Embed Size (px)
Citation preview

Aus der Klinik für Hals-Nasen-Ohrenheilkunde, Kopf- und
Halschirurgie am St. Elisabeth Hospital Bochum
Universitätsklinik der Ruhr-Universität Bochum
Direktor: Professor Dr. med. H. Hildmann
Untersuchungen zum osteoklastären Knochenabbau im Cholesteatom
Inaugural-Dissertation
zur Erlangung des Doktorgrades der Medizin
einer Hohen Medizinischen Fakultät der Ruhr-Universität Bochum
vorgelegt von Jörg Ebmeyer aus Tübingen
2002

Dekan: Prof. Dr. med. G. Muhr Referent: PD Dr. med. H. Sudhoff Koreferent: Tag der mündlichen Prüfung: 22. Oktober 2002

Meinem Vater in Dankbarkeit gewidmet

Inhaltsverzeichnis Seite
Abkürzungsverzeichnis
1. Einleitung 1
1.1 Historischer Überblick und Definition 1
1.2 Pathogenese des Cholesteatoms 3
1.2.1 Einteilung der Cholesteatome 3
1.2.1.1 Klassische Einteilung 3
1.2.1.2 Neuere Klassifikation 5
1.3 Klinisches Erscheinungsbild des Cholesteatoms 5
1.4 Typische Komplikationen des Cholesteatoms 6
1.5 Therapiemöglichkeiten des Cholesteatoms 7
1.6 Knochenabbau durch das Cholesteatom 7
1.7 Biologie des Osteoklasten 8
1.7.1 Funktion von Osteoblasten und Osteoklasten 8
1.7.2 Morphologische Grundlagen 8
1.8 Fragestellung und Zielsetzung 10
2. Material und Methoden 12
2.1 Ossikel 12
2.1.1 Präparate 12
2.1.2 Untersuchung der Präparate 12
2.2 Lichtmikroskopie 13
2.2.1 Färbung 13
2.2.1.1 Zellkulturen 13
2.2.1.2 Schädelknochenbiopsien 13
2.2.2 Mikroskop/Dokumentation 13
2.2.2.1 Dokumentation der Zellkulturen 13
2.3 Elektronenmikroskopie 14

2.3.1 Vorbereitung der Präparate 14
2.3.2 Critical Point Trocknung 14
2.3.2.1 Prinzip 14
2.3.2.2 Critical Point Trocknung der Präparate 14
2.3.3 “Sputtern” der Präparate 15
2.3.3.1 Prinzip 15
2.3.3.2 Sputtervorgang 15
2.4 Zellkulturen 16
2.4.1 Gewinnung der Zellen 16
2.4.2 Kultivierung der Zellen 16
2.4.3 Studien mit Zoledronat und Calzitonin in vitro 17
2.4.3.1 Zoledronat 17
2.4.3.2 Calzitonin 17
2.4.3.3 Auswertung der Zellkulturen 17
2.4.4 Dentin-Chips 18
2.5 Tierexperimente 18
2.5.1 Keratinimplantate 18
2.5.2 Zoledronat in vivo 20
2.5.3 Auswertung der Schädelknochenpräparate 20
2.6 Statistiken 21
3. Ergebnisse 22
3.1 Ossikel 22
3.1.1 Kontrollen 22
3.1.2 Knöchelchen aus Cholesteatom-erkrankten Ohren 22
3.1.3 Morphologie des Knochenabbaus 25
3.2 Zellkulturen 30
3.2.1 Wirkung von Zoledronat in vitro 30
3.2.1.1 TRAP-Färbung 30
3.2.1.2 TUNEL-Färbung 32
3.2.2 Wirkung von Calzitonin in vitro 33
3.2.2.1 Zellkulturen 33

3.2.2.2 Dentinscheibchen 34
3.3 Morphologie der Resorptionslakunen 37
3.4 Tierexperimente 41
3.4.1 Keratinpartikel-induzierte Osteolyse in vivo 41
3.4.2 Statistische Auswertung der Schädelpräparate 42
4. Diskussion 44
4.1 Morphologie der Ossikel im Cholesteatom 44
4.1.1 Oberflächliche Resorption 44
4.1.2 Perivaskuläre Knochenresorption 45
4.2 Vergleich der Knochenresorption in vivo und in vitro 45
4.3 Inhibition der Knochenresorption in vitro 46
4.3.1 Zoledronat in vitro 46
4.3.2 Calzitonin in vitro 47
4.4 Inhibition der Knochenresorption in vivo 48
4.4.1 Zoledronat in vivo 48
4.5 Vergleich der Wirkung von Zoledronat + Calzitonin 49
4.6 Klinische Relevanz 50
5. Zusammenfassung 51
6. Literaturverzeichnis 53
7. Danksagungen 60
8. Lebenslauf 61

Abkürzungsverzeichnis α-MEM minimal essential medium
CTR Calzitoninrezeptor
EDTA Ethylen-Diamino-Tetraessigsäure
hCT Humanes Calzitonin
M-CSF macrophage colony stimulating factor
Oc Osteoklast
ODF Osteoklasten-Differenzierungsfaktor (=OPGL=RANKL)
OPGL Osteoprotegerin-Ligand (=ODF=RANKL)
PBS Phosphate-buffered Saline
RANKL receptor-activated NFκB ligand (OPGL=ODF)
REM Raster-Elektronenmikroskopie
SEM Scanning Elektron Microscopy
TNF-α Tumornekrosefaktor α
TNF-β Tumornekrosefaktor β
TRAP tartrate resistent acid phosphatase
TUNEL TdT-mediated UTP nick end labelling

Einleitung 1
1. Einleitung
1.1 Historischer Rückblick und Definition
Das Cholesteatom ist ein häufig zu beobachtendes Krankheitsbild
des Mittelohres, das zu Schwerhörigkeit, einem Verlust des Gleich-
gewichtsorgans, einer Schädigung des Gesichtsnerven, aber auch
zu lebensbedrohlichen endokraniellen und septischen Komplikatio-
nen führen kann.
Du Verney war wahrscheinlich der erste, der eine Cholesteatom-
ähnliche Masse beschrieb. Er bezeichnete das Krankheitsbild 1683
als „Steatom“. Die Bezeichnung „Chole-steat-om“, also „Cholesterin-
Fett-Tumor“, prägte der deutsche Physiologe Johannes Müller 1838
aufgrund von histologischen Beobachtungen. Er beschrieb seinerzeit
„einen geschichteten perlförmigen Tumor aus Fett“.
Cruveilhier sprach 1829 wegen des makroskopisch perlförmigen
Erscheinungsbildes des Cholesteatoms eher deskriptiv von einer
„tumeur perlée“, einer „Perlgeschwulst“ des Felsenbeines. Dieser
Begriff wurde von Rudolf Virchow 1855 aufgegriffen. Bis in die
heutige Zeit hat sich allerdings aus unerfindlichen Gründen der
formal falsche und irreführende Begriff Cholesteatom gehalten.
Es handelt sich beim Cholesteatom um einen benignen Tumor der
Mittelohrräume, der aus zwiebelschalenartig angeordneten verhor-
nenden Plattenepithelzellen besteht. Das Cholesteatom wächst lang-
sam und lokal destruierend, wobei die Destruktion durch einen
Abbau des umliegenden Knochens bedingt ist. Das Cholesteatom
wächst weder aktiv infiltrierend wie ein maligner Tumor, noch setzt
es Metastasen.
Das Cholesteatom kann definiert werden als „chronische osteokla-
stische Knochenzerstörung als Folge von ortsfremdem, verhornen-
dem Plattenepithel in den normalerweise nur mit Schleimhaut ausge-
kleideten Mittelohrräumen mit bedrohlichen Komplikationsmöglichkei-
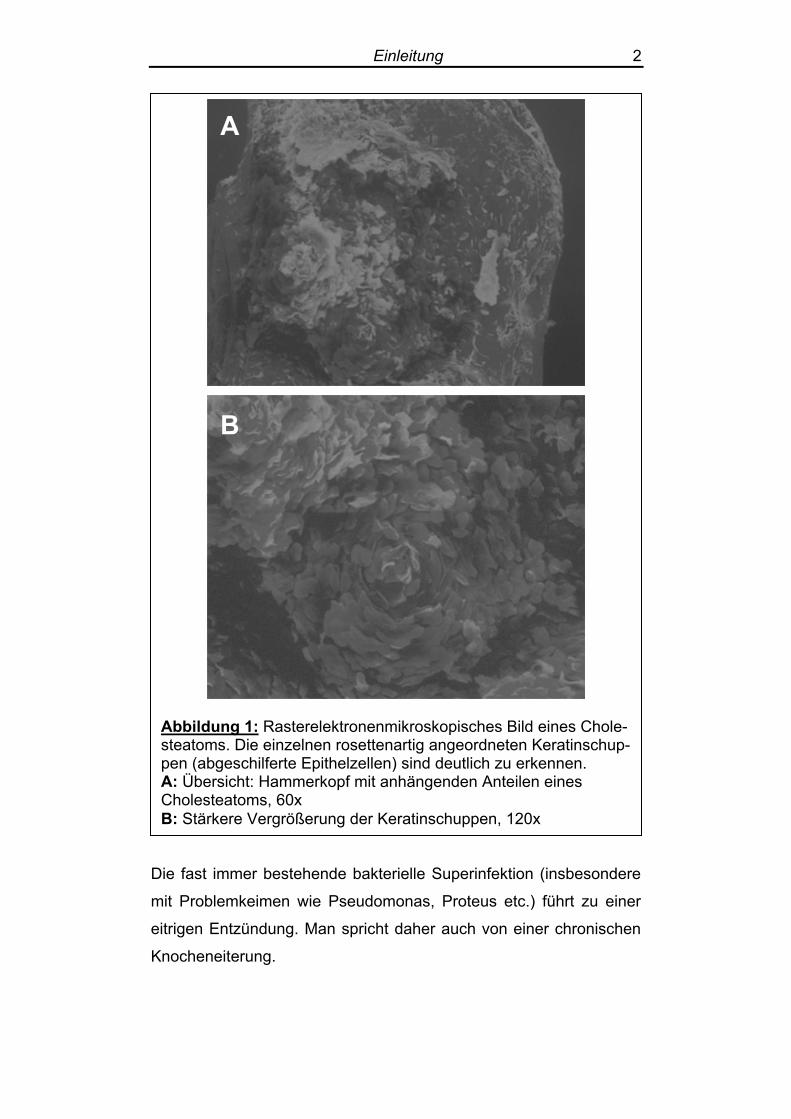
ten“ (Zenner 1997) (Abbildung 1).
Einleitung 2
Abbildung 1: Rasterelektronenmikroskopisches Bild eines Chole-steatoms. Die einzelnen rosettenartig angeordneten Keratinschup-pen (abgeschilferte Epithelzellen) sind deutlich zu erkennen. A: Übersicht: Hammerkopf mit anhängenden Anteilen eines Cholesteatoms, 60x B: Stärkere Vergrößerung der Keratinschuppen, 120x
B
A
Die fast immer bestehende bakterielle Superinfektion (insbesondere
mit Problemkeimen wie Pseudomonas, Proteus etc.) führt zu einer
eitrigen Entzündung. Man spricht daher auch von einer chronischen
Knocheneiterung.

Einleitung 3
1.2 Pathogenese des Cholesteatoms
Zur Pathogenese des Cholesteatoms existieren verschiedene Theo-
rien (Sudhoff et al. 1999). Generell kann unterschieden werden zwi-
schen kongenitalen (angeborenen) und erworbenen Cholesteatomen
(siehe 1.2.1).
Ein Cholesteatom entsteht in aller Regel dann, wenn Plattenepithel-
zellen in das Cavum tympani (sog. „Paukenhöhle“, Mittelohr) gelan-
gen.
Im Normalfall ist die Paukenhöhle von einem einschichtigen platten
bis kubischen Epithel ausgekleidet. Gelangt verhornendes Platten-
epithel in den Mittelohrraum (sei es aus dem äußeren Gehörgang,
durch Metaplasie oder durch Versprengung embryonaler Mesen-
chymreste), so beginnt dieses aus noch ungeklärten Gründen
unkontrolliert zu wachsen. Die so entstandene „Perle“ aus zwiebel-
schalenartig angeordneten verhornenden Plattenepithelzellen wächst
weiter und bildet die Cholesteatommatrix. Diese wird von der Chole-
steatomperimatrix unterschieden, die durch das subepitheliale Binde-
gewebe gebildet wird.
Im Bereich der Perimatrix kommt es zu einer chronisch inflammatori-
schen Reaktion. Im Rahmen dieser Entzündung kann es zum osteo-
klastären Abbau insbesondere der Gehörknöchelchen, aber auch der
angrenzenden Felsenbeinstrukturen kommen.
1.2.1 Einteilung der Cholesteatome
1.2.1.1 Klassische Einteilung
Aufgrund ihrer grundsätzlich unterschiedlichen Pathogenese unter-
scheidet man zwei verschiedene Gruppen von Cholesteatomen:
(1) Kongenitale (angeborene) Cholesteatome
(2) Erworbene Cholesteatome

Einleitung 4
Zu (1):
Kongenitale Cholesteatome sind definiert als Cholesteatome hinter
einem intakten Trommelfell ohne zurückliegende Infektionen oder
ohrchirurgische Eingriffe (Derlacki et al. 1965). Zur Entstehung kon-
genitaler Cholesteatome bestehen verschiedenste Theorien (Sudhoff
et al. 1999). Das Trommelfell bleibt intakt, es handelt sich um okkulte
Cholesteatome. Das ektopische Plattenepithel verhält sich meist
längere Zeit stumm, kann aber zu denselben zum Teil lebensbedroh-
lichen Komplikationen führen wie das erworbene Cholesteatom.
Diese Hypothese wurde erstmals von Habermann 1888 aufgestellt.
Zu (2):
Die erworbenen oder sekundären Cholesteatome treten weitaus
häufiger auf. Zu ihrer Pathogenese existieren ebenfalls verschiedene
Theorien, die teilweise sehr kontrovers diskutiert werden. Die vier
bedeutendsten Theorien sollen hier kurz dargestellt werden:
1. Aufgrund von Tubenventilationsstörungen bilden sich Retrak-
tionstaschen oder Invaginationen des Trommelfells, in denen sich
Keratinozyten von der Außenfläche des Trommelfells ansammeln
und so ein Cholesteatom bilden können (Wittmaack 1933). Am häu-
figsten betroffen ist die Pars flaccida, da sie der Teil des Trommel-
fells ist, der am wenigsten unter Spannung steht und somit die
größte Compliance aufweist.
2. Plattenepithel des äußeren Gehörgangs wächst durch eine Trom-
melfellperforation in die Pauke vor (Habermann 1888, Bezold 1890).
Die Trommelfellperforation kann dabei entweder entzündlich durch
eine chronische Otitis media oder traumatisch entstehen. Trauma-
tisch kann ein Cholesteatom auch durch Verschleppung von Platten-
epithel z.B. infolge von Schädelfrakturen entstehen.
3. Durch den Reiz einer chronischen Entzündung kommt es zu einer
Metaplasie von Zellen der Mittelohrschleimhaut zu Plattenepithelzel-
len (von Tröltsch 1864, Wendt 1873, Sadé 1971); diese Theorie ist
heute sehr umstritten.

Einleitung 5
4. Sporadisches papilläres Tiefenwachstum von Plattenepithelzellen
von der Außenseite des Trommelfells durch die Basalmembran führt
zur Bildung von kleinen „Keratinozytenzysten“, die sich zu einem
Cholesteatom ausdehnen können (Lange 1925, Ruedi 1978).
In den meisten Fällen wird sicherlich eine Kombination dieser
Theorien die Pathogenese des Cholesteatoms am besten erklären.
1.2.1.2 Neuere Klassifikation
Tos nahm 1993 eine neuere Klassifikation des Cholesteatoms vor,
wobei er sich mehr nach der Lokalisation des Prozesses richtete. Er
unterschied:
1. Attic-Cholesteatom
2. Sinus-Cholesteatom
3. Tensa-Cholesteatom
Das Attic-Cholesteatom wurde definiert als eine unübersichtliche
Retraktion der Pars flaccida (=Schrapnell-Membran), die sich in Attic
oder Aditus und evtl. auch in Antrum, Mastoid oder Mittelohrraum
ausdehnt.
Das Sinus-Cholesteatom bezeichnet eine posterior-superior gelege-
ne Retraktion oder Perforation der Pars tensa, die sich in den Sinus
tympanicus, die posterioren Anteile des Tympanons oder noch weiter
ausdehnt.
Unter dem Tensa-Cholesteatom versteht man eine Retraktion oder
Adhäsion der gesamten Pars tensa, die tympanale Öffnung der Eu-
stachischen Röhre mitbetreffend. Das Tensa-Cholesteatom kann
auch noch weiter in den Attic-Raum vordringen.
1.3 Klinisches Erscheinungsbild des Cholesteatoms
Primäre Cholesteatome können klinisch lange Zeit unauffällig bleiben
und sich erst durch auftretende Komplikationen bemerkbar machen.

Einleitung 6
Klassischerweise tritt ein Cholesteatom jedoch durch zunehmende
Schalleitungsschwerhörigkeit in Verbindung mit schleimig-eitriger,
fötider Otorrhoe in Erscheinung.
Oft berichten Patienten auch über ein periaurikuläres Druckgefühl.
Auch über Tinnitus und Halbseitenkopfschmerz wird gelegentlich
geklagt. Zusätzliches Auftreten eines oder mehrerer Symptome wie
Schwindel, Erbrechen, Ertaubung, Fazialisparese, Fieber, Schüttel-
frost oder Benommenheit muß als Alarmsignal für eine akute
Komplikation gewertet werden.
Ohrmikroskopisch können Cholesteatome sehr unterschiedlich in
Erscheinung treten: Die Bandbreite reicht von einer weißlichen
„Perle“, die durch das intakte Trommelfell hindurchschimmert im
Falle eines primären Cholesteatoms über unübersichtliche Retrak-
tionstaschen bis hin zu schuppig belegten randständigen Trommel-
fellperforationen. Sekundäre Cholesteatome gehen fast immer mit
einer schmierig-eitrigen, fötiden Otorrhoe einher.
1.4 Typische Komplikationen des Cholesteatoms
Die Komplikationsmöglichkeiten des Cholesteatoms sind sehr
schwerwiegend und kommen durch seine Tendenz zustande, den
umliegenden Knochen zu arrodieren bzw. zu resorbieren. Ein
weiterer wichtiger Faktor ist die stets vorhandene bakterielle
Superinfektion, häufig durch Problemkeime. Das Cholesteatom kann
durch Destruktion der Gehörknöchelchenkette zu einer Schallei-
tungsschwerhörigkeit führen, durch Fistelbildung zu einer Schädi-
gung des Innenohres, durch Zerstörung des knöchernen Fazialis-
kanals zu einer Fazialisparese, bei Einbruch in das Labyrinth zu
einer Labyrinthitis bzw. ausgeprägter Schwindelsymptomatik oder zu
einem Verlust des Hörvermögens. Weiterhin kann es durch eine
Arrosion des Sinus sigmoideus zu einer Sinusthrombose und
Thrombophlebitis führen. Ein Durchbrechen in die mittlere bzw.
hintere Schädelgrube kann zu einer Meningitis, Enzephalitis, Hirn-

Einleitung 7
und Kleinhirnabszessen führen. Durch ein Einschleppen von Keimen
in die Blutbahn kann es zu einer Sepsis kommen.
Solche bedrohlichen Komplikationen des Cholesteatoms entstehen
zwar meist erst nach längerem Krankheitsverlauf. Sie lassen sich
jedoch nur durch ein rasches und entschlossenes therapeutisches
Vorgehen verhindern.
1.5 Therapiemöglichkeiten des Cholesteatoms
Eine medikamentöse Therapie des Cholesteatoms ist derzeit nicht
möglich. Aufgrund der drohenden schweren Komplikationen ist eine
operative Sanierung absolut indiziert. Hierbei ist besondere Sorgfalt
auf die vollständige Entfernung des Cholesteatomsackes und des
entzündeten Knochens aus Mittelohr und Mastoid zu verwenden.
Gelingt dies nicht, ist mit hoher Wahrscheinlichkeit ein Rezidiv zu
erwarten.
In vielen Fällen ist eine Wiederherstellung bzw. zumindest eine
Verbesserung des Hörvermögens durch eine Tympanoplastik mög-
lich, wenn die Innenohrfunktion noch nicht nachhaltig beeinträchtigt
ist.
1.6 Knochenabbau durch das Cholesteatom
Zur genauen Pathophysiologie des Knochenabbaus durch das
Cholesteatom existieren verschiedene Theorien. Ging man früher
noch von einer Drucknekrose bzw. Atrophie des Knochens aus
(Erdheim 1905, Grünwald 1910, Rüedi 1957, Tumarkin 1958), misst
man heute der begleitenden Entzündungsreaktion die größte Bedeu-
tung zu. Ein Zusammenspiel verschiedenster Zytokine ist an dieser
Reaktion beteiligt (Amling et al. 1996, Ahn et al. 1990, Bujía et al.
1996, Chung et al. 1998, siehe auch 1.7.2). Den wichtigsten patho-
physiologischen Faktor stellt die entzündungsbedingte Chemotaxis

Einleitung 8
und Aktivierung von Osteoklasten dar, die den Abbau des
umliegenden Knochens bewirken und somit die bereits erwähnten
Komplikationen verursachen können (Chole 1984 + 1988).
1.7 Biologie des Osteoklasten
1.7.1 Funktion von Osteoblasten und Osteoklasten
Die Entwicklung des Skelettsystems, die Reparatur von Knochenbrü-
chen und die Umstrukturierung der Knochensubstanz im Sinne einer
biomechanischen Anpassung an veränderte Beanspruchungen
(„Remodeling“) sind essentielle Ansprüche, die der Knochen neben
seinen mechanischen Stütz- und Haltefunktionen erfüllen muß. Hinzu
kommt eine weitere sehr wichtige Aufgabe des Skelettsystems: Es
dient dem Körper als gigantischer Kalzium- und Phosphatspeicher.
Um all diese Aufgaben zu erfüllen, ist ein ständiger Umbau der
Knochen notwendig. Von außen kaum sichtbar finden stetige An-
und Abbauvorgänge statt. Die Zellen, die diese spezifischen
Aufgaben leisten, sind Osteoblasten und Osteoklasten. Während die
Osteoblasten für den Aufbau von Knochensubstanz und den Einbau
von Kalzium und Phosphat zuständig sind, ist die Aufgabe der
Osteoklasten der Abbau von Knochensubstanz. Sie setzen dabei
nicht nur Kalzium und Phosphat aus dem Knochen frei (Deminerali-
sation des Knochens), sondern sind auch in der Lage, die Knochen-
grundsubstanz, die vornehmlich aus Kollagen Typ I besteht, abzu-
bauen.
Eine Störung der Funktion von Osteoblasten und / oder Osteoklasten
bedingt entweder eine übermäßige Knochendichte (Osteopetrose,
Osteosklerose) oder einen Verlust von Knochenmasse (Osteopenie,
Osteoporose).
1.7.2 Morphologische Grundlagen
Osteoklasten sind terminal differenzierte, hochspezialisierte, amöbo-
id bewegliche vielkernige Riesenzellen und haben eine Größe von

Einleitung 9
10-100 µm. Typischerweise liegen sie in Einbuchtungen, den soge-
nannten Howship’schen Lakunen, die durch ihre knochenresorbie-
rende Tätigkeit entstanden sind. Sie besitzen ein stark basophiles
granuliertes Zytoplasma mit unterschiedlich großen Vakuolen. Die
bis zu 30 (oder sogar mehr) Zellkerne zeigen ein inhomogenes
Muster unterschiedlicher Größe, Form und Basophilie. Am apikalen
(dem Knochen zugewandten) Pol der Zelle ist die Plasmamembran
stark gefaltet und bildet einen Bürstensaum („ruffeled border“),
umrandet von einer dem Knochen sehr eng anliegenden Zone
(„sealing zone“). Hierbei handelt es sich um den resorbierenden Teil
der Zelle. Abgedichtet von der sealing zone dient der Bürstensaum
der Sekretion von knochenabbauenden Enzymen, Protonen, Prote-
inen etc., sowie der Resorption abgebauter Knochenbestandteile.
Die Zelle bildet so ein von der Umgebung abgeschlossenes
Resorptionskompartiment, in dem ein extrem saures Milieu herrscht
(Abbildung 2).
Abbildung 2: Schematische Darstellung eines Osteoklasten in einer Resorptionslakune („Howshipsche Lakune“).
Sealing Zone
Resorption
Sekretion
Knochen
Resorptionskompartiment
Ruffeled border
Nuklei
Sealing Zone
Osteoklasten entstehen durch die asynchrone Fusion von mononu-
kleären Vorläuferzellen aus dem Knochenmark (Nijweide 1986). Ihre
Differenzierung erfolgt aus der Monozyten-Makrophagen Linie

Einleitung 10
(Chambers 1985, 1988, 2000, Prallet 1992, Amling et al. 1996).
Insgesamt ist nur etwa 1% der Knochenoberfläche mit Osteoklasten
bedeckt, es handelt sich also um eine verhältnismäßig kleine Zellpo-
pulation. Die Reifung der Osteoklasten umfasst vier Stadien:
Proliferation, Differenzierung, Fusion und Aktivierung. Im Prolifera-
tionsstadium sind die Zellen monozytisch, teilen sich aktiv und
exprimieren keine Tartratresistente Saure Phosphatase (TRAP). In
der Differenzierungsphase hören die Zellen auf, sich zu teilen und
werden TRAP-positiv. Die Zellen wachsen und fusionieren in der
Fusionsphase zu mehrkernigen Riesenzellen. In der folgenden
Aktivierungsphase bildet sich die „Ruffeled Border“ und die Zellen
beginnen mit der Knochenresorption. Es ist bekannt, daß viele
Zytokine wie zum Beispiel Makrophagen-Koloniestimulierender
Faktor (M-CSF), Interleukin-1, Interleukin-6, Tumornekrosefaktor-α
(TNF-α), Transformierender Wachstumsfaktor-β (TGF-β), Osteokla-
sten-Differenzierungsfaktor (ODF, RANKL, OPGL), Prostaglandine
und Leukotriene die osteoklastäre Knochenresorption steigern
(Amling et al. 1996, Ahn et al. 1990, Bujía et al. 1996, Chung et al.
1998).
Biochemisch kennzeichnend für Osteoklasten ist die Expression von
Calzitoninrezeptoren (CTR), des Vitronektinrezeptors (ανβ3), einer
Protonenpumpe (H+ ATPase), sowie die Bildung von Carboanhydra-
se II und tartratresistenter saurer Phosphatase (TRAP) (Prallet et al.
1992).
1.8 Fragestellung und Zielsetzung
In der vorliegenden Arbeit soll der Knochenabbau im Cholesteatom
rasterelektronenmikroskopisch dargestellt werden. Hierzu sollen im
Rahmen cholesteatomsanierender ohrchirurgischer Eingriffe entnom-
mene Gehörknöchelchen untersucht und die Morphologie des Kno-
chenabbaus dargestellt werden.

Einleitung 11
Wie bereits in der Literatur nachzulesen (z.B. Chole et al. 1984,
Richardson et al. 1993), wird der Knochenabbau im Cholesteatom
mit großer Wahrscheinlichkeit durch die Aktivierung knochenabbau-
ender Zellen (Osteoklasten) bewirkt. Es soll in der vorliegenden
Arbeit nach morphologischen Anhaltspunkten für die Ursache des
cholesteatombedingten Knochenabbaus gesucht werden.
Zur Therapie des Cholesteatoms stehen bisher nur chirurgische
Maßnahmen zur Verfügung. Diese sind oft sehr aufwendig und mit
hohen Komplikations- und Rezidivraten behaftet. In der vorliegenden
Arbeit sollen daher Möglichkeiten zur medikamentösen Hemmung
des Knochenabbaus im Cholesteatom untersucht werden.
Der Einfluss und die Wirkungsweise der Substanzen Calzitonin und
Zoledronat auf Osteoklasten in vitro sollen untersucht und verglichen
werden.
Das Polypeptidormon Calzitonin dient dem Körper zur Modulation
des Kalziumhaushalts. Es bewirkt eine Hemmung der Kalziumfreiset-
zung aus dem Knochen. Dabei wirkt es über den Calzitoninrezeptor
direkt hemmend auf Osteoklasten.
Zoledronat ist ein Bisphosphonat der dritten Generation. Ältere
Bisphosphonate werden bereits zur Therapie der Osteoporose und
anderer mit Knochenabbau einhergehender Erkrankungen einge-
setzt. Ihre Wirkungsweise ist nicht vollständig geklärt. Sie scheinen
ebenfalls eine Hemmung der Osteoklastentätigkeit zu bewirken.
Durch die Untersuchung der Wirkungsweise der erwähnten Pharma-
ka sollen mögliche zusätzliche Therapieoptionen für Cholesteatome
aufgezeigt werden, die das chirurgische Vorgehen erleichtern und
die Komplikations- und Rezidivrate verringern könnten.

Material und Methoden 12
2. Material und Methoden
2.1 Ossikel
2.1.1 Präparate
Zur morphologischen Darstellung des Knochenabbaus im Chole-
steatom wurden 46 Gehörknöchelchen untersucht. 40 dieser Ossikel
stammten von 32 an Cholesteatomen erkrankten Patienten, die in
der Zeit von 1997-1999 an der HNO-Klinik der Universität Bochum
am Mittelohr operiert wurden. Das Alter der Patienten lag zwischen 6
und 86 Jahren, durchschnittlich bei 39,5 Jahren, die Geschlechtsver-
teilung ergab ein Verhältnis M:W von ca. 1:1.
Zum Vergleich wurden 6 Gehörknöchelchen aus Felsenbeinen nicht
am Ohr erkrankter Leichen untersucht. Hierbei handelte es sich um
jeweils 3 Hammer und 3 Ambosse. Bei den Gehörknöchelchen der
an Cholesteatomen erkrankten Patienten handelte es sich um 17
Hammer/Hammerköpfe und 23 Ambosse. Die Präparate wurden
direkt nach der Entnahme in Formalin oder Glutaraldehyd fixiert. Ein
Teil der Gehörknöchelchen wurde daraufhin zur Ablösung von
Weichteilstrukturen und Schleimhaut zunächst für 24 Stunden in
Natrium-Hypochlorid eingelegt (Wong et al. 1991). Danach erfolgte
die Aufbereitung für die rasterelektronenmikroskopische Untersu-
chung wie unter Punkt 2.4 beschrieben.
2.1.2 Untersuchung der Präparate
Sämtliche Präparate wurden rasterelektronenmikroskopisch bei
ansteigenden Vergrößerungen untersucht, wobei besonderer Wert
auf die Morphologie des Knochenabbaus gelegt wurde. Sämtliche
Auffälligkeiten und Befunde wurden fotografisch dokumentiert
(„Agfachrome RSX 200“ Diafilme) und archiviert.

Material und Methoden 13
2.2 Lichtmikroskopie
2.2.1 Färbung
2.2.1.1 Zellkulturen
Für die lichtmikroskopischen Untersuchungen an den Zellkulturen
wurden TRAP-Färbungen angefertigt. TRAP (Tartrate Resistent Acid
Phosphatase, Tartratresistente saure Phosphatase) ist ein Enzym,
das von ausgereiften Osteoklasten gebildet wird. In der TRAP-
Färbung stellt es sich braun dar und erlaubt so die Identifikation von
Osteoklasten. Es wurde das „Leucocyte Acid Phosphatase Kit“ von
Sigma Diagnostics, St. Louis, USA verwendet.
Zur Darstellung apoptotischer Zellen diente die TUNEL-Färbung
(TdT-mediated UTP nick end labelling, In Situ Cell Death Detection
Kit, Boehringer, Mannheim), in der sich apoptotische Zellen dunkel
anfärben.
2.2.1.2 Schädelknochenbiopsien
Die Schädelknochenbiopsien wurden nach der Fixierung zunächst in
0.35 mol/l Na-EDTA (pH 7.4) für mehrere Tage demineralisiert, dann
in aufsteigenden Acetonlösungen dehydriert und in EPON-Araldite
eingebettet. Drei 2µm Schnitte wurden mindestens 100µm voneinan-
der entfernt von jedem Präparat mit Hilfe eines Reichert Ultramicro-
toms angefertigt. Die Präparate wurden mit Toluidinblau gefärbt und
histologisch ausgewertet, wobei die Osteoklasten gezählt wurden.
2.2.2 Mikroskop/Dokumentation
2.2.2.1 Dokumentation der Zellkulturen
Die Untersuchung der Zellkulturpräparate erfolgte mit unserem
Zellkulturmikroskop (Olympus IMT 2). Von jedem Präparat wurden
bei identischer Vergrößerung, Blende und Belichtung fünf Bilder von
unterschiedlichen, zufällig ausgewählten Regionen der Kulturplatte
digital aufgenommen (Digitalkamera Olympus DP 50). Die digitale
Auswertung erfolgte mit Hilfe des Bildanalysesystems Zeiss Kontron
KS 300 wie unter Punkt 2.4.3.3 beschrieben.

Material und Methoden 14
2.3 Elektronenmikroskopie
2.3.1 Vorbereitung der Präparate
Nach der Entnahme wurden die Präparate direkt für mindestens 24
Stunden in Formalin oder Glutaraldehyd fixiert.
2.3.2 Critical Point Trocknung
2.3.2.1 Prinzip
Das Prinzip der Critical Point Trocknung besteht darin, daß sämt-
liches Wasser in einem Präparat durch eine Ersatzflüssigkeit ersetzt
wird. Diese wird daraufhin langsam in einen gasförmigen Zustand
gebracht und so dem Präparat auf schonende Weise wieder
entzogen.
Hierfür eignet sich insbesondere Kohlendioxid (CO2), da es sich
unterhalb von 31°C durch Druckerhöhung zu einer Flüssigkeit
verdichten läßt, die durch eine geringe Erhöhung der Temperatur
wieder zum Verdampfen gebracht werden kann.
2.3.2.2 Critical Point Trocknung der Präparate
Die Präparate wurden zunächst für 10 Minuten in 0,1 molarer PBS-
Lösung gewaschen und dann in einer aufsteigenden Alkoholreihe
dehydriert: Jeweils fünfminütiges Schwenken in Ethanolkonzentratio-
nen von 30%, 50%, 75%, 96% und 2mal hintereinander 100%.
Sodann wurden die Präparate in unserem „Balzers Union Critical
Point Dryer“ (Balzers Union Limited, Accessories for Electron
Microscopy) getrocknet: Die dehydrierten Präparate wurden in die
auf ca. 10 °C vorgekühlte Probenkammer gegeben. Diese wurde
unter hohem Druck mit flüssigem Kohlendioxid gefüllt. Nach
sorgfältigem Durchmischen mit einem Magnetrührer wurde das
Gemisch langsam auf 42°C erhitzt. Dadurch verdampfte das CO2
und konnte langsam abgelassen werden. Den Präparaten wurde so
sämtliche Flüssigkeit entzogen.

Material und Methoden 15
Die getrockneten Präparate wurden auf Probentellerchen geklebt
und zur Bedampfung mit Gold in unseren „Cressington 108auto
Sputter Coater“ gegeben.
2.3.3 „Sputtern“ der Präparate
2.3.3.1 Prinzip
Für die Elektronenmikroskopie müssen die Präparate mit einer
leitfähigen Oberfläche versehen werden. Hierfür werden die Proben
üblicherweise durch einen Induktionsstrom, der Moleküle aus einem
„Target“ herauslöst, mit Gold bedampft. Dieses Verfahren erlaubt es,
eine sehr dünne leitfähige Schicht auf das Gewebe zu bringen, ohne
wichtige Strukturen zu zerstören oder zu verdecken.
2.3.3.2 Sputtervorgang
Es wurde unser Bedampfungsgerät „Cressington 108auto Sputter
Coater“ verwendet. Die Präparate wurden in die Probenkammer des
Gerätes gegeben. Dann wurde zunächst ein Vakuum erzeugt und die
Kammer mit Argon, dem „Prozeßgas“, geflutet, so daß eine Argon-
Atmosphäre mit einem Druck von ca. 0.1 mbar entstand.
Durch einen Sputterstrom von 30 mA wurden aus dem Target (einem
massiven Gold-Block) Moleküle herausgelöst und auf die Oberfläche
des Präparates übertragen. Auf diese Weise bildete sich eine feine,
leitfähige Gold-Schicht auf dem Präparat.
Die so aufbereiteten Präparate wurden mit unserem Raster-Elektro-
nenmikroskop (Stereoscan 250, Cambridge Instruments) untersucht.

Material und Methoden 16
2.4 Zellkulturen
2.4.1 Gewinnung der Zellen
Zur Züchtung von Osteoklasten in vitro wurden Knochenmarkszellen
aus Tibia und Femur von 3-6 Wochen alten männlichen Mäusen
gewonnen.
Die Tiere wurden hierzu zunächst mit Ether betäubt und durch zer-
vikale Dislokation getötet. Daraufhin wurden Tibia und Femur her-
auspräpariert und in einer sterilen Schale auf Eis gesammelt. Für
jeden Versuchsansatz wurden die Knochen von drei Mäusen ver-
wendet.
Die Knochenmarkszellen wurden gewonnen, indem die Röhren-
knochen mit Hilfe einer Spritze mit α-MEM (minimal essential me-
dium) gespült wurden. Die zum Teil verklumpten Knochenmarks-
zellen wurden durch einen Cell Strainer gefiltert und vorsichtig über
Ficoll Hypaque Trennlösung pipettiert. Daraufhin wurden die Zellen
bei 1500 U/min für 15 min zentrifugiert. Aufgrund ihrer unterschied-
lichen Dichte sammelten sich Zellen unterschiedlichen Typs an
verschiedenen Stellen im Reagenzglas und so konnte die Mono-
zyten/Makrophagen-Fraktion selektiv pipettiert werden.
2.4.2 Kultivierung der Zellen
Die so gewonnenen Monozyten und Makrophagen wurden in 24-Well
Zellkulturplatten in α-MEM mit Zusatz von 10% fetalem Kälberserum
(FCS) und Antibiotika bei 37°C in einer Atmosphäre von 95% Luft
und 5% CO2 kultiviert.
Zur Stimulation der Osteoklastogenese wurde den Kulturen 30ng/ml
M-CSF (Macrophage Colony stimulating Factor) und 50ng/ml OPGL
(Osteoprotegerin-Ligand / Osteoklasten-Differenzierungsfaktor) zu-
gegeben. Medium und sämtliche Faktoren wurden jeden zweiten Tag
gewechselt.

Material und Methoden 17
2.4.3 Studien mit Zoledronat und Calzitonin in vitro
Um die Wirkung von Zoledronat bzw. Calzitonin in vitro zu untersu-
chen, wurden diese Substanzen den Zellkulturen in unterschiedlicher
Konzentration jeweils mit dem Mediumwechsel zugegeben.
2.4.3.1 Zoledronat
Zoledronat wurde den Zellkulturen jeden zweiten Tag in Konzentra-
tionen von 10-6 – 10-9 mol/l zugegeben. Zusätzliche Kontrollkulturen
wurden unter Zusatz von EDTA (Äthylendiamintetraessigsäure)
inkubiert, um eine Auswirkung des Calzium-komplexierenden
Effektes von Bisphosphonaten auszuschließen. Die Kulturen wurden
für 10 Tage inkubiert. Dann wurden die Zellen fixiert und TRAP-
Färbungen (siehe 2.2.1.1) durchgeführt. Danach erfolgte die
mikroskopische Auswertung.
2.4.3.2 Calzitonin
Calzitonin wurde den Zellkulturen jeden zweiten Tag in Konzentratio-
nen von 10-6 – 10-10 mol/l zugegeben. Die Kulturen wurden ebenfalls
für 10 Tage inkubiert, dann fixiert und TRAP-Färbungen durchgeführt
(siehe 2.2.1.1). Danach erfolgte die mikroskopische Auswertung.
2.4.3.3 Auswertung der Zellkulturen
Zur Auswertung der Zellkulturen wurden mit Hilfe einer digitalen
Kamera (Olympus DP 50) von jedem Präparat bei identischer
Vergrößerung, Blende und Belichtung fünf Fotos von unterschiedli-
chen, zufällig ausgewählten Regionen der Kulturplatte aufgenom-
men.
Unter Zuhilfenahme des digitalen Bildanalysesystems Zeiss Kontron
KS 300 wurde der prozentuale Anteil der von Osteoklasten be-
deckten Fläche an der Gesamtfläche des Bildausschnittes berech-
net. Die so gewonnenen Zahlen wurden für jedes Präparat gemittelt,
so daß Durchschnittswerte angegeben werden konnten.

Material und Methoden 18
2.4.4 Dentin-Chips
Als Substrat für die Knochenresorption dienten 400µm dicke Dentin-
Chips, die mit Hilfe eines Sägemikrotoms (Leica 1600) aus Walfisch-
zähnen geschnitten wurden. Sie wurden jeweils der Hälfte der
Kulturplatten-Öffnungen zugegeben. Auf den Dentin-Chips fand in
vitro osteoklastäre Knochenresorption statt. Die Auswertung erfolgte
elektronenmikroskopisch. Zunächst wurden die Chips nach Beendi-
gung der Kulturzeit im Ultraschallbad gereinigt und von sämtlichen
Zellen befreit. Daraufhin wurden sie für die Raster-Elektronenmikro-
skopie aufbereitet (siehe 2.3).
Zur Auswertung wurden unter identischen Vergrößerungs- und Be-
lichtungsverhältnissen fünf Bilder von unterschiedlichen Regionen
jedes Präparates digitalisiert. Mit Hilfe unserer Bildanalysesoftware
Zeiss Kontron KS 300 wurde der prozentuale Anteil der Resorptions-
lakunen an der Gesamtfläche der Dentinscheibchen berechnet.
2.5 Tierexperimente
Die tierexperimentellen in-vivo-Studien wurden im Labor von Prof.
Dr. R. Chole, St. Louis, USA, durchgeführt. Bei den Versuchstieren
handelte es sich um drei bis sechs Wochen alte männliche Mäuse
(C57/BL6). Die Versuchsprotokolle wurden vom „Institutional Animal
Care and Use Committee“ (IACUC) der Washington University in St.
Louis genehmigt und die Versuche erfolgten unter Berücksichtigung
der „PHS Policy on Humane Care and Use of Laboratory Animals“,
des „NIH Guide for the Care and Use of Laboratory Animals“ und des
„Animal Welfare Act (7 U.S.C. et seq.)“.
2.5.1 Keratinimplantate
Zur Stimulation eines osteoklastären Knochenabbaus wurde ein von
Chole et al. (2001) entwickeltes Modell zur Induktion von
osteoklastärem Knochenabbau benutzt: Den Versuchstieren wurden

Material und Methoden 19
sterilisierte Keratinpartikel subperiostal auf die Schädelkalotte
implantiert.
Zur Implantation erhielten die Mäuse eine Ether-Narkose. Die Haut
über der Kalotte wurde rasiert, desinfiziert und auf einer Länge von
etwa 1,5 cm inzidiert. Der Schädelknochen wurde freipräpariert und
das Periost stumpf nach lateral verschoben. Auf beiden Seiten der
sagittalen Schädelnaht wurden ca. 10-12 mg Keratinpartikel aufge-
bracht, die Haut readaptiert und mit Wundklemmen verschlossen.
Die Tiere vertrugen die Operation gut, es gab keine Mortalität und
keine sichtbare Morbidität.
Nach Beendigung des Experimentes wurden die Tiere in Ethernarko-
se getötet, die Schädelkalotte entfernt und mit einer 5 mm Haut-
Biopsiestanze beiderseits der Sagittalnaht eine Probe entnommen,
wobei die unmittelbar an die Sagittalnaht angrenzende Region
vermieden wurde (Abbildung 3). Die so gewonnenen Präparate
wurden halbiert und in 4% Formaldehyd und 0.05% Glutaraldehyd in
0.1 mol/l Phosphatpuffer (pH 7.4) bei Zimmertemperatur fixiert.
Abbildung 3: Schematische Darstellung der Präparation der Mäu-seschädel. (Mit freundlicher Genehmigung von Prof. Chole, St. Louis)

Material und Methoden 20
2.5.2 Zoledronat in vivo
Zur Untersuchung der Wirkung von Zoledronat in vivo wurden die
Versuchstiere in sechs Gruppen zu je fünf Tieren aufgeteilt. Gruppe
I: Kontrolle 1 (nicht operierte Tiere), Gruppe II: Kontrolle 2 (Operation
ohne Keratinimplantation, keine Injektionen: „SHAM Operation“),
Gruppe III: Kontrolle 3 (Keratinimplantation, tägliche intraperitoneale
Injektionen von Ca++/Mg++-freiem PBS), Gruppe IV-VI: Studiengrup-
pen (tägliche intraperitoneale Injektionen von 1, 3, bzw. 10µg Zole-
dronat pro kg Körpergewicht). Alle Tiere wurden vier Tage vor und
fünf Tage nach der Operation behandelt.
2.5.3 Auswertung der Schädelknochenpräparate
Die Auswertung der Schädelknochenpräparate erfolgte nachdem die
Objektträger mit den Schnitten mit neutralen Codes versehen worden
waren, so dass der Untersucher nicht wusste, um welches Präparat
es sich handelte.
Osteoklasten wurden in den Präparaten identifiziert und auf der
gesamten Länge jedes 5 mm langen Schädelknochen-Präparates
gezählt. Dem Knochen anliegende Zellen konnten anhand folgender
Kriterien eindeutig als Osteoklasten identifiziert werden:
1. mehrkernige (≥ 2 Zellkerne) Zellen
2. deutliche „ruffeled border“
3. granuliertes Zytoplasma
4. im Bereich des Kontaktes mit Knochen zumindest teilweise
aufgelöste Lamina limitans
Um als Osteoklast gezählt zu werden, musste eine Zelle mindestens
drei der vier oben genannten Kriterien erfüllen (Abbildung 19, S. 41). Die statistische Auswertung (Berechnung der Anzahl Osteoklasten
pro Millimeter Knochen) wurde mit Hilfe der Statistiksoftware von
SPSS Science Inc. durchgeführt.

Material und Methoden 21
2.6 Statistiken
Die Datenanalyse erfolgte mit Hilfe der Datenanalysesoftware SPSS
9.0 von SPSS Science Inc. Alle Ergebnisse wurden als Mittelwerte
mit ihrer Standardabweichung und dem Standardfehler berechnet.
Zur Varianzanalyse wurde eine einfaktorielle ANOVA Prozedur
verwendet. Die Berechnung der Signifikanz erfolgte mit Hilfe der
Scheffé-Prozedur, α war 0,05 in allen Analysen.

Ergebnisse 22
3. Ergebnisse
3.1 Ossikel 3.1.1 Kontrollen
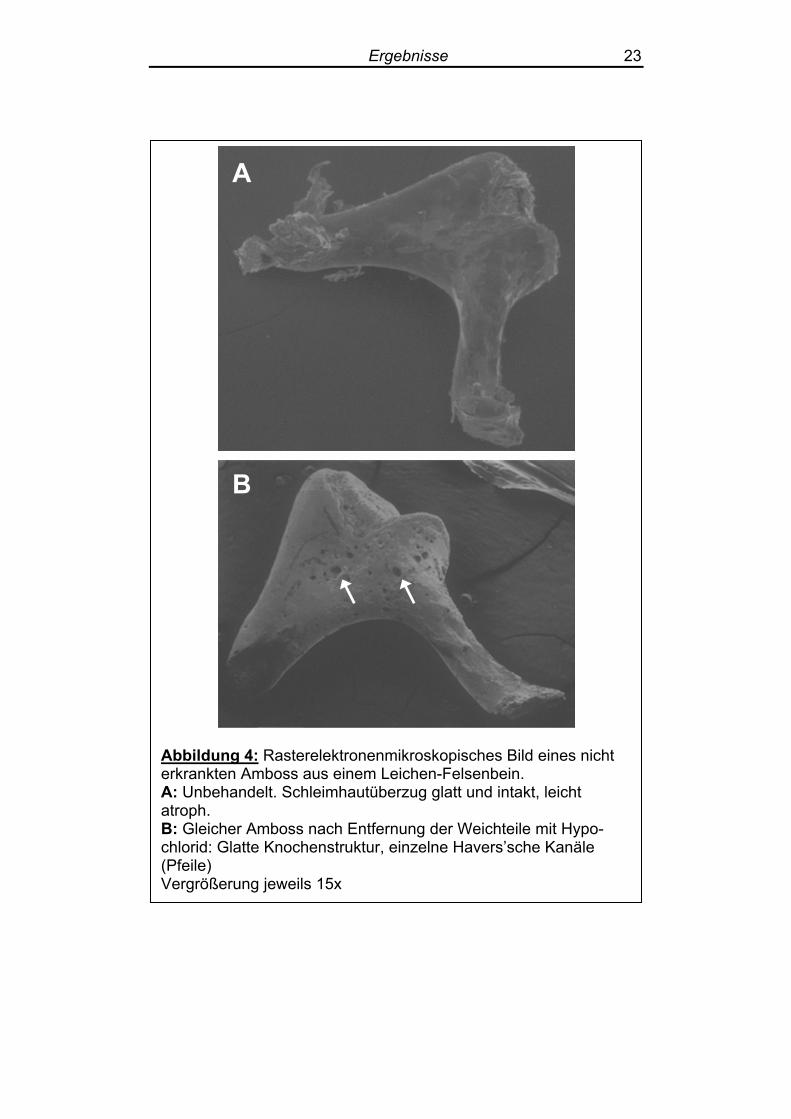
Die zur Kontrolle untersuchten gesunden Gehörknöchelchen zeigten
sich in der Raster-Elektronenmikroskopie unverletzt, mit einer allseits
glatten Oberfläche und größtenteils intakter, jedoch teils atrophischer
Schleimhaut (Abbildung 4 A). Die Schleimhautatrophie ist sicherlich
auf die Tatsache zurückzuführen, dass die Kontroll-Präparate aus
Leichen-Felsenbeinen stammten. Nach Entfernung der Weichteile
durch 24stündiges Einlegen in 0,1 molare Natriumhypochlorid-
Lösung zeigte sich eine glatte faserige Lammellenknochenstruktur,
die nur gelegentlich durch Havers’sche Kanäle unterbrochen war
(Abbildung 4 B).
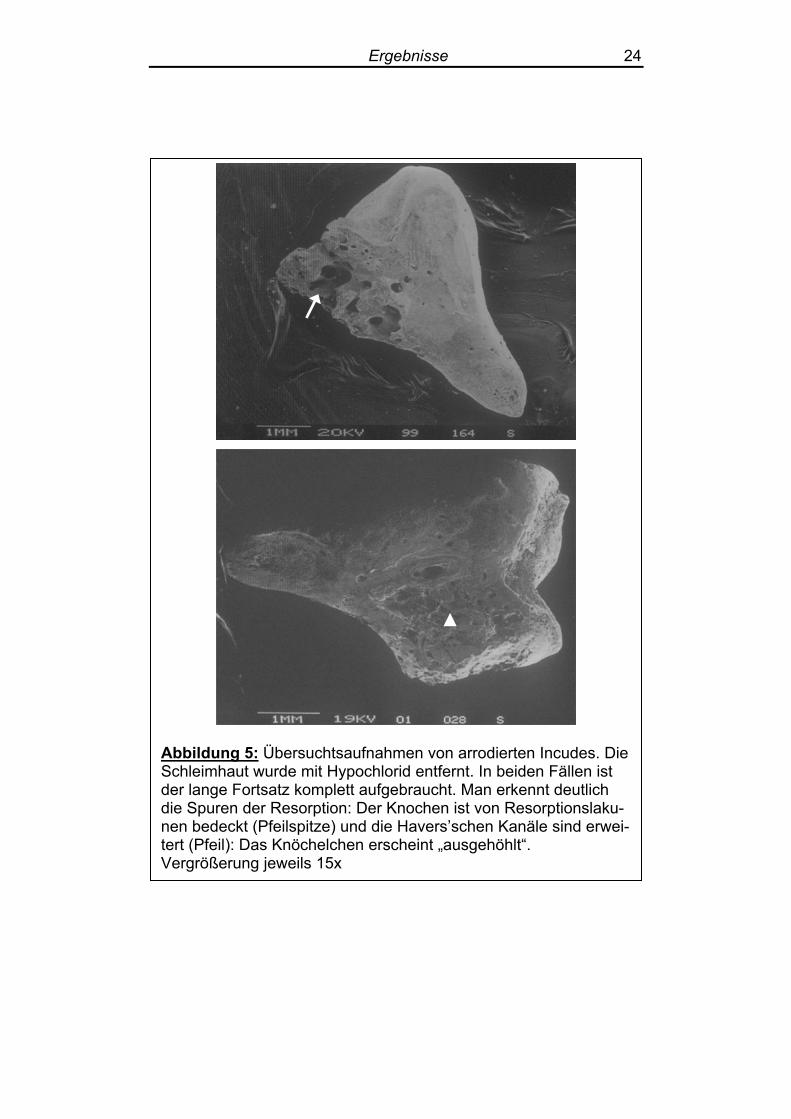
3.1.2 Knöchelchen aus Cholesteatom-erkrankten Ohren
An sämtlichen der untersuchten Gehörknöchelchen von Patienten
mit Cholesteatomen fanden sich deutliche Spuren des Knochenab-
baus (Abbildung 5). Die Schleimhaut war größtenteils intakt, stellenweise im Bereich der
Resorptionslakunen jedoch angedaut. Hier fanden sich gelegentlich
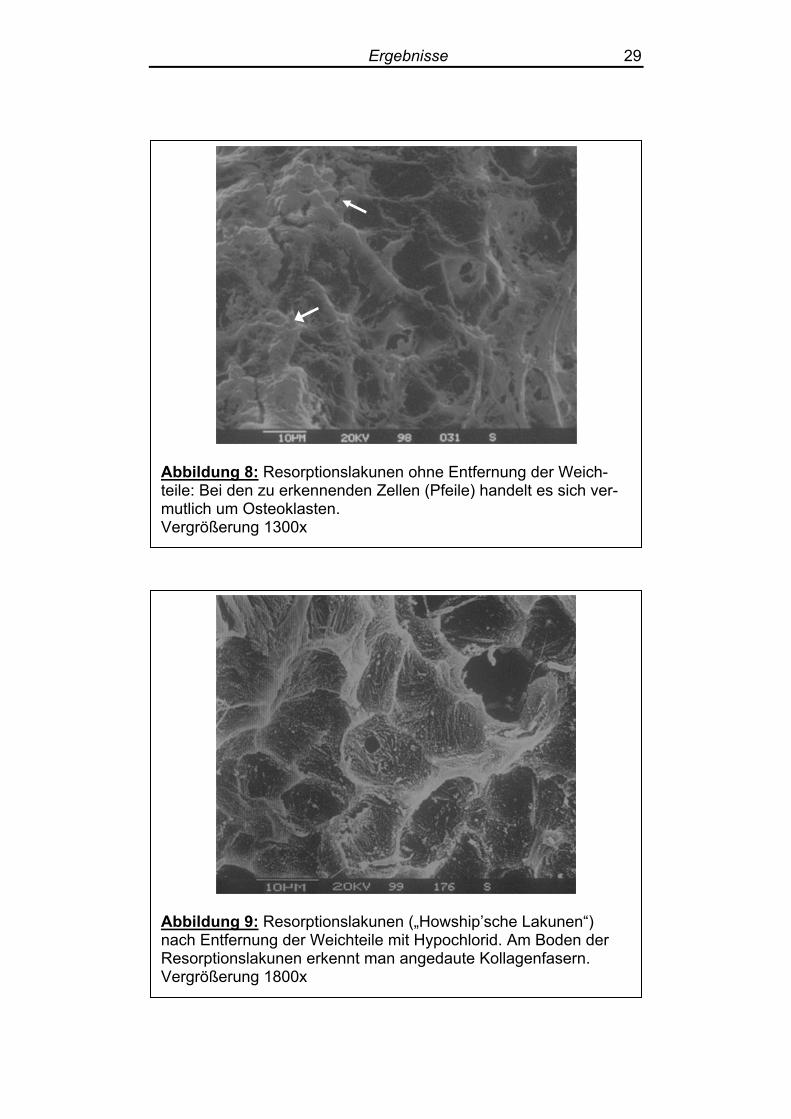
auch große, in den Resorptionslakunen gelegene Zellen, vermutlich
Osteoklasten (Abbildung 8). Auf einigen Gehörknöchelchen ließ sich die noch anhängende
Cholesteatommatrix darstellen. Deutlich zu erkennen war die schup-
pige Struktur des Cholesteatoms, hervorgerufen durch abschilfernde
Plattenepithelzellen (Abbildung 1). Im Randbereich des Cholesteat-
oms ließen sich deutlich die verschiedenen Stadien des Knochenab-
baus darstellen (siehe 3.1.3 „Morphologie des Knochenabbaus“,
Abbildung 6 + 7). Nach Entfernung der Weichteile konnte die Auswirkung des Chole-
steatoms auf die Knochenstruktur dargestellt werden.
Ergebnisse 23
Abbildung 4: Rasterelektronenmikroskopisches Bild eines nicht erkrankten Amboss aus einem Leichen-Felsenbein. A: Unbehandelt. Schleimhautüberzug glatt und intakt, leicht atroph. B: Gleicher Amboss nach Entfernung der Weichteile mit Hypo-chlorid: Glatte Knochenstruktur, einzelne Havers’sche Kanäle (Pfeile) Vergrößerung jeweils 15x
B
A
Ergebnisse 24
Abbildung 5: Übersuchtsaufnahmen von arrodierten Incudes. Die Schleimhaut wurde mit Hypochlorid entfernt. In beiden Fällen ist der lange Fortsatz komplett aufgebraucht. Man erkennt deutlich die Spuren der Resorption: Der Knochen ist von Resorptionslaku-nen bedeckt (Pfeilspitze) und die Havers’schen Kanäle sind erwei-tert (Pfeil): Das Knöchelchen erscheint „ausgehöhlt“. Vergrößerung jeweils 15x

Ergebnisse 25
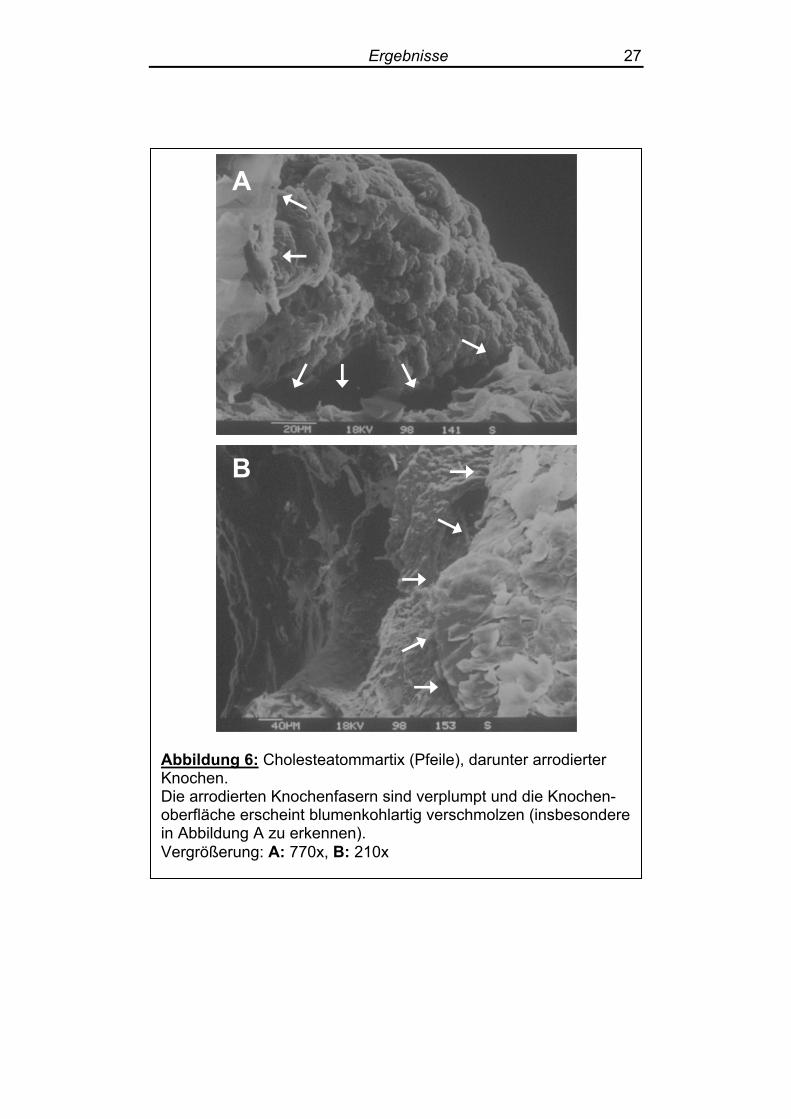
3.1.3 Morphologie des Knochenabbaus
Der Knochenabbau trat im Wesentlichen in vier verschiedenen
Formen in Erscheinung:
(1) Auflösung der regulären Kollagenstruktur
(2) Bildung von Resorptionslakunen
(3) Verschmelzung / Verklumpung von Kollagenfasern
(4) Erweiterung der Haversschen Kanäle
Zu (1):
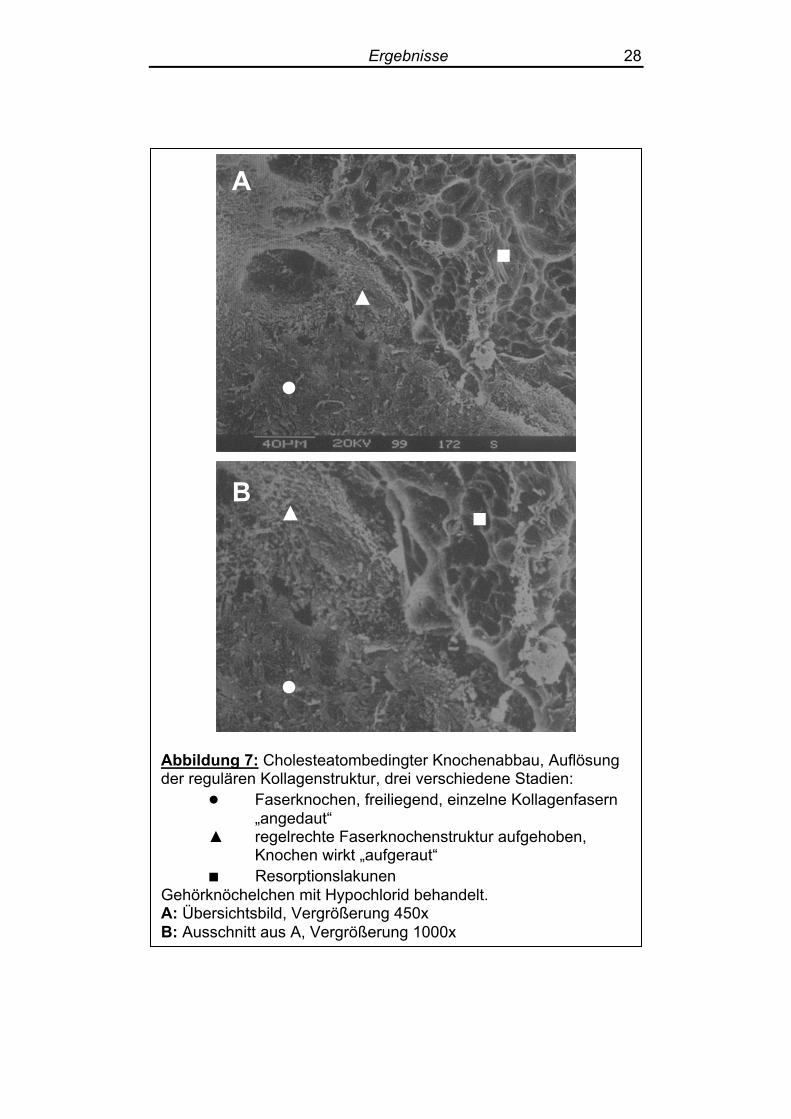
Die Auflösung der regulären Kollagenstruktur kann als Vorstufe der
Bildung von Resorptionslakunen gesehen werden. Zwei Stadien
ließen sich nebeneinander beobachten, bevor es zur Ausbildung von
Resorptionslakunen kam: Zunächst ließen sich bei insgesamt noch
intakter Faserknochenstruktur einzelne „angedaute“ Kollagenfasern
darstellen, die aus der regulären Oberflächenstruktur des Knochens
ausgelöst erschienen (Abbildung 7). Im zweiten Stadium war die reguläre Oberfläche aufgelöst, der Fa-
serknochen erschien aufgeraut (Abbildung 7).
Zu (2):
Die Ausbildung von Resorptionslakunen erfolgt über die oben be-
schriebenen Schritte. Resorptionslakunen stellten sich als rundliche,
grüppchenweise angeordnete, im Durchmesser 10-40 µm große,
konfluierende Kerben im Knochen dar (Abbildung 7-9). In seltenen
Fällen ließen sich große Zellen in den Resorptionslakunen darstellen,
bei denen es sich um Osteoklasten handeln dürfte (Abbildung 8). Bei stärkerer Vergrößerung zeigten sich am Boden der Resorptions-
lakunen einzelne arrodierte Kollagenfasern (Abbildung 9).
Zu (3):
Die Verschmelzung von Kollagenfasern war vor allem im direkten
Randbereich des Cholesteatoms zu beobachten und führte zu einer
blumenkohlartigen Verklumpung der Faserknochenoberfläche (Ab-

Ergebnisse 26
bildung 6). Die einzelnen Kollagenfasern waren hier nicht mehr
abgrenzbar.
Zu (4):
Ein weiteres sehr bedeutsames Erscheinungsbild des Knochenab-
baus im Cholesteatom ist die an sämtlichen untersuchten Gehörknö-
chelchen zu beobachtende deutliche Erweiterung der Haversschen
Kanäle (Abbildung 5). Die Gehörknöchelchen erschienen wie aus-
gehöhlt.
Ergebnisse 27
Abbildung 6: Cholesteatommartix (Pfeile), darunter arrodierter Knochen. Die arrodierten Knochenfasern sind verplumpt und die Knochen-oberfläche erscheint blumenkohlartig verschmolzen (insbesondere in Abbildung A zu erkennen). Vergrößerung: A: 770x, B: 210x
A
B
Ergebnisse 28
Abbildung 7: Cholesteatombedingter Knochenabbau, Auflösung der regulären Kollagenstruktur, drei verschiedene Stadien:
● Faserknochen, freiliegend, einzelne Kollagenfasern „angedaut“
▲ regelrechte Faserknochenstruktur aufgehoben, Knochen wirkt „aufgeraut“
■ Resorptionslakunen Gehörknöchelchen mit Hypochlorid behandelt. A: Übersichtsbild, Vergrößerung 450x B: Ausschnitt aus A, Vergrößerung 1000x
■
●
▲
A
B ■▲
●
Ergebnisse 29
Abbildung 8: Resorptionslakunen ohne Entfernung der Weich-teile: Bei den zu erkennenden Zellen (Pfeile) handelt es sich ver-mutlich um Osteoklasten. Vergrößerung 1300x
Abbildung 9: Resorptionslakunen („Howship’sche Lakunen“) nach Entfernung der Weichteile mit Hypochlorid. Am Boden der Resorptionslakunen erkennt man angedaute Kollagenfasern. Vergrößerung 1800x
Ergebnisse 30
3.2 Zellkulturen
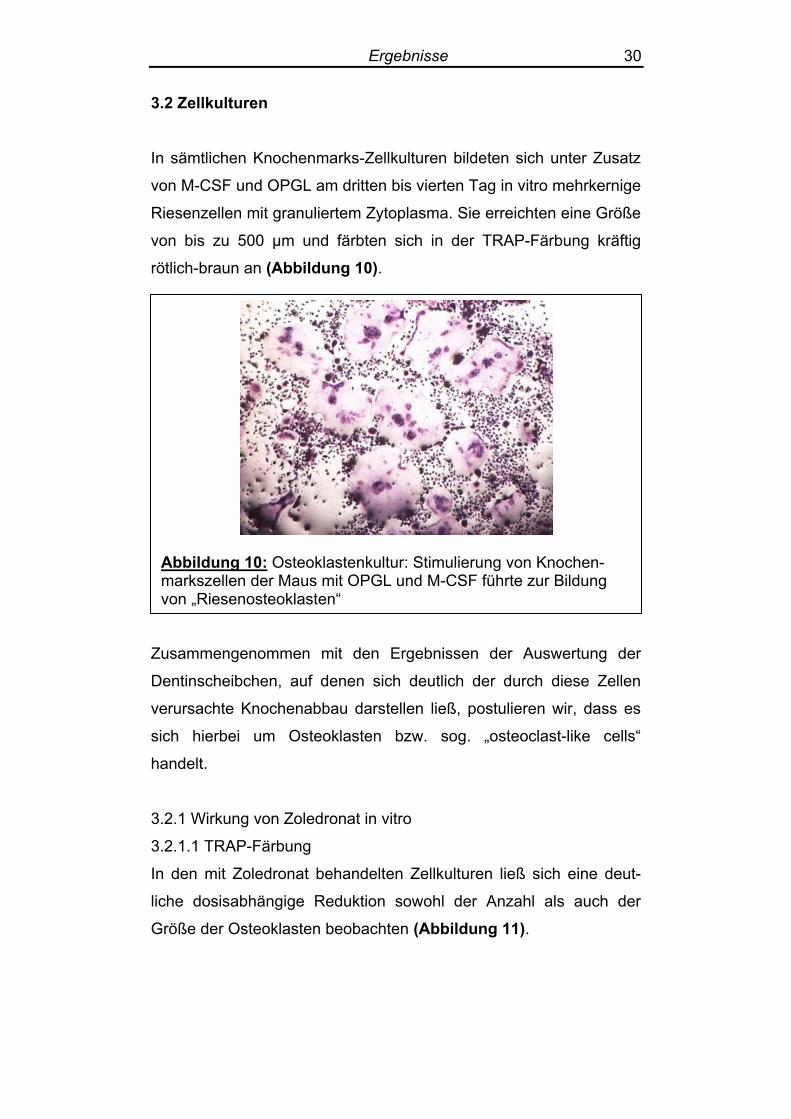
In sämtlichen Knochenmarks-Zellkulturen bildeten sich unter Zusatz
von M-CSF und OPGL am dritten bis vierten Tag in vitro mehrkernige
Riesenzellen mit granuliertem Zytoplasma. Sie erreichten eine Größe
von bis zu 500 µm und färbten sich in der TRAP-Färbung kräftig
rötlich-braun an (Abbildung 10).
Abbildunmarkszelle von „Riese
Zusammen
Dentinsche
verursachte
sich hierbe
handelt.
3.2.1 Wirku
3.2.1.1 TRA
In den mit
liche dosis
Größe der O
g 10: Osteoklastenkultur: Stimulierung von Knochen-n der Maus mit OPGL und M-CSF führte zur Bildungnosteoklasten“
genommen mit den Ergebnissen der Auswertung der
ibchen, auf denen sich deutlich der durch diese Zellen
Knochenabbau darstellen ließ, postulieren wir, dass es
i um Osteoklasten bzw. sog. „osteoclast-like cells“
ng von Zoledronat in vitro
P-Färbung
Zoledronat behandelten Zellkulturen ließ sich eine deut-
abhängige Reduktion sowohl der Anzahl als auch der
steoklasten beobachten (Abbildung 11).
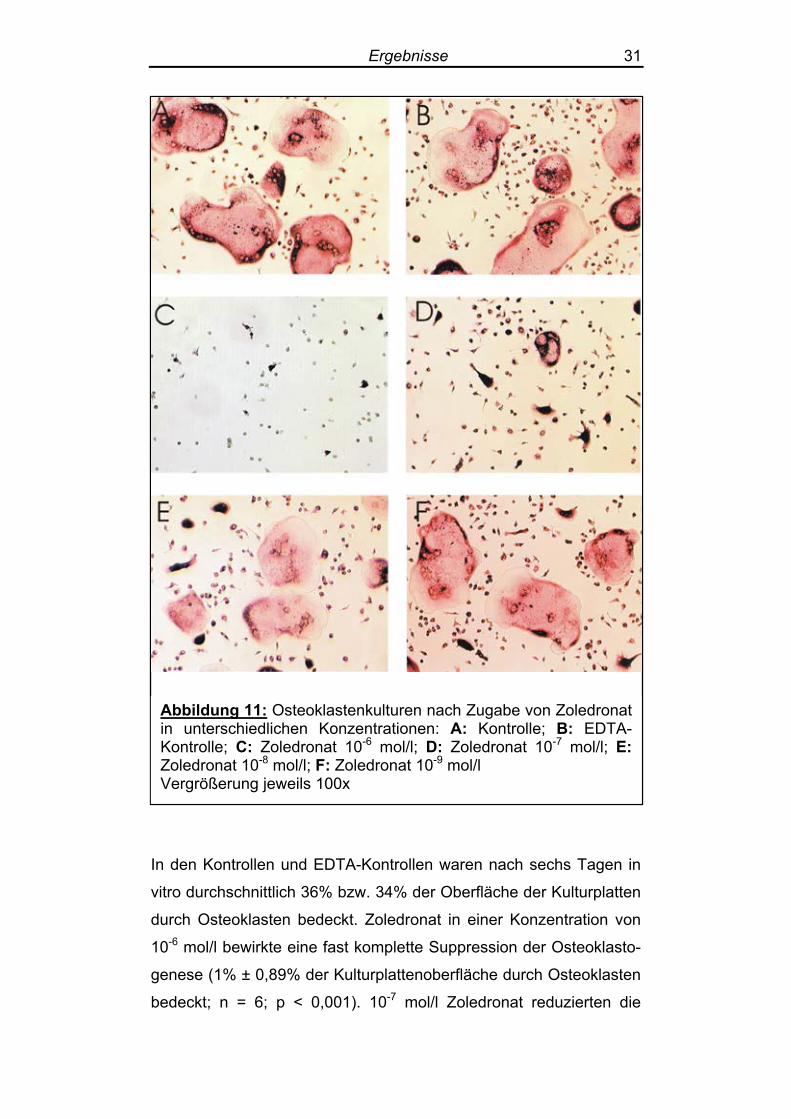
Ergebnisse 31
Abbildung 11: Osteoklastenkulturen nach Zugabe von Zoledronat in unterschiedlichen Konzentrationen: A: Kontrolle; B: EDTA-Kontrolle; C: Zoledronat 10-6 mol/l; D: Zoledronat 10-7 mol/l; E: Zoledronat 10-8 mol/l; F: Zoledronat 10-9 mol/l Vergrößerung jeweils 100x
In den Kontrollen und EDTA-Kontrollen waren nach sechs Tagen in
vitro durchschnittlich 36% bzw. 34% der Oberfläche der Kulturplatten
durch Osteoklasten bedeckt. Zoledronat in einer Konzentration von
10-6 mol/l bewirkte eine fast komplette Suppression der Osteoklasto-
genese (1% ± 0,89% der Kulturplattenoberfläche durch Osteoklasten
bedeckt; n = 6; p < 0,001). 10-7 mol/l Zoledronat reduzierten die
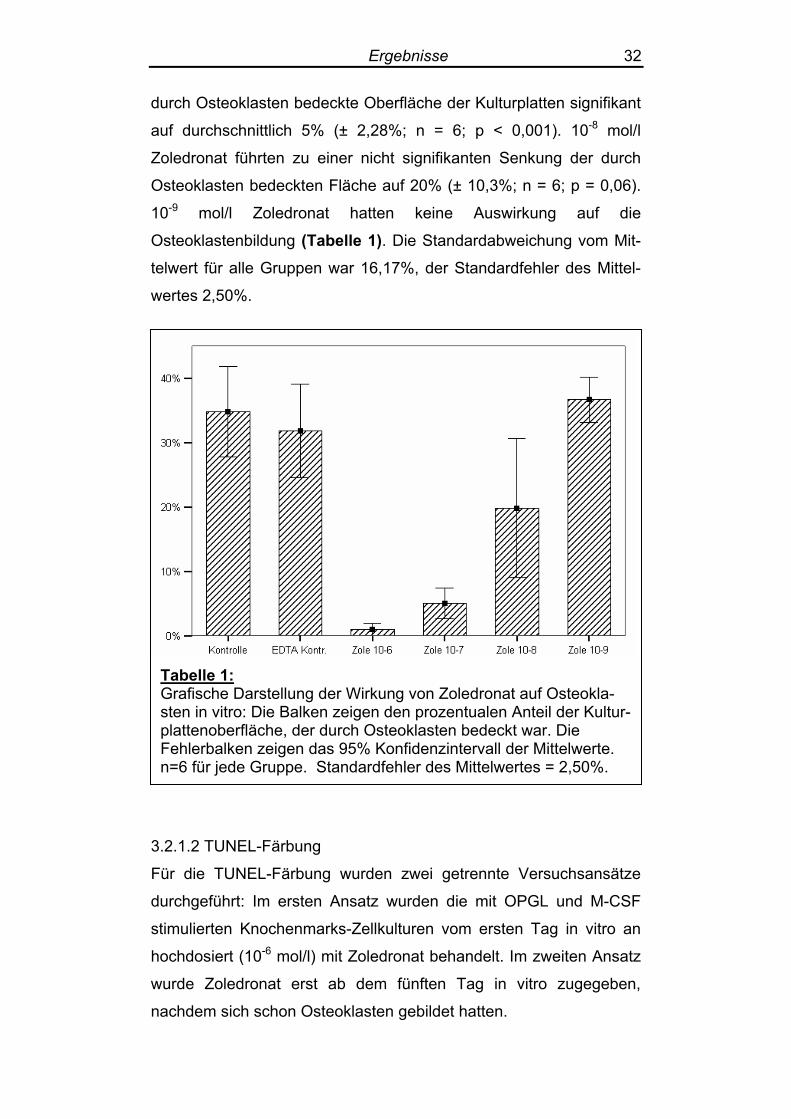
Ergebnisse 32
durch Osteoklasten bedeckte Oberfläche der Kulturplatten signifikant
auf durchschnittlich 5% (± 2,28%; n = 6; p < 0,001). 10-8 mol/l
Zoledronat führten zu einer nicht signifikanten Senkung der durch
Osteoklasten bedeckten Fläche auf 20% (± 10,3%; n = 6; p = 0,06).
10-9 mol/l Zoledronat hatten keine Auswirkung auf die
Osteoklastenbildung (Tabelle 1). Die Standardabweichung vom Mit-
telwert für alle Gruppen war 16,17%, der Standardfehler des Mittel-
wertes 2,50%.
Tabelle 1: Grafische Darstellung der Wirkung von Zoledronat auf Osteokla-sten in vitro: Die Balken zeigen den prozentualen Anteil der Kultur-plattenoberfläche, der durch Osteoklasten bedeckt war. Die Fehlerbalken zeigen das 95% Konfidenzintervall der Mittelwerte. n=6 für jede Gruppe. Standardfehler des Mittelwertes = 2,50%.
3.2.1.2 TUNEL-Färbung
Für die TUNEL-Färbung wurden zwei getrennte Versuchsansätze
durchgeführt: Im ersten Ansatz wurden die mit OPGL und M-CSF
stimulierten Knochenmarks-Zellkulturen vom ersten Tag in vitro an
hochdosiert (10-6 mol/l) mit Zoledronat behandelt. Im zweiten Ansatz
wurde Zoledronat erst ab dem fünften Tag in vitro zugegeben,
nachdem sich schon Osteoklasten gebildet hatten.

Ergebnisse 33
Es fanden sich TUNEL-positive Zellen in sämtlichen mit Zoledronat
behandelten Versuchsansätzen, sowohl in reifen Osteoklasten als
auch in Osteoklasten-Vorläuferzellen. (Abbildung 12). Daneben
fanden sich andere, vitale Knochenmarkszellen. In den Kontroll-
Kulturen zeigten sich keine oder nur vereinzelt TUNEL-positive
Zellen.
Abbildung 12: TUNEL-positive Zellen A: Osteoklasten-Vorläuferzellen B: reife Osteoklasten Vergrößerung jeweils 100X
3.2.2 Wirkung von Calzitonin in vitro
3.2.2.1 Zellkulturen
Die Osteoklastogenese wurde durch Zusatz von Calzitonin nicht
beeinflusst: Der durch Osteoklasten bedeckte Anteil der Kulturplat-
tenoberfläche änderte sich durch Zugabe von Calzitonin nicht
(Abbildung 13). Zwischen den Osteoklasten in den Kontrollen und den mit Calzitonin
behandelten Zellkulturen ließen sich weder licht- noch rasterelektro-
nenmikroskopisch morphologische Unterschiede feststellen (Abbil-dung 13 + 14).
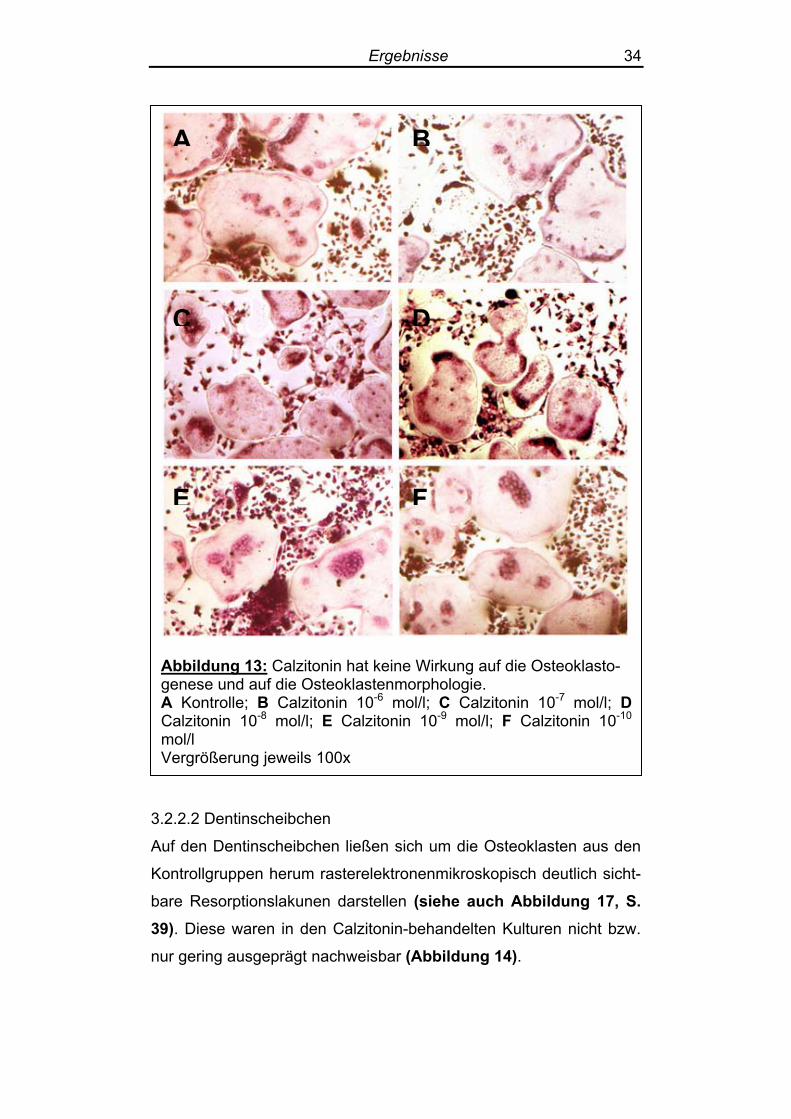
Ergebnisse 34
Abbildung 13: Calzitonin hat keine Wirkung auf die Osteoklasto-genese und auf die Osteoklastenmorphologie. A Kontrolle; B Calzitonin 10-6 mol/l; C Calzitonin 10-7 mol/l; D Calzitonin 10-8 mol/l; E Calzitonin 10-9 mol/l; F Calzitonin 10-10 mol/l Vergrößerung jeweils 100x
FE
DC
BA
3.2.2.2 Dentinscheibchen
Auf den Dentinscheibchen ließen sich um die Osteoklasten aus den
Kontrollgruppen herum rasterelektronenmikroskopisch deutlich sicht-
bare Resorptionslakunen darstellen (siehe auch Abbildung 17, S. 39). Diese waren in den Calzitonin-behandelten Kulturen nicht bzw.
nur gering ausgeprägt nachweisbar (Abbildung 14).
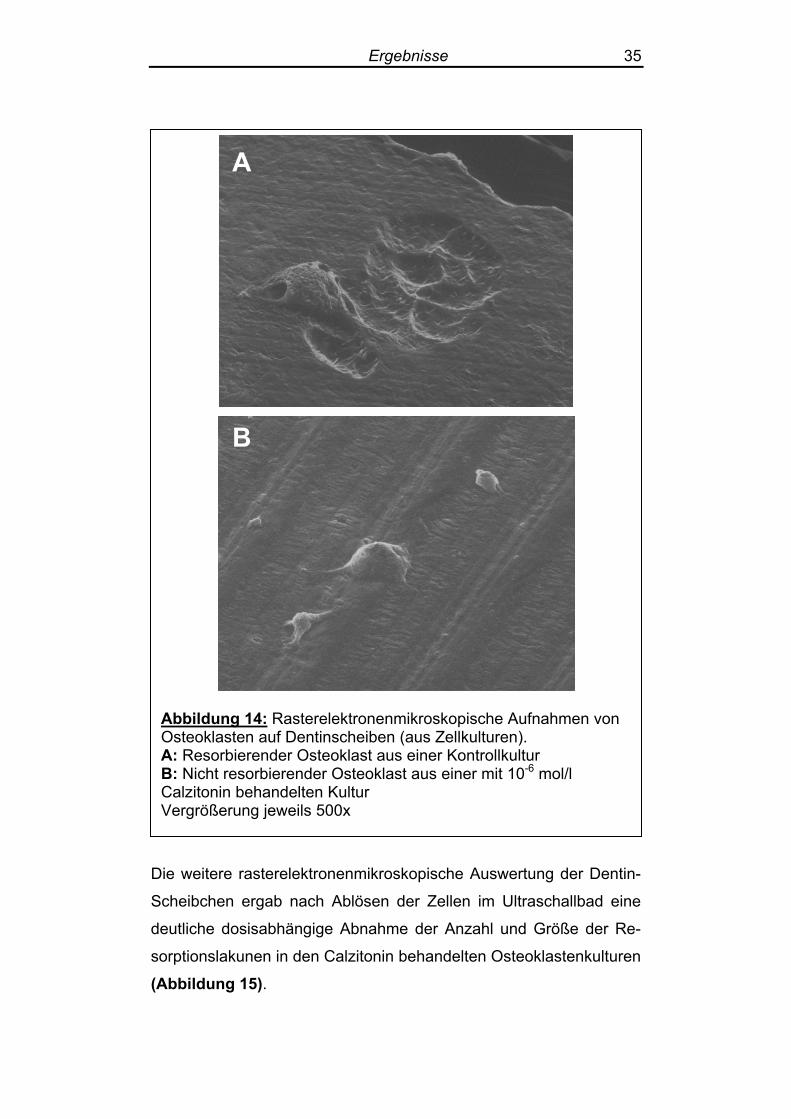
Ergebnisse 35
Abbildung 14: Rasterelektronenmikroskopische Aufnahmen von Osteoklasten auf Dentinscheiben (aus Zellkulturen). A: Resorbierender Osteoklast aus einer Kontrollkultur B: Nicht resorbierender Osteoklast aus einer mit 10-6 mol/l Calzitonin behandelten Kultur Vergrößerung jeweils 500x
B
A
Die weitere rasterelektronenmikroskopische Auswertung der Dentin-
Scheibchen ergab nach Ablösen der Zellen im Ultraschallbad eine
deutliche dosisabhängige Abnahme der Anzahl und Größe der Re-
sorptionslakunen in den Calzitonin behandelten Osteoklastenkulturen
(Abbildung 15).
Ergebnisse 36
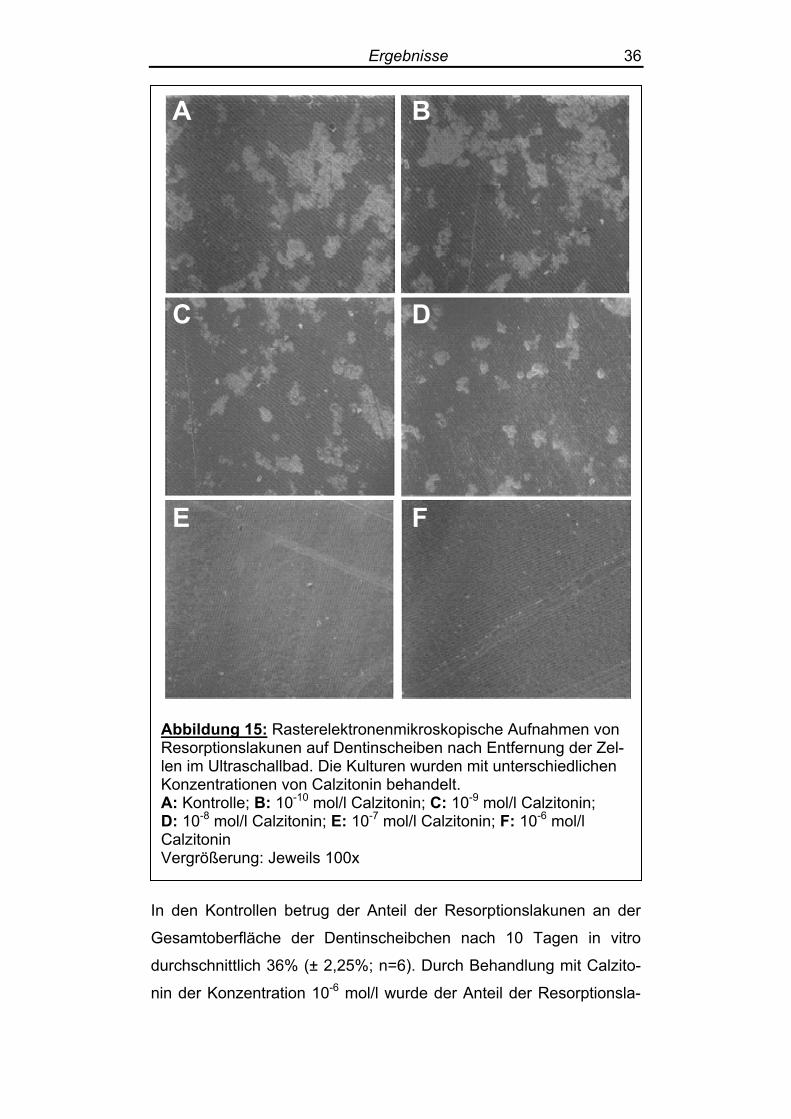
Abbildung 15: Rasterelektronenmikroskopische Aufnahmen von Resorptionslakunen auf Dentinscheiben nach Entfernung der Zel-len im Ultraschallbad. Die Kulturen wurden mit unterschiedlichen Konzentrationen von Calzitonin behandelt. A: Kontrolle; B: 10-10 mol/l Calzitonin; C: 10-9 mol/l Calzitonin; D: 10-8 mol/l Calzitonin; E: 10-7 mol/l Calzitonin; F: 10-6 mol/l Calzitonin Vergrößerung: Jeweils 100x
B
C D
E F
A
In den Kontrollen betrug der Anteil der Resorptionslakunen an der
Gesamtoberfläche der Dentinscheibchen nach 10 Tagen in vitro
durchschnittlich 36% (± 2,25%; n=6). Durch Behandlung mit Calzito-
nin der Konzentration 10-6 mol/l wurde der Anteil der Resorptionsla-
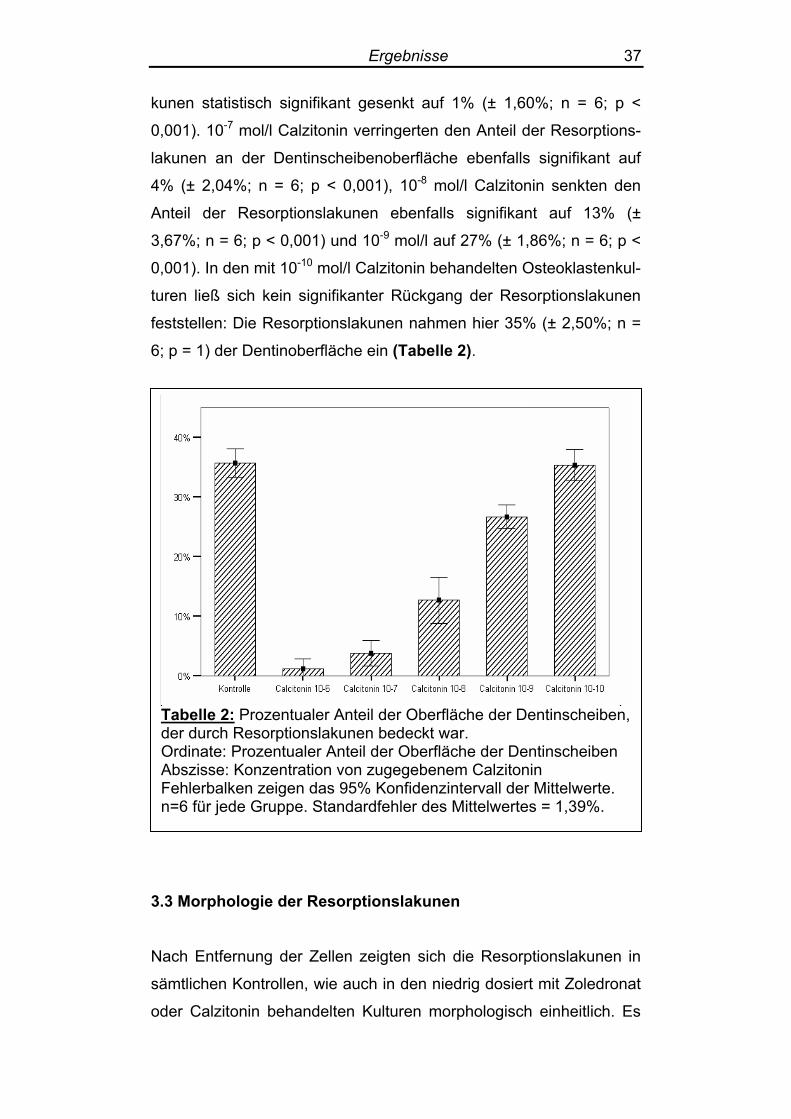
Ergebnisse 37
kunen statistisch signifikant gesenkt auf 1% (± 1,60%; n = 6; p <
0,001). 10-7 mol/l Calzitonin verringerten den Anteil der Resorptions-
lakunen an der Dentinscheibenoberfläche ebenfalls signifikant auf
4% (± 2,04%; n = 6; p < 0,001), 10-8 mol/l Calzitonin senkten den
Anteil der Resorptionslakunen ebenfalls signifikant auf 13% (±
3,67%; n = 6; p < 0,001) und 10-9 mol/l auf 27% (± 1,86%; n = 6; p <
0,001). In den mit 10-10 mol/l Calzitonin behandelten Osteoklastenkul-
turen ließ sich kein signifikanter Rückgang der Resorptionslakunen
feststellen: Die Resorptionslakunen nahmen hier 35% (± 2,50%; n =
6; p = 1) der Dentinoberfläche ein (Tabelle 2).
3 N
s
o
Tabelle 2: Prozentualer Anteil der Oberfläche der Dentinscheiben, der durch Resorptionslakunen bedeckt war. Ordinate: Prozentualer Anteil der Oberfläche der Dentinscheiben Abszisse: Konzentration von zugegebenem Calzitonin Fehlerbalken zeigen das 95% Konfidenzintervall der Mittelwerte. n=6 für jede Gruppe. Standardfehler des Mittelwertes = 1,39%.
.3 Morphologie der Resorptionslakunen
ach Entfernung der Zellen zeigten sich die Resorptionslakunen in
ämtlichen Kontrollen, wie auch in den niedrig dosiert mit Zoledronat
der Calzitonin behandelten Kulturen morphologisch einheitlich. Es

Ergebnisse 38
handelte sich um grüppchenweise dicht nebeneinander angeordnete
rundliche Kerben mit einem Durchmesser von 10-40 µm (Abbildung 16).
Abbildung 16: Resorptionslakunen auf Dentinscheibchen nach Entfernung der Zellen. Entstanden in Osteoklastenkultur. Vergrößerung: 100x, Inlay: 500x
Auf den mitsamt Zellen untersuchten Dentinscheibchen fanden sich
immer ein oder mehrere große Osteoklasten (bzw. osteoclast-like
cells) in den Resorptionslakunen (Abbildung 17).

Ergebnisse 39
Abbildung 17: Resorbierende Osteoklasten auf Dentinscheiben. Die Präparate stammen aus Osteoklastenkulturen. Vergrößerung: A, B: 650x; C: 750x; D: 300x
DC
BA
Die Resorptionszonen erschienen nie in die Tiefe ausgebreitet. Auch
in den Positivkontrollen zeigten sich die Resorptionslakunen flächig
verteilt.
Morphologisch wiesen die in der Osteoklastenkultur entstandenen
Resorptionslakunen eine große Ähnlichkeit zu den Resorptionslaku-
nen auf, die bereits auf den im Rahmen der Cholesteatomchirurgie
entnommenen menschlichen Gehörknöchelchen beschrieben wur-
den. Die in vivo entstandenen Resorptionslakunen erschienen jedoch
weniger oberflächlich ausgebreitet (Abbildung 18).
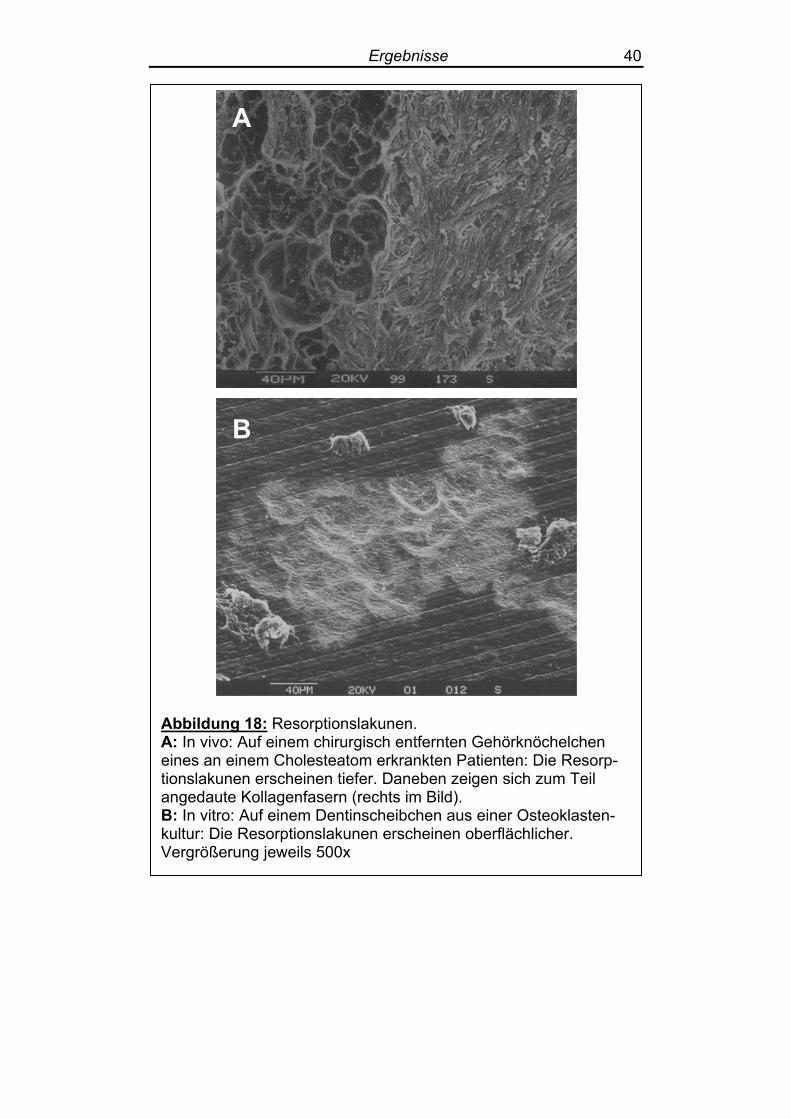
Ergebnisse 40
A
Abbildung 18: Resorptionslakunen. A: In vivo: Auf einem chirurgisch entfernten Gehörknöchelchen eines an einem Cholesteatom erkrankten Patienten: Die Resorp-tionslakunen erscheinen tiefer. Daneben zeigen sich zum Teil angedaute Kollagenfasern (rechts im Bild). B: In vitro: Auf einem Dentinscheibchen aus einer Osteoklasten-kultur: Die Resorptionslakunen erscheinen oberflächlicher. Vergrößerung jeweils 500x
B
Ergebnisse 41
3.4 Tierexperimente
3.4.1 Keratinpartikel-induzierte Osteolyse in vivo
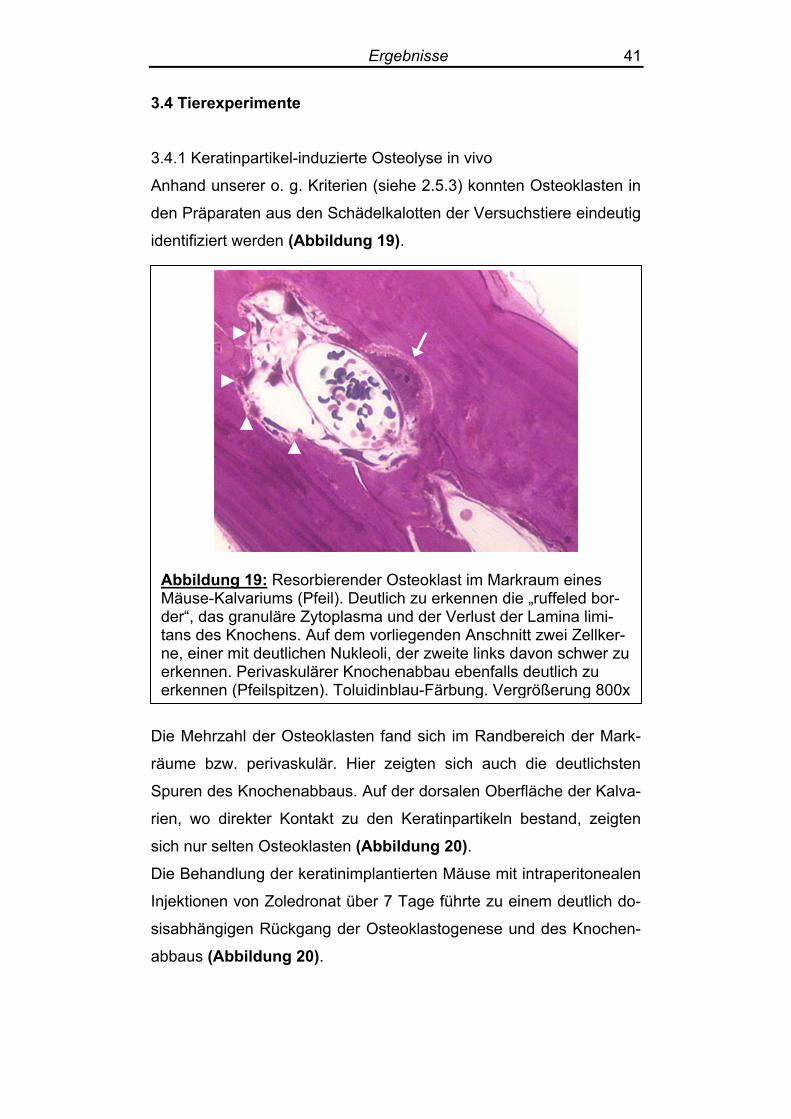
Anhand unserer o. g. Kriterien (siehe 2.5.3) konnten Osteoklasten in
den Präparaten aus den Schädelkalotten der Versuchstiere eindeutig
identifiziert werden (Abbildung 19).
Abbildung 19: Resorbierender Osteoklast im Markraum eines Mäuse-Kalvariums (Pfeil). Deutlich zu erkennen die „ruffeled bor-der“, das granuläre Zytoplasma und der Verlust der Lamina limi-tans des Knochens. Auf dem vorliegenden Anschnitt zwei Zellker-ne, einer mit deutlichen Nukleoli, der zweite links davon schwer zu erkennen. Perivaskulärer Knochenabbau ebenfalls deutlich zu erkennen (Pfeilspitzen). Toluidinblau-Färbung. Vergrößerung 800x
Die Mehrzahl der Osteoklasten fand sich im Randbereich der Mark-
räume bzw. perivaskulär. Hier zeigten sich auch die deutlichsten
Spuren des Knochenabbaus. Auf der dorsalen Oberfläche der Kalva-
rien, wo direkter Kontakt zu den Keratinpartikeln bestand, zeigten
sich nur selten Osteoklasten (Abbildung 20). Die Behandlung der keratinimplantierten Mäuse mit intraperitonealen
Injektionen von Zoledronat über 7 Tage führte zu einem deutlich do-
sisabhängigen Rückgang der Osteoklastogenese und des Knochen-
abbaus (Abbildung 20).
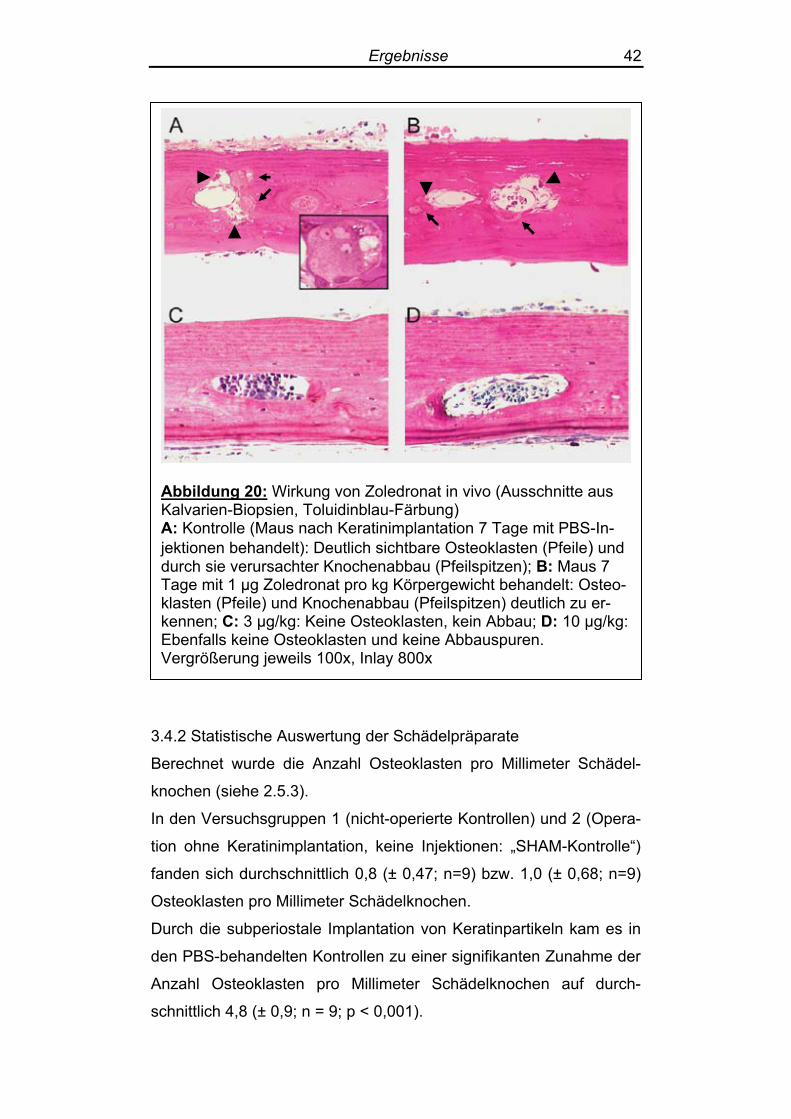
Ergebnisse 42
Abbildung 20: Wirkung von Zoledronat in vivo (Ausschnitte aus Kalvarien-Biopsien, Toluidinblau-Färbung) A: Kontrolle (Maus nach Keratinimplantation 7 Tage mit PBS-In-jektionen behandelt): Deutlich sichtbare Osteoklasten (Pfeile) und durch sie verursachter Knochenabbau (Pfeilspitzen); B: Maus 7 Tage mit 1 µg Zoledronat pro kg Körpergewicht behandelt: Osteo-klasten (Pfeile) und Knochenabbau (Pfeilspitzen) deutlich zu er-kennen; C: 3 µg/kg: Keine Osteoklasten, kein Abbau; D: 10 µg/kg: Ebenfalls keine Osteoklasten und keine Abbauspuren. Vergrößerung jeweils 100x, Inlay 800x
3.4.2 Statistische Auswertung der Schädelpräparate
Berechnet wurde die Anzahl Osteoklasten pro Millimeter Schädel-
knochen (siehe 2.5.3).
In den Versuchsgruppen 1 (nicht-operierte Kontrollen) und 2 (Opera-
tion ohne Keratinimplantation, keine Injektionen: „SHAM-Kontrolle“)
fanden sich durchschnittlich 0,8 (± 0,47; n=9) bzw. 1,0 (± 0,68; n=9)
Osteoklasten pro Millimeter Schädelknochen.
Durch die subperiostale Implantation von Keratinpartikeln kam es in
den PBS-behandelten Kontrollen zu einer signifikanten Zunahme der
Anzahl Osteoklasten pro Millimeter Schädelknochen auf durch-
schnittlich 4,8 (± 0,9; n = 9; p < 0,001).
Ergebnisse 43
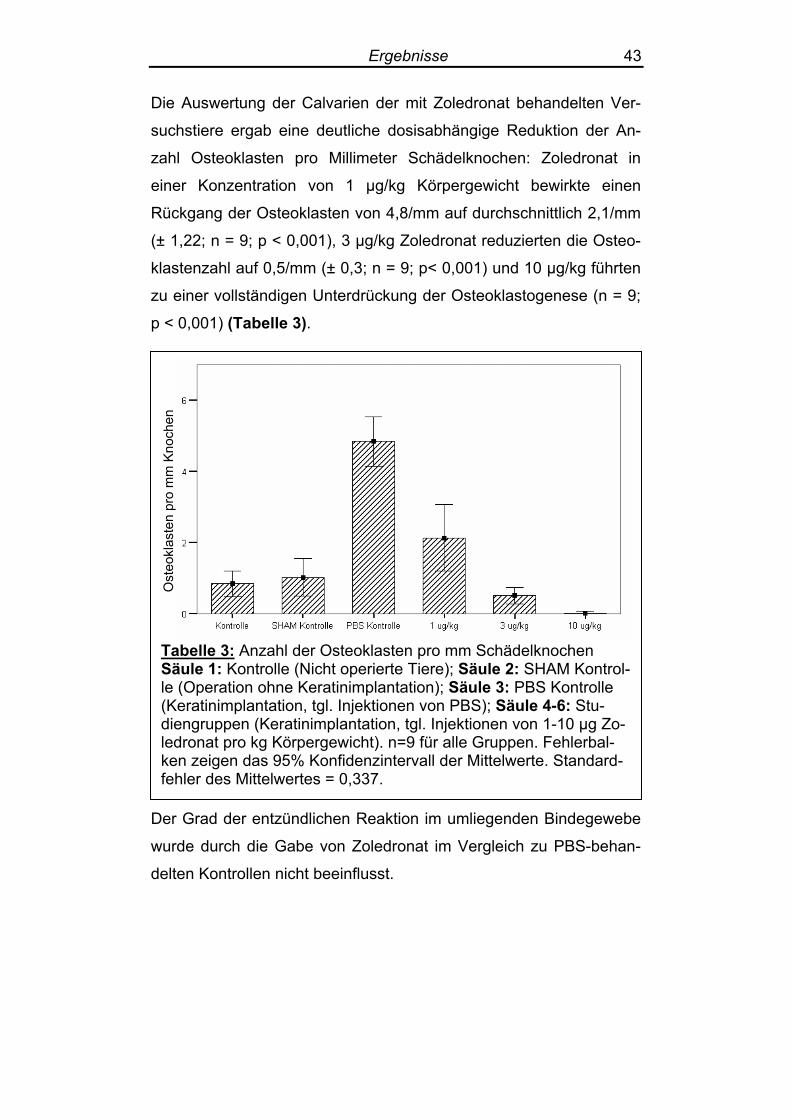
Die Auswertung der Calvarien der mit Zoledronat behandelten Ver-
suchstiere ergab eine deutliche dosisabhängige Reduktion der An-
zahl Osteoklasten pro Millimeter Schädelknochen: Zoledronat in
einer Konzentration von 1 µg/kg Körpergewicht bewirkte einen
Rückgang der Osteoklasten von 4,8/mm auf durchschnittlich 2,1/mm
(± 1,22; n = 9; p < 0,001), 3 µg/kg Zoledronat reduzierten die Osteo-
klastenzahl auf 0,5/mm (± 0,3; n = 9; p< 0,001) und 10 µg/kg führten
zu einer vollständigen Unterdrückung der Osteoklastogenese (n = 9;
p < 0,001) (Tabelle 3).
Tabelle 3: Anzahl der Osteoklasten pro mm Schädelknochen Säule 1: Kontrolle (Nicht operierte Tiere); Säule 2: SHAM Kontrol-le (Operation ohne Keratinimplantation); Säule 3: PBS Kontrolle (Keratinimplantation, tgl. Injektionen von PBS); Säule 4-6: Stu-diengruppen (Keratinimplantation, tgl. Injektionen von 1-10 µg Zo-ledronat pro kg Körpergewicht). n=9 für alle Gruppen. Fehlerbal-ken zeigen das 95% Konfidenzintervall der Mittelwerte. Standard-fehler des Mittelwertes = 0,337.
Ost
eokl
aste
n pr
o m
m K
noch
en
Der Grad der entzündlichen Reaktion im umliegenden Bindegewebe
wurde durch die Gabe von Zoledronat im Vergleich zu PBS-behan-
delten Kontrollen nicht beeinflusst.

Diskussion 44
4. Diskussion
4.1 Morphologie der Ossikel im Cholesteatom
Das Cholesteatom bewirkt einen Abbau von Knochen. Dieser kann
auf intraoperativ gewonnenen Gehörknöchelchen sehr deutlich dar-
gestellt werden. Solche Ossikel zeigen sowohl äußerlich als auch
innerlich Spuren des Knochenabbaus: Es finden sich Resorptionsla-
kunen verschiedener Ausprägung auf ihrer Oberfläche bis hin zum
vollständigen Abbau ganzer Teile des Gehörknöchelchens, in den
meisten Fällen des langen Ambossfortsatzes. Außerdem kommt es
im Cholesteatom zu einer Erweiterung der Haversschen Kanäle und
somit zu einer „Aushöhlung“ der Gehörknöchelchen. Diese Beobach-
tung deutet darauf hin, dass der Knochenabbau nicht nur in direkter
Nachbarschaft des Cholesteatoms an der Knochenoberfläche statt-
findet, sondern auch perivaskulär in den Markräumen der befallenen
Ossikel.
4.1.1 Oberflächliche Resorption
Die Resorption an der Knochenoberfläche läuft in zwei verschiede-
nen Stadien ab: Bevor es zur Ausbildung von Resorptionslakunen
kommt, findet eine oberflächliche Andauung der Kollagenfasern des
Knochens statt. Hierbei wird die reguläre Kollagenstruktur aufgelöst,
jedoch noch nicht resorbiert. Dies ist Anzeichen dafür, dass zunächst
die anorganische (mineralische) Knochenstruktur abgebaut wird.
Schließlich kommt es zur Resorption der organischen Knochensub-
stanz (Kollagen) und somit Bildung von Resorptionslakunen, an
deren Boden man noch angedaute Kollagenfasern erkennen kann.
Diese Beobachtungen stimmen mit den Ergebnissen von Uno et al.
(1995), sowie Wong et al. (1991) überein.
Weiterhin könnte auch die mit der Knochenresorption konkurrierende
Knochenneubildung eine Rolle bei der Entstehung der Morphologie
des Knochenabbaus spielen. So stellen Wong et al. (1991) die These

Diskussion 45
auf, dass es erst dann zur Ausbildung von Resorptionslakunen
kommt, wenn der Knochenabbau die konkurrierende Knochenneubil-
dung verdrängt.
4.1.2 Perivaskuläre Knochenresorption
Die Beobachtung des „perivaskulären Knochenabbaus“ von den
Markräumen her konnte sowohl an operativ entfernten Ossikeln als
auch im Tierexperiment gemacht werden. In unserem Maus-Modell
zum osteoklastären Knochenabbau in vivo zeigten sich die deutlich-
sten Spuren des Knochenabbaus und auch die meisten Osteoklasten
in den Markräumen der Kalvarien der Versuchstiere bzw. perivasku-
lär. Auf der dorsalen Fläche des Knochens, im direkten Kontakt zu
den implantierten Keratinpartikeln, fanden sich deutlich weniger
Osteoklasten und deutlich geringere Spuren des Knochenabbaus.
Diese Beobachtung unterstützt die These, dass der Knochenabbau
im Cholesteatom durch aus dem Blut stammende Zellen und Entzün-
dungsmediatoren bewirkt wird (Schilling et al. 1992, Ahn et al. 1990,
Cheshire et al. 1991, Sadé et al 1981) und spricht gegen die Theorie
einer Druckatrophie des Knochens (Erdheim 1905, Grünwald 1910,
Rüedi 1957, Tumarkin 1958). Die durch Grippaudo (1958), Pollock
(1959) und von Schulthess (1961) beobachtete chronische Osteo-
myelitis von Gehörknöchelchen im Cholesteatom passt ebenfalls zu
diesen Beobachtungen.
4.2 Vergleich der Knochenresorption in vivo und in vitro
Der Vergleich der Morphologie des Knochenabbaus im Cholesteat-
om und in der Osteoklasten-Zellkultur unterstützt ebenfalls die These
eines osteoklastären Knochenabbaus, da die Morphologie der in vivo
und in vitro entstandnen Resorptionslakunen nahezu identisch ist.
Auf der Oberfläche von im Cholesteatom arrodierten Ossikeln finden
sich regelmäßig Resorptionslakunen. Diese sind rund, grüppchen-
weise angeordnet und haben einen Durchmesser von 10-40 µm. Die

Diskussion 46
in der Osteoklastenkultur entstandenen Resorptionslakunen haben
eine fast identische Morphologie. Einziges Unterscheidungsmerkmal
ist, dass die in der Zellkultur entstandenen Resorptionslakunen
oberflächlicher erscheinen. Für diesen Unterschied bestehen mehre-
re Erklärungsmöglichkeiten. Zum einen haben wir in den Zellkulturen
ein anderes Substrat für die Knochenresorption verwendet. Hier wäre
es denkbar, dass das Wal-Dentin andere Resorptionseigenschaften
besitzt als Knochen. Zum anderen wäre es möglich, dass sich die
amöboide Beweglichkeit der Osteoklasten in der Zellkultur stärker
auswirkt, da natürliche Barrieren wie z.B. Schleimhaut hier fehlen.
4.3 Inhibition der Knochenresorption in vitro
Neuere Untersuchungen (Lacey et al. 1998, Yasuda et al. 1998)
haben gezeigt, daß Knochenumbau und Knochenresorption durch
ein Zusammenspiel zwischen dem Rezeptor-Aktivator von NF-κB
(=RANK) und seinem Ligand RANKL (auch OPGL, TRANCE oder
ODF) kontrolliert werden. Der OPGL-Rezeptor (RANK) wurde auf
dendritischen Zellen, Chondrozyten, Osteoklasten-Vorläuferzellen
und reifen Osteoklasten identifiziert (Suda et al. 1999).
Durch Stimulation mit OPGL und M-CSF (Macrophage Colony
Stimulating Factor) konnten aus Knochenmarks-Zellkulturen Osteo-
klastenkulturen etabliert werden. Beide Faktoren sind wirksame
Stimulatoren der Osteoklastogenese und Osteoklastendifferenzie-
rung. Sie führen zuverlässig zur Bildung von resorbierenden Osteo-
klasten aus Knochenmarks-Vorläuferzellen.
4.3.1 Zoledronat in vitro
Zoledronat ist ein Bisphosphonat der dritten Generation und nach-
gewiesenermaßen deutlich stärker wirksam als ältere Bisphospho-
nate (Cheer et al. 2001).
In den Zellkultur-Experimenten konnte gezeigt werden, dass Zoledro-
nat in vitro in Konzentrationen zwischen 10-9 und 10-6 mol/l eine

Diskussion 47
dosisabhängige Inhibition der Osteoklastogenese und der osteokla-
stären Knochenresorption verursacht. Weiterhin war zu beobachten,
dass in den mit Zoledronat behandelten Zellkulturen viele TUNEL-
positive Zellen nachzuweisen waren. Diese traten sowohl in Zellkul-
turen auf, in denen die Osteoklasten-Vorläuferzellen vor der Behand-
lung mit Zoledronat ausgereift waren, als auch in Kulturen, denen
Zoledronat gleich zu Beginn zugegeben wurde. Hier kam es gar nicht
zur Ausbildung reifer Osteoklasten, die Osteoklasten-Vorläuferzellen
zeigten sich TUNEL-positiv.
Diese Beobachtungen deuten darauf hin, daß die inhibierende
Wirkung von Zoledronat auf die Knochenresorption durch eine
Induktion von Apoptose in Osteoklasten und ihren Vorläuferzellen
zurückzuführen sein könnte. Ähnliche Beobachtungen machten auch
Jagdev et al. (2001) und Fromigue et al. (2000) an Brustkrebs-Zellen.
Untersuchungen von Coxon et al. (2000) zum molekularen Me-
chanismus der Wirkung von Bisphosphonaten deuten darauf hin,
dass diese Eigenschaft auf eine Hemmung der Protein-Geranylgera-
nylation zurückzuführen sein könnte.
Ergebnisse von Reinholz et al. (2000) zeigen, dass der inhibitorische
Effekt von Bisphosphonaten auf die Knochenresorption teilweise
auch auf eine Aktivierung von Osteoblasten zurückzuführen ist.
4.3.2 Calzitonin in vitro
In unseren Untersuchungen konnten wir nachweisen, dass humanes
Calzitonin in vitro in Konzentrationen von 10-10 bis 10-6 mol/l
dosisabhängig die OPGL / M-CSF-induzierte osteoklastäre Knochen-
resorption inhibiert. Diese Ergebnisse passen zu den Beobachtungen
von Galvin et al (1998). Auswirkungen auf die Osteoklastogenese
oder Osteoklastenmorphologie konnten in vitro weder licht- noch
raster-elektronenmikroskopisch nachgewiesen werden. Die Wirkung
von Calzitonin scheint am ehesten durch eine direkte Hemmung der
Resorptionstätigkeit der Osteoklasten über den Calzitonin-Rezeptor
bedingt zu sein (Galvin et al. 1998). Eine bereits praktizierte
Anwendung von Calzitonin besteht in der Therapie der Osteoporose.

Diskussion 48
Es existieren daher bereits ausführliche Daten zur systemischen
Anwendung dieses Polypeptidhormons (Weber et al. 1999). Unsere
Ergebnisse deuten darauf hin, dass eine lokale oder auch systemi-
sche Behandlung mit Calzitonin durchaus einen Stellenwert in der
adjuvanten Therapie des Cholesteatoms zusätzlich zur Chirurgie
haben könnte.
Weiterführende Studien sollten die bereits beschriebene Abnahme
der Wirkung bei langfristiger Calzitonin-Behandlung durch Abnahme
der Rezeptor-Dichte auf den Zielzellen (Samura et al. 2000, Galvin et
al. 1998) untersuchen. Außerdem sollte die Wirkung von Calzitonin
auf den cholesteatombedingten Knochenabbau in vivo weiter unter-
sucht werden. Hier sollte auch die Möglichkeit einer Lokaltherapie mit
Calzitonin in Betracht gezogen werden.
4.4 Inhibition der Knochenresorption in vivo
4.4.1 Zoledronat in vivo
Zur Untersuchung der Wirkung von Zoledronat in vivo zogen wir ein
kürzlich von Chole et al. (2001) beschriebenes Tiermodell zum Kno-
chenumbau im Cholesteatom heran: Sterilisierte Keratinpartikel
wurden subperiostal auf die Oberfläche des Os parietale von Mäusen
implantiert, was zu einer lokalen osteoklastären Knochenresorption
führte.
Dieses Modell konzentriert sich auf die osteoklastäre Knochenre-
sorption und bietet die Möglichkeit, die Funktion von Osteoklasten
und Möglichkeiten ihrer Inhibition in vivo zu untersuchen. Ein Chole-
steatom im eigentlichen Sinne wird nicht erzeugt. Die implantierten
Keratinpartikel sollen jedoch ein Cholesteatom simulieren. Sicherlich
spielt bei der in diesem Modell auftretenden Knochenresorption die
Entzündungsreaktion auf die implantierten Fremdkörper eine ent-
scheidende Rolle.
Der Nachteil eines Maus-Modells liegt also darin, dass die Maus
nicht spontan oder induziert „echte“ Cholesteatome entwickelt. Der

Diskussion 49
entscheidende Vorteil eines Maus-Modells liegt jedoch darin, dass
viele der Gene und Genprodukte, die entzündliche Prozesse steuern
in der Maus sehr gut identifiziert und charakterisiert sind. Außerdem
existieren Knockout-Mäuse oder transgene Mäuse für viele dieser
Gene, so dass gute Ansätze für weiterführende Studien gegeben
sind.
Es wurden bereits diverse Tiermodelle zur Untersuchung des Chole-
steatoms, seiner Entwicklung und Pathophysiologie entwickelt. Die
einzigen Säuger außer dem Menschen, von denen man weiß, daß
sie spontan Cholesteatome entwickeln, sind die mongolische Renn-
maus (Meriones unguiculatus) (Chole et al. 1981) und die Sandrenn-
maus (Psammomys obesus) (Feinmesser et al. 1988). Beide Modelle
bieten große Ähnlichkeiten zur Cholesteatomentstehung beim Men-
schen, haben aber den Nachteil, dass nur sehr wenig genetische
Informationen über die verwendeten Spezies zur Verfügung stehen.
4.5 Vergleich der Wirkungen von Zoledronat und Calzitonin
Die Wirkung dieser beiden unterschiedlichen Substanzen ist ähnlich;
beide reduzieren die osteoklastäre Knochenresorption signifikant.
Die Mechanismen, die dieser Wirkung zugrunde liegen, sind jedoch
sehr unterschiedlich. Zoledronat bewirkt durch eine Induktion von
Apoptose eine Hemmung der Osteoklastogenese, Calzitonin bewirkt
über den Calzitonin-Rezeptor eine Hemmung der Osteoklastenfunk-
tion. Die Wirkung von Calzitonin lässt aufgrund der vorbeschriebenen
Abnahme der Rezeptordichte bei längerer Behandlungsdauer nach
(Samura et al. 2000). Zur Langzeitbehandlung mit Zoledronat liegen
noch keine verwertbaren Ergebnisse vor, aufgrund des Wirkmecha-
nismus wäre ein ähnliches Phänomen jedoch unwahrscheinlich. Eine
systemische Anwendung von Zoledronat erscheint allerdings nicht
unproblematisch, da es wahrscheinlich nachhaltige Auswirkungen
auf den gesamten Knochenstoffwechsel und andere unerwünschte
Wirkungen hätte (Cheer et al. 2001).

Diskussion 50
Aufgrund der unterschiedlichen Wirkungsweisen der beiden Medika-
mente erscheint eine Kombinationstherapie zum Beispiel auch in
Form einer Lokaltherapie als adjuvante Behandlung des Cholesteat-
oms sinnvoll. Weiterführende Studien sollten diese Möglichkeit oder
auch die Kombination einer systemischen Anwendung von Calzitonin
und einer Lokaltherapie mit Zoledronat untersuchen.
4.6 Klinische Relevanz Die Ergebnisse deuten darauf hin, dass die Gabe von Zoledronat
und Calzitonin lokal oder systemisch, einzeln oder aber auch in
Kombination eine sinnvolle Ergänzung der chirurgischen Therapie
des Cholesteatoms sein könnte. Weiterführende Studien sollten sich
mit dieser Möglichkeit auseinandersetzen. Die zu erwartenden Ne-
benwirkungen wären jedoch abzuwägen.
Die chirurgische Therapie des Cholesteatoms bleibt jedoch nach wie
vor in jedem Falle indiziert.

Zusammenfassung 51
5. Zusammenfassung
Ziele der vorliegenden Arbeit sind die Darstellung des Knochenab-
baus im Cholesteatom, der Rückschluss von der Morphologie auf die
Pathogenese des Knochenabbaus und die Untersuchung von Mög-
lichkeiten seiner medikamentösen Inhibition.
Die Arbeit gliedert sich in einen morphologisch-deskriptiven und
einen experimentellen Teil. Für den morphologischen Teil wurden im
Rahmen cholesteatomsanierender Mittelohreingriffe entnommene
Gehörknöchelchen rasterelektronenmikroskopisch untersucht und
die Spuren der Knochenresorption dargestellt. Hierbei zeigte sich,
dass der Knochenabbau im Cholesteatom zum einen in verschie-
denen Stadien auf der Oberfläche der Ossikel abläuft. Dies führt über
die Ausbildung von Resorptionslakunen bis hin zur vollständigen
Destruktion von Teilen der Ossikel. Zum anderen tritt auch eine deut-
liche Erweiterung der Haversschen Kanäle auf, was auf eine Kno-
chenresorption in den Markräumen ausgehend von Blutgefäßen
hindeutet.
Im experimentellen Teil wurden Zellkulturen resorbierender Osteo-
klasten etabliert. Der Vergleich der auf diese Weise in vitro entstan-
denen Resorptionslakunen mit den durch das Cholesteatom in vivo
entstandenen Resorptionsspuren zeigte große morphologische Ähn-
lichkeiten. Dies deutet auf eine osteoklastäre Genese der Knochen-
destruktion im Cholesteatom hin.
Im nächsten Schritt sollten Möglichkeiten der medikamentösen
Inhibition der Knochenresorption untersucht werden. Hierzu wurde in
der Osteoklasten-Zellkultur die Wirkung des Polypeptidhormons
Calzitonin einerseits und des Bisphosphonats Zoledronat anderer-
seits untersucht. Es konnte gezeigt werden, dass sowohl Calzitonin
als auch Zoledronat in vitro wirksame Inhibitoren der Knochenresorp-
tion sind. Sie haben unterschiedliche Wirkungsmechanismen und
wären somit auch in Kombination sinnvoll einsetzbar.

Zusammenfassung 52
Im letzten Schritt sollte die Wirkung von Zoledronat in vivo untersucht
werden. Hierzu wurde ein Maus-Modell zum osteoklastären Kno-
chenabbau verwendet. Es zeigte sich, dass Zoledronat bei systemi-
scher Gabe in vivo eine deutliche dosisabhängige Reduktion des
osteoklastären Knochenabbaus bewirkt.
Die Ergebnisse deuten darauf hin, dass die Gabe von Zoledronat
und Calzitonin lokal oder systemisch eine sinnvolle Ergänzung der
chirurgischen Therapie des Cholesteatoms darstellen könnte.

Literatur 53
6. Literaturverzeichnis
1. Abramson, M, Moriyama H, Huang CC. Pathogenic factors in bone resorption in cholesteatoma. Acta Otolaryngol 1984, 97, 437-42.
2. Ahn, J, Huang C, Abramson M. Interleukin-1 causing bone
destruction in middle ear cholesteatoma. Otolaryngol, Head Neck Surg 1990, 103, 527-536.
3. Amling, M, Delling G. Zellbiologie des Osteoklasten und
molekulare Mechanisman der Knochenresorption. Pathologe 1996, 17, 358-367.
4. Arnold, W. [The reactions of human middle ear mucous
membrane (author's transl)]. Arch Otorhinolaryngol 1977, 216, 369-473.
5. Basle, MF, Chappard D, Grizon F, Filmon R, Delecrin J,
Daculsi G, Rebel A. Osteoclastic resorption of Ca-P biomaterials implanted in rabbit bone. Calcif Tissue Int 1993, 53, 348-56.
6. Bezold, F. Cholesteatom, Perforation der Membrana flaccida
und Tubenverschluß. Z Hals Nasen Ohrenheilk 1890, 20, 5-29.
7. Body, JJ. Clinical research update: zoledronate. Cancer
1997, 80, 1699-701. 8. Breuil, V, Cosman F, Stein L, Horbert W, Nieves J, Shen V,
Lindsay R, Dempster DW. Human osteoclast formation and activity in vitro: effects of alendronate. J Bone Miner Res 1998, 13, 1721-9.
9. Bucher, O, Wartenberg H. Cytologie, Histologie und
mikroskopische Anatomie des Menschen. Göttingen, H. Huber, 1997.
10. Bujía, J, Kim C, Ostos P, Sudhoff H, Kastenbauer E, Hültner
I. Interleukin-1 (IL-1) and IL-1-R antagonist (IL-1-RA) in middle ear cholesteatoma: an analysis of protein production and biological activity. Eur Arch Otorhinolaryngol 1996, 253, 252-255.
11. Chambers, TJ. The pathobiology of the osteoclast. J Clin
Pathol 1985, 38, 241-52.

Literatur 54
12. Chambers, TJ. The regulation of osteoclastic development
and function. Ciba Found Symp 1988, 136, 92-107. 13. Chambers, TJ. Regulation of the differentiation and function
of osteoclasts. J Pathol 2000, 192, 4-13. 14. Cheer, SM, Noble S. Zoledronic acid. Drugs 2001, 61, 799-
805. 15. Cheshire, I, Blight A, Ratcliffe W, Proops D, Heath D.
Production of Parathyroid-hormone-related protein by cholesteatoma cells in culture. Lancet 1991, 338, 1041-1043.
16. Chole, R, Henry K, McGinn M. Cholesteatoma: Spontaneous
occurrence in the Mongolian gerbil Meriones unguiculatus. Am J Otol 1981, 2, 204-210.
17. Chole, RA. Cellular and subcellular events of bone resorption
in human and experimental cholesteatoma: the role of osteoclasts. Laryngoscope 1984, 94, 76-95.
18. Chole, RA. Osteoclasts in chronic otitis media,
cholesteatoma, and otosclerosis. Ann Otol Rhinol Laryngol 1988, 97, 661-6.
19. Chole, RA, Hughes RM, Faddis BT. Keratin particle-induced
osteolysis: a mouse model of inflammatory bone remodeling related to cholesteatoma. J Assoc Res Otolaryngol 2001, 2, 65-71.
20. Chung, J, Yoon T. Different production of Interleukin-1 alpha,
Interleukin-1 beta and Interleukin-8 from cholesteatomatous and normal epithelium. Acta Otolaryngol (Stockh) 1998, 118, 386-391.
21. Coxon, FP, Helfrich MH, Van't Hof R, Sebti S, Ralston SH,
Hamilton A, Rogers MJ. Protein geranylgeranylation is required for osteoclast formation, function, and survival: inhibition by bisphosphonates and GGTI-298. J Bone Miner Res 2000, 15, 1467-76.
22. Cruveilhier, J. Anatomie Pathologique du Corps Humain, Vol
1 und 2. Paris: Baillière 1829. 23. Derlacki, EL, Clemis JD. Congenital cholesteatoma of the
middle ear and mastoid. Ann Otol Rhinol Laryngol 1965, 74, 706-27.

Literatur 55
24. DuVerney, J. Traité de l'Organe de l'Ouie, Contenant
Structure: Les usages et les Maladies des Toutes les Parties de l'Oreille. Paris: E Michallet 1683, 189.
25. Feinmesser, R, Ungar H, Adler J. Otic cholesteatoma in the
sand rat (Psammomys obesus). Am J Otol 1988, 9, 409-11. 26. Fromigue, O, Lagneaux L, Body JJ. Bisphosphonates induce
breast cancer cell death in vitro. J Bone Miner Res 2000, 15, 2211-21.
27. Galvin, RJ, Bryan P, Venugopalan M, Smith DP, Thomas JE.
Calcitonin responsiveness and receptor expression in porcine and murine osteoclasts: a comparative study. Bone 1998, 23, 233-40.
28. Habermann, J. Zur Entstehung des Cholesteatoms des
Mittelohres. Arch Ohrenheilk 1888, 27, 43-51. 29. Helms, J. [The theses of the "conservative" surgery of
cholesteatoma]. Vestn Otorinolaringol 1994, 40-2. 30. Helms, J. [Restorative interventions on tympanum and
mastoid. III]. Laryngorhinootologie 1998, 77, A53-8. 31. Helms, J. [Restorative interventions of the tympanic
membrane and mastoid. II]. Laryngorhinootologie 1998, 77, A41-51.
32. Helms, J. [Restorative interventions of the tympanum and
mastoid. I]. Laryngorhinootologie 1998, 77, A29-A39. 33. Hildmann, H, Muller-Greven D. [Behavior of the middle ear
mucosa in cholesteatoma and following tympanoplasty. An experimental histological study in rabbits]. Arch Otorhinolaryngol 1977, 216, 513-4.
34. Hildmann, H, Muller-Greeven D. [The behaviour of middle
ear mucosa with cholesteatoma and after tympanoplasty]. HNO 1978, 26, 19-24.
35. Hildmann, H, Karger B, Steinbach E. [Ear ossicle transplants
for reconstruction of sound transmission in the middle ear. A histologic long-term study]. Laryngorhinootologie 1992, 71, 5-10.
36. Hoppe, F. [Proliferation behavior of cholesteatoma]. HNO
1995, 43, 710-5.

Literatur 56
37. Jagdev, SP, Coleman RE, Shipman CM, Rostami HA, Croucher PI. The bisphosphonate, zoledronic acid, induces apoptosis of breast cancer cells: evidence for synergy with paclitaxel. Br J Cancer 2001, 84, 1126-34.
38. Lange, W. Über die Entstehung der Mittelohrcholesteatoma.
Z Hals Nas Ohrenheilk 1925, 11, 250-271. 39. Lucae, A. Beiträge zur Kenntnis der Perlgeschwülste des
Felsenbeins. Arch Ohr-Nas-Kehlkhk 1873, 7, 255. 40. Macri, JR, Chole RA. Bone erosion in experimental
cholesteatoma--the effects of implanted barriers. Otolaryngol Head Neck Surg 1985, 93, 3-17.
41. Milewski, C. [Role of perimatrix fibroblasts in development of
acquired middle ear cholesteatoma. A hypothesis]. HNO 1998, 46, 494-501.
42. Müller, J. Über den feineren Bau und die Formen der
krankhaften Geschwülste. Berlin: G Reimer 1838, 50. 43. Nijweide, PJ, Burger EH, Feyen JH. Cells of bone:
proliferation, differentiation, and hormonal regulation. Physiol Rev 1986, 66, 855-86.
44. Plester, D. Hereditary factors in chronic otitis with
cholesteatoma. Acta Otorhinolaryngol Belg 1980, 34, 51-5. 45. Plester, D, Helms J, Jahnke K, Kastenbauer ER, Kley W,
Zollner C. [Artificial materials in reconstructions of the middle ear]. Laryngol Rhinol Otol (Stuttg) 1985, 64, 224-7.
46. Prallet, B, Male P, Neff L, Baron R. Identification of a
functional mononuclear precursor of the osteoclast in chicken medullary bone marrow cultures. J Bone Miner Res 1992, 7, 405-14.
47. Reinholz, GG, Getz B, Pederson L, Sanders ES,
Subramaniam M, Ingle JN, Spelsberg TC. Bisphosphonates directly regulate cell proliferation, differentiation, and gene expression in human osteoblasts. Cancer Res 2000, 60, 6001-7.
48. Ruedi, L. Pathogenesis and surgical treatment of the middle
ear cholesteatoma. Acta Otolaryngol Suppl 1979, 361, 1-45. 49. Sadé, J. Cellular differentiation in the middle ear lining. Ann
Otol Rhinol Laryngol 1971, 80, 376.

Literatur 57
50. Sadé, J, Berco E, Buyanover D, Brown M. Ossicular damage in chronic middle-ear inflammation. Acta Otolaryngol (Stockh) 1981, 92, 273-283.
51. Samura, A, Wada S, Suda S, Iitaka M, Katayama S.
Calcitonin receptor regulation and responsiveness to calcitonin in human osteoclast-like cells prepared in vitro using receptor activator of nuclear factor-kappaB ligand and macrophage colony-stimulating factor. Endocrinology 2000, 141, 3774-82.
52. Schilling, V, Negri B, Bujía J, Schulz P, Kastenbauer E.
Possible role of Interleukin 1 alpha and Interleukin 1 beta in the pathogenesis of cholesteatoma of middle ear. Am J Otol 1992, 13, 350-355.
53. Schmidt, M, Grunsfelder P, Hoppe F. Induction of matrix
metalloproteinases in keratinocytes by cholesteatoma debris and granulation tissue extracts. Eur Arch Otorhinolaryngol 2000, 257, 425-9.
54. Schmidt, M, Grunsfelder P, Hoppe F. Up-regulation of matrix
metalloprotease-9 in middle ear cholesteatoma-- correlations with growth factor expression in vivo? Eur Arch Otorhinolaryngol 2001, 258, 472-6.
55. Steinbach, E, Hildmann H. [Re-use of autologous incus in
cholesteatoma and in chronic mucosal suppuration]. Z Laryngol Rhinol Otol 1972, 51, 659-64.
56. Steinbach, E. [Experimentally produced cholesteatoma
(author's transl)]. Laryngol Rhinol Otol (Stuttg) 1978, 57, 724-33.
57. Steinbach, E, Gruninger G. Experimental production of
cholesteatoma in rabbits by using non- irritants (skin tolerants). J Laryngol Otol 1980, 94, 269-79.
58. Steinbach, E. Experimental studies on cholesteatoma
formation. Acta Otorhinolaryngol Belg 1980, 34, 56-1. 59. Steinbach, E, Pusalkar A. Long-term histological fate of
cartilage in ossicular reconstruction. J Laryngol Otol 1981, 95, 1031-9.
60. Steinbach, E, Pusalkar A, Heumann H. Cholesteatoma--
pathology and treatment. Adv Otorhinolaryngol 1988, 39, 94-106.

Literatur 58
61. Steinbach, E. [Pathogenesis of cholesteatoma]. HNO 1995,
43, 1-2. 62. Sudhoff, H, Bujia J, Borkowshi G, Koc C, Holly A, Hildmann
H, Fisseler-Eckhoff A. Basement membrane in middle ear cholesteatoma. Immunohistochemical and ultrastructural observations. Ann Otol Rhinol Laryngol 1996, 105, 804-10.
63. Sudhoff, H, Liang J, Dazert S, Borkowski G, Michaels L.
[Epidermoid formation in the pathogenesis of congenital cholesteatoma-- a current review]. Laryngorhinootologie 1999, 78, 63-7.
64. Sudhoff, H, Hildmann H, Michaels L. Pathogenesis in
Cholesteatoma. The Hague, The Netherlands, Kugler Publications, 1999.
65. Sudhoff, H, Tos M. Pathogenesis of attic cholesteatoma:
clinical and immunohistochemical support for combination of retraction theory and proliferation theory. Am J Otol 2000, 21, 786-92.
66. Sudhoff, H, Dazert S, Gonzales AM, Borkowski G, Park SY,
Baird A, Hildmann H, Ryan AF. Angiogenesis and angiogenic growth factors in middle ear cholesteatoma. Am J Otol 2000, 21, 793-8.
67. Sudhoff, H, Tos M. Pathogenesis of attic cholesteatoma:
clinical and immunohistochemical support for combination of retraction theory and proliferation theory. Am J Otol 2000, 21, 786-92.
68. Tos, M. Manual of Middle Ear Surgery. Stuttgart/New York,
Thieme Verlag, 1993. 69. Uno, Y, Saito R. Bone resorption in human cholesteatoma:
morphological study with scanning electron microscopy. Ann Otol Rhinol Laryngol 1995, 104, 463-8.
70. Virchow, R. Über Perlgeschwülste (Cholesteatoma Joh.
Müller). Virchows Arch Pathol Anat 1855, 8, 371. 71. von Schulthess, G. Die cholesteatombedingte Arrosion der
Gehörknöchelchen. Laryngologie 1962, 2, 108-114. 72. Von Tröltsch, A. The Diseases of the Ear, Their Diagnosis
and Treatment. New York NY, William Wood & Co, 1864.

Literatur 59
73. Weber, K, Hoffmann A, Leb G. [Therapy of osteoporosis:
strategies for individualized treatment]. Wien Med Wochenschr 1999, 149, 489-92.
74. Wendt, H. Desquamative Entzündung des Mittelohres
('Cholesteatom des Felsenbeins'). Arch Ohr Nasen Kehlkopfheilk 1873, 14, 428-446.
75. Wittmaack, K. Wie entsteht ein genuines Cholesteatom?
Arch Ohren Nasen Kehlkopfhk 1933, 137, 306. 76. Wolfman, DE, Chole RA. Osteoclast stimulation by positive
middle-ear air pressure. Arch Otolaryngol Head Neck Surg 1986, 112, 1037-42.
77. Wong, J, Stoney P, Hawke M. Ossicular erosion by
cholesteatoma: investigation by scanning electron microscopy utilizing a new preparation technique. J Otolaryngol 1991, 20, 216-21.
78. Yoon, TH, Lee SH, Park MH, Chung JW, Kim HJ. Inhibition
of cholesteatomatous bone resorption with pamidronate disodium. Acta Otolaryngol 2001, 121, 178-81.
79. Zenner, HP. Hals-Nasen-Ohrenheilkunde. Weinheim
(Bundesrepublik Deutschland), Chapman & Hall GmbH, 1997.

Danksagung 60
7. Danksagungen
Mein herzlicher Dank gilt allen, die mich bei der Durchführung und
Fertigstellung der Arbeit unterstützt haben.
Herrn PD Dr. Holger Sudhoff danke ich besonders herzlich für die
Überlassung des Themas, die Einführung in die Methodik und
diverse Hilfestellungen bei der Durchführung.
Herrn Prof. Dr. Hildmann danke ich für gute Ratschläge und die
freundliche Unterstützung.
Sehr herzlicher Dank gebührt Jae Jung (St. Louis, MO, USA) für
unzählige wertvolle Tipps und praktische Hilfe.
Prof. Dr. R. Chole (St. Louis, MO, USA) danke ich für die freundliche
Aufnahme in seinem Forschungslabor.
Die Arbeit wurde von der DFG (Su 226 2/1) unterstützt.
Nicht zuletzt geht ein ganz besonderer Dank an meine Frau Umay
für ihre unendliche Geduld und moralische Unterstützung.

Lebenslauf 61
8. Lebenslauf
PERSÖNLICHE DATEN: Name: Jörg Ebmeyer Geburtsdatum/-ort: 23.02.1974 in Tübingen
Ehefrau: Umay Ebmeyer, geb. Dalkesen
Eltern: Dr. med. Wolf Ebmeyer Dr. med. Ingrid Ebmeyer, geb. Brenner
Geschwister: Michael Ebmeyer
Leoni Kuhn, geb. Ebmeyer Jana Ebmeyer Lennart Ebmeyer
B BISHERIGER WERDEGANG:
1980-1984 Frölenberg Grundschule, Bielefeld 1984-1993 Ratsgymnasium Bielefeld,
Abitur am 18. Juni 1993 1993-1994 Zivildienst beim Deutschen Roten Kreuz
Bielefeld 1994-2000: Studium der Medizin an der Ruhr-Uni-
Bochum: 10.09.1996: Physikum 28.08.1997: erster Abschnitt der Ärztlichen
Prüfung 14.09.1999: zweiter Abschnitt der
Ärztlichen Prüfung 16.10.2000: dritter Abschnitt der Ärztlichen
Prüfung
26.10.2000: Approbation zum Arzt im Praktikum
11/2000 – 12/2001: Arzt im Praktikum am St. Elisabeth Hospital Bochum, HNO-Universitätsklinik, Leiter: Prof. Dr. med. H. Hildmann
seit 12/2001: Arzt im Praktikum an der HNO-Klinik der
Julius Maximilian Universität Würzburg, Leiter: Prof. Dr. med. J. Helms