Embed Size (px)
Citation preview

(Aus dem Pharmakologischen Ins t i tu t der Universit/s P› Ungarn.)
Unte r suchungen liber Muske lkont rak t ion .
VIL MitteilungL
Dis Rolle des Kaliums in der Kontraktion.
Von
E. Ernst und L. Sehef�9
(Eingegangen ara g. Juni 1928.)
Einleitung, Als Endergebnis der vorhergehenden Mitteilung 2 haben wir dis An-
nahme gemaeht, daft infolge der Reizung eine Neubildung von Ionen stattfindet und dadurch 3 kardinale Erseheinungen der Muskelt/s Aktionsstrom, Kontraktion und Volumverminderung hervorgerufen werden. In Anbetracht der wiehtigen Rolle, welehe den neuentstandenen Ionen zugeschrieben wird, dr/~ngt sieh die Frage auf, weleher Muskel- bestandteil diese physiologisch so wiehtige Erscheinung zeigt, also welche Substanz diejenige ist, dis auf den Reiz hin in Ionenform fibergeht und dis genannten kardinalen Tatigkeitserseheinungen des Muskels bedingt. Vor der Beantwortung dieser Frage mus vorerst die normale Zusammen- setzung unseres Versuchsobjektes des Frosehgastrocnemius (Esculenta) betrachtet werden, wie sis in Tab. I a gegeben ist.
Tabelle 1 a. Prozentische Zusammensetzung des Froschmuskels.
20 I 1,o I o,19 I o,16 I o,0~~ I 0,35 Jo,o55 Io:023 Jo, o16
Es w/~re aber nich™ ganz richtig, wenn man ausschlie$1ieh auf Grund der Prozentzahlen auf die funktionelle Wichtigkeit einer Substanz fol gern woll~e, da betreffs der Wirkung eines Stoffes aueh der molaren Ko~�87 zentration bekanntlieh gro6e Bedeutung zukommt. Reehnen wir also obige Daten auf molare Konzentration um, so gewinnen wir ein ganz
1 Ausgefiihrt mit Unterstfitzung des ,,Orsz~gos Term› Alap" (Landesfond �9 ~aturwissenschaften).
Pfliigers Arch. il8, 13�9 (1927). Pfliigers Archiv f. d. ges. Physiol. Bd. 220. 43

656 E. Erns~ und L. Scheffer:
~nderes Bild. Es is t ~us der T~b. 1 b ersichtl ich, d~ft sich in dem Muskel unter allen anderen Bestandteilen Kal~um in der gr6[3ten molaren Konzen- tration be]indet.
Tabelle lb . Die Anzahl Gramm Mole der einze[nen Sto]]e in 1000 g Muskel.
~~~o~~ !Kohle~.ra~ I P I s ! ~ ~ ! ~ ~ l M g l ~ '/,oo ) 1/10o I 1/,o I '/~0 ll/00 I ,/,, I1/~0 I'/,00 1 1/~~0
In dieser Tabelle wurde dem Muskeleiweil~ ein mittleres 1Violekulargewieht von 20000, dem Glykogen ein solches von 1000 zugesohrieben. I)iese Stoffe zer- f•llen zwar und die so entst~ndenenen Zerfallsprodukte bedingen eine gr6Bere molare Konzentration, als in der Tabelle angegeben, das MaB des Zerf~lles und die daduroh bedingte Konzentrationszunahme is~ aber unbekannt. Ein anderer mSglioher Einwand gegen diese Art der tabell~rischen Zusammenstellung k6nnte darin bestehen, daf~ nicht die m olare, sondern die g.quivalente Konzentr~tion in Betr~eht gezogen we�9 miiItte. I)er Vergleich der Aquivalenzwerte wiirde aber einerseits wenig aussagen, wenn man z. B. der Monowlenz des Kaliums die etwa 30--40fache Valenz des Eiweil]es gegeniiberstellt; andererseits wgre eine solehe Zusammenstellung viel zu unsioher, da j~ z. ]3. es fraglioh ist, was fiir eine Valenz dem Element Phosphor gegeniiber dem Kalium zugewiesen werder~ sollte 1 (KH=PO a , K 2 I-IPO~, KaPOa, I�9 In Anbetracht der gesohilderten Schwierigkeiten schien es angebraoht, die molaren Konzentrationen einem Vergleioh zu unterwerfen.
Nich t nur jene Tats~ohe, dag das K a l i u m ira Muskel in der grSSten mola ren Konzent ra~ion vorkommt0, sloricht daffir, dag dem K u l i n m eine wicht ige Rol le in den Tgt igke i t se rsohe inungen des Muske]s zukommt , sondœ auoh die gesamte diesbeziigliche L i t e ra t~ r . So f inden wir, dal~ die A~s ich ten von Bernstein2, Bri~nings a, Overton 4 sohon vor e inem Vie r t e l j ah rhunder t da r in i ibe re ins t immten , dal~ be im Zust~r~dekommen des Ruhe- , Demark~t ions - und Akt ions t romes dem K~l inm die H a u p t - rol le zukommt . J a Macallum 5 geht soweit , daf~ er dem K~l ium eine wiohtige R011e bei dem K o n t r a k t i o n s p r o z e $ zusohreibt .
W e n n w�9 nun ~uf Grund des Gesagten dem K a l i u m eine gewisse Rol le ira Hervorb r ingen der Tgt igke i t se rsehe inungen des Muskels zu- e rkennen wollen, so k6nnen wir in A n b e t r a c h t der l r i iheren Ergebnisse unsere A n n a h m e nur in jener F o r m zum Ausd ruok br ingen, d~G auf den l~eiz hin, K ' - I o n e n en t s t ehen und diese die e rwghnten Tgt igkei t serschei - nungen hervorrufen. I) ie Annahrne l iber die ~Teubildung von K ' - Ionen schlieBt aber die wei tere A n n a h m e in sich, daft da.s K a l i u m ira ruhenden Muskel wenigstens tei lweise in n ich td i ssoz i ie r tem Z u s t a n d vor- handen ist .
1 Vgl. Fi~rth, Erg. Physiol. 17, 459 (1919). 2 P�9 Arch. 92, 542 (1902). 3 Ebenda 100, 391 (1903). 4 Eb› 1105, 185 (1904). 5 Erg. Physiol. 11, 651 (1911).

Un~ersuehungen fiber h�9 VIL 657
Es sind jedenfalls Meinungen l~ut geworden, naeh denen das Kalium nur ira Ionenzustand in der Zelle anwesend w~re, so z. B. die von Osterhautl; Macallum 2 betont, da• es keine ,,maskierten" Kaliumverbindungen gibt. Diesen und anderen ~hnlichen Meinungen kann jedoch entgegengehalten werden, daB die fiberwiegende MehrzahI der Autoren eine nichtdissoziierte Kaliumverbindung in der Zelle an- nimmt. So gibt Loeb 3 seiner Meinung Ausdruck, daB Kalium ira Muskel aus organisehen Verbindungen stetig in Ionen�9 iibergeffihrt wird. Um nur einige weitere Forseher zu nennen, h~ben Macdonald 4, Burridge 5, t~ona und Petow 6 die Existenz einer gebundenen Kaliumverbindung mit Bestimmtheit behauptet. Was ffir eine ehemische N~tur diese nichtdissoziierten Kaliumverbindungen h~ben, ob von einer ,,echten" ehemischen Bindung, wie dies z. B. Robertson 7 annimmt, oder von einer ,,Adsorptionsbindung" mehr die Rede sein kann s, ist hier ganz neben- s~ichlieh, zumal einerseits nach Roncato 9 einige Eigensehaften soleher Verbindungen ffir Adsorption, andere wieder fiir eine ehemisehe Vereinigung zeugen, andererseits mfissen aber diese 2 MSglichkeiten nicht unbedingt als Gegens~tze gelten, be- haupten doch Wedekind und Wilke TM, dal~ in geeigneten F~llen der chemisehen Bindung eine Adsorption vorausgeht.
W e n n nun Netter 11 auf dem S t a n d p u n k t s teht , dal~ , d a s K~l ium ira Inne rn (des Muskels) frei d issozi ier t vo rhanden ist", so da r f v ie l le icht demgegenf iber die Meinung HSbers ~~ ~ngef i ihr t werden, ,,dal~ in den Mus- keln neben e inem �9 Ante i l doch wohl aueh ein gr51~erer Teil der Salze in n ieh td issoz i ie r te r F o r m sein df i r f te" , Zwaardemal~er~a mein t sogar , d~i~ die Menge des , ,Depotkal iums" ira querges t re i f ten Muskel 30ma l grSl~er ~st als das dfffusible Ka l i um.
W e n n wir also annehmen , dal~ das Kal ium oder ein Teil desselben ira
Muskel in nichtdissoziiertem Zustand vorhanden ist, so is t d a t a i t nur die K o n d i t i o -sine- qua -non j ener unserer Ann~hme erffillt , nach we]cher auf den Reiz h in eine Neub i l dung von K ' - I o n e n s t a t t f i nde t . E ine wei tere Grund- l~ge zu unserer A n n a h m e ergeben die Un te r suehungen von Maeallum ~a
und ~nderen Forschern , welehe da r tun , dal~ da8 Ka l ium in den aniso-
1 Ronas Ber. 17, 273 (1923). 2 Erg. Physiol. 7, 605 (1908). 30ppenheimer, Handbuch der Biochemie 2 (I), ]11 (1910). 4 Macallum, Erg. Physiol. 11, 651 (1911). �87 J. of Physiol. 42, 359 (]911). 6 Biochem. Z. 137, 362 (1923).
Erg. Physiol, 10, 335 (1910). 8 Vgl. z. B. Ho]meister, Erg. Physiol. 10, 433 (1910). 9 Ronas ]~er. 31, 329 (1925).
10 Ebenda 34, 11 (1926). 11 Pflfigers Areh. 218, 329 (1927). 13 Physikalisehe Chemie der Zelle und Gewebe 1, 473 (1922). Dasselbe wird
in den Dars~ellungen von Tschermak (Allgemeine Physiologie I [1924]) und von Neuschlosz angenommen (ttandbueh der norm~len und pathologischen Physiologie 8 [I], 132 [1925]).
13 Erg. Physio]. 19, 386 (1921). 1y Erg. Physiol. 11, 636 (1911); siehe Abb. l l und 12a und b der Tafe].
43*

658 E, Ernst und L. Scheffer:
tropen Schichten enthalten isO, d~ diese Fests te l lung zur exper imentel len Pr i i fung unserer A n n a h m e den Weg weist. W e n n ngmlich in den aniso- t ropen Schichten auf den Reiz h in eine gewisse Menge dif�9 K u l i n m
ents teht , so ruf t dies ein K~llum-Konzentr~t ionsgef~l le von den aniso- t ropen Abschn i t t en nach ihren ~ul~eren Milieu hervor. Dieses Konzen- tr~tionsge�9 t re ib t n u n das K a l i u m nach aul~en u n d so k~nn un te r geeigneten Versuchsbedingungen ein K~l iumver lus t des gereizten Muskels e int re ten. I m �9 wollen wir diese Ann~hme ~u�9 ihre Richt igkei t experimentel l priifen.
Methodik.
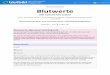
Die zur Kaliumbestimmung verwendeten Gastrocnemien wurden �9 ge- wogen, ira Vakuumexsicc~tor bei Z�9 oder bel 80--90 ~ fiber Schwe- fetsgure getrocknet, nachher wieder gewogen. Trotzdem man in der Literatur manche Hinweise ~uf die Unsicherhdt der trockenen Veraschung �9 bedienten wir uns doch dieser Methode. So beh~upten z. B. Costantino 2 und Dehn a, dag ein Verlust von Alkalisalzen bei der trockenen Veraschung unvermeidlich ist. Die Methode wurde von Stolte scheinbar vergebens verbessert, demi nach t~aab 4 ist ,die j~ stets etwas heikle trockene Veraschung" zu vermeiden 5. Wir haben die trockene Ver~schung ira elektrischen Ofen (Hereus) vorgenommen und diese Methode hat sich wegen der Einfachheit, Bequemlichkeit und Verlgglichkeit glgnzend bewghrt y Es ~~~rde mit 2 Ofen gearbeitet, beide ~fen wurden mit stgdti- schem Wechselstrom gespeist, mit beiden wurde je ein regulierbarer Widerstand in Ser�9 geschaltet und so konnten beide auf eine Temieratur eingestellt werden, bei welcher selbst nach Gliihen fiber eine Woche und noch mehr weder vom KC1, KC1 und NaC1-Gemisch 7 noch vom K1)Oa sein Verlust eintrat. Aus diesen S~lzen wurden die Kationen ebenso wie die Anionen miV einigen l)rozent Fehlern zuriick- gewmmen. Aber nicht nur die anorganischen S~lze, die schon als solche in die Platinschalen eingebrucht und geglfiht wurden, zeigten Konstanz ihres Gewichtes, sondern auch die Asche von Gastrocnemien ergab selbst nach 6t~gigem Glfihen einen normalen t™ Durch ein 6--Sstfindiges Gliihen wird der Muskel vollkommen verascht, ohne dag inzwischen irgendeine Manipulation mit der- selben ausgefiihrt werden mfii]te (siehe die Beispide).
Die Asche wurde gewogen, nach Zugabe von 2--3 Tropfen Eisessig ira Wasser gel6st und der I(aliumgehalt der L6sung nach Kramer-Tisdall bestimmt. Es wurde
1 Die Beweiskraft dieser Befunde wird noch dadurch verstgrkt, dag Woerdeman in den 2l~ttchen des etektrischen Organs ei ne ausgesprochene Lokalisation des Kaliums �9 seiner Meinung nach entsprechend den anisotropen Muskelteilen. R,onas Ber. l , 168 (1921). Vgl. jedoch Costantino, Biochem. Z. 3~, 52 (1911).
2 Biochem. Z. 37, 56 (1911). s Hoppe-Seylers Z. 144, 178 (1925).
Pflfigers Arch. 216, 542 (1927). Vgl. jedoch Egg und Klinke, Biochem. Z. 19|, 439 (1927).
�87 Vgl. Praktikum der physiologischen Chemie 3, 36 (1928) von Knipping und Rona.
7 Vgl. Egg und Klinke, a. ~. O. s Es wurde KI-I2PO ~ abgewogen, welches sich in dea ersten Minuten in dem
Ofen quan~itativ zu I™ a umwandelt (vgl. Schmidt, Pharmazeutische Chemie 1 (I), 366 [1906]).
Untersuchungen tiber l'Viuskelkontraktion. VIL 659
Beispiele zur Methodik.
Aus dem Versuch
~ r .
198
Nr. der Platin- schale
221 a 1
2 L
230a 1
Die ausgegliihte Substanz
Gewicht der Schale + Subs~anz in g
! Ana Anfang [
58,4mgKC1 15,6354 31,0 . . . . 13,3616 53,2 ,, N a C 1 15,6875
-~ 56,0 ,, KC1 I 44,0 ,, N a C 1 13,4326
~- 58,0 ,, KC1 / - - 447,3 , I™ 16,0254 235,3 . . . . ] 13,5659
Asche von 1,25 g - - Muskel
Nach Gliihen von
6Tg. 15,6360 i - - 13,3613i
24St. 15,6875
24 ,, 13,4326 I - -
~ ]5,9655-24S~-. 5 ,, 13,5354 2 ~ , 1-2s~. 15,582~ ~ ™
M
15,9637 13,5350 15,5825
11 i Kaliumgehalt des Muskels nach 6tagigem Gliihen 1,77% und 1,79%
bei allen Versuchen der Bestimmung des Faktors des Reagens, und der Selbstreduk- tion des Hypermangans besondere Achtung geschenkt. Die Methode hat sich in der hier angegebenen Art recht gut bewahrt.
Ergebnlsse.
D a unsere F rages t e l l ung sieh auf die Vergnderung des K a l i u m g e h a l t e s des Muskels rieh~et, so m u g t e -cor a l lem der no rmale K a l i u m g e h a l t des Muskels b e s t i m m t werdœ und zwar umso eher, da sieh aus den vorl iegen- den D a t e n ftir uns eine Uns ieherhe i t e rgibt . K a t z 1 ha~ zwar den K a l i u m - geha l t des f f ischen F rosehmuske l s zu 0,30% Urano 2 und Mitche l l und Wilson ~ dense lben zu 0,34% angegeben, diese Zah len s ind jedoeh als Mi t te lwer te zu betrach~en, und in der l e tz ten Zei~ s ind von Neuschlosz ~
u n d von Raab 5 sogar Kal iumwCrte des fr ischen Muskels mi~getei l t worden, die un t e r e inande r u m 100 und mehr Prozen~ differieren.
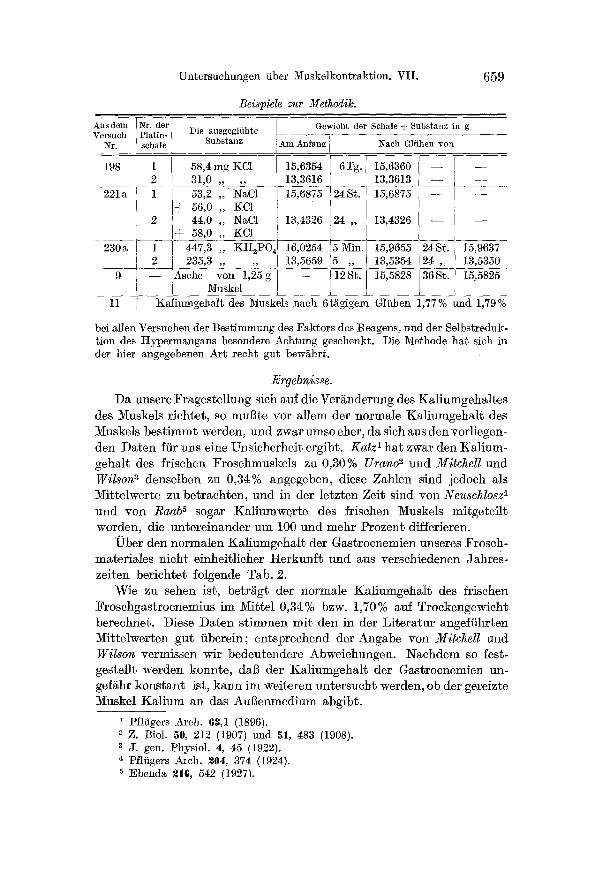
~ b e r den no rma len K a l i u m g e h a l t der Gas t rocnemien unseres Frosch- mate r i a les n ich t e inhei t l icher H e r k u n � 9 und aus verschiedenen Jah res - ze i ten berichte~ �9 Tab. 2.
Wie zu sehen is~, be t r / ig t der no rmale K a l i u m g e h a l t des fr ischen F roschgas t rocnemius ira Mi~tel 0,34% bzw. 1,70% auf Trockengewich t berechnet . Diese D a t e n s t immen m i t den in der L i t e r a t u r ange �9 Mi t t e lwer t en gu t i ibere in ; en t sp rechend der Angabe von Mitchel l und Wil son vermissen wir bedeu tendere Abweichungen . N a c h d e m so �9 ges te l l t werden konnte , dag der Kal iumgehM~ der Gas t rocnemien un- gefiihr k o n s t a n t is~, k a n n ira wei~eren un t e r such t werden, ob der gereizte Muskel K a l i u m an das AuBenmedium abg ib t .
1 Pfltigers Arch. 63,1 (1896). > Z. Biol. 50, 212 (1907) und 51, 483 (1908). a j . gen. Physiol. 4, 45 (1922). a Pfliigers Arch. 204, 374 (1924). 5 Ebenda 216, 542 (1927).
660 E. Ernst und L. Scheffer:
T~belle 2.
Kaliumgehalt der [rischen bzw. ausgetrockneten Gastrocnemien.
Ver- $11ohs-
ri [lrlIIII~‡
I 1
21 31 9
10 11 12 14 16 21 47 48 49 58 59 60 61 63 64 65 74 98
106 114 129 132 134 199 2OO 201 202 203 204 2O5 207 217 220 221 234 243 247
Muskelgewicht in g K-Gehalt der ~Iuskeln
in g frisch getrocknet g ewichtes
rechts I liaks rechts i links rechts I links rer I links
19,42 0,63 2,17 1,19
10,72 37,63 19,59
in % d. Trocken-
1,70 1,73 1,64
1,25 0,84 1,30 0,93 0,92 1,56 1,17
1,09
1,13
1,63
1,80 1,08 1,98 1,44 2,37 1,52 1,65 2,00
1,29 1,03
0,94 1,30 1,01
1,42 0,82 1,32 0,90
1,81
1,65 1,33 1,35 1,95 1,60 1,62 1,70 1,55 1,50
1,28
0,84
0,23 0,17 0,28 0,18 0,22 0,36 0,23 0,20 0,26 0,16 0,25
0,24
0,42 0,22 0,46 0,34 0,50 0,40 0,44 0,47 0,37 0,34
0,22
0,16 0,23 0,19
0,25 0,16 0,28 0,18
0,20 0,20 0,18
0,37
0,33 0,22 0,27 0,44
0,39 0,37 0,32 0,35
0,17
4,54 2,67 4,77 3,12
3,27 3,32
1,65 1,57 1,61 1,63 1,79 1,77 1,66 1,73 1,41 - - 1,44 - - 1,81 1,68 1,63 1,70 1,66 2,00 1,75 1,66 - -
- - 1,65 1,63
- - 1,72 1,03 1,65
0~242 1 ' 5 0 1 o,325~ I - - 1 , 6 8 ]
1,60 I 1,82 I 1,8o I
1,76 - - 1,81 - -
1,82 - - 1,53 - - 1,52 - - 1,58 - - 1,66 - - 1,47 - - 1,65 - - 1,50 - - 0,302 0,322 1,50 - -
1,62 2,00 1,73 1,85
Datum
15. VII. 1926 16. VIL 1926 17. VII. 1926 19. VIL 1926 22. VII. 1926 24. VIL 1926
7. IX. 1926 11. IX. 1926 14. IX. 1926 27. IX. 1926 22. X. 1926 23. X. 1926 23. X. 1926 10. I. 1927 11. I. 1927 11. I. 1927 12. I. 192�9 13. I. 1927 14. I. 1927 17. I. 1927 26. I. 1927 17. i I . 1927 4. I I I . 1927
16. III . 1927 10. VI. 1927 13. VI. 1927 14. VI. 1927 20. XII . 1927 20. XII . 1927 21. XII . 1927 22. XII . 1927 23. XII . 1927 27. XII . 1927 28. XII . 192�9 30. XII . 1927 16. I. 1928 23. I. 1928 24. I. 1928 27. I I . 1928 23. III . 1928 27. III . 1928
1 20 Gastrocnemlen von 10 Fr6schen. Auf Feuchtgewicht gerechnet.
Untersuchungen iiber Muskelkontraktion. VII. 661
Tabelle 3.
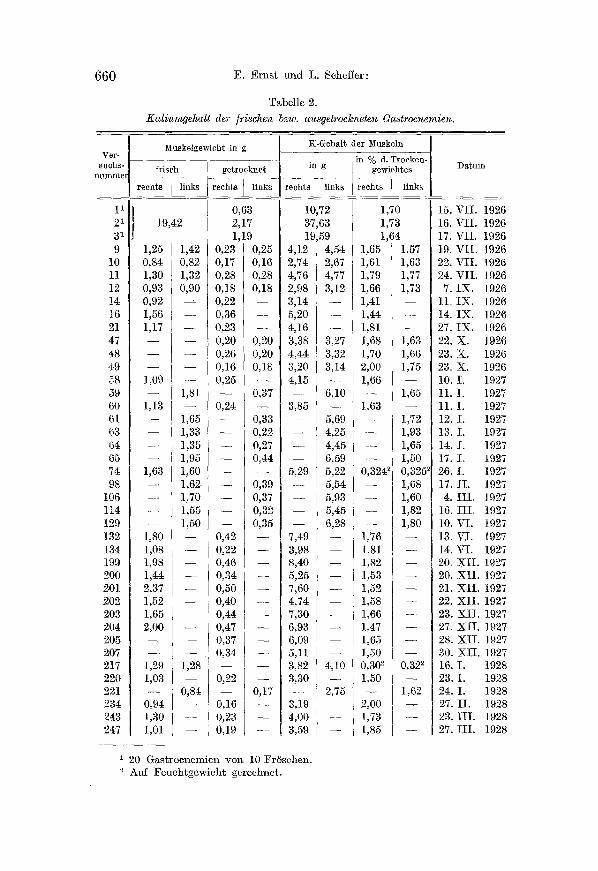
Kaliumgehalt von Gastrocnemien, welche mehrere Stunden lang mit einer Ringer- 15sung vou der Zusammensetzung 6,5promiIL NaC1 ~ O,lpromill. KCl und CaCl2
-~ 0,2promill. NaHCO a durchstrSmt wurden.
Versuchs- nummer
14 15 16 18 19 21 20 25 26 27 28 43 44 46 50 52 54 55 58 59 60 61 63 64 65 90
109 122 125 126 128 151 216 218 219 223 231 232 233
Muskelgewicht in g [~ K-Gehalt der Muskeln
frisch get_rocknet [ in in % C. Trockengew.
rechts links rechts links [ rechts links rech~s links
2,56
1,85 2,13
1,88 2,76
1,50 1,20 1,56 1,15
1,94
2,25 1,18 1,52
0,31
0,35 0,33
0,37
0,33 0,22 0,27 0,43 0,42
0,27
0,32 0,52
0,26 0,19 0,28 0,19
0,22
0,36
0,~3 0,25 0,36 0,26 0,30 0,27 0,30 0,31 0,22 0,29 0,33 0,31 0,37 0,25
0,24
0,3] 0,34
0,37
0,3~ 0,17 0,24
5,94
5,04 5,44
4,97 8,58
3,74 3,09 4,23 3,16
3,07
5,90
4,16 3,98 4,95 4,35 5,02 4,75 5.06
1,91
1,44 1,65
1,5~
1,4~ 1,77 1,6G 1,39 1,49
1,73
1,55 1,65
1,44 1,62 1,51 1,66
1,40
1,64
1,80 1,59 1,40 1,61 1,67 1.76 1,69 1,46 1,67 1,42 1,61 1,71 1,66 1,54
1,72
1,50 1,48
1,72
1,72 1,87 1,88
Zu diesem Zwecke wurden die FrSsche n~ch L~iwen-Trendelenburg prgpariert, von der Aorr aus gemeins~m oder von den Arteriae iliacae ~us sep~r~t durchstr6mt und elektrisch direkt gerdzt.
662 E. Ernst und L. Scheffer:
Es h~ndelte sich, wie gesagt, um die Priifung jener Annahme, daB infolge der 1%izung das Kal ium mobilisiert und mi t der Durchst r6mungs- fliissigkeit h inausgeschwemmt wird. U m dies beweisen zu k6nnen, muBte zuerst entschieden werden, ob die Durchs t rSmung ~llein nicht imstande ist, einen Teil des Kal ium aus den Muskel herauszubef6rdern. Diese Frag e beantwor ten die in Tab. 3 zusammengestel l ten Versuche, wo die Kal iumwerte jener Gastrocnemien zu finden sind, die mehrere Stun- den lang mit einer 1%inger16sung von der Zusammensetzung 6,50/00 NaC], 0,1 o/0 o KC1 und CaCl�87 0,2 ~ o NaHCOa durchs t rSmt wurden. Diese Zahlen zeigen, - - wie es auch Mitchell und Wilson fanden - - dM~ der ruhende l~uskel selbst nach einer mehrere S tunden lang fortgesetzten Durch- s t r6mung mit einer normMen Ringerl6sung seinen normalen KMium- gehalt f~st unverminder t beibehi~lt.
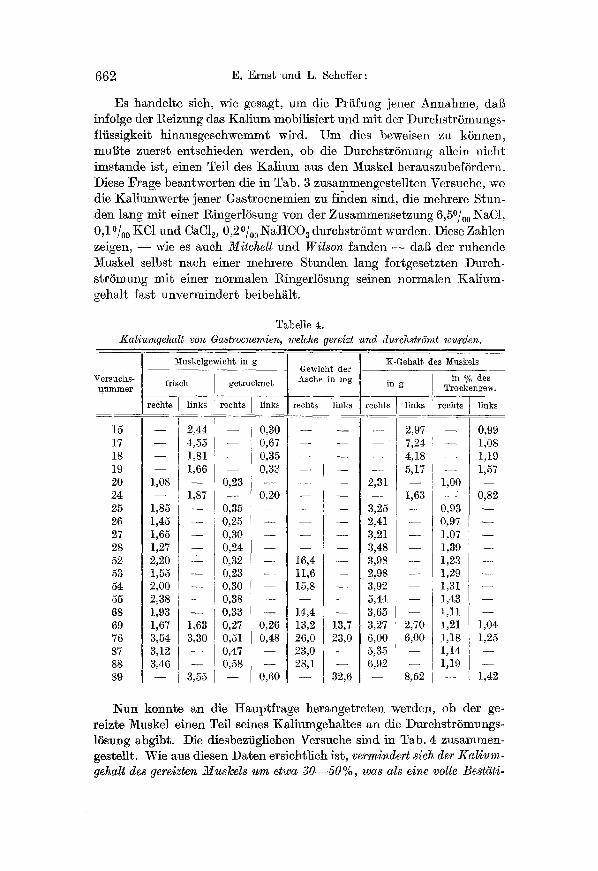
Tabelle 4. Kaliumgehalt vou Gay welche gereizt und durch~.tr6mt wurden.
Versuchs- nuramer
15 17 18 19 20 24 25 26 27 28 52 53 54 55 68 69 76 87 88 89
I fr isch getrocknet
I rechts links rechts links
1,08
1,85 1,45 1,65 1,27 2,20 1,55 2,00 2,38 1,93 1,67 3,54 3,12 3,46
Muskelgewicht in g
2,44 4,55 1,81 1,66
1,87 0,23
0,35 0,25 0,30 0,24 0,32 0,23 0,30 0,38 0,33 0,27 0,51 0,47 0,58
0,30 0,67 0,35 0,33
0,20
0,26 0,48
0,60
Gewicht der Asche in mg
rechts links
16,4 - - 11,6 15,8
14,4 13,2 13,7 26,0 23,0 23,0 - - 28,1
- - 32,6
K-Gehalt, des Muskels
in g in % des Trockengew.
1,63 3,30
3,55
rechts links rechts links
2,97 7,24 4,18 5,17
2,31 - -
- - 1,63 3,25 2,41 - - 3,21 3,48 3,98 2,98 - - 3,92 - - 5,44 - - 3,65 3,27 2,70 6,00 6,00 5,35 6,92 - - - - 8,52
1,00
0,93 0,97 1,07 1,39 1,23 1,29 1,31 1,43 1,11 1,21 1.18 1114 1,19
0,99 1/o8 1,19 1,57
0,82
1,04 1,25
1,42
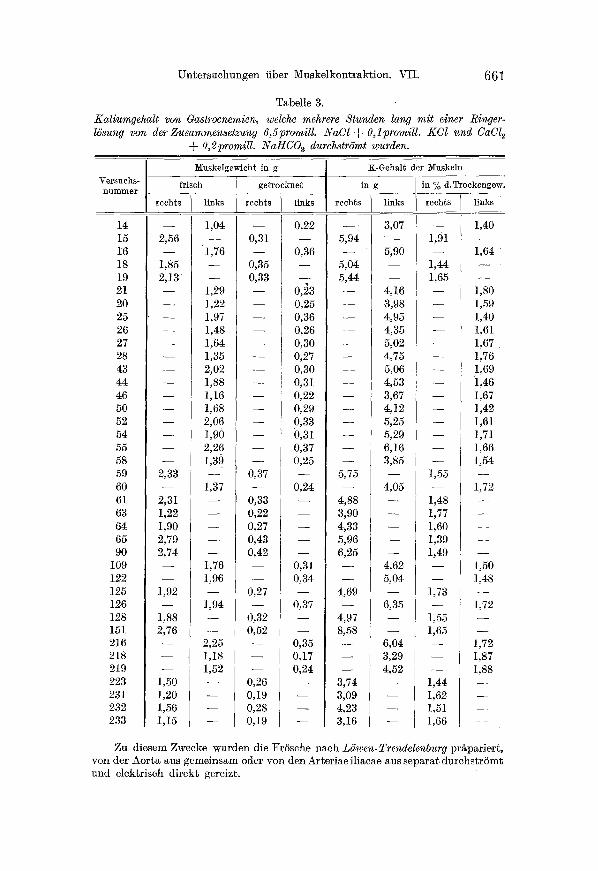
Nun konnte an die H~uptf rage herangetre ten werden, ob der ge- reizte Muskel einen Teil seines KMiumgeh~ltes un die Durchst rSmungs- 15sung abgibt. Die diesbezfiglichen Versuche sind in Tab. 4 zusammen- gestellt. Wie aus diesen D~ten ersichtlich ist, vermindert 8ich der Kal ium- gehatt des gereizten Muskels um etwa 30- -50%, was als eine volle Best~ti.

Untersuchungen tiber Muskelkontraktion. VII. 663
gung ]ener eingangs gegufierten Meinung scheint, nach welcher ein Teil des indi//usiblen Kaliums durch den Reiz mobilisiert und als solcher an die DurchstrSmungs/liissigkeit abgegeben wird.
11. Einwiin”
Diese Frage scheint jedoch eine viel verwickeltere zu sein, als dag sie sich so leicht erledigen liel]e. Es roui3 ngm]ieh auch jene MSglichkeit erwogen werden, ob der Kaliumverlust tats/~chlich als die prim/s Folge der Reizung - - verursacht durch die Ionisation des vorher indiffusiblen Kaliums - - oder aber als eine sekund~re Wirkung au�9 ist. Wir wollen 3 solche M0glichkeiten besprechen: 1. Der Kaliumverlust wird durch die Ermiidung bedingt. 2. Das Kalium wird von der Milchs/~ure herausgeschwemmt, welche bel der Reizung entstanden ist. 3. Der Reiz macht die Muskelmembranen fiir Kalium permeabel und 5ffnet dadurch dem aueh vor dem Reiz ionisierten I™ einen Weg nach aul3en.
Zu I. Mitchell und Wilson 1 behaupten auf Grund ihrer Versuche, dal~ die l~eizung keinen Kaliumverlust bedingt, derselbe wird erst dann er- reicht, falls solange gereizt wird, bis der Muskel seine Reizbarkeit verliert. Dem ist aber nicht so. Es sel an erster Stelle erwghnt, dal3 zu einem Kaliumver]ust keineswegs eine ,,extreme stage of fatigue" nStig ist. Es wird ja von Schenk 2 berichtet, dag bel stark ermfideten Menschen eine ErhShung des Kaliumgehaltes ira Blut zu finden ist, es ist also auch hier mit einiger Wahrseheinliehkeit Kaliumaussehwemmung aus den Muskeln anzunehmen, ohne dal3 von einer irreiersiblen Seh~digung des Muskels gesprochen werden kSnnte. Auch wir konnten in mehreren Versuchen einen bedeutenden Kaliumverlust beobachten (siehe Tab. 5), aber die .Mus- keln zeigten auch nach den Reizperioden noch eine Erregbarkeit. Es braucht also keine ,,totale" Ermfidung eingetreten zu sein, um einen K~lium- verlust im Muskel beobachten zu kSnnen.
Tabelle 5. Kaliumgehalt von Gastrocnemien, welche nach den ~eizperioden noch erregbar waren.
~ ~ I Muskelgewicht in g
> = ]ungereizt gereizt lungoreiz~ gereiz$
76 i - - 3,54 87 3,06 3,12 88 3,45 3,46
155 1,56 1,32 2191 1,52 1,76 2231 1,50 1,64
Gewich~ der [ K-Gehal~ de s Muskels Asche in mg I in - ! in Proz. des
1 ~ Trockengewiehtes
ungereiztl gereizt | ungereizt I ~ ; !~e�9
0,47 0,59 0,27 0,24 0,26
0,51 0,47 0,58 0,28 0,24 0,24
24,4 29,1
14,7 14,8
26,0 - - 23,0 7,18 28,1 9,20 - - 3,81
13,2 4,52 15,3 3,74
6,00 5,35 6,92 2,84 0,81 2,45
- - 1,18 1,53 1,14 1,56 1,19 1,41 1,01 1,88 0,34 1,44 1,02
J. gen. Physiol. 4, 45 (1922). z l-VItinch, med. Wschr. (1925) 2053.
'664 E. Ernst und L. Scheffer:
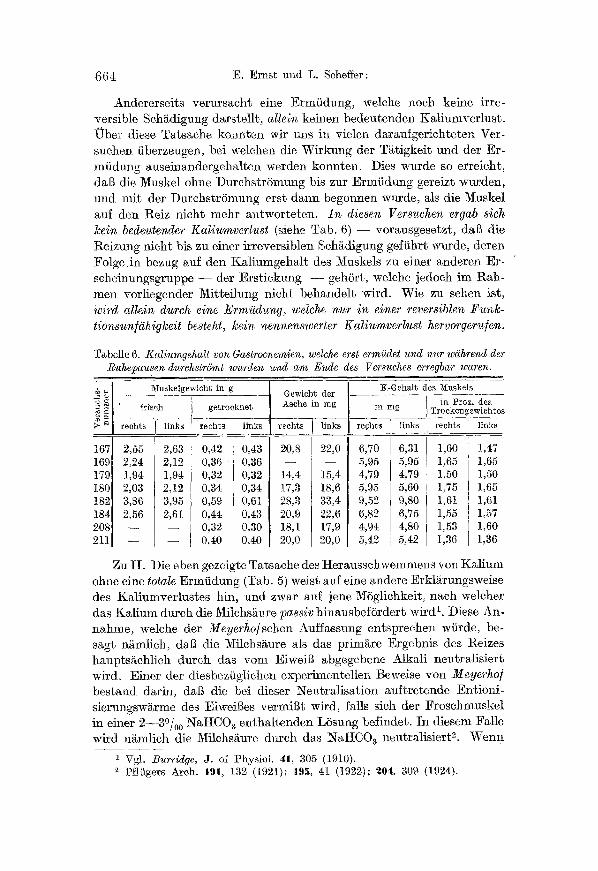
Andererseits verursacht eine Ermiidung, welche noch keine irre- versible Schadigung darstellt, allein keinen bedeutenden I™ Uber diese Tatsache kon~s wir uns in vielen darau~gerichteten Ver- suchen fiberzeugen, bel welchen die Wirkung der Tatigkeit und dcr Er- miidung auseinandergehalten werden konnten. Dies wurde so erreicht, dal~ die Muskel ohne Durchstr6mung bis zur Ermiidung gereizt w~rcden, und mit der DurchstrSmung erst dann begonnen wurde, als die Muskel auf den Reiz nicht mehr antworteten. In diesen Versuchen ergab sich kein bedeutender Kaliumverlust (siehe Tab. 6) - - vorausgesetzt, daB die Reizung nicht bis zu einer irreversib]en Sch~digtmg gefiihrt wurde, deren Folge in bezug auf derr Kaliumgehalt des Muskels zu einer anderen Er- scheinungsgruppe - - der E�9 --_- geh5rt, welche jedoch ira Rah- men vorliegender Mitteilung nicht behandelt wird. Wie zu sehen ist, wird allein dureh eine Ermi~dung, welche nur in einer reversiblen Funk- tionsun/š besteht, kein nennenswerter Kaliumverlust hervor9eru]en.
Tabelle 6. Kaliumgehalt von Gastrocnemien, welche erst e�9 und nur wdihrend der Ruhepausen durchstr6mt wurden und ara Ende des Versuches r waren.
Muskelgewicht in g
oe $ �9238
rechts links
167 2,55 2,63 169 2,24 2,12 179 1,94 1,94 180 2,03 2,12 182 3,86 3,95 184 2,56 2,6i 208 - - - - 211
! getrocknet
0,42 0,43 0,36 0,36 0,32 0,32 0,34 0,34 0,59 0,61 0,44 0,43 0,32 0,30 0,40 0,40
Gewicht der Asche in mg
re™ links
6,70 6,31 5,95 5,95 4,79 4,79 5,95 5,60 9,52 9,80 6,82 6,75 4,94 4,80 5,42 5,42
K-Gehalt des~fuskels
in Proz. des in mg Trockengewichtes
rechts
1,60 1,65 1,50 1,75 1,6i 1,55 1,53 1,36
�9 links
20,8 22,0
14,4 15,4 17,3 18,6 28,3 33,4 20,9 22,6 18,1 17,9 20,0 20,0
links
1,47 1,65 1,50 1,65 1,61 1,57 1,60 1,36
Zu I I . Die eben gezeigte Tatsa.che des Herausschwemmens von KMium ohne eine totale Ermfidung (Tab. 5) weist auf eine andere Erklgrungsweise des Kaliumverlustes hin, und zwar auf jene MSglichkei% nach we]cher dgs Kal ium durch die Milchsgure passiv hinausbef6rdert wird 1. Diese An- nahme, welche der Meyerho/schen Auffassung entsprechen wiirde, be- sagt n/s dal~ die Milchs~ure als das prim~re Ergebnis des Reizes haupts~chlich dureh das vom Eiweif~ o~bgegebene AlkMi neutrMisiert wird. Einer der diesbeziiglichen experimentellen Beweise von Meyerho[ bestand darin, dal~ die bel dieser Neutraiisation auftretende Entioni- sierungsw/trme des Eiweil~es vermi{~t wird, falls sich der Froschmuskel in einer 2--3~ NaHCOa enthaltenden L6sung befindet. In diesem Falle wird n~mlich die Milchs~ure durch dgs I~aHCOs neutralisiert ~. Wenn
1 Vg]. Burrid9e, J. of Physiol. 41, 305 (1910}. P�9 Arch. 191, 132 (1921); 195, 41 (1922); 204, 309 (1924).
Untersuchmlgen tiber :Viuskelkontraktion. VIL 665
Mso die Durchstr6mung des gereizten Froschmuskels mit einer solchen Ringerl6sung vorgenommen wird, deren NaHCO3-GehMt 2o/0o betr~gt, so diirfte der gereizte Muskel nicht KMium an die durchspfilende L6sung abgeben, da die Aviditgt der Milchs• durch das NaHCOs voll gedeckt wi~re. Die Versuche aber, die in Tab. 7 zusammengeste]lt sind, zeigen,
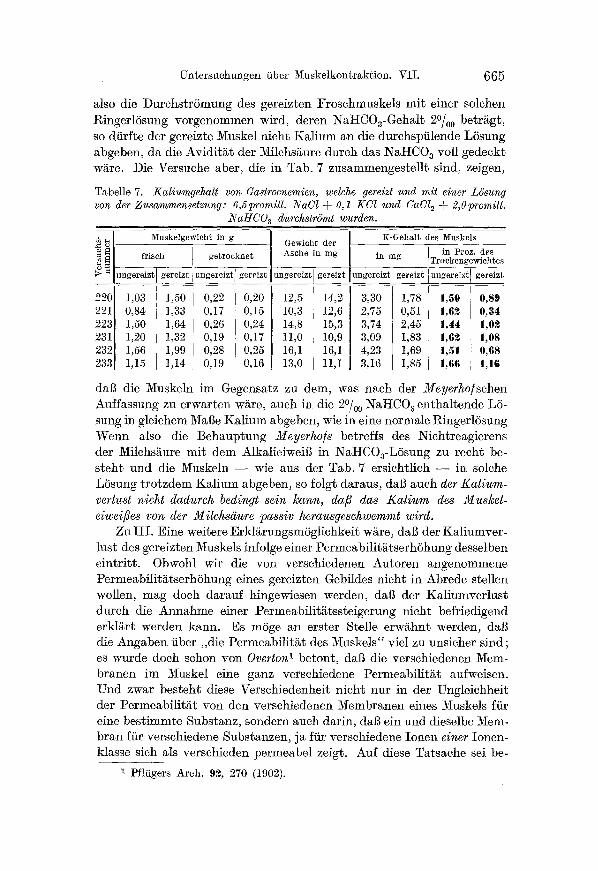
Tabelle 7. Kaliumgehalt von Gastrocnemien, welche gereizt und mit einer L6sung von der Zusammensetzung: 6,5promill. 1VaC1 ~- 0,1 KCl und CaCl 2 -~ 2,0promill.
NaHCOa durchstrSmt wurden.
[ ~uskelgeewicht in g
oe ~ lungerei~�9 g E } ~ œ i 2201 1,03 221[ 0,84 223~ 1,50 231[ 1,20 232[ 1,56 233[ 1,15
1,50 1,33 1,64 1,32 1,99 1,14
0,22 0,17 0,26 0,19 0,28 0,19
0,20 0,15 0,24 0,17 0,25 0,16
Gewicht der Asche in mg
K-Gehalt des Muskels I in Proz. des
in mg , Trockengewichtes ungereizt] gereizt
12,5 14,2 10,3 12,6 14,8 15,3 11,0 10,9 16,1 16,1 13,0 11,1
ungereizt] gereizt ]ungereizt] gereizt
1,78 0,51 2,45 1,83 1,69 1,85
3,30 2,75 3,74 3,09 4,23 3,16
1,50 0,89 1,62 0,34 1,44 1,02 1,62 1,08 1,51 0,68 1,66 1,16
daB die Muskeln im Gegensatz zu dem, was nach der Meyerho/schen Auffassung zu erwarten w~re, auch in die 2~ NattCO z enthaltende L6- sung in gleichem Mage KMium abgeben, wie in eine normale Ringerl6sung Wenn also die Behaup~ung Meyerho/s betreffs des Nichtreagierens der Milchsaure mit dem AlkalieiweiB in NiI-ICOa-L6sung zu recht be- steh~ und die Muskeln - - �87 aus der Tab. 7 ersichtlich - - in solche L6sung t rotzdem Kal ium abgeben, so folgt daraus, daB auch der Kalium- verlust nicht dadureh bedingt sein kann, dafi das Kalium des Muskel- eiwei/3es von der Milchsgure passiv herausgeschwemmt wird.
Zu I I I . Eine weitere Erkl~rungsm6glichkeit ware, dag der Kaliumver- lust des gereizten Muskels infolge einer Permeabilit~tserh6hung desselben eintritt. Obwohl wir die von verschiedenen Autoren angenommene Permeabilit~tserh6hung eines gereizten Gebildes nicht in Abrede stetlen wollen, mag doch darauf hingewiesen werden, daB der KMiumverlust durch die Annahme einer Permeabilitatssteigerung nicht be�9 erklgrt werden kann. Es m6ge an erster Stelle erw~hnt werden, daB die Angaben fiber ,,die Permeabiliti~t des Muske]s" r iel zu unsicher sind ; es wurde doch schon von Overton 1 betont, daB die verschiedenen Mem- br~nen im Muskel eine ganz verschiedene Permeabilit~t aufweisen. Und zwar besteht diese Verschiedenheit nicht nur in der Ungleichheit der Permeabil i tat von den verschiedenen Membranen eines Muskels fiir eine best immte Substanz, sondern auch darin, d~i~ ein und dieselbe Mem- bran fiir verschiedene Subs~anzen, ja ftir verschiedene Ionen einer Ionœ Masse sieh Ms versehieden loermeabel zeigt. Auf diœ Tatsache sel be-
l Pfliigers Arch. 92, 270 (1902).
666 E. Ernst und L. Scheffer:
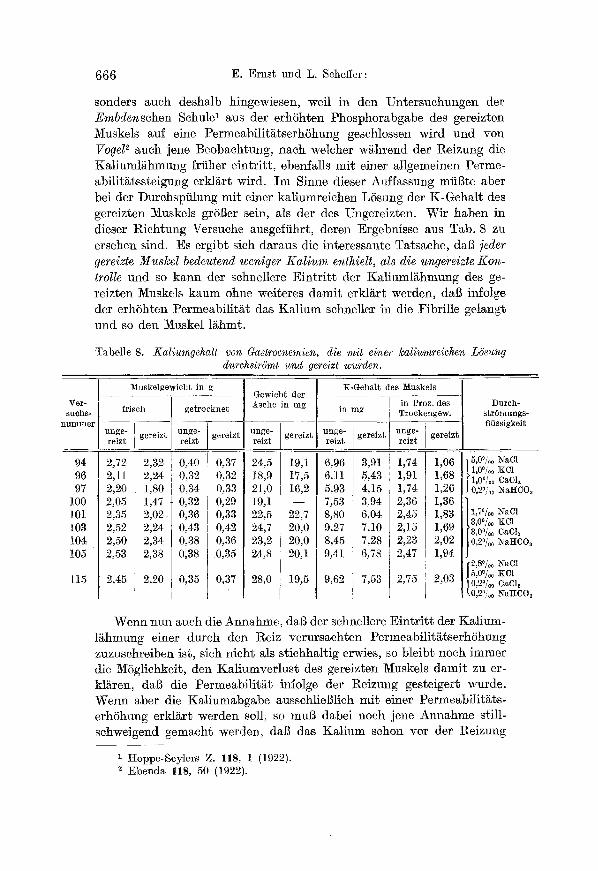
sonders aueh deshalb hingewiesen, weil in den Untersuchungen der Embdensehen SchulO aus der erh6hten Phosphorabgabe des gereizten Muskels auf eine Permeabilit~tserh6hung geschlossen wird und von Vogel 2 auch jene Beobaehtung, naeh welcher w~.hrend der l~eizung die KMiuml~hmung frfiher eintritt, ebenfalls mit einer allgemeinen Perme- ~bilitgtssteigung erkl~rt wird. Im Sinne dieser Auffassung mfil3tœ aber bei der Durchspiilung mit einer kaliumreichen L6sung der K-Gehalt des gereizten Muskels gr6Ber sein, als der des Ungereizten. Wir haben in dieser Riehtung Versuehe ausgeffihrt, deren Ergebnisse aus Tab. 8 zu ersehen sind. Es ergibt sich daraus diœ interessante Tatsache, daB ]eder gereizte Muskel bedeutend weniger Kalium enthielt, als die ungereizte Kon- trolle und se kann der sehnellere Eintritt der Kaliumlghmung des ge- rœ Muskels kaum ohne wœ234 datait erkl/~r~ werden, dag infolge der erh6hten Permeabilit~t das Ka]ium sehnœ in die Fibrille gelangt und se den Muskel l~hmt.
Tabelle 8. Kaliumgehalt von Gastrocnemien, die mit einer kaliumreichen L6sung durchstr6mt und gereizt wurden.
V@r- I suchs- I
nummer I
94 96 97
100 101 103 104 105
115
luskelgewicht in g
. . . . frisch getrocknet
un.ge- gereizt I unge- relz~ I reizt
2,72 2,11 2,20 2,05 2,35 2,52 2,50 2,53
2,45
gereizt
2,32 0,40 0,37 2,24 0,32 0,32 1,80 0,34 0,33 1,47 t 0,32 0,29 2,02./ 0,36 0,33 2,24 / 0,43 0,42 2,34 / o,3s 0,36 2,38 / 0,38 0,35
! 2,20 L0,35 0,37
Gewich~ der Asche in mg
unge- gereizt reizt
24,5 19,1 18,9 17,5 21,0 16,2 19,1 - - 22,5 22,7 24,7 20,0 23,2 20,0 24,8 20,1
28,0 19,5
K-Gehalt des Muskels
in mg
unge- gereizt reizt
6,96 3,91 6,11 5,43 5,93 4,15 7,53 3,94 8,80 6,04 9,27 7,10 8,45 7,28 9,41 6,78
9,62 7,53
in ~roz. des Trockengew.
unge- gereizt reizt
1,74 1,06 1,91 1,68 1,74 1,26 2,36 1,36 2,45 1,83 2.15 1,69 2]23 2,02 2,47 1,94
2,75 2,03
Dureh- str6mungs- itiissigkeit
5,0%o NaC1 1,0% o KC1 1,0%o CaCI.. 0,2%o );aHC03
1,7o/o~ NaC1 3,0o{o~ :KC1 3,0~ Ca™ 0,2%o NaHC03
2,8% o .N-aC1 5,0%o :KC1 0;2%o Cael~ 0,2%o N~ItCOs
Wenn nun auch die Annahme, dag der schnellere Eintritt der KMium- l~hmung einer durch den Rœ verursaehten Permeabilit~tserh6hung zuzusohreiben ist, sich nieht als stiehhalt�8 erwies, se bleibt noeh immer die M6glichkeit, dœ Kaliumverlust des gereizten Muskels datait zu er- kl~ren, daB die Permeubilit~t in�9 der l~eizung gesteigert wurde. Wenn ~ber diœ KMiumabgabe aussehliœ mi™ einer Perme~bilit~ts- œ erkl~r~ werden soll, se mu$ dubei noch jœ Annahme still- sehweigœ gemacht werdœ d~$ das I™ sehon ver der Reizung
1 ]=Ioppe-Seylers Z. 118, 1 (1922). 2 Ebœ 118, 50 (1922).
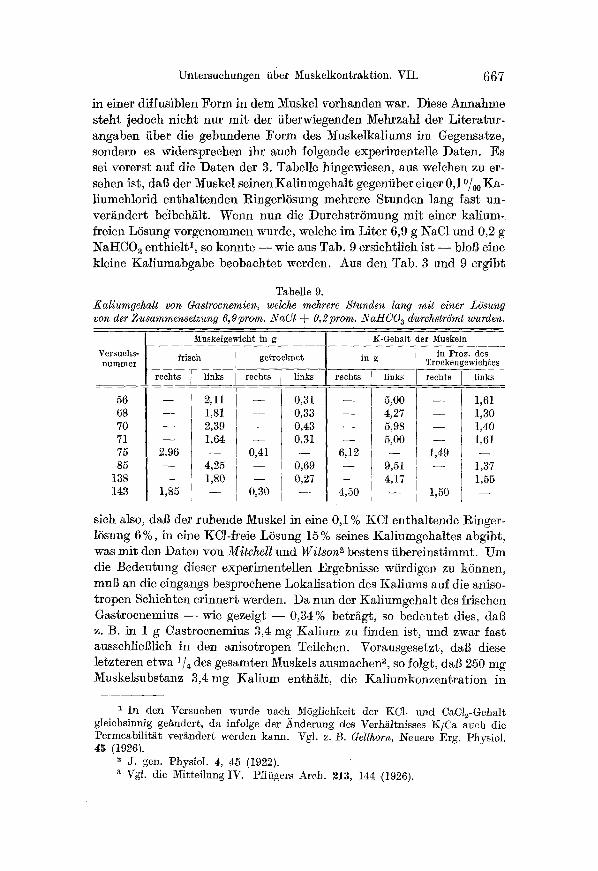
Untersuchungen liber Muskelkontraktion. VII. 667
in einer dif�9 Form in dem Muskel vorhanden war. Diese Annahme steht jedoch nicht nur mi t der tiberwiegenden Mehrzahl der Liter~tur- angaben iiber die gebundene Form des Muskelkaliums ira Gegensatze, sondern es widersprechen ihr auch �9 experimentelle Daten. Es sei vorerst auf die Daten der 3. T~belle hingewiesen, aus we]chen zu er- sehen ist, daI~ der Muskel seinen K~liumgehalt gegeniiber einer 0,1 ~ Ka- liumchlorid enth~ltenden RingerlSsung mehrere Stunden lang fast un- ver~ndert beibehi~lt. Wenn nun die DurchstrSmung mit einer kalium- freien L6sung vorgcnommen wurde, welche ira Liter 6,9 g NaC1 und 0,2 g NaHC03 enthielt 1, so konnte - - wie sus Tab. 9 ersichtlich ist - - blol~ eine kleine KaHumabgabe beobachtet werden. Aus den Tab. 3 und 9 ergibt
Tabelle 9. Kaliumgehalt von Gastrocnemien, welche mehrere Stunden lang mit einer L6sung von der Zusammensetzung 6,9prom. NaC1 @ 0,2 orom. 2VaHCO a durchstrSmt wurden.
~r in g t ™ d e r M u s k e l n
in Proz. des VersuehS-nummer frisch getrocknet in g Trockengewichtes
rechts links rechts links rechts I liuks rechts links I
56 68 70 71 75 85
138 143
2,96
1,85
2,11 1,81 2,39 1,64
4,25 1,80
m
0,41
0,30
0,31 0,33 0,43 0,31
0,69 0,27
6,12
4,50
1 5,00 4,27 5.9s 5100
9,51 4,17
1,49
1,50
1,61 1,30 1,40 1,61
1,37 1,55
sich also, da~ der ruhende Muskel in eine 0,1% KC1 enthaltende Ringer- 15sung 6%, in eine KCl-�9 LSsung 15 % seines Kaliumgehaltes abgibt, was mit den Daten von Mitchel l und Wil son 2 bestens fibereinstimmt. Um die Bedeutung dieser experimentellen Ergebnisse w/irdigen zu k5nnen, mul~ an die eingangs besprochene Lokalisation des Kaliums auf die aniso- tropen Schichten erinnert werden. Da nun der Ka]iumgehalt des frischen Gas~rocnemius - - w i e gezeigt - - 0,34% betr~gt, so bedeutet dies, dal~ z. B. in 1 g Gastrocnemius 3,4 mg Xalium zu finden ist, und zwgr f~st ausschliel~lich in den anisotropen Teilchen. Vorausgesetzt, dal~ diese letzteren etwa 1/4 des gesamten Muskels gusmachen s, so �9 dal3 250 mg Muskelsubstanz 3,4 mg Kal ium enth~tlt, die Kaliumkonzentrat ion in
1 In den Versuchen wurde nach MSglichkeit der KCI- und CaCl~-Gehalt gleichsinnig ge~ndert, da infolge der )~nderung des Verhgltnisses K/Ca auch die Permeabilitht ver~tndert werden kann. Vgl. z. B. Gellhorn, Neuere Erg. Physiol. 45 (1926).
J. gen. Physiol. 4, 45 (1922). a Vgl. die Mitteilung IV. Pflfigers Arch. 213, 144 (1926).

668 E. Ernst und L. Scheffer:
denselben betr~gt Mso e~wa 14~ Wenn wir noch bedenken, d~g der Muskel sich mit einer L6sung, welche eine Ka]iumkonzentrat ion von etwg 0,1~ aufweist, isoionisch zeigt, so kommen wir zu dem Sehlug,
dag die Kaliumkonzentrat ion der Fibrillen ungef~hr OE~ ~ 140mal
gr613er ist als die ihrer Umgebung. Dieser gewalt�9 Konzentr~~ions- untersehied glœ sieh aueh in mehreren Stunden eines Versuehes, selbst in viœ Jagren des Lœ nieht nus. Sollte eine so absolute an keinem anderen Objekt vorzufindende Kaliumimpermeabil i tgt eben nur bei der Muskelfibrille vorhanden sein ? Oder wiirde es doeh eher wahrsehœ erseheinœ dieses Nieh~ausgleiehen jenœ Umst~nd zuzusehreiben, dag der gr6gte Teil des Kaliums in der Fibrille in einer indiffusiblen Form vorhanden ist ~. Wir gl~uben, diese letztere Erkl~rungsweise um so mehr annehmen zu diirfen, da dureh dieselbœ aueh die nus Tab. 3 und 9 er- siehtliehe Tatsaehe erkl~rt werden kann, dag der ruhende Muskel an eine kMiumfreie bzw. -arme L6sung nur wenlg Kal ium abgibt.
Wir glauben also uns der von der Mehrheit der Autoren angenom- menen Auffassung, daB n~mlieh der gr6gere Teil des Kaliums sieh in der Fibrille in indi�9 Form befindœ ansehlieBen zn mfissœ Wenn dem aber so ist, dann ist es ohne weiteres klar, dag der Reiz ausschliefilieh dureh Permeabilitiitssteigerung das Kalium nus der Fibrille nicht heraus- be]6rdsrn lcann, dazu ist noeh die Mobilisierung bzw. Ionisierung des vorher indiffusiblen Kaliums n6tig 2.
In diesem Zusammenhang dfirfte es viel]eieht erlaubt sein, neeh guf eine andere Erscheinungsgruppe hinzuweisen, und zwar au�9 den Unter- sehied zwisehen der elektrisehen Leitfghigkeit des ruhenden bzw. ge- reizten Muskels. Es ist n~mlieh sei~ langem bekannt, dag der Mnskel und der Nerv bel Querdnrehstr6mung eine r iel kleinere Lei~f~higkeit zeigen als bei L~ngsdurehstrSmung. Diese Tatsaehe wurde anseheinend rieh~ig dahin gedeutet, daB der Strom bel der QuerdurehstrSmung in den Membranen viel mœ W~derst~nde findet als bel Lfi.ngsdurehstrS- mung. Es wm'de nun die Er�9 gemaeht, daB die Leitf~higkœ des Mnskels und des Nerven bei der t~eizung sieh vergr6Bert und diesœ Leit- fahigkeitserhShung datait erkl~.rt, daB die Permeabil i tat ildolge des Reizes sieh erhSht nnd so dem Passieren des Stromes weniger Wider- stand bereitet. Es wurde aber sehon von Hermann aueh jene interessante
1 Die Angaben, daB in einer isotonisehen KC1- (8~ oder K2S04-L6sung (16~ die Kaliumkonzentrution gr6Ber w~re, ~ls in der Fibrille, mfissen also ~ls irrti~mlich beze�9 werden.
2 Aueh Embden nimmt zur Erkl~rung der gesteigerten Phosphors~ureabgabe ~uger der l%rme~bilit~tszun~hme ,,eine ira Kontraktionsaugenbliek erfolgende Vermehrung d6r ~norganisehen Phosphors~ure ira Muskelinnern" ~n (ttolope - Seylers Z. 118, 1 [1922]).

Untersuehungen iiber Muskelkontraktion. VIL 669
Tatsaehe gezeigt ~, daB dœ Ileiz eine viel kleinere Leitf/~higkeitszunahme bei der Querdurehst rSmung Ms bei der L/~ngsdurehstr6mung hervor- ruft . Bestiinde nun die Wi rkung des l~œ ausehlieBlieh in einer Perme- abi l i tgtszunahme, so w~re zu erwarten, d~B der Querwiderstand dureh den l~eiz r iel mehr abnimmt, Ms der Lgngswiderstand, weil ja der gr613te Teil des Querwidœ - - die Impermeabi l i t~ t der Membranen - - durch den Reiz aufgehoben wurde. D~ Hermann - - wie gesagt - - das Entgegengesetzte davon fand, so kommen wir zu dœ Uberzeugung, dag aueh die Erscheinungen der Lei t �9 des gereizten Muskels mi t der Annahme der Permeabi l i t~tszunahme Mlein nieht befriedigend zu erklgren sind. Die eben angedeuteten Schwiœ lassen sich aber dureh die Annahme einer Neubi ldung von Ionen ebenfMls beheben. Selbst H6ber, einer der eifrigster~ Vork/~mpfer der Permeabilit/itsfrage, ~uBert si› ja dahin, dag die Leit�9 entweder durch das Freiwerden von Elektrolyten oder dureh t terantransloort derselben an die P lasmahau t bedingt ist 2.
! I I . C)ber den Mechanismus des K-Austr i t tes .
Kehren wir Mso zu unserer m'spriinglichen Annahme zurtick, laut Weleher auf den Reiz hin eine Neubi ldung vor~ Ionen stat tf indet , so stehen wir mi t dieser Annahme keinesfMls Mlein. Es wurde doeh schon von Howell und Duke ~ und von Macdonald 4 die Auffassung geguBert, dag in- folge des Reizes KMium aus der gebundenen Form frei wird und in den Ionenzus tand �9 5. Aueh Loeb ba t sieh dieser Annahme angesehlos- sen G. In diesem Zusammenhange sei aueh auf den Befund Boehms 7 hin- gewiesen, daB ngmlich das t Ierz in eine kMiumlose Ringerl6sung Kal ium abgibt, welche Ang~be Zwaardemaker s in zahlreichen Versuehen best~tigt 2and. Es mag �9 die Aulfassung von Burridge~ Erwghnung �9234 n a e h welcher ein Teil des �9 in einer indiffusiblen F o r m
1 Pflfigers Areh. 5; 239 (1872); 6, 560 (1872); �9 ~, 323(1873); I@, 215 (1875); |~, 154 (1876). Hermann spricht setbstverst/indlich nicht in der 1N'omenklatur der post~rrheniusschen Zeit, doch bedeutet das von ihm Gesagte dasselbe.
Physik, Chemie der Zelle und Gewebe ~, 718 (1924). Vgl. Cremer in Nagels Handbuch 4, 878 (1909). - - Verzdr (Pfliigers Arch. 158, 304 [1913]) erw/~hnt eben- fa.lls diese 2 MSgliehkeiten, wenn er sich auch zu der Annuhme der Perme~bi]it/~ts- ~nderung bekennt.
3 Amer. J. Physiol. ~1, 51 (1908). Burridge, J. of Physiol. 41, 304 (1910).
�87 Vgl. Hemmete~', Bioehem. Z. 63, 119 (1914), der dies in Zweifel zieht, ent- gegen den Ang~ben von Schein/inkel (Z. Biol. 8~, 286 [1925]) und von Granit und Bornsdor/ (Sk~nd. Areh. Physiol. [Berl. u. Lloz. ] 51, 311 [1927]).
60ppenheimers 1~Iandbueh der Biochemie ~ (I), 139 (1910). - - Vgl. Neuschlo~z, Pfliigers Arch. l~~, 410 (1923).
Areh. f. exper. Path. 7~, 230 (1914). s Erg. Physiol. 1~, 342 (1921) und 2~, 547 (1926).
J. o�9 Physiol. 4~, 359 (1911).

670 E. Ernst und L. Scheffer:
dureh impermeable Membranen eingeschlossen ist und an der Kon- traktion teilnimmt.
Wenn auch diese Erkl~rung des Kaliumverlustes mit der Ionisation des gebundenen Kaliums wahrscheinlich gemacht werden konnte, so fragt es sieh, wie das K-Ion den Muskel verl~{~t. Lange bevor eine theo- retisch ausgearbeitete Erkl~rungsweise zu Gebote gestanden w~re, hat Overton 1 angenommen, dag bel der Kontraktion ein Austausch zwischen Kalium und Natrium statffindet und dieser Auffassung haben sich seit- her mehrere Autoren angeschlossen ~. Diese Annahme �9 eine ganz &llgemeine, theoretisch auszezeiehnet ausgearbeitete Grundlage in den zahlreichen Arbeiten von Micha6li~ a. Diese berechtigen uns heutzutage a.nzunehmen, dag das Kal ium dis Mus~el]ibrille in der Weise verlgflt, dafl an seine Stelle Natr ium aus dem Auf ienmedium eintritt.
Ira Sinne dieser Annahme war also zu erwarten, dag der Na-Gehalt des Muskels entsprechend dem Kaliumverlust sich vermehrt. Aber se]bst, wenn dieser Fall sich verwirklichte, konnte er noeh immer nicht ffir einen Beweis des I™ gelten, da der vermehrte I~a-Gehalt des gereizten Muskels aueh datait erkl~rt werden k6nnte, daB der gereizte ~r in�9 seiner erh6hten Permeabilit~t mehr l~aC1 ans der Ringer- 16sung au�9 In diesem Falle mfigten selbstverst.~ndlich die Werte des iiber den Normalwert gefundenen Natriums und Chlors miteinander genau ~quivalent sein. Der Versueh 4 zeigte aber (s. Tab. 10), d~l~ in alIen F&llen mehr Natrium vom gereizten Muskel au�9 wird als Chlor, oder anders ausgedrfiekt, besteht ein Na-~bersehug. Ira Sinne
Tabelte 10. Kaliumverlust und Natriumi~berschu[3 der gereizten und durchstr6mten Gastroenemien. Aus der L6sung ist NaHCO S weggelay
V e f s u c h s - n u m m e r
235 236 237 238 240
K-Verlust i Na-Uberschu~
in mg-~quivaIent
0,059 0,032 0,061 - - 0,015 0,045 0,0].7 0,028 0,021 0,028 0�87
V e r s u c h s - n u m � 9
242 244 245 246
K-Verlust T Na-�8
in mg-Xquivalent
o,o28 ' o23o 0,021. 0,018 0,046 0,011 0,032 ' 0 008
1 Pfliigers Arch. 92, 381 (1902). 2 Vgl. Robertson, Erg. Physiol. 10, 327 (1910). - - H6ber, Physik, Chemie der
Zelle und Gewebe 2, 715 (1924). - - Abderhal” und Gellhorn, Pfliigers A_rch. 196, 584 (1922).
3 Biochem. Z. 146, 158, 161, 162, 163, 164, 169, 170, 1~2, 173 und J. gen. Physiol. 10 •. ll.
4 Es geh6rt nicht in den Rahmen vorliegender Mitteilung iiber die Einzel- heiten der angewandten 1V[ethoden zu berichten, zumal manche Teile der l~Iethodik von uns modi�9 worden sind, und so sei bezfig]ieh dieser auf unsere demn~tehst erscheinende l~Iitteilung hingewiesen.

Untersuchungen tiber Muskelkontraktion. VIL 671
unserer Annahme liber dan Austausch zwischen K und Na ira Muskel w~re es nun zu erw` gewesen, da~ das abgegebene Kalium und der Na-Uberschul3 in ihren Aquivalentwerten iibereinstimmen. Die Ver- �87 bestgtigen aber diese Erwartung ira allgemeinen nicht, was viel- leicht mit der Angabe von Embden und Lange 1 zu erklgren w~re, dal~ bai der Reizung des Muskels Chlor auch an Stalle v0n Phosphat in den Muskel gelangt, so dal3 aus der Mange des Chlors nicht mit Bestimmtheit der Natriumfibersehul~ festgestellt werden kann. Tdber diese Frage werden ira Gang befindliehe Versuche vie]leicht ngheren Aufsehlul3 geben, i iur erw~hnen mSehten wir noeh, dal~ die Annahme liber dan Austauseh des ionisierten Kaliums gegen Natrium mit den Versuchsergebnissen von Mitchell, Wilson und Stanton 2 gui iibereinstimmt, laut welehen Rubidium und C~sium nur vom gereizten Muskel verankert wird, nie aber vom ruhenden.
MiS dem hier Mitgeteilten war es uns nicht so sehr darum zu tun, die festgestellten Tatsachen in den Mittelpunkt zu stellen, ̀ vielmehr durch sic auf aine MSglich/ceit hinzuweisen, welche kurz in fo]gender Weise formuliert werden kann: Es /indet au/ dan Relz hin aine Neubildung von K-Ionen in den anisotropen Mus]~elschichten statt, und zwar aus einer in der Ruhe undissoziiert vorhandenen Kaliumverbindung. Aus diesem diHusibel gewordenen Kalium tritt wiihrend der Durchstr5mung ein Verlust ein, indem K. gegen iVa.Ion ausgetauscht wird.
Zusammenfassung. I. l. Der Normalgehalt des Frosehgastrocnemius (R. esc.) an Kalium
betrggt ira Mittel 1,70 % der Trockensubstanz. 2. DurchstrSmung mis normal-Ringer ftir die Dauer von 5--6 Stun-
den verursaeht einen 6proz., mit K-�9234 LSsung einen 15proz. K- Verlust.
3. Bel direkter Reizung erreicht der K-Verlust 30--50%. II. Der K-Verlust l~l~t sich allein weder dureh 1. Ermiidung noeh
durch 2. Milehs~turebildung oder 3. durch Permeabflit~tssteigerung des gereizten Muskels erkl~ren.
III . Der N` des gereizten Muskels nimmt mehr zu als der C1- Gehalt, woraus auf die MSgliehkeit gesehlossen wird, da~ ein K-Na- Aus~ausch im gereizten Muskel stattfindet.
1 Hoppe-Seylers Z. 130, 350 (1923). J. gen. Physiol. 4, 141 (1922).
Pltiigers Arehiv f. d. ges. Pllysiol. Bd. 220. 4~