -
Duroh derartige Untersuchungen fanden Henriques und ich, da6 der spezielle Anteil der Lunge an dem gesamten Sauerstoffver- brauch des Organismus von ganz unbedeutenden Werten an bis iiber _.
Bohr und Henriques, Archiv d. Physiol. 1897. S. 590. * Bohr, Nagels Eundbuch. I. I. S. 187.

OBER DIE SPEZIFISCRE TATIGKEIT DER LUNGE USW. 223
60 Prox. rariierte; im Mittel fanden etwa 33 Proz. des Umsatzes in der Lunge statt. Der respiratorische Quotient ( C 0 2 / 0 2 ) fur den in der Lunge erfolgten Umsatz war sehr variabel.
Nachdem das Vorhandensein einer derartigen spezifischen Lungen- tatigkeit mithin nachgewiesen worden war, suchten wir deren Abhangig- keit von rerschiedenen HuBeren Bedingungen naher zu bestimmen, indem wir besonders den Vorgang wahrend moglichst normaler Ver- hiiltnisse un tersuchten. Wir sahen deshnlb in einer folgenden Ver- suchsreihel von der Messung der Menge des die Lunge durch- striimenden Blutes ab und beschrinkten uns darauf, den respiratorischen Quotienten zu berechnen, und zwar einerseits mittels der Zusamrnen- setzung der ein- und der ausgeatmeten Luft, anderseits aus dem pro- zentigen Gehalt an Sauerstoff und Kohlensiure in den gleichzeitig entnommenen Blutproben aus dem rechten und dem linken Herzen.
Sind die mittels dieser beiden Berechnungen gefundenen W'erte des respiratorischen Quotienten voneinander verschieden , so muS in der Lunge selbst ein Umsatzvorgang stattgefunden haben, und zwar mit einem Quotienten, der von dem des respiratorischen Umsatzes in den Geweben verschieden ist; erweisen die auf diesen beiden ver- schiedenen Wegen berechneten Quotienten sich dagegen als gleichgrol3, so konnen wir, wie leicht ersichtlich, in dieser Weise iiber die hier zu untersuchende Funktion Sjchts erfahreo, obschon sie auch in diesem Falle sehr wohl rorhanden gewesen sein kann, sofern sie mit dem- selben Quotienten wie der Umsatz in den Geweben erfolgt ist. -
Yun findet man in fast allen Fallen Verschiedenheit der Quo- tienten; so war in drei verschiedenen Versuchen an normalen Hunden der Quotient im Blute stets niedriger als der in der Atemluft, und zwar in einer Ausdehnnng, die es nicht zullssig macht, die Ver- schiedenheit durch analytische Fehler zu erkliiren, wenn man beachtet, daB der Pehler der Blutgasbestimmungen bei den hierzu in Anwen- dung gebrachten exakten Methoden weniger als 0.05 wm pro 100 wm Slut betrlgt; flir die groBe Genauigkeit der Versuche sprechen auch in hohem Grade die in den einzelnen Proben dee Blutes gefundenen Werte des absorbierten Stickstoffes. hiittels Ferschiedener Eiogriffe lassen die Verhiltnisse der Quotienten sich indern; so ist nach Blutentziehung der aus den Rlutproben berechnete respiratorische Quotient griifler als der in der Atemluft gefundene, und unter Urntitinden kiinnen die Bus- schlage so groB werden, da6 man, um ihre Realitit zu bezweifeln,
Bohr und Henr iques , a. a. 0. S. 819.

224 CHRISTIAN BOHR :
gezwungeii sein murde, Fehler ron mehreren Kubikzen timeter Gas pro 100 cenl Blut vorauszusetzen.
Auch ein Vergleich der aus dem Blute mit den aus der Atem- luft berechneten respiratorischen Quotienten ergibt also mit Sicherheit, dab io der Lunge selbst ein Sauerstoffrerbrauch, bzm. eine Kohlen- siiurebildung vorgeht, und nach dem Ergebnisse samtlicher Versuche la& sich, was die Existenz einer solchen Funlition betrifft, wie es mir scheint,, kein berechtigter Zmeifel erheben.
Mit Bezug auf die Bedeutung, welche dieser spezifischen Lungen- tatigkeit fur den Haushalt des Organismus zuzuschreiben ist, brachten Versuche uber die GrijBe des Stoffwechsels nach einer hohen dorta- absperrung, die wir gleichzeitig veroffentlichten, auch iiber diesen Punkt Aufschlusse. Es erwies sich niimlicli, daB der Stoffwechsel sich nach einem derartigen Eingriffe eine Zeitlaug fast auf seiner anfiinglichen Hohe erhielt, obgleich die Hauptblutbahn nach dem groBten Teile der Gewebe des Organismus hierdurch Ferschlossen wurde ; der langsame, sickernde Kollateralkreislauf, der unter diesen Verhaltnissen durch die abgesperrten Partien stattfindet, genugt, um den Stoffmechsel fast auf normaler Hohe zu erhalten, ist hierfiir aber auch eine absolut not- mendige Bedingung. Hebt man namlich den Kreislauf durch die abgesperrten Partien v o 11 s t ii n d i g auf, indem man zugleich die Vena cava sperrt, so sinkt der Stoffwechsel sofort bis auf ganz minimale Werte. Da diejenige Menge Sauerstoff (hochstens 20 wm in 100 ”), die wahrend der Aortaabsperrung den betreffenden Geweben mittels des geringen Blutstromes zugefiihrt wird, durchaus nicht ge- nugt, um den gesamten beobachteten Sauerstoffverbrauch zd decken, hat man die Erkliirung der hier besprochenen Erscheinungen darin zu suchen, daB sich in den abgesperrten sauerstoffarmen Teilen inter- mediiire Stoffwechselprodukte bilden, die mit dem durchsickernden Blutstrom nach der Lunge gefubrt werden und hier, wo der Sauer- stoff reichlichen Zutritt findet, nnter Kohlensaurebildung in die schlieb lichen Produkte oxydiert werden.
Dieses Vermogen der Gewebe, unter Umstiinden trotz des Mangels an Sauerstoff den Stoffwechsel dnrch Bildung intermediarer Produkte zu unterhalten, macht uns die Bedeutung des in der Lunge selbst stattEndenden Umsatzprozesses klar, indem un ter anderen hierdurch offenbar einem MiBrerhaltnisse zmischen dem Stoff mechsel der Gewebe und der Sauerstoffzufuhr abgeholfen werden kann. Wahrend der Aortaabsperrung war es in der Tat Erstickungsblut, vas aus den Geweben den Lungen zugefiihrt und hier mit dem Sauerstoff in Be- ruhrung gebracht wurde, und bei den resultierenden Oxydationsprozessen

OBER DIE SPEZIFISCHE TATIGKEIT DER LUNGE usw. 225
mussen die Zellen der Lunge irgendwie eine mitwirkende Rolle gespielt haben, denn nur hierdurch wird es erklarlich, daB man in A. Schmid ts bekannten Versuchen, wo Erstickungsblut auBe r h a1 b des Organismus mit Sauerstoff geschuttelt murde, keine intermediaren Stoffwechselpro- dukte in nennenswerter Nenge nachzuweisen vermochte.1
Bus dem Folgenden wird hervorgehen, daB die nahere Betrachtung der Kreislaufverhaltnisse wahrend der Muskelarbeit uns zu der iuBerst wichtigen Polgerung bewegt, daB auch mihrend einer forcierten Arbeit ein Zustand entsteht, wahrenddessen der Organismus in reichem bTaBe die Fahigkeit der Gewebe verwertet, mittels intermediiirer, erst in der Lunge vollig oxydierter Stoffwechselprodukte die gesteigerten dnspriiche zu befriedigen, welche die Arbeit an die Energieentwicklung stellt.
Bevor wir aber hierzu ubergehen, miissen wir vorerst in Kiirze die Kritik betrachten, die an den von H e n r i q u e s und mir an- gestellten Versuchen und an den Schliissen, die wir aus diesen zogen, geiibt worden ist.
Diese Kritik steht von Anfvng an im Zusammenhang mit dem Einflusse, den unsere Anschauungen notwendigerweise auf die An- wendbarkeit einer bekannten Methode zum Messen des die Lunge dnrchstromenden Blutes erhalten mu6ten ; mittels dieser Methode sucht man nimlich die Menge des wlhrend gegebener Zeit die Lunge durch- strcimenden Blutes aus der Menge Sauerstoff zu berechnen, die wlhrend derselben Zeit ans der Einatmungsluft aufgenommen wurde, indem man diese Menge mit dem prozentigen Gehalt des Venen- nnd Arterien- blutes an dem genannten Gase zusammenhalt. Ein solches Verfahren ist berechtigt, solange man, mie es der Fall war, als Fick dasselbe vorschlug und Gr6hant und Quinqand es zum erstenmal in An- wendung brachten, keinen Grund for die Annahme hat, da6 in der Lunge selbst ein belangreicher Sauerstoffverbrauch stattfinde. Seitdem ein Umsatz letzterer Art in experimenteller Weise nachgewiesen wurde, ist indes der Methode ihre Grundlage vollig entzogen. Wenn Zun t z a gie nichtsdestoweniger zu denjenigen Bestimmungen der Geschwindig- keit des Kreislanfes bei Pferden benutzt, welche die Basis der von ihm berechneten Werte fir den Anteil des Herzens am Stoffwechsel bilden, so mu6 er von der Voraussetzung ansgehen, da6 den Egeb- nissen, zu denen Henriqnes nnd ich, wie oben beschrieben, gelangten, jede Realitiit abgehe. Eine solche Annahme scheint mir in der Tat aber jeglicher Berechtigung zu entbehren. Freilich fiihrt Znntz an, die von
Vgl. Bohr, Nagels Hudbuch. I. I. S. 186. Zuntz und Hagemann, Stoffwechsel dea Pferdes. Berlin 1898. S. 371 ff.
Skandin. Archiv. XXII. 15

226 CHRISTIAN BOHR:
uns dem Venenblut entnommenen Proben entspriichen nicht dein wid;- lichen Durchschnitt desselben; dies ist jedoch nicht zutreffend, wenn man, mie es in unseren Versuchen geschah, eine langere Zeit hindurch Blut- proben mit ganz regelmiBigen Zwischenraumen entnimmt ; bei einer solchen Anordnung nihssen die Proben wirkliche Durchschnittsproben nerden.
Nicht einmal die von Xun t z selbst gefundenen experimentellen Resultate - denn auch in seinen Versuchen' gibt es Versehiedenheit der respiratorischen Quotienten, die teils aus Blutproben, teils aus der gleichzeitig genommenen Atemluft berechnet wurden - lassen sich mit irgend welcher Wahrscheinlichkeit allein durch eine mangelhafte Probenentnahme erklaren, obschon die Proben hier auf einmal ge- nommen wurden und nicht, mas bei weitem vorzuziehen ist, nach und nach wahrend des Verlaufs eines verhaltnisma6ig liingeren Zeitraums, wie in den von H e n r i q u e s und mir angestellten Versuchen. Betreffs dieser Seite der Sache wie auch betreffs des Nachweises, daB gewisse unserer Versuchs- resultate iiberhaupt die Niiglichkeit einer Erklarung durch mangelhafte Probenentnahme ausschliefien, muB ich indes auf die ausfuhrliche Er- orterung dieser Punkte verweisen, die ich anderswo gegeben habe.a
Indes vermutet Zuntz , wie gesagt, dab die Proben in unseren Versuchen dem wirklichen Durchschnitt nicht entsprachen, und meint auf diese Vermutuiig hin, von den Ergebnissen, zu denen wir ge- langten, vollig absehen zu .konnen und imstande zu sein, die Striimungsgeschwindigkeit des Blutes in einer Weise zu berechnen, der die Berechtigung abgeht, wenn mit Recht auch nur bezweifelt werden kann, da6 nicht der gesamte respiratorische Stoffwechsel in den Gemeben geschieht, sondern daB auch in der Lunge selbst eine Umsetzung intermediarer Stoff mechselprodukte vorgeht. Auch Loewy wvendet noch nach dem Erscheinen unserer Arbeit die Unter- suchung der Blutproben und der Atemluft zur Bestimmung der Blut- striimung durch die Lunge an, ohne die Existenz der iifters genannten Vorgange in der Lunge zu beriicksichtigen. I n einer folgenden Ver- Gffentlichung von Loewy und Zuntz4 wird angefuhrt, die von uns gefundenen Verschiedenheiten der respiratorischen Quotienten des Blutes von denen der Atemluft seien gar zu gering, um ihnen irgend welche Bedeutung beizumessen. Wie ich oben gezeigt babe, verhalt sich dies,
* Zuntz und Hagemann, a. a. 0. S. 403. * Bohr, Nagels Eanizbuch. I. I. S. 191 u. 193. a Loewy und v. Schriitter, Blzctxirhlution beim Hensehen. Berlin 1905.
' Loewy und Zuntz in Michselis: Sauerslofftherapk. Berlin1906. 5.26. S. 2-3 und S. 87ff.

OBEX DIE SPEZIFISCHE TATIGKEIT DEB LUNGE usw. 227
bei der von uns angemandten Uethode, nicht richtig, jm Gegenteil ist auch den unter normalen VerhLltnissen nachgewiesenen Verschieden- heiten der Quotienten reelle Bedeutung beizulegen , geschweige denn den wihrend einer Anamie gefundenen (siehe oben), und letztere be- sitzen fur die prinzipielle Seite der Sache denselben Wert mie die normalen. In der jungsten Zeit hat Loewyl diese Frage mieder in seiner Respirationslehre behandelt; auch hier behauptet er die Ansicht, die Verschiedenheiten der respiratorischen Quotienten seien gar zu ge- ring, um ihnen Bedeutung beimessen zu konnen; mit Bezug auf eine andere unserer Versuchsreihen kommt er aber zu dem Schlusse, daB ,,eine rein kr i t ische Widerlegung dieser Bohr-Henriques'schen Ver- suche" zurzeit nicht miiglich sei. Da das Wort ,,kritisch" besonders hervorgehoben ist, muB wohl hiermit gemeint sein, daB eine andere Art der Widerlegung nicht ausgeschlossen sei; als solche kann aber neben der kritischen nur eine exper imente l le Widerlegung in Be- tracht kommen, die jedoch gar nicht vorliegt. Eine von anderen Forschern unternommene Kontrolle unserer Versuche wiirde naturlich auch uns sehr willkommen sein; ich bin indessen der tfberzeugung, daB sie die prinzipielle Seite unserer Anschauungen nur bestiitigen miirde. HBchstens kann Loewy am angefuhrfen Orte indes zugeben, da8 gewisse unserer Versuche eiuen Zweifel an der Richtigkeit der friiheren Ansichten erregen konnten; mir scheint es allerdings, als ob eine Sohatzung des gesamten experimeutellen Naterials, das wir in dieser Frage beigebracht haben, weiter als bis zu einem Zweifel fiihren miiSte, ich stelle aber nur fest, da8, wenn die Richtigkeit der iilteren Ansichten uberhaupt angezweifelt wird, dies unvermeidlich bewirkt, daB auch Zweifel an der Richtigkeit der hierauf basierenden Messungen der Stromungsgesohwindigkeit des Blutes entstehen muB, die nnr unter der Voraussetzung eine sichere Grundlage besitzen konnen, da6 der ge- samte Sauerstoffverbrauch beweislich in den Geweben, nioht aber auch zugleioh in den Lungen selbst erfolge.
Die obengenannte Bereohnung der Geschwindigkeit der Blut. strbmung mit-tels der Bestimmung teils der gesamten Sauerstoffaufnahme, teils des prozentigen Sauerstoffgehalts des Arterien- und des Venenblutes ist deshalb eine unsichere Xethode, deren Fehlerquelle in den einzelnen Fiillen sehr versohiedene Bedentung haben kann ; denn die intrapulmo- nare Oxydation ist, wie experimentelle Untersuohungen erwiesen haben, eine in Ausdehnung sehr variable Funktion. In Fillen, wo diese Oxy- dation nur gering ist, wird die Bestimmung der Blutxtriimung dem
1 Loewy-Oppenheimer, Bwchmzie. 1908. ,,Die Gase des K6rpera und der Gasweehsel". S. 94ff.
15 *

228 CHRISTIAN BOHR:
wirklichen Werte nahekommen , ist der intrapulmonare Sauerstoffrer- brauch dagegen betrachtlich, so wird man die Geschwindigkeit der Blutstromung gar zu groe finden, ja vielleicht sogar griiBer, als sich mit irgend welcher Wahrscheinlichkeit oder auch nur hloglichkeit an- nehmen liiBt. Wie wir sogleich sehen werden, ist dies eben der Pall, wenn man die auf dem soeben erwlhnten indirekten Wege gefundenen Werte fur die Geschwindigkeit der Blutstromung wahrend einer for- cierten Muskelarbeit naher untersucht, und dies ist wieder damit gleich- bedeutend, daB der intrapulmonare Sauerstoffverbrauch wlhrend der Muskelarbeit ein Faktor von eingreifender Bedeutung ist.
Betrachten wir in dieser Beziehung vorerst das Verhalten bei einem energisch arbeitenden Manne, wo der Sauerstoffverbrauch bis auf das Zehnfache des Wertes, den er wahrend der Ruhe hat, steigen kann; so findet L. Zuntzl durch Messungen an sich sclbst wiihrend der Ruhe einen Sauerstoffverbrauch von 263ccm pro Minute, wirhrend anstrengenden Gehens auf der Tretbahn aber von 2396-2695ccm pro Minute. Es erhebt sich nun die Frage, wie das Blut, vorausgesetzt, daf3 der gesamte Sauerstoffverbrauch in dcn Geweben des Korpers stattfindet, imstande ist, die wHhrend der Arbeit zehnfach vermehrte Sauerstoffmenge nach diesen zu befirdern. Nach Loewy und v. Schro t t e ra ist der Ver- brauch an Sauerstoff beim Obergange aus Arterienblut in Venenblut wlhrend der Ruhe auf durchschnittlich 6.5c"m pro 1 0 0 W m Blut anzu- setzen. Wiihrend der Arbeit ist die Ausnutzung des Sauerstoffs erheb- licher, llI3t sich jedoch nicht auf mehr als ca. 85 Proz. des Sauerstoff- gehalts des Arterienblutes schltzen; rechnet man letztereu als 19 Volum- proz., so werden in diesem Falle 16.2Wm Sauerstoff' fiir je lCOWm Blut verbraucht, mithin 2l/, ma1 mehr als wiihrend der Ruhe. Hieriiber hinaus kann die Sauerstoffzufuhr nur durch Vermehrung der pro Minute das Herz durchstrbmenden Blutmenge steigen ; da die Sauerstoffaufnahme bis auf das Zehnfache gestiegen ist und d g wie wir gesehen haben, die griiI3ere Ausnutzung des Sauerstoffs des Blutes nur eine Zunahme um d8s 2l/,fache bewirken kann, so muI3 unter der Voraussetzung, daB die game Oxydation in den Geweben geschieht, die Stromung des Blutes wiihrend der Arbeit 4 ma1 so groS geworden sein. Bei normalen Menschen nimmt nun die Pulsfrequenz selbst wlhrend sehr forcierter Arbeit durch- schnittlich nicht mehr als bis auf das Zweifache ihres Wertes wiihrend
L. Zuntz, Cfaswechsel und Energieumsatx des Radfahrers. Berlin 1899.
L o e w J und v. S c h r 6 t t e r, Blutzirkulation beim Memehen. Berlin S. 26-27.
1905. s. 77.

~ B E R DIE SPEZIFISCHE TATIGKEIT DER LUNGE usw. 229
der Ruhe zu (von 70 bis 140 pro Minute), folglich sollte wahrend der Arbeit das Schlagvolumen des Herzens ebenso wie die Pulsfreqenz bis auf das Doppelte steigen; eine solche gleichzeitige Verdoppelung der Puls- frequenz und des Schlagvolumens kommt mir aber allem Vorliegenden zufolge als sehr wenig wahrscheinlich vor. Die verschiedenen Forscher, die mittels d i rek ter MeBmethoden die Mengen des die Aorta pas- sierenden Blutes untersuchten, haben allerdings oft bei Zusammen- stellung der einzelnen Bestimmungen einer Versuchsreihe gefunden, da6 die maximalen Werte des Schlagvolumens um mehrere Nale groSer waren als die minimalen Werte; dies kann indes leicht davon her- riihren, daB einzelne Bestimmungen ein abnorm geringes Schlagrolumen ergeben haben (Herzschwlchung), was ja unter Bezugnahme darauf, daB die direkte Messung der Blutstriimung gewohnlich mit nicht un- bedeutenden Eingriffen verbunden ist, hiichst wahrscheinlich ist. Fiir un8 ist die Frage jedoch eine andere, namlich die, inwiefern miin sich aus den vorliegenden Versuchen eine Ansicht iiber den EinfluB bilden kann, den eine VergriiBerung der Pulsfrequenz durchschni t t l ich und normal auf das Schlagrolumen und mithin auf das Sekundvolumen des Blutes iiben wird. Hier scheinen die verschiedenen sowohl iilteren als jiingeren Untersiichungen l, sofern direkte MeSmethoden zur An- wendung kamen, darin iibereinzustimmen, da6 das Schlagvolumen bei geringerer VergrGBerung der Pulzfrequenz zwar zunehmen kann, daB es bei bedeutenderer VergroBerung derselben jedoch abnimmt, so da6 in diesem Falle das Sekundvolumen nicht einmal in demselben Verhiilt- nisse anwachst wie die Pulsfrequenz. Hierfiir sprechen auch Heringsa Versuche an Pferden ; bier fand er mittels seiner bekannten Injektions- methode, da6 eine durch Arbeit verursachte VergrOBerung der Puls- frequenz ron 36 bis auf 100 pro Minute die Striimungsgeschwindig- keit freilich steigerte, doch nicht einmal anniiherungsweise in demselben Grade, wie die Pulsfrequenz anwuchs. 1st dies das durchengige nor- male Verhiltnis, so wird man in unserem Beispiele mit dem arbeitenden Mann, bei dem die Pulsfrequenz bis auf das Doppelte steigt, die Blut- menge, die wiihrend des Arbeit pro Minute die Lunge passiert, nicht hiiher ansetzen kiinnen als auf das Doppelte der wiihrend der Ruhe passierenden Blutmenge; da nun die Ansnutzung des Sauerstoffes des Blutes, wie oben gefunden, hiichstens auf das 2lI2 fache geschiitzt werden darf, wird die gesamte Zufnhr von Sauerstoff zu den Geweben wiihrend der Arbeit hbchst 5mal giiBer werden als wiihrend der Ruhe. Da nun aber der
Vgl. Tigerstedt, Erghi s seder Physiologie. 1905. Bd. IV. 5.494 u. 505. * Hering, Archiv fi ph,ysiol. Heilkurrde. 1853. Bd. XII. S. 133.

2 30 CHRISTIAN BOHR:
gesamte Verbrauch Ton Sauerstoff auf das Zehnfache gestiegen ist, so mu8 wenigstens die Halfte des Sauerstoffoerbrauches intrapulmonar, durch in der Lunge selbst geschehene Umsetzungen stattgefunden haben.
Alles scheint mir, dem oben Entmickelten zufolge, dafur z u sprechen, daB die Sache sich wirklich SO rerhalt; ich hebe aber ausdriicklich hervor, daB die d i r e k t e n htessungen der Blutstriimung nicht in solcher Variation und in solchem Umfange oorliegen, daB sich durch unscr Beispiel mehr als ein hoher Grad der Wahrscheinlichlieit erzielen lieBe. Positive GemiBheit uber den groBen Anteil der intrapulmonaren Um- setzung an der wahrend der Arbeit eintretenden T'ermehrung des Stoff- wechsels lionnen wir dagegen durch Hetrachtung der Versuche geminnen, die Zuntz und H a g e m a n n l in ihrem gehaltreichen Werke uber den Stoffmechsel des Pferdes mitgeteilt haben. Diese Versuche eignen sich wegen ihrer Feststellung eines bedeutenden Steigcns des Stoffwechsels wihrend der Arbeit, in besonderem MaBe zu unserem Zwecke.
Die ron Z u n t z angefuhrten MaBe der Blutmenge, die beim Pferde unter rerschiedenen Verhaltnissen pro Minute den Herzrentrikel passiert, wurden in der ofters erwihnten Weise aus der totalen Sauerstoffauf- nahme des Tieres, mit dem prozentigen Sauerstoffgehalt des Venen- und des Arterienblutes zusammengehalten, berechnet. In untenstehender Tabelle sind die Resultate fur Ruhe, mlf3ige Arbeit und exzessire Arbeit angefiihrt; die Werte fiir die Blutmenge pro Minute und fur die Pulsfrequenz sind die ron Zuntz selbst (a. a. 0. S. 404) an- gegebenen Zahlen , die er zur Berechnung der Herzarbeit benutzt hat. Fur Ruhe und mBSige Arbeit sind die Zahlen die durchschnitt- lichen' Werte mehrerer Versuche (mittleres Gewicht des Tieres 347.6 kg);
fiir die exzessive Arbeit ruhren die Werte aus einem Versuche mit maximaler Arbeit her (Gemicht des Tieres 34'Ikg). AuBer diesen ron Z u n t z angegebenen Zahlen habe ich in einer dritten Kolonne das Schlagvolumen eines Herzventrikels hinzugefiigt, welches ich ganz einfach dadurch berechnete, dafi ich die Blutmenge pro Minute mit der Anzahl der Pulsschliige pro Minute dividierte. In einer vierten Kolonne ist die Sauerstoffaufnahme durch die Lunge pro Minute angefuhrt.
Minut- B1utmenge
1
Ruhe . . . . , . . . . 29.2 MaSige Arbeit . . . . ' 1 53.0 Exzessive Arbeit . . 1 245.5
Puls- Schlag- Sauerstoff- volumen aufnahme
fiequenz eines Herz- pro Minute Pro Minute ventrikels i l
2.08 5-01
22.94 70 3.51

OREB DIE SPEZIPISCHE TATIGKEIT DER LUNGE usw. 231
Wie bereits iifters erwahnt, werden die fur die Minut-Blutmenge berechneten Zahlen nur dann richtig sein konnen, wenn die gesamte in der Lunge aufgenommene Sauerstoffmenge mit dem Arterienblute in die Gewebe hinaus gefuhrt wird; erfolgt in der Lunge selbst ein Sauerstoffverhrauch, so werden die Zahlen zu groB. Umgekehrt : wenn die fiir die Minut-Blutmenge in der hier besprochenen Weise berechneten Zahlen zmeifelsohne zu groB sind, so muB ein Teil des Sauerstoff- verbrauches intrapulmonar geschehen sein. Dies ist nun ganz unzweifel- haft der Fall mit Bezug anf die exzessive Arbeit, wo die Blubmenge pro Minute 8.4mal so groB ist als miihrend der Ruhe.
Schon die Kurze der Zeit, die demnach im genannten Falle zu einem vollstiindigen Kreislauf verbraucht sein sollte, muB groBe Be- denklichkeiten erregen. Wird die Blutmenge auf 1/13 des Korper- gewichtes angesetzt, so betriigt sie bei einem 347 kg wiegenden Pferde 26.7 1; da das Minutvolumen 245.51 ist, sollte eih Kreislauf mit Passage des Herzens ond der GefiiSe nnd mit dem Aufenthalt in zwei Kapillarsystemen nur 6.5 Sek. beanspruoht haben. Dies ist um so weniger wahrscheinlich, als Her in g mittels seiner Injektionsmethode findet, daB bei einem Pferde, dessen Pulsfrequenz infolge anstrengenden Trabens von 36 biu 100 gestiegen ist, die Gmlaufszeit der am ge- schwindesten umlaufenden Partikelchen 17.5 Sek. betriigt. DaB die berechnete Minut-Blutmenge gar zu groB ist, wird jedoch auBer allen Zmeifel gestellt, menn man das Schlagvolumen betrachtet. Indem die Pulsfrequenz von 40 bis auf 70 anwichst, ist das Schlagyolumen wHhrend der exzessiven Arbeit um ca. 5mal groBer geworden als wiihrend der Ruhe und hat die Hohe von 3.5 1 erreicht. Dies ist aber, wie die anatomische Untersuchung erweist, unm6glich, da der Herz- ventrikel des Pferdes iiberhaupt nicht so vie1 zu fassen vermag, nicht einmal anniihernd. So gibt Colin2 an, dab der Herzventrikel bei Pferden der hier besprochenen Gewichtsklasse zirka 1 1 fabt; er unter- nahm seine Messungen an absolut frischen Herzen unmittelbar nach dem Tode des Tieres, also bevor Totenstarre eingetreten watS Dies ist selbstverstiindlich notwendig, denn wie derselbe Forscher durch systematischs Messungen nachgewiesen hat, erhiilt das einmal erstarrte Herz seine urspriingliche Schlaff heit nicht glnzlich wieder zuriick, mithin auch nicht sein volles diastoliscbes Volumen. Far die Richtig-
A. a. 0. Colin, Physwlogie compar6e des animaum. Paris 1888. T. 11. p. 461. Milne Edwards, Lqons sur la physiologie. Paris 1859. T. IV. S. 92.
Anmerkung.

232 CHRISTIAN BOHR:
keit der ron Colin angestellten Messungen an den ganz frischen Herzen spricht ubrigens auch der Umstand, daS er trotz der sehr rerschiedenen Dicke der Muskelwandungen in den meisten Fallen fast dieselben Zahlen fur das Volumen des rechten, mie fur das des linken Ventrikels findet. Das Schlagrolumen des Herzeus beim Pferde ist deshalb zireifrlsohne geringer als 11.
Um diese Frage noch ferner zu beleuchten, habe ich an Pferde- herzen einige Messungen nach einer Methode unternommen, die, indem sie es gestattet, von dem mehr oder weniger erstarrten Zustande des Herzens ahzusehen, das absolute anatomische Maximum derjenigen Menge Blnto angibt, melche das Herz iiberhaupt zu fassen rermag. Die Methode besteht in dem Xessen all der Fliissigkeit, die der Perikardial- sack fassen kann, menn man das Volumen des Herzfleisches abzieht; hierdurch erhilt man offenbar dasjenige Volumen, welches das ganze Herz zu fassen imstande ist, wenn es nach vorhergehender volliger Entleerung so stark erweitert wird, daB es die Perikardialhaut moglichst auftreibt. Diese Messung wird so ausgefuhrt, daS in das iintere Ende des Perikardiums eine R6hre eingebunden mird, durch die man so vie1 Wasser hineingiebt, mie das Perikardium iiberhaupt fassen kann; dies sei P Liter. Darauf nimmt man das Herz aus dem Perikardium heraus und mifit dessen Volumen (H) bis zur Perikardialgrenze durch Ver- senkung in Wasser; zuletzt zerschneidet man das Herz so, daS alle HBhlungen geoffnet werden, und durch Versenkung in Wasser findet man das Volumen (P) des eigentlichen Herzmuskels. Das Volumen der Hbhlungen des Herzens ist also H + P; addiert man diese GrciSe zu P, so erhalt man (P + H + P) als das groSte Volumen des Herzens, wenn dieses nach Zusammenpressung bis zur Verwischung jeder Hohlung gefiillt wird , bis die Perikardialhaut fernere Erweiterung verwehrt. E k e in dieser Weise an einem Pferdeherzen (Gewicht des Tieres ca. 500 kg) ausgefiihrte Bestimmung ergab P = 4-4, H = 3.6 und P = 3.3, also P + H + 3’ = 4.7 1. An einem anderen Pferde der- selben GrBSe fand ich etwas kleinere Zahlen, namlich P = 3.3, B = 4.0, P= 3-5, mithin P+ Hs P = 3.8 1. Das Mittel der beiden Be- stimmungen gibt 4.21. Fiir jede Half te des Herzens erhiilt man somit 2.1 1, was also das iiuI3erste anatomisch mijgliche maximale Volumen eines Vorhofes und einer Herzkmmer im Verein ist. Um die maximale Erweiterung eines Herzventrikels zu erfahren, miiSte man hiervon daher das Volumen des maximal gefiillten Vorhofes ab- ziehen, und da das Pferd im obenstehenden Arbeitsversuche 347 kg wog, also bedeutend weniger als diejenigen Pferde (500 kg), deren Herzen gewogen wurden, mu6te zu unserem Zwecke eine fernere Reduktion der

~ B E R DIE SPEZIFISCHE TATIGKEIT DER LUNGE usw. 233
Naximalzahl unternommen werden. Diese beiden Reduktionen wurden die Zahl sicherlich bedeutend herabsetzen; da es aber schwierig ist, fiir dieselben einen genauen numerischen Wert anzugeben, werden wir sie unterlassen und uns damit begniigen, die 2- 1 1 auf 2 1 abzurunden und diese als das anatomische MaximcllmaB des Herzrentrikels rechnen.
Es geht hieraus hervor, dab bei einem 3 4 7 k g wiegenden Pferde das Schlagvolumen eines einzigen Herzrentrikels aus anatomischen Griinden unmoglich 3.5 1 betragen kann; diejenige Menge Blut, die . der Ventrikel entleeren wiirde, wenn er, nachdem er erst dermafien angefiillt worden ware, daB er das Perikardium miiglichst weit auf- getrieben hatte, darauf bis auf den letzten Tropfen geleert wiirde, wiirde namlich weniger als 2 1 betragen, und miihrend dem Leben ist eine solche Anfullung und Entleerung ja durchaus nicht moglich.
Die von Zuntz nach der Fickschen Methode berechnete Minut- Blutmenge ist daher viel grofier, als sie in Wirklichkeit sein kann, und hieraus folgt notgedrungen, daB nicht die gesamte wahrend des hier besprochenen Arbeitsversuches aufgenommene Sauerstoffmenge die Lunge im Verein mit dem Arterienblute verlassen haben kann, und daB im Gegenteil ein bedeutender Teil derselben i n der Lunge selbst verbraucht worden ist.
Fiir den numerischen Wert dieses intrapulmonaren Umsatzes bei exzessiver Arbeit (siehe die Tab. S. 230) werden wir erst ein HuSerstes Unimum berechnen , indem wir den anatomischen Maximalwert des Volums des Herzventrikels (2 1) gleich dem Schlagvolumen setzen, ob- schon dieses selbstverstandlich viel kleiner sein muB. Wir erhalten dann fiir eine Pulsfrequenz von 70 eine Minut-Blutmenge von 140 1. Die groBte Ausnutzung des Sauerstoffes des Blutes beim Obergange aus Arterien- in Venenblut, die Zuntz bei arbeitenden Pferden beob- achtet hat, betrigt 87 Proz. des Sauerstoffgehaltes des Arterienblutes (14 Volumproz.) oder 12.2 Volumproz. Sauerstoff. J e 100 ccm Blut, die die Lunge passieren, konnen also in msximo 12.2- Sauerstoff auf- nehmen, oder, da die ubrigen Zahlen der Tabelle in Litern angegeben sind, jedes Liter Blut kann 0.122 1 Sauerstoff anfnehmen. Die 140 1 Blut, die pro Minute die Lunge passieren, verm8gen mithin 140 x 0.122 = 17.08 1 Sauerstoff aufzunehmen, und mehr Sauerstoff kann das Blnt wiihrend einer Minute den Geweben natiirlich nicht zufiihren. Nun betrlgt die Sauerstoffaufnahme des Tieres indes in toto 22.94 1 pro Minute, und hiervon miissen 22.94 f 17-08 = 5.86 1 Sauerstoff oder 25 Proz. der insgesamt anfgenommenen Sauerstoffmenge zu Umsltzen verbraucht worden sein, die in der Lunge selbst statt- gefunden haben. Da dies, wie gesagt, wegen der gar zu hohen An-

234 CHRISTIAN BOHR :
setzung des Wertes des Schlagrolumens ein entschieden minimaler Wert ist, geht aus den Zuntzschen Versuchen somit herror, daB bei exzessiver Nuskelarbeit ein bedeutender oxydatirer UmsatzprozeB in der Lunge selbst vorgeht.
Der auf diese Weise berechnete minimale Wert liegt selbstrer- standlich erheblich unterhalb des wirklichen Anteils des intrapulmo- naren Umsatzes an dem Sauerstoffrerbrauch. Dieser Anteil 1aBt sich annahernd dadurch berechnen, daB wir das Schlagrolumen gleich dem von Colin an ganz frischen Herzen gefundenen Volumen eines Herz-. ventriliels oder = 1 1 setzen, was iibrigens aller Wahrscheinlichlieit nach noch als Maximum zu betrachten ist. Dann erhalt nian fur 70 Pulsschlage pro Minute eine Ninut-Blutmenge von 70 1; dies wird (bei demselben Grade der Ausnutzung [S7 Proz.] des Sauerstoffs des Blutes, der oben angewandt wurde) fiir die pro Minute mit dem Ar- terienblut den Geweben zugefiihrte Saueratoffmenge 8.54 1 ergeben. Da die gesamte Sauerstoffaufnahme pro Minute 22.94 1 betriigt, wird der Anteil der Lunge am Sauerstoffverbrauche demnach 14.40 1 oder 63 Proz. des ganzen Verbrauches.
Rahrend der Ruhe und der ml6igen Arbeit lassen Berecli- nungen mie die obenstehenden, mo man m a x i m a l e Zalilen zugrunde zu legen sucht, sich selbstrerstandlich nicht durchfiihren; dies darf je- doch keineswegs als damit gleichbedeutend aufgefa6t werden, da6 an- zunehmen sei, es finde in diesen Fallen kein intrapulmonarer Sauer- stoffverbrauch statt. I m Gegenteil: die friiher in diesem Paragraphen hesprochenen Verschiedenheiten der aus der Atemluft von den aus den Blutproben berechneten respiratorischen Quotienten erweisen, da6 sogsr wlhrend der Ruhe gewohnlich ein iibrigens sowohl der Menge als der Art nach wechselnder intrapulmonarer Umsatz vorgeht.
D e r i n t r a p u l m o n a r e Sauers tof fverbrauch , dessen Exi - s t e n z d u r c h d i r e k t e Versuche dnrge leg t worden ist,' e r w e i s t s i c h a l so a l s e i n f u n d a m e n t a l e r F a k t o r des gesamten Respi- ra t ionsprozesses . Er e r m o g l i c h t es, wie d ie Versuche i iber d i e A o r t a a b s p e r r u n g zeigen, daB d e r Stoffwechsel d e r Ge- mebe s e l b s t be i b e d e u t e n d b e r a b g e s e t z t e r S a u e r s t o f f z u f u h r f o r t g e h e n k a n n ; wie s ich a u s d e n von Zuntz u n t e r n o m - m e n e n Arbei t sversuchen nachweisen 11Bt, s p i e l t d e r intr t t - p u l m o n a r e ProzeB i n d e s a u o h e ine wicht ige Rol le i m Stoff- wechse l w a h r e n d exzessiver A r b e i t u n d e r m o g l i c h t h i e r d i e s t a r k e Z u n ah m e d e s S a u e r Y t o ff v e r b r a u c h s.
Bohr und Henriques, Arch. de physiol. 1897. a. a. 0.

UBER DIE SPEZIFISCEE TATIGKEIT DER LUNGE usw. 235
Von der Beschaffenheit der infermediaren Stoffmechselprodukte, die ails den Gewebea nach der Lunge gefuhrt werden, urn hier eine Umsetzung zu erleiden, gebricht es uns zurzeit an Kenntnis. tfber die Art des Unisatzvorganges missen mir, daB, mas das Erstickungsblut betrifft, eine einfache Sauerstoffzufuhr nicht genugt, um die Oxydation hervorzurufen , daB dagegen die Zellen der Lunge eingreifen mussen,l dies geschehe nun durch Aufnahme und Verarbeitung der inter- mediiren Produkte oder, was vielleicht wahrscheinlicher ist, dadurch, da6 die Zellen mittels einer inneren Sekretion die Oxydation im Blute beeinflussen. DaI3 die Arbeit der Lungenzellen auch fur den unter anderen Verhiltnissen stattfindenden in trapulmonaren Umsatz eine notwendige Bedingung ist, und daB das Vorhandensein von Sauerstoff an und fiir sich nicht geniigt, scheint daraus hervorzugehen, daB das Venenblut wiihrend der Kuhe und gewv6hnlich auch wahrend der Arbeit einen betrachtlichen OberschuB an Sauerstoff enthalt.
Fur diese mie auch fiir eine ganze Reihe anderer Fragen, die sich an diese wichtige Funktion kniipfen, werden indes neue und er- weiterte Un tersuchungen erforderlich sein.
6 2. Die spezifiache Tatigkeit der Lnnge beim GCaswechsel dnrch
DaB die Lunge bei der Wanderung des Sauerstoffes aus der Alreolenluft in das Blut und der Wanderung der Kohlensiiure in der nmgekehrten Hichtung eine aktive Rolle spielt, geht aus den Werten hervor, die man durch Bestimmungen der Spannung dieser Gase an beiden Seiten der Alveolarwand findet. Wirkte die Lunge nur als eine inerte dunne Membran, die, alien Gasen durchdringlich, die Lungenluft Tom Blute trennte, so miiBten die niedrigsten Spannungs- werte stets in der Richtung der Wanderung der Gase zn finden sein, in betreff des Sauerstoffes also im Blute, in betreff der Kohlenslure in der Alveolenluft. Dies ist jedoch nicht der Fall; fuhrt man behufs der Anf klbung dieser VerhLltnisse eine Reihe Spannungsmessungen aus, so wird man unter den Bestimmungen fast stets einige finden, wo die Werte sich nicht anders als durch Intervention einer spezifi- schen Zellentgtigkeit erkliiren lassen.
Um dies nachzuweisen, geniigt es, die unmittelbaren Resultate von Versuchen ins Auge zu fassen, wo die Spannungen mittels d i r e k t e r tonometrischer Methoden gemessen wnrden; diem sind, weil die durch-
die Alveolarwand.
Bohr , Nagels Handbuch. I. I. S. 1S6.

236 CERISTIAN BOHR:
schaulichsten, als die ~icherst~eii zu betrachten, und in diesem Zusammen- hang sehe ich deshalb von den Methoden ab, wo man die Spannung durch Analyse des Luftgehalts eines abgesperrten Lungenteiles zu be- stimmen sucht, wie auch von den Bestimmungen, die man nach der Kohlenoxydmethode ausgefiihrt hat.
Durch tonometrische, mittels des Himataerometers angestellte, direkte Bestimmungen hat es sich nun bekanntlich erwiesen,’ daB die Werte der Sauerstoffspannung im Arterienblut der Hauptsache nach um die gleichzeitig fur die Alveolenluft gefundenen Werte herum sohwanken, bald etwas niedriger, bald auch etwas hoher als diese liegen. Zu iihnlichen Resultaten kam ich mit Bezug auf die Kohlen- saure; die Spannung dieses Gases im Blute lag in gewissen Ver- suchen freilich hoher, in anderen aber entschieden niedriger als die in der Alveolenluft gefundene. In denjenigen Fallen, wo die Kohlen- saurespannung in der Weise hBher oder die Sauerstoffspannung niedriger in der Alveolenluft als im Blute gefundeu wurde, miissen die Lungenzellen eine Tatigkeit entfaltet haben, die irgendwie das Resultat beeinfluBt hat; wlre von einer inerten Uembran die Rede, so konnten die Spannungen nimmerlnehr in der Richtung der Bewegung an- gewachsen sein. Es widerspricht selbstverstrindlich der Existenx einer derartigen Zellenthtigkeit nicht, da8 diese nicht immer die Sauerstoff- spannung des Blutes iiber die Spannung der Alveolenluft hinauf- steigert oder die Kohlensaurespannung unter dieselbe hinabsenkt. Es kann nicht angenommen werden, da6 die Aufgabe einer solchen Zellentatigkeit darin bestehen sollte, die Spannung immer bis auf eine gewisse Hohe zu bringen, sondern mu6 vielmehr darin zu suchen sein, die Spannung in jedem einzelnen Falle den komplizierten Anspriichen des Organismus anzupassen. Bei dem gro6en Anpassungsvermagen des Organismus an die auBeren Bedingungen ist es ganz natiirlich, daB die Gasspannungen im Blute hierbei nnter gewohnlichen Verhllt- nisuen, beim Einatmen atmosphiirischer Luft, um einen mittleren Wert herum schwanken , der mit dem der Alveolenluft zusammenfallt; be- merkenswert ist es aber, da6, wie Fredericq nachgewiesen hat, das- selbe der Fall ist, wenn die Zusammensetzuug der Alveolenluft sich wegen der Einatmung einer mehr oder weniger sauerstoffreichen Luft- mischung lndert ; eine Einstellung der Gasspannungen im Arterien- blute auf ungefiihr dieselbe Hohe wie in der Alveolenluft ist deshalb ah eine weseutliche Eigentiimlichkeit des Prozesses zu beachten, die
Bobr, Dies Archiv. 1890. B. 11. 8. 236. Fredericq, Zentralblatt f i Physwl. 1893. Bd. VII. S. 33.

OBER DIE SPEZIFISCHE ‘rATICfKEIT DER LUNQE USW. 237
vielleicht mit der Gefahr einer Riickdiffusion in Beziehung steht, die sonst stets, wie man sich nun auch den Vorgang der Gaswanderung denken mag, rorhanden sein wiirde.1
Spatere Versuche,2 bei denen die Kohlenslurespannungen des Blutes mittels Kr oghs Uikrotonometers bestimmt wurden, fiihrten zu ganz denselben Resultaten wie den oben angegebenen; auch hier fand man in mehreren zuverllssigen Versuchen die Kohlensiurespannung grri6er im Arterienblute als in der Lungenluft, und zwar in aus- gepragtem Ma6e; die spezifische Tatigkeit der Lunge bei der Kohlen- slureausscheidung wahrend der mit der Versuchsanordnung vereinbaren moglichst normalen Verhlltnisse la6t sich daher nicht bezweifeln. In dieser Untenuchungsreihe gelang es aber zugleich, durch Variation der Versuchsbedingungen ein Mittel zu finden, urn willkiirlich einen solchen Zustand hervorzurufen, da6 die Kohlensiurespannung im Blute - sogar im Venenblute - sich konstant niedriger erwies als in der Lungenluft. Dies erzielte man dadurch, da6 man jede der beiden Lungen fur sich atmen lie6 und zwar eine Luft, welche gleichzeitig fir jede Lunge verschiedenen Gehalt an Kohlenslure enthielt. Atmete z. B. die eine Lunge atmosphirische Luft, die andere ein Gasgemisch mit ca. 8 Proz. Kohlenslure, so fand in letzterer Lunge stet8 noch eine Kohlenslureausscheidung aus dem Blute statt, obschon das aus dem rechten Herzen zur Lunge stromende Blut eine Spannung hatte, die tief unter der der Alveolenluft lag.
DaB sich bei der Wanderung der Gase durch die Lungenwandung eine Zellentiitigkeit geltend macht, liBt sich mithin nicht bezweifeln. Hiermit steht es selbstverstlndlich nicht in Widerstreif, da6 durch die Alveolarwand eine Gasdiffussion erfolgt ; eine solche mu& wie die Auf- nahme indifferenter Gase (Stickstoff, Wasserstoff, Schwefelwasserstoff) zeigt, unzweifelhaft stattfinden, und fraglich ist nur, in welchem Um- fang die Gasdiffusion geschieht, und in welcher Beziehung sie zur spezifischen Zellentltigkeit der Lunge steht.
Fur die Auffassung der Wirkungsweise de r Lnngenzel len wiihrend der Gassekretion konnen wesentlich zwei Gesichtspunkte zur Geltung kommen, was ich bereits in meiner ersten ausfiihrlicheren Ver- Bffentlichung iiber die Lungenatmung hervorhob, wo 8s heik3 ,,Den EinfluS, den das Lungengewebe erweislich suf das Verhiiltnis der Gas- spaunungen im Blute und in der Lungenluft ausfibt, kann man sich
Bohr, Nagels Handbuch. I. I. S. 157. 2. Stiick. Bohr, Zenfralblatt f. Physiol. 1907. Bd. XXI. s. 367.
a Dies Archio. 1890. Bd. 11. S. 268.

2 38 CHRISTIAN \BOER:
entmeder als eine direkte Aufnahme ron Snuerstoff und Ahgabe ron Ihhlensiiure durch die Epithelzelleii der Alreolarwand rorstellen, oder auch als eine Einmirkung auf das Blut, durch welche die Spannungen in demselben in dem Augenblicke redndert merden, in melchem es die Lunge rerla6t; selbstrerstandlich schlieBt die eine dieser beiden Wirkungsmeisen nicht die andere aus." Fur beide Anschauungen sprechen gemichtige Momente; fur die direkte Aufnahme der Gase durch die Zellen z. B. die ron E h r l i c h nachgewiesene reduzierende Wirkung des Lungen- gewebes und mohl auch die Analogie zur Ssuerstoffausscheidung in der Schwimmblase der Fische. Anderseits ist eine Einwirkung der Lungen- zellen auf das Blut, modurch diese mittels einer inneren Sekretion Anderungen der Spannungsmerte der von ihnen aufgenommenen Gase hervorrufen, eine iiuDerst naheliegende Annahme. Wie ich in einer Reihe ron Arbeiten a nachgewiesen habe, sind die gasbindenden Stoffe des Blutes, besonders das Hamochrom, leicht veranderlich ; so ist sowohl die Menge des Sauerstoffs und der Kohlensiure, welche das Hamochrom bei maximalem Drucke zu binden rermag (spezifische Gasbindung), wechselnd, als auch die Form der Spannungskurve fur ein gegebenes Himochrom bei verschiedenen Einwirkungen variabel ist. Ich brauche hier nur an die Einwirkung zu erinnern, welche die Kohlenslure auf die Sauerstoffspannungskurve ubt, wie auch an die Anderung dieser Kurve schon beim 'Obergange des in den Blutkorperchen enthaltenen Hgmochroms in gelostes Hirnoglobin, die wahrscheinlich dnrch eine Anderung der Bindung zmischen dem Hamatin und dem Globin, eventuell dem Lecithin verursacht wird. DaB diese Labilitat des HImo- chroms einen innerhal b weiter Grenzen wirksamen Regulator der Gas- spannungen im Blute bilden kann, habe ich anderswo gezeigt.
Um die beiden genannten Anschauungen von der Art der Zellen- tltigkeit, die sich ubrigens sehr wohl als unter beiden Formen neben- einander wirksam denken la&, nlher zn prlzisieren, wollen wir sie durch ein Beispiel illustrieren. Hierzu benutzen wir einen Versuch (Versuch V, Abt. 2) aus der oben zitierten Untersu~hungsreihe,~ wo die getrennten Lungen je ihre Luftmischung atmeten, wihrend das ihnen zustromende Venenblut betreffs der Zusammensetzong identisch war. In diesem Versuche hat das Venenblut eine Kohlensaurespannung von 31mm; die rechte Lunge atmet eine Luft mit einem Gehalt ron
E h r 1 i c h , Sawrstoff bedurfnis des Organismus. Berlin 1886. 2 Dies Archio. 1892. Bd. 111. S. 47 u. 101.
Bohr, a. a. 0. und Nagels ffandbuch. I. I. S . 196ff. ' Zentralblatt f. Physiol. 1907. Bd. XXI. S. 372.

UBElt DIE SPEZIFISCHE TATIGKEIT DER LUNGE USW. 239
8.80 Yroz. CO, und ihre Alceolarspannung ist infolgedessen bis auf 69.5 mln CO, gestiegen. Nichtsdestoweniger wird Kohlensaure aus dem Blute in die Alveolenluft ausgeschieden, und zwar speziell 28 ccm pro Kilogramm uud Stunde. Hier mu8 offenhar eine Zellentatigkeit irgend einer Art eingegriffen haben. Man kann sich den Verlauf derselben nun so denken, daB die Kohlenslure ron den Lungenzellen gebunden, durch diese hindurchgefuhrt und an der Alveolarseite der Nembran gegen die hier herrschende hbhere Spannung ausgeschieden mjrd ; auf diese Weise mii6te nun simtliche Kohlensaure, die mittels der unzweifel- haft gleichzeitig stattfindenden Diffusion wegen der Druckdifferenz aus der Alveolenluft in das Blut eindringt, wieder zuriiclisezerniert und auBerdem noch ferner ein ffberschu6 von 28- pro Kilogramm und Stunde durch die Alveolarwand geftihrt werden. Man kann sich aber auch die Tltigkeit dcr Zellen als darin bestehend denken, daB sie die Kohlenslurespannung im Blute andert, wlhrend die Wanderung der Kohlensaure durch die Alveolarwand mittels Diffusion gepchieht; dann mii6ten in unserem Versuche die Zellen wlhrend der Passage des Blutes durch die Lunge dessen Kohlensiiurespannung von 31 mm bis auf mehr als 69.5 mm gesteigert und diese Erhohung der Spannung so reguliert haben, daB eine gewisse kleinere Menge Kohlensiure aus dem Blute fortwlhrend in die Alreolenluft diffundierte. In dem hier erbrterten Versuche wurde die Erscheinung ihre hinliingliche Erkllrung dadurch finden, daB die Kohlensiiurespannung wahrend der Passage durch die Lunge hijher wurde. In anderen Fallen mu8 man indes, um die Kohlensaureausscheidung mittels einer Regulierung der Spannung zii erkliiren, eine mehr komplizierte Wirkung der Zellenarbeit annehmen, welche die Kohlensiiurespannung im Blute erst wiihrend der Passage durch die Lunge steigerte und sie darauf, beror das Blut die Lunge verlieBe, wieder bedeutend herabsetzte. So im Versuch I1 der oben zitierten Untersuchungsreihe; hier fand die Atmnng in normaler Weise mit der ganzen Lunge statt und war die Einatmungsluft atmosphiirische Luft. Die Kohlenslurespannung in der Alreolenluft betrug 25.9 mm, im Blute des rechten Herzens ebenfalls 25.9 mm und im Arterienblute 16.8 mm; es wurden 363 CO, pro Kilogramm und Stunde aus- geschieden. Unter der Voraussetzung, da6 die Ausscheidung nicht da- durch erfolgt, da6 die Zellen direkt die Kohlensiiure duroh die Wand befijrdern, sondern dadnroh, da6 sie die Spannung regulieren, hat man hier anzunehmen, dab die Kohlensiinrespannung des veniisen Blntes erst von 25.9 mm (der Spannung dea Blutes im rechten Herzen) nm SO vie1
Zentralblatt f i Physiol. 1907. Bd. XXI. S. 369.

240 CHRISTIAN BOHR:
fiber die Spannung der Alveolenluft (ebenfalls 25.9 mm) gebracht wid, daB die ausgeschiedene Menge Kohlensawe megen der Druckdifferenz diffundieren kann , worauf diese Kohlensaurespannung mieder auf die Spannung des Blutes, als es die Lunge ~erlie6, also auf 16.8 mm, herabgesetzt wird.
Was den Sauerstoff betrifft, wird die Zellentatigkeit, wie wir oben sahen, dadurch dargelegt, daB die Sauerstoffspannung des Arterienblutes die Spannung dieses Gases in der Alveolenluft ubersteigen kann. Auch hier gibt es natiirlich die Moglichkeit, beide soeben genannte Arten spezifischer Zellentatigkeit anzunehmen. So ist es mbglich, daB die Lungenzellen den Sauerstoff binden und ins Blut befordern, wodurch die Spannung im letzteren zuweilen bis iiber die Alveolarspannung gesteigert werden konnte. Es ist aber auch denkbar, daB die Tatigkeit der Zellen in einer h d e r u n g der Sauerstoffspannung des Blutes he- etiinde, wodurch diese so reguliert wurde, daB die fiir eine hinlingliche Diffusion durch die Alveolarwand erforderlichen Druckdifferenzen ent- stiinden; dann hitte man anzunehmen, die Sauerstoffspannung des die Lunge passierenden Blutes lasse sich dergestalt regulieren, daB sie erst herabgesetzt werde, um darauf, bevor das Blut die Lunge verlasse, bis auf eine angemessene Hohe, bisweilen bis hoher als die Sauerstoff- spannung in der Alveolenluft gesteigert zu werden.
Wir mogen nun aber annehmen, daf3 die Zellentitigkeit in einer direkten Beforderung von Gasen durch die Alveolarwand oder aber in einer solchen h d e r u n g der Gasspannungen im Blute bestehe, daB durch sie eine fur den Diffusionsdurchgang geniigende Druckdifferenz erzeugt werde, so haben wir in beiden Fa l len mit einer spezi- fischen Lungenfunkt ion zu schaffen, die eine Regulierung der Gas- manderung ermoglicht, so daf3 diese nicht niitig hat, die Bedingungen zu befolgen, die durch den Untersohied zwisohen der Spannung der Alveolenluft und der Spannung in dem die Gewebe des Korpers ver- lassenden, der Lunge zustriimenden Venenbluto gegeben sind. In beiden Fallen ist der ProzeS deshalb den eigentlichen Driisensekretionen an die Seite zu stellen. DaS in dem einen Falle die Diffusion ein Glied des gesamten Sekretionsprozesses wird, was vielleicht auch bei anderen Driisensekretionen geschieht , verhdert den Kernpunkt nicht; dieser ist die spezifische Zellentitigkeit der Lunge wahrend der Gaswanderung und hierdurch die R e g u 1 i e r u n g der Sauerstoffaufnahme und der Kohlensiiureabgabe. Eine derartige Regulierung mittels der Lungen- zellen wird durch die sogenannte Difffusionshypothese ausgeschlossen, und hierin liegt biologisch gesehen der Hauptunterschied zwischen dieser und der Sekretionshypothese.

OUICR DIE SPEXIFISCHE TATIGKEIT DER LUNGE usw. 241
\Vie mir gesehen haben, erwiesen die Bestimmungen der Gas- spannungen im Blute und in der Lungenluft. sich als sehr belangreicli fur den Nachmeis einer in der Lunge stattfindenden Zellentatigkeit untl fur die Stellung der Fragen, die es in erster Linie zii untersuchen gilt; die Losung dieser Fragen, die fur die allgemeine Sekretionslehre so groBes Interesse darbieten, l i6t sich wohl schmerlich durch Spannungs- bestimmungen allein vollfuhren. Was am meisten not tut, um tiefer in die Sache eindrinpen z u kiinnen, scheint sichere Bestimmungen der GrBBe der einfachen Gasdiffusion durch die Lunge und eine erschiipfende Berechnung der physikalischen Bedingungen dieser Diffusion zu sein. Hierdurch erst hat man die Aussicht, sich eine begrundete Meinung fiber den U m f a n g des aktiven Prozesses und uber dessen variierenden Anteil an dem gesamten Gasmanderungsvorgange unter verschiedenen Verhaltniseen bilden zu konnen, und vielleicht darf man au6erdem auch hoffen, hierdurch Anhaltspunkte fiir die Beurteilung der Art der Zellentiitigkeit zu gewinnen, deren Existenz unbestreitbar durch die Spannungsbestimmungen dargelegt worden ist.
Die Wichtigkeit quantitativer Untersuchungen uber die GroBe der Lungendiffusion ist iibrigens schon langst erkannt und sowohl H u f n e r als auch Loewy und Zun t z haben derartige Untersuchungen angestellt, die im nachsten Abschnitt zu nlherer Besprechung kommen werden.
A bsc hn i t t 11.
lethodisches zur Feststellung der Diffusionsgr8Be der Base am der AlveoleniuPt ins Blut. Berechnung der im Organismus bei dem Gaswechsel unter verschiedenen Vcrhlltnissen in
Betracht kommenden Druckkrilfte.
Um die GroBe der Gasdiffusion berechnen zii konnen, die unter bestimmten gegebenen Bedingungen im Organismus zwischen der Alveolenluft und dem die Lunge durchstromenden Blnte stattfindet, muB man in enter Linie uotwendigerweise diejenige Menge Gas kennen, die wlhrend der Zeiteinheit die Alveolarwand, passieren wiirde, wenn der Spannungsuntersohied dieses Gases an beiden Seiten der Membran eine Druckeinheit (1 mm) betriige, die hierbei in Betracht kommenden Methoden merden wir im folgenden 0 1 nlher durchgehen.
AuBer dieser Konstanten muB selbstverstlndlich aber auch die unter den gegebenen Bedingungen wirklich existierende Differenz zmischen
Wandin. ArcbIv. XXII. 16

242 CHRISTIAN BOHB:
den Spannungen des betreffenden Gases an beiden Seiten der Alveolar- luft bekannt sein. Handelt es sich z. I3. um die Diffusion des Sauer- stoffs aus der Alreolenluft ins Blut, so muB man die Spannung dieses Gases an beiden genannten Stellen bestimmen. Xun rerhalt es sich bekanntlich so, daB, wahrend die Alreolenluft, solange der Organismus unter denselben Bedingungen verbleibt, eine leicht bestimmbare, einiger- maBen konstante Spannung hat, dies dagegen mit den1 Blute mihrend deren Passage durch die Lunge nicht der Fall ist. Dieses tritt als Venenbliit mit einer dementsprechenden Sauerstoffspannung in die Lunge ein; wahrend der Passage durch die Lunge nimmt es aus der Alveolenluft Sauerstoff auf, und hierbei steigt seine Sauerstoffspannung fortwahrend, so da6 es beim Verlassen der Lunge gewohnlich fast rnit Sauerstoff von der Spannung der Alreolenluft gesattigt ist. Um die fur die Diffusion mafigebende Differenz zwischen der Sauerstoffspannung der Lungenluft und der des Blutes bestimmen zu konnen, muB man deshalb aus den jeweilig wiihrend der Durchstromung der Lungen ver- sohiedenen, stets anwachsended Sauerstoffspannungen einen mi t t l e ren W e r t fiir das Blut berechnen. Der Wert dieser mittleren Spannung wird sich als verschieden erweisen, je nach dem gro6eren oder geringeren Sauerstoffgehalt des Venenblutes hei dessen Eintritt in die Lunge, wie auch die absolute GriiBe- der Alveolenluftspannung auf die mittlere Spannung Einflu6 iiben wird. Den Wert dieses Faktors unter normalen Verhlltnissen wie auch dessen Variation bei wechselnden Bedingungen der Atmung berechnen zu kiinnen, ist deshalb eine fur unsere Auf- gabe unumghgliche Voraussetzung, deren rationelle Lbsung im 5 2 be- sprochen werden wird. Die Voraussetzung, wo man, wie hier, die Diffusionsbedingungen in ihrer Reinheit zu untersuchen wiinscht, ist iiberall natiirlich die, da6 man von jedem der im Abschnitt I nach- gewiesenen Einfliisse einer spezihohen Zellentlitigkeit absieht.
$j 1. iiber die bei der leesung der Lungendiffnaion in Betracht kommenden Pethoden.
Diejenige Menge Gas (u), die nach eingetretenem Beharrungs- zustande wlhrend einer gegebenen Zeit (Q) durch eine Schicht Fliissig- keit oder eine von Fliissigkeit durchtriinkte Membran mit der Ober- flache 0 diffundiert, la& sich, wie E x n e r und S t e p h a n s Unter- suchungen nachgewiesen haben, bekanntlich als
ausdriicken, wo P und p die Spanoungen an den beiden Seiten der

~ B E R DIE SPEZIFISCHE TATIGKEIT DER LUNGE usw. 243
Nembran, d die Diclie der letzteren und u den Bunsenschen Ab- sorptionskoeffizienten des Gases in der Flussigkeit bezeichnen ; K ist eine fur das betreffende Gas und die betreffende Flussigkeit bei gegebener Temperatur spezifische Konstante. Wahlt man mit Stephan zur Einheit fur die Oberflache der Fussigkeitschicht 1 qcm, fur deren Dicke 1 cm und fur die Zeit 24 Stunden, so findet man K riicksichtlich der Diffusion der Kohlensaure durch Wasser von 16O gleich 1.38. Nach Exners Untersuchungen, die mit Bezug anf Wasser und die fur uns in Frage stehenden Gase durch Versuche von H u f n e r l bestatigt worden sind, ist k umgekehrt proportional xur Quadratwurzel des spezifischen Gewichtes der Gase (m); es ist deshalb zweckml6ig, wie von Loewy nnd Zun t z vorgeschlagen, * statt K eine andere, als Diffusionsfaktor (cl) bezeichnete Konstante einzufuhren, indem man c1 = kT/;;t setzt. Be- nutzt man die obeugenannten Stephanschen Einheiten, so findet man den Dift'usionsfitktor bei 1 6 O also gleich 9.154; fur die Berechnung der Diffusion durch so groSe Flachen, wie die Lunge darbietet, ist es ferner bequemer, mit Loewy und Zuntz (a. a. 0.) die Einheiten der Zeit und der Dicke der Fliissigkeitsschicht aus 24 Stunden und Zentimetern in Minuten und Millimeter umzuandern und den Diffusionsfaktor daher rnit 144 zu dividieren. &tan findet dam, da6 bei 16O der Wert des Diffusions- faktor (c) 0.0636 betragt. Die Anzahl Kubikzent'imeter (Oo 760mm) der verschiedenen Gase, die wahrend T Minuten durch eine dmm dicke Fliis- sigkeitsschicht mit der Oberfllche Oqcm dringt, wird nun, indem man die obigen Bezeichnungen benutzt,
a - c T. 0. (P c p ) d * 760 1/%
V =
Mittels einer Formel dieser Art hat Hiifner und spiiter Loewy und Zuntz die unter verschiedenen Verhlltnissen durch die Lungen statt- findende Gasdiffusion zu berechnen versucht. Hierzu ist es selbstredend notwendig, zu erfahren, welche Werte in dem speziellen Falle den in der Formel vorkommenden Konstanten (a, c , 0, d) beizulegen sind. Das bietet indes, wie wir im folgenden sehen werden, Schwierigkeiten dar, die hinsichtlich einiger der Konstanten, zurzeit wenigtens, als nniiber- windlich zu betrachten sind.
HiifnerS nimmt an, dak? das Lungengewebe, was die Diffusions-
' Ann. d. Physik u. Chemie. Neue Folge. 1897. Bd. LX. S. 134. Loewy und Zuntz in Michaelis H a h h df?r Sauerstofffherapie.
Archiv f. Physiologk. 1897. S. 122. Berlin 1906. S. 43.
16 *

244 CHRISTIAN BOHR:
bedingungen betrifft, als ein von Wasser durchsetztes Gewebsgerust betrachtet werden konne. Fiir ein solches wiirden dann der Absorp- tionskoeffizient (a) und der Diffusionsfaktor (c ) dieselben wie fur Wasser sein, und es wiirde, speziell betreffs der Lunge, nur die Bestimmung der Oberflache (0) und der egektiren Dicke (d) der Scheidemand zwischen der Aloeolenluft und dem Elute erubrigen. Die Grd3e der Oberflache schatzt Ht i fner auf 90qm mit 70'lm GePiBoberfliche, er bemerkt aber, da6 die Schitzung unsicher sei, so daB an und fur sich nichts uns verwehre, die Oberflache, mie einige Anatomen es tun, auf das Doppelte anzusetzen. Schon hierdurch gerat freilich einige Un- sicherheit in die Berechuung der diffundierenden Gasmenge, weit griiBere Schwierigkeiteu bereitet jedoch die Feststellung der Dicke der Schicht. Hii fner nimmt an, daB der Dickedurchmesser des zwischen Lunge und Blut geschalteten Gewebes durchschnittlich hochstens 4 p betrage. Er betrachtet indes, wie gesagt, dieses Gemebe in Analogie mit den yon ihm zu Diffusionsversuchen benutzten Hydrophanplatten als aus einem von Wasser durchsetzten Geriiste bestehend , das der Diffusion einen Widerstand darbiete; dieser Widerstand erscheine, teils indem das Geaebe als siebformiges Gebilde die Oberflache oermindere, teils ,,indem es nicht ron kurzen Kanilen, die direkt und gerade TO^ der einen Seite zur anderen gehen, sondern von mannigfach ausammenhangenden Hohlriumen durchsetzt ist, die den Weg verlangern, den die diffun- dierenden Molekiile zu durchlaufen haben." Dann wiirde die Diffusion durch das Gewebe unter sonst gleichen Verhiltnissen langsamer erfolgen als durch eine Wasserschicht von derselben Oberflriche und Dicke; die direkt gemessene Dicke (411) mii6te in der Berechnung deshalb durch eine gr6Bere ersetzt werden, die dadnrch entstunde, da6 man zur gemessenen Dicke den ,,Diffusionswasserwert" des Widerstandes addierte. Ob das Lungengewebe sich nun wirklich der Diffusion gegeniiber einer wasserdnrchtrinkten Hydrophanplatte analog verhalt, laBt sich wohl kaum mit Sicherheit entscheiden ; jedenfalls l i B t sich der even- tuelle Diffusionswiderstand in diesem Falle aber nicht messen. H u f n e r nimmt freilich an, das Gewebsgeruste erzeuge einen solchen Wider- stand, da6 d in der Berechnung passend auf das Zehnfache des gemes- senen Wertes, also auf 40p anzuschlagen sei; es wird aber kein geniigender Grund fur die Wahl eben dieser Zahl angefuhrt; seine Annahme ist ganz willkiirlich, und man kiinnte mit derselben Berech- tigung eine andere Zahl tviahlen. Da somit eine ganz willkurliohe GriiBe in die Bereohnung eingefuhrt wird, kann man dem Resultate,
Hufner, Annalen d. Physik u. Chemie. 1897. Rd. LK. S. 141.

~ B E R DIE SPEZIFISCHE TATIGKEIT DER LUNGE GSW. 245
demzufolge bei 110"" Druck 724cCm Sauerstoff pro Ninute durch die menschliche Lunge diffundieren sollten, keine Bedeutung beilegen.
Im Gegensatz hierzu suchen Loemy und Zun tz l als Grundlage fur die Berechnung der Diffusion durch die menschliche Lunge samt - l i che Konstanten der oben angefuhrten Diffusionsgleichung zu be- stimmen. Wghrend Hufne r , mie gesagt, annimmt, die Diffusion durch das Lungengewebe finde wie eine Diffusion durch Wasser statt, niir unter sonst gleichen Umstanden langsamer wegen des vom Cfewebs- geriiste erzeugten Wderstandes, nehmen Loewy und Zuntz auch diese Frage zur experimentellen Behandlung auf. Als Versuchsobjelit benutzten sie bei diesen Untersuchungen F r o s c h 1 u n g e n , indem sie die Geschwindigkeit maBen, mit welcher Kohlendure oder Stiok- oxydul aus den Hohlraumen der Lunge in die urugebende atmo- sph5rische Luft diffundiert; keiint man die au6ere Oberflache der Lunge und die Dicke der Schicht, welche die Begrenzung ihrer Hohlriiume nach au6en bildet, so kann man hieraus C C * C (das Produkt aus dem Absorptionskoeffizienten und dem Diffusionsfaktor) berechnen. Da6 man hierdurch nur das Produkt, der beiden Konstanten, nicht aber den IVert jeder Konstanten for sich findet, ist fur die Berechnung der Diffusion des betreffenden Gases durch dasselbe Medium unter verschiedenen Drucli- und Oberflichenverhaltnissen selbstverstandlich ohne Belang; auch f i r die Obertragung der Berechnung auf die Diffusion anderer Gase durch dasselbe Nedium hat dies keine Bedeutung, denn da c in diesem Falie konstant ist, so ist das Verhaltnis zmischen den Produkten a. c hinsichtlich zwei verschiedener Gase gleich dem Verhaltnisse zwischen den Absorptionskoeffizienten dieser Gase im Lungengewebe, einem Verhaltnisse, das mir wohl gleich dem bei ihrer Absorption ia Wasser gefundenen setzen diirfen.
Die von Loewy und Zuntz benutzte Methode bietet den Vorteil dar, da6 das eigentliche Messen des durchdiffundierten Gasvolumens bequem ist und durch mehrere unmittelbar aufeinanderfolgende Wieder- holungen des Versuches gesichert werden kann. DaB man nicht die Diffusion durch die die Lungenluft vom Blute trennende Wand, sondern die durch die ZiuSere Begrenzung der Lunge priift, kommt wohl auch h u m in Betracht, solange die Frage prinzipieller Natur ist und wesent- lich bezweckt, zu priifen, ob die Dihsion durch .ein tierisches Gewebe dieser Art sich hauptsachlich wie die Diffusion durch Wasser verhllt. Was die numerischen Werte betrifft, so fanden Laewy nnd Zuntz die Konstante t c ' c sowohl f i r CO, als fiir N,O hinsichtlich des Lungen-
Archiv f i Physiologie. 1904. S. 166.

246 CHRISTIAN BOHR:
gemebes doppelt so groB alu hinsichtlich des Wassers, und sie sind geneigt, dies mit einem entsprechend grofieren Werte des Absorptions- koeffizienten (.) fur die Losung der Gase im Lungengewebe, der rielleicht auf dem Gehalte dieses Gewebes an lipoiden Stoffen beruht, in Be- ziehung zu setzen.
Bevor man die somit fur die Konstante gefundenen Werte als hinlinglich sicher anerkennt, muB man indes folgendes in Betracht ziehen. In der Bestimmung von u.c kommen auBer der Menge des diffundierten Gases, die sich nach der angegebenen Methode zweifels- ohne mit hinllnglichet Genauigkeit bestimmen liBt, zugleich auch die Gro6e der iuBeren Oberflache der Lunge und die m i t t l e r e Dicke der Lungenwand vor. Die Messung der Oberflache der Lunge bietet zwar keine besonderen Schmierigkeiten dar ; dies ist dagegen im hijchsten Grade mit der Peststellung eines auch nur annlhernd richtigen Wertes f i r die mittlere Dicke der Wand der Fall, und zwar selbst, wenn man annehmen will, woriiber nlheres unten, daB die angewandten Fixations- mittel keine h d e r u n g der Dicke der Wand bewirken. Man hat nlmlich zu beachten, dal3 die Dicke der Wand bei weitem nicht die gleiche in der gesamten Fliiche ist; wie von Loewy und Zuntz her- vorgehoben, ist sie im Gegenteil sehr variabel und durchmeg starker in der NIhe der Septa, dunner auf der zwischen diesen liegenden Strecke. Unter solchen Verhiiltnissen zu einer auch nur annlhernd richtigen Bestimmung der m i t t l e r e n Dicke zu gelangen, diirfte lu6erst schwierig sein und erfordert jedenfalls eine sehr systematische und hochst umfangreiche Reihe von Messungen ; solche, sogar recht zahl- reichen Messungen der variierenden Dicke an verschiedenen nach Gut- achten gewiihlten Stellen zu unternehmen, oder die Mittelzahl aus einer Anzahl Messungen der Dicke an einem linearen Schnitte durch die Wand zu berechnen, wird offenbar nicht gentigen; es wird notwendig sein, drts Areal, dessen Dicke man zu messen wlinscht, in Zonen von annahernd derselben Dicke zu teilen und das VerhZiltnis xwischen den Arealen dieser Zonen zu bestimmen. Hierzu kommt, da6 die einzelnen ,,Alveolen" durch Aufblasen sicherlich in verschiedenem MaBe rtufgebllht werden und mithin rerschiedene Dicke der Wand erhalten, weshalb die Messung sich auf eine bedeutende Anzahl der- selben erstrecken mu6. Eine solche systematische Zonenmessung scheint von Loewy und Z u n t z nicht angestellt worden zu sein, wenigstens nicht in den Versuchen, wo die Anzahl der einzelnen Messungen an- gegeben wird (z. B. im Versuchc Seite 199, wo die Anzahl 30 betrigt). Zu den Schwierigkeiten der Messung der Wanddicke kommt nun ferner, da6 man nicht mit Sicherheit weil3, ob das Gewebe sich nicht mahrend

OBEH DIE SPEXIFISCHE TATIGKEIT DER LUNGE usw. 24 7
der Fixat ion verandert hat. Diese erfolgte teils mittels Formalins, teils mittels Sublimats, teils mittels Fullung der Lunge mit Gelatine, die darauf durch ermlrmte Kochsalzlosung wieder abgeschmolzen wurde. Nun ist die Wirkung dieser PrHparationsmittel bekanntlich je nach ihrer Konzentration und dem Zustand des Gewebes sehr verschieden, und es ist sehr wenig wahrscheinlich, daB die Zellen mahrend der Behandlung ihre Dimensionen unverandert erhalten haben. DaB die iu6ere Form des Organs sich bei der Formalinfixation nicht veriindert, hat in diesem Zusammenhang nichts zu bedeuten; so findet Krogh,' da6 die aufgeblahte Froschlunge xiur sehr wenig einschrumpft, menn sie iiber Schwefelsiure getrocknet wird, wahrend die Zellen hierdurch doch ganz einschrumpfen ; auch eine Saugetierlunge verlndert ihre Dimensionen nicht , wenn die GefiiBe mit einer hypotonischen Losung gefiillt werden, die die GefaBzellen zum Aufquellen bis zur vblligen Unwegbarmachung des GefiiBes bringt. Es wird uns durch die von Loewy und Zun tz angestellten Versuche daher nicht verbiirgt, da6 die angegebene mittlere Dicke der wahrend des Versuches tatsachlich vorhandenen mit geniigender Annaherung entspricht, und in diesem Falle wird dann auch in betreff der Konstante a . c eine entsprechende Unsicherheit herrschen, denn diese murde aus einer Gleichung berechnet, in welcher die Dicke (4 als Faktor vorkommt.
. Bei der von Loewy und Zuntz unternommenen Berechnung der Diffusion durch die menschliche Lunge mu6 man indes au6er der oben diskutierten Konstanten a. c zugleich auch die T,ungenoberfllche und die Dicke der Schicht, welche die Alveolenlnft vom Blute trennt, kennen. Auch die Bestimmung dieser beiden letztgenannten GrbBen kann nur mit groBer Unsicherheit geschehen, ja, man mu6 gestehen, daB es uns zurzeit, iiberhaupt an Mitteln gebricbt, die effektive Dicke der Wand rnit hinliinglicher Annlherung zu bestimmen. Was nun erst dieMessung der Lungenoberfliiche betrifft, so mu6 man hier, was gewohnlich dsnn auch geschieht, zwischen der Messung der gesamten Oberfliche und der Messung desjenigen Teiles derselben, den die GefiBe einnehmen, nnterscheiden. Die gesamte Oberfllche ISiSt sich wohl trotz der ziemlioh schwankenden Werte, die man auf anatomischem Wege gefunden hat, anniihernd auf 909" ansetzen, wie von Zuntzs angegeben, der in sehr sinnreicher Weise als Grundlage fiir die Berechnung derselben die Kapazitat der Lunge und den mittleren Durchmesser der dlveolen ver- wertete. Einen wie groBen Teil hiervon wieder die Oberfllche der
Dies Archin. 1904. Bd. XV. S. 401. Hermann, Handbuch d. Physiologie. Leipzig 1882. Bd. IV. 11. S. 90.

248 CHRISTIAN I ~ O H H :
GefiiBe bildet, ist aber unsicher, in der Regel nimmt man an, dab diese etwa 70 Proz. der geeamten Oberfliiche betriigt.
Was die Dicke der die Lungenluft rom Blute scheidenden Schicht betrifft,, so mird diese con Hi i fner auf Basis einer sehr summarischen Sohltzung auf hiichstens 4 p im Yittel angeschlagen, und dies findet Zustimmung bei Loewy und Zuntz. So, wie diese GriiBe gefunden murde, miissen mir sie indes als eincn Ausdruck der kiirzesten, direliten Entfernung zwischen der Oherfliiche der Lunge nnd dem Inhalt der Kapillaren betracht,en ; nun sind bekanntlich die Zmischenraume zmischen den IhpillargefiiBea im Vergleich hiermit groB, und rechnet man daher mit der gesam ten Lungenoberflliche als gasnufnehmend, so darf man nicht zugleich mit der kurzesten IMfernung nach den Kapillaren rechnen, indem eine groBe Menge der diffundierenden Molekiile sich in schriigen Bahnen bewegen werden, wodiirch die effek t i r e Schicht- dicke sich vergrijBert, vielleicht sogar in bedeutendem MaBe; um einen auch nur annahernd richtigen numerischen Wert derselben festzustellen, gebricht es uns jedoch an hinlanglichen Momenten.
Es ist aber noch eine andere Seite der Sache in Betracht zu ziehen, die die numerische Feststellung der Konstanten ganzlich un- sicher macht. Der Sauerstoff - und hier handelt es sich corzug- lich um dieses Gas - muS bei seiner Aufnahme in den Organismus nicht allein die Alveolarwand passieren und bis zum stromenden Plasma gelangen, ron dem wir anzunehmen haben, dab seine veFschiedenen Schichten wiihrend seiner Bemegung fortmahrend gleichmabig gemischt werden, sondern das Gas muB auch zum weit iibermiegenden Teil in die B l u t k o r p e r c h e n eindringen, ivo es rom Hamochrom gebunden aird. Dies ist ein Moment, das von Hi i fner nicht erwahnt mird; Loewy und Zuntz beriihren es allerdings, legen demselben aber keine be- sondere Bedeutung fur die Berechnung der Diffusion bei, indem sie annehmen, daB der Sauerstoff in der Substanz der Blutkorperchen mit besonderer Geschmindigkeit wandre, da er sich hier in einem Medium bemege, das ihn in ca. 50 fach groBerer Dichte als das Wasser absorbiere. Es ist indes auf Basis unserer jetzigen Kennt,nis von dieser Frage in der Tat ganz unmoglich, auch nur mit einiger Annaherung den Wert der Diffusionsgeschwindigkeit der Gase durch die teigige Liisung, die das Hiimochrom in den Blutkorperchen bildet, anzugeben. Ob die Eigenschaft des Hgmochroms als sauerstoff bindender Korper hierbei, wie Loewy und Z u n t z meinen, iiberhaupt in Betracht kommt, ist jedenfalls iiuberst zweifelhaft. Der Bun sensche Absorptionskoeffizient,
A. a. 0. S. 205. Anmerkung.

UBER DIE SPEZIFISCHE TATIQKEIT DER LUNGE usw. 249
der mohl besonders auf die Geschmindigkeit influieren miiBte, betragt fur die Blutkorperchen nur ca. 81 Proz. von dem des Wassers; anderer- seits ist aber der Wert des Diffusionsfaktors unbekannt, indem die Substanz der Blutkorperchen sich megen ihrer gro6en Konzentration und ihres Gehnlts an lipoiden Stoffen hinsichtlich der Diffusions- bedingungen nicht ohne weiteres mit dem Wasser zusammenstellen 156t. Die in den Lungen zirkulierenden Blutkorperchen bieten aller- dings eine bedeutende Gesamtoberflache dar, die man, wenn das in der Lunge enthaltende Blut auf 7 Proz. der gesamten Blutmenge an- gesetzt m i d , auf gegen 200 qm veranschlagen kaun; demungeachtet ist bei unserer mangelhaften Kenntnis der fur die Substanz der Blut- korperchen geltenden Diffusionsbedingungen die Moglichkeit keines- wegs ausgeschlossen, daB ein nicht uubedeutender Teil der zur Diffusion rerstreichenden Zeit gerade Ton der Aufnahme der Gase in die Blut- korperchen beansprucht mird.
Wie man sieht, hat man wegen der groBen Unsicherheit der Be- stimmungen samtlicher Konstanten durchaus keine Garantie, da6 die durch die Berechnung gefundenen Werte der Diffusionsgeschwindigkeit der Gase in der Lunge auch nur einigerma6en den wirklichen ent- sprechen, und man ist deshalb auch nicht berechtigt, die auf diese Weise berechneten Werte als Grundlage fernerer Folgerungen hin- sichtlich des respiratorischen Prozesses zu benutzen. Dies gilt selbst- rerstandlich auch von dem ron Loewy in seiner Respirationslehre auf diesem Wege gefundenen Resultate, lsut dessen der Sauerstoff bei einer Druckdifferenz von 35mm aus der Lungenluft in das Blut in einer Mengc von ca. 6000 cem (fir eine Lungenflache von 90 qm) oder von ca. 9500 (fur eine Oberfliicbe von 140 qm pro Minute diffun- dieren sollte.
Um die Diffusion durch die Lunge zu messen, eriibrigt daher wohl kein anderer Weg als der, der iibrigens auch unmittelbar als der naturlichste und sicherste zu betrachten ist, niimlich der direkte Venuch, durch den an einer b lu tdurchs t romten Lunge der quan- t i t a t ive Obergang von Gasen aus der Alveolenloft i n das B l u t un te r geeigneten Bedingungen bes t immt wird.
Es liegt nahe, die Probe anzustellen, derartige Versuche an der herausgenommenen Siugetierlnnge auszufiihren, wo die Kreislauf- und Atmungsverhiiltnisse dem Normalen mbglichst nahe zu bringen wlren ; man wiirde hier zugleich leichten Zutritt zu jeder erwiinschten Variation
Oppenheimer, Biochemie. Die Gase des K8rpers und der Cfastoechel. s. 106.

250 CHRISTIAN BOHR:
der Bedingungen haben. Nachdem ich - iibrigens nicht ohne Schwierig- keit - eine geeignete Technik ausgearbeitet hatte, habe ich denn auch eine Reihe solcher Versuche ausgefiihrt. Es gelang mir indes nicht, auf diese Weise brauchbare Resultate zu erzielen, indem das Priiparat sich als gar zu leicht rerinderlich erwies. Die Strijmungs-
. geschwindigkeit des Blutes lieB sich wlhrend der Versuche nicht lange genug auf angemessener Hohe erhalten, und, was die Sache besonders erschwerte, es wurden stets Partien der Lunge schnell durch Emphy- sem, Odem oder Atelektase verandert; deswegen wurden die Resultate selbstverstandlich unbrauchbar, indem die Aufnalime von Gasen sich in unbestimmbarem Grade verminderte.
Befriedigende Resultate lassen sich dagegen durch Bestimmung der Lungendiffusion an dem lebenden Tiere erzielen. Selbstredend ist es dann notwendig, ein Gas anzuwenden, bei dessen Aufnahme die Zellentltigkeit aller Wahrscheinlichkeit nach keine Rolle spielen kann, und wo zugleich das Blut mlhrend seiner Passage durch die Lunge nicht mit dem Gase viillig gesrittigt wird, denn alsdnnn wiirde die aufgenommene Menge natiirlich nur als ein minimaler Wert der Diffusion Bedeutung haben. Am Kohlenoxyd besitzen wir nun ein Gas, das die beiden genannten Bedingungen befriedigt und das au6erd dem den Vorteil darbietet, daS es ebenso wie der Sanerstoff vorxugs- weise in den Blutkorperchen und nur in geringer Menge im Plasma anfgenommen wird. Dies ist von Wichtigkeit fiir die Obertragung der gefundenen Werte vom Kohlenoxyd auf den Sauerstoff, welches Gas ja besonderes Interesse darbietet ; mit Bezug auf beide dieser Gase werden niimlich die oben erwlhnten besondere Diffusions- bedingungen fir Gasaufnahme in die Blutkorperchen zur Geltung gelangen.
Es trifft sich nun so giinstig, da6 in der Literatur bereits gut miteinander iibereinstimmende Versuche uber die Aufnahme des Kohlenoxyds in die Lunge vorliegen. Diese Versuche, die wir zum Teil Grehant , zum Teil Haldane verdanken, sind freilich nicht urn die Diffusion durch die Lunge zu messen angestellt worden; sie wurden indes unter Bedingungen ausgefiihrt, die, wie wir in einem folgenden Abschnitte sehen werden (Abschn. 111, 5 l), die Ansftihrung einer solchen Berechnung gestatten. Hierdurch kaun man nun in betreff des Kohlenoxyds zur Bestimmung derjenigen Menge Gas gelangen, die bei einer Spannungsdiferenz von l m m in der lebenden Lunge pro Minute aus der Alveolenluft ins Blut iibergeht. Diese GriiSe (B) liiI3t sich nach bekannten Regeln so umrechnen, daI3 sie auch fiir andere Gase, speziell Sauerstoff und Kohlenslure gilt (Abschn. 111, 0 2). Fiir diejenige

OBER DIE SPEZIFISCHE TATIG~KEIT DER LUNGE usw. 251
Menge Gas (v), die wlhrend T Minuten durch die Lunge diffundiert, hat man dann
wo Y die Spannung des Gases in der Alveolenluft, p dessen mi t t le re Spannung im Blute bezeichnet. Die Bedeutung dieser lehteren GroBe wurde bereits oben beriihrt; im untenstehenden 5 2 werden wir niher entwickeln und durch Beispiele erlautern, wie sie sich in einem ge- gebenen Falle berechnen IiiBt.
v = D ( P i p ) T,
5 2. Methode znr Berechnung der mittleren Gasspannung des Blntee wiihrend seiner Stromung dnrch die Lnngenkapillaren. Werte der mittleren Sanerstoff - und Kohlensiinreapannung im Blnte bei ver-
schiedenen Spannnngen dieser Gsse in der Alveolenluft.
Indem das Venenblut mit einer gewissen Partialspannung (pa) eines in ihm aufgelosten Gases - dies miige z. B. Sauerstoff sein - in die Lunge hineinstrGmt, wo die Partialspannung des Sauerstoffs in der Alveolenluft sich auf einem konstanten Werte P groSer als pa erhiilt, wird durch Diffusion aus der Alveolenluft Sauerstoff ins Blut aufgenommen, dessen Sauerstoffspannung hierdurch fortwahrend steigt, bis es mit der Sauerst,offspannung p , die Lunge verlgBt. Hierbei kann p , sich selbstyerstlndlich P mehr oder weniger nahern, kann aber nie gr6Ber als dieser Wert werden, da wir hier nur die Wirkung der Diffusion beriicksichtigen und von der spezifischen Zellentltigkeit glnzlich absehen.
Es handelt sich nun darum, einen mittleren Druck p' zu finden, so daS durch die Druckdifferenz P + p' ganz dieselbe Diffusion bewirlit werden wiirde, die tatslchlich in der Lunge stattgefunden hat, 80
die Druckdifferenzen zwischen Alveolenluft und Blut, wlhrend letzteres die Eapillaren durchstrtimte, unablissig variierten, von P + pa bis Y i p, abnehmend.
Der numerische Wert des mittleren Druckes in einem gegebenen Falle mu6 selbstverstiindlich zwischen den WerQen pa und pa liegen, kann aber nicht, wie von Loewy angenommen, gleich deren Hittelzahl
I' + p b gesetzt werden. Wie die folgende Berechnung zeigt, ist der Wert in mehr komplizierter Weise Ton den vorkommenden GroSen abhlngig.
Die Bereohnung des Wertes des mittleren Druckes, die sioh auf
(+I
L o e w y in Oppenheimers Biochemie. S. 105.

252 CEIRISTIAN BOER:
das allgemeine, oben im 0 1 besprochene DiffusionsgeseOz stutzt, 1aBt sich folgenderma6en ausfuhren. Es stelle Fig. 1 die Verhlltnisse in einem an den Alveolarraum grenzenden KapillargefiiBe cor; a b ist die das Blut (unterbrochene Linien) von der Alveolenluft (punktiert) schei- dende Schicht; das Blut tritt bei a in das Kapillargefi6 ein und ver- ll6t dasselbe bei b. Wir benutzen iibrigens bei der Berechnung, bei welcher eine rollstandige Mischung des Blutes in jedem Querschnitte corausgesetzt mird, folgende Bezeichnungen:
pro Liingeneinheit des Iiapillargefabes; 3 S wirksame Oberflache P Volumen L Lange des I<apillargefiiBes; z Linge des KapillargefiiBes bis zum betrachteten Punkte; v die mittlere Geschwindigkeit der Flussigkeit; y die Menge des pro Volumeinheit geliisten Gases an dem be-
trachteten Punkte;
Als Grundlage fur die obenstehende Berechnung sind ausschlieBlich die allgemeinen Diffusionsgesetze benutzt. Ich mochte bei dieser Oelegenheit die Aufmerksamkeit darauf leiten, daB bei der vou mir anderswo fruher be- nutzten In- und Evasionsberechnung wohl die theoretische Bebandlung als zutreffend und unter Umstanden mit Nutzen anwendbar anzusehen ist, dab aber die von mir angegebenen Invasionskonstanten sich als zu niedrig erwiesen baben, wie dies schon mit Recht von L o e w y (Oppenheimers Biochemie, Gase des KGrpers, S. 124) an der Hand biologischer Tatsachen hervorgehoben worden ist. - Ich war (einerseits mit Hilfe einer, der friiheren ( W i e d e m a n n s Ann. 1899. Bd. LXVIII. 8. 500) analogen, aber vielfach verbesserten Methodik, anderseits durch Versuche anderer Art, wo die Gasaufnabme in einer m8g- lichst dunnen , eine Glaswand hcrabstromenden Flussigkeitsschicht bestimmt wurde) zu ubereinstimmenden Resultaten gekommeo, und zwar zu den in N a g e l s Handbuch fur 0% und CO veroffentlichten. Indessen zeigten weitere Untersuchungen, daB die Grenze der Flussigkeitamischung, zu der ich ge- kommeii, keine definitive war, sondern daB duruh andere Methoden ein be- deutend gr6Berer Wert der Koeffizienten sich orreichen lieB. So wurde bei der Untersuchung des Vcrhaltens einer in strsmender Fliissigkeit angebrachten kleinen Gasblase fur die Invasionskoeffizienten ca. 3.5 ma1 groEere Werte ge- funden. Die numerischen Resultate, zu welchen ich bei der Theorie des Tono- meters und anderer lhnlicher Vorgiinge, wo die Aufnahme der Gase in Fliissig- keiten auBerbalb des Organismus geschieht , gekommen bin, bleiben hierbei wesentlich unberiibrt. Dagegen bewirkt die Erhshung der Koeffizienten, wie leicht ersichtlich, eine hedeutende Anderung in den von mir frither gefundenen Werte fur die Spannung der Gase in Oberfllchen, speziell auch in der Lungen- oberfliiche; diese mu6ten mit Hilfe der neuen Koeffizienten umgerechnet werden. Da man aber nicht sicher sein kann, da8 selbst bei den neuen Versuchen eine wahre Grenze der OberflBchenschicht erreicht worden ist, habe ich vor- derhand, so auch uberall in dieser Abhandlung von einer Auwendung der Invasionsberechnung abgesehen.

UBER DIE SPEZIFISCHE TATIGKEIT DEB LUNGE usw. 253
P der Partialdruck des Gases in der Alveolenluft; p der Gaspartialdruck der Fluseigkeit in einem betrachteten Quer-
schnitt des KapillargefaBes; in der Diffusions- R effektioer Diffusionskoeffizient = ~
d v %
Indem man ausdruckt, da6 diejenige Menge Gas (dy), um welche der Gehalt der Flusaigkeit an aufgelostem Gas (y) wahrend der Passage durch einen Ton zwei Querschnitten begrenzten, unendlich kleinen Teil
a - c
gleichung S. 243.
Fig. 1.
des KapillargefaBes (c. Fig. 1) zunimmt, gleich derjenigen Menge ist, die wegen der Druckdifferenz (P + p ) durch die dem Querschnitt ent- sprechende Oberflache (S d ~ ) diffundiert, hat man
(P - p ) n S d x d t = (y + dy) Pv d t - y Pv d t
(P - p ) n S d x = d y Pv und hieraus
oder
Nennt man die wirksame Oberfllche des KapillargefiiSes in dessen ganzer Ltinge 0, also 0 = S X, und die Menge des durohstrbmenden Blutes M; so da6 M = T'v, und erinnert man sich, da6 p eine Funktion von y, also p = f(v) ist, so erhilt man duroh Integration auf die ge- samte Ltinge des KapillargefaSes von a bis b
Y a
nS 1 - . A vu $ =J- d y .
Ya
1st nun das betxdende Gas, das Henrysohe Gesetz befolgend, einfaoh in der Fltissigkeit adgelGst, so hat man p = fQ = qy, wo q konstant ist. Die Integration ergibt dann
,

254 CHRISTIAN BOER:
Diese letztere Losung ist es indes nicht, was uns in dieser Frage interessiert. Wir sahen freilich, daB bei der Aufnahme des Sauerstoffs und der Kohlensiure ins Blut p eine Funktion von y ist, diese Funktion ist aber keine geradl in ige . Das Verhaltnis zwischen y und p wird dagegen durch die bekann ten Spannungskuryen fur die Aufnahme dieser Gase ins Blut ausgedruckt ; die Form dieser Spannungskurven haben fruhere Untersuchungen in einem fur unseren Zweck genugenden Um- fang ergeben; eine rationelle E'ormel derselben 1aBt sich zurzeit aber nicht mit Sicherheit aufstellen. Trotzdem konnen wir selbstversthdlich, da wir die Spannungskurven aus Versuchen kennen, in jedem einzelneii gegebenen Falle eine numerische Integration unternehmen, die uns den gesuchten speziellen Wert des mittleren Druckes geben wird. Es sei P die Sauerstoffspannung der Alveolenluft, y, = a und ya = b das Volumprozent des Sauerstoffs in dem eintretenden, bzw. austretenden Blute, also im Venen-, bzw. Arterienblute. Die Syannungskurve fur die Aufnahme des Sauerstoffs ins Blut ist bekannt, und aus dieser leiten wir diejenigen Spannungen ( p ) her, die den verschiedenen Volum- prozenten (y) des aufgelosten Sauerstoffs entsprechen. Diese GroBen seien in der 2., bzw. 1. Kolonne der untenstehenden Tabelle angefiihrt, in der 3. Kolonne seien die entsprechenden Werte P + p und in der 4. Kolonne angegeben. P 7 p
Y
0
1
a
i,
Man bildet
P
0
Pl
Pa
Pb
larauf eine
P f p
P
P t p ,
P + p a
P + pb
1 P s p
Kurve (Fig. 2), wo das Volumprozent des Sauerstoffs im Blute (y) die Abszisse, die; entsprechenden Werte von
die Ordinaten sind. Mittels des Planimeters miBb man das P + p Areal, das von der Abshsenachse und den Ordinaten an a und b, wie auch den dazmischen liegenden Kurvenstrecken begrenzt wird

OBER DIE SPEZIFISCHE TATIGKEIT DEE LUNGE usw. 255
(aa , b, b); dieses Areal (A) ist gleich dem gesuchten Integral. Man hat also (S. 253)
b
d y = A . m O 1 nl =JP - f(y)
a
Fig. 2.
Indem man ein Rechteck (a c d b) von demselben Areal wie A auf der Grundlinie b i u bildet, erhiilt dieses eine gewisse Hohe ac =
und man hat dann P + p" n O (b - a) = - A = - - -
P - p' bI
n 0 (P - p') = M ( b - u).
Da M(b - a) die wLhrend der StrBmung des Blutes aufgenommene Menge Sauerstoff ist, so ist P + p ' der Definition zufolge die mittlere Druckdifferenz, denn diese GriiBe ergibt, wie die Gleichung zeigt, durch Multiplikation mit dem effektiven DifTusionskoef6zienten und der Ober- fliiche gerade die wirklich aufgenommene Sauerstoffmenge. Der mittlere Druck p' des Sauerstoffs im Blute ist ja hiermit gegeben; bei den Diffusionsberechnungen werden wir indes besonders von der mittleren Druckdifferenz P + p' Gehrauch zu machen haben.
Wir schreiten darauf zur Anwendung dea oben entwickelten Ver- fahrens zur Berechnung des mittleren Druckes auf best immte Bei- spiele, die wir in den folgenden Abschnitten verwerten werden. Fiir die Relation zwischen Sauerstoffmenge und Sauersbffspannung im Blute benutzen wir die von mir friher angegebenen Werte beim Obergange des
1
und hieraus
~
* Nagels Handbuch. I. I. S. 199.

256 CHRISTIAN BOHR :
Blutes aus Arterien- in Venenblut oder umgekehrt, die in untenstehender Tabelle angefuhrt werden.
Sauerstoffmengen (y) und Sauerstoffspannungen ( p ) beim ubergangc aus Venen- in Arterienblut.
Volumproz. Y
1 2
4
6 7 8 9
10
._ __-___
L 3
' 5
Spaunung
~~
7.8 11.7 14.2 1 6 . 5 18.5 20.4 22.2 24.2 26.0 25.0
I
, Volumproz. 1 Spannung 1 1 P I Y
11 1 30.0 12 32-2
14 37.3 15 41.0 16 ~ 46.0
5 2 . 3
77 -0
_ ~_ ~. - - __
13 j 34.9
I " 15 1 62.0
l9 I 120.0
I
I
An der Hand der obenstehenden Tabelle berechnen wir vorervt die mittlere Druokdifferenz, wie sie sich unter normalen Durchschnitts- verhaltnissen stellt, wo die Sauerstoffspannung in der Alvoolen- lu f t auf 104"" angesetzt werden kann und das zur Lunge striimende Venenblut 12 Volumproz. Sauerstoff enthalt, w8hrend das Arterienblut anniihernd gesittigt ist und z. B. 19 Volumproz. Sauerstoff enthilt. Die Berechnung dieses Beispiels geben wir der Deutlichkeit wegen detailliert wieder, was vielleicht von Nutzen sein kann, da eine Be- rechnung des mittleren Drnckes auf mehrere Punkte der Respirations- lehre Anwendung finden mijchte.
Wir bilden nun erst fGr P = 104 eine Reihe Werte von P + p Ulld - den verschiedenen Werten von y entsprechend. Diese P + p ' Werte, die unter Benutzung der obenstehenden Tabelle am zweck- miibigsten fiir siimtliche ganze Volnmprozente gebildet werden, geben wir in Auswahl in nachstehender Tabelle (5. 257) wieder.
Die Werte fiir y ftihrt man als Abszissen, die entsprechenden Werte fiir 1 als Ordinaten auf in einem fiir die Ausmessung des Areals P t p bequemen MaSstabe, indem z. B. 1 Om der Abszisse 1 Volnmproz. Sauer- stoff 1 cm 'der Ordinate - . 1000 entspricht ; hierdurch entsteht eine Knrve, die in verkleinertem MaBstabe in Fig. 3 (S. 258) wiedergegeben ist. An dieser Kurve m a t man das Areal aus, das durch die Ordinaten fiir 12 und 19 Volumproz. Sanerstoff begrenzt wird, in Fig. 3 also das
P?P
OBER DIE SPEZIFISCHE TATIGKEIT DER LUNGE usw. 257
0 2 5 8 10 12 16
0 ' 1 1 ' 7 . 8 2 ~ 11.7 3 14.2
19 69.9 I9 67.5 19 63.7 19 59.7 19 56.5 19 52.9 19 42.1
4 5 6 8 12 16 18 19
16.5 18.5 20.4 24.2
104 96.2 92.3 89.8 87.5 85.5 83.6 79.8
32.2 71 -8 46.0 1 58.0 62.0 1 42.0 77.0 1 27.0
1 P + p
0.00962 0.01040 0 * 01083 0-01114 0.01143 0.01 170 0 * 01 196 0,01253 0.01393 0 * 01724 0.02381 0 -03704
Areal (12B (719); man findet dieses gleich 132.27qcm; hieraus bildet man das Rechteck (1 2 E P 19), dessen HBhe - 132'27 oder 18.9 betrggt;
diese Hiihe entspricht einem Werte fir - von - = 0.0189; P t p ' oder die mittlere Druckdifferenz ist in diesem Falle also -- - 5 2 ~ 9 ~ ~ .
Sind die Spannung der Alreolenluft und der Sauerstoffgehalt des Arterienblutes dieselben wie im obigen Beispiele, wird aber der Sauer- stoff des Blutes wiihrend der Passage desselben durch die Kapillaren des Kiirpers stjrker ausgenutzt, so daS daa in der Lunge ankommende Blut nur z. B. 2 Volumproz. Sauerstoff enthiilt, so zeigt eine ganz analoge Berechnung des Areals an derselben Knrve (Fig. 31 daJ3 die mittlere Druckdzerenz in diesem Falle griiSer ist, niimlich 67 5 mm.
Untenstehende Tabelle gibt eine Reihe mittlerer Druckdifferenzen bei verschiedenem Sauerstoffgehalte des Venenblutes.
Mittlere Druckdifferenz (P + p') fur P = 104 mm bei verachiedenem Sanerstoffgehalt dee Venenblnh.
- 19712 18.9
P 7 p 1000
0.0189
Mittlere Venenblut Arterienblnt Druckdifferene
Volnmproz. 0, Volnmproz. 0, ! I P s p '
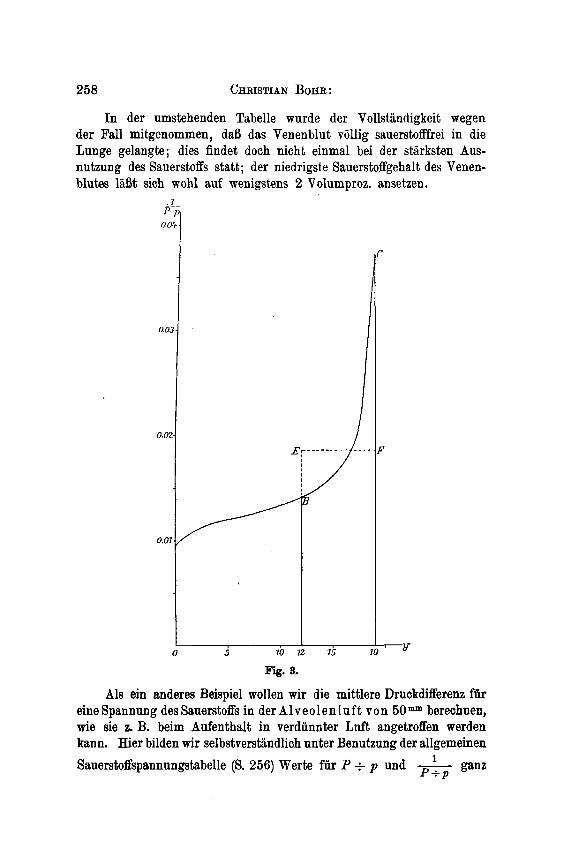
258 CHRISTIAN BOHR :
In der umstehenden Tabelle wurde der VollstLndigkeit wegen der Fall mitgenommen, daS das Venenblut viillig sauerstofffrei in die Lunge gelangte; dies findet doch nicht einmal bei der starksten Aus- nutzung des Sauerstoffs statt; der niedrigste Sauerstoffgehalt des Venen- blutes la& sich Kohl auf wenigstens 2 Volumproz. ansetzen.
0 7 0 12 15.
Fig. 3.
T
F
-Y
Als ein anderes Beispiel wollen wir die mittlere Druckdifferenz fiir eine Spannung des Sauerstoffs in der Alveolenluft von 50mm bereohnen, wie sie z. B. beim Anfenthalt in verdiinnter Luft angetroffen werden kann. Hier bilden wir selbstverstiindlich nnter Benutznng der allgemeinen Sauerstoffspannungstabelle (S. 256) Werte fiir P + p und ganz

OBER DIE SPEZIFISCHE TATIGKEIT DER LUNGE usw. 259
wie im vorhergehenden Beispiele, nur daB P hier gleich 50 mm ist. Da das Blut nach volliger Sattigung bei einer solchen Spannung 16 7 Volumproz. Sauerstoff aufnimmt, lrann dieses Volumprozent in unserem Beispiele naturlich nicht iiberstiegen werden ; man kann daher annehmen, daB das Arterienblut, wenn es fast gesattigt die Lunge verllBt, 16 Volumproz. enthiilt. Indem man die Berechnung sonst dem ausftthrlich wieder- gegebenen ersten Beispiele analog ausfuhrt, findet man folgende Werte fiir die mittlere Druckdifferenz:
Mittlere Druckdifferenz (P + p') fur P = 50 mm bei verschiedenem Sauersto5gehalt des Blutes.
Venenblut Volumproz. 0,
0 2 4 8
12 2
Arterienblut Volumproz. 0,
16 16 16 16 16 12
Mittlere Druckdifferenz
P i p'
19.7 18.0 16.7 13.8 10.4 25.5
Die mittlere Druckdifferenz ist in diesem Beispiel, wie sich vor- aussehen lieI3, niedriger als im vorigen. 1st die Slttigung mit Sauer- stoff des die Lunge verlassenden Blutes eine unvollstindigere, was wiihrend einer hinlinglich geschwinden Strbmung des Blutes denkbar ist, so wird die mittlere Druckdifferenz gr88er; wie obenstehende Tabelle zeigt, ist P s p ' , wenn das Venenblut 2 Volumproz. Sauerstoff, das Arterienblut aber einmal 12, ein andres Ma1 16 Volumproz. enthiilt,
Es kann noch von Interesse sein, zu sehen, wie sich die mittlere DruckMerenz stellt, wenn die Sauerstoffspannung der Alveolenluft eine besonders niedrige ist. Untenstehende Tabelle enthilt die Werte fir
, eine Alveolenluf tspannung von 30mm Sauerstoff. Bei vblliger Siittigung kann das Blut hier nur die 30- entsprechende Saueretoff- menge von 11 Volumproz. aufnehmen. Wie stark sich das Blut wiihrend der Passage darch die Lunge ' diesem Siittigungsgrade niihert, iibt natiirlich einen nicht geringen Einfld3 auf die Werte des mittleren Druckes. Die untenstehende Tabelle gibt AufschluS tibw deren GrbBe, wenn der Sauerstoffgehalt des Venenblutee von sehr niedrigea Werten (0 und 2 Volumproz.) an wiihrend des Lungenkreislaufs bis auf 8 oder 10 Volumproz. steigt, mithin bis zur SWigung von 73 Proz., bzw. 91 Proz. der vblligen Sttigung.
25.5, bzw. lSmm.
17*

260 CHRISTIAN BOHR:
Mittlere Druckdiffercnz (P + p') fiir P = 30 mm bei verschiedenem Sauerstoffgehalt dee Blutes.
-
Venenblut Volumproz. 0,
Spannung
21.0 32.2 34 46 50 64
Arterienblut Volumproz. 0,
Mittlere Druckdifferens
P i. p'
8 8
10 10
12.2 10.5 7 .9 6 . 9
Da das Venenblut niemals ganzlich sauerstofffrei wird, und da man nicht annehmen darf, daB die Sauerstoffsattigung des Arterien- blutes geringer als 73 Proz. (8 Volumproz.) ist, kann die mittlere Druck- differenz, wenn die Alveoknspannung wihrend der Atmung in ver- diinnter Luft bis auf 30- sinkt, nur auf ca. 10"" angesetzt werden.
Was die Kohlensiiure bet'rifft, so llBt sich auch deren mittlere Druckdifferenz unter' verschiedenen Verhaltnissen selbstredend auf analoge Weise berechnen. Da indes die Spannungskurven (die Relation zwischen aufgelBster Menge und Spannung) hier wahrscheinlich ziemlich schwankend sind, beschranken wir uns darauf, was fiir unseren Zmeck geniigen wird, den mittleren Druck f i r normale Atmung zu berechnen, indem wir die durchschnittliche Kohlensaurespannungskurve zugrunde legen. Fiir die hier in Betracht kommende Strecke der Kurve hat man
~~
Kohlensiiure Volumproz.
35 40 41 45 46 50
Setzt man die Kohlensiiurespannung der Alveolenluft z. B. auf 32 2 mm an, so kann das Arterienblut also nicht weniger als 40Volumproz. CO, enthalten; wir nehmen nun an, daB das Blut mit einem Gehalt von 46 Volumproz. in die Lunge gelangt und dieselbe mit 41 Volumproz. CO, verliil3t. Hieraus kann man nun mittels obenstehender Spannungs- tabelle ganz den friiheren Berechnungen analog die mittlere Druck-
Bohr, N a g e l s Handbuoh. I. I. S. 106.

OBEB DIE SPEZIFISCHE TATIGKEIT DER LUNGE usw. 261
50 46
differenz berechnen, nur bildet man hier die Differenz p' i P statt P i p ' . Der Wert findet sich unten angegeben, wie auch der der Vollstindigkeit wegen berechnete Wert des mittleren Druckes fiir den Fall, daS das Venenblut 50 Volumproz. enthllt.
Mittlere Kohleneiiuredruckdifferenz fur P = 32 - 2 -.
I
41 9.3
41 I 8.1
Mittlere
A bsc h n i t t IIL Messnng der -4rch die Lnngen stattfln,mden 6lasdifPnsion.
Wie oben entwichelt, erfordert eine auoh nur annlhernd genaue Bestimmung der Gasdiffnsion durch die Lunge die Anwendung des d i re k t en Experimentes, in welcbem man an dem lebenden Organismus diejenige Menge eines Gases mist, die bei bekannten Druckdifferenzen aus der Alveolenluft ins Blnt aufgenommen wird. Mit diesem speziellen Zweok vor Augen angestellte Untersnchungen liegen meines Wissens in der Literator nicht vor, wir werden aber im folgenden (§ 1) sehen, wie mehrere teils von Grbhant , teils von Haldane ausgefiihrte Ver- s u c h iiber die Aufnahme des Kohlenoxyds in das Blut unter solchen Bedingangen angestellt worden sind, daS eine Berechnung der Dif€usion des Kohlenoxyds durch die Lunge auf Basis derselben miiglich ist. Bus der 80 gefundenen GriiSe der Diffusion des Kohlenoxyds 1 s t sich darauf die Diffusion der uns zuniiohst interessierenden Gase, des Sauerstoffes and der Kohlensirnre in bekannter Weise bereohnen (siehe unten 6 2).
\
1. Bestimmung der Diffaeion der Kohlenoxydr am den Versaahen von Orbhant und von Haldane.
Wenn man an einem Versuchsindividunm, das eine kohlenoxyd- haltige Gasmischung atmet, diejenige Menge des genannten Gases

262 CHRISTIAN BOHR :
bestimmt, die nach Verlauf eiuer gewissen Zeit ins Blut aufgenommen worden ist, kann man, da das Kohlenoxyd im Organismus nicht um- gesetzt wird l, unter Urnstinden hieraus die DiffusionsgroBe (B) des Kohlenoxyds fur das betreffende Individuum berechnen, oder mit anderen Worten diejenige Nenge Gas, die pro Minute und pro lmm Druckdifferenz bei diesem Individuum aus der Alveolenluft ins Blut dringt. Es miissen hierbei gewisse Versuchsbedingungen erfiillt sein ; teils darf das Blut zu dem Zeitpunkte, wo die Probe behufs der Bestim- mung des Kohlenoxydgehaltes entnommen wird, noch lange nicht mit Kohlenoxyd unter den gegebenen Spnnnungsverhiltnissen gesattigt sein ; teils miissen die Bedingungen solche sein, daB die wahrend der Dauer des Versuches herrschende durchschnittliche Druckdifferenz zwischen der GO-Spannung der Alveolenluft und der des Blutes, oder was wir oben
' (Abschn. 11, 8 2) die mittlere Druckdifferenz nannten, bekannt ist. Eine selbstvershdliche Voraussetzung, fiir deren Richtigkeit vorllufig alles zu sprechen scheint, ist es ferner, dal3 das Kohlenoxyd durch einfache Diffusion ohne das Eingreifen irgend welcher Zellentitigkeit aufgenom- men wird.
Die zu solchen Berechnungen geeigneten, in der Literatur vor- liegenden Versuche wurden ubripens nach verschiedenen Methoden angestellt. Grbhant fiihrt seine Versuche an Hunden aus; das Kohlen- oxyd wird durch Auspumpen des Blutes wiihrend Erwlrmung mit Phosphorslure gewonnen und im Grisoumeter dosiert. Haldane be- nutzt sich selbst als Versuchvobjekt und bestimmt den Kohlenoxyd- gehalt des Blutes mittels seiner kolorimet-rischen Titriermethode. Nichts- destoweniger und obschon die mitt.lere Druckdifferenz bei Haldan e bis lOmal so groB ist wie bei GrBhant, gibt es, wie wir unten sehen werden, eine befriedigende tfbereinst,immung der Resultate, die sich aus den Versuchen der beiden genannten Forscher ableiten lassen.
Die Versuche von Grbhant,' die sich zur Berechnung der GriiSe der Diffusion, eignen, sind solche, wo die Einatmungsluft nur sehr geringe Mengen Kohlenoxyd enthiilt. Hier sind, wie die untenstehende Tabelle zeigt, die wiihrend der Dauer des Versuches ins Blut auf- genommenen Kohlenoxydmengen relativ gering und, was f i r unseren Zweck von grol3er Wiohtigkeit ist, sowohl der Spannung des Kohlen- oxyds in der Einatmnngslnft als namentlich auch bei derselben Span- nung der Zeitdauer viillig proportional.
. Haldane , Journ. of Physiology. 1900. Bd. XXV. S. 225. N. G r 6 h a n t , L'oxyde de carbone. Paris 1903. S . 67. Auch in den
Archives. de PhySiologie (5) . 1898. Bd. X. S. 315.

UBER DIE SPEZIFISCHE TATIGKEIT DEB LUNGE usw. 263
~~
CO-Gehalt Volumproz. CO des Blutes - der 1 Einatmungaluft nach 1 Stunde j nach 2 Stunden
'Is 0 000 1 0.44 0 . 8 8 1/80000 1 0.22 1 0.44
Diese Proportionalitat zeigt, da6 die kleinen ins Blut aufgenommenen Kohlenoxydmengen keine mehare Spannung ausgeiibt haben; sonst kiinnte unmoglich wlhrend doppelt so langer Zeit eine doppelt so gro6e Menge aufgenommen worden sein. Dies bedeutet wieder, da6 der bei der Diffusion treibende Druck, die mittlere Druckdifferen z, in diesem Falle gleich der Kohlenoxydspannung der Alveolenluft ist. Obenstehende Tabelle reprben tiert vier verschiedene Versuche, die, wie aus der er- wahnten Proportionalitit hervorgeht, samtlich dasselbe Resultat geben, und es ist mithin einerlei, welchen derselben man zur Durchfiihrnng der Diffusionsberechnung wahlt. W ir werden hierzn im folgenden den Trersuch verwerten, wo die Einatmungsluft - - 0.00166 Prozent Kohlenoxyd enthielt, wlhrend ins Blnt pro Stunde 0.22- dieses Gases aufgenommen wnrden.
Far 1 kg Hund betrlgt die Blutmenge die 1/13 des Kiirpergewiohtes ausmacht, 7'lcCm; pro Kilogramm wurde also ins Blnt aufgenommen
Diese 100
Menge wurde, wie oben gezeigt, bei einer mittleren DrnckdBerenz gleich der Kohlenoxjdspannnng der Alveolenluft anfgenommen, welche letztere GriiSe sich folgendermaBen annlhernd berechnen 1BBt. Durch direkte Bestimmungen wies G r 6 ha n t nach, daS in seinen Versnchen mit Hunden l/, der eingeatmeten Kohlenoxydmenge aufgenommen wird; in der Ausatmungsluft findet sioh also ein Kohlenoxydgehalt von 4/6
des Gehalts der Einatmungsluft oder in nnserem Falle '/, . 0.00166 = 0.00133 Proz. CO. Bei der Berechnnng der Alveolenluft kann der schiidliche Raum wiihrend der Atmung anniihernd anf des Atem- zuges angesetzt werden; der Sicherheit wegen schlagen wir denselben anf
an, wodurch die berechnete Diffusion ein wenig gr6Ser wird, dooh nur nm ca. 4 Proz. des Werfes. Sind Bnnd J die Prozente des Kohlenxoyds der Bus- bzw. der Einatmungslnft, so wird das Prozent der Alveolenlnft also
; hieraus h d e t man, da8 die Alveolenluft fiir die obengenannten 3 EaJ 2
-. '' 0.22 = 1-694mm CO pro Stunde oder 0.0282 pro Minute.
Werte = 0-00117 P~oz. CO 3 x 0*0013 i. 0.00166 2
A. a. 0. S. 65.

264 CERISTIAN BOHR:
enthalt, was bei einem Totaldruck von 7 10- die Kohlenoxydspannung der Alveolenluft = 0.00831 mm ergibt. Die DiffusionsgriiBe oder die- jenige Menge Kohlenoxyd, die pro 1 Minute und 1- Spannung von lkg Hund aufgenommen wird, erweist sich also als
In Haldanes Versuchen, die er an sich selbst anstellte, ist der Kohlenoxydgehalt der eingeatmeten Luft durchweg bedeutend groBer als in den soehen besprochenen Versuchen Gr6 han ts. Selbstverstindlich sind zur Berechnung der DiffusionsgroBe nur diejenigen Versuche von Haldane brauchbar, wo das Blut For der Probeentnahme noch weit von der den gegebenen Spannungen entsprechenden Sattigung mit Kohlenoxyd ist. Die von Haldane und Lorra in S m i t h in ihrer Arbeit iiber die Sauerstoffspannungen des Blutes veriiffentlichten Ver- suche, wo bis zur relativen Sattigung des Blutes mit Kohlenoxyd ge- atmet wird, kommen in diesem Zusammenhang deswegen nicht in Betracht, ebensowenig wie die daraus gewonnenen Resultate. Dagegen finden sich in einer friiheren Abhandlung von Haldanel Versuche, die eine Dflusionsberechnung gestatten, indem in diesen Proben des Blutes entnommen wurden zu einem Zeitpunkt, wo dasselbe noch weit von der SIttigung entfernt ist. In diesen Versuchen erfolgten die Bestimmungen des Sittigungsgrades mittels der kolorimetrischen Yethode, die sich als gute Resultate gebend erwies, wenn man die Einwirkung des Lichtes auf die Tritrierung beriicksichtigt. (Der Einfld3 des Lichtes auf die Spannungskurve des Kohlenoxyds ist dagegen, wie am dem Folgenden hervorgehen wird, fiir unsere Berechnungen 'ohne Belang.) In der zitierten Abhandlung wird angegeben, wie hoch die AtemgriiSe durohschnittlich war, namlich 7 1 pro Minute, wlhrend die GrBBe eines Atemzuges 08. 5OOwm betrug. Da der schldliohe h u m sich mit hinlsnglicher Anniiherung auf des Volumens des Atemzuges ansetzen ESt, sind mithin die erforder- liohen Daten zur Bereohnnng der Zusammensetzung der Alveolenluft gegeben, wenn man den prozentigen Gehalt der Ausatmungsluft an Kohlenoxyd kennt. Haldane gibt an, daB in seinen Versuchen mit grober Anniherung die Hilfte des eingeatmeten Kohlenoxyds wiihrend der Atmung ins Blut aufgenommen wird, so daB der Kohlenoxydgehalt der Ausatmungsluft nur halb so groB ist wie der der Einatmungsluft. Da indes in einer spIteren Abhandlunga die totale, im Blute der
' Journal of Physiology. 1895. Bd. XVIII. S. 430. Haldane and Smith, Jmm. of Physiology. 1900. Bd. XXV. S. 331.

OSEE DIE SPEZIFISCHE TATIQKEIT DER LUNGE usw. 265
Versuchsperson enthaltene Sauerstoffmenge mittels der Kohlenoxyd- methode auf 656 oder ca. 66OCcm bestimmt wird (a. a. O., S. 340, Ver- suchsperson J. S. H., Gewicht 78kg), ist es moglich, mit Bezug auf jeden einzelnen Versuch mittels des relativen Siittigungsgrades mit Kohlen- oxyd die gesamte wiihrend der Versuchsdauer ins Blot aufgenommene Kohlenoxy dmenge und hieraus mieder den Kohlenoxydgehalt der Aus- atmungsluft zu bestimmen ; dies gibt natiirlich groBere Genauigkeit als die Anwendung der obengenannten summarischen Angabe iiber die Zusammensetzung der Ausatmungsluft.
Au6er der Kohlenoxydspannung in der Alveolenluft ist bei der Difisionsberechnung ferner der mittlere Druck des Kohlenoxyds des Blutes wiihrend des Versuches in Anschlag zu bringen, da dieser hier nicht wie in Grhhants Versuchen, wo die Kohlenoxydspannungen seh r niedrig waren, als verschwindend b’etrachtet werden kann. Bus diesem Grunde wird es am richtigsten sein, nur solche Versuche zu benutzen, wo die Spannung des Kohlenoxyds im Blnte wegen dessen verhiiltnismii6ig geringen Siittigungsgrades mit diesem Gase absolut genommen so gering ist, da6 miigliche kleinere Unsicherheiten der Spannungsbestimmungen keinen EinfluS von Bedentung auf die Resul- tate erlangen (nlheres hieriiber in der folgenden detaillieren Be- rechnung der Versuche). Da in der Berechnung der mittleren Druck- differenz die Kohlenoxydspannung des Blutes als Subtrahendus vor- kommt, wird man zugleich ersehen, da6 man auch dafiir Sorge zu tragen hat, solche Versuche zur Berechnung zu wiihlen, daB die GriiBe, von welcher snbtrahiert wird (die Spannung der Alveolenluft), doch auch keinen gar zu geringen absoluten Wert erhllt; aus diesem Grunde l w e n sich diejenigen der Versuche (a. a. 0. IX, X, XI), wo der Kohlenoxydgehalt der eingeatmeten Luft sehr gering ist (0.025 bzw. 0-019 und 0-04 Proz.), und wo die Alveolenluft daher nur weniger als 0.01 Proz. CO enthiilt, nicht wohl zu einer Messung der Diffusion gebrauchen ; sie konnen bei der Berechnung freilich iihnliche Zahlen wie die iibrigen ergeben, kleine Schwankungen der verschiedenen Korrektionen spielen hier aber eine so entscheidende Roue, dal3 das Ergebnis in der Tat als durchaus unsicher zu betrachten ist. Wie untenstehende Berechnung zeigt, gilt dies dagegen nicht von den- jenigen Versuchen, wo die Einatmungslnft 0.11 Proz. CO und dariiber enthiilt. Einige dieser Versuche geben indes nicht die zu den Be- rechnungen erforderlichen Daten (a. a 0. I und VIII) oder nur eine einezelne Bestimmnng, die keine Sicherheit bietet, daS die Aufnahme des Kohlenoxyds regelmiiBig verlaufen ist (111). Zuriick stehen also fiinf Versuche (11, IV, V, VI, VII). In den Versuchen 11 nnd V ist

CHRISTIAN BOHR: 266
die Einatmungsluft wesentlich dieselbe (0.35 und 0.37 Proz. CO) und die Sattigung des Blutes nach halbstundigem Atmen ebenfalls wesent- lich iibereinstimmend (Versuch I1 nach 30 l/, Minute 39 Proz. Sattigung; Versuch V nach 29 Minuten 35 Proz. Sattigung); diese Sattigungsgrade sind aus den oben angefiihrten Griinden jedoch schon zu hoch, um sich zur Berechnung benutzen zu lassen, und die zu einem friiheren Zeitpunkte , nach Verlauf von 15 Minuten, in den beiden Versuchen entnommenen Proben stimmen nicht miteinander iiberein , indem der Versuch I1 23 Proz., der Versuch V aber nur 13 Proz. Sgttigung gibt; hier ist wahrscheinlich mit Bezug auf letzteren Versuch, der einen geringeren Wert der Diffusion gibt als samtliche anderen Versuche, ein zufalliger Fehler untergelaufen. Jedenfalls wird es am richtigsten sein, den Versuch V nicht mitzunehmen.
Im folgenden werden wir nun die GroBe der Diffusion auf Basis der Versuche 11, IV, VI und VII berechnen, indem wir simtliche Werte und Korrektionen angeben und das erste Beispiel ausfiihrlicher durchgehen. Das Kohlenoxydprozent der Einatmnngsluft ist der spater von Haldane gegebenen Mitteilung gemziB auf 90 Proz. des ange- fiihrten Wertes reduziert worden, welche Korrektion die berechnete DiffusionsgroBe selbstverstandlich steigert.
Versuch 11 (Haldane, a. a. 0. S. 435). Die Einatmungsluft enthelt 0 .35 Proz. CO; nach Verlauf von 15 %.
ist der Grad der Sgttigung des Blutes 23 Proz. Da die gesamte Sauerstohenge (oder Kohlenoxydmenge), die das
Blut aufzunehmen vermag, wie oben angegeben 660 wm beta%@, so 23 sind w!&rend der 15 ?huten 660 - = 151 '8 oder pro Minute 100
-- l5lS8 - 10.12 cem CO ins Blut aufgenommen worden. 15
Da die Atemgr6Be 71 pro Minute betriigt, so sind in 1 Minute 7000.0.35 = 23.5 wm CO eingeatmet woden, und da ins Blut pro Minute 10.12- aufgenommen wurden, so sind gleichzeitig 23.5 + 10.12 = 13-38 wm ausgeatmet worden. Dies gibt ein Proz. dieses Gases in der Aus- atmungsluft von = 0.191 Proz. CO. Die Alveolenluft ent- h< daher, den schlrdlichen R a m als 1/3 des Atemzuges gerechnet,
Von der prozentigen Spannung der Alveolenluft ist, urn die mittlere Druckdifferenz zu finden, die mittlere Spannung des Kohlen- oxyds im Blute wiihrend des Versuches zu subtrahieren. Denjenigen
100
13.38 * 100 7000
3~0,191 + 0.35 2 = 0.112 Proz. CO.
Haldane und S m i t h , Joum. of Physiology. 1896. Bd. XX. S. 509.

~ B E B DIE SPEZIFISCHE TATIGKEIT DEB LUNGE usw. 267
Wert der GroBe, der hier zugrunde gelegt werden mu8 (die prozentige Spannung des Kohlenoxyds im Blute bei verschiedener SBttigung des letzteren mit dem genannten Gas in einer Atmosphare mit ca. 21 Proz. Sauerstoff), findet man bekanntlich in verschiedener Hohe angegeben,l indem die von Hiifner fur eine gegebene Sattigung gefundenen Spannungen doppelt so groB sind wie die entsprechenden Spannungen in Haldanes Bestimmungen; so gibt fiir 23 Proz. Sattigung eine Interpolation der Hufnerschen Kurve eine Spannung, die 0.044 Proz. CO entspricht, eine Interpolation der Haldaneschen Kurve aber nur 0-022 Proz. CO. In den hier zur Diffusionsberechnung benutzten Versuchen erfolgt nun die Aufnahme des Kohlenoxyds zum Teil, wahrend das Blut in der Lunge noch mehr oder minder veniis kt, und dessen CO- Spannung fiir eine gegebene absorbierte Menge dieses Gases wird daher niedriger, als wenn das gesamte Blut bei dem Drucke der Atmosphhe mit Sauerstoff gesiittigt ist. Im folgenden sehen wir indes hiervon ab, indem dieser Umstand nur bewirken wird, daB man die Diffusion durch unsere Berechnung etwas gr66er findet, was fiir die Anwendung unserer Resultate nur von geringer Bedeutung ist. Aus demselben Grunde benutzen wir iiberall die Htifnersche, nicht aber die Haldanesche Kurve, da erstere die groBere Spannung im Blute, also die geringere mittlere Druckdifferenz und mithin den gr6Beren (im schlieBlichen Resultat bei diesem Versuche etma um 10 Proz. grooeren) Wert der Diffusion pro Minute und Millimeter gibt. Die in dieser Weise gefundenen Kohlenoxydspannungen diirfen selbstverstiindlich indes nicht mit ihrem vollen Betrag abgezogen werden, da die CO-Spannung im Anfang des Versuches 0 ist und erst bei dessen AbschluS den oben angegebenen Wert erreicht; nimmt man als un- gefihren Abzng die Hilfte des Wertes, so erhalt man in unserem hier behandelten Falle aus Hiifners Kurve 0.022 Proz. CO. Um m unter- suchen, ob es in diesem Falle zulissig sei, eben die Hilfte des Wertes abzuziehen, berechnete ich nach der oben (Abschnitt 11, Q 2) fiir die rationelle Berechnung der mittleren DruckdiEerenz angegebenen Methode den Wert von P + p' auf Basis der Hiifnerschen Kurve; ioh fand p'= 0.017 Proz., also fast denselben Wed, zu dem wir oben gelangten.
Wir finden nun die mittlere Dmckdifferenz, indem wir diese GriiBe von dem Kohlenoxydprozent der Alveolenlnft subtrahieren, wo- durch wir 0.112 + 0.017 = 0-095 Proz. CO erhalten. Dies entspricht, da der Gesamtdruck auf 710 mm angesetzt werden kann, einer Spannung (8) von 7100- = 0-674. Die DifFusionsgriiSe (die wir im folgenden 0.095
100
Vgl. Nagels Handbuch. I. I. S. 126.

268 CHRISTIAN BOHW
B nennen) oder die pro Minute und 1 mm Druckdifferenz diffundierende Anzahl Kubikzentimeter ist dann die pro Minute ins Blut aufge- nommene Kohlenoxydmenge, dividiert mit der mittleren Druckdifferenz oder - = 15.02. 10.12
0 - 674
Versuch IV (Haldane , a. a. 0. S. 436). Einatmungsluft 0.32 Proz. CO; nach Verlauf von 18 Minuten ist der
Pro Minute ins Blut aufgenommen 660 ~ = 9.533. Bei 7 1 Atmung pro Minute eingeatmet 23.4, ausgeatmet also
23.4 f 9.53 = 13-87ccm. Folglich in der Ausatmungsluft 0.198 und in der Alveolenluft 0.137 Proz. CO. CO-Spannung im Blute, wie im Versuch I1 berechnet = 0.026; die mittlere Druckdifferenz wird dann
0.137 f 0.026 = 0.111 Proz. CO,
Sittigungsgrad des Blutes 26 Proz. 26
100.18
0.111 100
S = 710- = 0*738mm
und B = - 9.533 = 12.10. 0.788
Versuch VI (Ha ldane , a. a. 0. S. 437). Einatmungsluft 0.108 Proz. CO; nach Verlauf von 15 Minuten ist
der Sllttigungsgrad des Blutes 8 Proz. Pro Minute ins Blut aufgenommen 660 ~ = 3*52ecm. Atem-
gr8Se pro Min. 7 1; eingeatmet pro Min. 7-56, ausgeatmet 7.56 + 3.52 = 4.04 -. Hiervon in der Ausatmungsluft 0.058, in der Alveolenluft 0.033 Proz. CO-Spannung im Blute, wie in Vers. I1 berechnet = 0.002. Mittlere Druckdifferenz = 0.033 + 0.002 = 0.031 Proz. CO.
8 100.15
s = 710 - o'031 0 . 2 2 0 ~ . D = ?!!? = 16.00. 100 0.22
Versuch VII (Haldane , a. a. 0. S. 438). Einatmungsly@ 0.19 Proz. CO; nach Ablauf von 20 Minuten ist der
Pro Minute ins Blut aufgenommen 660 __ = 5.61 "m. Atern- gr8Be 7 1; eingeatmet pro Min. 13-3, ausgeatmet 13.3 f 5-61 = 7-69 ecm. Hiervon in der Ausatmungsluft 0 1 10, in der Alveolenluft 0 - 07 0 Proz. CO-Spannung im Blute wie in Versuch I1 berechnet. = 0*010. Mittlere Druckdifferenz = 0.070 + 0.010 = 0.06 Proz. CO.
SMiguogsgrad des Blutes 17 Proz. 17
100.20
0.06 5.61 8 710 - = 0.426 mm. B = - = 13.17. 100 0.426
Die in den einzelnen Versuchen gefundenen Werte der Diffusions- grBBe B oder der pro Minute und lmm bei einem Manne von 7akg diffundierenden CO - Menge sind in untenstehender Tabelle angefiihrt.

OBER DIE SPEZIFISCHE TATIGKEIT DEB LUNGE usw. 269
I1 IV VI
VII
15-02 12-10 16-00 13-17
Mittel 14.07
Die GrBBe der D h i o n des Kohlenoxyds bei einem Manne von 78 kg erweist sich also als durchschnittlich gleich 14-07c"m. Es wird indes zweckmaBig sein, die GrBBe der Diffusion durch die Lunge nicht mit dem Gewicht,e, sondern mit der GrBBe der iiuBeren Kiirperober- fliiche in Beziehung zu setzen. Da ein Mann von 78 kg nach Vierordt- Meehs Formel eine Oberflache Fun 22476 qem besitzt, erhalten wir die Lungendiffusion, pro Minute und Millimeter fiir 1000qcm M e r e Kiirperoberfliche berechnet, gleich 0 - 63 eem.
Der auf Basis von Grhhants oben beschriebenen Versuchen fiir Hunde gefundene Wert muS des Vergleiches wegen ebenfalls so um- gerechnet werden, da6 er fiir 1000qcm ZinBere Oberfllche gilt, denn fiir die Diffusion durch die Lunge ist ebenso wie fiir den Stoffwechsel nicht das Gewicht, sondern die Kijrperoberfliiche zugrunde zu 1egen.l In Grkhants Versuchen betrug die DiffusionsgrijBe fiir 1 kg Hund 0.3394; das Gewicht des Hundes findet sich nicht angogeben, man dsrf aber sicherlich annehmen, daB derselbe wie die anderen von Grdhant zu diesen Versuchen benutzten Hunde etwa 10 kg wog; seine Oberfliiohe wiirde dann 5700 qcm ausmachen und die GriiBe der Lungendiffusion, pro 1000 qam KBrperoberfllche berechnet, wiirde 5.1 = 0-58 sein, also nahe dem aus Haldan es Versuchen fiir Menschen berechneten ,Werte. War der Hund in Grhhants Versuchen von etwas griil3erem oder kleinerem Gewichte, so wird die Diffusion pro 1000qCm Kiirper- oberfliiche wegen des Verhdtnisses zwischen Gewicht und Oberfiiiche selbstverstindlich etwm grBBer oder kleiner als die angegebene Zahl sein; jedenfalls findet man aber aus Grbhants Versuchen, die von den Ealdaneschen sowohl mit Bezug auf das Versuchstier als auf die analytische Methode und auf den Wert der alveolaren CO-Spannung sehr verschieden sind, eine Zahl, die nioht weit von der aus Haldanes Versuchen bereohneten lie&
Dieee letzteren , wo alle wesentlichen Versuchsbedingungen an- gegeben sind, und die am Menschen unternommen wurden, werden wir im folgenden unseren Betrachtungen zugrunde legen.
.
3.394
* Vgl. Haldane, Jam. of Physiology. 1895. Bd. XVIII. S. 447.

270 CHRISTIAN BOHR:
0 2. Die DiffusionagroBe f ir Sanerstoff und Kohlensanre.
Es ist eine ganz natiirliche Annahme, daB das Kohlenoxyd mittels eines einfachen Diffusionsprozesses ohne Beteiligung irgendwelcher Zellen- tatigkeit in die Lunge aufgenommen wird, und hierfiir spricht auch in hohem Grade, wie von Haldane und Smi th angefuhrt, die Art und Weise, wie dessen allmahliche Aufnahme wahrend der Atmung verlluft; wird eine Luftmischung von einem angemessen geringen Gehalt an Kohlenoxyd eingeatmet, so steigt namlich anfangs der Gehalt des Blutes an diesem Gase fortwihrend; nach Verlauf einer gewissen Zeit wird die Menge des Kohlenoxyds im Blute aber konstant, und der Grad der Slttigung mit Kohlenoxyd, den das Blut somit erreicht hat, steht in einem regelmiiBigen Verhiltnisse zur Spannung des Kohlenoxyds in der Einatmungsluft. Dieser Verlauf der Aufnahme des Kohlenoxy ds durch die Lunge spricht selbstverstandlich in hohem Grade dafiir, daB einfache Diffusion der mabgebende Faktor ist. Ob das Kohlenoxyd hierbei durch die gesam te Lungenoberflsiche diffundiert, oder ob vielleicht nur gewisse Teile der Gewebe und Zellen das Gas durchlassen, andere Teile aber nicht, wird auf diesem Wege natiirlich nicht aufgeklirt; wie es sich nun aber auch hiermit verhalten mag, so ist es doch fir die wahr- scheinlichste Annahme zu halten, daB diejenigen Stellen, an denen die Diffusion des Kohlenoxyds geschehen kann, auch fiir andere Gase, speziell fiir den 8auerstoff und die Kohlenslure, passabel sind. Absolut sicher ist dies allerdings nicht; es wke ja denkbar, daS die letzt- genannten Gase, die beim Respirationsprozesse eine besondere Rolle spielen, in irgendeiner Weise gebunden wiirden , wo das Kohlenoxyd freien Durchgang hsitte. Vorllnfig liegt jedoch nichts vor, was hierfiir sprechen kiinnte, und im folgenden gehen wir deshalb davon aus, dab alle Gase bei ihrer Diffusion durch die Gewehe der Lunge gleich- artigen Bedingungen nnterworfen sind.
In dieaem Falle kann man ans der oben gefundenen Dihsions- grBbe fiir das Kohlenoxyd bekanntlich die Diffusion des Sauerstoffs nnd der Kohlensiure berechnen, indem die Mengen der verschiedenen Gase, die unter sonst gleichen Bedingungen diffindieren, sich direkt wie die Absorptionskoeffizienten und umgekehrt wie die Quadratwurzeln des spezifischen Gewichtes verhalten. Der Absorptionskoeffizient 1 bei 37 O
fiir die Aufnahme in Wasser ist fiir das Kohlenoxyd 0.0184, fiir den Sanerstoff 0.0242 nnd fiir die Kohlensanre 0.588. Mit aller Wahr-
* Vgl. Winkler, Zeitschr. f. physik. C h b . 1892. Bd. IX. S. 171. - Bohr und Bock, Wiedemanns Annalen. 1891:Bd. XLIV. S. 318. - Bohr , Wiedemanns Annalm. 1899. Bd. LXVIII. S. 504.

~ B E R DIE SPEZIFISCHE TATIGKEIT DER LUNGE usw. 27 1
80 70 60 50
scheinlichkeit sind die Verhal tnisse der Absorptionskoeffizienten fiir das Lungengewebe und fiir Wasser identisch; die GrBBe der Diffusion durch die Lunge pro Minute und lmm Druck wird dam, fiir 1000qcm Kiirperoberflache berechnet, indem sie fiir das Kohlenoxyd = 0.63 gefunden wurde,
0 -0242 0.0184 *
fiir den Sauerstoff 0.63 ~- = 0.77,
22859 17.6 354.7 20939 16.1 324 * 9 18895 14.6 300 * 0 16740 12.9 259 * 8
fiir die Kohlenslure 0.63 0*568 ' = 15.52. 0.0184 p 4
Mittels dieser Zahlen ist in untenstehender Tabelle die Diffusions- gr68e bei Menschen von verschiedenem Gewicht sowohl fiir Sauerstoff als fiir Kohlenslure berechnet worden.
Diffusion durch die gesamte Lunge pro Minute und Imm
Sauerstoff 1 Kohlenslure
Gewicht
kg

272 CHRISTIAN BOHR:
fernere direkte Versuche uber das Verhalten und die moglichen Varia- tionen der Lungendiffusion angestellt wurden, und zu solchen Enter- suchungen hoffe ich spater einen Beitrag leisten zu konnen.
A bsc hni t t IV. Uber den Gaswechsel durch die Lwngen.
Im AnschluS daran, was in den vorigen Abschnitten teils iiber die Zellentiitigkeit der Lunge, teils uber die Gro6e der Diffusion durch dieses Organ entwickelt wurde, Ferden wir im folgenden in Kurze die Resultate besprechen, die sich hieraus mit Bezug auf den Gaswechsel der Lunge ableiten lassen.
3 1. Aueecheidnng der Kohlensanre. Die Ausscheidung von Kohlenslure geschieht unter Mitwirkung
einer Zellentiitigkeit ; nur wenn man ganz willkurlich von Resnltaten absehen wollte, die auf experimentellem Wege nach einer Methode gewonnen wurden, gegen welche keine Einwiirfe erhoben worden sind und wahrscheinlich auch nicht erhoben werden kiinnen, lie6e sich die Bedeutung der Zellentiitigkeit fur die Kohlensiiureansscheidung bestreiten. Ich werde in dieser Beziehung beispielsweise einen der Versuche aus einer friiheren Untersuchungsreihe anfiihren. Ein 15.7 kg wiegender Hund atmete mit natiirlicher Respiration und ohne daS andere Ein- griffe als die f i r den Respirationsversuch und die Blntentnahme er- forderlichen unternommen worden wiiren ; die Einatmungslnft war atmo- spharische Luft und es wurden pro Ki logram and Stnnde 363com Kohlensiiure abgegeben ; die Kohlensiiurespannungen in der Alveolenluft, im Blute an8 dem rechten Herzen (m folgenden als Venenblut be- zeichnet) und im Arterienblnte wurden gleichzeitig bestimmt, die beiden letzteren mittels des Kroghschen Tonometers; es ergab sich, daS die Spannung in der Alveolenluft 25.9 "", im Venenblute ebenfalls 25.9 mm nnd im Arterienblnte 16.8- betrug. Eine Stnnde spiiter, wiihrend die Einatmungslnft noch immer kohlensiiurefrei war, betrug die Span- nung in der Alveolenluft 24-l-, im Arterienblute nur 9.3"" CO,. Die Kohlensiiureausscheidung kann hier augenscheinlich nnmiiglich ohne ein Eingreifen der Lungenzellen erfolgt sein, da die Spannung des Venenblutes nicht hiiher, die des Arterienblutes aber niedriger ist als die Spannnng der Alveolenlnft. In einem anderen Versuche aus der- selben Reihe liegt die Kohlensiiurespannung der Alveolenluft z wisc hen
Bohr, ZatraZbZ. f i Physiol. 1907. Bd. XXI. S. 369. Versuch 11.

OBER DIE SPEZIFISCHE TATIGKEIT DER LUNGE usw. 273
der Spannung des Venen- und der des Arterienblutes; auch hier muB die Zellentiit#igkeit sich geltend gemacht haben. DaB in anderen Versuchen die Spannung im Arterienblute hijher a19 in der Alveolenluft gefunden wurde, zeigt uns nur, da6, wie bereits erwahnt, das Essentielle des Pro- zesses nicht die Herstellung einer konstanten Kohlenslurespannung im Arterienblute, sondern wohl vielmehr eine Anpassung samtlicher Ver- hlltnisse an die augenblicklichen Fordernngen des Organismus ist.
Der spezifische EinfluB der Lunge auf das sie durchstriimende Blut ist also eine experimentell festgestellte Tatsache, Ton welcher man bei der Erkllrung der Ausscheidung der Kohlensaure nicht ab- sehen kann, und welche die Entfernung dieses Gases aus dem Blute den eigentlichen Sekret ionen an die Seite stellt, insofern man hierunter e inen du rch Zellenti i t igkeit i n der Driise selbst regul ie r ten A bson der un gsv o r gan g versteht.
Eine hierbei stattfindende hde rung der Spannung wird nach den iiber die spezifische Kohlensaurehaltigkeit des Hlmochroms vorliegenden Tatsachen ohne Schwierigkeit als eine durch die Zellentiitigkeit be- werkstelligte Anderung des genannten Farbstoffes denkbar sein, durch welche die Spannung dersel ben Menge absorbierter Kohlensiiure sich lndert; auch eine Anderung der Alkaleszenz des Blutes kann hierbei in Betraoht kommen.
Es entsteht nun die Frage (vgl. Abschnitt I, 8 2), ob man dafiir zu halkn hat, da6 die Zellentitigkeit auf eine solche Reguliernng der Spannungen beschrjnkt ist, welche hinliingliche Bedingungen fiir die Diffusion des Gases aus dem Blute in die Alveolenluft erzeugt, oder ob es zur Erkliirung der Erscheinungen notwendig ist, eine mit Hilfe der Epithelzellen bewerkstelligte dire k t e Befijrderung der Kohlendure durch die Alveolarwand anzunehmen. Dies ist der Punkt, wo die Bestimmungen der quantitatiren Verhlltnisse der Diffusion Bedeutung erhalten. Legen wir nun die im 0 2 des vorhergeheiden Abschnitteg auf Basis der Haldaneschen Versuche berechneten Werte der Diffusions- gro6e zugrunde, denen gem56 die Diffusion bei einem Mensohen von 7 0 4 325wm pro Xinute und Millimeter betriigt, 80 when Wir, d8S die wBhrend der Ruhe durch die Lunge ansgeschiedene Menge Eohlen- siiure bei einer Druckdifferenz von ca. 1- durch die Lunge Mundieren kann. Die wihrend einer exzessiven Arbeit produzierk 10 ma1 so groh Menge Kohlensiiure wird folglich bei einer mittleren Druckdifferenz von 08. 10- dzundieren k8nnen.
Den vorliegenden Versuchen mfolge kann 88 keinem Zweifel unter-
1 Bohr, Dies Archiv. 1891. Bd.II1. S.66. Skandln. 15rchir. XW. 18

214 CHRISTIAN BOHR:
liegen, daS die spezifische Tltigkeit der Lunge Spannungsvariationen zu bewirken vermag, die die genannten 10"" nicht unerheblich iiber- schreiten, und es liegt deshalb keine Notwendigkeit fur die Annahme vor, dab der Spannungsregulierung iiebengeordnet eine direkte Be- forderung der Kohlenslure mittels der Epithelzellen stattfinden sollte; am nachsten liegt wohl die Annahme, daS die Zellentltigkeit bei der Kohlensaureausscheidung wesentlich darin besteht, die Spannung so einzustellen, daB das . Gas durch Diffusion die Lungenwand passieren kann; in diesem Falle hat man aber notgedrungen anzunehmen, daS die nicht selten in den Versuchen gefundene niedrige Kohlenslure- spannung des Arterienblutes, welche offenbar die Kohlensaureabgabe in den Geweben des Korpers erleichtert, erst dann erzeugt wird, wenn dils Blut im Begriffe steht, die Lunge zu verlassen.
Man hat sich daher zu denken, daB die Kohlensaurespannung im Blute wlhrend dessen Passage durch die Lungen von der Zellentatig- keit reguliert wird und hierbei, wie es im Wesen der Regulierung liegt, urn eine gegebene Mittellage auf- und abwarts schwingt, und es liegt nahe, in den von Henr iques l nachgewiesenen rhythmischen Schwankungen der KohlensBureausscheidung gerade einen Ausdruck der Spannungsregulierung zu erblicken. Wiire die mittlere Druck- Merenz, um welche sich diese Spannungsregulierung hewegt, dieselbe, die wir durch Berechnung aus der normalen durchschnittlichen Kohlen- slurespannungskurve finden (Abschnitt 11, 0 2), also ein wenig iiber 6 mm, so wiirde bei einem Manne von 70 b! die Ausscheidung duch Diffusion (vgl. Tab. S. 271) gleich 325 x 6 oder ca. 20OOwm Kohlensinre pro Mi- nute sein, also bedeutend mehr, als unter gew6hnlichen Verhiltnissen tatsichlich dnrch die Lunge ausgeschieden wird. Nimmt man nun an, da6 die Xohlensiiure durch Diffusion die Alveolarwand durchdringt, so muS, indem man von einer Regulation der Spannung absieht, die vijllige Ausgleichung zwischen den Kohlensiinrespannungen im Blute und in der Alveolenluft also schon sehr bald nach Eintritt des Blutes iu die Lunge stattfinden, nnd das Blut daher immer mit einer C0,-Spannung, die derjenigen der Alveolenlnft vollig gleichliommt, die Lunge verlassen. Die Kohlensiinrespannung des Arterienblutes muS daher unter dieser Vor- aussetzung immer genan gleich derjenigen der Alveolenluft gefunden werden ; wo eine aderhalb der Fehlergrenzen liegende Abweiohnng dieser beiden Spannungswerten voneinander sich zeigt, mnf3 daher immer eine durch spezifische Lungenwirksamkeit im Blute hervorgerufene Spannungslnderung angenommen werden, und dies ebensowohl, wenn
* IJcnriquea, DiaP Arahdo. 1892. Bd. IV. S. 206.

OBER DIE SPEZIFISCHE TATIGKEIT DEB. LUNGE usw. 275
die Arterienblutspannung nach oben als wenn sie nach un ten von der Alveolenspannung abweicht. Eben die im Verhiiltnisse zur Kohlenslure- ausscheidung so bedeutende Diffusionsgrobe dieses Gases macht diese SchluBfolgerung zwingend, imd um so mehr natiirlich ein je gr6Berer Wert fiir die Diffusion der Kohlensiure angenommen wird. Gewohnlich hat man nur diejenigen Werte der C0,-Spannung des Arterienblutes, als fiir eine Zellenwirksamkeit beweisend angesehen, die niedriger als die gleichzeitige Spannung der Alveolenluft waren ; nimmt man die GroBe der Diffusion in Betraoht, sieht man aber, daB auch solche Fiille, wo die C0,-Spannung in hinllnglichem Grade im Arterienblute hiiher als in der Alveolenluft sind, eine spezihhe Wirksamkeit der Lungen anzeigen. Hierdurch tritt ein solches aktives Moment bei fast siimtlichen bis jetzt vorliegenden Versuchen hervor, indem diejenigen Fiille, in welchen die C0,-Spannung des Arterienblutes mit derjenigen der Alveolenluft annlherungsweise ubereinstimmt, verhiltnismi6ig selten sind.
Die quant,itativen Verhaltnisse der Diffusion, mit der Menge der tatsiichlich pro Minute durch die Lunge ausgeschiedenen Kohlensanre zusammengehalten, zeigen uns a160, daB die durch den direkten tono- metrischen Versuch nachgewiesene Zellenmirksamkeit eine bei der Kohlenslureausscheidung auch unter gewohnlichen Verhaltnissen fort- wiihrend eingreifende Funktion ist.
0 2. Anfnahme des Sanerstoffea. Ebensowohl wie mit Bezug auf die Kohlenslureausscheidiing hat
man aucli hinsichtlich der Sauerstoffaufnahme eine Beteiligung der Lungenzellen nachgewiesen. So finden sich in einer friiher Ton mir veriiffentlichten Untersuchungsreihe a mehrere Versuche, wo die Sauer- stoffspannung des Arterienblutes hijher als die der Alveolenluft ist, und wo man deshalb eine spezifische Lungentltigkeit anznnehmen hat. Anders lassen sich wohl auch solche Versuche nicht erkliiren, wo, wie in einigen Flllen bei Zuntz und Hagemann , 3 die $auerstoffmenge des dem Tiere entzogenen Blutes grii6er befunden murde a19 die Sauer- stoffmenge desselben Blutes, naohdem dieses au6erhalb des Organismus mi t atmosphirischer Luff geschiittelt worden war; die Sauerstoffspannnng mu6 hier im Arlerienblute gyOI3er gewesen sein ala in der atmol sphlrisclien Luft; mithin ills0 auch grii6er als in der Alveolenluft. Aber
* Vgl. die Werte der CO,-Spannnngen bei B o h r , Dies Arcbiv. 1891.
* Dies Archie. 1890. Bd. 11. S. 256. Bd. 11. S. 263 u. 264; zsntralbl. 'f. Physiul. 1907. Bd. XXI. S. 367.
8toffweohsd a h Pf4rde.q. Berlin.1898. s. 402. 18*

276 CHRISTIAN BOHE:
Aufgenommene Sauerstoffmenge
I CfasrOeehSs~ und hhrg tk tmmtx des Radfahrers. Berlin 1899. 6. 26-27.

~ B E R DIE SPEZIFISCHE TATIQKEIT DEE LUNGE usw. 277
,, mittleres ,, 8 ,, schnellea :, 2
Ruhe Gang; langsamea Tempo
Sauerstoffgehalt des Venenblutes auf 2 Volumproz. ansetzen. An der Hand der Tabelle iiber die mittlere Druckdifferenz S. 257 erhalten wir unter diesen Voraussetzungen:
19 53 853
19 60 966 19 68 1095
Vergleicht man diese Zahlen mit der von Zuntz in seinen Geh- versuchen gefnndenen Sauerstoffaufnahme, so sieht man, daS, wiihrend die diffundierte Sauerstoffmenge den Sauerstoffverbrauch bei Ruhe nnd langsamen Gehen zn decken vermag, dies dagegen nicht der Pall ist, wenn der Sauerstoffverbrauch bei stjrkerer Arbeit noch ferner steigt. Schon beim Gehen in mittlerem Tempo ist die berechnete dnrchdiffun- dierte Sauerstoffmenge geringer als die wirklich aufgenommene, indem erstere ca. 70 Proz. der letzteren betriigt; beim schnellen Tempo tritt dieses Verhalten weit stirker hervor, indem die berechnete Sauerstoff- diffusion nur ca. 43 Proz. der aufgenommenen Sanerstoffmenge ausmacht.
Unter Voraussetzung einer DiffusionsgriiSe wie die zugrunde gelegte muS deshalb, in diesen Fillen wenigstens, eine Sauerstoffanfnahme unter Mitwirkung der Lnngenzel len stattgefunden haben, nnd es erhebt sich nun wieder, ebenso wie betreffv der Kohlensiinresekretion die Frage, ob man anzunehmen hat, daS diese Zellentitigkeit nur in einer Ein- stellung der Sauerstoffspannung des Blutes anf Werte besteht, die eine hinllingliche Diffusion aus der Alveolenluft ins Blut ermbglichen, oder ob eine direkte Befirderung des Sauerstoffv durch die Alveolenwand erfolgt ist. Eine Wirkung der Lungenzellen, mittels deren die Span- nungen reguliert werden, ist an und fiir sich als eine naheliegende Annahme zu hetrachten, seitdem die Verinderlichkeit des Hbochroms sowohl in betreff des spezifischen Sauerstoffgehalts als 8uch in betreff der Form der Spannungskurve nachgewiesen und die Bedentnng diems Verhaltene fiir die Regnlierung der Sanerstoffipannungen im Plasma entwickelt ist.' Die kage ist aber die, ob die Wirkung eher solahen Spannnngsreguliernng nicht gar zu begrenet ist, um hieraus die ganze Erscheinung zu erkken; 88 whe niisllich wohl denkbar, daS die
N a g e l e Hadbuch. L I. S. 196.

278 CHRISTIAN BOHIL:
Sauerstoffspannung des Blutes durch eine Regulierung wahrend der Stromung des Blutes durch die Lunge verschwindend klein gemacht werden kiinnte, unter Null kann sie selbstverstandlich aber niemals sinken. Die au6erste marimale Grenze der Wirkung einer Spannungs- regulierung erreicht man daher, wenn die Sauerstoffspannung des Blutes auf Null erhalten wird; dann wirkt die ganze Sauerstoffspannung der Alveolenluft als mittlere Druckdifferenz. Betrachten wir nun von diesem Gesichtspunkte aus den obengenannten Versuch mit schnellem Gehen, so betragt die Sauerstoffspannung der Alveolenluft hier 92 Die griiSte lllenge Sauerstoff, die durch Diffusion aufgenommen werden kann, wenn man nnnimmt, da6 die Zellentatigkeit in der Lunge die Sauerstoffspannung des Blutes wllirend der Passage desselben durch das Organ auf Null erhalt, ist dann 16.1 x 92 = 148lCCm pro Minute; in \Virklichkeit ist aber urn bedeutend mehr aufgenommen worden, nrimlich 2652ccm.
Es erweist sich also, daB selbst, wenn die Sauerstoffspannung dcs Blutes wlhrend der Passage desselbeii durch die Lunge trotz der statt- gehabten Sauerstoffaufnahme nicht zugenommen haben sollte, sondern durch eine Zellentiitigkeit auf dem niedrigen Werte Kull erhalten worden ist, nicht einmal hierdurch eine Druckdifferenz erzeugt murde, die hin- lringlich gr06 war, um die gesamte Menge des wirklich aufgenommenen Sauerstoffs mittels eines Diffusionsprozesses aus der Alveolenluft ins Blut iibergehen zu lassen. Es wird deshalb notwendig, anzunehmen, daS der Sauerstoff, zum Teil wenigstens, in irgendeiner Weise durch Zellenarbeit d i re k t durch die Alveolarwand befijrdert wird.
Zu ganz analogen Resultaten \vie den fur die Sauerstoffaufnahme wiihrend der Ruhe und der Arbeit bei gemiihnlichem atmosphlrischen Druck gefundenen gelangen wir, wenn wir die Verhiiltnhe wiibrend der Atmung in verdi innter Luf t mit der daraus folgenden Herab- setzung der Sauerstoffspannung in der Alveolenluft ontersuchen. Als Beispiel benutzen wir einen der in dem groBen Werke von Zuntz, Loewy, Miiller und Caspari (Hohenklima und Bergwanderung. Berlin 1906) angefiihrten Versuche. Die Versuchsperson (Kolmer, Gcwicht 65-72b) nahm wiihrend des Aufenthaltes auf dem Monte Rosa (Borometerdruok 08. 450mm) bei Ruhe 293ccm Sauerstoff pro Minute auf; hierbei war .die 0,-Spannung in der Alveolenluft 46 mm.
Wrihrend der Arbeit war bei einer Spannung der Alveolenluft von 57 m m die Menge aufgenommenen Sauerstoffs 1520 pro Minute. Bei der Berechnung der unter dieaen Bedingungen stattiindenden Sauerstoff-
A. a. 0. Tthelle XI P. XVIIJ. Vemuch v. 3.-S./IX. 1901.

~ B E R DIE SPEZIFISCHE TATIQKEIT DER LUNQE usw. 279
diffusion setzen mir die GroBe der Diffusion pro Minute und Millimeter bei dem gegen 70kg miegenden Manne auf 16“”” an. Die mittlere Druckdifferenz wiihrend des Ruheversuches lHBt sich aus der S. 259 fur 50 mm Alreolarspannung berechneten Tabelle ableiten. Rechnen wir, was die groI3ere mittlere Druckdifferenz gibt, daB das Venenblut besonders sauerstoffarm ist und nur 2 Volumproz. Saiierstoff en thllf, so ergibt die Tabelle eine Druokdifferenz von 18“”, mithin eine Dif- fusion pro Ninute von 16 x 18 = 288ccm, also fast ebensoviel, mie tat- sachlich aufgenommen wurde. Wlhrend der Arbeit betriigt die Sauer- stoffspmnung der Alreolenluft 57 mm (der Sauerstoffspannungskurre zu- folge einer maximalen Sauerstoffaufnahme ins Blut von 17.7 Volumproz. Sauerstoff entsprechend). Indem man fiir diese Alveolenlnftspannung ganz in derselben Weise wie oben (Abschnitt 11, 6 2) die mittlere Druck- differenz berechnet, erhllt man bei stiirkster Ausnutzung des Sauerstoffs im Blute (Venenblut 2 Volumproz., Arterienblut 17 Volumproz.) eine Druckdifferenz von 23 ””; folglich eine Diffusion von 16 x 23 = 368 - pro Minute. Nimmt man an, um die fiir die Differenz giinstigsten Voraussetzungen zu wiihlen , daB die Sauerstoffdttigung des Arterien- blutes nur 90 Proz. begrug, so erhalt man (Venenbliit 2 Volumproz, Arlerienblut 16 Volumproz.) eine mittlere Druckdifferenz = 28 mm und eine Diffusion pro Minute von 16 x 28 = 448 -. In Wirkliehkeit be- tr lgt aber, wie oben angefiihrt, die aufgenommene Sauerstoffmenge 1520 eem pro Minute. Bei der Sauerstoffaufnahme mu6 die Zellentiitig- keit daher eine vorwiegende Rolle gespielt haben. Priift man, ob diese Zellcntiitigkeit lediglich in einer Spannungsregulierung bestanden haben kann, und nimmt man an, um das Maximum der Wirkung einer solchen Regulierung zu erhalten, da6 die Sauerstoffspannung des Blutes wiihrend der Passage durch die Lunge fortwlhrend bis auf Null herabgedriickt iat, so wird die Druckdifferenz 57mm und die Menge, die diffundieren kann, also 16 x 57 = 912”“ pro Minute. Pa die wirklich aufgenommene Sauerstoffmenge 1520mm betriigtj miissen somit wenigstens 1520 P 912 = 6OSCCm oder 40 Proz. der gesamkn aufgenommenen Menge mittel8 direliter Wirknng der Zellen durch die Alveolarwand geschaf€t wor- den sein.
Schlie6lich betrachten wir noch in Eiirze die VwMtnisse wihrend der n i e d rigs t e n mi t Wohlbefinden vereinbarens auer s t o f f 8 p ann un g e n d e r A lveo len lu f t In dem oben zitierten Werkel findet sich ein Versuch mit Atmung in einem pneumatischen Kabinette (Druok 399-),
f Zuntz, Loewy, MUller und Csepsri, a. a. 0. Tebelle XVI. Ver~nch v. 11./vII. 1901.

280 CERISTIAN ROHR: DEER DIE SPEZIF. TATIGKEIT DER T , U N ~ E usw.
!TO die Versuchsperson (Waldenburg, Cewicht 57 kg) bei einer alreolaren Sauerstoffspnnnung yon 32"" 342"" Sauerstoff aufnimmt ; es wird an- gegeben, daB wlhrend des Versuches WohlbeEnden herrschte, gegen Ende aber Mudiglieit eintrat. Die mittlere Druckdifferenz fir eine Alveolen- luft mit 30"" O,-Spannung ist a,uf ca. 10"" anzusetzen, oder, fdlls man anuimmt, daB das die Lunge verlassende Blut nicht die der 30 mm Spannung entsprechenden 11, sondern nur 8 Volumproz. Sauer- stoff enthalt, auf hBchstens 12"" (siehe S. 260). Rechnen wir nun wegen der etwas hoheren Spannung der Alveolenluft (32"") die mittlere Druckdifferenz als 13mm, so ist dies ein Naxiuium. Die Diffusion pro Minute und Millimeter betragt bei einem Nanne von 57 kg 14ccm. Hieraus ergibt sich, daB die Diffusion 14 x 13 = 182"cm pro Ninute ist. Die wirklich aufgenommene Saueritoffmenge war, mie oben erwlhnt, 342"" pro Minute; man sieht, daB bei so niedrigen Sauerstoff- spannungen die Unzullnglichkeit der Diffusion auch in Ruherersuchen hervortritt.
Sowohl die Kohlenslureausscheidung als die Sauerstoffaufnahme miissen daher als sekretorische Vorglnge betrachtet werden, die durch Zellentiitigkeit der Lunge reguliert und nicht durch die Zusammen- setzung der Alveolenluft und die Zusammensetzung des zur Lunge strijmenden Blutes a l l e i n bestimmt werden. Eine mesentliche Seite dieser Regulierung ist in einer Einwirkung auf die Gasspannungen des die Lunge durchstrijmenden Blutes zu suchen, und was die Kohlensiure- ausscheidung betrifft, wird man sogar dahin gebrach t, eine solche Ein- stellung der Spannung fiir das Hauptmoment der Sekretion zu halten. Wenn aber die aus H a l d a n e s und G r b h a n t s Versuchen rtbgeleiteten Diffusionswerte auch nnr annlhernd richtig sind - und es scheiot mir kein Grnnd vorhsnden, dies zu bezweifeln, obschon, wie oben ge- sagt, eine variierte Wiederholung der Versnche, bevor die Sache als endgiiltig entschieden betrachtet wird, sehr erwiinscht wsire -, so muB, was die Sauerstoffanfnahme betrifft, neben der Spannungsregulierung eine direkte, den Sauerstoff befirdernde ZellentPtigkeit eine Rolle spielen, wahrscheinlich dem Prozesse analog, mittels dessen der Sauerstoff in der Schwimmblase sezerniert wird.





























